Изобретение относится к области биотехнологии и касается средств для получения рекомбинантного инсулина человека.
Инсулин - гормон поджелудочной железы - представляет собой глобулярный белок, состоящий из двух полипептидных цепей (A и B), связанных дисульфидными связями. Его недостаток в крови приводит к тяжкому заболеванию - инсулинозависимому сахарному диабету. Единственным способом терапии являются регулярные инъекции инсулина. Первоначально для этих целей использовался только инсулин животного происхождения, выделяемый из поджелудочной железы сельскохозяйственных животных, как правило свиней. Но такой подход не позволяет обеспечивать всех нуждающихся в регулярных инъекциях инсулина. В связи с этим были разработаны способы получения гормона методами генной инженерии, базирующиеся на создании на основе клеток E.coli штаммов-продуцентов гибридных белков - предшественников инсулина человека, которые в больших количествах накапливаются в клетках в виде тел включения. Гибридный белок состоит из лидерного пептида и пептида с аминокислотной последовательностью проинсулина, в котором A- и B-цепи соединены так называемым C-пептидом. Для получения биологически активного гормона необходимо отщепить лидерный пептид, ренатурировать проинсулин, образовав три необходимые дисульфидные связи, и удалить соединительный C-пептид.
Известен [FEBS Letters, 402, 1997, 124-130] способ получения генно-инженерного инсулина человека, основанный на культивировании штамма, продуцирующего гибридный белок, в котором аминокислотные последовательности лидерного пептида и проинсулина соединены через остаток метионина. Продуцирующий штамм несет в себе плазмиду, полученную следующим образом: в качестве матрицы использовалась коммерчески доступная библиотека кДНК из поджелудочной железы человека; кДНК гибридного белка получали с помощью полимеразной цепной реакции, при этом праймеры были синтезированы таким образом, чтобы ввести в амплифицируемый фрагмент сайты рестриктаз BamH 1 и Sal I на 5'- и 3'-концах, соответственно, для клонирования в экспрессирующем векторе pQE-31 (Qiagene); дополнительно был введен триплет, кодирующий метионин, непосредственно перед 5'-концом последовательности, кодирующей проинсулин; амплифицированный фрагмент после расщепления рестриктазами BamH I и Sal I лигировали в обработанный теми же рестриктазами и дефосфорилированный вектор с помощью Т4 ДНК-лигазы. Полученной плазмидой трансформировали компетентные клетки Е. coli штамм XL2-Blue MRF'. Трансформированный штамм-продуцент гибридного белка выращивали на среде LB, содержащей 100 мкг/мл ампициллина, индуцируя экспрессию гена изопропил -β-D- тиогалактопиранозидом (ИПТГ). По окончании культивирования из биомассы выделяли тела включения, растворяли гибридный белок в 70% муравьиной кислоте и отщепляли лидерный пептид действием бромциана. Затем пептид с аминокислотной последовательностью проинсулина подвергали окислительному сульфитолизу, а полученный S-сульфонат проинсулина очищали и ренатурировали. Далее очищенный, ренатурированный проинсулин превращали в биологически активный гормон обработкой трипсином и карбоксипептидазой B по методу Кемлера. Описанная технологическая схема является наиболее распространенной.
Недостатками описанного способа являются, во-первых, многостадийность технологического процесса, и, во-вторых, использование для отщепления лидерного пептида высокотоксичного реагента - бромциана.
Известно [Journal of Biotechnology, 48, 1996, 241-250] использование для получения инсулина человека плазмиды pTrpZZ-R-proinsulin, экспрессирующей гибридный белок, в котором два IgG-связывающих домена белка A и пептид с последовательностью проинсулина соединены через остаток аргинина, что позволяет отказаться в технологической схеме от использования высокотоксичного реагента - бромциана. В качестве хозяина для плазмиды в этом случае были использованы клетки E.coli штамм O17 и получение инсулина включало следующие стадии: культивирование штамма-продуцента, выделение тел включения, солюбилизацию гибридного белка, сульфитолиз и ренатурирование гибридного белка, его очистку аффинной хроматографией на IgG-Сефарозе и расщепление гибридного белка трипсином и карбоксипептидазой-B с получением инсулина, который далее подвергали хроматографической очистке.
Основными недостатками такого способа-прототипа являются использование аффинной хроматографии, которая нетехнологична при крупномасштабном производстве, и низкий выход на стадии ферментативного превращения гибридного белка в инсулин (44%).
Изобретение направлено на упрощение технологического процесса получения инсулина человека с одновременным увеличением выхода конечного продукта.
Для достижения данного технического результата предложен способ получения инсулина человека, включающий культивирование клеток E.coli, выделение тел включения, последовательно солюбилизацию, сульфитолиз и ренатурирование гибридного белка, очистку ренатурированного гибридного белка, его расщепление трипсиноподобным и карбоксипептидазо-B-подобным ферментами и выделение инсулина, при этом культивируют новый штамм E.coli JM-109-pRRproins, несущий новую плазмиду pRRproins, или второй новый штамм E.coli JM-109-pKRproins, несущий другую новую плазмиду pKRproins, a ренатурированный белок очищают методом ионообменной хроматографии. Для повышения степени чистоты целевого продукта гибридный белок после сульфитолиза перед ренатурированием целесообразно дополнительно очищать анионообменной хроматографией.
Штамм Е. coli JM-109-pRRproins, несущий плазмиду pRRproins, является продуцентом гибридного белка, в котором аминокислотные последовательности лидерного пептида, представляющего собой IgG- связывающий домен белка A, и проинсулина человека соединены двумя остатками аргинина.
Штамм Е. coli JM-109-pKRproins, несущий плазмиду pKRproins, является продуцентом гибридного белка, в котором аминокислотные последовательности лидерного пептида, представляющего собой IgG-связывающий домен белка A, и проинсулина человека соединены остатками лизина и аргинина.
Новая рекомбинантная плазмида pRRproins, экспрессирующая гибридный белок, в котором аминокислотные последовательности лидерного пептида, представляющего собой IgG-связывающий домен белка A, и проинсулина человека соединены двумя остатками аргинина, имеет размер 5025 п.о. и состоит из следующих элементов: EcoR I - Hpa I фрагмента (212 п.о.), кодирующего лидерный пептид, два связующих остатка аргинина и две первые аминокислоты B-цепи инсулина; Hpa I-Hind III фрагмента (257 п.о.), кодирующего остаток В3-В30 B-цепи инсулина, C-пептид и A-непь инсулина; Hind III - EcoR I фрагмента (4556 п.о.), несущего в себе tac-промотор и ген устойчивости к ампициллину.
Другая новая рекомбинантная плазмида pKRproins, экспрессирующая гибридный белок, в котором аминокислотные последовательности лидерного пептида, представляющего собой IgG-связывающий домен белка A, и проинсулина человека соединены остатками лизина и аргинина, имеет размер 5025 п.о. и состоит из следующих элементов: EcoR I - Hpa I фрагмента (212 п.о.), кодирующего лидерный пептид, связующие остатки лизина и аргинина и две первые аминокислоты B-цепи инсулина; Hpa I-Hind III фрагмента (257 п.о.), кодирующего остаток В3-В30 B-цепи инсулина, C-пептид и A-цепь инсулина; Hind III - EcoR I фрагмента (4556 п. о. ), несущего в себе tac-промотор и ген устойчивости к ампициллину.
На фиг. 1 приведена схема плазмиды pNY1, являющейся исходной для получения плазмид pRRproins и pKRproins. На фиг. 2 и фиг.3 показаны схемы конструирования плазмид pRRproins и pKRproins соответственно. На фиг.4 приведена схема технологического процесса получения инсулина человека.
Плазмиды pRRproins и pKRproins получают путем направленного мутагенеза известной плазмиды pNY1 [Антибиотики и химиотерапия, 39, 1994, N4, 3-7; Биоорганическая химия, 18, 1992, N12, 1478- 1486], кодирующей гибридный белок, в котором аминокислотные последовательности проинсулина человека и лидерного пептида, представляющего собой один IgG - связывающий домен белка A, соединены через остаток метионина. Плазмида pNY1 (фиг. 1) в составе штамма E. coli JM-109-I хранится в коллекции культур Государственного научного центра антибиотиков (ГНЦА) под номером 1864 и при необходимости выделяется из него методом Бирнбойма-Доли [Nucleic Acids Res., 7,1979,1513-1523].
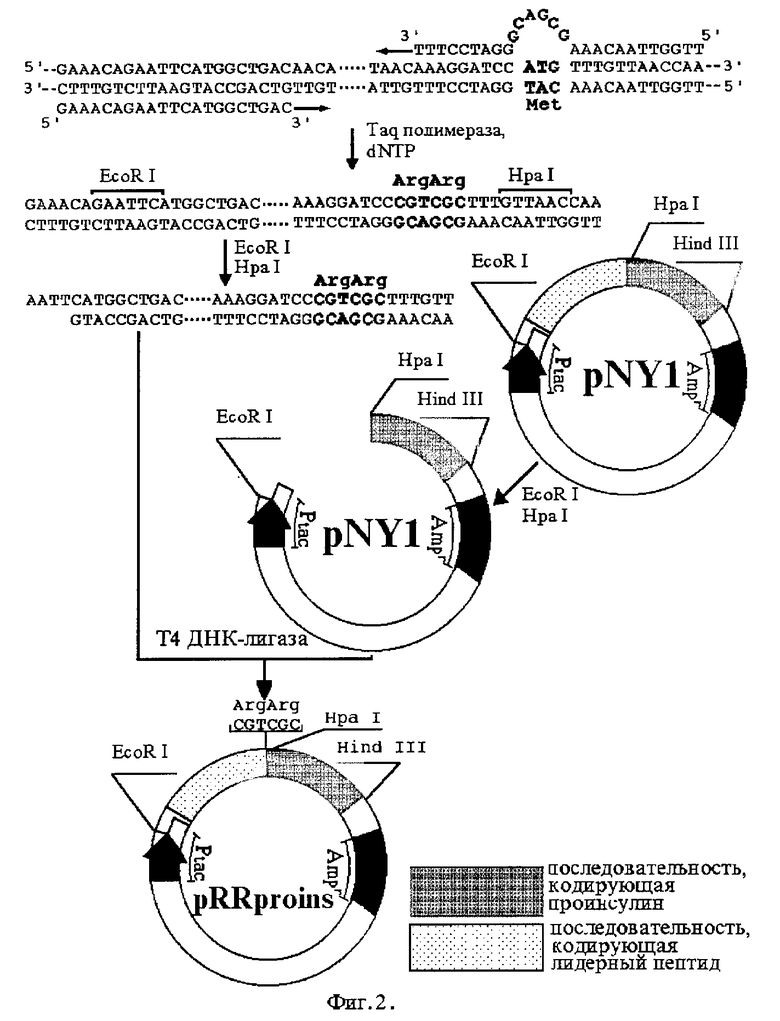
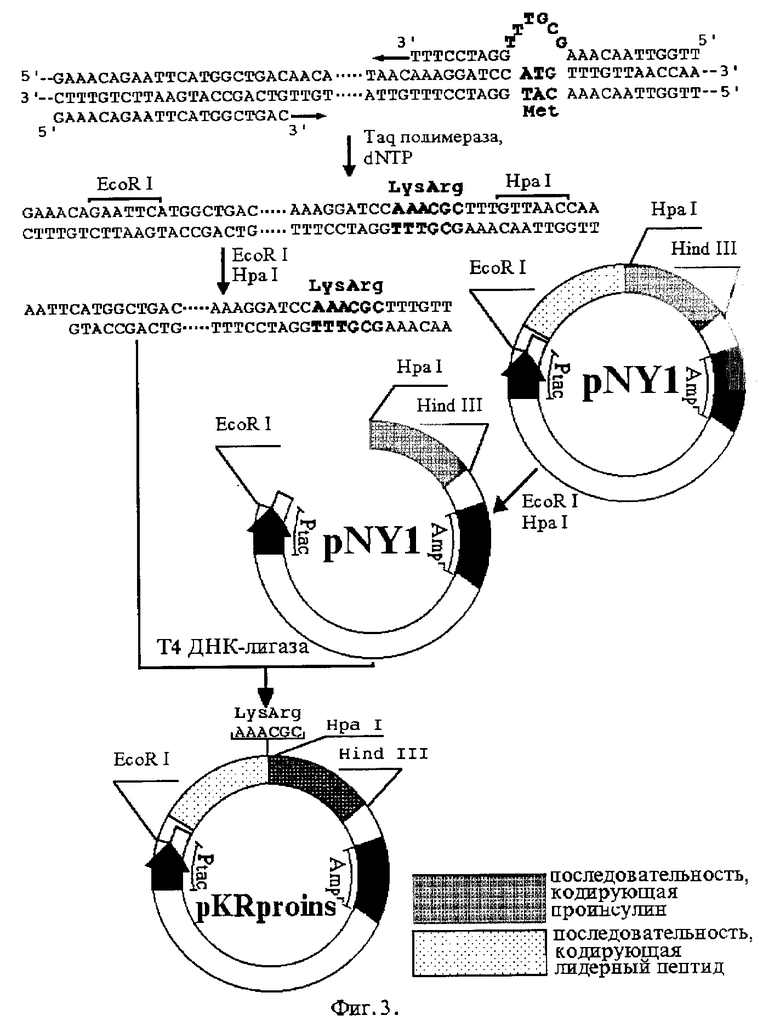
Цель мутагенеза заключается: для плазмиды pRRproins - в замене триплета ATG, кодирующего метионин, на гексануклеотид CGTCGC, кодирующий два аргинина (фиг. 2), а для плазмиды pKRproins - в замене того же триплета ATG на гексануклеотид AAACGC, кодирующий пару лизин- аргинин (фиг. 3). Фрагмент исходной плазмиды, кодирующий лидерный пептид, амплифицируют полимеразной цепной реакцией с помощью химически синтезированных олигонуклеотидов, один из которых вместо триплета CAT, соответствующего метионину (имеется в виду комплементарная цепь), содержит гексануклеотид GCGACG, соответствующий паре аргинин-аргинин (для плазмиды pRRproins) или гексануклеотид GCGTTT, соответствующий паре лизин-аргинин (для плазмиды pKRproins). Изменяемые сайты на фиг. 2 и фиг. 3 отмечены жирным шрифтом. Амплифицированный фрагмент расщепляют рестриктазами EcoR I и Hpa I и лигируют в плазмиду pNY1, из которой по тем же сайтам вырезана исходная последовательность, кодирующая лидерный пептид. Каждая из полученных рекомбинантных плазмид содержит структурный ген соответствующего гибридного белка, экспрессирующийся под контролем сильного tac-промотора, и ген устойчивости к ампициллину.
Штаммы-продуценты E. coli JM-109-pRRproins или E.coli JM-109-pKRproins получают путем трансформации клеток E. coli штамма JM-109 соответственно плазмидой pRRproins или pKRproins. После трансформации случайным образом отбирают колонии, выращенные на среде с ампициллином, выделяют из них плазмиды и подвергают их рестриктному анализу и секвенированию по методу Сенгера. Линию клеток, несущую плазмиду с заданной мутацией, несколько раз пересевают на среду с ампициллином и полученной моноклоновой культурой инокулируют 5 мл жидкой среды с ампициллином. Часть культуры используют для проверки индуцируемой экспрессии гибридного белка с помощью SDS-электрофореза в полиакриламидном геле (ПААГ). К оставшейся части добавляют глицерин и клетки хранят при минус 40oC.
Полученные штаммы-продуценты характеризуются следующими признаками:
Морфологические признаки. Клетки прямые, палочковидные, размером около 2 мкм, граммотрицательные, неспороносные, с хорошо различимыми телами включения после индукции синтеза гибридного белка.
Культуральные признаки. Клетки хорошо растут на обычной среде LB, содержащей бактотриптон и дрожжевой экстракт. На агаризованной среде LB через 24 часа роста при 37oC образуются крупные (3-4 мм) колонии желтоватого цвета. При росте в жидкой среде образуется равномерная муть желтого цвета.
Физиолого-биохимические признаки. Клетки растут при 15-39oC с оптимумом при 37oC при pH 6,8-7,0. Источник углерода - глюкоза. Источники азота - бактотриптон, бактопептон, дрожжевой экстракт и т.п.
Устойчивость к антибиотикам. Клетки обладают устойчивостью к 50-100 мкг/мл ампициллина.
Стабильность плазмиды в штамме. При поддержании клеток в течение 6 месяцев на агаризованной среде, содержащей ампициллин, не наблюдаются потери или перестройки плазмиды, влияющие на экспрессию гибридного белка.
B изобретенных штаммах гибридный белок синтезируется эндогенно в виде тел включения, и его содержание составляет 30-40% от общего количества клеточных белков.
Изобретенный способ получения инсулина человека осуществляется следующим образом (фиг.4):
- культивируют штамм E.coli JM-109-pRRproins или E.coli JM-109- pKRproins на любой подходящей среде, например на богатых средах LB medium. Superbroth, TY medium и т.п.;
- выделяют тела включения, например, суспендируя биомассу клеток в подходящем буферном растворе, дезинтегрируя ее и подвергая центрифугированию либо микро- или ультрафильтрации;
- солюбилизируют гибридный белок любым подходящим растворителем, не нарушающим его первичную структуру, например растворами мочевины, гуанидиния хлорида, фосфорной или муравьиной кислот;
- известными методами последовательно осуществляют сульфитолиз и ренатурирование гибридного белка;
- ренатурированный гибридный белок очищают ионообменной хроматографией;
- очищенный ренатурированный гибридный белок расщепляют трипсиноподобным и карбоксипептидазо-B-подобным ферментами; ферментативный протеолиз может осуществляться последовательным применением указанных ферментов либо чаще используемым одностадийным методом Кемлера [J. Biol. Chem., 246, 1971, 6786- 6791];
- выделяют инсулин из реакционного раствора любым известным методом, например препаративной обращенно-фазовой (ОФ) ВЭЖХ, катионообменной хроматографией на S-Сефарозе или Фосфо-Биохроме К, анионообменной хроматографией на TSK-DEAE-650M или Q-Биохроме A, гель-хроматографией на сефадексе G-50 или комбинацией этих методов.
Для получения высокоочищенного инсулина, соответствующего фармакопейным требованиям, целесообразно после сульфитолиза перед ренатурированием подвергнуть гибридный белок очистке с помощью анионообменной хроматографии, которая может быть выполнена, например, на таких сорбентах, как DEAE-Сефароза, DEAE-Сефароза FF, DEAE-Toyopearl, Mono Q HR, DEAE-Сферанит ОН, Q-Биохром.
Таким образом, использование в изобретенном способе либо штамма- продуцента, полученного на основе новой плазмиды pRRproins, либо штамма-продуцента, полученного на основе новой плазмиды pKRproins, дает возможность исключить использование в технологической схеме высокотоксичного реагента бромциана и использовать для очистки ренатурированного белка ионообменную хроматографию вместо аффинной хроматографии на IgG-сефарозе, что упрощает технологический процесс и делает его более пригодным для промышленного использования в крупномасштабном производстве. Одновременно данный новый способ позволяет увеличить выход конечного продукта с единицы биомассы сухих клеток за счет уменьшения молекулярной массы балластной части гибридного белка - лидерного пептида (м.в. гибридного белка в способе-прототипе - 25 kDa; м.в. гибридного белка в изобретенном способе - 16,8 kDa) и за счет большего уровня экспрессии (25% - в способе-прототипе, до 40% - в изобретенном способе). В изобретенном способе получения инсулина выход на стадии ферментативного расщепления составляет 65% при использовании штамма E.coli JM-109-pRRproins и 80% при использовании штамма E.coli JM-109-pKRproins, что существенно выше, чем в способе-прототипе (44%).
Пример 1. Получение плазмиды pRRproins.
Плазмиду pRRproins получают путем направленного мутагенеза известной плазмиды pNY1 (фиг. 1) в соответствии со схемой, приведенной на фиг. 2.
Плазмиду pNY1 выделяют из штамма-продуцента E.coli JM-109-I N1864, хранящегося в коллекции штаммов ГНЦА. Для этого коллекционный штамм рассевают на чашки с LB-агаром, содержащем 100 мкг/мл ампициллина, и отдельно стоящей колонией инокулируют 5 мл жидкой среды LB, содержащей 100 мкг/мл ампициллина. Культуру инкубируют при 37oC и интенсивном встряхивании в течение ночи. 1 мл полученной ночной культуры вносят в 500 мл среды LB с ампициллином и инкубируют при 37oC до насыщения (OD600≈4,0). Клетки осаждают центрифугированием при 6000g в течение 10 мин и ресуспендируют в 4 мл 25 мМ трис-HCl (pH 8,0), содержащем 50 мМ глюкозы и 10 мМ ЭДТА. К суспензии добавляют 1 мл раствора лизоцима (25 мг/мл) и инкубируют 10 мин при комнатной температуре. Далее добавляют 10 мл раствора 0,2М NaOH с 1% SDS и перемешивают до полного осветления суспензии. Выдерживают во льду 10 мин. К полученному раствору добавляют 7,5 мл 3М ацетата калия и снова выдерживают во льду 10 мин. Сформировавшийся осадок отделяют центрифугированием при 20000g, а из супернатанта высаживают плазмиду 0,6 объемами изопропанола. Кольцевую форму плазмиды получают равновесным центрифугированием в хлориде цезия в присутствии бромистого этидия (500000g, 14 час) и используют для направленного мутагенеза.
Для внесения в нуклеотидную последовательность плазмиды pNY1 мутации синтезируют олигонуклеотиды:
I - 5'GAAACAGAATTCATGGCTGAC 3' и
II - 5'TTGGTTAACAAAGCGACGGGATCCTTT 3'.
100 мкл реакционной смеси для полимеразной цепной реакции содержит 1 мкг плазмиды, 2,5 ед. Tag полимеразы и следующие компоненты в указанных концентрациях: 1 мкМ праймер I, 1 мкМ праймер II, 10 мМ трис-HCl (pH 8,4), 50 мМ KCl, 1,5 мМ MgCl2, 20 мкг/мл желатина, 0,8 мМ смесь всех четырех дезоксинуклеотидтрифосфатов. Реакцию осуществляют по следующей схеме: 94oC - денатурирование (1 мин), 45oC - отжиг (2 мин), 72oC - полимеризация (1 мин). Всего проводят 30 циклов амплификации. По окончании реакции 1 мкл смеси анализируют электрофорезом в 1% агарозном геле в 1xTBE (89 мМ трис-основание, 89 мМ H3BO3 2 мМ ЭДТА) в присутствии 0,5 мкг/мл бромистого этидия. Амплифицированный фрагмент очищают ионообменной ВЭЖХ на колонке DEAE-NPR (2,6х150 мм) в градиенте NaCl от 0 до 1М.
1 мкг плазмиды pNY1 расщепляют рестриктазами EcoR I и Hind III в следующих условиях: 6 мМ трис-HCl (pH 7,6), 6 мМ MgCl2, 6 мМ 2-меркаптоэтанол и 50 мМ NaCl, 2 ед. EcoR I, 2 ед. Hind III. Объем реакционной смеси - 30 мкл. Реакцию проводят 1 час, при 37oC и останавливают прогреванием при 65oC. EcoR I-Hind III фрагмент плазмиды pNY1 (4556 п.о.) выделяют электроэлюированием из 1% агарозного геля и дефосфорилируют бактериальной щелочной фосфатазой.
Аналогично расщепляют амплифицированный фрагмент и лигируют его в подготовленную плазмиду с помощью Т4-ДНК лигазы при 11,8oC в течение ночи. Полученной после лигирования смесью трансформируют клетки E.coli штамм TG-1 с целью отбора плазмиды, несущей заданную мутацию.
Трансформацию проводят следующим образом. Культуру выращивают до оптической плотности 0,6 Ед при 600 нм, центрифугируют и клетки аккуратно суспендируют в 0,1 М ледяном растворе CaCl2. Суспензию помещают на 40 мин в ледяную баню, собирают центрифугированием и ресуспендируют в 1/20 первоначального объема хлорида кальция. К 100 мкл полученных компетентных клеток добавляют 30 мкл смеси после лигирования, оставляют на 40 мин на ледяной бане и подвергают тепловому шоку (42oC, 2 мин). Клетки разводят в 10 раз средой LB, подращивают при 37oC в течение 1 часа и высевают на чашки с агаризованной средой LB, содержащей 100 мкг/мл ампициллина.
Случайным образом отбирают колонии, способные расти на среде, содержащей 100 мкг/мл ампициллина. Выделенные из отдельных колоний плазмиды подвергают рестриктному анализу и секвенируют по методу Сенгера. В результате отбирают линию клеток, несущих плазмиду pRRproins.
Пример 2. Получение плазмиды pKRproins.
Плазмиду pKRproins получают путем направленного мутагенеза известной плазмиды pNY в соответствии со схемой, приведенной на фиг.3. Для этого синтезируют олигонуклеотиды:
I - 5'GAAACAGAATTCATGGCTGAC 3'
II - 5'TTGGTTAACAAACGCTTTGCATCCTTT 3'.
Получение осуществляют способом, описанным в примере 1. В результате отбирают линию клеток, несущих плазмиду pKRproins.
Пример 3. Получение штамма E.coli JM-109-pRRproins.
Плазмидой pRRproins трансформируют компетентные клетки E.coli штамм JM-109 методом, описанным в примере 1. Отдельно локализованную колонию трижды пересевают на чашки со средой LB, содержащей 100 мкг/мл ампициллина. Полученной моноклоновой культурой инокулируют 5 мл жидкой среды LB и инкубируют в течение ночи, при интенсивном встряхивании, при 37oC. Для проверки способности полученного штамма к индуцированной экспрессии гибридного белка 300 мкл ночной культуры переносят в 2,7 мл подогретой среды LB и выращивают при 37oC до достижения поздней логарифмической фазы (≈1 ч 50 мин). К культуре добавляют ИПТГ до концентрации 0,1 мМ и инкубируют при 37oC еще 4 часа. Клетки собирают центрифугированием и промывают водой для удаления среды, осадок растворяют кипячением в течение 5 минут в присутствии SDS и 2-меркаптоэтанола и наличие индуцированной экспрессии гибридного белка проверяют SDS-электрофорезом в ПААГ. По результатам сканирования окрашенного Coomassie R-250 геля содержание гибридного белка составляет 35±5% от общего белка клетки.
Полученный штамм E.coli JM-109-pRRproins хранят в 20% глицерине при минус 40oC.
Пример 4. Культивирование штамма-продуцента - E.coli JM-109- pRRproins.
Штамм-продуцент выращивают в 15 мл жидкой среды, содержащей 10 г/л бактотриптона (Difco), 5 г/л дрожжевого экстракта (Difco), 5 г/л NaCl и 100 мкг/мл ампицилина, при 37oC, в течение ночи. Полученной ночной культурой инокулируют 10 колб, содержащих по 100 мл той же среды, в соотношении 1:100. Культуры инкубируют при 37oC и интенсивном встряхивании до достижения поздней логарифмической фазы (1 час 50 мин). Биосинтез гибридного белка индуцируют добавлением в среду ИПТГ до концентрации 0,1 мМ. Через 4 часа после индукции собирают 8,57 г биомассы клеток 70% влажности с содержанием гибридного белка 35±5% от общего белка в клетках (по данным SDS-электрофореза в ПААГ).
Пример 5. Получение и культивирование штамма E.coli JM-109-pKRproins.
Штамм E.coli JM-109-pKRproins получают путем трансформации штамма E.coli JM-109 плазмидой pKRproins. Получение, проверку и культивирование штамма проводят по методике, описанной в примерах 3 и 4.
Получают 8,17 г биомассы клеток 70% влажности с содержанием гибридного белка 35±5% от общего белка клетки.
Пример 6. Получение инсулина человека с использованием штамма E.coli JM-109-pRRproins.
Штамм-продуцент E.coli JM-109-pRRproins культивируют способом, описанным в примере 4.
Для выделения тел включения клетки суспендируют в 70 мл водного буферного раствора, содержащего 50 мМ трис-HCl (pH 7,5), 0,1 М NaCl и 0,1 М трилон Б, после чего разрушают ультразвуковым дезинтегратором. В полученной суспензии содержание общего белка, определяемого по методу Лоури, составляет 850 мг. Тела включения отделяют от водорастворимых примесей центрифугированием (15000 g, 30 мин).
Солюбилизируют гибридный белок, растворяя тела включения в течение 6 часов, при 30oC в 60 мл 8 М мочевины, содержащей 0,1 М NaCl, 0,01 М дитиотреитол, 0,01 М трилон Б, 50 мМ трис-HCl (pH 7,5). Нерастворившиеся частицы удаляют центрифугированием (15000 g, 30 мин).
Гибридный белок подвергают сульфитолизу, получая S-сульфонат гибридного белка, для чего в раствор добавляют 1,5 г сульфита натрия и 120 мг тетратионата натрия, доводят pH до 9,0 при помощи 2 М NaOH и оставляют раствор перемешиваться в течение 6 часов. S-сульфонат гибридного белка осаждают доведением pH раствора до 5,2 25% уксусной кислотой и разбавлением раствора в 3 раза дистиллированной водой. Сформировавшийся осадок отделяют центрифугированием (5000 g, 10 мин), растворяют в 25 мл 30 мМ трис-HCl (pH 7,5) и хроматографируют в том же буфере на колонке с DEAE-Сефарозой FF (1 х 25 см) в градиенте концентрации от 0 до 1 М NaCl в течение 4 часов. Скорость элюции - 40 мл/час. Фракции анализируют на наличие S-сульфоната гибридного белка ионообменной ВЭЖХ (колонка Protein РАК DEAE 5 PW (7,5 мм х 7,5 см) (Waters), 30 мМ трис-HCl (pH 7,5), 1 мл/мин, градиент NaCl от 0 до 1 М), используя заведомо чистый S-сульфонат. Фракции с S-сульфонатом гибридного белка объединяют, обессоливают на колонке 2,5 х 20 см с Сефадексом G-25 F в 50 мМ NH4HCO3 и лиофилизируют. В результате получают 177 мг S-сульфоната гибридного белка 90% чистоты, выход которого на этом этапе составляет 53% от гибридного белка в биомассе.
Далее S-сульфонат гибридного белка подвергают ренатурированию: его растворяют в 265 мл охлажденного до +4oC 50 мМ глицинового буфера (pH 10,5), добавляют 12,7 мг цистина и после его растворения добавляют 7,54 мкл 2-меркаптоэтанола. Раствор перемешивают в течение 24 часов, поддерживая pH 10,5 добавлением 0,2М NaOH. Увеличение концентрации ренатурированного гибридного белка в растворе контролируют обращенно-фазовой (ОФ) ВЭЖХ, используя заведомо чистый образец ренатурированного гибридного белка. По окончании реакции раствор концентрируют на установке Minitan с мембраной PTGC (Millipore, США) до 20 мл, обессоливают, как указано выше, и лиофилизируют. В результате получают 169 мг продукта, содержащего 48 мг ренатурированного гибридного белка (выход на стадии 30%; выход на этом этапе по отношению к гибридному белку в биомассе -15,9%).
Очистку ренатурированного гибридного белка проводят на колонке 1 х 11 см с S-Сефарозой. 167 мг образца растворяют в 15 мл 50 мМ CH3COONa (pH 4,2), содержащего 6 М мочевину, и хроматографируют в том же буфере в градиенте концентрации от 0 до 0,5 М NaCl в течение 4 часов при скорости потока 40 мл/час. Фракции с ренатурированным гибридным белком (по данным ОФ ВЭЖХ) объединяют, обессоливают и лиофилизируют. В результате получают 48 мг ренатурированного гибридного белка 90% чистоты. (Выход на данной стадии 90%, выход на этапе по отношению к гибридному белку в биомассе - 14,34%).
Ферментативный протеолиз осуществляют следующим образом: 45 мг полученного образца растворяют в 2 мл 50 мМ трис-HCl (pH 7,8), нагревают до 37oC в термостате, добавляют 9 мкл свежеприготовленного раствора трипсина (5 мг/мл, 35 Ед/мг) и 9 мкл раствора карбоксипептидазы В (5 мг/мл, 135 Ед/мг). Через 30 мин реакцию останавливают добавлением 100 мкл ледяной уксусной кислоты. Два основных продукта реакции идентифицируются сравнением со стандартными образцами методом ОФ ВЭЖХ как инсулин человека и C-пептид. Содержание инсулина в реакционной смеси составляет 8,8 мг, что соответствует 65% выходу на этой ферментативной стадии.
Гидролизат подвергают препаративной ОФ ВЭЖХ в двух циклах на картридже RAD РАК micro Bondapak C 18 в держателе Z-модуль фирмы Waters (США). В качестве подвижной фазы используют градиент концентрации ацетонитрила от 29,6% до 35,2% в течение 1 часа в 0,25 М ацетат-аммонийном буфере (pH 4,5). Скорость элюции - 1 мл/мин. Собирают и объединяют фракции с содержанием инсулина более 97% (по данным ОФ ВЭЖХ). После лиофилизации объединенных фракций получают 4,4 мг высокоочищенного инсулина человека (содержание основного вещества - 98%). Выход на стадии очистки - 50%, общий выход - 4,65%.
Дополнительно аутентичность полученного продукта инсулину человека подтверждают определением N-концевых аминокислотных последовательностей A- и B-цепей; сравнением пептидных карт полученного препарата и стандартного образца после обработки протеазой V8 из S.aureus и масс-спектрометрией (м. м. 5807).
Пример 7. Получение инсулина человека с использованием штамма E.coli JM-109-pKRproins.
Для получения инсулина человека используют биомассу, полученную при культивировании штамма E.coli JM-109-pKRproins по примеру 5.
Способ осуществляют по методике, описанной в примере 6. Содержание белка в суспензии, полученной после ультразвукового разрушения клеток, составляет 817 мг. S-сульфонат, полученный после очистки (169 мг), имеет степень чистоты 90%. Выход на этапе - 53%.
После ренатурации получают 152 мг продукта, содержащего 45,6 мг ренатурированного гибридного белка 90% чистоты (выход на стадии - 30%, выход на этом этапе по отношению к гибридному белку в биомассе - 15,9%).
В результате очистки получают 45 мг ренатурированного гибридного белка 90% чистоты. Выход на стадии - 90%, общий выход - 14,3%.
Выход инсулина на стадии ферментативного протеолиза - 80%, что соответствует 10,8 мг инсулина в реакционной смеси.
После очистки получают 5,4 мг высокоочищенного инсулина человека с содержанием основного вещества 97%. Выход на стадии очистки - 50%, общий выход - 5,72%.

Изобретение относится к биотехнологии. Созданы новые рекомбинантные плазмиды, экспрессирующие гибридный белок, в котором аминокислотные последовательности лидерного пептида, представляющего собой IgG - связывающий домен белка A, и проинсулина человека соединены двумя остатками аргинина (плазмида pRRproins) или остатками лизина и аргинина (плазмида - pKRproins). Инсулин человека получают способом, включающим культивирование штамма E/coli IM-109-pRRproins, несущего плазмиду pRRproins, или штамма E/coli IM-109-pKRproins, несущего плазмиду pKRproins, выделение тел включения, солюбилизацию, сульфитолиз и ренатурирование гибритного белка, очистку ренатурированного гибридного белка методом ионообменной хроматографии, его расщепление трипсиноподобным и карбоксипептидазо-B-подобным ферментами и выделение инсулина. Изобретение упрощает технологический процесс получения инсулина человека и увеличивает выход инсулина. 5 c. и 1 з.п. ф-лы, 4 ил.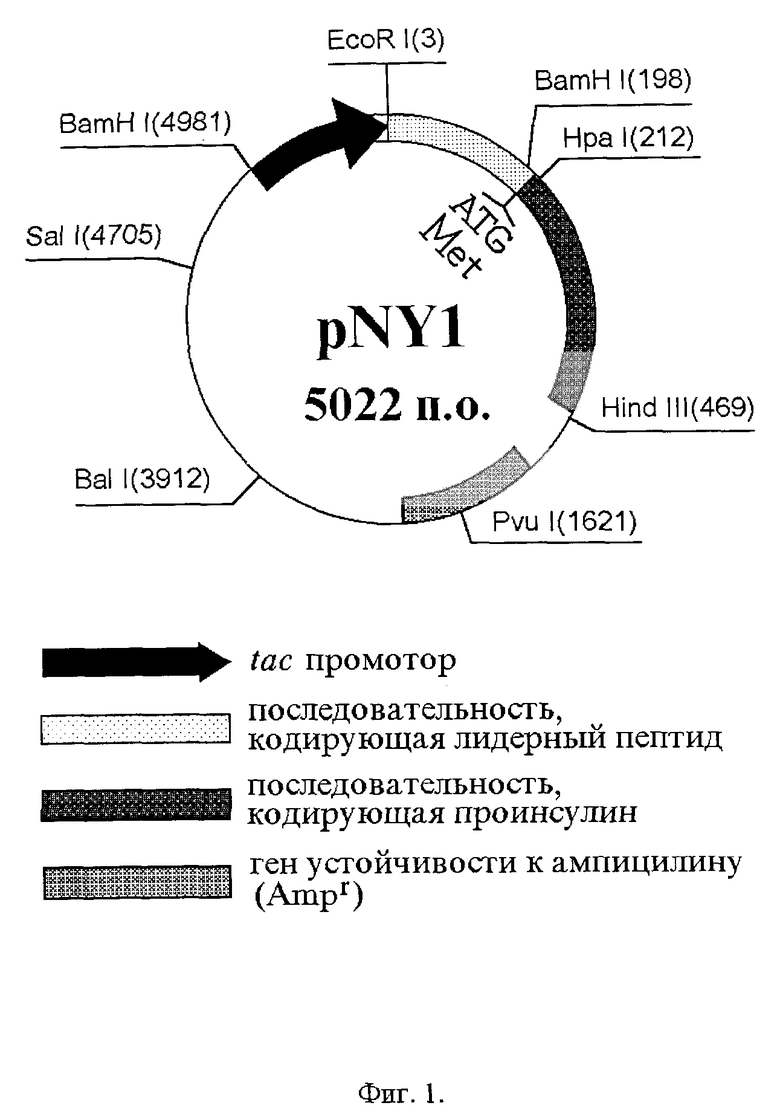
| J.of Biotechnology | |||
| Предохранительное устройство для паровых котлов, работающих на нефти | 1922 |
|
SU1996A1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ ПРЕПРОИНСУЛИН ЧЕЛОВЕКА | 1997 |
|
RU2115729C1 |
| Биоорганическая химия | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |
| Антибиотики и химиотерапия | |||
| Прибор для охлаждения жидкостей в зимнее время | 1921 |
|
SU1994A1 |
