Изобретение относится к молекулам ДНК, рекомбинантным векторам и клеточным культурам, предназначенным для использования в способах экспрессии стимулируемой солями желчи липазы (BSSL) в метилотрофных дрожжах Pichia pastoris.
Считается, что стимулируемая солями желчи липаза (BSSL; EC 3.1.1.1) (см. ссылку на работу Wang и Hartsuck, 1993) ответственна за основную часть липолитической активности человеческого молока. Отличительный признак такой липазы состоит в том, что необходимо наличие первичных солей желчи для обеспечения ее активности в отношении эмульгированных длинных цепей триацилглицеринов. BSSL к настоящему времени была обнаружена только в молоке людей, горилл, кошек и собак (Hernell с сотр., 1989).
BSSL приписывается решающая роль в процессе переваривания молочных липидов в кишечнике детей, кормящихся грудью (Fredrikzon с сотр., 1978). BSSL синтезируется в организме женщин в период лактации молочных желез и секретируется с молоком (Bläckberg с сотр., 1987). Считается, что она обеспечивает примерно 1% общего количества молочного белка (Bläckberg & Hernell, 1981).
Предполагается, что BSSL является основным лимитирующим скорость фактором в абсорбции жира и последующем росте, особенно в возрасте полового созревания, у детей с дефицитом собственного продуцирования BSSL и что добавка в рацион питания упомянутых выше детей рецептур, содержащих очищенный энзим, существенно улучшает пищеварение и способствует росту детей (US 4944944; Медицинский Исследовательский Фонд Оклахомы). Этот факт является весьма важным в клинической практике в области составления детских рецептур, содержащих относительно высокое процентное содержание триглицеридов и основанных на молочных белках растительного или нечеловеческого происхождения, так как новорожденные, питающиеся такими рецептурами, не способны переваривать жир в отсутствие добавляемой BSSL.
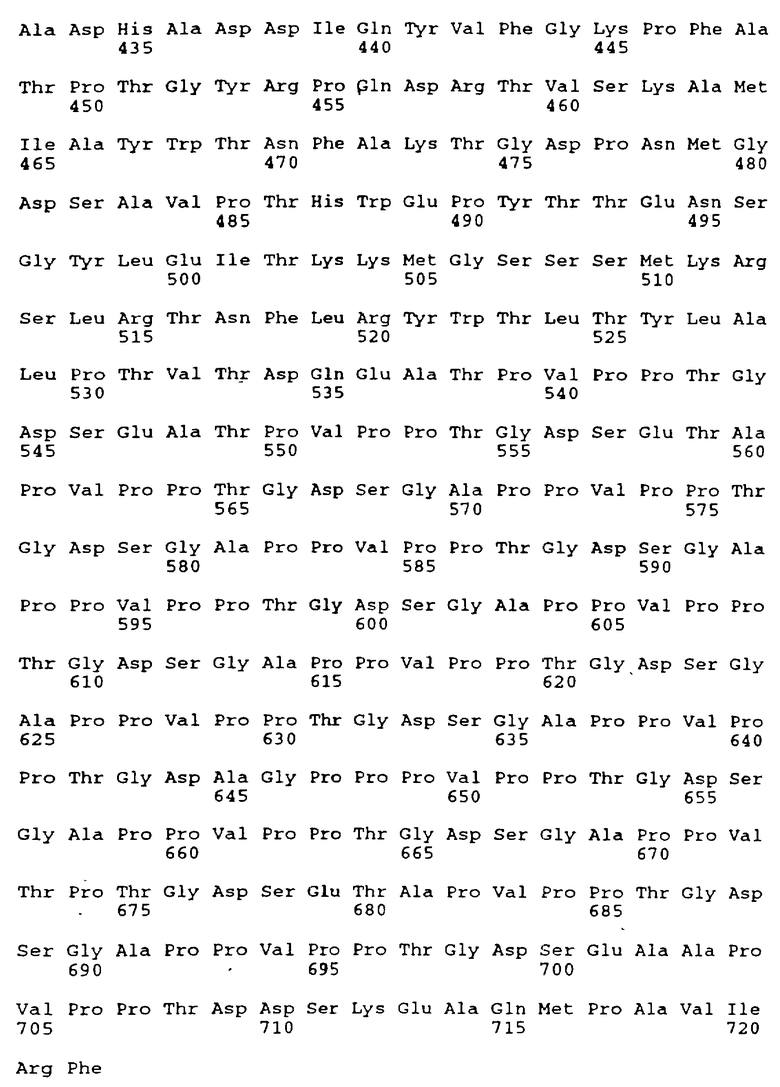
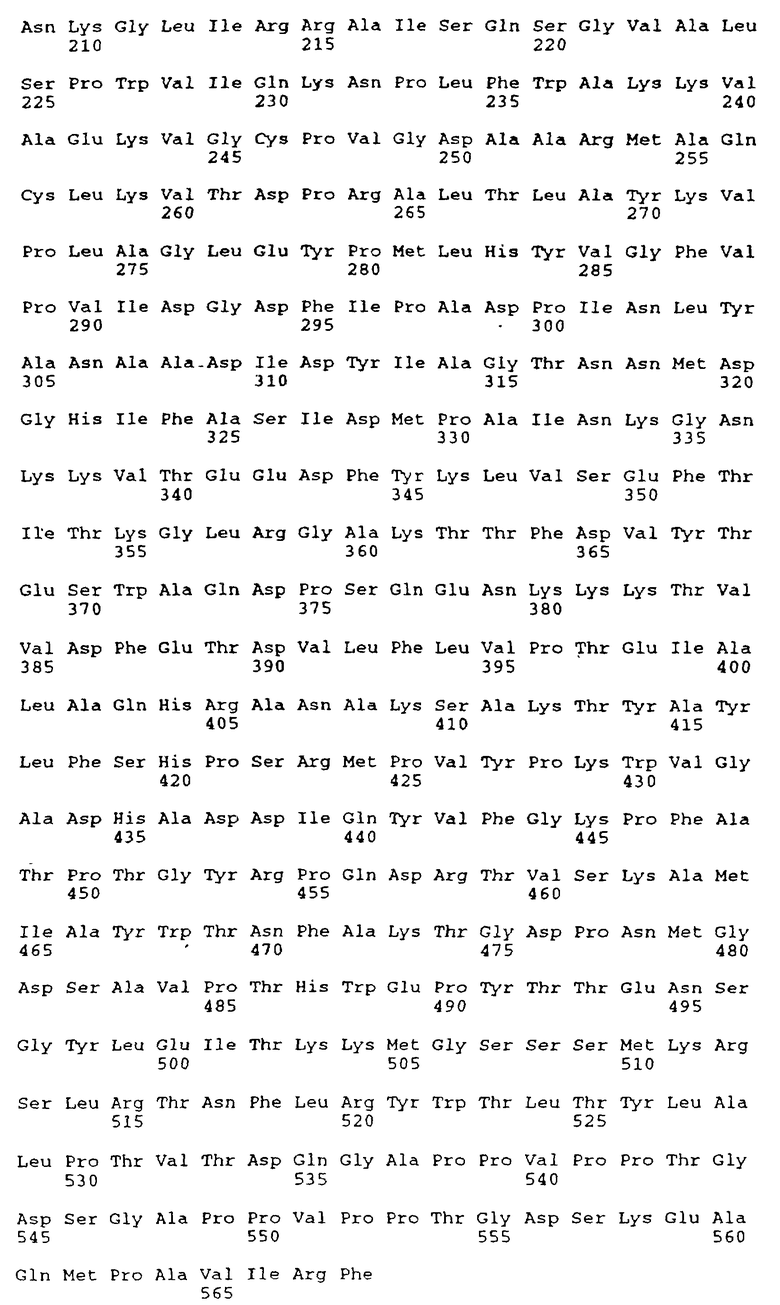
Структуры кДНК как молочной BSSL, так и карбонового сложного эфира гидролазы поджелудочной железы (СЕН) были охарактеризованы (Baba с сотр., 1991; Hui и Kissel, 1991; Nilsson с сотр., 1991; Reue с сотр., 1991) и был сделан вывод, что молочный энзим и фермент поджелудочной железы являются продуктами одного и того же гена, гена CEL. Последовательность кДНК (SEQ ID N: 1) CEL гена раскрыта в патенте США 5200183 (Медицинский Исследовательский Фонд Оклахомы); WO 91/18293 (Actiebolaret Astra); Nilsson с сотр. (1990); a также Baba с сотр. (1991). Выведенная аминокислотная последовательность белка BSSL, включающая сигнальную последовательность из 23 аминокислот, представлена как Последовательность SEQ ID N: 2 в Перечне последовательностей, приведенном в конце описания, тогда как последовательность нативного белка, состоящего из 722 аминокислот, представлена как SEQ ID N: 3.
C-терминальная область белка содержит 16 повторов из 11 аминокислотных остатков каждый, за которыми следует консервативный участок из 11 аминокислот. Нативный белок имеет высокую степень гликозилирования, и имеются сообщения о широком интервале наблюдаемых молекулярных весов. Вероятно, этот факт может быть объяснен изменением степени гликозилирования (Abouakil с сотр., 1988). N-терминальная половина белка гомологична ацетилхолинэстеразе и некоторым другим эстеразам (Nilsson с сотр., 1990).
Рекомбинантная BSSL может быть продуцирована экспрессией в таком подходящем хозяине, как E.coli, Saccharomyces serevisiae или клеточные линии млекопитающих. Для масштабирования системы экспрессии BSSL с целью достижения коммерчески приемлемой стоимости производства может рассматриваться применение гетерологических экспрессирующих систем. Как упоминалось выше, человеческая BSSL имеет 16 повторов из 11 аминокислот на C-терминальном конце. Для определения биологической значимости такого участка повторов конструировались различные мутанты человеческой BSSL, у которых частично или полностью отсутствовали участки повтора (Hansson с сотр., 1993). Так, например, вариант BSSL-C (SEQ ID N: 4) имеет делеции от аминокислотных остатков 536-568 и аминокислотных остатков 591-711. Исследования экспрессии с использованием линии клеток млекопитающего C127 и сектора экспрессии на основе вируса коровьей папиломы показали, что различные варианты могут экспрессироваться в активных формах (Hansson с сотр., 1993). Из исследований экспрессии был также сделан вывод, что повторы, обогащенные пролином, в человеческой BSSL не играют существенной роли в каталитической активности или активации BSSL солями желчи. Однако получение BSSL или ее мутантов в экспрессионной системе млекопитающего оказывается слишком дорогостоящим для рутинного терапевтического использования.
Эукариотическая система, например дрожжи, может обеспечить значительные преимущества по сравнению с использованием прокариотических систем в плане получения некоторых полипептидов, кодируемых рекомбинантной ДНК. Так, например, дрожжи обычно могут расти до более высокой плотности клеток, чем бактерии, и могут оказаться способными к гликозилированию экспрессированных полипептидов в том случае, когда такое гликозилирование важно для биологической активности. Однако применение дрожжей разновидности Saccharomyces cerevisiae в качестве организма-хозяина часто приводит к плохим уровням экспрессии и плохой секреции рекомбинантного белка (Cregg с сотр., 1987). Максимальные уровни содержания гетерологических белков в S.cerevisiae составляют величину порядка 5% от общего количества клеточного белка (Kingsman с сотр., 1985). Дополнительным недостатком применения Saccharomyces cerevisiae в качестве хозяина является то, что рекомбинантные белки проявляют тенденцию к сверхгликозилированию, которое может оказывать влияние на активность гликозилированных белков млекопитающих.
Pichia pastoris представляют собой метилотрофные дрожжи, которые способны расти на метаноле в качестве единственного источника углерода и энергии, поскольку для такого микроорганизма характерен высокорегулируемый путь утилизации метанола (Ellis с сотр., 1985). P. pastoris также способны подвергаться эффективной технологии ферментации с высокой плотностью клеточной популяции. В связи с этим технология рекомбинантной ДНК и эффективные способы трансформации дрожжей позволили использовать P.pastoris в качестве хозяина для экспрессии гетерологического белка в большом количестве с помощью экспрессионной системы на основе метанолоксидазного промотора (Cregg с сотр. , 1987).
Применение Pichia pastoris в качестве хозяина для экспрессии известно из литературы, например, следующих гетерологических белков: фактора некроза человеческих опухолей (EP-A-0263311); пертактиновых антигенов Bordetella (WO 91/15571); поверхностного антигена вируса гепатита В (Cregg с сотр., 1987); человеческого лизоцимного белка (WO 92/04441); апротинина (WO 92/01048). Однако успешная экспрессия гетерологического белка в активной, растворимой и секретированной форме зависит от большого числа факторов, например правильного выбора сигнального пептида, надлежащего конструирования участка слияния между сигнальным пептидом и зрелым белком, условий роста и т.д.
Цель настоящего изобретения состоит в преодолении упомянутых выше недостатков известных систем и разработке способа получения человеческой BSSL, который являлся бы экономически выгодным и давал выходы продукта, сравнимые или превосходящие производительность процесса в других микроорганизмах. Эта цель была достигнута разработкой способов экспрессии BSSL в клетках Pachia pastoris.
Таким образом, с помощью настоящего изобретения было показано, что человеческая BSSL и вариантная BSSL-C могут экспрессироваться в активной форме, секретируемой из P. pastoris. Нативный сигнальный пептид, а также гетерологический сигнальный пептид, являющийся произведенным белка инвертазы S. cerevisiae, были использованы для транслокации зрелого белка в культуральную среду в активной, надлежащим образом процессированной форме.
В соответствии с первым аспектом изобретение предусматривает ДНК-молекулу, содержащую:
(a) область, кодирующую полипептид, представляющий собой человеческую BSSL или ее биологически активный вариант;
(b) присоединенный к 5'-концу области, кодирующей полипептид, участок, кодирующий сигнальный пептид, способный управлять секрецией указанного полипептида из клеток Pichia pastoris, трансформированных с помощью указанной ДНК-молекулы; и
(c) оперативно связанный с кодирующими областями, указанными в (a) и (b), метанолоксидазный промотор Pichia pastoris или функционально эквивалентный промотор.
Под термином "биологически активный вариант" BSSL понимается полипептид, обладающий BSSL активностью и содержащий часть аминокислотной последовательности, представленной последовательностью SEQ ID N: 3 в Перечне последовательностей (Перечень последовательностей приведен в конце описания). Под термином "полипептид, обладающий BSSL активностью", в настоящем контексте понимается полипептид, обладающий следующими свойствами: (a) пригодностью для орального применения; (b) способностью активироваться специфическими солями желчи; и (c) способностью к действию в качестве неспецифической липазы в содержимом тонкого кишечника, т.е. способностью к гидролизу липидов независимо от их химической структуры и физического состояния (эмульгированного, мицеллярного, растворимого).
Указанный BSSL-вариант может представлять собой, например, вариант, содержащий менее 16 повторяющихся единиц, вследствие чего термин "повторяющаяся единица" понимается как повторяющаяся единица из 11 аминокислот, закодированных нуклеотидной последовательностью, обозначенной как "повторяющаяся единица" в разделе под заголовком "(IX) Отличительный признак" в "Информации для SEQ ID N: 1" в Перечне последовательностей. Главным образом BSSL-вариант может представлять собой вариант BSSL-C, в котором аминокислоты 536-568 и 591-711 подвергнуты делеции (SEQ ID N: 4 в Перечне последовательностей).
Следовательно, ДНК-молекула в соответствии с изобретением предпочтительно представляет собой молекулу ДНК, которая кодирует BSSL (SEQ ID N: 3) или BSSL-C (SEQ ID N: 4).
Однако ДНК-молекулы согласно настоящему изобретению не должны строго ограничиваться молекулами ДНК, кодирующими полипептиды с аминокислотными последовательностями, идентичными SEQ ID N: 3 или 4 в Перечне последовательностей. Наоборот, изобретение охватывает молекулы ДНК, кодирующие полипептиды, несущие такие модификации, как замены, небольшие делеции, инсерции или инверсии, причем такие полипептиды в значительной мере обладают биологическими активностями BSSL. Таким образом, настоящее изобретение включает молекулы ДНК, кодирующие BSSL-варианты, упомянутые выше, а также молекулы ДНМ, кодирующие полипептиды, аминокислотные последовательности которых по крайней мере на 90% гомологичны, предпочтительно по крайней мере на 95% гомологичны аминокислотной последовательности, представленной последовательностью SEQ ID N: 3 или 4 в Перечне последовательностей.
Сигнальный пептид, на который ссылались выше, может представлять собой пептид, идентичный или в значительной мере подобный пептиду с аминокислотной последовательностью, представленной как аминокислоты от -20 до -1 в SEQ ID N: 2 в Перечне последовательностей. С другой стороны, это может быть пептид, включающий сигнальный пептид инвертазы Saccharomyces cerevisiae.
В соответствии с другим аспектом изобретение предусматривает вектор, содержащий молекулу ДНК, упомянутую выше. Предпочтительно такой вектор представляет собой способный к репликации вектор экспрессии, который несет и способен управлять экспрессией в клетках вида Pichia ДНК-последовательности, кодирующей человеческую BSSL или ее биологически активный вариант. Такой вектор может представлять собой, например, плазмидный вектор pARC 5771 (NCIMB 40721), pARC 5799 (NCIMB 40723) или pARC 5797 (NCIMB 40722).
Согласно еще одному аспекту изобретение предусматривает культуру клеток-хозяев, содержащую клетки вида Pichia, трансформированные с помощью упомянутых выше молекулы ДНК или вектора. Предпочтительно клетки-хозяева представляют собой клетки Pichia pastoris такого штамма, как PPF-1 или GS115. В качестве указанных клеточных культур могут использоваться, например, культура PPF-1 [pARC 5771] (NCIMB 40721), GS115 [pARC 5799] (NCIMB 40723) или GS115 [pARC 5797] (NCIMB 40722).
В соответствии с еще одним аспектом изобретение предусматривает способ получения полипептида, представляющего собой человеческую BSSL или ее биологически активный вариант, заключающийся в культивировании клеток-хозяев согласно изобретению в условиях, при которых указанный полипептид секретируется в культурную среду, и выделении указанного полипептида из культурной среды.
ПРИМЕР 1: Экспрессия BSSL в Pichia pastoris PPF-1.
1.1. Конструкция pARC 0770.
Последовательность кДНК (SEQ ID N: 1), кодирующую BSSL-белок, включающий нативный сигнальный пептид (на который ниже ссылаются, как на NSP), клонировали в pTZl9R (Фармация) в виде фрагмента EcoRI-SacI. Клонирование кДНК NSP-BSSL в S.cerevisiae экспрессионный вектор pSCW 231 (полученный от профессора L. Prakash, Рочестерский университет, NY, США), представляющий собой вектор экспрессии дрожжей с низким числом копий, в котором экспрессия осуществляется под контролем конститутивного ADH1 промотора, проводили в две стадии. Вначале кДНК NSP-BSSL клонировали в pYES 2.0 (Инвитроген, США) в виде фрагмента EcoRI-SpbI из pTZ19R-SP-BSSL. Избыточные 89 пар оснований между EcoRI и NcoI в начале кодирующей последовательности сигнального пептида удаляли созданием слияния EcoRI/NcoI (89) и регенерацией EcoRI сайта. Полученный в результате клон pARC 0770 содержал ATG кодон, первоначально закодированный в NcoI сайте, за которым сразу же располагался регенерированный EcoRI сайт в рамке считывания информации с оставшейся NSP-BSSL последовательностью.
1.2. Конструкция pARC 5771 плазмиды.
Для конструирования подходящего экспрессионного вектора для экспрессии BSSL, кДНК фрагмент, кодирующий BSSL белок, совместно с его нативным сигнальным пептидом клонировали с экспрессионным вектором pDM 148 P.pastoris. Векторный pDM 148 (полученный от д-ра S. Subramani, UCSD) конструировали следующим образом: расположенную против хода транскрипции нетранслированную область (5'-UTR) и расположенную по ходу транскрипции нетранслированную область (3'-UTR) гена метанолоксидазы (MOX1) выделяли методом PCR и помещали тандемно в множественную клонирующую последовательность (MCS) вектора pSK+ E.coli (полученного от Стратаген, США).
Для надлежащего отбора предполагаемых трансформантов Р.pastoris ДНК последовательность, кодирующую ген ARG4 S.cerevisiae совместно с его собственной промоторной последовательностью, вставляли между 5'- и 3'-UTR в pSK-. Полученная в результате конструкция pDM 148 имела следующие отличительные признаки: в MCS области pSK- клонировали 5'-UTR МОХ, геномную последовательность ARG4 S.cerevisiae и 3'-UTR МОХ. Между 5'-UTR MOX и геномной последовательностью ARG4 располагали серии уникальных рестрикционных сайтов (SalI, ClaI, EcoRI, PstI, SmaI и BamHI) так, что кодирующая последовательность гетерологического белка может клонироваться для экспрессии под контролем MOX промотора в P.pastoris. Для облегчения интеграции такого полигенного экспрессирующего кластера в MOX1-локус в хромосоме Р.pastoris такой экспрессирующий полигенный кластер может быть отщеплен от оставшейся части pSK- вектора путем расщепления рестрикционным энзимом NotI.
5'-UTR MOX1 разновидности P. pastoris, клонированная в pDM 148, имела длину около 150 bp, тогда как 3'-UTR MOX1 из Р.pastoris, клонированная в pDM 148, имела длину около 1000 bp. Для инсерции NSP-BSSL кДНК-последовательности между 5'-UTR MOX1 и ARG4 кодирующей последовательностью S.cerevisiae в pDM 148, кДНК-вставку (SP-BSSL) выделяли из pARC 0770 путем расщепления EcoRI и BamHI (фрагмент ДНК размером примерно 2.2 kb) и клонировали между EcoRI и BamHI сайтами в pDM 148.
Полученная в результате конструкция pARG 5771 (NCIMB 40721) содержала 5'-UTR MOX1 P. pastoris с последующей NSP-BSSL кодирующей последовательностью, за которой следовала ARG4 генная последовательность S.cerevisiae и 3'-UTR MOX1 гена P.pastoris, тогда как ДНК сегмент от 5'-UTR MOX1 до 3'-UTR MOX1 клонировали в MCS pSK-.
1.3. Трансформация BSSL в P.pastoris PPF-1.
Для экспрессии BSSL в P.pastoris PPF-1 (his4, arg4; получено от Филлипс Петролеум Ко.) плазмиду pARC 5771 переваривали NotI и всю расщепленную смесь (10 мкг общей ДНК) использовали для трансформации PPF-1. Используемый протокол трансформации был практически таким же, как метод для дрожжевого сферопласта, описанный Cragg с сотр. (1987). Трансформанты регенерировали на минимальной среде с дефицитом аргинина так, чтобы можно было осуществлять селекцию Arg+ колоний. Регенерационный верхний агар, содержащий трансформанты, снимали и гомогенизировали в воде и дрожжевые клетки культивировали до образования примерно 250 колоний в расчете на пластину на минимальных глюкозных пластинах с дефицитом аргинина. Затем мутантные колонии идентифицировали методом реплик на минимальных метанольных пластинах. Примерно 15% от всего числа трансформантов образовывалось в виде Muts (медленный рост в метаноле) фенотипа.
1.4. Скрининг трансформантов у экспрессирующих BSSL.
С целью быстрого скрининга большого числа трансформантов на экспрессию липазы был разработан метод анализа липазы на платах. Использовали следующую методику для приготовления таких пластин: к раствору 2% агарозы (конечная концентрация) добавляли 10 х Na-холатный раствор в воде до конечной концентрации 1%. Липидный субстратный трибутин добавляли в смесь до конечной концентрации 1% (об./об.). Для поддержания роста трансформантов смесь дополняли 0.25% дрожжевого азотистого основания (конечное значение) и 0.5% метанола (конечная концентрация). Ингредиенты тщательно перемешивали и переливали на пластины до толщины 3-5 мм. Сразу после отвердевания смеси трансформанты полосками наносили на пластины и полученные пластины дополнительно инкубировали при +37oC в течение 12 часов. Липаза-продуцирующие клоны демонстрировали прозрачный гало вокруг клона. В типичном эксперименте 7 из 93 трансформантов идентифицировали, как BSSL продуцирующие трансформанты. Два клона (NN 39 и 86), продуцирующие самые крупные гало (ореолы) вокруг полоски колонии, отбирали для дальнейшей характеристики.
1.5. Экспрессия BSSL из PPF-1 [pARC 5771].
Два трансформанта NN 39 и 86, описанные в Разделе 1.4, отбирали и выращивали в BMGY жидкой среде (1% дрожжевого экстракта, 2% бактопептона, 1.34% дрожжевого азотистого основания без аминокислоты, 100 мМ KPO4 буфера, pH 6.0, 400 мкг/л биотина и 2% глицерина) в течение 24 часов при 30oC до достижения культурами значения A600, близкого к 40. Культуры осаждали и ресуспендировали в BMMY (2% глицерина, замененного 0.5% метанола в BMGY) среде при A600 = 300. Индуцированные культуры инкубировали при 30oC при встряхивании в течение 120 часов. Супернатанты культуры отбирали через различные промежутки времени для анализа на экспрессию BSSL путем осуществления анализа на активность энзима, SDS-PAGE анализа и вестерн-блоттинга.
1.6. Детекция ферментативной активности BSSL в культуральных супернатантах клонов NN 39 и 86.
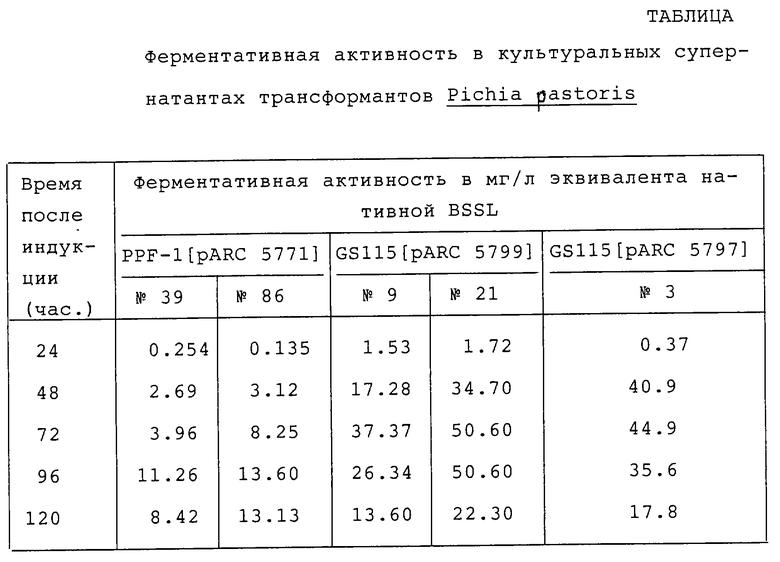
Для определения ферментативной активности в бесклеточном культуральном супернатанте индуцированных культур NN 39 и 86, как описано в Разделе 1.5, культуры центрифугировали и 2 мкл бесклеточного супернатанта анализировали на BSSL-энзимную активность в соответствии со способом, описанным Hernell и Olivecrona (1974). Как показано в таблице, было установлено, что обе культуры содержат BSSL-энзимную активность при максимальном значении активности на 96 час после индукции.
1.7. Вестерн-блоттинг анализ культуральных супернатантов PPF-1: pARC 5771 трансформантов (NN 39 и 86).
Для определения наличия рекомбинантной BSSL в культуральных супернатантах NN 39 и 86 PPF-1 [pARC 5771] трансформантов, культуры выращивали и индуцировали, как описано в Разделе 1.5. Культуры отбирали через различные промежутки времени после индукции и подвергали Вестерн-блоттинг анализу с использованием анти-BSSL поликлонального антитела. Полученные результаты указывают на присутствие BSSL в культуральном супернатанте в виде полосы в 116 кДа.
ПРИМЕР 2: Экспрессия BSSL в Pichia pastoris GS115.
2.1. Конструкция pARC 5799.
Поскольку 5'-MOX UTR и 3'-MOX UTR не выражены надлежащим образом и поскольку в pDM 148 векторе отсутствует какой-либо другой подходящий маркер (например, G418 устойчивый ген) для мониторинга числа копий BSSL, интегрированных в Pichia хромосоме, кДНК вставку нативной BSSL совместно с ее сигнальным пептидом клонировали в другой P. pastoris экспрессионный вектор, pHIL D4. Интеграционную плазмиду pHIL D4 получали от Филлипс Петролеум Компани. Плазмида содержала 5'-MOX1, сегмент алкогольоксидазного промотора размером примерно 1000 bp и уникальный EcoRI - клонирующий сайт. Она также содержала примерно 250 bp 3'-MOX1 участка, содержащего терминационную последовательность алкогольоксидазы после EcoRI-сайта. Район "терминации" сопровождался гистидинолдегидрогеназным геном HIS4 P.pastoris, содержащимся на 2.8-kb фрагменте для комплементации дефектного HIS4 гена в хозяине GS115 (см. ниже). 650-bp участок, содержащий 3'-MOX1 ДНК, был слит с 3'-окончанием HIS4 гена, что совместно с 5'-MOX1 участком необходимо для сайт-направленной интеграции. Бактериальный канамицин-устойчивый ген из pUC-4K (PL-Биокэмикал) подвергали инсерции по уникальному NaeI-сайту между HIS4 и 3'-MOX1 участку на 3'-окончании HIS4 гена.
Для клонирования NSP-BSSL-кодирующего фрагмента кДНК по уникальному EcoRI сайту pHIL D4, двухнитевой олиголинкер с положением расщепления BamHI-EcoRI лигировали с BamHI переваренной плазмидой pARC 5771, и полную NSP-BSSL-кодирующую последовательность объединяли в виде EcoRI-фрагмента размером 2.2 kb. Этот фрагмент клонировали в EcoRI-сайт pHIL D4 и правильно ориентированную плазмиду обозначали как pARC 5799 (NCIMB 40723).
2.2. Трансформация pARC 5799.
Для облегчения интеграции NSP-BSSL кодирующей последовательности по геномному локусу MOX1 в P.pastoris плазмиду pARC 5799 переваривали BglII и использовали для трансформации P.pastoris штамма GS115(his4) (Филлипс Петролеум Компани) в соответствии с протоколом, описанным в Разделе 1.5. Однако в этом случае селекцию проводили на His прототрофность. Трансформанты отбирали после серийно разбавленного посева регенерированного верхнего агара и непосредственно тестировали анализом культивирования липазы, как описано в Разделе 1.4. Два трансформированных клона (NN 9 и 21) отбирали на основе размера ореола на пластинке для анализа липазы и дополнительно проверяли на экспрессию BSSL. Было установлено, что такие клоны являются Mut+.
2.3. Определение BSSL энзимной активности в культуральных супернатантах GS115[pARC 5799] трансформантов NN 9 и 21.
Два этих трансформированных клона NN 9 и 21 GS115[pARC 5799] выращивали практически полностью следуя протоколу, описанному в Разделе 1.5. Культуральные супернатанты в различные периоды времени после индукции анализировали на BSSL-энзимную активность в соответствии с описанным в Разделе 1.6. Как показано в таблице, было установлено, что оба культуральных супернатанта содержат BSSL-энзимную активность, причем наивысшая энзимная активность наблюдалась через 72 часа индукции. Оба клона демонстрировали улучшенную экспрессию BSSL по сравнению с клонами PPF-1[pARC 5771].
2.4. SDS-PAGE и Вестерн-блоттинг анализ культуральных супернатантов GS115 [pARC 5799] трансформантов NN 9 и 21.
Культуральные супернатанты, собранные в различные периоды времени, как описано в Разделе 2.3, подвергали анализу SDS-PAGE и Вестерн-блоттингу. Из SDS-PAGE профиля было оценено, что около 60-75% общего белка, присутствующего в культуральных супернатантах индуцированных культур, представляет собой BSSL. Молекулярный вес такого белка составил величину около 116 кДа. Данные Вестерн-блоттинга также подтвердили, что основная часть белка, присутствующего в культуральном супернатанте, представляет собой BSSL. Такой белок, по-видимому, имеет тот же молекулярный вес, что нативная BSSL.
ПРИМЕР 3: Масштабирование BSSL экспрессии.
3.1. Масштабирование экспрессии BSSL из трансформированного клона GS115 [pARC 5799] (N 21).
Использовали ферментер Б. Брауна емкостью 23 л. Пять литров среды, содержащей 1% YE, 2% Пептона, 1.34 YNB и 4% вес./об. глицерина обрабатывали в автоклаве при 121oC в течение 30 минут и биотин (400 мкг/л, конечная концентрация) добавляли в ходе инокуляции после фильтрационной стерилизации. В качестве прививочного материала глицериновую массу GS115 [pARC 5799] (N 21) инокулировали в синтетическую среду, содержащую YNB (67%) плюс 2% глицерина (150 мл), и выращивание проводили в течение 36 часов при +30oC. Применяли следующие условия ферментации: температура составляла +30oC; pH 5.0 поддерживали с использованием 3.5 N NH4OH и 2 N HCl; концентрация растворенного кислорода составляла 20-40% от насыщения воздуха; в качестве противопенного агента использовали полипропиленгликоль 2000.
За ходом роста следили через регулярные интервалы времени, определяя значение оптической плотности (OD) при длине волны 600 нм. Величина A600 достигала максимума, равного 50-60, за 24 часа. К этому моменту протекала фаза периодического роста, о чем свидетельствовали повышенные уровни растворенного кислорода.
Фаза роста сразу сопровождалась фазой индукции. В ходе этой фазы загружали метанол, содержащий 12 мл/л PTM1 солей. Скорость подачи метанола составляла 6 мкл/ч в ходе первых 10-12 часов, после чего скорость постепенно повышали каждые 7-8 часов на 6 мл/ч, до максимального значения 36 мл/ч. Аммиак, используемый для регулирования pH, выполнял функции источника азота. Накопление метанола проверяли каждые 6-8 часов путем зондирования растворенного кислорода, и было установлено, что такая аккумуляция является ограниченной в ходе всей фазы индукции. Значение OD при 600 нм увеличивалось от 50-60 до 150-170 в ходе 86 часов подачи метанола. Дрожжевой экстракт и пептон добавляли каждые 24 часа до получения конечной концентрации 0.25% и 0.5% соответственно.
Образцы отбирали через 24-часовые промежутки времени и анализировали на BSSL-энзимную активность в бесклеточном бульоне. Бульон также подвергали анализу SDS-PAGE и Вестерн-блоттингу.
3.2. Белковый анализ секретированной BSSL из выращенной в ферментере культуры GS115 [pARC 5799] (N 21).
BSSL-энзимная активность в бесклеточном бульоне повышалась от 40-70 мг/л (что эквивалентно для нативного белка) к 24 часу до максимального значения 200-227.0 мг/л (что эквивалентно нативному белку) к концу 86-90 часа. SDS-PAGE анализ бесклеточного бульона показал полосу интенсивного окрашивания Кумасс голубым с мол. вес. порядка 116 кДа. Идентичность такой полосы была подтверждена Вестерн-блоттингом, осуществленным, как описано в Разделе 1.7, для нативного BSSL.
3.3. Очистка рекомбинантной BSSL, секретированной в культуральный супернатант GS115 [pARC 5799] (N 21) клонов.
P. pastoris клон GS115 [pARC 5799] выращивали и индуцировали в ферментере, как описано в Разделе 3.1. Для очистки рекомбинантной BSSL 250 мл культурной среды (индуцированной к 90 часу) центрифугировали с силой 12.000 х г в течение 30 минут для удаления всего мелкозернистого материала. Бесклеточный культуральный супернатант ультрафильтровали в устройстве Амикон с использованием мембраны, отсекающей фракции 10 кДа. Соли и низкомолекулярные белки, а также пептиды культурального супернатанта удаляли повторным разбавлением в ходе фильтрации. Буфер, используемый для такого разбавления, представлял собой 5 мМ Барбитола при pH 7.4. После концентрирования культурального супернатанта ретентат доводили до объема 250 мл с использованием 5 мМ Барбитола, pH 7.4 и 50 мМ NaCl и загружали в Гепарин-Сефарозную колонку (объем слоя 15 мл), которую предварительно уравновешивали с помощью того же буфера. Загрузку образца осуществляли с объемной скоростью 10 мл/ч. После загрузки колонку промывали 5 мМ Барбитола, pH 7.4 и 0.1 M NaCl (200 мкл промывного буфера) до тех пор, пока поглощение при длине волны 250 нм не становилось ниже уровня детекции. BSSL элюировали 200 мл Барбитольного буфера (5 мМ, pH 7.4) и линейным градиентом NaCl в интервале 0.1 М - 0.7 М. Фракции (2.5 мл) собирали и анализировали на элюированный белок путем мониторинга поглощения при 250 нм. Фракции, содержащие белок, анализировали на BSSL-энзимную активность. Соответствующие фракции анализировали на 8.0% SDS-PAGE для проверки профиля очистки.
3.4. Характеристика очищенной рекомбинантной BSSL, секретированной в культуральный супернатант GS115 [pARC 5799].
SDS-PAGE и Вестерн-блоттинг анализы фракций (описанных в Разделе 3.3), проявляющих максимальную BSSL-энзимную) активность, продемонстрировали, что рекомбинантный белок имеет чистоту примерно 90%. В соответствии с данными SDS-PAGE и Вестерн-блоттинг анализов молекулярный вес очищенного белка имел величину около 116 кДа. При перегрузке образцов для SDS-PAGE анализа полоса, соответствующая низкомолекулярному белку, может быть детектирована окрашиванием красителем Кумасс Бриллиантовым голубым, которая не проявляется при Вестерн-блоттинге. Очищенный белок подвергали N-терминальному анализу в автоматизированном белковом секвенаторе. Полученные результаты показали, что белок соответствующим образом процессирован из нативного сигнального пептида и что рекомбинантный белок имеет N-терминальную последовательность A K L G A V Y. Удельная активность очищенного рекомбинантного белка, как было установлено, аналогична активности нативного белка. Сведения о ферментативной активности в культуральных супернатантах трансформантов Pichia pastoris приведены в таблице.
ПРИМЕР 4: Экспрессия BSSL-C в Pichia pastoris GS115.
4.1. Конструкция pARC 5797.
кДНК кодирующую последовательность для BSSL варианта BSSL-C сливали по ее 5'-окончанию с кодирующей последовательностью сигнального пептида S.cerevisiae SUC2 генного продукта (инвертаза), сохраняя целостность открытой рамки считывания, инициированной по первому ATG кодону сигнального пептида инвертазы. Такую слитую генную конструкцию вначале клонировали в S.cerevisiae экспрессионный вектор pSCW 231 (pSCW 231 представляет собой вектор экспрессии дрожжей с низким числом копий, и экспрессия осуществляется под контролем конститутивного ADH1 промотора) между свитами EcoRI и BamHI с целью создания экспрессионного вектора pARC 0788.
кДНК слитого гена дополнительно субклонировали в P.pastoris экспрессионный вектор pDM 148 (описанный в разделе 1.2) путем высвобождения соответствующего 1.8-kb фрагмента с помощью EcoRI и BamHI переваривания pARC 0788 и субклонирования фрагмента в pDM 148 переваренную с EcoRI и BamHI. Сконструированный в результате pARC 5790 переваривали с BamHI и двухнитевой олигонуклеотидный линкер физической структуры BamHI-EcoRI-BamHI подвергали лигированию для создания сконструированной pARC 5796, главным образом, для выделения кДНК фрагмента слитого гена, следуя при этом стратегии, описанной в Разделе 2.1.
Наконец, 1.8-kb фрагмент, содержащий систему сигнальный пептид инвертазы/BSSL-C слитый ген, высвобождали из pARC 5796 путем EcoRI переваривания и клонировали в pHIL D4 по EcoRI сайту. В результате соответствующего рестрикционного анализа был идентифицирован вектор экспрессии, содержащий вставку в надлежащей ориентации, и он был обозначен как pARC 5797 (NCIMB 40722).
4.2. Экспрессия рекомбинантной BSSL-C из P.pastoris.
Для экспрессии рекомбинантной BSSL-C из P.pastoris Р. pastoris хозяин GS115 трансформировали с pARC 5797 по способу, описанному в Разделах 1.3 и 2.2. Трансформанты проверяли на продуцирование липазы по способу, описанному в Разделах 1.4 и 2.2. Единственный трансформант (N 3) отбирали на основе высокой липаза-продуцирующей способности с помощью метода детекции при анализе липазных пластин и дополнительно анализировали на продуцирование BSSL-энзимной активности в культуральном супернатанте, следуя, по существу, способу, описанному в разделах 1.6 и 2.3. Как следует из данных, представленных в таблице, культуральный супернатант GS115 [pARC 5797] (N 3) содержал BSSL-энзимную активность, и ее количество прогрессивно увеличивалось вплоть до 72 часа после индукции.
4.3. SDS-PAGE и Вестерн-блоттинг анализы культурального супернатанта GS115 [pARC 5797] трансформанта (N 3).
Культуральный супернатант, собранный в различные периоды времени, как описано в Разделе 4.2, подвергали SDS-PAGE и Вестерн-блоттинг анализам в соответствии с описанным в разделах 1.7 и 2.4. Из SDS-PAGE профиля было установлено, что примерно 75-80% общего количества внеклеточного белка представляет собой BSSL. В соответствии с данными SDS-PAGE анализа молекулярный вес белка составил примерно 66 кДа. При Вестерн-блоттинге было обнаружено, что лишь две полосы (дублет) в области 66 кДа являются иммунореактивными, и эти данные подтверждают экспрессию рекомбинантной BSSL-C.
СРАВНИТЕЛЬНЫЙ ПРИМЕР: Экспрессия BSSL в S.cerevisiae.
Были предприняты попытки экспрессии BSSL в Saccharomyces cerevisiae. BSSL плохо секретировалась в S. cerevisiae, и нативный сигнальный пептид функционировал неэффективно. Кроме этого, нативный сигнальный пептид не отщеплялся от зрелого белка в S.cerevisiae.
Депонирование микроорганизмов.
Следующие плазмиды, трансформированные в Pichia pastoris культуры, были депонированы в соответствии с Будапештским договором в Национальных коллекциях Промышленных и морских бактерий (NCIMB), Абердин, Шотландия, Великобритания. Дата депонирования - 2 мая 1995 г.
Штамм [плазмида] - N по NCIMB
PPF-1[pARC 5771] - 40721
GS115[pARC 5799] - 40723
GS115[pARC 5797] - 40722
Список литературы
Abouakil, N., Rogalska, E., Bonicel, J. and Lombardo, D. (1988) Biochim. Biophys. Acta. 961, 299-308.
Baba, Т., Downs, D., Jackson, K.W., Tang, J. and Wang, C-S (1991) Biochemistry 30, 500-510.
Bläckberg, L. and Hernell, O. (1981) Eur. J. Biochem. 116, 221-225.
Bläckberg,L.,Ängquist, K.A. and Hernell, O. (1987) FEBS Lett. 217, 37-41.
Cregg, J.M. et al. (1987) Bio/Technology 5, 479-485.
Ellis, S.B. et al. (1985) Mol. CelL Biol 5, 1111-1121.
Fredrikzon, В. , Hernell, О., Bläckberg, L. and Olivecrona, T. (1978) Pediatric Res. 12, 1048-1052.
Hansson, L., Bläckberg, L., Edlund, M., Lundberg, L., Strömqvist, M. and Hernell, O. (1993) J. Biol. Chem. 268, 26692-26698.
Hernell, O. and Olivecrona, T. (1974) Biochim. Biophys. Acta 369, 234-244.
Hernell, O. , Bläckberg, L. and Olivecrona, T. (1989) in: Textbook of gastroenterology and nutrition in infancy (Lebenthal, E., ed.) 347-354, Raven Press, NY.
Hernell, O. and Bläckberg, L. (1982) Pediatric Res. 16, 882-885.
Hui, D. Y. and Kissel, J. A. (1990) FEBS Letters 276, 131-134.
Kingsman, et al. (1985) Biotechnology and Genetic Engineering Reviews 3, 377-416.
Nilsson, J., Bläckberg, L., Carlsson, P., Enerbäck, S., Hernell, O. and Bjursell, G. (1990) Eur. J. Biochem. 192, 543-550.
Reue, К. , Zambaux, J., Wong, H., Lee, G., Leete, T.H., Ronk, M., Shively, J. E., Sternby, B., Borgström, B., Ameis, D. and Scholtz, M.C. (1991) J. Lipid. Res. 32, 267-276.
Wang, C-S, and Hartsuck, J.A. (1993) Biochim. Biphys Acta 1166, 1-19.











Изобретение относится к молекулам ДНК, рекомбинантным векторам и клеточным культурам, предназначенным для использования в способах экспрессии стимулируемой солями желчи липазы (BSSL) в метилтрофных дрожжах Pichia pastoris. Молекула ДНК содержит область кодирования полипептида, представляющего собой человеческую BSSL или ее биологический активный вариант с аминокислотной последовательностью, представленной в описании. К 5'-окончанию указанной области присоединен участок, кодирующий сигнальный пептид, способный управлять секрецией указанного полипептида из клеток Pichia pastoris, трансформированных указанной молекулой ДНК, с аминокислотной последовательностью, представленной в описании, а также оперативно связанный с указанными кодирующими областями метанолоксидазный промотор Pichia pastoris или функционально эквивалентный промотор. Описаны плазмидные векторы рARC 5771, рARC 5799, pARC 5797. Полипептид получают культивированием клеток-хозяев, трансформированных указанными плазмидами, с выделением его из культуральной среды. Изобретение позволяет увеличить выход целевого продукта по сравнению с известными способами. 5 с. и 3 з.п. ф-лы, 1 табл.
промотор метанолоксидазы Pichia pastoris или функционально эквивалентный промотор.
| RU 2060275 C1, 20.05.1996 | |||
| RU 94046360 A1, 10.06.1997 | |||
| Устройство для отделения сока от винограда | 1973 |
|
SU443063A1 |
| DE 4111321 A1, 17.10.1991 | |||
| US 5200183, 06.04.1993 | |||
| US 5306636, 26.04.1994. | |||

