Изобретение относится к области микробиологической и медицинской промышленности и может быть использовано для создания лекарств.
Создание высокоэффективного штамма-продуцента проинсулина - одна из важнейших биотехнологических задач, решению которой посвящены многочисленные исследования последних 10-15 лет.
Предшествующий уровень техники
Существует два принципиально отличных пути: первый путь - создание систем секретирующих проинсулин (Chan et al., Proc. Nat. Acad. Sci. USA 1981, 78, 5401-5405; Stahl & Christiansen, Gene 1988, 71, 147-156; Murby et al. , Biotechnol. Appl. Biochem. 1991, 14, 336-346; Kang & Yoon, J. Biotechnol. 1994, 36, 45-54); второй путь - интрацеллюлярное накопление конечного продукта экспрессии в тельцах включения (Williams et al. Science 1982, 215, 687-689; Guo et al. Gene 1984, 29, 251-254; Nilsson et al., J. Biotechnology 1996, 48, 241-250).
У каждого из этих путей есть свои преимущества и недостатки. В первом случае конечный продукт попадает во внеклеточное пространство. Это упрощает процесс выделения и очистки проинсулина. Однако уровень экспрессии конечного продукта в таких системах невысок. В системах второго типа экспрессия намного выше, однако локализация рекомбинантного белка в тельцах включения требует больших технологических затрат.
Другая проблема - какой именно препроинсулин экспрессируется. Она связана с выбором технологии получения инсулина.
Один путь - ничего не изменяя воспользоваться последовательностью гибридного белка, где проинсулин соединен с лидерным пептидом через метионин (McGregor, Ann. N. Y. Acad. Sci. 1983, 413, 231). Недостатком такого подхода является использование токсичного соединения (бромциана) для отщепления лидерной последовательности.
Другой путь - создание неметионинового линкера между лидерной последовательностью и проинсулином, который можно легко "разрезать" трипсином. Такой подход позволяет отказаться от бромцианового гидролиза и сократить технологическую цепочку получения конечного продукта.
Наиболее близким по технологической сущности и достигаемому результату к предложенному изобретению является создание и использование штамма E.coli, продуцирующего инсулин (Nilsson et al, Integrated production of human insulin and its c-peptide. J. Biotechnology 1996, 48, 41-50).
Известный способ, принятый за прототип изобретения, включает получение полного структурного гена проинсулина человека, содержащего на 5' конце лизин, аргинин или лизил-аргинин, с использованием вектора pRIT37, амплифицирование искусственного IgG-связывающего фрагмента с использованием вектора pEZZ318, перенос этого фрагмента в вектор pTrpBB с образованием вектора pTrpZZ, образование слитого белка, содержащего структурный ген проинсулина и ZZ фрагментов в качестве лидерного пептида, разделенные между собой лизином или аргинином, или лизил-аргинином и трансформацию клеток E.coli RRIΔ M15. Следует отметить, что ZZ-фрагмент является модификацией природного IgG-связывающего B-домена белка А. Введение в конструкцию дополнительного участка, ответственного за связывание IgG, дало возможность авторам приведенной работы модифицировать процедуру выделения и очистки конечного продукта так, чтобы наряду с инсулином было возможным выделять пептид C, который имеет, по их мнению, выраженную биологическую активность.
Однако конструкция, полученная авторами данной работы, содержит достаточно длинный лидерный пептид, что в конечном счете снижает выход инсулина. Помимо более тяжелого лидерного пептида, использование некоммерческих векторов при конструировании гибридного белка делает использование штамма-продуцента дорогим.
Раскрытие изобретения
Задачей настоящего изобретения является разработка простого способа получения штамма-продуцента гибридного белка AproIns, создание конструкции с более легким лидерным пептидом, что позволит сделать процесс выделения инсулина более технологичным и дешевым.
Поставленная задача решается описываемым способом получения штамма-продуцента гибридного белка AproIns для получения инсулина человека, включающим конструирование полного структурного гена проинсулина человека путем химико-ферментативного синтеза, его связывание через остаток аргинина с лидерным пептидом, сконструированным на основе IgG- связывающего домена белка A из S.aureus, при этом конструирование лидерного пептида осуществляют путем предварительного моделирования структуры, модифицированной искусственно внесенным лизином с расчетом оптимальной структуры по параметрам солюбилизации и свободной энергии молекулы белка, последующего химико-ферментативного синтеза выбранной структуры лидерного пептида, переноса в экспрессирующий вектор, предпочтительно в вектор рКК223-3 и трансформацию в клетки E. coli, например, XLI- Blue, JM109. TG1.
Кроме того, поставленная задача решается конструированием полного структурного гена проинсулина человека путем химического синтеза олигонуклеотидов A, B, C, D, E, F, см. I в конце текста.
и поэтапной ферментативной сборки модулей, вначале сборки первого модуля A/B, клонирование полученного модуля в вектор pUC18, затем сборки второго модуля C/D, клонирование его в вектор pUCproInsA/B и сборки третьего модуля E/F, клонирование его в вектор pUCproInsA/D.
Поставленная задача решается также химико-ферментативным синтезом выбранной структуры лидерного пептида следующим образом. Вначале путем синтеза были получены восемь олигонуклеотидов G, H, I, J, К, L, М, N, см. II в конце текста.
с последующим их фосфорилированием, отжигом и лигированием с вектором pUC18/EcoRI-Sall для получения плазмиды pUC-A.
Предложенное изобретение основано на следующих предпосылках. В экспрессионных бактериальных системах, где a priori не предусмотрена секреция рекомбинантного белка, встает необходимость растворения белковых агрегатов (в частности, "телец включения"). В таких случаях необходимо сразу предусмотреть особенности последующего выделения и очистки рекомбинантного белка. В связи с этим ДНК, кодирующая искомый белок, добавляется искусственным амино-терминальным лидерным пептидом (лидером), который должен отвечать определенным требованиям, т.е. все сводится к поиску наиболее оптимального сочетания особенностей самого лидера, возможностей его адаптации к условиям экспрессии, а также учету характеристик экспрессионной системы. При выборе лидера мы считали необходимым обратить внимание на следующие условия:
обеспечение лидером стабильной экспрессии белка;
влияние на правильный фолдинг белка;
солюбилизацию белковых агрегатов;
достаточную афинность связывания для последующего выделения и очистки;
возможность протеолитического расщепления на границе "лидер-белок" за счет искусственного привнесения сайта протеолиза, либо наличие такового в последовательности изначально;
влияние на секретируемость (в качестве сигнальной последовательности в секретирующих системах);
эффективность чрезмембранного транспорта (в секретирующих системах).
Таким образом, выбор лидера должен определяться тем, чтобы он, по возможности, максимально удовлетворял большинству требований. Наш выбор был сделан в пользу одного из Ig-связывающих доменов белка A из Staphylococcus aureus, который обладает сразу несколькими удачными с точки зрения биотехнологии особенностями, однозначно определившими его в качестве лидерной последовательности. Это, в первую очередь, надежная экспрессия слитого вместе с ним инсулина, отсутствие необходимых в таких случаях точечных замен для адаптации к белок-синтезирующему аппарату клетки-хозяина, возможность конечного выделения и очистки слитого белка за счет естественной афинности с Ig, а также отсутствие препятствий для корректного фолдинга зрелого белка.
Предлагаемый подход имеет следующие преимущества:
1. Использование химико-ферментативного конструирования структурного гена эукариотического белка упрощает способ;
2. Выбор наиболее выгодной конструкции лидерного пептида, что способствует высокому уровню экспрессии и эффективной солюбилизации гибридного белка;
3. Появляется возможность использования технологичных методов выделения и очистки конечного продукта.
Лучший вариант осуществления изобретения.
1. Моделирование структуры лидерного пептида.
Особенность одного из этапов технологии экспрессионных систем в E.coli предусматривает растворение телец включения, представляющих собой агрегаты рекомбинантного белка. После разрушения клеточной стенки (ультразвуком, лизоцимом и т. п. ) встает вопрос о растворении белковых агрегатов. Для этого необходимо повысить солюбилизирующие свойства поверхностей рекомбинантных белков. Это может быть достигнуто введением полярных аминокислот в те области белковой молекулы, которые, во-первых, расположены на поверхности и, следовательно, в определенной степени были бы доступны воде, во-вторых, ни коим образом не вносили бы искажения в структуру глобулы, и, в-третьих, не влияли бы на связывающие свойства молекулы с Ig. В этой связи модификация непосредственно инсулина отпадает по следующим причинам: рекомбинантный проинсулин должен совпадать с природным не только по структуре (как первичной, так и трехмерной), но и сохранить ту же (или близкую) биологическую активность. Остается лидерная последовательность, в которой областей для модификации остается немного. Из 58 аминокислот, свернутых в трехспиральный "бочонок" и образующих, таким образом, компактную структуру, стабилизируемую гидрофобными взаимодействиями, точечные замены могут быть произведены только в межспиральной петле. Модификация областей, образующих регулярные структуры (спирали) привела бы к искажению Ig-связывающего центра, что вызвало бы проблемы с очисткой и выделением. Поэтому дополнительная полярная аминокислота, а именно - лизин, была привнесена нами в указанную область.
2. Расчет структуры.
Для определения оптимальной позиции лизина предварительно был проведен расчет общей и доступной для воды поверхностей алгоритмом обкатки радиусом пробной сферы = 1.4  , что соответствует радиусу воды.
, что соответствует радиусу воды.
Поверхностей у белковой глобулы несколько. Более того, в силу подвижности белка некоторые атомы могут переходить из категории "доступных для воды" в категорию "образующих полости".
Подсчет доступной поверхности, анализ которой и был взят за основу в качестве характеристики солюбилизации белка, велся согласно формуле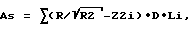
D=dZ/2+d'Z,
где Li - длина дуги данной секции, Zi - перпендикулярная дистанция от центра сферы до секции i; dZ - расстояние между секциями, и d'Z есть меньшая из величин dZ и R-Zi.
Суммарные результаты подсчета поверхностей представлены на фиг. 1.
Как видно из приведенных результатов, наибольшую поверхность, доступную для растворителя, имеют модели Loop6 и Loop8 с позициями лизина 23 и 25, соответственно. При этом не была нарушена структура связывающего центра лидерной части.
Необходимо отметить, что не всегда площадь поверхности, доступной растворителю, однозначно характеризует уровень солюбилизации белка. Наиболее точной характеристикой является доля полярных аминокислот от общего числа поверхностных остатков. Но во всех наших моделях, из-за сравнительно небольшой модификации и небольшого размера молекулы, это значение примерно одинаково, и составляет около 83% с небольшими отклонениями, что не позволяет установить границы отсечения. Однако из-за довольно устойчивой трехспиральной конформации, стабилизируемой гидрофобными связями, все полярные аминокислоты каждый раз оказывались на поверхности, вне зависимости от положения "лишнего" положительного заряда. Таким образом, величиной, характеризующей растворимость молекулы, являлась площадь доступной поверхности.
Кроме параметров, характеризующих солюбилизирующие свойства молекулы, нами проведен сравнительный анализ энергетических параметров моделей, отличающихся позицией лизина (Loop1-8), а также его отсутствием (Loop9), взятым в качестве шаблона.
Моделирование пространственной структуры лидерного пептида проводилось с учетом классических требований, предъявляемых к белковой глобуле в целом: полученная структура должна иметь энергетически стабильную конформацию. Структурная стабилизация молекулы обусловлена взаимодействиями между атомами различной природы, причем энергетические затраты на ее своеобразное "удержание" должны быть минимальными. Таким образом, чем меньше общая свободная энергия системы (в данном случае белковой глобулы), тем более стабильна данная система. Для подсчета общей энергии проводился подсчет отдельных энергий, ее составляющих. В данной работе общая энергия Uоб считалась по формуле
Uоб=Uнв+Uугл+Uторс+Uэл+Uвс,
где Uнв - энергия невалентных взаимодействий;
Uугл - энергия деформации валентных углов;
Uторс - энергия вращения вокруг валентных связей;
Uэл - энергия электростатических взаимодействий;
Uвс - энергия водородных связей.
Каждая из энергетических составляющих считалась наиболее распространенным способом. В частности, для описания Uнв использовался потенциал Леннарда-Джонса
Uнв=Ar-6 + Br-12,
где A и B - некие эмпирические параметры, r - расстояние между парой валентно-несвязанных атомов, а степени "-6" и "-12" характеризуют дисперсионные силы притяжения атомов.
Деформация валентных углов рассчитывалась по закону Гука
Uугл = 1/2KαΔα2,
где Kα - коэффициент упругости, Δα - отклонение валентного угла от своего "идеального" значения.
Торсионный потенциал имеет вид
Uторс = 1/2Uo(1-cosnϕ),
где U0 - потенциальный барьер, ϕ - двугранный угол вращения.
Электростатические взаимодействия обычно рассматриваются в виде взаимодействий точечных парциальных зарядов атомов
Uэл = eiej/εrij,
где ei, и ej - точечные заряды атомов; rij - расстояние между атомами; ε - диэлектрическая проницаемость.
Водородные связи учитывались в соответствии с потенциалом Поланда и Шераги
Uвс=(d/r12-C/r6)+S(r),
где d и C - эмпирические параметры; S(r) - электростатический вклад в энергию водородной связи, оцененный в монопольном приближении.
3. Выбор оптимальной структуры.
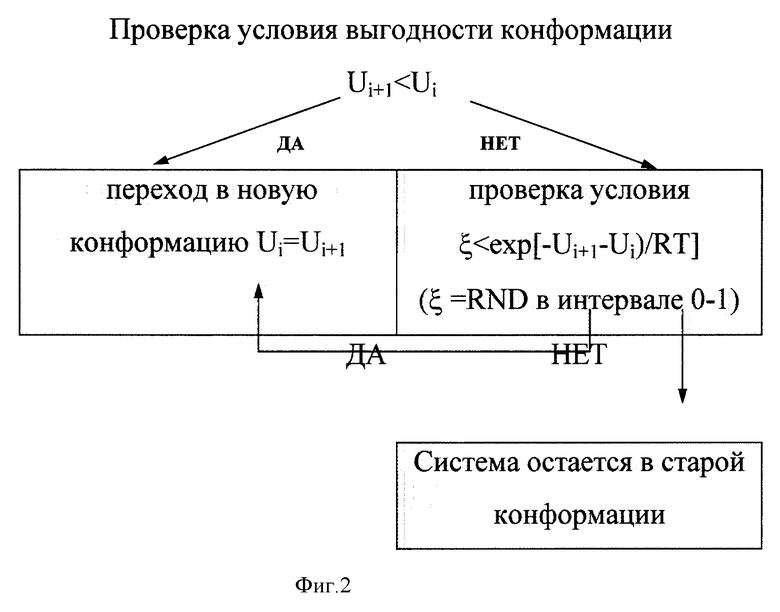
Выбор оптимальной в энергетическом смысле конформации определялся в соответствии с процедурой минимизации, согласно методу Монте-Карло. Суть метода заключается в сравнении значений энергии вновь полученной структуры, U(i+1, с уже имеющейся, Ui. При таком подходе система будет стремиться достигнуть своеобразного "глобального" минимума (фиг. 2).
Таким образом, наиболее оптимальной конструкцией с точки зрения растворимости и стабильности является модель Loop8 (фиг. 3). Пространственная структура полученного лидера показана на фиг. 4. Выбранную оптимальную структуру лидерного пептида получают путем химико-ферментативного синтеза из соответствующих модулей.
Участок структурного гена, кодирующий полипептидную цепь проинсулина, также сконструирован на основе модульного метода, суть которого состоит в химико-ферментативном синтезе отдельных участков гена и их соединения с помощью вводимых сайтов рестрикции.
4. Химико-ферментативный синтез структурного гена человеческого проинсулина.
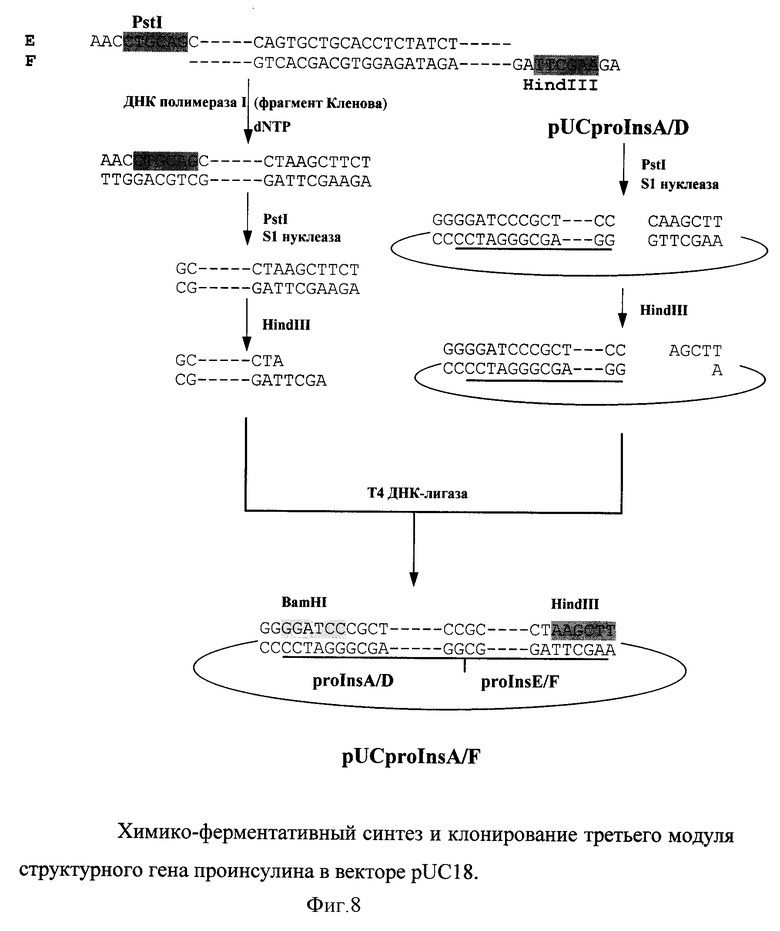
На фиг. 5 представлена полная нуклеотидная последовательность структурного гена гибридного белка AproIns. Горизонтальными линиями отмечены химически синтезированные олигонуклеотиды (A-F), которые использовали для сборки отдельных модулей (A/B, C/D и E/F) структурного гена проинсулина. Вертикальные стрелки указывают на места, куда были введены тетрануклеотидные последовательности для создания временных сайтов рестрикции.
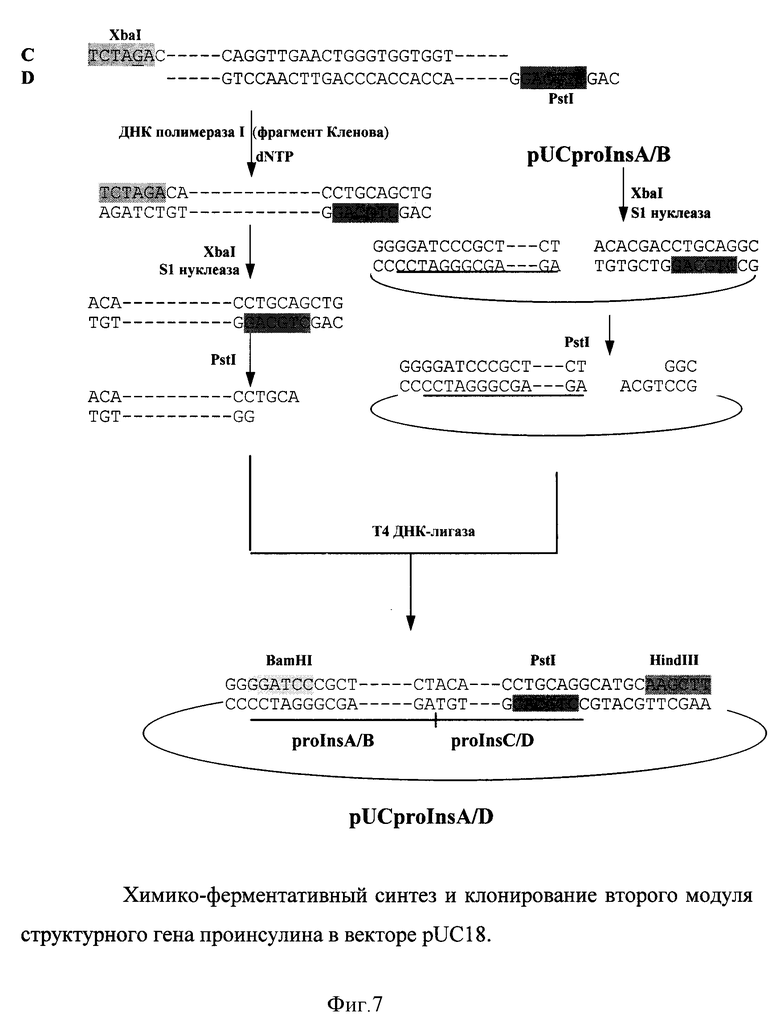
Для сборки первого модуля олигонуклеотиды A(55о) и B(55о) (см. фиг. 6) отжигали в растворе и выступающие концы достраивали фрагментом Кленова ДНК полимеразы I. На фиг. 6 подчеркнут тетрануклеотид (3'GATC5'), введенный для создания временного сайта рестриктазы XbaI.
Полученный двухцепочечный фрагмент расщепляли рестриктазой BamHI и лигировали в вектор pUC18, предварительно разрезанный по уникальным BamHI и HincII сайтам.
Второй модуль собирали аналогично первому из химически синтезированных олигонуклеотидов C(68о. ) и D(52o.) (фиг. 7). В олигонуклеотид D был введен тетрануклеотид 3'ACGT5' для создания временного сайта рестриктазы PstI.
Для удаления ранее введенного XbaI сайта полученный фрагмент и вектор с клонированным первым модулем расщепляли рестриктазой XbaI и обрабатывали S1 нуклеазой. Второй модуль лигировали в вектор pUCproInsA/B по тупому концу и PstI сайту.
Третий модуль был синтезирован из олигонуклеотидов E(66о.) и F(58o.) (фиг. 8). После расщепления по PstI сайту, ранее введенный тетрануклеотид 5'TGCA3' удалялся S1 нуклеазой. Модуль клонировали в векторе pUCproInsA/D после расщепления по Hind III сайту.
Таким образом был реконструирован полный структурный ген проинсулина.
5. Химико-ферментативный синтез лидерного пептида.
Химико-ферментативный синтез выбранной структуры лидерного пептида осуществлялся следующим образом. Синтезировано восемь олигонуклеотидов G, H, I, J, К, L, М, N, см. III в конце текста.
Последовательность ДНК, кодируемая этими олигонуклеотидами, включает транслируемую часть гена, ограниченную с 5'-конца сайтом рестрикции EcoRI и с 3'-конца - HpaI и SalI. Фосфорилированные олигонуклеотиды "отжигали", лигировали с вектором pUC18/EcoRI-SalI и лигазной смесью трансформировали клетки E.coli TG1. Из четырех полученных трансформантов выделяли плазмидную ДНК и последовательность клонированного гена определяли секвенированием по методу Сенгера с использованием стандартных прямого и обратного М13 праймеров. Таким образом была получена плазмида pUC-A.
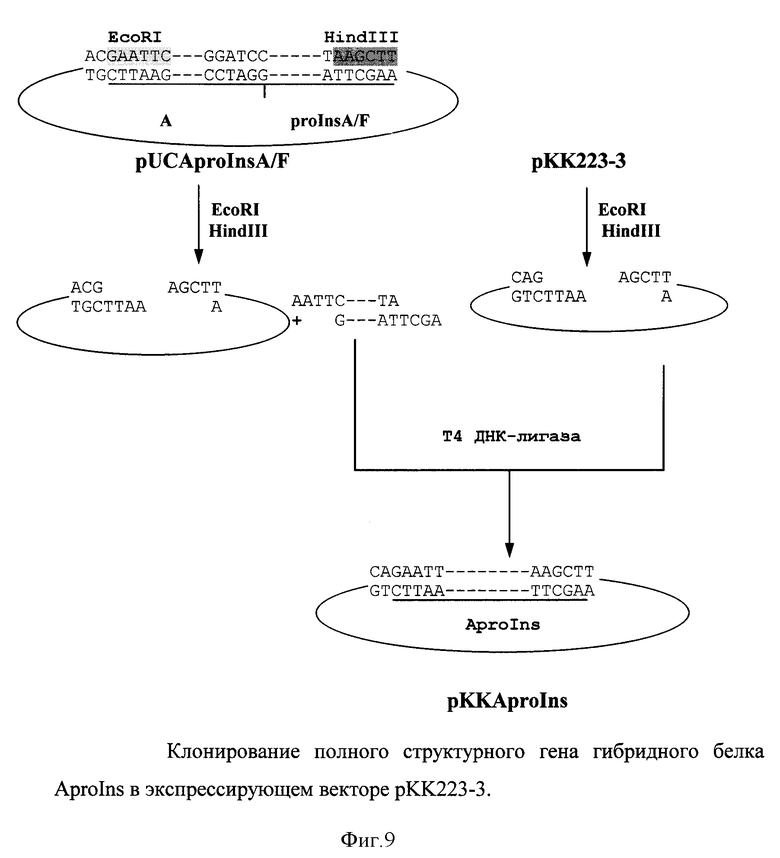
6. Клонирование структурного гена гибридного белка AproIns в эксперссируемом векторе рКК223-3.
EcoRI-HpaI фрагмент гена из плазмиды pUC-A выделяли из агарозного геля, лигировали с вектором pUCproInsA/F/EcoRI-HpaI и лигазной смесью трансформировали клетки E. coli. Правильность сборки гена, кодирующего гибридный белок, подтверждали секвенированием по методу Сергера. Далее фрагмент Aprolns вырезали рестриктазами EcoRI и HindIII, клонировали в экспрессирующий вектор рКК223-3 и трансформировали один из штаммов E.coli, в частности, в штаммы XL1-Blue, JM-109 и TG1 (фиг. 9).
В результате осуществления вышеизложенного способа нами получены штаммы - продуценты Escherichia coli AproIns - продуценты гибридного белка для получения инсулина.
7. Свойства штамма.
В качестве примера по данному изобретению ниже приведены свойства одного из полученных нами штаммов, а именно штамма Escherichia coli XLI-Blue/AproIns.
Полученный штамм-продуцент характеризуется следующими признаками:
Морфологические признаки. Клетки палочковидной формы, грамотрицательные, неспороносные.
Культуральные признаки. Клетки хорошо растут на простых питательных средах. На агаре "Дифко" клетки образуют круглые, гладкие колонии с ровными краями. При росте в жидкой питательной среде образуют интенсивную ровную муть.
Физико-химические признаки. Клетки растут в температурном диапазоне от 8 до 40oC, оптимум pH от 6,8 до 7,0. В качестве источников азота, углерода и других необходимых компонентов используются минеральные соли, казеин, пептон, дрожжевой автолизат и т.д.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к тетрациклину и ампициллину.
Штамм - продуцент E.coli XL1-Blue/AproIns отличается от штамма-реципиента E.coli XL1-Blue наличием рекомбинантной плазмидной ДНК pAproIns, которая придает дополнительную устойчивость к ампициллину.
Клетки E. coli XL1-Blue/AproIns являются продуцентом гибридного белка. При индукции изопропил- бета-D-тиогалактозидом происходит интенсивный синтез данного белка, который накапливается в виде телец включения. Выход гибридного белка составил более 30% от суммарного белка клеток.
Таким образом, предложенный способ создания штамма - продуцента гибридного белка AproIns для получения инсулина человека позволяет по простой и безопасной технологии с использованием коммерчески доступных исходных веществ получить штамм-гиперпродуцент, который может быть легко адаптирован к существующим технологиям выделения инсулина человека.











Изобретение относится к биотехнологии. Способ создания штамма E.coli - продуцента гибридного белка AproIns для получения инсулина человека включает конструирование полного структурного гена проинсулина с помощью химико-фармацевтического синтеза, моделирование и расчет оптимальной структуры IgG - связывающего домена белка А, синтез наиболее оптимальной структуры лидерного пептида, связывание его с образованием гибридного белка AproIns, перенос в экспрессирующий вектор и трансформацию клеток E.coli. Способ позволяет создать наиболее оптимальную структуру гибридного белка для получения инсулина человека по простой и безопасной технологии. 2 з.п.ф-лы, 9 ил.
и поэтапной ферментативной сборки модулей, вначале проводят сборку первого модуля A/B, клонирование полученного модуля в вектор pUC18, затем сборку второго модуля C/D, клонирование его в вектор pUCpro Ins A/B и сборку третьего модуля E/F, клонирование его в вектор pUCpro Ins A/D, а оптимальный лидерный пептид синтезируют вначале путем химического синтеза олигонуклеотидов G, H, I, J, K, L, M, N, см. II в графической части,
затем полученные олигонуклеотиды фосфорилируют и "отжигают", лигируют с вектором pUC18/EcoRI-SalI с получением плазмиды pUC-A.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| J.Biotechnology, 1996, 48, p | |||
| Одноколейная подвесная к козлам дорога | 1919 |
|
SU241A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ ПРЕПРОИНСУЛИН ЧЕЛОВЕКА | 1997 |
|
RU2115729C1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| US 5399490, 21.03.1995 | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Доклады АН СССР, 1983, 270, с | |||
| ПРИБОР ДЛЯ ОПРЕДЕЛЕНИЯ ОБЪЕМА ТВЕРДЫХ ТЕЛ | 1923 |
|
SU743A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| АППАРАТ ДЛЯ ОБОГАЩЕНИЯ РУД ПО МЕТОДУ ВСПЛЫВАНИЯ | 1915 |
|
SU279A1 |

