Это изобретение является областью исследований генной инженерии (манипуляции гена) при помощи технологии рекомбинантной ДНК (РН диагностик) и иммунизации/вакцинации. Более точно, изобретение относится к обнаружению, клонированию и анализу последовательности ДНК генома вируса куриной анемии (КАВ) и посредством этого предположению возможных применений.
КАВ вирус, который был классифицирован не так давно, вызывает инфекционную анемию у цыплят. Вирус впервые был выделен в Японии в 1979 году и получил такое имя потому, что вызывал серьезную анемию у молодых птенцов (Jnasa и др., 1979). Другими симптомами КАВ инфекции являются атрофия костного мозга и деструкция лимфоцитов в тимусе. Повреждения найдены в селезенке и печени.
Наиболее подвержены однодневные птенцы. У этих животных вялость, анорекция и протекающая анемия наблюдались через 4 - 7 дней после прививки КАВ, и половина животных умирала между 2 и 3 неделями после инфицирования. С повышением возраста природная резистентность также увеличивается. При заражении птенцов в возрасте семи дней они проявляют только текущую анемию после заражения, и при инфицировании 14-дневных животных анемия не следует.
Защита от КАВ и симптомов КАВ заболевания главным образом основывается на характере иммунных защитных механизмов. Vielitz (1989) разработал практически, достаточно эффективный метод предупреждения с помощью "контролированной экспозиции" с КАВ-инфицированными суспензиями печени несушек, таким образом потомство приобретало материнский иммунитет. В Германии этот метод иммунизации использовался на практике, но, оказывается, он не совершенно свободен от риска.
Эксперименты на животных, проводимые в отдельных птицефермах с Central Diergeneeskundig Institut (CDI) при Lelustac подтвердили защитную ценность материнских антител. Здесь "контролированная экспозиция" проводилась с КАВ, размноженным на тканевой культуре. Присутствие материнских антител против КАВ полностью предупреждало КАВ репликацию при заражении однодневных цыплят от таким образом вакцинированной матери животных. КАВ симптомы тоже не наблюдались. Такая пассивная защита была также достигнута у потомков иммунизированных плодов и также после впрыскивания в специфически патоген-свободных (СПС) птенцов желтковых экстрактов яиц с подобно иммунизированными плодами. Пассивная защита, принимая во внимание КАВ инфекцию, с помощью введения КАВ антител проводится до возраста 4 недели. Позже, пассивная защита, как было найдено, будет недостаточной. Эти эксперименты показали, что материнские антитела, получаемые вакцинацией матери животных, будут играть важную предупреждающую роль в практической ситуации.
Также экспериментально было продемонстрировано, что у птенцов, которые пережили КАВ инфекцию, наблюдается протекание истощения специфичной популяции лимфоцитов тимуса (Jeurissen и др., 1989). Атрофия тимуса возможно вызвана иммунодепрессией вводимого КАВ, приводящей к тому, что специфические вакцинации менее эффективны, например против Neweastle заболевания. КАВ был выделен несколько раз в стаях с увеличенными потерями благодаря Marek's заболеванию, Gemberos болезни (Jnfektion Bursal Disease Virus, IBDV, Juasa и др., 1980) и у животных с Blue Wing заболеванием в ассоциации с ретровирусами (Engstrom 1988a, Engstrom и др., 1988, б)). В экспериментальных двойных инфекциях было показано усиление свойств КАВ, принимая во внимание другие куриные вирусы, например вирус Marek's болезни, MDV De Boer и др., 1989 а/. Недавно мы наблюдали в наших собственных экспериментах резко усиленную реакцию после аэрозольной вакцинации одновременно вакцины Newcastle болезни и КАВ инфекции. Следовательно, КАВ приводит к иммуноподавляющим и усиляющим эффектам при других вирусных инфекциях. Эти свойства КАВ приводят к увеличению инцидента вспышек заразных заболеваний в практике.
Оказывается КАВ распространен во всем мире. Через некоторое время после начала исследования КАВ в Японии, выделение КАВ было осуществлено в Европе, а именно в Германии Von Bulow /1983/ и позже McNulty с сотр. /1990/ в Англии. В Нидерландах первые выделения КАВ из материала из США, Израиля и Туниса были осуществлены De Boer с сотр. /1988/. Имеющиеся литературные данные показывают, что выделения КАВ относятся к одному серотипу, но выделения КАВ нескольких областей были исследованы на внутреннее родство и возможные отличия в патогенности/. McNulty с сотр. 1990/. Распространение КАВ внутри стаи вероятно происходит путем инфицирования через испражнения и воздух. Вертикальное перемещение вируса к потомству тем не менее также играет важную роль в КАВ эпидемиологии. В различных странах наличие КАВ было доказано серологически.
В условиях тканевой культуры КАВ тяжело размножается. До сих пор КАВ приводил только к цитопатологичному эффекту в MDV трансформированных лимфобластоидных клеточных линиях лимфомы Mapeкa /MDCC-MSBI клетки/ или в трансформированных вирусом лейкемии лимфобластоидных клеточных линиях лимфоидного лейкоза /1104-Х5; Уиаза, 1983/.
Недавнее изучение Todd с сотр. /1990/ описывает вирусные частицы, имеющие диаметр 23,5 нм, которые концентрируются при плотности 1,33 - 1,34 г/мл в градиенте CsCl. Вирус имеет один доминирующий полипептид /Mв: 50,000/ и циклическую простую цепочку ДНК генома длиной 2,3 килобаз. Два малых вируса, свиной цирковирус и вирус, ассоциированный с Psittacine Beak и Feathe Disease сходны с КАВ кольцевой одноцепочечной ДНК, но имеют более маленький геном и более маленький диаметр вирусных частиц /Ritchie с сотр., 1989; Tischer с сотр. , 1982/. В течение долгого времени считалось, что КАВ относится к парвовирусам. Хотя большинство парвовирусов являются одноцепочечными ДНК-вирусами, они обладают линейной ДНК, большим геномом и вероятно также другой композицией вирусных полипептидов.
Краткое описание изобретения
Общепринято, что клеточные компоненты, входящие в репликацию и транскрипцию вируса, функционируют только, если ДНК имеет форму двойной спирали. Вирус, имеющий кольцевую одноцепочечную ДНК, может находиться в клетке в фазе двухцепочечной ДНК. Данные исследователи использовали этот факт.
Данные изобретатели охарактеризовали двухцепочечную КАВ ДНК, имеющую длину 2,3 килобаз в КАВ инфицированных 1104-Х5 и MDCC-MSBI клетках и клонировали ее в pIC-20H. ДНК была полностью секвенирована /фиг. 1/. В диагностическом тесте при помощи меченой клонированной КАВ ДНК КАВ нуклеиновые кислоты могли быть обнаружены в вирусе, печени и препаратах клеточной культуры. Найдено, что клонированный КАВ имеет все биологические и патогенные свойства КАВ дикого типа, как в клеточной культуре, так и в тестах на животных.
В реакциях удлинения цепи полимеразой и экспериментах по гибридизации показано, что клонированный полный КАВ геном является типичным представителем КАВ дикого типа.
С помощью Нозерн-анализа с 32P-мечеными ДНК-пробелами было продемонстрировано, что все природные изоляты содержат молекулу ДНК размером 2,3 кб. Рестрикционный анализ продемонстрировал, что клонированная КАВ ДНК соответствует ДНК при разных изолятах. С помощью дот-блок анализа продемонстрировано, что дигоксигенинмеченая клонированная CAV-ДНК специфично гибридизуется с ДНК различных изолятов данного типа. В реакции удлинения цепи полимеразол, использующей олигонуклеотиды, последовательность которых происходит из последовательности клонированного КАВ /фиг. 4/, КАВ ДНК специфически амплифицирована или узнана.
Детальное описание изобретения
Настоящее изобретение дает, во-первых, рекомбинантную генетическую информацию в форме меченых или немеченых ДНК или РНК, содержащих специфические нуклеотидные последовательности вируса куриной анемии /КАВ/, соответствующие или комплементарные нуклеотидной последовательности КАВ генома или его части.
Предпочитаемое воплощение настоящего изобретения содержит такую рекомбинантную генетическую информацию как включающую КАВ специфичную нуклеотидную последовательность, соответствующую или комплементарную нуклеотидной последовательности, показанной на фиг. 1, нуклеотидную последовательность, гомологичную ей, как минимум, на 60%, или ее часть.
Этот аспект изобретения включает нуклеиновую кислоту, выбранную из ДНК и РНК в разном возможном проявлении, т.е. как в форме свободных ДНК и РНК, так и форме ДНК и РНК, упакованных каким либо путем /т.е. в белках или в вирусных частицах/ или связанных с другим образом, например с носителем или с материалом, функционирующим как маркер. ДНК может быть как одноцепочечной, так и двухцепочечной и быть как линейной, так и кольцевой.
Характеристикой рекомбинантной генетической информации, относящейся к данному изобретению, является присутствие в ней КАВ-специфичной нуклеотидной последовательности. С практической точки зрения эта КАВ-специфичная последовательность необязательно должна охватывать весь геном КАВ, необходима только специфическая часть, которая полезна в большинстве случаев применения.
Первой предпочтительной возможностью является КАВ-специфичная нуклеотидная последовательность, соответствующая или комплементарная нуклеотидной последовательности, кодирующей КАВ белок и находящейся в КАВ геноме, или ее часть. Рекомбинантная ДНК, содержащая такую кодирующую последовательность, может быть использована, например, при обнаружении и РНК. КАВ в образцах, или, например, в процессе получения КАВ белков или их частей. Термин "ее или его часть", в принципе, означает каждую часть, которая еще может быть обозначена как КАВ-специфичная. На белковом уровне это будет эпитом для наибольшего количества применений, т. е. антигенный детерминант, распознаваемый антителами.
Другая возможность состоит в том, что рекомбинантная генетическая информация, относящаяся к изобретению, включает КАВ-специфичную нуклеотидную последовательность, соответствующую или комплементарную нуклеотидной последовательности, обладающей регуляторной функцией, находящуюся в КАВ геноме, или части ее. Одним примером является использование элементов КАВ промотор/усилитель в комбинации с последовательностями, кодирующими другие белки, не КАВ, например, для возможности экспрессии таких же КАВ белков у домашней птицы /цыплята/ и других животных, в которых регуляторные сигналы КАВ эффективны.
Как в вышеуказанном случае, так и, в основном, рекомбинантная генетическая информация, относящаяся к изобретению, может также содержать нуклеотидную последовательность, не содержащуюся в КАВ геноме. Такая "нуклеотидная последовательность, не принадлежащая КАВ геному" может быть получена, например, из нуклеотидной последовательности, полученной из прокариотического или эукариотического вектора экспрессии. Таким образом, изобретение включает возможность внедрения КАВ-специфичной последовательности в /вирусной или невирусной/ вектор, подходящий для экспрессии в эукариотических организмах или в плазмиду, подходящую для экспрессии в бактериях. Более того, также возможно, чтобы рекомбинантная генетическая информация из так называемой "нуклеотидной последовательности, полученной не из КАВ генома", относящаяся к изобретению, включала нуклеотидную последовательность, не имеющуюся в КАВ геноме, способную к регуляторной функции.
Тем не менее, "нуклеотидная последовательность, произведенная не из КАВ генома" может также содержать нуклеотидную последовательность, кодирующую белок, отличный от КАВ белка, например если КАВ регуляторные сигналы используются для экспрессии такого же КАВ белка /или части его/ в хозяине, доступном для КАВ вируса, или если рекомбинантную ДНК следует использовать для получения гибридного или слитого белка, в котором функции КАВ белка как носителя для эпитопа НЕ-КАВ белка или, наоборот, не КАВ белковые функции как носителя для эпитопа КАВ белка.
Если рекомбинантную генетическую информацию согласно изобретению используют в процессе детектирования комплементарных ДНК или РНК в образце, может быть необходимо присутствие метки. Метка при использовании здесь является маркером, подходящим для использования с ДНК или РНК, которая позволяет или помогает детектированию меченых ДНК или РНК. Известно много типов маркеров, пригодных для этой цели, таких как радиоизотопы /например, 32P/, молекулы ферментов /например, пероксидазы/, гаптены /биотин/, флюоресцентные вещества, краски, пигменты /неорганический фосфор/, и особенные маркера /например, частицы золота или селена/.
Во-вторых, изобретение касается использования рекомбинантной генетической информации, как описано выше, особенно в диагностических целях, целях иммунизации и вакцинации, или для получения КАВ или не КАВ белков.
Более точно, это относится к, например, использованию рекомбинантной генетической информации в соответствии с изобретением, в качестве КАВ-специфичной пробы или затравки в процессе детектирования КАВ-ДНК или РНК, например, в процессе ДНК/РНК слот-блоттинга, Южного блоттинга, Северного блоттинга, гибридизации in situ амплификации ДНК путем полимеразной цепной реакции SI-картирования и удлинения траймера, изобретение также охватывает диагностический набор для детектирования КАВ-ДНК или -РНК такими способами, как ДНК/РНК слот-блоттинг, Южный блоттинг, Северный блоттинг, гибридизация in situ, амплификации ДНК путем полимеразной цепной реакции SI картирования или удлинения праймеров, причем диагностический набор содержит генетическую рекомбинантную информацию, относящуюся к изобретению, как КАВ-специфичную пробу или затравку.
Дальнейшее имеет отношение к использованию рекомбинантной генетической информации согласно изобретению в качестве живой вирусной вакцины для защиты от КАВ и других патогенов, изобретение также ведет к расширению вакцинных препаратов для иммунизации против КАВ или других патогенов, которое заключается в том, что препарат содержит рекомбинантную генетическую информацию в соответствии с изобретением и дополнительно один или более носитель и добавку, пригодных для использования в живых вирусных вакцинах.
Также интересно использование рекомбинантной генетической информации согласно изобретению в качестве клонирующего вектора, т.е. использование КАВ-ДНК в качестве "эукариотической плазмиды" для птичьих систем, в которых фрагменты гена введены в полный или почти полный КАВ геном.
Использование рекомбинантной генетической информации в соответствии с изобретением в процессе получения КАВ белка, его части или отличного от КАВ белка in vitro или in vivo трансляцией также включено в изобретение. Тоже самое относится к прокариотическим или эукариотическим клеткам, содержащим рекомбинантную генетическую информацию, как описано выше, и, в частности, к таким прокариотическим или эукариотическим клеткам, которые способны экспрессировать как минимум один белок или его часть, кодируемый рекомбинантной генетической информацией, соответствующей изобретению. Эти различные возможности будут подробно описываться ниже в этом описании.
Следующий аспект изобретения касается КАВ белка или его части, полученных in vitro трансляцией рекомбинантной генетической информации, согласно изобретению содержащей нуклеотидную последовательность, кодирующую КАВ белок или его часть, также как и КАВ белок или его часть, полученные выделением из прокариотической или эукариотической клетки, содержащей рекомбинантную генетическую информацию, в соответствии с изобретением включающую нуклеотидную последовательность, кодирующую КАВ белок или его часть, и способную к экспрессии.
Белок согласно изобретению имеет широкое применение, в частности использование КАВ белка или его части для диагностики, иммунизации или вакцинации, или для продуцирования КАВ специфичных антител.
Более конкретно, изобретение включает использование КАВ белка или его части, как описано выше, в качестве реагента, связывающего КАВ-специфичные антитела в процессе иммуноанализа для детектирования КАВ-специфичных антител, например, при иммунопероксидазном окрашивании, ELISA или иммунофлуоресцентном анализе, и, соответственно, в диагностическом наборе для детектирования КАВ-специфичных антител в процессе иммуноанализа, такого как иммунопероксидазное окрашивание, ELISA или иммунофлуоресцентный анализ, в котором диагностический набор содержит КАВ белок или его часть в соответствии с изобретением как реагент для связывания с КАВ-специфичными антителами.
Изобретение также предусматривает использование КАВ белка или белковой части, как описано выше, в качестве субъединицы вакцины, осуществляющей защиту против КАВ также хорошо, как приготовление вакцины против КАВ, которая содержит КАВ белок или белковую часть в соответствии с изобретением, и желательно один или более носителей и добавок, пригодных для субъединичной вакцины.
Использование КАВ белка или его части, как описано выше, в процессе получения КАВ-специфичных поликлональных или моноклональных антител, также входит в объем изобретения. Все эти применения будут более детально описаны ниже.
Следующий аспект изобретения также относится к КАВ-специфичным антителам, продуцируемым с помощью КАВ белка или его части, как описано выше, также хорошо, как другое использование КАВ-специфичных антител, например, для целей диагностики, иммунизации или вакцинации, или для препаративных целей.
Более конкретно, интересно использование КАВ-специфичных антител в соответствии с изобретением в качестве связывающих реагентов КАВ белка в процессе иммуноанализа для детектирования КАВ белка, а также в диагностическом наборе для детектирования КАВ белка в процессе иммуноанализа, причем диагностический набор содержит КАВ-специфические антитела, в соответствии с изобретением реагенты, связывающие КАВ-белок.
Дальнейшим примером является использование КАВ-специфичных антител в соответствии с изобретением для пассивной иммунизации против КАВ инфекции, а также препаратов для пассивной иммунизации против КАВ, причем препарат содержит КАВ-специфичные антитела в соответствии с изобретением и, желательно, один или более носителей и добавок, пригодных для препаратов пассивной иммунизации.
Что касается препаративных применений, одним примером является использование КАВ-специфичных антител в соответствии с изобретением в процессе выделения и/или очистки КАВ белка.
Изобретение иллюстрируется следующими примерами.
Примеры.
Анализы низкомолекулярной ДНК, выделенной из КАВ-инфицированных клеток.
КАВ геном, выделенный из очищенного вирусного препарата, является кольцевой одноцепочечной ДНК, с длиной около 2300 баз /Todd с сотр., 1990/. Мы ожидали, что КАВ-инфицированные клетки содержат дополнительно циклическую двухцепочечную КАВ-ДНК. Двухцепочечная ДНК была разрезана рестриктазами и, следовательно, могла быть прямо клонирована в отличие от одноцепочечной ДНК. С этой точки зрения было проверено, содержит ли низкомолекулярная фракция КАВ-инфицированных клеток ДНК продукт, который отсутствует в неинфицированных клетках.
Низкомолекулярная ДНК была выделена из КАВ-инфицированных MDCC-MSBI и 1104-Х5 клеток и из неинфицированных 1104-Х5 клеток. ДНК была фракционирована на агарозо/этидий бромидном геле. Очень слабая группа ДНК, имеющая длину около 3 килобаз, обнаруживалась в геле. Этот специфичный ДНК-продукт отсутствовал в ДНК, выделенной из неинфицированных клеток.
В последующем эксперименте было показано, что специфическая ДНК присутствует только в КАВ-инфицированных клетках. ДНК, выделенная из инфицированных клеток, была разделена по размерам на агарозном геле. Была выделена ДНК, имеющая длину 2,7 - 3,5 кб. Эта ДНК фракция должна содержать специфическую вирусную ДНК дополнительно к другим клеточным ДНК. Выделенная ДНК была радиоактивно мечена и гибридизована в Саузерн-блоттинге с низкомолекулярной дНК из КАВ инфицированных клеток и из неинфицированных клеток. 3 кб - продукта ДНК гибридизируется в блоттинге с КАВ-инфицированными клетками и отсутствует в блоттинге с ДНК из неинфицированных клеток.
Расстояние 3 кб было определено с помощью ДНК маркеров, содержащих двухцепочечные линейные молекулы ДНК. Поведение циклической двухцепочечной молекулы ДНК в агарозном геле отлично от поведения линейных ДНК фрагментов. ДНК в 3 кб из КАВ-инфицированных клеток должна иметь линейную форму, что в действительности составляет 2,3 кб.
Если циклическая двухцепочечная ДНК переваривается рестриктазами, режущими только в одном месте ДНК молекулы, то должна образовываться линейная молекула ДНК, имеющая размер 2,3 кб.
То, что это предположение верно, было продемонстрировано раздельно инкубацией низкомолекулярной ДНК, выделенной из КАВ инфицированных 1104-Х5 клеток с шестью различными энзимами рестрикции /Bam HI, EcoRI, Hind III, Kpn I, Rst I и Xba I./ Низкомолекулярная ДНК, выделенная из КАВ-инфицированных 1104-Х5 клеток, и вышеуказанные фрагменты рестрикции были гибридизованы с радиоактивно меченой ДНК, как описано выше. Это показало, что обработка ферментами рестрикции Bam I, EcoRI, Rst I и Xba I приводит к образованию молекулы ДНК, имеющей размер 2,3 кб. ДНК из неинфицированных клеток, инкубированная с Bam HI, не содержала такой ДНК продукт. Фермент рестрикции Hind III разрезает ДНК в двух местах, тогда как Kpn I не разрезает.
Из вышерассмотренных экспериментов можно заключить, что в низкомолекулярной ДНК из КАВ-инфицированных клеток находится циклическая молекула ДНК в 2,3 кб, которая отсутствует в неинфицированных клетках, и что она и есть геном КАВ в форме циклической двухцепочечной молекулы ДНК.
Клонирование и субклонирование двухцепочечной КАВ-ДНК в бактериальном векторе.
Низкомолекулярная ДНК КАВ-инфицированных 1104-5Х клеток была отдельно инкубирована с Bam I, EcoRI, Rst I и Xba I. ДНК разделялась на агарозном геле с низкой точкой плавления. Из всех четырех препаратов была выделена молекула ДНК в 2,3 кб. Вектор клонирования pIC-20H был отдельно переварен теми же четырьмя ферментами рестрикции, которые режут низкомолекулярную ДНК. Линейный вектор был обработан кишечной щелочной фосфатазой теленка. Каждый 2,3 кб фрагмент ДНК лигирован к соответствующему рестрикционному сайту pIC-20H. Продукты лигирования были перенесены в E coli штамм HB101. Все 4 клона дали плазмиды, содержащие вставленную ДНК, имеющую размер около 2,3 кб. Дальнейший рестрикционный анализ показал, что, как минимум, 7 плазмид содержат такой же ДНК-фрагмент. Место интеграции вектора тем не менее отличается, так как используются различные ферменты для разрезания циклической молекулы. При использовании ферментов Bam I, EcoR I, Rst I и Xba I была составлена рестрикционная карта всех четырех КАВ ДНК клонов.
Четыре "различные" КАВ ДНК-плазмиды были радиоактивно мечены и гибридизованы путем Саузерн-блоттинга с BamH I - обработанной ДНК, выделенной из КАВ инфицированных или KAB неинфицированных клеток. Все тестируемые клоны гибридизовались только с 2,3 кб молекулой ДНК, присутствующей в ДНК КАВ-инфицированных клетках.
Биологическая активность двух KAB ДНК клонов.
Два KAB клона pIC-20H/КАВ-Eco RI и pIC-20H/KAB-PstI были обработаны ферментами рестрикции так, что КАВ ДНК была полностью вырезана из вектора. Линейные КАВ ДНК-молекулы были обработаны T4-ДНК лигазой. Линейные ДНК были, таким образом, были превращены в кольцевые. Теперь "клонированные" КАВ ДНК имели двухцепочечную кольцевую форму, также проявляемую КАВ ДНК дикого типа в инфицированных клетках. MDCC-MSBI и 1104-X5 клетки были трансфицированы "клонированными" кольцевыми КАВ ДК. Для клона pIC-20H/КАВ-EcoRI очень четкий цитопатогенный эффект (ЦПЭ) был найден для клеток обоих видов. Клон pIC-20H/КАВ-PstI проявлял четкий ИПЭ в клетках MDCC-MSBI и менее четкий ЦПЭ в клетках 1104-X5. Тем не менее супернатанты трансфицированных pIC-20H/КАВ-PstI 1105-X5 клеток проявляли ясный ЦПЭ в MDCC-MSBI клетках. Трансфекция ДНК, выделенной из КАВ-инфицированных клеток, также ведет к четкому ЦПЭ в МДСС-MSBI клетках, хотя в 1104-X5 клетках наблюдали менее четкий ЦПЭ. ЦПЭ не наблюдали после трансфекции MDCC-MSBI или 1104-X5 клеток ДНК pIC-20H вектора.
Саузерн анализ показал, что в клеточных лизатах MDCC-MSBI и 1104-X5 клеток, инфицированных вирусом, полученным клонированием КАВ ДНК, присутствовала КАВ ДНК.
Тест на нейтрализацию с MDCC-MSBI клетками показал, что ЦПЭ, вызванный клонированными ДНК в трансфицированных клетках, был результатом КАВ инфекции. Нейтрализующие антитела, направленные против КАВ, предотвращает ЦПЭ MDCC-MSBI клеток, инфицированных КАВ-потомками трансфицированных клеток.
Однодневных птенцов инъецировали внутримышечно супернатантом трансфицированных клеток. У птенцов супернатанты вызывали те же клинические симптомы, что и КАВ дикого типа: замедление роста, изменение костного мозга и уменьшение содержания гематокрита /анемия/, атрофия тимуса /истощение специфической популяции T клеток/ и смертность. Супернатанты клеток, трансфицированных вектором, не приводят к симптомам заболевания у контрольных птенцов.
Секвенирование двухцепочечной ДНК КАВ генома.
Полная двухцепочечная ДНК КАВ генома была полностью секвенирована методом Сангера /Sanger и сотр., 1977/ и методом Максама Гилберта. С помощью праймеров секвенирования M13 обратного секвенирования M13 последовательность ДНК размером 2300 п.о. была определена для pIC-20H/КАВ (BamHI, EcoRI; PstI; XbaI) клонов. Затем КАВ геном был субклонирован. ДНК последовательность для различных 5 субклонов ДНК КАВ генома была определена методом Сангера с помощью M13 затравки и/или методом Максама-Гилберта. Таким образом, последовательность ДНК обеих цепей КАВ генома была определена.
Длина КАВ (двухцепочечный) ДНК - 2319 по. Первое основание Eco RI сайта кольцевого КАВ генома пронумерован +1. Последовательность ДНК цепи, содержащая наибольшую открытую рамку считывания показана на фиг. 1 и названа (+) цепью. Состав оснований в этой цепи: 25,5% аденина; 28,7% цитозина; 27,7% гуанина; 18,1% тимина. Компьютерное изучение возможной гомологии КАВ генома с уже известными вирусными последовательностями показало, что эта ДНК прежде не была описана и не является частью ранее описанных групп вирусов. Первоначальная гипотеза о том, что КАВ является первовирусом просуществовала недолго, до тех пор пока последовательность и форма ДНК КАВ генома (кольцевая) не были изучены.
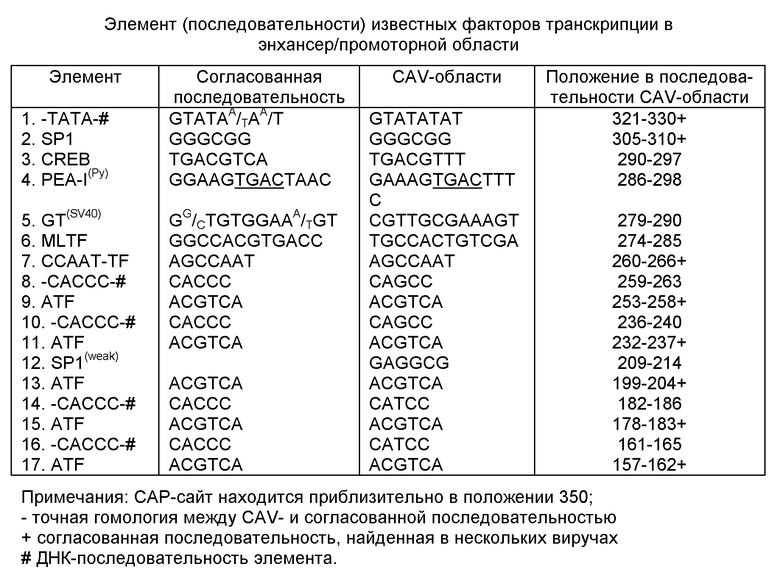
С помощью компьютерного изучения организация КАВ генома была охарактеризована. Открытые рамки считывания, элементы промотора/усилителя, сигнал и сайт полиаденилирования и "начало репликации" были предсказаны. Фиг. 2 показывает предполагаемые открытые рамки считывания длиной 300 б для обеих ДНК цепей КАВ. Фиг. 2а показывает открыто не рамки считывания, начиная с кодона АТГ. АТГ кодон является наиболее часто используемым инициирующим кодоном для белков. Замечено, что одна из двух ДНК цепей кодирует 3 белка, имеющих длину в 449 аминокислот (51,6 кДа), 216 аминокислот (24 кДа), и в 121 аминокислоту (13,3). Todd с сотр. (1990) обнаружил 50 кД белок в очищенном КАВ. Если используются все открытые рамки считывания, то около 80% вирусного генома транслируется. Некоторые области даже дублируются. Вполне возможно, что 3 открытые рамки считывания транслировались с I РНК. Предполагаемый старт РНК молекулы находится в положении 354 и поли(A) добавление - в позиции 2317. Только поли(A) сигнал находится в позиции 2287 + цепи.
Мало вероятно, чтобы открытые рамки считывания были использованы на другой ДНК цепи, так как эта цепь не содержит некоторые существенные регуляторные последовательности. Фиг. 2Б и 2С показывают открытые рамки считывания, использующие соответственно ЦТГ и ГТГ стартовые кодоны. Тем не менее, описано только несколько белков, для которых эти стартовые кодоны действительно используются (Напп с сотр., 1988).
Компьютерные исследования сходства между отдельным КАВ белками и уже известными белками обнаружили только ограниченные области гомологии по последовательностям, присутствующим в программах. Соответственно, трудно предположить, к какому типу белков относятся КАВ белки. Относительно высокие показатели были получены для белков вирусного капсида, ДНК связывающих белков и коагулирующих кровь белков. Результаты здесь не приводятся.
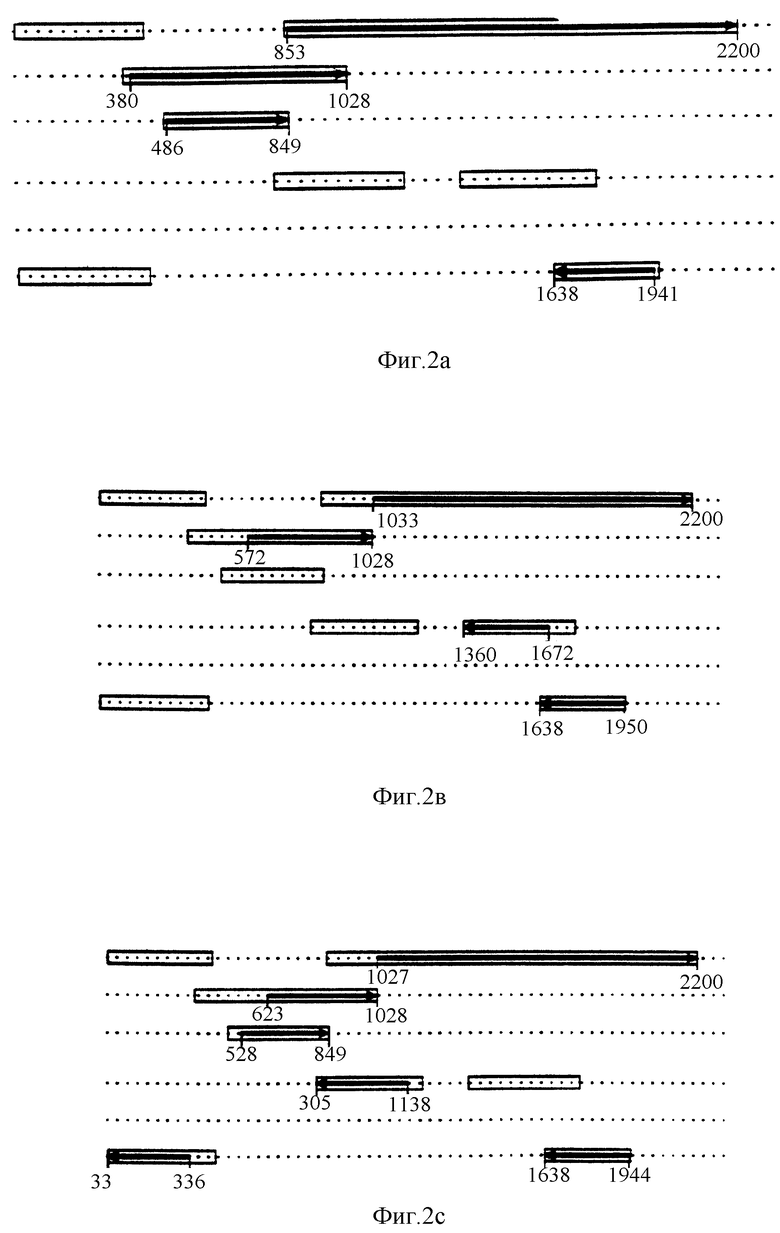
Экспрессия белков регулировалась промотор/усилитель элементами (Джонс, 1990). Эукариотический промотор имеет наиболее правое положение до старта транскрипции. КАВ последовательность содержит до сap-сайта основные элементы: TATA бокс, SPI бокс и CAAT бокс. Последовательность этих боксов прекрасно соответствует описанным для большинства эукариотических промоторов.
Вокруг позиции 285 могут быть сайты связывания для четырех различных факторов транскрипции: CREB, MLTF, CT и PEA-I.
Эукаритический ген также содержит элементы Энханера, детерминирующие длину эукариотического промотора. Предполагаемые энханирные элементы локализованы между позициями 144 и 260 и представляют собой пять прямых повторов в 21 нуклеотид каждый. Все повторы имеют 19 идентичных нуклеотидов. Только последние 2 нуклеотида отличны. Повтор 1 идентичен 2-му, 3-й равен 5-му. Повторы 1, 2 и 3 локализованы рядом друг с другом, также как 4 и 5. Между 3 и 4 существует "щель" в 12 нуклеотидов. Компьютерное изучение показало, что описанные энханиры (эукариотические), не содержат все последовательности, найденные для вероятных энханирных элементов КАВ. Все прямые повторы содержат АТФ элемент, который может быть включен в усиление транскрипции КАВ РНК. Прямые повторы содержат дважды последовательность ЦАТЦД и дважды последовательность ЦАГЦЦ. Последняя последовательность частично перекрывается с ЦААТ боксом. Эти четыре последовательности имеют только 1 неправильное спаривание с ЦАЦЦЦ боксом, описанным для β-глобина (таблица).
Фиг. 3 показывает, что приблизительно между позициями 55 и 135 и между позициями 2180 и 2270 в x-цепи ДНК присутствуют очень большие шпилечные структуры в (одноцепочечной) форме ДНК КАВ. Шпилечные структуры НК могут быть включены в репликацию КАВ ДНК. Шпильки между 2180 и 2270 могут присутствовать не только в КАВ ДНК, но также и в КАВ РНК и, вероятно, играют роль при стабилизации КАВ РНК.
Различные формы ДНК КАВ в инфицированных клетках.
Четыре различные КАВ ДНК - молекулы видны, обнаруживаются в Саузерн-блоттинге препарата ДНК КАВ инфицированных клеток. ДНК была гибридизована с радиоактивно меченой ДНК клона pIC-20H/КАВ-EcoRI. КАВ ДНК молекулы с точки зрения их измеренных длин и форм в неденатурированном агарозном геле и чувствительности к SI нуклеазе являются соответственно двухцепочечными открытыми кольцами (3 кб), суперскрученной двухцепочечной ДНК (2 кб), кольцевой одноцепочечной ДНК (0,8 кб) и одноцепочечной линейной ДНК (1,5 кб). Иногда линейная двухцепочечная ДНК КАВ обнаруживается (2,3 кб). Todd и сорт. (1990) измерили длину 0,8 кб для кольцевой одноцепочечной ДНК, выделенной из КАВ на основе электрофоретической подвижности в неденатурированном агарозном геле Детекция КАВ ДНК в вирусных препаратах.
Полная ДНК была выделена из КАВ и очищена в соответствии с методом, описанным Von Bulow (1989). Препарат ДНК был проанализирован анализом по Саузерну с меченой КАВ ДНК пробой, содержащей полную клонированную КАВ последовательность. ДН, выделенная из очищенного КАВ содержит молекулу ДНК размером 0,8 кб, определенную в неденатурированном агарозном геле. В Саузерн-анализе ДНК, выделенной из очищенного КАВ, с олигонуклеотидами, полученными из клонированной КАВ ДНК-последовательности, как пробы, было продемонстрировано, что вирус содержит отрицательную ДНК цепь. Исходя из этого можно заключить, что одноцепочечная ДНК КАВ в капсиде является отрицательной цепью.
Саузерн-анализ ДНК из КАВ природных изолятов.
Препараты ДНК были приготовлены из КАВ культуры, полученной из цыплят из выводков, в которых болезнь Марека протекала по возрастающей. ДНК препараты из КАВ культур, полученные в 12 компаниях в Нидерландах, были собраны неселективно из коллекции 60 образцов. Только в одной компании проявилась более высокая смертность благодаря болезни Марека. Более того, КАВ-изолят дня выделен из цесорки. КАВ культуры, исследуемые нами, были получены, главным образом, после того, как атрофия тимуса была установлена при исследовании.
Для целей изучения степени сходства клонированной КАВ ДНК (pIC-20H/КАВ-EcoRI) и ДНК различных природных КАВ изолятов MDCC-MSBI клетки были инфицированы выделенными недаммами КАВ. Был проведен Саузерн-анализ. Все ДНК препараты содержали ДНК молекулы, которые специфично гибридизовались с 32P-меченой клонированной КАВ ДНК. ДНК молекулы различных CAB природных изолятов имеют длины, соответствующие размерам клонированного КАВ, и являются двухцепочечными или одноцепочечными. Саузерн-блоттинг, прямо проведенный с тканевыми образцами-КАВ-инфицированных цыплят, обнаружил содержание молекул ДНК, гибридизованных с меченым pIC-20H/КАВ-EcoRI.
Рестрикционный анализ ДНК из КАВ природных изолятов.
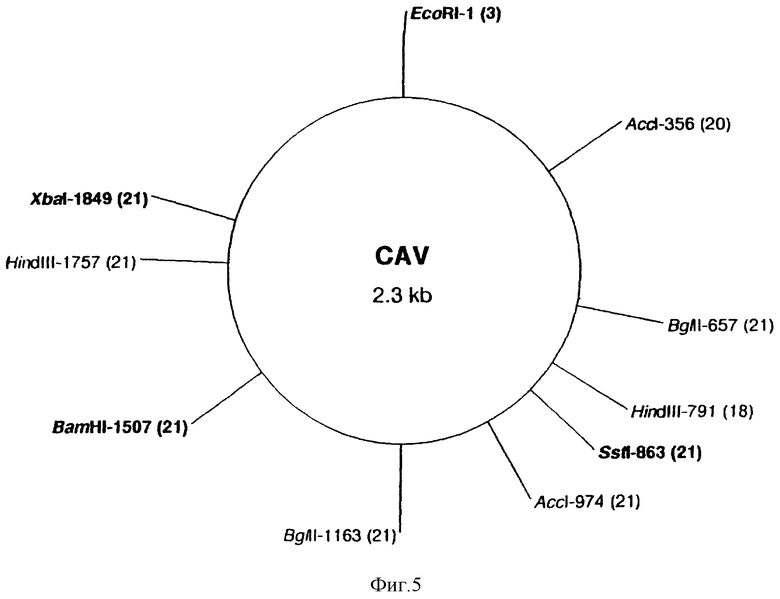
Родство ДНК из различных КАВ природных изолятов с клонированным КАВ геномом было затем проверено с помощью рестрикционного анализа. Препараты ДНК КАВ-изолятов и клонированная ДНК были по отдельности разрезаны семью ферментами рестрикции. Для всех ДНК одинаково проводили разрезание ферментами BamHI, BglI, SstI и XbaI. ДНК из большинства природных изолятов содержала два AccI сайта и/или два HindIII сайта, тогда как ДНК только нескольких изолятов содержала EcoRI сайт. Фиг. 5 суммирует карты ферментов рестрикции клонированного КАВ и различных природных изолятов. Приходящееся на сайте рестрикционного фермента количество природных изолятов, содержащих этот сайт, указано в скобках.
Полимерная цепная реакция (ПЦР) ДНК КАВ природных изолятов.
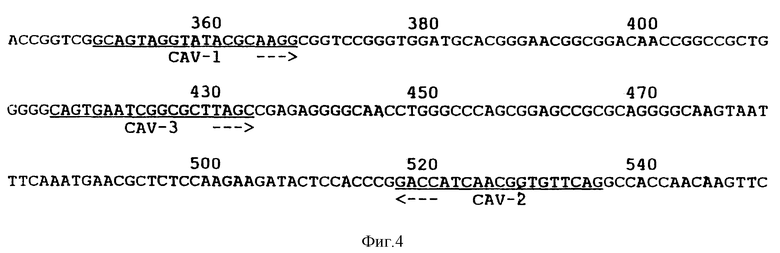
Олигонуклеотиды КАВ-1 и КАВ-2 (фиг. 4), полученные из клонированной КАВ ДНК-последовательности, были синтезированы. Использование этих олигонуклеотидов в ПРЦ привело к специфичному детектированию ДНК КАВ в природных изолятах. ДНК, выделенная из MDCC-MSBI клеток, инфицированных различными КАВ изолятами, и ДНК, выделенная из неинфицированных клеток, была амплифицирована. После амплификации ДНК была электрофоретически разделена по размерам на агароза/этидий бромидном геле. Амплифицированная нить 186 п.о. /т.е. теоретически ожидаемое значение/ была визуализована во всех ДНК-образцах клеток, инфицированных различными КАВ-изолятами. Эта специфическая нить не присутствует после амплификации ДНК, выделенной из неинфицированных клеток. Амплифицированные ДНК-нити всех природных изолятов показали одинаковую скорость миграции в агарозном геле. Этот результат подразумевает то, что большие вставки или делеции не происходят в этой части генома различных КАВ полевых изолятов. Саузерн-анализ с 32P-меченым олигонуклеотидом КАВ-3 /фиг.3/ показал, что амплифицированная 186 п.о. ДНК является КАВ-специфичной и что нет другой ДНК-нити, гибридизующейся с КАВ-3 пробой.
Была проверена чувствительность детекции КАВ ПЦР. ДНК была выделена из КАВ-инфицированных клеток, разбавлена, амплифицирована и анализирована на агароза/этидиум бромистом геле. После амплификации образцов, содержащих ДНК в количестве, соответствующем количеству ДНК в примерно 100 КАВ-инфицированных клеток, был обнаружен КАВ-специфический ДНК-фрагмент 186 п.о. Тем не менее, если амплифицированная ДНК подвергается Саузерн-анализу 32P-меченой КАВ-3 ДНК, в количестве ДНК, соответствующем количеству ДНК из 1 клетки, была найдена четко видимая КАВ-специфичная ДНК-нить. КАВ ПЦР является высоко чувствительным детектирующим методом, который до сих пор специфичен для проверки КАВ изолятов.
Дот-блот анализ ДНК КАВ природных изолятов с дигоксигенин-мечеными КАВ ДНК-пробами.
Дополнительно к ПЦР был разработан анализ по детектированию ДНК из КАВ природных изолятов. Этот тест основан на использовании не радиоактивных проб. КАВ ДНК-вставка клона pIC-20H/КАВ-EcoRI была помечена II-dVTP-дигоксигенином. Препараты ДНК из MDCC-MSBI клеток, отдельно инфицированных различными КАВ-изолятами, были нанесены на фильтры и проанализированы по их способности гибридизоваться с дигоксигенин-меченой ДНК-пробой. Препараты ДНК из MDCC-MSBI клеток, инфицированных различными КАВ-изолятами, гибридизуются с дигоксигенин-меченой ДНК-пробой, тогда как ДНК из неинфицированных клеточных культур не гибридизуется. Это тест, использующий не радиоактивно-меченую КАВ ДНК-пробу, следовательно, подходит для определения ДНК КАВ природных изолятов.
Применения
ДНК.
КАВ последовательности, например ДНК pIC-20H/КАВ-EcoRI плазмиды или ее части, могут быть использованы для демонстрации КАВ ДНК и/или РНК в препаратах для последующего исследования в научных и диагностических целях. ДНК может быть мечена радиоактивно или другим путем, например биотином/дигоксигенином. С помощью ДНК/РНК слот-блоттинга, Саузерн/Нозерн анализов и гибридизации in virto присутствие КАВ нуклеиновых кислот было установлено. Части КАВ последовательностей при использовании здесь также являются ДНК олигомерами.
Олигомеры, происходящие из КАВ последовательностей клона pIC-20H/КАВ-EcoRI, можно использовать в "Полимеразной Реакции Цепи" для выявления очень низких концентраций КАВ ДНК/РНК. ПЦР является высоко чувствительным методом, часто используемым для детекции вирусов.
Диагностические наборы для проведения вышерассмотренных исследований возможны в практике.
Для научных целей применение методик, подобных картированию и удлинению праймера с КАВ ДНК фрагментами, очень важно. С помощью этих двух методов КАВ РНК может быть количественно определена и далее охарактеризована.
Олигомеры в антисмысловой конфигурации могут быть использованы для изучения функций гена. Они могут также использоваться в качестве модели для изучения новых способов ингибирования вирусной репликации.
КАВ ДНК может быть использована как носитель в трансфекции для малых фрагментов гена, особенно, если патогенные свойства удалены делецией в КАВ геноме.
КАВ олигомеры в антисмысловой конфигурации могут быть экспрессированы в составе вирусных векторов, что дает возможность изучать КАВ репликацию или другие функции гена in vivo или in vitro.
РНК.
КАВ ДНК фрагменты, клонированные в Sp6/T7 векторах, приводят к образованию РНК продуктов. КАВ ДНК, полученные транскрипцией in vitro, можно использовать для синтеза КАВ белков in virto, in vivo. Таким образом, РНК молекулы, например, в экстракте пшеничных зародышей могут быть транслированы в белки /трансляция in vitro/. КАВ белки, полученные трансляцией in vitro, могут быть затем использованы, например, для мечения антител, направленных против КАВ в сыворотке цыплят /смотри ниже/. КАВ РНК молекулы могут быть перенесены в клетки путем микроинъекции, чтобы транслироваться там в белки. Таким образом, эффекты КАВ белков могут быть изучены на клеточном уровне. Также могут быть проанализированы белок/белок и/или белок/ДНК взаимодействия.
КАВ ДНК также могут быть использованы как пробы для определения нуклеиновых кислот в препаратах. Анализы могут быть проведены с помощью слот-блоттинга, Саузерн-, Нозерн- и гибридизационного анализа in situ. Эти методы могут быть использованы для развития диагностических тестов для КАВ.
Белки.
Все КАВ белки могут быть экспрессированы в прокариотических или с эукариотических системах. Это требует, чтобы КАВ открытые с рамки считывания были клонированы в подходящем векторе экспрессии. Для бактериальной системы существует вектор экспрессии, основанный на T7 промоторе, подходящем для экспрессии КАВ открытых рамок считывания. Бакуловирусная система, дрожжи и CHO-dnfr система являются возможными эукариотическими экспрессионными системами. Вирусные векторы, такие как ретровирусные векторы, также могут быть использованы.
КАВ белки или эпитопы, локализованные в них, могут быть использованы для обнаружения антител против КАВ. Таким образом, КАВ-инфицированные цыплята могут быть обнаружены. КАВ белки или эпитопы, локализованные в них, могут быть использованы в иммуноанализах, таких как иммунопероксидазный, ELISA и иммунофлуоресцентный анализы.
КАВ белки или эпитопы, локализованные в них, могут использоваться для обеспечения гуморального и/или клеточно связанного иммунитета против КАВ. КАВ белки, полученные экспрессией в эукариотических или прокариотических системах вектор/хозяин, могут использоваться для получения субъединичных вакцин.
С помощью КАВ белков или их эпитопов можно получить КАВ специфичные антитела, что дает возможность обнаружить КАВ-белки в препаратах КАВ-инфицированных птенцов /смотри ниже/.
Антитела.
В ряде инфекционных тестов на молодых птенцах можно было заключить, что материнские антитела могут обеспечивать эффективную пассивную защиту против КАВ инфекции. Материнские антитела были перенесены в молодых птенцов естественным путем, а также через инъекцию только родившимся птенцам яичных желтковых экстрактов, содержащих КАВ антитела. Пассивная защита от КАВ инфекции также достигается путем инъекции экстрактов яичных желтков из яиц несушек, которые были инфицированы прямо перед периодом вынашивания.
Вакцинация несушек КАВ белками, экспрессированными в одной или более экспрессионных системах, приводит к образованию материнских антител. Молодые птенцы этих несушек будут защищены от КАВ инфекции.
Диагностические тесты могут быть разработаны на основе антител против КАВ. Как поликлональные, так и моноклональные антитела могут быть использованы. С помощью КАВ-специфичных антител препараты могут быть проверены на присутствие КАВ белков.
Вышерассмотренные возможности применения КАВ антител в равной мере относятся как к антителам, полученным способами, описанными здесь, так и природным КАВ антителам.
Живые вирусные вакцины.
Обеспечение иммунной системы вирусными белками с помощью вектора живого вируса, вероятно, приводит к лучшему иммунному ответу, чем субъединичная вакцина. Одна или более КАВ открытых рамок считывания /полностью или частично/ могут быть клонированы в векторах живого вируса. Для домашней птицы могут только использоваться живые вирусные векторы, что само по себе показывает хорошую репликацию в системе птиц. Подходящими в качестве векторов для применения в цыплятах являются, например, вирус оспы птиц, векторы ретровирусов, векторы герпес вируса, вирус герпеса птиц серотипа 1,2 и 3, и вирус инфекционного ларинготрахеита и, возможно, также аденовирусы, такие как CELO. Иммунизация вышеуказанными векторами живых вирусов защищает против КАВ и вирусов переносчиков.
С помощью одной и более делеций в КАВ геноме можно разработать вакцины, которые иммунизируют молодых птенцов от КАВ инфекции. Делетированы должны быть патогенные детерминанты КАВ инфекции, но реплекативные и, следовательно, иммунизирующие свойства должны быть сохранены.
КАВ-геном может быть сам по себе пригодным в качестве животного вирусного вектора для экспрессии антигенов других вирусов. Это требует, чтобы КАВ геном был изменен так, чтобы дополнительно или вместо КАВ белков "посторонние" вирусные белки экспрессировались. КАВ векторы, следовательно, должны быть сконструированы так, чтобы имелась защита против "посторонних" вирусов исключительно или также против КАВ в зависимости от экспрессии вирусных белков с помощью рекомбинантного вектора в вакцинированном животном.
КАВ вакцины, полученные как субъединичная вакцина, делеционная вакцина или фрагмент гена в другом вирусном векторе, следует главным образом использовать для вакцинации несушек. Тем не менее вакцинация птенцов в ином возрасте, например, в комбинации с вакцинацией против болезни Марека также входит в возможное использование изобретения.
Элементы энхансер/промотор.
Элементы КАВ промотора и энхансера могут быть клонированы в ДНК векторы. Под контролем КАВ промотора/энхансера КАВ белки или "посторонние" белки могут быть экспрессированы как в клетках цыплят, так и в других клетках.
Возможно, КАВ промотор функционирует в клетках костного мозга. В качестве модельной системы для генной терапии "посторонние" белки были экспрессированы in vitro в клетках костного мозга с помощью КАВ элементов промотор/энхансер, обычно в комбинации с ретровирусными векторами. Генетически модифицированные клетки костного мозга могут затем трансплантироваться в костный мозг, в данном случае, цыпленка. Для очень маленьких генных фрагментов КАВ геном также подходит для использования в качестве вектора.
КАВ элементы энхансер/промотор могут быть также активны в других организмах. Указанные элементы могут быть также использованы в, например, мышиной системе в качестве модели для генной терапии.
Возможность использования полного или по существу полного генома КАВ в качестве вектора клонирования, т. е. как разной видности эукариотической плазмиды для систем птиц, реальна с точки зрения выявленной структуры КАВ генома.
Депонирование КАВ клона pIC-20H/КАВ-EcoRI
HB101 клетки, трансформированные плазмидой pIC-20H/КАВ-EcoRI были депонированы в Центральном бюро культур в Баарне, Нидерланды, 7 сентября 1990 года, под номером CBS 361.90.
Описание чертежей.
Фиг. 1 показывает нуклеотидную последовательность клонированной КАВ ДНК. Полная длина - 2319 оснований, первый гуанин EcoRI - сайта обозначен под N 1.
Фиг. 2 показывает предполагаемые открытые рамки считывания, имеющие длину более 300 оснований для обеих цепей ДНК. Предполагаемые открытые рамки считывания стартовых кодонов АТГ, ЦТГ и ГТГ показаны в фиг. 2A, 2B, 2C соответственно.
Фиг. 3 показывает предполагаемые шмилечные структуры КАВ генома, содержащие одноцепочечную ДНК.
Фиг. 4 показывает олигонуклеотиды, используемые в ПЦР. Также показана ДНК последовательность и позиции олигонуклеотидов в КАВ геноме. Положения нуклеотидов в КАВ геноме соответствуют положениям нуклеотидов, как представлено на фиг. 1.
Фиг. 5 показывает карту рестрикции клонированной КАВ ДНК. В скобках указано количество КАВ изолятов, содержащих соответствующий рестрикционый сайт в геноме.
Материалы и методы.
Клеточные культуры и вирусы.
КАВ изоляты были культурированы в трансформированных лимфобластоидных клеточных линиях из опухолей цыплят, индуцированных лейкозным вирусом птиц подгруппы A (1104-X-5), или вирусом заболевания болезни Марека (MDCC-MSBI). Клеточные культуры были инфицированы около 0,1-ITCI 50 на клетку. Клетки собирали после двух дней. Клетки были инфицированы вирусом клонированной КАВ ДНК, или природными изолятами. КАВ-Сих-1, впервые выделенный в Германии из выводка цыплят, пострадавших от болезни Марека (Von Vulow с сотр., 1983, 1985), был предоставлен доктором M.C.McNultu, Ветеринарная Исследовательская Лаборатория, Белфаст, Северная Ирландия. В январе 1988 года доктор Дж.П. Розенбергер, Университет of Delaware, Newark, США, передал нам два образца крови для определения вирулентности штамма T-1704, вызывающего болезнь Марека и его производного, MDV-Del-S, которое впервые пассировано в цыплятах. Мы получили КАВ-T-1704 и КАВ-DEL-S изоляты из SPF-цыплят, инфицированных MDV-штаммом T-1704 и его производным MDV-DEL-S. Голландские KAB изоляты были неселективно выбраны из 60 серий, которые все были культивированы в MDCC-MSBI клеточных культурах. Полевой природный материал был поставлен J.C. Vander Wijnaard, Gerondheidsdienst Brabant at Boxtrel и J.Naber, главным образом, потому что атрофия тимуса была установлена в течение вскрытия. KAB изоляты, полученные из наших собственных P-выводков, были добавлены к сериям. Выделение общей ДНК.
Вирусные и печеночные препараты были ресуспендированы в 20 мМ Трис HCl с pH 7,5, 2 мМ ЭДТА, 0,2% СДС, 0,6 мг/мл Протеинкиназы-K, и инкубированы в течение 1 часа при 37oC. Препараты были экстрагированы системой фенол/хлороформ/изоамиловый спирт /25:24:1/, и ДНК осаждалась этанолом. Осадок ДНК ресуспендировали в 100 мкл 10 мМ Трис HCl с pH 7,5; 1 мМ ЭДТА.
Экстракция и анализ низкомолекулярной ДНК.
Низкомолекулярная ДНК была выделена из KAB-инфицированных 1104-X5 и MDCC-MSBI клеток и неинфицированных 1104-X5 клеток в соответствии с методом, описанным Hirt /1967/. ДНК разделяли на агарозных гелях и после окрашивания с этидиум бромидом прямо анализировали с помощью УФ света или блоттировали на фильтре в соответствии с методом, описанным Southern /1982/. Блоты были гибридизованы с праймированной 32P-меченой ДНК, выделенной из низкомолекулярной ДНК КАВ-инфицированных 1104-X5 клеток, имеющей длину 2,7-3,5 кб.
Клонирование KAB ДНК.
Полный KAB ДНК геном был клонирован в бактериальном векторе pIC-20H. Части КАВ ДНК генома были клонированы в векторе pIC-19. Все этапы клонирования плазмид ДНК были проведены в основном в соответствии с методами, описанными Maniatis с сотр. /1982/.
Анализ последовательности КАВ ДНК.
КАВ ДНК плазмиды были очищены в CsCl-градиенте и путем Сефакрил-S500 /фармация/ хроматографии. Двухцепочечная ДНК была секвенирована с помощью T7 ДНК полимеразы /Фармация/, или с помощью Tag ДНК полимеразы /Промега/. Оба метода проводились согласно инструкциям, данным фирмами Фармация и Промега. Олигонуклеотиды были обработаны T4 нуклеотид киназой ("Фармации"). "Жесткие остановки" были секвенированы согласно методу, описанному Максамом и Гилбертом /1977/.
Образование кольцевой формы клонированного КАВ ДНК генома.
10 мкг плазмидной ДНК клонов, содержащих полный ДНК геном, были переварены ферментом рестрикции так, что только КАВ ДНК вставка была отделена от вектора. T4-ДНК лигазная обработка 2,3 кб линейной молекулы КАВ ДНК привела к образованию циклической двухцепочечной КАВ ДНК. Лигированные продукты анализировали на 0,8% агарозном геле.
ДЕАЕ - декстрановая трансфекция.
Для трансфекции 2 мкг 1104-X5 и MDCC-MSBI клеток лигированной КАВ ДНК суспендировали дважды в 25 мкл Милли-Q воды и смешивали с 260 мкл TBS буфера. К ДНК смеси добавляли 15 мкг 10 мг/мл ДЕАЕ декстрана и смесь инкубировали 30 минут при комнатной температуре.
1104-X5 клетки. 50 мм чашка тканевой культуры с 1-2•106 1104-X5 клетками на чашку была промыта дважды TBS буфером. TBS буфер полностью удаляли из клеточного монослоя и добавляли 300 мкл разбавленного раствора ДНК/ДЕАЕ-декстран. Клетки были инкубированы 30 минут при комнатной температуре. Смесь ДЕАЕ-декстран/ДНК была перенесена в 2 мл 25%-ного раствора DMSO/TBS и клеточный монослой инкубировался 2 минуты при комнатной температуре. Клетки промывали дважды ТБС буфером, а затем добавили субстрат тканевой культуры /RPM11640 или E-MEM/. Клетки инкубировали при 37oC и 5% CO2.
MDCC-MSBI клетки. Около 2•106 MDCC-MSBI клеток центрифугировали при 1500 об/мин в настольной центрифуге. Среда была замещена 5 мл ТБС буфера, и клетки осторожно ресуспендировали. Стадию промывки повторяли. Удаляли весь ТБС буфер, осадок клеток осторожно ресуспендировали в 300 мкл смеси ДЕАЕ-декстран/ДНК и инкубировали при комнатной температуре 30 минут. Добавляли 0,5 мл 25% раствора DMSO/TBS и суспензию инкубировали 3 минуты при комнатной температуре. Добавляли 5 мл ТБС-буфера и центрифугировали при 1500 об/мин в настольной центрифуге. Супернатант удаляли и добавляли 50 мл субстрата тканевой культуры. Клетки ресуспендировали и центрифугировали. Клетки были помещены в 5 мл среды для тканевой культуры и инкубированы при 37oC и 5% CO2. С целью контроля были использованы 2 мкг pIC-20H плазмиды для трансфекции.
Тест на нейтрализацию
MDCC-MSBI клетки инфицировали супернатантом MDCC-MSBI, а 1104-X5 клетки были трансфицированы клоном "КАВ ДНК". Было инфицировано около 2•104 клеток. Содержание вируса в этом инокуляте не было точно известно. До половины инфицированных клеточных культур к среде была добавлена поликлональная разбавленная 1:100 сыворотка с нейтрализующей активностью против КАВ.
С целью контроля были взяты серии "ячеек" с КАВ-инфицированными клетками MSBI, причем к субстрату не была добавлена антисыворотка, направленная против КАВ.
КАВ инфекция однодневных птенцов.
Супернатанты КАВ ДНК и контрольная ДНК, трансфицированная MDCC-MSBI и 1104-X5 клетками, были инъецированы внутримышечно однодневным птенцам. Спустя 6 дней после инфекции были проведены аутопсия 5 птенцов на группу, после чего значение гематокрита и общий вес тела были измерены в первую очередь. Для выделения вируса и иммуногистохимических исследований были собраны гепариновая кровь, тимус и костный мозг. Иммуногистохимическое исследование проводили с помощью пероксидазного окрашивания тимуса с использованием КАВ-специфических моноклональных антител CV-1-85.1. Через четырнадцать и двадцать восемь дней после инфицирования были проведена аутопсия еще 5 птенцов, и все вышеуказанные анализы проводились.
Полимеразная реакция цепи /ПЦР/.
Олигонуклеотиды были синтезированы в синтезаторе циклон ДНК /Biosearch США/. Последовательность происходит от КАВ ДНК последовательности, показанной на фиг. 1. ПЦР была проведена на ДНК из КАВ-инфицированных и неинфицированных MDCC-MSBI клеток. Конечная концентрация реагентов была: 50 мМ KC, 10 мМ Трис-HCl /pH 8,3/, 3 мМ MgCl2, 0,01% сывороточного альбумина теленка, 200 мкМ для каждого dNTP, 1 мкМ для каждого олигонуклеотида и 2 единицы Tag-дНК полимеразы /CETVS; США/, в 100 мкл. ДНК образцы были циклически инкубированы 30 раз при 93oC в течение минуты, при 55oC в течение минуты, и при 72oC в течение 3 минут в Перкин/Элмер/ Цетус термическом циклизаторе. Одна десятая часть амплифицированной ДНК была прямо проанализирована на 2% агарозном/этидиум бромидном геле, или Саузерн-блоттингом. Использованная ДНК проба являлась олигонуклеотидом, меченным по концам 32P в соответствии с Maniatis с сотр. /1982/.
Дот-Блоттинг.
Вставка КАВ ДНК в pIC-20H/КАВ-EcoRI была помечена дигоксигенин-11-dVTP /Boehringer, Mannheim Германия/ в соответствии с протоколом поставщика. Биомеченые PP фильтры были насыщены 1,5 М NaCl и 0,15 М цитратом натрия. Образцы ДНК были ресуспендированы в 10 мМ Трис HCl /pH 7,5/ и 1 мМ ЭДТА, прокипечены в течение 3 минут, охлаждены на льду и нанесены на фильтр. Фильтр сушили при комнатной температуре и инкубировали 30 минут при 65oC. Фильтры гибридизовали с дигоксигенин меченой ДНК. ДНК, меченную дигоксигенином, обнаруживали с помощью иммунологического окрашивания в соответствии с протоколом поставщика.



Изобретение относится к обнаружению, клонированию и анализу последовательности ДНК генома вируса куриной анемии (КАВ). КАВ инфицированные клетки 1104-Х5 и MDCC-MSBI, содержащие двухцепочечную КАВ ДНК, имеющую длину 2,3 килобаз, клонировали в рIС-20Н. ДНК была полностью секвенирована. Рекомбинантную ДНК, включающую ДНК-последовательность КАВ, используют для диагностики, вакцинации или продуцирования КАВ-белка. КАВ-белок, в свою очередь, применяют для диагностики, вакцинации или продуцирования КАВ-специфичных антител. 14 с. и 1 з.п. ф-лы, 5 ил., 1 табл.
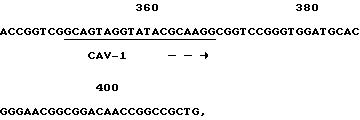
используемый в качестве праймера в полимеразной цепной реакции для идентификации природных изолятов СAV.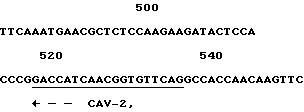
используемый в качестве праймера в полимеразной цепной реакции для идентификации природных изолятов СAV.
используемый в качестве гибридизационной пробы, 32Р, для идентификации природных изолятов СAV.

