Изобретение относится к вариабельной области (V-области) мышиных моноклональных антител к рецепторам человеческого интерлейкина-6 (IL-6R), человеческим/мышиным химерным антителам к человеческим IL-6R и измененным человеческим антителам, включающим человеческие антитела, в которых области, определяющие комплементарность (CDRS) V-области человеческих легких цепей (L-цепей) и V-области человеческих тяжелых цепей (H цепей) перенесены в CDRS мышиных моноклональных антител к человеческим IL-6R. Кроме того, настоящее изобретение представляет ДНК, кодирующую вышеупомянутые антитела или их части. Настоящее изобретение к тому же представляет векторы, особенно векторы экспрессии, содержащие вышеназванную ДНК, и клетки-хозяева, трансформированные или трансфицированные вышеназванным вектором. Настоящее изобретение дополнительно еще представляет способ получения химерных антител к человеческому IL-6R и способ получения измененных человеческих антител к человеческому IL-6R.
Интерлейкин-6 (IL-6) является полифункциональным цитокином, который продуцируется рядом клеток. Он регулирует иммунный ответ, реакции острой фазы и гематопоэз и может играть центральную роль в механизмах защиты организма-хозяина. Он действует на широкий ряд тканей, оказывая вызывающее рост, подавляющее рост и вызывающее дифференциацию воздействие зависимости от природы клеток-мишеней. Специфические рецепторы для IL-6 (IL-6R) экспрессируются на лимфоидных, так же как и на нелимфоидных клетках в соответствии с полифункциональными свойствами IL-6. Было высказано предположение, что ненормальная экспрессия IL-6 гена включена в патогенез многих заболеваний, особенно аутоиммунных заболеваний, мезангиального пролиферативного гломерулонефрита и плазмацитомы/миеломы (смотрите обзор Hirdno et al., Immunol. Today 11, 443-449, 1990). Наблюдается, что человеческие миеломные клетки продуцируют IL-6 и экспрессируют IL-6R. В экспериментах антитела против IL-6 подавляли in vitro рост миеломных клеток, показывая таким образом, что аутокринный регуляторный замкнутый контур управляет онкогенезом человеческих миелом (Kawano et al., Nature, 332, 83, 1988).
IL-6R присутствуют на поверхности различных животных клеток и специфически связываются с IL-6, и о числе молекул IL-6R на клеточной поверхности сообщалось (Taga et al, J. Exp. Med. 196, 967, 1987). Кроме того, кДНК, кодирующая человеческий IL-6R была клонирована, и о первичной структуре IL-6R сообщалось (Yamasaki et al., Science 241, 825, 1988).
Мышиные антитела высоко иммуногенны у людей и по этой причине их терапевтическое значение для людей ограничено. Полупериод присутствия мышиных антител in vitro у людей относительно короток. Кроме того, мышиные антитела невозможно применить с многократным дозированием без образования иммунного ответа, который не только препятствует ожидаемой эффективности, но также создает риск побочной аллергической реакции у пациента.
Чтобы разрешить эти проблемы были разработаны методы получения очеловеченных мышиных антител. Мышиные антитела могут быть очеловечены двумя путями. Более простым методом является создание химерных антител, когда V-области происходят из первоначальных мышиных моноклональных антител и C-области происходят из подходящих человеческих антител. Получаемые в результате химерные антитела содержат целые V-области исходных мышиных антител, и можно ожидать связывания антигена с той же самой специфичностью, что и у исходных мышиных антител. Кроме того, химерные антитела имеют существенное снижение процентного содержания белковой последовательности, происходящей из нечеловеческого источника, и поэтому ожидается, что они будут менее иммуногенны, чем исходные мышиные антитела. Хотя химерные антитела, как предсказано, хорошо связываются с антигеном и менее иммуногенны, иммунный ответ на мышиные V-области может все же присутствовать (Lo Buglio et al, Proc. Natl. Acad. Scl, USA 84, 4220-4224, 1989).
Второй способ очеловечивания мышиных антител более сложен, не более интенсивно снижает потенциальную иммуногенность мышиных антител. При этом методе области, определяющие комплементарность (CDRS) из V-областей мышиных антител пересаживаются в человеческие области для создания "видоизмененных" человеческих V-областей. Эти измененные человеческие V-области затем соединяются с человеческими C-областями. Единственными частями конечных измененных человеческих антител, образованными нечеловеческими протеиновыми последовательностями, являются CDRS, CDRS состоит из высоковариабельных протеиновых последовательностей. В них не обнаружено видоспецифических последовательностей. По этим причинам преобразованные человеческие антитела, несущие мышиные CDRS, не должны быть более иммуногенны, чем естественные человеческие антитела, содержащие человеческие CDRS.
Как видно из вышеприведенного, предполагается, что измененные человеческие антитела будут применяться для терапевтических целей, но преобразованные человеческие антитела к человеческим IL-6R неизвестны. Более того, не существует способа получения видоизмененных человеческих антител, универсально применимого для любых конкретных антител. Поэтому, чтобы получить вполне активные видоизмененные человеческие антитела к конкретному антигену, необходимы различные способы. Даже хотя и были получены мышиные моноклональные антитела к человеческому IL-6R, т.е. PM1 и MT18 (Японская патентная заявка N 2-189420), и авторы настоящего изобретения получили мышиные моноклональные антитела к человеческому IL-6R, т.е. AUK 12-20, AUK 64-7 и AUK 146-15, авторы настоящего изобретения не знают публикаций, которые представляют создание видоизмененных человеческих антител к человеческому IL-6R.
Авторы настоящего изобретения обнаружили также, что когда мышиные моноклональные антитела к человеческим IL-6R вводили безволосым мышам, которым трансплантировались миеломные клетки человеческой линии, рост опухоли заметно подавлялся. Это дает основание полагать, что антитела к рецепторам человеческого IL-6 применимы в качестве терапевтического средства для лечения миеломы.
Поэтому настоящее изобретение направлено на получение менее иммуногенных антител к человеческим IL-6R. Соответственно настоящее изобретение представляет видоизмененные человеческие антитела к человеческим IL-6R. Настоящее изобретение также представляет часть видоизмененного человеческого антитела, так же как и системы экспрессии для продукции видоизмененных человеческих антител и их частей и химерные антитела.
Более конкретно, настоящее изобретение представляет V-область L-цепи мышиных моноклональных антител к человеческим IL-6R и H-цепь V-области мышиных моноклональных антител к человеческим IL-6R.
Настоящее изобретение также представляет химерные антитела к человеческим IL-6R, включающие:
(1) L-цепь, содержащую человеческую C-область L-цепи и V-область L-цепи мышиного моноклонального антитела к IL-6R, и
(2) H-цепь, содержащую человеческую C-область H-цепи и V-область H-цепи мышиного моноклонального антитела к человеческому IL-6R.
Настоящее изобретение также представляет CDR V-области L-цепи мышиного моноклонального антитела к человеческому IL-6R, и CDR V-области H-цепи мышиного моноклонального антитела к человеческому IL-6R.
Настоящее изобретение, сверх того, представляет видоизмененную человеческую V-область L-цепи антитела к человеческому IL-6R, включающую:
(1) остовные области (FRS) человеческой V-области L-цепи и
(2) CDRS V-области L-цепи мышиных моноклональных антител к человеческим IL-6R и
видоизмененную человеческую V-область H-цепи антител к человеческим IL-6R, включающую:
(1) FRS человеческой V-области H-цепи и
(2) CDRS V-области H-цепи мышиных моноклональных антител к человеческим IL-6R.
Настоящее изобретение также представляет видоизмененную человеческую цепь антител к человеческим IL-6R, включающую:
(1) человеческую C-область L-цепи и
(2) V-область L-цепи, включающую человеческие FRS, и CDRS мышиных моноклональных антител к человеческим IL-6R и
видоизмененную H-цепь антитела к человеческим IL-6R, включающую:
(1) человеческую C-область H-цепи и
(2) V-область H-цепи, содержащую человеческие FRS и CDRS из мышиного моноклонального антитела к человеческим IL-6R.
Настоящее изобретение к тому же еще представляет видоизмененное человеческое антитело к человеческому IL-6R, содержащее:
(A) L-цепь, включающую
(1) человеческую C-область L-цепи и
(2) V-область L-цепи, включающую человеческие FRS L-цепи и CDRS L-цепи мышиных моноклональных антител к человеческим IL-6R, и
(B) H-цепь, содержащую
(1) человеческую C-область H-цепи и
(2) V-область H-цепи, включающую FRS H-цепи и CDRS H-цепи мышиного моноклонального антитела к человеческим IL-6R.
Настоящее изобретение, кроме того, представляет ДНК, кодирующую полипептиды любого из вышеупомянутых антител или его части.
Настоящее изобретение также представляет векторы, например векторы экспрессии, включающие вышеназванную ДНК.
Настоящее изобретение к тому же представляет клетки-хозяева, трансформированные или трансфицированные вышеназванными векторами.
Настоящее изобретение еще представляет процесс получения химерных антител к человеческим IL-6R и процесс получения видоизмененных человеческих антител к человеческим IL-6R.
Фиг. 1 представляет векторы экспрессии, включающие промоторно/энхансерную систему человеческого цитомегаловируса (HCMV), применимую для экспрессии пептида настоящего антитела.
Фиг. 2 является графиком, показывающим результаты ELISA для подтверждения способности настоящего химерного антитела AUK 12-20 связываться с человеческим IL-6R.
Фиг. 3 является графиком, показывающим результат определения способности настоящего химерного антитела AUK 12-20 подавлять связывание IL-6R с человеческим IL-6R.
Фиг. 4 является графиком, показывающим результат ELISA по связыванию настоящих химерных антител PM-1a и PM-1b с человеческим IL-6R.
Фиг. 5 является графиком, показывающим определения по ELISA способности настоящих химерных антител PM-1a и PM-1b подавлять связывание IL-6 с человеческим IL-6R.
Фиг. 6 является диаграммой строения первого варианта видоизмененной V-области H-цепи человеческого PM-1.
Фиг. 7 является диаграммой строения первого варианта видоизмененной человеческой V-области L-цепи PM-1.
Фиг. 8 представляет процесс создания плазмиды экспрессии HEF-12h-gγI, включающей человеческий промотор/энхансер фактора элонгации Iα (HEF-Iα ), применимого для экспрессии H-цепи.
Фиг. 9 представляет процесс создания плазмиды экспрессии HEF-12k-gk, включающей систему промотора/энхансера HEF-Iα, применимую для экспрессии L-цепи.
Фиг. 10 представляет процесс создания плазмиды экспрессии DHFR-RMh-gγI, включающей промотор/энхансер HCMV и ген дигидрофолатредуктазы (dhfr), связанный с дефектной SV40 промоторной/энхансерной последовательностью для амплификации, применимой для экспрессии H-цепи.
Фиг. 11 представляет процесс создания плазмиды экспрессии DHFR-ΔE-RVh-PM-1f, содержащей промотор/энхансер EFIα и ген dhfr, связанный с дефектной SV40 промоторной/энхансерной последовательностью для амплификации, применимую для экспрессии H-цепи.
Фиг. 12 является графиком, показывающим способность вариантов "a" и "b" видоизмененной V-области L-цепи человеческого PM-1 на связывание с человеческим IL-6R.
Фиг. 13 является графиком, показывающим способность варианта "f" видоизмененной V-области H-цепи человеческих PM-1 плюс варианта "a" видоизмененной V-области L-цепи PM-1 к связыванию с человеческим IL-6R.
Фиг. 14 является графиком, показывающим способность варианта "f" видоизмененной V-области H-цепи PM-1 плюс варианта "a" видоизмененной V-области L-цепи PM-1 подавлять связывание IL-6 с человеческим IL-6R.
Фиг. 15 представляет плазмиды экспрессии HEF-VL-gk и HEF-VH-gγI, включающие промотор/энхансер человеческого EFI-α, применимые для экспрессии L-цепи и H-цепи соответственно.
Фиг. 16 показывает процесс создания ДНК, кодирующей видоизмененную V-область L-цепи человеческих антител AUK12-20.
Фиг. 17 является графиком, показывающим результаты ELISA для подтверждения способности видоизмененной V-области L-цепи человеческих антител AUK 12-20 связываться с человеческими IL-6R. На фигуре "стандарт" AUK 12-20 (химерн. ) означает результат для химерных антител AUK 12-20, продуцируемых клетками CHO и очищенных в большом количестве.
Фиг. 18 является графиком, показывающим результат ELISA на способность видоизмененных человеческих антител AUK 12-20 (вариант "a" L-цепи + вариант "b" H-цепи) связываться с человеческими IL-6R.
Фиг. 19 является графиком, показывающим результат ELISA на способность видоизмененных человеческих антител AUK 12-20 (вариант "a" L-цепи + вариант "d" H-цепи) связываться с человеческими IL-6R.
Фиг. 20 показывает процесс химического синтеза видоизмененной V-области H-цепи человеческих антител sle 1220 H.
Фиг. 21 является графиком, показывающим результат ELISA на способность видоизмененных человеческих антител sle 1220 (вариант "a" L-цепи + вариант "a" H-цепи) подавлять связывание IL-6 с человеческими IL-6R.
Фиг. 22 является графиком, показывающим результат ELISA на способность видоизмененных человеческих антител sle 1220 (вариант "a" L-цепи + вариант "b" H-цепи) подавлять связывание IL-6 с человеческими IL-6R.
Фиг. 23 является графиком, показывающим результат ELISA на способность видоизмененных человеческих антител sle 1220 (вариант "a" L-цепи + вариант "c" H-цепи) подавлять связывание IL-6 с человеческими IL-6R.
Фиг. 24 является графиком, показывающим результат ELISA на способность видоизмененных человеческих антител sle 1220 (вариант "a" L-цепи + вариант "d" H-цепи) подавлять связывание IL-6 с человеческими IL-6R.
Наилучший способ осуществления изобретения
Клонирование ДНК, кодирующей мышиные V-области
Говоря точнее, чтобы клонировать ДНК, кодирующую V-области мышиных моноклональных антител к человеческим IL-6R, необходимо создание гибридомы как источника генов, которая продуцирует моноклональные антитела к человеческим IL-6R. В качестве такой гибридомы японская патентная заявка N 2-189420 описывает мышиную гибридому PM-1, которая продуцирует моноклональные антитела PM-1, и ее свойства. В справочных примерах 1 и 2 настоящей спецификации описывается процесс создания гибридомы PM-1. Авторы настоящего изобретения создали гибридомы AUK 12-20, AUK 64-7 и AUK 146-15, причем каждая продуцирует мышиные моноклональные антитела к человеческим IL-6R. Процесс создания этих гибридом описан в справочном примере 3 этой спецификации.
Чтобы клонировать желаемую ДНК, кодирующую V-области мышиных моноклональных антител, гибридомные клетки гомогенизируются и суммарную РНК получают по общепринятому методу, описанному Chirgwin et al, Biochemistry 18, 5294, 1977. Затем вся РНК используется, чтобы синтезировать одноцепочечные ДНК по методу, описанному J. W. Larrick et al., Biotechnology 7, 934, 1989.
Затем проводится специфическая амплификация соответствующей части кДНК по методу полимеразной цепной реакции (PCR). Для амплификации V-области к L-цепи мышиных моноклональных антител используются 11 групп олигонуклеотидных праймеров (мышиных каппа вариабельных; MKV), представленных в SEQ ID N: с 1 по 11 и олигонуклеотидный праймер (мышиных каппа константных; МКС), представленный в SEQ ID N: 12, в качестве 5'-терминальных праймеров и 3'-терминальных праймеров соответственно. MKV праймеры гибридизируются с ДНК-последовательностью, кодирующей лидерную последовательность мышиной L-цепи, и МКС праймер гибридизируется с ДНК последовательностью, кодирующей константную область мышиной L-цепи. Для амплификации V-области H-цепи мышиных моноклональных антител используются 10 групп олигонуклеотидных праймеров (мышиных тяжелых вариабельных; MHV), представленных в SEQ ID N: с 13 по 22, и олигонуклеотидный праймер (мышиных тяжелых константных; MHC), представленный в SEQ ID N: 23 в качестве 5'-терминальных праймеров и 3'-терминального праймера соответственно.
Заметим, что 5'-терминальные праймеры содержат нуклеотидную последовательность GTCGAC ближе к его 5'-концу, последовательность, которая представляет сайт расщепления рестрикционного фермента sal 1, и 3'-терминальный праймер содержит нуклеотидную последовательность, которая представляет сайт расщепления рестрикционного фермента Xma 1. Эти сайты расщепления рестрикционных ферментов используются для субклонирования фрагментов ДНК, кодирующих вариабельную область, в клонирующие векторы.
Затем продукт амплификации расщепляется рестрикционными ферментами sal 1 и Xma 1, чтобы получить фрагмент ДНК, кодирующий желаемую область V-мышиных моноклональных антител. С другой стороны подходящий клонирующий вектор, такой как плазмида pUC19, расщепляется теми же самыми рестрикционными ферментами sal 1 и Xma 1, и вышеуказанный фрагмент ДНК связывается с расщепленной pUC19, чтобы получить плазмиду, включающую фрагмент ДНК, кодирующий желаемую V-область мышиных моноклональных антител.
Секвенирование клонированной ДНК может быть осуществлено с помощью общепринятых методов.
Клонирование желаемой ДНК и ее секвенирование детально описаны в примерах от 1 до 3.
Области, определяющие последовательность (CDRS)
Настоящее изобретение представляет гипервариабельные или определяющие комплементарность области (CDRS)каждой V-области настоящего изобретения, V домены каждой пары L- и H-цепей из сайта связывания антигена. Домены L- и H-цепей имеют ту же самую структуру, и каждый домен содержит четыре основные области (FRS), чьи последовательности относительно сохранны, соединенные тремя CDRS (смотрите Kabat, E.A. Wu, T.T., Bilofsky, H, Reid-Miller, M. and Perry, H. , in "Sequences of Proteins of Immunological Interest", US Dept. Health and Human Services, 1983). Четыре FR главным образом принимают β-плоскую конформацию и CDR образуют петли, соединяющие FR и, в некоторых случаях, образующие часть β- плоской структуры. CDR удерживается в непосредственной близости к FR и CDR из другого домена, содействуют формированию сайта, связывающего антиген. CDR описаны в примере 4.
Создание химерного антитела
Перед разработкой видоизмененных человеческих V-областей антител к человеческим IL-6R необходимо подтвердить, что CDR, которые будут использованы на деле, образуют эффективную область связывания антигена. С этой целью создавались химерные антитела. Кроме того, аминокислотные последовательности V-областей мышиных против человеческих IL-6R антител, предсказанные исходя из нуклеотидных последовательностей клонированных ДНК 4 мышиных моноклональных антител, описанных в примерах 1 и 2, сравнили друг с другом и с V-областями из известных мышиных и человеческих антител. Для каждого из 4 мышиных моноклональных антител был клонирован набор типичных функциональных V-областей L- и H-цепей. Все четыре мышиные анти-IL-6R антитела, однако, имели относительно различные V-области. Четыре антитела не были просто незначительными вариациями одно другого. При использовании клонированных мышинах V-областей были созданы 4 химерных анти-IL-6R антитела.
Основной метод создания химерных антител включает соединение мышиных лидерной последовательности и последовательности V-области, как обнаружено при клонировании с применением PCR кДНК, с последовательностью, кодирующей человеческие C-области, уже присутствующей в векторах экспрессии клеток млекопитающих. Среди вышеуказанных 4 моноклональных антител создание химерных антител из моноклональных антител AUK 12-20 описано в примере 5.
Создание химерных антител из моноклональных антител PM-1 описывается в примере 6. кДНК, кодирующая лидерную и V-область L-цепи мышиных PM-1, была субклонирована с PCR в вектор экспрессии, содержащий геномную ДНК, кодирующую человеческую каппа C-область. кДНК, кодирующая лидерную и V-области H-цепи мышиных PM-1 была субклонирована с PCR в вектор экспрессии, содержащий геномную ДНК, кодирующую человеческую гамма-I C-область. При использовании специально разработанных праймеров PCR кДНК, кодирующие V-область мышиных PM-1, были адаптированы на их 5'- и 3'-концах (1) так, чтобы они легко встраивались бы в векторы экспрессии и (2) так, чтобы они функционировали бы соответствующим образом в этих векторах экспрессии. Модифицированные PCR V-области мышиных PM-1 затем встраивались в векторы экспрессии HCMV, уже содержащие желаемые человеческие C-области (фигура 1). Эти векторы пригодны для или кратковременной или постоянной экспрессии генетически разработанных антител в ряде линий клеток млекопитающих.
В дополнение к созданию химерных антител PM-1 с V-областями, идентичными V-областям, присутствующим в мышиных антителах PM-1 (вариант a), был создан второй вариант химерных антител PM-1 (вариант b). У химерных антител PM-1 (вариант b) аминокислота в положении 107 V-области L-цепи была заменена с аспарагина на лизин. При сравнении V-области L-цепи мышиных антител PM-1 с другими V-областями L-цепей было замечено, что появление аспарагина в положении 107 было необычным явлением. В V-областях L-цепей мыши наиболее типичной аминокислотой в положении 107 является лизин. Чтобы оценить значение наличия атипичной аминокислоты аспарагина в положении 107 V-области L-цепи мышиных антител PM-1, положение 107 изменялось на типичную аминокислоту лизин в этой позиции. Это изменение достигалось при использовании метода мутагенеза с PCR (M. Kamman et al., Nucl. Acids Res. (1989) 17:5404), чтобы произвести необходимые изменения в последовательностях ДНК, кодирующих V-область L-цепи.
Вариант (a) химерных антител PM-1 проявлял активность по связыванию с человеческими IL-6R. Вариант b химерных антител PM-1 связывается с человеческими IL-6R так же, как и вариант (a). Подобным же образом из 2 других моноклональных антител AUK 64-7 и AUK 146-15 были созданы химерные антитела. Все 4 химерных антитела хорошо связываются с человеческими IL-6R, показывая таким образом в функциональном исследовании, что нужные мышиные V-области были правильно клонированы и секвенированы.
Из 4 мышиных против-IL-6R антител антитела PM-1 были выбраны в качестве первого кандидата для разработки и создания видоизмененных человеческих антител к человеческим IL-6R. Выбор мышиных антител PM-1 основывался главным образом на результатах, полученных при изучении действия мышиных против-IL-6R антител на человеческие опухолевые миеломные клетки, трансплантированные безволосым мышам. Из 4 мышиных против-IL-6R антител антитела PM-1 показали наивысшую активность против опухолевых клеток.
Сравнение V-областей из мышиных моноклональных антител PM-1 с V-областями из известных мышиных и человеческих антител
Для того чтобы создать видоизмененные человеческие антитела, причем CDR мышиных моноклональных антител пересаживаются в человеческие моноклональные антитела, желательно, чтобы существовала высокая гомология между FR мышиных моноклональных антител и FR человеческих моноклональных антител. Поэтому аминокислотные последовательности V-областей L- и H-цепей из мышиных антител PM-1 сравнивались со всеми известными мышиными V-областями, какие находятся в базе данных OWL (или Leeds) белковых последовательностей.
Что касается V-областей мышиных антител, V-область L-цепи антитела PM-1 была наиболее подобна V-области L-цепи мышиного антитела musigkcko (Chen, H. -T. et al. , J. Biol. Chem. (1987) 262:13579-13583) с идентичностью 93,5%. V-область H-цепи антител PM-1 была наиболее подобна V-области H-цепи мышиных антител musigvhr 2 (F.J. Grant et al., Nucl. Acids Res. (1987) 15: 5496) с идентичностью 84,0%.
V-области мышиных PM-1 показывают высокий процент идентичности с известными мышиными V-областями, свидетельствуя таким образом, что V-области мышиных PM-1 являются типичными мышиными V-областями. Это представляет дополнительное непрямое доказательство, что клонированные последовательности ДНК являются правильными. Присутствует, обычно, более высокий процент идентичности между V-областями L-цепи, чем между V-областями H-цепи. Возможно это связано с меньшим числом различий, обычно наблюдаемым в V-областях L-цепи по сравнению с V-областями H-цепи.
Что касается V-областей человеческих антител, V-область L-цепи антител PM-1 была наиболее сходна с V-областью L-цепи человеческих антител Klhure, также упоминаемых как REI (W. Palm et al., Physiol. Chem. (1975) 356: 167-191) с идентичностью 72,2%. V-область H-цепи антител PM-1 была наиболее сходна с V-областью H-цепи человеческого антитела humighvap (VAP) (H.W. Schroeder et al. , Science (1987) 238: 791-793) с идентичностью 71,8%. Сравнение с человеческими V-областями наиболее важно для рассмотрения того, как разработать видоизмененные человеческие антитела исходя из мышиных антител PM-1. Процент идентичности с человеческими V-областями меньше, чем процент идентичности с мышиными V-областями. Это является непрямым доказательством того, что V-области мышиных PM-1 сходны с мышиными V-областями и не похожи на человеческие V-области. Эти данные также указывают на то, что для исключения проблем иммуногенности у больных людей наилучшим решением будет очеловечить V-области мышиных PM-1.
V-области мышиных антител PM-1 также сравнили с соответствующими последовательностями различных подгрупп человеческих областей, как определено E.A. Kabat et al. ((1987) Sequences of Proteins of Immunological Interest, Forth Edition, U.S. Department of Health and Humman servides, U.S. Government Printing (Office). Проведено сравнение FR-областей. Результаты представлены в таблице 1.
FR V-области L-цепи мышиных PM-1 наиболее похожи на FR из соответствующей последовательности V-областей человеческих L-цепей подгруппы I (HSG I) с идентичностью 70,1%. FR V-области H-цепи мышиных PM-1 наиболее сходны с FR соответствующих последовательностей V-областей человеческих H-цепей из подгруппы II (HSG II) с идентичностью 52,9%. Эти результаты подтверждают результаты, полученные при сравнении с известными человеческими антителами. V-область L-цепи в человеческих REI принадлежит к подгруппе I V-областей L-цепей и V-области H-цепи в человеческих VAP относятся к подгруппе II человеческих V-областей H цепи.
Исходя из этого сравнения с V-областями человеческих антител можно выбрать человеческие V-области, которые будут основой для разработки видоизмененных V-областей человеческих PM-1. Наилучшим было бы использование V-области человеческой L-цепи, которая принадлежит к подгруппе I (HSG II) для разработки видоизмененной V-области L-цепи преобразованных человеческих PM-1 и V-области человеческой H-цепи, которая относится к подгруппе II (HSG II) для разработки V-области H-цепи видоизмененных человеческих PM-1.
Разработка вариабельных областей видоизмененных человеческих PM-1
Первой стадией в разработке областей видоизмененных человеческих PM-1 был выбор человеческих V-областей, которые были бы основой разработки. FR V-области L-цепи мышиных PM-1 были наиболее сходны с FR V-областей человеческой L-цепи, относящихся к подгруппе I (таблица 1). Как обсуждалось выше, при сравнении V-области L-цепи мышиных PM-1 с известными V-областями L-цепи человека, они были наиболее сходны с V-областью REI L-цепи человека, члена подгруппы I V-областей человеческих L-цепей. При разработке V-областей L-цепи видоизмененных человеческих PM-1 были использованы FR из REI. Кроме того, FR REI использовались как стартовый материал для создания V-области L-цепи видоизмененных человеческих PM-1.
У этих человеческих FR на основе REI существует пять различий с FR первоначальных человеческих REI (положения 39, 71, 104, 105 и 107 по Kabat et al., 1987; см. таблицу 2). Три изменения в FR4 (положения 104, 105 и 107) базировались на J-области из другой человеческой каппа L-цепи и поэтому не представляют отклонения от человеческого (L. Riechmann et al., Nature (1988), 322: 21-25). Два изменения в положениях 39 и 71 были изменениями обратно к аминокислотам, которые присутствуют в FR V-области L-цепи крыс САМРАТН-1 (Riechmann et al., 1988).
Были разработаны два варианта V-области L-цепи видоизмененных человеческих антител PM-1. В первом варианте (вариант "a") человеческие FR были идентичны FR на основе REI, присутствующим в видоизмененных человеческих САМРАТН-1H (Riechmann et al., 1988) и мышиные CDR были идентичны CDR в V-области L-цепи мышиных PM-1.
Второй вариант (вариант "b") базировался на варианте "a" с заменой только одной аминокислоты в положении 71 человеческой FR3. Остаток 71 является частью канонической структуры CDR 1 V-области L-цепи, как определено C.Chothia et al., (J. Mol. Biol (1987) 196 : 901-917). Предсказывается, что аминокислота в этом положении непосредственно влияет на строение петли CDR 1 V-области L-цепи и поэтому может значительно влиять на связывание антигена.
В V-области L-цепи мышиных PM-1 в положении 71 находится тирозин. В модифицированных FR REI, использованных при разработке варианта "a" V-области L-цепи видоизмененных человеческих PM-1 находился фенилаланин. В варианте "b" V-области L-цепи видоизмененных человеческих PM-1 фенилаланин в положении 71 был заменен на тирозин, который обнаружен в V-области L-цепи мышиных PM-1. Таблица 2 показывает аминокислотные последовательности V-области L-цепи мышиных PM-1, FR REI которых модифицированы для использования в видоизмененных человеческих антителах CAMPATH-1H (Richmann et al., 1988) и двух вариантов V-области L-цепи видоизмененных человеческих PM-1.
FR V-области H-цепи мышиных PM-1 были наиболее сходны с FR V-областей H-цепи человека, принадлежащих к подгруппе II (таблица 1). Как обсуждалось выше, при сравнении V-области H-цепи мышиных PM-1 с известными V-областями H-цепей человека она наиболее сходна с V-областью VAP человеческой H-цепи, члена подгруппы II V-областей человеческих H-цепей. Последовательности ДНК, кодирующие FR V-области NEW человеческой H-цепи, другого члена подгруппы II V-областей человеческой H-цепи были использованы в качестве исходного материала для построения V-области H-цепи видоизмененных человеческих PM-1 и как основа для разработки V-области H-цепи видоизмененных человеческих PM-1.
Было разработано шесть вариантов V-области H-цепи видоизмененных человеческих PM-1. Во всех шести вариантах человеческие FR имели в основании FR NEW, присутствующие в видоизмененных человеческих CAMPATH-1H (Riechmann et al. , 1988), и мышиные CDR были идентичны CDR V-области H-цепи мышиных PM-1. Смесь аминокислотных остатков в человеческих FR (положения 1, 27, 28, 30, 48 и 71, см. таблицу 3), как было определено, имеют возможное побочное влияние на связывание антигена. B модели V-областей мышиных PM-1 остаток I V-области H-цепи является поверхностным остатком, который располагается близко к петлям CDR. Остатки 27, 28, 29 и 30 являются или частью канонической структуры CDR1 V-области H-цепи, как предсказано C.Chothia et al., Nature (1989) 34: 877-883, и/или, как наблюдается на модели V-областей мышиных PM-1, образуют часть первой структурной петли V-области H-цепи (Chothia, 1987). Остаток 48, как замечено на модели V-области мышиных PM-1, является спрятанным остатком. Изменения спрятанного остатка могут разрушать структуру V-области в целом и ее антиген-связывающий сайт. Остаток 71 является частью канонической структуры CDR2 V-области H-цепи, как предсказано Chothia et al., (1989). Шесть вариантов видоизменного человеческого антитела PM-1 включают различные комбинации изменений аминокислот в этих семи положениях человеческих FR NEW (см. таблицу 3).
Создание видоизмененных V-областей человеческих PM-1.
Первые варианты V-областей L- и H-цепей видоизмененных человеческих PM-1 каждый создавали, используя новый метод, основанный на PCR. В основном плазмидную ДНК, кодирующую видоизмененную V-область человека, которая уже содержала подходящие человеческие FR, модифицировали, используя праймеры PCR, чтобы заменить CDR, присутствующие в изначальной видоизмененной человеческой V-области CDR из мышиных антител PM-1. Исходный материал для построения видоизмененной V-области L-цепи человеческих PM-1 является плазмидной ДНК, видоизмененную V-область человеческой D1.3L-цепи. Видоизмененная V-область человеческой D1.3L-цепи создавалась на основе FR, присутствующих в V-области человеческой L-цепи REI. Исходным материалом для построения видоизмененной области H-цепи человеческих PM-1 была плазмидная ДНК, содержащая видоизмененную человеческую область D1.3H-цепи. Видоизмененная человеческая область D1.3H-цепи создавали на основе FR, присутствующих в V-области человеческой H-Цепи NEW (M.Verhoeyen et al., Science (1988) 239:1534-1536).
Если исходные плазмидные ДНК, содержащие желаемые человеческие FR, уже выбраны, праймеры PCR предназначались для того, чтобы сделать возможным замещением CDR мышиных PM-1 CDR мышиных D1.3. Для каждой видоизмененной области человеческих PM-1 были разработаны и синтезированы три праймера, содержащих последовательности ДНК, кодирующие CDR мышиных PM-1, и два праймера, расположенных по краям полной последовательности ДНК, кодирующей видоизмененную человеческую V-область. Используя пять праймеров PCR в серии PCR, получали продукт PCR, который состоял из человеческих FR, присутствующих в исходной видоизмененной человеческой V-области и CDR, присутствующих в V-области мышиных PM-1 (см. пример 7 и фигуры 7 и 8). Продукты PCR клонировали и секвенировали, чтобы убедиться в том, что полная последовательность ДНК варианта "a" видоизмененной V-области L- и H-цепей человеческих PM-1 кодировала правильную аминокислотную последовательность (SEQ ID N 55).
Остальные варианты видоизмененных областей человеческих PM-1 создавались с использованием небольших модификаций опубликованных методик мутагенеза с PCR (Kamman et al., 1989). Как описано для разработки видоизмененных областей человеческих антител PM-1, был создан один дополнительный вариант (вариант b) видоизмененной V-области L-цепи человеческих антител и получены пять дополнительных вариантов (варианты "b", "c", "d", "e" и "f") видоизмененной V-области H-цепи человеческих PM-1. Эти дополнительные варианты содержат серии незначительных изменений по сравнению с первыми вариантами. Разработаны праймеры, которые будут вводить необходимые изменения в последовательность ДНК. После серии PCR продукт PCR клонировали и секвенировали, чтобы убедиться, что изменения в последовательности ДНК такие, как запланировано. Последовательность варианта "f" V-области H-цепи видоизмененных человеческих антител PM-1 показана в SEQ ID N 54.
Если последовательности ДНК различных вариантов V-областей видоизмененных человеческих PM-1 подтверждались секвенированием, V-области видоизмененных человеческих PM-1 субклонировали в векторы экспрессии клеток млекопитающих, уже содержащие человеческие C-области. Видоизмененные V-области L-цепи человеческих антител PM-1 соединяли с последовательностями ДНК, кодирующими человеческую C-область. Видоизмененные V-области H-цепи человеческих PM-1 соединяли с последовательностями ДНК, кодирующими человеческие гамма-1 C-область. Чтобы достичь высоких уровней экспрессии видоизмененных человеческих антител PM-1, векторы экспрессии HCMV, как показано на фигуре 1, модифицировали. чтобы заменить промоторно-энхансерную область HCMV промотором-энхансером человеческого фактора элонгации (HEF-I) (см. фигуру 15).
Затем все комбинации вариантов (a) и (b) видоизмененной человеческой L-цепи с вариантами от (a) до (f) V-области H-цепи испытывали на связанные с человеческими IL-6R и в результате видоизмененные человеческие антитела, включающие вариант (a) L-цепи и вариант (f) H-цепи, показали способность связываться с IL-6R на том же самом уровне, что и химерные PM-1 (a) (фиг. 13), что описано в деталях в примере II.
Модификации последовательностей ДНК, кодирующих области видоизмененных человеческих PM-1 с целью улучшения уровней экспрессии.
При рассмотрении уровней видоизмененных человеческих PM-1 антител, которые продуцируются cos клетками, стало очевидно, что уровни экспрессии видоизмененных человеческих H-цепей были всегда примерно в 10 раз ниже, чем уровни экспрессии видоизмененных человеческих L-цепей или химерных L- или H-цепей. По-видимому существует проблема в кодировании ДНК видоизмененной V-области человеческой H-цепи, что служит причиной низких уровней экспрессии. Чтобы определить были ли низкие уровни экспрессии белка результатом низких уровней транскрипции, получали PHK из cos клеток, трансфицированных одновременно векторами, экспрессирующими видоизмененные L- и H-цепи человеческих PM-1. Первично-цепочечная кДНК синтезировалась как описано для клонирования с PCR V-областей мышиных PM-1. Используя праймеры PCR, разработанные для расположения по концам ДНК, кодирующей V-области видоизмененных L- и H-цепей, продукты PCR получали с кДНК, которая соответствовала V-области видоизмененной человеческой L-цепи или видоизмененной V-области человеческой H-цепи.
Для V-области видоизмененной L-цепи существовали два продукта PCR, один длиной 408 bp, как ожидалось, и более короткий продукт PCR длиной 299 bp. Продукт PCR правильного размера производился до уровня примерно 90% от общего выхода продукта PCR и более короткий продукт PCR производился на уровне примерно 10% от общего выхода. Для видоизмененной V-области человеческой H-цепи существовали также два продукта PCR, один длиной 444 bp, как ожидалось, и более короткий продукт PCR длиной 370 bp. В этом случае, однако, неправильный более короткий продукт PCR производился до преобладания в общем выходе продукта PCR, примерно в 90%. Продукт PCR правильного размера производился на уровне только примерно 10% от общего выхода продукта PCR. Эти результаты показали, что какое-то количество РНК, кодирующей видоизмененные человеческие V-области, содержит делеции.
Чтобы определить, какие последовательности были стерты, более короткие продукты PCR клонировали и секвенировали. Исходя из последовательностей ДНК стало ясно, что как для V-областей L-цепи, так и H-цепи специфические части ДНК стерты. Изучение последовательностей ДНК, находящихся на концах стертых последовательностей, показано, что эти последовательности соответствовали согласованным последовательностям для соединения донорно-акцепторных последовательностей (Breathnach, R. et al., Ann. Rev. Biochem (1981) 50:349-383). Объяснение для низких уровней экспрессии видоизмененных человеческих H-цепей состояло в том, что конструкция V-областей видоизмененных человеческих H-цепей неумышленно создала довольно эффективный набор донорно-акцепторных сайтов сплайсинга. Кажется также, что разработка V-областей видоизмененных человеческих L-цепей неумышленно создала довольно неэффективный набор донорно-акцепторных сайтов сплайсинга. Чтобы удалить донорно-акцепторные места сплайсинга, были произведены незначительные модификации в последовательностях ДНК, кодирующих варианты "a" и "f" соответственно видоизмененных V-областей L- и H-цепей человеческих PM-1 с использованием методов мутагенеза с PCR, описанных выше.
Другой возможной причиной сниженных уровней экспрессии, как полагали, являлось наличие интронов в лидерных последовательностях V-областей видоизмененных человеческих как L-, так и H-цепей (SEQ ID N0: 54 и 55). Эти интроны были первоначально получены из лидерной последовательности мышиной mu H-цепи (M.S.Neuberger er al., Nature 1985, 314: 268-270), которая была использована при построении видоизмененных человеческих D1.3 и V-областей. (Verhoeyen et al. , 1988). Так как видоизмененная D1.3 экспрессировалась в векторе клеток млекопитающих, в котором использовался мышиный иммуноглобулиновый промотор, наличие мышиного лидерного интрона было важным. Лидерный интрон содержит последовательности, которые важны для экспрессии из иммуноглобулиновых промоторов, но не с вирусных промоторов, подобных HCMV (M.S. Neuberger et al., Nucl. Acids Res. (1988) 16:6713-6724). Там, где видоизмененные человеческие L- и H-цепи PM-1 экспрессировались в векторах с использованием неиммуноглобулиновых промоторов, интроны в лидерных последовательностях утрачивались при клонировании с PCR кДНК, кодирующей видоизмененные человеческие V-области (см. пример 12).
Другой возможной причиной сниженных уровней экспрессии, как полагали, являлось наличие протяженности примерно на 190 bp нефункциональной ДНК в интроне между видоизмененной V-областью H-цепи человеческих PM-1 и человеческой гамма-I C-областью. Видоизмененная V-область H-цепи человеческих PM-1 строилась из последовательностей ДНК, первоначально полученных из видоизмененной V-области H-цепи человеческих BI-8 (P. T. Jones et al., Nature (1986) 321: 522-525). Эту первичную видоизмененную человеческую V-область конструировали из V-области H-цепи мышиных NP (M.S. Neuberger et al., Nature (1985); M. S. Neuberger et al., EMBO J. (1983) 2:1373-1378). Эта протяженность в примерно 190 bp, находящаяся в интроне между видоизмененной V-областью человеческой H-цепи и сайтом Bam HI для присоединения видоизмененных человеческих V-областей к вектору экспрессии, удалялась при клонировании с PCR кДНК, кодирующей видоизмененные человеческие V-областти.
Последовательности ДНК и аминокислот окончательных вариантов видоизмененных V-областей L- и H-цепей человеческих PM-1, которые изменены для улучшения уровней экспрессии, показаны в ID NOs: 57 и 56. Эти последовательности ДНК кодируют вариант "a" видоизмененной области цепи человеческих PM-1, как показано в таблице 2, и вариант "f" видоизмененной V-области H-цепи человеческих PM-1, как показано в таблице 3. При встраивании в векторы экспрессии HEF-1 (фигура 15) эти векторы временно продуцируют примерно 2 мкг/мл антител в трансфицированных cos клетках. Чтобы стабильно получать большие количества видоизмененных человеческих антител PM-1, был создан новый вектор экспрессии HEF-Iα, включающий dhfr (см. пример 10, фиг. 11). "Поврежденный" ген dhfr вводился в вектор HEF-Iα, экспрессирующий человеческие гамма-I H-цепи, что было описано для вектора HCMV, экспрессирующего человеческие гамма-I H-цепи. Вектор HEF-Iα, экспрессирующий видоизмененные L-цепи человеческих PM-1 и вектор HEF-Iα dhfr, экспрессирующий видоизмененные H-цепи человеческих PM-1 трансфицировались вместе в клетки CHO dhfr (-). Стабильно трансформированные линии клеток CHO отбирались на альфа-минимально необходимой среде (Alpha-Minimum Essential Midium [альфа-MEM]) без нуклеазидов и с 10% фетальной телячьей сыворотки (FCS) и 500 мкг/мл G418. До каких-либо шагов по амплификации генов, лини клеток CHO, как наблюдалось, продуцировали до 10 мкг/106 клеток/сутки видоизмененных человеческих антител PM-1.
Сравнение V-областей мышиных моноклональных антител AUK 12-20 с V-областями из известных человеческих антител
Гомология FR V-области kL-цепи мышиных моноклональных антител AUK 12-20 с FR V-области человеческой kL-цепи подгруппы (HSG) I до IV и гомология FR V-области H-цепи мышиных моноклональных антител AUK 12-20 с FR V-областей человеческой H-цепи подгруппы (HSG) I до III показаны в таблице 4.
Как видно из таблицы 4, V-область kL-цепи мышиных моноклональных антител AUK 12-20 гомологична в степени (64 по 68%) сходной с V-областью kL-цепи человека подгрупп (HSG) I по IV. При поиске в базе данных "LEEDS" на белок V-область L-цепи человеческих антител Len (M.Schneider et al., Physiol. Chem. (1975) 366: 507-557), принадлежащих к группе HSG-IV, проявляет наивысшую гомологию 68%. С другой стороны, человеческие антитела REI, использованные для построения видоизмененных человеческих антител из мышиных моноклональных антител PM-1, принадлежат к HSG-I, проявляют 62% гомологию с V-областью L-цепи мышиных моноклональных антител AUK 12-20. Кроме того, CDR в V-области L-цепи AUK 12-20 антител, особенно CDR 2, лучше соответствовали каноническим структурам CDR в REI, чем таковым в LEN.
Принимая во внимание вышеизложенное, нет необходимости выбирать человеческие антитела, используемые для очеловечивания V-области L-цепи мышиных моноклональных антител AUK 12-20, из антител, принадлежащих к HSG IV. Поэтому, как в случае очеловечивания V-области L-цепи мышиных моноклональных антител PM-1, для гуманизации V-области L-цепи мышиных моноклональных антител AUK 12-20 используются FR REI.
Как показано в таблице 4, V-область H-цепи антител AUK 12-20 проявляет наивысшую гомологию с HSG I. Более того, при поиске в базе данных "LEEDS" человеческие антитела HAX (S'tollar, B.O. et al., J. Immunol. (1987) 139: 2496-2501), также принадлежащие к HSG I, проявляют примерно 66% гомологии с V-областью H-цепи антител AUK 12-20. Соответственно, чтобы разработать человеческую V-область H-цепи антител AUK 12-20, используются FR человеческих антител HAX, принадлежащих к HSG I, и FR очеловеченной V-области H-цепи антител 425, которые имеют FR, состоящие из соответствующей последовательности HSG I (Ketteborough C.A. et al., Protein Engineering (1991) 4: 773-783). Заметим, что V-область H-цепи антител AUK 12-20 проявляет примерно 64% гомологии с вариантом "a" очеловеченной V-области H-цепи антител 425.
Разработка V-областей L-цепи видоизмененных человеческих антител AUK 12-20.
По вышеприведенной причине V-области L-цепи видоизмененных человеческих антител AUK 12-20 разрабатываются как указано в таблице 5, с использованием FR REI.
Разработка V-областей H-цепи видоизмененных человеческих антител AUK 12-20.
По вышеприведенной причине V-области H-цепи видоизмененных человеческих антител AUK 12-20 разрабатываются с использованием FR видоизмененных человеческих VHa 425. Обнаружено, однако, что нуклеотидная последовательность ДНК, кодирующей V-область H-цепи видоизмененных человеческих антител AUK 12-20, разработанная таким образом, имеет последовательность хорошо согласующуюся со сплайсинговой донорной последовательностью. Поэтому, как и в случае видоизмененных человеческих антител PM-1, существует возможность ненормального сплайсинга в видоизмененных человеческих антителах AUK 12-20. Поэтому нуклеотидная последовательность частично модифицировалась, чтобы убрать сплайсинговую донорноподобную последовательность. Модифицированная последовательность обозначается как вариант "a".
Кроме того, варианты от "b" до "d" V-области H-цепи видоизмененных человеческих антител AUK 12-20 были дополнительно разработаны. Аминокислотные последовательности вариантов "a" до "d" представлены в таблице 6.
Более того, варианты с "a" по "d" V-областей H-цепи видоизмененных человеческих антител AUK 12-20, разрабатываются как показано в таблице 7, с использованием FR человеческих антител HAX (J. Immunology (1987) 139: 2496 - 2501; антитела продуцировались клетками гибридомы 21/28, полученной из B клеток больного CKB; их аминокислотная последовательность описана на фиг. 6 и нуклеотидная последовательность ДНК, кодирующей аминокислотную последовательность, показана на фиг. 4 и 6 этого литературного источника).
Для продукции настоящих химерных или видоизмененных человеческих антител к человеческим IL-6R может использоваться любая система экспрессии, включая эукариотические клетки, например клетки животных как утвердившиеся линии клеток млекопитающих, клетки грибов, дрожжевые клетки, такие как клетки E. coli. Предпочтительно настоящие химерные или видоизмененные человеческие антитела экспрессируются в клетках млекопитающих, таких как cos клетки или CHO клетки.
В таких случаях может использоваться общепринятый промотор, применимый для экспрессии в клетках млекопитающих. Например, предпочтительно используется вирусная система экспрессии, такая как ранний промотор человеческого цитомегаловируса (HCMV). Примеры векторов экспрессии, содержащих промотор HCMV включают: HCMV-VH-HCγI, HCMV-VL-HCK, HCMV-12h-gγI, HCMV-12k-gk и тому подобные, полученные из pSV2neo, как показано на фиг. 1.
Другим примером промотора, применимого для настоящего изобретения, является промотор человеческого фактора элонгации I α(HEF-I). Векторы экспрессии, содержащие этот промотор, включают HEF-12h-gγI и HEF-12h-gk (фиг. 8 и 9), так же как и HEF-VH-gγI и HEF-VL-gk (фиг. 15).
Для амплификации гена dhrf в линии клеток-хозяев вектор экспрессии может содержать ген dhfr. Векторами экспрессии, содержащими ген dhfr, являются, например, DHFR -ΔE- PMh -gγI (фиг. 10). DHFR -ΔE- PVh-PM-1-F (фиг. 11) и тому подобные.
Подводя итог, можно сказать, что настоящее изобретение впервые представляет V-область L-цепи и V-область H-цепи мышиных моноклональных антител к человеческим IL-6R, так же как и ДНК, кодирующую V-область L-цепи, и ДНК, кодирующую V-область H-цепи. Они применимы для создания человеческо/мышиных химерных антител и видоизмененных человеческих антител к человеческим IL-6R. Моноклональными антителами являются, например, AUK 12-20, PM-1, AUK 64-7 и AUK 146-15, V-область L-цепи имеет аминокислотную последовательность, показанную, например, в SEQ ID NOs: 24, 26, 28 или 30; и V-область H-цепи имеет аминокислотную последовательность, показанную в SEQ ID NOs: 25, 27, 29 или 31. Эти аминокислотные последовательности кодируются нуклеотидными последовательностями, например, показанным в SEQ ID NOs: с 24 по 31 соответственно.
Настоящее изобретение также относится к химерным антителам к человеческим IL-6R, содержащим:
(1) L-цепь, включающую C-область человеческой L-цепи и область мышиной L-цепи; и
(2) H-цепь, включающую C-область человеческой H-цепи и V-область мышиной H-цепи. V-область мышиной L-цепи и V-область мышиной H-цепи и ДНК, кодирующая их, описаны выше. C-область человеческой L-цепи может быть любой C-областью человеческой L-цепи и, например, является человеческой Ck. C-область человеческой H-цепи может быть любой С-областью человеческой Н-цепи и, например, является человеческой CγI.
Для продукции химерных антител создаются два вектора экспрессии, т.е. один, содержащий ДНК, кодирующую V-область мышиной L-цепи и C-область человеческой L-цепи контролем области контроля экспрессии, такой как энхансер/промоторная система, и другой, содержащий ДНК, кодирующую область мышиной H-цепи и C-область человеческой H-цепи под контролем области контроля экспрессии, такой как энхансер/промоторная система. Затем векторы экспрессии трансфицируются вместе в клетки-хозяева, такие как клетки млекопитающих, и трансфицированные клетки культивируются in vitro или in vivo для продукции химерных антител.
Или же, иначе, ДНК, кодирующая V-область мышиной L-цепи и C-область человеческой L-цепи, и ДНК, кодирующая V-области мышиных H-цепей и C-область человеческой H-цепи, вводятся в единственный вектор экспрессии, и этот вектор используется для трансфицирования клеток-хозяев, которые затем культивируются in vivo или in vitro для продукции желаемых химерных антител.
Настоящее изобретение, кроме того, представляет видоизмененное антитело к человеческому IL-6R, содержащее:
(A) L-цепь, включающую
(1) C-область человеческой L-цепи, и
(2) V-область L-цепи, содержащую FR человеческой L-цепи, и CDR-цепи мышиного моноклонального антитела к человеческому IL-6R;
(B) H-цепь, включающую
(1) C-область человеческой H-цепи, и
(2) V-область H-цепи, содержащую FR человеческой H-цепи, и CDR H-цепи мышиного моноклонального антитела к человеческому IL-6R.
В предпочтительном воплощении CDR L-цепи имеют аминокислотные последовательности, представленные в любой из SEQ ID NOs: 24, 26, 28 и 30, причем протяженности аминокислотных последовательностей определены в таблице 9; CDR L-цепи имеют аминокислотные последовательности, показанные в любой из SEQ ID NOs: 25, 27, 29 и 31, причем протяженности аминокислотных последовательностей определены в таблице 9; FR человеческой L-цепи получают из REI, а FR человеческой H-цепи получают из NEW или HAX.
В предпочтительном воплощении V-область L-цепи имеет аминокислотную последовательность, показанную в таблице 2, как PVLa; и V-область H-цепи имеет аминокислотную последовательность, показанную в таблице 3, как PVHa, PVHb, PVHc, PVHd, PVHe или PVHf. Аминокислотная последовательность PVHf наиболее предпочтительна.
Для продукции видоизмененных человеческих антител созданы два вектора экспрессии, т.е. один, содержащий ДНК, кодирующую видоизмененную L-цепь, как определено выше, под контролем области контроля экспрессии, такой как энхансер/промоторная система, и другой, содержащий ДНК, кодирующую видоизмененную человеческую H-цепь, как определено выше, под контролем области контроля экспрессии, такой как энхансер/промоторная система. Затем векторы экспрессии вместе трансфицируются в клетки-хозяева, такие как клетки млекопитающих, и трансфицированные клетки культивируются in vitro или in vivo для продукции видоизмененных человеческих антител.
Или же иначе, ДНК, кодирующая видоизмененную человеческую L-цепь, и ДНК, кодирующая видоизмененную H-цепь, вводятся в единственный вектор экспрессии, и этот вектор используется для трансфицирования клеток-хозяев, которые затем культивируются in vivo или in vitro для продуцирования желаемых видоизмененных человеческих антител.
Химерные антитела из видоизмененных человеческих антител, полученные таким образом, могут быть выделены и очищены с помощью общепринятых методов, таких как аффинная хроматография на протеине A, ионообменная хроматография, гельфильтрация и тому подобное.
Настоящая химерная L-цепь или видоизмененная человеческая L-цепь может быть соединена с H-цепью, чтобы получить полное антитело. Подобным же образом химерная H-цепь или видоизмененная H-цепь может быть соединена с L-цепью, чтобы получить целое антитело.
Настоящее V-область мышиной L-цепи, V-область видоизмененной человеческой L-цепи, V-область мышиной H-цепи и V-область видоизмененной человеческой H-цепи являются по-существу областью, которая связывается с антигеном, человеческим IL-6R, и поэтому, как считают, применимы сами по себе или в качестве белка, соединяемого с другим белком, для получения фармацевтических препаратов или диагностических средств.
Кроме того, данные CDR V-области L-цепи и CDR V-области H-цепи и являются по-существу областями, которые связываются с антигеном, человеческими IL-6R, и поэтому, как считают, применимы сами по себе или в качестве белка, соединяемого с другим белком, для получения фармацевтических препаратов или диагностических средств.
ДНК, кодирующая V-область мышиной L-цепи настоящего изобретения, применима для построения ДНК, кодирующей химерную L-цепь, или ДНК, кодирующей видоизмененную человеческую L-цепь.
Подобным же образом, ДНК, кодирующая V-область мышиной H-цепи данного изобретения, применима для построения ДНК, кодирующей химерную H-цепь, или ДНК, кодирующей видоизмененную человеческую H-цепь. Кроме того, ДНК, кодирующая CDR V-области L-цепи настоящего изобретения, применима для построения ДНК, кодирующей V-область видоизмененной человеческой L-цепь, и ДНК, кодирующей видоизмененную человеческую L-цепь. Аналогично, ДНК, кодирующая CDR V-области H-цепи настоящего изобретения, применима для построения ДНК, кодирующей видоизмененную человеческую H-цепь.
Настоящее изобретение далее будет иллюстрироваться, но никоим образом не ограничиваться, следующими примерами.
Пример 1.
Клонирование ДНК, кодирующей V-область мышиного моноклонального антитела к человеческому IL-6R (I)
ДНК, кодирующая V-область мышиного моноклонального антитела к человеческому IL-6R, клонировалась следующим образом.
1. Получение суммарной РНК
Суммарную РНК из гибридомы AUK 12-20 получали по методу, описанному Chirgwin et al, Biochemistry 18, 5294 (1979). А именно, 2,1 • 108 клеток гибридомы AUK 12-20 полностью гомогензировали в 20 мл 0,4 М тиоцианата гуанидина (Fulka). Гемогенат наслаивался на пласт 5,3 М раствора хлорида цезия в центрифужной пробирке, которую затем центрифугировали в роторе Becкмaн SW 40 при 31000 об/мин при 20oC в течение 24 часов, чтобы осадить РНК. Осадок РНК промывался 80% этанолом и растворялся в 150 мкл 10 мМ Трис-HCl (pH 7,5), содержащего 1 мМ ЭДТА и 0,5% додецилсульфата натрия, и после добавления протеиназы (Boehringer) вслед за тем по 0,5 мг/мл инкубировали при 37oC в течение 20 минут. Смесь экстрагировали фенолом и хлороформ, и РНК осаждали этанолом. Затем осадок РНК растворяли в 200 мкл 10 мМ Трис-HCl (pH 7,5), содержащего 1 мМ ЭДТА.
2. Синтез однонитевой кДНК
Чтобы синтезировать однонитевую кДНК по методу, описанному J.W. Larrick et al., Biotechnology 7.934 (1989), примерно 5 мкг суммарной РНК, полученной как описано выше, растворяли в 10 мкл 50 мМ Трис-HCl (pH 8,3) буферного раствора, содержащего 40 мМ KCl, 6 мМ Cl2, 10 мМ дитиотреитола, 0,5 мМ dATP, 0,5 мМ dGTP, 0,5 мМ dCTP, 0,5 мМ dTTP, 0,35 мкМ олиго dT праймера (Amersham). 48 единиц RAV-2 обратной транскриптазы (RAV-2: Rous Associated virus 2; Amersham) и 25 единиц человеческого плацентарного ингибитора рибонуклеазы (Amersham), и реакционную смесь инкубировали при 37oC в течение 60 минут и непосредственно использовали для последующего метода полимеразной цепной реакции.
3. Амплификация кДНК, кодирующей V-область антитела путем метода PCR
Метод PCR осуществляли, используя Thermal Cecler Model PHC-2 (Techne)
(1) Амплификация кДНК, кодирующей вариабельную область мышиной к легкой (kL)-цепи.
Праймерами, использованными для метода PCR, были MKV (мышиные каппа вариабельные) праймеры, представленные в SEQ ID NO: от 1 до 11, которые гибридизуруются с лидерной последовательностью мышиной к L-цепи (S.T. Jones et al, Biotechnology, 9, 88, 1991) и МКС (мышиного каппа константного) праймера, представленного в SEQ ID NO: 12, который гибридизуется с мышиной C-областью L-цепи (S.T. Jones et al, Biotechnology, 9, 88, 1991).
Сначала 100 мкл раствора PCR, содержащего 10 мМ Трис-CHl (pH 8,3), 50 мМ КCl, 0,1 мМ dATP, 0,1 мМ dGTP, 0,1 мМ dCTP, 0,1 мМ dTTP, 1,5 мМ MgCl2, 2,5 единиц ДНК-полимеразы Ampli Tag (Perkin Elmer Cetus), 0,25 мМ каждой группы MKV праймера, 3 мкМ МКС праймера и 1 мкЛ реакционной смеси синтеза однонитевой ДНК нагревали при начальной температуре 94oC в течение 1,5 минут и затем при 94oC в течение 1 минуты, при 50oC в течение 1 минуты и 72oC в течение 1 минуты, в этом порядке. После того как этот температурный цикл повторялся 25 раз, реакционную смесь дальше инкубировали при 72oC в течение 10 минут.
(2) Амплификация кДНК, кодирующей V-область мышиной H-цепи.
В качестве праймеров для PCR использовали МHV (мышиные тяжелые вариабельные) праймеры от 1 до 10 представлены в SEQ ID NO: с 13 до 22 (S.T. Jones et al., Biotechnology 9, 88, 1991) и MHC (мышиные тяжелые константные) праймер, представленный в SEQ ID NO: 23 (S.T. Jones et al., Biotechnology, 9, 88, 1991). Амплификацию проводили по той же самой методике, которая описана для амплификации гена V-области L-цепи в разделе 3.(I).
4. Очистка и расщепление продукта PCR
Фрагменты ДНК, амплифицированные PCR, как описано выше, очищали, используя набор для очистки продукта QIAGEN PCR (QIAGEN Inc, US), и расщепляли с помощью 10 единиц рестрикционного фермента Sal I (GIBCO BRL) в 100 мМ Трис-HCl (pH 7,6), содержащего 10 мМ MgCl2 и 150 мМ NaCl, при 37oC в течение трех часов. Расщепляющую смесь экстрагировали фенолом и хлороформом, и ДНК выделяли путем осаждения этанолом. Затем осадок ДНК расщепляли 10 единицами рестрикционного фермента Xma I (New England Biolabs) при 37oC в течение двух часов, и полученные в результате фрагменты ДНК разделяли путем электрофореза в агарозном геле, используя агарозу с низкой температурой плавления (FMC Bio Products USA).
Агарозный участок, содержащий фрагменты ДНК длиной примерно 450 bp, вырезали и расплавляли при 65oC в течение 5 минут и затем добавляли равный объем 20 мМ Трис-HCl (pH 7,5), содержащего 2 мМ ЭДТА и 200 мМ NaCl. Смесь экстрагировали фенолом и хлороформом, и фрагмент ДНК выделялся путем осаждения этанолом и растворялся в 10 мкЛ 10 мМ Трис-HCl (pH 7,5), содержащего 1 мМ ЭДТА. Таким же путем получали фрагмент ДНК, содержащий ген, кодирующий V-область мышиной L-цепи, и фрагмент ДНК, содержащий ген, кодирующий V-область машиной H-цепи. Оба из вышеназванных фрагментов ДНК имели способный к соединению Sal I конец на их 5'-конце и способный к соединению Xma 1 конец на их 3'-конце.
5. Лигирование и трансформация
Примерно 0,3 мкг фрагмента ДНК Sal I - Xma I, содержащего ген, кодирующий V-область мышиной цепи L, полученного как описано выше, сшивали с примерно 0,1 мкг вектора pUC19, полученного путем расщепления плазмиды pUC19 с помощью Sal I и Xma I, в реакционной смеси, содержащей 50 мМ Трис-HCl (pH 7,4), 10 мМ MgCl2, 10 мМ дитиотреитола, 1 мМ спермидина, 1 мМ dATP, 0,1 мкг/мл бычьего сывороточного альбумина и 2 единицы Т4 ДНК-лигазы (New England Biolabs) при 16oC в течение 16 часов.
Затем 7 мкЛ вышеприведенной смеси лигирования добавляли к 200 мкЛ компетентных клеток E. coli DH5 α и клетки инкубировали в течение 30 минут на льду, одну минуту при 42oC и одну минуту опять на льду. После добавления 800 мкл среды SOC (Molecular Cloning: A Laboratory Manual, Sambroock et al., Cold Spring Habor Laboratory Press, 1989) клеточную суспензию инкубировали при 37oC в течение одного часа и высевали на чашку с агаром 2xYT (Molecular Cloning: A Laboratory Manual, Sambroock et al., Cold Spring Habor Laboratory Press, 1989), которую затем инкубировали при 37oC в течение ночи, чтобы получить трансформант E.coli. Этот трансформант культивировали в 5 мл 2xYT среды, содержащей 50 мкг/мл ампициллина, при 37oC в течение ночи, и плазмидную ДНК получали из культуры по щелочному методу (Molecular Cloning: A Laboratory Manual, Sambrook et al., Cоld Spring Habor Laboratory Press, 1989). Полученная таким образом плазмида, содержащая ген, кодирующий V-область мышиной L-цепи, извлеченный из гибридомы AUK 12-20, была обозначена p12-k2.
Тем же самым методом, который описан выше, была создана плазмида, содержащая ген, кодирующий V-область мышиной H-цепи, полученный из гибридомы AUK 12-20, из фрагмента ДНК Sal I - Xma I и обозначена p12-h2.
Пример 2.
Клонирование ДНК, кодирующей V-область мышиного моноклонального антитела (2)
По существу тот же самый метод, что описан в примере 1, применялся в отношении гибридом PM-1, AUK 64-7 и AUK 146-15, чтобы получить следующие плазмиды:
плазмиду pPM-k3, содержащую ген, кодирующий V-область к L-цепи, полученный из гибридомы PM-1;
плазмиду pPM-h1, содержащую ген, кодирующий V-область H-цепи, полученный из гибридомы PM-1;
плазмиду p64-k4, содержащую ген, кодирующий V-область к L-цепи, полученный из гибридомы AUK 64-7;
плазмиду p64-h2, содержащую ген, кодирующий V-область h-цепи, полученный из гибридомы AUK 64-7;
плазмиду p146-k3, содержащую ген, кодирующий V-область к L-цепи, полученный из гибридомы AUK 146-15, и
плазмиду p146-h1, содержащую ген, кодирующий V-область H-цепи, полученный из гибридомы AUK 146-15.
Заметим, что штаммы E. coli, содержащие вышеупомянутые плазмиды, были помещены на хранение в Национальную Коллекцию Промышленных и Морских бактерий, действующую в рамках Будапештского договора от 11 февраля 1991 года, и получили регистрационные номера, показанные в таблице 8.
Пример 3. Секвенирование ДНК
Нуклеотидные последовательности кДНК кодирующей области в вышеупомянутых плазмидах определяли, используя набор Sequenase(тм) вариант 2.0 (U.S. Biochemical Corp. USA).
Сначала примерно 3 мкг плазмидной ДНК, полученной как описано выше, денатурировали с помощью 2,0 N NAOH, ренатурировали с секвенирующим праймером и метили 35S-dATP по прописи поставщика. Затем меченную ДНК помещали на 6% полиакриламидный гель, содержащий 8 М мочевины, и после электрофореза гель фиксировали 10% метанолом и 10% уксусной кислотой, высушивали и подвергали ауторадиографии, чтобы определить нуклеотидную последовательность.
Нуклеотидная последовательность кДНК колирующей области каждой плазмиды представлена в SEQ ID NO: с 24 до 31.
Пример 4. Определение CDR.
Основные структуры V-областей L-цепи и H-цепи похожи одна на другую, причем 4 остовные области (FR) связываются через 3 супервариабельные области, т. е. области, определяющие комплементарность (CDR). В то время как аминокислотные последовательности в FR относительно хорошо сохранны, аминокислотные последовательности в CDR очень высоко вариабельны (Kabat, E.A. et al. , "Sequences of Proteins of Immunological Interest", US Dept. Heath and Human Services 1983).
На основе определенных выше аминокислотных последовательности V-областей мышиных моноклональных антител к человеческим IL-6R и по Kabat, E.A. et al., "Sequences of Proteins of Immunological Unterest", US Dept. Heath and Human Services, 1983, были определены CDR каждой V-области мышиных моноклональных антител к человеческим IL-6R, которые представлены в таблице 9.
Пример 5.
Подтверждение экспрессии клонированной кДНК (I)
(Создание химерных антител AUK 12-20)
Создание плазмиды экспрессии
Химерная L-цепь/H-цепь конструировалась из клонированной с PCR кДНК, кодирующей V-области к L-цепи и H-цепи AUK 12-20. Чтобы легко соединять кДНК, кодирующую V-область мышиных AUK 12-20 с ДНК, кодирующей человеческую C-область в векторе экспрессии в клетках млекопитающих, содержащем энхансер и промотор человеческого цитомегаловирусного ( HCMV) вектора экспрессии, необходимо ввести подходящие сайты расщепления для рестрикционного фермента в 5'- и 3' - концы мышиной кДНК.
Эта модификация 5' -и 3'- концов выполнялась методом PCR. Были разработаны и синтезированы два набора праймеров. Запаздывающий праймер V-области L-цепи (SEQ ID NO:32) и запаздывающий праймер V-области H-цепи (SEQ ID NO: 33) были разработаны так, что праймеры гибридизуются с ДНК, кодирующей начало лидерной последовательности, сохраняют последовательность ДНК, необходимую для эффективной трансляции (Kazak M.J. Mol. Biol, 196: 947 - 950, 1987) и образуют сайт Hind III для клонирования в HCMV вектор экспрессии. Ранний праймер V-области L-цепи (SEQ ID NO: 34) и ранний праймер V-области H-цепи (SWQ ID NO: 35) разрабатывались так, что праймеры гибридизируют с ДНК, кодирующей концевую часть J-области, сохраняют последовательность ДНК, необходимую для сплайсинга в C-область и формируют сайт Bam НI для соединения с человеческой C-областью в HCMV вектора экспрессии.
После амплификации путем PCR продукт PCR расщеплялся с помощью Hind III и Bam HI, клонировался в HCMV вектор, содержащий ДНК C-областей человеческих к и γI цепей, и секвенировался, чтобы подтвердить, что не были введены ошибки во время амплификации PCR. Получение в результате векторы экспрессии обозначаются как HCMV-12k-g к и HCMV-12h-gγI.
Структуры экспрессирующих плазмид представлены на фиг. 1. В плазмиде HCMV-VL-HCK VL-область может быть любой V-областью мышиной L-цепи. В этом примере была встроена область к цепи AUK 12-20, чтобы получить HCMV-12k. В плазмиде HCMV-VH-HCγI, VH область может быть любой V-областью мышиной H-цепи. В этом примере была встроена V-область H-цепи AUK 12-20, чтобы получить HCMV-12h-gγI.
Временная экспрессия в cos клетках
Чтобы проследить временную экспрессию химерных AUK 12-20 антител в cos клетках, векторы экспрессии, сконструированные как описано выше, были испытаны на cos клетках. Вектор ДНК вводили в cos клетки путем электропорации, используя Gene Putsar аппарат (Bio Rad). А именно, cos клетки суспендировали в фосфатно-буферном физиологическом растворе (PBS) до клеточной концентрации 1•107 клеток/мл, и к 0,8 мл аликвотной пробе суспензии добавляли 10 мкг (на каждую плазмиду) ДНК. Подводили импульсы с 1900 V и 25 мк.
После периода восстановления в 10 минут при комнатной температуре электропарированные клетки добавляли к 8 мл DMEM (GIBCO) (модифицированной по способу Дульбекко среды Игла), содержащей 10% фетальной бычьей сыворотки. После инкубации в течение 72 часов супернатант культуры отбирали, центрифугировали, чтобы удалить остатки клеток, и хранили асептически в течение короткого периода при 4oC или в течение длительного периода при -20oC.
Количественное определение химерных антител путем ELISA
Супернатант культуры трансфицированных cos клеток исследовался с помощью ELISA для подтверждения продукции химерных антител. Чтобы определить химерные антитела плату покрывали козлиными против-человеческого IgG цельномолекулярным (Sigma). Плату блокировали и в каждую ячейку добавляли серийно разведенный супернатент из культуры cos клеток. После инкубации и промывания добавляли связанный с щелочной фосфатазой козлиный против-человеческого IgG (специфическая γ- цепь, Sigma) в каждую ячейку. После инкубации и отмывания вслед за тем добавляли субстратный буфер. Реакционную смесь инкубировали и после прекращения реакции измеряли оптическую плотность при 405 нм. В качестве стандарта использовался очищенный человеческий IgG (Sigma).
ELISA для подтверждения способности связывать человеческие IL-6R.
Супернатант культуры трансформированных cos клеток исследовали с помощью ELISA, чтобы определить продуцируются ли антитела, которые могут связываться с антигеном. Для определения связывания с антигеном плату покрывали мышиными моноклональными антителами MT18 (см. пример 1) и после блокирования 1% бычьим сывороточным альбумином (BSA) добавляли растворимые рекомбинатные человеческие IL-6R (SP 344). После промывания добавляли серийно разведенный супернатант культуры клеток cos в каждую ячейку. После инкубации и отмывания добавляли связанный с щелочной фосфатазой козлиный против-человеческого IgG. Реакционную смесь инкубировали и после промывания добавляли субстратный буфер. После инкубации реакция закачивалась и определялась оптическая плотность при 450 нм.
Результат показан на фиг. 2. Трансфекция гена, кодирующего химерные антитела AUK 12-20, в cos клетки повторялась дважды. Образцы обоих супернатантов культур проявляли сильное связывание с IL-6R и оптическая плотность при 405 нм изменялась с разведением образца (концентрацией моноклональных антител) - характер зависимости показан на фиг. 2 пустыми кружочками и закрашенными кружочками, обнаруживая наличие антител к IL-6R в образце.
Определение способности подавлять связывание IL-6R с IL-6
Чтобы определить подавляют ли антитела, присутствующие в среде, связывание IL-6R с IL-6, плату покрывали моноклональными антителами MT18 (справочный пример 1). После блокирования добавляли растворимые рекомбинантные человеческие IL-6 (SR 344). После промывания добавляли серийно разведенные образцы из клеточной культуры cos в каждую ячейку с биотинированным IL-6.
После отмывания добавляли стрептоавидин, связанный с щелочной фосфатазой, и после инкубации и отмывания добавляли субстратный буфер. Реакционную смесь инкубировали и после прекращения реакции измеряли оптическую плотность при 405 нм; очищенные мышиные моноклональные антитела AUK 12-20 добавляли в качестве положительного контроля, и культуральную среду cos клеток, экспрессирующих несвязывающие антитела, использовали в качестве отрицательного контроля.
Результат показан на фиг. 3. Супернатант культуры cos клеток, трансфицированных генами, кодирующими химерные антитела AUK 12-20, проявляли связывание IL-6R с IL-6 при наивысших и вторых за наивысшими концентрациях. А именно, как показано закрашенными кружочками на фиг. 3, оптическая плотность при 405 нм изменялась в зависимости от разведения образца (концентрация антител), обнаруживая подавление связывания IL-6R с IL-6 антителами образца. Это, кроме того, подтверждается независимым соответствием изменения положительного контроля (открытые кружочки), зависящим от концентрации антител. Заметим, что отрицательный контроль не проявляет подавляющей активности (незакрашенные треугольники).
Пример 6.
Подтверждение экспрессии клонированной кДНК (2)
(Создание химерных антител PM-1)
Создание векторов экспрессии
Чтобы создать векторы, экспрессирующие химерные антитела PM-1, клоны кДНК pPM-k3 и pPM-h1, кодирующие V-область к L-цепи и H-цепи мышиных PM-1, соответственно модифицировали методом PCR и затем вводили в HCMV векторы экспрессии (см. фигуру 1). Были разработаны запаздывающие праймеры pmK-S (SEQ N0: 38) для V-области L-цепи и pmh-S (SEQ N0: 40) для V-области H-цепи, чтобы гибридизовать к ДНК последовательностям, кодирующим начало лидерных последовательностей, и чтобы получить согласующуюся последовательность Kozak и сайт рестрикции Hind III. Ранние праймеры pmk-a (SEQ N0: 36) для V-области L-цепи и pmh-a (SEQ N0: 39) для области H-цепи разработаны, чтобы гибридизовать к последовательностям ДНК, кодирующим концы J-областей и чтобы получить сплайсинговую донорную последовательность и сайт реструкции Bam HI.
Для V-области каппа L-цепи были синтезированы два ранних праймера. Хотя в большинстве каппа L-цепей лизин в положении 107 сохраняется, в каппа L-цепи мышиных PM-1 в положении 107 находится аспарагин. Чтобы исследовать эффект этого изменения на активность химерных антител PM-1 по связыванию антигена, был разработан ранний праймер pmk-b (SEQ N0: 37), чтобы изменить положение 107 с аспарагина на лизин. После реакции PCR продукты PCR очищали, расщепляли с помощью Hind III и Bam HI и субклонировали в вектор pUC19 (Yanishe-Perron et al., Gene (1985) 33: 103 - 109). После секвенирования ДНК фрагменты Hind III - Bam HI вырезались и клонировались в вектор экспрессии HCMV-VH-HCγI, чтобы получить HCMV-pmh-gγI для химерной H-цепи, и в вектор экспрессии HCMV-VL-HCK, чтобы получить HCMV-pmka-gk и HCMV-pmkb-gk для химерной L-цепи.
Трансфекция cos клеток
Векторы испытывали в cos клетках, чтобы обнаружить временную экспрессию химерных человеческих антител PM-1. HCMV-pmh-gγI и или HCMV-pmka-gk, или HCMV-pmkb-gk трансфицировали вместе в cos клетки с помощью электропорации, используя аппарат Gene Pulsar (Bio Rad), ДНК (10 мкг каждой плазмиды) добавляли к 0,8 мл аликвотным образцам из 1 •107 клеток/мл в PBS. Импульсы подводились при 1900 вольт, емкостью 25 микрофарад. После 10 мин периода восстановления при комнатной температуре электропорированные клетки добавляли к 20 мл модифицированной по Дюльбекко среды Игла (DMEM) (GIBCO), содержащим 10% фетальной телячьей сыворотки свободной от гамма-глобулина. После 72 часов инкубации среду собирали, центрифугировали, чтобы удалить клеточные остатки, и хранили в стерильных условиях при 4oC в течение коротких периодов времени или при -20oC в течение длительных периодов.
Экспрессия и анализ химерных антител PM-1
После 3 дней временной экспрессии среду от cos клеток отбирали и испытывали на наличие химерных антител PM-1. Сначала среду анализировали с помощью ELISA, чтобы определить продуцируются ли трансфицированными cоs клетками антитела подобные человеческим. Путем использования известных количеств очищенных человеческих IgG в качестве стандарта в этом исследовании можно также определить количество антител, подобных человеческим (в этом случае, химерные антитела PM-1), присутствующих в среде от cos клеток. Для определения человеческих антител платы покрывали козлиным противочеловечьим IgG (целые молекулы, Sigma). После блокирования образцы от cos клеток серийно разводили и добавляли в каждую ячейку. После инкубации и отмывания добавляли козий античеловеческий IgG, конъюгированный с щелочной фосфатазой (специфические гамма-цепи, Sigma). После инкубации и отмывания добавляли субстратный буфер. После инкубации реакцию останавливали и измеряли оптическую плотность при 405 нм. В качестве стандарта использовался очищенный человеческий IgG Sigma.
Среда от cos клеток, трансфицированных векторами, несущими гены химерных PM-1, была положительной при исследовании на экспрессию антител, подобных человеческим, и примерные их количества определялись, как описано.
Затем, та же самая среда от cos клеток, трансфицированных векторами, несущими гены химерных PM-1, исследовалась на способность связывать человеческие IL-6R. Для определения связывания с антигеном платы покрывали мышиными моноклональными антителами MT18 (справочный пример 1), антителами к человеческим IL-6R. После блокирования добавляли растворимые рекомбинантные человеческие IL-6R (SR 344). После отмывания образцы разводили серийно и добавляли в каждую ячейку. После инкубации и промывания добавляли козий противочеловеческий IgG (гамма-цепь специфическая, Sigma). После инкубации и промывания добавляли субстратный буфер. После инкубации реакцию останавливали и определяли оптическую плотность при 405 нм. Стандарта для этого исследования нет.
Два образца были после трансфекации генов, кодирующих химерные антитела с V-областями, идентичными областям, обнаруженным в мышиных антителах PM-1 (химерных антителах PM-1a, фигура 4). Один образец был после трансфекции генами, кодирующими химерные антитела с заменой единственной аминокислоты, в положении 107 V-области L-цепи, как описано выше (химерные антитела PM-1b, фигура 4). Все образцы продемонстрировали сильное связывали с IL-6R, которое снижается с разбавлением образца. Таким образом, химерные антитела PM-1, которые сконструированы, являются функциональными и могут хорошо связываться с их антигеном. Наиболее важно, что демонстрация функциональности химерных PM-1 является прямым доказательством того, что были клонированы и секвенированы правильные V-области мышиных PM-1. Химерные антитела PM-1 с любой из двух аминокислот в 107 положении цепи хорошо связывались с их антигенами IL-6R. Очевидно, что положение 107 в V-области L-цепи мышиных PM-1 не является самым решающим в связывании антигена и что аспарагин или лизин в этом положении будут функционировать удовлетворительно. Так как мышиные антитела PM-1 имеют аспарагин в этом положении V-области L-цепи, вся дальнейшая работа с химерными антителами PM-1 выполнялась с вариантом, который имеет V-области, идентичные областям, обнаруживаемым в мышиных антителах PM-1.
Чтобы стабильно получать большие количества химерных антител PM-1, был создан новый HCMV вектор экспрессии, включающий ген dhfr. Первым шагом в достижении более высоких уровней экспрессии химерных антител PM-1 была модификация вектора HCMV-Vн-HCγI (фигура 1) таким образом, чтобы этот вектор содержал ген dhfr, который должен экспрессироваться "поврежденным" промотором-энхансером SV40. Энхансерные элементы SV40 удалялись из вектора PSV2-dgfr (S. Subramani et al., Mol. Cell. Biol. (1981) 1:854-864) и ген dhfr, который должен экспрессироваться "поврежденным" промотором SV40, встраивался в вектор HCMV-VH-HCγI вместо neo-гена, экспрессируемого промотором-энхансером SV40. V-область мышиных PM-1 встраивали затем в этот новый HCMV-VH-HCγI- dhfr вектор. Создание вектора с улучшенной экспрессией описывается детально в примере 10.
CHO dhfr (-) клетки (G. Urlaub et al., Proc. Natl. Acad. Sci. USA (1980) 77: 4216-4220) трансфицировали одновременно двумя плазмидными ДНК, вектором HCMV-VL-HCk для экспрессии L-цепи химерных PM-1a (HCMV-pmka-gk) и вектором HCVM-VH-HCγI-dhfr для эксперссии H-цепи химерных PM-1 (DHFR -ΔE PMh-gγI ; пример 10). ДНК (10 мкг/мл каждой плазмиды) добавляли к 0,8 мл аликвотным образцам с 1 • 107 клеток/мл в PBS. Подводили импульс с напряжением 1900 вольт, емкостью 25 микрофарад. После 10-минутного периода восстановления при комнатной температуре электропарированные клетки добавляли к 10 мл минимальной необходимой среды альфа (Alpha minimum essential medium (альфа-MEM), содержащей нуклеозиды и 10% FCS. После инкубации в течение ночи среду заменяли на α-MEM без нуклеозидов и с 10% FCS - 500 мкг/мл G418 (GIBCO) для селекции dhfr+ и neo+ трансформированных клеток. После селекции отобранные клоны использовали для амплификации генов. После одного круга амплификации в 2 • 10-8 М метотрексате (МТХ) отбиралась линия клеток (PM-1k 3-7), которая продуцировала примерно 3,9 мкг/106 клеток/сутки химерных антител PM-1a.
Исследование ELISA на способность химерных антител подавлять связывание IL-6 с человеческими IL-6R
Антитела, продуцируемые трансфицированными клетками cos или устойчивыми линиями клеток CHO, исследовали, чтобы определить, могут ли эти антитела конкурировать с биотинилированным IL-6 за связывание с IL-6R. Платы покрывали мышиными моноклональными антителами MT18. После блокирования добавляли растворимые человеческие IL-6R (SR 344). После отмывания образцы из cos клеток серийно разводили и добавляли вместе с биотинилированными IL-6 в каждую ячейку. После отмывания добавляли стрептавидин, конъюгированный с щелочной фосфатазой. После инкубации и отмывания добавляли субстратный буфер. После инкубации реакцию останавливали и измеряли оптическую плотность при 405 нм. Результаты показаны на фиг. 5.
Пример 7.
Создание видоизмененных человеческих антител PM-1
Чтобы достичь переноса CDR более быстро и эффективно, был разработан метод секвенциального CDR-переноса путем PCR. Этот метод основан на методах PCR-мутагенеза (Kamman et al., 1989).
Чтобы получить матричную ДНК, содержащую выбранные человеческие FR, для переноса CDR, было необходимо реклонировать соответственно измененные V-области в подходящие векторы. Плазмидные ДНК alys II и F10 кодируют видоизмененные человеческие D1.3 L- и H-цепи и содержат FR из человеческих REI и NEW соответственно. Фрагмент примерно 500 bp Ncol-Bam HI, содержащий ДНК-последовательность, кодирующую V-область видоизмененной D1.3 L-цепи, вырезался из alys II и субклонировался в расщепленную Hind III - Bam HI pBR327, чтобы получить вектор V1 - lys - pBR327. Фрагмент Hind III-Bam HI из V1 - lys-pBR327 встраивался в pUC19, расщепленную в Hid III-Bam HI, чтобы получить вектор V1 - lys - pUС19. Фрагмент примерно в 700 bp Ncol-Bam HI, содержащий ДНК последовательность, кодирующую V-область видоизмененной человеческой D1.3 H-цепи, вырезался из F10 и субклонировался в сайт Hind III-Bam HI вектора pBR327, с использованием Hind III-Ncol адаптора и получением Vh - lys - pBR327. Затем из этого вектора вырезался фрагмент Hind III - Bam HI и субклонировался в расщепленный Hind III-BamHI вектор pUC19 с получением Vh - lys - pUC19.
Обратите внимание, что создание плазмиды alys II и последовательности ДНК, кодирующей V-область видоизмененной человеческой D1.3 L-цепи, использованных в матрице, описаны, ДНК последовательность, кодирующая V-область видоизмененной человеческой D1.3 H-цепи в плазмиде F10, использованной в качестве матрицы, описывается у V. Verhoey et al., Science 237: 1534-1536 (1988), фиг. 2.
Фигура 6 показывает на схеме праймеры и реакции PCR, использованные при создании первого варианта V-области H-цепи видоизмененных человеческих PM-1. Поздний праймер A (APCR1; SEQ N0: 41) и ранний праймер E (APCR4; SEQ N0 : 42) гибридизуются с ДНК последовательностями в векторе. Хотя для pUC19 вектора были в частности разработаны APCR1 и APCR4, могли бы использоваться и праймеры с универсальной последовательностью M13.
Мутагенный праймер B CDR1-пересадки (phV-1; SEQ N0: 43), праймер C CDR1-пересадки (phv - 2; SEQ N0: 44) и праймер CDR3-пересадки (phV-3; SEQ N0: 45) были длиной 40-60 bp, состоящими из ДНК последовательностей, кодирующих CDR V-области H-цепи мышиных PM-1 и человеческие FR в матричной ДНК, которая находится по краям CDR областей. В первой реакции PCR использовались ранний праймер APCR4 и поздний праймер D. Первый продукт PCR, который содержит CDR3 последовательность мышиных PM-1, очищали и использовали во второй реакции PCR в качестве раннего праймера с праймером C в качестве позднего праймера. Тем же путем второй и третий продукты PCR, которые содержат CDR2 и CDR3 мышиных PM-1, и все три CDR мышиных PM-1 соответственно использовали в качестве праймеров в следующем этапе PCR. Четвертый продукт PCR, который имеет полную V-область H-цепи видоизмененных человеческих PM-1, очищали, разрезали Hind III и Bam HI и сублонировали в вектор pUC19 для дальнейшего анализа.
Три мутагенных праймера phv-1, phv-2 и phv-3 были синтезированы для конструирования V-области H-цепи видоизмененных человеческих PM-1. Они были очищены на 12% полиакриламидном геле, содержащем 8 М мочевины. Мутагенный праймер phv-1 был разработан не только для пересадки CDR1 мышиных PM-1, но также для замен в положениях 27 и 30 в человеческих FR1, Ser на Tyr и Ser на Thr соответственно. Каждые 100 мкл при реакции PCR обычно содержали 10 мМ Трис-HCl (pH 8,3), 50 мМ KCl, 1,5 мМ MgCl2, 250 мкМ dNTP, 50 нг матричной ДНК (Vh - lys - pUC19), 2,5 ед Amply Tag (Perkin Elmer Cetus) и праймеры. Первая реакция PCR при содержании 1 мкМ каждой из phv-3 и праймеров APCR4 проводилась после первоначальной денатурации при 94oC в течение 1,5 мин и 30 циклов при 94oC в течение 1 минуты, при 37oC в течение 1 мин и 72oC в течение 1 минуты были повторены. Время перерыва между отжигом и стадиями синтеза было установлено в 2,5 мин. После завершения последнего цикла следовало финальное развитие реакции при 72oC в течение 10 мин. Продукт PCR в 523 bp очищали, используя 1,6% агарозный гель с низкой температурой плавления, и затем использовали в качестве праймера во второй реакции PCR.
Во второй реакции PCR в качестве праймеров использовали примерно 1 мкг очищенного первого продукта PCR и 25 пкМ мутагенного праймера phv-2. Условия PCR были теми же самыми, что описаны для первой реакции PCR. Таким же образом, продукт PCR 665 bp из второй реакции PCR и продукт PCR 737 bp из третьей реакции PCR использовались в качестве праймеров в третьей реакции PCR с праймером phv-1 и в четвертой реакции PCR с праймером APCR1 соответственно. Продукт PCR из четвертой реакции PCR очищали, разрезали Hind III и Bam HI и затем фрагмент примерно 700 bp, содержащий V-область H-цепи видоизмененных человеческих PM-1, субклонировали в вектор pUC19. Два из четырех секвенированных клонов имели последовательность ДНК, кодирующую правильную аминокислотную последовательность и были обозначены pUС-RVh-PM1a.
Чтобы создать другие варианты V-области H-цепи видоизмененных PM-1, были синтезированы пять мутагенных праймеров PCR. Каждая реакция PCR в основном проводилась при тех же условиях, что описаны выше. Для варианта "b" были использованы: мугатенный праймер phv-4 (Val-71 до Arg-71; номер дан по Kabat et al; см. таблицу 3) (SEQ ID N0: 46) и APCR4 в первой реакции PCR с матричной ДНК, pUC-RVh-PM1a. Продукт PCR из этой первой реакции PCR очищался и использовался в качестве раннего праймера во второй реакции PCR с праймером APCR1. Продукт PCR из второй реакции PCR очищали, используя 1,6% агарозный гель с низкой температурой плавления, расщепляли с помощью Hind III и Bam HI и субклонировали в вектор pUC19, получая pUC-RVh-PM-1b. Таким же образом получали вариант "c" (pUC-RVh-PM-1c), используя мугатенный праймер phv-nm (Asp-1 до Gln-1) (SEQ ID N0:47) и матрицу pUC-RVh-PM-1b; вариант "d" pUC-RVh-PM-1d) получали, используя мутагенный праймер phv-m6 (11e-48 до Met-48) (SEQ ID N0: 48) и матрицу pUC-RVh-PM-1b; вариант "e" (pUC-RVh-PM-1e) получали, используя мутагенный праймер phv-6 и матрицу pUC-RVh-PM-1c; вариант "f" (pUC-RVh-PM-1f) получали, используя мутагенный праймер phv-7 (Thr-28 до Ser-28, Phe-29 до 11e-29) (SEQ ID N0: 49) и матрицу pUC-RVh-PM-1b. Аминокислотная последовательность варианта "f" V-области видоизмененной человеческой H-цепи и нуклеотидная последовательность, ее кодирующая, показаны в SEQ ID N0: 54.
Фигура 7 представляет диаграммы праймеров и реакции PCR, использованные при создании первого варианта V-области L-цепи видоизмененных человеческих PM-1. Для построения первого варианта V-области L-цепим видоизмененных человеческих PM-1 были синтезированы праймер переноса CDR1 pkv-1 (SEQ ID N0: 50), праймер переноса CDR2 pkv-2 (SEQ ID N0: 51) и праймер переноса CDR3 pkv-03 (SEQ ID N0:52) и очищали на 12% полиакриламидном геле, содержащем 8 М мочевину. Реакции PCR проводили как описано выше. Первая реакция PCR содержала 1 мкМ каждого pkv-3 и APCR4 праймера. Продукт PCR 350 bp из первой реакции PCR очищали, используя 1,5% агарозный гель с низкой температурой плавления, и использовали в качестве раннего праймера во второй реакции PCR. Продукт PCR из второй реакции PCR очищали, расщепляли Bam HI и Hind III и фрагмент 500 bp, содержащий пересаженную к CDR ДНК, субклонировали в вектор pUC19 или секвенирования ДНК. Была идентифицирована плазмидная ДНК, имеющая правильную последовательность и использована в качестве матричной ДНК в последующей реакции PCR. В третьей реакции PCR было использовано 25 пкмолей мутагенных праймеров pkv-2 и APCR4. Продукт PCR из третьей реакции PCR очищали и использовали в качестве праймера с праймером pkv-1 в четвертой реакции PCR. Таким же образом, продукт PCR из четвертой реакции PCR использовали в качестве праймера с праймером APCR1 в пятой реакции PCR.
Продукт PCR 972 bp из пятой реакции PCR очищали, расщепляли Bam HI и Hind III и субклонировали в вектор pUC19 для секвенирования ДНК. Была определена проблема в области CDR2. Были необходимы дополнительные реакции PCR. В шестой и седьмой реакциях PCR продукт PCR из пятой реакции PCR, который клонирован в вектор pUC19, использовался в качестве матричной ДНК. В шестой реакции PCR праймерами были pkv-2 и APCR4. Продукт PCR из шестой реакции PCR очищали и использовали в качестве праймера с праймером APCR1 в седьмой реакции PCR. Продукт PCR седьмой реакции PCR очищали, расщепляли Bam HI и Hind III и фрагмент ДНК 500 bp субклонировали в вектор pUC19 для секвенированния ДНК. Два из пяти секвенированных клонов имели правильные последовательности. Клон был обозначен pUC-RVI-PM-1a. Последовательность показана в SEQ IN N0: 55.
Для создания другого варианта V-области L-цепи видоизмененных человеческих антител PM-1 был синтезирован мутагенный праймер pvK-mI (SEQ ID N0: 53). Реакции PCR были в основном такими, как описаны выше. В первой реакции PCR использовался мутагенный праймер pkv-mI (Phe-71 по Tyr-71) и праймер APCR4 с матричной ДНК pUC-RVI-PM-1a. Продукт PCR первой реакции PCR очищали и использовали в качестве праймера с праймером APCR1 во второй реакции PCR. Продукт PCR во второй реакции PCR очищали, расщепляли Bam HI и Hind III и субклонировали в вектор pUC19 для секвенирования ДНК. Клон обозначали pUC-RVI-PM-1b.
Пример 8.
Создание векторов, в которых используется ранний промотор человеческого цитомегаловируса (HCMV), для экспрессии генетически сконструированных антител в клетках млекопитающих (фиг. 1).
Фрагменты ДНК, кодирующие V-области L- и H-цепей химерных PM-1, были первоначально встроены в векторы HCMV (HCMV-VL-HCk и HCMV-VH -HCγI) разработанные для экспрессии или человеческих каппа L-цепей или человеческих гамма-I H-цепей в клетках млекопитающих (смотрите фигуру 1). Детальное описание создания HCMV векторов экспрессии публикуется у Maeda et al., Human Antibodies and Hybridomas (1991) 2:124-134; C.A. Kettleborough et al., Protein Engeneering (1991) 4: 773-783. Оба вектора базируются на psv-2neo (P.J. Southerm et al. , J. Mol. Aplln. Genet (1962) 1:327-341) и содержат промотор человеческого цитомегаловируса (HCMV) и энхансер (M.Boshart et al., Cell (1985) 41: 521-530) для высокого уровня транскрипции L- и H-цепей иммуноглобулина. Вектор экспрессии L-цепи содержит геномную ДНК, кодирующую человеческую каппа C-область (T.H. Rabbitts et al., Curr Top. Microbiol. Immunol. (1984) 113: 166-171), и вектор экспрессии H-цепи содержит геномную ДНК, кодирующую человеческую гамма-I C-область (N.Takahashi et al., Cell (1982) 29: 671-679). Вектор экспрессии HCMV является многосторонним и может быть использован как для временной, так и для стабильной экспрессии в ряде типов клеток млекопитающих.
Пример 9.
Создание векторов, в которых используется промотор человеческого фактора элонгации Iα(HEF-Iα) для экспрессии генетически сконструированных антител в клетках млекопитающих (фиг. 8 и фиг. 9)
Фактор элонгации человеческих полипептидных цепей Iα (HEF-Iα) является одним из наиболее распространенных белков. Он экспрессируется в большинстве клеток. Транскрипционная активность человеческого промотора-энхансера EF-Iα примерно в 100 раз сильнее, чем у раннего промотора-энхансера SV40 (D.W.Kim et al., Gene (1990) 91: 217-223, и T. Uetsuki et al., J. Biol. Chem. (1989) 264: 5791-5798). Промоторно-энхансерная область HEF-I α 2,5 kb состоит из ДНК примерно 1,5 kb, находящейся на 5'-конце гена, из 33 bp в первом экзоне, 943 bp в первом интроне и 10 bp в первой части второго экзона. Фрагмент Hind III-EcoRI примерно 2,5 kb вырезался из плазмидной ДНК pEF321-CAT (D.W.Kim et al. , Gene (1990) 91: 217-223 и T.Uetsuki et al., J.Biol. Chem. (1989) 264: 5791-5798) и клонировался в вектор ДНК pdKCR (M.Tsushiya et al., EMBO (1987) 6: 611-616) (K. O'Hate et al. , Proc. Natl. Acad. Sci USA Vol. 78, N 3, 1527-1531, 1981), чтобы заменить фрагмент в примерно 300 bp Hind III-EcoRI, содержащий последовательность раннего промотора-энхансера SV40, с получением таким образом pTEF-1. pTEF-1 разрезался EcoRI, заполненным полимеразой Кленова, и лигировали Hind III линкерами. Фрагмент Hind III-Smal примерно в 1,6 kb затем вырезался из модифицированного вектора ДНК pTEF-1.
Плазмида HCMV-12h-gγI(ΔE2) разрезалась EcoRI, заполнялась полимеразой Кленова и расщеплялась Hind III. Полученный в результате фрагмент примерно в 7 kb, содержащий последовательность ДНК, кодирующую человеческую гамма-I C-область, лигировали с полученным ранее (выше) фрагментом Hinf III-Smal 1,6 kb, содержащим промотор-энхансер HEF-Iα, получая HEF-12h-gγI. Область промотора-энхансера HEF-Iα в этом векторе была такой же, как и в pTEF-1, за исключением 380 bp ДНК, находящейся на конце 5'-области. Область H-цепи, присутствующая как фрагмент Hind III-Bam HI, была способна легко взаимозаменяться другими V-областями H-цепи. Фрагменты ДНК Hind III-Bam HI, содержащие область видоизмененной H-цепи, вырезались из pUC-RVh-PM-1a, pUC-RVh-PM-1b, pUC-RVh-PM-1c, pUC-RVh-PM-1d, pUC-RVh-PM-1e и pUC-RVh-PM-1f (пример 7) и встраивались в часть Hind III-Bam HI HEF-12h-gγI, чтобы получить экспрессию векторов RVh-PM-1a, RVh-PM-1b, RVh-PM-1c, RVh-PM-1d, RVh-PM-1e и RVh-PM-1f соответственно. Векторы экспрессии RVh-PM-1a, RVh-PM-1b, RVh-PM-1c, RVh-PM-1d, RVh-PM-1e и RVh-PM-1f так же, как и HEF-PMh-gγI содержали варианты "a", "b", "c", "d", ""e" и "f" V-областей H-цепи видоизмененных человеческих PM-1 соответственно.
Чтобы создать вектор экспрессии цепи, HEF-12k-gk, фрагмент PVUI-Hind III примерно 3,0 kb, содержащий область HEF-Iα промотора-энхансера, вырезался из HEF-12h -gγI и лигировался к фрагменту PVUI-Hind III примерно в 7,7 kb из HCMV вектора экспрессии L-цепи HCMV-12k-gk, сконструированного в примере 5, чтобы получить HEF-12k-gk. Так же как и вектор экспрессии HEF-12h-gγI H-цепи, V-область L-цепи в HEF-12k-gk, присутствующая в качестве Hind III-Bam HI фрагмента, является легко взаимозаменяемой другими V-областями L-цепи.
Фрагменты ДНК Hind III-Bam HI, содержащие V-область видоизмененной L-цепи, вырезали из pUC-RVI-PM-1a и pUC-RVI-PM-1b (пример 87) и встраивали в часть Hind III-Bam HI HEF-12k-gk, чтобы получить векторы экспрессии RVI-PM-1a и RVI-PM-1b соответственно. Векторы экспрессии RVI-PM-1a, RVI-PM-1b и HEF-PMK-gk содержат "a", "b"-области видоизмененной человеческой L-цепи и V-область цепи мышиных PM-1 соответственно.
Пример 10.
Создание векторов, в которых используется ген дигидрофолатредуктазы (dhfr), связанный с дефектной промоторно-энхансерной последовательностью SV40 для достижения высоких уровней экспрессии генетически сконструированных антител в клетках CHO (фиг. 10 и фиг. 11)
Чтобы удалить энхансерную последовательность из раннего промотора SV40, плазмидную ДНК pSV2-dhfr (S. Subramani et al., Mol. Cell. Biol. (1981) 1: 854-864) (АТСС 33694) расщепляли с помощью SphI и PVUII, заполняли полимеразой Кленова и подвергали самолигированию, чтобы получить pSV2-dhfr-ΔE (см. фигуру 10). Фрагмент EcoRI примерно 3,7 kb, содержащий промотор HCMV, V-область H-цепи и человеческую гамма-I C-область вырезался из HCMV-PMh -gγI путем частичного расщепления с помощью EcoRI. Этот фрагмент лигировали к EcoRI, расщепленному pSV2-dhfr -ΔE, чтобы получить DHFR-ΔE-PMh-gγI.
Сходный вектор создали на основе вектора экспрессии H-цепи, в котором используется HEF-Iα промотор-энхансер (см. фигуру 11). Фрагмент EcoRI примерно 3,7 kb, полученный из HCMV-12h-gγI лигировали с EcoRI-расщепленной pSV2-dhfr-ΔE , чтобы получить DHFR-ΔE-PMh-gγI cайт Bam HI, следующий за кДНК последовательностью в DHFR-ΔE-12h-gγI, удаляли путем частичного расщепления Bam HI, заполнения полимеразой Кленова и самолигирования. Фрагмент PVUI-Bam HI примерно 4 kb, содержащий кДНК dhfr, вырезали из модифицированной ДНК DHFR-ΔE-12h-gγI и лигировали к фрагменту PVUI-Bam HI примерно 3 kb из RVh-PM-1f-4 (созданного в примере 12), чтобы получить DHFR -ΔE- RVh-PM-1f.
Плазмиды с улучшенной экспрессией, которые получены как представлено выше, могут использоваться для продукции видоизмененных человеческих антител PM-1 настоящего изобретения.
Пример 11.
Экспрессия и анализ различных вариантов видоизмененных человеческих антител PM-1
HEF-Iα векторы, экспрессирующие L- и H-цепи видоизмененных человеческих PM-1, трансфицировали вместе в cos клетки. В качестве стандартного контроля HEF-Iα векторы, экспрессирующие и H-цепи химерных PM-1, также вместе трансфицировали в cos клетки. После 3 дней среду из трансфицированных cos клеток собирали и анализировали с помощью ELISA (1) на количество человеческих IgG антител, присутствующих в супернатанте, и (2) на способность этого человеческого IgG связываться с IL-6R. Затем те же образцы также испытывали с помощью ELISA на способность антител подавлять связывание человеческих IL-6 с человеческими IL-6R.
Оценка двух вариантов V-областей L-цепи видоизмененных человеческих PM-1 проводилась с cos клетками, трансфицированными одновременно одним из двух векторов, экспрессирующих L-цепи видоизмененных человеческих PM-1 (RVI-PM-1a или RVI-PM-1b), и вектором, экспрессирующим H-цепь химерных PM-1 (HCMV-PMh-gγI) . Клетки также одновременно трансфицировали векторами, экспрессирующими L- и H-цепи химерных PM-1 (HCMV-PMKa-gk и HCMV-PMh-gγI . Данные с использованием неочищенных супернатантов cos клеток показали, что вариант "a" L-цепи видоизмененных человеческих PM-1 был эквивалентен L-цепи химерных PM-1 в исследованиях по связыванию с IL-6R. Вариант "b" L-цепи видоизмененных человеческих PM-1, однако, фактически уничтожал связывание с IL-6R (фигура 12). Из этих результатов было сделано заключение, что изменение в положении 71 FR3 с фенилаланина (который присутствует в человеческих REI, модифицированных на CAMPATH-1H) на тирозин (который присутствует в природных человеческих REI и в мышиных PM-1) очень вредно для образования функционального сайта, связывающего антиген.
Вариант "a" V-области L-цепи видоизмененных человеческих PM-1 был выбран как наилучший вариант. В последующих экспериментах, оценивающих различные варианты V-областей H-цепи видоизмененных человеческих PM-1, всегда использовался вариант "a" V-области L-цепи видоизмененных человеческих PM-1.
Оценку шести вариантов V-областей H-цепи видоизмененных человеческих PM-1 проводили путем одновременного трансфицирования cos клеток одним из шести векторов, экспрессирующих H-цепи видоизмененных человеческих PM-1 (RVh-PM-1a, RVh-PM-1b, RVh-PM-1c, RVh-PM-1d, RVh-PM-1e или RVh-PM-1f) и вектора, экспрессирующего вариант "a" L-цепи видоизмененных человеческих PM-1 (RVI-PM-1a). Клетки также одновременно трансфицировали векторами, экспрессирующими L- и H-цепи химерных PM-1 (HEF-PMK-gk и HEF-PMh-gγI) . Предварительные данные с использованием неочищенных супернатантов cos клеток показали, что вариант "a" L-цепи видоизмененных человеческих PM-1 и вариант "f" H-цепи видоизмененных человеческих PM-1 были эквивалентны L- и H-цепям химерных PM-1 в исследованиях по связыванию с IL-6R.
Чтобы подтвердить эти предварительные данные, химерные и видоизмененные человеческие антитела РМ-1 из супернатантов cos клеток концентрировали и очищали, используя протеин A. А именно, среду с клеток cos концентрировали, используя устройство для разделяющей ультрафильтрации 100 к (Amicon). Концентрированную среду очищали, используя агарозу с протеином A (Affi-Gel Protin A MAPSII Kit, Bio Rad). Короче, концентрированную среду наносили на колонку агарозы с протеином A, которая уравновешивалась пятью объемами слоя связывающего буфера. Колонку промывали 15 объемами слоя связывающего буфера, за чем следовали 5 объемов слоя элюирующего буфера. Элюат концентрировали и буфер заменяли на PBS, используя микроконцентратор (Centricon 10, Amicon). Очищенные антитела использовали для дальнейшего анализа.
Анализ очищенных образцов химерных антител РМ-1 и видоизмененных человеческих антител РМ-1 с вариантом "a" V-области L-цепи и вариантами "a", "b", "c", "d", "e" и "f" V-области видоизмененной человеческой H-цепи был осуществлен. Вариант "a" L-цепи плюс вариант "f" H-цепи является явно наилучшим видоизмененным человеческим антителом РМ-1. Оно связывается с IL-6R так же, как и химерное антитело РМ-1 (фигура 13). Эти антитела также подавляют связывание человеческого IL-6 с IL-6R, так же как и оба мышиные и химерные антитела РМ-1 (фигура 14).
Пример 12.
Реконструирование V-областей видоизмененных человеческих РМ-1 для улучшения уровней экспрессии
Чтобы удалить интроны из последовательностей ДНК, кодирующих лидерные последовательности V-областей L- и H-цепи видоизмененных человеческих РМ-1 (смотрите SEQ ID N0s: 54 и 55), кДНК, кодирующую V-области, повторно клонировали, используя праймеры PCR. Векторы экспрессии L-и H-цепей RVI-PM-1a и RVh-PM-1f трансфицировали одновременно в cos клетки. После 48 часов получали суммарную РНК (Chirgwin et al, Biochemistry (1979) 18: 5294-5299) и 5 мкг суммарной РНК использовалось для синтеза первой нити кДНК, который описан для клонирования PCR V-областей мышиных антител. Были разработаны и синтезированы три PCR праймера. LEV-PI (SEQ ID N0: 60) и HEV-PI (SEQ ID N0: 58) содержат донорную последовательность спласинга и сайт Bam HI и были использованы в качестве ранних праймеров для V-областей L- и H-цепи соответственно. HEV-P2 (SEQ ID N0: 59) содержит согласующую последовательность Козака перед кодоном инициации ATG и сайт Hind III и использовался в качестве позднего праймера для обоих областей V как L-, так и H-цепи. Каждые 100 мкл из реакции PCR содержали 20 мМ Трис-HCl (pH 8,8), 10 мМ KCl, 10 мМ (NH4)2O4, 2 мМ MgSO4, 0,1% Тритон X-100, 0,1 мкг BSA, 250 мкм dNTP. 2,5 ед. V-епт ДНК Полимеразы (Biofabs, U.K), 50% кДНК из реакции синтеза первой нити и 100 пкмолей каждого из раннего и позднего праймера. В каждую пробирку PCR наслаивали 50 мкл минерального масла и затем подвергали цикличному процессу после первоначального растворения при 94oC в течение 1,5 мин, при 94oC в течение 1 мин, при 50oC в течение 1 мин и 72oC в течение 1 мин и затем при 72oC в течение 10 мин. Продукт PCR 408 bp, содержащий V-область L-цепи, и продукт PCR 444 bp, содержащий V-область H-цепи, очищали, используя 2,0% агарозные гели с низкой температурой плавления, расщепляли Bam HI и Hind III и субклонировали в вектор pUC19, чтобы получить pUC-RVI-RCIa-3 и PUC-RVh-PM-1f-3 соответственно.
Было обнаружено, что ДНК последовательности V-областей L- и H-цепи видоизмененных человеческих PM-1 содержат неподходящие донорные сплайсинговые и акцепторные сайты (смотрите SEQ ID N0s: 54 и 55). Сайты V-области в L-цепи используются нечасто (примерно 10% от мРНК), но сайты в V-областях H-цепи используются часто (примерно 90% от мРНК). Этот ненормальный сплайсинг приводил к низким уровням экспрессии видоизмененных человеческих антител PM-1. Чтобы исключить ненормальный сплайсинг в V-областях? донорные сайты спласинга удаляли, используя метод на основе PCR. Для V-области H-цепи синтезировали поздний праймер NEW-SPI (SEQ ID N0: 61) и ранний праймер NEW-SP2 (SEQ ID N0: 62), изменяя ДНК последовательность TGG GTG AGA на ДНК последовательность TGG GTT CGC. Условия для PCR реакций были описаны выше для клонирования кДНК, за исключением того, что матричной ДНК были 50 нг pUC-PVh-PM-1f-3 и праймерами были или HEV-P2 и NEW-SP2, или HEV-P1 и NEW-SP1.
Продукты PCR из двух реакций PCR очищали, используя 2,0% агарозный гель с низкой температурой плавления, и использовали в соединяющей реакции PCR. 98 мкл реакционной смеси PCR с содержанием 0,5 мкг каждого из первичных продуктов PCR и 5 ед Vent ДНК полимеразы инкубировали при 94oC в течение 2 мин, при 50oC в течение 2 мин и 72o в течение 5 минут, а затем 100 пкмолей каждого из HEV-P1 и HEV-P2 праймеров добавляли. В пробирку PCR наслаивали 30 мкл минерального масла и подвергали 25 циклам PCR при 94oC в течение 1 мин, 50oC в течение 1 мин, и 72oC в течение 1 мин и затем инкубировали при 72oC в течение 10 мин.
Подобным же образом донорный сайт сплайсинга в V-области L-цепи видоизмененных человеческих PM-1 удалялся при использовании праймеров PCR PEI-SPI (SEQ ID N0: 63) и PE-SP2 (SEQ ID N0: 64), что меняло ДНК последовательность CAG CTA AGG на ДНК последовательность CAG GAA AGG. Оба продукта PCR, фрагменты ДНК 408 bp для V-области L-цепи и фрагмент ДНК 444 bp для V-области H-цепи очищали с использованием 2,0% агарозного геля с низкой температурой плавления, расщепляли Hind III и Bam H1 и субклонировали в вектор pUC19 для получения pUC-RV1-PM1a-4 и pUC-RVh-PM1f-4 соответственно.
RVh-PM1f-4 создавался путем замещения фрагмента Hind III-Bam HI RVh-PM-1f фрагментом Hind III-Bam H1, вырезанным из pUC-RVh-PM-1f-4. Последовательность варианта "a". V-области L-цепи видоизмененных человеческих антител PM-1, причем интроны были удалены, показаны в SEQ ID N0: 57, и последовательность варианта "f" V-области H-цепи видоизмененных человеческих антител PM-1, где удалены интроны, показана в SEQ IN N0: 56.
Пример 13.
Создание ДНК, кодирующей V-область L-цепи видоизмененных человеческих антител AUK 12-20
Процесс создания ДНК, кодирующей V-область L-цепи видоизмененных человеческих антител AUK 12-20, показан на фиг. 16. Ген, кодирующий V-область L-цепи человеческих антител, включается в вектор pUC19, при использовании ферментов рестрикции Hind III и Bam H1. Готовятся восемь праймеров PCR (от A до H) и амплифицируются в первых 4 областях PCR, которые образуют ген, кодирующий V-область. Праймеры A и H имеют гомологию к ДНК последовательностям вектора pUC19. Праймеры B, C и D являются праймерами длиной 40 и 60 bp каждый, имеющими генную последовательность CDR, которая должна быть перенесена соответственно. Праймеры E, F и G обладают гомологией к последовательности ДНК длиной от 15 до 20 bp 5'-конца праймеров B, C и D соответственно. В четырех первых PCR используют пары праймеров A и E, B и F, C и G, так же как D и H соответственно.
Продукт PCR A-E кодирует FR1 и продукт PCR B-F кодирует CDR1 и FR2. 3'-концевая часть A-E фрагмента и 5'-концевая часть B-F фрагмента обладают гомологией на их отрезке в 15-20 bp, что позволяет объединить здесь фрагменты на более поздней стадии. Аналогично фрагмент B-F обладает гомологией с фрагментом C-G, который кодирует CDR2 и FR3. Фрагмент C-G, кроме того, обладает гомологией с фрагментом D-H, который кодирует CDR3 и FR4. Таким образом, эти четыре фрагмента могут соединяться с помощью их общей гомологии. После соединения этих 4 фрагментов в реакционной смеси PCR добавляют их праймеры A и H во второй реакции PCR, чтобы амплифицировать продукт, образованный при правильном соединении 4 фрагмента. Второй продукт PCR, полученный таким образом, имеет три пересаженные CDR-области и после расщепления Hind III и Bam HI, субклонируется в вектор pUC19.
В частности, в качестве матрицы использовалась плазмида pUC-RV1-PM1a, созданная путем встраивания ДНК, кодирующей вариант "a" V-области L-цепи видоизмененных человеческих антител PM-1, в плазмиду pUC19.
Последовательность праймеров от A до H приведена в табл. 10.
Поздние праймеры 1220-L1, 1220-L2 и 1220-L3 для CDR переноса очищали с помощью 12% полиакриламидного геля, содержащего 8 М мочевины, перед тем, как их использовать.
100 мкл реакционной смеси PCR содержали 20 мМ Трис-HCl (pH 8,8), 10 мМ KCl, 10 мМ (NH4)2SO4, 2 мМ MgSO4, 0,1% Тритон Х-100, 1 мкг БСА, 250 мМ dUTP, 5 единиц Vent ДНК-полимеразы (Biolabs, U.K.), 50 нг pUC-RV1-PM1a-4 ДНК и 100 пкмолей каждого из ранних и поздних праймеров. В каждую пробирку PCR наслаивали 50 мкл минерального масла и после первоначальной денатурации при 94oC в течение 1,5 минут проводили 30 циклов реакции при 94oС в течение 1 минуты, 50oC в течение 1 минуты и 72oC в течение 1 минуты, за чем следовала инкубация при 72oC в течение 10 минут.
Каждый из продуктов PCR, 252 bp (A-E), 96 bp (B-F), 130 bp (C-G) и 123 bp (D-H) очищали с помощью 2,0% агарозного геля с низкой температурой плавления (FMC. Biо Products, USA) А именно, часть агарозного геля, содержащего фрагмент ДНК, вырезалась, расплавлялась при 65oC в течение 5 минут и добавлялась к тому же объему 20 мМ Трис-HCl (pH 7,5), содержащего 2 мМ ЭДТА и 200 мМ NaCl. Смесь экстрагировали фенолом и хлороформом. Фрагмент ДНК выделяли путем осаждения этанолом, растворяли в 10 мМ Трис-HCl (pH 7,5), содержащего 1 мМ ЭДТА, и использовали для реакции соединения PCR.
Затем 98 мкл реакционной смеси PCR, содержащей 0,2 мкг каждого из продуктов первой PCR и 6 единиц Vent ДНК-полимеразы, инкубировали при 94oС в течение 2 минут, 50oC в течение 2 минут и 72oC в течение 5 минут для реакции соединения. Затем 100 пкмолей каждого из праймеров A (обратные) и H (универсальные) добавляли к реакционной смеси, чтобы довести ее до объема 100 мкл, и на реакционную смесь наслаивали 50 мкл минерального масла и подвергали 30 циклам реакции при 94oC в течение 1 минуты, 50oC в течение 1 минуты и 72oC в течение 1 минуты, за чем следовала инкубация при 72oC в течение 10 минут.
Второй продукт PCR длиной 558 bp, содержащий V-область L-цепи, в которую перенесены CDR L-цепи мышиных моноклональных антител AUK 12-20, очищали на 2,0% агарозном геле с низкой температурой плавления и после расщепления с помощью Bam HI и Hind III субклонировали в pUC19 вектор, чтобы получить pUC-RLL-1220a, и секвенировали. Полученная в результате аминокислотная последовательность V-области L-цепи и нуклеотидная последовательность, кодирующая аминокислотную последовательность, показаны в SEQ IN N0: 71.
Затем для создания вектора экспрессии L-цепи фрагмент Hind III-Bam HI ДНК, содержащий V-область L-цепи видоизмененных человеческих антител AUK 12-20 вырезался из вышеупомянутой плазмиды pUC-RLL-1220a и встраивался в сайт Hind III-Bam HI вектора HEF-12k-gk экспрессии L-цепи, чтобы получить вектор экспрессии RVL-1220a для варианта "а" V-области L-цепи видоизмененных человеческих антител AUK 12-20.
Пример 14.
Экспрессия и анализ L-цепи видоизмененных человеческих антител AUK 12-20
Временная экспрессия в cos клетках
Вектор экспрессии RVL-1220a для L-цепи видоизмененных человеческих антител AUK 12-20 и вектор экспрессии HEF-12h-gγI для H цепи химерных 12-20 антител (пример 5) трансфицировали вместе в cos клетки, чтобы оценить варианта "а" L-цепи видоизмененных человеческих антител AUK 1220. А именно, cos клетки суспендировали в фосфатно-буферном физрастворе (PBS) до концентрации 1 • 107 клеток в 1 мл и к 0,8 мл суспензии добавляли плазмидную ДНК (10 мкг каждой плазмиды). К суспензии подводили импульсы с напряжением 1900 V и емкостью 25 мкF, используя аппарат Gene Pulser (Bio Rad).
После восстановления при комнатной температуре в течение 10 мин электропорированные клетки добавляли к 8 мл среды DMEM (GIBCO), содержащей 10% бычьей фетальной сыворотки. После инкубации в течение 72 часов супернатант собирали, центрифугировали, чтобы удалить остатки клеток, и хранили в асептических условиях при 4oC в течение короткого периода или при -20oC в течение более длительного периода.
Определение антител, подобных человеческим, путем ELISA.
Супернатант трансфицированных cos клеток оценивался путем ELISA и подтверждалась продукция химерных антител. Чтобы определить антитела, подобные человеческим, плата покрывалась козьим против человеческого IgG (целые молекулы) (Sigma). После блокирования супернатант от cos клеток последовательно разводился и добавлялся в каждую ячейку.
Плату инкубировали и промывали и затем добавляли козий против человеческого IgG α-цепи специфические (Sigma), коньюгированный с щелочной фосфатазой. После инкубации и промывания добавляли субстратный раствор. После дополнительной инкубации реакцию останавливали и измеряли оптическую плотность при 405 нм. В качестве стандарта использовался очищенный IgG (Sigma).
ELISA для подтверждения способности связываться с человеческими IL-6R.
Супернатант от трансфицированных клеток cos оценивали путем ELISA, чтобы определить могут ли продуцируемые антитела, подобные человеческим, связываться с антигеном, человеческими IL-6R. Плату покрывали мышиными моноклональными антителами МТ18 (пример для справки 1). После блокирования 1% ВСА добавляли растворимые рекомбинантные человеческие IL-6R (SR 344) в плату. После промывания платы последовательно разводили супернатант от клеток cos и добавляли в каждую ячейку платы. После связывания и промывания добавляли козий против человеческого IgG, конъюгированный с щелочной фосфатазой, в ячейки и после дополнительной инкубации и промывания затем добавляли субстратный раствор. После инкубации реакцию прекращали и измеряли оптическую плотность при 405 нм.
Результат представлен на фигуре 17. Антитела, подобные человеческим, включают комбинацию варианта "а" L-цепи видоизмененных человеческих антител AUK 12-20 и H-цепи химерных 12-20 антител. Оптическая плотность при 405 нм изменялась до известной степени в зависимости от разведения, подтверждая, что образец содержит антитела к IL-6R. Кроме того, этого результат показывает, что вариант "а" L-цепи видоизмененных человеческих антител AUK 12-20 имеет способность связывать антиген такую же высокую, как и у L-цепи химерных антител AUK 12-20.
Пример 15.
Создание гена, кодирующего H-цепи видоизмененных человеческих AUK 12-20 антител с использованием согласующей последовательности HSG I.
По тому же методу, который описан в примере 13, CDR V-области H-цепи антител AUK 12-20 переносили в видоизмененные человечески VHa425, содержащие согласующие последовательности HSG I в качестве их FR (Kettleborough et at., Protein Engineering (1991) 4: 773-783). Сначала фрагмент ДНК Hind II-Bam HI, кодирующий видоизмененные человеческие VHa425 (фиг. 3 в литературе) вырезали из плазмиды HCMV-RVHa-425-γI и субклонировали в сайты Hind III-BamHI в pUC19 вектор, чтобы получить pUC-RVH-425a, который затем использовался в качестве матрицы. Было синтезировано 8 праймеров PCR (от A1 по H1). Праймер 1220-H1 был разработан, чтобы перенести CDRI и вызвать мутацию с Т-28 на S-28, и праймер 1220-H3 был разработан, чтобы перенести CDR3 и вызвать мутацию с S-92 на R-94. Праймеры 1220-H1, 1220-H2 и 1220-H3 очищали, используя 12% полиакриламидный гель, содержащий 8 М мочевину, перед их использованием. Нуклеотидная последовательность каждого праймера была следующей (см. табл. 11).
Условия PCR были теми же самыми, что и описанные в примере 13, за исключением того, что pUC-RVH-425a использовался в качестве матричной ДНК, и вышеупомянутые праймеры использовались для переноса CDR H-цепи. Чтобы провести первые реакции PCR использовали пары праймеров A1 и E1, B1 и F1, C1 и G1, а также D1 и H1, и соответствующие первые продукты PCR. 186 bp (A1-E1), 75 bp (B1-F1), 173 bp (C1-G1) и 105 bp (D1-H1) очищали с помощью агарозного геля 2% с низкой температурой плавления и использовали в последующей PCR реакции соединения. В соответствии с условиями, описанными в примере 13, 0,2 мкг каждого из первых продуктов PCR использовали для проведения второй реакции PCR (включая PCR реакцию соединения),чтобы получить продукт PCR 495 bp, содержащий ДНК, кодирующую V-область человеческой H цепи, в которую были пересажены CDR V-области H-цепи мышиных антител AUK 12-20, и продукт PCR очищали, используя 2,5% агарозный гель с низкой температурой плавления. После расщепления продукта PCR с помощью Bam HI и Hind III, полученный в результате фрагмент ДНК Bam HI-Hihd III субклонировали в pUC19 и секвенировали, чтобы получить pUC-RVH-1220a.
Было обнаружено, что ДНК последовательность кодирующая V-область H-цепи видоизмененных человеческих антител AUK 12-20, содержит последовательность, хорошо согласующуюся со сплайсинговой донорной последовательностью, которая может вызывать ненормальный сплайсинг, который представлял препятствие при продукции видоизмененных человеческих антител РМ-1. Поэтому эта последовательность ДНК модифицировалась PCR. Были синтезированы мутагенные праймеры SG1-SP1 (SEQ ID N0 : 97) и SG1-SP2 (SEQ ID N0 : 98). Эти праймеры превращают ДНК последовательность AAG GTG AGC в ДНК последовательность AAA GTC AGC. Условия реакции PCR были теми же, которые описаны выше, за исключением того, что 50 нг pUC-RVH-1220а использовались в качестве матричной ДНК, и SG1-SP1 и универсальный (SEQ ID N0 : 82) или SG1-SP2 и обратный (SEQ ID N0:83) использовались в качестве праймеров.
Продукты PCR из двух реакций PCR очищали с помощью 2% агарозного геля с низкой температурой плавления и использовали в PCR реакции соединения. 98 мкл реакционной смеси PCR, содержащей 0,2 мкг каждого из первых продуктов PCR и 5 единиц Vent ДНК-полимеразы, инкубировали при 94oC в течение 2 минут, при 55oC в течение 2 минут и при 72oC в течение 5 минут для реакции соединения. Затем 100 пкмолей каждого из универсального и обратного праймеров добавляли к реакционной смеси, на которую затем наслаивали 50 мкл минерального масла и подвергали 30 циклам второй реакции PCR, состоящим из инкубации при 94o в течение 1 минуты, при 50oC в течение 1 минуты и при 72oC в течение 1 минуты, после чего следовала инкубация при 72oC в течение 10 минут. Фрагмент ДНК в 495 bp, полученный во второй PCR, очищали с помощью 2% агарозного геля с низкой температурой плавления и субклонировали в вектор pUC19 и секвенировали, чтобы получить pUC-PVH-1220a-2.
Затем фрагмент ДНК Hind III-Bam HI, содержащий ДНК, кодирующую V-область H-цепи видоизмененных человеческих антител AUK 12-20, вырезали из pUC-RVH-1200a-2 и встраивали в сайты Hind III-Bam HI H-цепи вектора экспрессии HEF-12-gγI, чтобы получить вектор экспрессии RVH-1220a для варианта "а" H-цепи видоизмененных человеческих антител AUK 12-20.
Для создания генов, кодирующих варианты с "b" по "d", были синтезированы две пары мутагенных праймеров. Каждая реакция PCR проводилась по существу при тех же условиях, что описаны выше. Для создания варианта "b" использовали в двух первых реакциях PCR или универсальный праймер (SEQ ID N0 : 82) и мутагенный праймер 120H-mlb (SEQ ID N0:78), или обратный праймер (SEQ ID N0: 83) и мутагенный праймер 1220 H-mlb (SEQ ID N0:79), а также pUC-RVH-1220a в качестве матрицы. Первые продукты PCR в 202 bp и 232 bp очищали с помощью 2,0% агарозного геля с низкой температурой плавления и использовали во второй PCR (включая PCR реакцию присоединения) при тех же самых условиях, что описаны выше, чтобы получить продукт в 495 bp (вариант "b"). Продукт расщепляли с помощью. Hind III и Bam HI и субклонировали в вектор pUC19, чтобы получить pUC-RVH-1220b.
Аналогично использовали мутагенный праймер 1220H-m2 (SEQ ID N0:80). 1220H-m2b (SED ID NO: 81) и матрицу pUC-RVH-1220a в PCR, чтобы получить продукт PCR (вариант "с"). Продукт расщепляли с помощью Hind III-Bam HI и встраивали в сайты Hind III-Bam HI вектора рUC19, чтобы получить pUC-RVH-1220c. Кроме того, мутагенные праймеры 1220H-mla (SEQ ID N0:78). 1220H-mlb (SEQ ID N0:79) и матрица pUC-RVH-1220c использовались чтобы получить продукт PCR (вариант "d"), который затем расщепляли с помощью Hind III и Bam HI и встраивали в сайты Hind III-Bam HI вектора pUC19, чтобы получить pUC-RVH-1220d.
Обратите внимание, что аминокислотная последовательность варианта "b" V-области H-цепи видоизмененных человеческих антител AUK 12-20 и нуклеотидная последовательность, ее кодирующая в плазмиде pUC-RVH-1220b, показана в SEQ ID N0: 84, и аминокислотная последовательность варианта "d" V-области H-цепи видоизмененных человеческих антител AUK 12-20 и нуклеотидная последовательность, ее кодирующая в плазмиде pUC-RVH-1220, показана в SEQ ID N0:85.
Затем, чтобы создать векторы экспрессии, фрагменты Hind III-Bam HI, содержащие V-область H-цепи видоизмененных человеческих антител AUK 12-20, вырезали из pUC-RVH-1220b, pUC-RVH-1220c и pUC-RVH-1220d и встраивали в сайты Hind III-Bam HI вектора экспрессии HEF-12h-gγI H-цепи, чтобы получить RVH-1220b, RHH-1220c и RVH-1220d соответственно.
Пример 16.
Экспрессия и анализ различных вариантов видоизмененных человеческих антител AUK 12-20.
COS клетки одновременно трансфицировали одним из 4 векторов экспрессии для H-цепи видоизмененных человеческих антител AUK 12-20 (RVH-1220a, RVH-1220b, RVH-1220c или RVH-1220d) и вектором экспрессии VRL-1220a, чтобы оценить 4 варианта области H-цепи видоизмененных человеческих антител AUK 12-20. Для примера, cos клетки одновременно трансфицировали векторами экспрессии L-цепи и H-цепи химерных антител 12-20 (HEF-12h-gγI и FEF-12-gк). В исследовании на связывание с человеческими IL-6R видоизмененные человеческие антитела AUK 12-20, состоящие из L-цепи видоизмененных человеческих антител AUK 12-20 и варианта "b" H-цепи видоизмененных человеческих антител AUK 12-20, и видоизмененные человеческие антитела AUK 12-20, состоящие из L-цепи видоизмененных человеческих антител AUK 12-20, и варианта "d" H-цепи видоизмененных человеческих антител AUK 12-20, демонстрируют хорошее связывание, такое же как и химерные антитела 12-20. Эти результаты представлены на фиг. 18 и 19.
Пример 17.
Создание гена, кодирующего H-цепь видоизмененных человеческих антител sle 1220 при использовании человеческих антител HAX.
Человеческими антителами, имеющими наивысшую гомологию с V-областью H-цепи моноклональных мышиных антител AUK 12-20, являются HAX (J. Immunology (1987) 139: 2496 - 2501; антитела, продуцируемые гибридомой 21/28, полученной из B клеток больного СКВ; их аминокислотная последовательность показана на фиг. 6 и нуклеотидная последовательность для них показана на фиг. 4 и 5 этого литературного источника), по данным базы данных "Leeds" по белкам. V-область H-цепи видоизмененных человеческих sle 1220H-антител конструировали, используя FR антител HAX и CDR V-области H-цепи мышиных моноклональных антител AUK 12-20.
Всю ДНК, кодирующую вариант "a" V-области H-цепи видоизмененных человеческих антител sle 1220H синтезированы химическими методами. ДНК, кодирующая V-область H-цепи антител sle 1220H с полной длиной в 439 bp, представлена путем разделения ДНК на 6 олигонуклеотидов длиной от 90 до 94 bp с перекрыванием каждого другим на 21 bp (sle 1220Н от 1 до 6; SEQ ID N0: с 85 по 91 соответственно). При разработке олигонуклеотидов вторую структуру испытывали и для сайтов, представляющих структурные помехи, третий нуклеотид в кодоне заменяли без изменения аминокислоты, им кодируемой. Связь этих олигонуклеотидов и процесс создания двухнитевых ДНК показаны на фиг. 20.
Реакция, показанная на фиг. 20, проводится с использованием методики PCR. А именно, 6 синтетических олигонуклеотидов добавляли в одну пробирку для реакции PCR, чтобы провести первую реакцию PCR, тем самым два олигонуклеотида отжигаются и удлиняются и получаются дальнейшие 4 олигонуклеотида или полный олигонуклеотид.
Затем добавляют завершающие праймеры A (SEQ ID N0: 92) и B (SEQ ID N0: 93), чтобы провести вторую реакцию PCR, причем могут быть амплифицированы только правильные нуклеотиды, имеющие полную длину. Полученный в результате продукт расщепляется с помощью Bam HI и Hind III и субклонируется в вектор pUC19 с последующим секвенированием.
Более конкретно, 98 мкл реакционной смеси, содержащей 100 мМ Трис-HCl (pH 8,5), 50 мМ KCl, 0,1 мМ dATP, 0,1 мМ dGTP, 0,1 мМ dCTP, 0,1 мМ dTTP, 1,5 мМ MgCl2 и 2,5 ед. ДНК-полимеразы Amplitag (Perkin Elmer Cetus), а также 5 пкмолей каждого из олигонуклеотидов, денатурировали при 94oC в течение 1,5 минут и подвергали 3 циклам реакции путем инкубации при 92oC в течение 3 минут, 50oC в течение 2 минут и 72oC в течение 5 минут с последующей инкубацией при 72oC в течение 10 минут. Один мкл каждого из 50 мМ завершающих праймеров A и B добавляли к реакционной смеси, которую затем покрывали 80 мкл минерального масла и после денатурации при 94oC в течение 1,5 минут подвергали 30 циклам реакции путем инкубации при 94oC в течение 1 минуты, 50oC в течение 1 минуты и при 72oC в течение 1 минуты с последующей инкубацией при 72oC в течение 10 минут. Продукт PCR из 439 bp очищали с помощью 1,5% агарозного геля с низкой температурой плавления, расщепляли рестрикционными ферментами Bam HI Hind III и субклонировали в вектор pUC19 с последующим подтверждением последовательности. Клон, полученный таким образом, обозначали pUC-RUH-sle 1220Ha. Аминокислотная последовательность варианта "a" V-области H-цепи видоизмененных человеческих антител sle 1220H и нуклеотид ее кодирующий в плазмиде pUC-RVH-sle 1220Ha показаны в SEQ ID N0: 94.
Затем фрагмент ДНК Hind III-BamHI, содержащий ген, кодирующий V-область H-цепи видоизмененных человеческих антител 12-20 (sle 1220H) вырезали из pUC-RVH-sle 1220Ha и встраивали в сайты Hind III-Bam HI вектора экспрессии H-цепи HEF-12h-gγI, чтобы получить RVH-sle 1220Ha.
Для создания вариантов "b" и "d" V-области H-цепи видоизмененных человеческих антител sle 1220H были синтезированы два мутагенных праймера sle 1220Hm1 (SEQ ID N0: 95) и sle 1220Hm2 (SEQ ID N0: 96). В каждой PCR использовали Vent ДНК-полимеразу и реакционную смесь с составом, описанным в примере 13. В каждой реакции PCR реакционную смесь, содержащую pUC-RVH-sle 1220Ha в качестве матрицы, 50 пкмолей мутагенного праймера sle 1220Hm1 или sle 1220Hm2 и 50 пкмолей завершающего примера B, денатурировали при 94oC в течение 1,5 минут и подвергали 30 циклам реакции путем инкубации при 94oC в течение 1 минуты, при 50oC в течение 1 минуты и при 72oC в течение 1 минуты с последующей инкубацией при 72oC в течение 10 минут. Продукт из 235 bp или 178 bp очищали с помощью 1,5% агарозного геля с низкой температурой плавления, чтобы использовать в качестве праймера во второй реакции PCR. А именно, вторую реакцию PCR проводили, используя 50 пкмолей завершающего праймера A. 0,2 мкг продукта PCR и pUC-RVH-sle 1220Ha в качестве матрицы и полученный в результате продукт из 439 bp очищали при помощи 1,5% агарозного геля с низкой температурой плавления, расщепляли с помощью Bam HI и Hind III и субклонировали в вектор pUC19, чтобы получить pUC-RVH-sle 1220H или pUC-RVH-sle 1220Hc, который кодирует варианты "b" или "c" V-области H-цепи видоизмененных человеческих антител sle 1220 соответственно.
ДНК, кодирующая вариант "d" V-области H-цепи видоизмененных человеческих антител sle 1220H, создавалась следующим образом. В качестве матрицы использовали pUC-RVH-sle 1220Hb, 50 пкмолей каждого мутагенного праймера sle 1200Hm2 и завершающего праймера B использовалось для проведения 30 циклов первой реакции PCR. Полученный в результате продукт PCR из 176 bp очищали в 1,6% агарозном геле с низкой температурой плавления, чтобы использовать в качестве праймера во второй реакции PCR. Этот праймер и 50 пкмолей завершающего праймера A использовали во второй PCR, чтобы получить фрагмент ДНК из 439 bp. Полученный таким образом продукт PCR очищали, расщепляли BamHI и Hind III и субклонировали в вектор pUC19 для получения pUC-RVH-sle 1220Hd.
Затем, чтобы создать векторы экспрессии или различных вариантов V-области H-цепи видоизмененных человеческих антител sle 1220H, фрагменты Bam HI-Hind III, содержащие ДНК, кодирующую V-область H-цепи видоизмененных человеческих антител sle 1220, вырезались из pUC-RVH-sle 1220Hb, pUC-RVH-sle 1220Hс и pUC-RVH-sle 1220Hd и встраивались в сайты Hind III-Bam HI вектора экспрессии H-цепи HEF-12h-gγI для получения вектора экспрессии RVH-sle 1220Hb, RVH-sle 1220Hc и RVH-sle 1220Hd соответственно.
Каждый из четырех векторов, экспрессирующих H-цепь видоизмененных человеческих антител sle 1220H (RVH-sle 1220Ha, RVH-sle 1220Hb и RVH-sle 1220Hc или RVH-sle 1220Hd) и вектор RVL-1120a, экспрессирующий цепь видоизмененных человеческих антител AUK 12-20, трансфицировали вместе с cos клетки, чтобы оценить четыре варианта V-области H-цепи видоизмененных человеческих антител sle 1220H на способность подавлять связывание IL-6 с IL-6R. Результаты представлены на фиг. 21-24. Обратите внимание, что эти результаты получены после очистки продуцируемых антител протеином A.
Как видно из вышеприведенного, в соответствии с настоящим изобретением в химерной L-цепи или в видоизмененной человеческой L-цепи, или в химерной H-цепи или в видоизмененной человеческой H-цепи, и особенно в RF, одна из более чем одна аминокислота может быть заменена другой аминокислотой при сохранении способности связываться с человеческими IL-6R. Поэтому настоящее изобретение включает химерные антитела и видоизмененные человеческие антитела, химерную L-цепь и видоизмененную человеческую L-цепь, химерную H-цепь и видоизмененную человеческую L-цепь, видоизмененную V-область H-цепи и видоизмененную V-область H-цепи, где одна или более чем одна аминокислота заменена другой, а также ДНК, их кодирующую, поскольку они сохраняют свои природные свойства.
Исходные гибридомы, использованные в настоящем изобретении, были связаны следующим образом.
Справочник пример 1. Создание гибридомы MT18.
Чтобы создать гибридому, продуцирующую моноклональные антитела к человеческим IL-6R, в качестве иммуногена была создана линия мышиных T-клеток, экспрессирующая человеческие IL-6R на клеточной поверхности следующим образом. А именно, плазмиду pBSF2R.236, описанную в H патентной заявке N HI-9774, и pSV2neo трансфицировали в мышиные T-клетки линии CTLL-2 (AТСС TIB214) по общепринятому методу, и полученный в результате трансформант отсортировывали, используя G418, в соответствии с общепринятым методом, чтобы получить линию клеток, экспрессирующих примерно 30000 IL-6R на клетку. Эту линию клеток обозначили CTBC3.
Клетки CTBC3 культивировали в RPM1 1640 в соответствии с общепринятой методикой, культивированные клетки промывали четыре раза PBS буфером, и 1 • 107 клеток вводили внутрибрюшинно мышам C57BL/6 для иммунизации. Иммунизация проводилась один раз в неделю в течение 6 недель.
От иммунизированных мышей получали клетки селезенки и проводили слияние с миеломными клетками P3U1, используя полиэтиленгликоль, в соответствии с общепринятым методом, и слившиеся клетки отбирали следующим образом. IL-6R-отрицательную линию человеческих T клеток JURKAT (ATCC CRL 8163) одновременно трансфицировали плазмидами pBSF2R. 236 и pSV2neo, и трансформированные клетки отсортировывали, чтобы получить линию клеток, экспрессирующих примерно 100000 IL-6R на клетку. Клеточную линию обозначили NJBC8. Клон гибридомных клеток, продуцирующих антитела, которые распознают NJBC8, лизированные NP40, но не распознают JURKAT, лизированные NP40, был клонирован и обозначен MT18. Гибридома МТ18 была помещена на хранение в Fermentation Research Institute Agency of Industrial Science and Technology (FRI) по Будапештскому договору как FERM BP-2999 10 июля 1990 г.
Справочный пример 2. Создание гибридомы PM-1.
Чтобы создать гибридому, продуцирующую моноклональные антитела к IL-6R, были экстрагированы в качестве антигена человеческие IL-6R следующим образом 3 • 109 человеческих миеломных клеток U266 (клеток, продуцирующих IL-6R) лизировали в 1 мл 1% дигитонина, 10 мМ триэтаноламинового буфера (pH 7,4), 0,15 М NaCl и 1 мМ PMSF (фенилметилсульфонилфторида: Wako Pure Chemicals). С другой стороны, антитела MT18, продуцируемые гибридомой MT18, полученной в справочном примере 1, связывали с активированной цианогенбромидом сефарозой 4B (Pharmacia) по общепринятому методу. Эту конъюгированную с MT18-антителами сефарозу 4B смешивали с полученным выше лизатом клеток, чтобы связать солюбилизированные IL-6R с антителами MT18 на сефарозе 4B. Вещества, неспецифически связанные с сефарозой 4B отмывали, и IL-6R, связанные с сефарозой 4B через антитела MT18, использовали в качестве иммуногена.
Мышей BALB/c внутрибрюшинно иммунизировали полученным выше иммуногеном один раз в неделю в течение 4 недель. Затем, от иммунизированных мышей получали клетки селезенки и проводили их слияние с миеломными клетками P3UI, используя полиэтиленгликоль в соответствии с общепринятым методом. Слившиеся клетки отсортировывали следующим образом. Сначала супернатант культуры и 0,01 мл протеин G-сефарозы (Pharmacia) смешивали, чтобы адсорбировать иммуноглобулин супернатанта на протеин G-сефарозе. С другой стороны, лизировали 107 клеток 266, внутренне меченных 35S-метионином, и IL-6R очищали за счет аффинитета, используя сефарозу, конъюгированную с МТ18. Затем, IL-6R, меченные 35S-метионином, подвергали иммунопреципитации с протеин G-сефарозой, полученной выше, с которой был связан иммуноглобулин, и преципитат анализировали с помощью SDS/PAGE (электрофореза в полиакриламидном геле с додецилсульфатом натрия). В результате был выделен один гибридомный клон, продуцирующий антитела, которые специфически связываются с IL-6R, и обозначен PM-1. Гибридома PM-1 была помещена на хранение в FRI по Будапештскому договору как FERM BP-2998 10 июля 1990.
Справочный пример 3. Создание гибридом AUK 12-20, AUK 64-7 и AUK 146-15.
Как иммуноген, растворимые IL-6R (SR 344), был получен по методу, описанному Yasukawa, K., et al., J. Biochem 108, 673 - 676, 1990. А именно, плазмиду pECE dhfr 344, содержащую кДНК, кодирующую IL-6R, в которой 345-й кодон с N-конца был замещен стоп-кодоном, трансфицировали в клетки CHO (5E27), трансфицированные клетки культивировали в среде без сыворотки (SF-O среда, Sunko Juhyaku) и полученный в результате супернатант концентрировали с помощью системы HF-Labl (Toson) и очищали на колонке Blue-5PW и Pnenyl-5PW колонке. Очищенные растворимые IL-6R показали единственную полосу на SDS-PAGE.
Мышам-самкам BALB/cAnNCrj (Nippon CREA) подкожно вводили 10 мкг/мышь иммуногена в полном адъюванте Фрейнда (Bacto Adjuvant Complete H 37 Pa, Difco), с последующими вторым и третьим выделениями того же самого количества иммуногена в неполном адъюванте Фрейнда (Bacтo) через две или три недели после первого введения соответственно. Последняя иммунизация (четвертное введение) проводилась без адъюванта в хвостовую вену через неделю после третьей инъекции. От иммунизированных мышей получали образец сыворотки, разводили серийно буфером для разведения и исследовали с помощью ELISA по методу, описанному Goldsmith, P. K., Analytical Biochemistry 117, 53 - 60, 1981. А именно, покрытую SR344 (0,1 мкг/мл) плату блокировали 1% BSA и добавляли затем разведенный образец. Мышиный IgG, связанный с SR344 определяли, используя козий против мышиного IgG/щелочную фосфатазу (A/P) (ZYMED) и субстрат для щелочной фосфатазы (Sigma-104).
После подтверждения повышения анти-SR344 антител в сыворотке через три дня после последней иммунизации от 5 BALB/c мышей получали клетки селезенки. Клетки селезенки и миеломные клетки (P3UI) смешивали в соотношении 25 : 1, проводили слияние, используя PEG1500 и культивировали в 2000 ячейку при клеточной концентрации 0,7 до 1,1 • 106 клеток/ячейку. Супернатанты из ячеек отсортировывали на их способность связываться с SR344 (первый отбор характеризовался как определение распознавания R344) и на их способность подавлять связывание SR344 с интерлейкином-6 с помощью пробы подавления связывания IL-6/IL-6R (RBIA). Первый отбор дал 240 положительных ячеек, и второй отбор дал 36 положительных ячеек.
Вышеупомянутое определение распознавания R344 проводилось следующим образом. Козьим против мышиного Ig (Cappel) (1 мкг/мл) покрытые платы (Maxisorp, Nunc) блокировали 1% БСА и затем добавляли 100 мкл/ячейку супернатанта культуры гибридомы с последующей инкубацией при комнатной температуре в течение одного часа. После промывания платы в каждую ячейку добавляли 20 мкг/мл SR344 и проводили инкубацию при комнатной температуре в течение одного часа. Количество SR344, связанного иммобилизированными антителами, полученными из супернатанта, определяли путем добавления кроличьего анти-SR344 IgG (# 2,5 мкг/мл), козьего противокроличьего IgG-щелочной фосфатазы (A/P) (1 : 3000, Tago) и субстрата (1 мг/мл, Sigma-104) с последующим определением оптической плотности при 405 - 600 нм.
Вышеупомянутое RBIA проводилось следующим образом. Плату покрытую антитела MT18 заполняли 100 мкг/мл SR344 (100 мкл/ячейку), и инкубацию проводили при комнатной температуре в течение одного часа. После промывания платы одновременно в каждую ячейку добавляли 50 мкл/ячейку супернатанта гибридомы и 50 мкг/ячейку конъюгата биотина-интерлейкина-6, и ячейки инкубировали при комнатной температуре в течение одного часа. Количество биотин-IL-6, связанного c SR344, определяли путем добавления стрептавидина-A/P (1 : 7000, PIERCE) и соответствующего субстрата (Sigma-104) с последующим измерением оптической плотности при 405 - 600 нм.
В конце положительные клоны очищали дважды повторенным методом серийных разведений, и были получены три клона гибридомы, т.е. AUK 12-20, AUK 145-15 и AUK 64-7, которые подавляли связывание SR344 с IL-6, и клон гибридомы AUK 181-6, который не подавляет связывание SR344 с IL-6.
Возможность промышленного использования.
Настоящее изобретение представляет видоизмененные человеческие антитела к человеческим IL-6R, включая человеческие антитела, в которых CDR человеческих V-областей заменены CDR мышиных моноклональных антител к человеческим IL-6R. Так как большая часть видоизмененного человеческого антитела получается из человеческого антитела и мышиных CDR, которые менее антигенны, данные видоизмененные человеческие антитела менее иммуногенны для человека и поэтому перспективны для терапевтического применения.
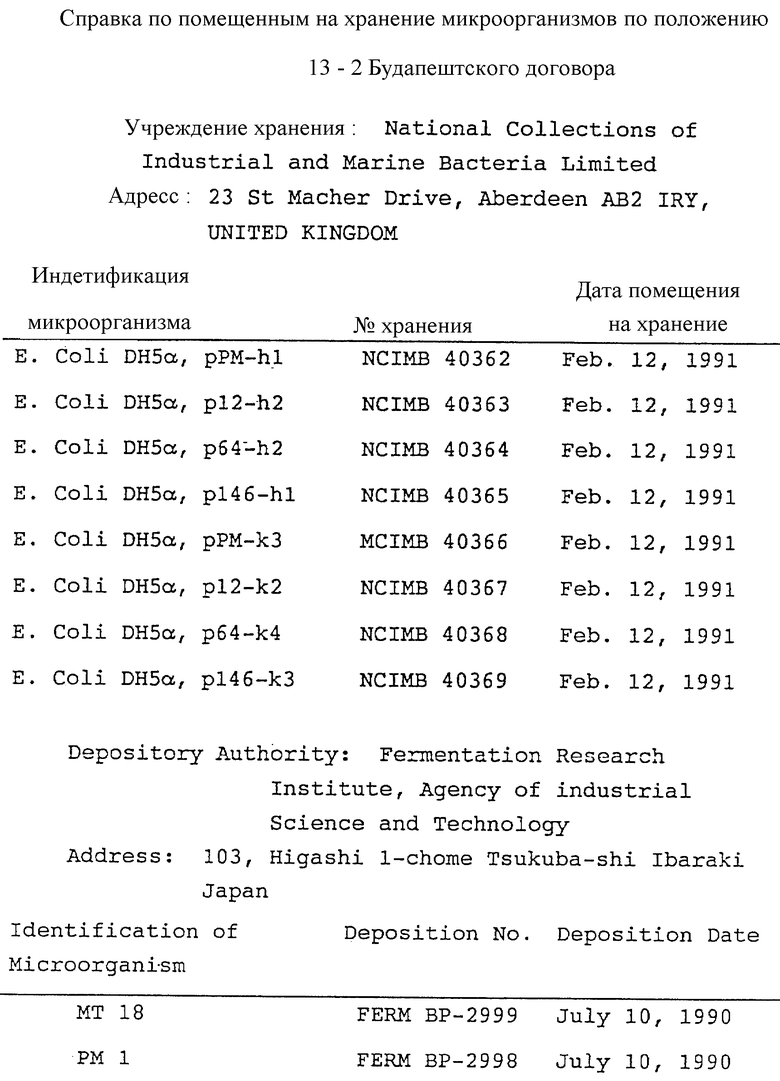
































































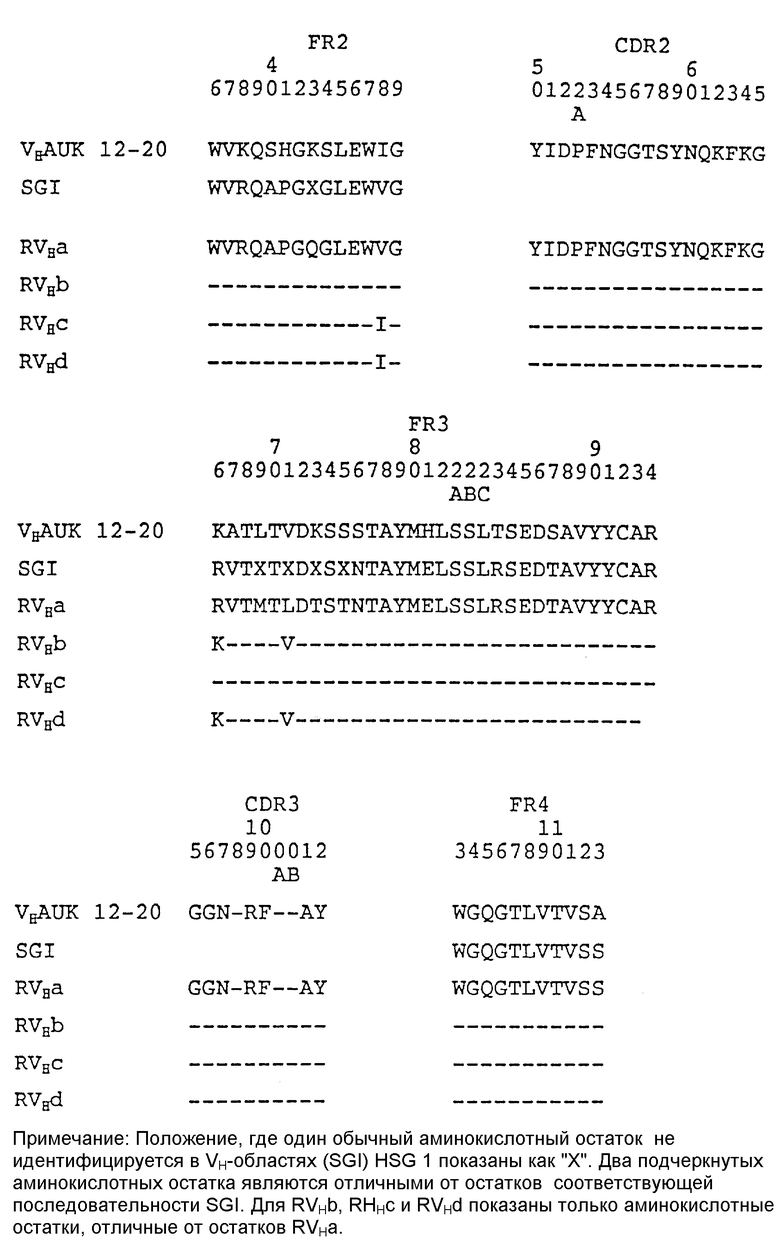



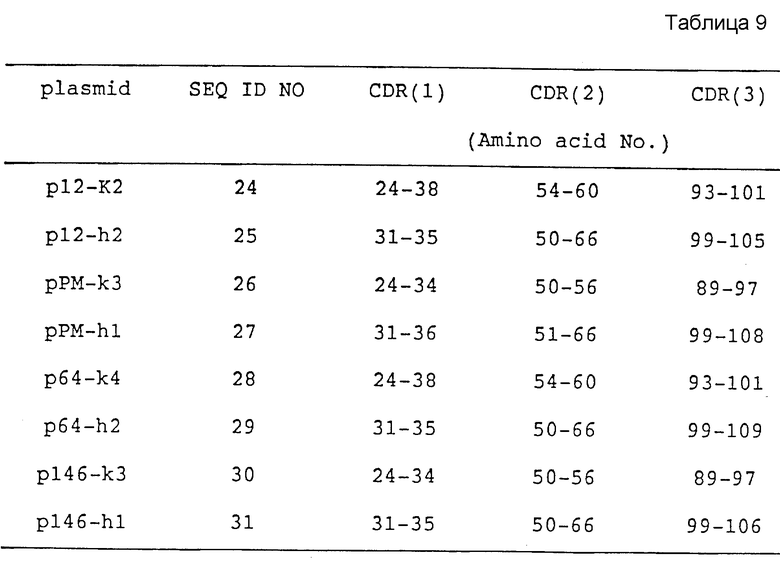
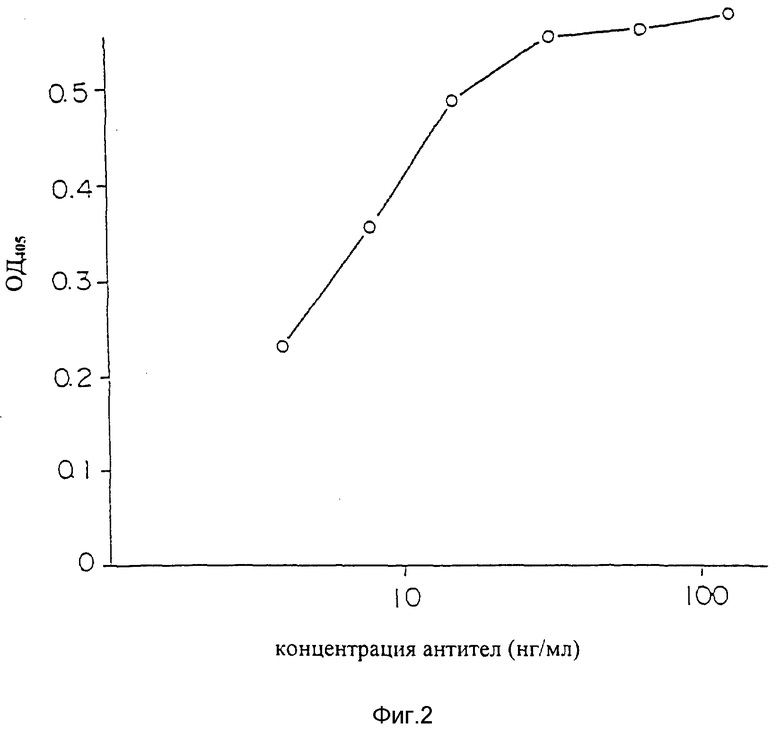
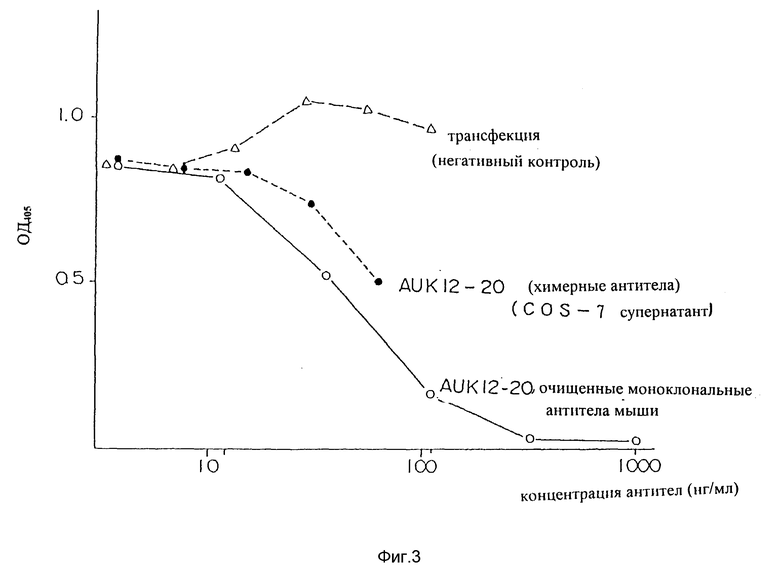
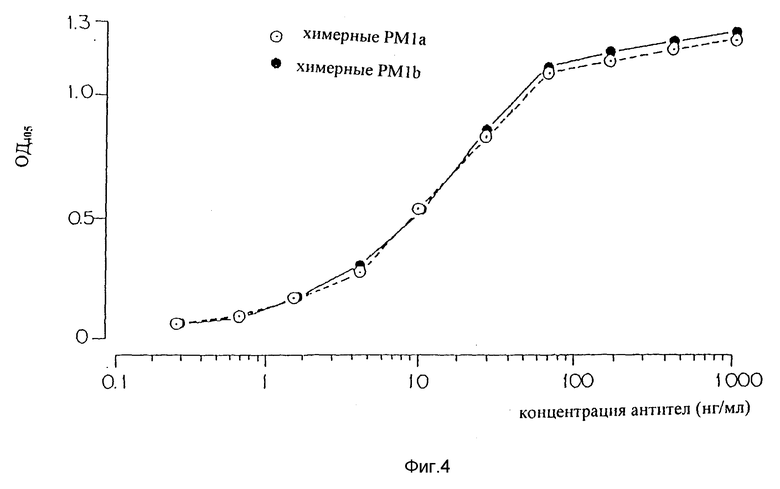
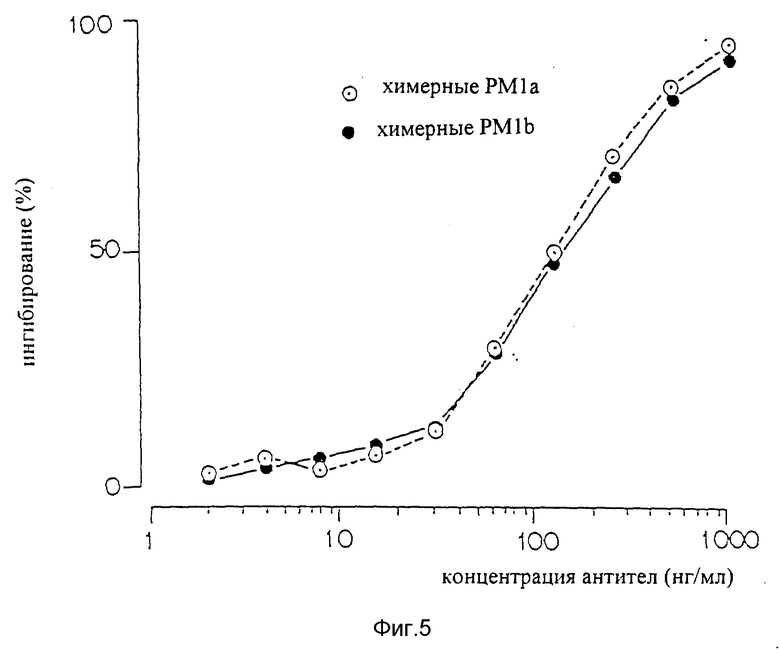
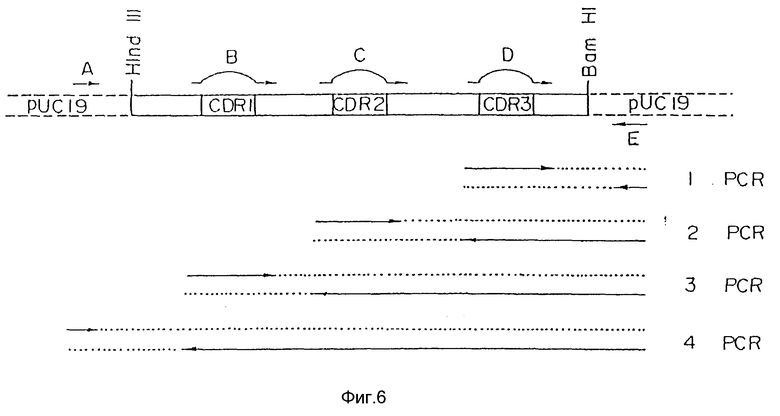
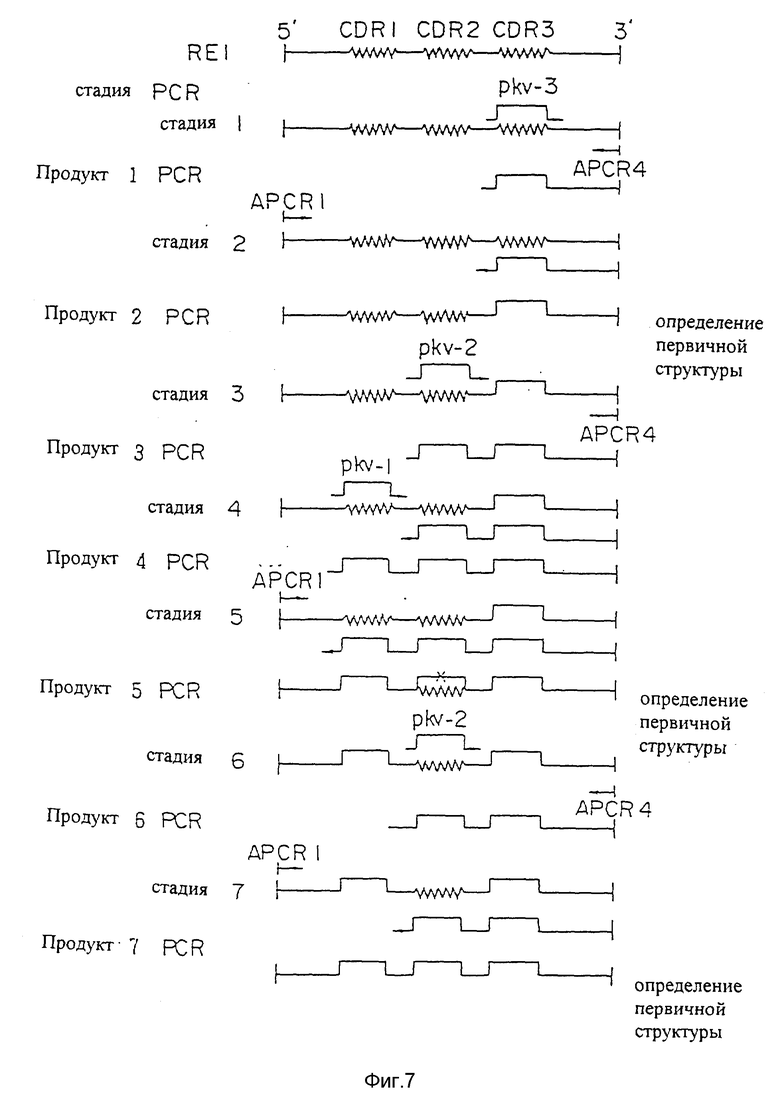
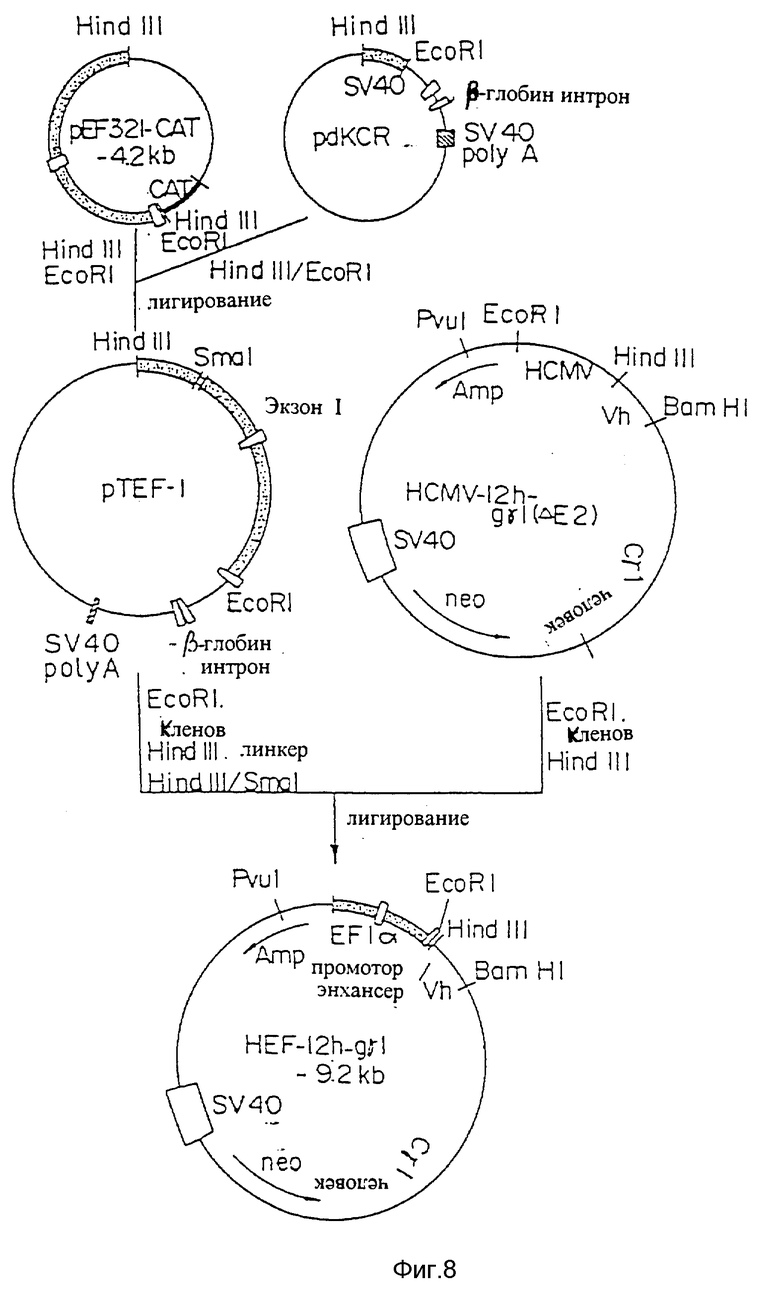
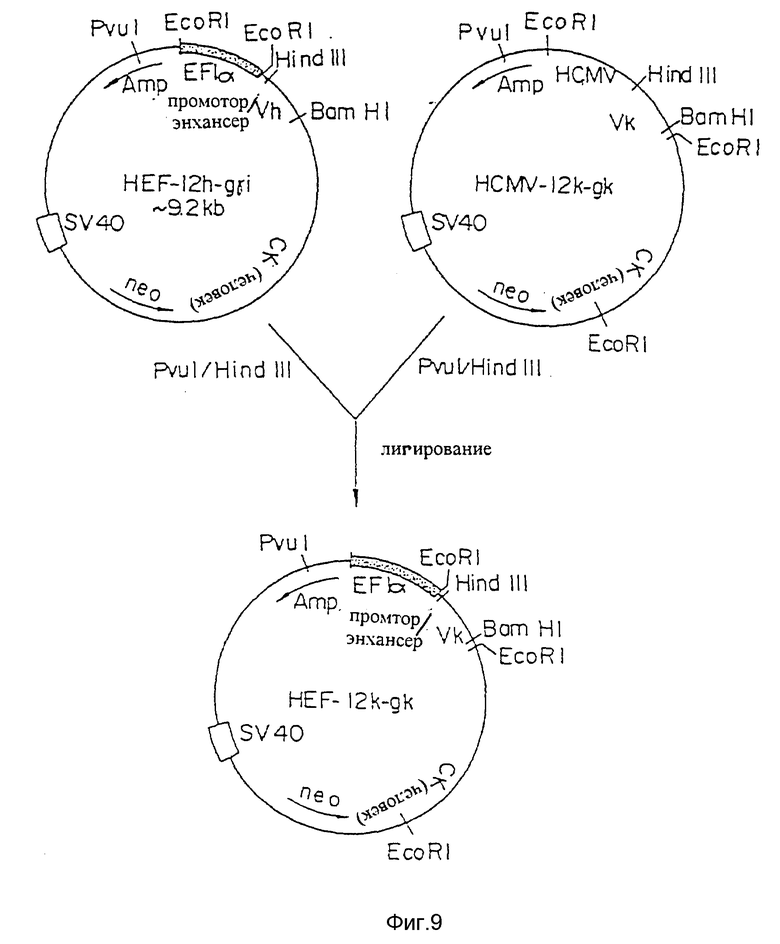
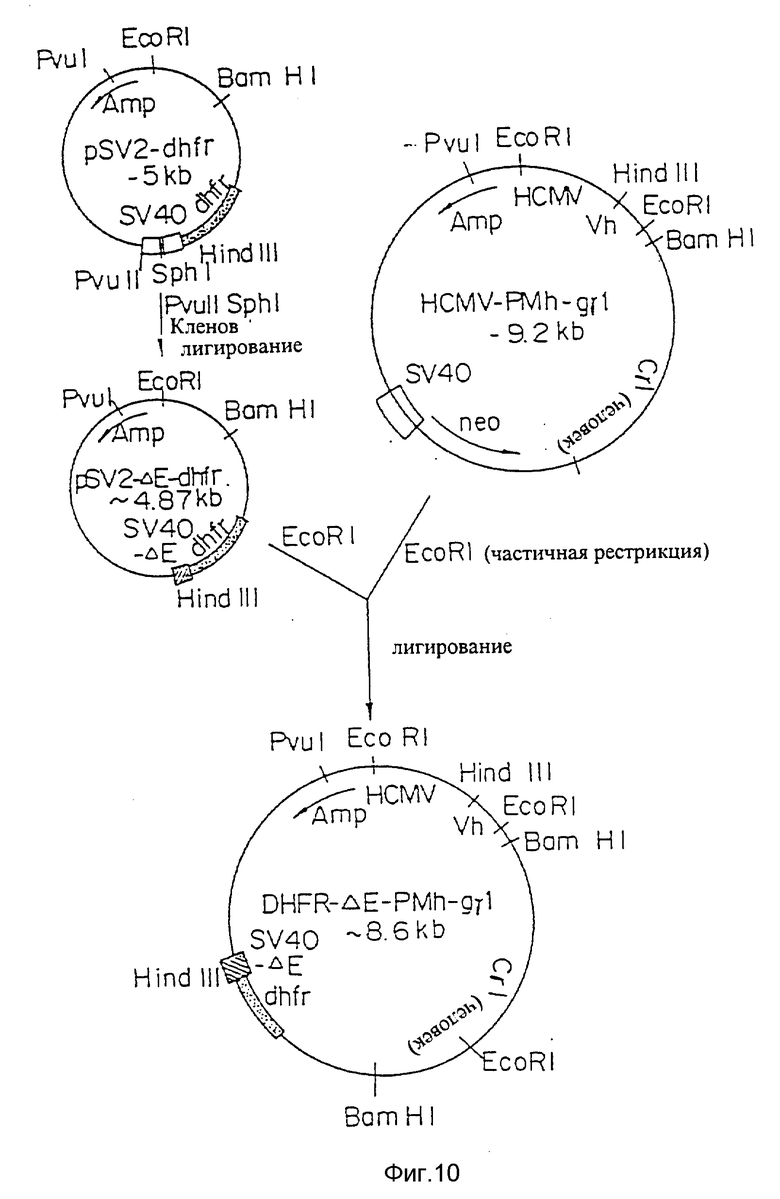
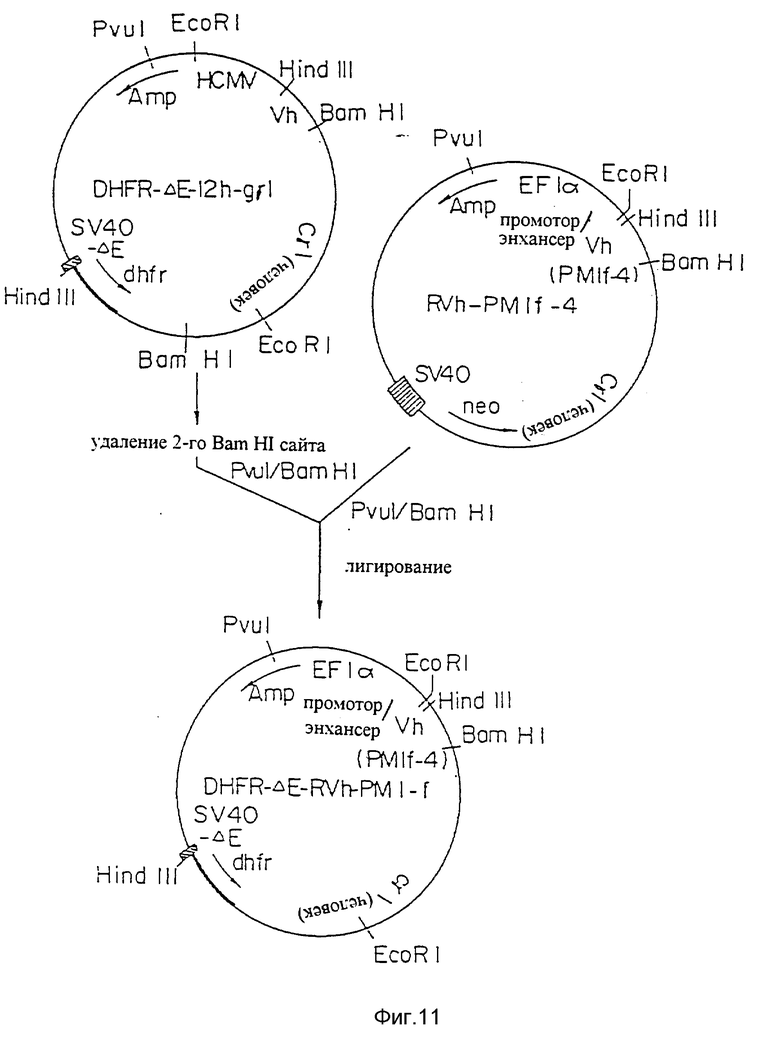
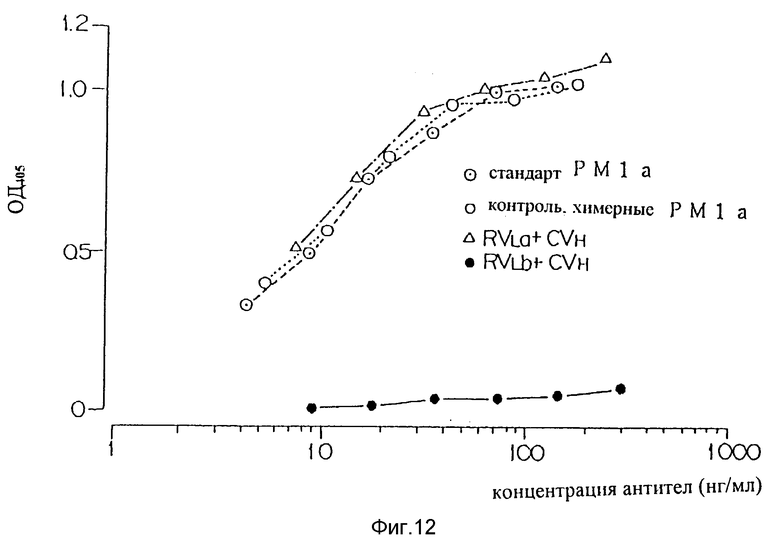
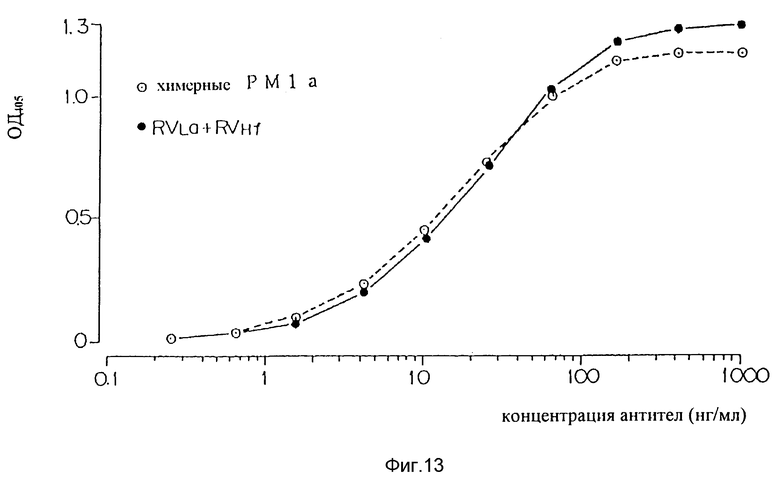
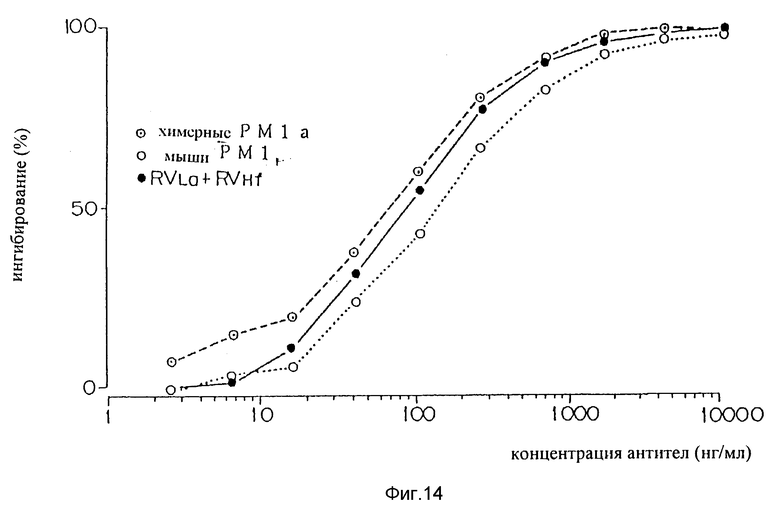
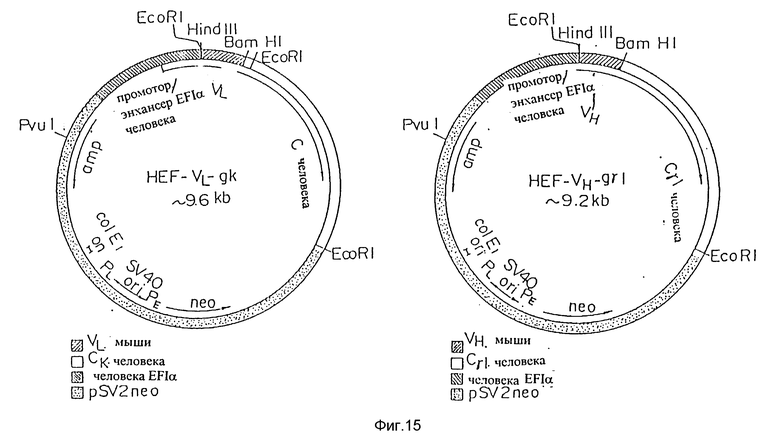
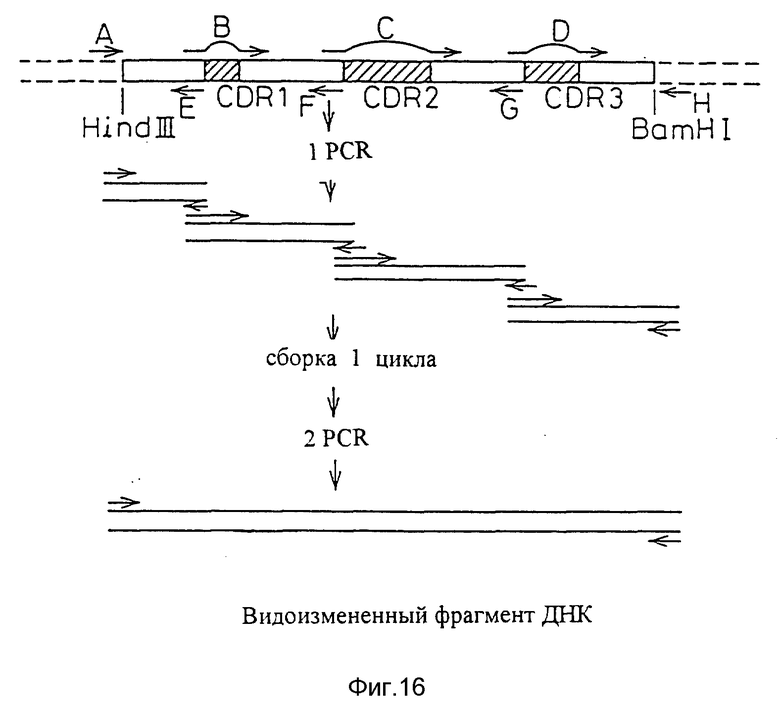
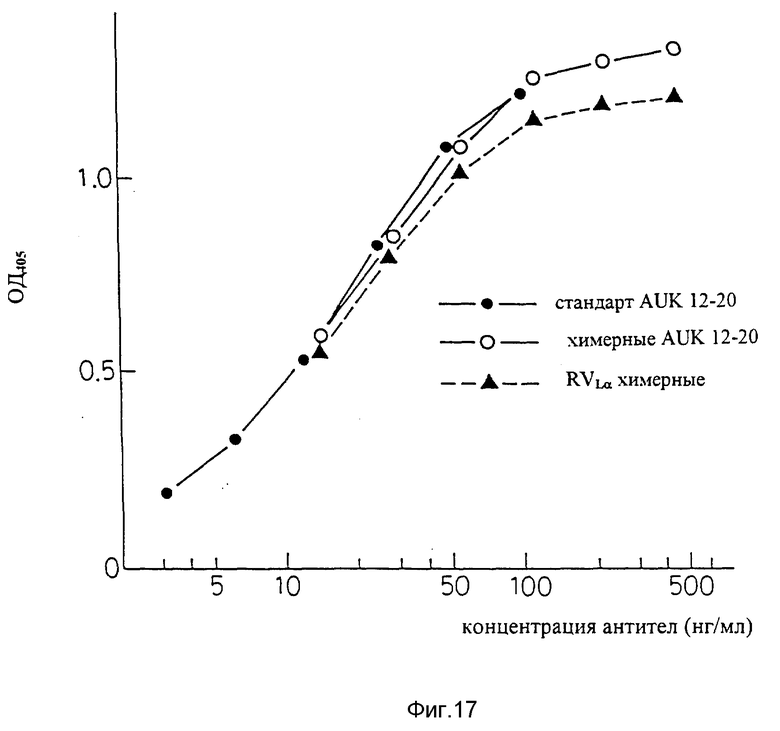
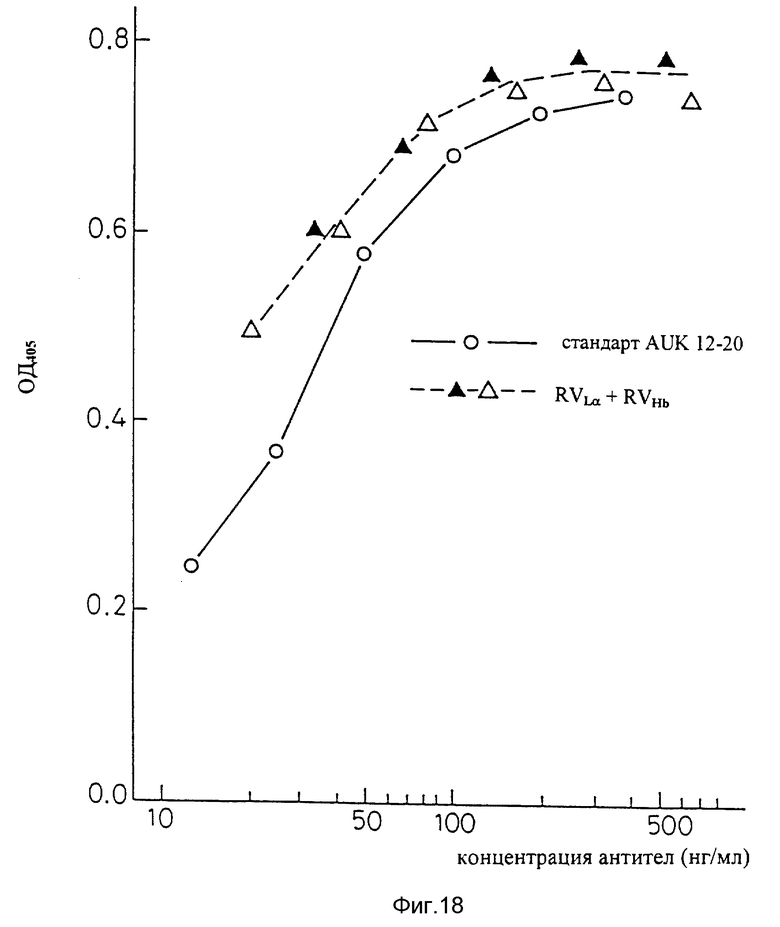
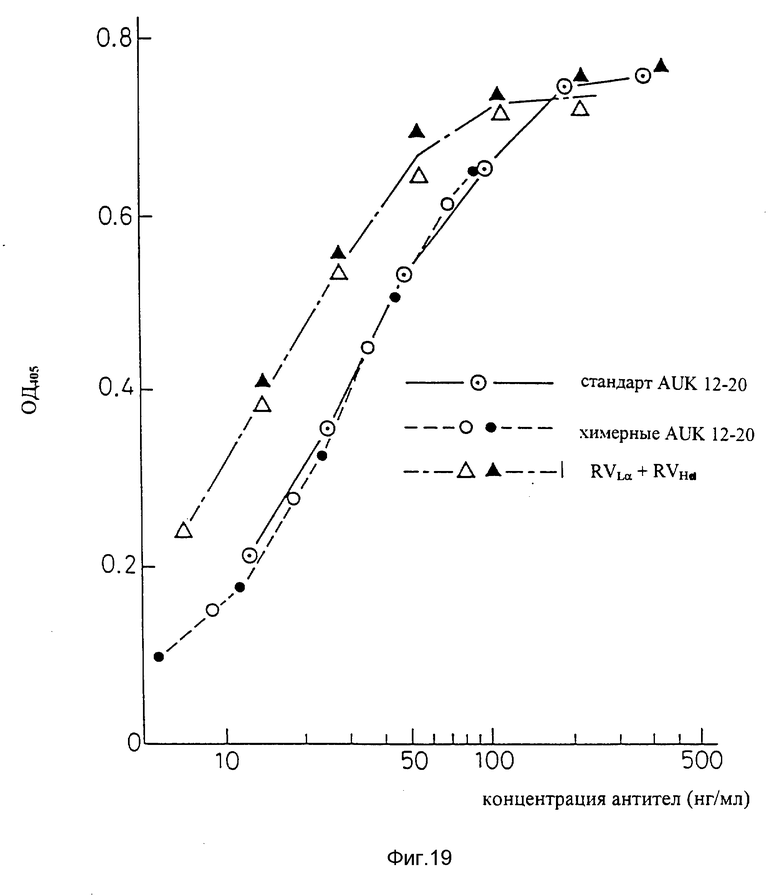
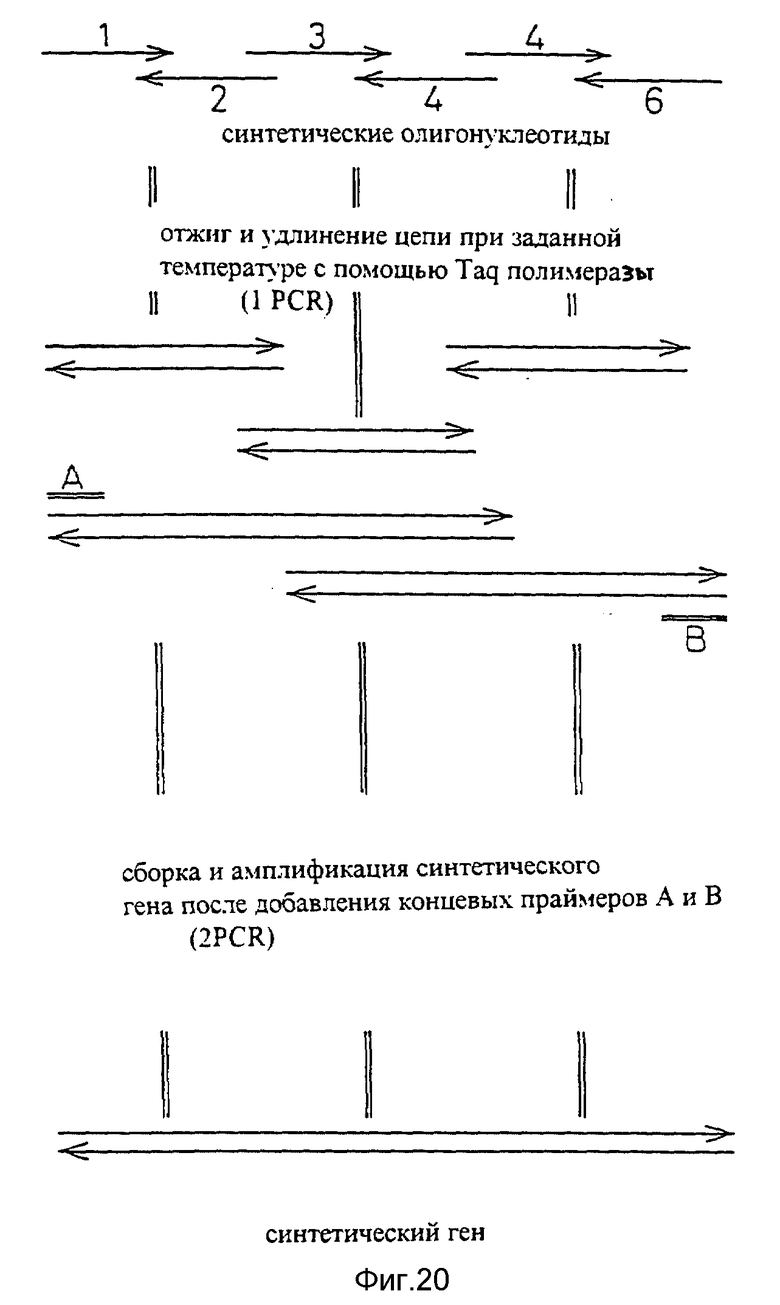
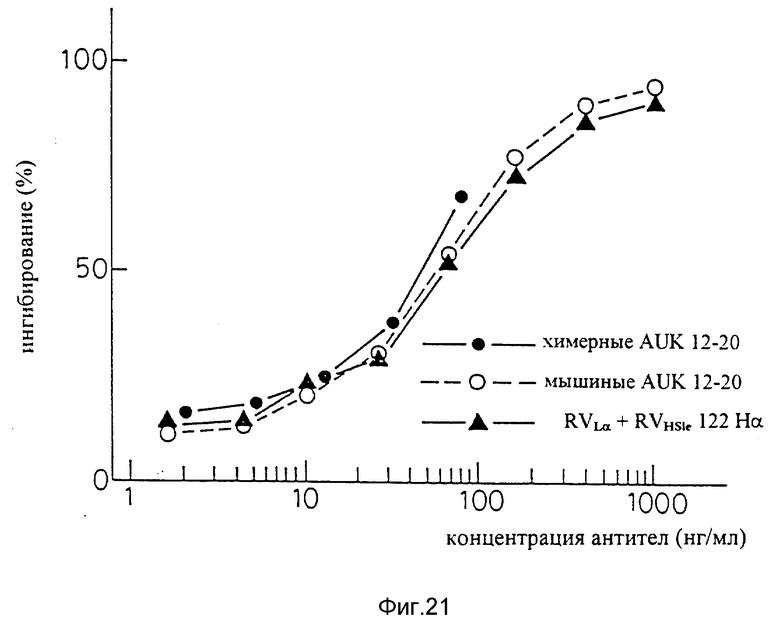
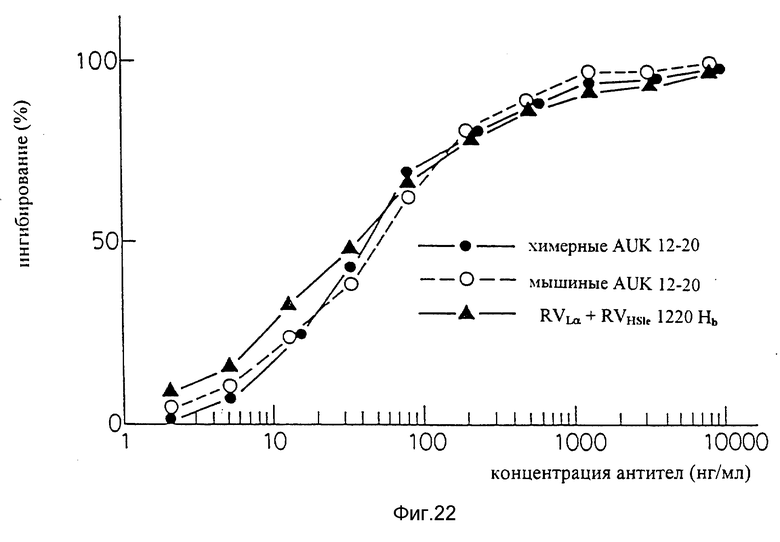
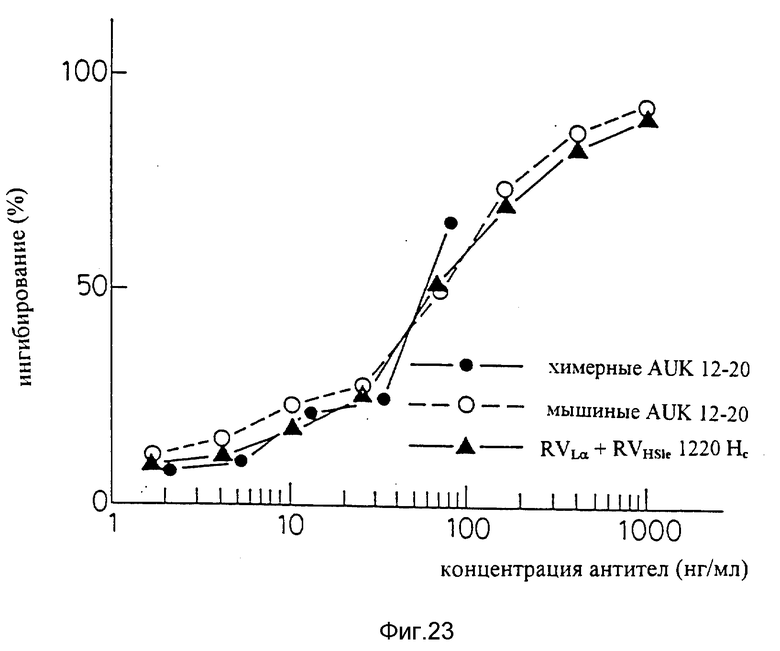
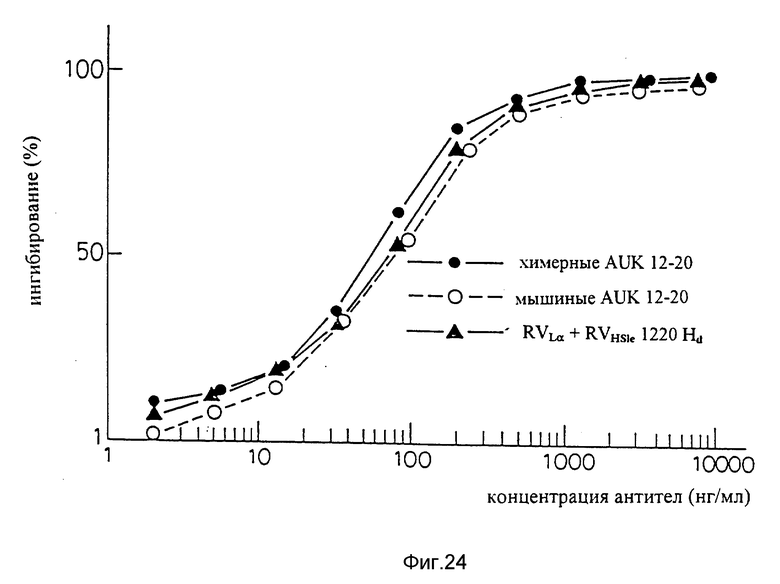
Изобретение представляет V-область L-цепи мышиных моноклональных антител к человеческим IL-6R и Н-цепь V-области мышиных моноклональных антител к человеческим IL-6R. Также представлены модифицированные антитела к человеческим IL-6R. Представлена также ДНК-последовательность, кодирующая вышеназванные антитела или их части. Антитела к рецепторам человеческого IL-6R применимы в качестве терапевтического средства для лечения миеломы. 18 с. и 6 з.п.ф-лы., 24 ил., 11 табл.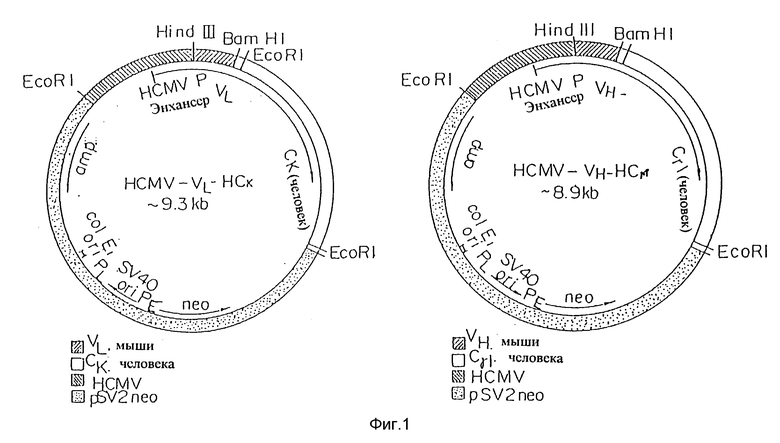
| 1972 |
|
SU413908A1 |

