В общем плане настоящее изобретение относится к гелонину иммунотоксина и, более конкретно, к молекулярной биологии гелонина, включая способ получения синтетического гена гелонина.
Одной из причин интереса в настоящее время к цитотоксическим веществам является их потенциональное применение в области специфических мишеневых опухолевых клет. Растительный токсин гелонин привлек такое внимание. Гелонин представляет собой гликопротеин (Мв. приблизительно 29-30000 КД), выделенный из семян Gelonium multiforum. Гелонин принадлежит к классу потенциальных растительных токсинов, инактивирующих рибосомы.
Другими членами этого класса инактивирующих рибосомы растительных токсинов являются цепи абрина, рицина и модецина. Гелонин, подобно абрину и рицину, ингибирует синтез протеина, повреждая 60S субъединицу рибосом млекопитающих. Хотя A цепь рицина (RTA) широко использовали в иммунотоксинах, гелонин, по-видимому, более стабилен по отношению к химической и физической обработкам, нежели RTA (Barbieri et al, Cahcer Surv. 1:489-520 (1982). Более того, сам гелонин не связывается с клетками и поэтому является нетоксичным (за исключением случаев высоких концентраций) и с ним безопасно работать в лаборатории. Инактивация рибосом обратима, по-видимому, не включает ко-факторов и происходит с эффективностью, которая предполагает энзиматический механизм действия гелонина.
Гелонин и рицин находятся среди большинства активных токсинов, которые ингибируют синтез протеина на основании веса протеина. Гелонин в 10-1000 раз более активен в ингибировании синтеза протеина, нежели цепь рицина A. Пептиды, подобные рицину и абрину, состоят из двух цепей: из A цепи, которая является токсичным звеном, и B цепи, которая функционирует как связывающаяся с клеткой. В отличие от рицина и абрина гелонин состоит из одной цепи, и так как у него нет В цепи для связывания с клетками, сам по себе он относительно нетоксичен для интактных клеток (Stirpe, et.al J. Biol. Chem. 255:6947-6953/1980). Клетки млекопитающих, по-видимому, не могут связывать и/или интернализировать нативную молекулу гелонина. Конъюгаты гелонина с мишеневыми для опухоли моноклональными антителами, такими как моноклональное антитело ZME, направленное на антиген, присутствующий на некоторых опухолевых клетках, таких как клетки меланомы, обеспечивают как специфический способ связывания гелонина с клеткой, так и способ интернализации комплекса гелонин-антитело. Одно из преимуществ использования гелонина токсина по сравнению с такими токсинами, как A цепь рицина, состоит в пониженной токсичности по отношению к здоровым тканям по сравнению с токсичностью цепи A рицина. Гелонин, соединенный с моноклональным антителом, направленным на антиопухолевый антиген, является активным и селективным иммунотоксическим агентом для лечения опухолей.
Некоторые исследователи используют гелонин в качестве цитотоксического агента, химически связанного с моноклональными антителами или с пептидными гормональными клеточными мишеневыми лигандами. Однако химические модификации гелонина и клеточные мишеневые фрагменты могут снизить мишеневую эффективность и цитотоксический потенциал самого гелонина. Более того, природные источники гелонина находятся в различных условиях при сборе и росте растений, которые могут влиять на цитотоксическую активность гелонина. Способность продуцировать синтетический гелониновый токсин, химическим путем или используя рекомбинатные технологии, обеспечивает богатый, репродуцируемый источник токсина. Поэтому весьма желательно получить синтетический ген гелонина и разработать способы получения синтетического гена с помощью рекомбинантной технологии.
Настоящее изобретение обеспечивает нуклеотидную последовательность для синтетического гена гелонина и способы его получения. ДНК последовательность синтетического гена гелонина представлена в Последовательности ID N 1 (SEQID N 1). В настоящем изобретении предложены также векторы экспрессии, содержащие такие ДНК последовательности, и клетки, трансформированные такими векторами.
В соответствии с одним вариантом настоящего изобретения предложен способ получения, клонирования и экспрессии синтетического гена, кодирующего гелонин. Вначале конструируют фрагмент двунитевой ДНК, кодирующий первичную аминокислотную последовательность, определенную для очищенного гелонинового протеина. Затем с этим ДНК фрагментом манипулируют для облегчения синтеза, клонирования и экспрессии или биохимического манипулирования с геном.
Далее конструируют, синтезируют, очищают и 5'-фосфорилируют набор синтетических олигонуклеотидов, способных связываться вместе с образованием нужного гена. Затем эти олигонуклеотиды отжигают и лигируют вместе до получения интактного гена. Этот синтетический ген лигируют вместе с подходящим клонирующим вектором, а затем определяют нуклеотидную последовательность клонированного гена. После этого проводят сайт-направленный мутагенез для коррекции любых нежелательных мутаций клонированного гена, и этот ген субклонируют в подходящий вектор экспрессии. Вектор экспрессии, несущий синтетический гелониновый ген, встраивают в подходящего хозяина экспрессии. После этого условия для хозяина экспрессии поддерживают такими, которые обеспечивают продуцирование гелонинового генного продукта, и протеин гелонина выделяют и очищают из клеток, экспрессирующих ген.
В соответствии с другим вариантом настоящего изобретения, предложена синтетическая ДНК, содержащая последовательность нуклеотидов и фрагментов и их производных, кодирующую протеин гелонин или полипептид, который ингибирует клеточный синтез протеина, но не связывает рецепторов поверхности клетки, причем эта нуклеотидная последовательность имеет формулу последовательности ID N 1. Предложен также вектор экспрессии, содержащий синтетическую ДНК гелонина, и клетки хозяина, содержащие и экспрессирующие синтетический ген гелонина.
В соответствии с еще одним вариантом настоящего изобретения предложен иммунотоксин, содержащий антитело, конъюгированное с растительным протеином токсина - гелонином.
Для целей настоящего изобретения термин "фрагмент" определяют как любую часть последовательности ID N 1, которая может продуцировать протеин, который ингибирует синтез клеточного протеина, но которая не связывается с поверхностью рецептора. "Производное" определяют как содержащее замещение - одной или более из отдельных нуклеиновых кислот, так что получается тот же самый протеин или полипептид.
Настоящее изобретение учитывает и адресовано к указанной ранее существенной необходимости и обеспечивает достаточное удовлетворение такой необходимости и его различных вариантов. Специалистам этой области будут очевидны другие цели и преимущества изобретения, а также методики настоящего изобретения. Для раскрытия целей изобретения приводится следующее описание вместе с прилагаемыми рисунками. Хотя эти описания детализированы для обеспечения адекватности и с целью лучшего понимания, они не должны ограничивать патент, цель которого заявить изобретение независимо от того, как впоследствии кто-либо сможет его замаскировать различными вариациями или добавлениями дальнейших усовершенствований.
С тем, чтобы сделать более понятными подробности способа (которым вышеуказанные характеристики, преимущества и цели изобретения, а также и все остальное, что станет ясно далее, достигается), более конкретное описание изобретения, кратко суммированного ранее, можно привести со ссылкой на его некоторые варианты, которые иллюстрируются прилагаемыми рисунками. Эти рисунки составляют часть описания. Однако следует учесть, что прилагаемые рисунки иллюстрируют лишь предпочтительные варианты изобретения, и поэтому их не следует рассматривать как ограничивающие его объем. Настоящее изобретение может допускать другие равноэффективные эквивалентные варианты.
Фиг. 1 изображает схематическую иллюстрацию способа химического синтеза олигонуклеотидов.
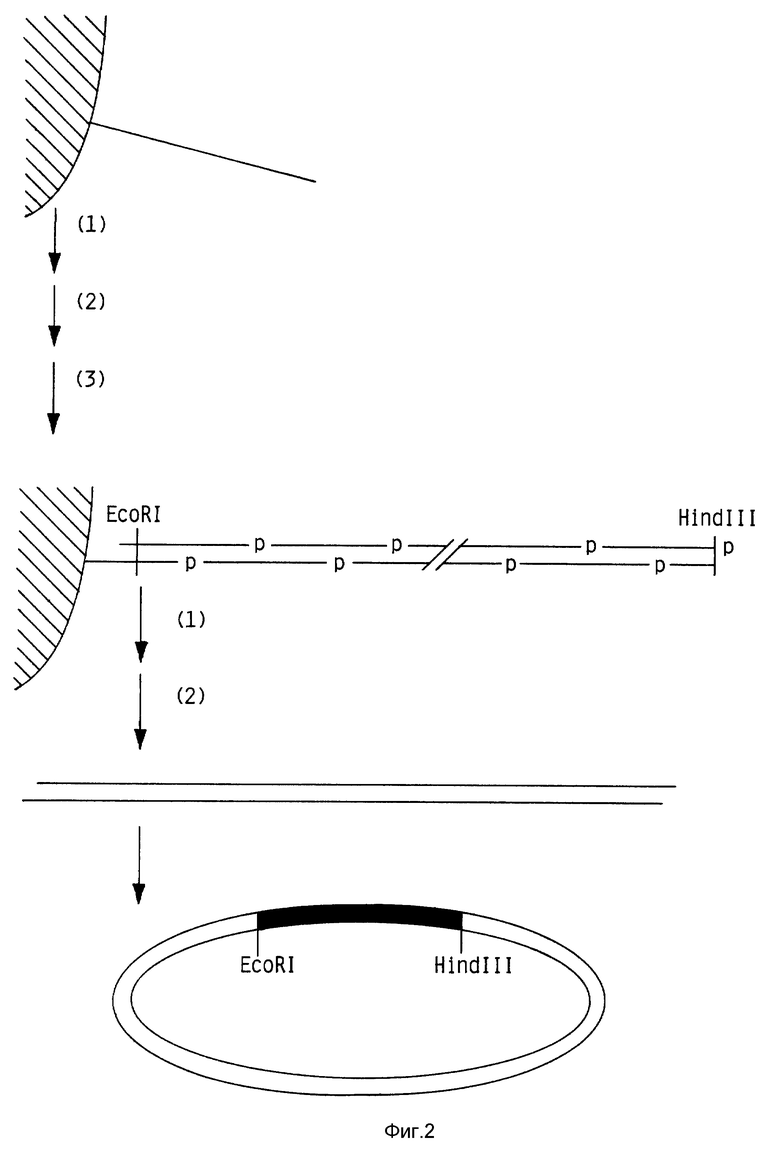
Фиг. 2 изображает процесс твердофазной сборки гена.
Фиг. 3 представляет схематическую диаграмму процесса сборки гена гелонина.
Фиг. 4 изображает схематическую иллюстрацию организации олигонуклеотидов в сборке гена гелонина.
Фиг. 5 изображает олигонуклеотидную последовательность, использованную для сборки гена гелонина.
Последние разработки в молекулярной биологии обеспечили возможность клонирования, экспрессии и генной инженерии множества генов, кодирующих протеины, представляющие ценности в биомедицине и сельском хозяйстве. Недавним заметным достижением в этой области является способность конструировать и продуцировать синтетические гены. Синтетические гены могут кодировать протеины известных аминокислотных последовательностей, а также новые протеины, которые не существуют в природе. Это особенно полезно в случае протеинов, представляющих терапевтическую ценность, таких как гелонин. Становится возможным конструировать, синтезировать, клонировать и экспрессировать ген на основании аминокислотной последовательности протеина. Конструирование, секвенирование, клонирование и экспрессирование возможны также, даже если недоступна специфическая информация природного гена, например, если ген до сих пор не был клонирован. Более того, синтез гена облегчает конструирование различных генных продуктов, обладающих свойствами, которые не найдены в природных протеинах. Так например, ген, кодирующий протеин, обычно находящийся только в растениях или животных, может быть сконструирован, синтезирован и клонирован в вектор, который может привести к продуцированию больших количеств протеина в клетках микробных хозяев.
Одно из преимуществ синтетических генов связано с разнообразием генетического кода. Большинство аминокислот может быть закодировано более чем одним "кодоном" из трех оснований внутри гена. Различные организмы имеют тенденцию использовать различные кодоны для их протеинов (S.Aota et. al., Nucleic Acids Res, v. 16, Supplement, p. r315-r391 (1987). Другими словами, кодоны, которые "предпочтительны" в одних организмах, отличаются от тех, которые предпочтительны в других организмах. Считают, что это явление связано с различиями в относительном избытке специфических изоакцепторных транспортных PHK молекул для данной аминокислоты. Можно сконструировать синтетические гены для включения предпочтительных кодонов для данной системы экспрессии, даже если генный продукт получен из "гетерологических" организмов хозяев, которые используют различный набор предпочтительных кодонов для некоторых аминокислот. Было показано, что синтетические гены, кодирующие протеины млекопитающих, дают заметно больше протеинового продукта в микробном хозяине, если выбранные кодоны в генной конструкции соответствуют тем, которые наиболее часто используются микробным хозяином. Это явление учитывалось при создании гена галонина (из растений), предназначенного для интродукции в экспрессирующего хозяина Escherichia coli. Для экспрессии в систему другого хозяина ген гелонина можно легко сконструировать заново (это может проделать любой специалист в этой области) для включения кодонов, оптимальных для другого хозяина.
Другое преимущество синтетических генов, реально обеспечиваемое многообразием генетического кода, состоит в том, что ДНК последовательность, кодирующую представляющий интерес ген, можно модифицировать (не изменяя закодированную аминокислотную последовательность) так, чтобы она содержала максимальное количество сайтов узнавания уникальных рестрикционных энзимов в этом гене. Существование множества однозначных рестрикционных сайтов в гене существенно облегчает биохимические манипуляции с этим геном. Так например, способность вырезать сегмент гена и заменять его другой ДНК последовательностью (что обеспечивается за счет близко расположенных уникальных рестрикционных сайтов) облегчает введение новой генетической информации в ген с помощью методик рекомбинатных ДНК. Удобные манипуляции, обеспечиваемые такими подходами, включают введение мутаций в определенные участки гена, исправление мутаций, возникающих в процессе синтеза гена, сборки и клонирования, и создания химерических или слитых генов, кодирующих протеины, которые объединяют функциональные домены отдельных протеинов.
Конструкция синтетического гена может также включать некодирующие "фланкирующие" ДНК последовательности, которые могут облегчить клонирование и экспрессию гена. Так например, последовательность, распознающую рестрикционную эндонуклеазу, можно включить во фланкирующие участки гена для обеспечения специфического лигирования гена в целевой вектор клонирования. Кроме того, генетические сигналы можно встроить в синтетический ген, который служит для контроля за экспрессией гена ин виво.
Кроме того, вновь сконструированный ген может облегчить введение в определенное место ин виво генного продукта в некоторые ткани или органы, что может повысить терапевтическое действие генного продукта. Так например, это облегчает направленное введение терапевтического агента в конкретный тип клеток за счет введения в соответствующий ген последовательности, которая кодирует полипептидный домен, который специфически связывается с рецептором на поверхности мишеневых клеток.
Конструкция синтетического гена может, кроме того, включать такие рассмотрения, которые связаны с химическим синтезом коротких нитей ДНК и сборкой олигонуклеотидов в более длинные дуплексные ДНК сегменты. В химическом синтезе ДНК важно избежать слабой эффективности связывания, связанной с последующим добавлением множества G или C остатков к цепи. Кроме того, желательно избегать ДНК последовательностей, которые содержат межнитевые вторичные структуры (структуры "шпильки") или межмолекулярные комплементарности, которые могут помешать правильной сборке гена во время отжига компонентов олигонуклеотидов. Этих целей можно достичь, выбирая альтернативные кодоны в генной конструкции.
Способ сборки генов включает следующие стадии. Вначале выделяют нуклеотидную последовательность, соответствующую двум нитям целевого кодирующего участка, обеспечивая хорошее спаривание комплементарных оснований между двумя нитями. Затем добавляют любые нужные фланкирующие последовательности. Так например, можно добавлять фланкирующие последовательности для включения сайтов распознавания рестрикционной эндонуклеазы соседних с кодирующей последовательностью. Затем ген разделяют на перекрывающиеся наборы однонитевых фрагментов. Однонитевые фрагменты химически синтезируют в приборе для автоматического синтеза ДНК. Степень комплементарного перекрывания между последовательными олигонуклеотидами вместе с синтетическим геном представляет вопрос выбора, но обычно составляет 6-20 оснований. После того как синтезированы все олигонуклеотиды, необходимые для синтетического гена, их предпочтительно очищают с помощью электрофореза на полиакриламидном геле или с помощью высокоэффективной жидкостной хроматографии. Затем очищенные олигонуклеотиды 5'-фосфорилируют под действием полинуклеотидкиназы и аденозин-5'-триптофазы. Затем нити отжигают вместе, либо в отдельной смеси в блоках 3-10 перекрывающихся олигонуклеотидов, либо поэтапно, добавляя олигонуклеотиды на твердофазный носитель. Концы собранного гена снабжают рестрикционными сайтами, которые используют при клонировании гена. Собранный ген обычно клонируют вначале в однонитевой вектор M13 для удобного ДНК секвенирования. При необходимости мутации исправляют олигонуклеотид направленным мутагенезом.
Нужные характеристики синтетического гена (оптимальное использование кодона, существование уникальных рестрикционных сайтов, исключение вторичной структуры и т. д. ) можно определить с помощью любой из ряда коммерческих программ анализа последовательностей ДНК для микрокомпьютеров.
Две последние разработки, обе из которых использованы в способе настоящего изобретения, обеспечивают более быстрый и экономичный синтез генов и создают новые возможности для конструирования протеина (K.L. Beattie et. al, Biotechnol, Appl. Biochem., 10, 510-521 (1988); K.L. Beattie and R.F. Fowler, Nature, 352, 548-549 (1991). Первая разработка, представленная на фиг. 1, представляет собой технологию для быстрого экономичного синтеза большого числа олигонуклеотидов. Эта технология позволяет получать все синтетические ДНК, необходимые для сборки гена, за один день. В соответствии с фиг. 1 обработанные нуклеозидами стекла с контролируемым размером пор помещают в отдельные ячейки синтеза, состоящие из тефлоновых цилиндров с пористыми концами, что позволяет жидкости протекать через стопку ячеек. Одновременное добавление A, G, C или T к цепям ДНК, присоединенным к CPG и остающимся внутри ячеек, осуществляется за счет последовательного потока реагентов через колонку за счет фосфорамидитного способа (L.I. McBride and М.Н. Caruthers, Tetrahedron Lett. , 24, 245-248 (1983)). После завершения каждого из циклов химических реакций ячейки сортируют по различным колонкам для обеспечения синтеза различных нуклеотидных последовательностей внутри каждой из ячеек. Изображено последовательное положение одной ячейки (заштрихована) в четырех циклах, что должно привести к добавлению AGCT к растущей цепи ДНК, содержащейся в ней.
Второй технологической разработкой, которая облегчает синтез, является создание средств для постадийного присоединения синтетических олигонуклеотидов на твердофазный носитель для получения гена. На фиг. 2 представлена конструкция целевого гена, который собирают из набора перекрывающихся комплементарных олигонуклеотидов. Сборку начинают со связывания одного конца нуклеотида с твердофазным носителем. 5'-фосфорилированные олигонуклеотиды добавляют последовательно (с молярным избытком) к связанной с носителем нити. На каждой стадии сборки гена несвязанную ДНК вымывают перед тем, как проводят следующую реакцию отжига. После завершения сборки все обрабатывают ДНК лигазой для лигирования одноцепочечных разрывов, затем ген отделяют от носителя, за счет расщепления по уникальному рестрикционному сайту, содержащемуся в связанном с носителем олигонуклеотиде. Освобожденную ДНК лигируют в подходящий вектор для секвенирования и экспрессии.
Множество вариаций существует для сред хозяина вектора при экспрессии синтетического гена, которая приводит к получению закодированного протеина. Они обсуждаются подробно в Methods Enzymology, Yol 152, 1987, Academic Press. Короче, специальные векторы экспрессии доступны для включения в бактериальные, грибковые, животные или растительные хозяева. Наиболее часто используют Escherichia coli для экспрессии "чужеродных" генов. Другим популярным хозяином для экспрессии являются дрожжи Saccharomyces cerevisial. Как было указано ранее, если представляющий интерес клонированный ген синтезирован химическим путем, предлагается использование оптимального кодона для хозяина целевой экспрессии для того, чтобы повысить уровень экспрессии сконструированного гена. Большинство векторов экспрессии содержат генетические контролирующие элементы, расположенные по соседству с сайтом клонирования, который обеспечивает высокие уровни экспрессии гена. Индуцируемые промоторы векторов экспрессии обычно получают из бактерий (например, тас, тгр) или вирусов (например, лямбда, SV 40). Элементы "сигнальной последовательности иногда включаются в вектор для управления транспортом генного продукта из клеток хозяина. Элементы сигнальной последовательности могут облегчить очистку и снизить протеолитическое разложение. В случае синтетических генов любой целевой генетический контрольный элемент может быть включен в собираемый дуплекс ДНК. Некоторые векторы экспрессии содержат кодирующую последовательность, следующую сразу за сайтом клонирования, так что вставка в рамку чужой кодирующей последовательности может быть обеспечена внутри собранной последовательности в случае синтетических генов. Продуцирование гена как протеина слияния обеспечивает ряд преимуществ, включая повышение экспрессии, более высокую стабильность, быструю афинную очистку с использованием связанного с носителем лиганда, который связывается с дополнительными полипептидными компонентами, и обеспечение направленного связывания генного продукта с клеткой (например, связывания с клетками, обладающими на своей поверхности рецепторами для дополнительных полипептидных компонентов). Для дальнейшего развития терапии на основе гелонина может быть использована генная инженерия, включающая эту последнюю особенность.
При клонировании и экспрессии ДНК последовательностей, кодирующих растительный токсин гелонин, можно использовать широкий круг векторов. Они включают, например, векторы, состоящие из сегментов хромосомных, нехромосомных и синтетических ДНК последовательностей, таких как различные известные производные SV 40, плазмиды из E.coli, включая col E1, pCR1, pBR322, pMB9 и их производные, широкий круг плазмид хозяев (например RP4, фаговые ДНК), например, различные производные фага лямбда, (например, NM 989) и другие ДНК фагов (например, M13 и нитевидные однонитевые ДНК фагов, плазмиды дрожжей, такие как 2mu плазмида или ее производные, и векторы, полученные из комбинаций плазмид и ДНК фагов, такие, как плазмиды, которые были модифицированы для использования ДНК фага или других последовательностей, контролирующих экспрессию.
Для каждого специфического клонирования или вектора экспрессии могут быть выбраны различные сайты для вставки ДНК последовательностей настоящего изобретения. Эти сайты обычно обозначаются по рестрикционным эндонуклеазам, которые разрезают их и которые хорошо известны специалистам. Хорошо известны также различные способы встраивания ДНК последовательностей в эти сайты для получения рекомбинатных молекул ДНК. Они включают, например, наращивание dG-dC или dA-dT, прямое лигирование, синтетические линкеры, реакции репарации, связанные с экзонуклеазой и полимеразой с последуюущим лигированием, или удлинение ДНК нитей ДНК полимеразой и соответствующим однонитевым темплатом с последующим лигированием. Естественно, следует учитывать, что векторы клонирования или экспрессии, пригодные для настоящего изобретения, не должны иметь сайты рестрикционных эндонуклеаз для включения выбранного ДНК-фрагмента. Вместо этого носитель можно присоединить к фрагменту другими способами.
Для экспрессии ДНК последовательности настоящего изобретения эти ДНК-последовательности оперативно связаны с одной или более из последовательностей, управляющих экспрессией, в векторе экспрессии. Такое оперативное связывание, которое можно осуществить до или после того, как выбранную ДНК-последовательность включают в вектор клонирования, обеспечивает экспрессию контролирующих последовательностей и промотирование экспрессии встроенной ДНК-последовательности.
Для экспрессии ДНК-последовательности настоящего изобретения можно использовать в этих векторах любую из разнообразных последовательностей регулирования экспрессии. Такие подходящие регулирующие экспрессию последовательности включают, например, ранний и поздний промотор SV 40, lac систему, trp систему, TAC или TRC систему, основные участки оператора и промотора фага лямбда, контрольные участки fd протеина оболочки, промотор для 3-фосфоглицераткиназы или других гликолитических промоторов, промоторы кислотной фосфатазы, например, Pho5, промоторы дрожжевых факторов альфа-спаривания и другие последовательности, которые, как известно, контролируют экспрессию генов прокариотных и эукариотных клеток или их вирусов, и различных их комбинаций. В клетках млекопитающих дополнительно возможно амплифицировать фрагменты экспрессии, за счет связывания гена с кодирующим дегидрофолатредуктазу и проводя селекцию по клеткам яичника китайского хомяка.
Вектор или носитель экспрессии, конкретные сайты, выбранные для встраивания выбранного ДНК-фрагмента и последовательность, контролирующая экспрессию, используемые в настоящем изобретении, определяют различными факторами. Так например, эти факторы включают ряд сайтов, соответствующих конкретному рестрикционному энзиму, размер подлежащего экспрессии протеина, такие характеристики экспрессии, как расположение кодонов начала и окончания относительно последовательности вектора. Другие факторы должны быть очевидны специалистам. Выбор вектора, экспрессия контрольной последовательности и встраивание сайта для конкретной последовательности протеина ингибитора фосфолипазы определяют соотношением этих факторов, причем не всегда этот выбор является одинаково эффективным в конкретном случае.
Рекомбинатную ДНК-молекулу, содержащую целевой ген, оперативно связанный с последовательностью, контролирующей экспрессию, можно затем использовать для трансформации широкого круга соответствующих хозяев с тем, чтобы обеспечить экспрессию этим хозяевам (трансформантам) гена или его фрагмента и получать полипептид или его часть, которую кодирует гибридная ДНК.
Для получения антигенов и ДНК-последовательностей настоящего изобретения можно использовать широкий круг хозяев. Эти хозяева включают, например, бактерии, такие, как E. coli, Bacillus and Streptomyces, грибки, такие, как дрожжи, животных, таких как CHO клетки, и клетки растений в культуре тканей. Выбор подходящего хозяина определяется целым рядом факторов, которые известны специалистам. Они включают, например, совместимость с выбранным вектором, токсичность ко-продуктов, простоту выделения целевого пептида, характеристики экспрессии, биобезопасность и стоимость. Исходя из одного только из этих факторов нельзя сделать абсолютного выбора хозяина для конкретной рекомбинантной ДНК молекулы или полипептида. Вместо этого следует рассматривать комплекс этих факторов, учитывая, что не все хозяева могут оказаться одинаково эффективными для экспрессии конкретной рекомбинатнтной молекулы ДНК.
Как было показано ранее, следует учитывать, что ДНК-последовательности, которые встраивают по выбранному сайту носителя клонирования или экспрессии, могут содержать нуклеотиды, которые не являются частью реального гена, кодирующего целевой полипептид, или могут включать только фрагмент полного гена этого протеина. Необходимо только, чтобы независимо от используемой ДНК-последовательности, трансформированный хозяин продуцировал протеин гелонин или полипептид с практически теми же функциональными активностями, что и гелонин. Так например, ДНК-последовательности настоящего изобретения можно слить в ту же самую считывающую рамку в векторе экспрессии, в части ДНК-последовательности, кодирующей по крайней мере один прокариотный или эукариотный протеин, или ДНК-последовательности, кодирующей по крайней мере одну эукариотную или прокариотную сигнальную последовательность или их сочетания. Такие конструкции могут служить при экспрессии нужной последовательности ДНК, для улучшения очистки или обеспечения секреции и, предпочтительно, вызревания целевого полипептида из клеток хозяина. В другом варианте ДНК-последовательность может включать ATG стартовый кодон, один или вместе с другими кодонами, слитый непосредственно с последовательностью, кодирующей первую аминокислоту целевого полипептида. Такие конструкции обеспечивают получение, например, метионильного или других пептидильных полипептидов, то есть часть изобретения. Затем N-терминальный метионин или пептид можно расщепить интра- или экстра-клеточно различными известными способами или можно полипептид использовать вместе с метионином или другим слитым и присоединенным к нему, в композициях и способах настоящего изобретения.
Примеры
Синтез и сборка гена гелонина
Пример 1. Связывание 5'-биотинилированных олигонуклеотидов с латексными микросферами, покрытыми стрептавидином.
Образец 0,2 мл DYNABEADS M280 (Denal Corp.) помещают в ампулу Эппендорфа емкостью 1,5 мл. Ампулу выдерживают над магнитной пластиной (Advanced maghetics, Ync) в течение нескольких минут для того, чтобы вызвать осаждение латексных микросфер с одной стороны, а затем жидкость сливают. Шарики дважды промывают 0,2 мл отжигающего буфера (состав приводится далее) при комнатной температуре, затем повторно суспендируют в 0,2 мл отжигающего буфера.
К суспензии шариков добавляют 1 нмоль 5'-биотинилированного олигонуклеотида. Спустя 30 минут, при комнатной температуре шарики дважды промывают 0,2 мл отжигающего буфера и снова суспендируют в 0,2 мл отжигающего буфера. Спектрофотометрический анализ несвязанного олигонуклеотида в промывках показывает, что приблизительно 300 пмолей олигонуклеотида связано с шариками.
Пример 2. Цикл отжига/промывки повторяют для дополнения каждого последующего нуклеотида.
Перед тем, как использовать в сборке гена, олигонуклеотиды очищают с помощью электрофореза на полиакриламидном геле и ферментативно, 5'-фосфорилируют, используя T4 полинуклеотидкиназу.
К 150 пмолям связанного с носителем олигонуклеотида добавляют 750 пмолей перекрывающего комплементарно олигонуклеотида, и отжиг ведут в 0,10 мл 50 мМ натрийфосфатного буфера, pH 7,5, 1M NaCl (буфер отжига) в течение 5 минут при 45oC, затем смесь охлаждают до комнатной температуры в течение 7 минут. Затем шарики дважды промывают 0,2 мл того же буфера при комнатной температуре. Этот цикл повторяют до тех пор, пока не добавляют последний олигонуклеотид в сборке.
Пример 3. Лигирование продукта и освобождение от носителя за счет переваривания рестрикционным энзимом.
После завершения сборки шарики промывают и снова суспендируют в 0,04 мл лигазного буфера. После добавления 0,005 мл ДНК лигазы (New England Biolabs, с высокой специфической активностью) эту смесь инкубируют при комнатной температуре в течение 2 часов, затем промывают и снова суспендируют в 0,04 мл рестрикционного переваривающего буфера. После добавления 10 единиц рестрикционной экдонуклеазы EcoR1, смесь инкубируют при 37oC в течение 90 минут. Жидкость сливают, а освобожденную ДНК осаждают этанолом и повторно суспендируют в 0,01 мл лигазного буфера.
Пример 4. Сборка гена гелонина.
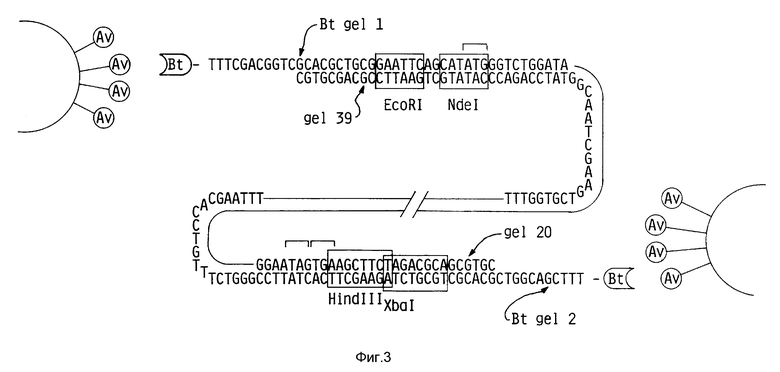
Этот ген собирают с обоих направлений, как показано на фиг. 3 и фиг. 4. Олигонуклеотидные последовательности представлены на фиг. 5. Сборку 5'-конца гена (приблизительно 500 bp N-терминального кодирующего участка) начинают со связанного с носителем Btgel1, и олигонуклеотиды добавляют в следующем порядке (Для каждого проводят цикл отжиг/промывка): gel39, gel1, gel38, gel2, gel37, gel3, gel36, gel4, gel35, gel5, gel34, gel6, gel33, gel7, gel32, gel8, gel31, gel9, gel30, gel10, gel29, gel11, gel28, gel12, gel27.
Сборку 3'-конца гена (приблизительно 300 bp C-терминального кодирующего участка) начинают со связанного с носителем Btgel2, и олигонуклеотиды добавляют в следующем порядке: gel20, gel19, gel18, gel22, gel17, gel23, gel16, gel24, gel15, gel25, gel14, gel26, gel13.
В соответствии с фиг. 3 5'-конец гена (N-терминальный кодирующий участок) освобождают от носителя, переваривая рестрикционной эндонуклеазной EcoR1, а 3'-конец гена (C-терминальный кодирующий участок) освобождают от носителя, переваривая рестрикционной эндонуклеазой Hind111. В соответствии с фиг. 4 два генных фрагмента, содержащие комплементарные по 20 пар оснований хвосты в олигонуклеотидах gel27 и gel13, отжигают вместе, а затем лигируют до получения интактного гена.
Пример 5. Клонирование синтетического гена гелонина.
Законченный ДНК продукт лигируют M13mp19RF ДНК, которую расщепили EcoR1 и Hind111, по стандартной методике, описанной в Molecular Cloning: Лабораторный справочник, EF. Sambrook et. al, 1989.
Пример 6. Секвенирование синтетического гена гелонина.
Последовательность синтетического гена в M13mp19 подтверждают дидеокси-секвенированием. В клонированном синтетическом гене обнаружены две мутации, обе с 5'-конца (N-терминальный кодирующий участок).
Пример 7. Сайт-направленный мутагенез для исправления мутаций в клонированном синтетическом гене.
Проводят олигонуклеотид-направленный мутагенез для исправления двух мутаций внутри гена гелонина по способу, прилагаемому с ин витро набором реагентов мутагенеза (Amersham Corp.).
Пример 8. Субклонирование синтетического гена в вектор экспрессии.
Синтетический гелониновый ген отщепляют от M13mp19 вектора под действием EcoR1 и Hind111, и содержащий ген фрагмент очищают на агарозном геле электрофоретически, а затем лигируют в EcoR1/Hind111-расщепленный вектор экспрессии pKK223-3 (Pharmacia).
Пример 9. Анализ экспрессии синтетического гена гелонина в Е.coli.
50 мл культуры E. coli JM105, содержащей синтетический ген гелонина, клонированный в pKK223-3, выращивают, индуцируют IPTG и проводят лизис до получения неочищенного экстракта. Этот экстракт анализируют с помощью электрофореза на акриламидном геле (наряду с контрольным экстрактом, полученным из клеток хозяина, содержащих вектор экспрессии без каких-либо вставок). Вестернблоттинг и функциональный анализ гелонина также проводят для подтверждения того, что протеин экспрессируется и активен.
И в заключение следует сказать, что настоящее изобретение и раскрытые здесь его варианты хорошо приспособлены для осуществления целей и получения указанных результатов. В способ и аппаратуру могут быть внесены некоторые изменения, не выходя за рамки духа и объема настоящего изобретения. Следует учитывать, что изменения возможны и что каждый элемент или стадия, указанные далее в следующих пунктах формулы изобретения, следует рассматривать как эквивалентные элементы или стадии для осуществления практически тех же целей и практически теми же способами. Изобретение следует рассматривать широко, независимо от того, в каком виде будут использованы его идеи. Поэтому настоящее изобретение хорошо адаптировано для решения задач и получения результатов и преимуществ, указанных ранее, а также других ему присущих.




Изобретение относится к биотехнологии. Получен синтетический ген, кодирующий белок гелонина с установленной нуклеотидной последовательностью SEQ ID NI, представленной в описании. Способ получения включает стадию отбора последовательности фрагмента ДНК, стадию синтеза, который включает разделение фрагмента ДНК на два перекрывающихся набора однонитевых фрагментов олигонуклеотидов. Синтезированные химически олигонуклеотиды выбирают из группы, состоящей из Btgel 1, Btgel 2 и gel 1 - gel 39. Затем их очищают, фосфорилируют на 5'-конце. Отжиг нитей олигонуклеотидов вместе с формированием дуплекса ДНК приводит к образованию синтетического гена. Получен также рекомбинантный штамм E.coli JM 105, трансформированный вектором экспрессии, обеспечивающим экспрессию белка гелонина. Вектор содержит фрагмент ДНК с последовательностью SEQ ID NI, лигированный в EcoRI/Hind III расщепленным вектором рКК 223-3. Данная рекомбинантная технология позволяет получить богатый репродуцирующий источник гелонина, являющийся иммунотоксическим агентом для лечения опухолей. 4 с. п.ф-лы, 5 ил.
| Gene | |||
| Кузнечная нефтяная печь с форсункой | 1917 |
|
SU1987A1 |
| Biochemical Journal | |||
| Устройство для видения на расстоянии | 1915 |
|
SU1982A1 |
| Procceding of the Natural Academy of Sciencens | |||
| Гребенчатая передача | 1916 |
|
SU1983A1 |
| Nature | |||
| Механизм для сообщения поршню рабочего цилиндра возвратно-поступательного движения | 1918 |
|
SU1989A1 |

