Изобретение относится к биотехнологии, а именно к новому полипептиду, усиливающему продукцию внеклеточных энзимов, фрагменту ДНК, который кодирует этот полипептид, рекомбинантной плазмидной ДНК, которая содержит данный фрагмент ДНК, и штамму, трансформированному рекомбинантной плазмидной ДНК.
Кроме того, изобретение относится к последовательности промотора гена, кодирующего новый полипептид, экспрессионной системе на основе Streptomyces и способу экспрессии чужеродных последовательностей ДНК в Streptomyces, кодирующих - секретируемые полипептиды и белки.
Виды Streptomyces являются хорошо известными продуцентами разнообразных внеклеточных ферментов, включая протеазы, фосфатазы, ксиланазы, целлюлазы, амилазы, липазы и нуклеазы.
Однако регуляторные механизмы, которые управляют экспрессией этих генов, все еще остаются неизвестными. В дополнение к специфическим регуляторным механизмам, таким как индукция амилазы декстринами или мальтотриозой и углеродно-метаболитное регулирование амилазы или агаразы, вероятно, происходит снятие подавления образования нескольких внеклеточных энзимов, поскольку после снижения питания у Streptomyces наблюдались одновременное продуцирование нескольких энзимов, разлагающих полимерные субстраты. Такие трансактивирующие регуляторные гены были обнаружены у Bacillus subtilis (J. Bacteriol. 169: 324-333, 1987), Bacillus natto (J, Bacteriol. 166: 20-28, 1986), Bacillus Licheniformis.
Позитивные регуляторные гены, воздействующие на синтез энзимов и/или секрецию, могли клонироваться при поиске повышенной секреции внеклеточных энзимов у слабо секретирующих штаммов, как S.Lividans.
Системы для экспрессии чужеродных последовательностей ДНК в Streptomyces ранее были описаны, например, в заявке на Европейский патент 148.552 и заявке VO-88/07079. Эти системы используют эндогенные промоторы внеклеточных энзимов, продуцируемых Streptomyces.
Изобретение, в первую очередь, относится к фрагменту ДНК, кодирующему SAF-полипептид, и SAF-полипептиду, который является фактором активации вторичного метаболизма. SAF-полипептид непосредственно или опосредованно изменяет экспрессию генов внеклеточных энзимов у Streptomyces путем взаимодействия с регулирующей областью структурного гена внеклеточных фрагментов.
Изобретение также относится к рекомбинантным плазмидным ДНК, определяющим экспрессию SAF-полипептида, штамму грибов Streptomyces lividans, содержащему рекомбинантную плазмидную ДНК pULAD3, и способу экспрессии SAF-полипептида.
Фрагмент ДНК, кодирующий SAF-полипептид получают путем молекулярного клонирования из бактерий Streptomyces griseus АТСС 10137; он имеет следующую структуру:
ATG TGC TCG GCC GTC GTC GCG GCG GCC GTG AGC TCG АТС ТСС CGC CGT CGC CGC CGG GCC AGC GCC АСС CGG CGC ТСС GCT GCG GTG AGC CCG CCC CAC ACT CCG TAG CGC TCG GGC TGT ACT AGC GCG TGC ТСС TGG CAC ТСС АСС АТС АСС GGA CAC CGG GCA CAG ACC CGC TTG GCC GCC TCC TCG CGG GCC AGC CGG GCG GCG GTC GGC TCC TTC GAC GGG GCG AAG AAC AGG CCG GCC TCG TCG CGG CGG CAG GCC GCA TCG GAA TGC CAG GGG CCC GCG TCG TCC TCC CGT GCG GGG GTC CGC TGG GCC GGA ACG GCG GCG AAC GGC AGA GGC TGA
Указанный фрагмент ДНК обнаружен у всех исследованных грибов Streptomyces, что указывает на важную роль, которую играет соответствующий ген в метаболизме стрептомицетов. SAF-полипептид содержит 113 аминокислот и имеет следующую структуру:
Met Cys Ser Ala Val Val Ala Ala Ala Val Ser Ser Met Ser Arg Arg Arg Arg Arg Ala Ser Ala Thr Arg Arg Ser Ala Ala Val Ser Pro Pro His Thr Pro Nyr Gly Ser Gly Cys Ile Ser Ala Cys Ser Trp His Ser Thr Ile Thr Gly His Arg Ala Gln Thr Arg Leu Ala Ala Ser Ser Arg Ala Ser Arg Ala Ala Val Gly Ser Phe Asp Gly Ala Lys Asn Arg Pro Ala Ser Ser Arg Arg Gin Ala Ala Ser Glu Cys Gln Gly Pro Ala Ser Ser Ser Arg Ala Gly Val Arg Trp Ala Gly Thr Ala Ala Asn Gly Arg Gly ter
Исследование транскрипции-трансляции in vitro при использовании E.coli показало, что в этом случае образуется белок правильного размера с мол. массой около 15000 дальтон. SAF-полипептид имеет сильный положительный заряд (18 положительно заряженных аминокислот) и не обнаруживает состава, характерного для лидерного пептида или трансмембранного белка и поэтому представляется маловероятным его непосредственное воздействие на секрецию белка. Положительно заряженный белок может легко взаимодействовать с ДНК. Действительно, в SAF-полипептиде обнаружена ДНК-связывающая область, типичная для нескольких регуляторных белков (Ann. Rew. Biochem. 53:293-321, 1984). Неожиданно было обнаружено, что saf-промотор более эффективен, чем природный промотор внеклеточных ферментов. Например, при экспрессии гена амилазы образуется больше амилазы, когда saf-промотор заменяет природный амилазный промотор. Таким образом, saf-промотор может быть использован вместо природного промотора для повышения экспрессии любого эндогенного полипептида или белка.
SAF-полипептид обладает плейстропным действием, т.е. воздействует более чем на один путь синтеза внеклеточных ферментов, а также на образование пигментов и на дифференциацию. В соответствии с изобретением достигается повышенное образование эндогенных (внеклеточных) белков.
Более широко фрагмент ДНК, кодирующий SAF-полипептид (saf-ген), и соответствующий полипептид могут быть использованы для увеличения экспрессии выбранных гетерологичных белков в стрептомицетах путем инсерции гена, кодирующего желаемый белок в подходящий сайт расщепления гена, кодирующего внеклеточный фермент, экспрессия которого усиливается за счет SAF. Затем хозяйские клетки стрептомицетов, содержащие saf-ген, трансформируют рекомбинантным геном и культивируют трансформированные клетки с получением секретируемого продукта. Предпочтительно, гетерологичная ДНК под контролем saf-промотора интегрируется в хромосомную ДНК и совместно с ней интегрируется вектор экспрессии, содержащий ДНК, кодирующую SAF-полипептид. Вектор экспрессии не требуется интегрировать в хромосомную ДНК.
SAF-ген имеет промотор без -10 и -35 консенсусных последовательностей и связан с петлевыми структурами, которые могут функционировать как терминаторы транскрипции. Для экспрессии SAF-полипептида используются различные рекомбинантные плазмидные ДНК. Одна из них содержит фрагмент ДНК плазмиды p1J702, включающий ген устойчивости к тиострептону и ген mel, кодирующий тирозиназу. Другая плазмидная ДНК содержит фрагмент плазмиды p1J699, включающий гены устойчивости к тиострептону, биомицину и канамицину. При этом обе плазмиды содержат последовательность ДНК из Streptomyces griseus, включающий фрагмент ДНК, кодирующий SAF-полипептид, и при необходимости промотор гена saf следующей структуры:
AGATCTCCTCGTCCCACCGGCTGTCGAAGCTCCGCGCCTACAGCGCCATCGACTT CGACCGGGCGAAGTAGGAAGCGGCCGGCGACAAAACGGGGAGGGGCGGGAAACGG TGGTCCGTTTCCCGCCCCTGCCCGTAGGCCGTGCGCGTCCCGCCGGTGCGCGGCG TTCAGCCCGCCGCCGCT
Рекомбинантная плазмидная ДНК pULAD3, определяющая экспрессию SAF-полипептида, содержит следующие конструктивные элементы:
последовательность ДНК из Str.griseus ATCC 10137 размером 1 кб, включающую фрагмент ДНК, кодирующий SAF-полипептид, и промотор гена SAF, структура которых указана выше;
фрагмент ДНК плазмиды p1J702, включающий ген устойчивости к тиострептону, и ген Меl, кодирующий тирозиназу.
Рекомбинантная плазмида ДНК pULAD300, определяющая экспрессию SAF-полипептида, содержит следующие конструктивные элементы:
последовательность ДНК из Str. griseus размером 1 кб, включающую фрагмент ДНК, кодирующий SAF-полипептид, и промотор гена SAF, структура которых указана выше;
фрагмент ДНК плазмиды pUC19, включающий промотор гена lac Z.
Для экспрессии SAF-полипептида клетки микроорганизма трансформируют генетической конструкцией, содержащей фрагмент ДНК, кодирующей SAF-полипептид. Трансформированные клетки выращивают в подходящей питательной среде и выявляют экспрессию SAF-полипептида. В частности, плазмидную ДНК pULAD3 вводят в клетки грибов Streptomyces lividans. Полученный штамм характеризуется культурально-морфологическими, биохимическими и физиологическими признаками, характерными для стрептомицетов, которые относятся к грамположительным, аэробным почвенным грибам.
Изобретение поясняется чертежами, на которых;
фиг. 1 - карты рестрикции фрагментов хромосомной ДНК клонов S. griseus ATCC 10137 по Bg1 П сайту плазмиды p1J702, которая содержит генетическую детерминанту для сверхпродукции щелочной фосфатазы;
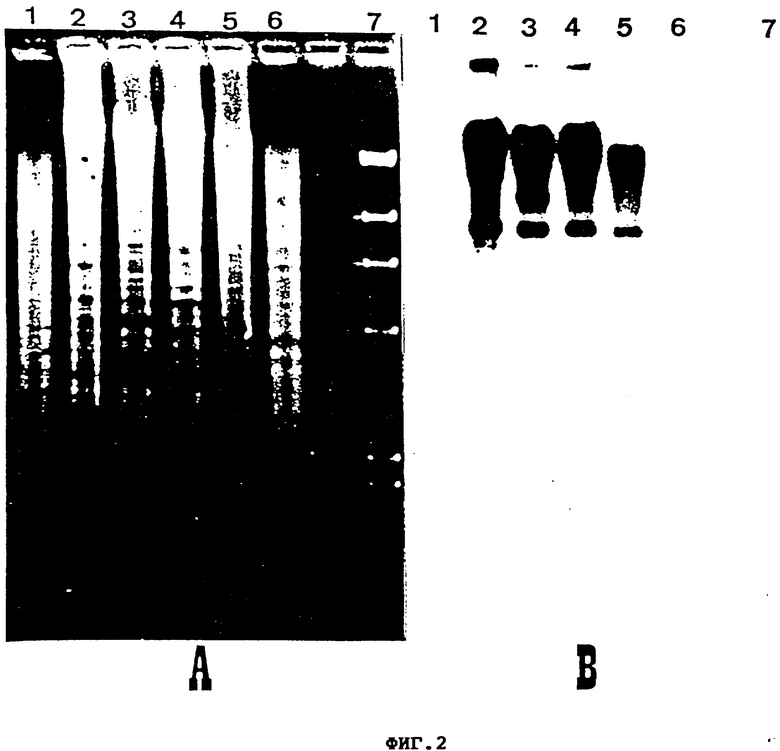
фиг. 2 - результаты анализа (Саузерн-гибридизация) гомологии между 1 кб Bg1 11 фрагментом p1LAD3 и геномной ДНК других видов Streptomyces; (A) фрагменты ДНК, полученные расщеплением BamH1 геномных ДНК (полосы 2 - 7). (B) Гибридизация ДНК, показанных на (A), с фрагментом величиной 1 кб в качестве образца/зонда/. Полосы: l. ДНК, переваренная Hind III с получением фрагментов размерами 23, 9.59, 6.68, 4.29, 2.28, 1.94 и 0.58 кб; 2. S.Lactamdurans; 3. S. acrimycini; 4. S. coeli-cobor 1157; 5. S. griseus 2212; 6. S. griseus 3570; 7. S. Lividans 1326;
фиг. 3 - опыты по испытанию на чашках Петри протеазной (A), амилазной (B) и липазной (C) активности S. Lividans, трансформированных p1J702 (слева) и трансформированных pULAD3 (справа);
фиг. 4 - эффект дозы гена на экспрессию saf-гена. Продуцирование протеазы (А) и амилазы (В) S.Lividans, трансформированными p1J702 (слева), S.Lividans, трансформированными pULAD3 (в середине), и S.Lividans, трансформированными pULAD30 (справа);
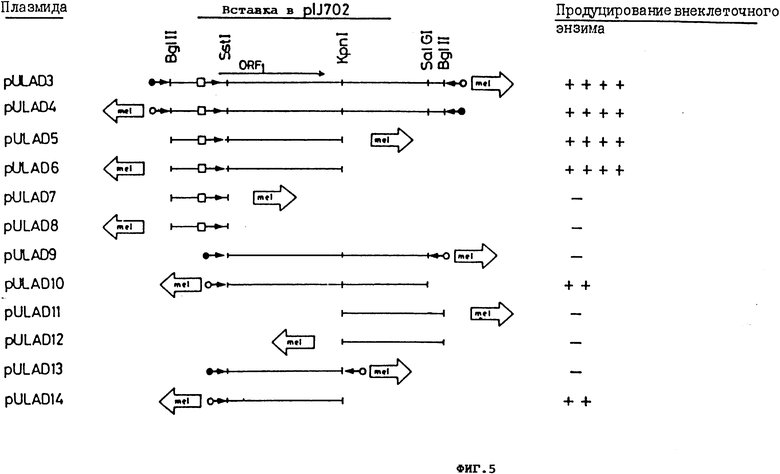
фиг. 5. - "Тримминг" saf-гена.1 кб Bg1 II фрагмента pULAD3 использовался в качестве исходного материала. Образование внеклеточного фермента для всех плазмидных конструкций определялось на чашках с твердой средой, как указано в описании: - означает продуцент дикого типа, два, три или четыре креста указывают различные степени сверхпродукции. Короткие открытые стрелки показывают локализацию и направление транскрипции гена тирозиназы (mel) в составе плазмиды p1J702. Закрытые кружки обозначают промотор mel-гена; открытые (светлые) кружки отражают активность (по часовой стрелке) предполагаемого промотора в составе плазмиды p1J702, а открытые квадраты обозначают saf-промотор. Направление транскрипции показано стрелками. ORF1 соответствует рамке считывания saf, выведенной на основе нуклеотидной последовательности (фиг.6);
фиг. 6 - 9 - нуклеотидная последовательность 1 кб Bg1 II фрагмента pULAD3 и выведенная на ее основе аминокислотная последовательность продукта saf-гена. Подчеркнут второй предполагаемый инициаторный кодон ATG. Инвертированные комплементарные повторяющиеся последовательности показаны сходящимися стрелками. Волнистая линия отмечает аминокислотную последовательность, подобную ДНК-связывающим областям известных ДНК-связывающих белков. Показаны подходящие сайты рестрикции. Нуклеотиды пронумерованы, начиная от сайта Bg1 II (нумерация справа);
фиг. 10 - сравнение участка выведенной аминокислотной последовательности продукта гена saf с ДНК-связывающими областями известных ДНК-связывающих белков. Данные взяты у Pabo и Sаuer, Ann. Rev. Biochem., 53:293-321, 1984 и Horinouchi и Beppu, Genetics of Industrial Microorganisms, p. 41, изд. Плива, Загреб, Югославия, 1986;
фиг. 11 - повторные нуклеотидные последовательности upstream и downstream по отношению к saf-гену;
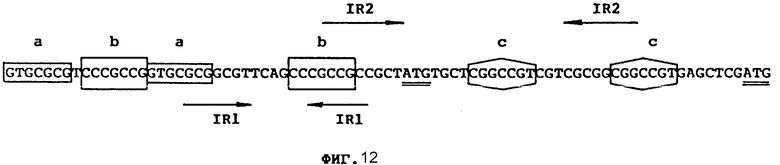
фиг. 12 - повторяющиеся нуклеотидные последовательности гена saf;
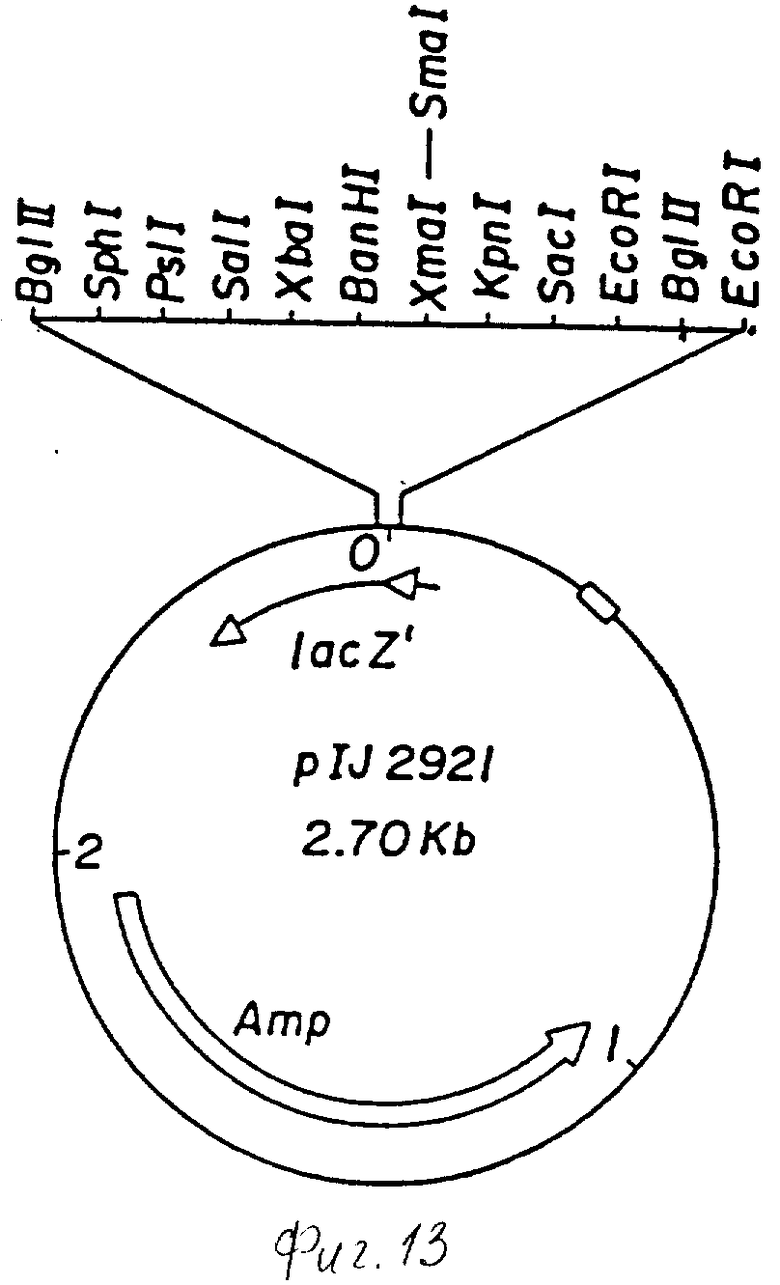
фиг. 13 - схематическое изображение плазмиды plJ2921.
Сведения, подтверждающие возможность осуществления изобретения
Штаммы бактерий и плазмиды. При осуществлении изобретения использовались штаммы Streptomyces и E.coli, которые приведены в Таблице 1. Плазмиды и фаги приведены в Таблице 2.
Среды и условия культивирования. Штаммы Streptomyces выращивали в R2YE, минимальной среде (ММ), TSB (Дифко) или YEME с добавлением 34% сахарозы и 5 мМ MgCl2 (Hopwood и др. Genetic Manipulation of Streptomyces. Лабораторный учебник, изд. John Innes Foundation, Норвич, Великобритания, 1985). Штаммы Streptomyces выращивали в тройных перегороженных флаконах при 28oC в ротационном шейкере при его вращении со скоростью 220 об/ми.
Штаммы E.coli выращивали в бульоне Luria (LB) (см. Миллер и др. EXPERIMENTS in Molecular genetics, стр. 433, Cold Spring Harbor Lab. Нью-Йорк, 1972) или Агаре Luria [LA) при 37oC.
Исследование ферментов на чашках Петри. Исследование Streptomyces на щелочную фосфатазу проводили на среде PRMM (ММ) без глюкозы, содержащей пониженный уровень (1 мМ) фосфотазы, с добавлением 30 мкг/мл 5-бром-4-хлор-3-индолил-фосфат-р-толуидина (ХР). Колонии, продуцировавшие щелочную фосфотазу, были синие, а неспособные к ее образованию - белые. Для E.coli использовали среду, которая содержит (на 1 л): 10 г бактопектона, 12 г основания Трисма; 10 г NaCl: 20 г агара Дифко; pH 7,5 и 40 мкг/мл ХР.
Амилазную активность колоний Streptomyces определяли на среде ММ (без глюкозы) с добавлением 1% крахмала. После 3 дней роста чашки обрабатывали парами иода. Зоны просветления вокруг колоний возникают вследствие разложения крахмала. Липазную активность измеряли путем выращивания Streptomyces в среде ММ с добавлением 2% (вес/об.) оливкового масла, 0,5 (вес./об.) Твина 80 и 0,5% (вес./об>) Твина 20. После 20 дней роста в чашки добавляли по 1 мл 1М CaCl2.
Образование липазы наблюдали в виде преципитата комплекса Ca2+ - жирная кислота (Odera и др.Fermental. Technol., 64:363-371, 1986).
Протеазную активность исследовали в среде ММ (без глюкозы) с добавлением 0,5% казеина и 10 мМ CaCl2. Ферментативную активность определяли как зоны осветления вокруг колоний.
Агарозную активность S. coelicolor определяли в среде ММ без глюкозы с заполнением чашек Грам-иодным раствором. Бета-галактозидазную активность наблюдали в виде синего окрашивания колоний, растущих на ММ без глюкозы с добавлением X-gal (36 мкг/мл) и IРTG (10 мкг/мл).
Выделение ДНК. Общую ДНК из Streptomyces получают, как описано Hopwood и др. , Лабораторный учебник; см. выше). Плазмидную ДНК из Streptomyces или E. coli выделяют по методу Кизера (Kieser и др. Plasmid, 12:19-36, 1984).
Процедуры клонирования. 10 мкг хромосомной ДНК штамма ATCC 10137 S.griseus и 0,5 мкг plJ702 полностью расщепляют Bg1 П и лигируют в течение 12 ч при 14oC с использованием Т4 ДНК-лигазы. Лигационную смесь используют непосредственно для трансформации. Субклонирование фрагментов ДНК проводят путем расщепления 1 - 2 мкг плазмидной ДНК адекватным рестрикционным ферментом (ферментами), а продукты реакции разделяют гель-электрофорезом в агарозе с низкой температурой (LMPA). Требуемые полосы ДНК экстрагируют методом, подразумевающим использование CTAB (Langridge и др., Annal. Biochem., 103: 264-272, 1980).
Методы трансформации. Штаммы Streptomyces трансформируют, как описано Хопвудом и др. в Лабораторном Руководстве (см. выше). После трансформации протопласты помещают на среду R2УЕ и оставляют регенерировать в течение 15 - 20 ч при 30oC. Затем в чашки добавляют 1 мл водного раствора тиострептона (300 мкг/мл), высушивают 1 или 2 ч и инкубируют в течение 2 или 3 дней. Трансформанты переносят на среду RPMM, содержащую тиострептон (50 мкг/мл) и ХР (30 мкг/мл).
Трансформацию E. coli осуществляют согласно Cohen и др. (Proc. Natl. Acad. Sci. США, 69:2110-2114, 1972). Трансформанты подвергают селекции на чашках, содержащих ампициллин (200 мкг/мл). В случае необходимости, в LA чашки добавляют X-gal (36 мкг/мл) и IPTG (10 мкг/мл).
Исследование гибридизации. Перенос ДНК с агарозного геля на нитроцеллюлозные фильтры и гибридизацию проводят, как описано Хопвудом и др. в Лабораторном Руководстве (см.выше). 7,2 кб Bg1 П-фрагмент плазмиды pULAD1 и 1 кб фрагмент Bg1 П плазмиды pULAD3 используют в качестве зондов. Гибридизацию осуществляют при 70oC в течение 24 ч. Фильтры промывают дважды по 30 мин в 2 г SSC, 0,1% SDS и затем еще два раза по 30 мин в 0,2 г SSC и 0,1% SDS при 70oC.
Анализ нуклеотидной последовательности. Нуклеотидную последовательность определяют методом терминации цепи по Сэнгеру (Sanger и др., Proc. Natl. Acad. Sci, США, 74:5463-5467, 1977). Фрагменты ДНК субклонируют в М13мр10 и М13мр11 для получения вставки в любой ориентации. Лигационные смеси трансформируют в компетентные клетки штамма JM103 E.coli и белые (гемолитические) бляшки скринируют для селекции вставок. Осуществляют секвенирование обеих цепей с использованием набора Amersham International plc. (Великобритания) и набора Секвеназа (Биохимическая корпорация Соединенных Штатов). Все фрагменты секвенируют с использованием dGTP, но при необходимости также используют dITP вместо dGTP. Реакционные смеси разделяют в 6%- или 8%-ных полиакриламидных гелях для секвенирования и затем экспонируют на рентгеновской пленке с целью авторадиографии.
Клонирование промоторов. Фрагменты с активностью инициации транскрипции отбирают с использованием мультикопийной плазмиды p1J486 (Ward и др. Mol. Gen. Genet., 203:468-478, 1986) Трансформанты с резистентностью к канамицину (Км) выделяют путем переноса колоний в среду ММ, содержащую 15 мкг/мл Км.
Транскрипция-трансляция in vitro. Плазмиды рULA300 и pUC19 транскрибируют и транслируют с использованием набора лабораторных средств для направленной трансляции прокариотической ДНК (Amersham Int. Plc.) L /35 S/-метионин используют в качестве радиоактивной метки. Для анализа меченых белков используют 12,5% полиакриламидные гели, содержащие додецилсульфат натрия.
Образование щелочной фосфатазы различными Streptomyces. На нескольких твердых средах исследуют продукцию щелочной фосфатазы десятью различными штаммами Streptomyces (см. Табл. 1). S.griseus IMRU3570 и S.griseus ATCC 10137 оказались лучшими продуцентами на всех средах. Лучшей твердой средой для образования щелочной фосфатазы была среда PRMM (содержащая 30 мкг/мл ХР). В этой среде S. griseus IMRU 3570 и S. griseus ATCC 10137 демонстрируют темно-синее окрашивание после 48 ч роста. S.Lividans 1326 и S. Coelicolor J12280 являются продуцентами щелочной фосфотазы: через 20 часов роста на PRMM, содержащей ХР, наблюдается только светло-голубое окрашивание.
Клонирование гена, вовлеченного в продуцирование щелочной фосфотазы. Общую ДНК от штамма ATCC 10137 S.griseus расщепляют Bgl П, лигируют с расщепленной Bg1 П плазмидой p1J702 и лигационную смесь вводят путем трансформации в протопласты S. Lividans 1326. Трансформанты переносят на среду PRMM, содержащую тиострептон (50 мкг/мл) и ХР (30 мкг/мл). Одну темно-синюю колонию обнаруживают среди 2.800 меланин-негативных трансформантов S.Lividans. Эта темно-синяя колония содержит производное p1J702, включающее 7,2 кб Bg1 П вставку и обозначенное pULAD1 (см. фиг. 1).
Плазмида pULAD1- S.Lividans нестабильна и при ретрансформации определяет образование белых и синих колоний. Все синие колонии, содержащие исходную плазмиду pULAD1, и белые колонии, содержащие дефектную форму (с делецией) плазмиды pULAD1, далее не исследуют. Поскольку нестабильность могла быть вызвана большим размером вставки в такой плазмиде, как p1J702, которая известна некоторой нестабильностью, то 7,2 кб вставку субклонируют в 5 кб Bg1 П фрагмент плазмиды p1J699 (Kieser и др.. Gene, 65:83-91, 1988). Получают две плазмиды, pULAD100 и pULAD101 со вставкой, ориентированной в противоположных направлениях. Обе плазмиды стабильны, и ген экспрессируется в S. Lividans в обеих ориентациях.
Локализация saf-гена.
7,2 кб Bg1 П фрагмент частично расщепляют с помощью Sau, ЗAl и лигируют с Bg1 П расщепленной плазмидой p1J702. Лигационной смесью трансформируют протопласты S.Lividans, и трансформанты переносят на среду PRMM, содержащую ХР. Плазмидную ДНК выделяют из нескольких синих колоний. Изучают подробно четыре небольшие плазмиды: pULAD2, pULAD3, pULAD16 и pULAD18, несущие детерминанту образования щелочной фосфатазы; эти плазмиды стабильны при ретрансформации в протопласты S. Lividans и включают вставки размером 2,4, l, 2,1 и 1,9 кб (фиг. 1) соответственно. Плазмида pULAD3 содержит самую маленькую вставку размером 1 кб, которая может быть вырезана с помощью Bg1 П. Будучи введенной в S. Lividans pULAD3 повышает образование щелочной фосфотазы, как и pULAD1. pULAD3 депонирована с регистрационным номером 1-859 19.05.89 в Национальной коллекции Культур Микроорганизмов, согласно Будапештскому Договору и Правилу 28 EPC.
Гибридизация с хромосомной ДНК нескольких Streptomyces. Общую ДНК из штамма ATCC 10137 S.Griseus, расщепленную BamH1 или Bg1 П, гибридизируют с 7,2 кб Вд1 П-фрагментом pULADI, меченным "ник-трансляцией" с (32 р) dCTP. Гомологичный этому зонду 7,2 кб Bg1 П фрагмент обнаруживается в ДНК штамма АТСС 10137 S.Griseus, расщепленный Bg1 П, как и ожидалось. В этом фрагменте находится сайт BamEH1, который определяет образование двух четких полос гибридизации (размером 7,9 кб и 9,4 кб), когда остальную ДНК расщепляют BamH1.
Для выяснения, тот ли же самый ген присутствует в других Streptomyces, общую ДНК: S. acrimycinii, S. coeli-color 1157, S. griseus 2212, S.griseus 3570, S. Lividans 1326 и S. Lactamdurans NRRL 3802 расщепляют BauH1 и гибридизуют c 1 кб Ba1 П фрагментом в качестве зонда. Гибридизацию с 9.5 кб с общей полосой наблюдают у первых четырех pULAD штаммов стрептомицетов (см. выше) тогда, как в ДНК S.Lividans и S. lactamdurans обнаруживают 4 и 5 слабых полос гибридизации, соответственно (фиг. 2). Кроме того, регистрируют гибридизацию ДНК S. albus G, S.coelicolor J12280, S. clavuligerus NRRL 3585 и S. fradiae ATCC 10475. Дальнейшие попытки охарактеризовать полосы гибридизации не предпринимаются.
Некомплементарность pho-мутантов E.coli.
Была сделана попытка установить некомплементарность E. coil phoA-мутантов Е15 и A 1046 при клонировании структурного гена щелочной фосфатазы штамма ATCC 10137 S. griseus. 7.2 кб Bg1 П фрагмент pULAD1 и 1 кб Bg1 П фрагмент pULAD3 субклонировали раздельно в обеих ориентациях в BamH1 расщепленной pUC 19. Все плазмидные конструкции тестировались в клетках E.coli М103 и затем использовались для трансформации E.coli Е15 (Sartly и др., J. Bacteriol, 145: 288-292, 1981) и E.coli AW1046. В В-ХР не было обнаружено никаких синих колоний; это позволяет предположить, что полный структурный ген щелочной фосфатазы не присутствует в 7.2 кб фрагменте S. griseus, или что этот ген не экспрессируется в E.coli. Структурный ген щелочной фосфатазы phoA или Bacillus Licheniformis Hullet, J. Bacteriol, 158:978-982, 1984) не гибридизовался с клонированным 1 кб фрагментом (данные не приведены). В дополнение, phoB (регулярный) E.coli мутант H2 (Kreuzer и др., Genetics, 81: 459-468, 1975) не был комплементирован ни с 1 кб Bg1 П фрагментом pULAD3, ни 7.2 Bg1 П фрагментом pULAD1.
Суперпродукция других внеклеточных энзимов.
Когда pULAD3 используют для ретрансформации протопластов S.Lividans, то наблюдается ясная отложенная (т.е. с задержкой) пигментация и споруляция. Как показано на фиг. 3, некоторые внеклеточные ферменты, включающие амилазу, протеазу и липазу, сверхпродуцируются S.Lividans, трансформированным с помощью pULAD3. Образование бета-галактозидазы также увеличивается, начинаясь примерно на 24 - 30 ч ранее, чем у нетрансформированного S. coelicolor. Благодаря плейотропным эффектам клонированного гена на внеклеточные ферменты, образование пигмента и дифференцирование, этот ген был назван saf (т. е. фактор активации вторичного метаболизма).
Эффект дозы гена. Количество копий plJ702 оценивают как около 100 - 200 на хромосому. Хотя точное количество копий pULAD3 не было определено, интенсивность обеих полос p1J702 и pULAD3 позволяет предположить сходное количество копий.
С целью изучения эффекта дозы гена субклонируют 1 кб BgIII фрагмент pULAD3 по сайту BamH1 плазмиды p1J61 с малым количеством копий (от 3 до 4 копий на клетку) [см. Хопвуд и др., Лабораторное Руководство]. Новую плазмиду обозначают pULAD30. На фиг. 4 показано, что образование внеклеточных ферментов S.Lividans, трансформированным pULAD30, отчетливо понижается по сравнению с S.Lividans, содержащим pULAD3. Кроме того, S.Lividans, включающие pULAD30, продемонстрировали сходный характер образования пигментов и споруляций как и не трансформированные S.Lividans.
Экспрессия в E.coli и тримминг гена.
Хотя pho мутанты E. coli не комплементировались saf-геном, 1 кб Bg1 П вставка pULAD3, будучи субклонирована в одном направлении в pUC19 (плазмида, обозначенная pULAD300) экспрессировалась в E.coli, вызывая измененную морфологию при росте на твердой среде, но не экспрессировалась при встраивании в обратном направлении (плазмида pULAD302). Плазмида pULAD300 содержит ORF (см. ниже) в направлении downstream от lacz промотора. Эти результаты позволяют предположить, что saf-ген экспрессируется в E.colie lacz промотора, входящего в состав pUC19.
Исследования транскрипции-трансляции в E. coli (с плазмидой pULAD300) проводят также in vitro, и продукты реакции наслаивают на 12,5%-ный полиакриламидный гель. Полоса мол. массы 15000 Da присутствует на дорожке, соответствующей pULAD300, и отсутствует в контрольной дорожке pUC19.
Для точной локации saf-гена продолжают расщепление 1 кб фрагмента плазмиды pULAD300. При этом используют уникальные сайты для ферментов рестрикции SstI/nt216/, Кpn I /nt648/ ц SalG1 /nt936/ в пределах 1 кб-фрагмента. Различные субфрагменты на 1 кб вставки pULAD300 клонируют на первой стадии в p1J2921, производное pUC18, содержащее модифицированный полимер, фланкированный Bg1 П сайтами (см. фиг. 13) и затем высвобождают с помощью Bg1 П липкие концы и клонируют в Bg1 П расщепленную плазмиду p1J702. Образование щелочной фосфатазы, амилазы и протеазы изучали у Streptomyces lividans со всеми плазмидными конструкциями. Результаты этих исследований показаны схематично на фиг. 5.
На основании приведенных результатов можно сделать следующие заключения:
1. 1 кб-фрагмент содержит полный saf-ген, включающий свой собственный промотор, поскольку он экспрессируется на сходном уровне в обеих ориентациях плазмиды p1J702 (плазмиды pULAD3 и pULAD4).
2. Указанный saf-ген, вероятно, расположен в 648- нуклеотидном Bg1 П-Кpn1 фрагменте (плазмиды pULAD5 и pULAD6)
3. Фрагмент Sst1-Kpn1 (432 нуклеотида), вероятно, содержит генетическую детерминанту для сверхпродукции внеклеточного фермента, но, по-видимому, утратил область промотора, поскольку экспрессируется только в одной ориентации (плазмида pULAD14 и pULAD10), но не в противоположной (плазмиды pULAD9 и pULAD13).
4. 215-нуклеотидный фрагмент Bg1 П-Sst1 содержит область промотора saf-гена.
5. Все эффекты на внеклеточные ферменты и дифференцировку сохраняются и утрачиваются вместе, указывая на то, что они обусловлены продуктом одного гена.
6. Неожиданным оказалось отсутствие экспрессии фрагмента Sst1-Kpn1 (saf-ген без промотора) с промотора тирозиназы mel-гена, присутствующего в p1J702, который тем не менее экспрессировался в противоположном направлении (по часовой стрелке). Это открытие позволяет предположить, что существует фрагмент с промоторной активностью, расположенный перед mel-геном в Bg1 П клонирующего сайта. Возможность функционирования OPF в обратном направлении (от Kpn1-Sst1) была отброшена по нескольким причинам:
A. Такая ORF не была обнаружена при анализе нуклеотидной последовательности.
B. Если бы такая ORF существовала, то ясно, что она имела бы свой собственный промотор, поскольку фрагмент BgnП-Kpn1 (1 - 648 н.п.) экспрессировался в обоих направлениях (плазмиды pULAD5 и pULAD6). Кроме того, выглядит бессмысленным, чтобы Kpn1-Sst1 фрагмент, который имеет промотор в области Kpn1, мог экспрессироваться только в одном направлении.
С. Экспрессия в E.coli всегда происходит, когда промотор lacZ находится перед предполагаемой ORF, и никогда - при его расположении в обратном направлении.
Промоторная активность ORF1 фрагмента ДНК в положении upstream
Из предыдущих экспериментов можно сделать вывод, что Bg1 П-Sst1 фрагмент содержит ген, который кодирует амино-гликозидфосфотрансферазу (neo), без промотора. Экспрессия этого гена придает S.Lividans резистентность к канамицину и неомицину. Когда этот фрагмент субклонируют в plJ486 (с Sst1 концом, ближайшим к neo-гену, то получают плазмиду p1J484:216. Наблюдают, что S.Lividans трансформированный p1J486:216, растет в ММ с содержанием канамицина более 100 мкг/мл, тогда как S.Lividans ) трансфорованный p1J486, не растет на ММ с добавлением канамицина в количестве 5 мг/мл. Эти результаты показывают, что указанный фрагмент имеет промоторную активность, и подтверждают существование предполагаемой ORF.
Другие заключения, которые можно сделать на основе нуклеотидной последовательности вероятной ORF, включают следующее:
1. Ген saf окружен двумя областями с инвертированными и комплементарными повторными последовательностями, которые могут образовывать в мРНК очень стабильные связи G = -38,4 Ккал (нуклеотиды t 637-697) [фиг. 8].
2. Секвенированный фрагмент содержит высокий G + С процент, что характерно для генов Streptomyces.
3. Перед геном saf между ATC-(нт 219) и повторяющимися структурами в положении upstream имеется очень интересная нуклеотидная область: в ней обнаружены 3 пары повторяющихся нуклеотидов (каждый составляет 7 нт) [фиг. 12]. Весьма вероятно, что эта область, особенно промоторная зона, играет очень важную роль в регуляции экспрессии гена saf.
4. Следует упомянуть о том, что SAF-полипептид содержит 18 суммарных положительных зарядов. Исходя из теоретических положений, это предполагает существование значительно большей аффинности к ДНК. Даже в выведенной аминокислотной последовательности можно было наблюдать область, сходную с областями ДНК-связывающих белков (Pabo и Sauer,1884) [фиг. 10].
Нуклеотидная последовательность saf-гена.
Нуклеотидную последовательность целой 1 кб Bg1 П вставки pULAD3 определяют с использованием плазмиды pULAD300 в качестве исходного материала. Поскольку М13mp10 и М13mp11 не имеют Kpn1 сайта, фрагменты с Kpn1-концами сначала субклонируют в pUCl9 и затем высвобождают в виде EcoR1-Hind III фрагментов и вводят в мр10 и мр11, расщепленные EcoRI и Hind III. Полная нуклеотидная последовательность активной области показывает наличие ORF1 из 339 нуклеотидов, которые кодируют предполагаемый SAF-полипептид (фиг. 6 - 9). При этом имеется только одна возможная открытая рамка считывания, содержащая Bg1 П-Kpn1 фрагмент, начинаясь от ATG-кодона инициации у нуклеотида 183 и заканчиваясь TGA стоп-кодоном у нуклеотида 524 (ORF1 на фиг. 5 - 9. Поскольку Sst1-Kpn1 вставка, содержащаяся в плазмиде pULAD14, демонстрирует более низкую степень активности, чем плазмиды, также содержащие область Sst1 сайта в положении upstream (pULAD3, pULAD4, pULAD5 и pULAD6), очень вероятно, что ATG-(нуклеотид 219) в пределах предполагаемой рамки считывания saf может функционировать как кодон инициации в pULAD10 и pULAD14. Никакие другие альтернативные триплеты инициации невозможны, поскольку TGA кодон терминации обнаружен в положении upstream по отношению к первому ATG. Длинная инвертированная повторяющаяся область, которая может образовывать очень стабильную стеблевую и петлевую структуру (G = - 53.6 Ккал), присутствует в положении upstream по отношению к saf-гену, от нт 90 до нт 135. Другая инвертированная повторяющаяся последовательность обнаруживается в положении downstream терминаторного триплета ORF1, простираясь от нт 637 до 697 (G = -38,4 Ккал).
Ген saf имеет высокое (G + С содержание (76,3%).
Описание раскрывает общую сущность изобретения; при этом любые возможные модификации не выходят за рамки изобретения. Ниже приведены штаммы и плазмиды, используемые для осуществления изобретения (табл. 1 и 2).










При осуществлении изобретения может быть получен фрагмент ДНК, кодирующий SAF-полипептид, являющийся фактором активации вторичного метаболизма. Указанный фрагмент получают методом молекулярного клонирования из клеток Streptomyces griseus АТСС 10137. Данный фрагмент входит в состав рекомбинантных плазмидных ДНК, обеспечивающих экспрессию SAF-полипептида. Кроме того, данным фрагментом трансформируют клетки микроорганизма, которые затем культивируют в питательной среде и выявляют экспрессию SAF-полипептида. Одна из указанных выше плазмид входит в состав штамма Streptomyces lividans. Данный штамм характеризуется признаками, типичными для стрептомицетов. 8 с. п. ф-лы, 13 ил.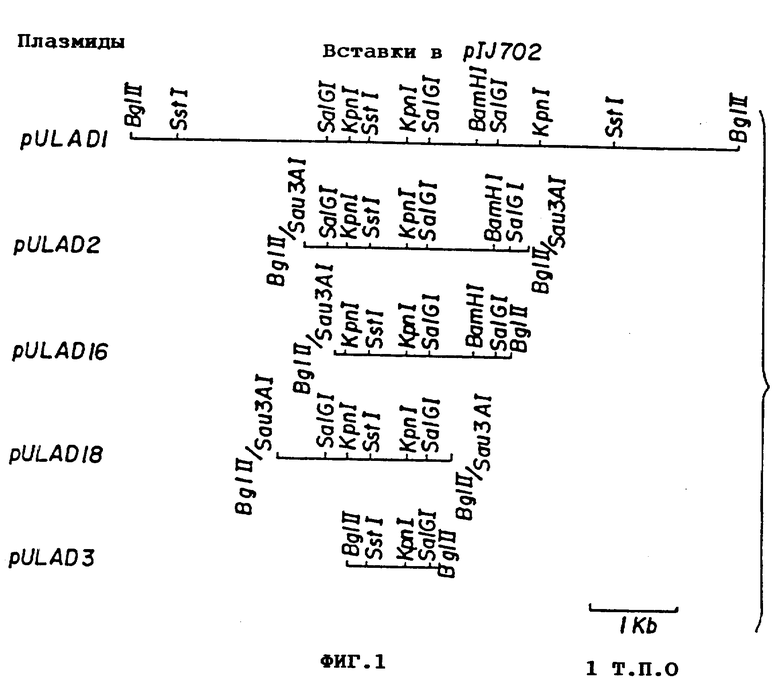
2. SAF-полипептид мол.м. около 15000 Да, кодируемый фрагментом ДНК, охарактеризованным в п.1 формулы, обладающий способностью активировать образование вторичных метаболитов стрептомицетов и имеющий последовательность аминокислот, установленную на основании последовательности нуклеотидов фрагмента ДНК, охарактеризованного в п.1 формулы, и представленную на с.
последовательность ДНК из Streptomyces griseus, включающую фрагмент ДНК, охарактеризованный в п. 1 формулы, и при необходимости промотор гена SAF структуры, представленной на с.432,
фрагмент ДНК плазмиды pIJ 702, включающий ген резистентности к тиострептону и ген Mel, кодирующий тирозиназу.
последовательность ДНК из Streptomyces griseus АТСС 10137 размером 1 кб, включающую фрагмент ДНК, охарактеризованный в п.1 формулы, и промотор гена SAF, представленную на с.432,
фрагмент ДНК плазмиды pIJ 702, включающий ген резистентности к тиострептону и ген Mel, кодирующий тирозиназу.
последовательность ДНК из Streptomyces griseus, включающую фрагмент ДНК, охарактеризованный в п. 1 формулы, и при необходимости промотор гена SAF, структуры, представленной на с.432,
фрагмент ДНК плазмиды pIJ 699, включающий гены резистентности к тиоострептону, биомицину и канамицину.
последовательность ДНК из Streptomyces griseus размером 1 кб, включающую фрагмент ДНК, охарактеризованный в п.1 формулы, и промотор гена SAF структуры, представленную на с.432,
фрагмент ДНК плазмиды pUC 19, включающий промотор гена lac Z.

