Изобретение относится к биотехнологии, конкретно к генетической инженерии, и представляет собой рекомбинантный аттенуированный штамм бактерий Salmonella typhimurium Т10 рКНВс, способный продуцировать коровый антиген вируса гепатита В (HBcAg). Предложенный рекомбинантный штамм микроорганизмов является кандидатом для конструирования живой вакцины против гепатита В.
Сальмонеллы обладают естественной способностью проникать через эпителий слизистой, преимущественно с помощью М-клеток, и размножаться как в локальных интерстинальных лимфоидных образованиях, так и парентеральной лимфоидной ткани [1] . Поэтому ослабленные, но способные к инвазии штаммы сальмонелл, являются привлекательным средством доставки гетерологичных антигенов к слизистой желудочно-кишечного тракта с последующим вовлечением в процесс иммунной системе [2, 3]. Данный подход может быть использован для создания вакцин против вирусов, проникающих в организм парентеральным путем и/или через слизистую генитального и ректального тракта.
Эффективность векторов на основе Salmonella проиллюстрирована многими экспериментами на животных, в которых использовались сальмонеллы, продуцирующие белки вирусов, бактерий и простейших. Антигены, продуцированные штаммами сальмонелл, стимулируют антиген-специфичные системные IgG, Т-хелперный ответ. CD8+I-класса или CD4+II-класса CTL ответ, а также наработку секреторных антител в слизистой оболочке [4]. Одним из ярких примеров является использование штаммов сальмонелл, экспрессирующих фрагмент гена С tetanus toxsoid [5] и обеспечивающих полную защиту мышей от парентерального заражения столбнячным токсином.
Рядом авторов описаны большие вариации иммунного ответа к различным антигенам, экспрессированным в сальмонеллах. Причиной этому могут служить различные факторы, в том числе уровень экспрессии, клеточная локализация антигена, путь введения и схема вакцинации [6].
В настоящее время создан ряд различных аттенуированных штаммов сальмонелл. Эти штаммы способны инвазировать лимфоидную ткань кишечного тракта, но обнаруживают пониженную патогенность даже при больших дозах [6, 7].
Известен штамм Salmonella typhi Δ cya Δ (crp-cdt) Δ asd, обеспечивающий синтез HBcAg-preS1 белок [8, прототип]. Были исследованы иммуногенные свойства этого штамма на добровольцах. Добровольцы были вакцинированы перорально или перректально живыми рекомбинантными штаммами сальмонелл, а образцы сывороток и слизистых обследованы на наличие специфических антител к антигенам гепатита В и сальмонелл. Оказалось, что рекомбинантный штамм индуцировал высокий титр антител к липополисахаридам сальмонелл и незначительный ответ на антигены гепатита В [8].
Недостатком данной конструкции является низкий гуморальный иммунный ответ на антигены вируса гепатита В.
Технической задачей изобретения является создание на основе аттенуированного штамма Salmonella typhimurium T10 рекомбинантного штамма, обеспечивающего конститутивный синтез корового антигена вируса гепатита В и вызывающего индукцию специфических антиВГВ антител при введении животным, т.е. являющегося кандидатом в живую вакцину против гепатита В.
Известен аттенуированный штамм Salmonella typhimurium Т10 (суа, CRP), обладающий пониженной вирулентностью [9]. На мышах, телятах и обезьянах показано, что вирулентность этого штамма снижена примерно в 1000 раз по сравнению с родительским штаммом, при сохранении инвазивной активности. Этот штамм по ряду основных свойств может быть использован для решения поставленной задачи, т.е. для создания на его основе вакцины против гепатита В.
Поставленная задача решается генетической трансформацией штамма Salmonella typhimurium Т10 (суа, CRP) плазмидой рКНВс [10], обеспечивающей конститутивный синтез коврового белка вируса гепатита В (HBcAg), и при введении штамма лабораторным животным вызывающая индукцию специфических антиВГВ антител и специфического клеточного ответа.
Сконструированный штамм получил обозначение S. typhimurium Т10/рКНВс и депонирован под номером В-887 в коллекции НИИ ККМ ГНЦ ВБ "Вектор".
Культурально-морфологические признаки.
Морфология: грамотрицательные палочки, средних размеров, расположены хаотично.
Культуральные свойства: растет на мясо-пептонном агаре, глюкозоминимальном агаре Девиса, средах Мак-Конки, Левина, Кауфмана, колонии в S-формс мелких размеров через 18 ч роста при 37oС. В мясо-пептонном бульоне, желчном бульоне, жидкой глюкозоминимальной среде А (Девиса) - диффузное помутнение через 18 ч роста. В цитраминимальной среде (Девиса) - отсутствие роста.
Резистентность к антибиотикам: на всех вышеперечисленных средах растет в присутствии ампицилина (100 мкг/мл).
Антигенная структура:
O4.12Нi1.2.
Биохимические свойства.
Не утилизирует цитрат на агаре Симмонса, не продуцирует индол, не расщепляет мочевину, не обладает фенилаланилтрансаминазой, реакция Фогес-Проскауэра - отрицательная; продуцирует сероводород в течение 48 ч, обладает лизин- и орнитилкарбоксилазными активностями и аргининдегидролазной активностью; ферментирует глюкозу, не ферментирует лактозу, сахарозу, мальтозу, сорбит, инозит, дульцит, адонит; в течение 48 ч ферментирует рамнозу, маннит, трегалозу. Желатину не разжижает.
Чувствительность к бактериофагам.
Чувствителен к бактериофагу Р22.
Плазмидный профиль.
Плазмида рКНВс с молекулярной массой 6032 kb. Данная плазмида получена авторами и обеспечивает синтез белка HBcAg вируса гепатита В в клетках сальмонелл [10].
Изобретение иллюстрируется следующими графическими материалами.
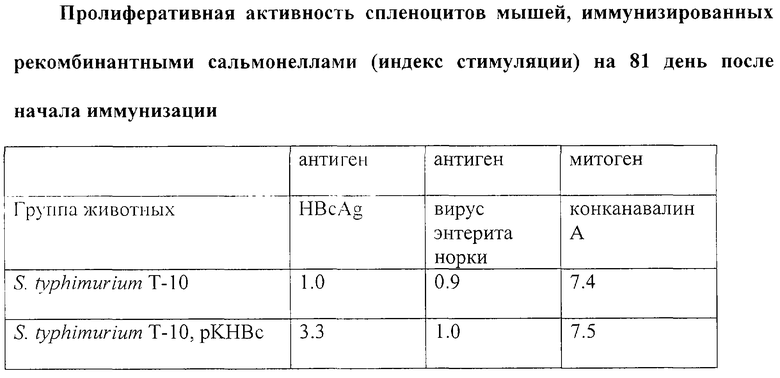
Фиг. 1. Иммуноблотинг белков Salmonella после разделения в 15% полиакриламидном геле (выявление белков с помощью кроличьей антисыворотки к денатурированному HBcAg и конъюгатом IgG козы к IgG кролика с щелочной фосфатазой).
1. Очищенный HBcAg.
2. Лизат клеток Salmonella typhimurium T10 рКНВс.
3. Лизат клеток Salmonella typhimurium T10, не содержащих плазмиду.
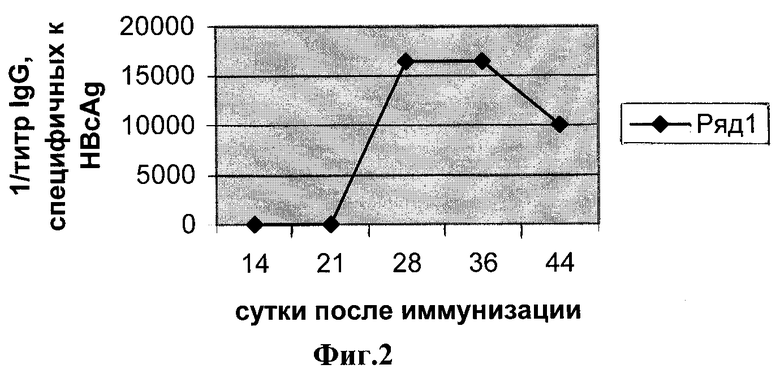
Фиг.2. Иммуноферментный анализ к HBcAg сывороток мышей, иммунизированных штаммом Salmonella typhimurium T10 рКНВс.
Для лучшего понимания сущности изобретения ниже следуют примеры его осуществления.
Пример 1.
Трансформация.
Электропорация. Культуру клеток Salmonella typhimurium T10 выращивают в среде YT до ОП=0,5, охлаждают, 1 мл культуральной жидкости центрифугируют 2 мин при 6000 об/мин на центрифуге Eppendorf. Осадок клеток ресуспендируют в 800 мкл охлажденной, бидистилированной воды и осаждают центрифугированием 1 мин при 6000 об/мин. Процедуру промывки повторяют 5 раз. Затем осадок ресуспендируют в 200 мкл 10% глицерина, центрифугируют 1 мин при 6000 об/мин и ресуспендируют в 40 мкл 10% глицерина.
К суспензии клеток добавляют 0,5 мкг плазмидной ДНК. Для электропорации используют электропоратор фирмы BIORAD. Затем добавляют по 300 мкл YT среды и инкубируют 1 ч при 37oС. Клетки рассевают на StI агар с ампициллином (50 мкг/мл). Выросшие на агаре с ампициллином колонии обозначают Salmonella typhimurium T10 рКНВс. Все манипуляции с плазмидной ДНК проводят согласно описанным методикам [11].
Культивирование
Клетки Salmonella typhimurium T10 рКНВс выращивают следующим образом: отдельную колонию сальмонеллы засевают в 50 мл среды YT, содержащей ампициллин (50 мкг/мл), и инкубируют 16 ч при 37oС; клетки осаждают центрифугированием.
Белковый электрофорез и иммунооблотинг
Клетки Salmonella typhimurium T10 рКНВс осаждают из 100 мкл культуры. Осадок растворяют в воде и добавляют 40 мкл литического буфера (1М трис-НСl рH 8,8, 5% ДСН, 20% глицерин, 10% меркаптоэтанол, 0,02% бромфеноловый синий) и выдерживают на кипящей водяной бане 5 мин. Электрофоретическое разделение белков проводят в 13%-ом полиакриламидном геле в присутствии 0,1% ДСН по Лэмли [12] . Перенос белков из геля на фильтр осуществляют по описанной методике [10] при напряжении электрического поля 10 В/см в течение 2-х ч. Фильтры помещают в 3%-ый раствор БСА в 0,01М трис-HCl, рН 7,5, 0,05% твин-20 на 1 ч, после чего инкубируют 1 ч с раствором антител кролика к HBcAg, разведенном в буфере 0,01М трис-HCl, рН 7,5, 0,15М NaCl, 0,05% твин-20, 0,05% NaN3, 0,001M ФМСФ. Этот же раствор используют для последующей отмывки фильтров и для инкубации с конъюгатом антител с фосфатозой. Результаты представлены на фиг. 1. В лизате клеток Salmonella typhimurium T10 рКНВс присутствует белок, с молекулярной массой, соответствующей рекомбинантному HBcAg и выявляемый антителами к рекомбинантному HBcAg. Это свидетельствует о продукции HBcAg в клетках Salmonella typhimurium T10 рКНВс.
Пример 2.
Культивирование
Для иммунизации клетки Salmonella typhimurium T10 рКНВс выращивают следующим образом: отдельную колонию сальмонеллы засевают в 50 мл среды YT, содержащей ампициллин (50 мкг/мл), и инкубируют 16 ч при 37oС; клетки осаждают центрифугированием и ресуспендируют в 1 мл физраствора. Концентрацию клеток определяют титрованием.
Иммунизация. Для иммунизации были использованы мыши линии BALB/c, весом 12-15 г, полученные из вивария ГНЦ ВБ "Вектор". Животные содержались на стандартном рационе.
Иммунизацию проводили двукратно, введением 107 клеток на мышь, перректально, с интервалом 3 недели. Забор крови производили через 14, 21, 28, 36 и 44 дней после первого введения сильмонелл.
Иммуноферментный анализ. Специфичность образующихся антител в сыворотках мышей анализируют с помощью ИФА. Для выявления антител против HBcAg используют очищенный рекомбинантный HBcAg. В каждом эксперименте берут образцы от трех животных. Эти образцы смешивают, после чего определяют титр специфических антител. Титр определяют как превышение титра сывороток мышей иммунизированных Salmonella typhimurium T10 рКНВс над титром сывороток мышей, иммунизированных исходным штаммом Salmonella typhimurium Т10. На каждую точку делают два повтора. Результаты представлены на фиг.2. Начиная с четвертой недели после иммунизации мышей клетками Salmonella typhimurium T10 рКНВс обнаруживается продукция IgG к HBcAg.
Реакция бласттрансформации. Реакцию бласттрансформации спленоцитов проводили через 81 день после первого введения сальмонелл по ранее описанной методике [13]. В качестве специфического антигена использовали HBcAg, в качестве неспецифического антигена - вирус энтерита норки. В лунку планшета вносили по 100 мкл каждого компонента. Концентрация антигенов была равна 2 мкг/мл. В качестве митогена использовали конканавалин А (производства "Sigma", США) в концентрации 5 мкг/мл. Клетки, стимулированные антигенами, инкубировали в течение 78 ч. За 18 ч до окончания срока инкубации вносили 3Н-тимидин (Санкт-Петербург) в дозе 2 мкКи на лунку. Клеточный ответ оценивали по показателю индекса стимуляции, который характеризует отношение пролиферации спленоцитов, индуцированных специфическими антигенами к спонтанной пролиферации спленоцитов (см таблицу). Формирование клона спленоцитов, сенсибилизированных к HBcAg, регистрировалось в группе животных, иммунизированных клетками S. typhimurium T10 pKHBc. Индекс стимуляции в группе S. typhimurium T10 pKHBc достоверно превосходил таковой как в контрольной группе S. typhimurium T10, так и при стимуляции спленоцитов гетерологичным антигеном - вирусом энтерита норок (данные представлены в таблице). Способность спленоцитов отвечать на стимуляцию HBcAg на 81-й день от начала иммунизации S. typhimurium T10 pKHBc свидетельствует о формировании специфической клеточной памяти (см. таблицу).
Таким образом. сконструированный штамм Salmonella typhimurium T10 рКНВс продуцирует HBcAg и при введении животным индуцирует выраженный специфический гуморальный и клеточный иммунный ответ к HBcAg. Предложенный рекомбинантный штамм может быть использован для конструирования живой вакцины против гепатита В.
Литература
1. Siebers A., Finlay В.В.М. Cells and the pathogenesis of mucosal and systemic infections. Trends Microbiol. 1996, 4: 22-29.
2. Hopkins S., Kraehenbuhl J-P., Schodel F. et al. A recombinant Salmonella typhimurium vaccine induces local immunity by four different routes of immunization. Infect. and Immun. 1995, 63: 3279-3286.
3. Международная заявка 98/48026, кл. MПК 6: C 12 N 15/74, опубл. 29.10.98г. и Бюллетень ИСМ, вып. 46, 10, 1999, с.124.
4. Fonts T.R., Tuskan R.G., Chada S. et al. Construction and immunogenicity of Salmonella typhimurium vaccine vectors that express HIV-1 gpl20. Vaccine. 1995, 13(17): 1697-1705.
5. Roberts M., Bacon A., Li J. et al. Prior immunity to homologous Salmonella serotypes suppresses local and systemic anti-fragment C antibody responses and protection from tetanus toxin in mice immunized with Salmonella strains expressing fragment C. Infect. and Immun. 1999, 67: 3810-3815.
6. Hormaeche C.E., Anjam Khan C.M., Mastroenii P. et al. Salmonella vaccines: mechanisms of immunity and their use as carriers of recombinant antigens. In: Ala Aldenen D.A., Hormaeche C.E. (ed.). Molecular and clinical aspects of bacterial vaccine development. England, Chichester, John Wiley& Sons Ltd., 1995, p. 119-154.
7. Международная заявка 14240, кл. МПK 6: C 12 N 15/31, опубл. 16.03.00г. и Бюллетень ИСМ, вып. 46, 3, 2001, с.17.
8. Nardelli-Haefliger D. , Kraehenbuhl J-P., Curtis R.III, Schodel F., Potts A. , Kelly S., de Grandi P. Oral and rectal immunization of adult female volunteers with a recombinant attenuated Salmonella typhi vaccine strain. Infect. Immun. 1996, Dec., p. 5219-5224.
9. Бойченко М.Н., Воробьев A.A., Тымчук С.Н. Перспектива получения мутантов суа и сrр сальмонeлл для использования в качестве вакцинных штаммов. Becтн. PAMH. 1995, (10): 37-39.
10 Карпенко Л.И., Рязанкин И.А., Чикаев Н.А., Колесникова Л.В., Ильичев А. А. N-конец корового белка вируса гепатита В спрятан внутри коровой частицы. Молекул. биология. 1997. N 31. С.919-924.
11. Маниатис Т. , Фрич Э., Сэмбрук Дж. Молекулярное клонирование. М.: Мир, 1984.
12. Luemmli E. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970. Vol. 227. P. 680-685.
13. Хоробрых В.В., Пронин A.В., Киркин А.Ф. и др. Методы постановки реакции бласт-трансформации в микромодификации. Иммyнoлогия. 1983, (3): 76-79.
Изобретение относится к биотехнологии, конкретно к генетической инженерии. Предложен новый рекомбинантный аттенуированный штамм бактерий Salmonella typhimurium T10/pKHBc, который получен генетической трансформацией штамма Salmonella typhimurium T10 плазмидой рКНВс. Штамм обеспечивает конститутивный синтез HBcAg (корового антигена вируса гепатита В). При введении лабораторным животным штамм вызывает индукцию специфических анти-ВГВ антител и специфический клеточный ответ. Новый штамм может использоваться для конструирования живой вакцины против гепатита В. 1 табл., 2 ил.
Рекомбинантный аттенуированный штамм бактерий Salmonella typhimurium, депонированный в НИИ ККМ под номером В-887, - продуцент корового антигена вируса гепатита В.
| NARDELLI-HAEFLIGER D | |||
| et al | |||
| "Oral and rectal immunization of adult female volunteers with a recombinant attenuated Salmonella typhi vaccine strain", Infect Immun., 1996, Dec; 64(12): 5219-24 | |||
| Способ крашения тканей | 1922 |
|
SU62A1 |
| US 4547367, 15.10.1985 | |||
| 0 |
|
SU251460A1 | |

