Связанные заявки
[0001] Согласно настоящей заявке испрашивается приоритет в соответствии с предварительной заявкой США №62/449228, поданной 23 января 2017 г., и предварительной заявкой США №62/541293, поданной 4 августа 2017 г. Полное содержание каждой из упомянутых выше заявок явным образом включено посредством ссылки.
[0002] Перечень последовательностей для настоящей заявки обозначен «Seq-List.txt», который создан 23 января 2018 г. и имеет размер 31 КВ. Полное содержание перечня последовательностей включено в данный документ посредством ссылки в полном объеме.
[0003] Настоящее изобретение было сделано при государственной поддержке согласно грантам №№ AI 093348, AI 056289 и AI 126712, присужденным Национальными институтами здоровья. Государство обладает определенными правами на настоящее изобретение.
Предшествующий уровень техники настоящего изобретения
[0004] Salmonella enterica вызывает значительно отягощенные заболевания у людей во всем мире. S. Typhi и Paratyphi А, В и С вызывают кишечную лихорадку (1) и представляют собой значительные угрозы для общественного здравоохранения (2-4). S. Typhi предположительно вызывает свыше 20,6 миллионов случаев, 433000 смертей в мире ежегодно (5, 6) и 12,2 миллионов лет жизни с поправкой на инвалидность (7). Помимо этих сероваров, нетифозная форма Salmonella (NTS) в последнее время все чаще распознается как основная причина инвазивных заболеваний (2, 8, 9), таких как сепсис и менингит, при этом отмечается 93,8 миллионов случаев и 681300 смертей ежегодно во всем мире (10, 11). NTS является основной причиной госпитализации и смерти в результате заболевания пищевого происхождения в США (12), ~1,2 миллионов случаев воспалительных диарейных заболеваний в год, приводя к 23000 госпитализациям и 450 смертям (12, 13) с экономическим ущербом примерно 3,31 миллиарда долларов вследствие преждевременной смертности, инвалидности, медицинских и производственных расходов и ежегодный ущерб 16782 лет жизни с поправкой на качество (14). Среди детей в возрасте <5 лет NTS представляет собой основного патогена и вызывает 4670 госпитализаций и 38 смертей (15). Заболевание, вызываемое NTS, в США главным образом объясняется сероварами, принадлежащими к трем сероварам В, D и С (16). Серовары Enteritidis, Typhimurium, Newport и Heidelberg связаны с наиболее частыми вспышками заболевания в США (17). Несмотря на то, что у подавляющего большинства пациентов развивается самоограничивающийся гастроэнтерит, характеризующийся воспалительной диареей, NTS может также вызывать системные заболевания и представляет собой единственную наиболее распространенную причину смерти в результате инфекции пищевого происхождения, ассоциированной с вирусами, паразитами или бактериями в США преимущественно у лиц с ослабленным иммунитетом (18). У детей младшего возраста и ВИЧ-инфицированных индивидуумов, NTS часто вызывает системную инфекцию, которая ассоциирована с высокой смертностью (19). Рост СПИДа во многих частях земного шара, главным образом в странах Африки к югу от Сахары, привел к существенному повышению частоты NTS-ассоциированной системной инфекции (20, 21). Бактериемия представляет собой наиболее тяжелый симптом и смертность у детей с бактериемией, которые попадают в клинику, может составлять около 25% (18, 21). Кишечная лихорадка и NTS в последнее время становятся все более сложными для лечения антибиотиками в связи с ростом множественной лекарственной резистентности Salmonella (22, 23), приводя к риску повышенного количества неизлечимых случаев (24, 25).
[0005] Кишечная лихорадка может быть предотвращена несколькими вакцинами (26, 27). Препараты убитых цельноклеточных сероваров Typhi и Paratyphi были эффективными в снижении частоты заболевания (28), однако были сняты с производства в связи с частыми побочными реакциями (29). Живой аттенуированный штамм S. Typhi Ту21а, полученный с помощью химического мутагенеза, придает лишь незначительный уровень защиты до трех лет против серовара Typhi, но не других релевантных сероваров (29, 30). Дополнительные генетически модифицированные штаммы Salmonella были исследованы в клинических испытаниях с некоторой долей успешности, однако ни один из них не был одобрен. Очищенный капсульный углевод Vi серовара Typhi индуцирует защитный иммунитет в течение нескольких лет против Typhi и возможно Paratyphi С, но не против Paratyphi А и В или Typhimurium, у всех из которых отсутствует указанная капсула (31). Конъюгация Vi с белковым антигеном повышает иммунные ответы у детей грудного возраста, основной восприимчивой к кишечной лихорадке популяции.
[0006] С целью охвата важного серовара Paratyphi А современные усилия сосредоточены на связывании О-антигена, углеводной части липополисахарида (LPS), с белковым антигеном (27). Эти две коммерческие вакцины главным образом используются для рынка вакцин для путешественников и никакой новой вакцины для широкого применения не было разрешено с 1990-х гг. (26). Несмотря на то, что три типа вакцин против S. Typhi в настоящее время являются коммерчески доступными, к сожалению, все еще отсутствует единственная разрешенная вакцина, доступная против S. Paratyphi А, при этом имеет место очень незначительная перекрестная защита, обеспечиваемая доступными вакцинами против S. Typhi, или же она отсутствует. Существуют вакцины против NTS сероваров Enteritidis и Typhimurium, которые являются эффективными у сельскохозяйственных животных, таких как домашняя птица и свиньи (32), однако недоступны у людей (33). Это свидетельствует о значительном ограничении существующих стратегий профилактики. Таким образом, лечение системного сальмонеллеза в последнее время стало более сложным, и существующие вакцины против Salmonella обеспечивают только при умеренных уровнях ограниченную продолжительность защиты и ограниченный охват клинически релевантных сероваров. Указанные ситуации создают неотложную медицинскую потребность в улучшенных вакцинах против Salmonella.
[0007] Применение рекомбинантных аттенуированных вакцин против Salmonella (RASV) в качестве вакцины или системы доставки гетерологичного антигена было изучено в связи с их способностью стимулировать системные иммунные ответы и иммунные ответы слизистой оболочки в локальных и отдаленных участках и преимуществами в качестве векторов для получения и презентации рекомбинантных вакцинных антигенов. RASV можно использовать для множества областей применения, в том числе без ограничения вакцинации против патогенов, которые вызывают заболевание, рак, хроническое респираторное заболевание и заболевания сердца. В последнее время RASV на основе регулируемых замедленных аттенуированных вакцин (RDA RASV) были разработаны для усиления иммунных ответов на RASV и переносимый защитный антиген. RDA RASV разрабатывают таким образом, что гены основных факторов вирулентности находятся под контролем индуцируемого промотора ParaBAD, индуцируемого арабинозой, не встречающейся в хозяине-млекопитающем. RDA RASV выращивают in vitro в присутствии арабинозы таким образом, что гены, опосредующие патогенный фенотип, экспрессируются, и RASV характеризуется свойствами дикого типа с целью инвазии в организм хозяев. Экспрессия патогенных генов прекращается вследствие отсутствия арабинозы in vivo, при этом продукты генов, разбавленные в результате репликации, приводят к образованию аттенуированного фенотипа, не вызывая заболевания. Поскольку они реплицируются изначально с полной вирулентностью, они колонизируют лимфоидные ткани до более высоких уровней с целью вызвать более сильные иммунные ответы, чем конститутивно аттенуированная RASV.
[0008] Для усиления иммуногенности также был разработан регулируемый замедленный синтез белка (RDPS). Повышенные уровни синтеза антигенов способствуют повышению вероятности того, что когнатные Т-клетки будут взаимодействовать с антиген-презентирующими клетками (АРС), приводя к эффективной пролиферации и продуцированию эффекторных молекул и Т-клеточной пролиферации in vivo. В то же время синтез антигенов высокого уровня накладывает метаболические требования, которые нарушают способность штаммов колонизировать эффекторные лимфоидные ткани. Система RDPS вызывает продуцирование рекомбинантного вакцинного антигена только после того, как RASV колонизирует лимфоидные ткани, поскольку клетки с RASV размножаются in vivo. На данную стратегию не влияет путь аттенуации.
[0009] Несмотря на то, что применение рекомбинантной Salmonella в виде живых вакцин с целью получения иммунного ответа у субъектов является перспективным, указанные организмы являются живыми и иногда патогенными. Соответственно, необходимо вводить регуляторные системы в бактерии для аттенуации и контроля экспрессии антигенов, которые экспрессируются указанными бактериями. Используемые в настоящее время средства аттенуации делают живые вакцинные штаммы восприимчивыми к средовым стрессовым факторам in vivo. Соответственно, меньше бактерий способны колонизировать клетку хозяина с целью достижения необходимого уровня иммуногенности. Таким образом, имеется потребность в новых штаммах рекомбинантных микроорганизмов, которые могут быть разработаны в виде живых вакцин, которые являются менее восприимчивыми к средовым стрессовым факторам in vivo и которые могут колонизировать клетки-хозяева с целью достижения более высоких уровней иммуногенности. Также существует потребность в новых средствах усиления безопасности живых аттенуированных вакцин in vivo.
Краткое раскрытие настоящего изобретения
[0010] В настоящем раскрытии предусмотрены штаммы рекомбинантных бактерий, в том числе Salmonella, которые зависят от трех сахаров для регуляции вирулентного фенотипа бактерий в результате контроля экспрессии нескольких генов вирулентности и антигена, представляющего интерес, а также регулируемого замедленного литического фенотипа, обеспечивая биологическое ограничение и усиление иммуногенных свойств. Другие свойства, которые можно регулировать с помощью одного или нескольких из сахаров, включают в себя кислотоустойчивость (например, во время пероральной иммунизации), как описано в публикации заявки на патент США №2014/0370057, полное содержание которой явным образом включено в данный документ посредством ссылки. Зависимость от трех сахаров усиливает безопасность рекомбинантных бактерий, учитывая невероятность того, что организмы будут иметь место со всеми тремя сахарами во встречающемся в природе среде. Неожиданным образом настоящее изобретение демонстрирует, что три различные сахара могли быть использованы для регуляции свойств рекомбинантных бактерий (например, экспрессии генов, кодирующих антиген, представляющий интерес, замедленного литического фенотипа и/или экспрессии генов вирулентности) без перекрестной интерференции какого-либо сахара в регулируемой сахарами активности любого другого сахара в результате катаболитной репрессии.
[0011] Указанные организмы могут быть использованы для безопасной и высокоэффективной доставки антигенных соединений субъекту с целью повышения эффективных защитных иммунных ответов. Такие рекомбинантные бактерии могут воздействовать на защитные антигены, синтезируемые клеточной поверхностью, и могут индуцировать защитные иммунные ответы с целью размножения сероваров Salmonella. Рекомбинантные бактерии могут быть использованы для усиления выживаемости бактерий в ответ на защитные стрессовые факторы хозяина, такие как желудочная кислота; для придания регулируемой замедленной аттенуации; для придания регулируемого замедленного лизиса in vivo (например, с помощью контроля экспрессии генов asdA и murA с высвобождением антигена, представляющего интерес, или ДНК-вакцины, кодирующей их); или для обеспечения слияния углеводных полимеров с углеводами и/или белками.
[0012] В частности, в данном документе раскрыты регулируемые тремя сахарами штаммы рекомбинантных аттенуированных вакцинных штаммов Salmonella (RASV). Эти штаммы доставляют несколько консервативных защитных поверхностных/секретируемых антигенов Salmonella с их природными конформациями с целью индукции защитного иммунитета против нескольких вирулентных сероваров Salmonella. В качестве примера RASV могут иметь регулируемый рамнозой синтез О-антигена в сочетании с регулируемым маннозой синтезом боковой цепи О-антигена с целью демонстрации консервативного внутреннего кора и регулируемым арабинозой продуцированием генерализованных модулей мембранных антигенов (GMMA) или везикул внешней мембраны in vivo с целью повышения продуцирования консервативных белков внешней мембраны (ОМР). RASV могут быть сконструированы в двух сероварах Salmonella, группе В S. Typhimurium и группе D S. Enteritidis для экспрессии консервативных иммуногенных генов с целью сведения к максимуму гуморальных, клеточных иммунных ответов и иммунных ответов слизистой оболочки против Salmonella. Раскрываемые RASV характеризуются рациональными конструктивными свойствами, отличающимися от других RASV, что повышает успешное применение. В частности, раскрываемые RASV обеспечивают безопасные и высокоэффективные вакцины против Salmonella низкой стоимостью и могут быть использованы для разработки RASV против S. Typhi или S. Paratyphi А для применения у человека.
[0013] В соответствии с одним аспектом в настоящем раскрытии предусмотрено рекомбинантное производное патогенной бактерии, содержащее а.) первый ген, регулируемый первым сахаром, который придает первый фенотип; b.) второй ген, регулируемый вторым сахаром, который придает второй фенотип; и с.) третий ген, регулируемый третьим сахаром, который придает третий фенотип; при этом первый, второй и третий фенотипы выбирают из группы, состоящей из: 1. регулируемой замедленной аттенуации; 2. регулируемой замедленной экспрессии антигена, представляющего интерес; 3. регулируемого замедленного лизиса in vivo; 4. регулируемого синтеза О-антигена; 5. регулируемого синтеза боковой цепи О-антигена; 6. регулируемого продуцирования генерализованных модулей мембранных антигенов (GMMA); 7. регулируемой повышенной выживаемости в ответ на стрессовое для хозяина условие; и 8. регулируемого продуцирования везикул внешней мембраны (OMV).
[0014] В соответствии с одним аспектом в настоящем раскрытии предусмотрено рекомбинантное производное патогенной бактерии, содержащее а.) первый ген, регулируемый первым сахаром, который придает первый фенотип; b.) второй ген, регулируемый вторым сахаром, который придает второй фенотип; и с.) третий ген, регулируемый третьим сахаром, который придает третий фенотип; при этом первый, второй и третий фенотипы выбирают из группы, состоящей из: 1. регулируемой замедленной аттенуации; 2. регулируемой замедленной экспрессии антигена, представляющего интерес; 3. регулируемого замедленного лизиса in vivo; 4. регулируемого синтеза О-антигена; 5. регулируемого продуцирования генерализованных модулей мембранных антигенов (GMMA); 6. регулируемой повышенной выживаемости в ответ на стрессовое для хозяина условие; и 7. регулируемого продуцирования везикул внешней мембраны (OMV).
[0015] В соответствии с одним вариантом осуществления каждый из первого сахара, второго сахара и третьего сахара представляют собой разный сахар. В соответствии с одним вариантом осуществления первый сахар, второй сахар или третий сахар не нарушает регуляцию гена, регулируемого другим сахаром.
[0016] В соответствии с одним вариантом осуществления первый сахар выбирают из группы, состоящей из арабинозы, маннозы, ксилозы, галактозы, рамнозы и мальтозы. В соответствии с одним вариантом осуществления второй сахар выбирают из группы, состоящей из арабинозы, маннозы, ксилозы, галактозы, рамнозы и мальтозы. В соответствии с одним вариантом осуществления третий сахар выбирают из группы, состоящей из арабинозы, маннозы, ксилозы, галактозы, рамнозы и мальтозы.
[0017] В соответствии с одним вариантом осуществления первый ген является функционально связанным с первым регулируемым сахаром промотором. В соответствии с одним вариантом осуществления второй ген является функционально связанным с вторым регулируемым сахаром промотором. В соответствии с одним вариантом осуществления третий ген является функционально связанным с третьим регулируемым сахаром промотором.
[0018] В соответствии с одним вариантом осуществления ген модифицируют с целью обеспечения обратимого синтеза сахарсодержащей молекулы, которая придает регулируемый сахаром фенотип. В соответствии с одним вариантом осуществления модифицированный ген представляет собой pmi. В соответствии с одним вариантом осуществления модифицированный ген представляет собой galE.
[0019] В соответствии с одним вариантом осуществления бактерия представляет собой грамотрицательную бактерию. В соответствии с одним вариантом осуществления бактерия принадлежит семейству Enterobacteriaceae.
[0020] В соответствии с одним вариантом осуществления фенотип представляет собой регулируемую замедленную аттенуацию, а ген, придающий указанный фенотип, представляет собой fur. В соответствии с одним вариантом осуществления фенотип представляет собой регулируемую замедленную экспрессию антигена, представляющего интерес, а ген, придающий указанный фенотип, кодирует антиген, представляющий интерес. В соответствии с одним вариантом осуществления фенотип представляет собой регулируемый замедленный лизис in vivo, при этом осуществление лизиса обеспечивается в цитозоле вследствие мутации в гене sifA. В соответствии с одним вариантом осуществления фенотип представляет собой регулируемый синтез О-антигена, а ген, придающий указанный фенотип, выбирают из группы, состоящей из waaG, rfaH, waaJ, wbaP, wzy, waaP, waaO, waaF, waaP, waaC, waaA, waaL и wbaP. В соответствии с одним вариантом осуществления фенотип представляет собой продуцирование генерализованных модулей мембранных антигенов (GMMA) или везикул внешней мембраны, а ген, придающий указанный фенотип, выбирают из группы, состоящей из ybgC, tolQ, tolA, tolR, tolB, paI и ybgF.
[0021] В соответствии с одним вариантом осуществления фенотип представляет собой регулируемый синтез боковой цепи О-антигена, а ген, придающий указанный фенотип, представляет собой tolR. В соответствии с одним вариантом осуществления первый фенотип представляет собой регулируемый синтез О-антигена, а второй фенотип представляет собой продуцирование GMMA или везикул внешней мембраны.
[0022] В соответствии с одним вариантом осуществления бактерия дополнительно содержит ген, кодирующий антиген, представляющий интерес, функционально не связанный с регулируемым сахаром промотором.
[0023] В соответствии с одним вариантом осуществления бактерия содержит делецию эндогенного гена синтеза О-антигена. В соответствии с одним вариантом осуществления делеция эндогенного гена синтеза О-антигена представляет собой частичную делецию указанного гена. В соответствии с одним вариантом осуществления делеция в эндогенном гене синтеза О-антигена представляет собой полноразмерную делецию указанного гена. В соответствии с одним вариантом осуществления ген синтеза О-антигена представляет собой waaL или wbaP.
[0024] В соответствии с одним вариантом осуществления бактерия содержит делецию в эндогенном гене фосфоманнозоизомеразы. В соответствии с одним вариантом осуществления делеция эндогенного гена фосфоманнозоизомеразы представляет собой частичную делецию указанного гена. В соответствии с одним вариантом осуществления делеция в эндогенном гене фосфоманнозоизомеразы представляет собой полноразмерную делецию указанного гена. В соответствии с одним вариантом осуществления ген фосфоманнозоизомеразы представляет собой pmi.
[0025] В соответствии с одним вариантом осуществления бактерия содержит делецию в эндогенном гене системы tol-pal. В соответствии с одним вариантом осуществления делеция в эндогенном гене системы tol-pal представляет собой частичную делецию указанного гена. В соответствии с одним вариантом осуществления делеция в эндогенном гене системы tol-pal представляет собой полноразмерную делецию указанного гена. В соответствии с одним вариантом осуществления эндогенный ген системы tol-pal выбирают из группы, состоящей из ybgC, tolQ, tolA, tolR, tolB, paI и ybgF.
[0026] В соответствии с одним вариантом осуществления первый ген, второй ген и/или третий ген расположены в плазмиде в бактерии. В соответствии с одним вариантом осуществления первый ген, второй ген и/или третий ген расположены в хромосоме в бактерии.
[0027] В соответствии с одним вариантом осуществления первый, второй или третий регулируемый сахаром промотор представляет собой регулируемый рамнозой промотор. В соответствии с одним вариантом осуществления регулируемый рамнозой промотор представляет собой rhaSR PrhaBAD».
[0028] В соответствии с одним вариантом осуществления первый, второй или третий регулируемый сахаром промотор представляет собой регулируемый арабинозой промотор. В соответствии с одним вариантом осуществления регулируемый арабинозой промотор представляет собой araC ParaBAD.
[0029] В соответствии с одним вариантом осуществления бактерия дополнительно содержит делецию в эндогенном гене relA. В соответствии с одним вариантом осуществления делеция эндогенного гена relA собой частичную делецию указанного гена. В соответствии с одним вариантом осуществления делеция эндогенного гена relA собой полноразмерную делецию указанного гена.
[0030] В соответствии с одним вариантом осуществления бактерия дополнительно содержит нуклеиновую кислоту, кодирующую репрессор LacI. В соответствии с одним вариантом осуществления репрессор LacI кодируется геном lacI. В соответствии с одним вариантом осуществления нуклеиновая кислота, кодирующая репрессор LacI расположена в плазмиде в бактерии. В соответствии с одним вариантом осуществления нуклеиновая кислота, кодирующая репрессор LacI расположена в хромосоме в бактерии.
[0031] В соответствии с одним вариантом осуществления бактерия дополнительно содержит делецию в эндогенном промоторе Pfur.
[0032] В соответствии с одним вариантом осуществления ген fur является функционально связанным с регулируемым арабинозой промотором. В соответствии с одним вариантом осуществления ген fur расположен в плазмиде в бактерии. В соответствии с одним вариантом осуществления ген fur расположен в хромосоме в бактерии.
[0033] В соответствии с одним вариантом осуществления бактерия дополнительно содержит делецию в гене, кодирующем аспартат-полуальдегиддегидрогеназу. В соответствии с одним вариантом осуществления ген, кодирующий аспартат-полуальдегиддегидрогеназу, представляет собой ген asd. В соответствии с одним вариантом осуществления ген, кодирующий аспартат-полуальдегиддегидрогеназу, представляет собой ген asdA.
[0034] В соответствии с одним вариантом осуществления ген, кодирующий антиген, представляющий интерес, расположен в плазмиде в бактерии. В соответствии с одним вариантом осуществления плазмида дополнительно содержит нуклеиновую кислоту, кодирующую аспартат-полуальдегиддегидрогеназу. В соответствии с одним вариантом осуществления аспартат-полуальдегиддегидрогеназа представляет собой AsdA. В соответствии с одним вариантом осуществления плазмида представляет собой низкокопийную плазмиду. В соответствии с одним вариантом осуществления плазмида представляет собой высококопийную плазмиду. В одном варианте осуществления плазмиду выбирают из группы, состоящей из pYA4589, pYA4595, pYA4763, pG8R15, pG8R16, pG8R17, pG8R18, pGR111, pG8R112, pG8R113 и pG8R114.
[0035] В соответствии с одним вариантом осуществления ген, кодирующий антиген, представляющий интерес, расположен в хромосоме в бактерии.
[0036] В соответствии с одним вариантом осуществления бактерия дополнительно содержит делецию в гене pagL. В соответствии с одним вариантом осуществления делеция гена pagL собой частичную делецию указанного гена. В соответствии с одним вариантом осуществления делеция гена pagL собой полноразмерную делецию указанного гена. В соответствии с одним вариантом осуществления мутация представляет собой ΔwaaL/ΔpagL::TT rhaSR PrhaBAD waaL.
[0037] В соответствии с одним вариантом осуществления бактерия дополнительно содержит антиген, представляющий интерес, функционально связанный с регулируемым репрессором промотором. В соответствии с одним вариантом осуществления промотор представляет собой регулируемый лактозой промотор. В соответствии с одним вариантом осуществления регулируемый лактозой промотор представляет собой LacI-регулируемый промотор. В соответствии с одним вариантом осуществления LacI-регулируемый промотор выбирают из группы, состоящей из Ptrc, Plac, PT7lac, Ptac, PompA lacO и Plpp lacO.
[0038] В соответствии с одним вариантом осуществления антиген, представляющий интерес, представляет собой антиген, происходящий из инфекционного агента. В соответствии с одним вариантом осуществления антиген, представляющий интерес, происходит из инфекционного агента, выбранного из группы, состоящей из вируса, бактерии, простейшего, приона, гриба и гельминта. В соответствии с одним вариантом осуществления антиген, представляющий интерес, происходит из бактерии. В соответствии с одним вариантом осуществления антиген, представляющий интерес, представляет собой антиген Salmonella. В соответствии с одним вариантом осуществления антиген Salmonella выбирают из группы, состоящей из FliC, FHC180, OmpC, OmpD, OmpF, SseB и SseI. В соответствии с одним вариантом осуществления антиген, представляющий интерес, представляет собой антиген из бактерии Clostridium. В соответствии с одним вариантом осуществления антиген представляет собой антиген С. perfringens. В соответствии с одним вариантом осуществления антиген представляет собой NetB, PlcC, их антигенные фрагменты, слитые белки, содержащие указанные антигены, или слитые белки, содержащие антигенные фрагменты антигенов.
[0039] В соответствии с одним вариантом осуществления антиген, представляющий интерес, представляет собой вирусный антиген. В соответствии с одним вариантом осуществления антиген, представляющий интерес, представляет собой антиген вируса гриппа. В соответствии с одним вариантом осуществления антиген вируса гриппа представляет собой гемагглютинин или нейраминидазу.
[0040] В соответствии с одним вариантом осуществления антиген, представляющий интерес, ассоциирован с раком. В соответствии с одним вариантом осуществления антиген, ассоциированный с раком, выбирают из группы, состоящей из MAGE-A, MAGE-C1, BAGE, GAGE, CAGE, XAGE, NY-ESO1, LAGE1 и сурвивина.
[0041] В соответствии с одним вариантом осуществления указанный антиген представляет собой белковый антиген, кодируемый последовательностью нуклеиновой кислоты, кодон-оптимизированной в отношении экспрессии в указанной бактерии.
[0042] В соответствии с одним вариантом осуществления бактерия дополнительно содержит делецию в гене sifA. В соответствии с одним вариантом осуществления делеция гена sifA собой частичную делецию указанного гена. В соответствии с одним вариантом осуществления делеция гена sifA собой полноразмерную делецию указанного гена. В соответствии с одним вариантом осуществления ген sifA является функционально связанным с регулируемым арабинозой промотором.
[0043] В соответствии с одним вариантом осуществления бактерия дополнительно содержит делецию в гене recF. В соответствии с одним вариантом осуществления делеция гена recF собой частичную делецию указанного гена. В соответствии с одним вариантом осуществления делеция гена recF собой полноразмерную делецию указанного гена.
[0044] В соответствии с одним вариантом осуществления бактерия дополнительно содержит делецию в гене recJ. В соответствии с одним вариантом осуществления делеция гена recJ собой частичную делецию указанного гена. В соответствии с одним вариантом осуществления делеция гена recJ собой полноразмерную делецию указанного гена.
[0045] В соответствии с одним вариантом осуществления бактерия принадлежит роду Salmonella. В соответствии с одним вариантом осуществления бактерия представляет бактерию Salmonella enterica. В соответствии с одним вариантом осуществления бактерия представляет собой бактерию Salmonella enterica subsp. enterica серовар Paratyphi А, бактерию Salmonella enterica subsp.enterica серовар Enteritidis, бактерию Salmonella enterica subsp. enterica серовар Typhi, бактерию Salmonella enterica subsp. enterica серовар Typhimurium, Salmonella enterica subsp. enterica серовар Dublin или Salmonella enterica subsp. enterica серовар Choleraesuis.
[0046] В соответствии с другим аспектом в данном документе раскрыта фармацевтическая композиция, содержащая рекомбинантную бактерию, раскрываемую в данном документе, и фармацевтически приемлемый носитель.
[0047] В соответствии с другим аспектом в данном документе раскрыт способ вызова иммунного ответа против антигена, представляющего интерес, у субъекта, при этом способ предусматривает введение субъекту эффективного количества фармацевтической композиции, раскрываемой в данном документе.
[0048] Другие аспекты и пункты настоящего изобретения описаны более подробно ниже.
Краткое описание чертежей
[0049] На Фиг. 1. изображены три вектора, содержащие регулируемые сахаром кассеты araC ParaBAD, phaRS-PrhaBAD и xylR-PxylA, для того, чтобы способствовать тому, чтобы конструкция производного суицидного вектора образовывала слияния регулируемой сахаром кассеты с геном, представляющим интерес, с целью замещения нативного промотора этим геном, представляющим интерес.
[0050] На Фиг. 2 изображены три плазмиды, в которых синтез GFP регулируется тремя различными сахарами.
[0051] На Фиг. 3А, 3В и 3С изображена мутация невосприимчивости к галактозе Δ(galE-ybhC)-851.
[0052] На Фиг. 4А, 4В, 4С, 4D, 4Е, 4F, 4G и 4Н изображены кривые роста штаммов Salmonella с другими мутациями galE в питательном бульоне с варьирующими концентрациями галактозы.
[0053] На Фиг. 5 изображена колонизация мутантов galE.
[0054] На Фиг. 6А, 6В, 6С, 6D, 6Е, 6F, 6G, 6Н, 6I, 6J, 6K, 6L, 6М, 6N, 6O, 6Р, 6Q и 6R изображены кривые роста штаммов Salmonella χ12341(pYA4763) и χ3761 в течение 24 часов в питательной среде с варьирующими указанными концентрациями сахара.
Подробное раскрытие настоящего изобретения
[0055] Для более легкого понимания настоящего раскрытия сначала определяются определенные термины. Эти определения необходимо трактовать с точки зрения оставшейся части настоящего раскрытия и как понимается специалистом в данной области. Если не определено иное, все технические и научные термины, используемые в настоящем документе, имеют такое же значение, как и обычно понимаемые специалистом в данной области. Дополнительные определения изложены по всему подробному описанию.
[0056] Используемый в данном документе термин «рекомбинантная бактерия» относится к бактериальной клетке, которая была генетически модифицирована по отношению к своему нативному состоянию. Например, рекомбинантная бактерия может содержать одну или несколько нуклеотидных вставок, нуклеотидных делеций, нуклеотидных реаранжировок и нуклеотидных модификаций. Указанные генетические модификации могут быть введены в хромосому бактерии или в альтернативном случае могут присутствовать во внехромосомной нуклеиновой кислоте (например, плазмиде). Рекомбинантные бактерии по настоящему раскрытию могут содержать нуклеиновую кислоту, расположенную в плазмиде. В альтернативном случае рекомбинантные бактерии могут содержать нуклеиновую кислоту, расположенную в бактериальной хромосоме (например, стабильно включенную в них). В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия является авирулентной. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия характеризуется сниженной вирулентностью. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия является невирулентной. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия является патогенной. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия является аттенуированной. В соответствии с другим вариантом осуществления рекомбинантная бактерия представляет собой рекомбинантное производное патогенной бактерии.
[0057] Используемый в данном документе термин «ген» относится к фрагменту нуклеиновой кислоты, который кодирует белок или его фрагмент или функциональную или структурную молекулу РНК и может необязательно содержать регуляторную последовательность, предшествующую (5' некодирующие последовательности) и расположенную после (3' некодирующие последовательности) кодирующей последовательности нуклеиновой кислоты. В соответствии с некоторыми вариантами осуществления «ген» не содержит регуляторные последовательности, предшествующие или расположенные после кодирующей последовательности.
[0058] В соответствии с одним вариантом осуществления ген представляет собой гетерологичный ген. В соответствии с другим вариантом осуществления нуклеиновая кислота представляет собой гетерологичную нуклеиновую кислоту. Используемые в данном документе термины «гетерологичный ген» или «гетерологичная нуклеиновая кислота» относятся к гену или последовательности нуклеиновой кислоты, присутствующих в рекомбинантной клетке, например, бактерии, которые обычно не встречаются в клетке дикого типа, например, бактерии, в природе. В соответствии с другим вариантом осуществления гетерологичный ген или гетерологичную нуклеиновую кислоту вводят в определенную клетку экзогенно. В соответствии с другим вариантом осуществления гетерологичный ген может представлять собой ген или его фрагменты, вводимые в не встречающуюся в природе клетку хозяина. В соответствии с другим вариантом осуществления термин «гетерологичный ген» включает в себя вторую копию нативного гена или его фрагмента, которые были введены в клетку хозяина помимо соответствующего нативного гена. Гетерологичная нуклеиновая кислота может также включать в себя в соответствии с некоторыми вариантами осуществления последовательность гена, которая встречается в природе в определенной клетке, однако которая была модифицирована, например, в результате регуляции с помощью другой промоторной последовательности, с целью экспрессии несвойственного количества нуклеиновой кислоты и/или полипептида, который она кодирует; и/или двух или более последовательностей нуклеиновой кислоты, которые не встречаются в том же самом отношении по отношению друг к другу в природе.
[0059] Используемый в данном документе термин «эндогенный ген» относится к нативному гену, который присутствует в своем естественном положении в геноме организма (например, бактериальной хромосоме).
[0060] Термин «промотор», используемый в данном документе, относится к последовательности нуклеиновой кислоты, которая способна контролировать экспрессию кодирующей последовательности гена. Промотор может содержать одну или несколько специфических последовательностей регуляции транскрипции с целью дополнительного усиления экспрессии и/или изменения пространственной и/или временной экспрессии нуклеиновой кислоты. Например, промотор может содержать одну или несколько нуклеиновых кислот, которые специфически распознаются белком-активатором транскрипции (например, энхансерным элементом), белком-репрессором транскрипции и т.п. Термин «функционально связанный», используемый в данном документе, означает, что экспрессия последовательности нуклеиновой кислоты находится под контролем промотора, с которым он пространственно соединен. Промотор может быть расположен в направлении 5' (выше) от последовательности нуклеиновой кислоты, находящейся под его контролем. Расстояние между промотором и последовательностью нуклеиновой кислоты, подлежащей экспрессии, может быть примерно таким же, что и расстояние между промотором и нативной последовательностью нуклеиновой кислоты, которую он контролирует. Как известно в данной области техники, вариация этого расстояния может быть адаптирована без потери функции промотора. Последовательности нуклеиновой кислоты промоторов, описанных в данном документе, известны в данной области техники и способы функционального связывания этих промоторов с геном (например, геном, кодирующим репрессор) известны в данной области техники.
[0061] В соответствии с некоторыми вариантами осуществления промотор для применения, описанного в данном документе, может регулироваться прямо или непрямо с помощью сахара. Например, в соответствии с некоторыми вариантами осуществления промотор является восприимчивым к уровню арабинозы, в других случаях называемый в данном документе «регулируемый арабинозой промотор». В самом общем виде арабиноза может присутствовать во время роста бактерии in vitro, при этом в типичном случае отсутствует в ткани хозяина. В соответствии с одним вариантом осуществления промотор происходит из системы araC-ParaBAD) Escherichia coli. Система araC ParaBAD представляет собой высокорегулируемую систему экспрессии, которая, как было показано, выступает в качестве сильного промотора, индуцируемого добавлением низких уровней арабинозы. Промотор araC-araBAD представляет собой двунаправленный промотор, контролирующий экспрессию последовательностей нуклеиновой кислоты araBAD в одном направлении и последовательность нуклеиновой кислоты araC в другом направлении.
[0062] Для удобства участок промотора araC-araBAD, который опосредует экспрессию последовательностей нуклеиновой кислоты araBAD и который контролируется продуктом последовательности нуклеиновой кислоты araC, называется в данном документе ParaBAD. Для применения, описанного в данном документе, может быть использована кассета с последовательностью нуклеиновой кислоты araC и промотором araC-araBAD. Указанная кассета называется в данном документе araC ParaBAD. Белок AraC представляет собой как положительный, так и отрицательный регулятор ParaBAD. В присутствии арабинозы белок AraC представляет собой положительный регуляторный элемент, который обеспечивает экспрессию из ParaBAD. В отсутствие арабинозы белок AraC приводит к репрессии экспрессии из ParaBAD. Другие кишечные бактерии содержат регулируемые арабинозой системы, гомологичные системе araC-araBAD Е. coli, в том числе, например, S. Typhimurium. Например, белок AraC Е. coli активирует только ParaBAD Е. coli (в присутствии арабинозы), но не ParaBAD S. Typhimurium. Таким образом, регулируемый арабинозой промотор может быть использован только в рекомбинантной бактерии, которая обладает аналогичным арабинозным опероном, без значительной интерференции между ними двумя, если промотор и оперон происходит от двух различных видов бактерий. В самом общем виде концентрация арабинозы, необходимая для индукции экспрессии, в типичном случае составляет менее чем приблизительно 2% (мас./мас.) в среде для культивирования. В соответствии с некоторыми вариантами осуществления концентрация составляет менее чем приблизительно 1,5%, 1%, 0,5%, 0,2%, 0,1% или 0,05% (мас./мас.) в среде для культивирования. В соответствии с другими вариантами осуществления концентрация составляет 0,05% или ниже, например, приблизительно 0,04%, 0,03%, 0,02% или 0,01% (мас./мас.). В соответствии с иллюстративным вариантом осуществления концентрация составляет приблизительно 0,05% (мас./мас.) в среде для культивирования.
[0063] В соответствии с другими вариантами осуществления промотор может быть восприимчивым к уровню мальтозы в среде, в других случаях называемый в данном документе «регулируемый мальтозой промотор». В соответствии с некоторыми вариантами осуществления рекомбинантных бактерий, описанных в данном документе, культивируют в среде, содержащей мальтозу. Ген malT кодирует MalT, положительный регулятор четырех восприимчивых к мальтозе промоторов (PpQ, PEFG, PKBM и PS). Комбинация malT и промотора mal создает высокорегулируемую систему экспрессии, которая, как было показано, выступает в качестве сильного промотора, индуцируемого в присутствии мальтозы. В отличие от системы araC-ParaBAD, экспрессия malT регулируется промотором (т.е., PT), который функционально не связан с другими промоторами mal. PT не регулируется MalT. Промотор malEFG-malKBM представляет собой двунаправленный промотор, который контролирует экспрессию последовательностей нуклеиновой кислоты malKBM в одном направлении и последовательностей нуклеиновой кислоты malEFG в другом направлении. Для удобства участок промотора malEFG-malKBM, который опосредует экспрессию последовательности нуклеиновой кислоты malKBM и который контролируется MalT, называется в данном документе PKBM, а участок промотора malEFG-malKBM, который опосредует экспрессию последовательности нуклеиновой кислоты malEFG и который контролируется MalT, называется в данном документе PEFG. Полная индукция PKBM требует присутствия MalT-связывающих сайтов Pefg. Для применения в векторах и системах, описанных в данном документе, генная кассета, содержащая последовательность нуклеиновой кислоты, кодирующей MalT и промотор mal может быть использована. Указанная генная кассета называется в данном документе malT-Pmal. В присутствии мальтозы белок MalT представляет собой положительный регуляторный элемент, который обеспечивает экспрессию, опосредуемую Pmal. В самом общем виде концентрация мальтозы, необходимая для индукции экспрессии, в типичном случае составляет менее чем приблизительно 1% (мас./мас.) в среде для культивирования. В соответствии с некоторыми вариантами осуществления концентрация составляет менее чем приблизительно 1,0%, 0,9%, 0,8%, 0,7%, 0,6%, 0,5%, 0,4%, 0,3% 0,2%, 0,1% или 0,05% (мас./мас.) в среде для культивирования. В соответствии с другими вариантами осуществления концентрация составляет 0,05% или ниже, например, приблизительно 0,04%, 0,03%, 0,02% или 0,01% (мас./мас.). В соответствии с иллюстративным вариантом осуществления концентрация составляет от приблизительно 0,2% до приблизительно 0,4% (мас./мас.) в среде для культивирования.
[0064] В соответствии с еще одними вариантами осуществления промотор, используемый в данном документе, может быть восприимчивым к уровню рамнозы в среде, в других случаях называемый в данном документе «регулируемый рамнозой промотор». По аналогии с системой araC-ParaBAD, описанной выше, система активатора-промотора rhaRS-PrhaB тщательно регулируется рамнозой. Экспрессия из промотора рамнозы (Prha) индуцируется до высоких уровней в присутствии рамнозы. В соответствии с некоторыми вариантами осуществления бактерии культивируют в присутствии рамнозы. Рамноза широко распространена у бактерий, но редко встречается у субъектов-людей. Оперон rhaBAD контролируется промотором PrhaBAD. Указанный промотор регулируется двумя активаторами, RhaS и RhaR, а соответствующие последовательности нуклеиновой кислоты принадлежат к одной транскрипционной единице, которая расположена в противоположном направлении от последовательностей нуклеиновой кислоты rhaBAD. В присутствии L-рамнозы RhaR связывается с промотором PrhaRS и активирует продуцирование RhaR и RhaS. RhaS совместно с L-рамнозой, в свою очередь, связывается с промоторами PrhaBAD и PrhaT и активирует транскрипцию структурных последовательностей нуклеиновых кислот. Полная индукция регулируемых арабинозой, мальтозой и рамнозой промоторов, описанных в данном документе, требует связывания комплекса Crp-сАМР, который является основным регулятором катаболической репрессии.
[0065] Несмотря на то, что как L-арабиноза, так и L-рамноза выступает непосредственно в качестве индукторов экспрессии регулонов, которые опосредуют их катаболизм, существуют важные различия в отношении регуляторных механизмов. L-арабиноза выступает в качестве индуктора с активатором AraC в положительном контроле арибинозного регулона. В то же время L-рамнозный регулон подвергается действию регуляторного каскада, и, тем самым, подвергается действию даже более высокого контроля, чем система araC-ParaBAD. L-рамноза выступает в качестве индуктора с активатором RhaR для синтеза RhaS, который, в свою очередь, выступает активатором в положительном контроле рамнозного регулона. В соответствии с настоящим раскрытием рамноза может быть использована для взаимодействия с белком RhaR, а затем белок RhaS может активировать транскрипцию последовательности нуклеиновой кислоты, функционально связанной С промотором PrhaBAD.
[0066] В соответствии с еще одними вариантами осуществления промотор может быть восприимчивым к уровню ксилозы в среде, называемый в данном документе «регулируемый ксилозой промотор». Как правило, концентрации ксилозы от 0,0002% до 0,63% (мас./мас.) в среде активируют экспрессию индуцируемого ксилозой промотора, описанного в данном документе (см., например, Bhavsar et al. (2001) Арр. Environ. Microbiol. 67(1): 403-10(34)). Система xylR-PxylA представляет собой хорошо известную индуцируемую систему активатора-промотора. Ксилоза индуцирует ксилозо-специфические опероны (например, xylE, xylFGHR и xylAB), которые регулируются XylR и системой циклического АМР-Crp. Белок XylR выступает в качестве положительного регулятора в результате связывания с двумя различными участками промоторов последовательности нуклеиновой кислоты xyl. Как и в случае системы araC-ParaBAD, описанной выше, могут быть использованы регуляторные системы xylR-PxylAB и /or xylR-PxylFGH. В соответствии с этими вариантами осуществления ксилоза, взаимодействующая с белком XylR, активирует транскрипцию последовательностей нуклеиновой кислоты, функционально связанных с этими двумя промоторами Pxyl.
[0067] Используемый в данном документе термин «экзогенный» относится к веществу (например, нуклеиновой кислоты или полипептиду), присутствующему в клетке, отличной от его природного источника. Термин «экзогенный» может относиться к нуклеиновой кислоте или белку, которые были введены с помощью способа, включающего участие человека в биологической системе, такой как клетка или организм, в которой они обычно не встречаются или в которой они встречаются в не определяемых количествах. Вещество может считаться экзогенным, если его вводят в клетку или предшественник клетки, который унаследует указанное вещество. В отличие от этого, термин «эндогенный» относится к веществу, которое является нативным по отношению к биологической системе или клетке.
[0068] Термин «фармацевтическая композиция», используемый в данном документе, относится к композиции, содержащей активный ингредиент (например, рекомбинантную бактерию, описанную в данном документе) с другими компонентами, такими как физиологически пригодный носитель и/или наполнитель.
[0069] Используемый в данном документе термин «фармацевтически приемлемый носитель» или «фармацевтически приемлемый наполнитель» относится к фармацевтически приемлемому веществу, композиции или основе, такой как жидкий или твердый наполнитель, разбавитель, наполнитель, вспомогательное средство (например, скользящее вещество, тальк, стеарат магния, кальция или цинка или стерическая кислота) или вещество для инкапсулирования растворителя, участвующего в переносе или транспорте исследуемого соединения из одного органа или части организма другому органу или части организма. Каждый носитель должен быть «приемлемым» с точки зрения совместимости с другими ингредиентами состава и не вредным для пациента. Некоторые примеры веществ, которые выступают в качестве фармацевтически приемлемых носителей, включают в себя: (1) сахара, такие как лактоза, глюкоза и сахароза; (2) крахмалы, такие как кукурузный крахмал и картофельный крахмал; (3) целлюлозу и ее производные, такие как кар боксиметил целлюлозу натрия, метилцеллюлозы, этилцеллюлозу, микрокристаллическую целлюлозу и ацетатцеллюлозу; (4) порошкообразный трагакант; (5) солод; (6) желатин; (7) скользящие средства, такие как стеарат магния, лаурилсульфат натрия и тальк; (8) наполнители, такие как масло какао и суппозиторные воска; (9) масла, такие как арахисовое масло, хлопковое масло, сафлоровое масло, кунжутное масло, оливковое масло, кукурузное масло и соевое масло; (10) гликоли, такие как пропиленгликоль; (11) полиолы, такие как глицерин, сорбит, маннит и полиэтиленгликоль (PEG); (12) сложные эфиры, такие как этилолеат и этиллаурат; (13) агар; (14) буферные средства, такие как гидроксид магния и гидроксид алюминия; (15) альгиновую кислоту; (16) апирогенную воду; (17) изотонический солевой раствор (например, фосфатно-буферный солевой раствор (PBS)); (18) раствор Рингера; (19) этиловый спирт; (20) рН-буферные растворы; (21) сложные полиэфиры, поликарбонаты и/или полиангидриды; (22) объемообразующие средства, такие как полипептиды и аминокислоты; (23) компонент сыворотки крови, такой как сывороточный альбумин, HDL и LDL; (24) С2-С12 спирты, такие как этанол; и (25) другие нетоксичные совместимые вещества, используемые в фармацевтических составах. Увлажняющие средства, красители, средства для регуляции высвобождения, покрывающие средства, средства для улучшения распадаемости таблеток, связующие вещества, подсластители, ароматизаторы, отдушки, ингибиторы протеаз, пластификаторы, эмульгаторы, стабилизирующие средства, загустители, пленкообразующие средства, солюбилизирующие средства, поверхностно-активные вещества, консерванты и антиоксид анты также могут присутствовать в составе. Термины, такие как «наполнитель», «носитель», «фармацевтически приемлемый наполнитель» и т.п. используются в данном документе взаимозаменяемо.
[0070] «Плазмида» или «вектор» включают в себя конструкцию на основе нуклеиновой кислоты, сконструированную для доставки в клетку хозяина или переноса между различными клетками хозяина. Нуклеиновая кислота, включенная в плазмиду, может быть функционально связана с последовательностью контроля экспрессии в случае, когда последовательность контроля экспрессии контролирует и регулирует транскрипцию и трансляцию этой полинуклеотидной последовательности.
[0071] Используемые в данном документе термины «белок» и «полипептид» используются в данном документе взаимозаменяемо с целью обозначения последовательности аминокислотных остатков, соединенных друг с другом с помощью пептидных связей между альфа-амино- и карбоксигруппами соседних остатков. Термины «белок» и «полипептид» относятся к полимеру из аминокислот, в том числе модифицированных аминокислот (например, фосфорилированных, гликированных, гликозилированных и т.д.) и аналогов аминокислот вне зависимости от их размера и функции. Термины «белок» и «полипептид», используемые в данном документе, относятся как к большим полипептидам, так и к малым пептидам. Термины «белок» и «полипептид» используются в данном документе взаимозаменяемо при обозначении продукта гена или его фрагментов. Таким образом, иллюстративные полипептиды или белки включают в себя продукты генов, встречающиеся в природе белки, гомологи, ортологи, паралоги, фрагменты и другие эквиваленты, варианты, фрагменты и аналоги вышеуказанного.
[0072] «Нуклеиновая кислота» или «последовательность нуклеиновой кислоты» может представлять собой любую молекулу, предпочтительно полимерную молекулу, содержащую единицы рибонуклеиновой кислоты, дезоксирибонуклеиновой кислоты или их аналог. Нуклеиновая кислота может как однонитевой, так и двунитевой. Однонитевая нуклеиновая кислота может представлять собой одну нить нуклеиновой кислоты из денатирурованной двунитевой ДНК. В альтернативном случае она может представлять собой однонитевую нуклеиновую кислоту, не происходящую из какой-либо двунитевой ДНК. В соответствии с одним аспектом нуклеиновая кислота может представлять собой ДНК. В соответствии с другим аспектом нуклеиновая кислота может представлять собой РНК. Подходящей молекулой нуклеиновой кислоты является ДНК, в том числе геномная ДНК или кДНК. Другие подходящие молекулы нуклеиновой кислоты представляют собой РНК, в том числе мРНК, рРНК и тРНК.
[0073] Изменения нативной аминокислотной последовательности могут быть выполнены с помощью любого количества методик, известных специалисту в данной области техники. Мутации могут быть введены, например, в определенных локусах с помощью синтеза олигонуклеотидов, содержащих мутантную последовательность, фланкированную сайтами рестрикции, обеспечивая лигирование с фрагментами нативной последовательности. После лигирования полученная в результате реконструированная последовательность кодирует аналог, имеющий необходимую аминокислотную вставку, замену или делецию. В альтернативном случае процедуры олигонуклеотид-направленного сайт-специфического мутагенеза могут быть использованы для получения измененной нуклеотидной последовательности, имеющей определенные кодоны в соответствии с необходимой заменой, делецией или вставкой. Методики получения таких изменений хорошо известны и включают в себя, например, таковые, раскрытые Walder et al. (35); Bauer et al. (36); Craik (37); Smith et al. (38); и патентами США №№4518584 и 4737462, которые включены в данный документ посредством ссылки в полном объеме. Любой цистеиновый остаток, не участвующий в поддержании надлежащей конформации полипептида, также может быть замещен, как правило, серином, для повышения устойчивости к окислению молекулы и предупреждения аберрантного сшивания. В отличие от этого, цистеиновая(цистеиновые) связь(связи) может(могут) быть добавлена(добавлены) к полипептиду с целью повышения его стабильности или облегчения олигомеризации.
[0074] Термин «статистически значимый» или «значимо» относится к статистической значимости и, как правило, означает разницу в два стандартные отклонения (2SD) или больше.
[0075] Используемый в данном документе термин «клетка хозяина» относится к клетке в организме, в которую рекомбинантную бактерию вводят с целью, например, индукции иммунного ответа. В соответствии с одним вариантом осуществления хозяин представляет собой птицу, лошадь или человека, а клетка хозяина относится соответственно к клетке птицы, клетке лошади или клетке человека.
[0076] За исключением того, что представлено в рабочих примерах, или в случае, если указано иное, все числа, выражающие количества ингредиентов или условия реакций, используемые в настоящем документе, необходимо понимать как модифицированные во всех случаях с помощью термина «приблизительно». Термин «приблизительно» при использовании в связи с процентами, может означать ±1%.
[0077] Формы существительного единственного числа необходимо понимать как означающие «по меньшей мере одно», если четко не указано противоположное.
[0078] Фраза «и/или» при использовании между элементами в перечне, означает либо (1) то, что только один приведенный элемент присутствует, либо (2) то, что более одного элемента перечня присутствует. Например, «А, В и/или С» означает, что выбор может представлять собой только А; только В; только С; А и В; А и С; В и С; или А, В и С. Фраза «и/или» может быть использована взаимозаменяемо с фразой «по меньшей мере одно из» или «одно или более из» элементов в перечне.
[0079] Диапазоны, предусматриваемые в данном документе, подразумеваются для всех значений в диапазоне. Например, диапазон от 1 до 50 включает в себя любое число, комбинацию из чисел или поддиапазон из группы, состоящей из 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49 или 50.
I. Рекомбинантные бактерии
[0080] В настоящем раскрытии предусмотрена в соответствии с некоторыми вариантами осуществления рекомбинантная бактерия, способная регулировать экспрессию по меньшей мере одной последовательности нуклеиновой кислоты, кодирующей ген, представляющий интерес. Рекомбинантная бактерия, описанная в данном документе, является особенно эффективной в вызове иммунного ответа (например, защитного иммунитета) против антигена, представляющего интерес, поскольку указанная бактерия содержит множественные рекомбинантные регуляторные системы, которые позволяют бактерии реплицироваться при введении и колонизировать лимфоидные ткани у субъекта с целью вызова сильных иммунных ответов. В то же время после нескольких циклов репликации in vivo бактерия в конечном итоге характеризуется аттенуированным фенотипом, который обеспечивает безопасное введение субъекту, например, вакцинной композиции. Рекомбинантные регуляторные системы бактерий, описанных в данном документе, зависят отчасти от множественных генетических регуляторных элементов, которые являются восприимчивыми к одному или нескольким сахарам (например, арабинозе, рамнозе, маннозе, мальтозе, ксилозе и галактозе), которые недоступны бактерии in vivo. Таким образом, фенотип рекомбинантных бактерий, описанных в данном документе, может быть изменен при введении субъекту. В соответствии с некоторыми вариантами осуществления субъекту вводят один или несколько сахаров до введения, после введения или совместно с введением рекомбинантной бактерии, описанной в данном документе, с целью активации и/или репрессии восприимчивой к сахару регуляторной системе бактерий. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия, описанная в данном документе, содержит по меньшей мере три регуляторные системы, при этом каждая из них зависит от разного сахара, что облегчает исходную инвазию клетки-хозяина в организме субъекта, замедленную аттенуацию и повышенную иммуногенность.
[0081] В соответствии с некоторыми вариантами осуществления рекомбинантную бактерию, описанную в данном документе, можно регулировать в отношении замедленной аттенуации in vivo. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия, описанная в данном документе, способна регулировать замедленную экспрессию нуклеиновой кислоты, кодирующей антиген, представляющий интерес. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия, описанная в данном документе, характеризуется регулируемым продуцированием генерализованных модулей мембранных антигенов (GMMA) или везикул внешней мембраны in vivo, которое может приводить к повышенному продуцированию консервативных белков внешней мембраны, присутствующих в бактерии и в конечном итоге повышенной иммуногенности. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия, описанная в данном документе, способна как к регулируемой экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей по меньшей мере один антиген, представляющий интерес, так к и регулируемой аттенуации. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия, описанная в данном документе, способна как к регулируемой экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей по меньшей мере один антиген, представляющий интерес, так к и регулируемому продуцированию GMMA или везикул внешней мембраны in vivo. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия, описанная в данном документе, способна как к регулируемому продуцированию GMMA или везикул внешней мембраны in vivo, так и к регулируемой аттенуации. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия, описанная в данном документе, способна к регулируемой экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей по меньшей мере один антиген, представляющий интерес, регулируемой аттенуации и регулируемому продуцированию GMMA или везикул внешней мембраны in vivo. В соответствии с некоторыми вариантами осуществления каждое из этих свойств прямо или косвенно регулируется с помощью избытка по меньшей мере одного сахара (например, арабинозы, рамнозы, маннозы, ксилозы, мальтозы и галактозы).
[0082] В соответствии с некоторыми вариантами осуществления бактерия, описанная в данном документе, представляет собой грамотрицательную бактерию. В соответствии с некоторыми вариантами осуществления бактерия представляет собой патогенную бактерию. В соответствии с некоторыми вариантами осуществления бактерия представляет собой авирулентную бактерию. В соответствии с некоторыми вариантами осуществления бактерия принадлежит семейству Enterobacteriaceae. В соответствии с некоторыми вариантами осуществления бактерия принадлежит роду, выбранному из: Alterococcus, Aquamonas, Aranicola, Arsenophonus, Brenneria, Budvicia, Buttiauxella, Candidatus Phlomobacter, Cedeceae, Citrobacter, Edwardsiella, Enterobacter, Erwinia, Escherichia, Ewingella, Hafnia, Klebsiella, Kluyvera, Leclercia, Leminorella, Moellerella, Morganella, Obesumbacterium, Pantoea, Pectobacterium, Photorhabdus, Plesiomonas, Pragia, Proteus, Providencia, Rahnella, Raoultella, Salmonella, Samsonia, Serratia, Shigella, Sodalis, Tatumella, Trabulsiella, Wigglesworthia, Xenorhbdus или Yersinia, Yokenella. В соответствии с некоторыми вариантами осуществления бактерия представляет собой патогенный вид Enterobaceteriaceae. В соответствии с некоторыми вариантами осуществления бактерию выбирают из группы, состоящей из Escherichia coli, Shigella, Edwardsiella, Salmonella, Citrobacter, Klebsiella, Enterobacter, Serratia, Proteus, Morganella, Providencia и Yersinia. В соответствии с некоторыми вариантами осуществления бактерия принадлежит роду Salmonella. В соответствии с некоторыми вариантами осуществления бактерия принадлежит роду Yersinia. В соответствии с некоторыми вариантами осуществления бактерия принадлежит роду Edwardsiella. В соответствии с некоторыми вариантами осуществления бактерия принадлежит роду, виду или штамму, широко используемому в качестве живой или аттенуированной вакцины.
[0083] В соответствии с некоторыми вариантами осуществления в настоящем раскрытии предусмотрен вид или подвид род Salmonella (например, S. enterica или S. bongori). Например, рекомбинантная бактерия может представлять собой серовар Salmonella enterica, в то числе, например, Paratyphi A, Enteritidis, Typhi, и Typhimurium. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия принадлежит серовару S. Typhimurium, S. Typhi, S. Paratyphi, S. Gallinarum, S. Enteritidis, S. Choleraesius, S. Arizonae, S. Newport, S. Heidelberg, S. Infantis, S. Cholerasiuis или S. Dublin.
[0084] Рекомбинантная бактерия, происходящая из Salmonella, может быть особенно пригодной для применения в качестве вакцины. Например, пероральное инфицирование штаммом Salmonella в типичном случае приводит к колонизации лимфоидной ткани, ассоциированной с кишечником (GALT), или пейеровых бляшек, что приводит к индукции генерализованного иммунного ответа слизистой оболочки против рекомбинантной бактерии. Дальнейшее проникновении указанной бактерии в мезентериальные лимфатические узлы, печень и селезенку может приводить к усилению индукции системных и клеточных иммунных ответов, направленных против указанной бактерии. Таким образом, применение рекомбинантной Salmonella для пероральной иммунизации приводит к стимуляции всех трех веток иммунной системы, что является особенно важным для иммунизации против агентов инфекционных заболеваний, которые колонизируются на поверхностях слизистой оболочки и/или проникают через них. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия, описанная в данном документе, используется для индукции иммунного ответа у домашней птицы (например, в виде вакцины). При использовании у домашней птицы рекомбинантная бактерия может быть введена с помощью спрея и, тем самым, инокулировать лимфоидную ткань, ассоциированную с конъюнктивой (CALT), посредством воздействия на глаза, назально-ассоциированую лимфоидную ткань (NALT) и лимфоидную ткань, ассоциированную с бронхами (BALT), посредством воздействия на дыхательные пути, и GALT посредством перорального воздействия. В соответствии с некоторыми вариантами осуществления рекомбинантную бактерию, описанную в данном документе, вводят только что вылупившимся цыплятам.
А. Аттенуация
[0085] В соответствии с некоторыми вариантами осуществления рекомбинантную бактерию, описываемую в данном документе, модифицируют таким образом, что экспрессию одного или нескольких генов, например, генов вирулентности, можно регулировать восприимчивым к сахару путем. В соответствии с некоторыми вариантами осуществления один или несколько эндогенных генов, например, генов вирулентности, удаляют из бактериальной хромосомы. В соответствии с некоторыми вариантами осуществления делеция представляет собой частичную делецию эндогенного гена. В соответствии с некоторыми вариантами осуществления делеция представляет собой полноразмерную делецию эндогенного гена. В соответствии с некоторыми вариантами осуществления ген, например, ген вирулентности, генетически изменяют с целью предупреждения транскрипции и/или трансляции гена, кодирующего белок. В соответствии с некоторыми вариантами осуществления эндогенный ген генетически изменяют с целью вставки терминатора транскрипции в открытую рамку считывания гена. В соответствии с некоторыми вариантами осуществления регуляторную область гена, например, гена вирулентности, генетически модифицируют с целью изменения (например, снижения) экспрессии гена. В соответствии с некоторыми вариантами осуществления промотор гена, например, гена вирулентности, изменяют с целью включения одного или нескольких регуляторных элементов (например, восприимчивого к сахару промотора).
[0086] В соответствии с некоторыми вариантами осуществления рекомбинантную бактерию, описываемую в данном документе, модифицируют с целью включения нуклеиновой кислоты, содержащей ген. В соответствии с некоторыми вариантами осуществления рекомбинантную бактерию модифицируют с целью включения нуклеиновой кислоты, содержащей ген, с помощью которого эндогенная копия гена в бактериальной хромосоме была изменена и/или удалена. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген, который на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентичен эндогенному гену в бактериальной хромосоме, который был удален и/или изменен. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген, который на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% гомологичен эндогенному гену в бактериальной хромосоме, который был удален и/или изменен. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген от вида, подвида, серовара или штамма бактерии, который отличается от вида бактерии рекомбинантной бактерии.
[0087] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген от вида, подвида, серовара или штамма бактерии, который является тем же самым, что и вид бактерии рекомбинантной бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген, который является функционально связанным с регулируемым промотором (например, регулируемым сахаром промотором). В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген, который является функционально связанным с регулируемым рамнозой промотором, регулируемым ксилозой промотором, регулируемым галактозой промотором, регулируемым арабинозой промотором, регулируемым маннозой промотором или регулируемым мальтозой промотором. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая указанный ген, расположена в плазмиде в бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая указанный ген, расположена в бактериальной хромосоме. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая указанный ген, расположена в локусе хромосомы, соответствующем локусу эндогенного гена, который был удален или изменен в бактериальной хромосоме. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота является кодон-оптимизированной (например, с целью повышения экспрессии нуклеиновой кислоты в рекомбинантной бактерии).
1. Гены синтеза О-антигена
[0088] В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит делецию в эндогенном гене синтеза О-антигена. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит делецию в эндогенном гене О-антиген-лигазы. В соответствии с некоторыми вариантами осуществления делеция представляет собой частичную делецию эндогенного гена О-антиген-лигазы. В соответствии с некоторыми вариантами осуществления делеция представляет собой полноразмерную делецию эндогенного гена О-антиген-лигазы. В соответствии с некоторыми вариантами осуществления эндогенный ген О-антиген-лигазы генетически изменяют с целью вставки терминатора транскрипции в открытую рамку считывания гена. В соответствии с некоторыми вариантами осуществления регуляторную область эндогенного гена О-антиген-лигазы генетически модифицируют с целью изменения (например, снижения) экспрессии гена. В соответствии с некоторыми вариантами осуществления промотор эндогенного гена О-антиген-лигазы изменяют с целью включения одного или нескольких регуляторных элементов (например, восприимчивого к сахару промотора). В соответствии с некоторыми вариантами осуществления промотор эндогенного гена О-антиген-лигазы изменяют с целью повышения расстояния между последовательностью Шайна-Дальгарно и старт-кодоном гена. В соответствии с некоторыми вариантами осуществления промотор эндогенного гена О-антиген-лигазы изменяют с целью снижения расстояния между последовательностью Шайна-Дальгарно и старт-кодоном гена. В соответствии с некоторыми вариантами осуществления последовательность Шайна-Дальгарно (SD), старт-кодон, второй кодон и/или третий кодон гена О-антиген-лигазы изменяют для повышения частоты адениновых нуклеиновых оснований с целью повышения эффективности трансляции гена. В соответствии с некоторыми вариантами осуществления последовательность Шайна-Дальгарно (SD), старт-кодон, второй кодон и/или третий кодон гена О-антиген-лигазы изменяют для снижения частоты адениновых нуклеиновых оснований с целью снижения эффективности трансляции гена. В соответствии с некоторыми вариантами осуществления ген О-антиген-лигазы представляет собой waaL (также известный как rfaL). О-антиген-лигаза waaL необходима для лигирования полисахарида с фрагментом кора LPS липидом А. Делеция waaL приводит к образованию интактного кора LPS с липидом А с отсутствием О-антигена или отдельных Сахаров, присоединенных к нему. В соответствии с некоторыми вариантами осуществления ген О-антиген-лигазы выбирают из группы, состоящей из waaG (также известного как rfaG), waal (также известного как rfaI), rfaH, waaJ (также известного как rfaJ), wbaP (также известного как rfbP), wzy (также известного как rfc), waaP, waaQ, waaF, waaP, waaC и waaA.
[0089] В соответствии с некоторыми вариантами осуществления рекомбинантную бактерию, описываемую в данном документе, модифицируют с целью включения нуклеиновой кислоты, содержащей ген О-антиген-лигазы. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая указанный ген О-антиген-лигазы, расположена в плазмиде в бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая указанный ген О-антиген-лигазы, расположена в хромосоме в бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген О-антиген-лигазы, расположена в локусе хромосомы, соответствующем локусу эндогенного гена О-антиген-лигазы, который был удален или изменен в бактериальной хромосоме. В соответствии с некоторыми вариантами осуществления рекомбинантную бактерию модифицируют с целью включения нуклеиновой кислоты, содержащей ген О-антиген-лигазы, с помощью которого эндогенная копия гена в бактериальной хромосоме была изменена и/или удалена. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген О-антиген-лигазы Salmonella.
[0090] Последовательность нуклеиновой кислоты иллюстративного гена waaL Salmonella представлена ниже:


[0091] Аминокислотная последовательность белка WaaL, кодируемого нуклеиновой кислотой под SEQ ID NO: 1, представлена ниже:

[0092] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген waaL Salmonella (представлен в виде SEQ ID NO: 1). В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген waaL, при этом ген waaL содержит последовательность нуклеиновой кислоты, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична последовательности нуклеиновой кислоты под SEQ ID NO: 1. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген waaL, при этом ген waaL содержит последовательность нуклеиновой кислоты, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% гомологична последовательности нуклеиновой кислоты под SEQ ID NO: 1.
[0093] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит последовательность нуклеиновой кислоты, кодирующую О-антиген-лигазу, где указанная О-антиген-лигаза содержит аминокислотную последовательность, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности под SEQ ID NO: 2. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит последовательность нуклеиновой кислоты, кодирующую О-антиген-лигазу, где указанная О-антиген-лигаза содержит аминокислотную последовательность, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% гомологична аминокислотной последовательности под SEQ ID NO: 2.
[0094] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген О-антиген-лигазы от вида, подвида, серовара или штамма бактерии, который отличается от вида бактерии рекомбинантной бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген О-антиген-лигазы от вида, подвида, серовара или штамма бактерии, который является тем же самым, что и вид бактерии рекомбинантной бактерии.
[0095] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген О-антиген-лигазы, который является функционально связанным с регулируемым промотором (например, регулируемым сахаром промотором). В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген О-антиген-лигазы (например, waaL), который является функционально связанным с регулируемым сахаром промотором. Предпочтительно рекомбинантные бактериальные штаммы, содержащие нуклеиновую кислоту, содержащую ген О-антиген-лигазы (например, waaL), который является функционально связанным с регулируемым сахаром промотором, будут синтезировать нормальный LPS в присутствии указанного сахара (например, рамнозы) in vitro, однако будут образовывать шероховатый LPS in vivo вследствие отсутствия сахара, который активирует промотор и, тем самым, экспрессию О-антиген-лигазы. Не ограничиваясь какой-либо определенной теорией, с помощью указанной стратегии бактерия будет демонстрировать консервативный кор LPS и характеризоваться повышенным продуцированием консервативных белков внешней мембраны (ОМР; например, поринов), которые могут приводить к повышенной иммуногенности и способствовать перекрестному иммунному ответу против антигена, представляющего интерес, синтезируемого в бактерии in vivo. В соответствии с некоторыми вариантами осуществления регулируемый сахаром промотор характеризуется повышенной активностью (например, повышенной транскрипцией) в присутствии конкретного сахара и сниженной активностью в отсутствие сахара. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген О-антиген-лигазы, который является функционально связанным с регулируемым рамнозой промотором (например, регулируемым сахаром промотором). В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген О-антиген-лигазы, который является функционально связанным с регулируемым арабинозой промотором (например, регулируемым сахаром промотором). В соответствии с некоторыми вариантами осуществления применение регулируемого рамнозой промотора (например, rhaSR PrhaBAD) может быть предпочтительным по сравнению с регулируемым арабинозой промотором, поскольку требуется сравнительно более высокая концентрация для активации регулируемого арабинозой промотора по сравнению с регулируемым рамнозой промотором (см., например, Giacalone et al. (2006) BioTechniques 40(3): 355-366 (39), полное содержание которой включено в данный документ посредством ссылки). В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит мутацию ΔwaaL/ΔpagL::TT rhaSR PrhaBAD waaL.
2. Гены деацилазы липида А
[0096] В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит делецию в эндогенном гене деацилазы липида А. В соответствии с некоторыми вариантами осуществления делеция представляет собой частичную делецию эндогенного гена деацилазы липида А. В соответствии с некоторыми вариантами осуществления делеция представляет собой полноразмерную делецию эндогенного гена деацилазы липида А. В соответствии с некоторыми вариантами осуществления эндогенный ген деацилазы липида А генетически изменяют с целью вставки терминатора транскрипции в открытую рамку считывания гена. В соответствии с некоторыми вариантами осуществления регуляторную область эндогенного гена деацилазы липида А генетически модифицируют с целью изменения (например, снижения) экспрессии гена. В соответствии с некоторыми вариантами осуществления промотор эндогенного гена деацилазы липида А изменяют с целью включения одного или нескольких регуляторных элементов (например, восприимчивого к сахару промотора). В соответствии с некоторыми вариантами осуществления ген деацилазы липида А представляет собой pagL. Было обнаружено, что бактерии, содержащие ген деацилазы липида A pagL продуцировали повышенные количества везикул внешней мембраны (см., например, Elhenawy et al. (2016) mBio 7(4): e00940-16 (40)). Делеция гена pagL Salmonella не приводит к нарушению вирулентности бактерий (see, e.g., Man et al. Proc. Nat'l. Acad. Sci. USA 111: 7403-8 (41)). He ограничиваясь какой-либо определенной теорией, в соответствии с некоторыми вариантами осуществления рекомбинантная бактерия, описанная в данном документе, содержит одну или несколько генетических модификаций, которые приводят к повышенной везикуляции (т.е., повышенному образованию везикул), что может быть частично предпочтительным в вызове иммунного ответа против антигена, представляющего интерес, который экспрессируется бактерией.
3. Гены фосфоманнозоизомеразы
[0097] В соответствии с некоторыми вариантами осуществления бактерия содержит делецию в эндогенном гене фосфоманнозоизомеразы. Фосфоманнозоизомераза, также известная как манноза-6-фосфат-изомераза, катализирует обратимое взаимопревращение фруктозо-6-фосфата в манноза-6-фосфат. Манноза-6-фосфат затем превращается в GDP-маннозу и используется для синтеза боковых цепей О-антигена. Бактерии с делециями гена фосфоманнозоизомеразы pmi являются невосприимчивыми к маннозе и частично аттенуированными (см., например, Collins et al. (1991) Infect. Immun. 59(3): 1079-85 (42)). Эти мутанты no pmi синтезируют уровни дикого типа боковых цепей LPS О-антигена при росте в средах, содержащих маннозу и одновременно являются аттенуированными, но высокоиммуногенными (см., например, Curtiss et al. (2007) "Induction of host immune responses using Salmonella-vectored vaccines." в Brogden KA, Minion FC, Cornick N, Stanton ТВ, Zhang Q, Nolan LK, Wannemuehler MJ, ed. Virulence Mechanisms of Bacterial Pathogens. 4th ed. Washington DC: ASM Press (43)). В соответствии с некоторыми вариантами осуществления делеция эндогенного гена фосфоманнозоизомеразы представляет собой частичную делецию. В соответствии с некоторыми вариантами осуществления делеция эндогенного гена фосфоманнозоизомеразы представляет собой полноразмерную делецию. В соответствии с некоторыми вариантами осуществления эндогенный ген фосфоманнозоизомеразы генетически изменяют с целью вставки терминатора транскрипции в открытую рамку считывания гена. В соответствии с некоторыми вариантами осуществления регуляторную область эндогенного гена фосфоманнозоизомеразы генетически модифицируют с целью изменения (например, снижения) экспрессии гена фосфоманнозоизомеразы. В соответствии с некоторыми вариантами осуществления промотор эндогенного гена фосфоманнозоизомеразы изменяют с целью включения одного или нескольких регуляторных элементов (например, восприимчивого к сахару промотора). В соответствии с некоторыми вариантами осуществления ген фосфоманнозоизомеразы представляет собой pmi.
[0098] В соответствии с некоторыми вариантами осуществления бактерия содержит делецию гена pmi. В соответствии с некоторыми вариантами осуществления бактерия содержит мутацию Δpmi-2426. Бактерия, содержащая мутацию Δpmi-2426, выращенная в присутствии маннозы, способна синтезировать полный О-антиген LPS. Нефосфорилированная манноза, которая представляет собой форму, требующуюся для захвата бактериями, недоступна in vivo. Таким образом, бактерия, содержащая мутацию Δpmi-2426, утрачивает способность синтезировать боковые цепи, специфические в отношении О-антигена LPS, in vivo, и число боковых цепей О-антигена, присоединенных к кору LPS, снижается приблизительно наполовину после каждого клеточного деления in vivo. LPS, который синтезируется, содержит коровую структуру, которая по сути аналогична у всех серотипов Salmonella enterica, за исключением S. Arizona. Это приводит к образованию бактерии, которая способна вызывать иммунный ответ против по меньшей мере двух серотипов Salmonella, при этом по сути не индуцируя иммунный ответ, специфический в отношении серотипа бактериального вектора. В соответствии с некоторыми вариантами осуществления указанная бактерия способна вызывать иммунный ответ против всех серотипов Salmonella, при этом по сути не индуцируя иммунный ответ, специфический в отношении серотипа бактериального вектора.
[0099] Рекомбинантная бактерия, описанная в данном документе, которая содержит делецию в мутации pmi, может также содержать другие мутации, которые способствуют тому, что манноза, доступная для бактерии во время роста in vitro, используется для синтеза О-антигена LPS. Например, бактерия может содержать мутацию Δ(gmd-fcl)-26. Эта мутация приводит к делеции двух последовательностей нуклеиновых кислот, которые кодируют ферменты для превращения GDP-маннозы в GDP-фукозу, способствуя тому, что манноза, доступная для бактерии во время роста in vitro, используется для синтеза О-антигена LPS, а не для продуцирования колановой кислоты. Аналогично, бактерия может содержать мутацию Δ(wcaM-wza)-8, которая приводит к делеции всех 20 последовательностей нуклеиновой кислоты, необходимых для продуцирования колановой кислоты, а также препятствует превращению GDP-маннозы в GDP-фукозу.
4. Гены UDP-галактозоэпнмеразы
[00100] UDP-Gal представляет собой предшественник для сборки боковых цепей О-антигена LPS, внешнего кора LPS, для колановой кислоты и других полисахаридных полимеров, имеющих галактозу в качестве компонента (44). UDP-Gal синтезируется посредством превращения глюкозо-1-Р в UDP-Glu с помощью фермента глюкозо-1-Р-уридилтрансферазы, кодируемого геном galU, при этом UDP-Glu превращался в UDP-Gal с помощью фермента UDP-галактозоэпимеразы, кодируемого геном galE (45, 46). Штаммы, выращенные в присутствии галактозы, могут синтезировать UDP-Gal в результате другого пути, в котором галактоза после захвата превращается в галактозо-1-Р с помощью галактозокиназы, кодируемой геном galK, который в свою очередь превращается в UDP-Gal с помощью фермента UDP-Gal-трансферазы, кодируемого геном galT (45). Штаммы с мутацией galE неспособны синтезировать внешней кор LPS и О-антиген LPS до тех пор, пока галактоза не поступает в среду для роста (47). В связи с указанными фактами и свойствами штаммы Salmonella с мутациями galE могут синтезировать LPS при выращивании с галактозой и являются инвазивными с колонизацией лимфоидных тканей, однако утрачивают эту способность in vivo в связи с недоступностью свободной галактозы, в результате чего они постепенно утрачивают компоненты LPS по мере из размножения в инфицированном или иммунизированном хозяине-животном. Аналогично мутантам по pmi, они постепенно становятся аттенуированными вследствие повышения восприимчивости к комплемент-опосредованной цитотоксичности и повышенному фагоцитозу и уничтожению макрофагами. В то же время поступление галактозы к таким мутантам по galE может приводить к клеточной смерти в результате лизиса, поскольку накопление Gal-1-Р и UDP-Gal является токсичным (30, 48, 49). В связи с этим, рост мутантов по galE в присутствии галактозы способствует отбору мутаций в генах захвата галактозы или в генах galK и galT, в результате чего токсические продукты не синтезируются. К сожалению, такие устойчивые к галактозе мутанты больше неспособны образовывать LPS и являются полностью аттенуированными, неинвазивными и неиммуногенными (30, 50). Для преодоления этих проблем с целью обеспечения применения мутаций по galE в вакцинных штаммах Salmonella были разработаны средства получения мутантов по galE с возможностью обратимого синтеза LPS, зависимого от присутствия или отсутствия галактозы, которые являются устойчивыми к галактозе, при этом отсутствует отбор мутантов, неспособных синтезировать UDP-Gal для синтеза LPS.
5. Регуляторные гены захвата железа
[00101] В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит делецию в эндогенном промоторе Pfur, который регулирует экспрессию гена fur. Fur приводит к репрессии транскрипции генов, участвующих в захвате железа в присутствии свободного железа. Если концентрации железа становятся низкими в бактерии, то синтез Fur прекращается, что приводит к конститутивной экспрессии генов, кодирующих белки захвата железа (например, регулируемые железом белки внешней мембраны (IROMP). В соответствии с некоторыми вариантами осуществления делеция представляет собой частичную делецию эндогенного промотора Pfur. В соответствии с некоторыми вариантами осуществления делеция представляет собой полноразмерную делецию эндогенного промотора Pfur. В соответствии с некоторыми вариантами осуществления эндогенный промотор Pfur генетически модифицируют с целью изменения (например, снижения) экспрессии гена fur. В соответствии с некоторыми вариантами осуществления эндогенный промотор Pfur генетически изменяют с целью включения терминатора транскрипции.
[00102] В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит нуклеиновую кислоту, содержащую ген fur (например, ген fur из того же самого вида бактерии, что и рекомбинантная бактерия).
[00103] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген fur, расположена в плазмиде в бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген fur, расположена в хромосоме бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген fur, расположена в локусе хромосомы, соответствующем локусу эндогенного гена fur, который был удален или изменен в бактериальной хромосоме. В соответствии с некоторыми вариантами осуществления рекомбинантную бактерию модифицируют с целью включения нуклеиновой кислоты, содержащей ген fur, с помощью которого эндогенная копия гена fur в бактериальной хромосоме была изменена и/или удалена.
[00104] Последовательность нуклеиновой кислоты иллюстративного гена fur Salmonella представлена ниже:
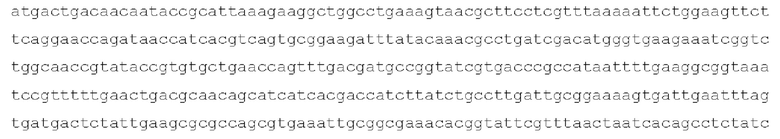

[00105] Аминокислотная последовательность белка Fur, кодируемого нуклеиновой кислотой под SEQ ID NO: 3, представлена ниже:

[00106] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген fur Salmonella (представлен в виде SEQ ID NO: 3). В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген fur, при этом ген fur содержит последовательность нуклеиновой кислоты, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична последовательности нуклеиновой кислоты под SEQ ID NO: 3. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген fur, при этом ген fur содержит последовательность нуклеиновой кислоты, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% гомологична последовательности нуклеиновой кислоты под SEQ ID NO: 3.
[00107] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит последовательность нуклеиновой кислоты, кодирующую белок Fur О, где указанный белок Fur содержит аминокислотную последовательность, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности под SEQ ID NO: 4. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит последовательность нуклеиновой кислоты, кодирующую белок Fur О, где указанный белок Fur содержит аминокислотную последовательность, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% гомологична аминокислотной последовательности под SEQ ID NO: 4.
[00108] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген fur от вида, подвида, серовара или штамма бактерии, который является тем же самым, что и вид бактерии рекомбинантной бактерии.
[00109] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген fur, который является функционально связанным с регулируемым промотором (например, регулируемым сахаром промотором). В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген fur, который является функционально связанным с регулируемым сахаром промотором. В соответствии с некоторыми вариантами осуществления регулируемый сахаром промотор характеризуется повышенной активностью (например, повышенной транскрипцией) в присутствии конкретного сахара и сниженной активностью в отсутствие сахара. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген fur, который является функционально связанным с регулируемым рамнозой промотором (например, регулируемым сахаром промотором). В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген fur, который является функционально связанным с регулируемым арабинозой промотором (например, регулируемым сахаром промотором). В соответствии с некоторыми вариантами осуществления регулируемый арабинозой промотор представляет собой araC ParaBAD. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит мутацию ΔPfur::ТТ araC ParaBAD fur.
6. Гены захвата колицина
[00110] Salmonella спонтанно высвобождает 50-90 нм пузырькообразные частицы внешней мембраны клеточной стенки, называемые генерализованными модулями мембранных антигенов (GMMA) или внешними мембранными везикулами, которые содержат обогащенный источник ассоциированных с внешней мембраной антигенов, которые сохраняют свою нативную конформацию и соответствующую ориентацию. Salmonella может быть генетически модифицирована с целью продуцирования большего количества GMMA или везикул внешней мембраны (например, в результате делеции гена tolR), которые могут быть легко очищены (например, с помощью центрифугирования и фильтрации в отсутствие детергента). GMMA или везикулы внешней мембраны содержат многочисленные патоген-ассоциированные молекулярные паттерны (РАМР), в том числе лиганды toll-подобных рецепторов (TLR), которые могут выступать в качестве самоадъювантов при вызове иммунных ответов. Рекомбинантные бактерии, которые не экспрессируют tolR, продуцируют больше GMMA или везикул внешней мембраны, которые могут быть особо предпочтительными в повышении презентации консервативных белков с целью содействия индукции, например, антител, перекрестно-реагирующих в отношении ОМР других сероваров Salmonella. Помимо этого, не ограничиваясь определенной теорией, повышенное продуцирование и высвобождение GMMA или везикул внешней мембраны, также будет приводить к повышенной презентации антигена, представляющего интерес, который экспрессируется рекомбинантной бактерией, описанной в данном документе. В соответствии с некоторыми вариантами осуществления антиген, представляющий интерес, представляет собой секретируемый антиген.
[00111] В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит делецию в эндогенном гене, кодирующем белок захвата колицина. Были описаны два типа колицинов. Колицины группы А представляют собой Tol-зависимые колицины, а колицины группы В представляют собой TonB-зависимые колицины (см., например, Cascales et al. (2007) Microbiol. Mol. Biol. Rev. 71(1): 158-229 (51), полное содержание которой включено в данный документ посредством ссылки). В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит делецию в эндогенном промоторе PtolR, который регулирует экспрессию гена tolR. Указанная делеция будет приводить к тому, что эндогенный ген tolR не будет экспрессироваться рекомбинантной бактерией, содержащей делецию. В соответствии с некоторыми вариантами осуществления эндогенный промотор Рюш. генетически модифицируют с целью изменения (например, снижения) экспрессии гена tolR. В соответствии с некоторыми вариантами осуществления эндогенный промотор PtolR генетически изменяют с целью включения терминатора транскрипции.
[00112] В соответствии с некоторыми вариантами осуществления рекомбинантную бактерию, описываемую в данном документе, модифицируют с целью включения нуклеиновой кислоты, содержащей ген tolR. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген tolR, расположена в плазмиде в бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген tolR, расположена в хромосоме бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген tolR, расположена в локусе хромосомы, соответствующем локусу эндогенного гена tolR, который был удален или изменен в бактериальной хромосоме. В соответствии с некоторыми вариантами осуществления рекомбинантную бактерию модифицируют с целью включения нуклеиновой кислоты, содержащей ген tolR, с помощью которого эндогенная копия гена tolR в бактериальной хромосоме была изменена и/или удалена.
[00113] Последовательность нуклеиновой кислоты иллюстративного гена tolR Salmonella представлена ниже:

[00114] Аминокислотная последовательность белка TolR, кодируемого нуклеиновой кислотой под SEQ ID NO: 5, представлена ниже:
MTDNNTALKKAGLKVTLPRLKILEVLQEPDNHHVSAEDLYKRLIDMGEEIGLATVYRV LNQFDDAGIVTRHNFEGGKSVFELTQQHHHDHLICLDCGKVIEFSDDSIEARQREIAAKH GIRLTNHSLYLYGHCAEGDCREDEHAHDDATK (SEQ ID NO: 6).
[00115] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген tolR Salmonella (представлен в виде SEQ ID NO: 5). В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген tolR, при этом ген tolR содержит последовательность нуклеиновой кислоты, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична последовательности нуклеиновой кислоты под SEQ ID NO: 5. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген tolR, при этом ген tolR содержит последовательность нуклеиновой кислоты, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% гомологична последовательности нуклеиновой кислоты под SEQ ID NO: 5.
[00116] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит последовательность нуклеиновой кислоты, кодирующую белок TolR, где указанный белок TolR содержит аминокислотную последовательность, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности под SEQ ID NO: 6. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит последовательность нуклеиновой кислоты, кодирующую белок TolR, где указанный белок TolR содержит аминокислотную последовательность, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% гомологична аминокислотной последовательности под SEQ ID NO: 6.
[00117] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген tolR от вида, подвида, серовара или штамма бактерии, который отличается от вида бактерии рекомбинантной бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген tolR от вида, подвида, серовара или штамма бактерии, который является тем же самым, что и вид бактерии рекомбинантной бактерии.
[00118] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген tolR, который является функционально связанным с регулируемым промотором (например, регулируемым сахаром промотором). В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген tolR, который является функционально связанным с регулируемым сахаром промотором. В соответствии с некоторыми вариантами осуществления регулируемый сахаром промотор характеризуется повышенной активностью (например, повышенной транскрипцией) в присутствии конкретного сахара и сниженной активностью в отсутствие сахара. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген tolR, который является функционально связанным с регулируемым рамнозой промотором (например, регулируемым сахаром промотором). В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген tolR, который является функционально связанным с регулируемым арабинозой промотором. В соответствии с некоторыми вариантами осуществления регулируемый арабинозой промотор представляет собой PBAD. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит мутацию ΔPtolR::TT araC ParaBAD tolR.
7. Гены эндосомального ускользания
[00119] В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия была генетически изменена таким образом, что бактерия является способной ускользать от эндосомального компартмента клетки хозяина. Рекомбинантная бактерия может характеризоваться временной задержкой ускользания от эндосомы после инвазии клетки хозяина. Способы выявления ускользания от эндосомального компартмента клетки хозяина хорошо известны в данной области техники и включают в себя, например, микроскопический анализ.
[00120] В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит делецию в эндогенном гене sifA. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит мутацию, которая изменяет функцию SifA. SifA представляет собой эффекторный белок, необходимый для образования индуцированных Salmonella филаментов и для сохранности вакуолярной мембраны, окружающей бактерию. Бактерии, содержащие делецию sifA, способны ускользать от эндосомы клетки хозяина (также называемой Salmonella-содержащей везикулой, или SCV) после клеточной инвазии. В соответствии с некоторыми вариантами осуществления делеция эндогенного гена sifA представляет собой частичную делецию. В соответствии с некоторыми вариантами осуществления делеция эндогенного гена sifA представляет собой полноразмерную делецию. В соответствии с некоторыми вариантами осуществления эндогенный ген sifA генетически изменяют с целью вставки терминатора транскрипции в открытую рамку считывания гена. В соответствии с некоторыми вариантами осуществления регуляторную область эндогенного гена sifA генетически модифицируют с целью изменения (например, снижения) экспрессии гена sifA. В соответствии с некоторыми вариантами осуществления промотор эндогенного гена sifA изменяют с целью включения одного или нескольких регуляторных элементов (например, восприимчивого к сахару промотора).
[00121] В соответствии с некоторыми вариантами осуществления рекомбинантную бактерию, описываемую в данном документе, модифицируют с целью включения нуклеиновой кислоты, содержащей ген sifA. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген sifA, расположена в плазмиде в бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген sifA, расположена в хромосоме бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген sifA, расположена в локусе хромосомы, соответствующем локусу эндогенного гена sifA, который был удален или изменен в бактериальной хромосоме. В соответствии с некоторыми вариантами осуществления рекомбинантную бактерию модифицируют с целью включения нуклеиновой кислоты, содержащей ген sifA, с помощью которого эндогенная копия гена sifA в бактериальной хромосоме была изменена и/или удалена.
[00122] Последовательность нуклеиновой кислоты иллюстративного гена sifA Salmonella представлена ниже:


[00123] Аминокислотная последовательность белка SifA, кодируемого нуклеиновой кислотой под SEQ ID NO: 7, представлена ниже:
MPITIGNGFLKSEILTNSPRNTKEAWWKVLWEKIKDFFFSTGKAKADRCLHEMLFAERAP TRERLTEIFFELKELACASQRDRFQVHNPHENDATIILRIMDQNEENELLRITQNTDTFS CEVMGNLYFLMKDRPDILKSHPQMTAMIKRRYSEIVDYPLPSTLCLNPAGAPILSVPLDN IEGYLYTELRKGHLDGWKAQEKATYLAAKIQSGIEKTTRILHHANISESTQQNAFLETM AMCGLKQLEIPPPHTHIPIEKMVKEVLLADKTFQAFLVTDPSTSQSMLAEIVEAISDQVFH AIFRIDPQAIQKMAEEQLTTLHVRSEQQSGCLCCFL (SEQ ID NO: 8).
[00124] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген sifA Salmonella (представлен в виде SEQ ID NO: 7). В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген sifA, при этом ген sifA содержит последовательность нуклеиновой кислоты, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична последовательности нуклеиновой кислоты под SEQ ID NO: 7. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген sifA, при этом ген sifA содержит последовательность нуклеиновой кислоты, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% гомологична последовательности нуклеиновой кислоты под SEQ ID NO: 7.
[00125] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит последовательность нуклеиновой кислоты, кодирующую белок SifA, где указанный белок SifA содержит аминокислотную последовательность, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности под SEQ ID NO: 8. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит последовательность нуклеиновой кислоты, кодирующую белок SifA, где указанный белок SifA содержит аминокислотную последовательность, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% гомологична аминокислотной последовательности под SEQ ID NO: 8.
[00126] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген sifA от вида, подвида, серовара или штамма бактерии, который отличается от вида бактерии рекомбинантной бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген sifA от вида, подвида, серовара или штамма бактерии, который является тем же самым, что и вид бактерии рекомбинантной бактерии.
[00127] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген sifA, который является функционально связанным с регулируемым промотором (например, регулируемым сахаром промотором). В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген sifA, который является функционально связанным с регулируемым сахаром промотором. В соответствии с некоторыми вариантами осуществления регулируемый сахаром промотор характеризуется повышенной активностью (например, повышенной транскрипцией) в присутствии конкретного сахара и сниженной активностью в отсутствие сахара. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген sifA, который является функционально связанным с регулируемым рамнозой промотором (например, регулируемым сахаром промотором). В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген sifA, который является функционально связанным с регулируемым арабинозой промотором. В соответствии с некоторыми вариантами осуществления регулируемый арабинозой промотор представляет собой PBAD. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит мутацию ΔsifA::TT araC PBAD sifA. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит мутацию ΔPsifA::TT araC ParaBAD sifA. При экспрессии нуклеиновой кислоты, содержащей ген sifA под контролем регулируемого арабинозой промотора, ускользание бактерии от эндосомы хозяина может быть замедлено. Поскольку арабиноза отсутствует в клетках-хозяевах, арабиноза не может индуцировать экспрессию гена sifA. Таким образом, если рекомбинантную бактерию культивируют в присутствии арабинозы до введения субъекту, экспрессия sifA будет постепенно снижаться с каждым циклом деления бактериальной клетки, тем самым, способствуя ускользанию бактерии от эндосомы клетки хозяина во время начальных циклов клеточного деления. Аналогичные мутации замедленного ускользания могут быть сконструированы с помощью других регулируемых промоторов, например, из регулируемых ксилозой или регулируемых рамнозой промоторных систем.
8. Гены GTP-пирофосфокиназы
[00128] В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит делецию в эндогенном гене relA, который кодирует GTP-пирофосфокиназу RelA. Включение делеции relA в рекомбинантную бактерию разделяет возникновение зависимого от роста лизиса и необходимость непрерывного синтеза белка. В соответствии с некоторыми вариантами осуществления делеция эндогенного гена relA представляет собой частичную делецию. В соответствии с некоторыми вариантами осуществления делеция эндогенного гена relA представляет собой полноразмерную делецию.
9. Другие способы аттенуации
[00129] Другие способы аттенуации известны в данной области техники. Например, аттенуация может осуществляться посредством изменения (например, удаления) последовательностей нуклеиновой кислоты, встречающихся в бактерии дикого типа. Например, если бактерия представляет собой Salmonella, неограничивающие примеры последовательностей нуклеиновой кислоты, которые могут быть использованы для аттенуации, включают в себя: последовательность нуклеиновой кислоты pab, последовательность нуклеиновой кислоты pur, последовательность нуклеиновой кислоты aro, asd, последовательность нуклеиновой кислоты dap, nadA, pncB, galE, pmi, fur, rpsL, ompR, htrA, hemA, cdt, cya, crp, dam, phoP, phoQ, rfc, poxA, galU, mviA, sodC, recA, ssrA, sirA, inv, hilA, rpoE, flgM, tonB, slyA и любую их комбинацию. Иллюстративные аттенуирующие мутации могут представлять собой aroA, aroC, aroD, cdt, cya, crp, phoP, phoQ, ompR, galE и htrA.
[00130] В соответствии с определенными вариантами осуществления вышеуказанные последовательности нуклеиновой кислоты могут быть помещены под контроль регулируемого сахаром промотора, при этом указанный сахар присутствует во время роста рекомбинантной бактерии in vitro, однако по сути отсутствует в организме животного-хозяина или человека-хозяина. Прекращение транскрипции указанных последовательностей нуклеиновой кислоты, приведенных выше, впоследствии будет приводить к аттенуированию и неспособности рекомбинантной бактерии индуцировать симптомы заболеваний.
В. Дополнительные мутации
[00131] В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит делецию в эндогенном гене recF, который кодирует белок, ответственный за репликацию и репарацию ДНК, RecF. В соответствии с некоторыми вариантами осуществления делеция эндогенного гена recF представляет собой частичную делецию. В соответствии с некоторыми вариантами осуществления делеция эндогенного гена recF представляет собой полноразмерную делецию. В соответствии с некоторыми вариантами осуществления эндогенный ген recF генетически изменяют с целью вставки терминатора транскрипции в открытую рамку считывания гена.
[00132] В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит делецию в эндогенном гене recJ, который кодирует экзонуклеазу RecJ. В соответствии с некоторыми вариантами осуществления делеция эндогенного гена red представляет собой частичную делецию. В соответствии с некоторыми вариантами осуществления делеция эндогенного гена recJ представляет собой полноразмерную делецию. В соответствии с некоторыми вариантами осуществления эндогенный ген recJ генетически изменяют с целью вставки терминатора транскрипции в открытую рамку считывания гена.
[00133] Бактерия также может быть модифицирована с целью получения системы сбалансированных леталей хозяин-вектор, хотя и другие типы систем также могут быть использованы (например, получение гетерозиготы по комплементации). Для системы сбалансированных леталей хозяин-вектор бактерия может быть модифицирована с целью контроля ее способности синтезировать различные обязательные компоненты, необходимые для синтеза ригидного пептидогликанового слоя своей клеточной стенки. В одном примере указанный компонент представляет собой диаминопимелиновую кислоту (DAP). Различные ферменты вовлечены в последующий синтез DAP.
[00134] В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит делецию в эндогенном гене asd. В соответствии с некоторыми вариантами осуществления делеция эндогенного гена asd представляет собой частичную делецию. В соответствии с некоторыми вариантами осуществления делеция эндогенного гена asd представляет собой полноразмерную делецию. В соответствии с некоторыми вариантами осуществления эндогенный ген asd генетически изменяют с целью вставки терминатора транскрипции в открытую рамку считывания гена. В соответствии с некоторыми вариантами осуществления промотор эндогенного гена asd изменяют с целью включения одного или нескольких регуляторных элементов (например, восприимчивого к сахару промотора). В одном примере бактерию модифицируют с помощью мутации ΔasdA с устранением способности бактерии продуцировать β-аспартат-полуальдегиддегидрогеназу, фермент, необходимый для синтеза DAP. Другие мутации, которые приводят к устранению синтеза DAP, включают в себя без ограничения dapA, dapB, dapC, dapD, dapE, dapF и asd (см., например, патент США №6872547, включенный в данный документ посредством ссылки). Другие модификации, которые могут быть использованы, включают в себя модификации способности бактерии синтезировать D-аланин или синтезировать D-глутаминовую кислоту (например, мутации ΔmurI), которые представляют собой уникальные компоненты пептидогликанового слоя бактериальной клеточной стенки.
[00135] Аналогично, в соответствии с различными вариантами осуществления может быть предусмотрена генная кассета araC ParaBAD с2, вставленная в последовательность нуклеиновой кислоты asd, которая кодирует аспартат-полуальдегиддегидрогеназу. Поскольку последовательность нуклеиновой кислоты araC транскрибируется в направлении, которое может приводить к интерференции экспрессии прилегающих последовательностей нуклеиновой кислоты и отрицательно влиять на функциональные характеристики вакцинного штамма, последовательность терминации транскрипции (ТТ), как правило, вставляют в направлении 3' по отношению к последовательности нуклеиновой кислоты araC. Хромосомную последовательность нуклеиновой кислоты asd в типичном случае инактивируют с целью обеспечения применения плазмидных векторов, кодирующих последовательность нуклеиновой кислоты asd дикого типа в системе сбалансированных леталей хозяин-вектор. Это способствует стабильной сохранности плазмид in vivo в отсутствие каких-либо характеристик устойчивости к лекарственным препаратам, которые являются недопустимыми в живых бактериальных вакцинах. В соответствии с некоторыми вариантами осуществления последовательность нуклеиновой кислоты asd дикого типа может кодироваться вектором, описанным в данном документе. Указанный вектор способствует регулируемой экспрессии последовательности, кодирующей антиген, посредством репрессируемого промотора.
С. Регуляторные системы репрессоров
[00136] В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит нуклеиновую кислоту (например, ген), которая функционально связана с регулируемым репрессором промотором с целью облегчения регулируемой экспрессии указанного гена. Таким образом, в соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит нуклеиновую кислоту, содержащую ген, кодирующий репрессор. В соответствии с некоторыми вариантами осуществления ген, кодирующий репрессор, является функционально связанным с регулируемым промотором. Способы хромосомной интеграции последовательности нуклеиновой кислоты, кодирующей репрессор, функционально связанный с регулируемым промотором, хорошо известны в данной области техники и подробно представлены в примерах. В соответствии с некоторыми вариантами осуществления последовательность нуклеиновой кислоты, кодирующую репрессор, не интегрируют в хромосомный локус, в результате чего способность бактерии колонизировать клетку-хозяина нарушается. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит нуклеиновую кислоту, кодирующую репрессор, который интегрируют в локус relA бактериальной хромосомы. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит нуклеиновую кислоту, кодирующую репрессор, который интегрируют в локус endA бактериальной хромосомы. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит по меньшей мере одну последовательность нуклеиновой кислоты, кодирующую репрессор. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит по меньшей мере две, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть или более нуклеиновых кислоты, кодирующих репрессор. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, кодирующая репрессор, присутствует в плазмиде в бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, кодирующая репрессор, расположена в бактериальной хромосоме. Если существует более одной последовательности нуклеиновой кислоты, кодирующей репрессор, то каждая последовательность нуклеиновой кислоты, кодирующая репрессор, может быть функционально связана с регулируемым промотором, в результате чего каждый промотор регулируется одним и тем же соединением или условием. В качестве альтернативы каждая последовательность нуклеиновой кислоты, кодирующая репрессор, может быть функционально связана с регулируемым промотором, при этом каждая из них регулируется различным соединением или условием.
[00137] Используемый в данном документе термин «репрессор» относится к биомолекуле, которая приводит к репрессии транскрипционой активности промотора. В соответствии с некоторыми вариантами осуществления репрессор синтезируется рекомбинантной бактерией в достаточно высоких количествах во время культивирования in vitro, в результате чего транскрипция нуклеиновой кислоты, которая является функционально связанной с регулируемым репрессором промотором, подвергается репрессии. Это может быть особенно предпочтительным, если, например, экспрессия продукта, кодируемого указанной нуклеиновой кислотой, задерживает рост бактерии in vitro и/или способность бактерии инфицировать и/или колонизировать субъекта. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, которая является функционально связанной с регулируемым репрессором промотором, экспрессирует антиген, представляющий интерес. В соответствии с некоторыми вариантами осуществления концентрация репрессора в клетке постепенно снижается с каждым циклом клеточного деления после того, как транскрипция гена, кодирующего репрессор, снижается или прекращается (например, in vivo). Применение определенного репрессора, описанного выше, может зависеть отчасти от вида, подвида, штамма или серовара рекомбинантной бактерии, подлежащей использованию. В соответствии с некоторыми вариантами осуществления репрессор происходит из такого же самого вида (например такого же самого вида бактерий или такого же самого фага), из которого происходит регулируемый репрессором промотор. В соответствии с некоторыми вариантами осуществления репрессор не происходит из такого же самого вида бактерии, что и вид бактерии, в котором репрессор экспрессируется. Например, в соответствии с некоторыми вариантами осуществления репрессор происходит из Е. coli, если рекомбинантная бактерия принадлежит роду Salmonella. Другие подходящие репрессоры включают в себя репрессоры, происходящие из бактериофага.
[00138] Последовательность нуклеиновой кислоты, кодирующая репрессор и регулируемый промотор, подробно описанные выше, может быть модифицирована для того, чтобы оптимизировать уровень экспрессии последовательности нуклеиновой кислоты, кодирующей репрессор. Оптимальный уровень экспрессии последовательности нуклеиновой кислоты, кодирующей репрессор, может быть оценена или может быть определена экспериментальным путем. При таком определении необходимо учитывать выступает ли репрессор в качестве мономера, димера, тримера, тетрамера или более высокого мультимера, а также необходимо учитывать число копий вектора, кодирующего антиген, представляющий интерес. В соответствии с иллюстративным вариантом осуществления уровень экспрессии оптимизируют, в результате чего репрессор синтезируется в благоприятной среде (т.е., рост in vitro) при уровне, который по сути ингибирует экспрессию нуклеиновой кислоты, кодирующей антиген, представляющий интерес, и по сути не синтезируется в неблагоприятной среде, тем самым, способствуя экспрессии нуклеиновой кислоты, кодирующей антиген, представляющий интерес.
[00139] В соответствии с некоторыми вариантами осуществления рекомбинантную бактерию, описываемую в данном документе, модифицируют с целью включения нуклеиновой кислоты, содержащей ген lacI, который кодирует репрессорный белок LacI. Экспрессия кодируемого lacI репрессора в рекомбинантной бактерии, описанной в данном документе, может быть использована для регуляции экспрессии гена, кодирующего антиген, представляющий интерес, экспрессируемый бактерией. Например, в соответствии с некоторыми вариантами осуществления экспрессия гена lacI регулируется регулируемым сахаром промотором (например, регулируемым арабинозой промотором). При культивировании в присутствии арабинозы рекомбинантная бактерия будет синтезировать репрессорный белок LacI, который в свою очередь приводит к репрессии экспрессии гена, кодирующего антиген, представляющий интерес, который является функционально связанным с восприимчивым к LacI промотору (например, Ptrc, Plac, PT7lac и Ptac). При введении субъекту и в отсутствии источника арабинозы синтез репрессора LacI прекращается, приводя к дерепрессии восприимчивого к LacI промотора и последующей экспрессии антигена, представляющего интерес. Концентрация LacI в клетке снижается приблизительно наполовину при каждом клеточном делении in vivo, приводя к постепенному снижению уровня репрессии и постепенному повышению синтеза антигена, представляющего интерес.
[00140] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген lacI, расположена в плазмиде в бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген lacI, расположена в хромосоме бактерии. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота, содержащая ген lacI, расположена в локусе хромосомы, соответствующем локусу эндогенного гена relA, который был удален или изменен в бактериальной хромосоме. В соответствии с некоторыми вариантами осуществления рекомбинантную бактерию модифицируют с целью включения нуклеиновой кислоты, содержащей ген lacI, с помощью которого эндогенная копия гена lacI в бактериальной хромосоме была изменена и/или удалена.
[00141] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген lacI Escherichia coli. Последовательность нуклеиновой кислоты гена lacI Е. coli представлена ниже:

[00142] Аминокислотная последовательность белка LacI Е. coli, кодируемого нуклеиновой кислотой под SEQ ID NO: 9, представлена ниже:
MKPVTLYDVAEYAGVSYQTVSRVVNQASHVSAKTREKVEAAMAELNYIPNRVAQQLA GKQSLLIGVATSSLALHAPSQIVAAIKSRADQLGASVVVSMVERSGVEACKAAVHNLLA QRVSGLIINYPLDDQDAIAVEAACTNVPALFLDVSDQTPINSIIFSHEDGTRLGVEHLVAL GHQQIALLAGPLSSVSARLRLAGWHKYLTRNQIQPIAEREGDWSAMSGFQQTMQMLNE GIVPTAMLVANDQMALGAMRAITESGLRVGADISVVGYDDTEDSSCYIPPLTTIKQDFR LLGQTSVDRLLQLSQGQAVKGNQLLPVSLVKRKTTLAPNTQTASPRALADSLMQLARQ VSRLESGQ (SEQ ID NO: 10).
[00143] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген lacI, при этом ген lacI содержит последовательность нуклеиновой кислоты, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична последовательности нуклеиновой кислоты под SEQ ID NO: 9. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген lacI, при этом ген lacI содержит последовательность нуклеиновой кислоты, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% гомологична последовательности нуклеиновой кислоты под SEQ ID NO: 9.
[00144] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит последовательность нуклеиновой кислоты, кодирующую белок LacI, где указанный белок LacI содержит аминокислотную последовательность, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности под SEQ ID NO: 10. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит последовательность нуклеиновой кислоты, кодирующую белок LacI, где указанный белок LacI содержит аминокислотную последовательность, которая на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 81%, на по меньшей мере 82%, на по меньшей мере 83%, на по меньшей мере 84%, на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% гомологична аминокислотной последовательности под SEQIDNO: 10.
[00145] В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген lacI, который является функционально связанным с регулируемым промотором (например, регулируемым сахаром промотором). В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген lacI, который является функционально связанным с регулируемым сахаром промотором. В соответствии с некоторыми вариантами осуществления регулируемый сахаром промотор характеризуется повышенной активностью (например, повышенной транскрипцией) в присутствии конкретного сахара и сниженной активностью в отсутствие сахара. В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген lacI, который является функционально связанным с регулируемым рамнозой промотором (например, регулируемым сахаром промотором). В соответствии с некоторыми вариантами осуществления нуклеиновая кислота содержит ген lacI, который является функционально связанным с регулируемым арабинозой промотором. В соответствии с некоторыми вариантами осуществления регулируемый арабинозой промотор представляет собой ParaBAD. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит мутацию ΔrelA::araC ParaBAD lacI TT.
D. Антигены
[00146] В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит нуклеиновую кислоту, кодирующую антиген, представляющий интерес. Используемый в данном документе термин «антиген» относится к биомолекуле, способной вызывать иммунный ответ у хозяина. В соответствии с некоторыми вариантами осуществления антиген может представлять собой белок или фрагмент белка. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит нуклеиновую кислоту (например, плазмиду), кодирующую антиген, представляющий интерес, при этом нуклеиновая кислота экспрессируется клеткой-хозяином (например, ДНК-вакцина). В соответствии с иллюстративным вариантом осуществления антиген вызывает защитный иммунный ответ у субъекта.
[00147] Используемый в данном документе термин «защитный» означает, что иммунный ответ способствует уменьшению любых симптомов, ассоциированных с инфицированием хозяина патогеном, от которого антиген произошел или против которого был сконструирован, чтобы вызывать ответ. Например, защитный антиген из патогена, такого как Salmonella, может индуцировать иммунный ответ, который способствует нормализации симптомов, ассоциированных с инфицированием Salmonella, или снижению заболеваемости и смертности, ассоциированных с инфицированием патогеном, или может снижать способность Salmonella инфицировать и колонизировать хозяина. Применение термина «защитный» в настоящем раскрытии не обязательно требует того, чтобы хозяин был полностью защищен от эффектов патогена.
[00148] В соответствии с некоторыми вариантами осуществления антиген, представляющий интерес, представляет собой антиген, происходящий из инфекционного агента. В соответствии с некоторыми вариантами осуществления антиген, представляющий интерес, происходит из инфекционного агента, выбранного из группы, состоящей из вируса, бактерии, простейшего, приона, гриба и гельминта. В соответствии с некоторыми вариантами осуществления антиген, представляющий интерес, происходит из бактерии. В соответствии с некоторыми вариантами осуществления антиген, представляющий интерес, представляет собой антиген Salmonella. В соответствии с некоторыми вариантами осуществления антиген Salmonella выбирают из группы, состоящей из FliC, FliC180, OmpC, OmpD, OmpF, SseB и SseI. В соответствии с некоторыми вариантами осуществления антиген, представляющий интерес, представляет собой вирусный антиген. В соответствии с некоторыми вариантами осуществления антиген, представляющий интерес, представляет собой антиген вируса гриппа. В соответствии с некоторыми вариантами осуществления антиген вируса гриппа представляет собой гемагглютинин или нейраминидизу, в случае доставки с помощью ДНК-вакцины. В соответствии с некоторыми вариантами осуществления антиген, представляющий интерес, ассоциирован с раком. В соответствии с некоторыми вариантами осуществления антиген, ассоциированный с раком, выбирают из группы, состоящей из MAGE-A, MAGE-C1, BAGE, GAGE, XAGE, NY-ESO1 (также известного как CTAG1B и LAGE2), LAGE1 (также известного как CTAG2) и сурвивина.
[00149] В качестве альтернативы антигены могут происходить из гамет, при условии, что они являются специфическими в отношении гамет, и могут быть сконструированы с целью блокирования оплодотворения. В соответствии с другим вариантом осуществления антигены могут представлять собой опухолевые антигены и могут быть сконструированы с целью снижения опухолевого роста. Особо подразумевается, что антигены из организмов, только что идентифицированные или только что ассоциированные с заболеванием или патогенным состоянием, или новые или возвращающиеся патогены животных или людей, в том числе таковые, известные в настоящее время или подлежащие идентификации в будущем, могут экспрессироваться бактерией, подробно описанной в данном документе. Кроме того, антигены не ограничены таковыми из патогенных организмов.
[00150] Иммуногенность бактерии можно усиливать и/или модулировать с помощью конструирования штаммов, которые также экспрессируют последовательности цитокинов, адъювантов и других иммуномодуляторов.
[00151] Некоторые примеры микроорганизмов, пригодных в качестве источника антигена, приведены ниже. Они могут включать в себя микроорганизмы для контроля чумы, вызываемой Yersinia pestis и другими видами Yersinia, такими как Y. pseudotuberculosis и Y. enterocolitica, для контроля гонореи, вызываемой Neisseria gonorrhoea, для контроля сифилиса, вызываемого Treponema pallidum и для контроля венерических заболеваний, а также глазных инфекций, вызываемых Chlamydia trachomatis. Виды Streptococcus как из группы А, так и из группы В, такие как виды, которые вызывают боль в горле или болезни сердца, Streptococcus equi, который вызывает мыт у лошадей, Streptococcus mutans, который вызывает образование кариозных полостей, и Streptococcus pneumoniae, Erysipelothrix rhusiopathiae, Neisseria meningitidis, Mycoplasma pneumoniae и другие виды Mycoplasma, Hemophilus influenza, Bordetella pertussis, Mycobacterium tuberculosis, Mycobacterium leprae, другие виды Bordetella, Escherichia coli, Brucella abortus, Pasteurella hemolytica и P. multocida, Vibrio cholera, виды Shigella, виды Borrellia, виды Bartonella, Heliobacter pylori, виды Campylobacter, виды Pseudomonas, виды Moraxella, виды Brucella, виды Francisella, виды Aeromonas, виды Actinobacillus, виды Clostridium (такие как С.perfringens), виды Rickettsia, виды Bacillus, виды Coxiella, виды Ehrlichia, виды Listeria и Legionella pneumophila представляют собой дополнительные примеры в объеме настоящего раскрытия, из которых последовательности нуклеиновой кислоты антигенов могут быть получены.
[00152] Также могут быть использованы вирусные антигены. Вирусные антигены могут быть использованы в микроорганизмах для доставки антигенов, направленных против вирусов, либо ДНК-, либо РНК-вирусов, например, из классов Papovavirus, Adenovirus, Herpesvirus, Poxvirus, Parvovirus, Reovirus, Picornavirus, Myxovirus, Paramyxovirus, Flavivirus или Retrovirus. Антигены также могут происходить из патогенных грибов, простейших и паразитов. В то же время, средства для доставки антигенов или последовательностей, кодирующих антиген, зависят от типа антигена и/или вируса.
[00153] В соответствии с определенными вариантами осуществления антиген может содержать В-клеточный эпитоп или Т-клеточный эпитоп. В качестве альтернативы антиген, в отношении которого иммунный ответ является желательным, может экспрессироваться в виде слияния с белком-переносчиком, который содержит сильный промискуитетный Т-клеточный эпитоп и/или выступает в виде адъюванта и/или облегчает презентацию антигена для усиления, во всех случаях, иммунного ответа в отношении антигена или его составной части. Это может быть осуществлено с помощью способов, известных в данной области техники. Слияние с фрагментом столбнячного токсина С, СТ-В, LT-B и кором вируса гепатита В является особенно пригодным для таких целей, хотя другие системы презентации эпитопов хорошо известны в данной области техники.
[00154] В соответствии с дополнительными вариантами осуществления последовательность нуклеиновой кислоты, кодирующая антиген, может содержать сигнал секреции.
[00155] Как указано выше, уровень синтеза антигена, представляющего интерес, может быть оптимизирован в результате модификации последовательности нуклеиновой кислоты, кодирующей репрессор и/или промотор. Используемый в данном документе термин «модифицировать» относится к изменению последовательности нуклеиновой кислоты репрессора и/или промотора, которое приводит к изменению уровня транскрипции последовательности нуклеиновой кислоты, кодирующей репрессор, или которое приводит к измерению уровня синтеза репрессора. Например, в соответствии с одним вариантом осуществления термин «модифицировать» может относиться к изменению старт-кодона последовательности нуклеиновой кислоты, кодирующей репрессор. В самом общем виде старт-кодон GTG или TTG, в отличие от старт-кодона ATG, может приводить к снижению эффективности трансляции в десять раз. В соответствии с другим вариантом осуществления термин «модифицировать» может относиться к изменению последовательности Шайна-Дальгарно (SD) последовательности нуклеиновой кислоты, кодирующей репрессор. Последовательность SD представляет собой сайт связывания рибосомы, как правило, расположенный на 6-7 выше от старт-кодона. Консенсусная последовательности SD представляет собой AGGAGG и вариации консенсусной последовательности могут приводить к изменению эффективности трансляции. В соответствии с еще одним вариантом осуществления термин «модифицировать» может относиться к изменению расстояния между последовательностью SD и старт-кодоном. В соответствии с еще одним вариантом осуществления термин «модифицировать» может относиться к изменению -35 последовательности для распознавания РНК-полимеразы. В соответствии с аналогичным вариантом осуществления термин «модифицировать» может относиться к изменению -10 последовательности для связывания РНК-полимеразы. В соответствии с аналогичным вариантом осуществления термин «модифицировать» может относиться к изменению числа нуклеотидов между -35 и -10 последовательностями. В соответствии с альтернативным вариантом осуществления термин «модифицировать» может относиться к оптимизации кодонов последовательности нуклеиновой кислоты, кодирующей репрессор, с целью изменения уровня трансляции мРНК, кодирующей репрессор. Например, кодоны, не имеющие высокого содержания А, расположенные изначально после старт-кодона последовательности нуклеиновой кислоты, кодирующей репрессор, могут не приводить к сведению к максимуму трансляции мРНК, кодирующей репрессор. Аналогично, кодоны последовательности нуклеиновой кислоты, кодирующие любые из белков, описанных в данном документе, могут быть кодон-оптимизированными, т.е., измененными, таким образом, чтобы имитировать кодоны из активно синтезируемых белков определенного организма. В соответствии с дополнительным вариантом осуществления термин «модифицировать» может относиться к изменению содержания GC последовательности нуклеиновой кислоты, кодирующей репрессор, с целью изменения уровня трансляции мРНК, кодирующей репрессор. Способы модификации последовательности нуклеиновой кислоты известны в данной области техники.
[00156] В соответствии с некоторыми вариантами осуществления более одной модификации или типа модификации может быть выполнено с целью оптимизации уровня экспрессии нуклеиновой кислоты, описанной в данном документе (например, нуклеиновой кислоты, кодирующей репрессор или антиген, представляющие интерес). Например, по меньшей мере одна, две, три, четыре, пять, шесть, семь, восемь или девять модификаций или типов модификаций могут быть выполнены с целью оптимизации уровня экспрессии нуклеиновой кислоты, описанной в данной документе. В качестве неограничивающего примера, если репрессор представляет собой LacI, то последовательность нуклеиновой кислоты LacI и промотора могут быть изменены таким образом, чтобы повысить уровень синтеза LacI. В соответствии с одним вариантом осуществления старт-кодон репрессора LacI может быть изменен с GTG на ATG. В соответствии с другим вариантом осуществления последовательность SD может быть изменена с AGGG на AGGA. В соответствии с еще одним вариантом осуществления кодоны lacI могут быть оптимизированы в соответствии с использованием кодонов для активно синтезируемых белков Salmonella. В соответствии с дополнительным вариантом осуществления старт-кодон lacI может быть изменен, последовательность SD может быть изменена и кодоны lacI могут быть оптимизированы.
[00157] В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия содержит нуклеиновую кислоту, которая расположена в плазмиде или векторе. Используемый в данном документе термин «вектор» относится к автономно реплицируемой единице нуклеиновой кислоты. Настоящее раскрытие может быть реализовано на практике с использованием любого типа вектора, в том числе вирусного, космидного, фазмидного и плазмидного векторов. Наиболее предпочтительным типом вектора является плазмидный вектор. В соответствии с некоторыми вариантами осуществления плазмида или вектор представляют собой высококопийные плазмиду или вектор. В соответствии с некоторыми вариантами осуществления плазмида или вектор представляют собой низкокопийные плазмиду или вектор.
[00158] Как хорошо известно в данной области техники, плазмиды и другие векторы могут иметь широкий спектр промоторов, последовательности множественного клонирования, терминаторы транскрипции и т.д., и векторы могут быть выбраны с целью контроля уровня экспрессии последовательности нуклеиновой кислоты, кодирующей антиген, с помощью контроля относительного числа копий вектора. В соответствии с некоторыми вариантами осуществления, в которых вектор может кодировать поверхностно расположенный адгезии в качестве антигена или антиген, способный стимулировать Т-клеточный иммунитет, может быть предпочтительным применение вектора с низким числом копий, например, двумя, тремя, четырьмя, пятью, шестью, семью, восемью, девятью или десятью копиями на бактериальную клетку. Неограничивающим примером низкокопийного вектора может быть вектор, содержащий ori pSC101.
[00159] В соответствии с некоторыми вариантами осуществления плазмида содержит последовательность нуклеиновой кислоты, кодирующую ген аспартат-полуальдегиддегидрогеназы (например, asdA). Эти плазмиды могут быть предпочтительно использованы с целью комплементации бактерии, которая содержит мутацию гена аспартат-полуальдегиддегидрогеназы (например, asdA). В соответствии с некоторыми вариантами осуществления плазмиду выбирают из группы, состоящей из pYA3342, pYA3337 и pYA3332.
[00160] В других случаях вектор с промежуточным числом копий может быть оптимальным для индукции необходимых иммунных ответов. Например, вектор с промежуточным числом копий может иметь по меньшей мере 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 копий на бактериальную клетку. Неограничивающим примером вектора с промежуточным числом копий может быть вектор, содержащий ori р15А.
[00161] В еще одних случаях высококопийный вектор может быть оптимальным для индукции максимального ответа антител. Высококопийный вектор может иметь по меньшей мере 31, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 100 копий на бактериальную клетку. В соответствии с некоторыми вариантами осуществления высококопийный вектор может иметь по меньшей мере 100, 125, 150, 175, 200, 225, 250, 275, 300, 325, 350, 375 или 400 копий на бактериальную клетку. Неограничивающие примеры высококопийных векторов могут включать в себя вектор, содержащий ori pBR или ori pUC.
[00162] Кроме того, число копий вектора может повышаться в результате отбора мутаций, которые повышают число копий плазмиды. Эти мутации могут возникать в бактериальной хромосоме, однако более вероятно возникнут в плазмидном векторе.
[00163] Предпочтительно векторы, используемые в данном документе, не содержат маркеров устойчивости к антибиотикам, с целью отбора в отношении сохранности вектора.
[00164] Промоторы для применения в вариантах осуществления, описанные в данном документе, известны в данной области техники. Специалисту в данной области техники будет понятно, что выбор репрессора обусловливает отчасти выбор промотора, подлежащего использованию, с целью регуляции экспрессии нуклеиновой кислоты, описанной в данном документе. Например, если репрессор представляет собой LacI, то промотор может быть выбран из группы, состоящей из восприимчивых к LacI промоторов, таких как Ptrc, Plac, PT7lac, Ptac, PompA lacO и Plpp lacO. Если репрессор представляет собой С2, то промотор может быть выбран из группы, состоящей из восприимчивых к С2 промоторов, таких как Р22 промоторы PL и PR. Если репрессор представляет собой С1, то промотор может быть выбран из группы, состоящей из восприимчивых к С1 промоторов, таких как λ промоторы PL и PR.
[00165] В соответствии с каждым вариантом осуществления промотор регулирует экспрессию последовательности нуклеиновой кислоты. В соответствии с некоторыми вариантами осуществления промотор содержит регуляторную последовательность, контролируемую репрессором, в результате чего экспрессия последовательности нуклеиновой кислоты подвергается репрессии, когда репрессор синтезируется (например, во время роста бактерии in vitro), однако экспрессия последовательности нуклеиновой кислоты, кодирующей антиген, является высокой, когда репрессор не синтезируется (например, in vivo). В самом общем виде концентрация репрессора будет снижаться с каждым клеточным делением после того, как экспрессия гена, кодирующего репрессор, прекращается. В соответствии с некоторыми вариантами осуществления концентрация репрессора снижается, в результате чего высокие уровни экспрессии последовательности нуклеиновой кислоты, которая подлежит регуляции, достигаются через приблизительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 делений бактерии. В соответствии с иллюстративным вариантом осуществления концентрация репрессора снижается достаточно для того, чтобы способствовать высокому уровню экспрессии последовательности нуклеиновой кислоты, кодирующей антиген, через приблизительно 5 делений бактерии in vivo.
[00166] В соответствии с определенными вариантами осуществления промотор может содержать другие регуляторные элементы. Например, промотор может содержать lacO, если репрессор представляет собой LacI. Это справедливо в случае промотора липопротеина Plpp lacO, который регулируется LacI, поскольку он имеет LacI-связывающий домен lacO. В соответствии с одним вариантом осуществления репрессор представляет собой репрессор LacI и промотор представляет собой Ptrc.
[00167] В соответствии с некоторыми вариантами осуществления экспрессия последовательности нуклеиновой кислоты, регулируемая репрессором, подвергается репрессии in vivo. Экспрессия может быть «подвергнута репрессии» или «частично подвергнута репрессии», если она составляет приблизительно 50%, 45%, 40%, 35%, 30%, 25%, 20%, 15%, 10%, 5%, 1% или даже менее чем 1% экспрессии в условиях без репрессии. Таким образом, несмотря на то, что уровень экспрессии в условиях «полной репрессии» может быть чрезвычайно низким, она, вероятно, подлежит выявлению с помощью чувствительных способов, поскольку репрессия никогда не может быть абсолютной.
[00168] В отличие от этого, экспрессия последовательности нуклеиновой кислоты, кодирующей антиген, должна быть высокой, когда экспрессия репрессора подвергается репрессии. Например, если репрессор не синтезируется во время роста рекомбинантной бактерии в организме хозяина, то экспрессия нуклеиновой кислоты под контролем репрессора будет высокой. Используемый в данном документе термин «высокий уровень» экспрессии относится к экспрессии, которая является достаточно выраженной для того, чтобы вызывать иммунный ответ в отношении антигена. Соответственно, число копий, коррелирующее с высоким уровнем экспрессии, может и будет варьировать в зависимости от антигена и типа необходимого иммунного ответа. Способы определения того, вызывает ли антиген иммунный ответ, например, с помощью измерения уровней антител или антигензависимых Т-клеточных популяций или уровней антигензависимых цитокинов известны в данной области техники, и способы измерения уровней экспрессии последовательностей, кодирующих антигены, с помощью определения уровней транскрибируемой мРНК или с помощью количественной оценки уровня экспрессии белка также известны в данной области техники.
[00169] В каждом из вышеуказанных вариантов осуществления рекомбинантная бактерия, способная к регулируемой экспрессии, также может быть аттенуированной. Термин «аттенуированный» относится к состоянию бактерии, при котором бактерия была ослаблена по отношению к своей приспособленности дикого типа в результате некоторой формы рекомбинантной или физической манипуляции. Оно включает в себя изменение генотипа бактерии с целью снижения ее способности вызывать заболевание. В то же время, способность бактерии колонизировать кишечник (в случае Salmonella) и индуцировать иммунные ответы, предпочтительно, по сути не нарушена.
[00170] В соответствии с иллюстративным вариантом осуществления рекомбинантная бактерия может быть аттенуированной, как описано выше. В этом случае как регулируемая аттенуация, так и регулируемая экспрессия последовательности, кодирующей антиген, могут зависеть от регулируемой сахаром системы. Соответственно, концентрация сахара (например, арабинозы), необходимая для оптимальной экспрессии регулируемой последовательности, кодирующей антиген, может быть не такой же самой, что и концентрация для оптимальной экспрессии или аттенуации. В соответствии с иллюстративным вариантом осуществления концентрация арабинозы для оптимизации как регулируемой аттенуации, так и регулируемой экспрессии последовательностей, кодирующих антиген, будет по сути одинаковой.
[00171] Соответственно, промотор и/или последовательность нуклеиновой кислоты, кодирующая белок для аттенуации, могут быть модифицированы для оптимизации системы. Способы модификации подробно описаны выше. Вкратце, последовательность связывания рибосомы SD может быть изменена и/или старт-кодон может быть изменен с ATG на GTG для последовательностей нуклеиновой кислоты fur и phoPQ, в результате чего уровни продуцирования Fur и PhoPQ являются оптимальными как для фенотипа регулируемой аттенуации, так и для регулируемой экспрессии, при выращивании штаммов в присутствии определенной концентрацией арабинозы. Специалисту в данной области техники будет понятно, что другие последовательности нуклеиновой кислоты, помимо fur и phoPQ, также могут быть изменены, как описано в данном документе, в сочетании с другими хорошо известными протоколами. Помимо этого, эти аттенуированные последовательности нуклеиновых кислот можно регулировать с помощью других систем с использованием общепризнанных протоколов, известных специалисту в данной области техники. Например, их можно регулировать с помощью промоторов, зависимых от добавления мальтозы, рамнозы или ксилозы, а не арабинозы.
II. Фармацевтические композиции
[00172] Рекомбинантная бактерия может быть введена хозяину в виде фармацевтической композиции. В соответствии с некоторыми вариантами осуществления фармацевтическая композиция может быть использована в виде вакцины для вызова иммунного ответа в отношении рекомбинантной бактерии, в том числе антигенов, которые могут быть синтезированы и доставлены указанной бактерией. В соответствии с иллюстративным вариантом осуществления иммунный ответ является защитным. Иммунные ответы в отношении антигенов хорошо изучены и широко описаны.
[00173] Фармацевтические композиции могут быть введены любому хозяину, способному вызывать иммунный ответ. Такие хозяева могут включать в себя всех позвоночных, например, млекопитающих, в том числе домашних животных, сельскохозяйственных животных, лабораторных животных и людей, а также различные виды птиц, в том числе домашних птиц и птиц сельскохозяйственного значения. Предпочтительно хозяин представляет собой теплокровное животное. В соответствии с одним вариантом осуществления хозяин представляет собой корову. В соответствии с некоторыми вариантами осуществления хозяин представляет собой лошадь. В соответствии с другим вариантом осуществления хозяин представляет собой птицу. В соответствии с другим вариантом осуществления хозяин представляет собой человека. Фармацевтическая композиция может быть введена субъекту в виде профилактического средства или для лечебных целей.
[00174] В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия является живой при введении хозяину в фармацевтической композиции, описанной в данном документе. Подходящие составы вакцинных композиций и способы введения подробно описаны ниже.
[00175] Фармацевтическая композиция, содержащая рекомбинантную бактерию, может необязательно содержать одно или несколько возможных вспомогательных веществ, таких как носители, консерванты, стабилизаторы, адъюванты и другие вещества.
[00176] В соответствии с одним вариантом осуществления фармацевтическая композиция содержит адъювант. Адъюванты необязательно добавляют для повышения способности вакцины вызывать, усиливать или продлевать иммунный ответ. В соответствии с иллюстративными вариантами осуществления применение живой аттенуированной рекомбинантной бактерии может выступать в качестве природного адъюванта. В соответствии с некоторыми вариантами осуществления рекомбинантная бактерия синтезирует и секретирует иммуномодулятор. Дополнительные вещества, такие как цитокины, хемокины и бактериальные последовательности нуклеиновой кислоты, встречающиеся в природе в бактериях, например, CpG, также могут представлять собой потенциальные вакцинные адъюванты.
[00177] В соответствии с некоторыми вариантами осуществления фармацевтическая композиция содержит буферный солевой раствор (например, фосфатно-буферный солевой раствор (PBS)).
[00178] В соответствии с некоторыми вариантами осуществления фармацевтическая композиция содержит продукт питания.
[00179] В соответствии с другим вариантом осуществления фармацевтическая композиция может содержать фармацевтический носитель (или наполнитель). Такой носитель может представлять собой любой растворитель или твердое вещество для инкапсуляции, которые являются нетоксичными в отношении неинокулированного хозяина и совместимыми с рекомбинантной бактерией. Носитель может придавать форму или консистенцию или может выступать в качестве разбавителя. Подходящие фармацевтические носители могут включать в себя жидкие носители, такие как физиологический раствор, и другие нетоксичные соли в физиологических или приблизительно физиологических концентрациях, а также твердые носители, которые не используются для людей, такие как тальк или сахароза, или корм для животных. Носители могут также включать в себя физиологические средства, увлажняющие и эмульгирующие средства, соли для варьирования осмолярности, инкапсулирующие средства, буферы и средства, усиливающие проникновение через кожу. Носители и наполнители, а также составы для парентерального и непарентерального введения лекарственного препарата изложены в Remington's Pharmaceutical Sciences 19th Ed. Mack Publishing (1995). При использовании для введения через бронхиолы фармацевтическая композиция предпочтительно представлена в форме аэрозоля.
[00180] В соответствии с некоторыми вариантами осуществления фармацевтическую композицию доставляют сельскохозяйственному животному (например, домашней птице). В соответствии с некоторыми вариантами осуществления фармацевтическую композицию доставляют в виде спрея (например, для применения в инкубаторах для доставки домашней птице). В соответствии с некоторыми вариантами осуществления фармацевтическую композицию доставляют в питьевой воде.
[00181] Необходимо соблюдать осторожность при использовании вспомогательных веществ для того, чтобы живая рекомбинантная бактерия не была уничтожена или имела способность эффективно колонизировать лимфоидные ткани, такие как GALT, NALT и BALT, нарушенные в результате использования вспомогательных веществ. Стабилизаторы, такие как лактоза или глутамат натрия (MSG), могут быть добавлены для стабилизации фармацевтической композиции в отношении ряда условий, таких как температурные изменения или процесс сублимационной сушки. Рекомбинантная бактерия также может быть введена совместно с глутаматом и/или аргинином, как описано в данном документе.
[00182] Дозы фармацевтической композиции могут и будут варьировать в зависимости от рекомбинантной бактерии, регулируемого антигена и предполагаемого хозяина, как будет понятно специалисту в данной области техники. В самом общем виде доза должна быть достаточной только для того, чтобы вызвать защитный иммунный ответ у большинства хозяев. С помощью стандартного эксперимента можно легко установить требуемую дозу. Типичные начальные дозы вакцины для перорального введения могут составлять от приблизительно 1×107 до 1×1010 CFU в зависимости от возраста хозяина, подлежащего иммунизации. Введение множественных доз также может быть использовано при необходимости для обеспечения необходимого уровня защитного иммунитета.
[00183] С целью стимуляции предпочтительного ответа клеток GALT, NALT или BALT введение фармацевтической композиции непосредственно в кишечник, носоглотку или бронх является предпочтительным, например, с помощью перорального введения, интраназального введения, желудочного зондирования или в форме аэрозолей, хотя другие способы введения рекомбинантной бактерии, такие как внутривенная, внутримышечная, подкожная инъекция или интрамаммарное, интрапениальное, интраректальное, вагинальное введение или другие способы парентерального введения возможны, например, для противоопухолевых областей применения.
[00184] В соответствии с некоторыми вариантами осуществления эти композиции составляют для введения с помощью инъекции (например, интраперитонеально, внутривенно, подкожно, внутридермально, внутримышечно и т.д.).
[00185] В соответствии с другим вариантом осуществления в настоящем раскрытии предусмотрен способ вызова иммунного ответа против антигена у хозяина. Способ предусматривает введение хозяину эффективного количества фармацевтической композиции, содержащей рекомбинантную бактерию, описанную в данном документе.
[00186] В соответствии с еще одним вариантом осуществления рекомбинантная бактерия может быть использована в способе вызова иммунного ответа против патогена у индивидуума, нуждающегося в этом. Способ предусматривает введение хозяину эффективного количества фармацевтической композиции, содержащей рекомбинантную бактерию, описанную в данном документе. В соответствии с дополнительным вариантом осуществления рекомбинантная бактерия, описанная в данном документе, может быть использована в способе для нормализации одного или нескольких симптомов инфекционного заболевания у хозяина, нуждающегося в этом. Способ предусматривает введение эффективного количества фармацевтической композиции, содержащей рекомбинантную бактерию, описанную в данном документе.
Примеры
[00187] Настоящее изобретение дополнительно иллюстрируется следующими примерами, которые никоим образом не должны носить ограничивающего характера. Содержание всех цитируемых ссылок, в то числе цитируемых литературных источников, выданных патентов и опубликованных заявок на патент, цитируемых во всей настоящей заявке, тем самым явно включены в данный документ посредством ссылки. Необходимо дополнительно понимать, что содержание всех фигур и таблиц, прикрепленных к данному документе, также явно включено в данный документ посредством ссылки.
Пример 1: Предпосылки
[00188] Защитный иммунитет в отношении Salmonella зависел от сочетанного действия определенных антител, В-клеток и Т-клеточных приобретенных иммунных реакций (52-56). Эффективное устранение первичной инфекции требовало ответа Th1 при содействии ограничивающей антителами бактериемии (55, 57). Ответы антител был важны для достижения защиты против инфекции, вызываемой Salmonella (58-60), как наблюдали в отношении защиты против S. Typhi у мышей (61) и людей (62-64). RASV индуцировали все три ветки иммунной системы (т.е., ответы антител слизистых оболочек и клеточные ответы, а также ответы системных антител и клеточные ответы). Все три ветки иммунной системы были важны для обеспечения защитного иммунитета в отношении Salmonella и всех патогенов, которые колонизируют или вторгаются через поверхность слизистых оболочек.
[00189] Salmonella имела ряд иммунологически родственных перекрестно-реагирующих антигенов. Они включали в себя коровый полисахарид LPS, который был одинаковым у большинства, если не у всех, сероваров S. enterica (65, 66), за исключением S. Arizonae (67, 68). Помимо этого, ОМР, несмотря на то, что характеризовались микрогетерогенностю, тем менее, имели общие антигенные детерминанты (69), а также регулируемые железом белки внешней мембраны (IROMP) (70), которые требовались для захвата железа (70), обязательной важной функции для успешности патогена в организме инфицированного животного.
[00190] Вакцины против Salmonella могли быть использованы для дисплея поверхностных антигенных детерминант дикого типа in vitro и во время начальной фазы инфицирования через поверхности слизистых оболочек у перорально иммунизированного хозяина, а затем для прекращения синтеза боковых цепей О-антигена LPS в результате мутации Δpmi (71-74) и для конститутивного синтеза IROMP во внутренних органах (75, 76) в результате мутации по типу делеции/вставки ΔPfur::TT araC ParaBAD fur (ΔPfur) (77). Штаммы S. Typhimurium с мутацией Δpmi были не полностью аттенуированными, однако характеризовались высокой иммуногенностью, эффективностью усиления индукции высоких титров антител в отношении перекрестных ОМР, IROMP (76) и консервативного кора LPS (78, 79). В то же время, кор LPS не полностью демонстрировался, поскольку еще два сахара связывались с кором LPS. Штаммы с мутацией Δpmi также усиливали продуцирование везикул внешней мембраны (OMV), которые доставляли рекомбинантные защитные антигены с целью повышения защитного иммунитета (80). Мутация ΔPfur способствовала тому, что экспрессия гена fur полностью зависела от присутствия арабинозы (75, 81, 82) и была невосприимчивой к концентрации железа для достижения высокого уровня конститутивного синтеза in vivo всех компонентов для захвата железа, в том числе иммунологически перекрестно-реагирующих IROMP. Иммунные ответы в отношении высокоиммуногенных IROMP были эффективными в предупреждении септицемической инфекции с участием энтеропатогенов (83). Антитела, индуцированные в отношении IROMP из одного серотипа бактерий, могли распознавать IROMP, синтезированные другими серотипами (84). Две инактивированные вакцины на основе сверхпродукции IROMP были разрешены для защиты против сальмонеллеза у домашней птицы (85, 86).
[00191] Живые Salmonella, доставляющие как поверхностные полисахариды, так и ОМР, иммунной системе, были более иммуногенными, чем гликоконъюгатные вакцины. Salmonella спонтанно высвобождала 50-90 нм пузырькообразные частицы внешней мембраны клеточной стенки (87-89). Эти пузырьки, называемые GMMA (генерализованные модули мембранных антигенов) или везикулами внешней мембраны, представляли собой обогащенный источник ассоциированных с внешней мембраной антигенов в их нативной конформации и надлежащей ориентации. GMMA или везикулы внешней мембраны обеспечивали значительные преимущества по отношению к рекомбинантным белкам, поскольку они содержали многочисленные патоген-ассоциированные молекулярные паттерны (РАМР), в том числе лиганды TLR, которые характеризовались потенциалом для функционирования в качестве самоадъювантов при иммунных ответах, которые они вызывали (90-95). GMMA или везикулы внешней мембраны также отличались от экстрагированных с помощью детергентов ОМР, которые утрачивали ряд компонентов внешней мембраны, например, липопротеины, и, тем самым, приводили к сниженной иммуногенности. GMMA или везикулы внешней мембраны в настоящее время исследуют в качестве вакцин для лечения менингококка (96, 97), Shigella (87) и Salmonella (27). Доклинические исследования с использованием кандидатных GMMA или везикул внешней мембраны свидетельствовали об удовлетворительной иммуногенности и широком перекрестном иммунитете против ряда штаммов (98). Прототипные менингококковые GMMA или везикулы внешней мембраны исследовали в одном клиническом исследовании фазы 1, при этом обнаружили отсутствие побочных эффектов (99), а прототипные GMMA Shigella или везикулы внешней мембраны планировали для испытания фазы 2. Продуцирование GMMA или везикул внешней мембраны могли усиливать в результате делеции гена tolR (87, 100, 101), как наблюдали у мутантов по tolR Salmonella и Shigella (87, 102). Делеции генов, таких как htrB (88) и msbB (103) для модификации липида А могли приводить к ослаблению реактогенности. Несмотря на то, что новые вакцины на основе GMMA или везикул внешней мембраны характеризовались снижением числа этапов очистки, поскольку они спонтанно высвобождались с помощью соответствующих вакцинных бактериальных штаммов, все еще требовались последующие процедуры, например, комплексное фильтрование тангенциальным потоком для очистки GMMA или везикул внешней мембраны (87, 104, 105). В отличие от этого, в настоящем раскрытии предусмотрены системы продуцирования GMMA или везикул внешней мембраны in vivo с целью отказа от последующих процедур очистки без нарушения эффективности.
[00192] Из поверхностно демонстрируемых или секретируемых защитных антигенов у Salmonella могли использовать 6 антигенов, FliC, OmpC, OmpD, OmpF, SseB и SseI (106-113). Эти антигены не были наиболее распространенными у Salmonella (106). Синтез FliC даже характеризовался дерегуляцией в системных участках (114-116). Доклинические исследования у мышей продемонстрировали, что иммунизация вышеуказанными антигенами могла приводить к защите против заражения Salmonella (57, 111, 117-120). OmpC и F индуцировали длительные ответы у мышей (121) и, как было показано, являлись безопасными и иммуногенными при испытании в исследовании фазы 1 у людей (122). OmpD представлял собой основную мишень для B1b-клеточного ответа антител, независимого от Т-клеток (57, 111), и был консервативным во всех сероварах Salmonella, за исключением серовара Typhi (123, 124). Субъединица транслокона SPI-2 SseB играла важную функцию для секреции эффектора T3SS и репликации Salmonella в клетке (125). Она представляла собой серодоминантную мишень адаптивного иммунитета у детей с бактериемией, вызванной Salmonella (120), и включала в себя многочисленные эпитопы для CD4 Т-клеточного иммунитета у людей-добровольцев (108, 120, 126). Другой эффектор SPI-2, SseI, играл роль в модулировании миграции инфицированных клеток и требовался для длительной системной инфекции (127-131). Сохранение надлежащей конформации таких антигенов было важным, что было обнаружено при неспособности рекомбинантных поринов Salmonella оказывать защитное действие у мышей (132). RASV способствовали доставке этих антигенов в их надлежащей конформации и ориентации при высоких уровнях продуцирования, в сочетании с самоадъювантными свойствами Salmonella, которые доставляли присущие им сигналы посредством лигандов TLR и других РАМР с целью индукции специфического в отношении Salmonella Т-клеточного и В-клеточного иммунитета.
[00193] В данном документе раскрыта инновационная платформа RASV для сверхпродуцирования защитных антигенов Salmonella in vivo. Указанная система представляла собой уникальную регулируемую тремя сахарами систему, с двойным отключением синтеза О-антигена при использовании рамнозы и маннозы и сверхпродуцированием GMMA или везикул внешней мембраны при использовании арабинозы. Она также включала системы RDA и RDPS. Эти системы не приводили к повышению вирулентности (в результате введения этих самоантигенов), поскольку гены антигена активно не экспрессировались in vivo (106). Сверхэкспрессия генов антигена также приводила к аттенуации штамма (133, 134), как показано при сверхэкспрессии гена флагеллина (133, 135). Также могли оценивать вирулентность штаммов с хромосомными мутациями и без них для этих генов антигена при переносе генов антигена экспрессионными плазмидами, как дополнительно описано ниже. В случае, если экспрессионная плазмида достаточно сильно повышала вирулентность, чтобы вызвать заболевание, штамм или плазмиду могли модифицировать с тем, чтобы обеспечить свойства аттенуации. Уровни экспрессии генов могли модифицировать в результате повышения или снижения, при необходимости, в результате переключения регулируемых сахарами промоторов, изменения промоторов и нуклеотидной последовательности Шайна-Дальгарно и разделения этих элементов и старт-кодонов регулируемого гена.
Пример 2: Материалы и способы
[00194] Бактериальные штаммы, среда и рост бактерий. Конструирование штаммов выполняли в вирулентном штамме S. Typhimurium χ3761 (75) и S. Enteritidis χ3550 (136). Для заражений использовали различные вирулентные серовары Salmonella дикого типа, в том числе S. Typhimurium χ3761 (В), S. Enteritidis χ3550 (D), S. Heidelberg χ3749 (В) (137), S. Choleraesuis χ3246 (С1) (138), S. Mantis χ3213 (С1) (139), S. Newport χ3240 (С2) (139), S. Dublin χ12323 (D) (140). Значения LD50 большинства этих штаммов находились между 103 и 105 у мышей и цыплят, за исключением того, что S. Heidelberg, S. Infantis и S. Newport часто не приводили к летальному заболеванию ни у мышей, ни у цыплят. Среду LB или планшеты с подходящими добавками, при необходимости, использовали для роста Salmonella (141, 142).
[00195] Молекулярные и генетические процедуры. Способы работы с ДНК и ПЦР были стандартными (143). Анализ последовательностей ДНК выполняли в лаборатории последовательностей ДНК UFL, в то время как синтез олигонуклеотидов и/или генных сегментов осуществляли коммерчески. Конструирование делеций или делеций/вставок у Salmonella осуществляли с помощью суицидных векторов или трансдукции Р22 (144-146).
[00196] Характеристика штаммов. Вакцинные штаммы полностью характеризовали на каждом этапе их конструирования и перед исследованиями иммунизации на наличие всех фенотипов и генотипов. Генетические свойства подтверждали с помощью ПЦР с использованием соответствующих зондов и/или анализов фенотипов. Метод поступления флуоресцентного красителя использовали для оценки проницаемости мембран мутантов. Штаммы сравнивали с векторными контрольными штаммами в отношении стабильности сохранности плазмид и синтеза антигенов, когда штаммы выращивали в присутствии арабинозы или других Сахаров и/или DAP в течение периода 50 поколений (147). LPS проверяли с помощью окрашивания серебром (148). Кривые роста определяли для каждого штамма. Другие эксперименты включали определение профилей ОМР (147) и IROMP (149), характеристику OMV (80) и GMMA(87, 150, 151), устойчивости сывороточных (152), желчных и микробных пептидов (136), а также прикрепление/инвазию в эпителиальные клетки INT-407 (153, 154). Каждый штамм с определяющей антиген плазмидой оценивали в отношении синтеза гетерологичного антигена с помощью вестерн-блоттинга.
[00197] Получение антигенов. Защитные антигены FliC, OmpC, OmpD, OmpF, SseB и SseI с С-терминальной His-меткой клонировали в векторы pBAD-His или рЕТ для синтеза в Тор10 или BL21 Е. coli и выделяли с помощью хроматографии с использованием никелевой колонки (Sigma). Очищенные белки использовали для анализов ELISA и ELISPOT и для приготовления антисыворотки у самок новозеландских кроликов. О-антигены LPS Salmonella получали коммерчески. Белки внешней мембраны S. Typhimurium (SOMP) очищали из штамма χ9424, который конструировали таким образом, чтобы он не был способен образовывать жгутики, все экспрессируемые in vitro антигены пилуса, О-антиген LPS и несколько капсул. Другие ОМР Salmonella очищали из соответствующих мутантов по О-антигену (147).
[00198] Статистические показатели. Программу SAS использовали для расчета статистических критериев, а анализ мощности для оценки количества животных.
Пример 3: Конструкция плазмид с регулируемым сахарами синтезом GFP с целью обеспечения определения того, способен ли штамм, неспособный метаболизировать сахар, усваивать этот сахар для обеспечения регуляции гена или последовательности гена в указанном штамме.
[00199] На Фиг. 1 в графическом виде изображены три плазмиды pYA3700, pYA5351 и pG8R74, которые имеют кассеты araC ParaBAD, rhaRS-PrhaB и xylR-PxylA соответственно в виде источников ДНК, кодирующих эти кассеты, с целью обеспечения получения суицидных векторов со слиянием определенной регуляторной кассеты с выбранным геном вместо промотора для указанного гена. Указанные манипуляции описаны в Примере 1, а штаммы с образующимися в результате мутациями типа делеция/вставка с экспрессией регулируемых сахарами генов описаны в следующих примерах.
[00200] Нуклеотидная последовательность кассеты rhaRS-PrhaB в pYA5351 представлена ниже:



[00201] Нуклеотидная последовательность кассеты xylR-PxylA в pG8R74 представлена ниже:


[00202] На Фиг. 2 описаны плазмиды pG8R115, pG8R116 и pG8R192, которые приводили к зависимому синтезу GFP в присутствии арабинозы, рамнозы или ксилозы соответственно. Указанные плазмиды могли электропорировать в любой штамм многочисленных видов бактерий с помощью отбора в отношении устойчивости к ампициллину, а затем подвергать скринингу в отношении синтеза GFP в присутствии сахара, представляющего интерес, и прекращения синтеза GFP в отсутствие указанного сахара. В случае, если наблюдали синтез GFP, то могли конструировать мутантные штаммы, в которых промотор гена, представляющего интерес, удаляли и замещали кассетой araC ParaBAD, rhaRS-PrhaB или xylR-PxylA таким образом, что экспрессия гена зависела от присутствия арабинозы или рамнозы или ксилозы соответственно. Следует отметить, что указанная способность была очень полезной в случае, если штамм или вид бактерии, представляющий интерес, были неспособны метаболизировать арабинозу, рамнозу или ксилозу или расти на них, в результате чего было неизвестно, могли ли эти сахара транспортироваться в бактериальные клетки, что было бы необходимым для использования присутствия этого сахара для экспрессии генов в указанных штамме или виде бактерии.
Пример 4: Выделение и характеристика штаммов с мутациями galE с целью обеспечения функционального обратимого синтеза LPS, зависимого от присутствия свободной галактозы. Конструирование вакцинных векторных штаммов с мутациями galE с целью обеспечения регулируемого синтеза UDP-Gal, необходимого для синтеза боковых цепей О-антигена LPS и внешнего кора LPS, и с целью обеспечения роста в присутствии галактозы без токсичности и потенциального отбора устойчивых к галактозе вариантов для снижения иммуногенности.
[00203] Ген galE кодировал UDP-галактозо-4-эпимеразу, которая взаимопревращала UDP-галактозу и UDP-глюкозу. В качестве части катаболизма галактозы galE связывали как с синтезом, так и с распадом галактозы. Все поверхностные липополисахариды (LPS) вида Salmonella содержали галактозные единицы в коре LPS и в боковой цепи О-антигена LPS. Поскольку UDP-галактоза представляла собой предшественник галактозных единиц в LPS, то мутант по galE синтезировал дефектный в отношении кора или «шероховатый» LPS до тех пор, пока не доставляли экзогенную галактозу (47). Мутант по galE S. Typhimurium был авирулентным и обеспечивал защиту против заражения вирулентной S. Typhimurium у мышей (47, 50, 155). Некоторые из мутантов по galE S. Choleraesius также были авирулентными (156, 157). Единственной разрешенной живой аттенуированной бактериальной вакциной, одобренной для применения у человека, была S. Typhi Ту21а с мутацией galE, а также другими мутациями (30, 158, 159). S. Typhi Ту21а была лишь частично аттенуированной и придавала умеренный защитный иммунитет против брюшного тифа в полевых исследованиях у человека (30, 47, 160-163).
[00204] В то же время единственная мутация galE в S. Typhi все еще способствовала вирулентности у людей и обеспечивала лишь умеренную защиту против брюшного тифа (30, 47, 161, 163, 164), как также наблюдали в случае с мутантами по galE S. Choleraesuis (165). Помимо этого, мутанты по galE Salmonella были восприимчивы к галактозе с целью индукции лизиса in vitro (48, 166, 167). Даже определенная делеция galE в S. Typhimurium придавала восприимчивость к индуцированному галактозой лизису, которая являлась нежелательным свойством вакцины. Разрешенный штамм Ту21а также характеризовался указанным нежелательным свойством. Мутанты по galE не имели фермента UDP-галактозо-4-эпимеразы, однако сохраняли способность усваивать галактозу из экзогенных источников посредством транспортеров галактозы (168, 169). При выращивании в присутствии галактозы галактокиназа и галактозо-1-фосфатуридилтрансфераза, кодируемые генами galK и galT соответственно, могли обеспечивать синтез UDP-галактозы из галактозы с использованием галактозо-1-фосфата и приводили к остановке клеточного роста и даже лизису. Точный механизм лизиса был неизвестен, однако гибель коррелировала с ростом и внутриклеточным накоплением метаболитов галактозы, особенно накоплением галактозо-1-фосфата и UDP-галактозы вследствие активности галактокиназы (30, 48). Накопление UDP-D-галактозы приводило к остановке роста вследствие низкой доступности СТР и UTP, что приводило с снижению синтеза РНК (49). Мутанты по galE росли слабо и их жизнеспособность была значительно снижена в результате лиофилизации (50). Авирулентность мутантов по galE главным образом была связана с неполным липополисахаридом клеточной стенки и индуцированным галактозой лизисом бактериальной клетки (30). Это способствовало отбору в отношении устойчивости к галактозе и Gal+ фенотипу (170). Лизис мог происходить при концентрациях галактозы всего 0,002% (166). Индуцированный галактозой лизис происходил в штамме Ту21а in vitro при концентрации галактозы ≥6 мМ (30, 47), в то время как Ту2 с определенной мутацией galE был даже более восприимчивым к ≥0,06 мМ галактозы (50, 163). Рост мутантов по galE в присутствии галактозы также приводил к отбору устойчивых к галактозе штаммов, которые утрачивали способность демонстрировать обратимую вариацию «шероховатый-гладкий» в зависимости от поступления галактозы (170). Глюкоза могла приводить к защите восприимчивых к галактозе штаммов galE от лизиса в результате катаболитной репрессии до той степени, когда лизис промежуточных продуктов галактозы достигал уровней, когда они не могли накапливаться (167, 171). Снижение активности галактокиназы также могло придавать штамму устойчивость к галактозе (48, 166, 167). Таким образом, с целью преодоления указанной проблемы и расширения использования мутаций galE, придающих обратимую вариацию «шероховатый-гладкий», которая выступает в качестве средства регулируемой замедленной аттенуации для вакцин, конструировали штамм с новой мутацией galE с повышенной устойчивостью к галактозе и в то же время проявлением регулируемой аттенуации, зависимой от присутствия добавленной галактозы, которая является недоступной in vivo. Затем оценивали включение указанной мутации в вакцинные штаммы S. Typhimurium.
[00205] Получали три штамма с различными мутациями galE. Штамм χ4094 характеризовался мутацией восприимчивости к галактозе galE496, как наблюдали в случае большинства мутантов по galE. Штамм χ4700 характеризовался неописанной мутацией в виде делеции Δ(galE-uvrB)-1005, которая способствовала тому, что штаммы характеризовались невосприимчивостью к галактозе. Штамм χ9792 характеризовался мутацией невосприимчивости к галактозе Δ(galE-ybhC)-851, которая приводила к делеции последовательностей из 11 генов из galE в ybhC (Фиг. 3А). Указанному штамму требовалась 0,001% галактоза в среде для роста для образования комплекса О-антигена LPS (Фиг. 3В) либо в питательном бульоне, либо в бульоне LB.
[00206] Для определения того, влияло ли добавление галактозы на рост штаммов Salmonella с различными мутациями galE, выполняли эксперименты с исследованием роста. В первом эксперименте оценивали конечные OD культур, полученных в течение ночи, с варьирующими концентрациями галактозы в бульоне LB или бульоне NB. Необходимо отметить, что бульон NB был лишен всех сахаров, в результате чего отсутствовали помехи в результатах вследствие следовых количеств галактозы. В среде LB OD культуры, полученной в течение ночи, χ4094 составлял 1,088 при использовании 0,001% галактозы, однако снижался до 0,159 при использовании 0,01% галактозы. OD χ4700 и χ9792 не подвергались значительному влиянию варьирующих концентраций галактозы. Аналогичные тенденции наблюдали, когда мутанты по galE выращивали в бульоне NB с варьирующими концентрациями галактозы (Фиг. 3С). В целом данные подтверждали, что χ9792 (Δ(galE-ybhC)-851)) не был таким восприимчивым к галактозе, как χ4094.
[00207] Во втором эксперименте оценивали рост штаммов во время 7-часового периода в среде для роста с варьирующими концентрациями галактозы (Фиг. 4А-4Н). Культуру каждого штамма, полученную в течение ночи, выращивали в бульоне NB без галактозы. Субкультурирование выполняли с помощью разведения при 1:100 в предварительно нагретых 3 мл бульона NB с варьирующими процентными концентрациями галактозы (0, 0,001, 0,01, 0,02, 0,05, 0,1, 0,2, 0,5). Культуры инкубировали при 37°С при встряхивании. Оптические плотности измеряли и фиксировали через каждый час. В отсутствие галактозы все штаммы росли аналогичным образом. Штамм χ4094 характеризовался мутацией восприимчивости к галактозе galE496. Штамм χ4700 характеризовался мутацией невосприимчивости к галактозе Δ(galE-uvrB)-1005. Штамм χ9792 характеризовался мутацией невосприимчивости к галактозе Δ(galE-ybhC)-851. Штамм χ11015 характеризовался мутацией невосприимчивости к галактозе Δ(galE-ybhC)-851 ΔgalP211. Штамм χ11141 характеризовался мутацией невосприимчивости к галактозе Δ(galE-ybhC)-851 ΔgalP211 ΔmglBAC. На Фиг. 4А-4Н показано, что мутации ΔgalP211 и ΔmglBAC способствовали достижению штаммом более высоких значений OD.
[00208] Как показано на Фиг. 4В, штамм χ4094 вырастал в течение 2 часов и затем начинал лизировать даже в присутствии 0,001% галактозы. При возрастающих концентрациях галактозы начальное время для лизиса снижалось. Как штамм χ4700, так и штамм χ9792 могли переносить галактозу до 0,5%, при этом нарушение роста отсутствовало (см. Фиг. 4А-4Н). Эти результаты продемонстрировали, что высокие концентрации галактозы не подавляли рост и колонизацию штамма с мутацией Δ(galE-ybhC). Повышение переносимости галактозы способствовало тому, что указанный штамм проявлял более высокую колонизацию тканей, чем штамм с мутацией восприимчивости galE496 в день 6 (Фиг. 5) после пероральной инокуляции мышей. Данные подтверждали, что мутацию Δ(galE-ybhC)-851 можно использовать в вакцинных штаммах для обеспечения обратимого фенотипа «шероховатый-гладкий», зависимого от присутствия галактозы в среде для роста, и она обеспечивала дополнительное средство регулируемой замедленной аттенуации in vivo, поскольку свободная нефосфорилированная галактоза не присутствует в тканях животных.
Пример 5: Конструкция и оценка штаммов RASV S. Typhimurium группы В с регулируемым рамнозой замедленным синтезом О-антигена, регулируемым маннозой замедленным синтезом боковых цепей О-антигена и регулируемым арабинозой продуцированием GMMA или везикул внешней мембраны, синтез защитных антигенов in vivo.
[00209] О-антиген-лигаза WaaL была необходима для лигирования полисахарида с фрагментом кора липида А. Мутация waaL приводила к образованию интактного кора с отсутствием О-антигена, присоединенного к нему (172, 173). Выполняли делецию waaL в опероне и помещали регулируемый рамнозой waaL (ΔRrhaBAD waaL) в ген pagL, поскольку мутация pagL не нарушала вирулентность Salmonella (174). Рамноза замещала арабинозу с достижением подавления синтеза О-антигена in vivo, поскольку относительно высокая концентрация рамнозы была необходима для активации указанного промотора (175). Штаммы RASV с регулируемым рамнозой waaL приводили к синтезу нормального LPS в присутствии рамнозы in vitro, однако образовывали шероховатый LPS вследствие отсутствия лигазы О-антигена in vivo. В результате указанной стратегии происходила демонстрация консервативного олигосахарида кора LPS и усиление продуцирования консервативных ОМР, в том числе поринов (176, 177), приводя к более эффективной презентации консервативных ОМР иммунной системе хозяина с целью усиления иммуногенности и облегчения получения перекрестного иммунного ответа против гетерологичных бактерий (173).
[00210] Мутантный штамм аттенуировали до 109 CFU и он обеспечивал защиту против заражения как S. Typhimurium, так и S. Enteritidis при 109 CFU (Табл. 1). В то же время данная мутация не была полностью аттенуированной, поскольку она по прежнему приводила к летальному исходу при 109. Таким образом, мутацию Δpmi также включали в качестве двойного отключения синтеза О-антигена в штамме χ12339. RASV с мутациями ΔPrhaBAD waaL и Δpmi нуждались как в рамнозе, так и в маннозе для образования полного О-антигена. С целью дополнительного повышения защиты мутацию ΔPfur включали для повышения синтеза IROMP in vivo с целью усиления индукции перекрестного иммунитета в отношении кишечных патогенов, как выполняли в штамме χ12362.

[00211] Все мутации были связаны с повышением презентации консервативных белков с целью облегчения индукции перекрестного иммунитета и достижения регулируемой замедленной аттенуации. Поскольку мутация tolR могла приводить к повышению продуцирования GMMA или везикул внешней мембраны (100, 101), кандидатные RASV дополнительно модифицировали с помощью введения регулируемой арабинозой мутации tolR (ΔPtolR:: ТТ araC ParaBAD tolR, в упрощенном виде обозначаемой ΔPtolR далее в данном документе) с целью повышения продуцирования GMMA или везикул внешней мембраны in vivo для сведения к максимуму индукции антител, перекрестно-реактивных в отношении ОМР других сероваров Salmonella. Кроме того, плазмиды, кодирующие защитные антигены, вводили в вакцинные штаммы с целью оценки защитного иммунитета. Поскольку кандидатные гены либо подвергались репрессии, либо экспрессировались при низких уровнях in vivo (106), сверхпродуцирование этих антигенов облегчало их презентацию (106, 108).
[00212] Конечный штамм нуждался в арабинозе, маннозе и рамнозе, чтобы функционировать в качестве дикого типа и достигать аттенуации in vivo постепенно (Табл. 2). Регулируемые рамнозой и маннозой гены вначале утрачивали свою функцию с целью демонстрации поверхностных антигенов, а затем отключение регулируемых арабинозой генов приводило к повышению продуцирования GMMA или везикул внешней мембраны.

[00213] Конструкция штаммов. Штамм χ12470 получали с использованием штамма χ12337 и последующего последовательного добавления мутаций Δpmi, ΔPfur и ΔPtolR (Табл. 2). Мутации ΔrelA::araC ParaBAD lacI TT (ΔrelA) в случае RDPS (180), ΔasdA в случае системы сбалансированных леталей и Δ(wza-wcaM) с целью устранения синтеза экзополисарахидов (Табл. 2) вводили в штамм χ12465 с целью облегчения его применения в качестве вектора. Мутацию ΔPtolR добавляли с целью получения штамма χ12473. После подтверждения конечного штамма с помощью фенотипического и ПЦР-анализа плазмиды Asd+, несущие ген отдельного антигена, вводили в штамм. Соответствующий ген антигена удаляли с помощью суицидного вектора (144, 181) из хромосомы с предупреждением потенциальной рекомбинации между генами в хромосоме и плазмиде. Тщательно изучали целостность мембран, продуцирование OMV (101), присутствие и стабильность всех фенотипических свойств штаммов. Регулируемые сахаром промоторы или SD или старт-кодон могли переключать с целью регуляции продуцирования лигазы О-антигена, Fur, TolR in vitro для приведения в равновесие иммуногенности и аттенуации (75).
[00214] Конструкция плазмид. Поскольку все антигены были поверхностно демонстрируемыми или секретируемыми, то природные последовательности генов экспрессировались с помощью вектора сбалансированных леталей Asd+ pYA3342 (Ptrc, ori pBR) (147). Присутствие RDPS приводило к репрессии экспрессии генов антигенов in vitro с помощью арабинозы, но к повышению экспрессии in vivo (180). Смещение в отношении низкокопийной плазмиды pYA3337 (Ptrc, ori pSC101) (182) или pYA3332 (Ptrc, ori p15A) (183) выполняли в случае, если сверхпродуцирование приводило к метаболическому грузу, свидетельством чего был значимо более медленный рост.
[00215] Все гены, за исключением fliC, использовали в соответствии с их природной последовательностью. Усеченный FliC180, в котором присутствовала делеция 180 аминокислот, кодирующих варьирующий в отношении антигенов серовар-специфический гипервариабельный домен антигена флагеллина, использовали с целью снижения индукции титров антитела в отношении серовар-специфических антигенов и повышения перекрестной защиты от консервативного домена флагеллина. Белок FliC180 сохранял консервативные N- и С-концевые участки, которые взаимодействовали с TLR5 с целью рекрутинга/стимуляции врожденных иммунных ответов (184, 185) и CD4-зависимых Т-клеточных эпитопов (186). Вначале исследовали отдельный антиген, после чего исследовали несколько антигенов с помощью плазмиды, кодирующей несколько антигенов в виде оперона (183, 187, 188) или с помощью нескольких генов, которые регулировались независимо (189-193). Мутацию recF включали с целью снижения рекомбинации между антигенами в плазмиде (194).
[00216] Оценка RASV, экспрессирующих гены защитных антигенов in vitro. Способность штаммов RASV синтезировать и секретировать защитный антиген анализировали с помощью проведения исследований по фракционированию клеток с целью определения количества антигена, присутствующего в цитоплазматических, периплазматических и супернатантных фракциях с помощью вестерн-блоттинга. Штаммы выращивали в бульоне Лурия (LB) до OD600 0,8 при 37°С и центрифугировали. Жидкость супернаната сохраняли для анализа секретируемых белков. Периплазматические и цитоплазматические фракции готовили с помощью метода опосредованного лизоцимом осмотического шока (147, 195, 196). Равные объемы периплазматических, цитоплазматических и супернатантных фракций и образцы общего лизата анализировали с помощью вестерн-блотов, зондированных соответствующим антителом. Эксперименты с использованием культивирования тканей выполняли для оценки транслокации антигенов в макрофагоподобные клеточные линии мыши, J774.A и/или P388D1 с помощью вестерн-блотов и иммунофлуоресценции (197-199).
[00217] Эксперименты на животных. Самок мышей BALB/c в возрасте от шести до восьми недель использовали и содержали в закрытом помещении BSL2 с клетками, закрытыми крышками фильтров. В стандартные эксперименты включали группы из пятнадцати мышей для заражения (с одним повтором) (200). Дополнительных мышей использовали для определения колонизации и извлечения селезенок для иммунологических анализов. Колонизацию и иммуногенность оценивали для всех конструкций, синтезирующих консервативные защитные антигены Salmonella.
[00218] Мышей иммунизировали перорально в день 0 дозой ~ 109 CFU RASV, бустировали такой же самой дозой через 1 неделю и перорально заражали на неделе 4 100 х LD50 вирулентным штаммом Salmonella в соответствии со стандартными процедурами (200). Значения LD50 штаммов дикого типа были известны или их оценивали. Заболеваемость и смертность фиксировали ежедневно. Вначале сравнивали штаммы, несущие каждый отдельный антиген с контролем PBS против заражения S. enteritidis. Если в данном исследовании наблюдали защитный эффект, то проводили последующие исследования с целью определения перекрестной защиты против других сероваров Salmonella. Кровь, РР, печень и селезенку извлекали из зараженных мышей для подсчета Salmonella в тканях с целью определения кинетики элиминации жизнеспособных Salmonella в виде функции времени после заражения и отслеживания иммунных ответов после заражения.
[00219] Измерение иммунных ответов, придаваемых RASV, синтезирующими защитные антигены. IgG сыворотки крови и SIgA слизистых оболочек из вагинальных смывов у иммунизированных мышей оценивали с помощью ELISA с использованием защитных антигенов, ОМР, IROMP и LPS из различных сероваров через 2 и 4 недели, а также титры IgG1 и IgG2a с целью разграничения ответов Th1 и Th2. Через 4 недели после иммунизации ответы спленоцитов на стимуляцию очищенными антигенами Salmonella или Salmonella определяли с целью измерения Т-клеточного иммунитета с помощью ELISPOT с определением профиля CD4 Т-клеток, которые продуцируют IL-4, IFN-γ и IL-17 (201). Поскольку количество секретируемого IgA, полученного в вагинальных смывах, могло точно не отражать ответ слизистых оболочек в кишечнике, количество клеток, секретирующих IgA, присутствующих в собственной пластинке кишечника, измеряли с помощью антигенспецифического ELISPOT IgA. Сыворотки крови собирали для анализа цитокинов с помощью мультиплексного анализа через 24, 48 и 72 часа после заражения с помощью системы Bio-plex Protein Array (BIO-RAD) в соответствии с инструкциями производителя (202). Цитокины IL-2, IL-4, IL-6, IL-10, IL-17А, IFN-γ, TNF-α, IL-21 и IL-23 измеряли в результате кокультивирования Т-лимфоцитов с антигенами Salmonella с целью определения путей Т-клеточной дифференцировки среди Th1/Th2/Th17/Tfh с помощью анализа Bioplex (202-204). В частности, IL-1β и IL-18 контролировали в случае размножения бактерий в печени и селезенке (41, 205, 206), TNF-α и IL-6 в случае индуцированных LPS цитокинов (207, 208). Проточную цитометрию использовали для определения распределения В- и Т-клеток памяти в РВМС и тканях мышей (209, 210) и Т-клеточной пролиферации с помощью окрашивания CFSE (211-218).
Пример 6: Конструкция и оценка штаммов RASV S. Enteritidis группы D с регулируемым рамнозой замедленным синтезом О-антигена, регулируемым маннозой замедленным синтезом боковых цепей О-антигена и регулируемым арабинозой продуцированием GMMA или везикул внешней мембраны, синтез защитных антигенов in vivo.
[00220] Штаммы RASV Enteritidis с такими же самыми свойствами, что и RASV Typhimurium, синтезирующие защитные антигены, конструировали и оценивали параллельно с RASV Typhimurium в качестве дополнительной стратегии. Суицидные векторы, используемые для S. Typhimurium, могли использовать для S. Enteritidis в связи с высокой гомологией между этими двумя сероварами. Поскольку отсутствовала модель для S. Typhi, данное исследование также облегчало аппроксимацию результатов в отношении S. typhi, поскольку они обе принадлежали Salmonella группы D. Это также могло облегчать применение в качестве бивалентной вакцины или для иммунизации типа прайм-буст против большинства инфекций NTS (219).
Конструкция вакцинных штаммов RASV Enteritidis
[00221] Аналогичные стратегии использовали для конструирования штаммов S. enteritidis с мутациями ΔwaaL, ΔPrhaBAD waaL, Δpmi, ΔPfur и ΔPtolR, происходящими из S. enteritidis χ3550, с целью получения вакцинного штамма В1. Мутацию ΔPtolR добавляли к штамму χ12457, происходящему из штамма χ3550 с мутациями ΔwaaL, ΔPrhaBAD waaL, Δpmi и ΔPfur, с целью получения вакцинного штамма В1. Вирулентность полученного штамма оценивали у мышей BALB/c. Мутации с целью снижения токсичности липида А вводили в случае, если штамм все еще был вирулентным (220, 221). При условии, что штамм был аттенуированным, как предполагалось, иммунизированных мышей заражали перорально 100 х LD50 штамма S. Typhimurium дикого типа χ3761. если наблюдали защитный эффект, то в последующих исследованиях определяли перекрестную защиту против других сероваров Salmonella. В предположении, что штамм был достаточно аттенуированным и обеспечивал некоторый защитный эффект, вводили ΔrelA и ΔasdA с получением штамма В2 с целью облегчения его применения в качестве вектора.
Оценка in vitro вектора для доставки антигенов RASV Enteritidis.
[00222] Наиболее эффективный вектор из Примера 3 вводили в штамм В2. Полученный в результате рекомбинантный штамм оценивали в отношении синтеза антигенов, стабильности плазмид и других характеристик in vitro и in vivo, в целом описанных в Примере 3.
Эксперименты на животных и измерение иммунных ответов, придаваемых защитными антигенами, синтезируемыми RASV Enteritidis.
[00223] Аналогичные процедуры и исследования проводили, как в Примере 3, за исключением того, что вначале заражали S. Typhimurium, а затем другими сероварами.
Пример 7: Улучшенные функциональные характеристики RASV в отношении индуцированного Clostridium perfringens некротического энтерита у цыплят-бройлеров при использовании штаммов, проявляющих фенотип регулируемого замедленного лизиса in vivo и другие свойства аттенуации и синтеза защитных антигенов, зависимые от свойств, регулируемых двумя сахарами по сравнению с тремя сахарами.
[00224] С целью определения защитных эффектов рекомбинантного бактериального штамма или RASV, содержащих системы свойств, регулируемых тремя сахарами, по сравнению с системами свойств, регулируемых двумя сахарами, осуществляли следующие эксперименты.
I. Сравнительная иммуногенностъ и защитный иммунитет χ11802 по сравнению с χ12341
[00225] Цыплят-бройлеров перорально иммунизировали одним из следующих штаммов Salmonella enterica:
χ11802 ΔPmurA25::TT araC PBAD murA ΔasdA27::TT araC PBAD c2 Δpmi-2426 Δ(wza-wcaM)-8 ΔrelA198::araC PBAD lacI TT ΔrecF126 (регулируемые арабинозой и маннозой фенотипы), содержащим pYA5112 (описанный в Jiang et al. (2015) Avian Diseases 59:475-85 (188), полное содержание которой включено в данный документ посредством ссылки), кодирующую оперон для синтеза PlcC и слияния NetB в качестве защитных антигенов С. perfringens;
χ12341 ΔPmurA25::TT araC PBAD murA ΔasdA27::TT araC PBAD c2 Δpmi-2426 ΔwaaL46 ΔpagL64::TT rhaRS PrhaBAD waaL Δ(wza-wcaM)-8 ΔrelA197::araC PBAD lacI TT ΔrecF126 ΔsifA26 (регулируемые арабинозой, маннозой и рамнозой фенотипы), содержащим pYA3681 в качестве контроля пустым вектором или pYA5112, кодирующую оперон для синтеза PlcC и слияния NetB в качестве защитных антигенов С. perfringens.
[00226] В исследование включали 48 клеток, начиная с 384 цыплят. Обработку повторяли в 6 блоках, каждый из которых состоял из 8 клеток. Исследование начинали, когда птиц размещали (день выведения) (DOT 0), при этом их распределяли в экспериментальные клетки. Никаких птиц не замещали во время исследования.
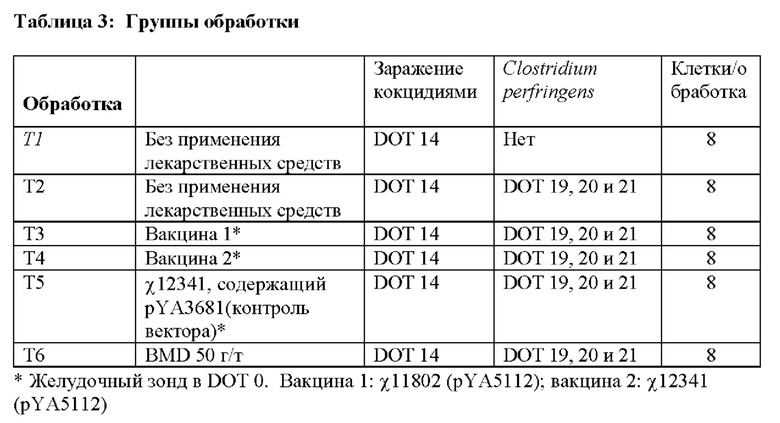
Рацион, используемый в эксперименте
[00227] Составляли стартерный корм для цыплят без содержания лекарственных средств, смешанный с кормовыми продуктами, широко используемыми в Соединенных Штатах Америки. Корм был характерным для местного коммерческого состава, а рассчитанные анализы соответствовали или превосходили требования NRC к стартерным кормам для цыплят. Состав корма включали в исходные данные. Корма для экспериментальной обработки готовили на основе указанного базового стартерного корма. Количества всех базовых кормов и исследуемых препаратов, используемых для приготовления партий для обработки, фиксировали. Корма для обработки смешивали для обеспечения однородного распределения соответствующего исследуемого препарата. Мешалку промывали для предупреждения перекрестной контаминации. Корм переносили в здание №2 и распределяли по клеткам с одинаковой обработкой. При внесении птиц кормили кормами для обработки. Указанный рацион (в форме смеси) скармливали во время указанного исследования. Корм взвешивали в DOT 0, а оставшийся корм взвешивали в DOT 14, 21 и 28.
Образцы корма
[00228] По одному образцу корма из начала, середины и конца каждой партии отбирали и смешивали с образованием комбинированного образца. Один образец извлекали из комбинации для каждой обработки и сохраняли в течение периода шести (6) месяцев после завершения исследования для потенциального анализа корма.
Животные
[00229] Однодневных петушков цыплят-бройлеров мужского пола получали от Cobb-Vantress, Кливленд, Джорджия. Порода представляла собой Cobb 500. Информацию о племенном стаде фиксировали. В инкубаторе у птиц определяли пол и они получали стандартные вакцинации. В исследовании использовали только цыплят, которые выглядели здоровыми. В начальный момент в каждой клетке было 8 цыплят (DOT 0). Всех птиц взвешивали в DOT 0, 14, 21 и 28.
Введение штаммов
[00230] Бактериальные штаммы вводили в DOT 0 каждому цыпленку в группах обработки 3, 4 и 5 через желудочный зонд в дозе ~5×108 CFU/цыпленок в объеме 0,1 мл.
Индукция заболевания
[00231] В DOT 14 всех птиц перорально инокулировали ~5000 ооцист Е. maxima.
[00232] Начиная с DOT 19 всем птицам (за исключением группы обработки 1) вводили бульонную культуру С. perfringens в дозе ~108 CFU/мл. В данном исследовании удаление корма отсутствовало. С помощью желудочного зонда птицам вводили 0,1 мл свежей бульонной культуры один раз в день в течение 3 дней (в DOT 19, 20 и 21).
Рост после заражения Clostridium perfringens
[00233] Используемым штаммом для заражения был Clostridium perfringens №6 (Hofacre, et al., 1998) (222). Его вводили в один (1) литр тиогликолятного бульона с добавлением 5% говяжьего экстракта и инкубировали при 37°С в течение 15 часов.
Количественное оценивание кишечных поражений вследствие некротического энтерита
[00234] Количественное оценивание кишечных поражений вследствие некротического энтерита выполняли, как описано в Hofacre, et al., 1998 (222). В DOT 21 трех птиц из каждой клетки через четыре (4) часа после третьего заражения Clostridium perfringens отбирали, умерщвляли, взвешивали и исследовали на наличие поражений вследствие некротического энтерита. Количественное оценивание было основано на шкале от 0 до 3 баллов, при этом 0 соответствовал норме, а 3 соответствовал наиболее высокой тяжести.
Анализ данных
[00235] Выполняли статистический анализ прироста массы в клетке, потребления корма, конверсии корма, баллов поражения и смертности вследствие NE. Результаты эксперимента показаны ниже в Табл. 4.

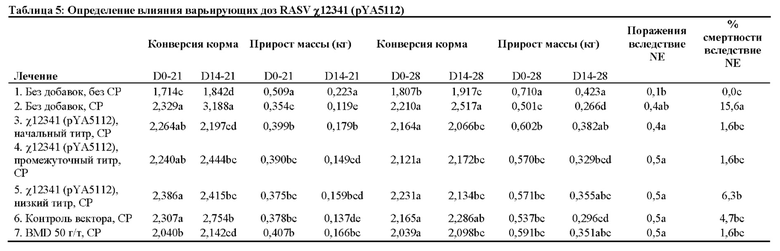
[00236] Как показано в Табл. 4, χ12341 (pYA5112) превосходил χ11802 (pYA5112) (а также контроль вектора и неиммунизированный контроль) по эффективности конверсии корма и приросту массы, а также более низким баллам поражения и смертности.
II. Влияние дозы RASV χ12341 (pYA5112).
[00237] Для оценки влияния дозирования RASV χ12341 (pYA5112), проводили следующий эксперимент с использованием низкого титра (5×107 CFU); промежуточного титра (1,5×108 CFU) или начального титра (описанного выше; 5×108 CFU) бактериального штамма RASV χ12341 (pYA5112).
Материалы и способы
A. Рацион, используемый в эксперименте
[00238] Составляли стартерный корм для цыплят без содержания лекарственных средств, смешанный с кормовыми продуктами, широко используемыми в Соединенных Штатах Америки. Корм был характерным для местного коммерческого состава, а рассчитанные анализы соответствовали или превосходили требования NRC к стартерным кормам для цыплят. Корма для экспериментальной обработки готовили на основе указанного базового стартерного корма. Количества всех базовых кормов и исследуемых препаратов, используемых для приготовления партий для обработки, фиксировали. Корма для обработки смешивали для обеспечения однородного распределения соответствующего исследуемого препарата. Мешалку промывали для предупреждения перекрестной контаминации. Корм распределяли по клеткам с одинаковой обработкой. Указанный рацион (в форме смеси) скармливали во время указанного исследования.
B. Животные
[00239] Однодневных петушков цыплят-бройлеров мужского пола получали от Cobb-Vantress, Кливленд, Джорджия. Порода представляла собой Cobb 500. Информацию о племенном стаде фиксировали. В инкубаторе у птиц определяли пол и они получали стандартные вакцинации. В исследовании использовали только цыплят, которые выглядели здоровыми. Размещение всех птиц, не используемых для распределения на группы, фиксировали. Бумаги или смывы дна всех боксов для птиц культивировали на наличие Salmonella.
Процедуры
а. Распределение птиц по группам и рандомизация клеток
[00240] Исследование начинали, когда птиц размещали (день выведения) (DOT 0), при этом их распределяли в экспериментальные клетки. Никаких птиц не замещали во время исследования.
b. Введение вакцин
[00241] Бактериальные штаммы вводили в DOT 0 каждому цыпленку в группах обработки 3, 4, 5 и 6 через желудочный зонд в дозе ~5×108 CFU/цыпленок в объеме 0,1 мл.
c. Взвешивания в клетках
[00242] Всех птиц взвешивали в DOT 0, 14, 21 и 28. Корм взвешивали в DOT 0, а оставшийся корм взвешивали в DOT 14, 21 и 28.
d. Индукция заболевания
[00243] В DOT 14 всех птиц перорально инокулировали ~5000 ооцист E. maxima. Начиная с DOT 19 всем птицам (за исключением группы обработки 1) вводили бульонную культуру С. perfringens в дозе ~108 CFU/мл. В данном исследовании корм не удаляли. С помощью желудочного зонда птицам вводили 0,1 мл свежей бульонной культуры один раз в день в течение 3 дней (в DOT 19, 20 и 21).
e. Рост после заражения Clostridium perfringens
[00244] Используемым штаммом для заражения был Clostridium perfringens №6 (как описано в Hofacre, et al., 1998) (222). Его вводили в один (1) литр тиогликолятного бульона с добавлением 5% говяжьего экстракта и инкубировали при 37°С в течение 15 часов.
f Количественное оценивание кишечных поражений вследствие некротического энтерита
[00245] В DOT 21 трех птиц из каждой клетки через четыре (4) часа после третьего заражения Clostridium perfringens отбирали, умерщвляли, взвешивали и исследовали на наличие поражений вследствие некротического энтерита. Количественное оценивание было основано на шкале от 0 до 3 баллов, при этом 0 соответствовал норме, а 3 соответствовал наиболее высокой тяжести. У всех из трех птиц для количественной оценки поражений отбирали кровь для хранения сыворотки крови.
g. Анализ данных и результаты
[00246] Выполняли статистический анализ прироста массы в клетке, потребления корма, конверсии корма, баллов поражения и смертности. Результаты эксперимента показаны ниже в Табл. 5. Как показано в Табл. 5, начальные и промежуточные дозы RASV превосходили низкую дозу или контроли.
III. Влияние способа иммунизации.
[00247] С целью определения влияния способа введения RASV χ12341 (pYA5112) цыплят иммунизировали либо высокой дозой (5×108 CFU) либо промежуточной дозой (1×108 CFU) бактериального штамма χ12341 (pYA5112) с помощью желудочного зонда, спрея или перорально (в питьевой воде). Вкратце в DOT 0 каждому цыпленку вводили перорально через желудочный зонд 0,1 мл бактериального штамма, ~5×108 CFU/цыпленок; в DOT 14 всех цыплят инокулировали перорально ~5000 ооцистами Е. maxima. Начиная с DOT 19 всем птицам (за исключением группы обработки 1) вводили бульонную культуру С. perfringens (CP) в дозе ~108 CFU/мл один раз в день в течение 3 дней (в DOT 19, 20 и 21). В DOT 21 трех птиц из каждой клетки через четыре (4) часа после третьего заражения Clostridium perfringens исследовали на наличие поражений вследствие некротического энтерита. Количественное оценивание было основано на шкале от 0 до 3 баллов, при этом 0 соответствовал норме, а 3 соответствовал наиболее высокой тяжести.
[00248] Как показано в Табл. 6, все способы иммунизации превосходили контрольную невакцинированную группу. Кроме того, группа с иммунизацией с использованием спрея характеризовалась удовлетворительными характеристиками по сравнению с группами с использованием желудочного зонда. Это является коммерчески важным, поскольку иммунизация с использованием спрея в инкубаторах является предпочтительным и наиболее экономичным средством иммунизации домашней птицы.
Пример 8: Неспособность высоких концентраций одного сахара нарушать способность двух других сахаров при низких концентрациях регулировать гены, необходимые для выживания штаммов RASV с фенотипом регулируемого замедленного лизиса in vivo
[00249] χ12341 (pYA4763) характеризовался облигатной потребностью в арабинозе для выживания, поскольку он имел регулируемые araC ParaBAD гены murA как в хромосоме, так и в плазмиде pYA4763. Поскольку продукт кодируемого гена murA, фермент, был фосфорилированным, и поскольку Salmonella не могла усваивать фосфорилированные сахара, штамм χ12341 (pYA4763) содержал зависимую от арабинозы летальную конструкцию. Таким образом, любой экзогенный сахар, который блокировал способность арабинозы усваиваться χ12341 (pYA4763) или приводить к транскрипции гена murA с помощью активации промотора ParaBAD, приводил к гибели клеток χ12341 (pYA4763). С целью определения того, нарушали ли высокие концентрации (т.е., 1,0%) любого из трех сахаров, используемых χ12341 (pYA4763), активности двух других сахаров при более низких концентрациях (т.е., 0,1% или ниже) с целью обеспечения выживания или дисплея регулируемого фенотипа, проводили следующие эксперименты. Эксперименты по изучению роста проводили с использованием бульона Purple (с целью предупреждения любого изменения значения рН вследствие утилизации сахаров).
[00250] Вкратце, рост штаммов Salmonella χ12341 (pYA4763) и χ3761 оценивали в течение 24-часового периода в среде для роста с варьирующими концентрациями сахаров. Вкратце, культуру каждого штамма, полученную в течение ночи, выращивали в буферном бульоне Purple + 0,05% арабиноза + 0,1% рамноза + 0,1% манноза. Субкультивирование выполняли с помощью разведения при 1:100 (Фиг. 6А, 6В, 6G, 6Н, 6М и 6N), 1:1000 (Фиг. 6С, 6D, 6I, 6J, 6О и 16Р) или 1:10000 (Фиг. 6Е, 6F, 6K, 6L, 6Q и 6R) в предварительно нагретом буферном бульоне Purple с варьирующими концентрациями арабинозы, рамнозы и маннозы. 200 мкл каждой культуры добавляли в отдельную лунку в 100-луночном планшете в двух повторностях для каждого штамма и условия использования сахара. Планшет помещали в автоматизированную микробиологическую систему анализа кривых роста Bioscreen С при 37°С и оставляли инкубироваться, при встряхивании, на 24 часа. Оптические плотности измеряли через каждые 30 минут и сравнивали с холостым образцом для подтверждения чистоты. На фигурах представлены данные при использовании всех условий (Фиг. 6A-6F), сравнение условий, при которых использовали 1% одного из трех сахаров (Фиг. 6G-6L) и сравнение различных концентраций арабинозы (Фиг. 6M-6R).
[00251] Как показано на Фиг. 6A-6R, наблюдали рост бактериального штамма χ12341 (pYA4763), а также штамма дикого типа S. Typhimurium UK-1 независимо от присутствия какого-либо одного сахара при концентрации 1,0% и двух других сахаров при концентрациях 0,1% или ниже. Необходимо отметить, что бульон Purple был лишен всех сахаров, в результате чего отсутствовали помехи в результатах вследствие следовых количеств арабинозы, маннозы и рамнозы. Эти результаты продемонстрировали, что высокие концентрации маннозы или рамнозы не подавляли способности низких концентраций арабинозы вызывать экспрессию гена murA, поскольку клеточной смерти не наблюдалось.
[00252] Вестерн-блоттинг могли проводить с целью анализа экспрессии генов, кодирующих продукты, регулируемые одним из регулируемых сахаром промоторов в штамме χ12341 (pYA4763).
Источники литературы
1. Lozano R, Naghavi M, Foreman K, Lim S, Shibuya K, Aboyans V, Abraham J, Adair T, Aggarwal R, Ahn SY, Alvarado M, Anderson HR, Anderson LM, Andrews KG, Atkinson C, Baddour LM, Barker-Collo S, Bartels DH, Bell ML, Benjamin EJ, Bennett D, Bhalla K, Bikbov B, Bin Abdulhak A, Birbeck G, Blyth F, Bolliger I, Boufous S, Bucello C, Burch M, Burney P, Carapetis J, Chen H, Chou D, Chugh SS, Coffeng LE, Colan SD, Colquhoun S, Colson KE, Condon J, Connor MD, Cooper LT, Corriere M, Cortinovis M, de Vaccaro KC, Couser W, Cowie ВС, Criqui MH, Cross M, Dabhadkar KC, et al. 2012. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet 380:2095-2128.
2. Tennant SM, Levine MM. 2015. Live attenuated vaccines for invasive Salmonella infections. Vaccine 33 Suppl 3:C36-C41.
3. Wain J, Hendriksen RS, Mikoleit ML, Keddy KH, Ochiai RL. 2015. Typhoid fever. Lancet 385:1136-1145.
4. Sahastrabuddhe S, Carbis R, Wierzba TF, Ochiai RL. 2013. Increasing rates of Salmonella Paratyphi A and the current status of its vaccine development. Expert Rev Vaccines 12:1021-1031.
5. Mogasale V, Maskery B, Ochiai RL, Lee JS, Mogasale VV, Ramani E, Kim YE, Park JK, Wierzba TF. 2014. Burden of typhoid fever in low-income and middle-income countries: a systematic, literature-based update with risk-factor adjustment. Lancet Glob Health 2:e570-580.
6. Crump JA, Luby SP, Mintz ED. 2004. The global burden of typhoid fever. Bull World Health Organ 82:346-353.
7. Murray CJ, Vos T, Lozano R, Naghavi M, Flaxman AD, Michaud C, Ezzati M, Shibuya K, Salomon JA, Abdalla S, Aboyans V, Abraham J, Ackerman I, Aggarwal R, Ahn SY, Ali MK, Alvarado M, Anderson HR, Anderson LM, Andrews KG, Atkinson C, Baddour LM, Bahalim AN, Barker-Collo S, Barrero LH, Bartels DH, Basanez MG, Baxter A, Bell ML, Benjamin EJ, Bennett D, Bernabe E, Bhalla K, Bhandari B, Bikbov B, Bin Abdulhak A, Birbeck G, Black JA, Blencowe H, Blore JD, Blyth F, Bolliger I, Bonaventure A, Boufous S, Bourne R, Boussinesq M, Braithwaite T, Brayne C, Bridgett L, Brooker S, et al. 2012. Disability-adjusted life years (DALYs) for 291 diseases and injuries in 21 regions, 1990-2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet 380:2197-2223.
8. Feasey NA, Dougan G, Kingsley RA, Heyderman RS, Gordon MA. 2012. Invasive non-typhoidal Salmonella disease: an emerging and neglected tropical disease in Africa. Lancet 379:2489-2499.
9. Chiu CH, Su LH, Chu C. 2004. Salmonella enterica serotype Choleraesuis: epidemiology, pathogenesis, clinical disease, and treatment. Clin Microbiol Rev 17:311-322.
10. Ao TT, Feasey NA, Gordon MA, Keddy KH, Angulo FJ, Crump JA. 2015. Global burden of invasive nontyphoidal Salmonella disease, 2010(1). Emerg Infect Dis 21.
11 Majowicz SE, Musto J, Scallan E, Angulo FJ, Kirk M, O'Brien SJ, Jones TF, Fazil A, Hoekstra RM, International Collaboration on Enteric Disease 'Burden of Illness S. 2010. The global burden of nontyphoidal Salmonella gastroenteritis. Clin Infect Dis 50:882-889.
12. Scallan E, Hoekstra RM, Mahon BE, Jones TF, Griffin PM. 2015. An assessment of the human health impact of seven leading foodborne pathogens in the United States using disability adjusted life years. Epidemiol Infect 143:2795-2804.
13. Scallan E, Hoekstra RM, Angulo FJ, Tauxe RV, Widdowson MA, Roy SL, Jones JL, Griffin PM. 2011. Foodborne illness acquired in the United States--major pathogens. Emerg Infect Dis 17:7-15.
14. Hoffmann S, Batz MB, Morris JG, Jr. 2012. Annual cost of illness and quality-adjusted life year losses in the United States due to 14 foodborne pathogens. J Food Prot 75:1292-1302.
15. Scallan E, Mahon BE, Hoekstra RM, Griffin PM. 2013. Estimates of illnesses, hospitalizations and deaths caused by major bacterial enteric pathogens in young children in the United States. Pediatr Infect Dis J 32:217-221.
16. Olsen SJ, Bishop R, Brenner FW, Roels TH, Bean N, Tauxe RV, Slutsker L. 2001. The changing epidemiology of Salmonella: trends in serotypes isolated from humans in the United States, 1987-1997. J Infect Dis 183:753-761.
17. Jackson BR, Griffin PM, Cole D, Walsh KA, Chai SJ. 2013. Outbreak-associated Salmonella enterica serotypes and food Commodities, United States, 1998-2008. Emerg Infect Dis 19:1239-1244.
18. Gordon MA. 2008. Salmonella infections in immunocompromised adults. J Infect 56:413-422.
19. Preziosi MJ, Kandel SM, Guiney DG, Browne SH. 2012. Microbiological analysis of nontyphoidal Salmonella strains causing distinct syndromes of bacteremia or enteritis in HIV/AIDS patients in San Diego, California. J Clin Microbiol 50:3598-3603.
20. MacLennan CA, Levine MM. 2013. Invasive nontyphoidal Salmonella disease in Africa: current status. Expert Rev Anti Infect Ther 11:443-446.
21. Graham SM, Molyneux EM, Walsh AL, Cheesbrough JS, Molyneux ME, Hart CA. 2000. Nontyphoidal Salmonella infections of children in tropical Africa. Pediatr Infect Dis J 19:1189-1196.
22. Crump JA, Medalla FM, Joyce KW, Krueger AL, Hoekstra RM, Whichard JM, Barzilay EJ, Emerging Infections Program NWG. 2011. Antimicrobial resistance among invasive nontyphoidal Salmonella enterica isolates in the United States: National Antimicrobial Resistance Monitoring System, 1996 to 2007. Antimicrob Agents Chemother 55:1148-1154.
23. Bhutta ZA, Threlfall J. 2009. Addressing the global disease burden of typhoid fever. JAMA 302:898-899.
24. Jean SS, Hsueh PR. 2011. High burden of antimicrobial resistance in Asia, hit J Antimicrob Agents 37:291-295.
25. Crump JA, Mintz ED. 2010. Global trends in typhoid and paratyphoid Fever. Clin Infect Dis 50:241-246.
26. Martin LB. 2012. Vaccines for typhoid fever and other salmonelloses. Curr Opin Infect Dis 25:489-499.
27. MacLennan CA, Martin LB, Micoli F. 2014. Vaccines against invasive Salmonella disease: current status and future directions. Hum Vaccin Immunother 10:1478-1493.
28. Engels EA, Falagas ME, Lau J, Bennish ML. 1998. Typhoid fever vaccines: a meta-analysis of studies on efficacy and toxicity. BMJ 316:110-116.
29. Anwar E, Goldberg E, Fraser A, Acosta CJ, Paul M, Leibovici L. 2014. Vaccines for preventing typhoid fever. Cochrane Database Syst Rev 1:CD001261.
30. Germanier R, Fuer E. 1975. Isolation and characterization of Gal E mutant Ту 21a of Salmonella typhi: a candidate strain for a live, oral typhoid vaccine. J Infect Dis 131:553-558.
31. Tacket CO, Ferreccio C, Robbins JB, Tsai CM, Schulz D, Cadoz M, Goudeau A, Levine MM. 1986. Safety and immunogenicity of two Salmonella typhi Vi capsular polysaccharide vaccines. J Infect Dis 154:342-345.
32. Desin TS, Koster W, Potter AA. 2013. Salmonella vaccines in poultry: past, present and future. Expert Rev Vaccines 12:87-96.
33. Gal-Mor O, Boyle EC, Grassl GA. 2014. Same species, different diseases: how and why typhoidal and non-typhoidal Salmonella enterica serovars differ. Front Microbiol 5:391.
34. Bhavsar AP, Zhao X, Brown ED. 2001. Development and characterization of a xylose-dependent system for expression of cloned genes in Bacillus subtilis: conditional complementation of a teichoic acid mutant. Appl Environ Microbiol 67:403-410.
35. Walder RY, Walder JA. 1986. Oligodeoxynucleotide-directed mutagenesis using the yeast transformation system. Gene 42:133-139.
36. Bauer CE, Hesse SD, Waechter-Brulla DA, Lynn SP, Gumport RI, Gardner JF. 1985. A genetic enrichment for mutations constructed by oligodeoxynucleotide-directed mutagenesis. Gene 37:73-81.
37. Craik CS. 1985. Use Of Oligonucleotides For Site-specific Mutagenesis BIOTECHNIQUES 3:12-19.
38. Smith M, Gillam S. 1981. Constructed Mutants Using Synthetic Oligodeoxyribonucleotides as Site-Specific Mutagens, p 1-32. In Setlow JK, Hollaender A (ed), Genetic Engineering: Principles and Methods Volume 3 doi:10.1007/978-1-4615-7075-2 1. Springer US, Boston, MA.
39. Giacalone MJ, Gentile AM, Lovitt ВТ, Berkley NL, Gunderson CW, Surber MW. 2006. Toxic protein expression in Escherichia coli using a rhamnose-based tightly regulated and tunable promoter system. Biotechniques 40:355-364.
40. Elhenawy W, Bording-Jorgensen M, Valguarnera E, Haurat MF, Wine E, Feldman MF. 2016. LPS Remodeling Triggers Formation of Outer Membrane Vesicles in Salmonella. MBio 7:e00940-00916.
41. Man SM, Hopkins LJ, Nugent E, Cox S, Gluck IM, Tourlomousis P, Wright JA, Cicuta P, Monie TP, Bryant CE. 2014. Inflammasome activation causes dual recruitment of NLRC4 and NLRP3 to the same macromolecular complex. Proc Natl Acad Sci U S A 111:7403-7408.
42. Collins LV, Attridge S, Hackett J. 1991. Mutations at rfc or pmi attenuate Salmonella typhimurium virulence for mice. Infect Immun 59:1079-1085.
43. Curtiss R, III., Zhang X, Wanda SY, Kang HY, Konjufca V, Li Y, Gunn B, Wang S, Scarpellini G, Lee IS. 2007. Induction of host immune responses using Salmonella-vectored vaccines, p 297-313. In Brogden KA, Minion FC, Cornick N, Stanton ТВ, Zhang Q, Nolan LK, Wannemuehler MJ (ed), Virulence mechanisms of bacterial pathogens, 4th ed. ASM Press, Washington D.C.
44. Raetz CR, Whitfield C. 2002. Lipopolysaccharide endotoxins. Annu Rev Biochem 71:635-700.
45. Frey PA. 1996. The Leloir pathway: a mechanistic imperative for three enzymes to change the stereochemical configuration of a single carbon in galactose. FASEB J 10:461-470.
46. Leloir LF. 1951. The enzymatic transformation of uridine diphosphate glucose into a galactose derivative. Arch Biochem Biophys 33:186-190.
47. Germanier R, Furer E. 1971. Immunity in experimental salmonellosis. II. Basis for the avirulence and protective capacity of galE mutants of Salmonella typhimurium. Infect Immun 4:663-673.
48. Fukasawa T, Nikaido H. 1959. Galactose-sensitive mutants of Salmonella. Nature 184(Suppl 15): 1168-1169.
49. Lee SJ, Trostel A, Le P, Harinarayanan R, Fitzgerald PC, Adhya S. 2009 Cellular stress created by intermediary metabolite imbalances. Proc Natl Acad Sci U S A 106:19515-19520.
50. Hone D, Morona R, Attridge S, Hackett J. 1987. Construction of defined galE mutants of Salmonella for use as vaccines. J Infect Dis 156:167-174.
51. Cascales E, Buchanan SK, Duche D, Kleanthous C, Lloubes R, Postle K, Riley M, Slatin S, Cavard D. 2007. Colicin biology. Microbiol Mol Biol Rev 71:158-229.
52. Mastroeni P, Simmons C, Fowler R, Hormaeche CE, Dougan G. 2000. Igh-6-/- (B-cell-deficient) mice fail to mount solid acquired resistance to oral challenge with virulent Salmonella enterica serovar typhimurium and show impaired Th1 T-cell responses to Salmonella antigens. Infect Immun 68:46-53.
53. Ugrinovic S, Menager N, Goh N, Mastroeni P. 2003. Characterization and development of T-Cell immune responses in B-cell-deficient (Igh-6-/-) mice with Salmonella enterica serovar Typhimurium infection. Infect Immun 71:6808-6819.
54. Mittrucker HW, Raupach B, Kohler A, Kaufmann SH. 2000. Cutting edge: role of В lymphocytes in protective immunity against Salmonella typhimurium infection. J Immunol 164:1648-1652.
55. Mastroeni P, Menager N. 2003. Development of acquired immunity to Salmonella. J Med Microbiol 52:453-459.
56. Pham OH, McSorley SJ. 2015. Protective host immune responses to Salmonella infection. Future Microbiol 10:101-110.
57. Cunningham AF, Gaspal F, Serre K, Mohr E, Henderson IR, Scott-Tucker A, Kenny SM, Khan M, Toellner KM, Lane PJ, MacLennan IC. 2007. Salmonella induces a switched antibody response without germinal centers that impedes the extracellular spread of infection. J Immunol 178:6200-6207.
58. MacLennan CA, Gondwe EN, Msefula CL, Kingsley RA, Thomson NR, White SA, Goodall M, Pickard DJ, Graham SM, Dougan G, Hart CA, Molyneux ME, Drayson MT. 2008. The neglected role of antibody in protection against bacteremia caused by nontyphoidal strains of Salmonella in African children. J Clin Invest 118:1553-1562.
59. Guzman CA, Borsutzky S, Griot-Wenk M, Metcalfe IC, Pearman J, Collioud A, Favre D, Dietrich G. 2006. Vaccines against typhoid fever. Vaccine 24:3804-3811.
60. MacLennan CA. 2014. Antibodies and protection against invasive Salmonella disease. Front Immunol 5:635.
61. Isibasi A, Ortiz V, Vargas M, Paniagua J, Gonzalez C, Moreno J, Kumate J. 1988. Protection against Salmonella typhi infection in mice after immunization with outer membrane proteins isolated from Salmonella typhi 9,12,d, Vi. Infect Immun 56:2953-2959.
62. Klugman KP, Gilbertson IT, Koornhof HJ, Robbins JB, Schneerson R, Schulz D, Cadoz M, Armand J. 1987. Protective activity of Vi capsular polysaccharide vaccine against typhoid fever. Lancet 2:1165-1169.
63. Acharya IL, Lowe CTJ, Thapa R, Gurubacharya VL, Shrestha MB, Cadoz M, Schulz D, Armand J, Bryla DA, Trollfors B, et al. 1987. Prevention of typhoid fever in Nepal with the Vi capsular polysaccharide of Salmonella typhi. A preliminary report. N Engl J Med 317:1101-1104.
64. Szu SC. 2013. Development of Vi conjugate - a new generation of typhoid vaccine. Expert Rev Vaccines 12:1273-1286.
65. Jansson PE, Lindberg AA, Lindberg B, Wollin R. 1981. Structural studies on the hexose region of the core in lipopolysaccharides from Enterobacteriaceae. Eur J Biochem 115:571-577.
66.  O, Westphal O, Staub AM, Nikaido H. 1971. Isolation and Chemical and Immunological Characterization of Bacterial Lipopolysaccharides, p 145-223. In Weinbaum G, Kadis S, Ajl SJ (ed), Bacterial Endotoxins, vol 4. in Microbial Toxins. Academic Press, Inc, New York.
O, Westphal O, Staub AM, Nikaido H. 1971. Isolation and Chemical and Immunological Characterization of Bacterial Lipopolysaccharides, p 145-223. In Weinbaum G, Kadis S, Ajl SJ (ed), Bacterial Endotoxins, vol 4. in Microbial Toxins. Academic Press, Inc, New York.
67. Kaniuk NA, Monteiro MA, Parker CT, Whitfield C. 2002. Molecular diversity of the genetic loci responsible for lipopolysaccharide core oligosaccharide assembly within the genus Salmonella. Mol Microbiol 46:1305-1318.
68. Olsthoorn MM, Petersen BO, Schlecht S, Haverkamp J, Bock K, Thomas-Oates JE, Holst O. 1998. Identification of a novel core type in Salmonella lipopolysaccharide. Complete structural analysis of the core region of the lipopolysaccharide from Salmonella enterica sv. Arizonae O62. J Biol Chem 273:3817-3829.
69. Malik M, Butchaiah G, Bansal MP, Siddiqui MZ, Bakshi CS, Singh RK. 2002 Antigenic relationships within the genus Salmonella as revealed by anti- Salmonella enteritidis monoclonal antibodies. Vet Res Commun 26:179-188.
70. Earhart CF. 1996. Uptake and metabolism of iron and molybdenum., p 1075-1090. In Neidhardt FC, Curtiss III R, Ingraham JL, Lin ECC, Low KB, Magasanik B, Reznikoff WS, Riley M, Schaechter M, Umbarger HE (ed), Escherichia coli and Salmonella Cellular and Molecular Biology, 2nd ed, vol 1. ASM Press, Washington, D.C.
71. Collins LV, Attridge S, Hackett J. 1991. Mutations at rfc or pmi attenuate Salmonella typhimurium virulence for mice. Infect Immun 59:1079-1085.
72. Rosen SM, Zeleznick LD, Fraenkel D, Wiener IM, Osborn MJ, Horecker BL. 1965. Characterization of the cell wall lipopolysaccharide of a mutant of Salmonella typhimurium lacking phosphomannose isomerase. Biochem Z 342:375-386.
73.  PH, Stocker BAD. 1969. Genetics of Polysaccharide Biosynthesis. Annual Review of Genetics 3:291-322.
PH, Stocker BAD. 1969. Genetics of Polysaccharide Biosynthesis. Annual Review of Genetics 3:291-322.
74. Stocker BAD, PH. 1971. Genetic Aspects of Biosynthesis and Structure of Salmonella Lipopolysaccharide, p 369-438. In Weinbaum G, Kadis S, Ajl SJ (ed), Bacterial Endotoxins, vol 4. in Microbial Toxins. Academic Press, Inc, New York.
75. Curtiss R, III, Wanda SY, Gunn BM, Zhang X, Tinge SA, Ananthnarayan V, Mo H, Wang S, Kong W. 2009. Salmonella enterica serovar Typhimurium strains with regulated delayed attenuation in vivo. Infect Immun 77:1071-1082.
76. Curtiss R, III, Zhang X, Wanda SY, Kang HY, Konjufca V, Li H, Gunn B, Wang S, Scarpellini G, S. LI. 2007. Induction of host immune responses using Salmonella-vectored vaccines, p 297-313. In Brogden KA, Minion FC, Stanton ТВ, Zhang Q, Nolan LK, Wannemuehler MJ (ed), Virulence Mechanisms of Bacterial Pathogens, 4th ed. ASM Press, Washington D.C.
77. Chamnongpol S, Dodson W, Cromie MJ, Harris ZL, Groisman EA. 2002. Fe(III)-mediated cellular toxicity. Mol Microbiol 45:711-719.
78. Nnalue NA. 1999. All accessible epitopes in the Salmonella lipopolysaccharide core are associated with branch residues. Infect Immun 67:998-1003.
79. Stanislavsky ES, Makarenko ТА, Kholodkova EV, Lugowski C. 1997. R-form lipopolysaccharides (LPS) of Gram-negative bacteria as possible vaccine antigens. FEMS Immunol Med Microbiol 18:139-145.
80. Muralinath M, Kuehn MJ, Roland KL, Curtiss R, III. 2011. Immunization with Salmonella enterica serovar Typhimurium-derived outer membrane vesicles delivering the pneumococcal protein PspA confers protection against challenge with Streptococcus pneumoniae. Infect Immun 79:887-894.
81. Englesberg E, Irr J, Power J, Lee N. 1965. Positive control of enzyme synthesis by gene С in the L-arabinose system. J Bacteriol 90:946-957.
82. Guzman LM, Belin D, Carson MJ, Beckwith J. 1995. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J Bacteriol 177:4121-4130.
83. Bolin CA, Jensen AE. 1987. Passive immunization with antibodies against iron-regulated outer membrane proteins protects turkeys from Escherichia coli septicemia. Infect Immun 55:1239-1242.
84. Lin J, Hogan JS, Smith KL. 1999. Antigenic homology of the inducible ferric citrate receptor (FecA) of coliform bacteria isolated from herds with naturally occurring bovine intramammary infections. Clin Diagn Lab Immunol 6:966-969.
85. Clifton-Hadley FA, Breslin M, Venables LM, Sprigings KA, Cooles SW, Houghton S, Woodward MJ. 2002. A laboratory study of an inactivated bivalent iron restricted Salmonella enterica serovars Enteritidis and Typhimurium dual vaccine against Typhimurium challenge in chickens. Vet Microbiol 89:167-179.
86. Woodward MJ, Gettinby G, Breslin MF, Corkish JD, Houghton S. 2002. The efficacy of Salenvac, a Salmonella enterica subsp. Enterica serotype Enteritidis iron-restricted bacterin vaccine, in laying chickens. Avian Pathol 31:383-392.
87. Berlanda Scorza F, Colucci AM, Maggiore L, Sanzone S, Rossi O, Ferlenghi I, Pesce I, Caboni M, Norais N, Di Cioccio V, Saul A, Gerke C. 2012. High yield production process for Shigella outer membrane particles. PLoS One 7:e35616.
88. Clementz T, Bednarski JJ, Raetz CR. 1996. Function of the htrB high temperature requirement gene of Escherichia coli in the acylation of lipid A: HtrB catalyzed incorporation of laurate. J Biol Chem 271:12095-12102.
89. Kulp A, Kuehn MJ. 2010. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu Rev Microbiol 64:163-184.
90. Dowling JK, Mansell A. 2016. Toll-like receptors: the swiss army knife of immunity and vaccine development. Clin Transl Immunology 5:e85.
91. Duthie MS, Windish HP, Fox CB, Reed SG. 2011. Use of defined TLR ligands as adjuvants within human vaccines. Immunol Rev 239:178-196.
92. Steinhagen F, Kinjo T, Bode C, Klinman DM. 2011. TLR-based immune adjuvants. Vaccine 29:3341-3355.
93. Lahiri A, Das P, Chakravortty D. 2008. Engagement of TLR signaling as adjuvant: towards smarter vaccine and beyond. Vaccine 26:6777-6783.
94. Ishii KJ, Akira S. 2007. Toll or toll-free adjuvant path toward the optimal vaccine development. J Clin Immunol 27:363-371.
95. Toussi DN, Massari P. 2014. Immune Adjuvant Effect of Molecularly-defined Toll-Like Receptor Ligands. Vaccines (Basel) 2:323-353.
96. Koeberling O, Delany I, Granoff DM. 2011. A critical threshold of meningococcal factor H binding protein expression is required for increased breadth of protective antibodies elicited by native outer membrane vesicle vaccines. Clin Vaccine Immunol 18:736-742.
97. Koeberling O, Seubert A, Granoff DM. 2008. Bactericidal antibody responses elicited by a meningococcal outer membrane vesicle vaccine with overexpressed factor H-binding protein and genetically attenuated endotoxin. J Infect Dis 198:262-270.
98. Pajon R, Fergus AM, Koeberling O, Caugant DA, Granoff DM. 2011. Meningococcal factor H binding proteins in epidemic strains from Africa: implications for vaccine development. PLoS Negl Trop Dis 5:e1302.
99. Zollinger WD, Babcock JG, Moran EE, Brandt BL, Matyas GR, Wassef NM, Alving CR. 2012. Phase I study of a Neisseria meningitidis liposomal vaccine containing purified outer membrane proteins and detoxified lipooligosaccharide. Vaccine 30:712-721.
100. Bernadac A, Gavioli M, Lazzaroni JC, Raina S, Lloubes R. 1998. Escherichia coli tol-pal mutants form outer membrane vesicles. J Bacteriol 180:4872-4878.
101. Henry T, Pommier S, Journet L, Bernadac A, Gorvel JP, Lloubes R. 2004. Improved methods for producing outer membrane vesicles in Gram-negative bacteria. Res Microbiol 155:437-446.
102. Berlanda Scorza F, Doro F, Rodriguez-Ortega MJ, Stella M, Liberatori S, Taddei AR, Serino L, Gomes Moriel D, Nesta B, Fontana MR, Spagnuolo A, Pizza M, Norais N, Grandi G. 2008. Proteomics characterization of outer membrane vesicles from the extraintestinal pathogenic Escherichia coli DtolR IHE3034 mutant. Mol Cell Proteomics 7:473-485.
103. Clementz T, Zhou Z, Raetz CR. 1997. Function of the Escherichia coli msbB gene, a multicopy suppressor of htrB knockouts, in the acylation of lipid A. Acylation by MsbB follows laurate incorporation by HtrB. J Biol Chem 272:10353-10360.
104. Meloni E, Colucci AM, Micoli F, Sollai L, Gavini M, Saul A, Di Cioccio V, MacLennan CA. 2015. Simplified low-cost production of O-antigen from Salmonella Typhimurium Generalized Modules for Membrane Antigens (GMMA). J Biotechnol 198:46-52.
105. Gerke C, Colucci AM, Giannelli C, Sanzone S, Vitali CG, Sollai L, Rossi O, Martin LB, Auerbach J, Di Cioccio V, Saul A. 2015. Production of a Shigella sonnei Vaccine Based on Generalized Modules for Membrane Antigens (GMMA), 1790GAHB. PLoS One 10:e0134478.
106. Barat S, Willer Y, Rizos K, Claudi B, Maze A, Schemmer AK, Kirchhoff D, Schmidt A, Burton N, Bumann D. 2012. Immunity to intracellular Salmonella depends on surface-associated antigens. PLoS Pathog 8:e1002966.
107. Kurtz JR, Petersen HE, Frederick DR, Morici LA, McLachlan JB. 2014. Vaccination with a single CD4 T cell peptide epitope from a Salmonella type III-secreted effector protein provides protection against lethal infection. Infect Immun 82:2424-2433.
108. Rollenhagen C, Sorensen M, Rizos K, Hurvitz R, Bumann D. 2004. Antigen selection based on expression levels during infection facilitates vaccine development for an intracellular pathogen. Proc Natl Acad Sci U S A 101:8739-8744.
109. Lee SJ, McLachlan JB, Kurtz JR, Fan D, Winter SE, Baumler AJ, Jenkins MK, McSorley SJ. 2012. Temporal expression of bacterial proteins instructs host CD4 T cell expansion and Th17 development. PLoS Pathog 8:e1002499.
110. McSorley SJ, Cookson ВТ, Jenkins MK. 2000. Characterization of CD4+ T cell responses during natural infection with Salmonella typhimurium. J Immunol 164:986-993.
111. Gil-Cruz C, Bobat S, Marshall JL, Kingsley RA, Ross EA, Henderson IR, Leyton DL, Coughlan RE, Khan M, Jensen KT, Buckley CD, Dougan G, MacLennan IC, Lopez-Macias C, Cunningham AF. 2009. The porin OmpD from nontyphoidal Salmonella is a key target for a protective B1b cell antibody response. Proc Natl Acad Sci U S A 106:9803-9808.
112. Yang Y, Wan C, Xu H, Aguilar ZP, Tan Q, Xu F, Lai W, Xiong Y, Wei H. 2013. Identification of an outer membrane protein of Salmonella enterica serovar Typhimurium as a potential vaccine candidate for Salmonellosis in mice. Microbes Infect 15:388-398.
113. Goh YS, Armour KL, Clark MR, Grant AJ, Mastroeni P. 2016. Igg Subclasses Targeting the Flagella of Salmonella enterica Serovar Typhimurium Can Mediate Phagocytosis and Bacterial Killing. J Vaccines Vaccin 7.
114. Cummings LA, Wilkerson WD, Bergsbaken T, Cookson ВТ. 2006. In vivo, fliC expression by Salmonella enterica serovar Typhimurium is heterogeneous, regulated by ClpX, and anatomically restricted. Mol Microbiol 61:795-809.
115. Winter SE, Winter MG, Godinez I, Yang HJ, Russmann H, Andrews-Polymenis HL, Baumler AJ. 2010. A rapid change in virulence gene expression during the transition from the intestinal lumen into tissue promotes systemic dissemination of Salmonella. PLoS Pathog 6:e1001060.
116. Cummings LA, Barrett SL, Wilkerson WD, Fellnerova I, Cookson ВТ. 2005. FliC-specific CD4+ T cell responses are restricted by bacterial regulation of antigen expression. J Immunol 174:7929-7938.
117. Simon R, Tennant SM, Wang JY, Schmidlein PJ, Lees A, Ernst RK, Pasetti MF, Galen JE, Levine MM. 2011. Salmonella enterica serovar enteritidis core О polysaccharide conjugated to H:g,m flagellin as a candidate vaccine for protection against invasive infection with S. Enteritidis. Infect Immun 79:4240-4249.
118. Simon R, Wang JY, Boyd MA, Tulapurkar ME, Ramachandran G, Tennant SM, Pasetti M, Galen JE, Levine MM. 2013. Sustained protection in mice immunized with fractional doses of Salmonella Enteritidis core and О polysaccharide-flagellin glycoconjugates. PLoS One 8:e64680.
119. Singh SP, Williams YU, Benjamin WH, Klebba PE, Boyd D. 1996 Immunoprotection by monoclonal antibodies to the porins and lipopolysaccharide of Salmonella typhimurium. Microb Pathog 21:249-263.
120. Lee SJ, Liang L, Juarez S, Nanton MR, Gondwe EN, Msefula CL, Kayala MA, Necchi F, Heath JN, Hart P, Tsolis RM, Heyderman RS, MacLennan CA, Felgner PL, Davies DH, McSorley SJ. 2012. Identification of a common immune signature in murine and human systemic Salmonellosis. Proc Natl Acad Sci USA 109:4998-5003.
121. Secundino I, Lopez-Macias C, Cervantes-Barragan L, Gil-Cruz C, Rios-Sarabia N, Pastelin-Palacios R, Villasis-Keever MA, Becker I, Puente JL, Calva E, Isibasi A. 2006. Salmonella porins induce a sustained, lifelong specific bactericidal antibody memory response. Immunology 117:59-70.
122. Salazar-Gonzalez RM, Maldonado-Bernal C, Ramirez-Cruz NE, Rios-Sarabia N, Beltran-Nava J, Castanon-Gonzalez J, Castillo-Torres N, Palma-Aguirre JA, Carrera-Camargo M, Lopez-Macias C, Isibasi A. 2004. Induction of cellular immune response and anti-Salmonella enterica serovar Typhi bactericidal antibodies in healthy volunteers by immunization with a vaccine candidate against typhoid fever. Immunol Lett 93:115-122.
123. Santiviago CA, Toro CS, Bucarey SA, Mora GC. 2001. A chromosomal region surrounding the ompD porin gene marks a genetic difference between Salmonella typhi and the majority of Salmonella serovars. Microbiology 147:1897-1907.
124. Santiviago CA, Fuentes JA, Bueno SM, Trombert AN, Hildago AA, Socias LT, Youderian P, Mora GC. 2002. The Salmonella enterica sv. Typhimurium smvA, yddG and ompD (porin) genes are required for the efficient efflux of methyl viologen. Mol Microbiol 46:687-698.
125. Chakraborty S, Mizusaki H, Kenney LJ. 2015. A FRET-based DNA biosensor tracks OmpR-dependent acidification of Salmonella during macrophage infection. PLoS Biol 13:e1002116.
126. Reynolds CJ, Jones C, Blohmke CJ, Darton TC, Goudet A, Sergeant R, Maillere B, Pollard AJ, Altmann DM, Boyton RJ. 2014. The serodominant secreted effector protein of Salmonella, SseB, is a strong CD4 antigen containing an immunodominant epitope presented by diverse HLA class II alleles. Immunology 143:438-446.
127. McLaughlin LM, Govoni GR, Gerke C, Gopinath S, Peng K, Laidlaw G, Chien YH, Jeong HW, Li Z, Brown MD, Sacks DB, Monack D. 2009. The Salmonella SPI2 effector SseI mediates long-term systemic infection by modulating host cell migration. PLoS Pathog 5:e1000671.
128. Worley MJ, Nieman GS, Geddes K, Heffron F. 2006. Salmonella typhimurium disseminates within its host by manipulating the motility of infected cells. Proc Natl Acad Sci U S A 103:17915-17920.
129. McLaughlin LM, Xu H, Carden SE, Fisher S, Reyes M, Heilshorn SC, Monack DM. 2014. A microfluidic-based genetic screen to identify microbial virulence factors that inhibit dendritic cell migration. Integr Biol (Camb) 6:438-449.
130. Lawley TD, Chan K, Thompson LJ, Kim CC, Govoni GR, Monack DM. 2006 Genome-wide screen for Salmonella genes required for long-term systemic infection of the mouse. PLoS Pathog 2:e11.
131. Thornbrough JM, Worley MJ. 2012. A naturally occurring single nucleotide polymorphism in the Salmonella SPI-2 type III effector srfH/sseI controls early extraintestinal dissemination. PLoS One 7:e45245.
132. Toobak H, Rasooli I, Talei D, Jahangiri A, Owlia P, Darvish Alipour Astaneh S. 2013. Immune response variations to Salmonella enterica serovar Typhi recombinant porin proteins in mice. Biologicals 41:224-230.
133. Pascual DW, Suo Z, Cao L, Avci R, Yang X. 2013. Attenuating gene expression (AGE) for vaccine development. Virulence 4:384-390.
134. Yang X, Suo Z, Thornburg T, Holderness K, Cao L, Lim T, Walters N, Kellerman L, Loetterle L, Avci R, Pascual DW. 2012. Expression of Escherichia coli virulence usher protein attenuates wild-type Salmonella. Virulence 3:29-42.
135. Yang X, Thornburg T, Suo Z, Jun S, Robison A, Li J, Lim T, Cao L, Hoyt T, Avci R, Pascual DW. 2012. Flagella overexpression attenuates Salmonella pathogenesis. PLoS One 7:e46828.
136. Kong Q, Liu Q, Roland KL, Curtiss R, III. 2009. Regulated delayed expression of rfaH in an attenuated Salmonella enterica serovar Typhimurium vaccine enhances immunogenicity of outer membrane proteins and a heterologous antigen. Infect Immun 77:5572-5582.
137. Curtiss R, III., Munson M. 1998. Cross-protective Salmonella vaccines.
138. Kelly SM, Bosecker BA, Curtiss R, III. 1992. Characterization and protective properties of attenuated mutants of Salmonella choleraesuis. Infect Immun 60:4881-4890.
139. Hassan JO, Curtiss R, III. 1994. Development and evaluation of an experimental vaccination program using a live avirulent Salmonella typhimurium strain to protect immunized chickens against challenge with homologous and heterologous Salmonella serotypes. Infect Immun 62:5519-5527.
140. Richardson EJ, Limaye B, Inamdar H, Datta A, Manjari KS, Pullinger GD, Thomson NR, Joshi RR, Watson M, Stevens MP. 2011. Genome sequences of Salmonella enterica serovar Typhimurium, Choleraesuis, Dublin, and Gallinarum strains of well- defined virulence in food-producing animals. J Bacteriol 193:3162-3163.
141. Kong Q, Liu Q, Jansen A, Curtiss R, III. 2010. Regulated delayed expression of rfc enhances the immunogenicity and protective efficacy of a heterologous antigen delivered by live attenuated Salmonella enterica vaccines. Vaccine 28:6094-6103.
142. Bertani G. 1951. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J Bacteriol 62:293-300.
143. Sambrook J, Russell DW. 2001. Molecular cloning: a laboratory manual, 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
144. Edwards RA, Keller LH, Schifferli DM. 1998. Improved allelic exchange vectors and their use to analyze 987P fimbria gene expression. Gene 207:149-157.
145. Schmieger H, Backhaus H. 1976. Altered cotransduction frequencies exhibited by HT-mutants of Salmonella-phags P22. Mol Gen Genet 143:307-309.
146. Kang HY, Dozois CM, Tinge SA, Lee TH, Curtiss R, III. 2002. Transduction-mediated transfer of unmarked deletion and point mutations through use of counterselectable suicide vectors. J Bacteriol 184:307-312.
147. Kang HY, Srinivasan J, Curtiss R, III. 2002. Immune responses to recombinant pneumococcal PspA antigen delivered by live attenuated Salmonella enterica serovar Typhimurium vaccine. Infect Immun 70:1739-1749.
148. Hitchcock PJ, Brown TM. 1983. Morphological heterogeneity among Salmonella lipopolysaccharide chemotypes in silver-stained polyacrylamide gels. J Bacteriol 154:269-277.
149. Chibber S, Bhardwaj SB. 2004. Protection in a mouse peritonitis model mediated by iron-regulated outer-membrane protein of Salmonella typhi coupled to its Vi antigen. J Med Microbiol 53:705-709.
150. Schertzer JW, Whiteley M. 2013. Bacterial outer membrane vesicles in trafficking, communication and the host-pathogen interaction. J Mol Microbiol Biotechnol 23:118-130.
151. Kuehn MJ, Kesty NC. 2005. Bacterial outer membrane vesicles and the host-pathogen interaction. Genes Dev 19:2645-2655.
152. Ho DK, Jarva H, Meri S. 2010. Human complement factor H binds to outer membrane protein Rck of Salmonella. Journal of Immunology 185:1763-1769.
153. Gahring LC, Heffron F, Finlay BB, Falkow S. 1990. Invasion and replication of Salmonella typhimurium in animal cells. Infection and Immunity 58:443-448.
154.  JE, Curtiss R, III. 1989. Cloning and molecular characterization of genes whose products allow Salmonella typhimurium to penetrate tissue culture cells. Proc Natl Acad Sci U S A 86:6383-6387.
JE, Curtiss R, III. 1989. Cloning and molecular characterization of genes whose products allow Salmonella typhimurium to penetrate tissue culture cells. Proc Natl Acad Sci U S A 86:6383-6387.
155. Moser I, Hohmann A, Schmidt G, Rowley D. 1980. Salmonellosis in mice: studies on oral immunization with live avirulent vaccines. Med Microbiol Immunol 168:119-128.
156. Nnalue NA, Stocker BA. 1987. Test of the virulence and live-vaccine efficacy of auxotrophic and galE derivatives of Salmonella choleraesuis. Infect Immun 55:955-962.
157. Germanier R. 1970. Immunity in Experimental Salmonellosis I. Protection Induced by Rough Mutants of Salmonella typhimurium. Infect Immun 2:309-315.
158. Kopecko DJ, Sieber H, Ures JA, Furer A, Schlup J, Knof U, Collioud A, Xu D, Colburn K, Dietrich G. 2009. Genetic stability of vaccine strain Salmonella Typhi Ty21a over 25 years. Int J Med Microbiol 299:233-246.
159. Germanier R, Furer E. 1983. Characteristics of the attenuated oral vaccine strain "S. typhi" Ту 21a. Dev Biol Stand 53:3-7.
160. Edelman R, Levine MM. 1986. Summary of an international workshop on typhoid fever. Rev Infect Dis 8:329-349.
161. Wahdan MH, Serie C, Cerisier Y, Sallam S, Germanier R. 1982. A controlled field trial of live Salmonella typhi strain Ту 21a oral vaccine against typhoid: three-year results. J Infect Dis 145:292-295.
162. Wahdan MH, Serie C, Germanier R, Lackany A, Cerisier Y, Guerin N, Sallam S, Geoffroy P, el Tantawi AS, Guesry P. 1980. A controlled field trial of liver oral typhoid vaccine Ty21a. Bull World Health Organ 58:469-474.
163. Hone DM, Attridge SR, Forrest B, Morona R, Daniels D, LaBrooy JT, Bartholomeusz RC, Shearman DJ, Hackett J. 1988. A galE via (Vi antigen-negative) mutant of Salmonella typhi Ty2 retains virulence in humans. Infect Immun 56:1326-1333.
164. Woodward ТЕ, Woodward WE. 1982. A new oral vaccine against typhoid fever. J Infect Dis 145:289-291.
165. Nnalue NA, Stocker BA. 1986. Some galE mutants of Salmonella choleraesuis retain virulence. Infect Immun 54:635-640.
166. Fukasawa T, Nikaido H. 1961. Galactose-sensitive mutants of Salmonella. II. Bacteriolysis induced by galactose. Biochim Biophys Acta 48:470-483.
167. Nikaido H. 1961. Galactose-sensitive mutants of Salmonella. I. Metabolism of galactose. Biochim Biophys Acta 48:460-469.
168. Postma PW. 1977. Galactose transport in Salmonella typhimurium. J Bacterid 129:630-639.
169.  N, Heine HG, Boos W. 1982. Cloning of mglB, the structural gene for the galactose-binding protein of Salmonella typhimurium and Escherichia coli. Mol Gen Genet 185:473-480.
N, Heine HG, Boos W. 1982. Cloning of mglB, the structural gene for the galactose-binding protein of Salmonella typhimurium and Escherichia coli. Mol Gen Genet 185:473-480.
170. Clarke RC, Gyles CL. 1986. Galactose epimeraseless mutants of Salmonella typhimurium as live vaccines for calves. Can J Vet Res 50:165-173.
171. Shuster CW, Rundell K. 1969. Resistance of Salmonella typhimurium mutants to galactose death. J Bacteriol 100:103-109.
172. Mulford CA, Osborn MJ. 1983. An intermediate step in translocation of lipopolysaccharide to the outer membrane of Salmonella typhimurium. Proc Natl Acad Sci U S A 80:1159-1163.
173. Nagy G, Palkovics T, Otto A, Kusch H, Kocsis B, Dobrindt U, Engelmann S, Hecker M, Emody L, Pal T, Hacker J. 2008. "Gently rough": the vaccine potential of a Salmonella enterica regulatory lipopolysaccharide mutant. J Infect Dis 198:1699-1706.
174. Merighi M, Ellermeier CD, Slauch JM, Gunn JS. 2005. Resolvase-in vivo expression technology analysis of the Salmonella enterica serovar Typhimurium PhoP and PmrA regulons in BALB/c mice. J Bacteriol 187:7407-7416.
175. Brenneman KE, Willingham C, Kong W, Curtiss R, III, Roland KL. 2013. Low-pH rescue of acid-sensitive Salmonella enterica serovar Typhi strains by a Rhamnose-regulated arginine decarboxylase system. J Bacteriol 195:3062-3072.
176. Singh SP, Williams YU, Klebba PE, Macchia P, Miller S. 2000 Immune recognition of porin and lipopolysaccharide epitopes of Salmonella typhimurium in mice. Microb Pathog 28:157-167.
177. Bentley AT, Klebba PE. 1988. Effect of lipopolysaccharide structure on reactivity of antiporin monoclonal antibodies with the bacterial cell surface. J Bacteriol 170:1063-1068.
178. Stevenson G, Andrianopoulos K, Hobbs M, Reeves PR. 1996. Organization of the Escherichia coli K-12 gene cluster responsible for production of the extracellular polysaccharide colanic acid. J Bacteriol 178:4885-4893.
179. Whitfield C. 2006. Biosynthesis and assembly of capsular polysaccharides in Escherichia coli. Annu Rev Biochem 75:39-68.
180. Wang S, Li Y, Scarpellini G, Kong W, Shi H, Baek CH, Gunn B, Wanda SY, Roland KL, Zhang X, Senechal-Willis P, Curtiss R, III. 2010. Salmonella vaccine vectors displaying delayed antigen synthesis in vivo to enhance immunogenicity. Infect Immun 78:3969-3980.
181. Sun W, Wang S, Curtiss R, III. 2008. Highly efficient method for introducing successive multiple scarless gene deletions and markerless gene insertions into the Yersinia pestis chromosome. Applied and Environmental Microbiology 74:4241-4245.
182. Baek CH, Wang S, Roland KL, Curtiss R, III. 2009. Leucine-responsive regulatory protein (Lrp) acts as a virulence repressor in Salmonella enterica serovar Typhimurium. J Bacteriol 191:1278-1292.
183.  MD,
MD,  LT, Kader R, Curtiss R, III, Clark-Curtiss JE. 2012. Live attenuated Salmonella vaccines against Mycobacterium tuberculosis with antigen delivery via the type III secretion system. Infect Immun 80:798-814.
LT, Kader R, Curtiss R, III, Clark-Curtiss JE. 2012. Live attenuated Salmonella vaccines against Mycobacterium tuberculosis with antigen delivery via the type III secretion system. Infect Immun 80:798-814.
184. Smith KD, Andersen-Nissen E, Hayashi F, Strobe K, Bergman MA, Barrett SL, Cookson ВТ, Aderem A. 2003. Toll-like receptor 5 recognizes a conserved site on flagellin required for protofilament formation and bacterial motility. Nat Immunol 4:1247-1253.
185. Murthy KG, Deb A, Goonesekera S, Szabo C, Salzman AL. 2004. Identification of conserved domains in Salmonella muenchen flagellin that are essential for its ability to activate TLR5 and to induce an inflammatory response in vitro. J Biol Chem 279:5667-5675.
186. Cookson ВТ, Bevan MJ. 1997. Identification of a natural T cell epitope presented by Salmonella-infected macrophages and recognized by T cells from orally immunized mice. J Immunol 158:4310-4319.
187.  MD, Yang J, Kader R, Alamuri P, Curtiss R, III., Clark-Curtiss JE. 2012. Live attenuated Salmonella vaccines displaying regulated delayed lysis and delayed antigen synthesis to confer protection against Mycobacterium tuberculosis. Infect Immun 80:815-831.
MD, Yang J, Kader R, Alamuri P, Curtiss R, III., Clark-Curtiss JE. 2012. Live attenuated Salmonella vaccines displaying regulated delayed lysis and delayed antigen synthesis to confer protection against Mycobacterium tuberculosis. Infect Immun 80:815-831.
188. Jiang Y, Mo H, Willingham C, Wang S, Park JY, Kong W, Roland KL, Curtiss R, III. 2015. Protection Against Necrotic Enteritis in Broiler Chickens by Regulated Delayed Lysis Salmonella Vaccines. Avian Dis 59:475-485.
189. Xin W, Wanda SY, Zhang X, Santander J, Scarpellini G, Ellis K, Alamuri P, Curtiss R, III. 2012. The Asd+-DadB+ dual-plasmid system offers a novel means to deliver multiple protective antigens by a recombinant attenuated Salmonella vaccine. Infect Immun 80:3621-3633.
190. Zhang X, Kong W, Ashraf S, Curtiss R, III. 2009. A one-plasmid system to generate influenza virus in cultured chicken cells for potential use in influenza vaccine. J Virol 83:9296-9303.
191. Zhang X, Curtiss R, III. 2015. Efficient generation of influenza virus with a mouse RNA polymerase I-driven all-in-one plasmid. Virol J 12:95.
192. Zhang X, Kong W, Wanda SY, Xin W, Alamuri P, Curtiss R, III. 2015. Generation of influenza virus from avian cells infected by Salmonella carrying the viral genome. PLoS One 10:e0119041.
193. Sanapala S, Rahav H, Patel H, Sun W, Curtiss R, III. 2016. Multiple antigens of Yersinia pestis delivered by live recombinant attenuated Salmonella vaccine strains elicit protective immunity against plague. Vaccine 34:2410-2416.
194. Zhang X, Wanda SY, Brenneman K, Kong W, Roland K, Curtiss R, III. 2011. Improving Salmonella vector with rec mutation to stabilize the DNA cargoes. BMC Microbiol 11:31.
195. Marvin HJ, ter Beest MB, Witholt B. 1989. Release of outer membrane fragments from wild-type Escherichia coli and from several E. coli lipopolysaccharide mutants by EDTA and heat shock treatments. J Bacteriol 171:5262-5267.
196. Witholt B, Boekhout M, Brock M, Kingma J, Heerikhuizen HV, Leij LD. 1976 An efficient and reproducible procedure for the formation of spheroplasts from variously grown Escherichia coli. Anal Biochem 74:160-170.
197. Juarez-Rodriguez MD, Arteaga-Cortes LT, Kader R, Curtiss R, III., Clark-Curtiss JE. 2012. Live attenuated Salmonella vaccines against Mycobacterium tuberculosis with antigen delivery via the type III secretion system. Infect Immun 80:798-814.
198. Konjufca V, Wanda SY, Jenkins MC, Curtiss R, III. 2006. A recombinant attenuated Salmonella enterica serovar Typhimurium vaccine encoding Eimeria acervulina antigen offers protection against E. acervulina challenge. Infect Immun 74:6785-6796.
199. Konjufca V, Jenkins M, Wang S, MD, Curtiss R, III. 2008. Immunogenicity of recombinant attenuated Salmonella enterica serovar Typhimurium vaccine strains carrying a gene that encodes Eimeria tenella antigen SO7. Infect Immun 76:5745-5753.
200. Shi H, Santander J, Brenneman KE, Wanda SY, Wang S, Senechal P, Sun W, Roland KL, Curtiss R, III. 2010. Live recombinant Salmonella Typhi vaccines constructed to investigate the role of rpoS in eliciting immunity to a heterologous antigen. PLoS One 5:e11142.
201. Czerkinsky CC, L. A. Nilsson, H. Nygren, O. Ouchterlony, and A. Tarkowski. 1983. A solid-phase enzyme-linked immunospot (ELISPOT) assay for enumeration of specific antibody-secreting cells. J Immunol Methods 65:109-121.
202. Li Y, Wang S, Scarpellini G, Gunn B, Xin W, Wanda SY, Roland KL, Curtiss R, III. 2009. Evaluation of new generation Salmonella enterica serovar Typhimurium vaccines with regulated delayed attenuation to induce immune responses against PspA. Proc Natl Acad Sci U S A 106:593-598.
203. Bonato VLD, V. M. F. Lima, R. E. Tascon, D. B. Lowrie, and C. L. Silva. 1998. Identification and characterization of protective T cells in hsp65 DNA-vaccinated and Mycobacterium tuberculosis-infected mice. Infect Immun 66:169-175.
204. Okamoto Y, H. Murakami, and M. Nishida. 1997. Detection of interleukin 6-producing cells among various organs in normal mice with an improved enzyme-linked immunospot (ELISPOT) assay. Endocr J 44:349-355.
205. Broz P, Newton K, Lamkanfi M, Mariathasan S, Dixit VM, Monack DM. 2010 Redundant roles for inflammasome receptors NLRP3 and NLRC4 in host defense against Salmonella. J Exp Med 207:1745-1755.
206. Rathinam VA, Vanaja SK, Waggoner L, Sokolovska A, Becker C, Stuart LM, Leong JM, Fitzgerald KA. 2012. TRIF licenses caspase-11-dependent NLRP3 inflammasome activation by gram-negative bacteria. Cell 150:606-619.
207. Riedemann NC, Guo RF, Sarma VJ, Laudes IJ, Huber-Lang M, Warner RL, Albrecht EA, Speyer CL, Ward PA. 2002. Expression and function of the C5a receptor in rat alveolar epithelial cells. J Immunol 168:1919-1925.
208. Montz H, Koch KC, Zierz R, Gotze O. 1991. The role of C5a in interleukin-6 production induced by lipopolysaccharide or interleukin-1. Immunology 74:373-379.
209. Buckner CM, Kardava L, Moir S. 2013. Evaluation of В cell function in patients with HIV. Curr Protoc Immunol Chapter 12:Unit 12 13.
210. Flores-Langarica A, Bobat S, Marshall JL, Yam-Puc JC, Cook CN, Serre K, Kingsley RA, Flores-Romo L, Uematsu S, Akira S, Henderson IR, Toellner KM, Cunningham AF. 2015. Soluble flagellin coimmunization attenuates Th1 priming to Salmonella and clearance by modulating dendritic cell activation and cytokine production. Eur J Immunol 45:2299-2311.
211. Quah BJ, Wijesundara DK, Ranasinghe C, Parish CR. 2014. The use of fluorescent target arrays for assessment of T cell responses in vivo. J Vis Exp doi:10.3791/51627:e51627.
212. Quah BJ, Wijesundara DK, Ranasinghe C, Parish CR. 2013. Fluorescent target array T helper assay: a multiplex flow cytometry assay to measure antigen-specific CD4+ T cell-mediated В cell help in vivo. J Immunol Methods 387:181-190.
213. Cretel E, Touchard D, Bongrand P, Pierres A. 2011. A new method for rapid detection of T lymphocyte decision to proliferate after encountering activating surfaces. J Immunol Methods 364:33-39.
214. Quah BJ, Parish CR. 2010. The use of carboxyfluorescein diacetate succinimidyl ester (CFSE) to monitor lymphocyte proliferation. J Vis Exp doi:10.3791/2259.
215. Parish CR, Glidden MH, Quah BJ, Warren HS. 2009. Use of the intracellular fluorescent dye CFSE to monitor lymphocyte migration and proliferation. Curr Protoc Immunol Chapter 4:Unit4 9.
216. Wallace PK, Tario JD, Jr., Fisher JL, Wallace SS, Ernstoff MS, Muirhead KA. 2008. Tracking antigen-driven responses by flow cytometry: monitoring proliferation by dye dilution. Cytometry A 73:1019-1034.
217. Hawkins ED, Hommel M, Turner ML, Battye FL, Markham JF, Hodgkin PD. 2007. Measuring lymphocyte proliferation, survival and differentiation using CFSE time-series data. Nat Protoc 2:2057-2067.
218. Brenchley JM, Douek DC. 2004. Flow cytometric analysis of human antigen-specific T-cell proliferation. Methods Cell Biol 75:481-496.
219. Tennant SM, Schmidlein P, Simon R, Pasetti MF, Galen JE, Levine MM. 2015. Refined live attenuated Salmonella enterica serovar Typhimurium and Enteritidis vaccines mediate homologous and heterologous serogroup protection in mice. Infect Immun 83:4504-4512.
220. Kong Q, Six DA, Roland KL, Liu Q, Gu L, Reynolds CM, Wang X, Raetz CR, Curtiss R, III. 2011. Salmonella synthesizing 1-dephosphorylated lipopolysaccharide exhibits low endotoxic activity while retaining its immunogenicity. J Immunol 187:412-423.
221. Kong Q, Six DA, Liu Q, Gu L, Wang S, Alamuri P, Raetz CR, Curtiss R, III. 2012. Phosphate groups of Lipid A are essential for Salmonella enterica serovar Typhimurium virulence and affect innate and adaptive immunity. Infect Immun 80:3215-3224.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Юниверсити оф Флорида
Кертис, Рой III
Ванг, Шифенг
<120> ИНДУКЦИЯ ЗАЩИТНОГО ИММУНИТЕТА ПРОТИВ АНТИГЕНОВ
<130> 126405-00120
<150> 62/449228
<151> 23 января 2017 г.
<150> 62/541293
<151> 4 августа 2017 г.
<160> 12
<170> Версия PatentIn 3.5
<210> 1
<211> 1215
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 1
atgctaacca catcattaac gttaaataaa gagaaatgga agccgatctg gaataaagcg 60
ctggtttttc tttttgttgc cacgtatttt ctggatggta ttacgcgtta taaacatttg 120
ataatcatac ttatggttat caccgcgatt tatcaggtct cacgctcacc gaaaagtttc 180
ccccctcttt tcaaaaatag cgtattttat agcgtagcag tattatcatt aatccttgtt 240
tattccatac tcatatcgcc agatatgaaa gaaagtttca aggaatttga aaatacggta 300
ctggagggct tcttattata tactttatta attcccgtac tattaaaaga tgaaacaaaa 360
gaaacggttg cgaaaatagt acttttctcc tttttaacaa gtttaggact tcgctgcctt 420
gcagagagta ttctgtatat cgaggactat aataaaggga ttatgccatt cataagctat 480
gcgcatcgac atatgtccga ttccatggtt ttcttatttc cagcattatt gaatatttgg 540
ctgtttagaa aaaatgcaat taagttggtt tttttggtgc ttagcgccat ctaccttttc 600
tttatcctgg gaaccctatc gcgaggggca tggttggcgg tgcttatagt aggtgttctg 660
tgggcaatac tgaaccgcca atggaagtta ataggagttg gtgccatttt attagccatt 720
atcggcgctt tggttatcac tcaacataat aacaaaccag acccagaaca tttactgtat 780
aaattacagc agacagatag ctcatatcgt tatactaacg gaacccaggg caccgcgtgg 840
atactgattc aggaaaaccc gatcaagggc tacggctatg gtaatgatgt gtatgatggt 900
gtttataata aacgcgttgt cgattatcca acgtggacct ttaaagaatc tatcggtccg 960
cataatacca ttctgtacat ctggtttagt gcaggcatat tgggtctggc gagcctggtc 1020
tatttatatg gcgctatcat cagggaaaca gccagctcta ccctcaggaa agtagagata 1080
agcccctaca atgctcatct cttgctattt ttatctttcg tcggttttta tatcgttcgt 1140
ggcaattttg aacaggtcga tattgctcaa attggtatca ttaccggttt tctgctggcg 1200
ctaagaaata gataa 1215
<210> 2
<211> 404
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 2
Met Leu Thr Thr Ser Leu Thr Leu Asn Lys Glu Lys Trp Lys Pro Ile
1 5 10 15
Trp Asn Lys Ala Leu Val Phe Leu Phe Val Ala Thr Tyr Phe Leu Asp
20 25 30
Gly Ile Thr Arg Tyr Lys His Leu Ile Ile Ile Leu Met Val Ile Thr
35 40 45
Ala Ile Tyr Gln Val Ser Arg Ser Pro Lys Ser Phe Pro Pro Leu Phe
50 55 60
Lys Asn Ser Val Phe Tyr Ser Val Ala Val Leu Ser Leu Ile Leu Val
65 70 75 80
Tyr Ser Ile Leu Ile Ser Pro Asp Met Lys Glu Ser Phe Lys Glu Phe
85 90 95
Glu Asn Thr Val Leu Glu Gly Phe Leu Leu Tyr Thr Leu Leu Ile Pro
100 105 110
Val Leu Leu Lys Asp Glu Thr Lys Glu Thr Val Ala Lys Ile Val Leu
115 120 125
Phe Ser Phe Leu Thr Ser Leu Gly Leu Arg Cys Leu Ala Glu Ser Ile
130 135 140
Leu Tyr Ile Glu Asp Tyr Asn Lys Gly Ile Met Pro Phe Ile Ser Tyr
145 150 155 160
Ala His Arg His Met Ser Asp Ser Met Val Phe Leu Phe Pro Ala Leu
165 170 175
Leu Asn Ile Trp Leu Phe Arg Lys Asn Ala Ile Lys Leu Val Phe Leu
180 185 190
Val Leu Ser Ala Ile Tyr Leu Phe Phe Ile Leu Gly Thr Leu Ser Arg
195 200 205
Gly Ala Trp Leu Ala Val Leu Ile Val Gly Val Leu Trp Ala Ile Leu
210 215 220
Asn Arg Gln Trp Lys Leu Ile Gly Val Gly Ala Ile Leu Leu Ala Ile
225 230 235 240
Ile Gly Ala Leu Val Ile Thr Gln His Asn Asn Lys Pro Asp Pro Glu
245 250 255
His Leu Leu Tyr Lys Leu Gln Gln Thr Asp Ser Ser Tyr Arg Tyr Thr
260 265 270
Asn Gly Thr Gln Gly Thr Ala Trp Ile Leu Ile Gln Glu Asn Pro Ile
275 280 285
Lys Gly Tyr Gly Tyr Gly Asn Asp Val Tyr Asp Gly Val Tyr Asn Lys
290 295 300
Arg Val Val Asp Tyr Pro Thr Trp Thr Phe Lys Glu Ser Ile Gly Pro
305 310 315 320
His Asn Thr Ile Leu Tyr Ile Trp Phe Ser Ala Gly Ile Leu Gly Leu
325 330 335
Ala Ser Leu Val Tyr Leu Tyr Gly Ala Ile Ile Arg Glu Thr Ala Ser
340 345 350
Ser Thr Leu Arg Lys Val Glu Ile Ser Pro Tyr Asn Ala His Leu Leu
355 360 365
Leu Phe Leu Ser Phe Val Gly Phe Tyr Ile Val Arg Gly Asn Phe Glu
370 375 380
Gln Val Asp Ile Ala Gln Ile Gly Ile Ile Thr Gly Phe Leu Leu Ala
385 390 395 400
Leu Arg Asn Arg
<210> 3
<211> 453
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 3
atgactgaca acaataccgc attaaagaag gctggcctga aagtaacgct tcctcgttta 60
aaaattctgg aagttcttca ggaaccagat aaccatcacg tcagtgcgga agatttatac 120
aaacgcctga tcgacatggg tgaagaaatc ggtctggcaa ccgtataccg tgtgctgaac 180
cagtttgacg atgccggtat cgtgacccgc cataattttg aaggcggtaa atccgttttt 240
gaactgacgc aacagcatca tcacgaccat cttatctgcc ttgattgcgg aaaagtgatt 300
gaatttagtg atgactctat tgaagcgcgc cagcgtgaaa ttgcggcgaa acacggtatt 360
cgtttaacta atcacagcct ctatctttac ggccactgcg ctgaaggcga ctgccgcgaa 420
gacgagcacg cgcacgatga cgcgactaaa taa 453
<210> 4
<211> 150
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 4
Met Thr Asp Asn Asn Thr Ala Leu Lys Lys Ala Gly Leu Lys Val Thr
1 5 10 15
Leu Pro Arg Leu Lys Ile Leu Glu Val Leu Gln Glu Pro Asp Asn His
20 25 30
His Val Ser Ala Glu Asp Leu Tyr Lys Arg Leu Ile Asp Met Gly Glu
35 40 45
Glu Ile Gly Leu Ala Thr Val Tyr Arg Val Leu Asn Gln Phe Asp Asp
50 55 60
Ala Gly Ile Val Thr Arg His Asn Phe Glu Gly Gly Lys Ser Val Phe
65 70 75 80
Glu Leu Thr Gln Gln His His His Asp His Leu Ile Cys Leu Asp Cys
85 90 95
Gly Lys Val Ile Glu Phe Ser Asp Asp Ser Ile Glu Ala Arg Gln Arg
100 105 110
Glu Ile Ala Ala Lys His Gly Ile Arg Leu Thr Asn His Ser Leu Tyr
115 120 125
Leu Tyr Gly His Cys Ala Glu Gly Asp Cys Arg Glu Asp Glu His Ala
130 135 140
His Asp Asp Ala Thr Lys
145 150
<210> 5
<211> 453
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 5
atgactgaca acaataccgc attaaagaag gctggcctga aagtaacgct tcctcgttta 60
aaaattctgg aagttcttca ggaaccagat aaccatcacg tcagtgcgga agatttatac 120
aaacgcctga tcgacatggg tgaagaaatc ggtctggcaa ccgtataccg tgtgctgaac 180
cagtttgacg atgccggtat cgtgacccgc cataattttg aaggcggtaa atccgttttt 240
gaactgacgc aacagcatca tcacgaccat cttatctgcc ttgattgcgg aaaagtgatt 300
gaatttagtg atgactctat tgaagcgcgc cagcgtgaaa ttgcggcgaa acacggtatt 360
cgtttaacta atcacagcct ctatctttac ggccactgcg ctgaaggcga ctgccgcgaa 420
gacgagcacg cgcacgatga cgcgactaaa taa 453
<210> 6
<211> 150
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 6
Met Thr Asp Asn Asn Thr Ala Leu Lys Lys Ala Gly Leu Lys Val Thr
1 5 10 15
Leu Pro Arg Leu Lys Ile Leu Glu Val Leu Gln Glu Pro Asp Asn His
20 25 30
His Val Ser Ala Glu Asp Leu Tyr Lys Arg Leu Ile Asp Met Gly Glu
35 40 45
Glu Ile Gly Leu Ala Thr Val Tyr Arg Val Leu Asn Gln Phe Asp Asp
50 55 60
Ala Gly Ile Val Thr Arg His Asn Phe Glu Gly Gly Lys Ser Val Phe
65 70 75 80
Glu Leu Thr Gln Gln His His His Asp His Leu Ile Cys Leu Asp Cys
85 90 95
Gly Lys Val Ile Glu Phe Ser Asp Asp Ser Ile Glu Ala Arg Gln Arg
100 105 110
Glu Ile Ala Ala Lys His Gly Ile Arg Leu Thr Asn His Ser Leu Tyr
115 120 125
Leu Tyr Gly His Cys Ala Glu Gly Asp Cys Arg Glu Asp Glu His Ala
130 135 140
His Asp Asp Ala Thr Lys
145 150
<210> 7
<211> 1011
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 7
atgccgatta ctatagggaa tggtttttta aaaagtgaaa tccttaccaa ctccccaagg 60
aatacgaaag aagcatggtg gaaagtttta tgggaaaaaa ttaaagactt ctttttttct 120
actggcaaag caaaagcgga ccgttgtcta catgagatgt tgtttgccga acgcgccccc 180
acacgagagc ggcttacaga gatttttttt gagttgaaag agttagcctg cgcatcgcaa 240
agagatagat ttcaggttca taatcctcat gaaaatgatg ccaccattat tcttcgcatc 300
atggatcaaa acgaagagaa cgaattgtta cgtatcactc aaaataccga tacctttagc 360
tgtgaagtca tggggaatct ttatttttta atgaaagatc gcccggatat tttaaaatcg 420
catccacaaa tgacggccat gattaagaga agatatagcg aaatcgtaga ctaccccctc 480
ccttcgacat tatgtctcaa tcctgctggc gcgccgatat tatcggttcc attagacaac 540
atagaggggt atttatatac tgaattgaga aaaggacatt tagatgggtg gaaagcgcaa 600
gaaaaggcaa cctacctggc agcgaaaatt cagtctggga ttgaaaagac aacgcgcatt 660
ttacaccatg cgaatatatc cgaaagtact cagcaaaacg catttttaga aacaatggcg 720
atgtgtggat taaaacagct tgaaatacca ccaccgcata cccacatacc tattgaaaaa 780
atggtaaaag aggttttact agcggataag acgtttcagg cgttcctcgt aacggatccc 840
agcaccagcc aaagtatgtt agctgagata gtcgaagcca tctctgatca ggtttttcac 900
gccattttta gaatagaccc ccaggctata caaaaaatgg cggaagaaca gttaaccacg 960
ctacacgttc gctcagaaca acaaagcggc tgtttatgtt gttttttata a 1011
<210> 8
<211> 336
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 8
Met Pro Ile Thr Ile Gly Asn Gly Phe Leu Lys Ser Glu Ile Leu Thr
1 5 10 15
Asn Ser Pro Arg Asn Thr Lys Glu Ala Trp Trp Lys Val Leu Trp Glu
20 25 30
Lys Ile Lys Asp Phe Phe Phe Ser Thr Gly Lys Ala Lys Ala Asp Arg
35 40 45
Cys Leu His Glu Met Leu Phe Ala Glu Arg Ala Pro Thr Arg Glu Arg
50 55 60
Leu Thr Glu Ile Phe Phe Glu Leu Lys Glu Leu Ala Cys Ala Ser Gln
65 70 75 80
Arg Asp Arg Phe Gln Val His Asn Pro His Glu Asn Asp Ala Thr Ile
85 90 95
Ile Leu Arg Ile Met Asp Gln Asn Glu Glu Asn Glu Leu Leu Arg Ile
100 105 110
Thr Gln Asn Thr Asp Thr Phe Ser Cys Glu Val Met Gly Asn Leu Tyr
115 120 125
Phe Leu Met Lys Asp Arg Pro Asp Ile Leu Lys Ser His Pro Gln Met
130 135 140
Thr Ala Met Ile Lys Arg Arg Tyr Ser Glu Ile Val Asp Tyr Pro Leu
145 150 155 160
Pro Ser Thr Leu Cys Leu Asn Pro Ala Gly Ala Pro Ile Leu Ser Val
165 170 175
Pro Leu Asp Asn Ile Glu Gly Tyr Leu Tyr Thr Glu Leu Arg Lys Gly
180 185 190
His Leu Asp Gly Trp Lys Ala Gln Glu Lys Ala Thr Tyr Leu Ala Ala
195 200 205
Lys Ile Gln Ser Gly Ile Glu Lys Thr Thr Arg Ile Leu His His Ala
210 215 220
Asn Ile Ser Glu Ser Thr Gln Gln Asn Ala Phe Leu Glu Thr Met Ala
225 230 235 240
Met Cys Gly Leu Lys Gln Leu Glu Ile Pro Pro Pro His Thr His Ile
245 250 255
Pro Ile Glu Lys Met Val Lys Glu Val Leu Leu Ala Asp Lys Thr Phe
260 265 270
Gln Ala Phe Leu Val Thr Asp Pro Ser Thr Ser Gln Ser Met Leu Ala
275 280 285
Glu Ile Val Glu Ala Ile Ser Asp Gln Val Phe His Ala Ile Phe Arg
290 295 300
Ile Asp Pro Gln Ala Ile Gln Lys Met Ala Glu Glu Gln Leu Thr Thr
305 310 315 320
Leu His Val Arg Ser Glu Gln Gln Ser Gly Cys Leu Cys Cys Phe Leu
325 330 335
<210> 9
<211> 1083
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 9
gtgaaaccag taacgttata cgatgtcgca gagtatgccg gtgtctctta tcagaccgtt 60
tcccgcgtgg tgaaccaggc cagccacgtt tctgcgaaaa cgcgggaaaa agtggaagcg 120
gcgatggcgg agctgaatta cattcccaac cgcgtggcac aacaactggc gggcaaacag 180
tcgttgctga ttggcgttgc cacctccagt ctggccctgc acgcgccgtc gcaaattgtc 240
gcggcgatta aatctcgcgc cgatcaactg ggtgccagcg tggtggtgtc gatggtagaa 300
cgaagcggcg tcgaagcctg taaagcggcg gtgcacaatc ttctcgcgca acgcgtcagt 360
gggctgatca ttaactatcc gctggatgac caggatgcca ttgctgtgga agctgcctgc 420
actaatgttc cggcgttatt tcttgatgtc tctgaccaga cacccatcaa cagtattatt 480
ttctcccatg aagacggtac gcgactgggc gtggagcatc tggtcgcatt gggtcaccag 540
caaatcgcgc tgttagcggg cccattaagt tctgtctcgg cgcgtctgcg tctggctggc 600
tggcataaat atctcactcg caatcaaatt cagccgatag cggaacggga aggcgactgg 660
agtgccatgt ccggttttca acaaaccatg caaatgctga atgagggcat cgttcccact 720
gcgatgctgg ttgccaacga tcagatggcg ctgggcgcaa tgcgcgccat taccgagtcc 780
gggctgcgcg ttggtgcgga tatctcggta gtgggatacg acgataccga agacagctca 840
tgttatatcc cgccgttaac caccatcaaa caggattttc gcctgctggg gcaaaccagc 900
gtggaccgct tgctgcaact ctctcagggc caggcggtga agggcaatca gctgttgccc 960
gtctcactgg tgaaaagaaa aaccaccctg gcgcccaata cgcaaaccgc ctctccccgc 1020
gcgttggccg attcattaat gcagctggca cgacaggttt cccgactgga aagcgggcag 1080
tga 1083
<210> 10
<211> 360
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 10
Met Lys Pro Val Thr Leu Tyr Asp Val Ala Glu Tyr Ala Gly Val Ser
1 5 10 15
Tyr Gln Thr Val Ser Arg Val Val Asn Gln Ala Ser His Val Ser Ala
20 25 30
Lys Thr Arg Glu Lys Val Glu Ala Ala Met Ala Glu Leu Asn Tyr Ile
35 40 45
Pro Asn Arg Val Ala Gln Gln Leu Ala Gly Lys Gln Ser Leu Leu Ile
50 55 60
Gly Val Ala Thr Ser Ser Leu Ala Leu His Ala Pro Ser Gln Ile Val
65 70 75 80
Ala Ala Ile Lys Ser Arg Ala Asp Gln Leu Gly Ala Ser Val Val Val
85 90 95
Ser Met Val Glu Arg Ser Gly Val Glu Ala Cys Lys Ala Ala Val His
100 105 110
Asn Leu Leu Ala Gln Arg Val Ser Gly Leu Ile Ile Asn Tyr Pro Leu
115 120 125
Asp Asp Gln Asp Ala Ile Ala Val Glu Ala Ala Cys Thr Asn Val Pro
130 135 140
Ala Leu Phe Leu Asp Val Ser Asp Gln Thr Pro Ile Asn Ser Ile Ile
145 150 155 160
Phe Ser His Glu Asp Gly Thr Arg Leu Gly Val Glu His Leu Val Ala
165 170 175
Leu Gly His Gln Gln Ile Ala Leu Leu Ala Gly Pro Leu Ser Ser Val
180 185 190
Ser Ala Arg Leu Arg Leu Ala Gly Trp His Lys Tyr Leu Thr Arg Asn
195 200 205
Gln Ile Gln Pro Ile Ala Glu Arg Glu Gly Asp Trp Ser Ala Met Ser
210 215 220
Gly Phe Gln Gln Thr Met Gln Met Leu Asn Glu Gly Ile Val Pro Thr
225 230 235 240
Ala Met Leu Val Ala Asn Asp Gln Met Ala Leu Gly Ala Met Arg Ala
245 250 255
Ile Thr Glu Ser Gly Leu Arg Val Gly Ala Asp Ile Ser Val Val Gly
260 265 270
Tyr Asp Asp Thr Glu Asp Ser Ser Cys Tyr Ile Pro Pro Leu Thr Thr
275 280 285
Ile Lys Gln Asp Phe Arg Leu Leu Gly Gln Thr Ser Val Asp Arg Leu
290 295 300
Leu Gln Leu Ser Gln Gly Gln Ala Val Lys Gly Asn Gln Leu Leu Pro
305 310 315 320
Val Ser Leu Val Lys Arg Lys Thr Thr Leu Ala Pro Asn Thr Gln Thr
325 330 335
Ala Ser Pro Arg Ala Leu Ala Asp Ser Leu Met Gln Leu Ala Arg Gln
340 345 350
Val Ser Arg Leu Glu Ser Gly Gln
355 360
<210> 11
<211> 4844
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 11
gggcgaattc gagctcggta ccctcgaggc tgaatttcat tacgaccagt ctaaaaagcg 60
cctgaattcg cgaccttctc gttactgaca ggaaaatggg ccattggcaa ccagggaaag 120
atgaacgtga tgatgttcac aatttgctga attgtggtga tgtgatgctc accgcatttc 180
ctgaaaattc acgctgtatc ttgaaaaatc gacgtttttt acgtggtttt ccgtcgaaaa 240
tttaaggtaa gaacctgacc tcgtgattac tatttcgccg tgttgacgac atcaggaggc 300
cagtatgacc gtattacata gtgtggattt ttttccgtct ggtaacgcgt ccgtggcgat 360
agaaccccgg ctcccgcagg cggattttcc tgaacatcat catgattttc atgaaattgt 420
gattgtcgaa catggcacgg gtattcatgt gtttaatggg cagccctata ccatcaccgg 480
tggcacggtc tgtttcgtac gcgatcatga tcggcatctg tatgaacata ccgataatct 540
gtgtctgacc aatgtgctgt atcgctcgcc ggatcgattt cagtttctcg ccgggctgaa 600
tcagttgctg ccacaagagc tggatgggca gtatccgtct cactggcgcg ttaaccacag 660
cgtattgcag caggtgcgac agctggttgc acagatggaa cagcaggaag gggaaaatga 720
tttaccctcg accgccagtc gcgagatctt gtttatgcaa ttactgctct tgctgcgtaa 780
aagcagtttg caggagaacc tggaaaacag cgcatcacgt ctcaacttgc ttctggcctg 840
gctggaggac cattttgccg atgaggtgaa ttgggatgcc gtggcggatc aattttctct 900
ttcactgcgt acgctacatc ggcagcttaa gcagcaaacg ggactgacgc ctcagcgata 960
cctgaaccgc ctgcgactga tgaaagcccg acatctgcta cgccacagcg aggccagcgt 1020
tactgacatc gcctatcgct gtggattcag cgacagtaac cacttttcga cgctttttcg 1080
ccgagagttt aactggtcac cgcgtgatat tcgccaggga cgggatggct ttctgcaata 1140
acgcgaatct tctcaacgta tttgtacgcc atattgcgaa taatcaactt cgttctctgg 1200
ccgaggtagc cacggtggcg catcagttaa aacttctcaa agatgatttt tttgccagcg 1260
accagcaggc agtcgctgtg gctgaccgtt atccgcaaga tgtctttgct gaacatacac 1320
atgatttttg tgagctggtg attgtctggc gcggtaatgg cctgcatgta ctcaacgatc 1380
gcccttatcg cattacccgt ggcgatctct tttacattca tgctgacgat aaacactcct 1440
acgcttccgt taacgatctg gttttgcaga atattattta ttgcccggag cgtctgaagc 1500
tgaatcttga ctggcagggg gcgattccgg gatttaacgc cagcgcaggg caaccacact 1560
ggcgcttagg tagcatgggg atggcgcagg cgcggcaggt tatcggtcag cttgagcatg 1620
aaagtagtca gcatgtgccg tttgctaacg aaatggctga gttgctgttc gggcagttgg 1680
tgatgttgct gaatcgccat cgttacacca gtgattcgtt gccgccaaca tccagcgaaa 1740
cgttgctgga taagctgatt acccggctgg cggctagcct gaaaagtccc tttgcgctgg 1800
ataaattttg tgatgaggca tcgtgcagtg agcgcgtttt gcgtcagcaa tttcgccagc 1860
agactggaat gaccatcaat caatatctgc gacaggtcag agtgtgtcat gcgcaatatc 1920
ttctccagca tagccgcctg ttaatcagtg atatttcgac cgaatgtggc tttgaagata 1980
gtaactattt ttcggtggtg tttacccggg aaaccgggat gacgcccagc cagtggcgtc 2040
atctcaattc gcagaaagat taatctagat aaataaaagc agtttacaac tcctagaatt 2100
gtgaatatat tatcacaatt ctaggataga ataataaaag atctctgcag gcatgcaagc 2160
ttgagtattc tatagtgtca cctaaatagc ttggcgtaat catggtcata gctgtttcct 2220
gtgtgaaatt gttatccgct cacaattcca cacaacatac gagccggaag cataaagtgt 2280
aaagcctggg gtgcctaatg agtgagctaa ctcacattaa ttgcgttgcg ctcactgccc 2340
gctttccagt cgggaaacct gtcgtgccag ctgcattaat gaatcggcca acgcgcgggg 2400
agaggcggtt tgcgtattgg gcgctcttcc gcttcctcgc tcactgactc gctgcgctcg 2460
gtcgttcggc tgcggcgagc ggtatcagct cactcaaagg cggtaatacg gttatccaca 2520
gaatcagggg ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa ggccaggaac 2580
cgtaaaaagg ccgcgttgct ggcgtttttc cataggctcc gcccccctga cgagcatcac 2640
aaaaatcgac gctcaagtca gaggtggcga aacccgacag gactataaag ataccaggcg 2700
tttccccctg gaagctccct cgtgcgctct cctgttccga ccctgccgct taccggatac 2760
ctgtccgcct ttctcccttc gggaagcgtg gcgctttctc atagctcacg ctgtaggtat 2820
ctcagttcgg tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc ccccgttcag 2880
cccgaccgct gcgccttatc cggtaactat cgtcttgagt ccaacccggt aagacacgac 2940
ttatcgccac tggcagcagc cactggtaac aggattagca gagcgaggta tgtaggcggt 3000
gctacagagt tcttgaagtg gtggcctaac tacggctaca ctagaagaac agtatttggt 3060
atctgcgctc tgctgaagcc agttaccttc ggaaaaagag ttggtagctc ttgatccggc 3120
aaacaaacca ccgctggtag cggtggtttt tttgtttgca agcagcagat tacgcgcaga 3180
aaaaaaggat ctcaagaaga tcctttgatc ttttctacgg ggtctgacgc tcagtggaac 3240
gaaaactcac gttaagggat tttggtcatg agattatcaa aaaggatctt cacctagatc 3300
cttttaaatt aaaaatgaag ttttaaatca atctaaagta tatatgagta aacttggtct 3360
gacagttacc aatgcttaat cagtgaggca cctatctcag cgatctgtct atttcgttca 3420
tccatagttg cctgactccc cgtcgtgtag ataactacga tacgggaggg cttaccatct 3480
ggccccagtg ctgcaatgat accgcgagac ccacgctcac cggctccaga tttatcagca 3540
ataaaccagc cagccggaag ggccgagcgc agaagtggtc ctgcaacttt atccgcctcc 3600
atccagtcta ttaattgttg ccgggaagct agagtaagta gttcgccagt taatagtttg 3660
cgcaacgttg ttgccattgc tacaggcatc gtggtgtcac gctcgtcgtt tggtatggct 3720
tcattcagct ccggttccca acgatcaagg cgagttacat gatcccccat gttgtgcaaa 3780
aaagcggtta gctccttcgg tcctccgatc gttgtcagaa gtaagttggc cgcagtgtta 3840
tcactcatgg ttatggcagc actgcataat tctcttactg tcatgccatc cgtaagatgc 3900
ttttctgtga ctggtgagta ctcaaccaag tcattctgag aatagtgtat gcggcgaccg 3960
agttgctctt gcccggcgtc aatacgggat aataccgcgc cacatagcag aactttaaaa 4020
gtgctcatca ttggaaaacg ttcttcgggg cgaaaactct caaggatctt accgctgttg 4080
agatccagtt cgatgtaacc cactcgtgca cccaactgat cttcagcatc ttttactttc 4140
accagcgttt ctgggtgagc aaaaacagga aggcaaaatg ccgcaaaaaa gggaataagg 4200
gcgacacgga aatgttgaat actcatactc ttcctttttc aatattattg aagcatttat 4260
cagggttatt gtctcatgag cggatacata tttgaatgta tttagaaaaa taaacaaata 4320
ggggttccgc gcacatttcc ccgaaaagtg ccacctgacg tctaagaaac cattattatc 4380
atgacattaa cctataaaaa taggcgtatc acgaggccct ttcgtctcgc gcgtttcggt 4440
gatgacggtg aaaacctctg acacatgcag ctcccggaga cggtcacagc ttgtctgtaa 4500
gcggatgccg ggagcagaca agcccgtcag ggcgcgtcag cgggtgttgg cgggtgtcgg 4560
ggctggctta actatgcggc atcagagcag attgtactga gagtgcacca tatgcggtgt 4620
gaaataccgc acagatgcgt aaggagaaaa taccgcatca ggcgccattc gccattcagg 4680
ctgcgcaact gttgggaagg gcgatcggtg cgggcctctt cgctattacg ccagctggcg 4740
aaagggggat gtgctgcaag gcgattaagt tgggtaacgc cagggttttc ccagtcacga 4800
cgttgtaaaa cgacggccag tgaattgtaa tacgactcac tata 4844
<210> 12
<211> 4228
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 12
gggcgaattc gagctcggta ccctcgagtc cataatcagg taatgccgcg ggtgatggat 60
gatgtcgtaa tattgggcac tccctttcag ttgctcaatt atgttatttc acactgctat 120
tgagataatt cacaagtgtg cgctcgctcg caaaataaaa tggaatgatg aaactgggta 180
attccgctag cttttgataa aaattttctc aaagccggtt acgtattacc ggttttgagt 240
ttttgcatga ttcagcagga aaagaaccat gtttactaaa cgtcaccgca tcacattact 300
gttcaatgcc aataaagcct atgaccggca ggtagtagaa ggcgtagggg aatatttaca 360
ggcgtcacaa tcggaatggg atattttcat tgaagaagat ttccgcgccc gcattgataa 420
aatcaaggac tggttaggag atggcgtcat tgccgacttc gacgacaaac agatcgagca 480
agcgctggct gatgtcgacg tccccattgt tggggttggc ggctcgtatc accttgcaga 540
aagttaccca cccgttcatt acattgccac cgataactat gcgctggttg aaagcgcatt 600
tttgcattta aaagagaaag gcgttaaccg ctttgctttt tatggtcttc cggaatcaag 660
cggcaaacgt tgggccactg agcgcgaata tgcatttcgt cagcttgtcg ccgaagaaaa 720
gtatcgcgga gtggtttatc aggggttaga aaccgcgcca gagaactggc aacacgcgca 780
aaatcggctg gcagactggc tacaaacgct accaccgcaa accgggatta ttgccgttac 840
tgacgcccga gcgcggcata ttctgcaagt atgtgaacat ctacatattc ccgtaccgga 900
aaaattatgc gtgattggca tcgataacga agaactgacc cgctatctgt cgcgtgtcgc 960
cctttcttcg gtcgctcagg gcgcgcggca aatgggctat caggcggcaa aactgttgca 1020
tcgattatta gataaagaag aaatgccgct acagcgaatt ttggtcccac cagttcgcgt 1080
cattgaacgg cgctcaacag attatcgctc gctgaccgat cccgccgtta ttcaggccat 1140
gcattacatt cgtaatcacg cctgtaaagg gattaaagtg gatcaggtac tggatgcggt 1200
cgggatctcg cgctccaatc ttgagaagcg ttttaaagaa gaggtgggtg aaaccatcca 1260
tgccatgatt catgccgaga agctggagaa agcgcgcagt ctgctgattt caaccacctt 1320
gtcgatcaat gagatatcgc aaatgtgcgg ttatccatcg ctgcaatatt tctactctgt 1380
ttttaaaaaa gcatatgaca cgacgccaaa agagtatcgc gatgtaaata gcgaggtcat 1440
gttgtaattc tagataaata aaagcagttt acaactccta gaattgtgaa tatattatca 1500
caattctagg atagaataat aaaagatctc tgcaggcatg caagcttgag tattctatag 1560
tgtcacctaa atagcttggc gtaatcatgg tcatagctgt ttcctgtgtg aaattgttat 1620
ccgctcacaa ttccacacaa catacgagcc ggaagcataa agtgtaaagc ctgggtgcct 1680
aatgagtgag ctaactcaca ttaattgcgt tgcgctcact gcccgctttc cagtcgggaa 1740
acctgtcgtg ccagctgcat taatgaatcg gccaacgcgc ggggagaggc ggtttgcgta 1800
ttgggcgctc ttccgcttcc tcgctcactg actcgctgcg ctcggtcgtt cggctgcggc 1860
gagcggtatc agctcactca aaggcggtaa tacggttatc cacagaatca ggggataacg 1920
caggaaagaa catgtgagca aaaggccagc aaaaggccag gaaccgtaaa aaggccgcgt 1980
tgctggcgtt tttccatagg ctccgccccc ctgacgagca tcacaaaaat cgacgctcaa 2040
gtcagaggtg gcgaaacccg acaggactat aaagatacca ggcgtttccc cctggaagct 2100
ccctcgtgcg ctctcctgtt ccgaccctgc cgcttaccgg atacctgtcc gcctttctcc 2160
cttcgggaag cgtggcgctt tctcatagct cacgctgtag gtatctcagt tcggtgtagg 2220
tcgttcgctc caagctgggc tgtgtgcacg aaccccccgt tcagcccgac cgctgcgcct 2280
tatccggtaa ctatcgtctt gagtccaacc cggtaagaca cgacttatcg ccactggcag 2340
cagccactgg taacaggatt agcagagcga ggtatgtagg cggtgctaca gagttcttga 2400
agtggtggcc taactacggc tacactagaa gaacagtatt tggtatctgc gctctgctga 2460
agccagttac cttcggaaaa agagttggta gctcttgatc cggcaaacaa accaccgctg 2520
gtagcggtgg tttttttgtt tgcaagcagc agattacgcg cagaaaaaaa ggatctcaag 2580
aagatccttt gatcttttct acggggtctg acgctcagtg gaacgaaaac tcacgttaag 2640
ggattttggt catgagatta tcaaaaagga tcttcaccta gatcctttta aattaaaaat 2700
gaagttttaa atcaatctaa agtatatatg agtaaacttg gtctgacagt taccaatgct 2760
taatcagtga ggcacctatc tcagcgatct gtctatttcg ttcatccata gttgcctgac 2820
tccccgtcgt gtagataact acgatacggg agggcttacc atctggcccc agtgctgcaa 2880
tgataccgcg agacccacgc tcaccggctc cagatttatc agcaataaac cagccagccg 2940
gaagggccga gcgcagaagt ggtcctgcaa ctttatccgc ctccatccag tctattaatt 3000
gttgccggga agctagagta agtagttcgc cagttaatag tttgcgcaac gttgttgcca 3060
ttgctacagg catcgtggtg tcacgctcgt cgtttggtat ggcttcattc agctccggtt 3120
cccaacgatc aaggcgagtt acatgatccc ccatgttgtg caaaaaagcg gttagctcct 3180
tcggtcctcc gatcgttgtc agaagtaagt tggccgcagt gttatcactc atggttatgg 3240
cagcactgca taattctctt actgtcatgc catccgtaag atgcttttct gtgactggtg 3300
agtactcaac caagtcattc tgagaatagt gtatgcggcg accgagttgc tcttgcccgg 3360
cgtcaatacg ggataatacc gcgccacata gcagaacttt aaaagtgctc atcattggaa 3420
aacgttcttc ggggcgaaaa ctctcaagga tcttaccgct gttgagatcc agttcgatgt 3480
aacccactcg tgcacccaac tgatcttcag catcttttac tttcaccagc gtttctgggt 3540
gagcaaaaac aggaaggcaa aatgccgcaa aaaagggaat aagggcgaca cggaaatgtt 3600
gaatactcat actcttcctt tttcaatatt attgaagcat ttatcagggt tattgtctca 3660
tgagcggata catatttgaa tgtatttaga aaaataaaca aataggggtt ccgcgcacat 3720
ttccccgaaa agtgccacct gacgtctaag aaaccattat tatcatgaca ttaacctata 3780
aaaataggcg tatcacgagg ccctttcgtc tcgcgcgttt cggtgatgac ggtgaaaacc 3840
tctgacacat gcagctcccg gagacggtca cagcttgtct gtaagcggat gccgggagca 3900
gacaagcccg tcagggcgcg tcagcgggtg ttggcgggtg tcggggctgg cttaactatg 3960
cggcatcaga gcagattgta ctgagagtgc accatatgcg gtgtgaaata ccgcacagat 4020
gcgtaaggag aaaataccgc atcaggcgcc attcgccatt caggctgcgc aactgttggg 4080
aagggcgatc ggtgcgggcc tcttcgctat tacgccagct ggcgaaaggg ggatgtgctg 4140
caaggcgatt aagttgggta acgccagggt tttcccagtc acgacgttgt aaaacgacgg 4200
ccagtgaatt gtaatacgac tcactata 4228
<---
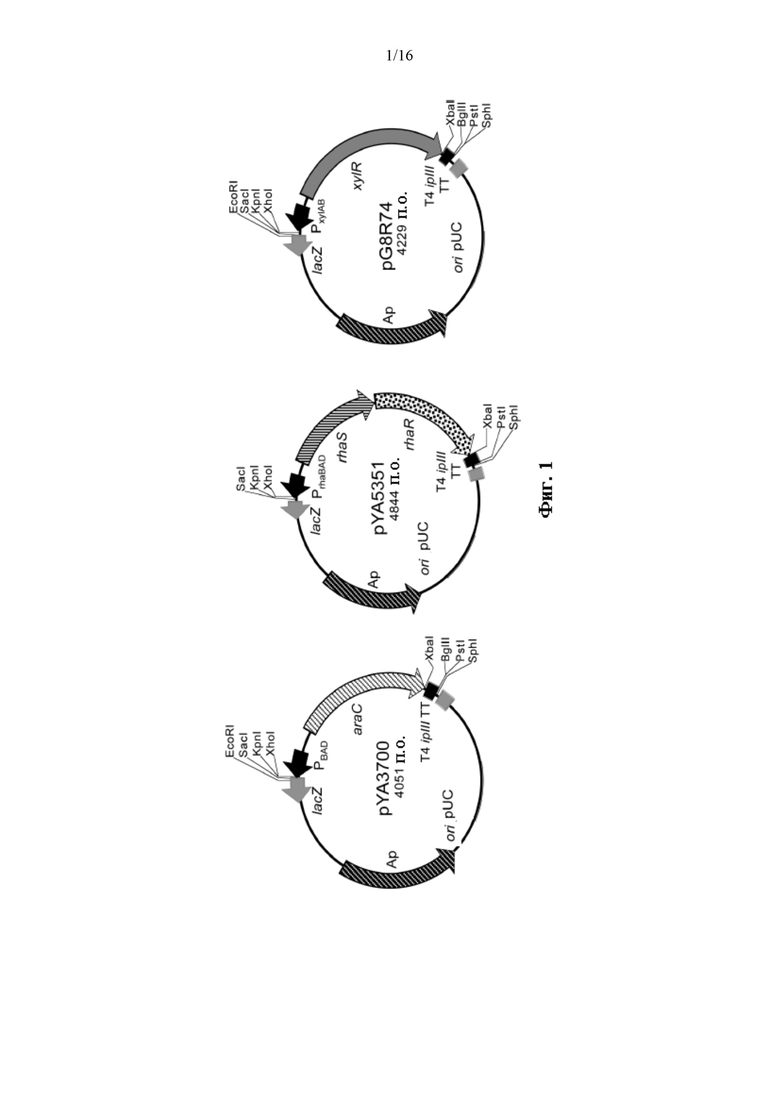
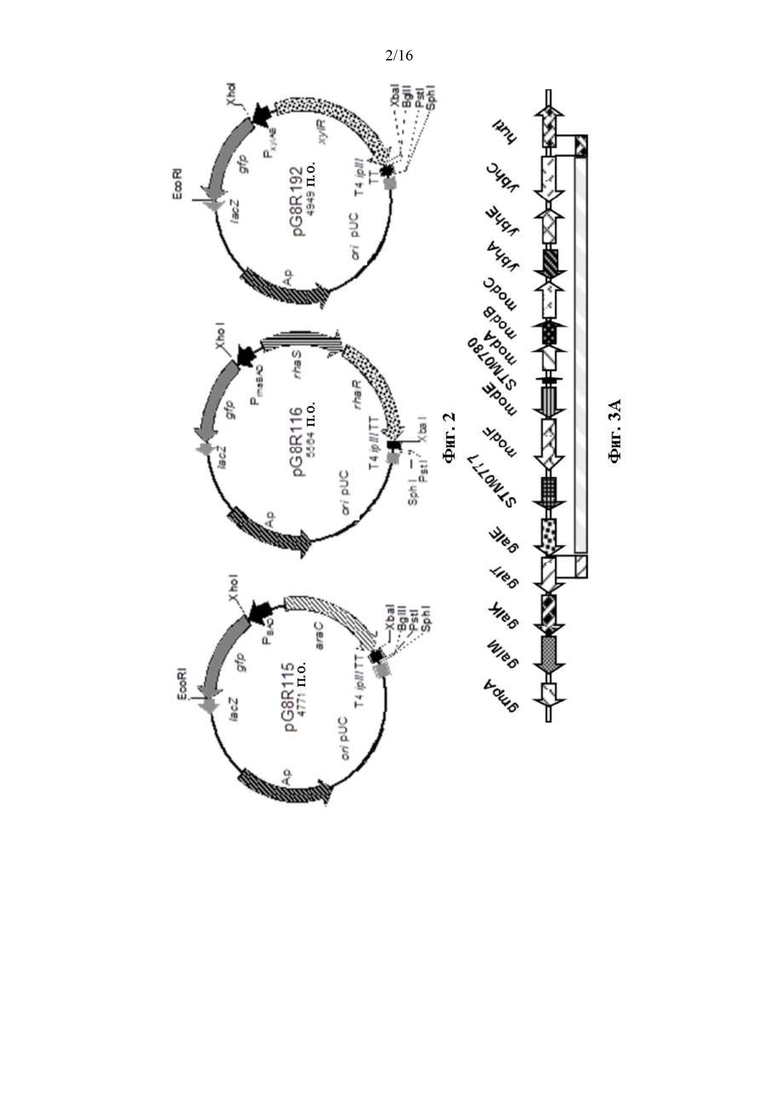

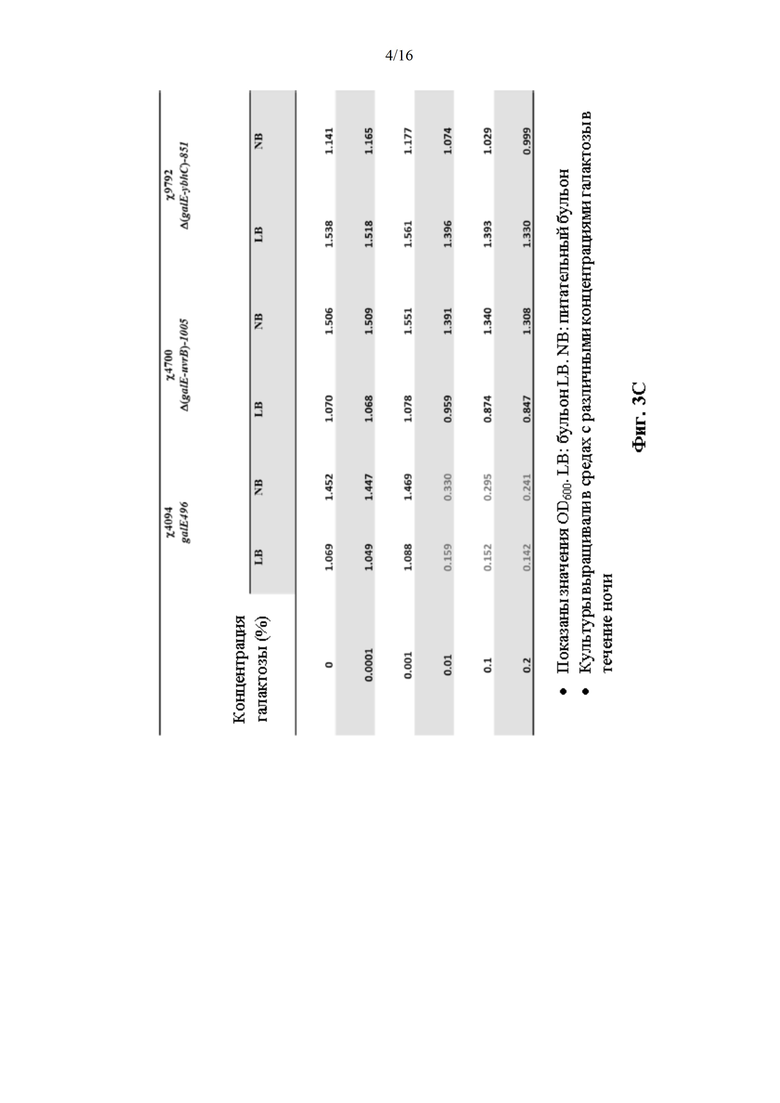
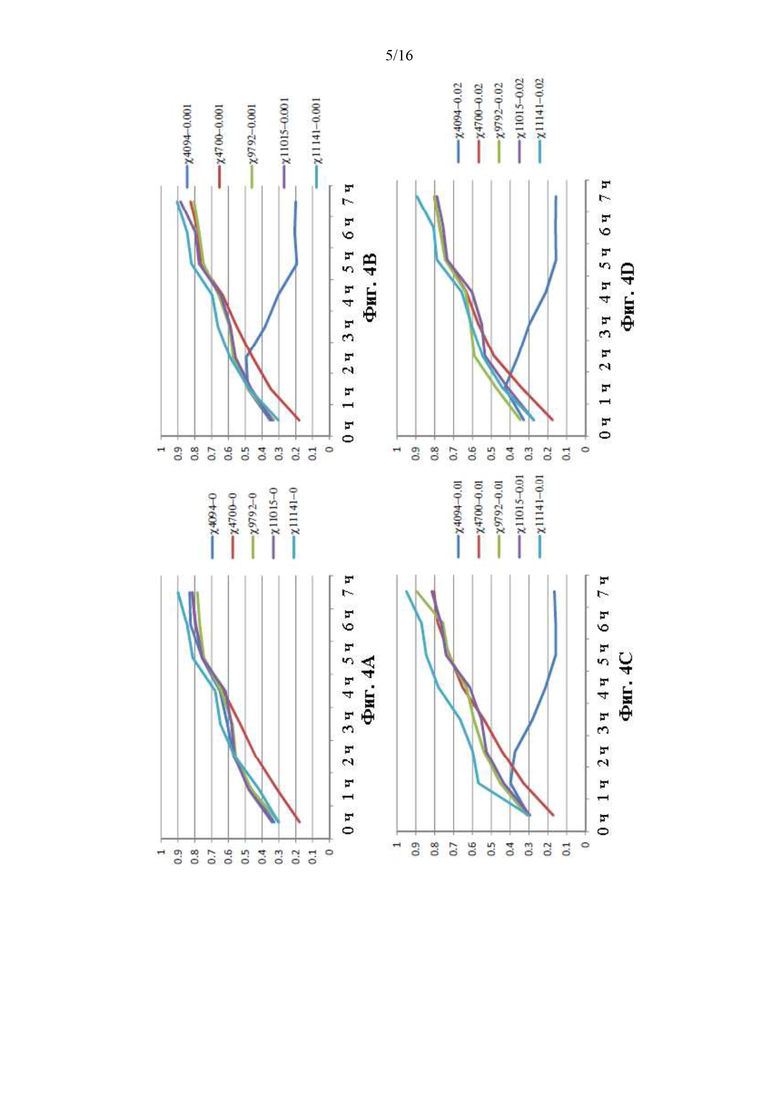

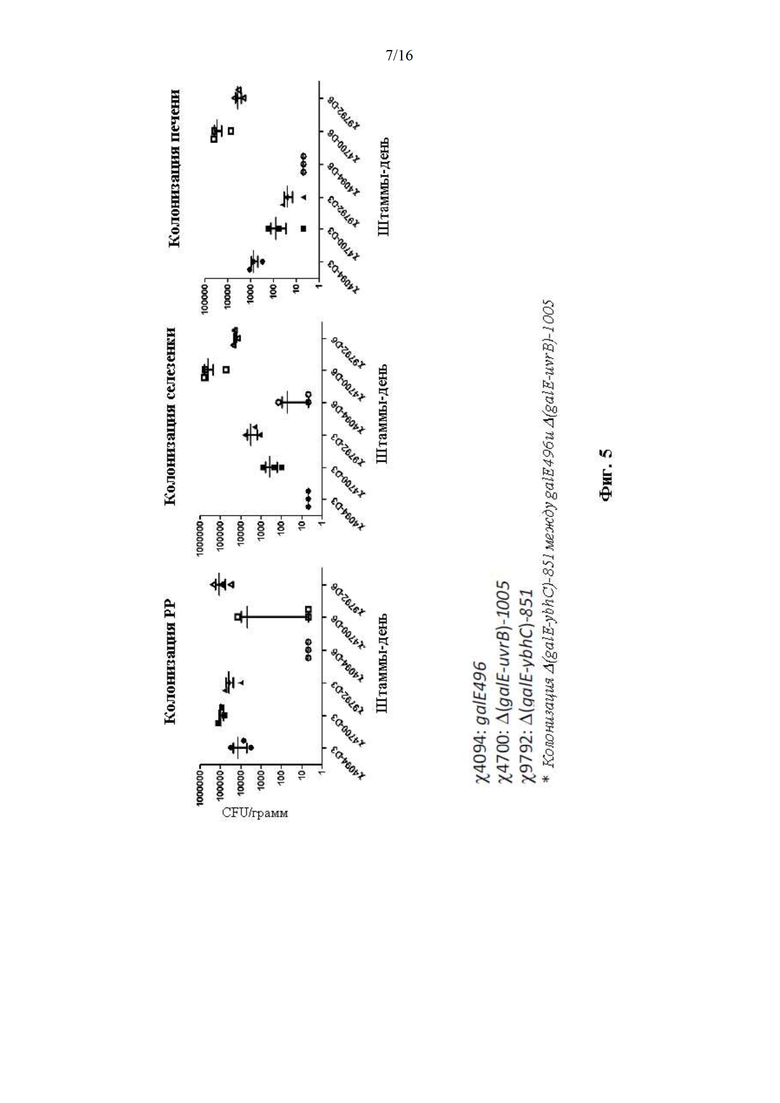

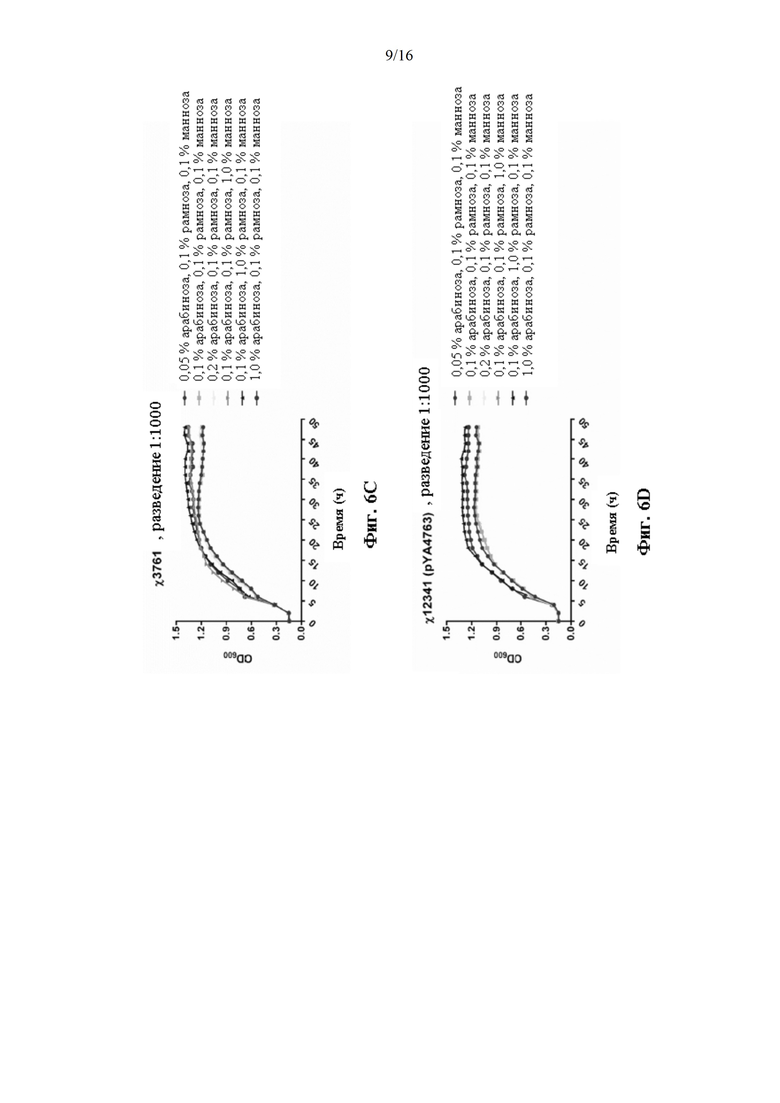

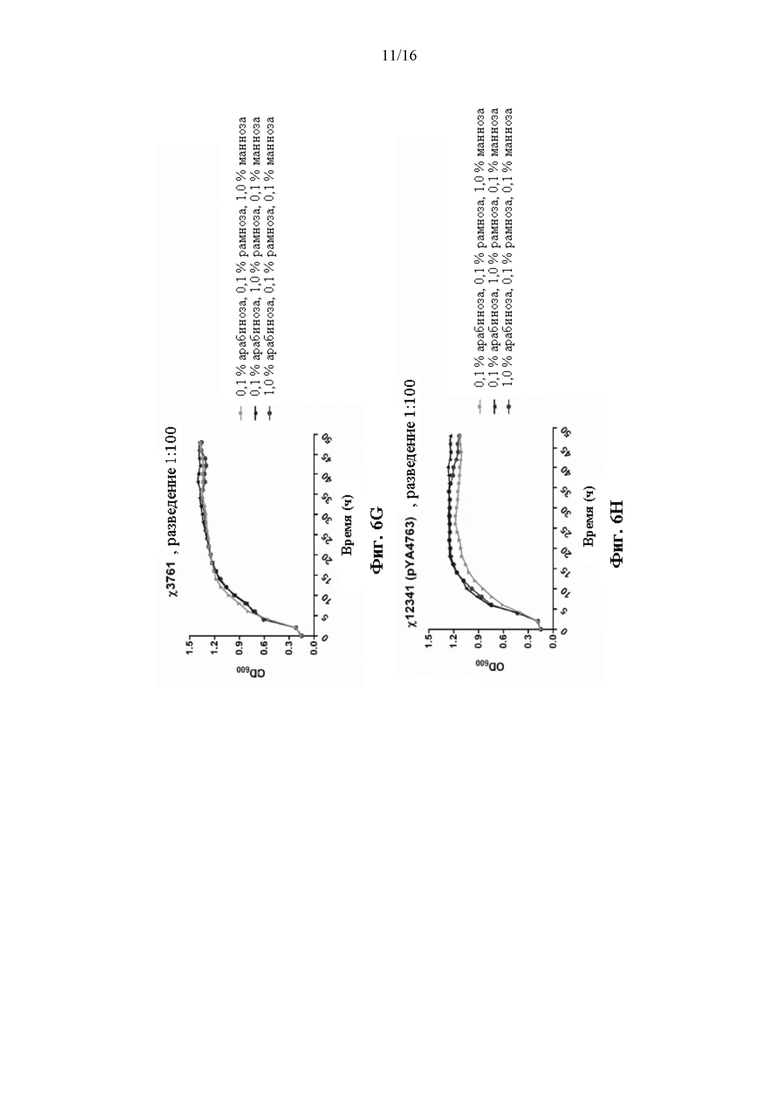





Изобретение относится к рекомбинантной патогенной бактерии и ее применению. Предложена рекомбинантная патогенная бактерия для применения в качестве системы доставки гетерологичного антигена. Указанная бактерия модифицирована для того, чтобы содержать: регулируемый araC ParaBAD ген murA; мутацию по типу делеция/вставка, которая инактивирует экспрессию гена asdA и осуществляет вставку гена с2; делецию в гене pmi; делецию в гене pagL; регулируемый rhaRS PrhaBAD ген waaL; делецию в гене wza-wcaM; мутацию по типу делеция/вставка, которая инактивирует экспрессию гена RelA и осуществляет вставку гена lacI; делецию в гене recF; и делецию в гене sifA. Также предложены рекомбинантная бактерия Salmonella enterica subsp. Enterica для применения в качестве системы доставки гетерологичного антигена, фармацевтическая композиция для вызова иммунного ответа против антигена, представляющего интерес, у субъекта, содержащая рекомбинантную бактерию и фармацевтически приемлемый носитель. Также предложен способ вызова иммунного ответа против антигена, представляющего интерес, у субъекта, предусматривающий введение субъекту эффективного количества указанной фармацевтической композиции. Изобретение обеспечивает регулируемую экспрессию одного или нескольких антигенов, представляющих интерес. 4 н. и 10 з.п. ф-лы, 6 ил., 6 табл., 8 пр.
1. Рекомбинантная патогенная бактерия для применения в качестве системы доставки гетерологичного антигена, где патогенная бактерия модифицирована для того, чтобы содержать:
регулируемый araC ParaBAD ген murA;
мутацию по типу делеция/вставка, которая инактивирует экспрессию гена asdA и
осуществляет вставку гена с2;
делецию в гене pmi;
делецию в гене pagL;
регулируемый rhaRS PrhaBAD ген waaL;
делецию в гене wza-wcaM;
мутацию по типу делеция/вставка, которая инактивирует экспрессию гена RelA и
осуществляет вставку гена lacI;
делецию в гене recF; и
делецию в гене sifA;
где
регулируемый araC ParaBAD ген murA представляет собой ΔPmurA25::TT araC PBAD murA;
мутация по типу делеция/вставка, которая инактивирует экспрессию гена asdA и
осуществляет вставку гена с2 представляет собой ΔasdA27::TT araC PBAD c2;
делеция в гене pmi представляет собой Δpmi-2426;
делеция в гене pagL и регулируемый rhaRS PrhaBAD ген waaL представляют собой ΔwaaL46 ΔpagL64::TT rhaRS PrhaBAD waaL;
делеция в гене wza-wcaM представляет собой Δ(wza-wcaM)-8;
мутация по типу делеция/вставка, которая инактивирует экспрессию гена RelA и
осуществляет вставку гена lacI, представляет собой ΔrelA197::araC PBAD lacI TT;
делеция в гене recF представляет собой ΔrecF126; и
делеция в гене sifA представляет собой ΔsifA26.
2. Бактерия по п. 1, где бактерия дополнительно содержит ген, кодирующий антиген, представляющий интерес, функционально связанный с регулируемым репрессором промотором.
3. Бактерия по п. 2, где регулируемый репрессором промотор представляет собой регулируемый лактозой промотор.
4. Бактерия по п. 3, где регулируемый лактозой промотор представляет собой Ptrc.
5. Бактерия по п. 2 или 3, где антиген, представляющий интерес, представляет собой антиген, полученный из инфекционного агента или опухолевого антигена.
6. Бактерия по п. 5, где антиген представляет собой антиген Clostridium perfringens.
7. Бактерия по п. 6, где антиген Clostridium perfringens представляет собой антиген NetB или его антигенный фрагмент, антиген PlcC или его антигенный фрагмент, или слитый белок, содержащий антиген NetB или его антигенный фрагмент и антиген PlcC, или его антигенный фрагмент.
8. Рекомбинантная бактерия Salmonella enterica subsp. Enterica для применения в качестве системы доставки гетерологичного антигена, при этом бактерия включает:
ΔPmurA25::TT araC PBAD murA;
ΔasdA27::TT araC PBAD c2;
Δpmi-2426;
ΔwaaL46 ΔpagL64::TT rhaRS PrhaBAD waaL;
Δ(wza-wcaM)-8;
ΔrelA197::araC PBAD lacI TT;
ΔrecF126;
ΔsifA26; и
оперон, кодирующий слитый белок, содержащий антиген NetB или его антигенный фрагмент и антиген PlcC или его антигенный фрагмент, функционально связанный с промотором Ptrc.
9. Рекомбинантная бактерия по п. 8, где бактерия содержит плазмиду pYA5112.
10. Фармацевтическая композиция для вызова иммунного ответа против антигена, представляющего интерес, у субъекта, содержащая рекомбинантную бактерию по любому из пп. 1-9 и фармацевтически приемлемый носитель.
11. Способ вызова иммунного ответа против антигена, представляющего интерес, у субъекта, при этом способ предусматривает введение субъекту эффективного количества фармацевтической композиции по п. 10.
12. Способ по п. 11, где субъект имеет некротический энтерит.
13. Способ по п. 11 или 12, где субъектом является цыпленок.
14. Способ по любому из пп. 11-13, где фармацевтическую композицию вводят субъекту с помощью спрея или пероральной иммунизации.
| WO 2009046449 A1, 09.04.2009 | |||
| WO 2010045620 A1, 22.04.2010 | |||
| WO 2010135563 A1, 25.11.2010 | |||
| RU 2009115177 A, 27.10.2010. |

