Изобретение относится к рекомбинантному полипептиду и к нуклеотидной последовательности, кодирующей полипептид, к системе экспрессии, способной экспрессировать этот полипептид, а также к фармацевтическим и косметическим композициям, содержащим этот полипептид, и к использованию полипептида для различных косметических или терапевтических целей.
Краткое описание изобретения
Кожа, как орган, представляет интерес с биологической, медицинской и косметической точек зрения. Существует множество кожных заболеваний, которые являются либо органоспецифическими, как например, псориаз и экземы, либо проявлениями таких общих заболеваний, как общие аллергические реакции. Тот факт, что существуют кожеспецифические заболевания, можно рассматривать как доказательство существования молекулярных механизмов, которые уникальны для кожи. Аналогично, исследования кожеспецифических молекулярных процессов имеют важное значение для понимания и лечения кожных заболеваний. По-видимому, разумно предположить, что некоторые из этих процессов, так или иначе, связаны с наиболее специализированными функциями кожи, то есть, с созданием физико-химического барьера между организмом и внешним окружением. Физико-химический кожный барьер локализован в самом верхнем слое кожи, роговичном слое (Stratum corneum).
Роговичный слой является наиболее специализированной структурой кожи. Он является конечным продуктом процессов дифференциации эпидермиса, то есть, расслоенным чешуйчатым эпителием, который представляет самую верхнюю часть кожи. Большинство клеток эпидермиса состоит из кератиноцитов в различных стадиях дифференциации. Наиболее глубоко расположенные кератиноциты, базальные клетки, находятся на базальной мембране в контакте с дермой /собственно кожей/, которая представляет соединительную ткань кожи, и являются единственными кератиноцитами, обладающими способностью к делению. Часть базальных клеток непрерывно покидает базальную мембрану и претерпевает процесс дифференциации, в результате которого клетки становятся строительными блоками роговичного слоя. В этом процессе кератиноциты претерпевают ряд адаптационных изменений. Происходит увеличение содержания цитоскелета, состоящего из эпидермис-специфических цитокератинов. Промежуточные нити соприкасающихся клеток соединены с функциональными фрагментами за счет возрастающего количества десмосом. Наиболее резкие изменения происходят при переходе от самого верхнего слоя живых клеток, stratum granulosum, к мертвому роговичному слою, stratum corneum, в процессе, который обычно носит название кератинизация. Ковалентно связанные протеины расположены близко к внутренней части плазменной мембраны, образуя очень устойчивую клеточную оболочку. Кроме того, богатое липидами вещество, образующееся в органеллах кератиноцит-специфических клеток, секретируется во внеклеточное пространство, и, образуя липидные пластинки, которые окружают клетки роговичного слоя, составляет барьер, проницаемый для гидрофильных веществ. И наконец, все межклеточные структуры, за исключением плотно упакованных цитокератиновых нитей, исчезают.
Клетки Stratum corneum, корнеоциты, являются, таким образом, мертвыми клетками. Это означает, что регуляция различных процессов в роговичном слое должна быть результатом "программирования" на той стадии, когда кератиноциты все еще являются живыми клетками. Превращения эпидермиса, которые обычно протекают в течение примерно четырех недель, причем клетки являются частью роговичного слоя примерно в течение двух недель, завершаются отпадением клеток с поверхности кожи в процессе шелушения /десквамации/. Этот процесс является примером "программирования" роговичного слоя. Необходимым условием для функционирования роговичного слоя в качестве физико-химического барьера является требование того, чтобы отдельные его клетки удерживались вместе за счет механически устойчивых структур, то есть десмосом. Деградация десмосом, которая необходима для десквамации, должна регулироваться таким образом, чтобы обеспечить такое шелушение с поверхности кожи, которое уравновешивалось бы образованием нового роговичного слоя, без нарушения барьерных функций ткани.
Нарушение кератинизации
В основе большого числа патологических состояний кожи различной степени тяжести лежит нарушение процесса кератинизации. При псориазе наблюдается, помимо типичного хронического воспаления, избыточное продуцирование незрелых клеток роговичного слоя, что приводит к типичному для этого заболевания шелушению. Существует группа врожденных заболеваний кожи, которые характеризуются утолщением роговичного слоя, что приводит к образованию "рыбьей чешуи" так называемого ихтиоза. У некоторых индивидуумов, страдающих ихтиозом, наблюдается пониженная скорость десквамации. Хотя и менее серьезное нежели ихтиноз, заболевание "сухая кожа" /ксеродерма/ также характеризуется роговичным слоем, с которого отшелушиваются корнеоциты, не так, как в обычных условиях, в виде отдельных клеток или мелких агрегатов клеток, но в виде крупных, макроскопических чешуек; это заболевание весьма обычно для старческого возраста, а также среди атопических индивидуумов с пониженной устойчивостью к кожным поражениям и склонностью к развитию характерных форм эндогенной экземы. В случае акне заболеваний наблюдается нарушенная кератинизация в протоках сальных желез, что приводит к образованию комедонов в закупорке желез. Образование комедонов прогрессирует и, как считают, приводит к образованию воспалительных угревых язвочек. Протеолитические энзимы участвуют в кератинизации.
Существует несколько стадий в процессе кератинизации и во время превращений роговичного слоя, где протеолитические энзимы, по-видимому, играют важную роль. Естественно, исчезновение всех внутриклеточных структур, за исключением цитокератиновых нитей, наблюдающееся в процессе перехода от живых до ороговевших эпидермальных слоев, должно включать протеолиз. Превращение профилаггрина в филаггрин, протеин, который, как считают, функционирует в специальном типе аггрегаций нитей цитокератина в процессе кератинизации, может катализироваться специфической протеиназой. В роговичном слое филаггрин далее разлагается до низкомолекулярных компонентов, которые, вероятно, важны как "природные увлажнители". Кроме того, существуют протеолитические модификации цитокератиновых полипептидов в процессе кератинизации. И наконец, протеолитические акты, по-видимому, играют решающую роль в деградации межклеточных когезивных структур в роговичном слое в процессах, которые обычно приводят к десквамации.
Когезия и десквамация клеток роговичного слоя
/Stratum corneum/
Роль десмосом
Межклеточная когезия в роговичном слое, так же как и в живых частях эпидермиса, в значительной степени осуществляется за счет десмосом. Десмосома состоит из двух симметричных половинок, каждая из которых образована двумя соприкасающимися клетками. Каждая половинка десмосомы имеет одну внутриклеточную часть, связанную с нитями цитокератина, и одну часть, состоящую из гликопротеинов, закрепленных внутриклеточно и с трансмембранной и внеклеточной частями. Внеклеточные части этих протеинов, десмоглеины, представляют собой адгезивные молекулы, и за счет их взаимодействия друг с другом во внеклеточном пространстве образуется когезивная структура. Деградация десмосом, по-видимому, протекает несколько другими путями в роговичном слое ладоней и подошв /стоп/ по сравнению с роговичным слоем, не относящимся к ладоням-подошвам. В последних тканях около 85% десмосом исчезает вскоре после того, как клетки становятся полностью ороговевшими. Оставшиеся десмосомы, которые расположены, предпочтительно, на ворсинчатых краях чрезвычайно уплощенных клеток, по-видимому, остаются интактными вплоть до того уровня, где происходит десквамация. В роговичном слое, который не относится к ладоням и подошвам, корнеоциты гораздо менее уплощены, и не происходит деградации десмосом в более глубоких слоях ткани. В обоих типах тканей десквамация связана с деградацией десмосом. В случае ихтиозной кожи, так же как и в случае "сухой кожи", число десмосом в поверхностных слоях роговичного слоя, как было показано, повышается.
Межклеточные липиды роговичного слоя
Различия в деградации десмосом, относящихся к ладоням и подошвам роговичных слоев и не относящихся к ним, могут быть связаны с различными количествами внеклеточных липидов в этих двух типах тканей. Содержание липидов значительно выше в роговичных слоях, не относящихся к ладоням-подошвам. В результате эффективность этой ткани в качестве барьера, проницаемого для воды и других гидрофильных веществ, гораздо выше проницаемости роговичного слоя ладоней и стоп. Так как десмосомы занимают значительный объем и так как интактные десмосомы предотвращают расширение межклеточного пространства, именно деградация десмосом может быть тем механизмом, за счет которого большее внеклеточное пространство становится доступным для липидов. Внеклеточные липиды роговичного слоя связаны с десквамацией также и некоторыми другими путями. Так как они составляют основную часть внеклеточного пространства, можно ожидать, что они оказывают значительное влияние на активность энзимов, которые действуют в этом месте, например, энзимов, ответственных за деградацию десмосом. Действительно, различные нарушения метаболизма липидов, как было показано, по-видимому, вызывают некоторые типы ихтиоза. По-видимому, также, сами липиды в некоторой степени вносят вклад в клеточную когезию роговичного слоя. Более того, секреция липидов в межклеточное пространство роговичного слоя, по-видимому, связана с секрецией также и ряда энзимов. Предшественники липидов хранятся в верхних живых кератиноцитах в специфических органеллах. Эти так называемые ламелларные /пластинчатые/ тела, как было показано, содержат ряд гидролитических энзимов. Таким образом предполагается, что энзим, ответственный за деградацию десмосом в роговичном слое, синтезируется, возможно в неактивной про-форме, кератиноцитами, хранится в ламелларных телах и секретируется в межклеточное пространство роговичного слоя в процессе кератинизации, где он может быть активирован, и его активность далее регулируется, наряду с другими факторами, составом внеклеточных липидов.
Нарушений клеточной когезии в жизнеспособном эпидермисе
Существует ряд кожных заболеваний, при которых нарушается когезия между кератиноцитами в неороговевших жизнеспособных эпидермальных слоях. Эти заболевания характеризуются феноменом, именуемым акантолизом, то есть разрушением десмосомных контактов между кератиноцитами, которые в остальных проявлениях кажутся нормальными. Этот процесс, по-видимому, осуществляется за счет протеиназ, которые до сих пор не были идентифицированы. Аквантолиз, в своих острых формах, приводит к образованию волдырей и является характеристикой автоиммунных заболеваний pemphigus vulgaris и pemphigus foliaceus наследственного доброкачественного familiar pemphigus /болезнь Hayley-Hayley's/ и наследственного dyskeratosis follicularis /болезнь Darier's/.
Эпидермис принимает активное участие в иммунологических и воспалительных реакциях
В дополнение к своим функциям в качестве создания физико-химического барьера между внутренностями организма и внешней средой, эпидермис также функционирует в качестве активного иммунологического барьера. Кератиноциты обладают способностью продуцировать, а также регулировать на большое число цитокинов и других медиаторов воспалений, за счет которых они коммутируют и взаимодействуют с клетками иммунологической и воспалительной систем. Это имеет огромное значение при защите хозяина от микробной инфекции и в заживлении ран. Современные исследования показали также, что кератиноциты, по-видимому, принимают активное участие во многих воспалительных кожных заболеваниях, таких, как псориаз и экземы.
Эпидермальные протеиназы могут играть важную роль в воспалительных процессах
Одним из цитокинов, продуцируемых кератиноцитами, является интерлейкин 1 /Il-1/. Il-1 существует в двух формах, Il-1 α и Il-1 β , причем обе они присутствуют в эпидермисе. Если Il-1 α после синтеза является полностью активным, Il-1 β синтезируется в виде неактивной 31 кД про-формы. Про- Il-1 β превращается в активную 17 кД форму за счет специфической протеиназы, присутствующей, например, в моноцитах, но до сих пор не обнаруженной в нормальном эпидермисе. Ряд сериновых протеиназ со специфичностью к химотрипсин-подобному субстрату /химотрипсин поджелудочной железы и катепсин G нейтрофильных гранулоцитов/ может, однако, служить в качестве активаторов про- Il-1 β.
Как предложил Норрис /Norris 1990/, протеолитические энзимы, присутствующие в межклеточном пространстве роговичного слоя, могут в определенных условиях оказаться способными превращать неактивные формы цитокинов в активные формы. Особый интерес в этом контексте представляет биологически неактивный про-интерлейкин-1 β , который, как было показано, продуцируется кератиноцитами /Migutani et at., 1991/. До сих пор не было обнаружено энзимов со способностью превращать про-интерлейкин-1 β в активный интерлейкин-1 β. Однако, так как химотрипсин и катепсин G /химотрипсиноподобный энзим/ обладают способностью катализировать превращение неактивного 31 кД рекомбинантного про-интерлейкина 1 β в полностью активную 17 кД форму, возможно, что и эпидермальные химотрипсин-подобные энзимы могут катализировать это превращение.
Подробное описание изобретения
Настоящее изобретение относится к энзиму, который был назван химотрипсиновым энзимом strarum corneum /роговичного слоя// SCCE/, который рассматривается как энзим, ответственный за деградацию десмосом в роговичном слое. Доказательство, представленное в примере 1, демонстрирует, что отшелушивание клеток с поверхности ороговевших поверхностных слоев кожи включает деградацию протеинов десмосом, и что ответственным за это энзимом, по-видимому, является химотрипсино-подобная сериновая протеиназа, которую можно ингибировать ионами цинка.
В примере 2 описывается открытие химотрипсинового энзима Stratum corneum /SCCE/, протеиназы, которая удовлетворяет критериям ответственности за деградацию внутриклеточных когезивных структур в роговичном слое ин витро и, возможно, также ин виво. Пример 3 описывает частичную характеризацию активности химотрипсинового энзима stratum corneum /SCCE/ с помощью хромогенных субстратов.
Полученные результаты демонстрируют, что SCCE отличается энзимологически от остальных химотрипсиновых протеиназ. Характер ингибирования и способность разлагать два различных субстрата трипсиноподобных энзимов существенно отличается для SCCE по сравнению с бычьим химотрипсином и катепсином G человека. По-видимому, SCCE также отличается от химазы тучных клеток человека. Последний энзим катализирует деградацию Suc-Ala-Ala-Pro-Phe-pNa эффективно /см. Schwartg et al., 1987, и Schelter et al., 1989/ - чего не делает SCCE- и не ингибируется апротинином SBTJ /Shelter et al., 1983 и Wintroub et al. 1986/, тогда как SCCE ингибируется.
В примере 4 описана частичная очистка SCCE из KCl экстрактов корнеоцитов с помощью афинной хроматографии на несолюбилизированном ингибиторе трипсина сои /SBTJ/ и определение N-терминальной аминокислотной последовательности SCCE. Для невосстановленных образцов были получены хорошие выходы на стадиях 1-6, но снизились до 0 на стадиях 7 и 9, и выходы на последующих стадиях были заметно снижены. Для восстановленных образцов на стадиях 7 и 9 не детектировались производные аминокислот, но на последующих стадиях, на которых детектировались производные, сильного падения выходов не наблюдалось. Эти результаты дают возможность предположить, что в положениях 7 и 9 существуют цистены. Однако, не представилось возможности определить карбоксиметилированный цистеин на стадиях 7 и 9 после восстановительной обработки иодоуксусной кислотой /100 мМ/. Полученная последовательность /фиг. 13, послед. ID N 3/ была идентична как для восстановленного, так и для невосстановленного образцов.
Пример 5 описывает получение SCCE-специфических моноклональных антител и поликлональных SCCE-специфических антител цыпленка и кролика, а также иммуногистохимические исследования с моноклональными антителами.
Пример 6 описывает клонирование и секвенирование кДНК, кодирующей SCCE человека. Вначале, кДНК библиотеку, полученную на мРНК, полученной из эпидермльных кератиноцитов взрослого человека, скринируют анти-SCCE кроличьими поликлональными антителами. Одно из этих антител дает высокий фоновый сигнал, и его исключают из интенсивных исследований за счет скринирования на ранней стадии. Используя другое поликлональное антитело /Д-5/, ряд иммунореактивных бляшек обогащают как превосхищающие истинно положительные бляшки. Не наблюдалось реакционноспособности с моноклональными антителами мoAb 4 и мoAb 9 ни для одной бляшки. Интенсивная характеристика рестрикционных энзимов и характеризация PCR одиннадцати выделенных бляшек не выявила никакого сходства между различными бляшками. На основании невозможно получить "отпечатки пальцев" вероятной кДНК последовательности SCCE, стратегию пришлось модифицировать.
Несмотря на предпочтительное детектирование SCCE иммунореактивного материала в супрабазальных кератиноцитах, для скринирования кДНК SCCE использовали кДНК библиотеку, полученную из культивируемых кератиноцитов человека. Можно ожидать, что такая библиотека должна содержать кДНК только из базальных кератиноцитов. Эта попытка была основана на наблюдении слабого, но, вероятно, значительного иммуноокрашивания с использованием SCCE моноклональных антител также из базальных кератиноцитов.
Бляшки были скринированы с использованием синтетического 17-мерного олигонуклеотидного зонда, сконструированного на основании наиболее надежной части аминокислотной последовательности, Ile-Ile-Asp-Gly-Ala-Pro /последовательность ID N 10 aa 1-aa 6/ экспериментально определенной аминотерминальной последовательности нативного SCCE энзима, как описано в примере 4. Из-за неопределенности внутри экспериментально определенной аминокислотной последовательности, этот гексапептид, как считают, представляет одну из наиболее надежных частей. Кроме того, возможные кодоны, кодирующие этот гексапептид, приводят к наименьшей из возможных дегенерации ДНК зонда. Более длинная экспериментально определенная последовательность Gln-Val-Ala-Leu-Leu-Ser-Gly-Asn-Gln-Leu /последовательность ID N 3, aa 15-aa 24/ была исключена из-за высокой степени дегенерации последовательности, кодирующей эту пептидную последовательность. Четырнадцать позитивных бляшек были идентифицированы при первичном скринировании. Эти позитивные бляшки были снова скринированы с использованием тех же зонда и метода, которые были описаны ранее. После процедуры повторного скринирования были идентифицированы две позитивные бляшки. Эти две отобранные бляшки были еще раз очищены, а размер вставок был определен с помощью PCR с использованием SYМ 1600 и SYM 1601, которые оказались комплементарны двум плечам фага, в качестве праймеров, и выделенных фагов в качестве матриц. Этот клонированный фрагмент был подвергнут частичному анализу последовательности.
Трансляция полученной ДНК последовательности привела к аминокислотной последовательности, которая была гомологична экспериментально определенной протеиновой последовательности. Однако, в этой последовательности не было трансляционного стартового кодона. Для выделения полной длины кДНК, полученный ДНК фрагмент выделили на агарозном геле и использовали в качестве зонда, обеспечив гибридизацию в жестких условиях. Для получения полной длины кДНК, библиотеку кДНК рескринировали дважды с помощью зонда, используя те же методы, которые были описаны ранее, за исключением того, что гибридизацию вели в жестких условиях при 65oC. Эти эксперименты привели к идентификации и выделению 45 отдельных позитивных бляшек, которые были вначале скринированы в PCR анализе с использованием SYM 1600 или SYM 1601 в сочетании с SYM 3208 в качестве PCR праймеров для идентификации бляшек, содержащих полную 5' открытую считывающую рамку.
После дальнейшего скринирования и секвенирования полученные PCR амплифицированные фрагменты, полученные из этих фагов, были клонированы, как было подробно описано в примере 6, и полученные результаты указывают, что один из фагов 205.2.1 содержит полной длины вставку. Была определена полная нуклеотидная последовательность кДНК фрагмента. Нуклеотидная последовательность /последовательность ID N 1/ содержит открытую считывающую рамку, достаточную для кодирования полной аминокислотной последовательности протеина SCCE предшественника, состоящего из 253 аминокислот, включая сигнальный пептид и преполипептид /последовательность ID N 2/.
Пример 7 описывает определение в эпидермисе человека двух SCCE мРНК-видов, которые способны гибридизоваться с кДНК зондами, полученными на базе SCCE кДНК последовательности.
Пример 8 описывает экспрессию рекомбинантного SCCE в E.coli. Полученный результат показывает, что представляется возможность продуцировать рекомбинантный SCCE в бактериях.
Пример 9 описывает экспрессию рекомбинантного SCCE человека в клетки млекопитающих. Получают три протеина, которые демонстрируют реакцию со всеми жизнеспособными поликлональными антителами SCCE кролика и цыпленка, так же как и с депонированными моноклональными антителами. Рекомбинантные протеины, которые реакционноспособны по отношению к антителам, выработанным против нативного SCCE, демонстрируют кажущийся молекулярный вес, который примерно на 1 кД больше, чем у нативного очищенного SCCE человека. Рекомбинантный протеин не демонстрирует какой-либо протеолитической активности.
Выделение, активация и дальнейшая характеризация рекомбинантного SCCE описаны в примере 10. Неактивную проформу рекомбинантного SCCE можно активировать протеолитическим расщеплением трипсином или эндопептидазой LYS-C. Предполагается, что ряд других протеаз, которые расщепляют пепетид после основной аминокислоты, таких как эндопротеиназа LYS-C, папаин и плазмин, могут быть способны активировать про- SCCE в активный SCCE. Было обнаружено, что сигнальный пептид состоит из 22 аминокислот и основан на N-терминальной аминокислотной последовательности нативного активного SCCE, пропептида, состоящего из семи аминокислот. Более того, было показано, что полученный рекомбинантный SCCE существует в двух N-гликозилированных формах и одной негликозилированной форме, что аналогично результатам, полученным для активного нативного SCCE.
Под термином "stratum corneum" химотрипсиновый энзим /SCCE/" /химотрипсиновый энзим роговичного слоя/ или "полипептид, обладающий SCCE активностью" в широком смысле подразумевают сериновую протезу или ее проформу, которую можно активировать за счет протеолитического расщепления, причем указанный энзим в его активной форме ингибируется теми же самыми ингибиторами и точно таким же образом, что и спонтанная клеточная диссоциация, которую можно вызвать в модельной системе с образцами ороговевшего слоя кожи, инкубируемыми при нейтральных или близких к нейтральным pH при физиологических температурах.
Более конкретно, термин "полипептид, обладающий SCCE активностью" определяет таким образом полипептид, который отличается от химотрипсина и катепсина G и который в его активной форме способен разлагать MeO-Suc-Arg-Pro-Tyr-pNA /S= 2586/, причем это разложение полипептидом ингибируется апротинином, химостатином и сульфатом цинка практически по способу примеров 3 и 13, что проиллюстрировано на фиг. 9 и в таблице 5.
Еще более конкретно термин "полипептид, обладающий SCCE активностью" включает полипептид, который способен вызвать протеолитическое расщепление протеина десмосом - десмоглеина 1 во время ин витро инкубирования роговичного слоя пяточной части.
Такой полипептид обычно также реагирует с антителами, выработанными против нативного SCCE, который был выделен из экстракта диссоциированных клеток роговичного слоя пяточной поверхности. Примеры таких антител представляют поликлональные антитела, полученные как указано в 5.2, и моноклональные антитела TE4b и TE9b, полученные как указано в примере 5.1. Моноклональные антитела получают за счет гибридом TE4o и TE9o, депонированных в Европейской коллекции клеточных культур животных, Porton Down, Salisbury, Wiltshire SR 4 OJP, United Kingdom под регистрационными номерами ECACC 93061817 и EC ACC 93061816, соответственно, в соответствии с требованиями Будапештского соглашения.
Клонирование и секвенирование кДНК, кодирующей SCCE человека, описано в примере 6. Нуклеотидная последовательность, содержащая открытую считывающую рамку, достаточную для кодирования полной аминокислотной последовательности протеина предшественника SCCE, состоящая из 253 аминокислот, включая сигнальный пептид и преполипептид, была таким образом найдена. Нуклеотидная последовательность представлена как последовательность ID N 1, а полученная аминокислотная последовательность "химотрипсинового энзима роговичного слоя /SCCE/" представлена как последовательность ID N 2.
Под термином "про- SCCE или его аналог или вариант его" подразумевают полипептид, содержащий аминокислотную последовательность ID N 2 или его аналог или вариант указанной последовательности, который получают, когда нуклеотидная последовательность изобретения экспрессируется в подходящую систему экспрессии, и который после протеолитической активации приводит к образованию сериновой протеиназы, которую можно ингибировать теми же самыми ингибиторами, что и спонтанную клеточную диссоциацию, которую можно вызвать в модельных системах с образцами ороговевшего слоя кожи, инкубируемыми при нейтральных или почти нейтральных pH при физиологических температурах, то есть, около 37oC. Вообще, протеин будет реагировать с антителами, выработанными против очищенного нативного или рекомбинантного SCCE.
Под термином "SCCE или его аналог или вариант его" подразумевают полипептид, содержащий аминокислотную последовательность ID N 2 или аналог или вариант указанной последовательности, который продуцируется, когда нуклеотидная последовательность настоящего изобретения экспрессируется в подходящую систему экспрессии, и который является сериновой протеиназой, которую можно ингибировать теми же самыми ингибиторами, что и спонтанную диссоциацию клеток, которую можно индуцировать в модельных системах с образцами ороговевших слоев кожи, инкубируемых при нейтральных или почти нейтральных значениях pH при физиологических температурах, т.е. около 37oC. Вообще, протеин должен реагировать с антителами, вырабатываемыми против нативного или рекомбинантного SCCE.
Под термином "его аналог или вариант" подразумевают полипептид, последовательность которого в точности не соответствует аминокислотной последовательности ID N 2, но которая все еще сохраняет "SCCE активность", как определено ранее. Вообще, такие полипептиды представляют полипептиды, которые отличаются, например, до некоторой степени по составу аминокислот или включают пост-трансляционные модификации, например, гликозилирование или фосфорилирование, по сравнению с SCCE протеином, описанным в примерах.
Термин "аналог" или "вариант" используют в настоящем контексте для того, чтобы обозначить протеин или полипептид с аналогичным составом аминокислот, или последовательностью такой, как характеристическая последовательность аминокислот ID N 2, полученная из SCCE протеина, как указано в примере 6, позволяя небольшие вариации, которые изменяют аминокислотную последовательность, например, делеции, сайт-направленный мутагенез, вставки дополнительных аминокислот, или их сочетания, для создания SCCE протеиновых аналогов. Эти модификации могут привести к инверсным и полезным новым свойствам аналога. Аналоговые полипептиды или протеины можно получить из животных или человека, или они могут быть частично или полностью синтезированы. Аналоги можно также получить за счет использования методик рекомбинантных ДНК.
Таким образом, важный вариант настоящего изобретения относится к полипептиду, в котором, по крайней мере, один аминокислотный остаток замещен другим аминокислотным остатком и/или в котором, по крайней мере, один аминокислотный остаток был исключен или добавлен, так что в результате получен полипептид, содержащий аминокислотную последовательность, отличную от аминокислотной последовательности, представленной в последовательности ID N 2, или субпоследовательность указанной аминокислотной последовательности, как определено далее, но практически обладающей SCCE активностью, как указано ранее.
Интересный вариант изобретения относится к полипептиду, который является аналогом или субпоследовательностью полипептида настоящего изобретения, содержащим от 50 до 250 аминокислот, например, по крайней мере, 70 аминокислот, по крайней мере, 100 аминокислот, по крайней мере, 150 аминокислот или, по крайней мере, 200 аминокислот.
Наиболее важным вариантом изобретения является полипептид, содержащий аминокислотную последовательность -7- 224 в последовательности ID N 2 /про-SCCE/ и полипептид, содержащий аминокислотную последовательность 1-224 в последовательности ID N 2 /SCCE/.
Термин "энзиматически активное вещество" обозначает полипептидную последовательность, которая содержит только часть полипептидной последовательности, представленной в последовательности ID N 2, и которая обладает энзиматической активностью. Сюда входят полипептидные субпоследовательности, которые были аналогизированы за счет модификаций, как здесь указано. Специфический полипептид /или полипептиды/, который обладает энзиматической активностью, считается особенно интересным.
Предсказанная аминокислотная последовательность ID N 2 была сравнена с аминокислотными последовательностями энзимов химотрипсина человека /Joulta et al., 1989/, катепсина G человека /Salvesen et al., 1987/ и химазы тучных клеток человека /Caughey et al., 1991/. Хотя предсказанная аминокислотная последовательность содержит консервативные активные участки сериновых протеаз, степень гомологичности очень низка, около 33-38%. В этом отношении кажется, что SCCE имеет только умеренное сходство с известными ранее сериновыми протеиназами. С другой стороны, SCCE является типичной сериновой протеиназой в отношении гистидинового, аспарагинового и серинового остатков активных сайтов и консервативных участков, расположенных вблизи от этих сайтов. Это справедливо также для большинства цистеиновых остатков и других высоко консервативных участков сериновых протеиназ. В глубине "кармана" /pouch/ первичной специфичности /остаток 189 в химотрипсине/ расположен сериновый остаток химотрипсина человека, химаза тучной клетки и простато-специфический антиген и аланиновый остаток катепсина G человека. В SCCE, с другой стороны, это положение занято аспарагиновым остатком. Это может объяснить тот обнаруженный факт, что хотя SCCE несомненно обладает химотрипсиновой активностью, его относительная активность по отношению к различным хромогенным пептидным субстратам отличается от химотрипсина и катепсина G, так же, как и ингибирующая эффективность химостатина, низкомолекулярного ингибитора химотрипсин-подобных энзимов.
Сравнение последовательностей химотрипсина человека, катепсина G человека и химазы тучных клеток человека с последовательностью SCCE представлено в последующем сравнительном анализе первичных структур этих последовательностей, представленных в конце описания.
Сравнительный анализ первичных структур осуществляют вручную.
Обозначения:
HUMCTRP = химотрипсин человека /Jouita et al., BBRC 158:569-575, 1989/
HUMCHTG = катепсин G человека /Salvesen et al., Biochemistry, 26:2289-2293, 1987/
HUMCHYM = химаза тучных клеток человека /Caughey et al., J. Biol. Chem., 226:12956-12963, 1991/
Гомологичность:
HUMCTRP/SCCE: 85/224 = 38%
HUMCHTG/SCCE: 74/224 = 33%
HUMCHYM/SCCE: 74/224 = 33%
HUMCHTG/HUMCHYM: 109/226 = 48%
Нумерация относится к химотрипсиногену
Подчеркнуты:
Консервативные участки вблизи активного сайта /Ile 15, His 57 ASP 102, Ser 195 химотрипсина/ и карманы первичной специфичности химотрипсина /Ser 189, Ser 213 - Trp 215, Gly 226 химотрипсина/.
Звездочки: возможные сайты N-гликозилирования в SCCE.
Таким образом, важный вариант настоящего изобретения относится к полипептиду, содержащему аминокислотную последовательность, в которой последовательная нить из 20 аминокислот гомологична, по крайней мере, на 80% нити аминокислот той же самой длины, выбранной из аминокислотной последовательности ID N 2.
Полипептидные последовательности настоящего изобретения, гомологичные, по крайней мере, на 80%, например, на 85%, или, по крайней мере, на 90%, полипептидам, представленным в последовательности ID N 2, составляют важный вариант изобретения. Так как последовательность, представленная как последовательность ID N 2, по-видимому, совершенно уникальна, в объем настоящего изобретения входят полипептиды, для которых степень гомологичности с аналогичной последовательной нитью из 20 аминокислот, выбранных из аминокислотной последовательности ID N 2, может быть не более 55%, хотя предпочтительно, не более 70%. Такие последовательности можно получить из аналогичных протеинов из других видов, например, таких других млекопитающих, как мыши, крысы, кролики, морские свинки, свиньи или коровы. Так как небольшие части последовательности могут демонстрировать существенное сходство с другими сериновыми протеазами, в объем настоящего изобретения включены также пептиды со степенью гомологичности, по крайней мере, 95%, и наиболее предпочтительно, по крайней мере, 99% гомологичности с аналогичной последовательной нитью из 20 аминокислот, выбранных из аминокислотной последовательности, представленной в последовательности ID N 2.
Под термином "гомологичность последовательности" подразумевают идентичность в последовательности аминокислот в сегментах двух или более аминокислот в паре по отношению к идентичности и положению аминокислот полипептидов.
Таким образом, термин "гомологичный" использован здесь для иллюстрации степени идентичности аминокислотной последовательности данного полипептида и аминокислотной последовательности, представленной в последовательности ID N 2. Аминокислотную последовательность, подлежащую сравнению с аминокислотной последовательностью, представленной в последовательности ID N 2, можно вывести из нуклеотидной последовательности, такой, как ДНК или РНК последовательность, например, полученные за счет гибридизации, как будет указано далее, или можно получить обычными методами аминокислотного секвенирования. Степень гомологичности предпочтительно определяют на аминокислотной последовательности зрелого полипептида, то есть, без учета какой-либо лидерной последовательности. Обычно используют только кодирующие участки при сравнении нуклеотидных последовательностей для определения их внутренней гомологичности.
В настоящем контексте термин "полипептид, который распознается, по крайней мере, одним из депонированных антител" должен включать аминокислотную последовательность, которая содержит аминокислоты, составляющие практически последовательную нить в линейной или пространственной конформации любой последовательности полипептида, представленного в последовательности ID N 2, которая распознается по крайней мере одним из депонированных антител. Как хорошо известно специалистам, интересными могут быть также вторичные или третичные конформации; они могут обладать полезными свойствами и могут составлять эпитопы. Депонированные антитела TE4b и TE9b, по-видимому, распознают конформационные эпитопы SCCE.
Было показано, что поликлональные кроличьи антитела, полученные по способу примера 5.2.2, обеспечивают гранулярное окрашивание stratum granulosum и, скорее, диффузное окрашивание нижнего stratum corneum /роговичного слоя/, наблюдаемое с помощью иммунофлюоресцентной микроскопии.
Антитела, выработанные против очищенного нативного или рекомбинантного SCCE, обычно связываются с верхними базальными клетками кератинизированного /ороговевшего/ чешуйчатого эпителия человека, но не с эпителиальными клетками неороговевшего чешуйчатого эпителия. Эти антитела связываются также с межклеточным пространством ороговевшего слоя кожи человека.
Антитела, реакционноспособные с полипептидами настоящего изобретения, пригодны для использования с различными целями, как перечислено далее:
Для выделения протеинов: Антитела можно использовать для выделения полипептида /полипептидов/ или его аналога из биологических образцов с использованием афинной хроматографии или иммуноосаждения.
Для диагностики и терапии: Моноклональные антитела против полипептида /полипептидов/ или его аналогов можно использовать для диагностики и терапии болезненных состояний у животных и человека. Диагностическим агентом может быть антитело со специфичностью для полипептида настоящего изобретения. Это антитело можно соединить с другим протеином или твердым носителем и/или можно использовать в текстах агглютинации или в тестах проявления окраски. Такие антитела можно также использовать для количественного определения SCCE полипептидов или их аналогов в биологических образцах с использованием стандартных методик гистохимии и иммунохимии.
В одном из аспектов настоящее изобретение относится к нуклеотидной последовательности, кодирующей полипептид изобретение, как определено ранее. В частности, изобретение относится к нуклеотидной последовательности, содержащей практически последовательность, представленную в последовательности ID N 1. Другие важные варианты относятся к нуклеотидной последовательности, кодирующей полипептид, содержащий субпоследовательность аминокислотной последовательности ID N 2, такой как нуклеотидная последовательность, которая кодирует полипептид, содержащий аминокислотную последовательность -7-224 последовательности N 2, или полипептид, который содержит аминокислотную последовательность 1-224 последовательности ID N 2.
Настоящее изобретение относится также к нуклеотидной последовательности, которая гибридизуется с нуклеотидной последовательностью, представленной в последовательности ID N 1 в жестких условиях, как указано в примере 7 и в примере 9.
В другом аспекте настоящее изобретение относится к нуклеотидной последовательности, содержащей нуклеотидную последовательность, представленную в последовательности ID N 1, или ее аналог или ее субпоследовательность, которая
1/ гомологична последовательности, представленной в последовательности N 1, по крайней мере, на 90%, и/или
2/ кодирует полипептид, аминокислотная последовательность которого, по крайней мере, на 80% гомологична аминокислотной последовательности, представленной в последовательности ID N 2, и/или
3/ кодирует полипептид, который связан моноклональным антителом, продуцированным гибридомной клеточной линией TE4b, которая депонирована 18 июня 1993 г. в ECACC под регистрационным номером ECACC 93061817 или моноклональным антителом, продуцированным гибридомной клеточной линией TE9b, которая депонирована 18 июня 1993 г. в ECACC под регистрационным номером 93061816, и/или
4/ кодирует полипептид, который связан поликлональной антисывороткой, выработанной против нативного SCCE, который был выделен из экстракта диссоциированных клеток роговичного слоя пяточной поверхности.
В объем настоящего изобретения входит также модифицированная нуклеотидная последовательность, которая отличается от указанной ранее нуклеотидной последовательности тем, что, по крайней мере, один нуклеотид исключен, замещен или модифицирован, или по крайней мере один дополнительный нуклеотид встроен, так что в результате получена нуклеотидная последовательность, которая кодирует полипептид, обладающий SCCE активностью.
В настоящем описании и формуле изобретения термин "субпоследовательность" обозначает последовательность, предпочтительный размер которой составляет, по крайней мере, 15 нуклеотидов, более предпочтительно, по крайней мере, 18 нуклеотидов, и наиболее предпочтительно, по крайней мере 21 нуклеотид. В ряде вариантов настоящего изобретения субпоследовательность или аналог нуклеотидной последовательности настоящего изобретения содержит, по крайней мере, 48 нуклеотидов, например, по крайней мере, 75 нуклеотидов или, по крайней мере, 99 нуклеотидов. "Субпоследовательность" должна удовлетворять по крайней мере одному из критериев 1/-4/, приведенных ранее, или должна гибридизоваться с нуклеотидной последовательностью, представленной в последовательности ID N 1.
Хорошо известно, что мелкие фрагменты подходят для PCR методик, как описано здесь. Также фрагменты и последовательности могут, наряду с другими применениями, быть использованы в качестве зондов при идентификации мРНК фрагментов нуклеотидной последовательности настоящего изобретения, как указано в примере 7.
Термин "аналог" по отношению к ДНК фрагментам изобретения должен обозначать нуклеотидную последовательность, которая кодирует полипептид, идентичный или практически идентичный полипептиду, кодируемому ДНК фрагментом изобретения. Хорошо известно, что одну и ту же аминокислоту можно кодировать различными кодонами, причем использование кодона связано, наряду с остальным, с предпочтительностью рассматриваемого организма, экспрессирующего нуклеотидную последовательность. Так, один или более из нуклеотидов или кодонов ДНК фрагмента настоящего изобретения может быть заменен другими, которые будучи экспрессированы, приводят к получению полипептида, идентичного или практически идентичного полипептиду, кодирующему рассматриваемый ДНК фрагмент.
Также, термин "аналог" используют в контексте настоящего изобретения для обозначения ДНК фрагмента или ДНК последовательности аналогичного нуклеотидного состава или последовательности как характеристики ДНК последовательности ID N 1, кодирующей аминокислотную последовательность, составляющую SCCE полипептиды, обеспечивая небольшие
вариации, которые не оказывают значительного вредного воздействия на энзиматическую активность аналога, по сравнению с активностью нативного SCCE протеина, как указано в примере 3. Под термином "значительное вредное воздействие" подразумевают, что энзиматическая активность аналога должна составлять, по крайней мере, 50%, более предпочтительно, по крайней мере 60%, еще более предпочтительно, по крайней мере, 70%, например, по крайней мере, 75% энзиматической активности нативного SCCE, если это определяют, например, по способу примера 3. Аналогичную ДНК последовательность или ДНК фрагмент можно получить из такого организма, как например, организм животного или человека, или получить частично или полностью синтетическим способом. Аналог можно также получить за счет использования методик рекомбинантных ДНК.
Более того, термины "аналог" и "последовательность" должны включать такие вариации в последовательности, как замещения, вставки /включая интроны/, добавления и перестройка одного или более из нуклеотидов, причем эти вариации не должны оказывать никакого вредного воздействия на полипептид, кодируемый ДНК фрагментом или его субпоследовательностью.
Термин "замещение" подразумевает замену одного или более из нуклеотидов в полной нуклеотидной последовательности одним или более из отличных нуклеотидов, "добавление" означает добавление одного или более из нуклеотидов с любого конца полной нуклеотидной последовательности; "включение" означает введение одного или более из нуклеотидов внутрь полной нуклеотидной последовательности; "делеция" означает, что один или более из нуклеотидов удален из полной нуклеотидной последовательности либо с любого конца полной последовательности, либо из любой его подходящей точки, а "перегруппировка" означает, что два или более нуклеотидных остатка поменялись местами внутри ДНК или полипептидной последовательности, соответственно. ДНК фрагмент может, однако, также быть модифицирован в результате мутагенеза либо до, либо после введения его в организм.
Термины "фрагмент", "последовательность", "субпоследовательность" и "аналог", в том смысле, как использованы в настоящем описании и формуле изобретения в отношении фрагментов, последовательностей, субпоследовательностей и аналогов в соответствии с настоящим изобретением, должны, естественно, рассматриваться как не содержащие этих вещей в их природном окружении, но существующие скорее, например, в выделенной, очищенной, ин витро или рекомбинантной форме.
В одном варианте настоящего изобретения, детектирование и/или количественное определение SCCE полипептидной мРНК можно получить за счет экстрагирования РНК из клеток или тканей и превращения его в кДНК или последующего использования в полимеразной цепной реакции /PCR/; PCR праймер /праймеры/ можно синтезировать на основании ДНК фрагмента настоящего изобретения, такого, как ДНК фрагмент, представленный в последовательности ID N 1. Этот способ для определения и/или количественного определения можно использовать в качестве диагностического метода для диагностики болезненных состояний, в которых SCCE мРНК экспрессируется в больших или меньших количествах нежели обычно.
В объем настоящего изобретения включен также диагностический агент, содержащий нуклеотидный зонд, который способен детектировать нуклеотидную последовательность изобретения, а также способ диагностики заболеваний, в которых CCE экспрессия нарушена, и/или заболеваний, при которых SCCE ген мутирован; причем метод включает проведение PCR анализа образца, полученного от пациента, у которого подозревается заболевание, при котором присутствуют более высокие количества SCCE, нежели обычно, или присутствуют мутированные формы SCCE, в котором образец подвергают контакту с диагностическим агентом, как описано ранее, обеспечивая нуклеотидный последовательности возможность быть амплифицированной и определяя наличие какой-либо идентичной или гомологичной нуклеотидных последовательностей в образце.
Полипептиды настоящего изобретения можно получить, используя рекомбинантную ДНК методику. Важный вариант настоящего изобретения относится к экспрессирующей системе, содержащей нуклеотидную последовательность настоящего изобретения.
Организм, который используют для получения полипептида настоящего изобретения, может быть высшим организмом, например, животного, или низшим организмом, например, микроорганизмом, таким, как E.coli. Независимо от типа используемого организма, ДНК фрагмент настоящего изобретения вводят в организм либо непосредственно, либо с помощью подходящего вектора. В другом варианте, полипептиды можно получить в клеточных линиях млекопитающих, вводя ДНК фрагмент или его аналог, или его субпоследовательность настоящего изобретения либо непосредственно либо с помощью вектора экспрессии.
ДНК фрагмент или аналог, или его субпоследовательность можно также клонировать в подходящий стабильный вектор экспрессии, а затем ввести в подходящую клеточную линию. Клетки, продуцирующие нужные полипептиды, затем селектируют по уровням продуктивности в условиях, подходящих для используемого вектора или клеточной линии. Отобранные клетки затем выравнивают далее, и они образуют очень важный и непрерывный источник нужных полипептидов.
Примером специфического аналога ДНК последовательности настоящего изобретения является ДНК последовательность, которая включает ДНК последовательность, представленную в ДНК последовательности ID N 1, или ее часть, и которая наиболее адаптирована для экспрессии в E coli. Эта ДНК последовательность является последовательностью, которая, будучи встроена в E. coli вместе с подходящей регулярной последовательностью, приводит к экспрессии полипептида, содержащего практически аминокислотную последовательность, представленную в последовательности ID N 2, или ее часть. Так, ДНК последовательность содержит специфические кодоны, распознаваемые E. coli. Пример такого варианта представлен в примере 8.
В настоящем контексте термин "ген" используют для обозначения ДНК последовательности, которая включена в продуцирование полипептидной цепи и которая включает участки, предшествующие и следующие за кодирующим участком /5'-прямом направлении и 3'-обратном направлении последовательностей/, так же как прерывающие последовательности, интроны, которые расположены между отдельными кодирующими сегментами, экзоны или в 5'-прямом направлении или 3'-обратном направлении участки. Участок в 5'-прямом направлении содержит регуляторную последовательность, которая контактирует экспрессию гена, обычно промотор, 3'-участок в обратном направлении содержит последовательности, которые участвуют в терминации транскрипции гена, и необязательно последовательности, ответственные за полиаденилирование транскрипта и 3'-нетранслируемого участка.
Наряду с вышеперечисленным настоящее изобретение относится к экспрессирующей системе, содержащей нуклеотидную последовательность, как описано ранее, кодирующую полипептид настоящего изобретения, систему, которая содержит 5'-фланкирующую последовательность, способную осуществлять экспрессию указанной нуклеотидной последовательности.
В частности, настоящее изобретение относится к реплицируемому вектору экспрессии, который обеспечивает экспрессию нуклеотидной последовательности настоящего изобретения. Конкретный вариант настоящего изобретения относится к реплицируемому вектору экспрессии, обозначенному pS 507, который был депонирован 11 мая 1993 г. в коллекции Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSM) под регистрационным номером DSM 8282 в соответствии с требованиями Будапештского соглашения, и к векторам экспрессии, которые экспрессируют нуклеотидные последовательности, которые отличаются от нуклеотидной последовательности указанного депонированного вектора экспрессии, но которые кодируют тот же самый полипептид или его аналог или его вариант, которые обладают SCCE активностью.
Другими словами, объем изобретения включает также реплицируемый вектор экспрессии, как было указано ранее, в котором экспрессируемая нуклеотидная последовательность является последовательностью, которая отличается от нуклеотидной последовательности депонированного вектора тем, что, по крайней мере, один нуклеотид был исключен, заменен или модифицирован, или, по крайней мере, один дополнительный нуклеотид был встроен таким образом, чтобы нуклеотидная последовательность, которая кодирует полипептид, обладала бы SCCE активностью.
Более того, настоящее изобретение относится к плазмиде, обозначенной pS 500, которая была депонирована 11 мая 1993 г. в коллекции Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSM) под регистрационным номером DSM 8281 в соответствии с требованиями Будапештского соглашения, и к плазмидам, содержащим нуклеотидную последовательность, которая отличается от нуклеотидной последовательности, представленной в нуклеотидной последовательности ID N 1, но которая кодирует полипептид, представленный в последовательности ID N 2, или ее аналог или ее вариант, которые обладают SCCE активностью, или которая гибридизуется с нуклеотидной последовательностью, представленной в последовательности ID N 1, или ее частью в жестких условиях гибридизации.
В объем настоящего изобретения входит также нечеловеческий организм, который содержит экспрессионную систему настоящего изобретения. Организмы, которые можно использовать в этом аспекте изобретения, включают микроорганизмы, такие как бактерии родов Bacillus, Escherichia или Salmonella, такие дрожжи, как Saccharomyces, Pichia, простейшие, или клетки, полученные из многоклеточных организмов, таких как грибы, клетки насекомых, клетки растений, клетки млекопитающих или клеточные линии. Если организм является бактерией, предпочтительно, чтобы бактерия относилась к роду Escherichia, например, E.coli.
Кроме того, настоящее изобретение относится к плазмидному вектору, содержащему ДНК последовательность, которая кодирует полипептид настоящего изобретения или полипептид слияния, как здесь определено. В одном из наиболее важных вариантов ДНК фрагмент, или его аналог, или его субпоследовательность настоящего изобретения или фрагмент ДНК слияния настоящего изобретения может содержаться в реплицируемом векторе экспрессии, который способен реплицироваться в организме хозяина или в клеточную линию.
Такой вектор может быть, в частности, плазмидой, фагом, космидой, мини-хромосомой или вирусом. В интересном варианте изобретения этот вектор может быть вектором, который при введении в клетку-хозяина оказывается интегрированным в клеточный геном.
Если используют высший организм, для получения полипептидов можно использовать трансгенную методику. Примерами подходящих животных являются овцы, рогатый скот, свинья и т.д. ДНК фрагмент, который кодирует полипептид настоящего изобретения, экспрессируется в нужную ткань под контролем ткане-специфических регуляторных элементов. Полученный протеин можно затем подвергнуть пост-трансляционным модификациям для получения полипептида настоящего изобретения.
Трансгенные млекопитающие /это не относится к человеку/ настоящего изобретения получают за счет введения "трансгена" в эмбриональную мишеневую клетку выбранного животного. В одном аспекте изобретения трансгеном является ДНК последовательность, которая способна продуцировать нужный фенотип, когда она содержится в геноме клеток трансгенного млекопитающего /не человека/. В специфических вариантах трансген содержит ДНК последовательность, кодирующую полипептид настоящего изобретения, причем этот трансген способен, будучи экспрессирован, продуцировать полипептид.
Включение экспрессирующей системы в зародышевую линию /germline/ млекопитающего можно осуществить, используя любую подходящую методику, например, описанную Hogan et al., 1986, или в WO 93/04172.
В конкретном аспекте изобретения нуклеотидная последовательность изобретения может содержать другую нуклеотидную последовательность, кодирующую полипептид, отличающийся от или идентичный полипептиду настоящего изобретения, слитый в рамке с нуклеотидной последовательностью последовательности, представленной в последовательности ID N 1, или ее аналогом, кодирующим SCCE полипептид, с целью получения слитого полипептида, причем этот полипептид составляет еще один интересный аспект настоящего изобретения, см. пример 8. Если используют технологию рекомбинантных ДНК, слитые ДНК последовательности могут быть встроены в подходящий вектор или геном. В другом варианте одну из нуклеотидных последовательностей встраивают в вектор или геном, уже содержащий другую нуклеотидную последовательность. Полипептид слияния можно также получить, встраивая две нуклеотидные последовательности отдельно и предоставляя возможность протекания экспрессии. Организм хозяина, который может быть как эукариотного, так и прокариотного происхождения, выращивают в условиях, которые обеспечивают экспрессию слитых последовательностей. Затем слитый полипептид очищают, и полипептид настоящего изобретения отделяют от его партнера слияния, используя подходящие методики.
Таким образом, один из аспектов настоящего изобретения относится к способу получения полипептида настоящего изобретения, который включает стадии:
/a/ встраивания нуклеотидной последовательности настоящего изобретения в вектор экспрессии,
/b/ трансформирования подходящего организма хозяина вектором, полученным на стадии /a/,
/c/ культивирования организма хозяина, полученного на стадии /b/ в условиях, подходящих для экспрессии полипептида,
/d/ сбора полипептида и
/e/ необязательной пост-трансляционной модификации полипептида.
В объем настоящего изобретения входит также описанный ранее способ, в котором полученный полипептид выделяют способом, включающим одну или более из стадий, подобных афинной хроматографии, с использованием иммобилизованного нативного или рекомбинантного SCCE полипептида или антител, реакционноспособных с указанным полипептидом, и/или процедурам хроматографической обработки и электрофореза.
Полипептид, полученный как указано ранее, можно пост-трансляционно модифицировать, например, за счет термообработки, химической обработки /формальдегид, глутаральдегид и т.д./ или энзиматической обработки /пептидазы, протеиназы и протеин-модифицирующие энзимы/. Этот полипептид при продуцировании в организме может быть процессирован другими способами, нежели те, которые происходит в его продуцировании в природном окружении. Например, гликозилирование часто достигается, когда полипептид экспрессируется такой клеткой высшего организма, как дрожжи или предпочтительно млекопитающие. Обычно гликозилирование связано с аминокислотными остатками Asn, Ser, Thr или гидроксилизином. Может оказаться выгодным /а может и нет/ удалить или изменить характеристики процессинга, создаваемого рассматриваемым организмом хозяина.
После экспрессии в соответствии с настоящим изобретением полипептида в организм или клеточную линию, полипептид можно использовать как он есть, или его можно вначале выделить из организма или клеточной линии. Если полипептид экспрессирован в виде секретированного продукта, его можно очистить непосредственно. Если полипептид экспрессирован в виде ассоциированного продукта, может понадобиться частичное или полное разрушение хозяина перед очисткой.
Примеры процедур, использованных для очистки полипептидов, включают:
/i/ иммуноосаждение или афинную хроматографию с антителами,
/ii/ афинную хроматографию с подходящим лигандом,
/iii/ другие хроматографические процедуры, такие как гельфильтрация, ионообмен или высокоэффективная жидкостная хроматография, или модификации вышеперечисленных,
/iv/ электрофоретические процедуры, подобные электрофорезу в полиакриламидном геле, электрофорезу в денатурированном полиакриламидном геле, электрофорезу в агарозном геле и изоэлектрическому фокусированию,
/v/ любые другие специфические методики солюбилизации и/или очистки.
Настоящее изобретение относится также к практически чистому SCCE полипептиду. В данном контексте термин "практически чистый" означает, что рассматриваемый полипептид практически не содержит других компонентов, то есть, других полипептидов или углеродов, которые могли образоваться в процессе получения и/или выделения полипептида, или каким-либо другим образом оказаться вместе с полипептидом. Степень чистоты протеина можно оценить с помощью электрофореза на SDS геле, как описано в примере 3.
Полипептид можно выделить по способу примера 4 с помощью SBTJ афинной хроматографии или с помощью афинной хроматографии на иммобилизованных антителах, например, на антителах TE4b, способом, известным специалистам. Высокая степень чистоты полипептида настоящего изобретения может быть выгодна если полипептид собираются использовать в фармацевтических или косметических композициях. Кроме того, благодаря высокой степени чистоты, практически чистый полипептид можно использовать в меньших количествах, нежели полипептид обычной более низкой степени чистоты, для большинства целей.
В одном из аспектов настоящего изобретения чистый полипептид можно получить из подходящей клеточной линии, которая экспрессирует полипептид настоящего изобретения, как описано в примере 9. Кроме того, полипептид настоящего изобретения можно получить хорошо известными способами жидкостного или твердофазного пептидного синтеза, используя последовательное сочетание отдельных аминокислот полипептидной последовательности. В другом варианте полипептид можно синтезировать за счет соединения отдельных аминокислот, образующих фрагменты полипептидной последовательности, которые затем соединяют для получения целевого полипептида. Эти способы, таким образом, составляют следующий интересный аспект изобретения.
Очень важный аспект изобретения относится к фармацевтической композиции, косметической композиции или композиции для лечения кожи, которая содержит полипептид, обладающий SCCE активностью, и фармацевтически и/или косметически приемлемый эксципиент. Композиция может содержать очищенный нативный протеин или рекомбинантный полипептид настоящего изобретения, или проэнзим или протеин слияния - формы полипептида настоящего изобретения, которые можно активировать за счет протеолитического расщепления.
Проэнзимную форму полипептидов настоящего изобретения можно классифицировать как "пролекарственную форму", то есть, соединение, которое после соответствующего протеолитического расщепления превращается в активную форму.
Особенно, но не исключительно, настоящее изобретение относится к композициям, пригодным для поверхностного нанесения, например, нанесения на кожу.
Фармацевтическими композициями настоящего изобретения, подходящими для поверхностного нанесения, могут быть кремы, мази, лосьоны, линименты, гели, растворы, суспензии, пасты, пластыри, спреи, шампуни, мыла, кондиционеры для волос или порошки.
Поверхностное нанесение предусматривает нанесение на или вблизи с частями тела, на которых наблюдаются рассматриваемые патологические изменения, например, на такие внешние части тела, как участки кожной поверхности. Нанесение может быть просто распределением композиции по поверхности или может включать любые приспособления, предназначенные для усиления и закрепления контакта композиции и патологических повреждений, например, закрывающие повязки, такие как изолирующие пластыри с нанесенной композицией настоящего изобретения. Эти композиции могут пропитывать или могут быть распределены на тампонах, пластырях, бинтах, марле, губчатых материалах, кусочках ваты и т. д. В некоторых случаях можно использовать композиции в форме инъекций непосредственно в пораженный участок или вблизи него.
Композицию для поверхностного нанесения настоящего изобретения могут содержать 1-80% активного соединения по весу в расчете на полный вес препаратов, например, 0,001-25% вес/вес активного соединения, например, 0,1-10%, 0,5-5% или 2-5%. В состав композиции может входить более одного активного соединения; так в объем настоящего изобретения включены композиции, содержащие SCCE, про- SCCE или ингибитор SCCE в сочетании с другими фармацевтическими и/или косметическими соединениями.
Композиция настоящего изобретения обычно наносят 1-10 раз в день, в зависимости от типа, степени и расположения пораженного участка.
Для поверхностного нанесения препарат может быть приготовлен в соответствии с обычной фармацевтической практикой с фармацевтическими эксципиентами, которые обычно используют для поверхностных нанесений. Природа носителя, который используют для приготовления каждой конкретной композиции, зависит от предполагаемого способа нанесения композиции. Помимо воды можно использовать в композициях и другие носители, которые включают такие твердые вещества или жидкости, как мягчительные средства, растворители, увлажнители, загустители и порошки. Примерами такого типа носителей, которые можно использовать либо отдельно, либо в смесях, состоящих более чем из одного носителя, являются следующие:
Умягчающие средства: стеариловый спирт, монорицинолеат глицерина, моностеарат глицерина, пропан-1,2-диол, бутан-1,3-диол, цетиловый спирт, изопропилизостеарат, стеариновая кислота, изобутилпальмитат, изоцетилстеарат, олеиловый спирт, изопропиллаурат, гексиллаурат, децилолеат, октадекан-2-ол, изоцетиловый спирт, цетилпальмитат, диметилполисилоксан, ди-н-бутилсебакат, изопропилмиристат, изопропилпальмитат, изобутилстеарат, бутилстеарат, полиэтиленгликоль, триэтиленгликоль, ланолин, касторовое масло, ацетилированные ланолиновые спирты, петролеум, минеральное масло, бутилмиристат, изостеариновая кислота, пальмитиновая кислота, изопропиллинолеат, лауриллактат, миристиллактат, децилолеат, миристилмиристат.
Растворители: вода, метиленхлорид, изопропанол, касторовое масло, моноэтиловый эфир этиленгликоля, монобутиловый эфир диэтиленгликоля, моноэтиловый эфир диэтиленгликоля, диметилсульфоксид, тетрагидрофуран, растительные и животные масла, глицерин, этанол, пропанол, пропиленгликоль и другие гликоли или спирты, фиксированные масла.
Такие увлажняющие агенты, как глицерин, сорбитол, натрий-2-пирролидон-5-карбоксилат, растворимый коллаген, дибутилфталат, желатин.
Такие порошки, как мел, тальк, каолин, крахмал и его производные, смолы, коллоидная двуокись кремния, полиакрилат натрия, химически модифицированный магний-алюминий-силикат, гидратированный алюмосиликат, карбоксивиниловый полимер, натрий-карбоксиметилцеллюлоза, моностеарат этиленгликоля.
Такие желирующие или способствующие набуханию агенты, как пектин, желатин и их производные, такие производные целлюлозы, как метилцеллюлоза, карбоксиметилцеллюлоза или окисленная целлюлоза, целлюлозная смола, гуаргам, смолы акации, карайи и трагаканта, бентонит, агар, альгинаты, карбомеры, желатин, bladderwrack, ceratonia, дикстран и его производные, смола ghatti, гекторит, смола ксантам, шелуха ispaghula.
Такие полимеры, как полиакриловая кислота или полимеры полигликолевой кислоты, или их сополимеры, парафин, полиэтилен, полиэтиленоксид, полиэтиленгликоль, полипропиленгликоль, поливинилпирролидон.
Такие поверхностно-активные агенты, как неионные поверхностно-активные агенты, например, гликоль и сложные эфиры глицерина, простые и сложные эфиры макроголя, простые и сложные эфиры сахара, такие как сорбитановые эфиры, такие ионные поверхностно-активные агенты, как аминомыла, металлические мыла, сульфатированные жирные спирты, алкилэфирсульфаты, сульфатированные масла и амфолитические поверхностно-активные агенты и лецитины.
Такие буферирующие агенты, как соли натрия, калия, алюминия, магния или кальция /такие как хлориды, карбонаты, бикарбонаты, цитраты, глюконаты, лактаты, ацетаты, глюцептаты или тартраты/.
Для поверхностного нанесения pH композиции, в принципе, может меняться в очень широком интервале значений, например, от 3 до 9. В предпочтительном варианте изобретения предпочтителен интервал pH, который соответствует протеолитической активности полипептида, например, pH от около 4 до 8. Указанные ранее буферирующие агенты можно использовать для установления нужных значений pH.
Препараты настоящего изобретения могут также содержать такие другие добавки, как стабилизирующие агенты, консерванты, агенты, облегчающие растворение, окрашивающие агенты, хелатирующие агенты, геле-образующие агенты, основания мазей, регуляторы pH, антиоксиданты, отдушки и агенты, защищающие кожу, и т. д. Если композиция приготовлена в форме мыла или шампуня, она может, кроме того, содержать вспенивающие агенты, оттеночные агенты и/или кондиционеры.
Типичные консерванты включают парабены, формальдегид, Kathon СG, Bronidox, Bronopol, парахлор-мета-крезол, хлоргексидин, бензалконийхлорид и т.д.
Если композиции настоящего изобретения имеют форму шампуня или мыла, можно использовать обычные ингредиенты, а типичные основы мыла и шампуня включают такие компоненты, как бетаин, натрийлаурилсульфат, ионилфенол, имидазол, сульфосукцинат, обезжиривающие агенты, увлажнители и кондиционеры.
Более того, может оказаться выгодным создать препараты с модифицированным выделением, в которых активное соединение включено в полимерную матрицу, или в наночастицы, или в липосомы, или мицеллы, или адсорбиравано на ионообменных смолах, или носителем является полимер.
Композиции можно получить в соответствии с обычной фармацевтической практикой, и они могут быть:
Полутвердыми композициями: гелями, пастами, смесями.
Жидкими композициями: растворами, суспензиями, эмульсиями.
Как было указано, фармацевтическая композиция настоящего изобретения может содержать сам полипептид настоящего изобретения или его функциональное производное, или сочетание таких соединений. Примеры подходящих функциональных производных включают фармацевтически приемлемые соли, особенно те соли, которые пригодны для использования на коже. Примеры включают фармацевтически приемлемые соли аминофункций, например, соли кислот, которые дают анионы, которые фармацевтически приемлемы, особенно для применения на коже. Примеры включают фосфаты, сульфаты, нитраты, иодиды, бромиды, хлориды, бораты, а также анионы, полученные из карбоновых кислот, которые включают ацетаты, бензоаты, стеараты и т.д.
Другие производные аминофункций включают амиды, имиды, мочевины, карбаматы и т.д.
Другие подходящие производные включают производные карбоксильной группы полипептида настоящего изобретения, включая соли, сложные эфиры и амиды. Примеры включают соли фармацевтически приемлемых катионов, например, соли лития, натрия, калия, магния, кальция, цинка, алюминия, железа /2 и 3/, аммония и низшего /C1-6/-алкиламмония. Сложные эфиры включают низшие алкилсложные эфиры.
Примеры композиций в примере 11 иллюстрируют примеры фармацевтических косметических композиций и композиций для ухода за кожей в соответствии с настоящим изобретением, но они никоим образом не ограничивают объем композиций настоящего изобретения.
Показано, что косметические композиции для ухода за кожей, содержащие нативные или рекомбинантные SCCE, активны против акне, ксеродермы или других гиперкератозных состояний, таких как мозоли и keratosis pilaris.
Существуют различные стадии акне вульгарис. Считают, что желательно вводить SCCE на тех стадиях, когда наблюдается распространенная кератинизация в протоках сальных желез, что приводит к образованию комедонов и закупорке проток, поэтому может быть выгодно вводить вещество, которое ингибирует SCCE на стадиях, когда воспалительные акне поражения представляют преимущественную характеристику.
Таким образом, один из аспектов настоящего изобретения относится к использованию полипептида для лечения или профилактики акне, ксеродермы и других гиперкератинозных состояний, таких как мозоли и keratosis pilaris.
На основании приведенных ранее научных наблюдений можно считать, что фармацевтические композиции, содержащие нативные или рекомбинантные SCCE, пригодны для лечения или профилактики различных ихтиозов, акне, псориазов или других воспалительных кожных заболеваний, таких, как экземы с гиперкератозом, микробные инфекции, и для залечивания ран, особенно при поверхностном нанесении.
Другой аспект изобретения относится к использованию полипептида, обладающего SCCE активностью, для приготовления фармацевтических композиций для лечения или профилактики различных ихтиозов, акне, псориазов или других воспалительных кожных заболеваний, сопровождающихся гиперкератозом, таких как экземы.
Другой аспект изобретения относится к способу лечения и/или предотвращения различных ихтиозов, акне, псориазов и других воспалительных кожных заболеваний, сопровождающихся гиперкератозом, таких как экземы, причем способ включает введение нуждающемуся в лечении пациенту терапевтически или профилактически эффективного количества полипептида, обладающего SCCE активностью. Обработка может быть профилактической, паллиативной или лечебной.
Предполагается, что "каскадная система" протеолетических энзимов существует в кожном окружении, которая аналогична системе активации плазминогена. Предполагается, что SCCE является одним из конечных продуктов этой системы. Поэтому предполагают, что активность SCCE можно ингибировать с помощью "SCCE ингибитора".
В случае ряда кожных заболеваний, таких, как автоиммунные заболевания-пузырчатки или акантолитические заболевания, например, семейная пузырчатка или болезнь Dariеr's, существует когезия между кератиноцитами и ороговевшим жизненным эпидермальным слоем /см. нарушения клеточной когезии в живом эпидермисе/. Считают, что этот процесс осуществляется за счет протеиназ, и поэтому на него можно повлиять, вводя соединение, которое способно ингибировать энзиматическую активность нативного SCCE. Может также оказаться выгодным вводить SCCE ингибитор для лечения псориаза и других воспалительных кожных заболеваний в состояниях, когда воспалительная компонента является преобладающей характеристикой.
В другом аспекте настоящее изобретение относится к использованию SCCE ингибитора, который оказывает ингибирующее действие на энзиматическую активность нативного SCCE, для приготовления фармацевтической композиции для лечения или профилактики автоиммунных заболеваний-пузырчаток или акантолитических заболеваний, таких как семейная пузырчатка или болезнь Darier's.
В настоящем контексте термин "SCCE ингибитор" относится к существующему или новому соединению, которое может взаимодействовать с энзиматической активностью полипептидной последовательности или субпоследовательности изобретения или его аналога таким образом, что его SCCE активность снижается. Такое снижение можно определить, например, осуществляя эксперимент в соответствии со схемой примера 3.2, используя потенциальный SCCE ингибитор в качестве ингибитора. Указанные соединения могут быть органическими молекулами, небольшими пептидами или крупными полипептидами или производными любых указанных ранее. Такой подход может найти применение в программе скринирования лекарств для определения SCCE ингибиторов.
Таким образом, следующий вариант настоящего изобретения относится к способу идентификации соединения, которое оказывает воздействие на энзиматическую активность нативного SCCE, включающему использование рекомбинантного полипептида настоящего изобретения.
В частности, настоящее изобретение относится к способу идентификации соединения, которое обладает ингибирующим действием на энзиматическую активность нативного SCCE.
В другом аспекте настоящее изобретение относится к способу идентификации соединения, которое способно повысить энзиматическую активность нативного или рекомбинантного SCCE.
Важным применением рекомбинантных полипептидов настоящего изобретения является скринирование лекарств. Полипептиды настоящего изобретения можно использовать в скринирующей лекарства системе. Настоящее изобретение относится также к способу идентификации соединения, которое способно превращать проэнзимную форму SCCE в активный SCCE, содержащий полипептид настоящего изобретения.
В концепцию настоящего изобретения входит также использование аминокислотной последовательности, как указано ранее, для определения трехмерной структуры SCCE полипептида для использования в создании вещества, способного связывать SCCE полипептид, особенно для использования в создании лекарственного вещества, которое связывает активный сайт энзима.
И наконец, важным аспектом настоящего изобретения являются различные способы регулирования активности, демонстрируемой SCCE полипептидом. Эта активность может иметь важное значение для различных болезненных состояний, как было указано ранее.
ДНК или РНК фрагмент, комплементарный, по крайней мере, части мРНК, соответствующей полипептиду настоящего изобретения или его аналога, может быть эффективным в прекращении трансляции SCCE vHYR в клетках человека, и тем самым, в ингибировании синтеза полипептида /полипептидов/. Этот подход, который может оказаться интересным при болезненных состояниях, в которых наблюдается более высокая нежели обычно экспрессия SCCE, таких как автоиммннуные заболевания-пузырчатки или акантолитические заболевания, такие как семейная пузырчатка и болезнь Dariеr's, обычно более известен как антисмысловая олиготерапия и, поэтому настоящее изобретение включает также и этот подход.
Описание чертежей
Фиг. 1
Однополярная клетка, отшелушившаяся с роговичного слоя пяточной поверхности ин витро.
Поверхность ткани, которая ин виво смотрела вверх, на фиг. 1 расположена этой поверхностью вниз. Следует отметить, что наблюдается прогрессивная клеточная диссоциация на этой поверхности в процессе инкубирования, а на остальных поверхностях клеточной диссоциации не наблюдается.
Фиг. 2
Временная зависимость и действие апротинина на выделение клеток из роговичного слоя пяточной поверхности, инкубированного без /кружки/ и с /треугольники/ апротинином. Приведены средние значения /суммарные значения для четырех образцов тканей/ и интервал достоверности.
Фиг. 3
Анти-десмоглеиновые /анти-DG1/ реактивные компоненты в сплошном роговичном слоем пяточной поверхности и в диссоциированных клетках.
А. Окрашенный Coomassie - blue SDS-PAGE.
В. Иммуноблот. 1-3: Связанные ткани, неразбавленные /1/, разбавленные 1/3 /2/, разбавленные 1/9 /3/. 4-5: диссоциированные клетки, неразбавленные /4/, разбавленные 1/3 /5/.
Отмечаются только очевидно интактные DG 1 с Мг 160 кД в цельной ткани, и только продукты разложения DG с Мг 95 и 80 кД в диссоциированных клетках.
Фиг. 4
Временная зависимость и влияние апротинина на деградацию десмоглеина 1 /DG 1/ в роговичном слоем пяточной поверхности, претерпевающей отшелушивание клеток ин витро. Денситометрическое сканирование иммуноблотов экстрактов роговичного слоя пяточной поверхности, инкубированной в отсутствие /A/ и в присутствии /B/ апротинина /15 мкМ/. Пик на 160 кД соответствует интактным DG 1. Пики на 95 и 80 кД соответствуют продуктам деградации этого протеина /см. фиг. 3/. Отмечается эффективное ингибирование апротинином деградации DG1.
Фиг. 5.
Влияние ионов цинка /A/, химостатина и леупептина /B/ на деградацию десмоглеина 1 /DG 1/ в клетках роговичного слоя пяточной поверхности, отшелушивающихся ин витро. Отмечается ингибирование трансформации анти- DG1 позитивных компонентов из 160 кД в 95 и 80 кД за счет ионов цинка и химостатина, но не за счет леупептина.
Фиг. 6.
Активность гидролиза пептидов, связанная с клетками роговичного слоя пяточной поверхности. За гидролизом двух субстратов наблюдают, измеряя изменения в поглощении на 405 нм. Среднее для трех инкубирований. Квадраты = S-2586; кружки = S-2288.
Фиг. 7
pH-зависимость корнеоцит-связанной S-2586 гидролизующей активности. Среднее для трех инкубирований. Квадраты = ацетат натрия, кружки = фосфат натрия, треугольники = Tris-HCl.
Фиг. 8
Зимография, демонстрирующая казеинолитическую активность в экстрактах диссоциированных клеток роговичного слоя пяточной поверхности. См. также текст примера 2.2 для подробностей эксперимента.
A: Сравнение энзима из пяточных корнеоцитов, экстрагированных образцов буфера Лаэмли без восстанавливающего агента /Sc/S/ и KCl /Sc/k/ с бычьим химотрипсином, 0,125 нг /с/ и бычьим трипсином, 0,5 нг /т/. Перед электрофорезом KCl-экстракт диализуют против 5 мМ Tris-HCl, pH 6,8, и SDS и глицерин добавляют до конечных концентраций, как и в образцовом буфере; 10 мкл добавляют во все полосы. Маркеры молекулярного веса приведены слева.
B: Зависимость от pH инкубационного буфера. Источник энзима = Д - экстракты диссоциированных пяточных корнеоцитов. Буферы для предварительной обработки Triton X= 100 и инкубирования: 0,1 М ацетат натрия /pH 4,0 и pH 5,5/, и 0,1 М Тris-HCl /pH 7,0 и pH 8,0/. Остальные условия как и для A.
C: Действие ингибиторов. Sс = SDS - экстракт пяточных корнеоцитов; с = бычий химотрипсин; т - бычий трипсин. Ингибиторы присутствуют во время предварительной обработки Triton X-100 и последующего инкубирования. Конечная концентрация леупептина составляет 160 мкМ, апротинина 15 мкМ, химостатина 40 мкМ и ионов цинка /в виде сульфата/ 100 мкМ. Леупептин и химостатин добавляют в виде растворов в диметилсульфоксиде /ДMCO/. Буфер для предварительной обработки и инкубирования = 0,1 М Tris-HCl, pH 8 при конечной концентрации ДMCO 1% /объем/объем/. Остальные условия как и в A.
Фиг. 9
Сравнение SCCE, бычьего химотрипсина и катепсина G человека в отношении действий ингибиторов /A; апротинин; B; химостатин, C; сульфат цинка/ и субстратной специфичности /Д/. Для A-C активность энзима без ингибиторов принимают за 100%. B Д активность энзима с MeO-Suc-Arg-Pro-Tyr-pNA принимают за 1 произвольную единицу.
Фиг. 10.
Афинная хроматография на ковалентно связанном ингибиторе трипсина сои/SBTJ. Пунктирная линия: поглощение на 280 нм отражает концентрацию протеина в элюате. Сплошная линия с незаполненными квадратами: активность гидролиза пептида с S-2586 в качестве субстрата, приведенная как изменение поглощения на 405 нм. Сплошная линия с пустыми кружками: Активность гидролиза пептида с S-2288 в качестве субстрата, приведенная как изменение поглощения на 405 нм, умножения на 10.
Фиг. 11.
SDS-PAGE и Coomassie-blue - окрашивание /A/ и зимография после SDS-PAGE с сополимеризованным казеином /B/ фракций 2,6 и 10 из хроматограмм, представленных на фиг. 10. Маркеры молекулярного веса представлены слева. В B: 1: группа казеинолитических компонентов, которые можно ингибировать леупептином /160 мкМ/. C: группа казеинолитических компонентов, которые можно ингибировать химостатином /40 мкМ/.
Фиг. 12.
SDS-PAGE SCCE, очищенного с помощью афинной хроматографии на SBTJ-Affigel 15/ 12,5% гель.
1: невосстановленный образец.
2: Восстановленный образец.
Маркеры молекулярного веса представлены слева.
Фиг. 13.
N-терминальная аминокислотная последовательность SCCE.
Звездочки означают неопределенности для предполагаемых цистеинов в положениях 7 и 9. Знаки вопросов означают положения, в которых нельзя определить аминокислотные производные, но на последующих стадиях нет снижения выхода.
Фиг. 14.
Характеризация моноклональных антител TE4b и TE9b за счет иммуноосаждения и иммуноблоттинга.
a: Coomassie - окрашенный 12,5% SDS-PAGE, невосстанавливающие условия.
Полоса 1: Маркеры молекулярного веса.
Полоса 2: KCl - экстракт, приготовленный по способу примера 3.1, диссоциированных пяточных корнеоцитов.
Полоса 3: SCCE, очищенный по способу примера 4.1 с помощью афинной хроматографии на сделанном нерастворимым соевом трипсиновом ингибиторе.
b: зимография иммуноосаждений в 12,5% SDS-PAGE c 0,1% сополимеризованным термоденатурированным казеином, Coomassie - окрашенный гель.
Полоса 1: Маркеры молекулярного веса.
Полоса 2: KCl - экстракт, диализованный по способу a.
Полосы 3-6: Солюбилизованные иммуноосадки с /слева направо /мoab TE4b /20 мкг/, мoab TE9b /10 мкг/, мoab PZ /10 мкг/, и буферированный фосфатом физиологический раствор. Moab PZ представляет собой мышиное моноклональное антитело Jg-Gl- карра типа к протеину зоны беременности, и был использован как неродственный негативый контроль.
C: Иммуноблот из 12,5% SDS-PAGE /невосстанавливающие условия/.
Полоса 1: биотинилированные маркеры молекулярного веса, детектируемые конъюгатами щелочная фосфатаза-авидин /BioRad/. Электрофоретически обработанный образец в полосах 2, 4 и 6 тот же, что и в полосе 2 в a, и в полосах 3, 5 и 7 тот же, что в полосе 3 в a.
Полосы 2 и 3: Первое антитело = мoab TE4b, 0,2 мкг на мл.
Полосы 4 и 5: Первое антитело = мoab TE9b, 0,1 мкг/мл.
Полосы 6 и 7: Первое антитело = мoab PZ, 0,1 мкг/мл.
Острия стрелок в a-c указывают относительные молекулярные веса /сверху вниз/ 93, 66, 45, 31, 22 и 14 кД, соответственно.
Фиг. 15.
Фиг. 15 демонстрирует плазмиду pS 500. Эта плазмида содержит полной длины кДНК SCCE человека, клонированную pS C19. Подробнее см. пример 6.
Фиг. 16.
Норзернблоттинг с мРНК, полученными из эпидермиса человека. Поли-T-очищенные РНК, соответствующие примерно 100 г полный РНК, нанесены в каждой полосе.
1: Гибридизацию осуществляют с зондом, полученным из 1070 bp Hinc 2/Hinc 2 фрагментом SCCE-кДНК.
2: гибридизацию ведут с зондом, полученным из 655 bp Hinc 2/Bgl 2 фрагментом SCCE-кДНК.
Фиг. 17.
a. Coomassie - окрашенный SDS-PAGE, 12,5% гель. 1 и 2: PBS-Triton X-100 нерастворимый осадок обработанных ультразвуком TG 2 клеток, трансформированных pS 510 и pS 511 соответственно, и индуцированных iPTG. 3: SCCE, выделенные из роговичного слоя пяточной поверхности человека.
b. Иммуноблоты с цыплячьей пре-иммунной сывороткой /1-3/ и цыплячьими анти- SCCE /4-6/, 1 и 4: TG 2 клетки, трансформированные pS 510 и индуцированные IPTG 2 и 5: TG 2-клетки, трансформированные pS 511 и индуцированные IPTG. 3 и 6: SCCE, выделенные из роговичного слоя пяточной поверхности человека.
Образцы подготовлены кипячением в образцовом буфере с меркаптоэтанолом.
Фиг. 18.
На фиг. 18 представлена круговая карта вектора экспрессии pS 507 сконструированного по способу примера 9. За счет вектора pS 507, осуществляется экспрессия рекомбинантного SCCE человека в клетки млекопитающих.
Фиг. 19.
На фиг. 19 представлен анализ экспрессии рекомбинантного человеческого SCCE гена pS 507 в клетках млекопитающих.
Полоса 1: РНК клеток C127.
Полоса 2: РНК из выделенного клона, 1:24, клеток C127, трансфецированных pS 507.
Полоса 3: РНК из смеси популяций клонов C127, трансфецированных pS 507.
Полосы 4 и 5: РНК из C127 клеток, трансфецированных вектором экспрессии, pS 147, который идентичен pS 507, за исключением того, что в нем отсутствует SCCE кДНК последовательность. Маркеры размеров указаны слева.
Фиг. 20.
SDS-PAGE с последующим иммуноблоттингом SCCE, экспрессированных в C127 клетки.
Полосы 1 и 6: Предварительно окрашенные маркеры молекулярного веса /Bio-Rad, 106, 80, 49,5, 32,5, 27,5 и 18,5 кДа/.
Полоса 2: pS 507/C127, смесь, T-склянки.
Полоса 3: pS 507/C127, смесь, роллер A.
Полоса 4: pS 507/C127, смесь, роллер B.
Полоса 5: Негативный контроль pS 522/C127.
Полоса 7: Нативный SCCE, полученный из роговичного слоя.
Полоса 8: pS 507/C127, клон 24, T-склянки.
Полоса 9: pS 507/C127, клон 24, роллер A.
Полоса 10: pS 507/C127, клон 24, роллер B.
Фиг. 21.
SDS-PAGE /a/ и иммуноблоттинг /B/ рекомбинантного SCCE, выделенного из клеточной C127 культуральной среды за счет клеток, несущих плазмиду pS 507.
Полосы 1 и 5: Предварительно окрашенные маркеры молекулярных весов /Bio-Rad, 106, 80, 49,5, 32,5, 57,5 и 18,5 кДа/.
Полоса 2: Клеточная среда перед очисткой.
Полоса 3: Несвязанный материал, собранный с хроматограмм.
Полоса 4: Связанный материал, элюированный буфером с
низким pH.
Фиг. 22.
Нативный SCCE и активный рекомбинантный SCCE, анализированные на их активность на казеиновом полиакриламидном геле.
Полосы 1 и 10: Маркеры молекулярных весов /Pharmacia 14-94 кДа/.
Полоса 2: Нативный SCCE.
Полоса 3: Нативный SCCE.
Полосы 4-6: Рекомбинантный SCCE, расщепленный в течение 1 часа, 3 часов и в течение ночи, соответственно /460 нг/ячейку/.
Полоса 7: Трипсин в том же количестве, что и в образцах в полосах 4-6, но без APMSF.
Полоса 8: Трипсин в том же количестве, что и в образцах в полосах 4-6, в присутствии того же количества APMSF, что и в образцах.
Фиг. 23
Иммуноблот N-гликозидазы F® обработанных нативного и рекомбинантного SCCE. Образцы разделяют на 8-18% SDS-PAGE, и иммуноблоттинг проведен, как указано ранее.
Полоса 1: Маркер молекулярного веса. Молекулярные массы, считая сверху: 106, 80, 49,5, 32,5, 27,5 и 18,5 кДа.
Полосы 2 и 3: рекомбинантный SCCE, 0,3 и 3 мкг, соответственно.
Образцы в полосах 3 и 5 обработаны N-гликозидазой F®.
Примеры
Пример 1
Доказательство того, что клетки, отшелушивающиеся с поверхности ороговевшего поверхностного слоя кожи, содержат разложившиеся десмосомальные протеины, и что энзимом, который, по-видимому, ответственен за это, является химотрипсиноподобная протеиназа, которую могут ингибировать ионы цинка.
1.1 Десквамация в роговичном слое
Целью исследования было выяснить природу механизма, ответственного за клеточную когезию и диссоциацию поверхностных клеток /десквамацию/ в ороговевшем слое кожи, Stratum corneum. Чешуйки роговичного слоя толщиной 0,3 - 0,6 мм срезают параллельно поверхности кожи с пяточной поверхности добровольца с нормальной кожей. Этот кусочек ткани погружают в буферированный фосфатом физиологический раствор с 0,1% натрийазидом на три часа при комнатной температуре и слабо прикрепленные клетки с внешней поверхности /ин виво/ соскребают. Кусочки толщиной 1 мм, нарезанные перпендикулярно поверхности ткани, помещают затем в среду, содержащую 0,1 М Tris-HCl pH 8, 5 мМ ЕДТА, 0,1% азида натрия и 0,45% агарозы, непосредственно перед гелеобразованием среды за счет наличия агарозы. После инкубирования в течение 0,5 и 15 часов при 37oC, кусочки геля с тканью замораживают на сухом льду. 20 мкм замороженные кусочки нарезают перпендикулярно поверхности кожи в криостате, укрепляют и исследуют в фазовом контрастном микроскопе. Наблюдается непрерывное однополярное отшелушивание клеток в кусочков роговичного слоя пяточной поверхности, инкубируемых ин витро /фиг. 1/. Клетки отшелушиваются только с той поверхности тканей, которые ин виво были наружными. Таким образом, наблюдаемый процесс имитирует десквамацию.
1.2. Влияние температуры, pH и энзимных ингибиторов
Затем был разработан способ количественного определения отшелушивания клеток, который позволяет исследовать влияние различных параметров, таких, как температура, pH и энзимные ингибиторы. Цилиндрики роговичной оболочки пяточной поверхности с диаметром 3 мм получили с помощью устройства для биопсии из чешуек ткани, взятой и замороженной из слабо прикрепленных поверхностных клеток, как было указано ранее в 1.1. Эти цилиндрики /с определенной площадью поверхности, которая ин виво была обращена наружу/ инкубируют в 0,5 мл среды, содержащей 0,1 М Tris-HCl pH 8, 5 мМ ЕДТА, 0,1% азида натрия и апротинин /4•10-6 моль/л/ или без него /Bochringer-Mannheim, Germany/ в 1.5 мл склянках Эппендорфа при 37oC в течение 5, 10 и 20 часов, а затем перемешивают в течение 10 секунд в смесителе Vortex для отделения диссоциированных поверхностных клеток. Оставшуюся ткань удаляют и помещают в свежую среду для продолжения инкубирования. Ампулы с отделившимися клетками центрифугируют в течение 2 минут при 5000 g для сбора клеток. Клеточные осадки промывают один раз 0,5 мл буферированного фосфатом физиологического раствора, а затем обрабатывают при 60oC в течение 1,5 часа 0,6 мл гидроксида натрия. Растворимый в щелочи протеин определяют в соответствии с Lowry et al., 1951, и берут за меру количества отделившихся клеток. Полученные результаты представлены на фиг. 2.
Аналогичным образом исследуют влияние различных потенциальных ингибиторов /апротинина, ингибитора соевого трипсина, пепстатина /Bochringer-Mannheim, Germany/ и иодоацетамида /Sigma, St. Lоuis, MO/. Отдельные цилиндрики ткани в 2 мм приготавливают и инкубируют со средой с различными потенциальными ингибиторами /или без них/ в концентрациях, указанных в таблице 1 далее, в течение 16 часов при 38oC, и отделившиеся клетки подсчитывают, как было указано ранее. Следует отметить, что ЕДТА /которая представляет собой ингибитор металлопротеиназ/ была включена в инкубационную среду для достижения оптимальных скоростей отторжения клеток.
Было обнаружено, что ингибиторы серинпротеиназы, апротинин и ингибитор соевого трипсина эффективно ингибируют отшелушивание клеток /фиг. 2 и таблица 1/. Так как эти два вещества были ингибиторами, но ингибиторами металлопротеиназ /ЕДТА/, тиолпротеиназ /иодоацетамид/, а аспарагиновых протеаз /пепстатин/ не были, был сделан вывод, что серинпротеиназа принимает участие в наблюдаемом процессе. Пришли также к выводу, что клеточная когезия в роговичном слое зависит от протеиновых структур, и что механизм, аналогичный наблюдаемому ин витро, должен также работать и в процессе десквамации ин виво / Lundstrom and Egelrud, 1988/.
В дальнейших исследованиях клеточного отшелушивания ин витро с роговичного слоя пяточной поверхности было обнаружено, что этот процесс можно разделить на две отдельные стадии. Первая стадия имеет место независимо от того, присутствует ли ЕДТА в инкубационной среде или нет. Вторая стадия происходит только в присутствии ЕДТА. Первую стадию можно ингибировать химостатином и ионами цинка, помимо апротинина. Вторую стадию можно ингибировать апротинином и химостатином /Lundstrom and Egelrud, 1990 a/. Химостатин является низкомолекулярным ингибитором протеиназ с химотрипсино-подобной специфичностью к субстратам. Кроме того, было обнаружено /Lundstrom and Egelrud, 1990 a/, что леупептин, низкомолекулярный ингибитор протеиназ с трипсиноподобной специфичностью к субстрату, не оказывает влияния на ин витро отшелушивание клеток.
Наиболее вероятно, что протеиновые структуры, которые ответственны за клеточную когезию в роговичном слое, и таким образом являются возможными кандидатами на деградацию в описанном ранее десквамационноподобном клеточном отшелушивании, являются десмосомами. Десмосомы состоят из двух симметричных половинок, которые расположены в непрерывных клетках. Две половины соединены во внеклеточном пространстве трансмембранными протеинами, называемыми десмоглеинами.
1.3 Судьба десмоглеина 1 /DG1/ в процессе отшелушивания клеток ин витро
Исследуют судьбу десмоглеина 1 /DG1/ в процессе отшелушивания клеток ин витро с роговичного слоя пяточной поверхности. Роговичный слой пяточной поверхности инкубируют, как было указано ранее, в 1.1, и все еще когезивную ткань отделяют от диссоциированных клеток. Клетки и ткань экстрагируют в буфере, содержащем 0,1 М Tris-HCl pH 9, 9 М мочевины, 2% додецилсульфата, 1% меркаптоэтанола, 1 мл буфера на 20 мг ткани, в течение 15 часов при 37oC. Экстракты подготавливаются для электрофореза на полиакриламидном геле в 7,5% гелях в присутствии натрийдодецилсульфата /SDS-PAGE/ в соответствии с указаниями Laemmli /Laemmli, 1970/, с последующим электрофоретическим переносом /Towbin et al., 1979/ на нитроцеллюлозную мембрану /Bio-Rad, Richmond, CA/, которая была зондирована кроличьей поликлональной антисывороткой, полученной против DG1 выделенных с бычьих морд /Gorbsky et al., 1985/. Связанные антитела детектируют с помощью щелочной фосфатазы, конъюгированной с козьими антикроличьими иммуноглобулинами /Dio-Rad, Richmond, CA/, /Вlake, et al., 1984/.
Полученные результаты представлены на фиг. 3. Количества образца, добавляемого к иммуноблотам, устанавливают таким, чтобы установить примерно одинаковые концентрации протеинов (что можно определить визуально по окрашенным Coomassie-blue SDS-PAGE гелям) для цельных тканей и диссоциированных клеток. Проводят несколько разбавлений экстрактов для того, чтобы обеспечить полу-количественное сравнение количеств различных анти- DG1 реактивных компонентов в цельном роговичном слое и диссоциированных клетках.
Если все еще цельную ткань и диссоциированные клетки экстрагируют отдельно, обнаруживается, что тогда как все еще цельная ткань содержит только, по-видимому, интактные DG1, диссоциированные поверхностные клетки содержат только предполагаемые продукты разложения этого протеина /фиг. 3/.
На фиг. 4 и 5 представлены влияния апротинина, ионов цинка, химостатина /Bochringer-Mannheim, Germany/ и леупептина /Bochringer-Mannheim, Germany/ на деградацию DG1 в процессе ин витро клеточного отшелушивания с роговичного слоя пяточной поверхности.
Во-первых, исследовали временную зависимость и влияние апротинина на деградацию десмоглеина 1 /DG1/ в отшелушивании клеток роговичного слоя пяточной поверхности ни витро. Экстракты роговичного слоя пяточной поверхности инкубируют, как указано в 1.2 /но без выделения цельной ткани от диссоциированных клеток перед экстрагированием/ в присутствии апротинина /15 мкл/ или без него, а затем экстрагируют спустя 0, 6, 12 или 24 часа. SDS-PAGE и иммуноблоттинг проводят как указано ранее. Денситометрическое сканирование иммуноблотов проводят на Shimadgu CS-900 сканирующем устройстве с летящим пятном /Shimadgu, Kyoto, Japan/ в отраженном свете на 560 нм в зигзагообразном режиме. Полученные результаты приведены на фиг. 4. Отмечается эффективное ингибирование апротинином деградации DG1.
Затем исследуют влияние ионов цинка, химостатина и леупептина на деградацию десмоглеина 1 /DG1/ в процессе отшелушивания клеток роговичного слоя пяточной поверхности ин витро. Схема эксперимента аналогична описанной ранее. Инкубирование ведут в течение 24 часов с сульфатом цинка в концентрациях 0, 1 или 5 мМ, и с химостатином или леупептином в концентрациях 330 мкМ. Полученные результаты демонстрируют ингибирование трансформации анти-DG1 позитивных компонентов из 160 кД в 95 кД за счет ионов цинка и химостатина, но не за счет леупептина.
Из этих результатов, очевидно, следует что апротинин, ионы цинка и химостатин оказались ингибиторами, а леупептин - нет. Так, характер ингибирования деградации DG1 оказался таким же, что и для отшелушивания клеток роговичного слоя пяточной поверхности ин витро /Lundstrom and Egelrud, 1990 b/.
Считалось важным продемонстрировать, что механизмы, аналогичные тем, которые ответственны за отшелушивание клеток с роговичного слоя ладоней-подошв, проявляются также в роговичном слое кожи с других участков тела, нежели взятые с ладоней и подошв. Takahashi et al. / Takahashi et al., 1987/ сообщил, что смесь детергентов N,N-диметилдодециламиноксида /Sigma, St. Lоuis, MO/ и натрийдодецилсульфата /Bio-Rad, Richmond, CA/ в молярном отношении 8:2 вызывает клеточную диссоциацию в роговичном слое, взятом не с поверхностей ладоней-подошв, подготовленном за счет трипсинизации целого эпидермиса. Чтобы избежать загрязнений за счет экзогенного трипсина, биопсин нормальной кожи человека с глутеальных участков инкубируют при pH 8 с вышеуказанной смесью детергентов и ЕДТА /Egelrud and Lundstrom, 1990/. Было обнаружено, что в таких условиях ороговевшие слои диссоциируют до отдельных клеток. Добавление апротинина к инкубационной среде предотвращает такую клеточную диссоциацию. Был сделан вывод, что клеточная когезия и в роговичных слоях, взятых не с ладоней-подошв, зависит от структур тканей, то есть, десквамация в такой ткани зависит от протеолиза, и что эта ткань содержит протеиназу, которая может катализировать такой протеолиз. Так как клеточная диссоциация включает лишь роговичный слой, а не более глубокие слои, не неороговевшие слои эпидермиса, был сделан вывод, что ответственная за этот процесс протеиназа находится в более глубоких слоях в неактивном или ингибированном состоянии. Пример 2
Открытие химотрипсинового энзима роговичного слоя /SCCE/: протеиназы, которая удовлетворяет критериям ответственности за деградацию внутриклеточных когезивных структур в роговичном слое ин витро, и возможно также ин виво.
Из экспериментов примера 1 был сделан вывод, что протеиназа, ответственная за диссоциацию однополярных поверхностных клеток в модели ин витро десквамации роговичного слоя пяточной поверхности, должна обладать следующими свойствами:
1. Она должна присутствовать в роговичном слое.
2. Она должна быть сериновой протеиназой.
3. Она должна обладать химотрипсино-подобной специфичностью к субстрату и характером ингибирования, аналогичным тому, который наблюдают при отшелушивании клеток ин витро, и связанному с деградацией десмоглеина 1.
4. Она должна быть локализована внеклеточно в роговичном слое.
5. Она должна быть зависима от pH, что позволяло бы ей быть активной в физиологических условиях, так как pH роговичного слоя составляет примерно 4,5 - 6.
6. Так как отшелушивание клеток с роговичного слоя пяточной поверхности ин витро протекает непрерывно во время пролонгированного времени инкубирования, даже если объем инкубационной среды очень велик по сравнению с объемом инкубируемых кусочков ткани, или если инкубационная среда повторно меняется в процессе инкубирования, кажется разумным предположить, что ответственный энзим связан с тканью таким образом, что это не позволяет экстрагировать его в инкубационную среду в процессе инкубирования.
Нижеследующие два эксперимента привели к открытию SCCE /Egelrud and Lundstrom, 1991/:
2.1 Энзиматическая активность связана с диссоциированными корнеоцитами пяточной поверхности
Диссоциированные клетки роговичного слоя пяточной поверхности /корнеоциты/ получают за счет инкубирования роговичного слоя пяточной поверхности по способу примера 1. Клетки фильтруют через найлоновую сетку с размером отверстий 10 мкМ, а затем трижды промывают десятью объемами 0,1 М Tris-HCl pH 8, 5 мМ ЕДТА и трижды 0,1 M Tris-HCl pH 8. Затем эти клетки инкубируют с двумя типами хромогенных протеиназных субстратов S-2288 или S-2586 /Kabai Diagnostica, Stockholm, Sweden/:
Ile-Pro-Arg-p-нитроанилид /S-2288/ расщепляется широким кругом сериновых протеиназ с аргинино-специфичностью /например, трипсин/. Arg-Pro-Tyr-p-нитроанилид /S-2286/ является субстратом для химотрипсино-подобных протеиназ.
В полном объеме 120 мкл каждая реакционная смесь содержит 0,07 M Tris-HCl pH 8, 0,1% азида натрия, 1, 2, 5, 5 или 10 мкл 25% суспензии промытых корнеоцитов с пяточной поверхности и 1,04 мМ /S-2586/ или 1,25 мМ /S-2288/ субстрата. После инкубирования в течение 5 часов при 37oC в микротитровальных пластинах реакцию останавливают, добавляя 125 мкл 10% уксусной кислоты. Клеткам дают осесть, и по 200 мкг каждой надосадочной жидкости переносят в новые ячейки. За гидролизом двух субстратов следят, измеряя изменения в поглощении на 405 нм, после того, как клетки переносят в Behring Elisa Processor /Behruigwerke, Marburg, Germany/.
Как показано на фиг. 6, существует энзиматическая активность, связанная с диссоциированными корнеоцитами пяточной поверхности, которая катализирует гидролиз S-2586. При сравнении активность по отношению к S-2288 оказывается низкой.
pH-зависимость гидролитической активности S-2586 исследуют после этого. Схема эксперимента соответствует представленной ранее, используют 10 мкл 25% суспензии промытых корнеоцитов пяточной поверхности и буферы с различными pH /ацетат натрия, фосфат натрия или Tris-HCl/. Конечная концентрация буфера составляет 0,07 М. Полученные результаты представлены на фиг. 7, из которого следует, что активность оптимальна при pH 7 - 8, но достаточно значительна и при pH 5,5.
В дальнейшем эксперименте исследуют влияния ЭДТА, ионов металлов и ингибиторов протеиназ на гидролиз S-2586 при pH 8 за счет протеиназы, связанной с супендированными клетками роговичного слоя пяточной поверхности. Схема эксперимента представлена ранее и в таблице 2.
Примечания к табл. 2:
a/ Вещество присутствует в аналитической смеси;
b/ 25% суспензия клеток роговичного слоя пяточной поверхности, приготовленная как указано в тексте, была предварительно инкубирована в течение 1 часа при комнатной температуре в присутствии 1 мМ PMSF /Sigma, St. Louis, MO/, растворенного в 2-пропаноле /конечная концентрация 2-пропанола = 4% объем/объем/. Контрольные суспензии предварительно инкубируют только с 4% 2-пропанолом.
c/ Ингибитор, растворенный в диметилсульфоксиде /ДМСО/. Все среды, включая контроли, содержат 5% /объем/объем/ ДМСО.
Как видно из таблицы 2, фенилметилсульфонилфторид /PMSF, общий ингибитор сериновых протеиназ/, апротинин, ингибитор соевого трипсина, химостатин /ингибитор химотрипсина/, и ионы цинка, но не леупептин /ингибитор трипсина/ ингибируют S-2586 гидролитическую активность. Характер ингибитора поэтому весьма аналогичен характеру, наблюдаемому при отшелушивании клеток ин витро и связанной деградацией десмоглеина 1.
2.2 Зимография диссоциированного роговичного слоя пяточной поверхности
Было обнаружено, что энзим, ответственный за гидролитическую активность S-2586, раскрытую в 2.1, может быть солюбилизирован, если корнеоциты экстрагируют 1 M KCl в 0,1 M Tris-HCl pH 8. Поэтому были проведены эксперименты с зимографией. По этой причине KCl экстракты корнеоцитов приготовили для электрофореза по способу Лаэмли /Laemmli, 1970/, но без восстанавливающего агента в образцовом буфере и не нагревая образцы. Приготовили также образцы за счет экстрагирования диссоциированных корнеоцитов пяточной поверхности с образцовым буфером Лаэмли без восстанавливающего агента при комнатной температуре. Для зимографии адаптировали модифицированную процедуру Horie et al. /Horie et al., 1984/. Электрофорез на полиакриламидном геле ведут в присутствии натрийдодецилсульфата /SDS-PAGE/ по способу Лаэмли в 12,5% гелях с 1% сополимеризованным термоденатурированным казеином. После электрофореза эти гели погружают в буфер, содержащий 2% Triton X-100 на 1 час при комнатной температуре для удаления SDS, а затем инкубируют при 37oC в течение 15 часов. Затем гели окрашивают Cocmassie-blue. Отдельные казеинолитические энзимы проявились как прозрачные полосы на голубом фоне. См. также пояснения к фиг. 8 для подробностей эксперимента.
Полученные результаты представлены на фиг. 8. Экстракты корнеоцитов пяточной поверхности содержат один основной казеинолитический энзим с кажущимся молекулярным весом около 25 кД. Наблюдаются также меньшие казеинолитические энзимы с молекулярными весами около 30 кД. /Эти меньшие компоненты на фиг. 8 видны не отчетливо. Позднее было обнаружено, что их можно ингибировать леупептином, а не химостатином/. 25 кД энзим обладает значительной активностью при pH 5,5 - 8. Его можно ингибировать аротинином, ионами цинка и химостатином, но не леупептином. Таким образом, он имеет такой же характер ингибирования, что и указанная ранее гидролитическая активность S-2586. В экспериментах с гельхроматографией исключения /не показаны/ 25 кД казеинолитический энзим, как было обнаружено, проявляется вместе с S-2586 гидролитической активностью.
В последующих экспериментах /не представлены/ с описанной ранее методикой в 2.2, было обнаружено, что роговичный слой не с ладоней и подошв содержит энзим со свойствами, практически идентичными свойствам 25 кД протеиназы, связанной с корнеоцитами пяточной поверхности, которые с этого момента будем называть химотрипсиновым энзимом роговичного слоя /SCCE/ /Lundstrom and Egelrud, 1991/.
Оказалось возможным также получить доказательства того, что SCCE связан с корнеоцитами пяточной поверхности таким образом, что это позволяет ему быть активным во внеклеточном пространстве роговичного слоя. Это было сделано первой демонстрацией того, что диссоциированные корнеоциты проницаемы для пероксидазы хрена /Мг 44 кД/. Было показано затем, что фибриноген человека /Мг 340 кД/ может деградировать под действием суспензии корнеоцитов, и что эту деградацию можно ингибировать теми же ингибиторами, что и SCCE. Отсюда можно сделать вывод, что деградация фибриногена связана с солюбилизированным энзимом /Egelrud, 1992/.
Пример 3
Частичная очистка химотрипсинового энзима роговичной оболочки /SCCE/ и анализ протеиназы с хромогенными субстратами
3.1 Получение KCl экстрактов корнеоцитов пяточной поверхности
Получение диссоциированных корнеоцитов пяточной поверхности по способу примера 1 было проведено в большем масштабе, и были получены KCl-экстракты промытых корнеоцитов пяточной поверхности, содержащие SCCE, как описано в примере 2.
Получение KCl экстрактов корнеоцитов пяточной поверхности схематически представлено в таблице 3 далее. Гиперпластичный роговичный слой пяточной поверхности собирают с помощью общества Swedish Pedicyrists.
Использовали только материал, полученный с помощью ножниц или щипцов. Материал не брали с ног, пораженных заболеваниями, связанными с отшелушиванием. Перед посылкой роговичный слой сушили на воздухе и упаковывали в пластиковые пакеты. В лаборатории материал хранился при -20oC до использования.
Для каждой последующей стадии афинной хроматографии собирают экстракты 1 и 2 (из двух получений по 50 г каждое) роговичного слоя пяточной поверхности
3.2 Анализы протеиназ хромогенными субстратами
Проводят сравнение SCCE, бычьего химотрипсина и катепсина G человека в отношении влияния ингибиторов апротинина, химостатина, сульфата цинка и специфичности субстрата.
Исходный раствор Meo-Suc-Arg-Pro-Tyr-pNA /S-2586/ приготавливают в дистиллированной воде, раствор Suc-Ala-Ala-Pro-Phe-pNA /Bochringer, Mannheim, Germany/ в 1-метил-2-пирролидоне /конечная концентрация растворителя в инкубационных смесях 4%/ и химостатина в диметилсульфоксиде /конечная концентрация растворителя в инкубационных смесях 1%/. Катепсин G из гнойной мокроты человека получают от E.Lotti, Geneva, Switzerland. Источником SCCE служит KCl-экстракт диссоциированных корнеоцитов роговичного слоя пяточной поверхности, полученных как описано ранее. Источники ингибирования апротинина, химостатина и сульфата цинка описаны ранее.
Инкубирование ведут при 37280C в микротитровальных пластинах. Полный объем инкубирования составляет 135 мкл. Каждая инкубационная смесь содержит Tris-HCl pH 8,0 /конечная концентрация 0,08 M/, KCl /концентрация 0,2 M/, 100 мкл раствора субстрата, 25 мкл источника энзима /соответствующим образом разбавленного в 0,1 M Tris-HCl pH 8,0, 1,0 M KCl/ и 10 мкл раствора ингибитора.
На фиг. 9 A-C, Meo-Suc-Arg-Pro-Typ-pNA /S-2586, начальная концентрация 1,2 мМ/ использован в качестве субстрата. В Д начальная концентрация обоих субстратов составляет 1,2 мМ.
К концу инкубирования /1,5 часа/ добавляют по 125 мкл 10% уксусной кислоты в каждую ячейку, и поглощение считывают на 405 нм, используя в качестве сравнения инкубационные смеси без добавления энзимов. Количество добавляемых энзимов устанавливают таким, чтобы изменения поглощения на 405 нм к концу инкубаций составляло бы 0,3 - 0,7.
Полученные результаты суммированы на фиг. 9, A - D. Для того чтобы изучить влияние ингибиторов, в качестве субстрата используют S-2586. Эффективность апротинина в качестве ингибитора SCCE и химотрипсина оказалась высокой и примерно одинаковой для этих двух энзимов. С другой стороны, влияние на катепсин G гораздо меньше /фиг. 9 A/. Химостатин приводит к ингибированию всех трех энзимов, но концентрация ингибитора, которая обеспечивает 50% ингибирование более чем на три порядка величины выше для SCCE, нежели для химотрипсина и катепсина G /фиг. 9 B/. Сульфат цинка оказывается эффективным ингибитором SCCE, но не химотрипсина и катепсина G /фиг. 9 C/. Активность трех энзимов против субстратов Meo-Suc-Arg-Pro-Tyr-pNA /S-2586/ и Suc-Ala-Ala-Pro-Phe-pNA сравнивают /см. фиг. 9 D/. Так как цель состояла в определении сходства или различий между исследованными энзимами, эти эксперименты проводили только с одной исходной концентрацией для каждого субстрата. Тогда как Suc-Ala-Ala-Pro-Phe-pNA являются, по-видимому, гораздо лучшим субстратом, нежели S-2586 для химотрипсина и катепсина G, для SCCE справедливо обратное.
Пример 4
Очистка N-терминальной аминокислотной последовательности, определяющей химотрипсиновый энзим роговичного слоя
4.1. Выделение SCCE из KCl-экстрактов корнеоцитов с помощью афинной хроматографии на несолюбилизированном ингибиторе соевого трипсина /SBTI/.
На фиг. 10 представлены результаты афинной хроматографии на SBTI. Афинный гель приготавливают, связывая 50 мг SBTI /Bochringer, Mannheim, Germany/ c 12 мл осажденного Affigel 15 /Bio-Rad, Richmand, CA/, в соответствии с рекомендациями изготовителя. Остальные активные группы на геле были блокированы этаноламином. Объединенные KCl-экстракты из 100 г /сухой вес/ роговичного слоя пяточной поверхности /полный объем 700 мл/ пропустили через 0,8х2 см слой SBTI-Affigel 15, набитый в стеклянную колонку, при скорости 42 мл/час с непрерывной записью поглощения элюата на 280 нм. Колонку промывают 0,1 M Tris-HCl pH 8, 1 M KCl, до тех пор, пока поглощение элюата не становится менее 0,01, а затем 10 мл 0,1 M Tris-HCl pH 8. Поэтапное элюирование связанного материала осуществляют HCl при 1, 10 и 100 мМ. Элюент заменяют, когда поглощение элюата снижается ниже 0,01. Фракции по 3 мл собирают в тестовые ампулы, которые содержат Tris-HCl pH 8, полный объем 0,4 мл, в количестве, рассчитанном таким образом, чтобы этого было достаточно для установления pH элюата выше 7. pH каждой фракции немедленно проверяют и, при необходимости, устанавливают около 7 за счет небольших объемов 1 M Tris-HCl, pH 8. Анализы пептидной гиролизующей активности с S-2586 /субстрат для SCCE/ и S-2288 /субстрат для трипсино-подобных энзимов/ проводят по способу примера 2.1. Начальная концентрация обоих субстратов в аналитических смесях составляет 1,1 мМ. Примерно 90% S-2586 гидролизующей активности связана с гелем. После поэтапного элюирования промытого геля с 10 - 100 мМ HCl, примерно 60% от полной S-2586 гидролизующей активности в нанесенном KCl-экстракте можно выделить. Из полной S-2288 гидролизующей активности около 20% оказывается связанной с афинностью геля и 10% можно выделить в элюате.
Фиг. 11 демонстрирует анализ элюата из SBTI-афинной хроматографии с электрофорезом на полиакриламидном геле в присутствии SDS /SDS-PAGE/ и с зимографией. Обрабатывали на 12,5% гелях невосстановленные образцы. См. также Egelrud and Lundstrom, 1991 и пример 2, 2.2 для подробностей эксперимента. Перед подготовкой для электрофореза образцы концентрируют примерно в 20 раз в A с помощью центрифужной фильтрации на Ultrafrec MC-фильтрах /срезах 10 кД: Millipore, Bedford, MA/ и разбавляют в 10 раз в B.
Фиг. 12 представляет сравнение с помощью SDS-PAGE невосстановленного и восстановленного образцов из SBTI-афинной хроматографии.
Как следует из фиг. 10 и 11, SBTI-афинная хроматография дает протеин, который имеет степень чистоты более 90% /о чем можно судить по Coomassie-blue-окрашенным SDS-PAGE гелям/ с кажущимся молекулярным весом около 25 кД в невосстановленной форме и около 28 кД в восстановленной форме. Кроме того, существует небольшая Coomassie-blue-позитивная компонента с кажущимся молекулярным весом примерно на 1 кД больше, нежели основная компонента. При зимографии гелей наблюдается одна основная и одна небольшая полоса с теми же самыми электрофоретическими подвижностями, что и у двух полос, детектируемых на Coomassie-blue-окрашенных гелях с невосстановленными образцами. Обе эти казеинолитические компоненты можно ингибировать химостатином. В дополнение, зимография показывает небольшие компоненты с кажущимися молекулярными весами около 30 кД, которые можно ингибировать леупептином. Был сделан вывод, что основным выделенным протеином является SCCE.
4.2. Анализ N-терминальной аминокислотной последовательности SCCE
200 мкл фракции из хроматограммы с SBTI-Affigel 15, A1 нм 0,2, приготавливают из SDS-PAGE с восстановлением или без него, и обрабатывают на 12,5% полиакриламидном геле /толщина 1 мм, ширина щели 73 мм/. После электрофореза выделенные протеины переносят электрофоретически на Immobilon фильтр /Millepore/ и окрашивают Coomassie-blue по способу Matsuidaira, 1987. Основную протеиновую полосу вырезают и обрабатывают в Applied Biosystems 477A импульсном жидкофазном анализаторе аминокислотных последовательностей с подключенным PTH 120A анализатором /Applied Biosystems Inc., Foster Gity, CA, USA/. Секвенирование осуществляют в соответствии с установленными циклическими программами и химикалиями от изготовителей. Начальный и повторные выходы, рассчитанные с учетом стандартных протеинов, составляют 25 и 97% соответственно.
Выходы производных аминокислот соответствуют только одному секвенированному пептиду. Для невосстановленных образцов выходы были хорошими на стадиях 1 - 6, но падали до 0 на стадиях 7 и 9. Выходы на последующих стадиях были заметно снижены. Хотя для восстановленных образцов на стадиях 7 и 9 не были обнаружены производные аминокислот, но для последующих стадий, где можно было детектировать производные, падения выходов не наблюдалось. Эти результаты дают возможность предположить, что в положениях 7 и 9 имеются цистеины. Однако, оказалось невозможным обнаружить карбоксиметилированный цистеин на стадиях 7 и 9 после восстановления и обработки иодоуксусной кислотой /100 мМ/. Полученная последовательность /фиг. 13, последовательность 1D N 3/ была идентична для восстановленных и невосстановленных образцов.
Пример 5
5.1. Получение SCCE-специфических моноклональных антител
Мышам BALB/c /Bomholtgaard, Denmark/ вводят примерно 30 мкг нативного SCCE, очищенного по способу примера 4.1, в полном адъюванте Фрейнда /Difco Laboratories, Detroit, MI/ в виде подкожных инъекций. Инъекции с тем же количеством SCCE в неполном адъюванте Фрейнда /Difco Laboratories, Detroit, MI/ повторяют через месяц. Спустя 4 месяца после первой инъекции одной мыши внутривенно вводят бустерные инъекции в 3 последующих дня, по 30 мкг антигена за инъекцию. Гибридомы получают по способу, описанному Carlsson et al., 1985, с клетками SP2/0 миеломной клеточной линии /ATCC CRL 1581/. Идентификацию антител, реагирующих с очищенными SCCE-препаратами, ведут с помощью ELISA. Надосадочные культуральные жидкости позитивных клонов анализируют далее с помощью иммуноблоттинга после SDS-PAGE. Клоны, которые продуцируют антитела, реагирующие с SCCE в этом тесте, размножают в мышиной асцитной жидкости, и антитела выделяют с помощью протеин A афинной хроматографии и классифицируют по способу, описанному в Carlsson et al., 1985. Получают два подходящих антитела, мoab Te4b и мoab TE9b, причем оба они классифицированы как LgGo-каппа.
Характеризация мoab Te4b и мoab TE9b с помощью иммуноосаждения и иммуноблоттинга представлена на фиг. 14.
Фиг. 14a /полоса 2/ представляет Coomassie blue - крашенный SDS-PAGE гель с концентрированным KCl-экстрактом диссоциированных корнеоцитов пяточной поверхности, полученный по способу примера 3.1. Образец диализуют в течение 4 часов против 0,1 M ацетата натрия, pH 4, и концентрируют приблизительно в 100 раз за счет ультрафильтрации перед проведением электрофореза. фиг. 14a /полоса 3/ представляет получение SCCE, очищенного по способу примера 4.1.
Фиг. 4b представляет результаты эксперимента иммуноосаждения, в котором антитела были инкубированы с KCl-экстрактом корнеоцитов, а затем выделены за счет инсолюбилизированного протеина A. Повторно солюбилизированные и диссоциированные комплексы антитело-антитело были проанализированы с помощью зимографии по способу примера 2.
250 мкл KCl-экстракта диссоциированных корнеоцитов пяточной поверхности, которые были в 5 раз сконцентрированы за счет ультрафильтрации, диализуют против буферированного фосфатом физиологического раствора, к которому был добавлен альбумин бычьей сыворотки /Sigma, St. Louis, MO/ до конечной концентрации 10 мг/мл, смешивают с 10 мкл раствора антител или буферированного фосфатом физиологического раствора, и инкубируют в течение 15 часов при 4oC. 25 мкл осажденной протеин A Сефарозы /Pharmacia, Uppsala, Sweden/ добавляют затем в ампулы, и инкубирование продолжают при осторожном встряхивании при комнатной температуре в течение 2 часов. Гель выделяют центрифугированием и пять раз промывают 1 мл 0,05% Tween 20 /Sigma, St. Lоuis, MO/ в 0,05 M Tris-HCl, pH 7,5, 0,5 M NaCl. После окончательной промывки гель экстрагируют 100 мкл образцового буфера Лаэмли без восстанавливающего агента в течение 1 часа при комнатной температуре. Эти экстракты очищают центрифугированием и наносят на гель.
Мoab TE4b и TE9b, оба осаждают казеинолитический энзим, с тем же самым Мг, что и очищенный SCCE, и соответствующий основной казеинолитический энзим в KCl-экстракте. Антитела не осаждают меньшие казеинолитические энзимы в экстракте с Мг около 30 кДа, которые, как было показано, ингибируются леупептином, ингибитором трипсино-подобных сериновых протеиназ. В дополнение к 25 кДа казеинолитическому энзиму антитела, по-видимому, связывают небольшой протеинолитический компонент с Мг около 80 кДа. Этот компонент обычно присутствует в KCl-экстрактах корнеоцитов пяточной поверхности, полученных из ткани, которая была высушена перед приготовлением, но не был обнаружен в препаратах из свежей ткани /T.Egelrud, неопубликованные наблюдения/.
Он не присутствует в SCCE-препаратах, выделенных с помощью афинной хроматографии, и может присутствовать в продуктах агрегации. Не удалось обнаружить соответствующую компоненту, реагирующую с антителами на иммуноблотах.
На иммуноблотах SDS-PAGE гелей, проведенных в невосстанавливающих условиях, /фиг. 14c/ мoab TE4b и TE9b реагируют с компонентой, присутствующей в KCl-экстрактах корнеоцитов пяточной поверхности /фиг. 14 с полосы 2 и 4/ и в очищенных SCCE-препаратах /фиг. 14c, полосы 3 и 5/, причем оба они имеют то же Мг, что и основной выделенный протеин и основная казеинолитическая компонента на зимограммах. Эти антитела оказались нереакционноспособными с образцами, которые были восстановлены в присутствии SDS, что предполагает, что они направлены против конформационно-зависимых эпитопов.
Помимо основного протеина с Мг около 25 кД в невосстановленной форме, очищенный SCCE-препарат содержит небольшую Coomasie blue компоненту с Мг около 26 кД /невосстановленный; см. пример 3/. На зимограммах соответствующая казеинолитическая компонента присутствует и может быть ингибирована химостатином способом, аналогичным способу для основной 25 кД казеинолитической компоненты /см. пример 3/. При более высоких концентрациях мoab TE4b и TE9b /результаты не представлены/ также можно видеть эту небольшую компоненту, реагирующую с антителами на иммуноблотах. Аналогичные результаты /не представлены/ были получены с поликлональными кроличьими антителами, выработанными против Coomassie blue-позитивной компоненты, выделенной с помощью препаративного электрофореза. Точное соотношение между двумя протеинами с SCCE-подобной активностью и кажущейся иммунологической перекрестной реакцией неизвестно.
5.2. Антитела, специфические для поликлональных SCCE.
5.2.1. Цыплячьи анти-SCCE
45 мкг SCCE, выделенных с помощью SBTI-афинной хроматографии по способу примера 4.1, в 0,2 мл 0,1 M Tris-HCl термоденатурируют в течение 60 минут при 60C и гомогенизируют в равном объеме адъюванта Фрейнда /Difco Laboratories/.
Полученную эмульсию вводят подкожно в Derco цыплят, приблизительно 20-недельного возраста, у которых отбирали образцы крови для приготовления пре-иммунной сыворотки. Цыплятам ввели дополнительные подкожные инъекции эмульсий, полученных как описано ранее, но с неполным адъювантом Фрейнда и с 30 мкг очищенного, термоденатурированного SCCE /полный объем каждой эмульсии 250 мкл/ после 3, 5 и 7 недель. У цыплят собрали кровь через 2 недели после последней инъекции. Эту кровь немедленно смешивают с 2 объемами раствора Alsever's /на 100 мл: 1,87 г глюкозы, 0,8 г цитрата натрия, 0,62 г хлорида натрия, лимонная кислота до pH 6,1/ и центрифугируют. Цыплячьи анти-SCCE, выбранные для дальнейших исследований, используют в разбавлениях 1/2000 в экспериментах с иммуноблоттингом. См. фиг. 17, пример 8 для иллюстрации специфичности антисыворотки.
5.2.2. Кроличий анти-SCCE
SCCE, выделенный с помощью SBTI-афинной хроматографии, подвергают SDS-PAGE без восстановления, как описано в примере 4.1, на гелях с толщиной 15 мм в соответствии с данными Лаэмли, 1970. Основная протеиновая полоса, которая как ранее было показано, является SCCE, была визуализирована с помощью метода окрашивания хлоридом меди, в соответствии с работой Lee et. al., 1987, и ее вырезают. После удаления хлорида меди с ЕДТА по способу Lee et. al. , срезы геля гомогенизируют в буферированном фосфатом физиологическом растворе. Образцы гомогенизированных кусочков геля суспендируют в равных объемах адъюванта Фрейнда. Приблизительно 30 мкг чистого SCCE, полученного таким образом в полном адъюванте, вводят подкожно кролику. Через 3, 5 и 7 недель инъекцию повторяют с тем же количеством SCCE, но с неполным адъювантом. Кровь у кролика отбирают через две недели после последней инъекции.
Полученный кроличий анти-SCCE /Д-5/ используют в разбавлении 1/500 - 1/1000 в экспериментах с иммуноблотами со щелочной фосфатазой, конъюгированной с анти-кроличьими иммуноглобулинами в качестве второго антитела.
Во всех экспериментах иммуноблоттинга связанные вторые антитела определяют по способу Blake et. al., 1984 (относится к примерам 5, 8 и 9).
Кроличьи анти-SCCE Bo-1 получают таким же образом, но с SCCE, которые были восстановлены перед SDS-PAGE, в качестве антигена.
5.3. Иммуногистохимические исследования с моноклональными антителами
В иммуногистохимических исследованиях с SCCE-специфическими моноклональными антителами, SCCE можно детектировать в высоких супрабазальных клетках кератинизирующего чешуйчатого эпителия человека /эпидермис, внутренняя оболочка корней волосяных фолликул, твердая пластина/, но не в некератинизированном чешуйчатом эпителии /внутренняя корневая оболочка волосяных фолликул, слизь губ и с внутренней поверхности щек/. SCCE можно специфически экспрессировать в кератинизирующий чешуйчатый эпителий. Кроме того, было обнаружено, что SCCE экспрессируется в высокие супербазальные клетки эпидермиса человека, реконструированные ин витро и выращенные на границе раздела воздух-вода. Если к среде добавить ретинольную кислоту в концентрации, которая стимулирует пролиферацию кератиноцитов, но ингибирует образование роговичного слоя, SCCE больше не экспрессируется. Это предполагает, что SCCE экспрессия может быть частью программы дифференциации эпидермиса.
Результаты иммуноэлектромикроскопических экспериментов с SCCE-специфическими моноклональными антителами соответствуют роли SCCE в десмосомальной деградации и, следовательно, в десквамации. Антитела, специфически меченные ламелларными телами, претерпевают секрецию в межклеточное пространство между самыми гранулярными клетками и самыми нижними клетками роговичного слоя, тогда как в роговичном слое антитела распознают эпитопы в тесной связи с десмосомами во внеклеточном пространстве.
Пример 6
Клонирование и секвенирование кДНК, кодирующей SCCE человека
Рестрикционные энзимы получают от Promega, Madison, MI, и TAQ-полимеразу от Perkui-Elmer-Cefus, Norwalk, CT. Библиотеку кДНК кератиноцитов λ gt II человека
получают из мРНК, полученных из кератиноцитов эпидермального происхождения взрослого человека, полученных от Clontech Laboratories, Polo Alto, CA (N в каталоге HL 1045 b). Вначале библиотеку скринируют анти-SCCE кроличьей поликлональной сывороткой D-5 и Bo-1 (см. пример 5.2.2.). Так как Bo-1 поликлональная анти-SCCE сыворотка дает высокие фоновые сигналы, ее исключают из интересных скринирований на ранней стадии. Используя Д-5 антисыворотку, ряд иммунореактивных бляшек были обогащены как предполагаемые для истинно положительных бляшек. Никакой реакционной способности не наблюдается с моноклональными антителами мoab 4 и мoab 9 ни для одной из бляшек. Интенсивная характеризация с помощью рестрикционных энзимов и PCR характеризация одиннадцати выделенных бляшек показали, что нет сходства между различными бляшками. Наличие такого частичного сходства указывает, что бляшки содержат гомологичные ДНК вставки из одной и той же кДНК последовательности. Так как не удалось определить "отпечаток пальцев" возможной SCCE кДНК последовательности, стратегию пришлось модифицировать.
Бляшки были скринированы в E.coli у 1090 (Clontech) за счет гибридизации бляшек с использованием в качестве зонда дегенеративного синтетического олигонуклеотида. Олигонуклеотидный зонд был сконструирован на основании экспериментально определенной аминотерминальной последовательности нативного SCCE энзима, как указано в примере 4.2. Наиболее надежная часть аминокислотной последовательности, Ile-Ile-Asp-Gly-Ala-Pro (последовательность ID N 3 aa 1 - aa 6) была выбрана для конструирования синтетического 17-мерного олигонуклеотидного зонда 5'-ATHATHGAYGGNGCNCC-3' /H=A или C или T; Y=C или T; N=A или C или G или T/, обозначенного SYM3067, последовательность ID N 4. Олигонуклеотидный зонд синтезируют, используя Весkmаn 200A ДНК синтезатор, используя фосфорамидитную методику в соответствии с инструкциями.
E. coli Y 1090 бактерии выращивают в течение ночи в LB в среде /Sambrook et al. , 1989/, содержащей 0,2% мальтозы и 10 мМ MgSO4, 0,4 мл культуры смешивают затем с разбавленным библиотечным фаговым источником и адсорбируют в течение 20 минут при 37oC. Инфицированную культуру смешивают с 6 мл мягкой агарозы /0,75% агарозы в LB и 10 мМ MgSO4/. Мягкую агарозную смесь выливают на 10 150 мм A пластин. Пластины инкубируют в течение 5 часов и помещают на ночь при 4oC. В целом, все пластины содержат приблизительно 4•105 бляшек.
Для иммобилизации бляшек каждую пластину покрывают NEN Du Pont Colony /Plague Screen мембранами /Du Pont. Wilmington, DE/ на две минуты. Полученные мембраны погружают дважды на 2 минуты в 0,5 M NaOH, дважды по 2 минуты в Tris-HCl pH 7,5 и оставляют сохнуть. Затем эти мембраны используют для гибридизационных экспериментов, как будет указано далее. Эти мембраны предварительно гибридизуют в 10% сульфате декстрана, 1M NaCl, 1% SDS растворе, содержащем 100 мг/мл денатурированной ДНК спермы сельди /Sigma, St. Louis MO/, в течение 5 часов при 65oC. В качестве зонда используют SYM 3067, [λ-32P] dATP, меченый с использованием T4 полинуклеотидной киназы /Promega, Madison, W1/ и его добавляют к прегибридизационной смеси. Гибридизацию ведут в течение 12 - 18 часов при 42oC.
После гибридизации мембраны промывают четыре раза по 5 минут в 2xSSC при комнатной температуре, два раза по 30 минут в 2xSSC, 1% SDS при 42oC, и, наконец, в 0,1 SSC при комнатной температуре в течение 30 минут. Эти мембраны авторадиографируют на рентгеновской пленке /Hyperfilm-MP, Amersham, UK/. В первичном скринировании идентифицируют 14 позитивных бляшек. Эти позитивные бляшки повторно скринируют, используя тот же зонд и методику, что и описанные ранее. После процедуры повторного скринирования идентифицируют две положительные бляшки. Две отобранные бляшки еще раз очищают, и размеры вставок определяют за счет PCR, используя в качестве праймеров SYM 1600 и SYM 1601, и выделенные бляшки в качестве матрицы. Эти два праймера комплементарны левому и правому плечам λ gt 11 фага, соответственно. Амплифицированный ДНК фрагмент примерно 0,9 kb, полученный из фага A 6.2.2, переваривают затем EcoR1 и клонируют в EcoR1 переваренный pUC19 /Pharmacia, Uppsala, Sweden/, pS 496. Этот клонированный фрагмент подвергают затем частичному анализу последовательности, используя праймеры последовательностей, комплементарных pU C19. Нуклеотидную последовательность определяют, используя T7 секвенирующие наборы /Pharmacia, Uppsala, Sweden или USB, Cleveland, Ohio/.
Трансляция полученной ДНК последовательности приводит к аминокислотной последовательности, которая гомологична экспериментально определенной протеиновой последовательности.
Однако, эта последовательность не содержит трансляционного стартового кодона. Для выделения полной длины кДНК, полученный ДНК фрагмент выделяют на агарозном геле и используют в качестве зонда, обеспечивающего гибридизацию в жестких условиях, этот зонд метят 32P за счет мультипраймерной ДНК метящей системы /Amersham, UK/ следующим способом. Воду добавляют в количестве 3 мл на грамм геля и помещают в баню с кипящей водой на 7 минут для того, чтобы расплавить гель и денатурировать ДНК. Затем ампулу переносят в водяную баню при 37oC, по крайней мере на 10 минут. В реакционную смесь для введения метки добавляют объем ДНК/агарозного раствора, содержащий примерно 25 нг ДНК. /Все это в соответствии с инструкциями поставщика/.
Для получения полной длины кДНК, библиотеку кДНК повторно скринируют дважды, используя те же способы, что и описанные ранее, за исключением того, что гибридизацию ведут в жестких условиях при 65oC. В результате этих экспериментов идентифицируют и выделяют 45 индивидуальных позитивных бляшек, которые вначале скринируют с помощью PCR, используя SYM 1600 /5'-GTG GCG ACG ACT CCT GGA-GCC-3'; последовательность ID N 5/, или SYM 1601 /5'-ACA CCA GAC CAA CTG GTA ATG-3' /последовательность ID N 6/ в сочетании с SYM 3208 в качестве PCR праймеров для идентификации бляшек, содержащей полную 5'- открытую считывающую рамку. SYM 3208, 5'-TGGGTGGGAGCCTCTTGCACA-3', последовательность ID N 7, по крайней мере, частично комплементарна 5' части SCCE кДНК, была сконструирована на основании Информации о ДНК последовательности, полученной из pS 496. После этого скринирования, были отобраны 4 фага для дальнейшего анализа. Для анализа последовательности, полученные PCR амплифицированные фрагменты, полученные из этих фагов, были клонированы в pUC19, как было указано ранее. Полученные результаты показывают, что один из этих фагов, 205.2.1 содержит полной длины вставку.
ДНК из фагового изолята 205.2.1 получают по способу Sambrook et al., 1989, и ДНК препарат переваривают EcoR1. Переваренную ДНК выделяют электрофорезом на агарозном геле, и фрагмент около 1 kb выделяют и клонируют в EcoR1 переваренную pUC19. Полученную плазмиду обозначают pS 500 /фиг. 15/. Полную нуклеотидную последовательность кДНК фрагмента определяют как указано ранее. В качестве праймера для реакций секвенирования используют специфические олигонуклеотиды, комплементарные pUC19 или SCCE последовательностям. Нуклеотидная последовательность /последовательность ID N 1/ содержит открытую считывающую рамку, достаточную для кодирования полной аминокислотной последовательности SCCE протеина-предшественника, состоящего из 253 аминокислот, включая сигнальный пептид и преполипептид /последовательность ID N 2/.
Как было обнаружено, другой фаг, названный 106.1.2, содержит SCCE кДНК последовательность, в которой отсутствует 5'-нетранслируемая последовательность и первые три кодона. Эту вставку выделяют как 954 bp EcoR1 фрагмент и клонируют в EcoR1 линеаризованную pUC19, что приводит к получению плазмиды pS 498. Эту плазмиду частично секвенируют.
Третий фаг, обозначенный 108.1.2, как было обнаружено, содержит SCCE кДНК последовательность, в которой также отсутствует 5'-нетранслируемая последовательность и семь нуклеотидов транслируемого участка. Эта кДНК вставка имеет более длинный вариант 3'-нетранслируемого участка, простирающийся от 1057 bp в обратном направлении от стоп кодона. Этот 1884 bp EcoR1 фрагмент выделяют и клонируют в EcoR1 линеаризованную pUC19. Полученную плазмиду полностью секвенируют и обозначают pS 501.
Пример 7
Определение SCCE мРНК в эпидермисе человека
Получение полной РНК из эпидермиса человека
Это осуществляют в соответствии с методикой Chomcgynski and Sacchi, 1987. Абдоминальную кожу здорового человека получают при пластической хирургии. Немедленно после удаления ее охлаждают на льду. В течение менее 15 минут эпидермис выделяют, соскребая скальпелем, погружают в раствор D /Chomcgynski and Sacchi, 1987/ и гомогенизируют в стеклянном гомогенизаторе. Затем все делают по схеме Chomcgynski. Осажденную полную РНК хранят при -20oC в 70% этаноле до дальнейшего анализа.
Получение информационной РНК
500 мкг полной РНК эпидермиса обрабатывают с помощью набора Poly A Tract /Promega/ в соответствии с инструкциями поставщиков.
РНК-электрофорез и блоттинг
Агарозные гели /1,4%/ приготавливают с 0,66М формальдегидом в 1хMOPS буфере и 0,6 г/мл этидиумбромида /Sigma, St. Loo MO/. мРНК, соответствующую 100 мкг полной РНК, растворяют в РНК-образцовом буфере /50% формамида, 2,2М формальдегида, 3% Ficoll 1•MOPS/ и нагревают при 60oC в течение 5 минут перед нанесением. РНК-маркеры /BRL, Geithersburg, MD/ обрабатывают аналогичным образом. После электрофореза гели погружают в дистиллированную воду на 5 минут, а затем в 50 мМ NaOH на 30 минут, и в 0,1M Tris-HCl pH 7,5 на 30 минут. Осуществляют блоттинг на мембраны Gene - Screen Plus /NEN Du Pont, Wilmington, DE/ с Vacu-Gene оборудованием /Pharmacia, Uppsala, Sweden/ в течение 1 часа в 10•SSC. Затем мембраны промывают 3•SCC, сушат в течение ночи и прогревают в течение 2 часов при 80oC. РНК визуализируют на мембранах в УФ-свете.
кДНК зонды
Плазмиду pS501, полученную как указано в примере 6, переваривают Hinc II и Bgl II. Эта кДНК содержит один Hincll-сайт у bp N 1060 и один Bgl 11 сайт у bp N 1715. 1070 bp фрагмент /Hinc II-сайт в pUC19 множественного клонирования сайт-эндогенный Hinc II-сайт/ содержит SCCE кодирующий участок кроме 7 bp с 5' конца и нетранслируемый участок, включая сайт полиаденилирования у bp 944-951, который является общим для всех SCCE-кДНК, которые были выделены. Фрагмент 655 bp Hinc II-Bgl II, который не содержит поли-A-хвоста, является уникальным для SCCE кДНК 108-1-2. Эти фрагменты были выделены с помощью электрофореза на агарозе и были использованы для получения 32PdCТP-меченых зондов с помощью набора Multiprime для введения меток в ДНК /Amersham, Buckinghamshire, UK/.
Гибридизация
Мембраны кипятят в течение 30 минут в 1% SDS в 1•TE и предварительно гибридизуют при 60oC в 1% SDS, 1 M NaCl, 10% декстрансульфате, ДНК спермы сельди 0,1 мг/мл в течение 3 часов. Гибридизацию ведут в том же растворе при 60oC в течение ночи. Промывки осуществляют 2х30 минут при 60oC в 1% SDS в 2•SCC и в течение 3 часов в 0,1•SCC при комнатной температуре. Затем эти мембраны подвергают авторадиографии.
Результаты:
Можно продемонстрировать наличие в эпидермисе человека двух видов мРНК с размерами около 1,2 kb и 2,0 kb, соответственно /фиг. 16/. Это находится в хорошем соответствии с доказательствами, полученными в экспериментах по клонированию, где были обнаружены два типа кДНК.
Пример 8
Экспрессия рекомбинантного SCCE в E.coli
Конструирование плазмид pGEX-2T/SCCE
1. Смысловые PCR праймеры
1.a.
CGTGGATCCATCGAAGGTCGTATTATTGATGGCGCCCCATGT /SYM 3367, послед. ID N 8, подчеркнута 3'-часть, кодирующая II-терминальные аминокислоты IIDGAPC активного нативного SCCE, 5'-часть с Bam H1-сайтом, и дополнительная последовательность, кодирующая сайт фактора Xa, IEGR.
1. b. CGTGGATCCATCGAAGGTCGTTTGGAAACTGCAGGAGAAGAA /SYM 3368, последовательность ID N 9, подчеркнуты 3'-часть, соответствующая парам оснований 76-96 в полной SCCE-кДНК последовательности, кодирующей аминокислотную последовательность LETAGEE, 5'-часть как и в 1a.
2. Антисмысловые PCR праймеры TGATCCTCTGAGCTCTCCTG /SYM 3371; комплементарен парам оснований 285 - 304 в полной SCCE-кДНК последовательности, последовательность ID N 1, с Sac1-сайтом у bp 294/.
Последовательность pS 498 /пример 6/ PCR-амплифицируют праймерами 1a/2 и 1b/2. Полученные продукты очищают экстракцией фенолом и осаждают этанолом переваривают Bam H1/ Sac1 и очищают с помощью электрофореза в агарозе. Затем их клонируют в TG2-клетки в pGEX-2T /Pharmacia/, переваривают Bam H1/EcoR1 вместе с 3' 673 парами оснований SCCE 106-1-2, полученными при переваривании pS 498 Sac1 и EcoR1. Из бактериальных клонов, которые были использованы для исследования экспрессии плазмид /pS 510, кодирующая нативный II-терминал, следующий за сайтом фактора Xa, и pS 511, кодирующая пропептид, следующий за сайтом фактора Xa/, были выделены, и нуклеотидные последовательности, соответствующие вставкам, полученным из PCR-продуктов, были проверены способом дидеокси-цепной терминации с использованием набора T7 для секвенирования /Pharmacia, Uppasala, Sweden/.
Исследования экспрессии
Культуры (ведущиеся в течение ночи)
TG клеток с pS 510 и pS 511 в LB среде, содержащей 50 мкг/мл Carbenicillin /Sigma St. Louis MO/, разбавляют в 10 раз свежей средой и выращивают в течение 3 часов при 37oC. IPTG /Sigma, St. Lоuis, MO/ добавляют до конечной концентрации 0,1 М, и культуры выращивают при 37oC еще в течение 3 часов. Бактериальные осадки обрабатывают ультразвуком в PBS 1% Triton X-100 /Sigma, St. Louis, MO/. После центрифугирования при 10000 х g в течение 15 минут, надосадочную жидкость и осадки анализируют с помощью SDS-PAGE и иммуноблоттинга с поликлональной SCCE-специфической цыплячьей анти-сывороткой.
Большие количества IPTG-индуцируемых протеинов с Мг примерно 50 кД /pS 510/ и 52 кД /pS 511/ с SCCE-подобной иммунорективностью были обнаружены в осадках, не растворимых в PBS-Triton X-100 /см. фиг. 17/.
Надосадочные жидкости после обработки ультразвуком в PBS-Triton X-100 содержат GST/SCCE протеины слияния того же размера, что и в нерастворимых осадках, но их количество мало по сравнению с количеством в нерастворимых осадках.
Эти результаты показывают, что можно экспрессировать SCCE как протеин слияния с GST, а последовательности, соответствующие специфическим протеазным сайтам расщепления в пре-про- SCCE аминокислотной последовательности, сделают возможным повторить этот эксперимент с целью получить рекомбинантный SCCE в бактериях. Полученный протеин можно солюбилизировать в мочевине или гуанидинийгидрохлориде, а затем очистить с помощью ионообменной хроматографии благодаря высокой изоэлектрической точке SCCE. Очищенный протеин можно ренатурировать, диализуя против буферов с низкой концентрацией денатурирующего агента, а затем расщепить фактором Xa для высвобождения GST полипептида из SCCE или про-SCCE. Протеины слияния GST/SCCE можно также использовать в качестве иммуногенов для получения SCCE-специфических антител и в качестве иммуносорбентов для очистки антител.
Пример 9
Экспрессия рекомбинантного SCCE человека в клетки млекопитающих
Для создания вектора экспрессия для получения рекомбинантного SCCE человека, кДНК последовательно выделяют из плазмиды pS500 в виде 897 bp EcoR1/Dral фрагмента. Этот фрагмент субклонируют в EcoR1 и Sma1 переваренную pUC19, получая pS502. Затем плазмиду pS502 переваривают EcoR1 и Sal 1 для выделения кДНК последовательностей SCCE в виде 0,9 kb фрагмента, который снова субклонируют в pUC19 вариант, в котором отсутствует Hind111 сайт, получая плазмиду pS503. Этот pUC19 вариант получают при переваривании pUC19 с помощью Hind111, заполняя, используя энзим Кленова и снова лигируя. Для облегчения клонирования в вектор экспрессии, Hind111 сайт вводят в 5'-конец кДНК SCCE. Этого добиваются, переваривая pS503 EciR1 и встраивая линкер, который превращает сайт в Hind111, SYM3603
5'-AATTGTGGAAGCTTCCAC-3', последовательность ID N 10. Полученную плазмиду, которая несет кодирующую протеин часть кДНК SCCE с Hind111 сайтом у 5' конца и Sal 1 сайтом у 3' конца, соответственно, обозначают pS505.
Окончательный вектор экспрессии получают, лигируя три различных ДНК фрагмента. Вначале, pS505 переваривают Hind 111 и Sal 1, и выделяют фрагмент 0,9 kb.
Затем, для получения дистальной части регуляторного элемента, расположенного в обратном направлении, мышиного металлотионеина, последовательности вируса бычьей папилломы, кроличий бета-глобулиновый геномный фрагмент, обеспечивающий сигналы процессинга мДНК, и плазмидные последовательности, pML2d, для обеспечения селекции и репликации в E.coli /Waldenstrom et al., 1992/, вектор pS147 переваривают SaC1 и Sal 1, и выделяют фрагмент около 12 kb.
В-третьих, для выделения проксимальной части промотора мышиного металлотионеина, плазмиду pS42, в которой нативный Bgl 11 расположен в лидерной последовательности, был превращен в Hind 111 сайт, переваривают Sac 1 и Hind 111, и выделяют фрагмент приблизительно 220 bp.
Лигирование этих трех фрагментов приводит к получению вектора экспрессии SCCE pS507 /см. фиг. 18/.
Вектор экспрессии pS507 совместно трансфецируют с вектором, содержащим ген устойчивости к неомицину, управляемый 5' длинным концевым повтором вируса саркомы Harley, и с сигналами полиаденилирования SV40 /Lusky and Botchan, 1984/, в мышиные C127 клетки /ATCC CRL 1616/. Эксперименты по трансфекции осуществляют в соответствии со способом кальций-фосфатного осаждения /Graham and Van der bb, 1973/.
Клетки культивируют в Ham's F12 /Dulbecco's модифицированной среде Eagle /DMEM; Gibco, BRL, Gаitersburg, MD/ /1:1/, дополненной 10% сывороткой плода теленка /HyClone, Logan, UT/. Клоны клеток, устойчивых к неомицину, отбирают с помощью 1,5 мг/мл G418 /Gibco-BRL/, и через 10 - 15 дней после селекции устойчивые клоны клеток идентифицируют и выделяют из основных пластин, и отправляют для дальнейшего анализа.
Для анализа экспрессии рекомбинантных SCCE генов, приготавливают полную РНК из выделенных клеточных линий. Полную РНК получают из C127 клеток и выделяют на 1% формальдегид-агарозном геле, переносят на нитроцеллюлозную мембрану и гибридизуют с SCCE зондом, меченым 32P. Этот зонд представляет собой 1070 bp Hinc 11 фрагмент SCCE кДНК, выделенной Hinc 11 перевариванием pS500, и агарозным электрофорезом. Экспериментальная процедура соответствует указаниям Ausubel et al., 1992. Эксперименты по Норзернблоттингу и гибридизации с 32P-меченой SCCE кДНК показывают, что рекомбинантную SCCE мРНК можно детектировать в нескольких клеточных линиях, содержащих SCCE вектор, pS507. Гибридизация не обнаружена в контрольных образцах, полученных из C127 клеточной линии, содержащей идентичный вектор, за исключением SCCE кДНК /фиг. 19/. Размер 1,4 kb соответствует ожидаемому размеру.
Образцы кондиционной клеточной культуральной среды собирают и анализируют в иммуноблоттинге. SDS-PAGE осуществляют по способу Лаэмли /1970/ и для иммуноблоттинга в качестве детектирующего антитела используют цыплячий анти-нативный SCCE. Меченый щелочной фосфатазой анти-цыплячий IgG /Sigma, St. Louis, MO/ используют для энзимной метки. Полученные результаты представлены на фиг. 20.
Для анализа экспрессии рекомбинантного SCCE, получают полную РНК из C127 клеток, трансфектированных вектором экспресии pS507. В качестве контрольных образцов получают полную РНК как из нетрансфектированных C127 клеток, так и из C127 клеток, трансфектированных вектором экспрессии pS147. Вектор pS147 аналогичен вектору pS507 за исключением того, что он содержит кДНК для желчными солями стимулированной липазы человека /Nilsson et al., 1990/ вместо SCCE кДНК человека. РНК получают по способу Ausubel et al., /1992/. Эксперименты Норзернблоттинга и гибридизации 32P-меченой кДНК SCCE показывают, что рекомбинантная мРНК SCCE примерно 1,4 kb детектируется в C127 клетках, содержащих SCCE вектор, pS507 /фиг. 19/. Гибридизация не обнаружена в контрольных образцах, полученных из C127 клеточных линий, содержащих pS147 или нетрансфектированные C127 клетки. Длина рекомбинантной мРНК SCCE соответствует ожидаемой величине.
Образцы кондиционной клеточной культурной среды собирают и анализируют за счет SDS-PAGE и иммуноблоттинга. Блоты проявляют по способу Blake et al., 1984. Полученные результаты /см. фиг. 20/ показывают, что C127 клетки, содержащие pS507, продуцируют три протеина, которые демонстрируют реакцию со всеми доступными поликлональными кроличьими и цыплячьими SCCE антителами, а также с анти- SCCE моноклональными, полученными по способу примера 5. Рекомбинантные SCCE реактивные протеины демонстрируют кажущийся молекулярный вес, который примерно на 1 кДа больше, чем у очищенного нативного SCCE человека. Рекомбинантный протеин не демонстрирует какой-либо протеолитической активности. При сравнении предполагаемой SCCE аминокислотной последовательности с экспериментально полученным NH-2 концом нативного SCCE человека и с последовательностями других химотрипсино-подобных протеаз, можно сделать вывод, что рекомбинантный SCCE, полученный в C127 клетках, является его про-энзимной формой. Данные последовательности показывают, что этот про-энзим можно активировать протеолитическим расщеплением со стороны C-конца лизина в последовательности . . .AQGDKIIDGAP..., где подчеркнутая последовательность является NH-2 последовательностью активного нативного SCCE человека /последовательность ID N2, aa 5 - aa 6/.
Пример 10
Выделение и характеризация рекомбинантного SCCE
Выделение
1,5 мг моноклонального антитела TE4b, направленного против нативного SCCE, соединяют с 1,5 мл СПВг-активированной сефарозы /Pharmacia-LKB Biotech., Uppsala, Sweden/, используя способ, рекомендованный изготовителем. 40 мл среды, содержащей SCCE, фильтруют через 0,45 мкм фильтр, а затем вводят в колонку. Колонку промывают несколько раз 10 мМ фосфата натрия, 150 мМ NaCl, pH 7,2, а затем элюируют 0,1 М глицин-HCl, pH 2,5. Элюированный протеин немедленно нейтрализуют, добавляя 0,1 объем 1 M Tris-HCl, pH 8,0.
Активация
Рекомбинантный SCCE /4,6 мкг в 200 мл/, очищенный на геле, соединенном с моноклональным антителом /см. ранее/, переваривают 4,6 мкл 0,1 мг/мл трипсина /массовое отношение 10:1/ при 37oC. Образцы /50 мкл отбирают через 20 минут, 1 час, 3 часа и 20 часов, и 5 мкл 10 мкМ /4-амидинофенил/метансульфонил /APMSF; Boehringer Mannheim, Germany/ добавляют для окончания реакции.
Активность расщепленного SCCE анализируют с помощью невосстанавливающего SDS-PAGE на казеиновых гелях по способу примера 2.2 Идентичность полученных отщепленных форм SCCE была проанализирована с помощью восстанавливающего SDS-PAGE с последующим иммуноблоттингом, с использованием цыплячьего анти-нативного SCCE с последующим введением метки щелочной фосфатазы в анти-цыплячий IgG и нитроблютетразолия и 5-бром-4-хлор-3-индолилфосфатазы в качестве субстрата для щелочной фосфатазы.
N-терминальное секвенирование
Афинно очищенный /как указано ранее/ SCCE, примерно 35 мкг, был подвергнут слот-блоттингу с использованием установки Bio-Dot SF. /Bio Rad, Richmond, CA/ на мембране Immobilon /Millipore, Bedford, MA/. Затем мембрану промывают несколько раз дистиллированной водой для удаления всего Tris и глицина. Часть мембраны, на которой был связан протеин, вырезают и секвенируют, используя Applied Biosystems /Foster City, CA/, 477A импульсный жидкофазный секвенатор с подключенным PTH 120A анализатором. Секвенирование осуществляют, используя установленные циклические программы и химикалии поставщиков. Последовательность, полученная в первых шести положениях, представляет собой glu-glu-ala-gln-gly-asp, что соответствует аминокислотам -7 -2 последовательности ID N 2. Как можно сделать заключение из этого результата, сигнальный пептид состоит из 22 аминокислот и основан на N-терминальной аминокислотной последовательности нативного активного SCCE, а пропептид состоит из семи аминокислот.
Дегликозилирование
Очищенный рекомбинантный SCCE /5 мкг/ и нативный SCCE /20 мкг/ кипятят в течение 3 минут в 20 мкл 0,5% SDS и 0,1 M β -меркаптоэтанола. Образцы разбавляют натрийфосфатным буфером, pH 8,6 и Nonidet P-40 до конечных концентраций 0,2 M и 1,25%, соответственно. Добавляют N-гликозидазу F® /Boehringer Mannheim/ для рекомбинантного протеина 0,6 единиц, а для нативного протеина 1,2 единиц энзима, и реакционную смесь инкубируют в течение ночи при 37oC. Конечная концентрация SDS в анализируемом образце составила 0,17%. Обработанные N-гликозидазой F® SCCE анализируют на 8 - 18% SDS-PAGE с последующим иммуноблоттингом, как указано ранее. Полученные результаты демонстрируют восстановление кажущегося молекулярного веса двух верхних полос, тогда как кажущийся молекулярный вес самой нижней полосы не изменяется /фиг. 23/. Это показывает, что рекомбинантный SCCE, продуцированный в C127 клетках, существует в двух N-гликозилированных формах и одной не-гликозилированной форме. Этот результат аналогичен с тем, который наблюдается для активного нативного SCCE /фиг. 23/.
Пример 11
Композиции, содержащие SCCE
Такие композиции можно получить в соответствие с обычными фармацевтическими методиками, включая тщательное смешивание активных соединений с другими ингредиентами. Все проценты даны по весу.
Композиции, которые содержат более одного активного соединения, также входят в объем изобретения. Нижеследующие примеры следует рассматривать как включающее также более одного активного соединения. Аналогично, термин "SCCE" можно заменить на "про-SCCE". Таким образом, в объем настоящего изобретения входят композиции, которые содержат как SCCE, так и про- SCCE.
SCCE = нативный или рекомбинантный химотрипсиновый энзим роговичного слоя, необязательно в сочетании с другими активными соединениями
q.S. = guantum satis (сколько нужно)
Крем масло/вода, %:
SCCE - 0,01 - 20
Полисорбат 80 - 0,5
Эмульгируемый воск - 5
Минеральное масло - 4
Диметикон - 1
Глицерилстеарат - 6
Антиоксидант - q.S.
ЕДТА - 0,1
Консервант - q.S.
Глицерин 85% - 4
Пропиленгликоль - 7
Агент, регулирующий pH - 0,01 - 10
Вода - 65 - 76
Примеры изменяемых факторов: Антиоксиданты, хелатирующие агенты, консерванты, эмульгирующие системы /соотношение между масляной фазой и водной фазой и содержание эмульгирующего агента и других относящихся к этому эксципиентов/.
Крем вода/масло, %:
SCCE - 0,01 - 20
Цетиловый спирт - 0,5
Ланолин - 5
Белый петролатум - 10
Минеральное масло - 45
Антиоксидант - q.S.
EДТА - 1
Агент, регулирующий pH - 0,01 - 10
Консервант - q.S.
Вода - 15 - 25
Примеры изменяемых факторов: антиоксиданты, хелатирующие агенты, консерванты, эмульгирующие системы /соотношение между масляной фазой и водной фазой и содержание эмульгирующего агента и других относящихся к этому эксципиентов/
Мазь, %:
SCCE - 0,01 - 20
Ланолин - 15
Петролатум - 58 - 68
Минеральное масло - 15
Диметикон - 2
Антиоксидант - q.S.
Примеры изменяемых факторов: антиоксиданты.
Линимент, %:
SCCE - 0,01 - 20
Эмульгирующий воск - 4
Глицерилстеарат - 3
Минеральное масло - 15
Полисорбат 80 - 0,6
Глицерин 85% - 3
Пропиленгликоль - 5
Антиоксидант - q.S.
ЕДТА - 0,1
Консервант - q.S.
Вода - 59 - 69
Примеры изменяемых факторов: антиоксиданты, хелатирующие агенты, консерванты, эмульгирующие системы /соотношение между масляной фазой и водной фазой и содержание эмульгирующих агентов и других относящихся к этому эксципиентов/
Гель, %:
SCCE - 0,01 - 20
Триэтаноламин - 1 - 5
Этиловый спирт - 10
Цетиловый спирт - 10
Целлюлозная смола - 5
ЕДТА - 0,1
Консервант - q.S.
Вода - 59 - 74
Примеры изменяемых факторов: гелеобразующие агенты, антиоксиданты, хелатирующие агенты, консерванты.
Водный раствор, %:
SCCE - 0,01 - 20
Агент, регулирующий pH - 0,01 - 10
Цетиловый спирт - 4
Пропиленгликоль - 5
Консервант - q.S.
Антиоксидант - q.S.
ЕДТА - 0,1
Вода - 37 - 89
Примеры изменяемых факторов: антиоксиданты, хелатирующие агенты, консерванты, обезжиривающие агенты, увлажнители.
Раствор в этиловом спирте, %:
SCCE - 0,01 - 20
Агент, регулирующий pH - 0,01 - 10
Пропиленгликоль - 5
Цетиловый спирт - 3
Антиоксидант - q.S.
ЕДТА - 0,1
Этиловый спирт - 50 - 95
Примеры изменяемых факторов: антиоксиданты, хелатирующие агенты, увлажнители.
Суспензия, %:
SCCE - 0,01 - 20
Карбомер - 0,5
Целлюлозная смола - 0,5
Полисорбат 80 - 0,1
Пропиленгликоль - 5
Аскорбиновая кислота - 0,05
Цетиловый спирт - 4
Полисорбат - q.S.
ЕДТА - 0,1
Агент, регулирующий pH - 0,01 - 10
Вода - 72 - 80
Примеры изменяемых факторов: антиоксиданты, хелатирующие агенты, консерванты, суспендирующие агенты.
Паста, %:
SCCE - 0,01 - 20
Петролатум - 45 - 55
Оксид цинка - 40
Минеральное масло - 5
Антиоксидант - q.S.
Примеры изменяемых факторов: антиоксиданты, основания паст.
Пластырь, %:
SCCE - 0,01 - 20
Cutina zm - 70 - 80
Миристиловый спирт - 5
Касторовое масло - 2
Белый пчелиный воск - 10
Белый петролатум - 3
Антиоксидант - q.S.
Примеры изменяемых факторов: антиоксиданты, основа пластыря.
Спрей /ручной/, %:
SCCE - 0,01 - 20
Агент, регулирующий pH - 0,01 - 10
Этиловый спирт - 30
Глицерин 85% - 5
Пропиленгликоль - 5
Цетиловый спирт - 3
Антиоксидант - q.S.
ЕДТА - 0,1
Консервант - q.S.
Вода - 22 - 57
Примеры изменяемых факторов: антиоксиданты, хелатирующие агенты, консерванты, обезжиривающие агенты, увлажнители.
Раствор для аэрозольного спрея, %:
SCCE - 0,01 - 20
Агент, регулирующий pH - 0,01 - 10
Изопропилмиристат - 3
Пропиленгликоль - 5
Этиловый спирт - 48 - 92
Пропеллант - q.S.
Примеры изменяемых факторов: обезжиривающие агенты, увлажнители.
Спрей - пенный аэрозоль, %:
SCCE - 0,1 - 20
Воск - 3
Этиловый спирт - 50 - 55
Антиоксидант - q.S.
ЕДТА - 0,1
Вода - 20 - 35
Пропеллант - q.S.
Примеры изменяемых факторов: пропелланты, антиоксиданты, хелатирующие агенты.
Спрей-аэрозольная эмульсия, %:
SCCE - 0,01 - 20
Производные целлюлозы - 1 - 3
Твин® 60 - 1,0
Глицерилстеарат - 2,5
Сорбат калия - 0,2
Антиоксидант - q.S.
ЕДТА - 0,1
Консервант - q.S.
Агент, регулирующий pH - 0,01 - 10
Вода - 50 - 95
Пропеллант - q.S.
Примеры изменяемых факторов: антиоксиданты, хелатирующие агенты, консерванты, эмульгирующие системы /соотношение между масляной фазой и водной фазой, и содержание эмульгирующих агентов и других, относящихся к этому эксципиентов/.
Шампунь, %:
SCCE - 0,01 - 20
Натрийлаурилсульфат - 40
Цетиловый спирт - 3
Вспенивающий агент или кондиционер - 3
Хлорид натрия - 2
Антиоксидант - q.S.
ЕДТА - 0,1
Консервант - q.S.
Вода - 43 - 53
Примеры изменяемых факторов: основания шампуней, антиоксиданты, хелатирующие агенты, консерванты, увлажнители, кондиционеры.
Шампунь для тела, %:
SCCE - 0,01 - 20
Натрийлаурилсульфат - 40
Цетиловый спирт - 4
Вспенивающий агент или кондиционер - 3
Перламутровый агент - 10
Консервант - q.S.
Антиоксидант - q.S.
ЕДТА - 0,1
Вода - 40 - 50
Примеры изменяемых факторов: основания шампуней, антиоксиданты, хелатирующие агенты, консерванты, добавки, кондиционеры.
Медицинское мыло, %:
SCCE - 0,01 - 20
1-гидроксиэтан-1,1-дифосфорная кислота - 0,2
Глицерин - 0,8
Натриевое мыло кокосового масла и таллового масла - 88 - 98
Добавки - 0,7
Примеры изменяемых факторов: увлажнители, основания для мыла.
Порошок, %:
SCCE - 0,01 - 20
Тальк - 65 - 70
Каолин - 6
Диоксид титана - 2
Карбонат кальция - 8
Стеарат магния - 3
Кукурузный или пшеничный крахмал - 5 - 10
Пример изменяемых факторов: основания порошков, консерванты, массовые соотношения.
Кондиционер для волос, %:
SCCE - 0,01 - 20
Цетиловый спирт - 2,2
Алкилтриметиламмонийхлорид - 1,25
Октилдодеканол - 1
Лимонная кислота - 1
ЕДТА - 0,1
Консервант - q.S.
Антиоксидант - q.S.
Вода - До 100
Примеры изменяемых факторов: кондиционеры, консерванты, хелатирующие агенты, антиоксиданты.
Аналогичным способом можно приготовить композиции для поверхностного нанесения, содержащие соединение, которое способно ингибировать или повышать активность SCCE.
Пример 12
Активность по перевариванию десмосом рекомбинантным SCCE
Корнеоциты, содержащие интактные десмосомы, удаляют с кожи, срезая полоски до более глубоких слоев роговичного слоя, и чешуйки отделяют гексаном и сушат в аликвотах. Аликвоты корнеоцитов /3 мг/ экстрагируют 1 M хлоридом натрия при 4oC для солюбилизации характеристических протеаз, а затем интенсивно промывают с инкубационным буфером /0,1 M Tris-HCl pH 8,0/ для удаления энгенной протеолитической активности из препаратов. Инкубирование ведут в 0,1 M Tr pH 8,0 с 10 мкг рекомбинантного SCCE /или без него/ в течение 24 часов при 37oC. Доказательством десмосомального переваривания за счет энзима является определение уровней десмосомного маркерного протеина десмоглеина 1 /DG1/. Его выделяют из чешуек, экстрагируя 8 M мочевины /2% SDS/ β-меркаптоэтанольным буфером с последующей очисткой DG-1 гликопротеина с использованием конканвалин A-афинной хроматографии. Элюат конканвалина A фракционируют за счет SDS-PAGE и электрофоретически переносят на PDVF мембрану для иммуноблоттинга, DG1 идентифицируют, используя специфическую антисыворотку, и определяют, используя возрастание хемилюминесценции. Полученные результаты приведены в таблице 4.
Результаты, представленные в таблице 4, показывают, что рекомбинантный SCCE способен разлагать межклеточные когезивные структуры в роговичном слое в анализе ин витро.
Пример 13
Влияние ингибиторов на активность рекомбинантного SCCE
Исследуют влияние ингибиторов апротинина, химостатина и сульфата цинка на S-2586 гидролизующую активность г SCCE. Схема эксперимента представлена в примере 3.2. Результаты представлены в таблице 5.
Как представлено ранее в таблице 5, апротинин, химостатин и ионы цинка ингибируют S-2586 гидролизующую активность рекомбинантного SCCE таким же образом, как и нативного энзима.









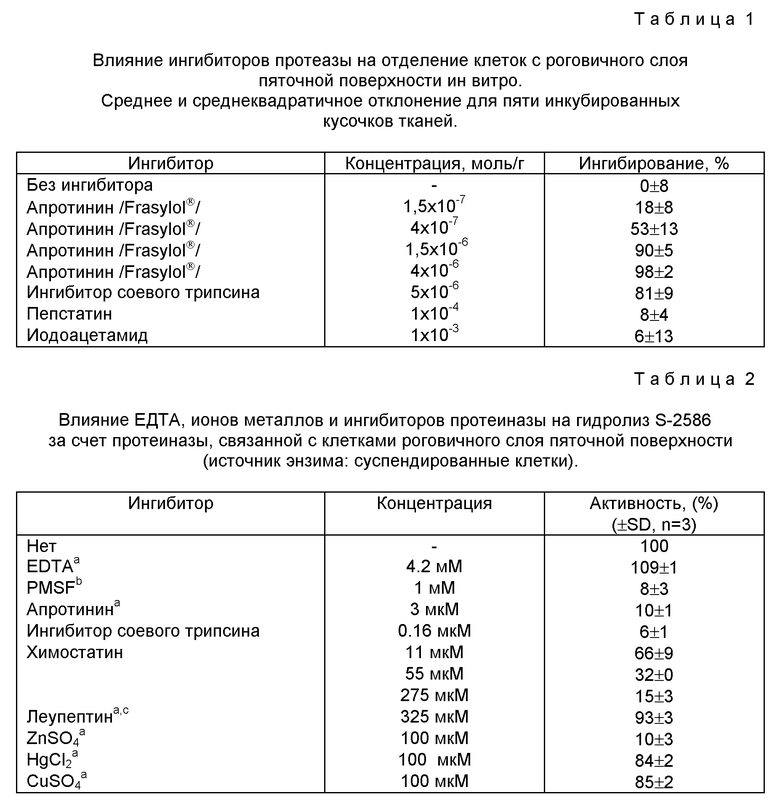
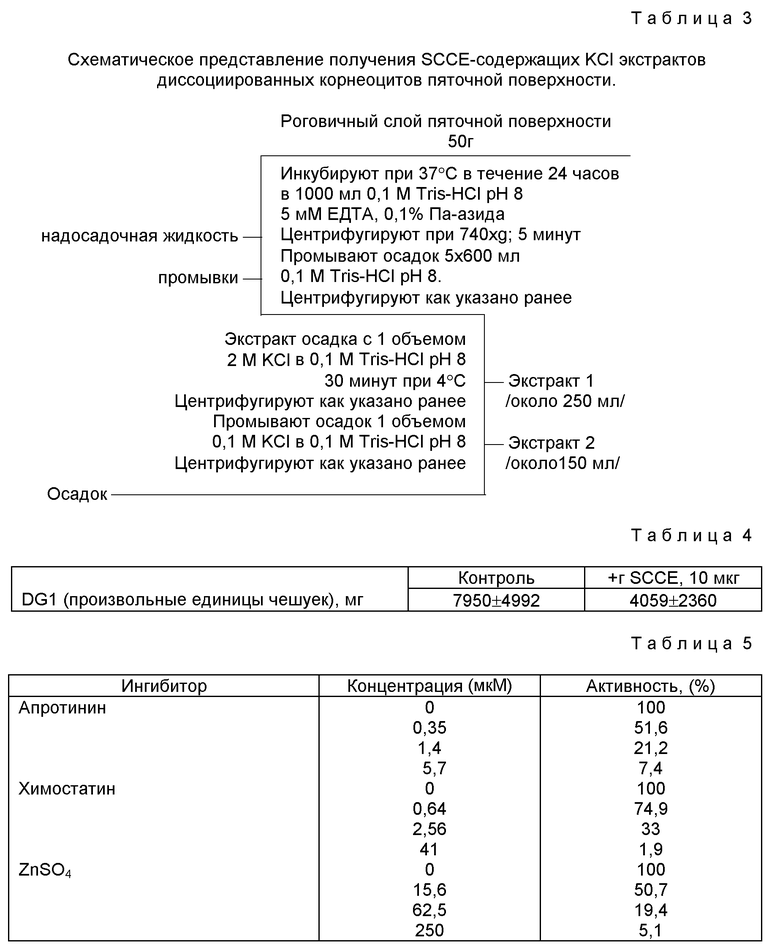

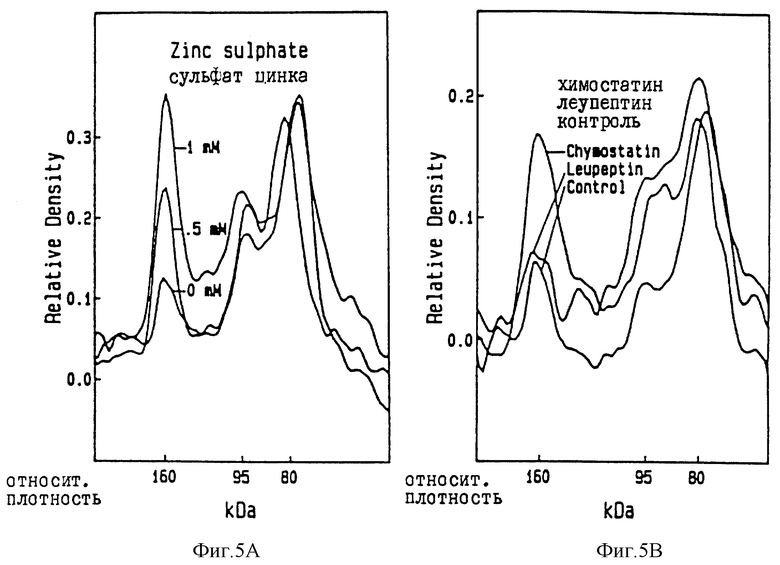
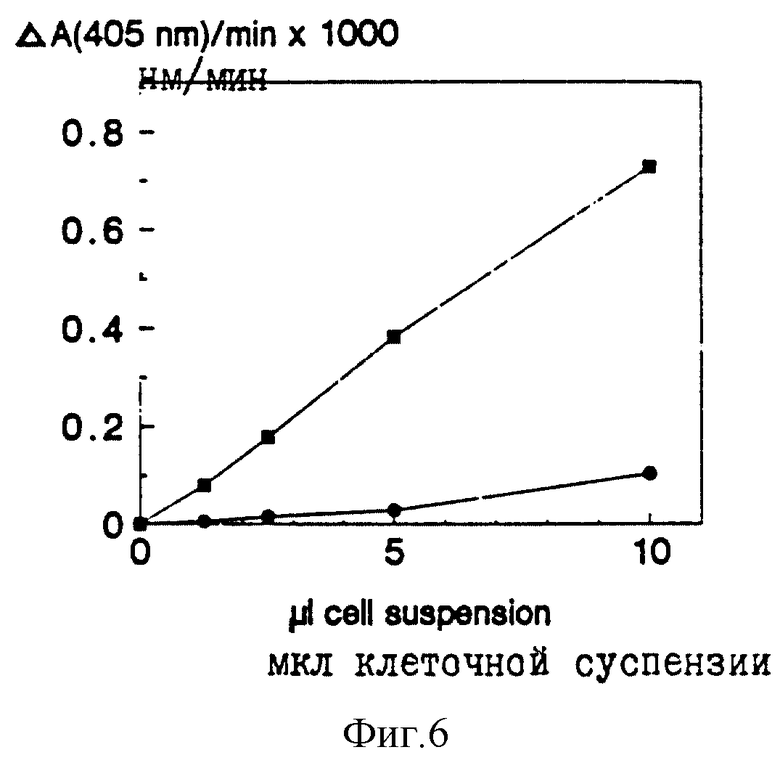
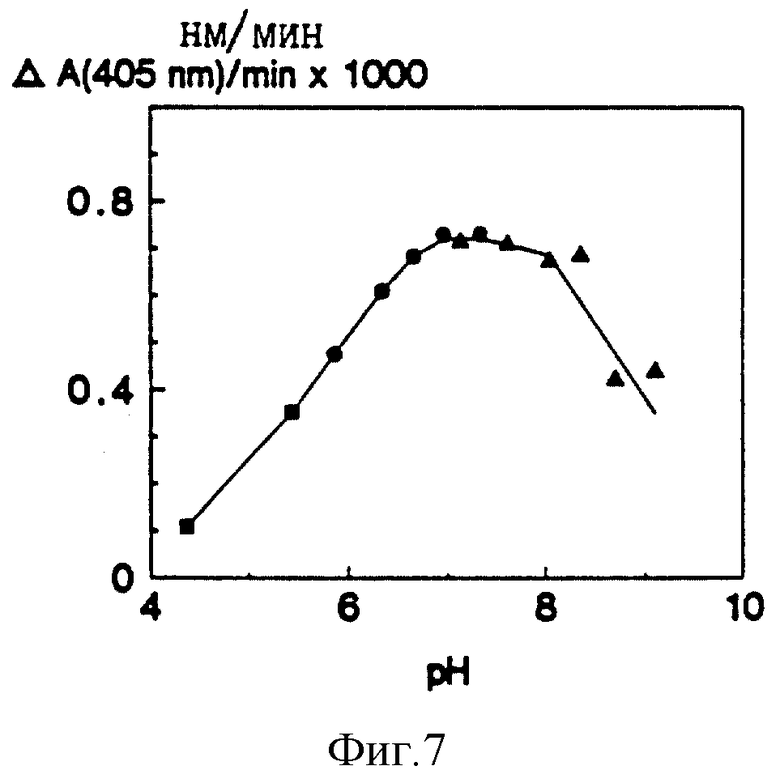
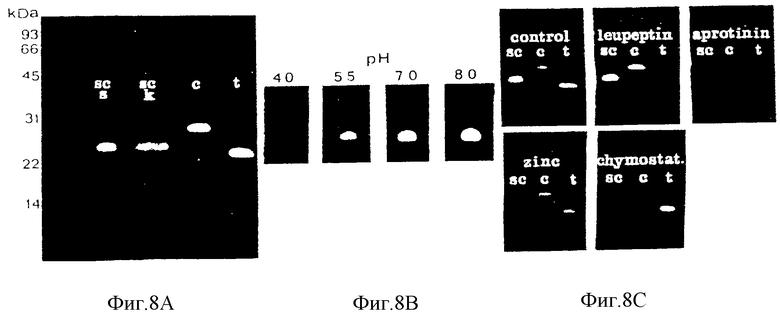
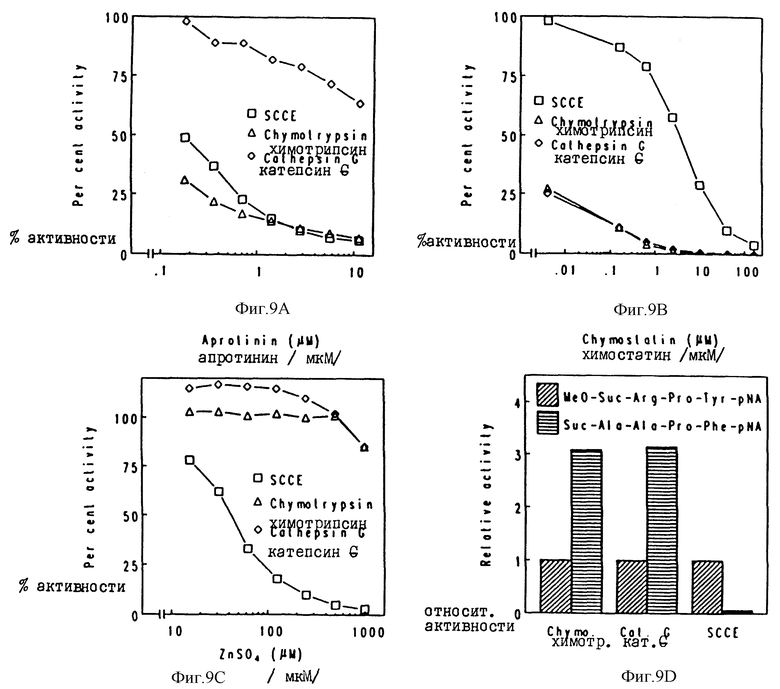
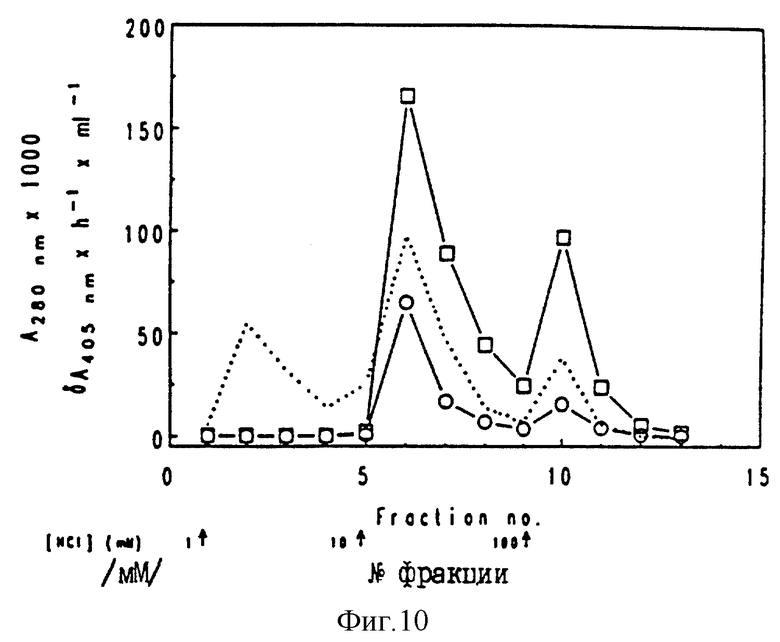

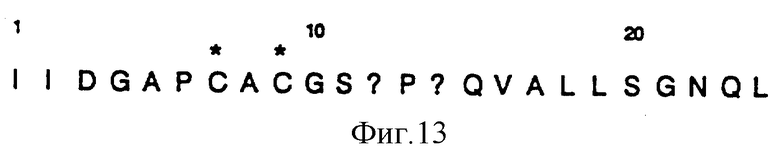

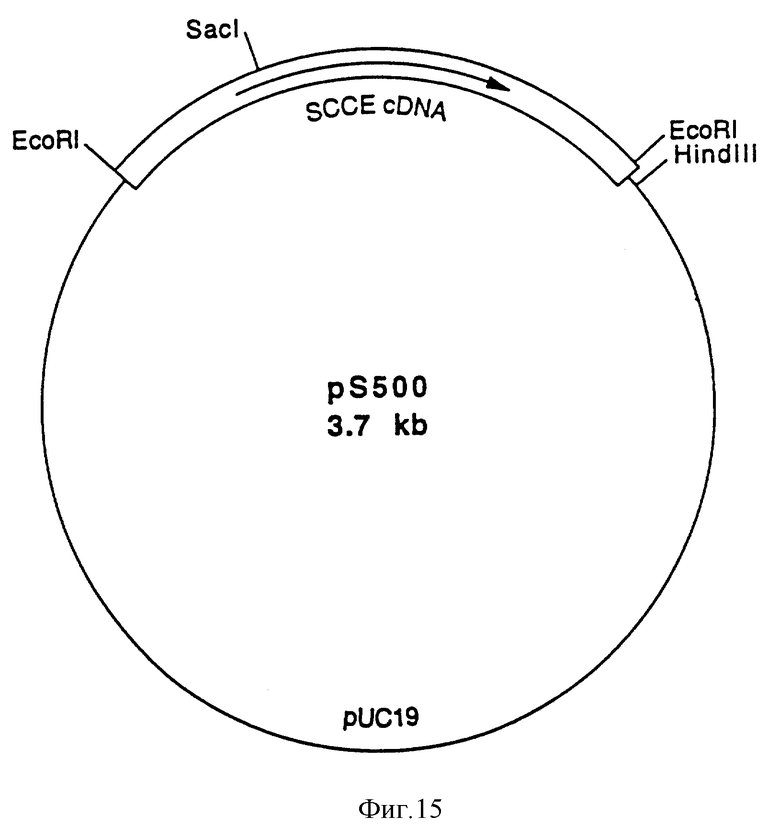


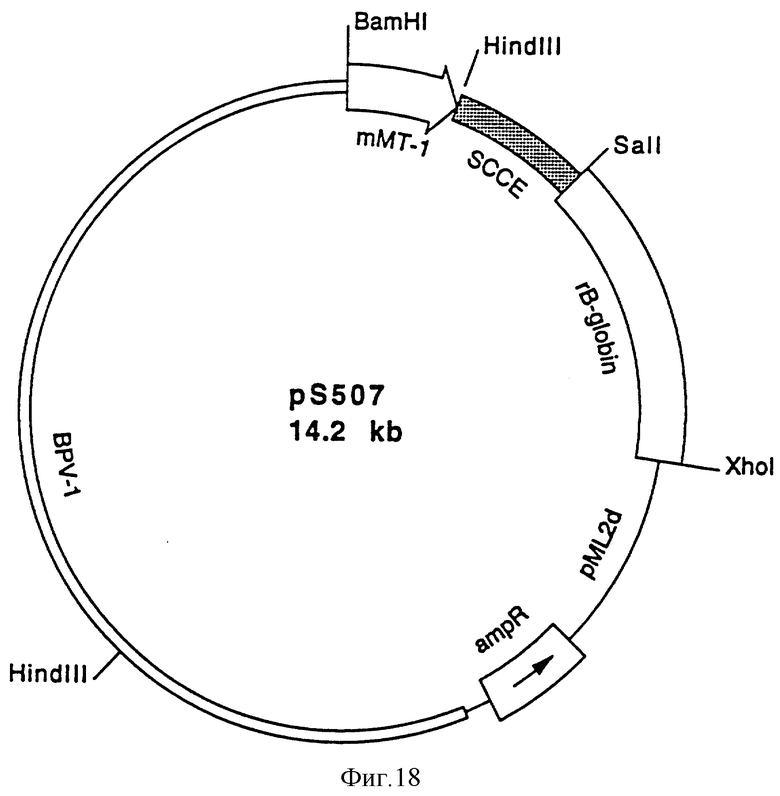




Изобретение относится к биотехнологии и медицине и может быть использовано для получения рекомбинантного химотрипсинового энзима роговичного слоя (SCCE). Рекомбинантный полипептид, обладающий активностью SCCE, получают путем культивирования мышиных клеток С127, трансформированных вектором экспрессии, содержащим нуклеотидную последовательность (SEQ IDN1), кодирующую указанный полипептид. В фармацевтическую композицию для лечения нарушений, связанных с кератинизацией, включают 0,00l - 25% нативного или рекомбинантного SССЕ. Изобретение позволяет получить энзим, активный в отношении роговичного слоя кожи, и использовать его в составе фармацевтических и косметических композиций. 7 с.п. ф-лы, 23 ил., 5 табл.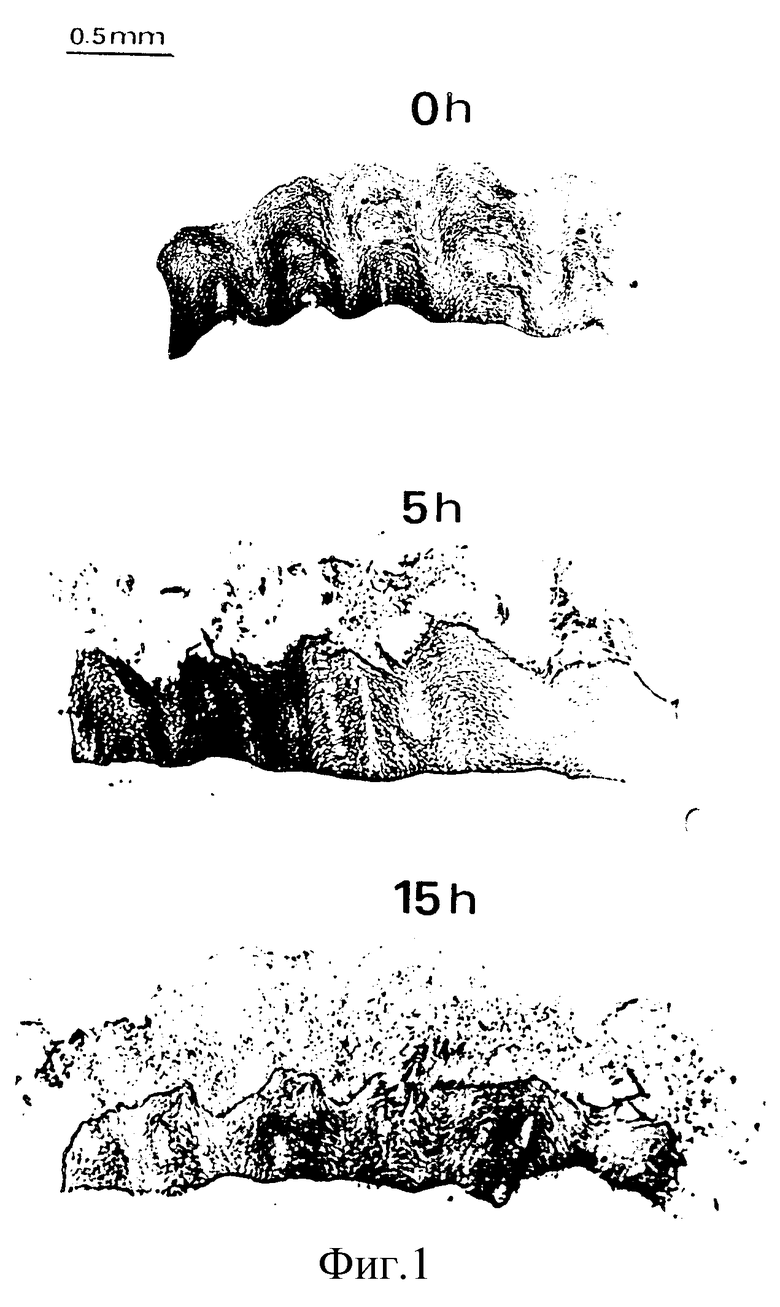
| Egelrud T | |||
| et al., Archives of Dermatogical Research, v.283, N 2, p.108-112, 1991 | |||
| Lundstrom A | |||
| et al., Actadermato - Venereologica, v.71, N 6, p.471-474, 1991 | |||
| Способ управления проводкой бумажного полотна на бумагоделательной машине | 1973 |
|
SU492614A1 |
| ГАЗОНАПОЛНЕННЫЙ ПРИБОР С ТЕЛОМ НАКАЛА | 0 |
|
SU231425A1 |
| СПОСОБ ЛЕЧЕНИЯ ИНФИЦИРОВАННЫХ РАН | 1992 |
|
RU2036658C1 |

