Изобретение относится к "очеловеченным" антителам и их связывающим белкам, способным связываться с Т-клетками, имеющими определенные вариабельные бета-цепи, а в частности с субпопуляциями клеток, экспрессирующих Vβ 5.2 и/или 5.3 и Vβ 8.1 человека. Настоящее изобретение также относится к получению таких антител; к фармацевтическим композициям, содержащим эти антитела; и к терапевтическому использованию этих антител, например, для лечения аутоиммунных заболеваний.
Т-клетки играют главную роль в дифференцировке и регуляции эффекторных механизмов в иммунной системе (Paul et а1. (1987) Science 195:1293-1300). Распознавание антигена и молекул главного комплекса гистосовместимости Т-клетками должно быть специфическим и точно регулируемым, поскольку неправильная иммунная регуляция способствует развитию аутоиммунитета. В некоторых лабораториях проводились исследования заболеваний, которые, по-видимому, обусловлены неправильной иммунной регуляцией, например такие как аутоиммунные состояния и некоторые формы иммунодефицита; причем в патогенезе указанных заболеваний участвуют Т-клетки.
Существуют такие ситуации, в которых, как сообщалось, происходит клональная или олигоклональная экспансия отдельного комплекса Т-клеточного рецептора. Наиболее ярким примером является состояние малигнизации (злокачественного развития), которое приводит к Т-клеточному лейкозу или к образованию лимфомы. В условиях развития Т-клеточного лейкоза или лимфомы Т-клеточный рецептор действует как уникальный опухолевый маркер, поскольку в этих случаях происходит стабильная перестройка Т-клеточных рецепторов и их представление на поверхности клетки. Другая ситуация, где конкретный рецепторный комплекс Т-клеток играет активную роль, возникает у реципиента трансплантированного органа, Т-лимфоциты которого имеют Т-клеточные рецепторы, делающие их агрессивными против молекул главного комплекса гистосовместимости (МНС) конкретного донора, например донора трансплантата костного мозга.
Что более важно, некоторые исследователи сообщали о селективном гене V-области т-клеточного рецептора антигена, экспрессируемом при некоторых аутоиммунных состояниях. Так, например, Grunwald и др. сообщают, что у пациентов, страдающих саркоидозом, экспрессия продукта Vα 2.3-гена в CD4+Т-клетках в бронхоальвеолярной промывной жидкости была преимущественной по сравнению с лимфоцитами периферической крови (Grunwald et а1., (1992) Eur.J.Immunol. 22: 129). При болезни Кавасаки преимущественная экспансия Vβ2 и Vβ8 в Т-клетках наблюдалась в начальной стадии болезни (Abe et а1., (1992) Proc. Natl, Acad. Sci. USA 89:4066).
C этой точки зрения было также проведено широкое изучение ревматоидного артрита. Некоторые исследователи отмечали предпочтительную экспансию субпопуляций Т-клеток; так, например, DerSimonian и др. ((1993) J.Exp.Med. 177-1623) наблюдали предпочтительную экспансию Vα 12.1-несущих Т-клеток в CD8+-лимфоцитах периферической крови; Stamenkovic и др. ((1988) Proc.Nati. Acad. Sci. USA 85:1179) сообщают, что, как показал Саузерн-блот-анализ, синовиальные мембрано-инфильтрующие Т-клетки, культивированные в 1L2, были олигоклональными; Paliard и др. ((1991) Science 253: 34) высказывают предположение, что суперантиген активирует Vβ 14+-Т-клетки, включая аутореактивные Т-клетки, которые клонально размножаются и мигрируют в синовиальную жидкость пациентов, страдающих ревматоидным артритом; Howell и др. ((1991) Proc.Nati. Acad.Sci. USA 88:10921) областей Т-клеток: Vβ 3, 14 и 17 в 2R+-клетках синовиальной жидкости пациентов, страдающих ревматоидным артритом; Uematsu и др. (Proc.Natl.Acad. Sci. USA 88:8534) показали использование гена V-области олигоклональных Т-клеток в Т-клетках синовиальной жидкости пациентов с ревматоидным моноартритом; а в заявке на Международный патент N W090/06758 указывается на причастность Vβ 3, 9 и 10 к ревматоидному артриту.
Кроме того, было также проведено тщательное изучение кишечных воспалительных заболеваний. Некоторыми группами исследователей, например Posnett и др. ((1990) J. Clin. Invest 85:1770); Spencer и др. ((1991) J.CIin.Pathol, 44: 915); Trejjdosiewicz и др. ((1991) Clin. Exp. Immunol. 84:440); и Van Kerckhove и др. ((1992)J.Exp.Meo.175:57), было отмечено, что кишечные воспалительные заболевания ассоциируются с размножением Т-клеточных популяций или преимущественной экспрессии V-области Т-клеточного рецептора. Другие исследователи (van Shooten et а1., (1992) Proc. Natl.Acad.Sci. USA 89:112 44; Wang et al. , (1993) Proc. Natl.Acad.Sci. USA 90:188) сообщали о преимущественной экспрессии гена V-области Т-клеток в бактерии Mycobacterium leprae.
Было установлено, что у человека, экспансия Т-клеток, несущих Vβ 8.1, в воспалительной ткани ассоциируется с некоторыми аутоиммунными заболеваниями, включая болезнь Крона (Posnett et al., (1990) J.Clin.Invest. 85:1770-1776), болезнь Кавасаки (Abe et al., (1992) Proc.Nail. Acad.Sci. USA 89: 4066-4070, Abe et al., (1993) J.Exp. Mad. 177:791-796) и ревматоидный артрит (Brennan et al., (1988) Clin.Exp.Immunol. 73: 417-423).
Другим аутоиммунным заболеванием, относящимся к объектам интенсивных исследований, является рассеянный склероз (MS). Рассеянный склероз представляет собой иммунопатологическое заболевание, характеризующееся инфильтрацией и демиелинизацией мононуклеарных клеток центральной нервной системы. Хотя патогенез рассеянного склероза неизвестен, однако, по-видимому, в развитии этого заболевания играют роль как генетический, так и экологический факторы. Главными элементами наследственной предрасположенности к данному заболеванию является связь этого заболевания с конкретными гаплотипами главного комплекса гистосовместимости класса II, а в частности HLA-DR21 DQWL (Terasaid et а1. (1976) Science 1933:1245-1247; Но et а1. (1982) Immunogenetics 15: 509-517; Spielman et а1. (1982) Epidemiol.Rev. 4:45-65; Francis et al. , (1986) Lancet 1:211; Elian et а1. (1987) Disease Markers 5:89-99; Urban et. al. (1988) Cell 54:577-592, Vandenbark et а1. (1989) Nature 341: 541-544; Howell et а1. (1989) Science 246:668-670).
Было показано, что Т-клетки, выделенные из спинномозговой жидкости пациентов, страдающих рассеянным склерозом, используют ограниченный набор генов V-области. Обнаружение in vivo-активированных Т-клеток, специфичных к основному белку миелина, у пациентов, страдающих рассеянным склерозом, свидетельствует о причастности MBP-реактивных Т-клеток к патогенезу этого заболевания (Wucherpfennig et а1. (1990) Science 248:1016-1019). При определении участия Vβ Т-клеточного рецептора MBP-реактивных клеточных линий посредством полимеразной цепной реакции (PCR) для амплификации кДНК с использованием праймеров Vβ T-клеточного (TCP) была выявлена преимущественная экспрессия лимитированного числа Vβ-генов (Wucherpfennig et а1., (1990), см. выше) - Vβ 17 и в меньшей степени Vβ 12 часто используются в распознавании области антигенной детерминанты аутоантигена MBP человека (MBP - основной белок миелина)) (Oksen-berg et.al. Nature 362:68). Результаты некоторых исследований также позволяют предположить, что в пораженных тканях головного мозга пациентов, страдающих рассеянным склерозом, происходит экспрессия ограниченного числа генов Vα Т-клеточного рецептора (Oksenberg et al., (1990) Nature 345: 344:346). Анализ популяции Т-клеток с использованием количественной PCR и окрашивания моноклональным антителом (mAb) показал, что Vβ 5.2 и/или 5.3 используются преимущественно MBP-специфическими Т-клетками, выделенными от пациента с рассеянным склерозом, по сравнению с контролем (Oksenberg и др. (1993), см. выше) (реарранжированные Vβ 5.2 гены были обнаружены в головном мозге пациентов с определенным HLA-фенотипом) и Kotzin и др. (1991) Proc.NatI. Acad. Sci.Sci. USA 88:9196 (смешение в сторону экспрессии вериабельной области β-цепи 5.2 и в меньшей степени Vβ 6.1 наблюдалось для MBP-специфических клонов от пациентов с рассеянным склерозом).
В настоящее время не существует эффективных способов лечения рассеянного склероза (Harrison's Principles of Internal Medicine, 12-th ed. Wilson et al., McGraw Hill, Inc., 1991). Применяемое терапевтическое лечение направлено на улучшение состояния в периоды обострения, предупреждения рецидивов или прогрессирования заболевания и ослабления симптомов.
Однако было обнаружено, что экспрессия гена вариабельной области Vβ 8.2 ассоциируется с экспериментальным аллергическим энцефаломиелитом (ЕАЕ) в мышиной модели человеческого рассеянного склероза. Было продемонстрировано, что обработка Vβ 8.2-специфическим моноклональным мышиным антителом (mAb) может предупреждать и ослаблять симптомы заболевания (Achaorbea et а1., (1988) Cell 54:263; и Urban et al. (1988) Cell 54:577). Таким образом, существует крайняя необходимость в разработке антитела или "антитело-подобной" молекулы, подходящей для лечения этого заболевания.
Антитела обычно состоят из двух тяжелых цепей, связанных друг с другом дисульфидными связями, и двух легких цепей, ассоциированных с N-концом каждой тяжелой цепи соответственно. Каждая тяжелая цепь имеет у своего N-конца вариабельный домен, а за ним, у своего С-конца, константный домен. Каждая легкая цепь также имеет у своего N-конца вариабельный домен, за которым следует константный домен. Вариабельные домены каждой пары легкой и тяжелой цепей образуют антигенсвязывающий центр. Вариабельные домены на легкой и тяжелой цепях имеют одинаковую общую структуру, и каждый домен содержит четыре каркасных участка, которые имеют относительно консервативные последовательности и которые перемежаются с тремя гипервариабельными (или определяющими комплементарность) участками (CDR). Указанные четыре каркасных участка имеют до некоторой степени β-складчатую конформацию, а гипервариабельные участки образуют петли, соединяющие β-складчатую структуру. Эти гипервариабельные участки находятся в непосредственной близости друг к другу, образуя антигенсвязывающий центр. Гипервариабельные и каркасные участки антител могут быть определены по системе нумерации Кэбета (Kabat и др. (1987) "Sequences of Proteins of Iimmunological Interest" US Dept. of Health and Human Services, US Government Printing Office) в сочетании рентгеновской кристаллографией, как описано в WO91/09967.
Для продуцирования антитела, которое может быть направлено против конкретного антигена, обычно используют метод Kohler Milstein (Kohler et al., (1976) Nature 256:495-497). B общих чертах этот метод предусматривает иммунизацию мышей антигеном; слияние клеток селезенки, полученных от иммунизированных мышей, с мышиными миеломными клетками; и отбор из продуцированных таким образом гибридом одной или несколько гибридом, секретирующих моноклональное антитело, специфичное к данному антигену-мишени.
Указанные мышиные моноклональные антитела (mAb) можно с успехом использовать в терапии. Однако такие антитела имеют в основном мышиное происхождение, а поэтому сами по себе они являются антигенными для человека. Если это антитело вводить человеку несколько раз, то его иммунная система будет вырабатывать иммунный ответ против указанного mAb, что в результате приведет к неэффективности этого антитела.
Поэтому Winter и его сотрудниками (см., например, Reichmann et а1., (1988) Nature 332:323-327; и Verhoeyen et al. (1988) Science 239; 1534-1536) было высказано предположение о возможности "прививки" гипервариабельных областей мышиного моноклонального антитела к каркасным участкам антитела человека с получением антитела с CDR-вставками, обладающего связывающими свойствами донорного мышиного mAb и физиологической совместимостью с иммунной системой человека благодаря наличию акцепторного каркаса, происходящего от антитела человека.
Однако функция молекулы антитела зависит от ее трехмерной структуры, которая, в свою очередь, зависит от ее первичной аминокислотной последовательности. Изменение аминокислотной последовательности антитела может неблагоприятным образом повлиять на его активность. Аналогичным образом, фрагменты антитела могут не сохранять соответствующую трехмерную структуру, которая благоприятствует связывающей активности. Кроме того, изменение ДНК-последовательности, кодирующей антитело, может повлиять на способность клетки, содержащей эту ДНК-последовательность, осуществлять экспрессию, секрецию или сборку молекулы антитела. Точно определить конкретные остатки, составляющие гипервариабельные участки (CDR), довольно трудно, и, кроме того, эти остатки не обязательно будут соответствовать всем остаткам гипервариабельных участков, определенных по системе нумерации Кэбета. Имеются также критические каркасные остатки, которые играют важную роль в расположении гипервариабельных участков, взаимодействующих с антигеном, либо которые участвуют во взаимодействиях между тяжелыми и легкими цепями. В некоторых случаях может оказаться необходимым изменить определенные каркасные остатки таким образом, чтобы они соответствовали донорным остаткам в определенных положениях, сообщая тем самым антителу с чужеродными CDR менее "человеческий" характер.
Были опубликованы различные предложения по поводу идентификации остатков гипервариабельных участков и каркаса, которые необходимо заменить на донорные остатки для продуцирования нужного антитела с чужеродными CDR (см., например, Queen et al., W090/07861; Kurrle et al, EP-A-O 403156; Adair et al., W091/09967; Queen et al., W092/11018; Bendig et al., WO92/01568). Как видно из этих документов, продуцирование нужного антитела с чужеродными CDR в каждом конкретном случае не является точно предсказуемым процессом.
Несмотря на проблемы, возникающие при попытке продуцировать специфическое антитело с чужеродными CDR, авторам настоящего изобретения в предпочтительном его варианте удалось получить антитело с чужеродными гипервариабельными участками (CDR), сконструированное на основе человеческих каркасных участков и имеющее антигенсвязывающий центр, специфичный к вариабельным участкам бета-цепи, которые являются характерными для определенных субпопуляций, Т-клеток. Неожиданно было обнаружено, что "очеловеченные" антитела настоящего изобретения обнаруживают такую же хорошую, если не лучше, аффинность связывания с Т-клетками-мишенями, как и их мышиные прототипы. Антитела настоящего изобретения или их части имеют те преимущества, что по сравнению со своими мышиными прототипами они являются менее иммуногенными, что способствует снижению неблагоприятных реакций организма пациента при лечении с использованием указанных антител.
Настоящее изобретение относится к "очеловеченным" антителам, обладающим специфичностью в отношении селективного связывания с определенными субпопуляциями Т-клеток; причем указанные антитела являются в высокой степени чувствительными к связыванию с этими субпопуляциями и обнаруживают такую же, если не лучше, специфичность и аффинность связывания, как и их мышиные прототипы (mAb), от которых происходят гипервариабельные участки рассматриваемых антител.
Настоящее изобретение также относится к способам получения таких "очеловеченных" антител, предусматривающим использование новых кДНК, кодирующих определенные остатки гипервариабельных и каркасных участков, и встраивание этих остатков в каркасные области тяжелой и легкой цепи иммуноглобулина человека. В других своих вариантах настоящее изобретение относится к фармацевтическим композициям и терапевтическим методам с использованием указанных антител.
В одном из особо предпочтительных вариантов осуществления настоящего изобретения мышиные моноклональные антитела (mAb) 16G8, которые распознают Vβ 8.1 человека, были "очеловечены" путем встраивания некоторых CDR и выбранных каркасных остатков, происходящих от мышиного mAb, в каркасные области тяжелой цепи KOl и легкой цепи REl. Для этого кДНК, кодирующие "очеловеченные" тяжелые (IgG1) и легкие (К) цепи в экспрессирующих векторах клеток млекопитающего, вместе с селективными маркерами Neo и DHFR соответственно трансфецируют в клеточную линию DHFR-фибробластов китайского хомячка (CHO) с последующей селекцией и амплификацией. Секретированные "очеловеченные" mAb (обозначенные "TM29") сохраняют специфичность в отношении Vβ 8.1 Т-клеточного рецептора TCR с аффинностью, сравнимой с аффинностью прототипов мышиного mAb (16G8). Таким образом, настоящее изобретение продемонстрировало, что "очеловечение" антител против TCR может быть осуществлено при сохранении нужной специфичности и аффинности в отношении данной субпопуляции клеток.
В другом своем варианте настоящее изобретение относится к использованию этих "очеловеченных" антител в качестве терапевтического средства для лечения аутоиммунных заболеваний, а в частности болезни Крона.
В другом из особо предпочтительных вариантов осуществления настоящего изобретения мышиное моноклональное антитело ТМ23, которое распознает Vβ 5.2 и 5.3 человека, "очеловечивают" путем встраивания CDR в каркасы тяжелой цепи NEWM и легкой цепи REl. Для этого кДНК, кодирующие "очеловеченные" тяжелые (IgG1) и легкие (К) цепи в экспрессирующих векторах клеток млекопитающего вместе с селективными маркерами Neo и DHFR соответственно трансфецируют в клеточную линию NHFR-фибробластов китайского хомячка (CHO) с последующей селекцией и амплификацией. Продуцированные таким образом "очеловеченные" mAb, обозначенные "ТМ27", сохраняют специфичность в отношении Vβ 5.2 и 5.3 TCR с аффинностью, сравнимой с аффинностью прототипов мышиного mAb (ТМ23). В других своих вариантах настоящее изобретение относится к использованию ТМ27 в качестве терапевтического средства для лечения рассеянного склероза (MS) у человека.
На фиг. 1 представлены различные вариабельные последовательности легкой цепи и тяжелой цепи для ТМ27;
на фиг. 2 представлены различные вариабельные последовательности легкой цепи и тяжелой цепи для ТМ29;
на фиг. 3 представлены результаты конкурентного анализа, проводимого для сравнения ТМ27L/NSO и ТМ27L/CHO с 4H11; и
на фиг. 4 представлены графики Скэтчарда, описывающие эксперимент, проиллюстрированный на фиг. 3.
Ниже приводится объяснение некоторых терминов, используемых в описании настоящего изобретения.
Используемый в настоящей заявке термин "антитело" относится к Ig или к любому его фрагменту, к мономерам или димерам легкой или тяжелой цепи и к одноцепочечным антителам, таким как одноцепочечные FV-фрагменты, в которых вариабельные домены тяжелой и легкой цепи соединены пептидным линкером; причем все перечисленные антитела могут быть натуральными или продуцированными с использованием техники рекомбинантных ДНК или какого-либо другого метода при условии, что полученное в результате антитело имеет по крайней мере один антигенсвязывающий центр. При этом остальная часть антитела не обязательно должна содержать лишь последовательности, происходящие от Ig. Так, например, может быть сконструирован ген, в котором ДНК-последовательность, кодирующая часть цепи Ig человека, соединена с ДНК-последовательностью, кодирующей аминокислотную последовательность полипептидной эффекторной или репортерной молекулы. Таким образом, термин "антитело" включает в себя понятие "гибридное антитело" (см. ниже).
Используемый в настоящей заявке термин "связывающий белок" относится к конструкции, имеющей конкретную аминокислотную последовательность, происходящую от антитела или комбинации антител, причем указанная последовательность имеет такую стерическую конфигурацию, что она образует по крайней мере один антигенсвязывающий центр.
Используемое в настоящей заявке сокращение "mAb" означает моноклональное антитело, продуцированное гибридомой или производной клеточной линией.
Используемый в настоящей заявке термин "рекомбинантное антитело" относится к антителу, продуцированному способом, предусматривающим использование техники рекомбинантных ДНК.
Термин "CDR" или "участок, определяющий комплементарность" или "гипервариабельный участок", относится к тем частям вариабельных участков тяжелой или легкой цепи антитела, которые, будучи расположены в непосредственной близости друг от друга в трехмерном пространстве, образует антигенсвязывающую поверхность.
Термин "химерное антитело", используемый в настоящем описании, означает антитело, в котором вариабельные домены, в целом происходящие из антитела от первого вида млекопитающих, соединены по крайней мере с одним константным доменом, происходящим из антитела от другого вида млекопитающего.
Используемый в настоящем описании термин "антитело с чужеродными CDR" относится к антителу или его фрагменту, в котором CDR, происходящие из антитела от первого вида млекопитающего, встроены в вариабельные каркасные участки, в основном происходящие от второго вида млекопитающего. Некоторые выбранные аминокислоты в указанных участках могут также происходить от указанного первого вида млекопитающего.
Используемый в настоящем описании термин "гибридное антитело" относится к белку, содержащему по крайней мере связывающую часть Ig, соединенную пептидной связью по крайней мере с частью другого белка. При этом следует отметить, что для описания таких конструкций некоторые специалисты могут также использовать термин "химерное антитело", однако в настоящем описании эти конструкции называются гибридными антителами, а термин "химерные антитела" имеет смысл, определенный выше.
Термин "очеловеченное антитело", используемый в настоящем описании, относится к антителу, содержащему в одном или двух своих вариабельных доменах по крайней мере один, а предпочтительно два или три гипервариабельных участка (CDR), происходящих из антитела от первого вида млекопитающего; при этом следует отметить, что такие антитела могут также содержать в сочетании с некоторыми гипервариабельными аминокислотными последовательностями определенные выбранные аминокислоты каркасной области, происходящие от указанного первого вида млекопитающего. Остальные Ig-части антитела происходят от одного или нескольких антител другого вида. Вариабельные домены могут быть получены с помощью техники рекомбинантных ДНК или с помощью пептидного синтеза.
Термин "экспрессирующие векторы" относится к векторам, способным экспрессировать содержащиеся в нем ДНК-последовательности, т.е. к векторам, в которых кодирующие последовательности соответствующим образом присоединены к другим последовательностям, обеспечивающим их эффективную экспрессию. Полезным, но не всегда необходимым (т.е. клетки насекомых) элементом эффективного экспрессирующего вектора является последовательность, кодирующая маркер, т.е. последовательность, кодирующая векторную последовательность, сообщающую клетке, содержащей данный белок, фенотипические свойства (например, резистентность к неомицину, резистентность к метионинсульфоксимину или триптофен-прототрофию), которые позволяют затем легко идентифицировать эти клетки. В целом термин "экспрессирующий вектор" является функциональным определением, а поэтому под этим термином подразумевается любая ДНК-последовательность, способная к эффективной экспрессии конкретной кодирующей ДНК-последовательности. В настоящее время в качестве таких векторов часто используются плазмиды. Таким образом, термины "плазмида" и "экспрессирующий вектор" часто являются взаимозаменяемыми. Однако настоящее изобретение включает в себя и другие формы экспрессирующих векторов, которые обладают эквивалентными функциями и которые со временем становятся все более известными, например ретровирусы, аденовирусы, in-vitro-системы (Baranov et а1., (1989) Gene 84:2:463) и т.п.
Как уже было установлено ранее, ДНК-последовательности после их правильного присоединения (т.е. такого присоединения, которое обеспечивало бы их экспрессию) к регулирующей экспрессию последовательности могут быть экспрессированы в клетках-хозяевах. Указанные экспрессирующие векторы обычно способны реплицироваться в организмах-хозяевах либо в виде эписом, либо в виде интегральной хозяйской хромосомной ДНК.
Термин "рекомбинантные клетки-хозяева" относится к клеткам, котоые были трансформированы векторами, сконструированными с использованием техники рекомбинантных ДНК. Благодаря такой трансформации, хозяйская клетка способна продуцировать нужный продукт в достаточных количествах, а не в тех небольших количествах (чаще всего в количествах, меньших, чем детектируемые количества), которые обычно продуцируются нетрансформированным хозяином. Антитела и связывающие белки настоящего изобретения могут продуцироваться рекомбинантными клетками-хозяевами в количествах, достаточных для проведения дополнительных экспериментов, или в коммерчески приемлемых количествах, например около 100 граммов или более.
В описании способов выделения антител или связывающих белков из рекомбинантных хозяев термины "клетка" и "клеточная культура", используемые для обозначения источника антитела, являются (если это не указано особо) взаимозаменяемыми. Другими словами, выделение из "клеток" может означать либо центрифугирование целых клеток с последующим выделением белка, либо выделение из клеточной культуры, содержащей среду и суспендированные клетки, либо, как это возможно в случае клеточных линий миеломы, выделение из асцитной культуры.
"Очеловеченные" антитела или их связывающие белки настоящего изобретения имеют аминокислотные последовательности, которые содержат все или часть гипервариабельных участков (CDR), происходящих в основном от моноклонального антитела, обладающего специфичностью к выбранным субпопуляциям Т-клеток, а в частности к субпопуляции Vβ 8.1 или субпопуляции Vβ 5.2/5.3. В предпочтительных вариантах осуществления настоящего изобретения предпочтительным моноклональным антителом является мышиное антитело. Поскольку антитела настоящего изобретения называются "очеловеченными" антителами, то в предпочтительных вариантах осуществления изобретения каркасная аминокислотная последовательность вариабельных доменов антител или их частей происходит в основном от человека. Очевидно, что такое "очеловечение" антител осуществляют для снижения их иммуногенности при терапевтическом введении человеку. Для использования в настоящем изобретении предпочтительными являются каркасные области NEWM или KOL, происходящие от тяжелой цепи иммуноглобулина человека, и каркасные области REl, происходящие от легкой цепи иммуноглобулина человека. При этом некоторые выбранные каркасные остатки могут иметь также мышиное, а не человеческое происхождение. Это необходимо, как очевидно, для получения соответствующей трехмерной структуры молекулы антитела в целях повышения специфичности и аффинности связывания с конкретной Т-клеточной субпопуляцией.
Следует отметить, что в объем настоящего изобретения входит любая часть "очеловеченных" антител (исходя из широкого определения термина "антитело"), которая обладает специфичностью и аффинностью связывания в отношении выбранных субпопуляций Т-клеток. Таким образом, в объем настоящего изобретения входят связывающие белки, происходящие от указанных антител, а также фрагменты, сохраняющие связывающую способность на уровне, по крайней мере подходящем для терапевтического использования, как описано ниже.
Каждый специалист может оценить параметры связывания различных конструкций настоящего изобретения с помощью стандартных анализов на связывание, проводимых с использованием специфических Т-клеток или Т-клеточных рецепторных белков, имеющих выбранную вариабельную область, являющуюся мишенью. Так, например, может быть осуществлен анализ на связывание с mAb-прототипом, на связывание с различными клеточными линиями, анализ Скэтчарда, и т.п., и полученные результаты можно сравнить с результатами, полученными для mAb-прототипа. Альтернативно для этих целей могут быть привлечены различные системы in vivo-моделей, например таких, как описаны ниже, либо для оценки терапевтической эффективности конструкций настоящего изобретения могут быть приведены клинические испытания; причем в этой связи следует отметить, что какая-либо конкретная конструкция может оказаться весьма подходящим терапевтическим средством, даже если ее характер связывания с антигеном-мишенью отличается от характера связывания с mAb-прототипом.
В особо предпочтительных вариантах осуществления настоящего изобретения аминокислотная последовательность "очеловеченного" антитела или его связывающего белка содержит всю вариабельную легкую цепь или ее часть, выбранную из группы, включающей в себя:
Легкая цепь ТМ 29:
1 DIQMTQSPSSLSASVGDRVTITCRASSS VNYIYWYQQTPGKAPLLIYYT 50
51 SNLAPGVPSRFSGSGSGTDYTFTISSL QPEDIATYYCQQFTSSPFTFGQG 100
101 TKLQIT
Легкая цепь TM 27:
1 DIQMTQSPSSLSASVGDRVTITKSASQG INYLNWYQQTPGKAPKLLIYY 50
51 TSSLHSGVPSRFSGSGSGTDYTFTISS LQPEDIATYYCQQYSKLPRTFGQ 100
101 GTKLQIT
отдельно или в комбинации со всей вариабельной тяжелой цепью или ее частью, выбранной из группы, включающей в себя:
Тяжелую цепь TM 29:
1 EVQLVESGGGVVQPGRSLRLSCSSSGFT FSNFGMHWVRQAPGKGLEWVAY 50
51 ISSGSSTIYYADTLKGRFTISRDNSKN TLFLQMDSLRPEDTGVYFCARRG 100
101 EGAMDYWGQGTPVTVSS 117
Тяжелую цепь TM 27:
1 QVQLQESGPGLVRPSQTLSLTCTVSGFS LTAYGVNWVRQPPGRGLEWLGM 50
51 IWGDGNTDYNSALKSRVTMLKDTSKNQ FSLRLSSVTAADTAVYYCARDRV 100
101 TATLYAMDYWGQGSLVTVSS 120
Для сохранения трехмерной конформации CDR очень важно выбрать каркасные остатки, находящиеся в непосредственной близости от отдельных CDR (Foote & Winter (1992) j.Mol., Biol.224:487-499). Так, например, в некоторых предпочтительных вариантах осуществления изобретения были сконструированы модифицированные тяжелые цепи. В одних вариантах вместо человеческих остатков 66-69 включительно и 73 была введена мышиная последовательность Leu-Ser-Ile-Ser (66-69) и Asp (73).
В других предпочтительных вариантах в положения 78 и 79 вводили мышиные остатки валин и фенилалин соответственно либо в положение 92 вводили мышиный аргинин.
Эти замещения проиллюстрированы на фиг. 1, где гипервариабельные участки показаны жирным шрифтом и подчеркнуты. Такие предпочтительные конструкции антитела (включая их связывающие белки) связываются с Vβ 5.2/5.3.
Другие предпочтительные антитела и связывающие белки настоящего изобретения могут быть получены путем модификации тяжелых цепей в остатках 23 и 24. В эти положения были введены соответствующие мышиные Ala-, в других проиллюстрированных конструкциях остаток 75 заменяли на мышиный пролин, а остатки 88, 89 и 91 заменяли на мышиный аланин, метионин или тирозин соответственно. Эти конструкции антител или их связывающих белков, которые более полно показаны на фиг. 2, связываются с Vβ 8.1-Т-клетками.
Особенно предпочтительными для использования в настоящем изобретении являются антитела со встроенными чужеродными CDR, состоящие в основном из легкой цепи TM27 и тяжелой цепи TM27, где остаток 48 является мышиным лейцином или изолейцином. Предпочтительными также являются антитела со встроенными чужеродными CDR, состоящие в основном из легкой цепи TM29 в комбинации с тяжелой цепью TM29.
"Очеловеченные" антитела или связывающие белки настоящего изобретения могут быть продуцированы различными способами, предусматривающими экспрессию в трансфецированных клетках, предпочтительно таких как дрожжевые клетки, CHO, клетки насекомых или миеломные клетки. Наиболее предпочтительными клетками-хозяевами являются клетки яичника китайского хомячка (CHO).
Для конструирования антитела со встроенными чужеродными CDR или связывающего белка настоящего изобретения, сначала необходимо определить последовательность вариабельного домена антитела, имеющего нужные связывающие свойства. Подходящими клетками-источниками для таких ДНК-последовательностей являются клетки птиц, млекопитающих или других позвоночных, таких как куры, мыши, крысы и кролики, а предпочтительно мыши. Последовательности вариабального домена (VH и VL) могут быть определены, исходя из кДНК тяжелой и легкой цепи, синтезированной из соответствующей мРНК с использованием стандартной техники, в основном известной специалистам. Гипервариабельные участки могут быть затем определены по методу Кэбета (см. выше). CDR могут быть определены с помощью структурного анализа с использованием рентгеновской кристаллографии или техники молекулярного моделирования. Затем могут быть сконструированы составные CDR, содержащие все остатки, соответствующие гипервариабельным участкам, вместе с некоторыми выбранными остатками из каркасных участков. Полученные составные CDR могут быть затем перенесены в качестве "антигенсвязывающего центра", а оставшаяся часть антитела, если необходимо сконструировать полный иммуноглобулин (Ig), будет включать в себя константные домены тяжелой и легкой цепей и остальные каркасные остатки. Последние части могут происходить от антител человека различных классов.
Константные домены могут быть выбраны так, чтобы они обладали нужными эффекторными функциями, подходящими для использования сконструированного таким образом антитела. Так, например, иммуноглобулины человека изотипов IgG, IgG1 и IgG3 являются эффективными для фиксации комплемента и клеточноопосредованного лизиса. Для других целей могут быть более подходящими другие изотипы, такие как IgG2 и IgG4 или иммуноглобулины других классов, таких как IgM и IgE.
Для лечения человека в целях минимизации антиглобулинового ответа во время терапии, особенно предпочтительно использовать изотипы человека. ДНК-последовательности константного домена человека; предпочтительно в сочетании с их каркасными участками, могут быть получены в соответствии с хорошо известными процедурами. В качестве примера может служить САМРАТН 1Б от Burrougs Wellcome Ltd.
В соответствии с предпочтительными вариантами осуществления настоящего изобретения некоторые антитела с чужеродными CDR имеют определенные модификации в человекоподобном каркасном участке (другими словами, за пределами CDR вариабельных доменов), в результате чего эти антитела обладают удовлетворительной аффинностью связывания. Такая аффинность связывания составляет предпочтительно от около 10-5 М до около 10-12 M, а более предпочтительно по крайне мере около 10-8 М. Наиболее предпочтительно, чтобы афинность связывания была приблизительно равна или превышала аффинность связывания антитела-прототипа (в предпочтительных вариантах таким прототипом-антителом является антитело мыши).
При конструировании антител с чужеродными CDR в соответствии с настоящим изобретением VH- и/или VL-генные сегменты могут быть модифицированы путем мутагенеза. Любому специалисту ясно, что аналогичным способом могут быть также модифицированы (см., например, PCT/US 89/00297) другие нуклеотиды, кодирующие аминокислотные остатки или последовательности, содержащиеся в Fc-фрагменте или в других областях антитела. В этих целях могут быть использованы, например такие методы, как добавление, делеция, или неконсервативные замещения ограниченного числа различных нуклеотидов, или консервативного замещения многих нуклеотидов при условии, что будет сохранена правильная рамка считывания.
Для получения конечной конструкции могут быть использованы замещения, делении, инсерции или другие субкомбинации. Поскольку существует 64 возможных последовательностей кодонов и всего 20 известных аминокислот, то генетический код является вырожденным в том смысле, что различные кодоны могут кодировать одинаковые аминокислоты. Однако генетический код однозначен, т.е. каждый кодон кодирует только одну аминокислоту. Иначе говоря, для каждой аминокислоты существует по крайней мере один кодон, и каждый кодон кодирует одну конкретную кислоту и никакую другую. Совершенно очевидно, что для получения правильной аминокислотной последовательности в конечном продуцированном полипептиде необходимо, чтобы была сохранена правильная рамка считывания.
Техника осуществления добавлений, делеций или замещений в заранее определенных аминокислотных участках, имеющих известную последовательность, хорошо известна специалистам. Примерами такой техники является олигонуклеотидоопосредованный сайт-направленный мутагенез и полимеразная цепная реакция (PCR.).
Сайт-направленный мутагенез с использованием нуклеотидов в основном предусматривает гибридизацию олигонуклеотида, кодирующего нужную мутацию, с одноцепочечной ДНК, содержащей мутируемую область, и использование одной цепи в качестве матрицы для удлинения олигонуклеотида в целях продуцирования цепи, содержащей мутацию. Эта техника в различных ее видах описана Zoller Smith (1982) Nuc. Acids Res., 10:6487) Norris et al. ((1983) Nuc.Acids Res., 11:5103), Zoller & Smith ((1984) DNA 3:479), Kramer et al. ((1982) Nuc.Acids Res. 10:6475).
Полимеразная цепная реакция предусматривает экспоненциальную амплификацию ДНК in vitro с использованием сайт- специфических олигонуклеотидов. Если необходимо, то эти олигонуклеотиды могут иметь изменения в последовательности. Техника с использованием полимеразной цепной реакции (PCR) описана Mulli и Foloona ((1987) Meth.Enz. 155:335). Примеры осуществления мутагенеза с использованием PCR описаны Higuchi и др. ((1988) Nuc.Acids Res., 16: 7351), Ho и др. ((1989) Gene, 77:51), Но и др. (Engineering Hybridization Restriction Genes without the Use of Restriction Enzymes: Gene Splicing by Overlap Extention), Horton и др. ((1989) Gene, 77:61).
Нуклеотидные последовательности настоящего изобретения, способные к экспрессии нужных антител или связывающих белков этих антител могут быть получены из различных полинуклеотидов (геномной ДНК, кДНК, РНК или синтетических олигонуклеотидов). В целях настоящего изобретения предпочтительно, чтобы полинуклеотидная последовательность представляла собой гибрид из кДНК и геномной ДНК. Эта полинуклеотидная последовательность может кодировать различные компоненты Ig (например V-, J-, D- и C-домены). Наиболее широко применяемым в настоящее время методом продуцирования такой полинуклеотидной последовательности является соединение подходящих геномной и кДНК-последовательностей, однако могут быть также использованы кДНК-последовательности (см., ЕР-А-О 239400; и Reichmann et а1. (1988) Nature 332:323).
Некоторые подходящие экспрессирующие векторы и хозяйские клетки описаны в US-A-A 816567.
Векторы и методы, описанные в настоящей заявке, являются вполне подходящими для использования в клетках-хозяевах широкого ряда прокариотических или эукариотических организмов.
Разумеется, в основном для клонирования ДНК-последовательностей в целях конструирования векторов, используемых в настоящем изобретении, предпочтительными являются прокариотические клетки. Например, особенно предпочтительными являются различные плазмидные клонирующие векторы, происходящие от E. coli, такие как Bluescript М13 (Stratagene, San Diego, California), pUCl9c (Genbank Accession N vB0026), pBP322 (описан ниже), и т.п. Эти примеры носят, разумеется, чисто иллюстративный характер и не ограничивают настоящего изобретения.
Прокариотические клетки могут быть также использованы для экспрессии. Так, например, в этих целях могут быть использованы вышеуказанные штаммы E. coli, бациллы, такие как Bacillus subtilis, и другие энтеробактерии, такие как Salmonella Typhimurium или Serratia marcescens и различные виды Pseudomonas.
В ocновном плазмидные векторы, содержащие последовательность репликона и регуляторную последовательность, происходящие от видов, совместимых с клеткой-хозяином, используются в сочетании с этими хозяевами. Обычно вектор имеет сайт-репликации а также маркерную последовательность, обеспечивающую фенотипическую селекцию в трансформированных клетках. Так, например, E.coli обычно трансформируют одним из многих производных плазмиды рВР322, происходящей от вида E.coli (Bolivar. (1977) Gene 2.95). Плазмида pBR322 содержит гены резистентности к ампициллину и тетрациклину, что обеспечивает легкую идентификацию трансформированных клеток. Плазмида рВR322, ее производные или другие микробные плазмиды могут также содержать или могут быть модифицированы так, чтобы они содержали промоторы, которые могут быть использованы микробными микроорганизмами для экспрессии рекомбинантных белков. При конструировании рекокбинантных ДНК обычно используют лактозные промоторные системы (Chang et а1. (1978) Nature 275:615; Itakura et al. (1978) Science 198: 1056; Goeoell et al. (1979) Nature 281:544) и триптофановые промоторы системы (Goeoell et al. (1980) Nuc Acids Res. 8.4057; и ЕР-А-О 036776). Хотя указанные промоторы применяются наиболее часто, однако уже открыты и используются другие микробные промоторы, и детали, относящиеся к их нуклеотидным последовательностям, опубликованы в литературе, что позволяет специалистам осуществить их функциональное лигирование в плазмидные векторы (Sieben-list et al. (1980) Cell 20:269).
Помимо прокариотов могут быть также использованы эукариотические микробы, такие как дрожжи. Из эукариотических микроорганизмов чаще всего используются Saccharomyces cerevisiae, или обыкновенные пекарские дрожжи, хотя существует множество других хорошо известных штаммов. Для экспрессии в Saccharomyces обычно используют, например, плазмиду УРр7 (Stinchcomb et а1. (1979) Nature 282: 39; Kingsman et al. (1979) Gene 7:141; Tschemper et al. (1980) Gene 19: 157). Эта плазмида уже содержит ген trp1, который сообщает селективный маркер мутантному штамму дрожжей, а именно неспособность этого штамма к росту в присутствии триптофана, например АТСС N 44076 или РЕР4 (Jones, (1977) Genetics 8:85:12). Наличие повреждения trp1, характеризующего геном дрожжевой хозяйской клетки, дает возможность эффективно обнаруживать трансформантов посредством их культивирования в отсутствие триптофана.
Подходящими промоторными последовательностями в дрожжевых векторах являются промоторы гена 3-фосфорглицераткиназы или других ферментов, таких как энолаза, глицеральдегид-3- фосфатдегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфофруктоназа, глюкозо-6-фосфат-изомераза, 3-фосфоглицератмутаза, пируваткиназа, триозофосфат-изомераза, фосфоглюкозоизомераза и глюкокиназа (Hess et al ((1968) J. Adv. Enzyme Reg. 7:149; Holland en. al. (1978) Biochemistry 17:4900).
При конструировании подходящих экспрессирующих плазмид в 3'-конец последовательности экспрессирующего вектора, нужной для экспрессии, могут быть также лигированы терминаторные последовательности, ассоциированные с указанными генами, для осуществления полиаденилирования мРНК и терминации. Другими промоторами, которые имеют дополнительные преимущества, а именно регулируют транскрипцию в условиях роста, являются промоторные участки алкогольдегидрогеназы 2, изоцитохрома С, кислой фосфатазы, расщепляющих ферментов, ассоциированных с метаболизмом азота, и вышеуказанной глицеральдегид- 3-фосфатдегидрогеназы, фермента, ответственного за утилизацию мальтозы и галактозы. При этом подходящим является любой плазмидный вектор, содержащий совместимый с дрожжами промотор, сайт инициации репликации и терминаторные последовательности.
Помимо микроорганизмов в качестве хозяев могут быть также использованы культуры клеток, происходящих от многоклеточных организмов. В принципе любая такая клеточная культура является подходящей независимо от того, является ли данный организм позвоночным или беспозвоночным. Однако за последнее время интерес к клеткам позвоночных сильно возрос, и в настоящее время размножение клеток позвоночных в культуре (тканевой культуре) стало рутинной процедурой (Kruse и Patterson, (1973), Tissue Culture, Academic Press).
Примерами таких подходящих хозяйских клеточных линий являются VEPO, Hela, клетки яичника китайского хомячка (CHO), W138, BHK, Cos-7, MDCK и клетки миеломы и миеломы Os (поставляемые фирмой Celltech, Slough U.K.). Экспрессирующие векторы для таких клеток могут содержать (если это необходимо) сайт инициации репликации, а также промотор, расположенный перед экспрессируемым геном, любые необходимые сайты связывания рибосомы, сигналы для сплайсинга мРНК, сайты полиаденилирования и сайты терминации транскрипции.
Для использования в клетках млекопитающих в экспрессирующих
векторах часто используются регуляторные элементы, происходящие от вирусного материала. Так, например, широко используемые промоторы происходят от цитомегаловирусов человека (HCM), вируса полиномы, аденовируса 2, и наиболее часто используемый промотор происходит от вируса обезьян 40 (SV 40). Ранний и поздний промоторы вируса SV40 являются особенно ценными, поскольку оба эти промотора могут быть легко получены из вируса в виде фрагмента, который также содержит сайт инициации репликации вируса SV40 (Fiers et al. (1978) Nature 273:113). Кроме того, также возможно, а часто желательно, использовать промоторную или регуляторную последовательности, обычно ассоциируемые с последовательностью нужного гена, при условии, что такие регуляторные последовательности совместимы с клеточной системой хозяина.
Сайт инициации репликации может быть введен либо путем конструирования вектора, содержащего экзогенный сайт инициации, например, происходящий от SV40 или другого вирусного источника (такого как вирус полиомы, аденовирус, VSV или BPV), либо он может быть введен посредством механизма репликации в хромосомах клеток-хозяев. Если вектор интегрирован в хромосому клетки-хозяина, то часто бывает достаточно второго механизма.
Векторы, содержащие ДНК-сегменты, представляющие конкретный интерес (например, последовательности, кодирующие тяжелую и легкую цепи, и последовательности, регулирующие экспрессию), могут быть перенесены в клетку-хозяина хорошо известными методами в зависимости от типа клетки-хозяина. Например, для прокариотических клеток часто используется кальций-хлоридная трансфекция, тогда как для других клеток-хозяев обычно используют обработку кальций-фосфатом, липотрансфекцию или электропорацию (Maniatis et аl. (1990) Molecular Cloning: A Laboratory Manual, 2nd Edition, Cold Shring Harbor Press).
После экспрессии конструкции настоящего изобретения могут быть очищены стандартными методами, включая, например, преципитацию сульфатом аммония, аффинную хроматографию, колоночную хроматографию и гель-электрофорез (Scopes, (1982) Protein Purification, Springer Verlag, NY). Аффинности связывания экспрессированных таким образом конструкций могут быть определены стандартными методами, которые более подробно описаны ниже.
В основном очищенные антитела со встроенными чужеродными CDR или связывающие белки антител имеют предпочтительно гомогенность по крайней мере 90-95%, причем для фармацевтического использования наиболее предпочтительной является гомогенность 98-99% или более. Полученные таким образом частично или полностью, если это необходимо, гомогенные антитела со встроенными чужеродными CDR могут быть затем использованы в диагностических или терапевтических целях (включая эктракорпоральное использование), либо в целях разработки и осуществления методов анализа, иммунофлуоресцентного окрашивания и т. п. (Lefkovite & Pernis (1979 1981), Immunological Methos, Volumes 1 and 11, Academic Press, NY).
Антитела с чужеродными CDR настоящего изобретения или связывающие белки этих антител предназначены в основном для лечения Т-клеточноопосредованных расстройств. Так, например, такими расстройствами являются аутоиммунные заболевания, а именно диабет типа 1, рессеянный склероз, ревматоидный артрит, системная красная волчанка, болезнь Крона и тяжелая псевдопаралитическая миастения.
Предпосылкой терапевтического использования Т-клеточных антител настоящего изобретения явилась корреляция между специфическим иммунопатологическим заболеванием и преимущественной экспрессией конкретного генного продукта, а именно конкретной V-области Т-клеточного рецептора антигена, или усиленной экспансией конкретного гена V-области Т-клеточного рецептора антигена. В этом смысле, особенно ценными являются V-области Т-клеточного рецептора антигена отчасти потому, что в данном случае можно регулировать иммунный ответ у пациента путем конкретных терапевтических мероприятий с использованием Т-клеточного рецептора антигена. Более конкретно, присутствие или экспрессия конкретного локуса вариабельной области коррелирует, как было показано, с конкретными иммунными заболеваниями. При определении конкретного генного локуса V-области, ассоциированного с конкретным иммунным расстройством, можно проводить лечение пациента путем ингибирования "атаки" Т-клетками, несущими конкретную V-область.
Используемый в настоящем описании термин "лечение" включает в себя понятия "предупреждение", "подавление" или "лечение" заболевания. Термин "предупреждение" заболевания означает мероприятия, предусматривающие введение протективной композиции до возникновения заболевания. Так, например, в мышиной модели EAE (экспериментальной аллергический энцефаломиелит) успешное введение протективной композиции перед введением инъекции энцефалитогена, индуцирующего это заболевание, приводит к "предупреждению" заболевания.
Термин "подавление" означает мероприятия, предусматривающие введение композиции после индуцирования заболевания, но до появления его клинических симптомов. И аналогично, в мышиной модели EAE успешное введение протективной композиции после инъекции энцефалитогена, но до появления нейрологических симптомов приводит к "подавлению" заболевания.
Термин "лечение" означает мероприятия, предусматривающие введение протективной композиции после появления симптомов заболевания. В уже приводимом выше примере EAE успешное введение протективной композиции после инъекции энцефалитогена и после появления клинических симптомов приводит к "лечению" заболевания.
Системы мышиных моделей, которые могут быть использованы для исследования эффективности антител или связывающих белков этих антител при лечении заболеваний, являются доступными специалистам. Исследования системной красной волчанки (SLE), проводимые с помощью экспериментов на восприимчивых мышах, описаны Knight и др. ((1978) j. Exp. Med., 147:1653) и Reinersten и др. ((1978) New Eng. , J. Med. 299:515). Эксперименты по исследованию тяжелой псевдопаралитической миастении (MG) проводили на мышах-самках SJL/J путем индуцирования заболевания растворимым белком AchP, взятым от другого вида, как описано Lindstrom и др. ((1988) Adv.Immunol. 42:233-284). Артрит индуцировали у восприимчивого штамма мышей путем инъекции коллагена типа II, как описано Stuart и др. ((1984), Ann. Rev. Immunol. 42: 233-284). Стимулированный артрит индуцировали у восприимчивых крыс путем инъекции микобактериального белка теплового шока, как описано Van Eden и др. ((1988) Nature 331: 171-173. Тиреодит у мышей индуцировали путем введения тироглобулина, как описано Maron и др. ((1980) J.Exp.Meo 152:1115-1120). Инсулинзависимый сахарный диабет (IDD M) возникает естественным путем, либо он может быть индуцирован у некоторых видов мышей, например так, как описано Kanasawa и др. ((1984) Diabetologia 27:113). EAE у мышей и крыс служит моделью MS человека. В этой модели заболевания, характеризующиеся димиелинизацией, индуцируют путем введения основного белка миелина, как описано Paterson ((1986) Textook of Immunopathology (Mischer и др., изд.)), Grune и Stratton (New York, pp. 179-213), McFarlin и др.((1973) Science 179: 478-480); и Satoh ((1987) - J. Immunol. 138: 179-184).
Антитела со встроенными чужеродными CDR настоящего изобретения или связывающие белки этих антител могут быть также использованы в комбинации с другими антителами, в частности mAb, реагирующими с другими маркерами на клетках человека, ответственных за указанные заболевания. Так, например, подходящими Т-клеточными маркерами являются маркеры, сгруппированные в так называемые "дифференцировочные кластеры" (этот термин был дан на 1-ом Международном симпозиуме по дифференцировке лейкоцитов (Bernhard et al. (1984) Leukocyte Typing, Springer Verlag, NY).
B основном антитела с чужеродными CDR настоящего изобретения или связывающие белки этих антител используются в очищенной форме вместе с фармакологически приемлемыми носителями. Обычно такими носителями являются водные или спиртовые/водные растворы, эмульсии или суспензии, включая физиологический раствор и забуференные среды. Носителями для парентерального введения являются раствор хлорида натрия, растворы Рингера, содержащие декстрозу; декстрозу и хлорид натрия; и лактат. Подходящие физиологически приемлемые адъюванты для поддержания комплекса в суспензии могут быть выбраны из загустителей, таких как карбоксиметилцеллюлоза, поливинилпирролидон, желатин и альгинаты.
Носителями для внутривенного введения являются жидкие и питательные наполнители и электролитные наполнители, такие как растворы Рингера, содержащие декстрозу. При этом могут также присутствовать консерванты и другие добавки, такие как антимикробные средства, антиоксиданты, хелатообразующие агенты и инертyые газы (Маcк (1982) Remington's Pharmaceutical Sciences, 16th Edition).
Конструкции настоящего изобретения могут быть введены как в виде отдельных композиций, так и в сочетании с другими агентами. Такими агентами могут быть различные иммунотерапевтические лекарственные средства, такие как циклоспорин, метотрексат, адриамицин или цисплатин; и иммунотоксины. Фармацевтические композиции могут представлять собой смеси ("коктейли") различных цитотоксичных или других агентов в сочетании с антителами с чужеродными CDR настоящего изобретения или связывающими белками этих антител; или даже комбинации конструкций настоящего изобретения и антитела, содержащие чужеродные CDR и имеющие различные специфичности.
Фармацевтические композиции настоящего изобретения могут быть введены любым из известных способов, обычно применяемых специалистами. Например, для лечения, включая различные виды иммунотерапии без каких-либо ограничений, антитела и связывающие белки настоящего изобретения могут быть введены любому пациенту в соответствии со стандартной техникой. Такое введение может быть осуществлено любым подходящим способом, например парентерально, внутривенно, внутримышечно, внутрибрюшинно или также, если это приемлемо, путем непосредственного вливания с помощью катетера. Доза и режим введения зависят от возраста, пола и состояния пациента, а также от других одновременно вводимых лекарственных средств, возможных противопоказаний и других параметров, обычно принимаемых в расчет лечащим врачом.
Конструкции настоящего изобретения могут быть лиофилизованы с последующим хранением и восстановлением в случае использования в соответствующем носителе. Было показано, что такие методы являются эффективными при использовании стандартных иммуноглобулинов, при этом может быть использована любая техника лиофилизации и восстановления. Следует отметить, что лиофилизация и восстановление препарата могут приводить к различной степени активности антитела (например, при использовании стандартных иммуноглобулинов антитела IgM имеют тенденцию к большой потере активности, чем антитела IgG), а поэтому для компенсации этих потерь используемые уровни антител могут быть скорректированы.
Композиции, содержащие антитела настоящего изобретения или их смесь, могут быть введены в профилактических и/или терапевтических целях. При некоторых терапевтических применениях термин "терапевтически-эффективная доза" означает адекватное количество, необходимое для достижения по крайней мере частичного ингибирования, суппрессии, модуляции, цитолиза или других измеряемых параметров воздействия на популяции выбранных клеток. Количества, необходимые для получения эффективных доз, зависят от тяжести заболевания и общего состояния иммунной системы пациента, но в основном доза антитела с чужеродными CDR или связывающего белка этого антитела составляет в пределах от 0,005 до 5,0 мг на кг веса тела, а обычно используют дозу от 0,05 до 2,0 мг/кг. Для профилактических целей композиции, содержащие конструкции настоящего изобретения, или их смесь могут быть также введены в немного меньших дозах.
Композиция, содержащая конструкцию настоящего изобретения, может быть использована в профилактических и терапевтических мероприятиях для альтерации, инактивации, цитолиза или удаления нужной Т-клеточной популяции-мишени у млекопитающих.
В другом варианте осуществления настоящего изобретения рассматриваемые конструкции могут быть использованы экстракорпорально или in vitro для селективного цитолиза, истощения или другого эффективного удаления целевых клеточных популяций из гетерогенного собрания клеток. Так, например, кровь, взятая у млекопитающего, может быть экстракорпорально объединена с антителами, имеющими чужеродные CDR вставки, или со связывающими белками таких антител, в результате чего из крови млекопитающего могут быть удалены ненужные клетки, а после этого кровь возвращается млекопитающему в соответствии со стандартной техникой.
Помимо терапевтического применения, композиции настоящего изобретения могут быть использованы в других целях.
В одном из вариантов осуществления изобретения рассматриваемые антитела или связывающие белки могут быть использованы в исследовательских целях. Связывающие Т-клеточный рецептор белки или антитела настоящего изобретения могут быть также использованы в структурных или функциональных исследованиях Т-клеточного рецептора. Помимо субстратов или связывающих доменов для специфических форм Т-клеточных рецепторов, эти конструкции антител могут служить в качестве инструмента для конформационных исследований в целях установления нативной конфигурации различных участков Т-клеточного рецептора. В еще одном варианте осуществления изобретения Т-клеточные антитела или связывающие белки могут быть использованы в качестве диагностических зондов. В одном из конкретных вариантов осуществления изобретения конструкции настоящего изобретения могут быть использованы для определения количества или присутствия подсемейства V-участков некоторых Т-клеток в биологическом образце. Один из таких анализов описан в Rittershaus - W092/08981, опубл. 26 Мая 1992 под названием "Therapeutical and Diagnostic Methods Using Total Leukocyte Syrface Antigene". По существу, настоящее изобретение относится к методам диагностики иммунопатологических заболеваний, таких как рассеянный склероз, основанным на обнаружении конкретных субпопуляций Т-клеточного рецептора антигена в биологическом образце.
Антитела, содержащие чужеродные CDR, или связывающие белки таких антител могут быть подвергнуты мечению в соответствии с известной техникой. Указанные конструкции могут также иметь и другие in vivo-применения. Так, например, они могут быть использованы для селективной обработки клеток периферической крови, из которой необходимо удалить лишь нужные Т-лимфоциты, или аналогично для обработки клеточной культуры, из которой необходимо удалить нежелательные Т-лимфоциты.
В нижеследующих примерах приводится более подробное описание некоторых аспектов настоящего изобретения, однако эти примеры не должны рассматриваться как некое ограничение изобретения.
Пример 1. TM27 - Краткое описание процедур.
1. Мышей BALC-с иммунизировали клеточной линией HPB-All Т-клеточного лейкоза (один раз внутрибрюшинно и два раза внутривенно).
2. От этих иммунизированных мышей брали клетки селезенки, и эти клетки подвергают слиянию с мышиными иммортализованными клетками миеломы клеточной линии P3X53AG8.653.
3. Отбор соответствующих клонов проводили в HAT-среде.
4. Скрининг отобранных таким образом клонов на соответствующее связывание осуществляли путем иммунопреципитации радиактивно меченых HBP-All.
5. Субклонирование осуществляли путем серийного разведения.
6. Тест на специфичность проводили путем PBL-стимуляции и исследования модуляции CD3.
7. Продуцированные гибридомой антитела обозначали 4Н11.
8. Из этого клона получали PHK и выделяли мРНК. После чего получали кДНК.
9. кДНК секвенировали и проверяли правильность полученной последовательности.
10. Выделенные кДНК (H и К) клонировали в векторы М13.
11. Выбранные каркасные участки, происходящие от антитела человека, были введены посредством мутагенеза, как описано в примере 2.
12. "Очеловеченные" антитела (≈ 1 мг каждое) были очищены из О-трансфектантов для предварительной оценки.
Было продуцировано пять различных антител TM27:
ТМ27: содержит лейцин в положении 48 тяжелой цепи.
TM271: содержит изолейцин в положении 48 тяжелой цепи.
TM27.1: FS в положениях 78-79 заменены на VF.
TM27.2: VTML/T заменены на LSIS/N в положениях 67-70/73.
TV27.3: V заменен на R в положении 92.
Целью настоящей работы является "очеловечение" мышиного моноклонального антитела (ТМ23), направленного против Vβ 5.2/5.3 TCP (Т-клеточного рецептора) человека, и продуцирование подходящей клеточной линии, экспрессирующей "очеловеченное" антитело. Это "очеловеченное" моноклональное антитело против Vβ 5.2/5.3 TCP обозначали ТМ27.
Мышиное антитело.
Гибридому 4Н11 получали путем слияния мышиной миеломной клеточной линии P3X63AG8.653 с клетками селезенки, взятыми от мышей Balb/c, иммунизированных клеточной линией Т-клеточного лейкоза HBP-А11. Полученные гибридомы продуцировали моноклональное антитело ТМ23 со специфичностью к Vβ 5.2/5.3 Т-клеточного рецептора (TCR) человека. Изотип-IgG2a/каппа.
"Очеловечение" антител.
кДНК, кодирующую тяжелую и легкую цепи ТМ23, выделяли из 4H11-кДНК-библиотеки, сконструированной с использованием 3'-концевых праймеров, специфичных к мышиным IgG2a и каппа-области. Выделенные кДНК-клоны были подтверждены ДНК-секвенированием и внутренним аминокислотным секвенированием. Гипервариабельные участки (CDR-участки, определяющие комплементарность) тяжелой и легкой цепей были идентифицированы путем анализа первичной структуры консервативных мышиных каркасных последовательностей. С помощью PCR были выделены ДНК-фрагменты: L, V, (D), J (L - лидерный, V - вариабельный, D - фрагмент изменчивости, J - соединительный фрагмент), и клонированы в экспрессирующие векторы клеток CHO, а именно pTCSIL C DHFR и pTCSL CgI NeoAp, несущие кДНК каппа- и константной области IgG1 человека соответственно в целях экспрессии мыш. V/чел. C-химерного антитела.
Исходя из сравнения аминокислотных последовательностей, выведенных из кДНК-клонов, было установлено, что тяжелая цепь ТМ23 принадлежит подгруппе Кэбета 1В. Кроме того, было установлено, что VK-последовательность принадлежит к VKV (по Кэбету), поэтому для "очеловечения" тяжелой и легкой цепей соответственно были выбраны каркасные участки REl и NEWM. Для генерирования полных геномных конструкций L-V(D)J перегруппированные фрагменты V(D)J (за исключением части 5'V и 3'V) были клонированы в векторы М13, несущие лидерные и интронные последовательности в геномной конфигурации.
Экспрессия в миеломной клеточной линии.
После конструирования V(D)J-областей с чужеродными CDR посредством мутагенеза ДНК-фрагменты клонировали в экспрессирующие векторы миеломы, несущие константную область каппа и IgG1 человека соответственно. Было сконструировано пять "очеловеченных" тяжелых цепей с различными мутациями. Каждая из этих пяти конструкций тяжелых цепей была ко-трансфенирована вместе с "очеловеченной" легкой цепью в миеломную) клеточную линию NSO. "Очеловеченные" антитела были очищены из супернатантов культуры размноженных чел. IgG1/k-положительных трансфектантов в целях проведения анализа. Результаты оценки показали, что TM27L с наименьшими мышиными аминокислотными последовательностями окрашивал Vβ 5.3-Т-клетки так же хорошо, если не лучше, как и другие "очеловеченные" варианты. Поэтому вариант TM27L, обозначенный TM27, был выбран для дальнейших исследований.
Экспрессия в клетках CHO.
Лидерные и вариабельные области тяжелой и легкой цепи ТМ27 выделяли из миеломных конструкций с помощью PCR для клонирования в кДНК-конфигурации в экспрессирующие системы клеток CHO, pTCSL CqINeoAp (тяжелая цепь) и pTCSL C DHFR (легкая цепь), как описано выше. Затем CHO-клеточные трансфектанты отбирали, и чел. IgG1/к-положительные клетки клонировали. Один из наиболее высокоэффективных продуцентов, а именно TM27L-662-35, был размножен для создания клеточных заготовок и получения мелкомасштабных препаратов "очеловеченных" Ab. Экспрессированное в CHO антитело TM27 было охарактеризовано в отношении улучшения его продуцирования, и полученные результаты были сравнимы с результатами, полученными для миеломных клеточных линий и для исходного TM23. Клоны клеток CHO подвергали амплификации и клонированию для повышения экспрессии антитела и демонстрации пригодности этого штамма для повышения продуктивности.
Подробное описание.
Иммунизация мышей BALB/c клетками HPB-A.
β F1- (моноклональное тело против константной области β-цепи Т-клеточного рецептора человека) - иммунопреципитат Т-клеточной линии лейкоза человека HPB-ALL (3•107), полученный у Dr. Michael Brenner (Harvard Medical School), смешивали с полным адъювантом Фрейнда, инъецировали внутрибрюшинно (i.p.) мышам BALB/c. Через 5 недель мышам вводили три дня подряд бустер-дозу 3 • 107 клеток в физиологическом растворе (первую - внутрибрюшинно, а две последующие внутривенно (i.v.))
Слияние клеток селезенки с P3X63AG8.653.
Слияние проводили через три дня после последней внутривенной бустер-инъекции в соответствии с традиционной техникой. Клетки селезенки в 5-кратном избытке подвергали слиянию с клетками P3X63AG8.653 (мышиная миеломная клеточная линия, не секветирующая Ig и происходящая от мыши BALB/c; АТСС CRL 1580) с использованием 50%-ного полиэтиленгликоля 1500 (BDH, Dorest, England).
HAT-селекция гибридных клеток.
Гибридные клетки высевали на 96-луночные планшеты, содержащие в качестве фидера (питающего подслоя) 2 • 105 тимоцитов BALB/c на лунку, и отбирали на среде RPMl-1640, содержащей 15% FBS (фетальной коровьей сыворотки), и HAT (6 мМ гипоксантина, 50 мМ аминоптерина и 2 мМ тимидина).
Клетки инкубировали в атмосфере воздуха с повышенной влажностью в присутствии 5% CO2 при 37oC.
Скрининг гибридом.
Через 13-14 дней лунки с культурой скринировали с помощью ELISA и иммунопреципитации меченных 125I клеток HPB- ALL. В условиях иммунопреципитации CD3-молекулы диссоциировали из α- и β-цепей TCR. Гибридомы, которые реагировали с HPB, размножали в колбах Т-75 для последующего клонирования.
Клонирование нужных гибридом.
Гибридомы, обладающие специфичностью к TCAR HPB-ALL (TCAR - Т-клеточный рецептор антигена), клонировали путем серийного разведения. Клетки высевали на 96-луночные планшеты для культивирования с 2-кратным серийным разведением при клеточной плотности от 2 до 0,015 клеток на лунку. Колонии с наилучшим ростом и с наиболее высокой продуктивностью отбирали для повторной вышеописанной процедуры, которую проводили еще два раза. После этих двух дополнительных процедур отбора одну из конечных колоний идентифицировали как 4H11, mAb, экспрессированные клоном 4Н11, вызывали иммунопреципитацию молекул, идентичных α- и β-цепям TCR, осаждаемым антителом mAb βF1, которое реагирует с константной областью β-цепи TCR. Этот результат позволяет предположить, что указанное mAb обладает специфичностью к β-цепи TCR, экспрессированной на Т-клетках HPB-ALL.
Для удобства полученное антитело обозначали TM23.
Характеризация моноклонального антитела ТМ23, продуцированного гибридомой 4Н11.
Изотипирование моноклонального антитела ТМ23 проводили с использованием коммерчески доступного набора (Zymed, South San Francisco, CA). Результаты показали, что это антитело принадлежит к мышиному изотипу IgG2a/k.
Для подтверждения реактивности против TCR моноклональное антитело скринировали на его способность к ко-модуляции CD3-молекулы с поверхности клеток HPB-ALL. Проведенные ранее исследования показали, что моноклональные антитела к α- или β-цепи TCR способны к ко-модуляции CD3 с клеточной поверхности в том случае, когда эти клетки были предварительно инкубированы с mAb против TCR. При предварительном инкубировании клеток HPB-ALL с TM23 наблюдалось значительное снижение реактивности с mAb Leu-4 против CD3 по сравнению с реактивностью Leu-4, наблюдаемой в необработанном контроле.
Была проведена оценка нормальных PBL (лимфоцитов периферической крови) на реактивность с TM23. Из исследованных PBL-клеток от 1,6% до 3,0% этих клеток реагировали с TM23. Аналогичные результаты были получены и с некоторыми другими mAb, которые, как было показано, реагируют с детерминантами специфических V-областей.
Для того чтобы определить, связывается ли ТМ23 с тем же самым эпитопом, что и lCl (mAb, которое реагирует с подсемействами Vβ 5.2 и 5.3) (Boylston et а1., 1986, J. immunol. 137:741), была исследована способность одного mAb блокировать связывания другого антитела. При инкубировании клеток HPB-ALL сначала с TM23 ингибировалось связывание флуоресцеин-коньюгированного lCl. Аналогично, когда клетки ингибировали сначала с lCl, то ингибировалось связывание с TM23. Этот результат позволяет предположить, что антитела TM23 и lCl связываются с одним и тем же эпитопом либо с эпитопами, находящимися в непосредственной близости друг к другу.
В другом исследовании, проведенном для того чтобы определить, имеют ли TM23 и lCl одинаковую реакционную способность, линии PBL получали путем стимулирования другим mAb. Затем каждую линию оценивали на ее реактивность с каждым антителом mAb. Если линию PBL получали путем стимулирования антителом TM23, то 1C1 и 4H11 реагировали с аналогичным процентом клеток. Тот же самый результат был получен в случае получения линии PBL путем стимулирования антителом lCl.
Выращивание.
4H11-клон TM23.17-31-91 культивировали и размножали в среде MEM/F12 (1: 1) + 1% фетальной телячьей сыворотки, 100 мг/мл пенициллина, 100 мг/мл стрептомицина и 12,5 мМ глутамина.
Получение кДНК, кодирующей тяжелую и легкую цепи TM23.
Около 10x клеток 4H11 были выделены путем центрифугирования, после чего эти клетки один раз промывали охлажденным льдом PBS. Полную PHK получали с использованием набора для выделения полной PHK (Promega RNAgents Total Isolation-A Kit, Promega, кат. N Z 5110). Poly (A)+-PHK получали с использованием системы для выделения мРНК (Promega PolyATract mRNA Isolation Systev, Promega, кат. N: Z 5200).
Мышиные IgG2a и каппа-специфическую кДНК-библиотеку получали с использованием набора BRL Superscript Kit (BRL кат. N 82У8 А) и IgG2a-праймера:
5' ATATGCGGCCGCTCATTTACCCGGAGT CCGGGAGAAGCTCTTAGT 3'
и каппа-праймера:
5'ATATGCGGCCGCTTAACACTCATTCCTC TTGAAGCTCTTCACAAT 3'
Эти два праймера содержат рестрикционные Xho1-сайты и последовательности, комплементарные 3'-концам последовательностей, кодирующих мышиные IgG2a и каппа-области, соответственно, кДНК клонировали в плазмиду ps PORT (поставляемую в наборе) и трансформировали в компетентные клетки DH5α E.coli путем электропорации. Плазмидные клоны, несущие вставки с нужным размером, отбирали для секвенирования.
Идентификация кДНК-клонов, кодирующих тяжелые и легкие цепи TM23.
Минипрепараты плазмидной ДНК со вставками нужного размера (клоны легкой цепи N 1, 6, 19, 20, 25 и 26; клоны тяжелой цепи N 4, 9, 11, 15, 22 и 34) подвергали двухцепочечному ДНК-секвенированию с использованием секвенирующего набора (USB кат. 70770), который содержит Т7-праймер 5' к ДНК-вставке. Было показано, что последовательности клонов легкой цепи 6, 20 и 25 и клона тяжелой цепи N 34 являются мышиными клонами Ig. Клоны легкой цепи N 6, 20 и 25 имеют идентичные последовательности, а клон 6 имел точечную мутацию и был нефункциональным клоном. Клоны N 20 и 34 были секвенированы с использованием праймеров, специфических к константной области каппа.
5' GACATTGATGTCTTTGGGGTAGAAGTT GTT 3'
и IgG2a
5' GGTCACTGGCTCAGGGAAATAACCCTT GAC 3'
И аналогично, результаты показали, что клон N 6 является нефункциональным клоном, а клоны N 20 и 34 являются истинными клонами для легкой и тяжелой цепи соответственно. кДНК и аминокислотные последовательности, выведенные из двух клонов, показаны на фиг. 1.
Вследствие N-концевого блокирования микроаминокислотное секвенирование осуществляли от расщепленных цианистым бромидом (CNBr) пептидных фрагментов, выделенных из полиакриламидных гелей. Аминокислотная последовательность внутренней V-области, полученная в результате этого секвенирования, совпадала с последовательностью, выведенной исходя из кДНК. Полученные результаты подтвердили, что выделенные кДНК-клоны кодируют тяжелую и легкую цепь TM23.
Гипервариабельные области тяжелой и легкой цепей были идентифицированы по методу Кэбета и Wu (см. выше) и представлены на фиг. 1, где они выделены жирным шрифтом и подчеркнуты.
PCR-выделевие ДНК V-области тяжелой (H) и легкой (L) цепей и ДНК-секвенирование.
Продуцирование химерного антитела TM23.
Выделение мышиной VH-ДНК и VK-ДНК.
Вариабельные области тяжелой и легкой цепей TM23 были адаптированы для инсерции в экспрессирующие векторы. ДНК- последовательность, кодирующую мышиную VH TM23, амплифицировали из ТМ23М VHM и IgG2a посредством полимеразной цепной реакции с использованием праймеров: Н1ВАСК
(5' AGGTSMAPCTGCAGSAGTC WGG 3'), который содержит сайт для рестрикции фермента Pst 1 и VHIFOR
5'TGAGGAGACGGTGACCGTGGTCCCTTGG CCCCAG 3', который содержит BstEII-сайт. M означает C или A, S означает C или G; R означает A или G, а W означает A или T. Эти сайты были введены у 5'- и 3'-концов ДНК TM23 H. Амплифицированную ДНК разрезали ферментами Pst1 и BstEII и лигировали в Pst1 и BstEII-сайты двухцепочечной репликативной формы (RF) ДНК вектора HPCPI M13, разрезанного теми же самыми ферментами.
VHPCRI М13 представляет собой вектор фага M13, происходящий от Ml3-HuVHNP (Jones et а1. (1986) Nature 321; 522-525) и содержащий промотор иммуноглобулина, сигнальную последовательность и соответствующие сайты сплайсинга (Orlandi et а1. (1989) Proc. Natl, Acad. Sci. USA 86:3833-3837).
Лигированную ДНК трансформировали в компетентный штамм TGl E.coli (К12, (lac-pro), supE, thi, hsdD5 F' tra D36, proA+B+, lacO4 lacZ M15). Одноцепочечную ДНК получали из рекомбинантных бляшек.
ДНК-последовательность, кодирующую мышиную VK TM23, амплифицировали из TM23M и VK.MuCK посредством PCR с использованием олигонуклеотидных праймеров: VKlBACK:
5' GACATCCAGCTGACCCAGTCTCCA 3', который содержит PvulI-сайт и VKIFOR
5' GTTAGATCTCCAGCTTGGTCCC 3', который содержит BqlI-сайт в сочетании с двумя перекрывающимися внутренними праймерами, сконструированными для удаления HindIII-сайта в CDP3, а именно oilgo 943:
5' CCGAGGAAGTTTACTATACTG 3'
и oligo 944:
5' CAGTATAGTAAACTTCCTCGG 3'
(Horton и др. (1990). Таким образом, PvulI- и BglII- сайты были введены в 5'- и 3'-концы ДНК K TM23. Амплифицированную ДНК гидролизовали ферментами PvulI и BqlII и лигировали в PrulI и BclI-рестриктированный VKPCRl M13. ДНК разрезали ферментами BqlII и BclI для продуцирования "липких" концов, которые могут быть лигированы вместе.
M13 KPCRI содержит те же самые промотор, сигнальную последовательность и сайты сплайсинга, что и M13 VHPCRI (Оrlandi и др. (1989)).
Лигированную ДНК трансформировали в TG1 Е.coli, и от образованных рекомбинантных бляшек получали одноцепочечную ДНК.
В результате были получены PCR-продукты нужного размера, а именно 359 пар оснований (п. о. ) для VH и 288 п.о. для VK. ДНК VH вводили в вектор M13VHPCRI, в результате чего получали М13-ТМ23М и VH. ДНК VK вводили в вектор М13-VKPCRI, в результате чего получали M13 TM23MuUK.
Подтверждение ДНК-последовательностей.
Была подтверждена правильность клонирования ДНК-последовательностей, и было установлено, что при PCR не было введено ложных мутаций. Последовательность одноцепочечной ДНК фага М13, состоящая из 823 п.о. -VH-области, включая промотор, сигнальную последовательность, сайты сплайсинга и 5'- и 3'-нетранслируемые последовательности, была получена дидезокси-методом (Sanger и др., 1977) с использованием ДНК-полимеразы Т7 или Секвеназы.
Последовательность одноцепочечной М13-фаговой ДНК, состоящая из 630 п.о. -VK-области, включая сигнальную последовательность, сайты сплайсинга и 5'- и 3'-нетранслируемые последовательности, была получена дидезокси-методом, как описано выше.
Были идентифицированы ДНК-клоны, содержащие нужную последовательность для VH ТМ23, адаптированную для инсерции в экспрессирующий вектор. Для дальнейших исследований был отобран клон N 2. Введение рестрикционных сайтов, необходимых для инсерции VH в экспрессирующий вектор, приводило к замене одной аминокислоты на N-конце, а именно лизин (K) в положении 5 был заменен на глутамин (G); и к замене 1 аминокислоты на C-конце, а именно серии (S) в положении 108 был заменен на треонин (Т).
Были идентифицированы ДНК-клоны, содержащие нужную последовательность для VK ТМ23, адаптированную для инсерции в экспрессирующий вектор. Один из этих клонов, клон N 4, был отобран для дальнейших исследований. При удалении HindIII-сайта аминокислотная последовательность не изменяясь, однако введение рестрикционных сайтов, необходимых для инсерции VK в экспрессирующий вектор, приводило к замене трех аминокислот на N-конце, а именно: метионин (M) в положении 4 был заменен на лейцин (L), треонин (Т) в положении 7 был заменен на серин (S) и теонин (Т) в положении 8 был заменен на пролин (Р); а также К заменен одной аминокислоты на C-конце, а именно: валин (V) в положении 105 был заменен на глутамин (Е) (эти замены были обусловлены использованием праймеров).
Клонирование ДНК-фрагментов V-области в вектор М13.
Был осуществлен перенос клонированной мышиной вариабельной области в экспрессирующие векторы, содержащие гены человеческой константной области, для последующей экспрессии антитела в клетках млекопитающих.
Отбор изотипа IgG "очеловечивания" антитела.
Изотин IgG1 человека для получения TM27 отбирали из следующих соображений:
1. IgG1 обладает способностью опосредовать антителозависимую клеточноопосредованную цитотоксичность (ADCC) и комплиментзависимую цитотоксичность (CDC). Поэтому этот изoтип может быть более эффективным для супрессии Vβ 5.2/5.3- Т-клеток.
2. IgG1-l более часто используется для "очеловечения" антител.
Отбор каркасных участков, происходящих от антитела человека, в целях продуцирования CDR-трансплантата для "очеловеченного" антитела ТМ23.
Конструирование "очеловеченных" последовательностей вариабельной области TM23.
Проводили отбор человеческих каркасных последовательностей для получения CDR TM23 и определения главных мышиных остатков, которые должны быть замещены в человеческом каркасе. Для этого было проведено автоматизированное (компьютерное) сравнение аминокислотных последовательностей TM23-VH и VK с известными последовательностями других мышиных антител и VH- и VK-последовательностей человеческих антител.
В этих целях была использована программа ALLIGN от DNA STAR Ltd. London W13 9BL, UK, на процессоре Compaq 386. С помощью программы ALLICN сравнивали две первичные белковые последовательности с использованием методов Lipman-Pearson и Needleman-Wunsch. Сначала методом Lipman-Реarson определяли области с наиболее высокой гомологией. Затем методом Needleman-Wunsch проводили конечное сравнение первичных последовательностей.
При сравнительном анализе особое внимание уделяли таким факторам, как длина гипервариабельных участков (CDR), а также остаткам, которые, как предполагается, являются критическими для поддержания конформации CDR, например остатки в положениях 71 и 94 и остатки, близкие к CDR (Tramontano и др. (1990); Foote и Winter (1992)).
В результате сравнения консенсусных последовательностей в соответствии с критерием Кэбета мышиные VH TM23 причисляли к подгруппе Кэбета VK. Причем не было обнаружено аминокислотных последовательностей, которые были бы необычны для этих подгрупп в каркасных участках TM23.
Каркасные последовательности для VH и VK были предоставлены антителами человека NEWM и REl соответственно. В случае VH в дополнение к CDR были использованы остатки 27-30, включительно, и 71, Как предполагается, эти каркасные остатки играют важную роль для сохранения конформации CDR ((Foote Winter (1992)). Остаток 71 может фиксировать относительное расположение между CDR1 и CDR2 в зависимости от того, присутствует ли объемная боковая цепь (лизин или аргинин) или меньшая боковая цепь (валин или аналин) (Tramontano и др. (1990)). Поэтому валин 71 в NEWM-VH был заменен на лизин как в VH TM23. Остатки 27-30 образуют часть структурной, а не гипервариабельной, антигенсвязывающей петли, как было определено Chothia и Lesk (1987). Каркасные остатки 47-49 могут быть также важными для структуры антитела (Foote и Winter (1992)). Поэтому были сконструированы альтернативные очеловеченные тяжелые цепи: одна цепь с изолейцином (1) в положении 48, как в NEWM; а другая цепь с лейцином (L) в положении 48 для того, чтобы были сохранены остатки 47-49 от мышиной H-области. Сравнение мышиной VH-последовательности с "очеловеченным" вариантом проиллюстрировано в примере 2.
В человеческом каркасе RE1, выбранном для VK TM23, не делали каких-либо изменений. Сравнение мышиной VK-последовательности с "очеловеченным" вариантом также проиллюстрировано в примере 2.
Описание экспресирующего вектора pTCSNeo млекопитающих.
Плазмида pTC S Neo содержит уникальный Xhol-сайт клонирования, в который были встроены нужные гены. Транскрипция встроенного гена осуществляется под контролем регуляторных последовательностей, состоящих из энхансера SV40, расположенного выше главного позднего промотора аденовируса 2 (Ad2-MLP). А сам Ad2-MLP расположен выше лидерной последовательности, состоящей из трех частей. Инсертированный ген был фланкирован сигналом полиаденилирования, происходящего от каппа-цепи (Ig) мышиного иммуноглобулина, у 3'-конца. Отбор клеток млекопитающих, несущих нужные гены, обеспечивался присутствием гена резистентности к неомицину (neor). Плазмида также содержала бактериальный сайт инициации репликации и ген β-лактамазы (Ampr).
Конструирование экспрессирующего вектора pTCS Neo.
Два комплементарных олигонуклеотида, имеющих антисмысловую последовательность:
5' CGACATCGATGATATCGAATTCGCGGC CGCCAGCTGGGGCCCTCTAGAC 3'
и смысловую последовательность:
5'CGATGCTAGAGGGCCCCAGCTGGCGGCC GCGAATTCGATATCATCGATG 3'
подвергали отжигу и линировали с pTCSNeo, который был гидролизован ферментом Xhol и застроен dT. Полученная плазмида pTCSLNeo с полилинкером, состоящим из множества рестрикционных сайтов, расположенных в следующем порядке: 5' Xhol Xbal/ Apal/ PvulI/ Notl/ EcoRI/ EcoRV/ Clal 3', была подходящей для инсерции нужного гена. Ген neor может быть с успехом заменен на другой селективный маркерный ген.
Конструирование экспрессирующего вектора pTCSLDHFR*.
Ген neor в плазмиде pTCSLNeo удаляли путем гидролиза ферментами HindIII и BainHI, и снова лигировали с генным фрагментом мышиной мутантной дигидрофолатредуктазы (DHFR*), который был продуцирован путем HindIII- и BamHI-гидролиза плазмиды pSV2-DHFR*, полученной у Dr.Paul Berg (Department of Biochemistry, Stanford University Medical School).
Описание плазмиды pSV 184 H-Neo/DNS-VkCk.
Плазмида pSV184 H-Neo/DNS-VkCk (полученная у Dr.Lee Herzenberg, Stanford University Medical School - см. Thesis of Jeffery Dangl, стр. 48-67) представляет собой химерную конструкцию из мышиной V-области и человеческой C-области, в которой перегруппированный ген V-J сегментов каппа-цепи, выделенный из Dansyl Chloride- специфической мышиной гибридомы, слитый с чел. геномным CK-фрагментом (pSV184 Δ H-Neo-CK).
Выделение константной области каппа-цели иммуноглобулина человека с помощью полимеразной цепной реакции
Для выделения константной области осуществляли полимеразную цепную реакцию с использованием негидролизованной плазмиды pSV184 Δ H-Neo/DNS-VKCK(pSV Huk), которая содержала константную область каппа-цепи человека; и двух праймеров (один - прямой, а другой - обратный). Олигомеры (праймеры) были получены от Орег on. Реагенты для PCR-реакции были взяты из набора для ДНК-амплификации GeneAmp с AmpliTaq (Derkin-Elmer-Cetus). PCR-реакции проводили при трех различных концентрациях Mg2+. Были выбраны аликвоты PCR-продуктов от первого образца Mg2+ (концентрация 1,5 мМ). Для последующего клонирования были сконструированы праймеры, фланкирующие C-область и содержащие рестрикционные сайты. Так, например, амплифицированную ДНК гидролизовали ферментом EcoRI (у 3' -конца). Сайт у 5'-конца (PvulI) не был гидролизован. Затем гидролизованный ДНК-фрагмент подвергали электрофорезу на 1%- агарозном геле, и 300 п.о. -полосу вырезали. ДНК экстрагировали из геля, и небольшую аликвоту анализировали на 1%-ном агарозном геле для количественной и качественной оценки. Была выявлена очищенная 300 п.о.-полоса, а количество ДНК составляло по оценкам ≈ 140 нг.
Константная область каппа-цепи человека, клонированная в вектор pBSKS+ и секвенированная.
EcoRl-гидролизованный ДНК-фрагмент константной области каппа-цепи человека вставляли в вектор pBS KS+, которая была гидролизована ферментами Smal и EcoRI, с использованием ДНК-лигазы Т4. Смесь для лигирования использовали для трансформации компетентных клеток DH5α E.coli. Единичные колонии собирали и использовали для инокуляции 24 жидких культур. От каждой из культур получали ДНК. Каждый ДНК-образец гидролизовали ферментами PvulI (у 5'-конца CK) и ЕсоRI (у 3'-конца CK) и анализировали на 2%-ном агарозном геле. Из-за трудности разделения полос низкого диапазона (300 п.о.) ДНК-образцы гидролизовали только ферментом PvulI. После анализа на 2%-ном агарозном геле 19 образцов из 24 имели правильную CK-вставку. Образцы NN 1-5 (из 19 "правильных" образцов) были выбраны для секвенирования с использованием праймеров T7 и T3 (последовательности в векторе, фланкирующем вставку). Другие реагенты для секвенирования были взяты из набора Sequenase Version 2.0 kit (USB). Было выявлено, что образец N 5 содержит правильную последовательность для константной области каппа-цепи человека.
Константная область каппа-цепи человека, клонированная в экспрессирующий вектор pTCSLDHFR* млекопитающего с образованием вектора pTCSLC D HFR*.
Константную область каппа-цепи человека выделяли из вектора pBSKS (образец N 5) с использованием рестриктирующих ферментов PvulI и ЕсоRI. Однако исходя из вышеуказанной проблемы, связанной с разделением полос, был осуществлен второй гидролиз конструкции pBS/чел. CK. Второй гидролиз осуществляли ферментами EcoRI и Xbal (с 5'-конца PvulI-сайта), в результате чего получали фрагмент ≈ 300 п.о. Затем этот фрагмент подвергали гель-очистке и гидролизовали ферментом PvulI для удаления оставшегося небольшого фрагмента вектора у 5'-конца CK. Этот конечный фрагмент подвергали гель-очистке, а затем анализировали на 1%-ном агарозном геле для количественной и качественной оценки. В результате наблюдалась очищенная полоса СK-ДНК с ≈ 280 п.о. , которая по количественным оценкам составляла ≈ 135 н.г.
Благодаря присутствию в экспрессирующем векторе pTCSLDHFR* трех EcoRl-сайтов, было установлено, что лучше всего клонировать только в PvulI-сайт. Поскольку CK-ДНК получали путем гидролиза ферментами PvulI - EcoRI, то EcoRI-конец репарировали (с образованием тупого конца) с использованием фрагмента Кленова. Репарированные затупленные по концам CK-ДНК
лигировали в вектор pTCSLDHFR*, который был гидролизован ферментом PvulI и обработан фосфатазой (с помощью) телячьей кишечной фосфатазы). Эту смесь для лигирования использовали для трансформации компетентных клеток DH5α Е. соli. Единичные колонии собирали и использовали для инокуляции 24 суспензионных культур. От каждой культуры получали ДНК. Все ДНК-образцы гидролизовали ферментами PvulI и HindIII и анализировали на 1%-ном агарозном геле. В результате этого анализа было выявлено, что 5 из 24 образцов содержали CK-вставку в правильной ориентации. Для последующих исследований был выбран образец N 7.
Замена мутантного гена DHFR геном DHFR дикого типа.
Для экспрессии вектора pTCSLC DHFR* в клетках CHO (dhfr) DUX BII, ген DHFR* заменяли на ген DHFR. ДНК от образца N 7 pTCSLC DHER*, а также pSV2-DHFR (плазмида, полученная у Dr. Paul Berg Department of Biochimistry, Staford University Medical School) гидролизовали рестриктирующими ферментами HindIII и BglII для выделения соответствующих генных фрагментов. После анализа было установлено, что дополнительный сайт в векторе pTCSLCKDHFR также удаляет часть polyA-сайта, который препятствует осуществлению векторной функции. Поэтому, гидролиз повторяли с использованием HindIII и BamHI. (DHFR*) pTCSLCK-часть вектора обрабатывали фосфотазой. Затем оба выделенные фрагменты (pTCSL C и DHFR) подвергали гель-очистке и анализировали на 1%-ном агарозном геле для количественной и качественной оценки. В результате наблюдались очищенные обе полосы, которые имели аналогичную концентрацию, составляющую по оценкам 40-50 нг/мл. Ген DHFR лигировали в вектор pTCSL CK с использованием ДНК-лигазы Т4. Смесь для лигирования использовали для трансформации компетентных клеток DH5 E.coli.
Единичные колонии собирали и использовали для инокуляции 24 суспензионных культур. От каждой культуры получали ДНК. Эти ДНК анализировали путем гидролиза рестриктирующими ферментами HindIII и BamHI. Из 24 культур 23 имели "правильные" полосы, указывающие на инсерцию гена DHFR в вектор pTCSLCK. Для дальнейшего использования выбирали образец N 22.
In vitro-мутагенез для конструирования V-областей с чужеродными CDR и каркасом человека.
Конструирование генов "очеловеченной" вариабельной области.
Ген "очеловеченной" VH-области конструировали посредством сайт-направленного мутагенеза M13- VHPCRI-матрицы, которая содержит синтетический VH-ген с каркасными участками белка NEWM человеческой миеломы (см. Jones и др. 1986). Одноцепочечную ДНК-матрицу получали от M13-VHPCRI, культивированного в штамме PZ1032 E.coli так, чтобы эта ДНК-матрица содержала урацил вместо тимина. Были синтезированы мутагенные олигонуклеотидные праймеры, кодирующие CDR TM23 и приблизительно 15 п.о., которые полностью соответствовали фланкирующей последовательности с какой-либо стороны. Если TM23 CDR-последовательность соответствовала матричной последовательности, то были использованы более короткие олигонуклеотидные праймеры. В качестве праймеров использовали:
CDR1:
5' GGCTGTCTCACCCAGTTTACACCATAG GCGGTTAATGAGAAGCCAGACACGG 3'
CDR2:
5' TGCTGGTGTCCTTTCAGCATTGTCACT CTGGATTTGAGAGCTGAATTATAGTCTGTA TTTCCATCACCCCATATCATTCCAAT/cCCA CTCAAGACC 3'
и CDR3:
5' CCAGTAGTCCATAGCATAGAGGGTAGC CGTAACCCTATCTCTTGCACAATAATAG 3
Олигонуклеотидный праймер для CDR был удлинен у 3'-конца для введения нужных остатков в положения 27-30. Для CDR2 был сконструирован смешанный олигонуклеотид, удлиненный у 3'-конца CDR либо G, либо Т у основания 402 VH-области, что дало 2 альтернативных кодона таких, что в положении 48 VH стал либо L, либо I. Олигонуклеотиды (10 пМ каждый) фосфорилировали 5-ю единицами полинуклеотидкиназы Т4 в полном объеме 50 мкл в течение 1 часа при 37oC. 1 пМ каждого мутагенного праймера и 1 пМ секвенирующего М13-праймера 3 к V-гену гибридизировали с 0,5 мкг матрицы в полном объеме 20 мкл путем нагревания до 90oC в течение 30 секунд с последующим быстрым охлаждением до 70oC и медленным охлаждением до 37oC. Затем гибридизированные праймеры удлиняли и лигировали с участием ДНК-полимеразы Т7 и ДНК-лигазы Т4, после чего инкубировали в течение 1 часа при комнатной температуре. Урацилсодержащую ДНК-матрицу удаляли путем обработки ДНК одной единицей урацил-ДНК-гликозидазы в течение 1 часа при 37oC, в результате чего мутированная нить оставалась интактной. Эту ДНК амплифицировали с помощью PCR-реакции, а затем для проверки, имеет ли продуцированная ДНК нужный размер, подвергали электрофорезу на агарозном геле. ДНК разрезали ферментами HindIII и BamHI и лигировали в HimdIII- и BamHI-сайты Ml3mpl9.
Аналогичным образом, реконструированный ТМ23-VK-ген (ТМ23H и VK) создавали посредством мутагенеза М13-фаговой матрицы, которая содержит ДНК-последовательность VK-гена с каркасными участками белка человека REI Bence - Jones. Эта матрица происходила от вектора VKPCRI М13, который был использован при конструировании химерной легкой цепи с рестрикционными PvulI- и BclI-сайтами, использованными для удаления клонированием. Используемыми праймерами были:
CDR1:
5' TCTGCTTGGTACCAGTTTAAATAATTG CTAATGCCCTGACTTGCACTACAGGTGATG GTC 3'
CDR2:
5' TCTGCTTGGCACACCTGAGTGTAAACT TGATGTGTAGTAGATCAGCAGCTT 3'
и CDR3:
5'CCCTTGGCCGAACGTCCGAGGAAGTTTA CTATACTGCTGGCAGTAGTAG 3'
Были получены PCR-продукты нужного размера (823 п.о. для обеих VH и 620 п.о. для VK) и клонированы в вектор М14 p19.
Идентификация ДНК-последовательностей "очеловеченной" вариабельной области.
Были подтверждены: инсерция нужных CDR; отсутствие ложных мутаций и правильное конструирование "очеловеченных" VH- и VK-генов. Полные вариабельные области секвенировали дидезокси-методом с использованием ДНК-полимеразы Т7 или секвеназы. Полученные последовательности сравнивали со сконструированными последовательностями. Клоны, имеющие ожидаемую последовательность для двух VH, отбирали. Эти клоны были обозначены M13 TM23NMVH и М13 TM23NMVHL. Клон, имеющий ожидаемую последовательность для реконструированной области VK (N 11), отбирали и обозначали M13 - TM23HuVK.
Клонирование перегруппированных V-областей в экспрессирующие векторы миеломы.
(Клонирование "очеловеченных" вариабельных областей ТМ23 в экспрессирующие векторы)
Клонированные "очеловеченные" вариабельные области переносили в экспрессирующие векторы, содержащие гены человеческой константной области, для последующей экспрессии антитела в клетках млекопитающих. Два "очеловеченных" VH-гена были выделены из M13 NTM23VM H и M13 TM23NMVHL в виде HindIII-BamHI-фрагментов и лигированы в HindIII- и BamHI-сайты экспрессирующего вектора тяжелой цепи, pSVgpt. Hul oGl.
"Очеловеченный" VK-ген выделяли из М13 TM23HVK в виде HindIII-BamHI-фpaгмeнтa и лигировали в HindIII- и BamHI-сайты экспрессирующего вектора легкой цепи pSVhVg. HuCK.
Рестрикционный анализ показал, что две альтернативные "очеловеченные" VH были правильно вставлены в вектор тяжелой цепи, в результате чего были образованы плазмиды: pSVgtpTM23NMVH. HulgGI и pSVgptNMVHL.HulgGI.
Аналогичным образом, "очеловеченная" VK-область была правильно встроена в вектор легкой цепи, в результате чего была образована плазмида pSVhygTM23HuVK.Huck.
Ко-трансфекция конструкций H- и L -цепей в клетки NSO.
Ко-трансфекция генов "очеловеченных" тяжелой и легкой цепей.
"Очеловеченные" экспрессирующие плазмиды были перенесены в клеточные линии млекопитающих: NSO и P3-8.653F8879.
"Очеловеченные экспрессирующие плазмиды были совместно трансфецированы в клетки NSO и P3-8.653F8879 с использованием той же самой процедуры, которая была использована для химерных экспрессирующих плазмид.
Каждая из двух "очеловеченных" плазмид тяжелой цепи, pSVgptTM23NMVH. Huckl и pSVgptNMVHL.HulgGI, были совместно трансфецированы с "очеловеченной" плазмидой легкой цепи pSVhygTM23HuVK.Huck, в результате чего были продуцированы клеточные линии, экспрессирующие два альтернативных "очеловеченных" антитела TM23HuVH/HuVK и ТМ23HuVHL/HuVK. Указанные экспрессирующие плазмиды были также ко-трансфецированы с химерными зкспрессирующими плазмидами, в результате чего были продуцированы клеточные линии, экспрессирующие гибридные антитела ТМ23HuVH/MuVK, TM23HuVHL/MuVK и TM23Mu VH/HuVR. Эти гибридные антитела являются ценным инструментом для обнаружения причины любого дефекта связывания тяжелой или легкой цепи в "очеловеченном" антителе.
Через 14 дней после трансфекции обнаружились колонии клеток NSO для всех плазмидных комбинаций. Что касается клеток P3-8.653F8879, то из всех засеянных чашек наблюдалась лишь одна колония, а поэтому эти клетки более не использовались.
Отбор чел. IgG1/каппа-положительных клонов.
Отбор клеток, продуцирующих человеческие антитела.
Были выделены клеточные линии NSO, продуцирующие "очеловеченные" и гибридные ("очеловеченные"/химерные) антитела. Образцы супернатанта, взятые из лунок, содержащих клеточные колонии, скринировали с помощью ELISA.
Клетки ТМ23HuVH/HuVK: -NSO были отобраны из 4 лунок, в которых супернатант культуры давал наиболее высокий уровень сигнала при считывании в ELISA-анализе, D5, D10; C6 и E11. Лунка 5 содержала 2 видимые колонии, а другие лунки - только одну. Однако клеточные линии, продуцированные из этих лунок, не обязательно являются клональными. Клеточные линии из этих лунок, ТМ23HuVH/HuVK D5, D10, C6 и E11, культивировали и замораживали в жидком азоте.
Клетки TM23HuVHL/HuVK:- NSO были отобраны из 4 лунок, в которых культуральный супернатант давал наиболее высокий уровень сигнала при считывании в ELISA (B2, B7, D8 и E9). Лунка E9 содержала 2 видимые колонии, а все остальные лунки - одну. Однако клеточные линии, продуцированные из этих лунок, являются не обязательно клональными. Клеточные линии из этих лунок, ТМ23HuVH/MuVK B2, B7, D8 и E9, культивировали и замораживали в жидком азоте.
Клетки TM23HuVH/HuVK: - NSO были отобраны из 4 лунок, в которых культуральный супернатант давал наиболее высокий уровень сигнала при считывании в ELISA (D9, E6, F10 и G10). Лунки F19 и G10 содержали 2 видимые колонии, а остальные - только одну. Однако клеточные линии, продуцированные из этих лунок, не обязательно являются клональными. Клеточные линии из этих лунок, ТМ23HuVH/MuVK D9, E6, F10 и G10, культивировали и замораживали в жидком азоте.
Клетки TM23HuVHL/MuVKL:- NSO были отобраны из 4 лунок, в которых культуральный супернатант давал наиболее высокий уровень сигнала при считывании в ELISA (B8, С6, D2 и D8). Лунки C6 и D8 содержали 2 видимые колонии, а остальные - только одну. Однако клеточные линии, продуцированные из этих лунок, не обязательно являются клональными. Клеточные линии из этих лунок, TM23HuVHL/MuVK B8, C6, D2 и D8, культивировали и замораживали в жидком азоте.
Клетки TM23MuVH/HuVK: - NSO были отобраны из 4 лунок, в которых культуральный супернатант давал наиболее высокий уровень сигнала при считывании в ELISA (B9, G4, G6 к G11). Все лунки содержали по крайней мере 4 видимые колонии. Все клеточные линии, продуцированные из этих лунок, не обязательно являются клональными. Клеточные линии из этих лунок, ТМ23MuVH/HuVK B9, G4, G6 и G11, культивировали и замораживали в жидком азоте.
Синтез одноцепочечной кДНК для экспрессии в клетках CHO.
Получение цитоплазматической РНК.
Получали PHK для обратной транскрипции. Колбу с клетками NSO ТМ23 Hu VHL/Hu VK, замороженными в жидком азоте, быстро оттаивали при 37oC. Затем клетки постепенно разводили 10 миллилитрами модифицированной по способу Дульбекко среды Игла/среды Хема F12 (DMEM/F12), в которую были добавлены антибиотики к 5%-ная фетальная коровья сыворотка, не содержащая гамма-глобулина (GG-лишенная FBS). Клетки центрифугировали при 1500 об/мин в течение 5 минут, после чего среду удаляли и ресуспендировали в 20 мл свежей среды в 75-см2 колбе. Культуру размножали до 30 мл в 175-см2 колбе. Активно растущие клетки собирали путем центрифугирования и три раза промывали холодным забуференным фосфатом физиологическим раствором (PBS). Цитоплазматическую PHK получали из клеток по методу Favoloro и др. (1980). Всего было получено 350 мкг цитоплазматической РНК.
Синтез кДНК.
Получали одноцепочечную кДНК для VH- и VK-областей. Одноцепочечную кДНК для вариабельной области синтезировали в реакционных смесях, содержащих 5 мкг PHK, 250 мкМ дезоксинуклеотидтрифосфатов (dNTPS), 15 ед. ингибитора рибонуклеазы (Pharmacia RNAguard) и 25 пМ олигонуклеотидного праймера. В качестве праймеров использовали олиго-1109 (для VH):
5' GGGGAAGACCGATGGGCCCTTGGTGGA GGCTGAGGAGACGGTGACC 3'
и олиго-1095 для VK:
5' AGATTTCAGCTGCTCATCAGATGGCGG GAAGATGAAGACAGATGG 3'
Затем реакционные смеси нагревали до 70oC в течение 10 мин и медленно охлаждали до 37oC. После этого добавляли 100 ед. обратной транскриптазы вируса мышиного лейкоза Молони, и реакционные смеси инкубировали в течение 30 минут при 37oC.
кДНК амплифицировали с помощью PCR с использованием вышеуказанных праймеров в комбинации с праймером олиго 1096
для VH:
5 GCCGCTCGACCCTCACCATGGGATGGAG CTGTATCATCCTCTTCTTGGTAG 3
и олиго 1094 для VK:
5 GCCGCTCGAGCCTCACCATGGGATGGAG CTGTATCATCCTCTTCTT CGTAGCAACAGCTACAGGTGTCCACTCCGAC ATCCAGATGACCCAGAG 3.
PCR-продукты анализировали с помощью гель-электрофореза на агарозном геле, и в этом анализе были выявлены ожидаемые размеры ДНК-полос (приблизительно 450 п.о.), которые затем очищали от геля.
Рестрикционный гидролиз.
Для клонирования в векторы CHO, PCR-продукты гидролизовали. VH-PCR-продукт гидролизовали ферментами Xhol и Ара1. VK-PCR-продукт гидролизовали ферментами Xhol и PvulI. Образцы гидролизованных ДНК подвергали электрофорезу на 0,8% агарозном геле. В результате наблюдались полосы предполагаемого размера (450 п.о.).
Выделение V-областей (VHTCS и S и VKS) из кДНК посредством PCR-амплификации VHTCS.
Получали одноцепочечные кДНК-образцы для ТМ27-VK и ТМ27 VH, а также олигонуклеотидные праймеры, используемые для PCR- амплификации этих молекул. Осуществляли PCR с использованием кДНК для TM27-VH соответствующих праймеров и полимеразы Tag. Продукт подвергали электрофорезу на агарозном геле и очищали с использованием набора Geneclean II (Bio101). Выход продукта составлял по оценкам 1,5 нг/мл.
Трансфекция клеток COS плазмидой ps RINeo4HII (легкая цепь) и плазмидой pBJ 14H11 (тяжелая цепь) для выделения кДНК для C-области.
Векторы экспрессии в клетках млекопитающих, PSRINeo и PBHI, были получены от pcDL-SRa296 (Takebe et al., (1988) Mol. Cell. Biol. 8:466), как описано Lin et al., ((1990) Science 249:677), где Xhol-сайт, расположенный между P-промотором и сайтом полиаденилирования 40, был заменен, для клонирования нужного гена, полилинкером, содержащим следующие рестрикционные сайты: 5' Xhol/Xbal/ Sfil/Notl/ EcoPI/ EcoPV/ HindIII/ Clal 3'. B psRlNeo между геном резистентности к ампициллину и SP-промотором был введен ген резистентности к неомицину кДНК, кодирующую лидерную последовательность и V-D-J-область тяжелой цепи ТМ23, начала лигировали с геномной константной областью тяжелой цепи, а затем вставляли в Xhol- и Notl-гидролизованный вектор pBJl, в результате чего генерировали плазмиду pBJ14H11 (тяжелая цепь). Аналогичным образом, кДНК легкой цепи, кодирующую лидерную и V-J-области, сначала лигировали с геномным фрагментом каппа, а затем инсертировали в Xhol/Notl-гидролизованный вектор pSRlNeo, в результате чего генерировали плазмиду pSRlNeo4H11 (легкая цепь). Эти две плазмиды pBJ14Н11 (тяжелая цепь) и pSRlNeo4Н11 (легкая цепь) затем совместно трансфецировали в клетки COS-7 (SV40-трансформированная клеточная линия почек Африканской зеленой мартышки, АТСС CRL 1651) для проведения кратковременной экспрессии.
Выделение константной области IgG1 человека.
кДНК константной области иммуноглобулина IgG1 человека получали из роlу(А)+-РНК, выделенной из трансфецированных клеток COS с помощью полимеразной реакции с использованием праймеров к константной области IgG1, а именно:
прямой праймер:
5' GCGTGACAAGGGCCCATCGGTCTTCCC CCTGGCACCCTC 3',
и обратный праймер:
5' GCGTGACAAGAATTCTCATTTACCCGG AGACAGGGAGACGCTCTT 3'
PCR-ДНК-фрагмент константной области, содержащей EcoRI-сайт у 3'-конца (фланкирующий кодирующую область) и Apal-сайт у 5'-конца (в кодирующей области), клонировали в плазмиду pBS KS+ для субклонирования и секвенирования. Полная константная область была секвенирована с использованием праймеров T3 и Т7 в комбинации с внутренними праймерами для константной области. Клон, содержащий правильную последовательность, был затем субклонирован в плазмиду pTCSLNeo, в результате чего получили плазмиду pTCSLDglNeo.
Конструирование плазмиды pTCSLCgNeo.
Клон, имеющий правильную последовательность в pBS KS+, вырезали с использованием рестриктирующих ферментов EcoRI и Apal, и лигировали в плазмиду PTCSNeo, гидролизованную теми же самыми ферментами. Полученную в результате этого плазмиду pTCSL CglNeo использовали для последующих модификаций в целях конструирования плазмиды pTCSL CgNeoApa.
Удаление Араl-сайта из плазмиды pTCSL СK lNeo для получения плазмиды pTCSLCKlNeoApa.
Вектор экспрессии в CHO, а именно pTCSLCKlNeo, содержал два сайта для рестриктирующего фермента Apal; один сайт был расположен в области полилинкера, используемого для инсерции клонированных фрагментов; а второй сайт был расположен непосредственно у 3'-конца гена neor. Для облегчения последующих стадий клонирования Apal-сайт, расположенный непосредственно возле гена neor, был элиминирован. Плазмиду pTCSLClNeo частично гидролизовали ферментом Apal, и небольшую аликвоту подвергали электрофорезу на агарозном геле для того, чтобы проверить, присутствуют ли линейные молекулы (т.е. плазмиды), разрезанные только в одном из двух Apal-сайтов. Затем гидролизованный материал обрабатывали ДНК-полимеразой Т4, которая селективно удаляла одноцепочечные участки "липких концов", образованные рестриктазой Apal, не расщепляя при этом двухцепочечную ДНК. Затем плазмидам снова придавали кольцевую форму путем лигирования ее концов и трансформировали в компетентные клетки HB101 E.coli. Тридцать трансформантов собирали и использовали для инокуляции суспензионных культур. От каждой культуры получали ДНК, которую затем гидролизовали рестриктазой Apal, и подвергали электрофорезу на агарозном геле. К сожалению, при этих гидролизах плазмидная ДНК подвергалась деградации. Гидролиз ферментом Apal повторяли на 21 образце из 30. В результате было выявлено, что два изолята (N 4 и 12) были линеаризованы ферментом Apal, тогда как век остальные были рестриктированы в двух сайтах. Затем эти два клона-кандидата были гидролизованы рестриктирующими ферментами Apal и HindIII для того, чтобы определить, какой именно из двух вышеуказанных Apal-сайтов присутствует. В этом эксперименте, по желанию, был удален Apal-сайт, расположенный непосредственно у 3'конца близ гена neor в изоляте N 12 pTCSLC INeoApa. Затем были проведены дополнительные рестриктирующие гидролизы, которые подтвердили структуру этого изолята.
Клонирование PCR-выделенных V-областей каппа- и лямбда-цепи человека в векторы CHO, такие как pTCSLCK DHFR и pTCSLCgINeoApa соответственно.
Клонирование Vb-фрагмента ТМ27 в pTCSLCKDHFR.
кДНК, кодирующую V-область каппа-цепи для ТМ27, подвергали гель-очистке, а затем гидролизовали соответствующими рестриктирующими ферментами. Небольшую аликвоту ДНК-образца анализировали с помощью электрофореза на 1%-ном агарозном геле для проведения количественной и качественной оценки. Дополнительную (контаменантную) полосу наблюдали при ≈ 800 п.о., а концентрация по оценкам составляла 1,25 нг/мл. V-область лигировали в вектор pTCSLCK DHFK, который был гидролизован рестриктирующими ферментами Xhol и Pvoll, и обработан фосфатазой. Лигирующую смесь использовали для трансформации компетентных клеток DH5α Е.соli. Выход колоний трансформанта был очень низким. Единичные колонии отбирали и использовали для инокуляции двенадцати суспензионных культур. ДНК, полученную от каждой культуры, гидролизовали ферментом Xhol и ферментом PvulI, а затем анализировали с помощью электрофореза на 1%-ном агарозном геле. Таким образом было получено два положительных образца, #3, и #4. Каждый из этих положительных образцов был секвенирован.
Клонирование VH-фрагмента ТМ27 в pTCSLCK lNeoApa
PCR-продукт, соответствующий PCR-продукту VH-области ТМ27, подвергали гель-очистке, а затем гидролизовали соответствующими рестриктирующими ферментами (Xhol и Apal). Однако, поскольку было замечено, что Apal-гидролиз может быть неполным, то его повторяли. Небольшую аликвоту анализировали с помощью электрофореза на агарозном геле для проведения количественной и качественной оценки. Концентрация полосы, соответствующей VH-области, по оценкам составляла 0,4 нг/мл. Как и в случае V-фрагмента, дополнительную полосу наблюдали при ≈ 800 п.о., что давало возможность предположить, что чистота этого материала была не такой, как ожидалось. VH-область лигировали в вектор pTCSLCKINeo-Ара, который был гидролизован рестриктирующими ферментами Xhol и Apal, и обработан фосфатазой. Лигирующую смесь использовали для трансформации компетентных клеток DH5α Е.Соli. Было выделено очень небольшое количество трансформантов. Шесть колоний отбирали и использовали для инокуляции суспензионных культур. Минипрепарат ДНК получали от каждой культуры, гидролизовали ферментами Xhol и EcoRI, а затем анализировали посредством электрофореза на агарозном геле. В результате первого лигирования был выделен один клон-кандидат pTCSL Hu VH4HCKlNeoApa (изолят # 6). В целях получения еще нескольких клонов-кандидатов (в случае, если один изолят # 6 не будет содержать правильную последовательность в последующих тестах) осуществляли дополнительные лигирования с использованием: 1) VH-PCR-продукта, повторно очищенного на легкоплавкой агарозе для удаления высокомолекулярной контаминирующей полосы, и 2) VH-PCR-продукта, синтезированного с использованием кДНК и праймеров. В результате вышеописанных реакций лигирования было проанализировано 8 дополнительных трансформантов и выделено 5 клонов-кандидатов: изоляты #9, 10, 11, 12 и 14.
V-Области клонированной последовательности на обеих цепях для подтверждения предполагаемой ДНК-последовательности.
Праймеры, сконструированные для облегчения ДНК-секвенирования V-областей, вставленных в векторы pTCSLCKlNeoApa и pTCSL CKDHFR, были получены от фирмы Operon Research. Прямой праймер был локализован внутри pTCS-векторных последовательностей и был использован для секвенирования V-вставок в обоих векторах pTCSLCKINeoApa и pTCSLCKDHFR. Обратные праймеры были расположены внутри CK или СKI-последовательностей и были использованы для секвенирования и H-вставок соответственно.
Двухцепочечное секвенирование V-области каппа-цепи человека.
Два образца от ТМ23 (N 3 и N 4) секвенировали по обеим цепям. Обратную цепь полностью секвенировали между рестрикционными сайтами (3' - 5'; PhulI - Xhol) с использованием праймера HDCK5RR. Последовательность прямой цепи была определена с использованием двух различных праймеров, PTCSFOR и PTCSFWO. Полноразмерная последовательность между рестрикционными сайтами (5' - 3'; Xhol - Prull) была получена путем объединения выходов последовательностей от каждого из праймеров. Наблюдалось присутствие небольшой зоны компрессии в прямых нитях, которые не разрешались ни с помощью праймеров, ни с помощью метода секвенирования, который устраняет такие компрессии. Однако было установлено, что обратная нить имела правильную последовательность в этой зоне. Было также обнаружено, что оба образца # 3 и # 4 имели правильную последовательность, а образец #4 отбирали для трансфекции клеток млекопитающего.
Двухцепочечное секвенирование V-области гамма-цепи ТМ27.
VH-фрагменты pTCSLHuVH4HC lNeApa (изоляты # 6 и 9) были полностью секвенированы по обеим нитям. VH-фрагмент ("очеловеченный") изолята # 6 соответствовал ожидаемой последовательности на протяжении всей кодирующей области. Наблюдалось замещение одной пары оснований (C на T) в положении -2 (соответствующему инициирующему кодону ATG), которая, как ожидалось, не влияет на экспрессию. VH-фрагмент изолята # 9 содержал два замещения в кодирующей области. Четыре дополнительных клона-кандидата не были секвенированы.
Котрансфекция клеток COS и CHO ТМ27-плазмидами и контрольными плазмидами.
Реципиентными клетками были клетки DUX BII dhfr CHO и COS-1. Клеточные линии DUX BII CHO получали от Bi ogenl с.
Для трансфекции CHO 5 микрограмм ТМ27-содержащих плазмид pTCSLHuVK 4HCKDHFR (изолят # 4), pTCSL HuVH4HCKlNeoApa (изолят # 6), и векторы pTCSLDHFRApa и pTCSL NeoApa, - линеаризировали путем гидролиза рестриктирующим ферментом Aat II. ТМ27-содержащие плазмиды (меченые ТМ27L) подвергали копреципитации в этаноле и ресуспендировали в H2O. Аналогично, плазмидные векторы (меченый контроль) подвергали копреципитации, а затем ресуспендировали в H2O. Каждую из совместно осажденных плазмид трансфецировали в 107 клеток CHO путем электропорации с использованием импульсного генератора BioRad Gene при 250 B и 960 мФ. Трансфецированные клетки культивировали в минимальной поддерживающей среде (αMEM) с добавлением тимидина, аденозина и дезоксиаденозина (неселективная среда) в течение двух дней, а затем проводили отбор.
Для трансфекции COS 5 микрограмм неразрезанных ТМ27 плазмид и векторных плазмид подвергали копреципитации в этаноле, а затем ресуспендировали в H2О. Плазмиды были трансфецированы в 4,1 • 106 клеток COS путем электропорации. Трансфецированные клетки культивировали в модифицированной по методу Дульбекко среде Игла (DMEM) в течение 3 дней, а затем супернатанты собирали для последующего анализа.
Анализ супернатантов трансфектантов COS путем ELISA-анализа на IgG1 человека и окрашивание.
Продуцирование IgG человека клетками COS оценивали c использованием ELISA на IgG1 человека с помощью супернатантов клеточных культур. Планшеты для микротитрования сенсибилизировали мышиным античел. IgG1 Fc-антителом в PBS и блокировали в блокирующем буфере (1% BSA в PBS). Супернатанты добавляли в 100 мкл образцов; соответствующие разведения в блокирующем буфере анализировали, если это необходимо. Захваченный IgG детектировали с помощью конъюгированного с пероксидазой хрена козьего антитела к Igk человека, и окрашивали о-фенилендиамином (OPD). Супернатанты от TM27L-трансфецированных клеток COS показали на уровни чел. IgG в 95 нг/мл. Чел. IgG от контроль-трансфецированных клеток COS не являются детектируемыми. Проточную цитометрию осуществляли на супернатантах от ТМ27L-трансфецированных клеток COS при ≈ 6-кратной концентрации. Интенсивность окрашивания на клетках HPB, как показала среднеканальная флуоресценция, была приблизительно в 9 раз ниже, чем интенсивность, полученная с очищенными TM27L (очищенные из клеток NSO) при ≈ 2,5 мг/мл. На клетках Jurkat окрашивание было на фоновом уровне.
Отбор CHO-трансфектантов.
В целях отбора CHO-трансфектанты помещали в среду MEM, не содержащую нуклеотидных добавок (для отбора DHFR+ на pTCSL HuVK4HCKDHFR), и среду, не содержащую Genetican (G418: для отбора neo на pTCSLD HuVH4HCKINeoApa). Клетки высевали в 24-луночные планшеты при 2,0 • 105 клеток/мл.
Анализ супернатантов CHO-трансфектантов, проведенный с помощью ELISA на IgG1 человека.
После выдерживания в течение 12 дней в селективной среде супернатанты от конфлуэнтных лунок собирали и анализировали с помощью ELISA на чел. IgG1/каппа. Концентрации в ТМ27L-супернатантах находились в пределах от 182 до 236 нг/мл. В большинстве случаев концентрация составляла более 200 нг/мл. В контрольных супернатантах чел. IgG1/каппа не обнаруживалась. Клетки от двенадцати высокопродуцирующих ТМ27-лунок переносили для размножения в Т25-сосуды. Единичные сосуды были заморожены в качестве резервных культур, а затем их хранили в присутствии жидкого азота. Клетки от двух контрольных лунок были также перенесены для размножения в Т25-сосуды и заморожены.
Первое культивирование при серийном разведении CHO-трансфектантов.
Четыре наиболее высокопродуцирующие лунки отбирали для культивирования при серийном разведении в 96-луночных планшетах: лунки # 1B1 (232,6 нг/мл), 1D6 (233,3 нг/мл), 2B2 (235,5 нг/мл), и 2C1 (236,0 нг/мл). Для каждой лунки в трех планшетах клетки культивировали в следующих селективных средах: одну при 1 клетку/лунку и две при 0,5 клеток/лунку.
Супернатанты от первой серии клонирования были скринированы посредством ELISA на чел. IgG1/каппа. Клетки от 90 высокопродуцирующих лунок переносили для размножения в 24-луночные планшеты, и культивировали до состояния сплошности. Супернатанты от 12 из этих культур анализировали с помощью ELlSA на присутствие чел. IgG1/каппа и с помощью проточной цитометрии. Концентрации в ELISA на присутствие чел. IgG1/каппа составляли от 211 до 1048 нг/мл. Все образцы окрашивали клетки HPB (среднеканальная флуоресция составляла от 46,5 до 169,9) и не окрашивали клетки Jurkat. Шесть наиболее высокопродуцирующих клона размножали и замораживали (как резервные) в жидком азоте. Два из шести наиболее высокопродуцирующих клона (1B7-C7 и 2B2-H9) отбирали для проведения последующих серий клонирования.
Две первичных культуры контрольных конструкций также вводили посредством одной серии культивирования при серийном разведении в 96-луночные планшеты. Один клон каждой из культур отбирали и размножали. Супернатанты анализировали, и в анализе ELISA на чел. IgG1/каппа не было обнаружено заметного сигнала (стр. 640-83). Клетки замораживали и хранили в жидком азоте.
Размножение неклонированных культур в роллер-флаконе для очистки антитела.
Две первичные культуры (1B1 и 2В2) размножали для продуцирования достаточного количества материала для очистки антитела ТМ27. Клетки переносили в 2-литровые роллерные флаконы для размножения и адаптировали для более низких концентраций сыворотки (Т25 в 10% сыворотке ---> Т75 в 10% сыворотке --->3 х Т75 в 5% сыворотке ---> 3 х 2-литровые флаконы в 1% сыворотке) (стр. 640-24). 2-литровые флаконы оставляли на 1 день и подпитывали (путем замены 1/2 объема свежей средой с 1%-ной сывороткой) на 2-ой, 5-ый и 8-ой дни. Первый сбор проводили на 10-ый день; второй сбор проводили на 12-ый день, а третий сбор проводили на 15-ый день. Собранные супернатанты подвергали центрифугированию и хранили при -20oC до очистки.
Очистка антитела на колонке с белком A.
Супернатанты от культур 2В2 собирали и объединяли (полный объем 2,7 литров), а затем фильтровали через найлоновые мембраны 0,22 микрон. ELISA-анализ на чел. IgG1 показал, что полное количество исходного материала составляло 1090 мг. Антитело очищали на колонке с Prosep-А (Белок А), элюированной 0,1 М цитратом при pH 5,0, 4,0 и 3,0. Элюаты диализовали в PBS. Количества очищенного антитела в элюированных фракциях измеряли с помощью чел. IgG1-ELISA. pH 3,0-фракция содержала наибольшее количество очищенного антитела, тогда как pH 4,0-фракция содержала небольшие количества антитела. Чел. IgG1-ELISA-анализ повторяли для получения более точных концентраций. Полный выход ТМ27 (pH 3,0 + pH 4,0-фракции) составлял 960 мг.
Очистка антител из супернатантов культуры трансфектантов NSO на колонке с Белком А.
Антитела, содержащиеся в супернатантах культуры трансфектантов NSO, были очищены на колонке с белком А способом, в основном описанным выше.
Характеризация антитела ТМ27, продуцированного миеломной клеточной линией NSO.
Антитела (mAb) ТМ27, очищенные из культуральных супернатантов NSO-трансфектантов посредством хроматографии на колонке с Белком А, использовали для окрашивания клеток HPB-ALL ( Vβ 5.2) и клеток Jurkat ( Vβ 8.1) в качестве негативного контроля по сравнению с мышиными mab, ТМ23 и химерными ТM27. ТМ27 окрашивались положительно на клетках HPB-ALL, но не на клетках Jurkat.
Для определения специфичности ТМ27 нормальные Т-клетки PBL окрашивали ТМ27 и для сравнения ТМ23 (mAb). Оба антитела ТМ27 и ТМ23 окрашивали приблизительно 3,0 % от общего числа Т-клеток PBL. Был проведен конкурентный анализ, в котором фиксированную концентрацию FTC-меченых mAb ТМ23 добавляли к различным концентрациям немеченых mAb 4Н11 или ТМ27. Эту смесь антител использовали для окрашивания клеток HBP-ALL. Оба немеченых ТМ23 и ТМ27 блокировали окрашивание FITC-меченым ТМ23. Для того чтобы определить, способно ли антитело ТМ27 к ко-модуляции с антигеном CD3, клетки HPB-ALL инкубировали с различными концентрациями ТМ27 или ТМ23 в течение ночи, а затем окрашивали моноклональным антителом против CD3. Полученные результаты показали, что ТМ27, а также ТМ23 могут вызывать эндоцитоз TCP/CD3-комплексов.
Для определения афинности связывания антитела ТM27 на клетках HPB-ALL осуществляли анализ по Скэтчарду. Результаты обоих анализов, т.е. анализа по Скэтчарду и конкурентного анализа (с mAb ТМ23), показали, что ТМ27 сохранило почти такую же самую аффинность (Kd = 2,0•10-8М-1) и может с успехом конкурировать с mAb 4Н11.
Было также проведено сравнение TM271L/NSO и TM271L/CHO в конкурентном анализе с 4Н11, и результаты этого анализа представлены на фиг. 3. Был проведен анализ по Скэтчарду, результаты которого проиллюстрированы на фиг. 4.
Второе культивирование при серийном разведении CHO-трансфектантов.
Культивирование при серийном разведении проводили так, как описано выше. Лунки скринировали с помощью чел. IgG1-ELISA-анализа. Клетки из шести лунок каждого (1В1-С7 и 2В2-Н9) размножали в 24-луночных планшетах, и анализировали с помощью чел. IgG1-ЕLISA. Два наиболее высокопродуцирующих клона от каждой концентрации размножали и замораживали в жидком азоте. Наиболее высокопродуцирующие клоны от каждой концентрации отбирали для последующей серии субклонирования.
Третье культивирование при серийном разведении CHO-трансфектантов.
Культивирование при серийном разведении проводили так, как описано выше. Лунки скринировали с помощью ч. IgG1-ELISA-анализа. Клетки из шести лунок каждой концентрации (1В1-С7 и 2В2-Н9) размножали в 24-луночных планшетах и анализировали с помощью ч. IgG1-ELISA. Три наиболее высокопродуцирующих клона от каждой концентрации размножали во флаконах Т25 и замораживали в жидком азоте. Единичный клон от каждой концентрации размножали в колбах Т75. На этой стадии из среды удаляли Geneticin, что приводило к устранению селекции на neоr. Селекция на HFP сохранялась. Супернатанты анализировали с помощью ч. IgG1-ELISA. Клон 1В1-С7 продуцировал 4,69 мг/106 клеток в день. Клон 2В2-Н9 продуцировал 2,65 мг/106 клеток в день. Поэтому в качестве конечного клона был выбран изолят С7, а изолят Н9 служил в качестве альтернативного клона.
Получение замороженного клеточного материала.
Конечный клон, отобранный, как описано выше, размножали во флаконах Т225 для получения достаточного количества клеток в целях создания банка замороженного клеточного материала. Затем клетки собирали из этих флаконов и объединяли. Объединенная суспензия содержала 98,3% жизнеспособных клеток. Полное число жизнеспособных клеток составляло 1,58 • 10x, что оказалось достаточным для получения нужного числа 72 флаконов при концентрации 2 • 106 кл/флакон. Клетки были заморожены и обозначены "ТМ27L-662-35". Кроме того, альтернативный клон H9 размножали во флаконе Т22 для создания небольшого банка замороженных клеток. Клетки, собранные из этого флакона, были на 97,3% жизнеспособны. Суспензия содержала достаточное количество клеток для получения 9,7 флаконов при концентрации 2 х 10 кл./флакон (стр. 662-89). Эти клетки замораживали в жидком азоте и обозначали "TM27L-662-89".
Анализ на восстановление жизнеспособности клеток после замораживания и анализ на загрязнение микоплазмой.
Один флакон замороженного клеточного материала ТМ27L-662-35 оттаивали и восстанавливали в DMEM. Как было определено путем окрашивания трипановым синим, жизнеспособность клеток составляла 88%. Конфлюэнтную культуру, культивированную из этого флакона, анализировали на загрязнение микоплазмой с использованием набора Bionique Testing Laborato2 ries. B этом варианте не было обнаружено какого-либо загрязнения. Кроме того, оттаивали один флакон замороженного клеточного материала TM27L-662-89. Как было определено путем окрашивания триптановым синим, жизнеспособность клеток составляла 93,8%. Конфлюэнтную культуру, культивированную из этого флакона, анализировали на загрязнение микоплазмой, как указано выше. В этом анализе не было обнаружено какого-либо загрязнения. Обе культуры размножали во флаконах Т75, и супернатанты от клеточных культур анализировали с помощью чел. IgG1-ЕLISA-анализа. Клон TM27L-662-35 продуцировал 3,24 мкг/106 кл/день, а клон TM27L-662-89 продуцировал 2,52 мкг/106 кл./день. Во втором анализе клон TM27L-662-35 продуцировал 3,03 мкг/106 кл/день, а клон TM27L-662-89 продуцировал 2,74 мгк/106 кл./день.
Экспрессия клона TM27L-662-35 в роллер-флаконе для очистки антитела.
Культуру ТМ2L -662-35 размножали для продуцирования достаточного количества материала для очистки антитела TM27L. Клетки переносили в 2-литровые роллер-флаконы для размножения, и адаптировали для более низкой концентрации сыворотки (Т25 при 10% сыворотке ---> Т75 при 10% сыворотке ---> 3 х Т75 при 5% сыворотке---> 3 х 2-литровые флаконы при 1% сыворотке), 2-литровые флаконы оставляли на 1 день, и подпитывали (путем замены 1/2 объема свежей средой с 1% сывороткой) на 2-й, 5-й, 7-й и 9-й день. Первый сбор проводили на 12-й день, второй сбор проводили на 14-й день, и третий сбор проводили на 16-й день. Собранные супернатанты подвергали центрифугированию, фильтровали и хранили при -70oC до очистки.
Очистка антитела на колонке с белком А.
Супернатанты от культур TM27L-662-35 объединяли с получением общего объема 5,1 литров. Антитело связывали на колонке с Ргоsер (белок А) и элюировали 0,1 M цитратом при pH 5,0 и 3,0. 3,0-элюат диализовали в PBS. Количество очищенного антитела в исходном материале и элюированные фракции оценивали с помощью чел. IgG1/каппа-ЕLISA-анализа. pH 3,0-фракция содержала приблизительно 3,9 мг антитела в 25 мл. Этот материал концентрировали на концентраторе Centriprep-30 (Amicon), и повторяли чел. IgG1/каппа-ELISA.
Полный выход TM27L составлял ≈ 3,2 мг при 1,6 мг/мл.
Предварительная характеризация очищенного антитела
Примерно 2,5 мг материала от каждой из pH 3,0- и pH 4,0-фракций подвергали проточной цитометрии, и ТМ27 выделяли из клеток HSO. Как было определено с помощью среднеканальной флуоресценции, интенсивность окрашивания на клетках HPB составляла: 283,46 для pH 3,0-фракции; 321,98 для pH 4,0-фракции; 506,79 для NSO-происходящих ТМ27. На клетках Jurkat окрашивание для всех образцов наблюдалось на фоновом уровне.
Примеры 2-5 см. в конце описания.
Пример 6. Общая методика для ТМ29.
* Гибридомы получали, в основном, так, как описано в Примере 1, и кДНК выделяли (как в Примере 5).
* Выделенные кДНК (H и каппа) (из гибридомы 16G8) клонировали в векторы М13.
* Чел. каркасную область (VHKOL/VKRE1) вводили посредством мутагенеза.
* V-области со встроенными CDR клонировали в миеломные экспрессирующие векторы.
* "Очеловеченные" антитела (≈ 1 мг каждого) очищали из NSO-трансфектантов для предварительной оценки.
Было получено четыре различных ТМ29:
ТМ29
ТМ29.1 с заменой SS на AA (23-24).
TM29.2 с заменой S на P (75).
ТМ29.3 с заменами GV/F на AM/У (92-93/95).
Подробное описание эксперимента - ТМ29
Выделение константной области каппа-цепи человека с помощью PCR-метода
Для выделения константной области полимеразную цепную реакцию осуществляли с помощью двух праймеров (одного прямого и одного обратного), с использованием негидролизованной плазмиды pSV184DH-Neo/DNS-VKCK (pSVHuk, содержащей константную область каппа-цепи человека). Олигомеры (праймеры) получали от Operon. Реагенты для полимеразной цепной реакции брали из набора реагентов для ДНК-амплификации GeneAmp с использованием Amplitaq (Perkin-Elmer Cetus). Полимеразные цепные реакции проводили с тремя различными концентрациями Mg2+. Аликвоты PCR-продуктов анализировали с помощью электрофореза на 1% агарозном геле. PCR-продукт выбирали из первого образца Mg2+ (концентрация 1,5 мМ).
В целях клонирования праймеры, фланкирующие C-область, конструировали так, чтобы они содержали рестриктирующие сайты. Таким образом, амплифицированную ДНК гидролизовали ферментом EcoRI (у 3'-конца). Сайт у 5'-конца (PvulI) не гидролизовали. Гидролизованный ДНК-фрагмент подвергали электрофорезу на 1%-ном агарозном геле, и 300 п.о.-полосу вырезали. Затем ДНК экстрагировали из геля, и небольшую аликвоту анализировали с помощью электрофореза на 1% агарозном геле для проведения количественной и качественной оценки. Выделенную 300 п.о.-полосу очищали, и полное количество ДНК оценивали (приблизительно 140 нг).
Клонирование в вектор pBS KS+ и секвенирование константной области каппа-цепи человека.
Гидролизованный ферментом EcoRI ДНК-фрагмент константной области каппа-цепи человека вводили в вектор pBS KS+, который был гидролизован ферментами Smal и EcoRI, с использованием ДНК-лигазы ТЧ. Эту лигирующую смесь использовали для трансформации компетентных клеток DH5α E.coli. Одиночные колонии отбирали и использовали для инокуляции 24 суспензионных культур. ДНК получали от каждой культуры. Каждый образец ДНК гидролизовали ферментом PvulI (5'-конец CK) и ферментом EcoRl (3'-конец CK), а затем анализировали посредством электрофореза на 2% агарозном геле. Из-за трудности разделения полос с более низким диапазоном (300 п.о.) образцы ДНК гидролизовали лишь ферментом PvulI. После анализа, проведенного с помощью электрофореза на 2% агарозном геле, 19 из 24 имели правильную CK-вставку. Образцы N 1 - 5 (из 19 правильных образцов) отбирали для секвенирования с использованием праймеров Т7 и Т3 (последовательности в векторе, фланкирующем вставку). Другие реагенты для секвенирования брали из набора Sequenase Version 2,0 (USB). Рассмотренные образцы N 5 имели правильную последовательность константной области каппа-цепи человека.
Клонирование константной области каппа-цепи человека в экспрессирующей вектор млекопитающего - pTCSLD HFR* с получением pTCSLCKDHFR*.
Константную область каппа-цепи человека выделяли из вектора KS+ (образец 5) с использованием рестриктирующих ферментов PvuII и EcoRl. Однако из-за трудности разделения полос, второй цикл гидролиза осуществляли с использованием конструкции, состоящей из pBS/CK человека. Этот гидролиз осуществляли с помощью ферментов EcoRI и Xbal (локализация 5'-сайт PvulI), и получали фрагмент (около 300 п.о.). Этот фрагмент очищали посредством гель-фильтрации и подвергали гидролизу ферментом PvulI для удаления оставшегося небольшого фрагмента вектора у 5'-конца CKС. Полученный конечный фрагмент подвергали очистке с помощью гель-фильтрации, а затем анализировали посредством электрофореза на 1% агарозном геле для проведения количественной и качественной оценки. Выделенная ДНК-CK представляла собой очищенную полосу (≈ 280 п.о.), а полная количественная оценка показала ≈ 135 нг.
Из-за присутствия трех EcoRI-сайтов в векторе экспрессии pTCSLDHFR*, наиболее предпочтительно осуществлять клонирование лишь в PvulI-сайт. После того как был получен PvulI-ЕсоRI-ДНК-фрагмент (CK-вставка), EcoRI-конец репарировали (с получением тупого конца) с использованием фрагмента Кленова. Репарированную и затупленную по концам ДНК-CK лигировали в вектор pTCSLDHFR*, который гидролизовали ферментом PvulI, и обрабатывали фосфатазой (использовали кишечную фосфатазу теленка). Эту лигирующую смесь использовали для трансформации компетентных клеток DH5α Е.cоli. Единичные колонии отбирали и использовали для инокуляции 24 суспензионных культур. ДНК получали из каждой культуры. Все образцы ДНК гидролизовали ферментом PvulI и ферментом HindIII, а затем анализировали с помощью электрофореза на 1% агарозном геле. Было установлено, что 5 из 24 образцов содержали правильную вставку CK. Образец N 7 был отобран для дальнейшего использования.
Замена мутантного гена DHFR на ген DHFR дикого типа.
В целях экспрессии вектора pTCSLCKDHFR в клетки CHO- (dhfr) DUX BII, ген DHFR заменяли на ген DHFR. ДНК от образца N 7 pTCSLCk - DHFR, а также pSV2-DHFR (плазмиды, полученной из Dr.Paul. Berg; Отделение биохимии медицинской школы Стенфордского университета) гидролизовали рестриктирующими ферментами HindIII и BglII для выделения соответствующих генных фрагментов. В результате анализа было установлено, что дополнительный BglII-сайт в векторе pTCSLCK - DHFR удалял также часть Poly(A)-сайта, препятствующего реализации функции вектора. Поэтому гидролиз повторяли с использованием ферментов HindIII и BamHI. Часть (DHFR)pTCSCK вектора обрабатывали фосфатазой. Затем оба выделенных фрагмента (pTCSL CK и DHFR) очищали с помощью гель-фильтрации и анализировали посредством электрофореза на 1%-ном агарозном геле для проведения количественной и качественной оценки. Обе выделенных полосы подвергали очистке и оценивали аналогичным способом (концентрация ≈ 40-50 нг/мл). Ген DHFR лигировали в вектор pTCSLCK с использованием ДНК-лигазы Т4. Лигирующую смесь использовали для трансформации компетентных клеток DH5 E. coli. Единичные колонии отбирали и использовали для инокуляции 24 суспензионных культур. ДНК получали от каждой культуры. После этого ДНК анализировали путем гидролиза рестриктирующими ферментами HindIII и BamHI. 23 культуры из 24 имели правильные полосы, что указывало на введение гена DHFR в вектор pTCSLCK. Образец N 22 отбирали для дальнейшего использования.
Клонирование PCR-выделенной V-области каппа-цепи человека в вектор pTCSLCK-DHFRD CHO.
кДНК V-области каппа-цепи для 16G8 очищали с помощью гель-фильтрации, а затем гидролизовали соответствующими рестриктирующими ферментами. Небольшую аликвоту образца ДНК анализировали с помощью электрофореза на 1% агарозном геле для определения количественной и качественной оценки. Дополнительную (с примесью) полосу наблюдали при ≈ 800 п.о., а концентрацию оценивали при 1,25 нг/мл. V-область лигировали в вектор pTCSLCK - DHFR, который гидролизовали рестриктирующими ферментами Xhol и PvulI, а затем обрабатывали фосфатазой. Лигирующую смесь использовали для трансформации компетентных клеток DH5α Е. coil. Выход колоний-трансформантов был очень низок. Единичные колонии отбирали и использовали для инокуляции 12 суспензионных культур. ДНК получали от каждой культуры, а затем гидролизовали ферментами Xhol и PvulI и анализировали посредством электрофореза на 1% агарозном геле. Были выделены четыре положительных образца: N 1, N 4, N 7, и N 10. Каждый положительный образец секвенировали. PCR-выделенную V-область гамма-цепи ТМ29 клонировали в вектор pTCSLCg-lNeoApa CHO.
Двухцепочечное секвенирование V-области каппа-цепи человека.
Четыре образца от 16G8 (N 1, N 4, N 7, и N 10) секвенировали по обеим цепям. Обратную связь полностью секвенировали на участке между рестрикционными сайтами (3' - 5'; PvulII - Xhol) с использованием праймера HDCK5PR. Образцы N 1 и N 4 имели правильную последовательность, образец N 7 не имел последовательности, а образец N 10 имел неправильную последовательность. Для образцов N 1 и N 4 последовательность с прямой цепью определяли с использованием двух различных праймеров, PTCSFOR и PTCSFWD. Полноразмерную последовательность на участке между рестрикционными сайтами (5' - 3'; Xhol-PvulI) получали путем объединения последовательностей, полученных на выходе из каждого из праймеров. В прямых цепях наблюдалась небольшая область сжатия, которая не разрешалась ни с одним из праймеров, ни с использованием какого-либо метода секвенирования, который бы устранял такие сжатия. Однако рассмотренные обратные цепи имели правильную последовательность в этой области. Полученные образцы N 1 и N 4 имели правильную последовательность вместе с образцом N 4, отобранным для использования в целях трансфекции клеток млекопитающего.
Ко-трансфекция в клетки Сos-1 и CHO плазмид, кодирующих легкую и тяжелую цепь ТМ29
Клетки Cos-1 и CHO(dnfr)DUX BII использовали в котрансфекции плазмид, кодирующих легкую и тяжелую цепь ТМ29. Клеточную линию DUX BII CHO получали методом Dr. Lawrence Chaisin (Колумбийский университет) и закупали у фирмы Biogen Inc.
Для трансфекции клеток Cos-1 5 мг каждой негидролизованной плазмиды, кодирующей ТМ29 (pTCSLHuVK-16GCK - DHFR и pTCSLHuVH-16GCK -lNeoApa; ТМ-29-4 и ТМ29-26 соответственно) осаждали в этаноле и ресуспендировали в H2O при концентрации 1 мг/мл. После этого плазмиды ко-трансформировали в 3,8 • 106 клеток путем электропорации, с использованием импульсного генератора BioRao Gene при 250 В и 960 mF. Трансфецированные клетки ресуспендировали в модифицированной по методу Дульбекко среде Игла (DMEM), и вводили в культуру, которую оставляли на три дня. Контрольные плазмидные векторы (pTCSLDHFPApa и pTCSLNeoApa) получали и ко-трансфецировали аналогично плазмидам, кодирующим ТМ29. Затем трансфецированные клетки ресуспендировали и культивировали аналогично ТМ29- трансфецированным клеткам Cos.
В целях трансфекции клеток DUX В11 CHO 5 мг каждой плазмиды, кодирующей ТМ29 (как описано выше), гидролизовали рестриктирующим ферментом Aat II, и получали линейный фрагмент ДНК, который осаждали в этаноле и ресуспендировали в H2О при концентрации в 1 мг/мл. Затем плазмиды ко-трансфецировали в 5 • 106 клеток путем электропорации, как описано выше. Трансфецированные клетки ресуспендировали в минимальную поддерживаемую среду (DMEM), содержащую тимидин, аденозин и дезоксиаденозин (неселективная среда), а затем помещали в культуру, которую оставляли на три дня. Полученные контрольные плазмидные векторы (описанные выше) ко-трансфецировали аналогично плазмидам, кодирующим ТM29. После этого трансфецированные клетки ресуспендировали, и культивировали аналогично ТМ29-трансфецированным клеткам CHO.
Анализ TM29-тpaнcфeциpoвaного COS-клеточного супернатанта с помощью ELISA человеческого IgG1k.
После отсеивания в культуре в течение 3 дней были получены ТМ-29-трансфецированные клетки COS в виде сплошной массы. Поэтому супернатант собирали, а клетки отбрасывали.
Продуцирование человеческого IgG клетками определяли из супернатанта культуры с использованием ELISA человеческого IgGK. IgG супернатанта, связанный с Fc-фрагментом мышиного античеловеческого IgG1, которым сенсибилизировали 96-луночный планшет, детектировали с помощью конъюгированного с пероксидазой хрена козьего человеческого антитела IgK, а развитие окраски осуществляли с использованием о-фенилендиамина (OPD). Концентрации IgG супернатанта определяли путем сравнения измерений оптической плотности с измерениями кривой, построенной для очищенных человеческих антител IgG1K в известной концентрации. Супернатант от ТМ29-трансфецированных клеток COS давал концентрацию человеческого IgG в 201,3 нг/мл. Супернатант от ДНК-трансфецированных контрольных клеток Cos давал концентрацию человеческого IgG в нг/мл.
Отбор ТМ29-трансфецированных клеток CHO.
После отстаивания в культуре в течение 3 дней ТМ29-трансфецированные клетки CHO достигали состояния сплошности. Поэтому клетки обрабатывали 0,25%-ным трипсином (в сбалансированном солевом растворе Хэнкса) для отделения их из флаконов с культурами. Эти клетки собирали, а затем центрифугировали, и супернатант удаляли. Клеточный осадок ресуспендировали в селективной среде αMEM при концентрации в 2•105 клеток/мл, а затем высевали в 24-луночный планшет в концентрации 1 мл/лунку. Селективная среда αMEM имела двойную избирательную способность, причем в ней отсутствовали нуклеотидные добавки, a FBS была заменена на диализованную FBS для отбора на присутствие, DHFR+ на pTCSL HuVK- 16GCK-DHFR, и был добавлен генетицин (G-418) для отбора на присутствие neor на pTCSL HuVH-16GCK-lNeoApa-.
Оценка человеческого IgGIK с помощью ELISA-анализа супернатанта от ТМ29-трансфецированных клеток CHO и с помощью проточной цитометрии.
После выдерживания в течение 14 дней в селективной среде супернатанты от TM29-трансфецированных клеток CHO собирали из каждой лунки и анализировали с помощью ELISA-анализа человеческого IgG1K. Концентрация человеческого IgG в супернатантах от ТМ29-трансфецированных клеток CHO находилась в пределах от 144,6 нг/мл до 291,9 нг/мл, а средняя концентрация составляла 240,5 нг/мл. Концентрация человеческого IgG в супернатантах от контрольных ДНК-трансфецированных клеток CHO составляла 0 нг/мл.
Супернатанты из трех наиболее высокопродуцирующих лунок ТМ29-трансфецированных клеток CHO (2С3, 2С4, 2В6) и одной лунки для контрольных ДНК-трансфецированных клеток CHO (3D4) подвергали обработке с помощью проточной цитометрии. Три ТМ29-образца, 2С3, 2С4 и 2В6, окрашивали позитивно клетки Jurkat (среднеканальная флуоресценция составляла 186,61-290,01) и не окрашивали позитивно клетки HPB (среднеканальная флуоресценция составляла 11,77-13,82). Контрольный образец 3D4 не окрашивал позитивно клетки Jurlat (среднеканальная флуоресценция составляла 11,96).
Клонирование ТМ29-трансфецированных клеток CHO путем серийного разведения.
Три наиболее высокопродуцирующих лунки ТМ29-трансфецированных клеток CHO (2С3, 291,9 нг/мл; 2С4, 289,7 нг/мл; 2В6, 289,5 нг/мл) отбирали для клонирования путем серийного разведения. Для каждой из лунок концентрации в селективной среде αMEM и высевали в 96-луночный планшет (при концентрации 2 клетки/лунку, 1 клетка/лунку и 0,5 клетки/лунку). В течение десяти дней после клонирования за ростом колоний наблюдали в пронумерованных лунках каждого (0,5 клетки/лунку) планшета для 2С3, 2С4 и 2В6. Поэтому супернатанты от всех лунок каждого из планшетов с 0,5 клет. на лунку собирали и анализировали путем ELISA-анализа на человеческий IgG1K. Все лунки, способствующие клеточному росту, были позитивными (ELISA). Все 24 наиболее высокопродуцирующие лунки (10 из 2С3 и 14 из 2В6) отбирали для последующего размножения в 24-луночный планшет и культивировали до сплошности. Супернатанты собирали от каждой лунки и анализировали с помощью ELISA на человеческ. IgG1K. Концентрация человеческого IgG составляло 0,7 мг/мл - 4,8 мг/мл, со средней концентрацией 2,9 мг/мл. Супернатанты от 6 наиболее высокопродуцирующих лунок для ТM29-трансфецированных клонов CHO (1С7, 1D9, 1G2, 2G1, 2G10, 2Н2а) анализировали с помощью проточной цитометрии. Все 6 образцов окрашивали позитивно клетки Hurkat (среднеканальная флуоресценция составила 313,68 - 356,87) и не окрашивали позитивно клетки HPB (среднеканальная флуоресценция составила 7,59-8,16). Каждый из шести образцов помещали в колбу размером 25 см2, а затем замораживали и хранили в атмосфере жидкого азота. Три наиболее высокопродуцирующие лунки (2C3-1G2, 2C3-2G10, 2С3-2Н2а) были отобраны для субклонирования.
Одну лунку для контрольных ДНК-трансфецированных клеток CHO (3D4) клонировали путем серийного разведения, аналогично ТМ29-лункам. При разведении концентрации клеток в 10 раз ниже, чем это необходимо, рост колоний не наблюдался уже через 10 дней после клонирования. Поэтому свежую культуру инициировали из замороженного сосуда (3D4). Клонирование путем серийного разведения повторяли с использованием этой новой культуры. Через 16 дней после клонирования шесть из наиболее конфлуэнтных лунок из планшетов с 0,5 клеток/лунку (1В5, 1С11, 1D6, 2Е7, 2F3, 2F9) отбирали для размножения в 24-луночном планшете. Для последующего размножения содержимое каждой лунки переносили в 25 см2 колбу, а затеи в 75 см2 колбу и культивировали до состояния сплошности. Супернатанты собирали из каждой колбы и анализировали с помощью ELISA на присутствие IgG1K человека. Концентрация IgG человека составляла 0 нг/мл. Каждую колбу замораживали и хранили в присутствии жидкого азота.
Размножение неклонированных ТM29-трансфецированных культур клеток CHO в роллер-флаконе.
Три наиболее высокопродуцирующие лунки для ТМ29-трансфецированных клеток CHO (2С3, 291,9 нг/мл; 2С4, 189,7 нг/мл; 2В6, 289,5 нг/мл) также отбирали в качестве неклонированных культур для размножения в 25 см2 колбе. В это время один сосуд с каждой из культур ТМ29 (а также один сосуд с контрольной ДНК-трансфецированной клеточной культурой CHO) замораживали и хранили в присутствии жидкого азота. Для размножения каждую из неклонированных культур переносили в 75 см2 колбу. Из одной 75 см2 колбы каждую культуру переносили в 375 см2 колбы, снижая при этом концентрацию сыворотки в среде αMEM от 10 до 5%. Каждую из 75 см2 колб для обеих 2С3 и 2В6 затем переносили в 2-литровый роллер-флакон. Все содержимое 375 см2 колбы для 2С4 замораживали и хранили в присутствии жидкого азота. В роллер-флаконе, содержащем одну из культур, концентрацию сыворотки в среде αMEM снова понижали от 5 до 1%. После отстаивания в течение 9 дней в культуре 2В6-флаконы имели полную сплошность, в то время как 2С3-флаконы имели около 25% сплошности. Супернатанты от всех 2В6 - и 2С3-флаконов собирали на 9 день, на 14 день и на 16 день, после чего клетки отбрасывали. Собранные супернатанты центрифугировали и хранили при температуре -20oC. Аликвоту каждого из трех собранных супернатантов для 2В6 и 2С3 анализировали с помощью ESILA на присутствие IgG1K человека. Затем проводили полную количественную оценку IgG человека для 2В6. Таким образом, она составляла 820 мг для 2В6, и 1923 мг для 2С3.
Субклонировавие ТM29-трансфецированных клонов клеток CHO путем серийного разведения.
Три наиболее высокопродуцирующих лунки для ТМ29-трансфецированных клонов клеток CHO (2C3-1G2, 4,8 мг/мл; 2С3- 2H2a, 4,1 мг/мл; 2C3-2C10, 4,0 мг/мл) отбирали для субклонирования путем серийного разведения. Уже через 14 дней после субклонирования рост колоний наблюдался в пронумерованных лунках каждого из планшетов с 0,5 клеток/лунку для 1G2, 2G10 и 2H2а. Поэтому супернатанты от тех лунок, где наблюдался рост колоний более быстро, собирали и анализировали с помощью ELISA на присутствие IgG1K человека. Все лунки, усиливающие клеточный рост, были позитивными в ELISA-анализе, и эффективность клонирования составляла 100%. Все 24 наиболее высокопродуцирующие лунки (2 из 2Н2а, 5 из 2G10, 17 из 1G2) отбирали для перенесения их в 24-луночный планшет и культивировали до состояния сплошности. Супернатанты от каждой лунки собирали и анализировали с помощью ELISA-анализа на присутствие чел. IgG1K. Концентрация IgG человека составляла 1,2 мг/мл - 9,2 мг/мл при средней концентрации 4,2 мг/мл. Супернатанты от четырех наиболее высокопродуцирующих лунок для ТМ29-трансфецированных субклонов CHO (2G5, 2H12, 1Н6 2Е9) также анализировали посредством проточной цитометрии. Все четыре образца окрашивали позитивно клетки Jurkat (среднеканальная флуоресценция составляла 211,51 - 292,45) и не окрашивали позитивно клетки HPB (среднеканальная флуоресценция составляла 8,36 -8,80). Каждый из этих четырех образцов переносили в 25 см2 колбу.
Второе субклонирование ТМ29-трансфецированных клонов клеток CHO путем серийного разведения.
Два наиболее предпочтительных ТМ29-трансфецированных клона клеток CHO (2C3-1G2-2G5 и 2C3-1G2-1H6) на основании общих результатов, полученных от ELlSA-анализа и проточной цитометрии, отбирали для второго субклонирования путем серийного разведения. В это время один сосуд с каждым из клонов (а также один сосуд с каждым из 2C3-1G2-2H12 и 2С3-1G2-2Е9) замораживали, а затем хранили в присутствии жидкого азота. Уже через 12 дней после субклонирования рост колоний наблюдался в пронумерованных лунках каждого из планшетов с 0,5 клетками/лунку для 2G5 и 1H6. Поэтому супернатанты от лунок, обеспечивающих рост колоний, собирали, а затем анализировали с помощью ELISA на присутствие IgG1K человека. Все лунки, обеспечивающие рост клеток, были позитивными в ELISA-анализе, в результате чего эффективность клонирования составляла 100%. Все 24 наиболее высокопродуцирующие лунки (20 из 2С5, и 4 из 1Н6) отбирали для их перенесения в 24-луночный планшет (# 646-101) и культивировали до состояния сплошности. Супернатанты собирали из каждой лунки и анализировали с помощью ELISA-анализа на присутствие IgG1K человека. Вследствие потери активности стандартным IgG1K человека, используемым в данном анализе, концентрации для неизвестных образцов не могли быть определены. Поэтому наиболее высокопродуцирующие лунки были оценены по их оптической плотности. Шесть наиболее высокопродуцирующих лунок (5 из 2G5 и 1 от 1Н6) отбирали для их перенесения в 25 см2 колбы с последующим перенесением в 75 см2 колбы, а затем культивировали до состояния сплошности. Супернатанты собирали из каждой колбы и анализировали с помощью ELlSA-анализа на присутствие IgG1K человека. Концентрация IgG человека составляла 14,6 мг/мл - 21,2 мг/мл, со средней концентрацией 18,6 мг/мл. После этого супернатанты из 6 колб для ТМ29-трансфецированных субклонов клеток CHO (1В5, 1D1, 1F11, 2В10, 2F5, 2F9) анализировали с помощью проточной цитометрии (≈ 646-122). Все шесть образцов окрашивали позитивно клетки Jurkat (среднеканальная флуоресценция составляла 304,51 - 383,0) и не окрашивали позитивно клетки HPB (среднеканальная флуоресценция составляла 8,67-10,25).
Получение замороженной исходной клеточной культуры для ТМ29-трансфецированного клона клеток CHO.
ТМ29-трансфецированный клон клеток CHO, отобранный для получения замороженного исходной клеточной культуры, представлял собой 2C3-1G2-1H6-2F5. Этот клон переносили из одной 75 см2 колбы в другую 225 см2 колбу, в результате чего получали достаточное число клеток для создания банка замороженных клеток, предназначенного для их исследования. Банк замороженных клеток получали с использованием SOP-TMB1-001-00. Эти клетки обрабатывали трипсином, собирали от каждой колбы и объединяли. Объединенная клеточная суспензия содержала 3,2 • 10x клеток с 99% жизнеспособностью. Таким образом, были получены 72 флакона, каждый из которых содержал 2 • 106 клеток в объеме 1 мл. Все флаконы для клонов метили ТM29-646-132.
Примеры 7-13 см. в конце описания.
| название | год | авторы | номер документа |
|---|---|---|---|
| Н- И L-ЦЕПИ МОНОКЛОНАЛЬНОГО АНТИТЕЛА РМ1 (МОНАТ) К РЕЦЕПТОРУ IL-6R ЧЕЛОВЕКА И ИХ V-ОБЛАСТИ, МОДИФИЦИРОВАННАЯ МОНАТ, ЕГО Н- И L-ЦЕПИ И ИХ V-ОБЛАСТИ, CDR- ПОСЛЕДОВАТЕЛЬНОСТИ, ДНК-ПОСЛЕДОВАТЕЛЬНОСТИ | 1992 |
|
RU2139351C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА IGA2m1-ИЗОТИПА В КЛЕТКАХ МЛЕКОПИТАЮЩИХ | 2022 |
|
RU2801178C1 |
| Способ получения димерной формы иммуноглобулина IgA1-изотипа в клетках млекопитающих | 2023 |
|
RU2822890C1 |
| РЕКОМБИНАНТНОЕ АНТИТЕЛО, СПЕЦИФИЧНОЕ К CD4, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ПСОРИАЗА | 1992 |
|
RU2170256C2 |
| Способ получения димерной формы мутантного иммуноглобулина IgA2m1-изотипа в клетках млекопитающих | 2023 |
|
RU2822889C1 |
| ШТАММ КЛЕТОК ЯИЧНИКОВ КИТАЙСКОГО ХОМЯЧКА - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО АНТИТЕЛА ПРОТИВ ФАКТОРА НЕКРОЗА ОПУХОЛИ АЛЬФА ЧЕЛОВЕКА | 2014 |
|
RU2556816C1 |
| СТРОЕНИЕ ЭКСПРЕССИОННОГО ВЕКТОРА, НОВЫЕ СПОСОБЫ ПОЛУЧЕНИЯ КЛЕТОК-ПРОДУЦЕНТОВ И ИХ ПРИМЕНЕНИЕ ДЛЯ РЕКОМБИНАНТНОГО ПОЛУЧЕНИЯ ПОЛИПЕПТИДОВ | 2012 |
|
RU2628703C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBiPr-ABIgA2m1F16-ht ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА А ИЗОТИПА IGA2m1 | 2016 |
|
RU2671477C2 |
| CD3-ЭПСИЛОН-СВЯЗЫВАЮЩИЙ ДОМЕН С МЕЖВИДОВОЙ СПЕЦИФИЧНОСТЬЮ | 2008 |
|
RU2561457C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ ХИМЕРНОЕ АНТИТЕЛО ПРОТИВ ФАКТОРА НЕКРОЗА ОПУХОЛИ-АЛЬФА ЧЕЛОВЕКА, ЛИНИЯ ЭУКАРИОТИЧЕСКИХ КЛЕТОК-ПРОДУЦЕНТ ХИМЕРНОГО АНТИТЕЛА И СПОСОБ ПОЛУЧЕНИЯ ХИМЕРНОГО АНТИТЕЛА | 2013 |
|
RU2555533C9 |




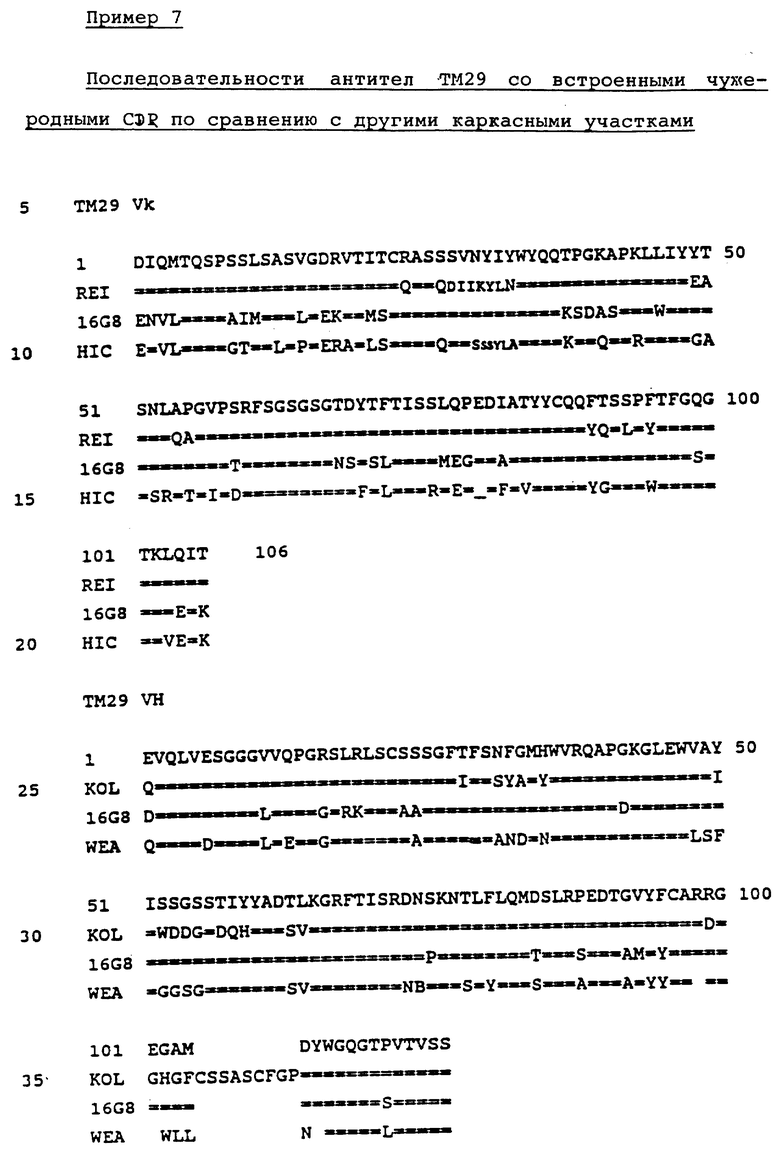







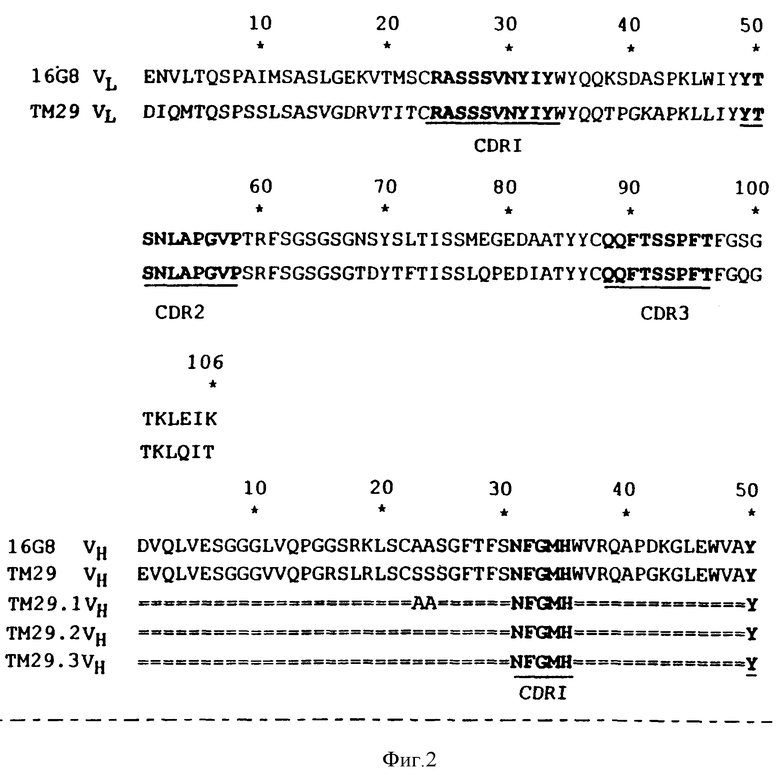

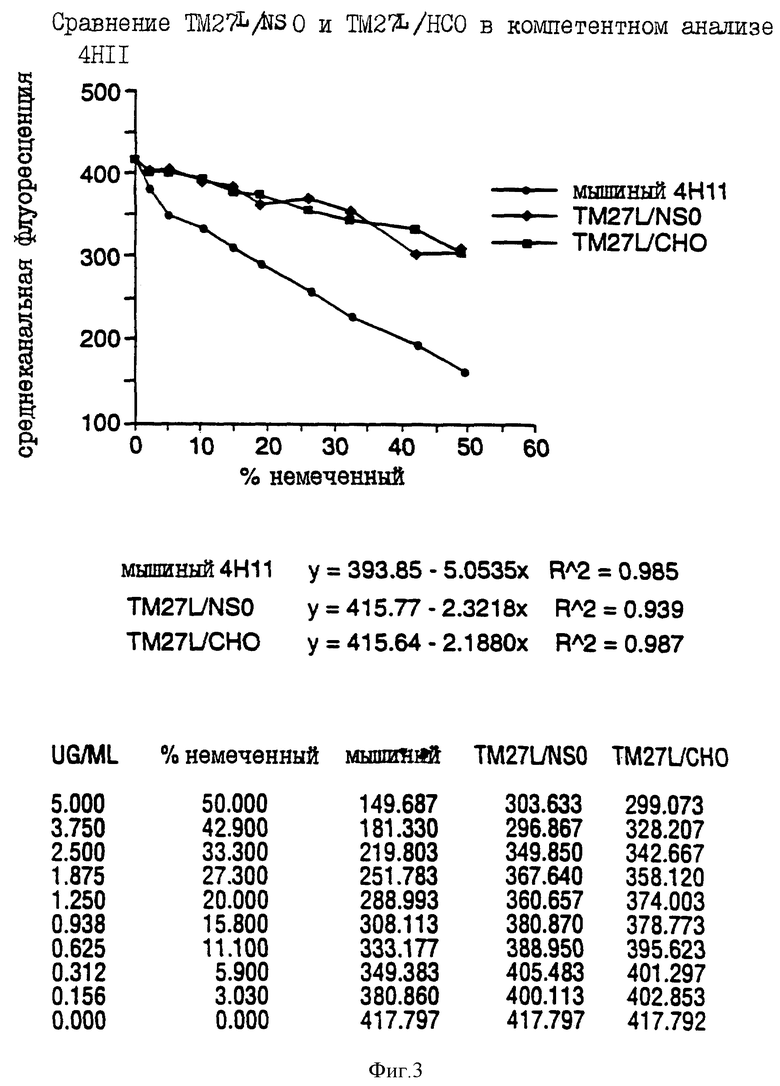
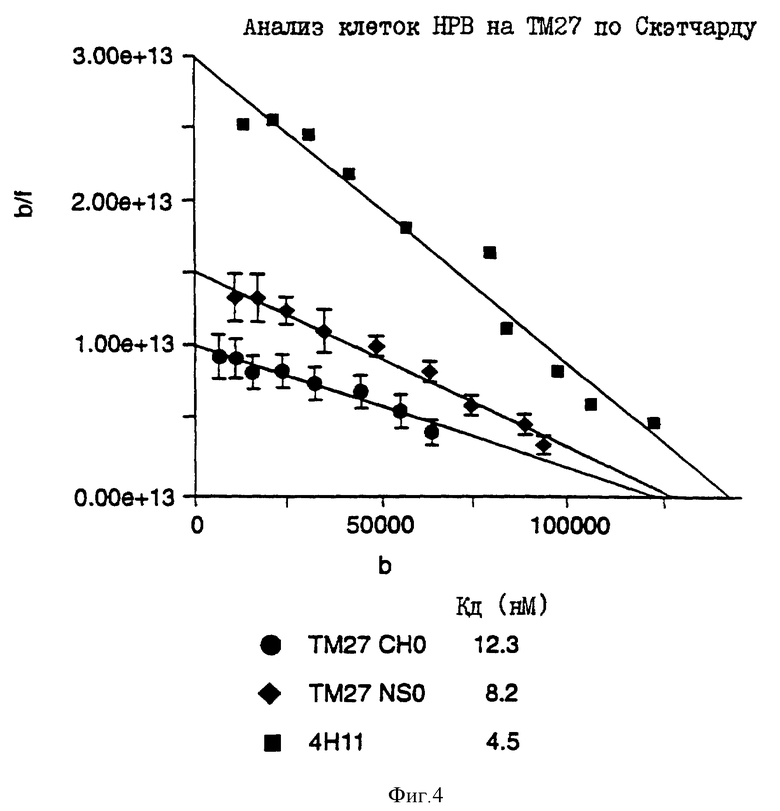
Изобретение относится к гуманизированным антителам и к их связывающим белкам, способным связываться с Т-клетками, имеющими определенные вариабельные бета-цепи. В частности, с субпопуляциями клеток, экспрессирующих Vβ 5,2 и/или 5,3 и Vβ 8,1 человека. Изобретение характеризует получение таких антител, фармацевтические композиции, содержащие эти антитела, и терапевтическое использование этих антител. Использование изобретения позволяет создать новые лекарства для лечения аутоиммунных заболеваний. 9 с. и 10 з.п.ф-лы, 4 ил.

и кроме того содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотной последовательности TM27 VH (SEQ 1D N:4) :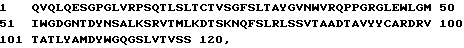
последовательности TM27 VH, в которой аминокислотный остаток 48 заменен на изолейцин (I) (SEQ ID N:6), последовательности TM27 VH, в которой аминокислотные остатки 78 и 79 представляют собой валин (V) и фенилаланин (F) (SEQ ID N:7), последовательности TM27 VH, в которой аминокислотные остатки 67 - 70, имеющие последовательность VTML, заменены на LSIS соответственно, а аминокислотный остаток 73 представляет собой аспаргин (N) (SEQ ID N:8), и последовательности TM27 VH, в которой аминокислотный остаток 92 представляет собой аргинин (R) (SEQ ID N:9).
и дополнительно включает аминокислотную последовательность, выбранную из группы, состоящей из последовательности TM29 VH: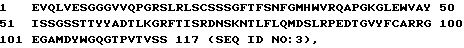
последовательности TM29 VH, в которой оба аминокислотных остатка 23 и 24 заменены на аланиновые (A) остатки (SEQ ID N:44), последовательности TM29 VH, в которой аминокислотный остаток 75 заменен на пролин (P) (SEQ ID N:45) и последовательности TM29 VH, в которой аминокислотные остатки 92 и 93 заменены на аланин (A) и метионин (M) соответственно, а аминокислотный остаток 95 заменен на тирозин (Y) (SEQ ID N:46).
и аминокислотную последовательность TM27 VH (SEQ ID N:4):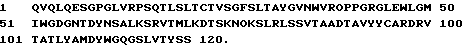
10. Гуманизированное антитело, имеющее аминокислотную последовательность, включающее гипервариабельные участки (CDR) моноклонального антитела, обладающего специфичностью к выбранной субпопуляции T-клеток, экспрессирующих вариабельную область Vβ 8.1 бета-цепи T-клеточного рецептора антигена в сочетании с выбранными вариабельными каркасными остатками, также происходящими из указанного моноклонального антитела, где гуманизированное антитело включает аминокислотную последовательность TM29 Vk:
и аминокислотную последовательность TM29 VH: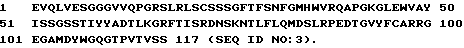
11. Гуманизированное антитело или его связывающий белок по п.1 или 2 для использования в терапевтических целях.
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| Огнетушитель | 0 |
|
SU91A1 |
| Способ получения динуклеотидов | 1983 |
|
SU1404512A1 |
