Изобретение относится к фрагментам ДНК, содержащим гены устойчивости к антрациклиновым антибиотикам, к рекомбинантным векторам, содержащим указанные фрагменты ДНК и к хозяевам, трансформированным указанными векторами.
Антрациклины группы даунорубицинов, такие, как доксорубицин, карминомицин и аклавиномицин, наряду с другими средствами широко используются в противоопухолевой терапии. Они являются поликетидами, полученными при помощи различных линий Streptomyces (S. peucetins, S.coeruleorubidus, S. galilaeus, S. griseus, S. griseorubes, S. Viridochromogenes, u S.bifurcus).
Доксорубицин получают только посредством S.percetius вид calsius. Тип линии S. percetius вид caesius LMRU 3920 (сокращенно "S. peucetius 3920") является доступным и описан в патенте США N 3590028. S. peucetius 3920 депонирован в Институте микробиологии университета Rutges, США, под номером IMRU 3920. Этот штамм и его мутанты, получаемые обычно с помощью классических методов мутагенеза, являются устойчивыми к высоким уровням доксорубицина.
Изучение механизмов, вызывающих устойчивость к указанным веществам, необходимо по двум основным причинам:
а) Имеется множество примеров, когда гены, вовлеченные в биосинтез вторичных метаболитов, образуют кластеры, по крайней мере, с одним геном устойчивости: например, к окситетрациклину (Rhodes P.M. Hunter I.S., Friend E.J. u Warren M., 1984, Trans. Biochem Soc.12, 586-587), эритромицину (Stanzak R. Matsushima P. , Baltz R. H. u Rao R.N, Biotechnology, Vol. 4, March 1986, 229-232), тилозину (Fayerman J.T., Biotechnology Vol. 4, Sept. 1986, 786-789) и тетраценомицину (Motamedi H. Hutchinson C.R.Proc. Natl. Acad. Sci. USA, Vol. 84, 4445-4449, 1987). Клонирование биосинтетических генов может быть полезно с точки зрения изменения путей метаболизма для получения разных молекул или для устранения препятствий на путях биосинтеза, что приводит, таким образом, к увеличению продуктивности штамма.
b) Устойчивость сама по себе подразумевается в регуляторных механизмах, и следовательно, изменяя уровни устойчивости (т.е. увеличивая число аллелей в генотипе), можно тем самым повысить продуктивность штамма. Этот факт давно известен специалистам и нашел свое воплощение в классических методах мутагенеза и произвольного скрининга, и при этом он вновь был положен в основу методов рДНК (Graveri R. и Davies J.E., The Journal of Antibiotics, Jan. 1986, 128-135).
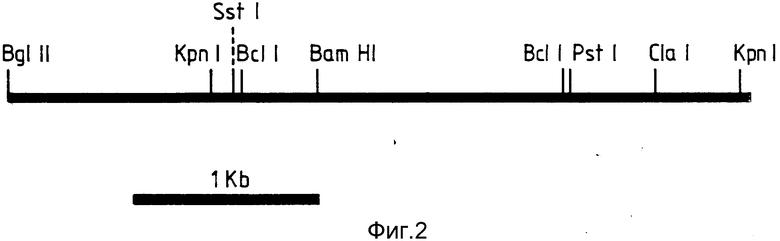
В настоящем изобретении были выделены два сегмента ДНК, которые включают гены устойчивости к доксорубицину. Таким образом, настоящее изобретение обеспечивает фрагмент геномной ДНК, кодирующей устойчивость к доксорубицину, изолированный из штаммf Streptomyces peucetius DSM 4592 или происходящих от него штаммов, содержащий рестрикционные сайты, показанные на фиг.1 или 2.
В дальнейшем, для удобства, ДНК-сегменты, показанные на фиг.1 и 2, будут называться ДНК-вставкой. Настоящее изобретение также относится: к рекомбинантным векторам, обладающим способностью трансформировать клетку-хозяина и содержащим ДНК-вставку или происходящий от нее фрагмент рестрикции, содержащий ген устойчивости к доксорубицину; и к клеткам - хозяевам, трансформированным указанными векторами.
На фиг. 1 представлен анализ рестрикционной карты первой ДНК настоящего изобретения. Она является вставкой в рекомбинантную плазмиду FICE 1 (Rec l). Эта вставка имеет Sau 3AI - концы и вводится в Bgl II - сайт плазмиды pIJ 702. Один Bgl II - сайт восстанавливается после лигирования; на фиг. 2 - анализ рестрикционной карты второй ДНК настоящего изобретения. Она является вставкой в рекомбинантную плазмиду FICE 2 (Rec 2). Эта вставка имеет Sau 3АI концы и вводится в Bgl II - сайт плазмиды pIJ 702. Один сайт Bgl II восстанавливается после лигирования.
Карты, изображенные на фиг. 1 и 2, дают достаточное количество рестриктирующих сайтов для точного распознавания сегментов.
ДНК-вставки и рестрикционные фрагменты настоящего изобретения содержат ген, кодирующий устойчивость к доксорубицину. Для экспрессии такого гена ДНК может иметь свою собственную последовательность регулирования транскрипции и, в частности, свой собственный промотор, который является непосредственно связанным с геном и который распознается РНК-полимеразой клетки-хозяина альтернативно, ДНК- вставка или рестрикционный фрагмент могут быть лигированы к другой последовательности регуляции транскрипции правильным образом или клонированы в вектор у сайта рестрикции, расположенного соответственно рядом с последовательностью регуляции транскрипции в векторе.
ДНК-вставка или фрагмент рестрикции, несущие ген устойчивости к доксорубицину, могут быть клонированы в векторе клонирования рекомбинантной ДНК. Может быть использован любой реплицирующий и интегрирующий и интегрирующий агент, содержащий молекулу ДНК, к которой могут быть добавлены один или более дополнительных сегментов ДНК. Однако обычно вектором является плазмида. Предпочтительной плазмидой является плазмида с большим числом копий pIJ 702 (Katz et al. , J. Gen Microbiol, 1983, 129, 2703-2714). Для введения ДНК-вставки или ее рестрикционного фрагмента в вектор может быть использована любая подходящая техника. Указанное введение может быть осуществлено путем лигирования ДНК в линеаризованный вектор у соответствующего сайта рестрикции. Для этого могут быть использованы прямая комбинация "липких концов" или гомополимерных концов или молекула линкера или адаптора.
Рекомбинантный вектор используют для трансформации соответствующей клетки-хозяина, при этом обычно используют клетки, обладающие способностью проявлять устойчивость доксорубицину. Клетки-хозяева могут быть доксорубицин-восприимчивыми, т.е. не могут расти в присутствии доксорубицина, или доксорубицин-устойчивыми, но при этом могли бы быть полезными вследствие более высокой устойчивости к доксорубицину. Хозяином может быть микроорганизм. Поэтому могут быть трансформированы штаммы S. pencetins, а в частности S. pencetins вид calsins, которые продуцируют доксорубицин, или другие штаммы Streptomyces, которые продуцируют антрациклины. Устойчивость, или повышенная устойчивость к доксорубину дает возможность продуцировать больше доксорубицина клетками указанных штаммов. Таким образом может быть достигнута устойчивость к более высоким концентрациям доксорубицина. Трансформанты штаммов S. pencetins обычно получают путем трансформации протопластов. Таким образом, доксорубицин может быть получен путем культивирования трансформированного штамма S. pencetins с последующим выделением таким образом полученного доксорубицина.
ДНК - вставки получают из геномной ДНК S. pencetins М76. S. pencetins М76 является мутантом штамма S. pencetins 3920, который способен превращать даунорубицин в доксорубицин при высоких уровнях. S. pencetins М76 депонирован в Немецкой коллекции микроорганизмов (Dentsche Sammlung von Microorganismen (DSM), ФРГ, 11 мая 1988 г. под номером допуска D.S.M. 4592. При этом можно также использовать штамм, происходящий из S. pencetins М76, который, как правило, также будет способен превращать даунорубицин в доксорубицин. Таким образом, ДНК - вставки могут быть получены путем: (a) получения библиотеки геномной ДНК S. pencetins М76, или происходящего из него штамма; (b) скрининга этой библиотеки на устойчивость к доксорубицину; (c) получения ДНК-вставки из рекомбинантного вектора, который является частью указанной библиотеки и который при скрининге отобран как положительный на устойчивость к доксорубицину, и (d) необязательно, получения из ДНК-вставки рестрицкционного фрагмента, содержащего ген, кодирующий устойчивость к доксорубицину.
В стадии (a) библиотека может быть получена путем частичного переваривания геномной ДНК штамма S. pencetins М76 или происходящего из него штамма. Предпочтительно использовать рестриктирующий фермент Mb 01. Полученные таким образом фрагменты могут быть фракционированы по размерам. Предпочтительными являются фрагменты размером от 4 до 6 kb. Эти фрагменты лигируют в линеаризованный вектор, такое как pIJ 702. Хозяйские клетки трансформируют смесью для лигирования. Обычно клетки-хозяева являются доксорубицин-восприимчивыми, например, восприимчивыми при концентрации 50 мкг или менее, или, предпочтительно, 30 мкг или менее на 1 мл. Могут быть также трансформированы, например, протопласты S. lividans ТК 23.
В стадии (b) полученные описанным выше способом трансформанты скринируют на устойчивость к доксорубицину. Доксорубицин-устойчивые клоны идентифицируют путем культивирования в среде, содержащей доксорубицин. Указанные клоны выделяют, а рекомбинантные векторы, содержащиеся в них, экстрагируют. После переваривания рекомбинантных векторов соответствующими рестриктирующими ферментами в стадии (c), ДНК штамма S. pencetins М76, вставленная в каждый вектор, может быть идентифицирована. Классифицирована по размерам и картирована. Таким образом, может быть проконтролировано, что вектор содержит ДНК-вставку настоящего изобретения.
Далее две или несколько перекрывающихся вставок, которые целиком или частично охвачены ДНК настоящего изобретения, могут быть выделены. Они могут быть сплавлены вместе путем расщепления в общем рестриктирующем сайте и последовательного легирования с целью получения ДНК настоящего изобретения, сокращенной в длину, с использованием соответствующих рестриктаз, если это необходимо. Рестрикционные фрагменты ДНК-вставки, которая содержит ген, кодирующий устойчивость к доксорубицину, могут быть получены в стадии (d) также путем расщепления ДНК-вставки соответствующим рестриктирующим ферментом.
И, наконец, ДНК настоящего изобретения может быть мутирована способом, не воздействующим на ее способность к приобретению устойчивости к доксорубицину. Это может быть достигнуто, например, путем сайт-направленного мутагенеза. Указанная мутированная ДНК также входит в объем настоящего изобретения.
Приведенный ниже пример иллюстрирует осуществление настоящего изобретения. В указанном примере обозначения TSR, DoxoR и DoxoS означают тиострептон-устойчивый, доксорубицин-устойчивый и доксорубицин-восприимчивый фенотипы, соответственно.
Пример 1. Материалы и методы
Бактериальные штаммы и плазмиды:
Streptomyces pencetins М76, нитеобразный стрептомицет, продуцирующий даунорубицин и доксорубицин и устойчивый к доксорубицину (MIC 250 мкг/мл), и некоторые биосинтетические мутанты, восприимчивые к доксорубицину; S. lividans ТК 23, восприимчивый к доксорубицину.
Плазмида pIJ 702 с большим числом копий была получена из Коллекции культур Джона Иннеса, Норидж, Великобритания.
Среды и буферы
TSB, содержащий 30 г триптического соевого бульона (DIFCO) на литр дистиллированной воды; среда JЕМЕ, содержащая 5 г дрожжевого экстракта (DIFCO), 10 г экстракта солода (DJFCO), 340 г сахарозы, 5 мМ MgCl2 • 6H2O и различные концентрации глицина на литр дистиллированной воды.
Восстановительная среда R2JE описана Chater K.F., Hopwood D.A., Kieser J. и Thompson C.J. (1982) в работе "Gene cloning in streptomyces" 69-95, P. H. Hofschneides and W. Goebbel (ed) "Gene cloning in Organisms other than E. coli", Springer-Verlag, Berlin. Эта среда была получена при следующем составе:
сахароза - 103 г;
смесь микроэлементов - 2 мл;
2,5% K2SO4 - 10 мл;
0,5% KH2PO4 - 10 мл;
MgCl2 • 6H2O - 10,1 г;
1M CaCl2 - 20 мл;
глюкоза - 10 г;
пролин - 3 г;
казаминокислоты 0,1 г;
0,25M TES pH 7,2 - 100 мл;
агар - 22 г;
10% дрожжевой экстракт - 50 мл.
Среда P описана Baltz R.H., J. Gen Microbiol. 107: 93-102 (1978).
Стрептомицеты поддерживались на твердой среде, описанной в патенте США N 3590028, пример 2.
Условия культивирования: для жидких культур оба вида Streptomyces культивировали в 50 мл JЕМЕ + TSB (1 : 1) при 28oC на роторном шейкере при 280 об/мин. Питательную среду засевали гомогенизированными мицелиями. Гомогенизацию осуществляли путем перемешивания мицелиев в пробирке, содержащей стеклянные шарики.
Трансформация протопласта
Мицелии выделяли из 35 мл жидкой культуры (дополненной 0,5% гицином) путем центрифугирования (10 мин, 1500 • г), дважды промывали 10,3% сахарозой, ресуспендированной в 10 мл среды P, содержащей 1 мг/мл лизоцима (SIGMA), и инкубировали в течение 60 мин при 30oC при реципрокном встряхивании (230 об/мин). После образования протопласта суспензию фильтровали через вату, один раз промывали средой P и ресуспендировали в 1 мл среды P. В основном, получали 108 протопластов.
Для каждой трансформации, 200 мкл среды P, содержащей около 2 • 107 протопластов, смешивали с 10 мкл необходимого количества ДНК в Te (Трис-HCl, 10 мМ; ЭДТК, 1 мМ, pH 8,0), и с 800 мкл 25% полиэтиленгликоля (PEG) 1000 в среде P. Протопласты осаждали при помощи центрифугирования, ресуспендировали в 1 мл среды P и высевали на R2JE. После инкубирования в течение 24 ч при 28oC трансформанты отбирались путем полива пластинок с протопластами 3 мл мягким буфером NA (8 г питательного бульона DIFCO и 5 г агара на литр), содержащим антибиотик. Число трансформантов составляло около 1 • 104 - 1 • 107 на мкг ДНК, в соответствии с используемыми штаммами.
Выделение плазмиды и геномной ДНК
Выделение плазмиды и геномной ДНК из стрептомицетов осуществлялось с использованием техники, описанной Hepwood D. A. et al., 1985, "Genetic Manipulation of Streptomyces a laboratory Manual." The John Innes Foundation".
Получение геномной библиотеки S. pencetins М76
Все рестриктирующие ферменты, щелочная фосфатаза телячьей вилочковой железы и лигаза Т4 были получены из BRL (Bethesda, MD) и использовались в соответствии с инструкциями производителя. Геномную ДНК S. pencetins М76 частично переваривали Mbol, и фрагменты размером в пределах от 4 до 6 kb выделяли путем электроэлюирования из агарозного геля. Эти фрагменты лигировали к pIJ 702, линеаризованной Bgl II и обрабатывали фосфатазой. Смесь для лигирования использовали для трансформации S. lividans ТК 23-протопластов, восприимчивых к 30 мгк/мл доксорубицина.
2. Результаты
Клонирование фрагментов ДНК, имеющих устойчивость к доксорубицину в восприимчивых штамма Streptomyces.
Частично Mb 01-переваренную геномную ДНК S. pencetins М76 вставляли в Bgl II-сайт pIJ 702. Смесь для лигирования использовали для трансформации протопластов S. lividans ТК 23. Трансформанты выбирали yf тиострептон-устойчивость и белый цвет, свидетельствующий о инсерционной инактивации гена меланина pIJ 702.
Затем тиострептон-устойчивые белые колонии скринировали на устойчивость к доксорубицину (100 мкг/мл). После чего их высевали на среду R2JE, инкубировали в течение 24 часов при 28oC в присутствии мягкого NA, содержащего 500 мкг/мл доксорубицина; а полученные таким образом два клона TSR и DoxoR идентифицировали.
Экстракция плазмидной ДНК из указанных двух клонов обнаружила присутствие вставок длиной 5,7 kb и 4,4 kb. Эти две рекомбинантные плазмиды, обозначенные, соответственно, FICE 1 и FICE 2, снова использовались для трансформации протопластов S. lividans ТК 23. В обоих случаях трансформация показала, что DoxoR - признак наряду с DoxoR - признаком сообщается с высокой степенью эффективности.
Экспрессия DoxoR-признака в DoxoS-мутантах S.pencetins
Затем две рекомбинантные плазмиды вводили в некоторые производные от S. pencetins М76, являющихся DoxoS-мутантами (MIC 50 мкг/мл). Эти трансформанты показали комплементацию DoxoS -признака. Они могли быть культивированы на доксорубицине при 1500 мкг/мл, что представляет собой уровень устойчивости к доксорубицину, превышающий уровень родительского штамма S.pencetins М76, донора клонированных генов (MIC 250 мкг/мл). Повышенный уровень устойчивости в тансформантах можно объяснить большим числом копий рекомбинантных плазмид (pIJ 101-репликон, Katz et al., 1983).
Рестрикционно-ферментный анализ клонированных фрагментов
Поскольку фенотип, сообщенный двумя клонированными фрагментами, был один и тот же, то проводили исследования для того, чтобы определить: одна или две различные функции способны передавать DoxoR-признак. На рисунках 1 и 2 изображены рестрикционные карты происходящих от S. pencetins М76 вставок FICE 1 и FICE 2. Большая часть каждой карты имеет размеры фрагментов, полученных путем однократного и двойного переваривания с использованием различных комбинаций ферментов. Длины интервалов между соседними сайтами получали путем непосредственного измерения соответствующих фрагментов при двойном и однократном переваривании.
Отсутствие видимого соответствия между картами и двумя клонированными фрагментами позволяет предположить, что устойчивость передается двумя различными генами.
Изобретение относится к биотехнологии. Получены фрагменты геномной ДНК, кодирующие устойчивость к доксорубицину, с установленными картами рестрикции. Фрагменты получены расщеплением ДНК штамма Streptomyces peucetius DSM 4592 или происходящих от него штаммов ферментом MboI. Полученные фрагменты фракционируют до размеров 4 kb - 6 kb, лигируют с линеаризованным вектором pIJ 702. Этой смесью трансформируют протопласты Streptomyces lividans TK 23, чувствительные к концентрации доксорубицина не более 30 мкг/мл. Клоны, устойчивые к доксорубицину, идентифицируют по росту в присутствии доксорубицина и выделяют из них указанный фрагмент. Клонирование биосинтетических генов позволяет изменять пути метаболизма, уровни устойчивости и тем самым повышать продуктивность штамма. 3 с.п.ф-лы, 2 ил.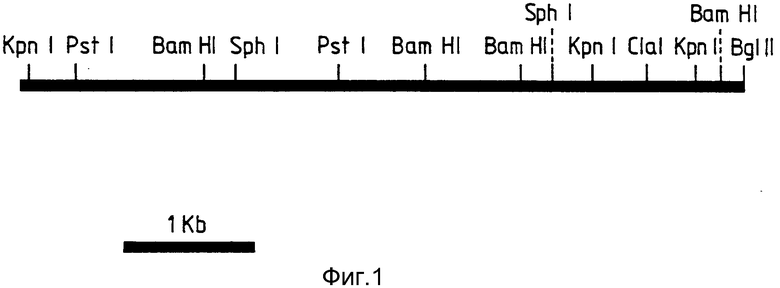
\ \\1 1. Фрагмент геномной ДНК, кодирующий устойчивость к доксорубицину, выделенный из штамма Streptomyces peucetius DSM 4592 или происходящих из него штаммов с картой рестрикации: I \\\6 ЫЫЫ1 \\\2 2. Фрагмент геномной ДНК, кодирующий устойчивость к доксорубицину, выделенный из штамма Streptomyces peucetius DSM 4592 или происходящих от него штаммов с картой рестрикции: II \ \ \6 ЫЫЫ2 \\\2 3. Способ получения фрагмента геномной ДНК, кодирующего устойчивость к доксорубицину, по п.1 или 2, отличающийся тем, что геномную ДНК штамма Streptomyces peucetius DSM 4592 или происходящих от него штаммов частично расщепляют ферментом MboI, полученные фрагменты фракционируют до размеров 4kb - 6kb, лигируют с линеаризованным вектором pIJ 702, полученной смесью трансформируют протопласты Streptomyces lividans TK 23, чувствительные к концентрации доксорубицина не более 30 мкг/мл, клоны, устойчивые к доксорубицину, идентифицируют по росту в присутствии доксорубицина и выделяют из них указанный фрагмент.
| US 3590028, 29.06.71 | |||
| БЕСКОНТАКТНОЕ ПРОМЕЖУТОЧНОЕ РЕЛЕEGZCGIQCHAI^ пл7:;1т;;з -^ т:::::;ч^г:;;пBiijAljDi^KA | 0 |
|
SU173327A1 |

