Эта заявка является частичным продолжением заявки США, серийный номер 07/735069, поданной 25 июля 1991 г., озаглавленной "Индукция ответов цитотоксических Т-лимфоцитов", Syamal Raychaudhuri и William H. Rastetter. Это изобретение относится к методам и композициям, пригодным для индукцирования опосредованных цитотоксическими Т-лимфоцитами, ответов у людей и у домашних или сельскохозяйственных животных.
Цитотоксические Т-лимфоциты (ЦТЛ), как считают, являются главным механизмом защиты хозяина при ответе на ряд вирусных инфекций и опухолевый или раковый рост. Эти клетки удаляют инфицированные или трансформированные клетки путем распознавания антигенных фрагментов в соединении с различными молекулами (названных МНС молекулами класса 1) на инфицированных или трансформированных клетках. ЦТЛ могут индуцироваться экспериментально путем цитоплазматической нагрузки определенными растворимыми антигенами в специфических клетках. Иммунизация одним растворимым антигеном обычно недостаточна для специфической индукции цитотоксических Т-лимфоцитов.
Один из методов, при помощи которого может индуцироваться ответ ЦТЛ, включает применение методик рекомбинантной инженерии для внедрения определяющих компонентов рассматриваемого антигена в геном слабого инфекционного агента. Целью такой стратегии является создание антиген-специфических ответов цитотоксических Т-лимфоцитов на желаемый эпитоп, путем вызова у хозяина легкой, самоограничивающей инфекции. Химерные векторы были описаны при использовании вирусов коровьей оспы, полно-, адено- и ретровирусов, так же, как и бактерий, таких как Listeria и БЦЖ. Например, Takahashi et al., 85 Proc. Natl. Acad. Sci., USA, 1988, описывает рекокмбинантый вакцинный вирус, экспрессирующий оболочечный ген HIV др. 160, в качестве потенциального средства для индукции цитотоксических Т-лимфоцитов.
Второй метод, с помощью которого может индуцироваться клеточно опосредованный ответ, включает использование адъювантов. Поскольку эта область, как видно, изобилует спорами по применению адъювантов, неясно индуцировался ли клеточно опосредованный иммунитет при таком способе, и включал ли такой клеточно опосредованный иммунитет ответ цитотоксических Т-лимфоцитов. Следующие, однако, являются характерными публикациями в этой области знаний.
Stover et al., 351 Nature 456, 1991 (не признан прототипом по отношению к настоящей заявке) описывает ответ ЦТЛ на батагалактозидазу при использовании рекомбинантной БЦЖ, содержащей ген бетагалактозидазы. Никакого подобного ответа не обнаружено при использовании неполного адъюванта Фрейнда и бета-галактозидазы.
Mitchell et al. , 8 J. Clinical Oncology 856, 1990 (который не признан прототипом для настоящего изобретения) описывает лечение больных с метастатической меланомой адъювантом, названным "ДЕТОКС" и лизатами аллогенной меланомы, вводимыми пять раз за шестинедельный период. У небольшой части больных наблюдалось увеличение числа цитологических Т-клеток. Авторы описывают необходимость повышения уровня продукции цитотоксических Т-лимфоцитов и предлагают комбинированную терапию адъювантом с интерлейкином-2 также, как и предварительное лечение циклофосфамидом, чтобы снизить уровень специфических для опухоли Т-супрессорных клеток, которые могут присутствовать. ДЕТОКС включает детоксифицированный эндотоксин (монофосфорильный липид A) из Salmonella minnesota каркасы клеточных стенок Mycobactirium phlei скваленовое масло и эмульгатор.
Allison и Gregoriadis, 11 Immunlogy Today 427, 1990 (который не признан прототипом для настоящего изобретения) отмечает, что единственным адъювантом "разрешенным для применения" в человеческих вакцинах являются соли алюминия (алюм), которые не вызывают стойкого клеточно опосредованного иммунитета.
Allison и Gregoriadis утверждают, что "существует потребность в разработке адъювантов с эффективностью полного адъюванта Фрейнда, но без его различных побочных эффектов, таких, как гранулемы". Они далее утверждают, что существует три возможных стратегии, например, использование липосом; использование адъювантов, называемых имуностимлуирующими комплексами (ISCOM, которые включают сапонин или Quil A (тритерпеноид с двумя углеводными цепями), холестерол и фосфатидилхолин), которые разрешены для использования в противогриппозной вакцине для лошадей (Morein et al., Immunological Adjuvants and Vaccines, Plenum Press, 153); и использование эмульсии (SAF) сквалена или скважина (с или без плюрониевого агента) и мурамил-дипептида (MDP), SAF, как сказано, вызывает клеточно опосредованный иммунитет у мышей, хотя "долго считали, что субъеденичные антигены не могут вызвать ответов цитотоксических Т-лимфоцитов (ЦТЛ); Takahashi et al., 344 Nature 873, 1990, описывают индукцию ограниченных классом II хелперов и цитотоксических Т-лимфоцитов путем использования ISOM при единственной подкожной иммунизации у мышей. Они сообщают, что адъювант Фрейнда, неполный адъювант Фрейнда и физиологический раствор с фосфатным буфером не индуцируют активность цитотоксических Т-лимфоцитов против мишеней, в которых они заинтересованы. Они заявляют, что в противоположность результатам, полученным с другими формами экзогенных растворимых белковых антигенов, они показали, что возможно примировать антиген-специфические МНС классом 1 ограниченные, C 8+ 4- ЦТЛ путем имунизации экзогенным интактным белком при использовании ISCOM. Они также сообщают, что описанные эксперименты наводят на мысль, сто возможно вызвать продукцию человеческих ЦТЛ путем использования белков HIV, содержащих ISCOM, и что вакцины на основе ISCOM могут достичь давно желанной цели индукции как ЦТЛ, так и антител с помощью очищенного белка.
Byars и Allison, 5 Vaccines 223, 1987 описывают использование SAF-1, который включает ТВИН 80, PLURONIC L12 и скален или сквален с мурамил-дипептидом или без него, и предполагают, что их данные показывают, что рецептура с мурамил-дипептидом будет пригодна для ветеринарных вакцин и вакцин для людей. Вторичные инъекции адъюванта давали без мурамил-дипептида. Мурамил-дипептид, как сказано, значительно увеличивает продукцию антител по сравнению с использованием одъюванта без мурамил-дипептида. Клеточно опосредованный иммунитет определялся как гиперчувствительность замедленного типа путем кожных проб, чтобы определить индукцию Т-хелперных клеток. Такая гиперчувствительновсть была сильнее и более длительна, когда адъювант снабжался мурамил-дипептидом. Аналогичные адъюванты описаны Allison et al., патент США 4770874 (где сообщается, что комбинация мурамил-дипептида и плюрониевого полиола необходима, чтобы вызвать сильный клеточно опосредованный или гуморальный ответ на яичный альбумин); Allison и др., патент США 4772466; Murphy - Cord и др., 246 Science 1293, 1989 (где сообщается, что использование комбинированных адъювантов с мурамил-дипептидом могло усиливать индукцию как гуморальной, так и клеточной ветвей иммунного ответа); Allison and Byars, 87 Vaccines 56, 1987 (где заявляется, что клеточно опосредованный иммунитет вызывается SAF (с мурамил-дипептидом), как показано по гиперчувствильности замедленного типа, пролиферативным ответам Т-клеток на антиген, по продукции интерлейкина-2 и по специфическому генетически ограниченному лизису мишеневых клеток, несущих иммунизирующий антиген); Allison and Byars, Immunophamacology of Infectious Diseases; Vaccine Adjuvants and Modulators of Non-Specific Resistance 191 - 201, 1987; Morgan et al., 29, J. Medical Virology 74, 1989; Kenney et al., 121 J. Immunological Methods 157, 1989; Allison and Byars, 95 J. Immunological Methods 157, 1986 (где показано, что соли алюминия и эмульсии минерального масла увеличивают образование антител, но не усиливает клеточно опосредованный иммунитет; и рецептуры с мурамил-дипептидом, как показано, вызывают клеточно опосредованный иммунитет); Byars et al. , 8 Vaccine 49, 1990 (не признано прототипом для настоящей заявки), где заявляется, что их рецептура адъюванта заметно увеличивает гуморальные ответы и, в меньшей степени, усиливает клеточно опосредованные ответы на гемаглютинин гриппа);
Allison and Byars, 28 Molecular Immunology 279, 1991 (не признана прототипом для настоящей заявки; которая сообщает, что функция мурамил-дипептида состоит в индукции экспрессии цитокинов и усилении экспрессии основных генов гистосовместимости (МНС); и что получены лучшие антительyые и клеточные ответы, чем с другими адъювантами, и что предполагают установить эффективна ли подобная стратегия у людей); Allison and Byars, Technology Advances in Vaccine Development 401, 1988 (которая описывает клеточно опосредованный иммунитет при использовании SAF); Epstein et al., 4 Advance Drug Delivery Reviews 223, 1990 (которая представляет обзор по различным адъювантам, использованным при получении вакцин); Alison and Byars, 95 J. Immolological Methods 157, 1986 (которая устанавливает, что добавление мурамил-дипептида к адъюванту заметно усиливает клеточно опосредованные ответы на ряд антигенов, включая моноклональные иммуноглобулины и вирусные антигены); и Morgan et al., 29 J. Medical Virology 74, 1980 (которая описывает использование SAF-1 для приготовления вакцины из вирусов Эпштейна-Барра).
Kwak et al., Idiotipe Networks in Biology and Medicine, Elsevier Science Publishers, p. 163, 1990 (не признана прототипом настоящей заявки) описывает использование SAF без мурамил-дипептида в качестве адъюванта для идиотипа B-клеточной лимфомы у людей. В частности, эмульсия плюрониевого L 121, скалана и 0.4% ТВИН-80 в физиологическом растворе с фосфатным буфером вводилась вместе с идиотипом. Они сообщают, что "добавление адъюванта должно, к тому же, усиливать гуморальные ответы и может также облегчить индукцию клеточных ответов.
Другие иммунологические препараты включают липосомы (Allison et al., патенты США 4053585 и 4117113); циклические пептиды (Dreesman et al., патент США 4778784); полный адъювант Фрейнда (Asherson et al., 22 Immunology 465, 1972; Berman et al., 2 International J. Canser 539, 1967; Allicon. 18 Immunopotentiation 73, 1973; и Allison, Non-Specific Factors Influencing Host Resistance 247, 1973); ISCOM (Letvin et al., 87 Vaccines 209, 1987); адъюванты, содержащие неионные блокирующие полимерные агенты, соединенные с минеральным маслом, поверхностно активным агентом и ТВИН-80 (Hunter and Bennett, 133 J. Immunology 3167, 1984; и Hunter etl al., 127 J. Immunology 1244, 1981); адъюванты, составленные из минерального масла и эмульгирующего средства с убитыми микобактериями или без них (Sancher - Percador et al., 141 J. Immunology 1720, 1988); и другие адъювант, такие как липофильное производное мурамил-трипептида и мурамил-дипептид, ковалентно связанный с рекомбинантным белком (id).
Кратное изложение изобретения
Заявитель открыл безопасный и обладающий рядом преимуществ способ и композиции, с помощью которых ответы ЦТЛ могут индуцироваться у людей и домашних или сельскохозяйственных животных. Способ включает использование антигенной рецептуры, которая обладает слабой токсичностью для животных или вообще ее не имеет, и в которой отсутствует иммуностимулирующий пептид (например, мурамил-дипептид), присутствие которого снижало бы желаемый клеточный ответ. Кроме того, методология проста для использования и не требует объемной работы, для изменения существующих клеток при помощи методик рекомбинантной ДНК, чтобы сделать их иммуногенными. Это открытие удивительно, так как не ожидалось, что подобные ответ ЦТЛ мог бы вызываться при применении такой антигенной рецептуры с отсутствием иммуностимулирующих пептидов или их эквивалента. Данные, полученные заявителем, дают возможность использования таких антигенных рецептур при широком спектре болезненных состояний или в качестве профилактических средств. Например, введение таких антигенных рецептур может использоваться при лечении вирусных заболеваний, при которых важен ответ ЦТЛ, например при лечении HIV инфекции или группа; оно может быть также расширено, чтобы использоваться при лечении бактериальных инфекций, рака, паразитарных инфекций и тому подобного. В качестве профилактического средства антигенная рецептура с подходящим антигеном применена для предупреждения инфекции, вызываемой вирусами, ответственными за вышеупомянутые вирусные заболевания, особенно для профилактики HIV инфекции, а также для профилактики у больных с риском рака, например, после резекции первичной опухоли.
Таким образом, в первом аспекте, изобретение показывает способ индуцирования ответа ЦТЛ у людей или домашних (например, кошки или собаки) или важных для сельского хозяйства животных (например, лошади, коровы или свиньи) на антигены, кроме антигена B-клеточной лимфомы или яичного альбумина. Способ включает стадии получения антигена, по отношению к которому желателен ответ ЦТЛ и получение нетоксичной антигенной рецептуры (формы), которая включает, состоит или по существу состоит из стабилизирующего детергента, образующего мицеллы средства и биодеградируемого и биосовместного масла. В этой антигенной рецептуре предпочтительно отсутствует какой-либо иммуностимулирующий пептидный компонент или имеются достаточно низкие уровни такого компонента, которые не снижают желаемый клеточный ответ. Эта рецептура предпочтительно представляют собой стабильную эмульсию масло-в-воде. То есть каждый из различных компонентов выбирается так, чтобы эмульсия оставалась в эмульгированном состоянии в течение периода по крайней мере одного месяца и, предпочтительно, в течение более чем одного года без разделения фаз. При этом способе антиген и антигенная рецептура смешиваются вместе для образования смеси (предпочтительно путем микропсевдоожижения), и эта смесь вводится животному в количестве достаточном, чтобы индуцировать ответ ЦТЛ в животного. Необходимо лишь одно такое введение.
Под "стабилизирующим детергентом" подразумевается детергент, который позволяет компонентам эмульсии сохраняться в виде стабильной эмульсии. Такие детергенты включают полисорбат, 80 (ТВИН) (сорбитан-моно-9-октадеценоат-поли-(окси-1,2-этандиил); производимый ICI Americas, Wilmington, DE), ТВИН 40, ТВИН 20, ТВИН 60, Zwittergent 3-12, TEEPOL HB7, и SPAN 85. Эти детергенты даются обычно в количестве примерно 0.05 до 0.5%, предпочтительно, в количестве примерно 0.2%.
Под "образующим мицеллы веществом" подразумевается вещество, способное стабилизировать эмульсию, образованную с другими компонентами, так что образуется мицеллоподобная структура. Такие средства предпочтительно вызывают некоторое раздражение в месте инъекции, чтобы побудить макрофаги вызвать клеточный ответ. Примеры таких средств включают полимерные сурфактанты, описанные в BASF Wyandotte publications, например, Schmolka, 54 J. Am. Oil. Chem. Soc. 110, 1977, и Hanter et al., 129 J. Immunol 1244, 1981, обе включены здесь в виде ссылки, PLURONIC L 62 LF, L 101, и L 64, L 121, PEG 1000, и TETRONIC 1501, 150R1, 701, 901, 1301, и 130R1. Химические структуры таких агентов хорошо известны в данной области. Предпочтительно агент выбирается так, чтобы иметь гидрофильоно-липофильное соотношение (HLB) между 0 и 2, как определено у Hunter И Bennet, 133 Journal of Immunology 3167, 1984. Вещество предпочтительно дается в количестве между 0.001 и 10%, наиболее предпочтительно в количестве между 0.001 и 5%.
Масло выбирается так, чтобы способствовать сохранению антигена в эмульсии масло-в-воде, то есть обеспечивать носитель для желаемого антигена, и предпочтительно, имеет температуру плавления меньше, чем 65oC, такую, что эмульсия образуется или при комнатной температуре (примерно от 20 до 25oC), или как только температура эмульсии снижается до комнатной температуры. Примеры таких масел включают сквален, сквалан, EICOSANE, тетратетраконтан, глицерол и арахисовое масло или другие растительные масла. Масло предпочтительно дается в количестве между 1 и 10%, наиболее предпочтительно между 2.5 и 5%. Важно, чтобы масло было биодеградируемым и биосовместимым так, чтобы организм мог разрушить масло через некоторое время и так, чтобы не появлялось никаких побочных эффектов, таких, как гранулемы, при использовании масла.
Важно, чтобы в вышеупомянутой рецептуре пептидный компонент, в частности, мурамил-дипептид (MDP) отсутствовал. Такой пептид будет влиять на индукцию ответа ЦТЛ, если он дается в количестве больше, чем примерно 20 микрограмм на нормальную человеческую дозу введения препарата. Предпочтительно, чтобы такие пептиды полностью отсутствовали в антигенном препарате, несмотря на явную стимуляцию ими гуморального отдела иммунной системы. То есть заявитель обнаружил, что, хотя такие пептиды могут усиливать гуморальный ответ, они неблагоприятны, когда желателен ответ цитотоксических Т-лимфоцитов.
В других связанных аспектах, антигенная рецептура (препарат) создается из только двух из вышеназванных трех компонентов и используется с любым желаемым антигеном (термин, который включает белки, полипептиды и их фрагменты, которые являются иммуногенными), за исключением яичного альбумина (или других альбуминов, например, HSA, BSA и овальбумин), чтобы индуцировать ответ ЦТЛ у вышеназванных животных или людей.
Заявитель полагает, что вышеприведенные препараты обладают значительными преимуществами над предшествующими препаратами (включая ISCOMS, DETOX и SAF) для применения у людей. В отличие от таких препаратов, настоящий препарат включает образующее мицеллы вещество и, в то же время, не имеет в составе пептидов, остовов клеточных стенок или компонентов бактериальных клеток. Настоящий препарат также индуцирует ответ ЦТЛ, который или не наблюдался при применении предшествующих препаратов, или значительно выше по сравнению с этими препаратами.
Под "нетоксичный" подразумевается, что наблюдается небольшое побочное действие или его отсутствие у лечившегося животного или человека. Специалисты в области медицины или ветеринарии поймут, что этот термин имеет широкое значение. Например, по существу здорового животного или человека может допускаться только легкая токсичность, тогда как у человека, страдающего заболеванием в терминальной стадии (с предполагаемой продолжительностью жизни меньше, чем, примерно, три года) может допускаться существенно большая токсичность.
В предпочтительных примерах осуществления антигенный препарат состоит по существу из двух из трех этих детергента, вещества и масла; способ состоит по существу в единственном введении смеси (антиген плюс антигенная рецептура) человеку или животному; человек или животное инфицированы вирусом и испытывают один или более симптомов (которые в основном определяются врачами - специалистами в соответствующей области) инфекции, вызванной вирусом; и антигенный препарат является нетоксичным для людей и животных.
В других предпочтительных примерах осуществления антиген выбирается из HIV антигенов: gp 160, gag, pol, Nef, Tat и Rev; малярных антигенов; белка CS и спорозонтного поверхностного белка 2; поверхностных антигенов гепатита B: Pre - S1, Pre - S2, HBc Ag, и HBc Ag; антигенов вируса гриппа: HA, NP и NA; поверхностных антигенов гепатита A; антигенов вируса герпеса: EBV gp 340, EBV gp 85, HSV gB, HSV gD, HSV gH, раннего белкового продукта HSV, цитомегаловирусного gB, цитомегаловирусного gH и IE белка gP72; антигенов респираторно-синцитиального вируса: белка F, белка G и белка N; и опухолевых антигенов CEA карциномы, карциномного муцина, P21 карциномы, P53 карциномы, MPG меланомы, p97 меланомы и онкогенного продукта Neu карциномы, продукта гена p53 карциномы, антигена меланомы, называемого MAGE, и мутированного белка p21 ras, присутствующего в ряде злокачественных опухолей.
В смежном аспекте изобретение описывает композицию, включающую, состоящую или, по существу, состоящую из антигена, смешанного с антигенной рецептурой, описанной выше, и антиген выбирается из этих антигенных частей, перечисленных выше.
В других смежных аспектах изобретение описывает способы лечения больного, инфицированного вирусом HIV, страдающих малярией, гриппом, гепатитом, больных раком, инфицированных вирусом герпеса, инфицированных респираторным синцитиальным вирусом, путем введения композиции, включающей соответствующий антиген (например, выбранный из перечисленных выше), смешанный с одной из вышеназванных антигенных рецептур.
Другие характерные черты и преимущества изобретения будут очевидны из последующего описания предпочтительных его воплощений и из формулы изобретения.
Описание предпочтительных примеров осуществления
Фиг. 1A и 1C являются графическим представлением данных сравнения индукции ЦТЛ различными овальбуминовыми препаратами; E : T представляет отношение клеток эффекторов к клеткам-мишеням на всех фигурах.
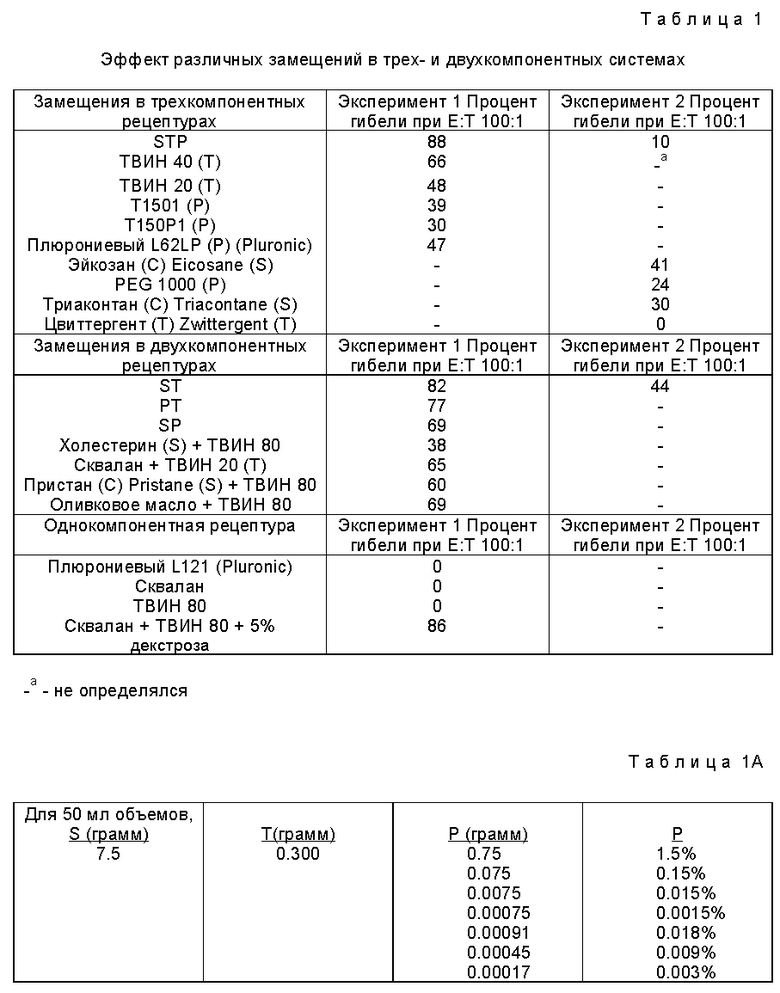
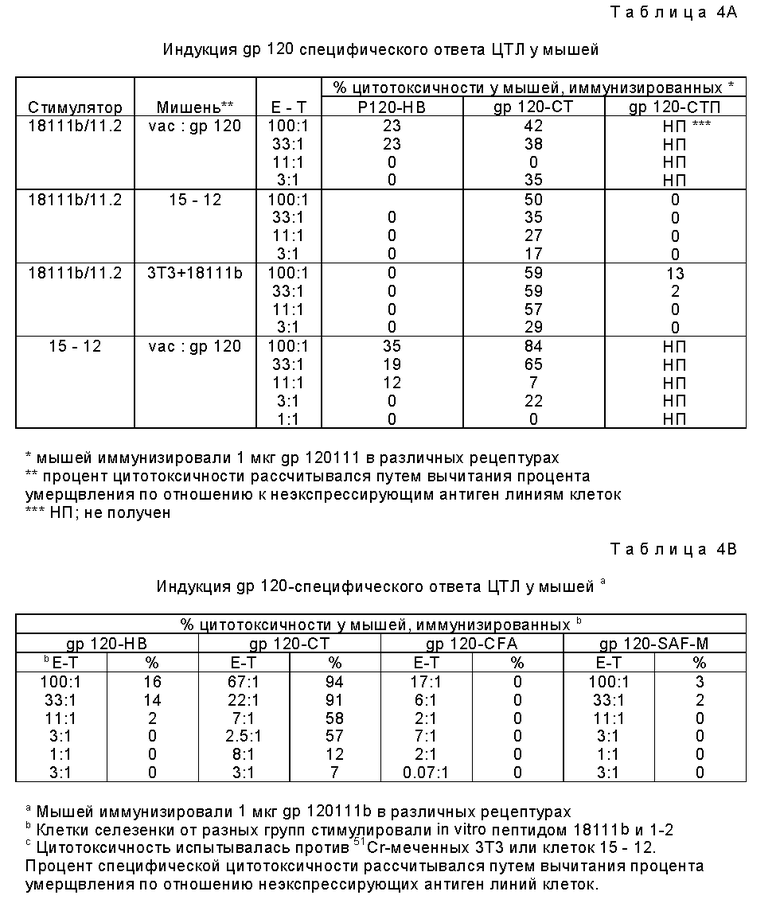
Фиг. 2A и 2B являются графическим представлением данных сравнения индукции ЦТЛ различными β-алактозидазными препаратами.
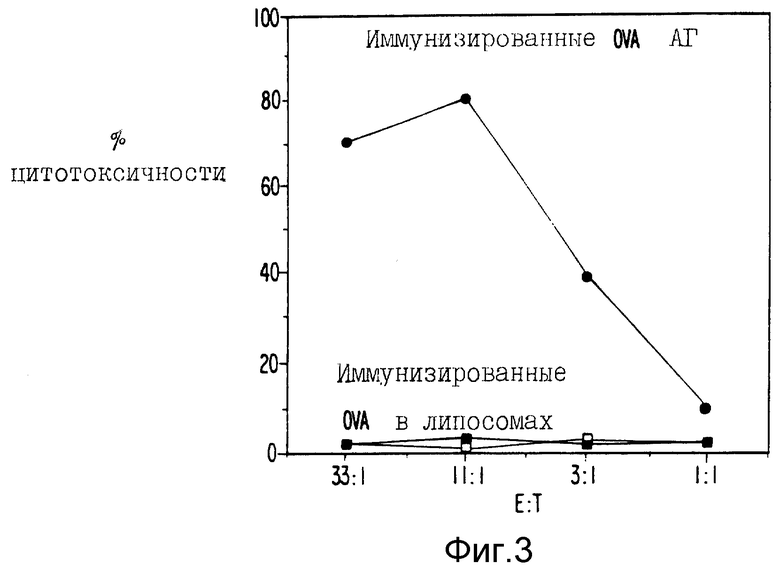
Фиг. 3 являются графическим представлением данных сравнения индукции ЦТЛ овальбумином в липосомах и в антигенном препарате.
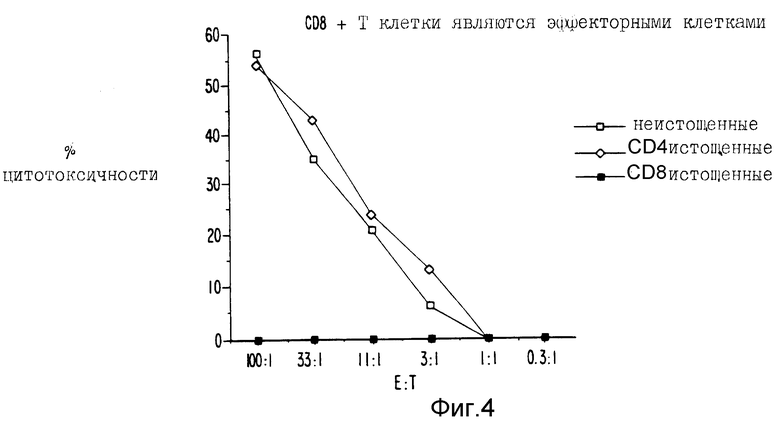
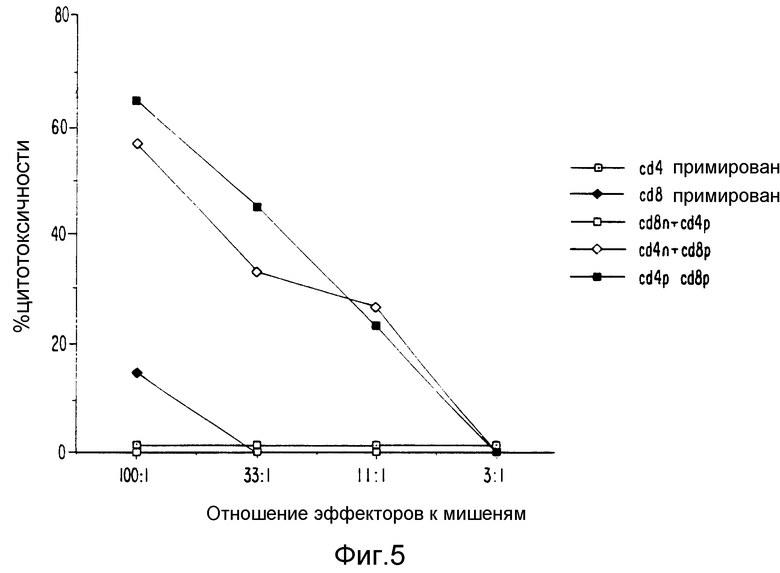
Фиг. 4 и 5 являются графическим представлением данных, показывающих действие CD4 и CD8 клеточного истощения на индукцию ЦТЛ.
Фиг. 6 являются графическим представлением данных, показывающих индукцию ЦТЛ смесью плюронцевого агента и ТВИН и антигена.
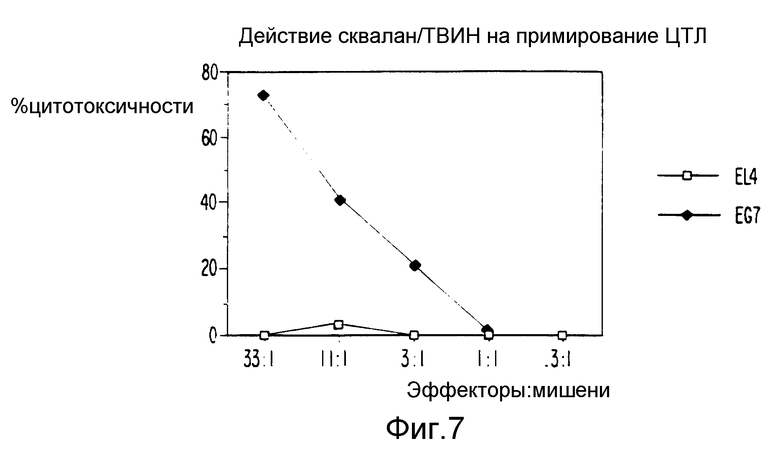
Фиг. 7 являются графическим представлением данных, показывающих индукцию ЦТЛ смесью сквалана и ТВИН и антигена.
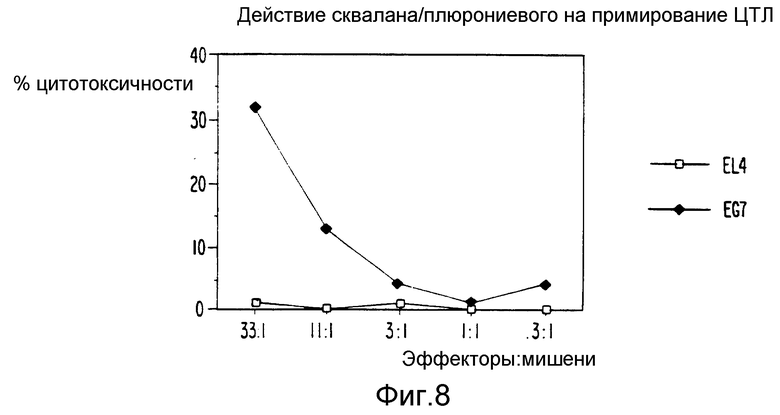
Фиг. 8 являются графическим представлением данных, показывающих индукцию ЦТЛ смесью скавалана и плюрониевого агента и антигена.
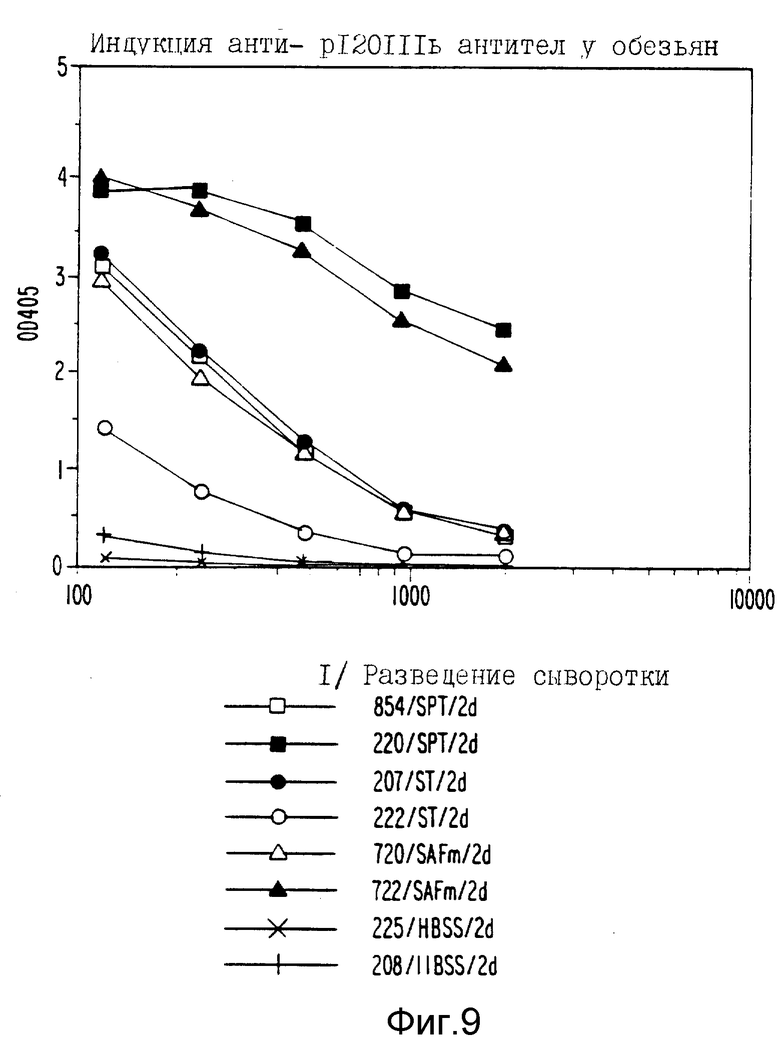
Фиг. 9 являются графическим представлением индукции анти- gp 120111b антител у обезьян при помощи различных антигенных рецептур; и
Фиг. 10A - 10B являются графическим представлением данных сравнения gp 120-специфического ответа ЦТЛ у обезьян, иммунизированных вирусом коровьей оспы - gp 120 и gp 120-AF.
Антигенная рецептура
Антигенные рецептуры, применяемые в этом изобретении, в основном описаны выше. Специалисты в этой области поймут, что легко приготовить эквивалентные рецептуры и можно ожидать, то они будут иметь эквивалентные свойства в индукции ответа ЦТЛ. Такие рецептуры легко испытываются на их свойства с использованием методик, эквивалентных описанным в примерах ниже.
Далее следуют примеры изобретения с использованием антигенной рецептуры (AF), составленной из примерно 15% сквалана (0.6 ТВИН 80) и (0.0045 - 3.75% плюрониевого) в фосфатно-буферном физрастворе (Imed STP). В частности, эмульсия AF включала: 150 мг сквалана, 0.045 - 37.5 мг полоксамера 401 (PLURONIC L 121), 6 мг полисорбата 80 (TWEEN 80), 0.184 мг хлорида калия, 0.552 мг моноосновного фосфата калия, 7.36 мг хлорида натрия, 3.3 мг диосновного фосфата натрия (безводного) на 1 мл воды, pH 7.4. Эта эмульсия микропсевдоожижалась с использованием стандартной методики (Microfluidies Model M110F) в модуле обратного давления при 11 - 14000 psi (фунтов на квадратный дюйм) с постепенным возвратом к атмосферному давлению, с охлаждением и сохранением в тающем льду.
В других примерах антиген смешивался с мкропсевдоожиженной смесью сквалана (S), плюрониевого (P) и ТВИНа 80 (Т), чтобы достичь конечной концентрации в 5% сквалана, 0.2% ТВИН 80 и 0.0015 - 1.25% плюрониевого, соответственно. Чтобы определить суб-компоненты, необходимые для индукции специфического иммунного ответа, сквален-ТВИН 80, плюрониевый-ТВИН 80 или сквалан-плюрониевый готовили в тех же самых концентрациях, что и для трехкомпонентной смеси. Плюрониевый, сквалан или ТВИН 80 готовили также отдельно, чтобы определить действие отдельного компонента на индукцию ЦТЛ. Производились также замещения ТВИН 20, ТВИН 40 или Zwittergent вместо ТВИНИ 80, чтобы определить действие различных производных ТВИН на индукцию ЦТЛ в ova системе. Замены Сквалана на Eicosone или Triacontone в трехкомпонентной рецептуре и замены вместо плюрониевого сополимера на PEG 1000, Pleuronic L 62 LF и Tetronocs 1501 и 150 RL также производились в той же самой трехкомпонентной рецептуре. В качестве двухкомпонентных рецептур, смешивали различные аналоги в разных сочетаниях и испытывали на ova специфическую индукцию ЦТЛ. Ими были смесь холестерола - ТВИН 80, Сквален - ТВИН 20, Pristane - ТВИН 80 или оливковое масло - ТВИН 80. Для изучения стабилизации смесь Сквалана - ТВИН 80 после микропсевдоожижения смешивали с декстрозой до конечной концентрации в 5%. Во всех случаях комбинации наполнителей смешивали в приборе для микропсевдоожижения для получения стабильной эмульсии. В некоторых экспериментах двухкомпонентные рецептуры смешивали с разными концентрациями MDP для индукции гуморального ответа и ответа ЦТЛ. Таблица 1 представляет полный список различных рецептур, использованных при этом исследовании (см. в конце описания).
В качестве контроля адъюванта использовалась синтексная рецептура адъюванта (микропсевдооожиженная; SAFm) и состояла из двух частей. Часть I состоит из фосфатно-буферного физраствора, содержащего конечную концентрацию сквалана в 5%, 1.25% плюрониевого и 0.2% ТВИН 80 (носителя или 1-SAF). Часть II состоит из N-ацетилмурамил-L-треонил-D-изоглютамина (Thr - MDP), производного компонента клеточной стенки микобактерий. Для целей иммунизации антиген смешивают с микропсевдоожиженным носителем (часть I), чтобы получить гомогенную эмульсию. MDP добавляется, чтобы приготовить SAFm и недолго интенсивно перемешивается. Концентрацию MDP в смеси изменяли, чтобы определить существует ли оптимум концентрации для индукции ЦТЛ. В качестве контроля для адъюванта мышей иммунизировали растворимыми антигенами, смешанными с алюминиевыми солями в соответствии с руководством производителя (Pierce Chemical, Rockford, IL) или с полным адъювантом Фрейнда (CFA).
Антигенная рецептура STP используется для индукции ответов цитотоксических лейкоцитов у мышей. Специалистам будет ясно, что такая мышиная модель является показателем того, что в эквивалентных экспериментах или при лечении будут подобным же образом вызываться ответы цитотоксических Т-лимфоцитов у людей, домашних животных и сельскохозяйственных животных. Количество антигенной рецептуры и антигена, пригодное для того, чтобы вызвать желаемый клеточный ответ, может быть определено эмпирически стандартными методами, хорошо известными специалистам в этой области, без ненужного экспериментирования. Таким образом, если желательно минимизировать побочные эффекты лечения такой смесью, специалисты могут определить минимальный уровень такой смеси, необходимый для введения человеку, домашнему или сельскохозяйственному животному, чтобы вызвать ответ ЦТЛ и тем самым вызвать иммунитет к желаемому антигену. При нормальном использовании такая смесь будет вводиться с помощью любой из ряда стандартных процедур, но особенно предпочтительна внутримышечная инъекция в месте, которое позволит эмульсии оставаться в стабильной форме в течение периода в несколько дней или несколько недель.
Следующие материалы и методы использовались в примерах, представленных ниже, если нет других указаний:
Мыши
Мыши-самки C57BL/6 (H-2b) и BALB/c (H-2) были куплены в Harlen Sprague (Can Diego, California).
Антигены
Овальбумин (ova, сорт Vii; Sigma Chimical Co., St. Louis, MO) использовался в нативной форме. β-галактозидаза ( β-gal, сорт VIII; BRL) использовалась в нативной форме и после кипячения в 1 М NaOH в течение 2 мин, чтобы получить щелочной перевар. Рекомбинантный gp120 был куплен у American Biotechnology.
Опухолевые клетки и трансфектанты
Использовали опухолевые клетки 1a линий EL4 (C57BL/6, H-2b тимома) и P815 (DBA/2, H-2d мастоцитома). Получение ovа-продуцирующего трансфектанта EL4, EG7-ova, описано ранее Moore et al, 54 CeJJ 777, 1988. Трансфектант, продуцирующий β-gel, P13.1, был получен путем электропорации 107 P815 клеток в 1 мл фосфатно-буферного физраствора (PBS) с 10 мг Pst I нормализованном pCH110 (Pharmacia LKB Biotechnology Inc. , Pisсataway., NJ) и 1 мг Pvu I нормализованном pSV 2 neo (Southern et al., 1 J. Mol. Appl. Genet 327, 1982) с последующей селекцией в среде с 400 мкг/мл антибиотика G418. Трансфетакт C3-4 получали из BALB/c гибридомы Igm 662 путем трансфекции плазмидой, кодирующей ген β-gal лигированные с третьим и четвертым экзоном тяжелой цепи Igm (Rammensee et al. , 30 Immunogenetics 296, 1989). Экспрессирующие p160111b фибробласты 3Т3, 15 - 12 были предоставлены доктором Germain из NIH (Bethesda, MD). Kb трансфицированная линия L-клеток была предоставлена доктором Carbone, Monash University, Australia. Db и Ld трансфицированные линии L-клеток были предоставлены доктором Ted Hensen, Washington University, St. Louis.
Иммунизация
Мышей иммунизировали внутривенно 200 мкл суспензии 25 • 106 спленоцитов после цитоплазматической нагрузки, как описано Moore et al., выше, и Carbone et al, J. Exp. Med. 169:603, 1989). Для иммунизации ova-антигенной рецептурой или β-gal-антигенной рецептурой, 30 мкг каждого белкового антигена на мышь вводилось в подушечку лапы и основание хвоста, подкожно. Каждая инъекция содержала 67 мкл микропседоожиженной антигенной рецептуры (изготовленной по стандартным процедурам) и 30 мкг белкового антигена в конечном объеме 200 мкл. До конечного объема доводили с помощью HBSS, смотрите Whittaker manual (Welkersville, MD). MDP давали в концентрациях между 0 и 300 мкг. Где указано, мышей иммунизировали растворимыми антигенами в CFA или в алюм в общем объеме 200 мкл.
Стимуляция in vitro популяций эффекторов
Клетки селезенки (30 • 106) от нормальных или иммунизированных мышей, которые были примированы по крайней мере 14 дней ранее, инкубировали с 1.5 • 106 EG7-ova (облученных 20 000 рад) для получения ответов на ova, или с 1.5 • 106 клеток С3-4 (облученных 20 000 рад) для получения ответа на β-gal в 24 ячеистых платах при 37oC в атмосфере с 7% CO2 воздухе. Все культуры тканей получали в полной среде, состоящей из среды IMDM, смотрите Whittaker Manual (Welkersville, MD), дополненной 10% плодной телячьей сыворотки (FCS), 2 мМ глютамина, гентамицинов и 2 • 10-5 М 2-меркаптоэтанола. Для экспериментов истощения in vitro примированные in vivo или стимулированные in vitro клетки селезенки обрабатывали моноклональными антителами (mAbs) RL. 172 (анти-CD 4) или mAbs 3.168 (анти-CD8) для удаления CD4+ или CD8+T-клеток (Sarmiento et al., 125 J. Immunol. 2665, 1980, и Ceredig et al., 314 Nature 98, 1985). mAb RL, 172 и mAb 3.168 были получены от Dr. Jonathan Sprent из Scripps Clinic and Research Foundation. La Jolla, CA.
Клетки селезенки (230 • 106) от нормальных и иммунизированных мышей были примированы по крайней мере за 21 день ранее и инкубировались с 1.5 • 106 клеток 15 - 12 (обработанных 200 мкг митомицина C в течение 45 минут на 108 клеток) или с 500 мкг пептида 18111b, содержащего доминантный эпитоп ЦТЛ Balb/c мышей в полной среде IMDM (Irvine Scientific, Santa Ana, CA), содержащей 10% заранее отобранной FCS (ICN Flow; ICN Biochemicals, Inc., Costa Mesa, CA), 2 мМ глютамина, гентамицин и 2 • 10-5 М 2-меркаптоэтанола. Для стимуляции пептидами in vitro клетки селезенки культивировали в полной IMDM, содержащей 5% ConA супернатанта.
Для экспериментов истощения примированные in vivo или стимулированные in vitro клетки селезенки обрабатывали mAbs RL.172 (анти-CD4) или mAbs 3.168 (анти-CD8) в присутствии низкотоксичного кроличьего комплемента (Cederlane Laboratories, Ltd. , Hornby Ontario, Canada) для удаления CD4+ или CD8+ T-клеток (22, 23). mAb RL. 172 и mAb 3.168 были даром от Dr. Jonatham Sprent из Scripps Clinic and Research Founsation, La Jolla, CA.
Исследование цитотоксичности
Клетки-мишени (1 • 106) метили 10 мк Ci (51Cr) хромата натрия в течение 60 минут. Для получения пептидных импульсных мишений 50 мкл раствора пептида 1 мг/мл в HBSS добавляли во время мечения мишений 51Cr. После отмывания, 104 меченных мишеней и серийные разведения эффекторных клеток инкубировали в 200 мкл RP10 в течение 4 часов при 37oC. 100 мкл супернатанта собирали и определяли специфический лизис как: процент специфического лизиса = 100 • ((выделение при действии ЦТЛ - спонтанное выделение) / (максимальное выделение - спонтанное выделение)). Спонтанное выделение в отсутствие цитотоксических T-лимфоцитов (ЦТЛ) было < 25% максимального выделения с помощью детергента во всех экспериментах.
Определение антительных ответов у мышей и обезьян
Каждую ячейку 96-ячеечных плат с U-образным дном (Costar, Cambridge, MA) покрывали 150 нг ova или gp 120 в 50 мкл HBSS и инкубировали в течение ночи при 4oC. Для определения анти-gp120 и анти-ova антительных ответов у мышей платы блокировали 1% BSA в течение 1 час. Добавляли серийно разведенную сыворотку в объеме 25 мкл на ячеку и инкубировали в течение 2 часов. Платы отмывали и добавляли 50 мкл на ячейку разведения 1:1000 козьего противомышеного IgG, конъюгированного с HRPO (Alabama), в 1% BSA. После 1 часа инкубации платы отмывали и добавляли 100 мкл субстрата на ячейку. Через 10 - 15 минут определялась OD405 (оптическая плотность при 405 нм). Для определения анти-gp120 антительного ответа у обезьян все стадии были такими же, за исключением того, что как блокирование плат, так и разведение сыворотки выполнялись с 5% нормальной козьей сывороткой в сбалансированном солевом растворе Хэнкса.
Пептидный синтез
Синтетические пептиды, соответствующие аминокислотным последовательностям 253-276 (Перечисление последовательностей No. 1: EQLESIINFEKLTEWTSSNVMEER; где используется стандартный однобуквенный код для представления каждой аминокислоты) овальбумина (ova 253 - 276), аминокислотных последовательностей 84 - 102 основного белка миелина (МВР 84 - 102) (Перечисление последовательностей No. 2: DENPVVHFFKNIVTPRTPP), и синтетические пептиды, соответствующие аминокислотным последовательностям 308 - 322 (18111b последовательность) gp120111b собирали путем твердофазного пептидного синтеза с использованием синтезатора Applied Biosystems 430A. Аминокислоты соединялись посредством заранее образованных симметричных ангидридов, за исключением аспарагина, глютамина и аргинина, которые соединялись в виде гидроксибензотриазольных эфиров. Эффективность соединения контролировалась с помощью нингидриновой реакции по методу Kaiser et al. 34 Anal. Biochem. 595, 1970. Пептиды освобождались от носителя с помощью HF, в соответствии с процедурой "низкий-высокий", описанной Tam, et. al., 21 J. Am. Chem. Soc. 6442, 1983, и пептиды экстрагировали из смолы 10% уксусной кислотой. После лиофилизации пептиды обессоливались на Sephadex G-25-колонке, и образцы пептидов затем очищали ВЭЖХ с обращением фазы на Vydac препаративной C-18 колонке. Очищенные пептиды (98%) растворяли в HBSS при колоночной концентрации 10 мг/мл и разводили до желаемой концентрации полной средой.
Расширение CNBr
Образцы белка (например, в-галактозидазы) обрабатывали 100-кратным молярным избытком бромида циана в растворе 100 мМ трифторуксусной кислоты. Давали пройти реакции в течение 18 часов при комнатной температуре (примерно 20oC) с ротацией. После предписанного времени реакции пептидные фрагменты отделяли от реагентов, применяя аппарат SEP-PAR (C-18 (Waters), элюировали 95% ацетонитрилом и лиофилизировали.
Щелочное расщепление
Образцы белка (например, β-галактозидазы) обрабатывали 1N NaOH и кипятили в течение 2 минут, и полученные в результате пептидные фрагменты отделяли от реагентов, используя аппарат C-18 SEP-PAK (Waters), и элюировали 95% ацетонитрилом и лиофилизировали.
Пример 1
Примирование ограниченных ЦТЛ класса 1
Moore et al. , 113 UCLA Symp. Mol. Cell. Biol. 1989 и Carbone и Bevan, 171 J. Exp. Medicine 377, 1991, показали, что мыши, иммунизированные клетками селезенки, цитоплазматически нагруженными растворимым ova, примировали для ova специфического ответа ЦТЛ, ограниченных классом 1. Экспрессирующий ova трансфектант EL4, EG7-ova, использовался для стимуляции in vitro примированных in vivo селезеночных лимфоцитов и также использовали в качестве мишени для ova специфического, опосредованного ЦТЛ, разрушения клеток. Это исследование также показало, что CD8+ эффекторы, индуцированные EG7-ova трансфектантом или клетками селезенки, цитоплазматически нагруженными ova, распознают детерминанту, картированную пептидом ova 258 - 273 в контексте H-2Kb, лизируют EG7-ova и также умерщвляет клетки EL4, покрытые оva 258 - 276. Таким образом, чтобы оценить, может ли индуцироваться растворимым антигеном эндогенный метаболизм ограниченных классом 1 CD8+ Т-клеток, использовались вышеописанная система, чтобы определить можно ли использовать определенные антигенные рецептуры, чтобы включить растворимый антиген в ограниченный метаболизм класса 1.
a) ova
Мышей C57 BL/6 иммунизировали однократно различными количествами ova (30 мкг - 1 мг на мышь) с антигенной рецептурой или без нее. Мыши получали подкожную инъекцию и в основание хвоста. Клетки селезенки брали от иммунизированных мышей по крайней мере через две недели после иммунизаций и стимулировали in vitro EG-7-ova трансфектантами. Такие низкие концентрации, как 30 мкг были также эффективны, как и доза в 1 мг. Поэтому исследования ЦТЛ обычно проводились с клетками селезенки от мышей, примированных 30 мкг ova. После пяти дней культивирования in vitro с EG7-ova примирование оценивали по наличию ova специфических эффекторов, способных к лизису EG7-ova.
У мышей, которым вводили растворимый ova в HBSS в такой высокой концентрации, как 1 мг, не обнаружено доказательств примирования ЦТЛ (фиг. 1A). Однако у мышей, имуннизированных 30 мкг ova в антигенной рецептуре, описанной выше (показанной как AF на фигурах) был показан значительный специфический ответ ЦТЛ на трасфектант (фиг. 1C). Кроме того, степень умерщвления EG7-ova иммунизированным ova-AF-клетками селезенки была сравнима с таковой клетками селезенки от мышей, иммунизированных клетками селезенки, нагруженными ova (фиг. 1B).
То, что специфичность примирования ЦТЛ in vivo была антигенспецифичной, было показано по отсутствию способности клеток селезенки от иммунизированных β-галактозидазой мышей проявлять вторичный ответ ЦТЛ in vitro при стимуляции EG7-ova. Не наблюдалось никакой ova специфической индукции ЦТЛ.
b) β-галактозидаза
Сходные результаты получены при использовании другого растворимого белкового антигена, β-gal. Для исследования β-gal-специфического ответа ЦТЛ использованной мишенью был происходящий от BALB/c, экспрессирующий β-gal трансфектант C3-4. Иммунизация мышей BALB/c растворимой β-gal создавало фон ответа ЦТЛ. Поэтому для определения специфичного ответа ЦТЛ отбор проб был отсрочен на по крайней мере восемь недель до того, как лимфоциты селезенки отбирали и культивировали в течение пяти дней в присутствии облученных трансфектантов C3-4.
Фиг. 2B показывает, что 30 мкг β-галактозидазы в AF индуцировали сильный специфический ответ ЦТЛ против трансфектанта. При соотношении эффекторов-к-мешеням (E : T) 3 : 1 иммунизированные β-gal-AF мыши продемонстрировали примерно 80% специфического умерщвления C3-4. Однако только 20% умерщвления той же самой мишени достигалось с эффекторами, выделенными от мышей, иммунизированные β-gal в HBSS, при том же самом соотношении E : T (фиг. 2A). Так как ни EL4 ни P815 не экспрессируют генные продукты МНС класса H, а лизис показывает сингенную рестрикцию, эти ova и β-gal специфические эффекты относятся к ограниченным классом 1 МНС.
Чтобы продемонстрировать полезность антигенонй рецептуры, мышей иммунизировали растворимым ova, инкапсулированным в два типа липосом, одни из которых были липосомы, чувствительные к pH. Через одну неделю клетки селезенки стимулировали in vitro, как описано выше, и испытывали против меченных 51Cr EG7-ova или EL4. Фиг. 3 показывает характерный результат, демонстрирующий, что ova в липосомах не могли примировать мышей для значительной индукции ЦТЛ. Сходные результаты наблюдались, когда иммунизировали ova в алюм.
Пример 2
Распознавание эпитопа ЦТЛ
Carbone и Bevan, выше, показали, что ЦТЛ, индуцированные у мышей C57 BL/6, трансфектантом EG7-ova и цитопламатически ova-нагружеными спленоцитами, распознают клетки EL4, покрытые пептидом ova 258 - 276. Чтобы определить индуцирует ли растворимый овальбумин в AF сходные ответы ЦТЛ, клетки селезенки получали от иммунизированных мышей и стимулировали in vitro EG7-ova. Эффекторы испытывали против клеток EL4, покрытых пептидом ova 253 - 276, или контрольным пептидом, полученным из основного белка миелина (МВР 84 - 102). Результаты показывают, что ova-AF примировал ЦТЛ со специфичностью, сходной со специфичностью ЦТЛ, примированных трансфектантами или цитоплазматически нагруженным ova (фиг. 1A, 1B, 1C). Примировали EG7-ova эффекторные клетки эффективно лизировали EG7-ovа и нетрансфицированные клетки EL4, покрытые 50 мкг/108 клеток ovа пептидом, но не лизировали клетки EL 4, покрытые 50 мкг/108 клеток пептида МВР.
В β-галактозидазной системе Carbone и Bevan, выше показали, что трансфектант, экспрессирующий β-gal, и спленоциты, цитоплазматически нагруженные растворимой β-галактозидазой, индуцировали ЦТЛ, которые лизировали экспрессирующие β-gal трансфектантные и нетрансфектантные клетки P815, покрытые расщепленной щелочью β-галактозидазой. Растворимая β-галактозидада индуцирует ЦТЛ, имеющие сходную специфичность, с той, которая получалась при иммунизации с AF (фиг. 2).
Пример 3
ЦТЛ эффекторами являются CD8+ Т-клетки
То, что растворимые белковые антигены в AF индуцируют CD8+ эффекторные Т-клетки, было показано следующим образом. Спленоциты от иммунизированных мышей культивировали в течение пяти дней с облученными трансфектантами in vitro. Затем клетки собирали и истощали от CD4+ или CD8+ Т-клеток путем использования моноклональных анти-CD4 или анти-CD8 антител плюс комплемент. Истощенные популяции затем испытывали против 51Cr-EL7-ova в ova системе или 51Cr-P13.1 в β-gal системе. Данные, представленные на фиг. 4, показывают, что в ova системе истощения CD8+ Т-клеточной популяции устраняло цитолитическую активность, приданную целой популяции эффекторных клеток. Однако истощение CD4+ Т-клеточной популяции не имело какого-либо эффекта на лизис EG-7-ova.
Подобным же образом в β-gal системе истощение CD8+ Т клеток устраняло цитолитическую активность клеток селезенки, иммунизированных β-gal-антигенной рецептурой (данные не представлены).
Пример 4
Растворимый ova в AF примирует CD8+ Т-клеток
Чтобы продемонстрировать, что ova-AF примирует CD8+ Т клеточные популяции in vivo и является определяющим для вторичного ответа in vitro, популяции CD4+ или CD8+ истощались в селезенках от мышей, иммунизированных ova-AF или от простых мышей. Эти обработанные популяции затем стимулировали in vitro AG7-ova один или в комбинации CD4+ и CD8+ Т-клеток от мышей, иммунизированных ova-AF, или в разных сочетаниях CD4+ или CD8+ Т-клеток от мышей, иммунизированных ova-AF, с CD4+ или CD8+ клетками от простых мышей. Фиг. 5 показывает, что примированные CD8+ клетки являются существенными для проявления вторичного ответа ЦТЛ in vitro. Эти данные также показывают, что для эффективного вторичного ответа ЦТЛ in vitro необходимы CD4+ Т-клетки. CD4+ клетки не нужны для примирования.
Вышеприведенные примеры показывают действие антигенной рецептуры на индукцию ответов, ограниченных классом 1 ЦТЛ против растворимых белковых антигенов. Опосредованное антигенной рецептурой примирование ЦТЛ, индуцированное растворимым антигеном, и по активности было сходно с таковым, индуцированным трансфектантами и спленоцитами цитоплазматически нагруженными расторимыми ova или β-gal. В овальбуминовой системе EG7-ova, цитоплазматически нагруженные ova спленоциты и ova-AF индуцировали:
(a) ограниченные классом 1 CD8+ ЦТЛ;
(b) ЦТЛ, которые распознают мишени, сенсибилизированные ova 253 - 276 синтетическим пептидом; и
(c) должно живущие ЦТЛ после только одной иммунизации.
В β-галактозидазной системе β-gal-AF индуцировал ЦТЛ, которые распознают трансфектант C3-4, экспрессирующий β-gal, а также нетрансфицированные клетки P815, сенсибилизированные, расщепленной щелочью β-gal. Это аналогично тому, что наблюдалось с ЦТЛ, индуцированными иммунизацией клетками селезенки, цитоплазматически нагруженными β-галактозидазой. Индукция ova-специфических ЦТЛ антигенной рецептурой является уникальной, потому что ни ova, инкапсулированный в чувствительных к pH липосомах, ни в алюм (данные не показаны), не могли индуцировать примирование ЦТЛ in vivo.
Эти примеры показывают, что антигенная рецептура, использованная выше, и ее эквиваленты применимы для лечения людей и в разработке вакцин для индукции ЦТЛ при различных видах рака и вирусных заболеваний.
Пример 5
Этот пример является специфическим примером для демонстрации использования вышеописанной AF при получении примирования ограниченных классом 1 ЦТЛ при помощи растворимого gp120 из HIV.
Экспрессирующая gp1600 IIIB линия клеток (15 - 12) была получена в происходящей из фибробласта Balb/c клеточной линии 3T3. Она получена от докторов Ron Germain и Jay Berzofsky, National Institute of Health, Bethesda, M.D. Экспрессирующая gp160 линия клеток использовалась для стимуляции in vitro примированных in vivo лимфоцитов селезенки, а также использовалась в качестве мишени для индукции gp160 специфичных ЦТЛ. Во многих экспериментах пептид 188IIIb, который содержит доминантный эпитоп ЦТЛ использовался для стимуляции in vitro. Для повторной стимуляции пептидом в культуре в среду добавлялся IL-2. Мышей Balb/c иммунизировали один раз 1 мкг gp120 на мышь с AF или без нее. Инъекции мышам производили подкожно и в основание хвоста. Клетки селезенки брали от иммунизированных мышей через три недели после иммунизации и стимулировали in vitro облученными трансфектантами gp160 или пептидом 18IIIb. После пяти дней культивирования in vitro примирование оценивалось по наличию специфичных эффекторов, способных лизировать трансфектанты gp160 и нетрансфицированные линии клеток. В некоторых экспериментах в качестве мишеней использовались инфицированные vac: gp160 P815 клетки. Результаты показаны в таблице 4A, когда ответ ЦТЛ потенцирован AF и gp120. Необходимо отметить, что в системе gp120 оптимальной антигенной рецептурой для gp120 специфичной индукции ЦТЛ (после одной иммунизации 1 мкг gp120 в AF) является та, которая не содержит или содержит минимальные количества плюрониевого. Однако, когда мышей иммунизировали многократно 5 мкг gp120 в AF, содержащей более высокие концентрации плюрониевого (3.75%), наблюдалась существенная индукция ЦТЛ (данные не представлены).
Следующий пример демонстрирует применение антигенных рецептур этого изобретения с использованием только одного или двух компонентов. Эти примеры показывают, что ответы ЦТЛ могут индуцироваться только с двумя из трех вышеназванных компонентов.
Пример 6
Определение решающих компонентов, необходимых для индукции ЦТЛ.
Чтобы определить необходимы ли все отмеченные выше компоненты для антиген-специфичной индукции ЦТЛ, мышей иммунизировали овальбумином в микропсевдоожиженной рецептуре с различными комбинациями двух из трех компонентов, представленных в вышеназванных AF, замещая третий компонент PBS. Два компонента, использованных комбинаций, были следующими:
сквалан / ТВИН в PBS,
сквалан / плюрониевый в PBS или
плюрониевый / ТВИН в PBS.
Другой набор групп включался, когда мышей иммунизировали ova оформленным в однокомпонентной системе, т.е. сквалана в PBS плюрониевого в PBS или ТВИН в PBS и только.
Вышеприведенные трехкомпонентные антигенные рецептуры состоят из:
0.300 г ТВИН 80 (Aldrich, WI),
1.875 г плюрониевого L121 (BASF, NJ), и 7.5 г сквалана (Aldrich, WI), доводимых до 50 мл PBS.
Двухкомпонентные рецептуры составляли:
сквалан / ТВИН : 0.300 г ТВИН 80 и 7.5 г сквалана, доводимых до 50 мл PBS;
Плюрониевый / ТВИН :
1.875 г плюрониевого 121 и 0.300 г ТВИН 80, доводимых до 50 мл PBS.
Плюрониевого / сквалана :
1.875 г плюрониевого 121 и 7.5 г сквалана, доводимых до 50 мл PBS.
Трехкомпонентными рецептурами с изменяемой концентрацией плюрониевого были:
Концентрации сквалана и ТВИН сохранялись как в предыдущих, но концентрации плюрониевого изменялись (см. талб.1А).
Образцы затем обрабатывались в приборе для микропседоожижения, модель 110Т, Microfludics Corp., помещали во флаконы и хранили при 4oC до использования.
Овальбумин (ova, Sigma, MO) взвешивали и доводили до концентрации раствора 0.3 мг/мл в HBSS (Whittaker, выше).
Стандартный раствор 0.3 мг/мл соединяли с двухкомпонентными рецептурами в следующих количествах:
5 частей раствора овальбумиа 0.3 мг/мл,
3.3 части 2-компонентной рецептуры и
1.7 части HBSS.
Подобным же образом β-gal и HIV gp120 смешивали с AF.
Рецептура энергично перемешивалась и сохранялась на льду до инъецирования. Все растворы соединяли непосредственно перед инъекцией.
Каждая мышь получала 200 мкл одной рецептуры, содержащей 30 мкл ova путем подкожной инъекции и введения в основание хвоста. Мышам давали отдохнуть в течение по крайней мере от двух до четырех недель до изъятия селезенки.
Через две недели после иммунизаций подготавливали клетки селезенок и стимулировали in vitro облученными EG7-ova. После пяти дней культивирования оценивалось наличие ova-специфических ЦТЛ путем испытания в отношении 51Cr-EG7-ova или 51Cr-EL4 в 4-часовом исследовании выделения 51Cr. Данные, показанные на фигурах 6 - 8, демонстрируют, что овальбуминовая сформированная в приборе для микропсевдоожижения двухкомпонентная система может примировать ova специфические ЦТЛ in vivo.
Далее мы оценили относительное значение отдельных компонентов по их способности индуцировать ЦТЛ в комбинации с белковыми антигенами. Для целей иммунизации растворимый антиген смешивался с составляющими, прошедшими микропсевдоожижение, чтобы получить стабильную гомогенную эмульсию с размерами частиц в пределах от 250 до 300 нм. Чтобы к тому же определить компоненты рецептуры сквален - ТВИН 80 - плюрониевый (СТП), ответственные за индукции ЦТЛ, мы иммунизировали мышей ova в смеси сквалана - ТВИН 80 (СТ), плюрониевого - ТВИН 80 (ПТ) или смеси сквалана - плюрониевого (СП) и в качестве контроля, в сквалане (С), ТВИН 80 (Т) или плюрониевом (П). Мышей также иммунизировали ova-SAFm (содержащим 70 мкг MDP) или ova-алюм в качестве контролей адъюванта. Для положительного контроля мыши были иммунизированы клетками селезенки, цитоплазматически нагруженными растворимым ova. Были также использованы других комбинации и заместители, и результаты представлены в таблице 1. Результаты показывают, что 30 мгк ova в комбинации с СТП или СТ примируют ответ ЦТЛ, ограниченных классом 1, у мышей. Примирование ova специфически ЦТЛ ova в СТП или ova в СТ по-видимому происходит лучше, чем примирование, индуцированное клетками селезенки, цитоплазматически нагруженными растворами ova. Ova в ПТ или в СП мог индуцировать ova специфические ответы ЦТЛ у мышей, но неустойчиво или плохо. В отличие от SAFm, добавлением MDP к СТ рецептуре не нарушало ova специфической индукции ЦТЛ у мышей (таблица 2). Не происходило ova-специфической индукции ЦТЛ, ни когда мышей иммунизировали ova, смешанным с отдельными компонентами, С, П или Т, ни когда мышей иммунизировали ova-SAFm или ova-алюм. У мышей, иммунизированных таким количеством ova, как 1 мг в (a) HBSS, в (b) SAFm или (c) адсорбированным на алюм, не примировались ova специфические ЦТЛ.
Пример 7
Компоненты, необходимые для продукции ova специфических антител
Мышей иммунизировали три раза через 2-недлеьные интервалы 30 мкг ova в HBSS, СТП, СТ, ПТ или СП. В качестве положительного контроля мышей также имунизировали ova-AF, так как известно, что SAFm индуцируют сильный антительный ответ. Через семь дней после второй и третьей иммунизации мышей обескровливали и сыворотки испытывали на ova специфический антительный ответ. Результаты показаны в таблице 3. Они указывают на то, что мыши, иммунизированные ova в СТП, СТ или в SAFm проявляют сходные анти-ova ответы после трех иммунизаций.
Пример 8
HIV gp120 специфическая индукция ЦТЛ
HIV gp120 IIIB использовался в качестве третьей антигенной системы для определения индукции ЦТЛ с СТП или 2- и 3-компонентными вариантов рецептуры. Мышей иммунизировали 1 мкг gp120 IIIb в HBSS, СПТ, ПТ или в СТ. Для контроля мышей иммунизировали 1 мкг gp120 IIIb в SAFm или CFA (полный адъювант Фрейнда) (таблица 4B). Через три недели после иммунизации клетки селезенки подготавливали и стимулировали in vitro клетками трансфектанта 15 - 12, обработанными митомицином, или пептидом 18IIIb. После пяти дней культивирования полученные в результате эффекторные клетки испытывали против вируса коровьей оспы: gp160 IIIB или исходных инцифированных вирусом коровьей оспы клеток P815 в качестве мишеней. Результаты показывают, что gp120-сквалан-ТВИН 80 постоянно индуцирует gp120 специфический ответ ЦТЛ у мышей (таблица 4A и 4B). CFA или SAFm были, однако неспособны индуцировать gp120 специфические ЦТЛ (таблица 4B). В отдельном исследовании gp120 в СТ с изменяемыми дозами плюрониевого от 0.0015% до 1.5% испытывались на индукцию ЦТЛ.
Пример 9
Индукция gp120 специфического гуморального ответа у мышей
Для индукции gp120 специфических гуморальных ответов мышей иммунизировали 1 мкг gp120IIIb три раза через двухнедельные интервалы. Животных обескровливали и сыворотку крови испытывали на наличие IgG антител, определяя gp120IIIb в твердофазном исследовании по ELISA. Результаты эксперимента 1 показывают, что gp120-в СТ или ПТ являются лучшими иммуногенами, чем gp120-HBSS, gp120 SAFm (таблица 5) или gp120-СТП. Однако результаты эксперимента 2 показывают, что gp120 в СТ иил СТП (содержащих концентрации плюрониевого 1.5% или 3.75%) могут вызывать ответы с высоким титром антител.
Пример 10
gp120 специфические антительные ответы у обезьян
Обезьяны (две на группу) были иммунизированы gp120-SAFm, gp120-СТП, gp120-СТ или gp120-HBSS. Для контроля группу обезьян иммунизировали рекомбинантным вирусом коровьей оспы, содержащим gp160 IIIb. Обезьян иммунизировали через двухнедельные интервалы и кровь брали через две и три недели после второй иммунизации. Неименные и иммунные сыворотки от каждой обезьяны разводили серийно и оценивали на анти-gp120 активность путем ELISA, как описано в материалах и методах. Данные (фигура 9) показывают, что у обезьян, иммунизированных gp120-СТП или gp120 SAFm, индуцировались сходные ответы. У одной обезьяны, иммунизированной gp120-СТ, индуцировался анти-gp120 ответ, сходный с ответом в группе, иммунизированной gp120-SAFm или gp120-СТП. У одной обезьяны, иммунизированной gp120-СТ, не индуцировался сильный анти-gp120 ответ после двух иммунизаций.
Пример 11
gp120 специфические ответы ЦТЛ у обезьян
Обезьян иммунизировали 30 мкг - 50 мкг HIV gp120 в AF или в HBSS многократно. Для контроля обезьян также иммунизировали рекомбинантным gp160 в вирусе коровьей оспы. Наши предварительные результаты показывают, что у 1/2 обезьян, иммунизированных gp120-AF, и у 1/2 обезьян, иммунизированных gp160: вирус коровьей оспы, показано преимущественное умерщвление инфицированных vac: gp160 аутологических мишеневых клеток (фиг-ы 10A и 10B).
(2) ИНФОРМАЦИЯ ПО ПОСЛЕДОВАТЕЛЬНОСТИ ID N: 1
(I) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: - 24
(B) ТИП: - аминокислотная
(C) ЧИСЛО НИТЕЙ: - единственная
(D) ТОПОЛОГИЯ: - линейная
(II) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: SEQ ID NO: 1:
Glu Gln Leu Glu Ser Ile Ile Asn Phe Glu Lys Leu Thr Glu Trp Thr - 16
20 Ser Ser Asn Val Met Glu Glu Arg - 24
(2) ИНФОРМАЦИЯ ПО ПОСЛЕДОВАТЕЛЬНОСТИ ID N: 2:
(I) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: - 19
(B) ТИП: - аминокислотная
(C) ЧИСЛО НИТЕЙ: - единственная
(D) ТОПОЛОГИЯ: - линейная
Asp Glu Asn Pro Val Val His Phe Phe Lys Ang Ile Val Thr Pro Arg - 16
Thr Pro Pro - 19н


Изобретение относится к медицинской иммунологии. Сущность изобретения состоит в создании антегенной композиции, содержащей антиген, стабилизирующий детергент и дополнительные компоненты. Композиция обладает способностью при введении в организм индуцировать цитотоксический эффект Т-лимфоцитов. Описан также способ индуцирования такого ответа. Изобретение содержит также способы лечения инфекций вирусной и невирусной природы. Технический результат изобретения заключается в расширении арсенала средств и методов борьбы с инфекцией различного происхождения. 3 с. и 35 з.п.ф-лы, 5 табл., 9 ил.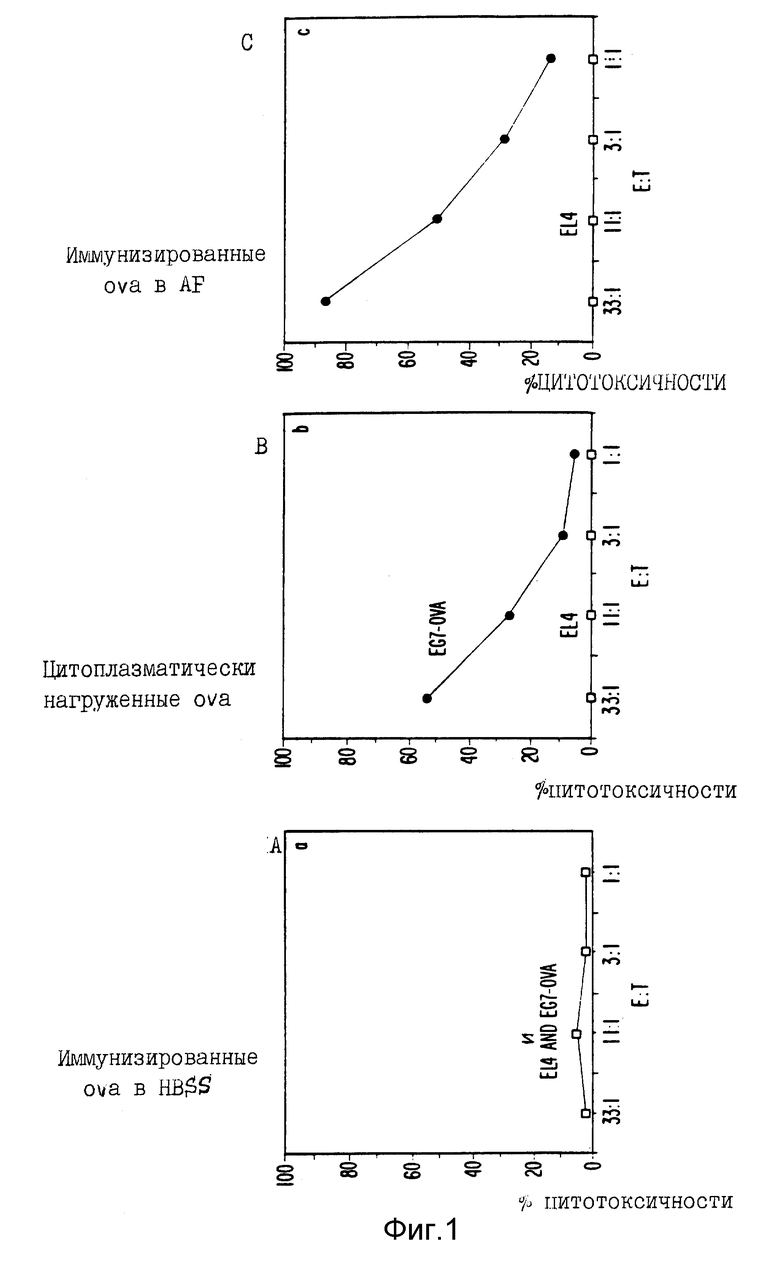
| СПОСОБ ФУТЕРОВКИ ДЕТАЛЕЙ ФУРМЕННОГО ПРИБОРА ДОМЕННОЙ ПЕЧИ | 0 |
|
SU202975A1 |
| EP 0405315, 02.01.91 | |||
| Торфодобывающая машина с вращающимся измельчающим орудием | 1922 |
|
SU87A1 |

