Изобретения относятся к области медицины и биотехнологии.
Объектами применения изобретений могут быть клетки любого рода организмов (в том числе и одноклеточных), культуры клеток и тканей, а также многоклеточные организмы в целом.
Указанные выше изобретения могут быть использованы, в частности:
- в медицине, для коррекции нарушений энергетического метаболизма клеток, клеточных систем, тканей, органов и многоклеточных организмов в целом;
- в биологии, для исследования особенностей энергетики клеточных систем, например, для синхронизации клеточной культуры (получения популяции микроорганизмов, находящихся в одной и той же фазе развития);
- в генной инженерии, для увеличения выхода синтезируемых микроорганизмами белков.
Известны различные способы регулирования внутриклеточного энергопроизводства клеток с помощью химических и биохимических средств: лекарств и препаратов, естественных (гормонов) или синтезированных. Одни из них усиливают энергопроизводство клеток, другие снижают. В частности, цитрат (лимонная кислота) ингибирует гликолиз. Добавление цитрата в питательный раствор, содержащий бактериальную культуру, снижает энергопроизводство этих клеток.
Большинство широко известных антибиотиков: пенициллин, стрептомицин, тетрациклин и др. также угнетают энергопроизводство клеток, препятствуя размножению болезнетворных микроорганизмов.
Недостатками известных способов являются высокая степень опасности ошибок дозирования, побочные вредные воздействия и их последствия, трудности введения в организм и, особенно, выведения.
В частности, известен способ регулирования энергопроизводства клеток, заключающийся в воздействии антибиотиками на клеточные системы или многоклеточные организмы в целом. Эти вещества понижают энергопроизводство всех клеток, подверженных их воздействию, включая как болезнетворные микроорганизмы и клетки воспаленной ткани, так и клетки здоровых тканей. При достаточном количестве введенного в организм антибиотика энергопроизводство снижается, размножение болезнетворных микроорганизмов прекращается и иммунная система организма их уничтожает. Негативным фактором при этом является угнетение по той же причине жизнедеятельности здоровых клеток, наблюдающееся до полного выведения антибиотиков из организма даже после уничтожения патогенных очагов. Вредные последствия наблюдаются особенно при передозировке антибиотика.
Таким образом, недостатком этого способа является принципиально неустранимое остаточное последействие антибиотиков на здоровые клетки даже после ликвидации имунной системой патогенных очагов, и связанное с этим стрессорное состояние организма, особенно выраженное при передозировке.
Наиболее близкими к заявляемому способу являются способы регулирования внутриклеточного энергопроизводства, заключающиеся в размещении живых организмов или отдельных клеток в герметичной камере, заполненной средой, содержащей кислород и другие средства жизнеобеспечения, и повышении давления в заполняющей камеру среде до заданного уровня (см., например, В.И. Бураковский и Л. А. Бокерия "Гипербарическая оксигенация в сердечно-сосудистой хирургии". - М. : Медицина, 1974, с. 81 - 114). В этом способе регулирование внутриклеточного энергопроизводства осуществляют за счет изменения содержания газов - энергетических субстратов и метаболитов в среде, окружающей живые организмы или отдельные клетки, или в дыхательной смеси.
Основным фактором, определяющим интенсивность внутриклеточного энергопроизводства в известном способе, является повышенное или пониженное в зависимости от цели (по сравнению с атмосферными условиями) содержание кислорода в дыхательной смеси.
Общеизвестно применение кислорода для дыхания ослабленных или потерявших сознание больных. Для этого используют кислородные подушки - резиновые емкости с патрубками, через которые кислород подается из подушки к пациенту.
Повышение концентрации кислорода в дыхательной смеси вызывает увеличение насыщенности кислородом межклеточной жидкости и цитоплазмы клеток. Повышенное содержание кислорода (гипербарическая оксигенация-ГБО) по сравнению с нормой влечет за собой увеличение скорости окисления пищевых субстратов в клетках и увеличение энергопроизводства. Кислород внутрь клеток поступает диффузионным путем.
По закону Фика его поток Q записывается выражением: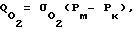
где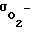 проницаемость клеточных мембран для кислорода;
проницаемость клеточных мембран для кислорода;
Pм - напряжение кислорода в межклеточной жидкости;
Pк - напряжение кислорода в цитоплазме.
В приведенных способах-прототипах пытаются увеличить градиент (Pм - Pк) за счет увеличения величины Pм, зная, что Pк гомеостатируется и удерживается в пределах 0,5 - 2 мм рт.ст.
Установлено, что Pк гомеостатируется за счет изменения величины проницаемости 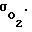 Пpи увеличении Pм величина
Пpи увеличении Pм величина 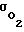 автоматически снижается и в некотором (физиологическом) диапазоне изменения Pм поток кислорода
автоматически снижается и в некотором (физиологическом) диапазоне изменения Pм поток кислорода 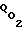 и напряжение кислорода Pк изменяется столь незначительно, что их можно считать неизменными.
и напряжение кислорода Pк изменяется столь незначительно, что их можно считать неизменными.
Только после исчерпания всего адаптационного резерва величина 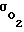 перестает изменяться (уменьшаться) и при дальнейшем увеличении Pм поток
перестает изменяться (уменьшаться) и при дальнейшем увеличении Pм поток 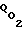 начинает существенно возрастать. Но при выходе Pм за этот физиологический диапазон, при котором возможна адаптация клетки к росту Pм и удержанию Pк на прежнем уровне, существенно возрастает перекисное окисление липидов наружного слоя мембраны. По этой причине при длительном превышении величиной Pм физиологического уровня (≈ 100 мм рт.ст.) возможна кислородная интоксикация клеток.
начинает существенно возрастать. Но при выходе Pм за этот физиологический диапазон, при котором возможна адаптация клетки к росту Pм и удержанию Pк на прежнем уровне, существенно возрастает перекисное окисление липидов наружного слоя мембраны. По этой причине при длительном превышении величиной Pм физиологического уровня (≈ 100 мм рт.ст.) возможна кислородная интоксикация клеток.
При регулировании энергетики клеток посредством дыхания гипоксическими смесями газов (смеси с пониженным содержанием кислорода, ниже, чем в атмосферном воздухе) понижение интенсивности энергопроизводства клеток происходит за счет уменьшения концентрации кислорода в межклеточной жидкости.
Здесь имеет место также неустранимый недостаток - опасность гибели клеток из-за недостатка кислорода. При уменьшении Pм срабатывает адаптационный механизм, клетки приспосабливаются к пониженному содержанию кислорода в окружающей среде за счет увеличения 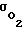 Эволюция обусловила появление эффективного адаптационного механизма с широким диапазоном изменения величины
Эволюция обусловила появление эффективного адаптационного механизма с широким диапазоном изменения величины 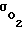 (на порядки). Вблизи капилляров Pм ≈ 100 мм рт.ст., а в так называемом летальном Кроговском углу - в наиболее удаленном от капилляров месте оно может составлять всего 10 - 20 мм рт.ст. и меньше.
(на порядки). Вблизи капилляров Pм ≈ 100 мм рт.ст., а в так называемом летальном Кроговском углу - в наиболее удаленном от капилляров месте оно может составлять всего 10 - 20 мм рт.ст. и меньше.
Однако интенсивность энергопроизводства у всех клеток определенного типа тканей одинакова. Работают клетки синхронно, в одинаковом режиме. Для того чтобы данный способ сработал, необходимо настолько уменьшить содержание кислорода в дыхательной смеси, насколько это необходимо для выхода из адаптационного диапазона. Это чревато глубокой гипоксией клеток в летальном углу и их гибелью.
Как отмечалось, регулирование связано с опасностью гибели клеток при повышенном Pк из-за перекисного окисления липидов мембраны, а при пониженном Pк - из-за недостатка кислорода и, следовательно, минимального достаточного для поддержания жизни клетки энергопроизводства. Кроме того, регулировочные характеристики (зависимости интенсивностей энергопроизводства от величины Pм) для разных типов клеток различны, что связано с различием их адаптационных возможностей; регулировочные характеристики существенно нелинейны: в физиологическом (адаптационном) диапазоне поток кислорода 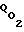 и, следовательно, интенсивность энергопроизводства весьма слабо зависят от Pм, медленно возрастая с ее ростом. За границей указанного диапазона даже незначительное изменение величины Pм может привести к катастрофическим последствиям и к гибели клеток как при больших величинах Pм (из-за перекисного окисления липидов), так и при низких (из-за нехватки энергии для поддержания жизни).
и, следовательно, интенсивность энергопроизводства весьма слабо зависят от Pм, медленно возрастая с ее ростом. За границей указанного диапазона даже незначительное изменение величины Pм может привести к катастрофическим последствиям и к гибели клеток как при больших величинах Pм (из-за перекисного окисления липидов), так и при низких (из-за нехватки энергии для поддержания жизни).
Общим недостатком известного способа также является его непрямое действие, т.е. воздействие не непосредственно на клетки, а через дыхательные пути и межклеточную жидкость. Это, во-первых, снижает их эффективность в применении к высокоорганизованным организмам. Приспособительные реакции уже при транспортировке газов кровью от легких к клеткам-потребителям и наоборот сводят к минимуму полезный результат воздействия; во-вторых, приводит к неоднородному, из-за особенностей строения кровеносной системы, в основном, сети ее капилляров, воздействию на одинаковые клетки, например, при дыхании гипоксическими смесями часть клеток, которые наиболее удалены от капилляров, будут уже испытывать кислородную недостаточность, в то время как другая их часть еще способна адаптироваться к новым условиям и продолжает нормально функционировать.
Таким образом, собственно регулирование внутриклеточного энергопроизводства по известным способам: сжатым воздухом, гипербарической оксигенацией и гипоксическими смесями возможно только в узких пределах (интервалах) на границах физиологического диапазона, внутри которого срабатывает клеточный адаптационный механизм поддержания постоянным величины Pк за счет изменения 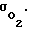
Известны традиционные способы лечения онкологических заболеваний, основанные на химико-терапевтическом и радиобиологическом воздействии, заключающиеся в ведении в организм или непосредственно в опухоль алкирующих агентов, антиметаболитов, антибиотиков и других биологически активных веществ, которые, угнетая клеточное энергопроизводство и прочие жизненно важные процессы, замедляют рост новообразований и вызывают гибель злокачественных клеток, способствуя выздоровлению (см. Л.Ф. Ларионов. Химиотерапия злокачественных опухолей. М., 1962; или М.М. Маевский (под ред.) Противоопухолевые антибиотики. М., 1962).
Неустранимым недостатком известных способов является вредное побочное воздействие на клетки здоровых тканей и возможность их гибели.
Наиболее близким к этому заявляемому изобретению является способ лечения онкологических заболеваний, заключающийся в размещении живых организмов в герметичной камере, заполненной средой, содержащей кислород и другие средства жизнеобеспечения, и повышении давления в заполняющей камеру среде до заданного уровня, причем повышение давления осуществляют сжатым воздухом или дополнительно закачиваемым в камеру кислородом (В.И. Бураковский и Л.А. Бокерия. Гипербарическая оксигенация в сердечно-сосудистой хирургии. - М.: Медицина, 1974, с. 81 - 114).
Недостатком этого способа является его низкая эффективность, обусловленная его неустранимой противоречивостью: использование для лечения сжатого воздуха приводит, с одной стороны, к увеличению напряжения кислорода в цитоплазме и, следовательно, к увеличению энергопроизводства клеток, что способствует их делению, но с другой стороны, повышенное давление уменьшает поток кислорода в клетки, понижая энергопроизводство и препятствуя делению клеток. Одно компенсирует другое и эффекта нет.
Традиционные способы предотвращения бактериального разложения пищевых продуктов при их хранении связаны с добавлением к ним химических веществ, угнетающих жизнедеятельность микроорганизмов, с охлаждением, пастеризацией с последующим охлаждением и помещением в бескислородную среду.
Недостатком способов, связанных с добавлением в продукты химических веществ, угнетающих жизнедеятельность микробов, является вредное воздействие таких веществ на организм после потребления пищевых продуктов.
Недостатком способов, использующих охлаждение, является температурная деструкция пищевых продуктов и частичная потеря товарных качеств, присущих свежим продуктам, не испытавшим охлаждения.
Наиболее близким к заявляемому изобретению является способ предотвращения бактериального разложения пищевых продуктов, заключающийся в помещении их в герметичную камеру, заполненную средой, с минимально возможным содержанием кислорода. Из-за недостатка кислорода большинство микроорганизмов, вызывающих бактериальное разложение, не могут размножаться, что и обеспечивает возможность хранения продуктов.
Недостатком этого известного способа является неконтролируемое размножение анаэробных бактерий. Их вредное действие сокращает срок хранения продуктов.
Известны способы предотвращения клеточного деления при использовании микроорганизмов при производстве белка, заключающиеся в том, что в питательный раствор, в котором культивируются бактерии, после достаточного их размножения добавляют вещества, угнетающие их энергопроизводство и препятствующие их дальнейшему размножению (об этих веществах уже говорилось выше). Энергопроизводство клеток устанавливают на уровне, достаточном для производства товарного белка, но недостаточном для клеточного деления. Известный способ существенно повышает эффективность затрат, снижая себестоимость конечного продукта.
Недостатком известного способа является опасность передозировки препарата, когда энергопроизводство клеток угнетается настолько, что существенно снижается скорость синтеза бактериями белкового продукта и, соответственно, эффективность производства.
Известны медикаментозные терапевтические способы лечения сахарного диабета, заключающиеся в том, что с помощью медикаментов стараются скорректировать развивающуюся патологию клеток органов, пораженных диабетическим процессом, в том числе и клеток поджелудочной железы.
Недостатком этих известных способов является неизбежное вредное воздействие на здоровые клетки.
Наиболее близким к заявляемому изобретению является способ, заключающийся в размещении больных организмов в герметичной камере, заполненной средой, содержащей кислород и другие средства жизнеобеспечения, и повышении давления в заполняющей камеру среде до заданного уровня за счет нагнетания в камеру сжатого воздуха или кислорода (см. В.И. Бураковский и Л.А. Бокерия "Гипербарическая оксигенация в сердечно-сосудистой хирургии". - М.: Медицина, 1974, с. 81 - 114).
Недостаток известного способа заключается в его недостаточной эффективности. Предполагалось, что основным действующим фактором является повышенное по сравнению с атмосферой содержание кислорода в дыхательной смеси, вызывающее интенсификацию окислительных процессов в клетке и рост клеточного энергопроизводства. Это должно было благотворно сказаться на функционировании клеток и улучшении самочувствия больного. Но этого на практике не наблюдается по причинам, приведенным выше: срабатывает адаптационный механизм поддержания постоянным напряжения кислорода в цитоплазме, и заметного усиления энергопроизводства клеток не наблюдается.
Использование заявленного в качестве изобретения способа регулирования внутриклеточного энергопроизводства обеспечивает возможность получения следующего "технического" результата:
- расширение диапазона регулирования внутриклеточного энергопроизводства;
- обеспечение одинакового (однородного) воздействия на клетки избранной системы (образований) в организме;
- обеспечение непосредственного воздействия на те или иные клетки организма;
- исключение последействия и вредных побочных воздействий на клетки;
- закрепление полезного эффекта воздействия на клетки.
Указанный технический результат обеспечивается при использовании способа регулирования внутриклеточного энергопроизводства, заключающегося в размещении живых организмов или отдельных клеток в герметичной камере, заполненной средой, содержащей кислород и другие средства жизнеобеспечения, и изменении давления в заполняющей камеру среде до заданного уровня, отличающийся тем, что повышение или понижение давления в заполняющей камеру среде, осуществляют путем по меньшей мере одноразового увеличения или уменьшения содержания в ней биологически неактивных составляющих, при этом предельный заданный уровень давления среды в камере устанавливают по достижении требуемой интенсивности внутриклеточного энергопроизводства, а поддержание в камере заданного уровня давления среды производят в течение времени, необходимого для закрепления в организмах или отдельных клетках требуемой интенсивности внутриклеточного энергопроизводства.
Следует особе отметить, что снижение интенсивности энергопроизводства клеток достигается за счет того, что по мере роста давления в среде, окружающей клетки, проницаемость их липидных мембран для мелких молекул и ионов уменьшается, ограничивая потоки внутрь клеток таких важнейших субстратов энергетического метаболизма, как кислород и (или) глюкоза. Это немедленно сказывается на интенсивности энергопроизводства клеток, понижая ее.
Очевидно, что ограничение потоков в клетку кислорода и(или) глюкозы, приведет к уменьшению интенсивности ее энергопроизводства. Наоборот, по мере уменьшения давления в среде, окружающей клетки (при неизменном парциальном давлении кислорода в этой среде), проницаемость их липидных мембран для мелких молекул и ионов увеличивается, что приводит к увеличению потоков внутрь клеток субстратов энергетического метаболизма (глюкозы и кислорода) и увеличению интенсивности энергетического метаболизма.
Таким образом, варьирование величиной общего давления в среде, окружающей клеточные системы, осуществляемое за счет нагнетания в камеру биологически инертных газов, обеспечивает расширение диапазона регулирования внутриклеточного энергопроизводства, которое можно реализовать известными способами повышением или понижением содержания только кислорода в дыхательной смеси.
Давление, поддерживаемое в камере, действует на все клетки организмов, помещенных в нее, непосредственно и одинаково.
Его действие прекращается немедленно после установления в камере атмосферного давления. В организм никаких посторонних химических веществ не вводится, поэтому в клетках и окружающих клеточные системы среде их не остается. Последействие не может иметь место (если приняты известные меры, исключающие кессонные явления), так как воздействие давления немедленно прекращается после его снятия.
Всегда можно выбрать такое парциальное давление кислорода в дыхательной смеси, которое исключает вредное побочное воздействие на клетки, такое как перекисное окисление липидов мембран при повышенном 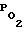 и гипоксия клеток при пониженном.
и гипоксия клеток при пониженном.
Ничто не мешает воздействовать давлением на клетки столько времени, сколько требуется для срабатывания адаптационного механизма, наработки в клетках необходимых белков и закрепления полезного эффекта. Ориентировочно, это занимает не менее 2-4 дней.
Опасность передозировки воздействия также исключается, так как неспецифическая проницаемость клеточных мембран определяется двумя взаимно независимыми факторами прохождения части сквозь мембрану: во-первых, по сквозным порам в ней и, во-вторых, непосредственно сквозь липиды, растворяясь в них.
Давление действует на составляющую проницаемости, зависящую только от первого фактора, то есть от количества и размеров сквозных пор в мембране. Даже когда сквозных пор в ней нет, вторая составляющая проницаемости обеспечивает тот минимальный поток субстратов энергетического метаболизма, который достаточен для поддержания жизнедеятельности клеток.
Использование заявляемого изобретения (способа регулирования внутриклеточного энергопроизводства) в качестве способа лечения онкологических заболеваний обеспечивает возможность получения следующего "технического" результата:
- исключение вредного побочного воздействия на клетки здоровых тканей и возможности их гибели;
- повышение эффективности лечения (по сравнению с прототипом).
Указанный технический результат обеспечивается при использовании способа регулирования внутриклеточного энергопроизводства в качестве способа лечения онкологических заболеваний, заключающегося в размещении больных организмов в герметичной камере, заполненной средой, содержащей кислород и другие средства жизнеобеспечения, и изменении давления в заполняющей камеру среде до заданного уровня, причем повышение или понижение давления в заполняющей камеру среде, осуществляют путем по меньшей мере одноразового увеличения или уменьшения содержания в ней биологически неактивных составляющих, при этом предельный заданный уровень давления среды в камере устанавливают по достижении требуемой интенсивности внутриклеточного энергопроизводства, а поддержание в камере заданного уровня давления среды производят в течение времени, необходимого для закрепления в злокачественных клетках требуемой интенсивности внутриклеточного энергопроизводства.
Как уже отмечалось выше, изменение давления в окружающей организмы среде влияет только на размеры и количество сквозных пор в липидных мембранах клеток, т. е. только на одну из составляющих неспецифической проницаемости мембран. При парциальном давлении кислорода в дыхательной смеси, близком к атмосферному, как того требует заявляемый способ, другая составляющая проницаемости, связанная с растворением молекул в липидах мембраны, обеспечивает потоки энергетических субстратов, в том числе и кислорода, достаточные для жизнедеятельности клеток здоровых тканей. Поэтому вредное воздействие и возможность гибели здоровых клеток исключается.
Однако энергопроизводство как злокачественных, так и здоровых клеток в условиях предлагаемого способа можно настолько понизить, повышая давление в среде, окружающей организм, что энергии не будет хватать для деления клеток, что и обеспечивает лечебный эффект.
В предлагаемом способе лечения онкологических заболеваний в отличие от прототипа нет "противоречий". Все операции заявляемого способа направлены на ограничение потоков в клетки важнейших энергетических субстратов и поддержании энергопроизводства клеток на возможно низком уровне, что повышает эффективность лечения.
Использование заявляемого изобретения в качестве способа предотвращения бактерийного разложения пищевых продуктов при их хранении обеспечивает возможность получения следующего "технического" результата:
- отказ от применения химических веществ, угнетающих жизнедеятельность микроорганизмов, и исключение таким образом вредного воздействия таких веществ на организм после потребления пищевых продуктов;
- отказ от применения охлаждения и, следовательно, исключение температурной деструкции пищевых продуктов и частичной потери товарных качеств, присущих свежим продуктам, не испытавшим охлаждения;
- угнетение размножения в пищевых продуктах анаэробных бактерий и продление сроков хранения пищевых продуктов.
Следует подчеркнуть, что при повышении давления ограничиваются потоки внутрь клеток всех мелких молекул, способных проходить сквозь мембрану по сквозным порам в ней. Это относится не только к молекулам кислорода, но и сахарам, которые необходимы для поддержания жизнедеятельности анаэробных микроорганизмов. Поэтому при повышении давления уменьшаются потоки сахаров внутрь анаэробных клеток и угнетается их жизнедеятельность, что обеспечивает указанный "технический" результат.
Использование заявляемого изобретения в качестве способа предотвращения клеточного деления при производстве микроорганизмами белков обеспечивает возможность получения следующего "технического" результата:
- отказ от применения препаратов, препятствующих размножению микроорганизмов, и исключение ошибок дозирования, снижающих эффективность производства;
- повышение технологичности производства.
Последнее связано с особенностями воздействия давления. Оно действует как в среде, окружающей организмы, так и внутри подвергаемых воздействию давления клеток одинаково и равномерно. Исключается необходимость перемешивать растворы, в которых содержатся микроорганизмы, для улучшения воздействия.
Использование заявляемого изобретения в качестве способа лечения сахарного диабета обеспечивает возможность получения следующего "технического" результата:
- отказ от медикаментов, оказывающих вредное воздействие на здоровые клетки;
- повышение, по сравнению с прототипом, эффективности лечения за счет корректировки энергетического метаболизма клеток, захваченных диабетическим процессом, т.е. перевод их энергопроизводства с усвоения преимущественно углеводов на усвоение жиров и белков, осуществляемый по предлагаемому способу за счет ограничения потоков глюкозы в клетки при избыточном давлении в камере. Это позволяет отказаться от медикаментов того же назначения, угнетающих углеводный путь клеточного производства энергии и исключить вредное побочное воздействие таких препаратов на здоровые клетки.
Повышенная по сравнению с прототипом эффективность лечения может быть достигнута благодаря более широким возможностям регулирования потоков кислорода и сахаров внутрь больных клеток, предоставляемых предлагаемым способом. Повышение давления уменьшает потоки сахаров в клетки, а его уменьшение по сравнению с атмосферным, наоборот, увеличивает эти потоки. В зависимости от фазы развития диабетического процесса и самочувствия больного может быть показано то или другое.
Повышение давления в окружающей живой организм среде за счет увеличения или уменьшения в ней биологически неактивных составляющих является основным фактором, определяющим вышеуказанные полезные свойства предлагаемого способа, обусловленные уменьшением или увеличением неспецифической проницаемости цитоплазматических мембран и, следовательно, ограничением или, соответственно, увеличением потоков сквозь них мелких молекул метаболитов: глюкозы, углекислого газа, кислорода, а также других физиологически активных газов и субстратов.
На фиг. 1, 2, 3, 4 и 5 представлены графические материалы, поясняющие сущность изобретения.
На фиг. 1 - кривая распределения по размерам R сквозных пор в мембранах клеток. Это - зависимость плотности вероятности 1/N•dN/dt обнаружить среди всех N сквозных пор пору с поперечным размером R. Площадь под кривой равна 1.
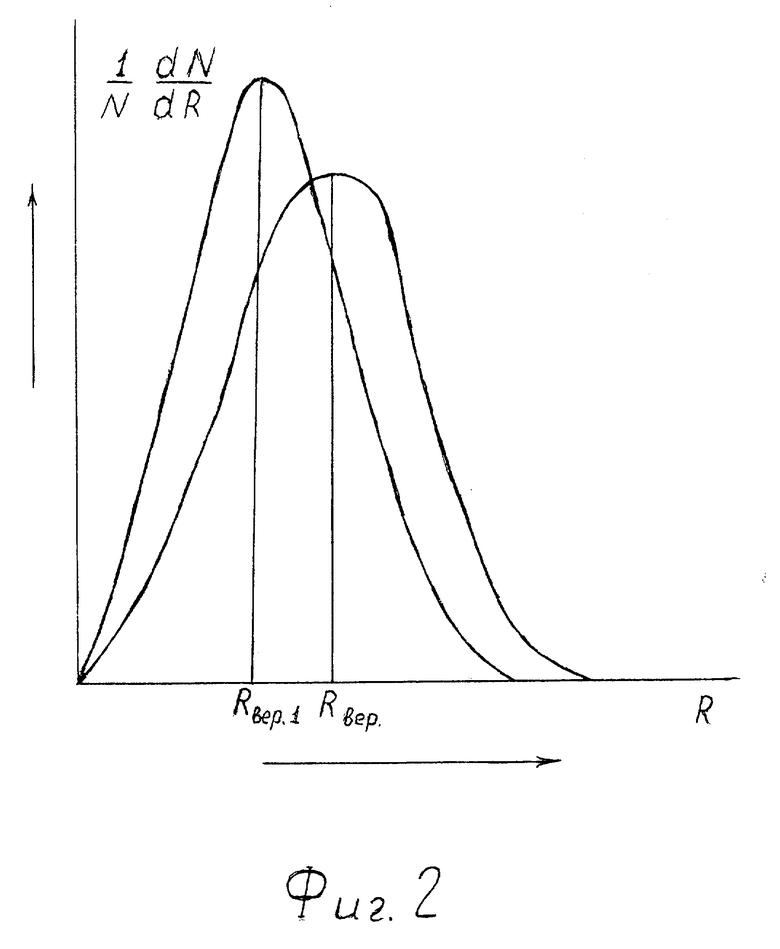
На фиг. 2 - кривые распределения по размерам R сквозных пор в мембранах клеток при двух значениях давления P в среде, окружающей организм.
Кривая с максимумом в точке Rвер относится к меньшему значению давления P.
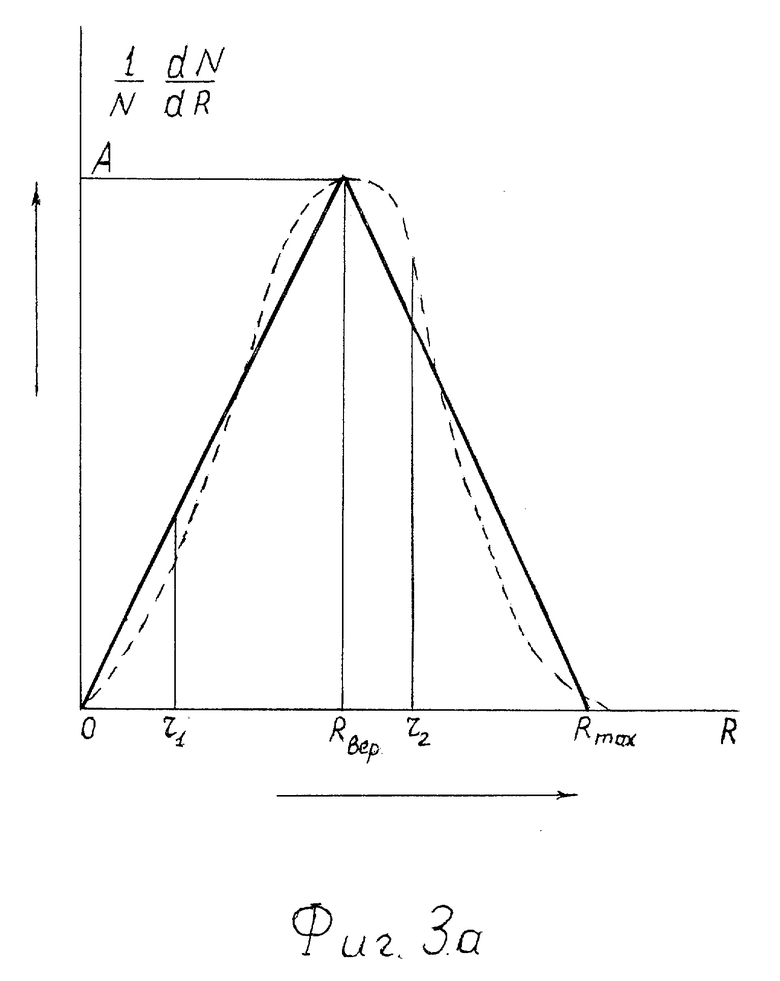
На фиг. 3а - аппроксимация треугольником кривой распределения по размерам R сквозных пор в мембранах клеток.
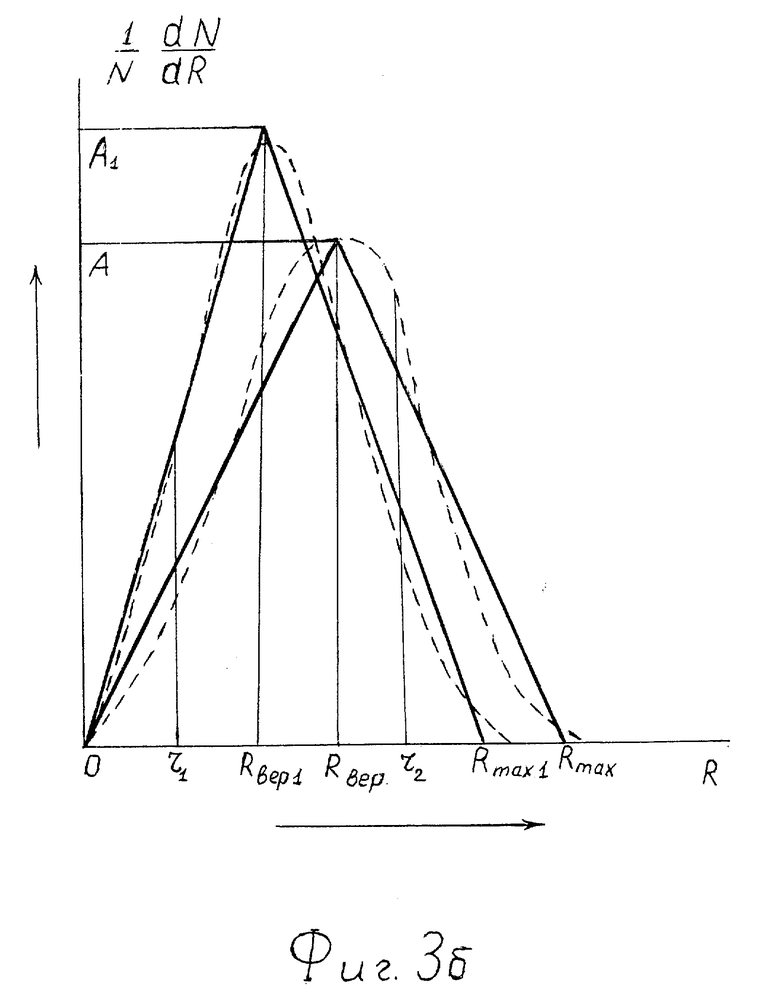
На фиг. 3б - аппроксимация треугольниками кривых распределения по размерам R сквозных пор в мембранах клеток при двух значениях давления в среде, окружающей организмы. Треугольнику меньшей высоты соответствует меньшее значение давления P.
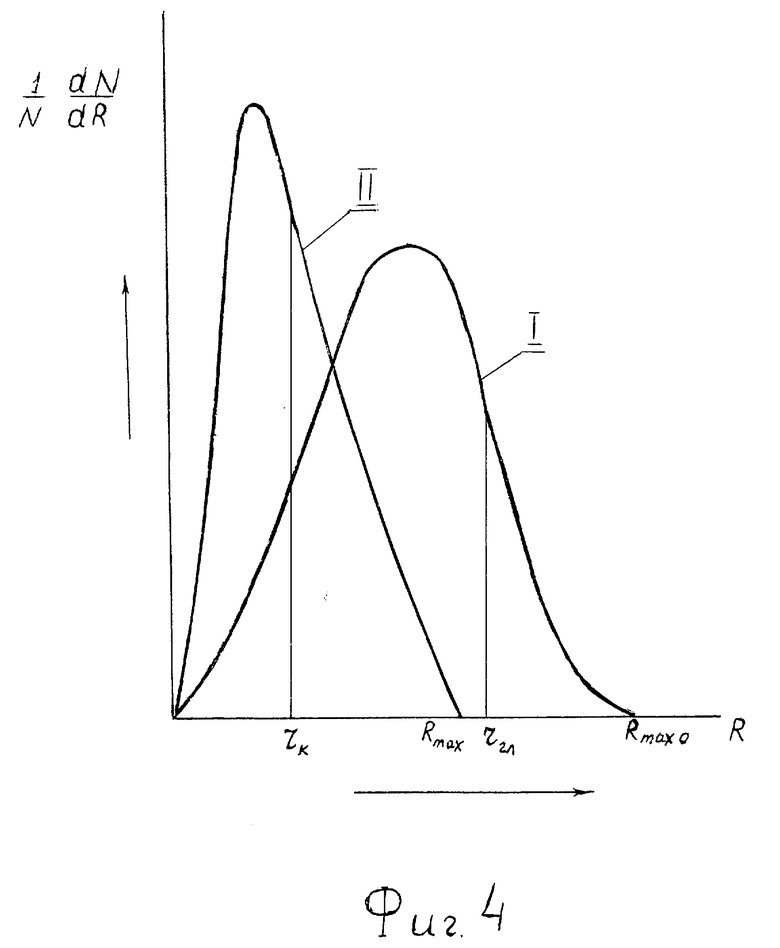
На фиг. 4 - деформация кривой распределения по размерам R сквозных пор в мембранах клеток при росте давления. Кривая I - при меньшем давлении, молекулы кислорода (rк - линейный размер молекулы O2) и глюкозы (rгл - линейный размер ее молекулы) проходят по сквозным порам. Кривая II - при более высоком давлении, кислород еще проходит по сквозным порам, а глюкоза нет, так как линейные размеры молекулы глюкозы rгл больше максимального линейного размера пор.
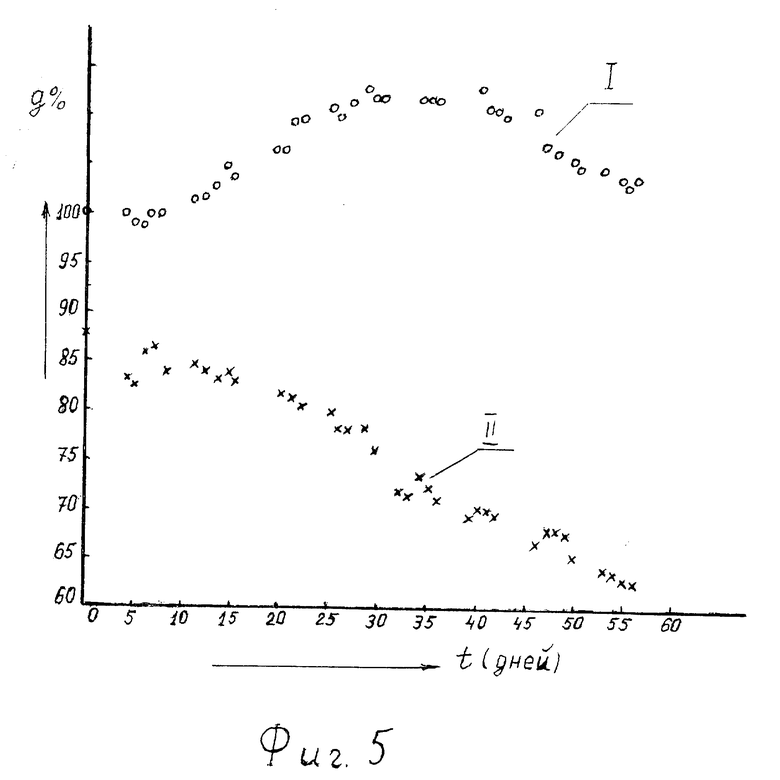
На фиг. 5 - временной ход зависимости веса подопытных мышей в % к исходному: • - контрольной группы (кривая I); x - испытавших воздействие давлением (кривая II).
В подтверждение вышеуказанного можно привести следующие теоретические выкладки.
Мелкие молекулы (глюкоза, углекислый газ, кислород и др.) и ионы, проникают сквозь клеточные оболочки, в основном, через мельчайшие сквозные поры в липидной оболочке. Скорость их проникновения Q подчиняется закону Фика, уже упоминавшемуся выше:
Q = σ(ne- ni),
где
Q - скорость проникновения (иначе поток) частиц сквозь мембрану, частиц/с);
σ - суммарная проницаемость всех оболочек клеток для рассматриваемых частиц, см3/с;
ne - концентрация частиц вне клеток, частиц/см3;
ni - концентрация частиц внутри клеток, частиц/см3.
Сквозные поры в липидном матриксе имеют размеры, сравнимые с размерами липидов в плоскости мембраны, т.е. 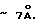 Распределение 1/N•dN/dR среднего размера R пор имеет колоколообразный вид, где N - число пор в мембране клетки (фиг. 1).
Распределение 1/N•dN/dR среднего размера R пор имеет колоколообразный вид, где N - число пор в мембране клетки (фиг. 1).
С ростом давления вследствие всестороннего сжатия гидрофобных липидных хвостов просвет (размер) пор уменьшается. Максимум кривой 1/N•dN/dR сдвигается влево (см. фиг. 2). Наиболее вероятный размер пор Rвер1 уменьшается и выполняется неравенство Rвер1<Rвер.
Сквозь мембрану по сквозным порам проходят лишь те частицы, размер которых меньше размера R поры. Поэтому проницаемость мембраны σ в формуле (2) для таких частиц пропорциональна суммарной площади сквозных пор с размером, большим r, т.е. величине: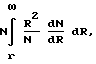
т.е.. 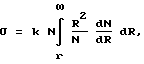 (3)
(3)
где
k = const.
Аппроксимируем кривую 1/N•dN/dR прямоугольником OAR, как показано на фиг. 3а. Тогда можно вычислить проницаемость σ для частиц с размерами r1, меньшими Rвер, и σ2- с размерами r2, большими Rвер. Т.к. площадь под кривой 1/N•dN/dR = 1, то позаботимся, чтобы площадь треугольника была также равна 1.
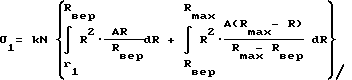 (4)
(4)
обозначим Rвер = a > 0 и Rmax - Rвер = b > 0. Тогда (4) можно записать в виде: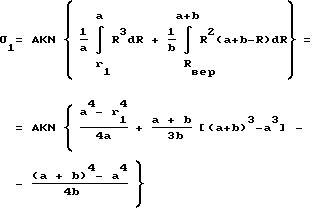
Т.к. A/2(a+b) = 1, т.е.. A = 2/(a+b), то
Для простоты рассмотрения предположим, что кривая 1/N•dN/dR симметрична относительно нормали к оси абсцисс, проходящей через R = Rвер, тогда a = b = a0, и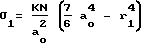 (5)
(5)
где
r1 < a0
Аналогично: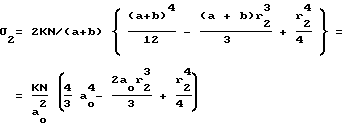 (6)
(6)
где
a0 < r2 < 2a0.
Обобщая, соотношения (5) и (6) можно записать в виде: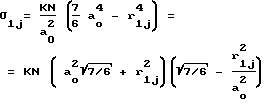 (7)
(7)
где
σj- проницаемость мембраны клеток для мелких частиц j-типа, линейные размеры которых rj < a0.
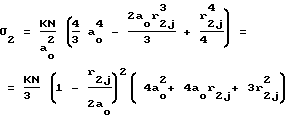 (8)
(8)
σ2j- проницаемость мембраны клеток для частиц j-го типа, линейные размеры которых r2j: a0 < r2j < 2a0.
При повышении давления кривая 1/N•dN/dR деформируется, вершина A смещается влево, в положение A1, а Rmax переходит в положение Rmax1, Rвер - в Rвер1 (фиг. 3б), и a0 уменьшается так, что
Rmax - Rвер = Rвер = a0 > Rmax1 - Rвер1 = Rвер1 = a01 (9)
Часть пор смыкается и исчезает так, что число пор N1 уменьшается:
N > N1 (10)
Сравнивая проницаемость мембраны для частиц j-го сорта σj без действия давления с проницаемостью мембраны σj1 под давлением для обоих случаев: во-первых, когда rj < a0 и, во-вторых, когда a0 < r2 < 2a0 констатируем, что σj > σj1/ Действительно, для первого случая: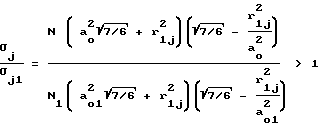 (11)
(11)
поскольку согласно неравенству (10) N/N1 > 1, а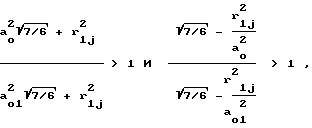
т.к. согласно соотношению (9) a0 > a01.
Для второго случая аналогично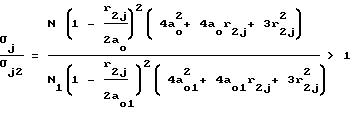 (12)
(12)
поскольку согласно неравенству (10) N/N1 > 1, а
и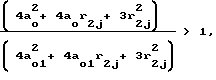
т.к. a0 > a01.
Таким образом, при повышении давления проницаемость мембран для частиц, способных проходить по сквозным порам, уменьшается. Допущение симметрии функции распределения 1/N•dN/dR сквозных пор по размерам не влияет на окончательный вывод. Доказательство уменьшения величины проницаемости σ при росте общего давления P для случая указанной несимметрии аналогично приведенному, но более громоздко и поэтому менее наглядно.
Выше некоторого значения общего давления P размер r становится больше размера Rmax и частицы не могут проходить по сквозным порам. Но это не означает, что они совсем не могут проходить сквозь мембрану. Остается другой путь неспецифического трансмембранного транспорта - растворение в липидах и дрейф, раздвигая их жировые "хвосты". В физиологических условиях данная составляющая неспецифической проницаемости значительно меньше, чем рассмотренная выше, и практически не зависит от давления.
Поскольку молекула глюкозы значительно крупнее молекулы кислорода, то для нее такая ситуация реализуется при гораздо меньших давлениях, чем для кислорода. На этом и основывается возможность регулирования углеводного обмена.
Подняв давление выше того значения, при котором молекулы глюкозы уже не могут проходить внутри клеток по сквозным порам в цитомембранах, можно существенно ограничить поток глюкозы в клетки и свести ее потребление к минимуму. Проницаемость мембран для глюкозы упадет в десятки раз, в то время как адаптационные механизмы организма способны поднять концентрацию глюкозы в крови и межклеточной жидкости только в несколько раз. Согласно соотношению (2) до поднятия давления
Qгл - Qгл= σгл(ne-ni), ((ne - n1),
где
Qгл - поток глюкозы в клетки;
σгл - проницаемость мембран для глюкозы;
ne - концентрация глюкозы в межклеточной жидкости;
ni - концентрация глюкозы в клетках.
Звездочками обозначим эти же величины после поднятия давления, тогда
Q
и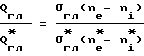 (13)
(13)
Запасов сахара в клетках хватает на 2-3 часа их интенсивного использования (глюкоза запасается в виде гликогена). В течение этого времени концентрация глюкозы в цитоплазме поддерживается почти на том же уровне, но по истощении запаса гликогена падает.
Т.к. ne >> ni, то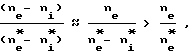
и во втором сомножителе соотношения (13) числитель в несколько раз меньше знаменателя, в то время как числитель первого сомножителя в десятки раз больше знаменателя. В результате получается, что величина Q
Однако, следует следить за тем, чтобы общее давление не оказалось чрезмерно большим и с кислородом не случилось то же самое, что и с глюкозой.
На фиг. 4 показана ситуация, когда при некотором давлении P кислород еще попадает внутрь клеток по сквозным порам в мембране, а глюкоза нет (кривая II). Реализуется ситуация
rк < Rmax < rгл,
где
rк - размер молекулы кислорода;
rгл - размер молекулы глюкозы, а при атмосферном давлении (кривая I)
rгл < rк < Rmax,
При дальнейшем повышении давления в некотором диапазоне его значений в клетках осуществляется авторегулирование проницаемости мембраны для кислорода. Молекулярный механизм этой авторегулировки изучен авторами предлагаемого изобретения и материалы отправлены в печать. В указанном диапазоне с ростом давления и уменьшением размеров сквозных пор автоматически растет их число, так что проницаемость мембраны для кислорода не меняется (этот адаптационный механизм работает так, чтобы внутри клетки сохранялась постоянной концентрация кислорода  и напряжение кислорода
и напряжение кислорода 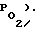 Кислород в клетки будет поступать в нормальных количествах, а поток глюкозы, и, следовательно, ее потребление уменьшится в несколько раз. Для покрытия энергодефицита, вызванного уменьшением утилизации углеводов, клетки вынуждены в большей степени метаболировать жиры. Их запасов без пополнения хватает на полгода. В результате возрастания вклада жиров в суммарное энергопроизводство за счет уменьшения вклада углеводов производство кислых продуктов метаболизма в клетках снижается и pH клеточной среды увеличивается. (Известно, что при производстве единицы макроэргов при утилизации жиров вырабатывается значительно меньше кислых продуктов, чем при утилизации углеводов). Установлено, что в этих условиях активность ферментов жирового пути энергетического метаболизма возрастает (при росте pH цитоплазмы), а активность ферментов углеводного пути - падает. Оба указанных обстоятельства (уменьшение утилизации сахаров и увеличение pH цитоплазмы) приводят к тому, что энергопрозиводство клеток осуществляется преимущественно посредством утилизации жиров.
Кислород в клетки будет поступать в нормальных количествах, а поток глюкозы, и, следовательно, ее потребление уменьшится в несколько раз. Для покрытия энергодефицита, вызванного уменьшением утилизации углеводов, клетки вынуждены в большей степени метаболировать жиры. Их запасов без пополнения хватает на полгода. В результате возрастания вклада жиров в суммарное энергопроизводство за счет уменьшения вклада углеводов производство кислых продуктов метаболизма в клетках снижается и pH клеточной среды увеличивается. (Известно, что при производстве единицы макроэргов при утилизации жиров вырабатывается значительно меньше кислых продуктов, чем при утилизации углеводов). Установлено, что в этих условиях активность ферментов жирового пути энергетического метаболизма возрастает (при росте pH цитоплазмы), а активность ферментов углеводного пути - падает. Оба указанных обстоятельства (уменьшение утилизации сахаров и увеличение pH цитоплазмы) приводят к тому, что энергопрозиводство клеток осуществляется преимущественно посредством утилизации жиров.
Очевидна возможность коррекции предлагаемым способом нарушений углеводного обмена при ожирении и сахарном диабете, а именно, перевод энергетического метаболизма клеток больных с преимущественно углеводного пути на преимущественно жировой.
Однако встает вопрос: а не вернутся ли клетки в прежнее состояние после сброса давления?
При принудительном переводе с помощью давления клеток с углеводного пути энергопроизводства на преимущественно жировой цитоплазма подщелачивается, активируются не только ферменты жирового метаболизма, но и увеличивается также их скорость воспроизводства, в то время как скорость воспроизводства ферментов углеводного метаболизма снижается. Как известно, постоянная времени воспроизводства белков τвоспр≈ 2-4 дня, а их распада - τрасп.- значительно больше - 1-3 месяца (иногда больше).
Если клетки продержать под давлением в условиях принудительного жирового метаболизма неделю или полторы (3-4τвоспр), за это время усиленного воспроизводства в клетках накопится значительно больше ферментов жирового пути метаболизма, а ферментов углеводного пути, наоборот, уменьшиться, по сравнению с тем, что было до поднятия давления и, следовательно, доля ферментов жирового пути в суммарном количестве ферментов энергетического метаболизма возрастет.
Таким образом, время выдержки организмов под повышенным давлением определяет закрепление по крайней мере на несколько месяцев, полезный эффект сдвига энергопроизводства с преимущественно углеводного на преимущественно жировой путь метаболизма.
При дальнейшем увеличении давления уже для молекул кислорода реализуется ситуация, подобная описанной выше для глюкозы. Начиная с некоторого значения давления P будет выполняться условие rк > Rmax, где rк - размер молекулы кислорода, (фиг. 5) и кислород будет проходить по сквозным порам.
При более высоких давлениях, чем, те, при котором rк = Rmax, молекулы кислорода могут поступать внутрь клеток только непосредственно сквозь липиды мембраны, раздвигая жировые хвосты этих молекул. Составляющая проницаемость мембраны (также неспецифическая), обусловленная таким механизмом, на несколько порядков меньше, чем максимальная возможность проницаемость, которая связана с наличием сквозных пор при нормальных условиях и практически не зависит от давления.
При изменении концентрации кислорода ne в межклеточной жидкости адаптационные механизмы клеток удерживают концентрацию кислорода 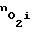 практически постоянной за счет изменения проницаемости мембраны
практически постоянной за счет изменения проницаемости мембраны 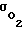 , которая меняется в зависимости от количества сквозных пор N.
, которая меняется в зависимости от количества сквозных пор N.
Если 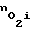 по какой-либо причине (внешней или внутренней) уменьшается, то автоматически увеличивается число пор N и, следовательно,
по какой-либо причине (внешней или внутренней) уменьшается, то автоматически увеличивается число пор N и, следовательно, 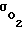 и поток кислорода внутрь клеток из межклеточной жидкости. Величина
и поток кислорода внутрь клеток из межклеточной жидкости. Величина 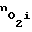 восстанавливается до прежнего значения. И наоборот, при повышении
восстанавливается до прежнего значения. И наоборот, при повышении 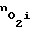 число пор N уменьшается, поток кислорода внутрь клеток уменьшается и концентрация
число пор N уменьшается, поток кислорода внутрь клеток уменьшается и концентрация 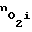 уменьшается. Такой адаптационный механизм может срабатывать только тогда, когда молекулы кислорода могут проходить по порам, однако, когда с ростом давления размеры пор настолько уменьшаются, что молекула кислорода не сможет поместиться в самой крупной поре (с Rmax), то адаптационный механизм перестает срабатывать. Т.к. в мембране клетки может быть не больше некоторого количества пор N (это связано со строением бислоя липидов), то по мере возрастания давления адаптационные возможности клеток к изменению внешней концентрации кислорода
уменьшается. Такой адаптационный механизм может срабатывать только тогда, когда молекулы кислорода могут проходить по порам, однако, когда с ростом давления размеры пор настолько уменьшаются, что молекула кислорода не сможет поместиться в самой крупной поре (с Rmax), то адаптационный механизм перестает срабатывать. Т.к. в мембране клетки может быть не больше некоторого количества пор N (это связано со строением бислоя липидов), то по мере возрастания давления адаптационные возможности клеток к изменению внешней концентрации кислорода 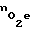 уменьшаются. При выполнении равенства rk = Rmax клетка теряет способность к адаптации. С этого момента в выражении:
уменьшаются. При выполнении равенства rk = Rmax клетка теряет способность к адаптации. С этого момента в выражении: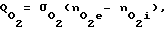
величина 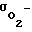 - величина постоянная и даже несколько меньшая, чем в нормальных условиях у покоящейся клетки. Об этом свидетельствует опыт глубоководных водолазов в мягких скафандрах. Несмотря на специальный обогрев скафандров (до 80oC) водолазы при таких погружениях мерзли.
- величина постоянная и даже несколько меньшая, чем в нормальных условиях у покоящейся клетки. Об этом свидетельствует опыт глубоководных водолазов в мягких скафандрах. Несмотря на специальный обогрев скафандров (до 80oC) водолазы при таких погружениях мерзли.
Если бы 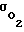 при высоких давлениях была бы много меньше проницаемости при нормальных условиях в покое и поток кислорода был бы настолько мал, что не обеспечивал бы воспроизводство энергии, необходимой для поддержания жизни, то клетки погибли бы, погибли бы и сами водолазы. С другой стороны, поскольку водолазы мерзли, то в клетках был явный дефицит энергопроизводства и поток кислорода в клетки был несколько ниже того, который реализуется в нормальных условиях в покоящихся клетках. Следовательно, величина
при высоких давлениях была бы много меньше проницаемости при нормальных условиях в покое и поток кислорода был бы настолько мал, что не обеспечивал бы воспроизводство энергии, необходимой для поддержания жизни, то клетки погибли бы, погибли бы и сами водолазы. С другой стороны, поскольку водолазы мерзли, то в клетках был явный дефицит энергопроизводства и поток кислорода в клетки был несколько ниже того, который реализуется в нормальных условиях в покоящихся клетках. Следовательно, величина  при высоких давлениях несколько меньшая, чем у покоящейся клетки в нормальных условиях, но достаточно высока для поддержания жизни.
при высоких давлениях несколько меньшая, чем у покоящейся клетки в нормальных условиях, но достаточно высока для поддержания жизни.
Из приведенного следует, что можно подобрать такое давление, которое обеспечит такой поток кислорода внутрь клеток, которого достаточно для поддержания ее состояния в покое, либо в состоянии активности, но не будет хватать для поддержания интенсивного энергопроизводства, необходимого для клеточного деления. Поэтому регулирование интенсивности энергопроизводства предлагаемым способом можно использовать для предотвращения клеточного деления в практических целях.
При уменьшении давления в камере ниже атмосферного картина будет обратной той, которая приведена выше. Энергопроизводство в клетках будет возрастать вследствие возрастания потоков кислорода и глюкозы в цитоплазму, что вызывается увеличением размера сквозных пор к мембране клеток.
Таким образом заявляемым способом можно как увеличить, так и уменьшить энергопроизводство клеток, обеспечив его регулирование в более широком диапазоне, чем способами - аналогами, включая прототипы.
Практически осуществимость заявляемых изобретений на современном техническом уровне не вызывает сомнений. Для осуществления операций, указанных в предлагаемом способе, пригодно выпускаемое серийно оборудование: барокамеры, компрессоры, трубопроводы, системы жизнеобеспечения при высоких давлениях и т.п. Характеристики необходимого оборудования, требуемого для реализации предлагаемого способа, не являются рекордными. Например, требуемые рабочие давления не превышают 30 атм, в то время как промышленно освоено серийное оборудование с рабочим давлением в несколько сотен и даже тысяч атмосфер.
Возможность достижения технического результата с помощью таких средств по каждому из заявляемых изобретений, помимо технического обоснования, приведенного выше, подтверждается нижеописанными опытами с линейными мышами линии C57BL/KS.
В опытах со здоровыми мышами указанной линии выяснено, что даже сравнительно недолгое (3-4 дней) воздействие повышенного давления приводит к снижению их веса. Это объясняется тем, что давление ограничивает поток энергетических субстратов в клетки, и для поддержания необходимого уровня энергопроизводства приходится расходовать их запасы в клетках, в основном, жиры, что приводит к уменьшению массы клеток и потере животными общего веса.
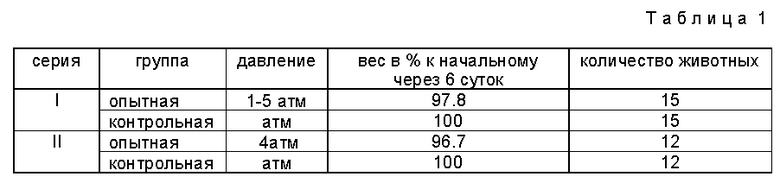
Опыты проводили следующим образом. В две одинаковые камеры объемом ≈ 3 л каждая, рассчитанные на давление до 45 атм, на 6 суток помещали мышей по 3 шт. в каждую. Мыши в одной из камер (контрольная группа) содержались при атмосферном давлении. В другую камеру подавалась газовая смесь из кислорода и аргона в таком соотношении, чтобы при поддержании давления в одной серии опытов 1,5 атм, а в другой 4 атм парциальное давление кислорода в смеси было близким к атмосферному. В камеры в одинаковых количествах помещали также корм и воду, следовательно, мыши контрольных и опытных групп содержались в одинаковых (за исключением давления) условиях. Проток газовой смеси и поглотители препятствовали накоплению углекислого газа в камерах. Перед началом опыта и через каждые 1-2 суток в течение опыта и после животных взвешивали, для чего давление в камере с опытными мышами со всеми предосторожностями приходилось сбрасывать. За поведением животных в процессе эксперимента можно было наблюдать через иллюминаторы в верхних крышках камер. Освещение внутри камеры осуществлялось электрическими лампочками малой мощности. Поведение мышей опытной и контрольной групп во время опытов ничем не отличалось. Животные ели корм, пили воду и двигались по камере с одинаковой интенсивностью. Взвешивание оставшегося после опытов корма показало, что животные в разных камерах питались примерно с одинаковой интенсивностью.
Результаты опытов представлены в табл. 1.
При более высоких давлениях интенсивность кормежки опытных мышей падала и сравнение становилось невозможным.
Результаты убедительно показывают, что как и ожидалось, с ростом давления растет эффект - уменьшение веса животных становится значительней.
Установлено, что наиболее интенсивно вес животных уменьшается в первые два дня, а к четвертым суткам стабилизируется.
Таким образом, данные опыты (вместе с фактами, приведенными выше, о самочувствии водолазов на больших глубинах) неопровержимо свидетельствует о возможности регулирования внутриклеточного энергопроизводства по заявляемому способу.
Возможность применения описанного выше способа регулирования внутриклеточного энергопроизводства в качестве способа лечения онкологических заболеваний можно продемонстрировать следующим образом.
Общепризнанным лечебным эффектом считается замедление роста злокачественного образования в результате уменьшения скорости деления клеток, регрессия опухоли и, как результат, увеличение сроков жизни больных организмов.
В опытах на линейных мышах C57BL/KS с привитой меланомой B16 проверили возможность достижения лечебного эффекта по заявляемому способу.
Опыты проводили по той же схеме, что и описанные выше, в тех же камерах. Контрольная группа мышей содержалась в камере при атмосферном давлении, в то время как опытная - при повышенном давлении в другой, внутри которой парциальное давление кислорода в газовой смеси было близко к атмосферному. Проток газовой смеси и поглотители исключали накопление углекислого газа в камерах.
Мышам подкожно прививали меланому B16. Эта опухоль характеризуется быстрым развитием (время жизни животного с опухолью данного типа 21 ± 5 дней) и метастазированием, в основном, в легкие, печень и селезенку, что выражено на 20 день после прививки у 100% животных. По мере развития и роста опухоли количество некротических клеток в ней повышается и к концу жизни мыши оно составляет около 3/4 ткани новообразования.
Уже при сравнительно низких давлениях (5-8 атм) обнаружился положительный эффект. После выдержки в камере в течение недели срок жизни мышей увеличивался в среднем в полтора раза, замедлялся рост опухоли, почти прекращалось метастазирование (у более чем половины опытных мышей метастазы вообще не наблюдалось), происходили существенные гистологические изменения: в злокачественной ткани проросли сосуды, отсутствовали некротические участки.
Особенно ярко положительный эффект при указанном давлении был выражен в опытах с опухолями 2 - 2.5 -недельного развития.
Однако в опытах с опухолями, находящимися на той же стадии развития, но подвергнутых воздействию более высокого давления (до 20 атм.), хотя и более кратковременному (22-22 часа), получены другие результаты: некротическая ткань в опухоли составляла более 80-90%, некоторая часть мышей через 1-3 дня после опыта погибала, по-видимому, из-за общего заражения крови продуктами разложения некротической ткани. Выжившие мыши жили примерно в 1,5 раза дольше, чем животные контрольной группы. Метастазирование опухоли у выживших мышей также практически отсутствовало.
Результаты опытов с опухолями 2-2,5 дневного развития представлены в табл. 2.
Приведенная выше теория позволяет объяснить различие результатов этих двух серий опытов. По мере роста давления в среде, окружающей пораженные клетки, все более ограничиваются потоки в них важнейших энергетических субстратов: сначала глюкозы, а потом и кислорода, что приводит к уменьшению интенсивности клеточного энергопроизводства, замедлению и даже прекращению деления клеток, требующих больших энергозатрат. При более низких давлениях (5-8 атм) практически полностью перекрывается поток глюкозы внутрь всех клеток-потребителей, как нормальных, так и злокачественных. Известно, что клетки злокачественных новообразований питаются преимущественно глюкозой. Это объясняет замедление их развития в первой серии опытов с привитыми опухолями. При этом наиболее ослабленные клетки погибают. К ним следует отнести, например, клетки, утратившие способность к сцеплению с соседними клетками, которые, отрываясь, попадают в лимфу и затем в кровь и дают метастазы. Однако энергетика большинства клеток опухоли за неделю замедленного деления перестраивается на жировой путь (за это время нарабатывается необходимое количество ферментов жирового пути энергетического метаболизма). Замедление роста опухоли обеспечивает прорастание кровеносных сосудов, улучшающих снабжение клеток кислородом. Иммунная система справляется с утилизацией погибших клеток и после опыта при гистологическом анализе селезенка оказывается чистой. В ней практически нет погибших клеток, как и в самих опухолях.
После воздействия высокого давления (порядка 20 атм) из-за резкого ограничения потоков как глюкозы, так и кислорода погибает гораздо больше клеток опухоли. Практически большинство клеток опухоли не выдерживают нехватки кислорода, в то время как клетки нормальных тканей способны перенести воздействие высокого давления без потерь. Иммунная система не справляется с массой погибших клеток. Вскрытие и гистологический анализ опытных мышей (в том числе и погибших), показали, что их селезенки гипертрофированны, капилляры селезенок буквально забиты погибшими клетками и возросшее кровяное давление раздувало этот орган.
Наблюдаемое в первой серии опытов (как показали морфологический и гистологический анализы) замедление деления и выравнивание клеток по своему развития к одному уровню подтверждает возможность получения по предлагаемому способу синфазных культур клеток.
В других сериях опытов с выдержкой мышей с привитыми опухолями более суток под высоким давлением (порядка 20 атм) обнаружено, что в этих условиях пораженные клетки вообще перестают делиться и почти все погибают, так что опухоль превращается в сплошной гнойник.
Таким образом, в этих сериях опытов показана возможность применения предлагаемого изобретения в качестве способа лечения онкологических заболеваний.
В привитых опухолях меланомы B 16 через 2-2,5 недели развития, как и в большинстве зрелых опухолей данного типа (см. "Биология злокачественного роста". -М.: Наука, 1965, 256 с.), среди делящихся есть клетки как с аэробным, так и анаэробным энергопроизводством. Так как злокачественные клетки по своим основным признакам близки к простейшим микроорганизмам, то вышеприведенные примеры доказывают возможность применения предлагаемого способа регулирования внутриклеточного энергопроизводства в качестве способа предотвращения бактериального разложения пищевых продуктов при их хранении и в качестве способа предотвращения клеточного деления при использовании микроорганизмов для производства белков.
Доказательством возможности применения способа регулирования внутриклеточного энергопроизводства в качестве способа лечения сахарного диабета являются нижеописанные опыты.
Характерный признак сахарного диабета - нарушение углеводородного обмена. Патологическим процессом могут быть охвачены практически любые органы и ткани организма. Сложившийся диабет проявляется в устойчивом повышении уровня глюкозы в крови и появлением сахара в моче. Как правило, на этой стадии развития болезни в патологический процесс вовлечена также поджелудочная железа и инсулиновая недостаточность ярко выражена.
Часто на ранних стадиях болезни наблюдается усиление потребления глюкозы клетками в ущерб утилизации жиров. В результате больные в это время отличаются излишним весом, иногда значительным.
Цель опытов состояла в том, чтобы выяснить возможность применения заявляемого способа для ограничения потоков глюкозы в клетки и коррекции нарушений углеводного обмена у таких больных.
В опытах со здоровыми мышами линии C57BL/KS надежно установлено, что в результате воздействия давления поток Q глюкозы в клетки значительно снижается.
Описание опыта. До помещения опытных мышей в камеру высокого давления их кровь исследовалась на содержание глюкозы. Для этого кончики их хвостов надрезались и в каплях крови с помощью глюкозиметра Reflolux S фирмы Bochringer Mannheim GmbH определялась концентрация глюкозы.
Далее мыши выдерживались под давлением 1-2 атм в течение 2-х, 4-х и 6-ти часов. После сброса давления со скоростью, исключающей баротравмы, вновь замерялась концентрация глюкозы в их крови. Точно такие же операции, но без подъема давления проводились с мышами из контрольной группы. Двухчасовой интервал был выбран для того, чтобы исключить влияние стрессов у мышей при проведении измерений, так как ранее было выяснено, что если проводить измерения чаще, чем через 40-60 мин, то не исключены случаи, когда из-за стрессов уровень сахара в крови исследуемой мыши возрастал от измерения к измерению. 2-х часовой интервал между измерениями надежно гарантировал от влияния стрессов.
В контрольной группе, над которой производили те же самые операции, что и над опытной, т.е. сажали в камеру, но давление не поднимали, уровень глюкозы в крови практически во всех измерениях был один и тот же и близок к норме. У опытных мышей после сброса давления после двухчасовой выдержки уровень глюкозы был ниже исходного на 20-40%, после четырехчасовой возрастал до исходного, а после 6-ти часов превосходил исходный на ≈ 20%. Все изменения уровня глюкозы были за пределами ошибок измерений. Опыты проводили натощак; мышей в течение всего опыта не кормили.
Результаты следует интерпретировать однозначно: давление влияет на уровень глюкозы в крови. Для выяснения вопроса, как это реализуется, запишем необходимые соотношения для потоков глюкозы в организме натощак. В этом случае глюкоза в кровь поступает из печени, где в клетках производится из жиров. Их запасов хватает на несколько месяцев без пополнения.
Выходящий из клеток печени поток глюкозы Q (г/сек) поступает сначала в кровь, где ее концентрация сначала поддерживается на определенном уровне.
Далее этот поток из крови попадает в клеки-потребители, где полностью усваивается. Скорость усвоения глюкозы в клетках при малых концентрациях пропорциональна ее концентрации nкл в клетках так, что
Q = kкл • nкл, (14)
где kкл = const.
С другой стороны, по закону диффузии, во-первых,
Q = σпеч (nпеч - nкр), (15)
и, во-вторых,
Q = σкл (nкр - nкл), (16)
где σпеч и σкл- суммарная проницаемость для глюкозы мембран клеток печени и клеток - потребителей глюкозы соответственно;
nпеч - концентрация глюкозы в цитоплазме клеток печени.
Производя в соотношениях (14), (15) и (16) преобразования, получим систему
Решая эту систему, получим (17)
(17)
и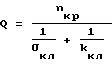 (18)
(18)
Из соотношения (18) видно, что при уменьшении nкр, как наблюдалось при выдержке мышей под давлением в течение 2 часов величина Q уменьшается даже если σкл остается постоянной. Это следует из того, что kкл зависит от количества ферментов и их активности, а они за столь малое время выдержки измениться не могут. Поэтому kкл также постоянная величина.
Концентрацию глюкозы в крови мыши nкр авторы могли измерить только вне камеры после быстрого (5-10 минут) сброса давления. Дальнейшие измерения показали, что nкр со временем растет и приходит в норму через 30-40 минут. Отсюда следует вывод: после 2-часовой выдержки под давлением nкр в камере была несколько меньше измеренной, и поток Q несколько меньший, чем полученный из соотношения (18) подстановкой значения nкр, измеренного на опыте.
Для дальнейшего анализа опытов следует учитывать, что в клетки-потребители поступает не вся глюкоза, вырабатываемая печенью, а только часть. Другая часть утилизуется непосредственно в цитоплазме печеночных клеток.
Q = Qпр - Qпеч, (19)
где Q - поток глюкозы из печени в кровь;
Qпр - скорость производства клетками печени глюкозы из жиров и белков;
Qпеч - скорость утилизации глюкозы клетками печени.
Qпеч = kпеч • nпеч, (20)
где kпеч = const.
kпеч - зависит от количества ферментов, утилизирующих глюкозу и их активности.
nпеч - концентрация глюкозы в клетках печени.
Присоединяя соотношения (19) и (20) к уравнениям системы (1), находим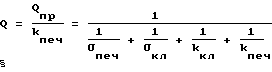 (21)
(21)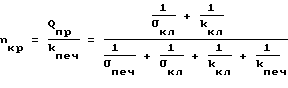 (22)
(22)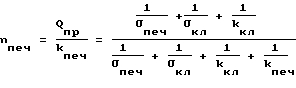 (23)
(23) (24)
(24)
Из соотношения (21) видно, что второй множитель правой части с ростом давления и уменьшением σпеч и σкл должен убывать. Скорость производства глюкозы в клетках печени Qпр не зависит от давления, а зависит только от количества ферментов реакций превращения жиров и белков в глюкозу и от активности этих ферментов. Последняя может повышаться под воздействием гормонов - глюкагона. Однако глюкагон не выделяется в кровь из α - клеток поджелудочной железы, пока не исчерпаны резервы (в виде гликогена) глюкозы в клетках, которых хватает примерно на 2,5 часа. Поэтому в течение этого времени Qпр будет сохраняться постоянным.
Обратимся к соотношению (23). Здесь в течение 2,5 часов Qпр постоянно. Величины kпеч и kкл также постоянны, даже при действии глюкагона. Обозначив величину 1/σпеч+ 1/σкл+ 1/kкл через x, получим из (18) формулу Михаэлиса-Ментен, в которой x при возрастании давления возрастает; растет значение всего выражения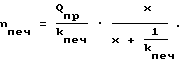
Следовательно, с ростом давления растет концентрация глюкозы в клетках печени. Ясно, что запасы глюкозы (гликоген) в клетках печени не расходуется.
В клетках-потребителях глюкозы, напротив, ее концентрация, как это видно из соотношения (24), при росте давления и уменьшении σкл и σпеч должна падать. Уменьшение притока глюкозы Q из печени в клетки-потребители некоторое время (2,5 часа) компенсируется распадов гликогена и выброса глюкагона в кровь не происходит.
Обратимся теперь к соотношению (22). Проницаемость мембраны клеток печени и клеток-потребителей глюкозы при увеличении давления уменьшается в одинаковое число раз (из-за одинакового строения липидного матрикса), так что всегда выполняется соотношение: σпеч = λσкл, где константа λ > 0.
Поэтому соотношение (22) можно записать в виде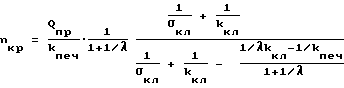 (25)
(25)
Т. к. Qпр, kпеч, Kкл и λ _ - положительные константы, σкл при росте давления уменьшается, вызывая увеличение суммы 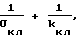 а опыт показал, что величина nкр - уменьшается, то величина 1/λkкл- 1/kпеч должна быть больше нуля, т.е. должно выполняться условие
а опыт показал, что величина nкр - уменьшается, то величина 1/λkкл- 1/kпеч должна быть больше нуля, т.е. должно выполняться условие
1/λkкл> 1/kпеч (26)
Только при выполнении (26) концентрация глюкозы в крови nкр при увеличении давления будет падать. Таким образом, выяснено, что выражение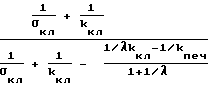
в соотношении (25) и выражение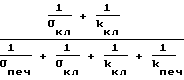
в соотношении (22) с ростом давления уменьшаются.
После того как в клетках-потребителях исчерпаются резервы глюкозы (через 2,5 часа), в кровь поступает гликоген. Скорость производства глюкозы Qпр клетками печени растет. Поэтому после 4-часовой выдержки мышей при повышенном давлении уровень глюкозы в крови повышается до нормального (отношение Qпр/kпеч в (22) и (25) растет, что компенсирует описанное выше уменьшение последних сомножителей этих выражений).
При 6-часовой выдержке мышей при повышенном давлении концентрация глюкозы в крови превышает норму приблизительно на 20%. В этом случае в соотношениях (22) и (25) увеличение первого сомножителя, связанное с ростом скорости производства глюкозы Qпр, уже не компенсируется соответственно уменьшением последнего сомножителя, связанного с уменьшением величин 
Однако в этом опыте невозможно установить, уменьшилась ли после 6-часовой выдержки под давлением скорость усвоения глюкозы Q клетками-потребителями или гормональный сигнал, подняв скорость выработки глюкозы Qпр печенью, довел ее поток Q в кровь до нормы.
Чтобы ответить на этот вопрос авторы провели опыты с мышами той же линии, но диабетическими мутантами (гомозиготные мыши db/db). Эти мыши характеризуются ожирением уже с 3-4-недельного возраста. В 4-месячном возрасте достигают максимальной массы тела 50-60 г. Затем на фоне тяжелого сахарного диабета масса тела быстро падает и в 5-7-месячном возрасте животные погибают от диабетической комы.
Прежде чем перейти к описанию опыта и обслуживанию его результатов, сделаем несколько предварительных замечаний.
Выше показано, что для установления факта ограничений потока Q глюкозы в клетки-потребители давлением, измерение уровня сахара в крови nкр недостаточно информативно из-за приспособления организма к влиянию давления. Но адаптационные возможности клеток небеспредельны. Известно, что реакции производства глюкозы - это насыщающиеся реакции как относительно концентраций гормонов (гликогена), так и концентраций глюкозы в клетках печени. Существуют предельные значения Qпрmax и nпогmax, выше которых данные величины подниматься не могут. Из формул (19) и (21) следует, что начиная с некоторых значений σ
При давлениях выше P* клетки, удовлетворяя потребность в энергии, вынуждены перейти на утилизацию жиров. Переход с углеводного на жировой метаболизм проще всего наблюдать у тучных животных, контролируя их вес. Поэтому опыты проводились с линейными мышами, отличающимися именно тучностью.
После выдержки мышей в течение недели под тем же давлением, что и в первом опыте, животные потеряли около 12,5% от исходного веса, несмотря на то, что в камере ели тот же самый корм и с той же интенсивностью, что и контрольные. В контрольной группе вес повышался с той же скоростью, что и до опыта.
Таким образом, на опыте доказано, что повышенное давление ограничивает поток небольших молекул внутрь клеток, в частности глюкозы.
Дальнейшее наблюдение за опытами и контрольными животными показало, что полученный эффект - переход клеток на жировой путь энергетического метаболизма - закрепляется и сохраняется по крайней мере на 1,5-2 месяца. Степень закрепления полученного эффекта зависит от времени нахождения животных под высоким давлением.
Типичный временной ход изменения веса опытных и контрольных мышей показан на фиг. 5. Кривая I (обозначена кружками) - мыши из контрольной группы, кривая II (обозначена крестиками) - мыши из опытной группы. По оси абсцисс отложено время t(дни) от момента снятия избыточного давления, а по оси ординат g(t)% - вес животных в процентах от начального (до помещения опытных мышей в камеры и поднятия давления), усредненный по группам отдельно опытной и контрольной. Видно, что даже за 2 месяца опытные животные не успевают набрать вес, равный начальному (до воздействия давлением). Более того, у животных опытной группы он продолжал падать, приближаясь к нормальному (g= 50-55%), характерному для данной линии мышей.
Таим образом, результаты приведенных опытов свидетельствуют о возможности применения заявляемого изобретения в качестве способа лечения сахарного диабета.
Столь быстрое уменьшение веса организма при сравнительно быстром избыточном давлении открывает возможность использовать предложенный способ для избавления от избыточного веса в лечебных целях.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ БЛОКИРОВАНИЯ ДЕЛЕНИЯ РАКОВЫХ КЛЕТОК | 2005 |
|
RU2296591C2 |
| СПОСОБ ОЦЕНКИ СОСТОЯНИЯ КИСЛОРОДНО-ТРАНСПОРТНОЙ ФУНКЦИИ КРОВИ У СУБЪЕКТА И ЕЕ ОТКЛОНЕНИЙ ОТ НОРМЫ | 2018 |
|
RU2703510C1 |
| СПОСОБ ЛЕЧЕНИЯ БОЛЬНЫХ С СЕРДЕЧНО-СОСУДИСТЫМИ ЗАБОЛЕВАНИЯМИ | 2009 |
|
RU2392919C1 |
| СПОСОБ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2019 |
|
RU2738165C1 |
| Состав для профилактики и лечения стресса у собак и кошек экзогенной этиологии | 2019 |
|
RU2735863C1 |
| СПОСОБ ЛЕЧЕНИЯ СОЛИДНЫХ ЗЛОКАЧЕСТВЕННЫХ НОВООБРАЗОВАНИЙ И ИХ МЕТАСТАЗОВ | 2010 |
|
RU2437692C2 |
| Способ лечения пациентов с ишемической болезнью сердца | 2023 |
|
RU2818733C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ СКОРОСТИ ВНУТРЕННЕГО ПОТРЕБЛЕНИЯ КИСЛОРОДА ЭРИТРОЦИТАМИ И УСТРОЙСТВО ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 1995 |
|
RU2146051C1 |
| СПОСОБ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ В ЭКСПЕРИМЕНТЕ | 2011 |
|
RU2484539C1 |
| СПОСОБЫ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, АССОЦИИРОВАННЫХ С ВПЧ | 2019 |
|
RU2799784C2 |

Изобретение может быть применено при лечении онкологических заболеваний и сахарного диабета, а также при хранении пищевых продуктов и микробиологическом синтезе. Объект обработки помещают в герметичную камеру, заполненную средой. Камера дополнительно содержит средства жизнеобеспечения. Среда, заполняющая камеру, содержит кислород и биологически неактивные компоненты. Периодически изменяют давление в камере путем изменения содержания биологически неактивных компонентов. Использование изобретения позволяет управлять ростом клеток организма. 5 с. и 3 з.п.ф-лы, 5 ил., 2 табл.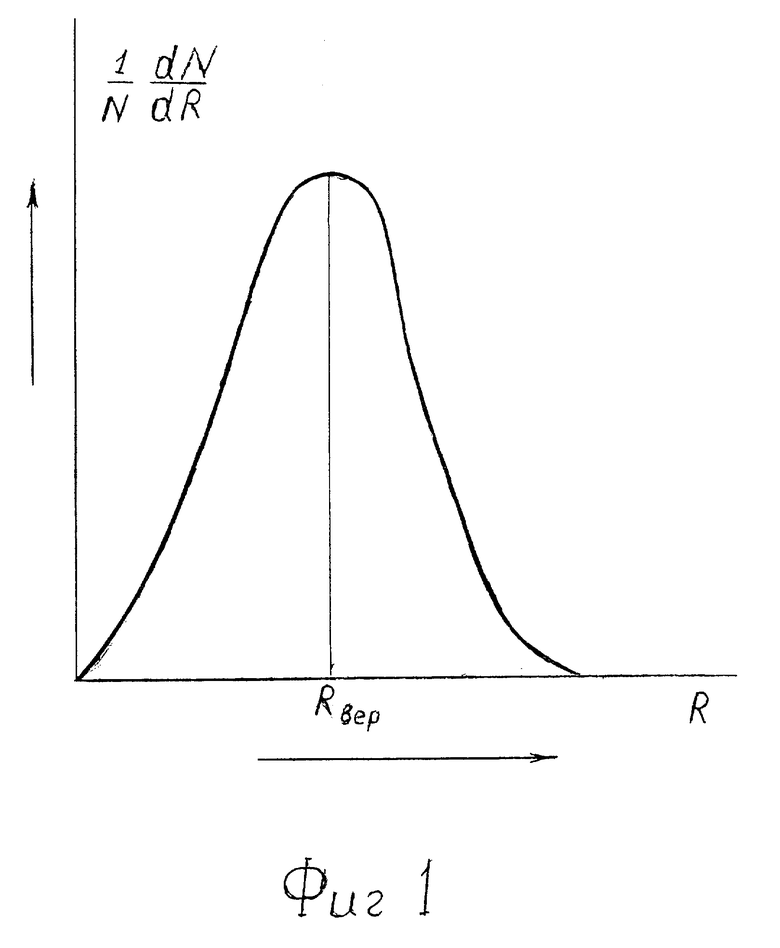
| Бураковский В.И | |||
| и др | |||
| Гипербарическая оксигенация в сердечно-сосудистой хирургии | |||
| - М.: Медицина, 1974, с.81-114 | |||
| Способ получения белка | 1973 |
|
SU591154A3 |
| Огнетушитель | 0 |
|
SU91A1 |

