Изобретение относится к материалам и методам сдерживания распространения инфекции вируса гепатита ни A, ни B. Более конкретно оно касается диагностических полипептидов, диагностических реагентов, диагностических наборов и диагностических способов для обнаружения этиологического агента гепатита вируса ни A, ни B, т.е. гепатита вируса C.
Библиографические ссылки в описании к заявке
Barr et al. (1986), Biotechniques 4:428.
Botstein (1979), Gene 8:17.
Brinton, M. A. (1986) in the Viruses: The togaviridae and Flaviviridae (Series eds. Fraenkel-Conrat and Wagner, vol. eds. Schlesinger and Schlesinger, Plenum Press), p. 327-374.
Broach (1981) in: Molecular Biology of the Yeast Saccharomyces. Vol. 1, p.445, Cold Spring Harbor Press.
Broach et al. (1983), Meth. Enz. 101:307.
Chang et аl. (1977), Nature 198:1056.
Chirgwin et аl. (1979), Biochemistry 18:5294.
Chomczynski and Sacchi (1987), Analytical Biochemistry 162:156.
Clewell et al. (1969), Proc. Natl. Acad. Sci. USA 62:1159.
Clewell (1972), J. Bacteriol. 110:667.
Cohen (1972), Proc. Natl. Acad. Sci. USA 69:2110.
Cousens et al. (1987), Gene 61:265.
De Boer et al. (1983), Proc. Natl. Acad. Sci. USA 292:128.
Dreesman et al. (1985), J. Infect. Disease 151:761.
Feinstone, S.M. and Hoofnagle, J.H. (1984), New Engl. J. Med. 311:185.
Fields & Knipe (1986), Fundamental Virology (Raven Press, N.Y.).
Fiers et al. (1978), Nature 273:113.
Gerety, R. J. et al., in Viral Hepatitis and Liver Disease (Vyas, B.N., Dienstag, J. L. , and Hoofnagle, J.H., eds, Grune and Stratton, Inc., 1984) pp. 23-47.
Goeddel et al. (1980), Nucleic Acids Res. 8:4057.
Graham and Van der Ed (1978), Virology 52:546.
Grunstein and Hogness (1975), Proc. Natl. Acad. Sci. USA 73:3961.
Grych et al. (1985), Nature 316:74.
Gubler and Hoffman (1983), Gene 25:263.
Hammerling et al. (1981), Monoclonal Antibodies and T-cell Hybridomas.
Hess et al. (1968), J. Adv. Enzyme Reg 7:149.
Hinnen et al. (1978), Proc. Natl. Acad. Sci. 75:1929.
Hitzeman et al. (1980), J. Biol. Chem. 255:2073.
Holland et al. (1978), Biochemistry 17:4900.
Holland (1981), J. Biol. Chem. 256:1385.
Houghton et al. (1981), Nucleic Acids Res. 9:247.
Hunyh, T. V. et al. (1985) in DNA Cloning Techniques; A Practical Approach (D. Glover, Ed., IRL Press, Oxford, U.K.) pp. 49-78.
Immun. Rev. (1982) 62:185.
Iwarson (1987), British Medical J. 295:946.
Kennett et al. (1980) Monoclonal Antibodies.
Laemmli (1970), Nature 227, 680.
Lee et al. (1988), Science 239:1288.
Maniatis, T. et al. (1982) Molecular Cloning; A Laboratory Manual (Cold Spring Harbor Press, Cold Spring Harbor, N.Y.).
Mayer and Walker, eds. (1987), Immunochemical Methods in Cell and Molecular Biology (Academic Press, London).
Maxam et al. (1980), Methods in Enzymology 65:499.
MacNamara et al. (1984), Science 226:1325.
Messing et al. (1981), Nucleic Acids Res. 9:309.
Messing (1983), Methods in Enzymology 101:20-37.
Methods in Enzymology (Academic Press).
Michelle et al. Int. Symposium on Viral Hepaatitis. Monath (1986) in The Viruses: The Togaviradae and Flaviviridae (Series eds. Fraenkel-Conrat and Wagner, vol. eds. Schlesinger and Schlesinger, Plenum Press), p.375-440.
Nagahuma et al. (1984), Anal. Biochem. 141:74.
Neurath et al. (1984), Science 224:392.
Nisonoff et al. (1981), Clin. Immunol. Immunopathol. 21:397-406.
Overby, L.R. (1985), Curr. Hepatol. 5:49.
Overby, L.R. (1986), Curr. Hepatol. 6:65.
Overby, L.R. (1987), Curr. Hepatol. 7:35.
Peleg (1969), Nature 221:193.
Pfefferkorn and Shapiro (1974), in Comprehensive Virology, Vol. 2 (Fraenkel-Conrat & Wagner, eds. Plenum, N.Y.) pp. 171-230.
Prince, A.M. (1983), Annu. Rev. Microbiol. 37:217.
Rice et al. (1986) in The Viruses: The Togaviridae and Flaviviridae (Series eds. Fraenkel-Conrat and Wagner, vol. eds. Schlesinger and Schlesinger, Plenum Press), p. 279-328.
Roehrig (1986) in The Viruses: The Togaviridae and Flaviviridae (Series eds. Fraenkel-Conrat and Wagner, vol. eds. Schlesinger and Schlesinger, Plenum Press).
Sadler et al. (1980), Gene 8:279.
Saiki et al. (1986), Nature 324:163.
Sanger et al. (1977), Proc. Natl. Acad. Sci. USA 74:5463.
Schlesinger et al. (1986), J. Virol. 60:1153.
Schreier, M., et аl. (1980) Hybridoma Techniques.
Scopes (1984), Protein Purification, Principles and Practice, Second Edition (Springer-Verlag, N.Y.).
Shimatake et аl. (1981), Nature 292:128.
Steimer et аl. (1986), J. Virol. 58:9.
Stollar (1980), in The Togaviruses (R. W. Schlesinger, ed. Academic Press, N.Y.), pp. 584-622.
Taylor et аl. (1976), Biochem. Biophys. Acta 442:324.
Towbin et аl. (1979), Proc. Natl. Acad. Sci. USA 76, 4350.
Tsu and Herzenberg (1980) in Selected Methods in Cellular Immunology (W. H. Freeman and Co.) pp. 373-391.
Vytdehaag et аl. (1985), J. Immunol. 134:1225.
Valenzuela, P. et al. (1982), Nature 298:344.
Valenzuela, P. et аl. (1984), in Hepatitis В (Millman, I. et аl., ed. Plenum Press) pp. 225-236.
Warner (1984), DNA 3:401.
Wu and Grossman (1987), Methods in Enzymology Vol. 154, Pecombinant DNA, Part E.
Wu (1987), Methods in Enzymology, Vol 155, Pecombinant DNA, Part F.
Zoller (1982), Nucleic Acids Res. 10:6487.
Ссылки на патенты
Патенты США NN 4341761, 4399121, 4427783, 4444887, 4466917, 4472500, 4491632 и 4493890.
Предпосылки для создания изобретения
Гепатит ни A, ни B является передающейся болезнью из семейства заболеваний, индуцированных, по-видимому, вирусом, и которые отличаются от других форм, обусловленных вирусом болезней печени, включая и те заболевания, которые вызваны известными вирусами гепатита, т.е. вирусом гепатита A, вирусом гепатита B и вирусом дельта-гепатита, а также от гепатитов, вызванных цитомегаловирусом или вирусом Эсстеина-Барра. Гепатит ни A, ни B был впервые идентифицирован у посттрансфузионных больных. Передача от человека шимпанзе и серия перепрививок у шимпанзе свидетельствуют о том, что гепатит ни A, ни B обусловлен переносимым инфекционным агентом или агентами. Однако переносимый возбудитель, ответственный за гепатит ни A, ни B, до настоящего времени не идентифицирован, и ряд агентов, которые являются причиной болезни, неизвестны.
Эпидемиологические данные наводят на мысль, что могут быть три типа гепатита ни A, ни B: эпидемический тип, связанный с водой; эпидемический тип, переносимый с кровью или иглой; и спорадически появляющийся (коммунально приобретенный) тип гепатита. Однако ряд возбудителей, которые могут вызывать гепатит ни A, ни B, неизвестен.
Клинический диагноз и идентификация гепатита ни A, ни B прежде всего осуществлялись путем исключения других вирусных маркеров. Среди методов, используемых для определения предполагаемых антигенов и антител гепатита ни A, ни B, агар-гелевая диффузия, радиоиммуноэлектрофорез, иммунофлюоресцентная микроскопия, иммуноэлектронная микроскопия, радиоиммунологические тесты, ферментосвязанный иммуносорбционный тест. Однако ни один из этих методов анализа не показал достаточной чувствительности, специфичности и воспроизводимости, чтобы быть использованным в качестве диагностического теста на гепатит ни A, ни B.
До настоящего времени не существует ни ясности, ни согласия относительно идентичности и специфичности систем антиген - антитело, связанных с возбудителями гепатита ни A, ни B. Это обусловлено, по меньшей мере частично, первичной или сопутствующей инфекцией вируса гепатита В с вирусом гепатита ни A, ни B у больных и известной сложностью растворимых и в виде частиц антигенов, связанных с вирусом гепатита B, а также интеграцией ДНК вируса гепатита В в геном клеток печени. В дополнение не исключена возможность, что гепатит ни A, ни B вызывается не одним инфекционным агентом, а также возможность того, что гепатит ни A, ни B диагностирован ошибочно. Кроме того неясно, что определяют серологические анализы в сыворотке больных гепатитом ни A, ни B. Было принято без доказательства, что агар-гелевая диффузия и радиоиммуноэлектрофоретические анализы определяют аутоиммунные реакции или взаимодействия неспецифических белков, которые иногда происходят между пробами сыворотки, и что они не представляют собой специфических реакций антиген - антитело. Иммунофлюоресценция, ферментосвязанный иммуносорбент и радиоиммунологический анализы, по-видимому, определяют низкие уровни материала, аналогичного ревматоидному фактору, который часто присутствует в сыворотке больных гепатитом ни A, ни B, а равно как и у больных с другими заболеваниями печени и болезнями, не связанными с печенью. Некоторую иммунологическую реактивность, определенную анализами, может представлять антитело к определенным хозяином цитоплазменным антигенам.
Имеется несколько кандидатов на вирус гепатита ни A, ни B. Смотрите, например, обзоры Prince (1983), Feinstone и Hoofnagle (1984) и Overby (1985, 1986 и 1987 гг.) и статью Iwarson (1987). Но нет доказательств, что какой-либо из этих кандидатов представляет собой этиологический агент гепатита ни A, ни B.
Потребность в чувствительных, специфических методах выявления и идентификации носителей вируса гепатита ни A, ни B и зараженной вирусом ни A, ни B крови и продуктов крови весьма значительная. Посттрансфузионные гепатиты появляются примерно в 10% случаев больных с переливанием крови и до 90% из этих случаев насчитывается больных гепатитом ни A, ни B. Основной проблемой при этом заболевании является частое развитие хронического поражения печени (25-55%).
Лечение больного, а также профилактика передачи гепатита ни A, ни B с кровью или продуктами крови или при тесном контакте с больными требуют надежных диагностических и прогностических средств для определения нуклеиновых кислот, антигенов и антител, связанных с вирусом гепатита ни A, ни B. В дополнение имеется необходимость также в эффективных вакцинах и иммунотерапевтических лечебных средствах для профилактики и/или лечения этой болезни.
Сущность изобретения
Изобретение касается выделения и определения характеристик недавно открытого этиологического агента гепатита ни A, ни B, вируса гепатита C. Более конкретно изобретение касается семейства реплик кольцевой ДНК частей генома гепатита вируса C. Эти реплики кольцевой ДНК были выделены методом, который включает новую операцию отсеивания продуктов экспрессии их пулов кольцевой ДНК, созданных из частиц возбудителя (агента) в инфицированной ткани с сыворотками больных гепатитом ни A, ни B, для определения вновь синтезированных антигенов, производных от генома до этого выделенного и неохарактеризованного вирусного агента, и селекцию клонов, которые вырабатывают продукты, иммунологически взаимодействующие только с сыворотками инфицированных индивидов в сравнении с неинфицированными лицами.
Исследования природы генома вируса гепатита C, используя пробы, полученные из кольцевой ДНК вируса гепатита C, а также последовательность информации, содержащейся в кольцевой ДНК вируса гепатита C, говорят о том, что вирус гепатита C является флавивирусом или флавиподобным вирусом.
Части последовательности оснований кольцевой ДНК, производные от вируса гепатита C, полезны как пробы для определения присутствия вируса в образцах и для выделения встречающихся в природе вариантов вируса. Эти фрагменты кольцевой ДНК позволяют также оценить полипептидные последовательности антигенов вируса гепатита C, закодированные в геноме (геномах) вируса гепатита C, и позволяют продуцирование полипептидов, которые полезны в качестве стандартов реагентов в диагностических тестах и/или в качестве компонентов вакцин. Антитела, как поликлональные, так и моноклональные, направленные против эпитопов вируса гепатита C, содержащихся в этих полипептидных последовательностях, также полезны для диагностических тестов, в качестве терапевтических средств, для отсеивания противовирусных агентов и для изоляции (выделения) возбудителя вируса ни A, ни B, от которого получены эти фрагменты кольцевой ДНК. В дополнение при использовании проб, полученных от этих частей кольцевой ДНК, возможно выделить и оценить последовательность других частей генома вируса гепатита C, получая таким образом дополнительные пробы и полипептиды, полезные для диагностики и/или лечения, как в профилактическом, так и в лечебном воздействии на гепатит ни A, ни B.
Аспектами изобретения являются: очищенный вирус гепатита C; приготовление полипептидов из очищенного вируса гепатита C; очищенный полипептид вируса гепатита C; очищенный полипептид, содержащий эпитоп, иммунологически идентифицируемый эпитопом, содержащимся в вирусе гепатита C.
Включенными аспектами изобретения являются: рекомбинантный полипептид вируса гепатита C; рекомбинантный полипептид, содержащий последовательность, полученную от генома вируса гепатита C или от кольцевой ДНК вируса гепатита C; рекомбинантный полипептид, содержащий эпитоп вируса гепатита C и конденсированный полипептид, полипептид слияния, содержащий полипептид вируса гепатита C.
Аспекты изобретения, касающиеся комплектов диагностических средств, следующие: образцы для анализа на присутствие полинуклеотидов, производных от вируса гепатита C, включающие пробу, содержащую последовательность нуклеотидов вируса гепатита C примерно 8 или более нуклеотидов, в подходящей упаковке; образцы для анализа на присутствие антигена вируса гепатита C, содержащие антитело, направленное против антигена вируса C, подлежащего определению, в подходящей упаковке; образцы для анализа на присутствие антител, направленных против антигена вируса гепатита C, включающие полипептид, содержащий эпитоп вируса гепатита C, присутствующий в антигене вируса гепатита C, в соответствующей упаковке.
Другими аспектами изобретения являются: полипептид, содержащий этипоп вируса гепатита C, фиксированный на твердом субстрате, и антитело к эпитопу вируса гепатита C, фиксированное на твердом субстрате.
Следующими аспектами изобретения являются: метод получения полипептида, содержащего эпитоп вируса гепатита C, включающий инкубирование клеток хозяина, трансформированных экспрессионным вектором, содержащим последовательность кодирования полипептида, содержащего эпитоп вируса гепатита C в условиях, допускающих экспрессию упомянутого полипептида, и полипептид, содержащий эпитоп, полученный этим метолом.
Изобретение включает также способ определения нуклеиновых кислот в образце, содержащем реагирующие нуклеиновые кислоты образца с пробой на полинуклеотид вируса гепатита C в условиях, позволяющих образование полипептидного дуплекса между пробой и нуклеиновой кислотой вируса гепатита C образца, и определение полинуклеотидного дуплекса, который содержит проба.
Иммунологические исследования также включены в объем изобретения. Они включают иммунологический анализ для определения антигена вируса гепатита C, включающий инкубирование образца, подознительного на содержание антигена вируса гепатита C, с антителом пробы, направленным против антигена вируса гепатита C, подлежащего определению в условиях, позволяющих образование комплекса антиген - антитело; и определение комплекса антиген - антитело, содержащего антитело пробы. Иммунологический анализ для определения антител, направленных против антигена вируса гепатита C, включающий инкубирование образца, подозрительного на содержание антител антивируса гепатита C, с полипептидом пробы, которая содержит эпитоп вируса гепатита C, в условиях, допускающих образование системы антитело - антиген, и определение системы антитело - антиген, содержащей антиген пробы.
В объем изобретения включены также вакцины для лечения инфекции вируса гепатита C, включающие иммуногенный пептид, содержащий эпитоп вируса гепатита C, или инактивированный препарат вируса гепатита C, или аттенуированный препарат вируса гепатита C.
Другим аспектом изобретения является культура ткани растущих клеток, инфицированных вирусом гепатита C.
Еще иным аспектом изобретения является способ получения антител к вирусу гепатита C, включающий введение больному выделенный иммуногенный полипептид, содержащий эпитоп вируса гепатита C в количестве, достаточном для продуцирования иммунной реакции.
И еще одним аспектом изобретения является способ выделения кольцевой ДНК, полученной из генома неидентифицированного инфекционного агента, включающий: (a) получение клеток хозяина, трансформированных векторами экспрессии, содержащимися в пуле кольцевой ДНК, приготовленном из нуклеиновых кислот, выделенных из ткани, инфицированной агентом, и выращивание клеток хозяина в условиях, допускающих экспрессию полипептида(ов), закодированных в кольцевой ДНК; (b) взаимодействие продуктов экспрессии кольцевой ДНК с содержащим антитело компонентом тела больного, инфицированного упомянутым инфекционным возбудителем в условиях, позволяющих иммунную реакцию, и определение комплексов антитело - антиген, образованных в результате взаимодействия; (c) выращивание клеток хозяина, осуществляющих экспрессию полипептидов, которые образуют комплексы антитело - антиген на операции (b) в условиях, допускающих их выращивание как индивидуальных клонов, и выделение упомянутых клонов; (d) выращивание клеток из клонов, полученных на операции (с) в условиях, допускающих экспрессию полипептида(ов), закодированных в кольцевой ДНК, и взаимодействие продуктов экспрессии с содержащими антитело компонентами тела индивидов, отличных от больного на операции (а), которые инфицированы инфекционным агентом, и с контрольными лицами, не инфицированными агентом, и определение комплексов антитело - антиген, образованных в результате взаимодействия; (e) выращивание клеток хозяина, которые осуществляют экспрессию полипептидов, которые образуют комплексы антитело - антиген с содержащими антитело компонентами тела инфицированных индивидов и лиц, подозрительных на инфекцию, но не с компонентами лиц в контрольной группе, в условиях, допускающих как развитие отдельных клонов, так и выделение упомянутых клонов; и (f) выделение кольцевой ДНК из клонов клеток хозяина, полученных на операции (e).
На фиг. 1 показана двунитчатая последовательность нуклеотидов кольцевой ДНК вируса гепатита C, вставленная в клон 5-1-1, и предполагаемая последовательность аминокислот полипептида, закодированного в ней.
На фиг. 2 представлены гомологи перекрытия последовательностей кольцевой ДНК вируса гепатита C в клонах 5-1-1, 81, 1-2 и 91.
На фиг. 3 показана сложная последовательность оснований кольцевой ДНК вируса гепатита C, полученная от перекрытия клонов 81, 1-2 и 91, и последовательность аминокислот, закодированная в ней.
На фиг. 4 показана последовательность двунитчатых нуклеотидов кольцевой ДНК вируса гепатита C, вставленной в клон 81, и предполагаемая последовательность аминокислот, закодированная в ней.
На фиг. 5 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне 36, сегмент которой перекрывает кольцевую ДНК вируса гепатита ни A, ни B в клоне 81, и последовательность полипептидов, закодированная в клоне 36.
На фиг. 6 представлена объединенная открыто читаемая структура кольцевой ДНК вируса гепатита C в клонах 36 и 81 и полипептиды, закодированные в ней.
На фиг. 7 показана последовательность оснований кольцевой ДНК вируса гепатита C в клоне 32, сегмент которой перекрывает клон 81, и закодированный в ней полипептид.
На фиг. 8 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне 35, сегмент которой перекрывает клон 36, и закодированный в ней полипептид.
На фиг. 9 представлена объединенная открыто читаемая структура кольцевой ДНК вируса гепатита C в клонах 35, 36, 81 и 32 и закодированный в ней полипептид.
На фиг. 10 показана последовательность оснований кольцевой ДНК вируса гепатита C в клоне 37b, сегмент которой перекрывает клон 35, и закодированный в ней полипептид.
На фиг. 11 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне 33b, сегмент которой перекрывает клон 32, и закодированный в ней полипептид.
На фиг. 12 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне 40b, сегмент которой перекрывает клон 37b, и закодированный в ней полипептид.
На фиг. 13 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне 25c, сегмент которой перекрывает клон 33b, и закодированный в ней полипептид.
На фиг. 14 представлена последовательность нуклеотидов и закодированный в ней полипептид открыто читаемой структуры, которая простирается через кольцевую ДНК вируса гепатита C в клонах 40b, 37b, 35, 36, 81, 32, 33b и 25c.
На фиг. 15 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клонах 40b и 33c, и закодированные в ней аминокислоты.
На фиг. 16 представлена последовательность оснований (кольцевой) ДНК вируса гепатита C в клоне 8h, сегмент которой перекрывает клон 33c, и закодированные в ней аминокислоты.
На фиг. 17 представлена последовательность оснований в кольцевой ДНК вируса гепатита C в клоне 7е, сегмент которой перекрывает клон 8h, и закодированные в ней аминокислоты.
На фиг. 18 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне 14c, сегмент которой перекрывает клон 25c, и закодированные в ней аминокислоты.
На фиг. 19 представлена последовательность оснований в кольцевой ДНК вируса гепатита C в клоне 8f, сегмент которой перекрывает клон 14c, и закодированные в ней аминокислоты.
На фиг. 20 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне 33f, сегмент которой перекрывает клон 8f, и закодированные в ней аминокислоты.
На фиг. 21 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне 33g, сегмент которой перекрывает клон 33f, и закодированные в ней аминокислоты.
На фиг. 22 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне 7f, сегмент которой перекрывает последовательность оснований в клоне 7e, и закодированные в ней аминокислоты.
На фиг. 23 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне 11b, сегмент которой перекрывает последовательность оснований в клоне 7f, и закодированные в ней аминокислоты.
На фиг. 24 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне 14i, сегмент которой перекрывает последовательность оснований в клоне 11b, и закодированные в ней аминокислоты.
На фиг. 25 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне 39c, сегмент которой перекрывает последовательность оснований в клоне 33g, и закодированные в ней аминокислоты.
На фиг. 26 представлена сложная последовательность оснований кольцевой ДНК вируса гепатита C, полученная от объединения кольцевых ДНК в клонах 14i, 11b, 7f, 7e, 8h, 33с, 40b, 37b, 35, 36, 81, 32, 33b, 25c, 14c, 8f, 33f и 33g, а также представлена последовательность аминокислот полипептида, закодированная в протяженной открыто читаемой структуре в полученной последовательности оснований.
На фиг. 27 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне 12f, сегмент которой перекрывает клон 14i, и закодированные в ней аминокислоты.
На фиг. 28 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне 35f, сегмент которой перекрывает клон 39c, и закодированная в ней последовательность аминокислот.
На фиг. 29 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне 19g, сегмент которой перекрывает клон 35f, и закодированные в ней аминокислоты.
На фиг. 30 представлена последовательность оснований в клоне 26g, сегмент которой перекрывает клон 19g, и закодированные в ней аминокислоты.
На фиг. 31 представлена последовательность оснований в клоне 15e, сегмент которой перекрывает клон 26g, и закодированные в ней аминокислоты.
На фиг. 32 представлена последовательность оснований в сложной кольцевой ДНК, которая получена объединением клонов с 12f до 15e в направлении 5' к 3'; представлены также аминокислоты, закодированные на непрерывной открыто читаемой структуре.
На фиг. 33 представлена фотография западных пятен, полученная на конденсированном белке, пероксиддисмутазе - ни A, ни B5-1-1 с сывороткой шимпанзе, инфицированной вирусами гепатита ни A, ни B, A и B.
На фиг. 34 представлена фотография западных пятен, полученная на конденсированном белке, пероксиддисмутазе - ни A, ни B5-1-1 с сывороткой человека, инфицированного вирусом гепатита ни A, ни B, вирусом гепатита A, вирусом гепатита B и от контрольной группы лиц.
На фиг. 35 представлена диаграмма существенных признаков вектора рAB24.
На фиг. 36 представлена предполагаемая последовательность аминокислот карбоксиокончания конденсированного полипептида C100-3 и закодированная в ней последовательность нуклеотидов.
На фиг. 37A представлена фотография окрашенного синим красителем полиакриламидного геля, который идентифицирует полипептид C100-3, экспрессированный в дрожжах.
На фиг. 37B представлено западное пятно полипептида C100-3 с сывороткой инфицированного вирусом ни A, ни B человека.
На фиг. 38 представлена ауторадиография северного пятна РНК, выделенной из печени инфицированной вирусом BB-ни A, ни B шимпанзе, исследованной вирусом BB-ни A, ни B кольцевой ДНК клона 81.
На фиг. 39 представлена ауторадиография нуклеиновой кислоты вируса ни A, ни B, обработанной рибонуклеазой A или дезоксирибонуклеазой I и исследованной кольцевой ДНК вируса BB-ни A, ни B клона 81.
На фиг. 40 представлена ауторадиография нуклеиновых кислот, экстрагированных из частиц вируса ни A, ни B, отловленных в инфицированной анти-ни A, ни B5-1-1 плазме, и исследованных меченной фосфором-32 кольцевой ДНК вируса ни A, ни B из клона 81.
На фиг. 41 представлена ауторадиограмма фильтров, содержащих выделенные аминокислоты вируса ни A, ни B, исследованные меченными фосфором-32 и плюс окрашенными пробами для ДНК, полученными из кольцевой ДНК вируса ни A, ни B в клоне 81.
На фиг. 42 представлены гомологи между полипептидом, закодированным в кольцевой ДНК вируса гепатита C и NS-белком флавивируса Денгье.
На фиг. 43 представлена гистограмма распределения инфекции вируса гепатита C в выбранных наугад образцах, как определено иммуноферментным твердофазным анализом.
На фиг. 44 представлена гистограмма распределения инфекции вируса гепатита C в случайных образцах, полученная с использованием двух конфигураций сопряжения иммуноглобулин - фермент в иммуноферментном твердофазном анализе.
На фиг. 45 представлены последовательности в первичной смеси, полученной из сохраненной последовательности в NS1 флавивирусов.
На фиг. 46 представлена последовательность оснований кольцевой ДНК вируса гепатита C в клоне k9-l, сегмент которой перекрывает кольцевую ДНК на фиг. 26, и закодированные на ней аминокислоты.
На фиг. 47 представлена последовательность оснований сложной кольцевой ДНК, полученной при соединении клонов k9-1 до 15e в направлении 5' к 3'; представлены также аминокислоты, закодированные в открыто читаемой протяженной структуре.
Способы осуществления изобретения
I. Определения
Термин "вируса гепатита C" предназначался разработчиками в данной области для дотоле неизвестного этиологического агента ни A, ни B. Следовательно, употребленный в описании "вируса гепатита C" касается возбудителя гепатита ни A, ни B, который ранее относился к вирусу ни A, ни B и/или BB-ни A, ни B. Термины "вируса гепатита C, вирус гепатита ни A, ни B и вирус гепатита BB-ни A, ни B" используются здесь как равнозначные. Как расширение этой терминологии болезнь называлась вирус гепатита C, прежде названная гепатит ни A, ни B, гепатит C. Термины гепатит ни A, ни B и гепатит C могут быть использованы здесь как равнозначные.
Термин "вируса гепатита C", используемый в описании, означает виды вирусов, вызывающих гепатит ни A, ни B, и ослабленные штаммы и дефектные вирусные частицы, производные от этих штаммов. Как показано будет ниже, геном вируса гепатита C состоит из РНК. Известно, что РНК-содержащие вирусы имеют относительно высокие уровни спонтанной мутации, т.е. по опубликованным данным порядка 10-3-10-4 на нуклеотид (Fields и Knipe, 1986). Поэтому имеют место множественные штаммы внутри вида вируса гепатита C, определенные ниже. Составы и методы, описанные здесь, делают возможным репродукцию, идентификацию, детектирование и выделение различных родственных вирусов. Более того, они позволяют также обеспечить получение диагностических средств и вакцин для различных штаммов и полезны в процессе отбора противовирусных агентов для фармакологического использования, так как они ингибируют репликацию вируса гепатита C.
Приведенная здесь информация, хотя и получена на одном штамме вируса гепатита C, вызываемого далее CDC/HCV1, является достаточной для того, чтобы вирусный таксономист идентифицировал другие штаммы, которые принадлежат этому виду вирусов. Как описывалось уже, мы открыли, что вирус гепатита C относится к флавивирусу или флавиподобному вирусу. Морфология и состав частиц флавивируса известны и описаны Brinton (1986). Вообще, принимая во внимание морфологии, флавивирусы содержат центральный нуклеокапсид, окруженный липидным двуслоем. Вирионы сферической формы и имеют диаметр около 40-50 нм. Их ядра имеют диаметр около 25-30 нм. Вдоль наружной поверхности оболочки вириона имеются выросты длиной примерно 5-10 нм с концевыми выпуклостями диаметром около 2 нм.
Вирус гепатита C кодирует эпитоп, который иммунологически тождественен эпитопу в геноме вируса гепатита C, от которого происходят кольцевые ДНК, записанные в нем. Эпитоп является единственным в своем роде к вирусу гепатита C, если сравнить с другими известными флавивирусами. Уникальность эпитопа может быть определена его иммунологической реактивностью с вирусом гепатита C и отсутствием иммунологической реактивности с другими видами флавивирусов. Способы определения иммунологической реактивности известны в данной области, например радиоиммунологическим анализом, иммуноферментным твердофазным анализом, гемагглютинацией, и несколько примеров подходящих методов анализа даны в описании.
В дополнение к описанному выше применимы следующие параметры, либо в отдельности, либо в сочетании при идентификации штамма как вируса гепатита C. Поскольку штаммы вируса гепатита C эволюционно родственны, то следует ожидать, что полная гомология геномов на нуклеотидном уровне будет около 40% или более, предпочтительно около 60% или более и еще более предпочтительно около 80% или более; и в дополнение, что будут иметь место соответствующие близкие последовательности по меньшей мере около 13 нуклеотидов. Соответствие между геномной последовательностью предполагаемого вируса гепатита C и последовательностью оснований кольцевой ДНК вируса гепатита C CDC/CHl может быть определена методами, известными в данной области. Например, они могут быть определены непосредственным сравнением последовательной информации полинуклеотида от предполагаемого вируса гепатита C и последовательности(ей) кольцевой ДНК вируса гепатита C, записанной в нем. Например, они могут быть, таким образом, определены гибридизацией полинуклеотидов в условиях, в которых образуются стабильные дуплексы между гомологическими участками, например те, которые должны быть использованы до ферментации с последующей ферментацией однотяжевой специфической нуклеазой(ами), сопровождаемой определением размеров ферментационных фрагментов.
По причине эволюционного родства штаммов вируса гепатита C предполагаемые штаммы вируса гепатита C тождественны по своей гомологии на полипептидном уровне. Вообще штаммы вируса гепатита C более чем на 40% являются гомологическими, предпочтительно более чем на 60% гомологи и даже еще более предпочтительно более чем на 80% гомологи на полипептидном уровне. Методы определения гомологии последовательности аминокислот известны в данной области. Например, последовательность аминокислот может быть определена непосредственно и сравнена с последовательностями, предусмотренными при этом заранее. Например, также нуклеотидная последовательность геномного материала предполагаемого вируса гепатита C может быть определена (обычно через промежуточную кольцевую ДНК): закодированная в ней последовательность аминокислот может быть определена и соответствующие участки сравнены.
Как использовано в описании, полинуклеотид, "производный от" (полученный из) указанной последовательности, например кольцевой ДНК вируса гепатита C, в особенности из тех, которые приведены в примерах на фиг. 1-32, или от генома вируса гепатита C, относится к полинуклеотидной последовательности, которая содержит примерно последовательность по меньшей мере 6 нуклеотидов, является предпочтительно по меньшей мере последовательностью около 8 нуклеотидов и более предпочтительно по меньшей мере 10-12 нуклеотидов и еще более предпочтительно по меньшей мере 15-20 нуклеотидов соответственно, т.е. гомологично или комплементарно к участку указанной последовательности нуклеотидов. Предпочтительно, чтобы последовательность участка, из которого происходит полинуклеотид, являлась гомологичной или комплементарной последовательности, которая уникальна для генома вируса гепатита C. Будет или нет эта последовательность уникальна к геному вируса гепатита C, можно определить методами, известными специалистам в данной области. Например, последовательность можно сравнить с последовательностями в банке данных, т.е. в банке генов, чтобы определить, присутствует ли она у неинфицированного хозяина или в других организмах. Последовательность можно также сравнить с известными последовательностями других вирусных агентов, включая тех, которые известны как возбудители гепатита, т.е. вируса гепатита A, вируса гепатита B и вируса гепатита D, и с другими членами семейства флавивирусов. Соответствие или несоответствие взятой последовательности другим последовательностям может быть также определено гибридизацией в соответствующих строгих условиях. Методы определения комплементарности последовательностей нуклеиновых кислот посредством гибридизации известны в данной области и обсуждаются ниже. См. также, например, Maniatis и др. (1982). В дополнение несоответствия двойных полинуклеотидов, образованных при гибридизации, можно определить известными методами, включающими, например, ферментацию нуклеазой, такой как S1, которая специфически сбраживает однотяжевые участки в двойных полинуклеотидах. Области, из которых может быть "произведена" типичная последовательность ДНК, включают, но этим не ограничиваясь, например, области, кодируемые специфическими эпитопами, а также нетранскрибированные и/или нетранслированные области.
Производный нуклеотид не обязательно является физически произведенным от показанной последовательности нуклеотидов, а может быть получен любым образом, включающим, например, химический синтез или репликацию ДНК или обратную транскрипцию или транскрипцию, которая основана на информации, даваемой последовательностью оснований в участке (в участках), от которых происходит полинуклеотид. В дополнение сочетания участков, соответствующих таковым известной последовательности, могут быть модифицированы путями, известными в данной области, соответствующими намечаемому использованию.
Аналогично последовательность полипептида или аминокислот, "производных" от соответствующей последовательности нуклеиновых кислот, например последовательности, представленные на фиг. 1-32, или от генома вируса гепатита C, относится к полипептиду, имеющему последовательность аминокислот, идентичную последовательности полипептида, закодированного в последовательности или в ее части, при этом часть содержит по меньшей мере 3-5 аминокислот, и более предпочтительно по меньшей мере 8-10 аминокислот, и еще более предпочтительно по меньшей мере 11-15 аминокислот, или который иммунологически тождественен полипептиду, закодированному в последовательности.
Рекомбинантный или производный полипептид не обязательно является транслированным от известной последовательности нуклеиновых кислот, например последовательностей, представленных на фиг. 1-26, или от генома вируса гепатита C: он может быть получен и иным образом, например химическим синтезом, или экспрессией рекомбинантной экспрессионной системы, или выделением из мутированного вируса гепатита C.
Термин "рекомбинантный полинуклеотид", использованный в описании, имеет в виду полинуклеотид геномного, полусинтетического, синтетического происхождения или происходящий от кольцевой ДНК, и благодаря своему происхождению и обработке: (1) не связан со всем или с частью полинуклеотида, с которым он связан в природе или в форме банка данных; и/или (2) присоединен к полинуклеотиду иначе, чем полинуклеотид, с которым он связан в природе.
Термин "полинуклеотид", использованный в описании, относится к полимерной форме нуклеотидов любой длины как рибонуклеотидам, так и к дезоксирибонуклеотидам. Этот термин относится только к первичной структуре молекулы. Таким образом, этот термин включает двунитчатую и однонитчатую ДНК, а также двунитчатую и однонитчатую РНК. Он также включает модифицированные, например метилированием и/или покрытием, и немодифицированные формы полинуклеотида.
Использованный в описании термин "вируса гепатита C, содержащий последовательность, соответствующую кольцевой ДНК", означает, что вирус гепатита C содержит полинуклеотидную последовательность, которая гомологична или комплементарна последовательности в соответствующей ДНК; степень гомологии или комплементарности к кольцевой ДНК будет приблизительно 50% или более и предпочтительно по меньшей мере около 70% и даже еще более предпочтительно будет по меньшей мере около 90%. Последовательности, которые соответственны, будут насчитывать по меньшей мере 70 нуклеотидов, предпочтительно по меньшей мере около 80 нуклеотидов и даже еще более предпочтительно по меньшей мере около 90 нуклеотидов в длину. Соответствие между последовательностью вируса гепатита C и кольцевой ДНК можно определить методами, известными в данной области, включающими, например, прямое сравнение материала последовательности с описанными кольцевыми ДНК или гибридизацией и ферментацией однонитчатой нуклеазой с последующим определением размера ферментационных фрагментов.
Термин "очищенный вирусный полинуклеотид" относится к геному вируса гепатита C или его фрагменту, который в значительной мере свободен, т.е. содержит менее чем 50%, предпочтительно менее 70% и даже еще более предпочтительно менее 90% полипептидов, с которыми вирусный полинуклеотид связан в природе. Технология очистки вирусных полинуклеотидов от вирусных частиц известна в данной области и включает, например, разрыв частицы chao-тропным агентом и разделение полинуклеотида(ов) и полипептидов ионообменной хромаографией, хроматографией по сродству и седиментацией по плотности.
Термин "очищенный вирусный полипептид" относится к полипептиду вируса гепатита C или его фрагменту, который в значительной мере свободен, т.е. содержит менее 50%, предпочтительно менее 70% и даже более предпочтительно менее 90% клеточных компонентов, с которыми вирусный полипептид естественно связан. Методы очистки вирусных полипептидов известны в данной области и примеры этих методов будут рассмотрены ниже.
"Рекомбинантные клетки хозяина", "клетки хозяина", "клетки", "линии клеток", "культуры клеток" и другие такие термины означают микроорганизмы или линии высших кариотных клеток, выращенные как одноклеточные формы, относящиеся к клеткам, которые могут быть или были использованы как реципиенты для рекомбинантного вектора или других переносов ДНК и включают прогению первоначальных клеток, которые были заражены вирусной нуклеиновой кислотой. Следует понимать, что прогения отдельной родительской клетки может необязательно быть полностью идентичной по морфологии или по геномному или общему комплементу ДНК, как первоначальный родитель вследствие случайной или преднамеренной мутации. Потомки родительской клетки, которые достаточно аналогичны родительским, должны характеризоваться релевантным свойством, таким как наличие нуклеотидной последовательности, кодирующей желаемый белок, включены в потомство, подпадающее под это определение, и входят в объем упомянутого выше термина.
"Репликон" является любым генетическим элементом, т.е.
плазмида, хромосома, вирус, который ведет себя как автономная единица полинуклеотидной репликации в клетке, т.е. способен к репликации под своим собственным контролем.
"Вектор" является репликоном, к которому присоединен другой полинуклеотидный сегмент с тем, чтобы осуществить репликацию и/или экспрессию (синтез) присоединенного сегмента.
"Контрольная последовательность" относится к полинуклеотидным последовательностям, которые необходимы для осуществления экспрессии кодируемых последовательностей, с которыми они связаны. Природа таких контрольных последовательностей различна в зависимости от организма хозяина: у прокариотов такие контрольные последовательности обычно включают промотор, место связывания рибосомы и терминаторы; у эукариотов вообще такие контрольные последовательности включают промоторы, терминаторы и в некоторых случаях - гены-усилители. Термин "контрольная последовательность" имеет в виду включение, как минимум, всех компонентов, присутствие которых необходимо для экспрессии, и может также включать дополнительные компоненты, присутствие которых выгодно, например основные последовательности.
"Операбельно связан" относится к положению, при котором описанные компоненты находятся в отношении, позволяющем им осуществлять функцию предназначенным образом. Контрольная последовательность, "операбельно связанная" с кодирующей последовательностью, соединена таким образом, что экспрессия кодирующей последовательности достигается в условиях, совместимых с контрольными последовательностями.
"Открытая читаемая структура" является областью полинуклеотидной последовательности, которая кодирует полипептид; эта область может находиться в части кодирующей последовательности и (общей) всей кодирующей последовательности.
"Кодирующая последовательность" является последовательностью полинуклеотидов, которая транскрибирована в информационную РНК и/или транслирована в полипептид, будучи поставлена под контроль соответствующих регулярных последовательностей. Границы кодирующей последовательности определены запускающим трансляцию кодоном при 5'-терминале и останавливающим трансляцию кодоном при 3'-терминале. Кодирующая последовательность включает, но не ограничивается этим, информационную РНК, кольцевую ДНК и рекомбинантные полинуклеотидные последовательности.
"Иммунологически идентичны с/как" относится к наличию эпитопа(ов) и полипептида(ов), которые также присутствуют и являются уникальными к определенным полипептиду(ам), обычно к белкам вируса гепатита C. Иммунологическая идентичность может быть определена связыванием антител и/или конкуренцией в связывании; эти методы известны среднему специалисту в данной области и также пояснены ниже. Уникальность эпитопа может быть также определена компьютерными исследованиями известных банков данных, т.е. генного банка, для полинукеотидных последовательностей, которые кодируют эпитоп, и сравнениями последовательности аминокислот с другими известными белками.
Использованный в описании термин "эпитоп" относится к антигенной детерминанте полипептида: эпитоп должен содержать три аминокислоты в пространственной конформации, которая уникальна для эпитопа, обычно эпитоп содержит по меньшей мере пять аминокислот и более обычно содержит по меньшей мере 8-10 аминокислот. Методы определения пространственной конформации аминокислот известны в данной области и включают, например, рентгеновскую кристаллографию и ядерный магнитный резонанс в двух измерениях.
Полипептид является "иммунологически реакционноспособным" с антителом, если он связывает антитело благодаря распознаванию антителом специфического эпитопа, заключенного в полипептиде. Иммунологическая реактивность может быть определена связыванием антитела, более конкретно кинетикой связывания антитела, и/или конкуренцией в связывании, используя в качестве конкурента(ов) известный полипептид(ы), содержащий эпитоп, против которого направлено антитело. Методы определения, является ли полипептид реакционноспособным с антителом, известны в данной области.
Использованный в описании термин "иммуногенный полипептид, содержащий эпитоп вируса гепатита C" включает встречающиеся в природе полипептиды вируса гепатита C или его фрагменты, а также полипептиды, полученные другими средствами, например химическим синтезом или экспрессией полипептида в рекомбинантном организме.
Термин "полипептид" относится к молекулярной цепи аминокислот и не относится к специфической длине продукта; таким образом, пептиды, олигопептиды и белки включены в определение полипептида. Этот термин не относится к постэкспрессионным модификациям полипептида, например гликолизациям, ацетиляциям, фосфориляциям и тому подобным.
Термин "трансформация", использованный в описании, относится к вставке экзогенного полинуклеотида в клетку хозяина, независимо от метода, используемого для введения, например прямое поглощение, трансдукция или f-скрещивание. Экзогенный полинуклеотид может быть сохранен как неинтегрированный вектор, например плазмида, или альтернативно может быть интегрирован в геном хозяина.
"Обработка", использованная в описании, относится к профилактике и/или терапии.
Термин "индивид", использованный в описании, относится к позвоночным, в частности к особям вида млекопитающих, и включает, но не ограничивается, домашними животными, спортивными животными, приматами и людьми.
Термин "плюс-тяж" (нить) нуклеиновой кислоты, использованный в описании, содержит последовательность, которая кодирует полипептид.
"Минус-нить" содержит последовательность, комплементарную к последовательности "плюс-нити".
Термин "положительный нитчатый геном" вируса, использованный в описании, является таковым, в котором геном, или ДНК, или РНК является однонитчатой и который кодирует вирусный полипептид(ы). Примеры положительных нитчатых РНК-вирусов включают тогавирусы, коронавирусы, ретровирусы, пикорнавирусы и кальцивирусы. Включены также флавивирусы, которые прежде классифицировались как тогавирусы. Смотрите Fields и Knipe (1986).
Использованный в описании термин "антитело, содержащее компонент тела", относится к компоненту тела индивида, который является источником антител, представляющих интерес. Антитело, содержащее компоненты тела, известны в данной области и включают, но не ограничиваются, например, плазмой, сывороткой, спинно-мозговой жидкостью, лимфой и внешними секрециями дыхательного, желудочно-кишечного и мочеполового трактов, а также слезы, слюна, молоко, клетки белой крови и миеломы.
Использованный в описании термин "очищенный вирус гепатита C" касается препарата вируса гепатита C, который выделен из клеточных составляющих, с которыми обычно связан вирус, и из других типов вирусов, которые могут присутствовать в инфицированной ткани. Методы выделения вирусов известны специалистам в данной области и включают, например, центрифугирование и аффинную хроматографию; методы получения очищенного вируса гепатита C описаны ниже.
II. Описание изобретения
При осуществлении настоящего изобретения будут использованы, если не указано специально, традиционные методы молекулярной биологии, микробиологии, рекомбинантной ДНК и иммунологии, известные специалистам в данных областях. Такие методы полностью разъяснены в литературе. Смотрите следующие источники: Maniatis, Fitsch & Sambrook, Molecular Cloning; A Laboratory Manual (1982); DNA Cloning, Volumes I and II (D.N. Glover ed. 1985); Oligonucleotide Synthesis (M. J. Gait ed. 1984); Nucleic Acid Hybridization (B.D. Harries & S.J. Higgins eds. 1984); Transcription and Translation (B.D. Hames & S. J. Higgins eds. 1984); Animal Cell Culture (R.I. Freshney ed. 1986); Immobilized Cells and Enzymes (IRL Press, 1986); В. Perbal, A Practical Guide to Molecular Cloning (1984); the series. Methods in Enzymology (Academic Press, Inc.); Gene Transfer Vectors for Mammalian Cells (J.H. Miller and M. P. Calos eds. 1987, Cold Spring Harbor Laboratory), Methods in Enzymology Vol. 154 and Vol. 155 (Wu and Grossman, and Wu, eds., respectively), Mayer and Walker, eds. (1987), Immunochemical Methods in Cell and Molecular Biology (Academic Press, London), Scopes, (1987), Protein Purification: Principles and Practice, Second Edition (Springer-Verlag, N.Y.), and Handbook of Experimental Immunology, Volumes I-IV (D.M. Weir and C.C. Blackwell eds, 1986).
Все патенты, патентные заявки и публикации, упомянутые ранее и ниже, включены в библиографический перечень.
Полезные материалы и процессы согласно настоящему изобретению сделали возможным получение семейства близко гомологических последовательностей нуклеотидов, обособленных из пула кольцевой ДНК, производных от последовательностей нуклеиновых кислот, присутствующих в плазме инфицированных вирусом гепатита C шимпанзе. Это семейство нуклеотидных последовательностей происходит не от человека или шимпанзе, поскольку оно не гибридизирует геномную ДНК от неинфицированных индивидуумов ни человека, ни шимпанзе, поскольку нуклеотиды этого семейства последовательностей присутствуют только в печени и плазме шимпанзе с инфекцией вируса гепатита C и поскольку последовательность не присутствует в банке генов. В дополнение семейство последовательностей не показывает существенной гомологии с последовательностями, содержащимися в геноме вируса гепатита В.
Последовательность одного члена семейства, содержащегося в клоне 5-1-1, имеет одну непрерывную открыто читаемую структуру, которая кодирует полипептид примерно с 50 аминокислотами. Сыворотка лиц, инфицированных вирусом гепатита C, содержит антитела, связанные с этим полипептидом, тогда как сыворотка от неинфицированного человека не содержит антител к этому полипептиду. И, наконец, в то время как сыворотка от неинфицированных шимпанзе не содержит антител к этому полипептиду, антитела вырабатываются у шимпанзе вследствие острой инфекции вируса ни A, ни B. Кроме того, антитела к этому полипептиду не определяются у шимпанзе и людей, инфицированных вирусом гепатита A и вирусом гепатита B. По этим критериям последовательность является кольцевой ДНК к вирусной последовательности, при этом вирус вызывает или связан с гепатитом ни A, ни B; эта последовательность показана на фиг. 1. Как обсуждалось выше, последовательность кольцевой ДНК в клоне 5- 1-1 отличается от последовательностей других выделенных кольцевых ДНК тем, что она содержит 28 дополнительных пар оснований.
Состав других идентифицированных членов семейства кольцевой ДНК, которые были выделены с использованием в качестве пробы синтетического эквивалента последовательности к фрагменту кольцевой ДНК в клоне 5-1-1, представлен на фиг. 3. Член семейства кольцевой ДНК, который был выделен с использованием синтетической последовательности, производной от кольцевой ДНК в клоне 81, представлен на фиг. 5, и состав этой последовательности с последовательностью клона 81 показан на фиг. 6. Другие члены семейства кольцевой ДНК, включая таковых, присутствующих в клонах 12f, 14i, 11b, 7f, 7e, 8h, 33c, 40b, 37b, 35, 36, 81, 32, 33b, 25c, 14c, 8f, 33f и 33g, 39c, 35f, 19g, 26g и 15e описаны в разделе IV.A. Состав кольцевых ДНК в этих клонах описан в разделе IV.A.19 и показан на фиг. 32. Состав кольцевой ДНК показывает, что она содержит один непрерывный открыто читаемый участок структуры и таким образом кодирует полипротеин. Эти данные согласуются с предположением, рассмотренным выше, что вирус гепатита C является флавивирусом или флавиподобным вирусом.
Наличие этого семейства кольцевых ДНК, представленных на фиг. 1-32 включительно, позволяет составление проб ДНК и полипептидов, полезных в диагностике гепатита ни A, ни B, вызванного инфекцией вируса гепатита C, и при массовой проверке донорской крови и крови реципиентов, а также продуктов крови на инфекцию. Например, из последовательностей возможно синтезировать олигомеры ДНК с 8-10 нуклеотидами или больше, полезные в качестве гибридизационных проб для определения наличия вирусного генома, например в сыворотке лиц, подозрительных на носителей вируса, или для проверки крови больного на присутствие вируса. Семейство последовательностей кольцевой ДНК позволяет также проектирование и получение специфических полипептидов вируса гепатита C, полезных в качестве диагностических средств для определения наличия антител, образующихся при гепатите ни A, ни B. Антитела к очищенным полипептидам, полученным из кольцевых ДНК, могут быть также использованы для определения вирусных антигенов у инфицированных лиц и в крови.
Знание этих последовательностей кольцевой ДНК делает возможным также проектирование и получение полипептидов, которые могут быть использованы в качестве вакцин против вируса гепатита C, а также для получения антител, которые в свою очередь могут быть использованы для защиты от болезни и/или лечения инфицированных вирусом гепатита C индивидов.
Кроме того, семейство последовательностей кольцевой ДНК позволяет дальнейшее определение свойств генома вируса гепатита C. Полинуклеотидные пробы, полученные из этих последовательностей, могут быть использованы для проверки пулов кольцевой ДНК на дополнительное перекрытие последовательностей кольцевой ДНК, которые в свою очередь могут быть использованы для получения еще последовательностей с перекрытым генетическим кодом. Несмотря на то что геном разделен на сегменты и сегментов недостает в общих последовательностях, этот метод может быть использован для получения последовательности целого генома. Однако если геном разделен на сегменты, то другие сегменты генома могут быть получены повторением процедуры серологического скрининга лямбда-gt11, использованной для выделения описанных здесь клонов кольцевой ДНК, или альтернативно выделением генома из очищенных частиц вируса гепатита C.
Семейство последовательностей кольцевой ДНК и полипептидов, производных от этих последовательностей, а также антитела, направленные против этих полипептидов, полезны также при выделении и идентификации вирусного агента(ов) BB ни A, ни B. Например, антитела, направленные против эпитопов вируса гепатита C, содержавшихся в полипептидах, полученных из кольцевых ДНК, могут быть использованы в способах, основанных на аффинной хроматографии для выделения вируса. Альтернативно антитела могут быть использованы для идентификации вирусных частиц, выделенных другими методами. Вирусные антигены и геномный материал выделенных вирусных частиц могут быть затем исследованы далее для определения их характеристик.
Информация, полученная от дельнейших исследований генома(ов) вируса гепатита C, а также дальнейшие характеристики антигенов вируса гепатита C и характеристики генома позволяют проектирование и синтез дополнительных проб и полипептидов, и антител, которые могут быть использованы для диагностики, профилактики и лечения гепатита ни A, ни B, индуцированного вирусом гепатита C, и для скрининга инфицированной крови и родственных крови продуктов.
Наличие проб на вирус гепатита C, включая антигены и антитела, и полинуклеотидов, производных от генома, от которого получено семейство последовательностей кольцевой ДНК, позволяет также развитие систем культуры тканей, которые найдут основное использование в выяснении биологии вируса гепатита C. Это в свою очередь может привести к усовершенствованию новых режимов лечения, основанных на противовирусных соединениях, которые преимущественно ингибируют репликацию или инфекцию вируса гепатита C.
Метод, использованный для идентификации и выделения этиологического агента гепатита ни A, ни B, является новым, и он может быть применен для идентификации и/или выделения до сих пор неохарактеризованных агентов, которые содержат геном и которые связаны с различными болезнями, включая болезни, вызываемые вирусами, вироидами, бактериями, грибками и паразитами. В этом методе пул кольцевой ДНК был создан из нуклеиновых кислот, присутствующих в инфицированной ткани инфицированного индивида. Пул был создан в векторе, позволяющем экспрессию полипептидаз, закодированных в кольцевой ДНК. Клоны клеток хозяина, содержащие вектор, которые производит иммунологически реактивный фрагмент полипептида этиологического агента, были выбраны иммунологическим просеиванием продуктов экспрессии пула с антителом, содержащим компонент тела от другого индивида, ранее инфицированного предполагаемым возбудителем. Операции метода иммунологического скрининга включали взаимодействие продуктов экспрессии, содержащей векторы кольцевой ДНК с антителом, содержащим в компоненте тела второго инфицированного индивида, и определение образования комплекса антитело - антиген между продуктом(ами) экспрессии и антителами второго инфицированного индивида. Выделенные клоны подвергали дальнейшему иммунологическому отсеиванию взаимодействием их продуктов экспрессии с антителом, содержащимся в компонентах тела других индивидуумов, инфицированных предполагаемым агентом, и с контрольными индивидами, неинфицированными предполагаемым агентом, и определяли образование комплексов антиген - антитело с антителами от инфицированных индивидов; и выделяли векторы, содержащие кольцевые ДНК, которые кодировали полипептиды, иммунологически взаимодействующие с антителами от инфицированных индивидов и индивидов, подозрительных на инфекцию, но не с контрольными индивидами. Инфицированные индивиды, используемые для построения пула кольцевой ДНК и для иммунологического скрининга, необязательно должны быть одного и того же вида.
Выделенные кольцевые ДНК как результат настоящего метода и продукты их экспрессии и антитела, направленные против продуктов экспрессии, полезны для получения характеристик и/или улавливания этиологического агента. Как описано более детально ниже, этот метод успешно использовали для выделения семейства кольцевых ДНК, полученных из генома вируса гепатита C.
II.A. Получение последовательности кольцевой ДНК
Депонированную сыворотку от шимпанзе с хронической инфекцией вируса гепатита C и содержащую высокий титр вируса, т.е. по меньшей мере 106 инфекционных доз шимпанзе/мл (ИДШ/мл), использовали для выделения вирусных частиц; нуклеиновые кислоты, выделенные из этих частиц, использовали в качестве матрицы при конструировании пула кольцевой ДНК к вирусному геному. Процесс выделения частиц предполагаемого вируса гепатита C и конструирование пула кольцевой ДНК в лямбда-gt11 описан в разделе IV.A.1. Лямбда-gt11 является вектором, который был специально разработан для экспрессии встроенных кольцевых ДНК как (конденсированных) полипептидов слияния с бета-галактозидазой и для просеивания большого числа рекомбинантного фага специфической антисывороткой, выведенной против определенного антигена. Пул лямбда-gt11 кольцевой ДНК, выработанный из пула кольцевой ДНК, содержащего кольцевую ДНК со средним размером приблизительно 200 пар оснований, исследовали на закодированные эпитопы, которые должны связываться специфически с сыворотками, полученными от больных, перенесших ранее бытовой гепатит ни A, ни B. Huynh, T. V. et al., (1985). Было просеяно примерно 106 фагов и идентифицировано пять положительных фагов, очищено и затем испытано на специфичность связывания с сыворотками от различных людей и шимпанзе, прежде инфицированных возбудителем вируса гепатита C. Один из фагов, а именно 5-1-1, связывал пять из восьми испытываемых сывороток человека. Это связывание казалось избирательным для сывороток больных с предшествующими инфекциями гепатита ни A, ни B, поскольку семь сывороток нормальной донорской крови не показали такого связывания.
Последовательность кольцевой ДНК в рекомбинантном фаге 5-1-1 была определена и представлена на фиг. 1. Полипептид, закодированный этой клоновой кольцевой ДНК, которая находится в той же трансляционной структуре, что и N-концевая бета-галактозидазная часть молекулы (конденсированного) полипептида слияния, показан над нуклеотидной последовательностью. Эта трансляционная открыто читаемая структура, таким образом, кодирует эпитоп(ы), специфически распознаваемые сывороткой больных, перенесших инфекции гепатита ни A, ни B.
Наличие кольцевой ДНК в рекомбинантном фаге 5-1-1 позволило выделение других клонов, содержащих дополнительные сегменты и/или альтернативные сегменты кольцевой ДНК к вирусному геному. Пул кольцевой ДНК лямбда-gt11, описанный выше, исследовали с использованием синтетического полинуклеотида, производного от последовательности клоновой кольцевой ДНК 5-1-1. Этот скрининг дал три других клона, которые были идентифицированы как 81, 1-2 и 91; кольцевые ДНК, содержащиеся в этих клонах, были упорядочены, т.е. определена их последовательность. См. разделы IV.A.3 и IV.A.4. Гомологи между четырьмя самостоятельными клонами показаны на фиг. 2, на которой гомологи указаны вертикальными линиями. Последовательности нуклеотидов, присутствующих уникально в клонах 5-1-1, 81 и 91, показаны мелкими буквами.
Клоновые кольцевые ДНК, присутствующие в рекомбинантных фагах в клонах 5-1-1, 81, 1-2 и 91, являются высокогомологичными и отличаются только двумя участками. Во-первых, нуклеотидный номер 67 в клоне 1-2 является тимидином, в то время как остальные три клона содержат цитидиновый остаток в этом положении. Это замещение, однако, не изменяет природы закодированной аминокислоты.
Второе отличие между клонами заключается в том, что клон 5-1-1 содержит 28 пар оснований в своем 5'-терминале, которые не присутствуют в других клонах. Дополнительная последовательность может быть 5'-концевым артефактом клонирования; 5'-концевые артефакты клонирования обычно наблюдаются в продуктах методов, связанных с кольцевой ДНК.
Синтетические последовательности, производные от 5'-участка и 3'-участка кольцевой ДНК вируса гепатита C в клоне 81 были использованы для отсеивания и выделения кольцевых ДНК из пула кольцевой ДНК вируса гепатита ни A, ни B лямбда-gt11, которые перекрывают кольцевую ДНК клона 81. (Раздел IV.A.5). Последовательности полученных кольцевых ДНК, которые находятся в клоне 36 и клоне 32, соответственно представлены на фиг. 5 и 7.
Аналогично этому синтетический полинуклеотид, базирующийся на 5'-участок клона 36, был использован для отсеивания и выделения кольцевых ДНК из пула кольцевой ДНК вируса гепатита ни A, ни B лямбда-gt11, которые перекрывали кольцевую ДНК клона 36 (раздел IV.A.8). Очищенный клон рекомбинантной, содержащей фаг кольцевой ДНК, который гибридизировал синтетическую полинуклеотидную пробу, был назван клоном 35, и последовательность кольцевой ДНК вируса ни A, ни B в этом клоне представлена на фиг. 8.
При использовании метода выделения перекрывающих последовательностей кольцевой ДНК были получены клоны, содержащие дополнительные восходящие и нисходящие последовательности кольцевой ДНК вируса гепатита C. Выделение этих клонов описано ниже в разделе IV.A.
Анализ нуклеотидных последовательностей кольцевых ДНК вируса гепатита C в выделенных клонах показывает, что композиционная кольцевая ДНК, закодированная в выделенных клонах, содержит один длинный участок открыто читаемой структуры. На фиг. 26 представлена последовательность композиционной ДНК из этих клонов вместе с полипептидом предполагаемого вируса гепатита C, закодированным в ней.
Описание метода реабилитации последовательностей кольцевой ДНК представляет собой в основном исторический интерес. Полученные последовательности (и их комплементы) представлены в описании, и последовательности или любая их часть могут быть получены синтетическими методами или сочетанием методов синтеза с восстановлением частичных последовательностей, используя методы, аналогичные описанным здесь методам.
Штаммы лямбда-gt11, реплицированные из пула кольцевой ДНК вируса гепатита C и из клонов 5-1-1, 81, 1-2 и 91, депонированы в Американском собрании типов культур (АТСС) N 12301 Parklawn Dr., Rockville, Maryland 20852, и им присвоены возрастающие номера (в терминах Budapest Treaty) представленные в табл. A.
При принятии и регистрации этой заявки как патента Соединенных Штатов все ограничения по наличию этих депозитов будут безусловно устранены и доступ к указанным депо будет обеспечен на период рассмотрения упомянутой заявки лицу, определенному на это уполномоченным, который указан в 37 CFR 1.14 и 35 USC 1.22. Кроме того, эти депонированные штаммы будут сохранены в течение 30 лет, считая от даты депонирования, или спустя пять лет после последнего обращения за депонированными культурами, или в течение времени действия патента США, если не дольше, эти и другие депонированные материалы, упомянутые в описании, приведены только для удобства, а не потому, что требуются для осуществления настоящего изобретения ввиду прилагаемого описания. Последовательности кольцевой ДНК вируса гепатита C во всех депонированных материалах приведены в библиографической ссылке.
Описание выше "перехода" генома при выделении перекрывающих последовательностей кольцевой ДНК из пула лямбда-gt11 вируса гепатита C обеспечивает один метод, которым могут быть выделены кольцевые ДНК соответствующего целому геному вируса гепатита C. Однако, помимо приведенной в описании информации, существуют другие методы выделения кольцевых ДНК, очевидные для специалистов в данной области. Некоторые из этих методов описаны в разделе IV.A ниже.
II.B. Получение вирусных полипептидов и их фрагментов
Наличие последовательностей кольцевой ДНК либо выделенных с использованием последовательностей ДНК на фиг. 1-32, как обсуждено ниже, а также последовательностей кольцевой ДНК на этих фигурах позволяет конструирование векторов экспрессии, кодирующих антигенно активные участки полипептида, закодированного в одной из нитей. Эти антигенно активные участки могут быть получены из внешнего покрова или оболочки антигенов, или из сердцевины антигенов, включая, например, полинуклеотидсвязывающие белки, полинуклеотидную полимеразу(ы) и другие вирусные белки, требующиеся для репликации и/или сборки вирусных частиц. Фрагменты, кодирующие желаемые полипептиды, получены из клонов кольцевой ДНК традиционным ограниченным сбраживанием или синтетическими методами и лигированы в векторы, которые могут, например, содержать части последовательностей слияния, таких как бета-галоктозидаза или пероксиддисмутаза, предпочтительно пероксиддисмутаза. Методы и векторы, полезные для получения полипептидов, содержащих последовательности слияния пероксиддисмутазы, описаны в публикации Европейского патентного ведомства N 0196056 от 1 октября 1986 года. Векторы, кодирующие полипептиды слияния (синтетические полипептиды) пероксиддисмутазы и полипептиды вируса гепатита C, т. е. ни A, ни B5-1-1, ни A, ни B81 и C100-3, который закодирован в композиционных кольцевых ДНК вируса гепатита C, описаны в разделах IV.B.1, IV. B. 2 и IV.B.4 соответственно. Любая желаемая часть кольцевой ДНК вируса гепатита C, содержащая открыто читаемую структуру в одной из чувствительных нитей, может быть получена как рекомбинантный полипептид, такой как зрелый белок или белок слияния; альтернативно полипептид, закодированный в кольцевой ДНК, может быть получен химическим синтезом.
ДНК, кодирующая желаемый полипептид либо в слитой или в зрелой форме, содержащий или не содержащий сигнальную последовательность, чтобы разрешить секрецию, может быть лигирован в векторы экспрессии, пригодные для любого традиционного хозяина. Как эукариотный, так и прокариотный типы хозяина используют теперь при формировании рекомбинантных полипептидов, и кратко некоторые общие контрольные системы и линии клеток хозяина описаны в разделе III. A ниже. Затем полипептид выделяют из лизированных клеток или их культуральной среды и очищают до степени, необходимой для его последующего использования. Очистка может быть проведена известными в данной области методами, например, солевое фракционирование, хроматография на ионообменных смолах, центрифугирование и тому подобные. См., например, "Методы в энзимологии для различных способов очистки белков. "Такие полипептиды могут быть использованы как диагностические средства, или те из них, которые способствуют нейтрализации антител, могут быть использованы для приготовления вакцин. Антитела, образующиеся против этих полипептидов, могут быть также использованы как диагностические средства или для пассивной иммунотерапии. В дополнение, как описано ниже в разделе II.J, антитела к этим полипептидам полезны при выделении и идентификации частиц вируса гепатита C.
Антигены вируса гепатита C могут быть выделены из вирионов вируса гепатита C. Вирионы могут быть выращены в инфицированных вирусом гепатита C клетках в культуре ткани или в инфицированном хозяине.
II.C. Получение антигенных полипептидов и соединение их с носителем
Антигенный участок полипептида, в общем, относительно мал - типично 8-10 аминокислот или менее в длину. Фрагменты менее пяти аминокислот могут характеризовать антигенный участок. Эти сегменты могут соответствовать участкам антигена вируса гепатита C. Следовательно, используя кольцевые ДНК вируса гепатита C как основу, можно получить кодируемые дезоксирибонуклеиновой кислотой короткие сегменты полипептидов вируса гепатита C рекомбинантной экспрессией, либо как белки слияния, либо как обособленные полипептиды. В дополнение, короткие аминокислотные последовательности могут быть удобно получены химическим синтезом. В случае, когда синтезированный полипептид имеет точную конфигурацию, чтобы дать корректный эпитоп, но слишком мал, чтобы быть иммуногенным, полипептид может быть связан с подходящим носителем.
Ряд методов получения такой связи известен в данной области, включая образование бисульфидных связей с использованием N-сукцинимидил -3-(2- пиридилтио)-пропионата и сукцинимидил -4-(N- малеимидометил)-циклогексан -1-карбоксилата, поставляемых Pierce Company, Rockford, Illinois (если в пептиде отсутствует сульфгидрильная группа, то это можно получить добавлением цистеинового остатка). Эти реагенты образуют бисульфидную связь между ними и пептидными цистеиновыми остатками на одном белке и амидную связь через эпсилон-амино на лизин, или другую свободную аминогруппу в другом белке. Разнообразие таких дисульфид/амидобразующих средств известно. См., например, Immun. Rev., (1982), 62:185. Другие бифункциональные связующие агенты образуют тиоэфиры скорее, чем бисульфидную связь. Многие из этих тиоэфиробразующих средств имеются в продаже и включают реакционноспособные сложные эфиры 6-малеимидокапроновой кислоты, 2-бромуксусной кислоты, 2-йодуксусной кислоты, 4-(N-малеимидометил) -циклогексан-1- карбоновой кислоты и тому подобные. Карбоксильные группы могут быть активированы сочетанием их с сукцинимидом или с натриевой солью 1-гидроксил- 2-нитро-4-сульфоновой кислоты. Перечисленный выше список не является исчерпывающим, и могут быть использованы модификации приведенных в перечне соединений.
Может быть использован любой носитель, который сам не вызывает образование антител, вредных для хозяина. Подходящие носители обычно являются крупными, медленно метаболизирующимися макромолекулами, такими как белки, полисахариды, такие как функционализированная латексом сефароза, агароза, целлюлоза, целлюлозные шарики и тому подобные; полимерные аминокислоты, такие как полиглутаминовая кислота, полилизин и тому подобные; сополимеры аминокислот и неактивные вирусные частицы (см., например, раздел II.D.). Особенно полезными белковыми субстратами являются альбумины сыворотки, гемоцианин, молекулы иммуноглобулина, тироглобулин, овальбумин, тетанусанатоксин и другие белки, известные специалистам в данной области.
II. D. Получение гибридных частиц иммуногенов, содержащих эпитопы вируса гепатита C
Иммуногенность эпитопов вируса гепатита C может быть также повышена получением их в системах млекопитающих или ферментных системах, слитых или собранных с образующими частицы белками, таких как, например, система, связанная с поверхностным антигеном гепатита B. Полученный в ней эпитоп вируса ни A, ни B связан непосредственно с образующим частицы белком, кодирующим последовательности, продуцирующие гибриды, которые являются иммуногенными по отношению к эпитопу вируса гепатита C. В дополнение все полученные векторы включают эпитопы, специфические к вирусу гепатита B, имеющие различную степень иммуногенности, такие как, например, пре-S пептид. Таким образом, частицы, построенные из образующего частицы белка, которые включают последовательности вируса гепатита C, иммуногенны в отношении к вирусу гепатита C и вирусу гепатита B.
Поверхностный антиген гепатита, как было установлено, был образован и собран в частицы в S.cerevisiae (Valenzuela P. и соавт., 1982 г.), а также в, например, клетках молочной железы (Valenzuela P., и соавт., 1984 г.). Образование таких частиц, как было показано, повышает иммуногенность мономерной субъединицы. Сборки могут также включать иммунодоминатный эпитоп поверхностного антигена гепатита B, содержащий 55 аминокислот предповерхностного (пре-S) участка. (Neurath и соавт., 1984 г.). Сборки частицы пре-S поверхностного антигена гепатита B, синтезированные в дрожжах, описаны в публикации Европейского патентного ведомства N 174444 от 19 марта 1986 г.; гибриды, включающие гетерологические вирусные последовательности для ферментной экспрессии, описаны в публикации Европейского патентного ведомства N 175261 от 26 марта 1986 г. Обе заявки принадлежат настоящему заявителю и приведены в библиографической ссылке. Эти сборки частиц могут быть также осуществлены в клетках млекопитающих, таким как яйцеклетки китайского хомяка, используя SV 40-дигидрофолятредуктазный вектор (Michelle et al., 1984 г.).
В дополнение, части образующего частицы белка, кодирующие последовательность, могут быть замещены кодонами, кодирующими эпитоп вируса гепатита C. В этом замещении участки, не требующиеся для посредничества в агрегации элементов (единиц) для образования иммуногенных частиц в дрожжах или у млекопитающих, могут быть стерты, устраняя таким образом дополнительные антигенные локализации вируса гепатита B от конкуренции с эпитопом вируса гепатита C.
II.E. Получение вакцин
Вакцины могут быть получены из одного или более иммуногенных полипептидов, полученных из кольцевой ДНК вируса гепатита C, а также из последовательностей кольцевой ДНК на фиг. 1-32 или из генома вируса гепатита C, к которому они относятся. Наблюдаемая гомология между вирусом гепатита C и флавивирусами дает информацию, касающуюся полипептидов, которые наиболее вероятно подходят для вакцин по своей эффективности, а также участков генома, на которых они закодированы. Общая структура генома флавивирусов описана Rice и соавт., 1986 г. РНК генома флавивирусов, по-видимому, является только вирусоспецифичным штаммом информационной РНК и транслирована в три вирусных структурных белка, т.е. C, М и E, а также как и в два крупных неструктурных белка, NV4 и NV5 и сложный ряд более мелких неструктурных белков. Известно, что основные нейтрализующие эпитопы для флавивирусов в оболочке E-белка (Roehrig, 1986 г.). Соответствующий E-ген вируса гепатита C и полипептидный кодирующий участок могут быть предсказаны на основе гомологии с флавивирусами. Таким образом, вакцины могут состоять из рекомбинантных полипептидов, содержащих эпитопы E вируса гепатита C. Эти полипептиды могут быть получены в бактериях, дрожжах или в клетках млекопитающих или альтернативно могут быть выделены из вирусных препаратов. Следует также ожидать, что и другие структурные белки могут также содержать эпитопы, которые образуют защитные антитела антивирусу гепатита C. Таким образом, полипептиды, содержащие эпитопы E, C и М, могут быть также использованы отдельно или в сочетании в вакцинах вируса гепатита C.
В дополнение к изложенному выше было показано, что иммунизация неструктурным белком 1 (NS1) приводит к защите от желтой лихорадки (Schlesinger и соавт. , 1986 г.). Это верно даже если иммунизация не приводит к образованию нейтрализованных антител. Таким образом, особенно с этим белком, по-видимому, высококонсервированы среди флавивирусов, это лучше, чем NS1 вируса гепатита C защищает против инфицирования вирусом гепатита C. Кроме того, это также показывает, что неструктурированные белки могут обеспечивать защиту против вирусной патогенности, даже если они не влияют на продуктивность нейтрализации антител.
Ввиду вышесказанного поливалентные вакцины против вируса гепатита C могут состоять из одного или более структурных белков и/или одного или более неструктурных белков. Эти вакцины могут содержать, например, рекомбинантные полипептиды вируса гепатита C и/или полипептиды, выделенные из вирионов. В дополнение возможно использование инактивированных вируса гепатита C в вакцинах; инактивация может быть осуществлена при получении вирусного лизата или иными средствами, известными в данной области, чтобы вызвать инактивацию флавивирусов, например, обработкой органическими растворителями или детергентами или обработкой формалином. Кроме того, вакцины могут быть получены из ослабленных штаммов вируса гепатита C. Получение аттенуированных штаммов вируса гепатита C описано ниже.
Известно, что некоторые белки флавивирусов содержат высококонсервированные участки, т.е. ожидается некоторая иммунологическая перекрестная реактивность между вирусом гепатита C и другими флавивирусами. Возможно, что поделенные эпитопы между флавивирусами и вирусом гепатита C вызовут образование защитных антител против одного или более расстройств, обусловленных этими патогенными возбудителями. Таким образом, возможно проектирование многоцелевых вакцин, основываясь на этих знаниях.
Получение вакцин, содержащих иммуногенный полипептид(ы) в качестве активных ингредиентов, известно специалистам в данной области. Типично такие вакцины готовят в форме препаратов для инъекции либо в виде жидких растворов или суспензий; твердые формы, пригодные для растворения или суспендирования в жидкости до проведения инъекции, могут быть также приготовлены. Препарат может быть эмульгирован или белок инкапсулирован в липосомы. Активные иммуногенные ингредиенты часто смешивают с носителями, которые являются фармацевтически приемлемыми и совместимыми с активным ингредиентом. Подходящими наполнителями являются, например, вода, физиологический раствор, декстроза, глицерин, этанол и тому подобные и их сочетания. В дополнение, если необходимо, вакцина может содержать небольшие количества вспомогательных веществ, таких как смачивающие или эмульгирующие средства, pH буферные средства и/или добавки, повышающие эффективность вакцины. Примеры адъювантов, которые могут быть эффективными, включают, но не ограничиваются этим: гидроокись алюминия, N-ацетил-мурамил-L- треонил-D-изоглутамин, N-ацетил-нор-мурамил-L -аланил-D-изоглутамин (CGP 11637, называемый далее нор-MDP), N-ацетил-мурамил-L- аланил-D -изоглутаминил-L-аланин-2-(1'-2' -дипальмитоил-sn- глицеро-3- гидроксифосфорилокси)-этиламин (CGP 19835A, называемый далее как MTP-PE) и RIBI, который содержит три компонента, экстрагированных из бактерий: монофосфориловый липид A, димиколат трехалозы и скелет клеточных оболочек (MPL + TDM + CWS) в 2% эмульсии сквалена в твине 80. Эффективность адъюванта может быть определена измерением количества антител, направленных против иммуногенного полипептида, содержащего антигенную последовательность вируса гепатита C, полученную от введения этого полипептида в вакцины, которые включали также различные адъюванты.
Вакцины традиционно вводили парентерально, инъекциями, например либо подкожно, либо внутривенно, или внутримышечно. Дополнительные формы медицинских препаратов, пригодных для введения другими путями, включают суппозитории и в некоторых случаях - формы для введения перорально. Суппозитории, традиционные связующие и носители могут включать, например, полиалкиленгликоли или триглицериды; такие суппозитории могут быть получены из смесей, содержащих активный ингредиент в пределах от 0,5 до 10%, предпочтительно 1-2%. Формы препаратов для перорального введения включают такие обычно применяемые эксципиенты, как например маннит фармацевтической чистоты, лактоза, крахмал, стеарат магния, сахарин натрия, целлюлоза, карбонат магния и тому подобные. Этим композициям придают форму растворов, суспензий, таблеток, пилюль, капсул, формы с длительным освобождением активного ингредиента или порошков и содержит 10-95% активного ингредиента, предпочтительно 25-70%.
Белки могут быть приготовлены как вакцины в нейтральной форме или в форме соли. Фармацевтически приемлемые соли включают кислотно-аддитивные соли (образованы со свободными аминогруппами пептида) и которые образованы с неорганическими кислотами, такими как, например, хлористо-водородная или фосфорная кислоты, или такими органическими кислотами, как уксусная, щавелевая, винная, малеиновая и тому подобные. Соли, образованные со свободными карбоксильными группами, могут быть также получены от неорганических оснований, таких как, например, гидроокиси натрия, калия, аммония, кальция или железа и такими органическими основаниями, как изопропиламин, триметиламин, 2-этиламиноэтанол, гистидин, прокаин и тому подобные.
II.F. Дозирование и введение вакцин
Вакцины вводили способом, совместимым с дозированными формами препаратов, и в таких количествах, которые были бы диагностически и/или терапевтически эффективными. Количество вводимой вакцины, которое обычно находится в пределах от 5 до 250 микрограмм антигена на дозу, зависит от субъекта, подлежащего лечению, функциональной активности иммунной системы субъекта синтезировать антитела и степени желаемой защиты. Точные количества активного ингредиента, потребные для введения, могут зависеть от заключения лечащего врача и могут быть строго индивидуальными для каждого субъекта.
Вакцина может быть предписана для введения в однократной дозе или предпочтительно в множественных дозах. При назначении многократной дозы первичный курс вакцинации может заключать 1-10 отдельных доз с последующим введением других доз в известном интервале времени, потребного для поддержания и/или усиления иммунной реакции, например с 1-4-месячным интервалом для второй дозы и, если необходимо, последующие дозы(а) - после нескольких месяцев. Режим дозирования будет также, по меньшей мере частично, зависеть от заключения лечащего врача.
В дополнение вакцина, содержащая иммуногенные антигены (антиген) вируса гепатита C, может быть введена в сочетании с другими иммунорегулирующими средствами, например иммуноглобулином.
II.G. Получение антител против эпитопов вируса гепатита C
Иммуногенные полипептиды, полученные как описано выше, использовали для продуцирования антител как поликлональных, так и моноклональных. Если необходимы поликлональные антитела, то выбранное млекопитающее (т.е. мышь, кролик, коза, лошадь и так далее) иммунизировали иммуногенным пептидом, несущим эпитоп(ы) вируса гепатита C. Сыворотку иммунизированного животного собирали и обрабатывали известными методами. Если сыворотка, содержащая поликлональные антитела к эпитопу вируса гепатита C, содержит антитела к другим антигенам, то поликлональные антитела могут быть очищены
иммуноаффинной хроматографией. Методы получения и обработки поликлональной антисыворотки известны в данной области, см., например, Mayer и Walker 1987 г.
Альтернативно поликлональные антитела могут быть выделены из млекопитающего, которое ранее было инфицировано вирусом гепатита C. Пример очистки антител к эпитопам вируса гепатита C из сыворотки от неинфицированного индивида, основанный на аффинной хроматографии и использовании полипептида сплавления пероксида дисмутазы и полипептида, закодированного в кольцевой ДНК клона 5-1-1, представлен в разделе V.E.
Моноклональные антитела, направленные против эпитопов вируса гепатита C, могут быть также легко получены специалистом в данной области. Общая методология получения моноклональных антител гибридизацией хорошо известна. Жизнедеятельные линии клеток-продуцентов антител могут быть созданы слиянием клеток, а также и другими методами, такими как прямая трансформация B лимфоцитов онкогенной ДНК или трансфекция вирусом Эпстайна-Барра. См. также M. Schreier и соавт., 1980; Hammerling и соавт., 1981; Kennett и соавт. 1980 г. ; см. также патенты США NN 4341761, 4399121, 4427783, 4444887, 4466917, 4472500, 4491632 и 4493890. Перечень моноклональных антител, продуцированных против эпитопов вируса гепатита C, может быть исследован на различные свойства, т.е. на изотип, сродство к эпитопу и так далее.
Антитела, как моноклональные, так и поликлональные, которые направлены против эпитопов вируса гепатита C, особенно полезны в диагностике, а нейтрализованные антитела полезны в пассивной иммунотерапии. Моноклональные антитела, в частности, могут быть полезны при образовании антиидиотипичных антител.
Антиидиотипичные антитела являются иммуноглобулинами, которые несут "внутреннее изображение" антигена инфекционного агента, от которого желательна защита. См., например, Nisonoff A. и соавт., 1981, и Dreesman и соавт., 1985 г.
Метолы вырабатывания антигенотипичных антител известны в данной области. См., например, Grzych (1985), MacNamara и соавт. (1984) и Uytdehaag и соавт. (1985). Эти антигенотипичные антитела могут быть также полезны при лечении гепатита ни A, ни B, а также для выяснения иммуногенных участков антигенов вируса гепатита C.
II.H. Диагностические олигонуклеотидные пробы и комплекты
Используя раскрытые в заявке части выделенной кольцевой ДНК вируса гепатита C за основу, включая и части кольцевой ДНК, представленные на фиг. 1-32, можно получить олигомеры примерно с 8 нуклеотидами и более, которые образуют гибриды с геномом вируса гепатита C и полезны для идентификации вирусного агента(ов), дальнейшей характеризации вирусного генома(ов), а также при определении вируса(ов) у заболевших индивидов. Пробы на полинуклеотиды (природные или производные) имеют длину, которая позволяет определение уникальных вирусных последовательностей гибридизацией. В то время как 6-8 нуклеотидов могут быть пригодной для работы длины, последовательность 10-12 нуклеотидов, однако, предпочтительна и оптимальной, по-видимому, является последовательность примерно 20 нуклеотидов. Предпочтительно, чтобы эти последовательности получали от участков, в которых отсутствует гетерогенность. Эти пробы могут быть получены с использованием обычных методов, включая методы автоматизированного синтеза олигонуклеотидов. Среди полезных проб, например, можно назвать клон 5-1-1 и дополнительные клоны, раскрытые в описании, а также различные олигомеры, полезные в апробации пулов кольцевой ДНК, представленных ниже. Комплемент к любой уникальной части генома вируса гепатита C будет удовлетворительным. Для использования в качестве проб желательна полная комплементарность, хотя она может быть необязательной, так как длина фрагмента увеличивается.
Для использования таких проб в качестве диагностических обрабатывают образец, который подлежит анализу, такой как кровь или сыворотка, если необходимо, для экстрагирования содержащихся в нем нуклеиновых кислот. Полученную из образца нуклеиновую кислоту можно подвергнуть гельэлектрофорезу или другому методу разделения по размерам; альтернативно образец нуклеиновой кислоты может быть радиографирован без разделения по размерам. Затем пробы метят. Подходящие метки и методы меченых проб известны в данной области и включают, например, радиоактивные метки, инкорпорированные при синтезе белка или ферментации, биотин, флюоресцентные пробы и хемолюминесцентные пробы. Нуклеиновые кислоты, экстрагированные из образца, обрабатывают затем мечеными пробами в условиях гибридизации с соответствующими мерами предосторожности. Пробы можно получить полностью комплементарными к геному вируса гепатита C. Поэтому соблюдают условия особой строгости, чтобы предотвратить ложные находки. Однако особо строгие условия должны соблюдаться только если пробы комплементарны к участкам вирусного генома, которые утратили гетерогенность. Строгость условий проведения гибридизации определяется рядом факторов в процессе самой гибридизации и при процедуре промывки, включающих температуру, ионную силу, длительность времени и концентрацию формамида. Эти факторы описаны, например, Maniatis, T., (1982 г.).
Обычно следует ожидать, что последовательности генома вируса гепатита C будут присутствовать в сыворотке инфицированных индивидов на относительно низких уровнях, т.е. примерно 102-103 последовательностей в мл. Этот уровень может потребовать использование методов усиления при гибридизационных исследованиях. Такие методы известны в данной области. Например, система "Bio-Bridge" Enzo биохимической корпорации использует концевую дезоксинуклеотидную трансферазу для добавления немодифицированных 3'-поли-dT-хвостов к пробе ДНК. Снабженная хвостами поли-dT-проба гибридизируется в последовательность нуклеотидной мишени, а затем в модифицированную биотином поли-A последовательность. Заявка по PCT N 84 (03520) и заявка на Европейский патент N 124221 описывают исследования по гибридизации ДНК, в которых: (1) аналит ренатурирован пробой однонитчатой ДНК, которая комплементарна меченому ферментом олигонуклеотиду; и (2) полученный хвостатый дуплекс гибридизируется в меченный ферментом олигонуклеотид. Заявка на Европейский патент N 204510 описывает исследования по гибридизации ДНК, в которых анализируемая ДНК контактирует с пробой, имеющей хвост, такой как поли-dT хвост, увеличенная нить которой имеет последовательность, которая гибридизирует к хвосту пробы, такой как последовательность поли-A, и которая способна связывать множество меченых нитей. Особенно желаемый метод может сначала вовлекать увеличение последовательностей мишени вируса гепатита C в сыворотках примерно в 10000 раз, т.е. примерно до 106 последовательностей на миллилитр. Это можно осуществить, например, по методу Saiki и соавт., (1986 г.). Увеличенная последовательность(и) может быть затем определена, используя гибридизационный анализ, который описан в продолжающейся заявке на патент США N 2300-0171, зарегистрированной 15 октября 1987 года на имя преемников по настоящей заявке и приведенной в библиографической ссылке. В этих исследованиях гибридизации, которые должны определять последовательности на уровне 106/мл, используются полимеры нуклеиновых кислот, которые связываются с анализируемой однонитчатой нуклеиновой кислотой и которые связывают также множество однонитчатых меченых олигонуклеотидов. Анализ фазы расслоения подходящего раствора, который может быть использован с мечеными полинуклеотидными пробами, и методы получения проб описаны в публикации о Европейском патенте N 225807, опубликованном 16 июня 1987 года, принадлежащем правопреемникам настоящей заявки, и который приведен в библиографической ссылке.
Пробы могут быть упакованы в диагностический комплект. Диагностические комплекты включают пробы ДНК, которые могут быть мечены; альтернативно пробы ДНК могут не иметь метки, а ингредиенты для введения метки могут быть включены в комплект. Комплект может содержать надежно упакованные реагенты и материалы, необходимые для конкретной гибридизации, например стандарты и инструкции по проведению теста.
II.I. Иммунологический анализ и диагностические комплекты
Полипептиды, которые взаимодействуют иммунологически с антителами сыворотки, содержащей вирус гепатита C, например, полипептиды, полученные от клонов, описанных в разделе IV.A, или закодированные в них и их композиты (см. раздел IV. A) и антитела, образованные против специфических эпитопов вируса гепатита C в этих полипептидах (см., например, раздел IV.E) являются полезными для проведения иммунологического анализа на определение наличия антител против вируса гепатита C, или присутствие вируса и/или вирусных антигенов в биологических образцах, включающих, например, образцы крови или сыворотки. Осуществление иммунологического анализа имеет дело с вариациями и их разнообразие известно специалистам в данной области. Например, в иммунологическом анализе можно использовать один вирусный антиген, например полипептид, полученный от любого из клонов, содержащих кольцевую ДНК вируса гепатита C, описанных в разделе IV.A, или от композиционных кольцевых ДНК, полученных от кольцевых ДНК в этих клонах, или от генома вируса гепатита C, от которого получена кольцевая ДНК в этих клонах; альтернативно в иммунологическом анализе можно использовать комбинацию вирусных антигенов, полученных из этих источников. Можно использовать, например, моноклональное антитело, направленное против одного вирусного антигена, моноклональные антитела, направленные против различных вирусных антигенов, поликлональные антитела, направленные против того же вирусного антигена или поликлональные антитела, направленные против различных вирусных антигенов. Анализы могут быть основаны, например, на конкуренции или непосредственной реакции или на анализах типа сендвич. В анализах могут быть использованы также, например, твердые подложки или могут быть с использованием иммунопреципитации. Большинство анализов включают использование меченых антител или полипептидов; метки могут быть, например, флюоресцентные, хемолюминесцентные, радиоактивные или молекулы красителя. Анализы, в которых усиливают сигналы от пробы, также известны и примерами таких анализов является использование биотина и авидина, и иммунологические анализы с мечеными ферментами и промежуточные анализы, такие как иммуноферментный твердофазный анализ.
Модель флавивируса для вируса гепатита C позволяет предсказания относительно вероятной локализации диагностических эпитопов для структурных белков пириона. Участки C, пре-М, М и E вероятно все содержат эпитопы значительного потенциала для детектирования вирусных антигенов и в особенности для диагностики. Аналогично этому локализации неструктурных белков, следует ожидать, содержат важные диагностические эпитопы (т.е. NS 5, кодирующий предполагаемую полимеразу, и NS 1, кодирующий предполагаемый комплементсвязывающий антиген). Рекомбинантные полипептиды или вирусные полипептиды, которые включают эпитопы от этих специфических локализаций, могут быть использованы для определения вирусных антител в инфекциях донорской крови и у инфицированных пациентов.
В дополнение антитела, направленные против белков E и/или М, могут быть использованы в иммунологических анализах для определения вирусных антигенов у больных с вирусом гепатита C, вызывающим гепатит ни A, ни B, и в инфекциях крови доноров. Кроме того, эти антитела могут быть чрезвычайно полезны при определении доноров и больных в острой фазе.
Комплекты удобны для иммунодиагностики и содержат соответствующим образом меченые реагенты, и укомплектованы необходимыми материалами, включающими упаковки полипептидов согласно изобретению, содержащих эпитопы вируса гепатита C или анализа, направленные против эпитопов вируса гепатита C в подходящих емкостях вместе с остальными реагентами, потребными для проведения анализа, а также ряд инструкций по проведению анализа.
II. J. Дальнейшая характеристика генома вируса гепатита C, вирионов и вирусных агентов, используя пробы, полученные от кольцевой ДНК, к вирусному геному
Информация о последовательностях кольцевой ДНК вируса гепатита C в клонах, описанных в разделе IV.A и показанных на фиг. 1-32 включительно, может быть использована для получения дальнейшей информации о последовательности генома вируса гепатита C и для идентификации и выделения агента вируса гепатита С и таким образом будет помогать в характеристике последовательности, включая природу генома, структуру вирусной частицы и природу антигенов, из которых она состоит. Эта информация, в свою очередь, может привести к дополнительным полинуклеотидным пробам, полипептидам, полученным из генома вируса гепатита C, и антителам, направленным против эпитопов вируса гепатита C, которые будут полезны для диагностики и/или лечения гепатита ни A, ни B, вызванного вируса гепатита C.
Информация о последовательности кольцевой ДНК в упомянутых выше клонах полезна для составления проб для выделения дополнительных последовательностей кольцевой ДНК, полученных из еще неопределенных участков генома(ов) вируса гепатита C, от которых получены кольцевые ДНК в клонах, в разделе IV. A. Например, меченые пробы, содержащие последовательность примерно 8 или более нуклеотидов и предпочтительно 20 или более нуклеотидов, которые получены из участков, примыкающих к 5'-окончаниям или 3'-окончаниям семейства последовательностей кольцевой ДНК вируса гепатита C, показанных на фиг. 1, 3, 6, 9, 14 и 32, могут быть использованы для выделения перекрывающихся последовательностей кольцевой ДНК от пулов кольцевой ДНК вируса гепатита C. Эти последовательности, которые перекрывают кольцевые ДНК в упомянутых выше клонах, но которые содержат также последовательности, производные от участков генома, от которых кольцевая ДНК в упомянутых выше клонах не происходит, могут быть использованы для синтеза проб для идентификации других перекрывающих сегментов, которые не обязательно перекрывают кольцевые ДНК в клонах, описанных в разделе IV.A. Несмотря на то что геном вируса гепатита C разделен на сегменты и общим последовательностям недостает сегментов, возможно проследить полный вирусный геном(ы), используя метод выделения перекрывающих кольцевых ДНК, полученных от вирусного генома(ов). Хотя и маловероятно, если геном является сегментированным геномом, которому недостает общих последовательностей, но последовательность генома может быть определена серологическим скринингом пулов кольцевой ДНК вируса гепатита C лямбда-gt11, как использовано для выделения клона 5-1-1, определения последовательностей в изолятах кольцевой ДНК и использования выделенных кольцевых ДНК для выделения перекрывающих фрагментов с использованием метода, описанного для выделения и определения последовательностей клонов, приведенных в разделе IV.A. Альтернативно характеристика геномных сегментов может исходить от вирусного генома(ов), выделенных из очищенных частиц вируса гепатита C. Методы очистки частиц вируса гепатита C и определение их в процессе очистки приведены в описании ниже. Способы выделения полинуклеотидных геномов из вирусных частиц известны в данной области, и один процесс, который может быть использован, приведен в примере к разделу IV.A.1. Выделенные геномные сегменты могут быть затем клонированы и определена их последовательность. Таким образом с помощью полученной здесь информации можно провести клонирование и определение последовательности генома(ов) вируса гепатита C, независимо от его природы.
Методы создания пулов кольцевой ДНК известны в данной области и обсуждены ниже и рассматривались выше; метод конструирования пулов кольцевой ДНК вируса гепатита C в лямбда-gt11 описан ниже в разделе IV.A. Однако пулы кольцевой ДНК, полезные для отбора проб нуклеиновых кислот, могут быть созданы также в других векторах, известных в данной области, например лямбда-gt10 (Huynk и соавт., 1985 г.). Кольцевая ДНК, полученная из вируса гепатита C, определенная пробами, полученными от кольцевых ДНК на фиг. 1-32 и от проб, синтезированных из полинуклеотидов, производных от этих кольцевых ДНК, может быть выделена из клона ферментацией выделенного полинуклеотида подходящим ограничивающим ферментом(ами) и определена ее последовательность. См., например, раздел IV. A.3 и раздел IV.A.4, методы, использованные для выделения и определения последовательности кольцевой ДНК вируса гепатита C, которая перекрывает кольцевую ДНК вируса гепатита C в клоне 5-1-1, разделы IV. A. 5 - IV.A.7, методы, использованные для выделения и определения последовательности кольцевой ДНК вируса гепатита C, которая перекрывает таковую в клоне 81, и раздел IV.A.8 и IV.A.9 - для выделения и определения последовательности в клоне, который перекрывает другой клон (клон 36), который перекрывает клон 81.
Информация о последовательностях, полученная из этих перекрывающихся кольцевых ДНК вируса гепатита C, полезна для определения областей гомологии и гетерогенности в вирусном геноме(ах), которые могут указывать на присутствие различных штаммов генома и/или популяций дефектных частиц. Она также полезна для составления гибридизационных проб для определения вируса гепатита C или антигенов вируса гепатита C, или нуклеиновых кислот вируса гепатита C в биологических образцах, и в процессе выделения вируса гепатита C (описано ниже, используя методы, описанные в разделе II.G). Кроме того, перекрывающиеся кольцевые ДНК могут быть использованы для создания векторов экспрессии для полипептидов, полученных из генома(ов) вируса гепатита C, которые также кодируют полипептиды, закодированные в клонах 5-1-1, 36, 81, 91 и 1-2 и в других клонах, описанных в разделе IV.A. Способы создания этих полипептидов, содержащих эпитопы вируса гепатита C, и антител, направленных против эпитопов вируса гепатита C, содержащихся в них, а также их использование являются аналогичными таковым, описанным для полипептидов, полученных от последовательностей кольцевой ДНК вируса гепатита ни A, ни B, содержащихся в клонах 5-1-1, 32, 35, 36, 1-2, 81 и 91, рассмотренных выше и ниже.
Закодированные в семействе кольцевых ДНК последовательности, содержащиеся в клонах 5-1-1, 32, 35, 36, 81, 91, 1-2 и других клонах, описанных в разделе IV.A, являются антигеном(ами), содержащими эпитопы, которые, по-видимому, уникальны к вирусу гепатита C, т.е. антитела, направленные против этих антигенов, отсутствуют у индивидов, инфицированных вирусом гепатита A или вирусом гепатита B, и у индивидов, не инфицированных вирусом гепатита C (см. серологические данные, представленные в разделе IV.B). Кроме того, сравнение информации с последовательностями этих кольцевых ДНК с последовательностями вируса гепатита A, вируса гепатита B, вируса гепатита D и с геномными последовательностями в банке генов показывает, что существует минимальная гомология между этими кольцевыми ДНК и последовательностями нуклеотидов тех источников. Таким образом, антитела, направленные против антигенов, закодированных в кольцевых ДНК этих клонов, могут быть использованы для идентификации частиц вируса BB-ни A, ни B, выделенных у инфицированных индивидуумов. B дополнение они полезны также для выделения возбудителя(ей) гепатита ни A, ни B.
Частицы вируса гепатита C могут быть выделены из сыворотки индивидов, инфицированных вирусом BB-ни A, ни B, или из культуры клеток любым известным методом в данной области, включая, например, методы, основанные на распределении по размерам, такие как седиментации, или методы исключения, или методы, основанные на плотности, такие как ультрацентрифугирование в градиентах плотности, или преципитация агентом, таким как полиэтиленгликоль, или хроматография на различных материалах, таких как анионо- или катионообменные материалы, которые связывают вследствие гидрофобности, а также аффинные колонии. В процессе выделения присутствие вируса гепатита C может определяться гибридизационным анализом экстрагированного генома, используя пробы, полученные из кольцевых ДНК вируса гепатита C, описанных выше, или иммунологическим анализом (см. раздел II.I), используя в качестве проб антитела, направленные против антигенов вируса гепатита C, закодированных в семействе последовательностей кольцевой ДНК, показанных на фиг. 1-32, а также направленных против антигенов вируса гепатита C, закодированных в перекрывающихся последовательностях кольцевой ДНК вируса гепатита C, описанных выше. Антитела могут быть моноклональными или поликлональными и может быть желательной очистка антител до их использования в иммунологическом анализе. Процесс очистки поликлональных антител, направленных против антигена(ов), закодированных в клоне 5-1-1, описан в разделе IV.E; аналогичный процесс очистки может быть использован для антител, направленных против других антигенов вируса гепатита C.
Антитела, направленные против антигенов вируса гепатита C, закодированных в семействе кольцевых ДНК, представленных на фиг. 1-32, а также антигенов, закодированных в перекрывающихся кольцевых ДНК вируса гепатита C, которые фиксированы на твердых подложках, полезны для выделения вируса гепатита C при иммуноаффинной хроматографии. Методы проведения иммуноаффинной хроматографии известны в данной области, включая и методы фиксации антител на твердых подложках, так что они сохраняют свою избирательную иммунологическую активность; методы могут предусматривать адсорбцию антител к подложке (см. , например, Kurstak в Enzyme Immunodiagnosis, страницы 31-37), а также методы, в которых антитела ковалентно связаны с подложкой. Вообще методы аналогичны таковым, используемым для ковалентного связывания антител с твердой подложкой, которые в общих чертах описаны в разделе II.C; однако пространственные группы могут быть включены в бифункциональные связующие агенты с тем, чтобы локализация связывания антигена оставалась доступной для антитела.
В процессе проведения очистки присутствие вируса гепатита C может быть определено и/или подтверждено гибридизацией нуклеиновой кислоты, используя в качестве пробы полинуклеотиды, полученные из семейства последовательностей кольцевой ДНК вируса гепатита C, представленных на фиг. 1-32, а также из последовательностей перекрывающихся кольцевых ДНК вируса гепатита C, описанных выше. В этом случае фракции обрабатывают в условиях, вызывающих разрыв вирусных частиц, например, детергентами в присутствии хелатообразующих средств, а присутствие вирусной нуклеиновой кислоты определяют методом гибридизации, описанным в разделе II.H. Дальнейшее подтверждение того, что выделенные частицы являются агентами, вызывающими вирус гепатита C, может быть получена инфицированием шимпанзе выделенными вирусными частицами с последующим определением того, что являются ли симптомы гепатита ни A, ни B результатом инфекции.
Вирусные частицы очищенного препарата могут быть исследованы далее. Очищают геномную нуклеиновую кислоту. Основываясь на ее чувствительности к рибонуклеазе, а не к дезоксирибонуклеазе I, становится очевидным, что вирус включает геном РНК. См. пример из раздела IV.C.2 ниже. Количество нитей и циклизация или нециклизация могут быть определены известными в данной области методами, включающими, например, наблюдение под электронным микроскопом, миграцией в плотные градиенты и седиментационными характеристиками. Основываясь на гибридизации захваченного генома вируса гепатита C с отрицательными нитями кольцевых ДНК вируса гепатита C, оказывается, что вирус гепатита C может включать положительный нитчатый геном РНК (см. раздел IV.H.1). Подобные методы описаны, например, Methods in Enzymology. B дополнение очищенная нуклеиновая кислота может быть блокирована и определена ее последовательность известными методами, включающими обратную транскрипцию, поскольку материалом генома является РНК. См. , например, Maniatis (1982 г.) и Glover (1985 г.). Использование нуклеиновой кислоты, полученной из вирусных частиц, позволяет проследить весь геном, сегментирован он или нет.
Исследование гомологии полипептида, закодированного в непрерывной открыто читаемой структуре объединенных клонов с 14i по 39c включительно (см. фиг. 26), показывает, что полипептид вируса гепатита C содержит участки гомологии с соответствующими белками в консервированных участках флавивирусов. Пример тому описан в разделе IV.H.3. Эта находка имеет много важных ответвлений. Во-первых, это свидетельство в сочетании с результатами, показывающими, что вирус гепатита C содержит положительный нитчатый геном, размер которого составляет примерно 10000 нуклеотидов, согласуется с предположением, что вирус гепатита C является флавивирусом или флавиподобным вирусом. Вообще вирионы флавивируса и его геномы имеют относительно стойкую структуру и организацию, которые известны. CM., Rice и соавт., (1986 г.) и Brinton М.A. (1988 г.). Таким образом, структурные гены, кодирующие полипептиды C, пре-М/М и E могут быть локализованы в 5'-окончании генома вверху в направлении клона 14i. Кроме того, использование сравнения с другими флавивирусами может предсказать точную локализацию последовательностей, кодирующих эти белки.
Выделение последовательностей вверх от последовательностей в клоне 14i может быть проведено различными путями, которые, располагая приведенной в описании информацией, будут очевидны для специалиста в данной области. Например, метод "блуждающего" генома может быть спользован для выделения других последовательностей, являющихся 5'-окончаниями к последовательностям в клоне 14i, но которые перекрывают этот клон; это в свою очередь ведет к изоляции дополнительных последовательностей. Этот метод достаточно обширно описан ниже в разделе IV.A. Например, известно также, что флавивирусы имеют консервированные эпитопы и участки последовательностей консервированных нуклеиновых кислот. Полинуклеотиды, содержащие законсервированные последовательности, могут быть использованы в качестве проб, которые связывают геном вируса гепатита C, разрешая таким образом его выделение. В дополнение эти законсервированные последовательности в сочетании с последовательностями, полученными от кольцевой ДНК вируса гепатита C, представленным на фиг. 22, могут быть использованы для конструирования затравок в системах, которые увеличивают последовательности генома вверх от последовательностей в клоне 14i, используя реакцию роста полимеразной цепи. Такой пример описан ниже.
Строение вируса гепатита C может быть также определено и его компоненты выделены. Морфология и размеры могут быть определены, например, электронной микроскопией. Идентификация и локализация специфических вирусных полипептидных антигенов, таких как покрытие или оболочка антигенов или внутренние антигены, такие как белки, связывающие нуклеиновые кислоты, сердцевина антигенов и полинуклеотидная полимераза(ы) могут быть также определены, например, из того, присутствуют ли антигены как основные или незначительные компоненты вируса, а также использованием антител, направленных против специфических антигенов, закодированных в выделенных кольцевых ДНК как пробах. Эта информация полезна для составления вакцин; например, может быть предпочтительно включить внешний антиген в препарат вакцины. Поливалентные вакцины могут быть образованы, например, из полипептида, полученного из генома, кодирующего структурный белок, например, белок E, а также из полипептида от другой части генома, например, неструктурного или структурного полипептида.
II. K. Системы клеточных культур и системы модельных животных для репликации вируса гепатита C
Предположение, что вирус гепатита C является флавивирусом или флавиподобным вирусом, предлагает также информацию о методах выращивания вируса гепатита C. Термин "флавиподобный" означает, что вирус показывает значительное число гомологии к известным законсервированным участкам флавивирусов и, что большая часть генома является одной открыто читаемой структурой. Метолы выращивания флавивирусов известны специалистам в данной области (см. например, обзоры Brinton (1986) и Stollar, V. (1980)). Вообще подходящие клетки или культуры клеток для выращивания вируса гепатита C могут включать клетки, поддерживающие репликацию флавивирусов, например следующие: линии клеток почек обезьяны (т.е. MK2, VERO); линии клеток почек свиньи (т.е. PS), линии почечных клеток молодого хомяка (т.е. BНК), линии макрофаговых клеток мурены (т. е. P388D1, MK1, Mm1), линии макрофаговых клеток человека (т.е. U-937), лейкоциты периферической крови человека, слипшиеся моноциты человека, гепатоциты и линии клеток гепатоцитов (т.е. HUH7, HEPG2), зародыши и зародышевые клетки (т.е. фибробласты зародышей цыплят) или клетки, полученные от беспозвоночных, предпочтительно от насекомых (т.е. линии клеток дрозофилы) и более предпочтительно от членистоногих, например, линии клеток москитов (т.е. A. Albopictus, Aedes aegypti, Cutex tritaeniorhynchus) или линии клеток клещей (т.е. RML-14, Dermacentor раrumapertus).
Возможно, что будут выращены первичные гепатоциты и затем инфицированы вирусом гепатита C; или альтернативно культуры гепатоцитов могут быть получены из печени инфицированных индивидов (т. е. человека или шимпанзе). В последнем случае это будет примером клетки, которая инфицирована in vivo и перевита in vitro. В дополнение могут быть использованы различные методы оживления, чтобы получить линии клеток, производные от культур гепатоцитов. Например, первичные культуры печени (до и после обогащения популяции) могут быть слиты в многообразие клеток для поддержания стабильности. Например, культуры могут быть также инфицированы трансформированными вирусами или заражены чужеродными трансформированными генами, чтобы получить перманентные и полуперманентные линии клеток. В дополнение, например, клетки в культурах печени могут быть сплавлены до востановленных линий клеток (т.е. HepG2). Методы слияния клеток известны в данной области и включают, например, использование агентов слияния, таких как полиэтиленгликоль, вирус Сендан и вирус Эпстайна-Барра.
Как обсуждалось выше, вирус гепатита C является флавивирусом или флавиподобным вирусом. Поэтому вероятно, что инфекция линий клеток вирусом гепатита C может быть осуществлена методами, известными в данной области при инфицировании клеток флавивирусами. Эти методы включают, например, инкубирование клеток с вирусными препаратами в условиях, позволяющих вирусу проникновение в клетку. В дополнение возможно получить вирусную продукцию заражением клеток вирусными полинуклеотидами. Известно, что тогавирусы и флавивирусы, как РНК-содержащие вирусы, инфекционны в различных линиях клеток позвоночных (Pfefferkorn и Shapiro, 1974) и в линии клеток москитов (Peleg, 1969). Методы заражения клеток культуры ткани дуплексами РНК, положительными нитчатыми РНК и кольцевыми ДНК (и другими ДНК) известны в данной области и включают, например, методы, которые используют электропорацию и преципитацию ДЕАЕ-декстраном или фосфатом кальция. Обильный источник РНК вируса гепатита C можно получить проведением in vitro транскрипции кольцевой ДНК вируса гепатита C на полный геном. Трансфекция этим материалом или клонированной кольцевой ДНК вируса гепатита C проявится в вирусной репликации и воспроизведении вируса in vitro.
В дополнение к культивируемым клеткам могут быть использованы системы модельных животных для репликации вирусов: животные системы, в которых известны флавивирусы, знакомы специалистам в данной области (см., например, обзор Monath, 1986). Таким образом, репликация вируса гепатита C происходит не только у шимпанзе, а также и у мармозетки и у сосунков мышей.
II.L. Проверка (скрининг) противовирусных агентов на вирус гепатита C
Наличие культуры клеток и систем модельных животных для вируса гепатита C делает также возможным проверку антивирусных агентов, которые ингибириуют репликацию вируса гепатита C, и особенно тех агентов, которые преимущественно способствуют росту клеток и увеличению их численности, ингибируют в то же время вирусную репликацию. Эти методы скрининга известны специалистам в данной области. Вообще противовирусные агенты испытывают при различных концентрациях на их действие по предотвращению вирусной репликации в системах культур клеток, которые поддерживают репликацию вируса, и затем на ингибирование инфекционности или вирусной патогенности (и на низший уровень токсичности) в модельной системе животного.
Методы и композиции, предусмотренные в описании для определения антигенов вируса гепатита C и полинуклеотидов вируса гепатита C, полезны для испытаний антивирусных агентов тем, что они представляют альтернативу и вообще более чувствительные средства для детектирования кипения агента на вирусную репликацию, чем метод розеток или тест на ингибирующую дозу - 50%. Например, описанные здесь полинуклеотидные пробы вируса гепатита C могут быть использованы для количественной оценки вирусной нуклеиновой кислоты, продуцированной в культуре клеток. Это можно осуществить, например, гибридизацией или конкурентной гибридизацией инфицированных нуклеиновых кислот клетки с меченым полинуклеотидом вируса гепатита C пробы. Например, антитела, направленные против вируса гепатита C, могут быть использованы для идентификации и количественной оценки антигена(ов) вируса гепатита C в клеточной культуре, используя иммунологический анализ, описанный выше. В дополнение, поскольку может быть желательно количественно оценить антигены вируса гепатита C в инфицированной культуре клеток конкурентным анализом, то полипептиды, закодированные в кольцевых ДНК вируса гепатита C, описанные в заявке, полезны при проведении этих конкурентных анализов. Вообще рекомбинантные полипептиды вируса гепатита C, полученные от кольцевых ДНК вируса гепатита C, должны быть помечены и ингибирование связывания этих меченых полипептидов с полипептидами вируса гепатита C благодаря антигену, продуцированному в системе культуры клеток, может быть зарегистрировано. Кроме того, эти методы особенно полезны в случаях, когда вирус гепатита C может быть способен к репликации в линии клеток, не вызывая их гибели.
II.М. Получение ослабленных штаммов вируса гепатита C
B дополнение к описанному выше, используя системы тканевых культур и/или систем модельных животных, можно выделить ослабленные линии вируса гепатита C. Эти линии могут быть пригодны для вакцин или для выделения вирусных антигенов. Ослабленные линии могут быть выделены после многократных пассажей в культуре клеток и/или модельного животного. Определение ослабленного штамма в инфицированных клетках или у индивида осуществляется методами, известными в данной области, и может включать, например, использование антител к одному или более эпитопам, закодированным в вирусе гепатита C как в пробе, или использование полинуклеотида, содержащего последовательность вируса гепатита C по меньшей мере из 8 нуклеотидов, как пробу. Альтернативно или в дополнение ослабленный штамм может быть получен с использованием информации генома вируса гепатита C, приводимой в описании, и использованием рекомбинантных методов. Вообще можно попытаться уничтожить участок генома, кодирующий, например, полипептид, относящийся к патогенности, но который допускает репликацию вируса. В дополнение строение генома должно допускать экспрессию генома, ведущего к образованию нейтрализующих антител к вирусу гепатита C. Этот видоизмененный геном может быть затем использован для трансформации клеток, допускающих репликацию вируса гепатита C, и клетки выращивают в условиях, позволяющих вирусную репликацию. Ослабленные штаммы вируса гепатита C полезны не только для целей, связанных с получением вакцин, но и как источник коммерческого продуцирования вирусных антигенов, поскольку обработка этих вирусов потребует менее строгих защитных мер для пользователей, участвующих в получении вирусов и/или вирусной продукции.
III. Общие методы
Общие методы, использованные для экстрагирования генома из вируса, получение и испытание пула кольцевой ДНК, определение последовательностей в клонах, конструирование векторов экспрессии, трансформация клеток, проведение радиоиммунологического анализа и иммуноферментного твердофазного анализа для выращивания клеток в культуре и тому подобные известны в данной области и лабораторные справочники доступно описывают эти методы. Однако для общего руководства ниже приводятся некоторые источники, в настоящее время доступные для проведения таких методов, и материалы, полезные при их осуществлении.
III.A. Хозяева и контрольные последовательности экспрессии
Прокариотные и эукариотные клетки-хозяева могут быть использованы для экспрессии желаемых кодирующих последовательностей, если используются соответствующие контрольные последовательности, совместимые с определенным хозяином. Среди прокариотных хозяев наиболее часто употребляют Е. coil. Контрольные последовательности экспрессии для прокариотов включают промоторы, необязательно содержащие части гена-оператора, и локализации связывания рибосом. Векторы переноса, совместимые с прокариотными хозяевами, обычно получают, например, из pBR322, плазмиды, содержащей опероны, придающие устойчивость к ампициллину и тетрациклину, и различные pUC векторы, которые также содержат последовательности, передающие маркеры антибиотической устойчивости. Эти маркеры могут быть использованы для получения трансформантов селекцией. Обычно используемые контрольные прокариотные последовательности включают беталактамазу (пенициллиназу) и системы лактозных промоторов (Chang и соавт., 1977 г. ), систему триптофанового промотора (trp) (Goeddel и соавт., 1980 г.) и лямбда-производный PL промотор и локализация связывания рибосомы N гена (Shimatake и соавт. , 1981 г.) и гибридный tac (промотор De Boer и соавт., 1983 г. ), полученный из последовательности trp и lac UV5 промоторов. Предыдущие системы особенно совместимы с E.coli; если необходимо, то могут быть использованы другие прокариотные хозяева, такие как штаммы Bacillus или Pseudomonas с соответствующими контрольными последовательностями.
Эукариотные хозяева включают дрожжи и клетки млекопитающих в системах культур. Saccharomyces cerevisiae и Saccharomyces carlsbergensis наиболее обычно используемые дрожжевые хозяева и удобными являются грибковые хозяева. Совместимые дрожжевые векторы несут маркеры, которые допускают селекцию удачных трансформантов передачей прототропии аутотропным мутантам или устойчивость к тяжелым металлам штаммам диких типов. Совместимые дрожжевые векторы могут применять двухмикронный источник репликации (Broach и соавт., 1983 г. ), сочетание CEN3 и ARS1 или другие средства для обеспечения репликации, такие как последовательности, которые приводят к внедрению соответствующего фрагмента в геном клетки хозяина. Контрольные последовательности для дрожжевых векторов известны в данной области и включают промоторы синтеза гликолитических ферментов (Hess и соавт., (1968 г.); Holland и соавт. (1978 г.)), включая промотор для 3-фосфоглицератиназы (Hitzeman, 1980 г.). Могут быть также включены прекращающие синтез агенты (терминаторы), такие как производные от анолазного гена (Holland, 1981 г.). Особенно полезными контрольными системами являются системы, которые содержат глицеральдегид-3- фосфатдегидрогеназный промотор (GAPDH) или спиртовой дигидрогенозный регулирующий промотор, терминаторы, также полученные из глицеральдегид-3-фосфатдегидрогеназы и, если необходимо вызвать секрецию, то головная последовательность из дрожжевого альфа-фактора. В дополнение участки транскрипционного регулирования и транскрипционного инициирования, которые операбельно связаны, могут быть такими, что они не естественно соединены в организме дикого типа. Эти системы детально описаны в Европейском патенте N 120551, опубликованном 3 октября 1984 г; в Европейском патенте N 116201, опубликованном 22 августа 1984 г., и в Европейском патенте N 164556, опубликованном 18 декабря 1985 года; все патенты принадлежат настоящему заявителю и приведены в библиографической ссылке.
Линии клеток млекопитающих, полезных в качестве хозяев для экспрессии, известны в данной области и включают многие оживленные линии клеток, получаемые из собрания типов культур Америки (АТСС), например клетки HeLa, клетки яичника китайского хомяка (CHO), клетки почек детеныша хомяка (BНК) и ряд других линий клеток. Подходящие промоторы также известны в данной области и включают вирусные промоторы, такие как промоторы из вакуолизирующего обезьяньего вируса (OB-40), вируса саркомы Роуза (ВСР), аденовируса и вирус бычьей папилломы (ВБП). Клетки млекопитающих могут также нуждаться в конечных последовательностях и поли A дополнительных последовательностях; могут быть также включены усиливающие последовательности, которые увеличивают экспрессию, и последовательности, которые вызывают увеличение числа генов, могут быть также желательны. Эти последовательности известны в данной области. Векторы, пригодные для репликации в клетках млекопитающих, могут включать вирусные репликоны или последовательности, которые обеспечивают целостность соответствующих последовательностей, кодирующих эпитопы вируса ни A, ни B в геноме хозяина.
III.B. Трансформации
Трансформация может быть осуществлена любым известным методом введения полинуклеотидов в клетку хозяина, включая, например, упаковку полинуклеотида в вирус и перенос в клетку-хозяина генетического материала вируса или непосредственное поглощение полинуклеотида. Процесс трансформации зависят от хозяина, подлежащего трансформированию. Например, трансформация клеток хозяина E. coli посредством лямбда-gt11, содержащим последовательности вируса гепатита BB-ни A, ни B, рассмотрена в этом разделе ниже. Бактериальная трансформация может быть при непосредственном поглощении, обычно включающем обработку хлористым кальцием или хлористым рубидием (Cohen (1972 г.); Maniatis (1982 г.)). Трансформация дрожжей прямым поглощением может быть проведена с использованием метола Hinnen с соавт., 1978 г. Трансформация клеток млекопитающих прямым поглощением может быть выполнена с использованием метода осаждения фосфатом кальция (метод Graham and Van der Eb, 1978 г.) или различных известных его модификаций.
III.С. Построение векторов
При конструировании векторов применяют способы, известные в данной области. Местоспецифическую сегментацию ДНК проводят обработкой подходящими ограничивающими ферментами в условиях, которые обычно определяются изготовителями этих коммерчески доступных ферментов. В общем, примерно 1 микрограмм плазмиды или последовательности ДНК расщепляется 1 единицей фермента примерно в 20 микролитрах буферного раствора при инкубации 1-2 часа при 37oC. После инкубации с ограничивающим ферментом белок удаляют экстракцией смесью фенол-хлороформ, а ДНК выделяют осаждением с этанолом. Расщепленные фрагменты могут быть разделены с использованием полиакриламида или методом электрофореза на агарозовом геле согласно общим методам, описанным в Methods in Enzymology (1980), 65:499-560.
Фрагменты расщепления с острыми концами могут стать с рваными концами, используя полимеразу 1 ДНК E.coli (Кленов) в присутствии соответствующего дезоксинуклеотидтрифосфата, находящегося в смеси. Может быть использована также обработка S1 нуклеазой, приводящая к гидролизу частей любой однонитчатой ДНК.
Лигации могут быть выполнены с использованием стандартного буфера и стандартных температурных условий с использованием лигазы ДНК T4 и аденозинтрифосфата и меньше лигазы, чем лигации с рваными краями. Если фрагменты вектора используют как часть лигационной смеси, то фрагмент вектора часто обрабатывают щелочной фосфатазой бактерий или кишечной щелочной фосфатазой теленка для удаления 5'-фосфата и таким образом предотвращения повторной лигации вектора; альтернативно рассасывание ограничивающим ферментом нежелательных
фрагментов может быть использовано для предотвращения лигации.
Лигационные смеси преобразуют в подходящих клонирующих хозяев, таких как E. coli, и эффективные трансформанты отбирают, например, резистентностью к антибиотикам, и исследуют истинную структуру.
III.D. Построение желаемых последовательностей ДНК
Синтетические олигонуклеотиды могут быть получены с использованием автоматического синтезатора, как описано (1984). Если желают, то синтетические нити могут быть помечены фосфором-32 при обработке полинуклеотидной киназой в присутствии меченного фосфором-32 аденозинтрифосфата, используя стандартные условия реакции.
Последовательности ДНК, включающие таковые, выделенные из пулов кольцевой ДНК, могут быть модифицированы методами, включающими, например, локально направленный мутагенез, как описано Zoller (1982). Коротко говоря, подлежащую модифицированию ДНК упаковывают в фаг, как однонитчатую последовательность, и обращают в двунитчатую ДНК полимеразой ДНК, используя как затравку синтетическую олигонуклеотидную комплементарность к части ДНК, которая должна быть модифицирована, и имеющая желаемую модификацию, включенную в ее собственную последовательность. Полученная двунитчатая ДНК трансформирована в фаг, поддерживающий бактерию-хозяина. Культуры трансформированных бактерий, которые содержат репликации каждой нити фага, помещали в чашки с агаровой средой, чтобы получить пятно. Теоретически 50% новых пятен содержат фаг, имеющий мутированную последовательность, а остальные 50% имеют исходную последовательность. Репликаты пятен гибридизируют в меченую синтетическую пробу при температурах и условиях, которые допускают гибридизацию с правильной нитью, но не с немодифицированной последовательностью. Последовательности, которые были идентифицированы гибридизацией, выделяли и из них образовывали клоны.
III.E. Гибридизация с пробой
Пулы ДНК могут быть опробированы по методу Grunstein и Hogness (1975). Кратко, в этом методе ДНК, подлежащую испытаниям, фиксировали на нитроцеллюлозных фильтрах, денатурировали и предварительно гибридизировали с буфером, содержащим 0-50% формамида, 0,75 моля хлористого натрия, 75 мМ нитрата натрия, 0,02% (от веса по объему) каждого: альбумина бычьей сыворотки, поливинилпирролидона и Ficoll натрия (pH 6,5), 0,1% додецилсульфата натрия и 100 микрограмм на миллилитр носителя денатурированной ДНК. Процентное содержание формамида в буфере, а также время и температурные условия предварительной гибридизации и последующей операции гибридизации зависят от соблюдения требуемой строгости. Олигомерные пробы, которые требуют для проведения более мягкие условия строгости, обычно используют низкое процентное содержание формамида, требуют более низких температур и более продолжительного времени гибридизации. Пробы, содержащие более 30 или 40 нуклеотидов, такие как пробы, полученные от кольцевой ДНК или геномных последовательностей, обычно требуют применения более высоких температур, т.е. около 40-42oC и высоких процентов, т.е. 50% формамида. В ходе предварительной гибридизации к буферу добавляли меченную фосфором-32 5'-олигонуклеотидную пробу, и фильтры инкубировали в этой смеси в условиях гибридизации. После промывки обработанные фильтры подвергали ауторадиографии для определения локализации гибридизированной пробы; ДНК в соответствующих локализациях на исходных агаровых пластинках использовали как источник желаемой ДНК.
III.F. Подтверждение конструкции и определение последовательности
Для традиционных построений векторов лигационную смесь трансформировали в штамм E. coli HB101 или другой подходящий хозяин и эффективные трансформанты отбирали по антибиотической резистентности или другим маркерам. Плазмиды от трансформантов готовили потом по методу Clewell и соавт. (1969), обычно сопровождая увеличением хлорамфениколом (Clewell, 1972 г.). ДНК выделяли и анализировали обычно определением ограничения активности фермента и/или определением последовательности. Определение последовательности можно провести дидезоксиметодом Sanger и соавт. (1977 г.), как далее описано Messing и соавт. (1981 г.), или по методу Maxam и соавт. (1980 г.). Проблемы, связанные со сжатием полос, которое наблюдается иногда в участках, богатых GC, преодолевают использованием T-деазогуанозина согласно Барру и соавторов (1986).
III.G. Иммуносорбентный анализ с ферментной меткой
Иммуноферментный анализ с ферментной меткой (ELISA) может быть использован для измерения концентраций либо антигена, либо антител. Этот метод зависит от слияния фермента либо с антигеном, либо с антителом и использует активность связывания фермента как количественную метку. Чтобы измерить антитела, известный антиген фиксируют на твердой фазе (т.е. на микропластике или пластиковой чашке), инкубируют с разведениями испытываемой сыворотки, промывают, инкубируют с иммуноглобулином, меченным ферментом, и снова промывают. Подходящие ферменты для введения метки известны в данной области и включают, например, пероксидазу ложечницы. Активность фермента, связанного с твердой фазой, измеряют добавлением специфического субстрата и определяют образование продукта или использование субстрата колориметрическим методом. Активность связанного фермента есть прямая функция количества связанных антител.
Чтобы измерить антиген, известное специфическое антитело фиксируют к твердой фазе и добавляют испытываемый материал, содержащий антиген; после инкубирования твердую фазу промывают и добавляют второе меченное ферментом антитело. После промывки добавляют субстрат и активность фермента определяют колориметрически и относят к концентрации антигена.
IV. Примеры
Представленные ниже примеры реализации настоящего изобретения имеют чисто иллюстративные цели и не ограничивают объема изобретения. В свете настоящего описания многочисленные осуществления в объеме изобретения будут очевидны для обычных специалистов в данной области. Представленные процедуры, например, в разделе IV.A могут быть при желании повторены, так как методы доступны для конструирования нуклеотидных последовательностей, основываясь на информации, представляемой изобретением. Экспрессия описана на примере Е. coli, однако и другие системы, описанные более полно в разделе III.A, доступны. Могут быть получены дополнительные эпитопы, производные от геномной структуры и использованы для образования антител, как описано ниже.
IV. A. Получение, выделение и определение последовательностей кольцевой ДНК вируса гепатита C
IV.A.1. Получение кольцевой ДНК вируса гепатита C
Источником возбудителя гепатита ни A, ни B был пул плазмы, полученной от шимпанзе с хронической инфекцией гепатита ни A, ни B. Шимпанзе был экспериментально инфицирован кровью другого шимпанзе с хроническим гепатитом ни A, ни B, индуцированным инфекцией вирусом гепатита C в зараженной пробе концентрата фактора 8, полученного из пула сывороток человека. Пул плазмы шимпанзе был образован объединением многих отдельных образцов плазмы, содержащих высокие уровни активности аланинаминотрансферазы; эта активность является результатом повреждения печени, вследствие инфекции вирусом гепатита C. Поскольку 1 мл 10-6 разбавления этой депонированной сыворотки, введенной внутривенно, вызывал гепатит ни A, ни B у другой шимпанзе, то ее инфекционная доза для шимпанзе была по меньшей мере 106/мл, т.е. имела высокий инфекционный вирусный титр.
Пул кольцевой ДНК из пула плазмы с высоким титром генерировали следующим образом. Сначала выделяли вирусные частицы из плазмы: 90 мл аликвотной пробы разбавляли 310 мл раствора, содержащего 50 мМ трис-HCl, pH 8,0, 1 мМ этилендиаминтетрауксусной кислоты, 100 мМ хлористого натрия. Дебрис удаляли центрифугированием в течение 20 минут при 15000 g и 20oC. Вирусные частицы в полученном надосадочном слое обращали затем в сферы центрифугированием в роторе Beckman SW 28 при 28000 оборотов в минуту в течение пяти чесов при 20oC. Чтобы освободить вирусный геном, частицы разрывали суспендированием шариков в 15 мл раствора, содержащего 1% додецилсульфата натрия, 10 мМ этилендиаминтетрауксусной кислоты, 10 мМ этилендиаминтетрауксусной кислоты, 10 мМ трис-HCl буфера, pH 7,5, также содержащего 2 мг/мл протеиназы K, с последующим инкубированием при 45oC в течение 90 минут. Нуклеиновые кислоты выделяли добавлением 0,8 микрограмма MS2 1H бактериофага в качестве носителя и экстрагировали смесь 4 раза смесью фенол-хлороформ в отношении 1:1 (фенол, насыщенный 0,5 М трис-HCl, pH 7,5; 0,1% (по объему) бета-меркаптоэтанола, 0,1% (от веса по объему) оксихинолона, с последующим экстрагированием хлороформом два раза. Водную фазу концентрировали с 1-бутанолом перед осаждением с 2,5 объемами абсолютного этанола при -20oC и оставляли на ночь. Нуклеиновую кислоту выделяли центрифугированием в роторе Beckman SW41 при 40000 оборотов в минуту в течение 90 минут при 4oC и растворяли в воде, которая была обработана 0,05% (по объему) диэтилпирокарбонатом и автоклавировали.
Нуклеиновую кислоту, полученную на предыдущей операции (< 2 микрограмм), денатурировали 17,5 мМ CH3HgOH; кольцевую ДНК синтезировали с использованием этой денатурированной нуклеиновой кислоты в качестве матрицы и клонировали в EcoRI локализацию фага лямбда-gt11, используя методы, описанные Huynh (1985 г. ), за исключением того, что случайные затравки заменяли олиго(dT) 12-18 в процессе синтеза первой нити кольцевой ДНК обратной транскриптазой (Taylor и соавт., 1976 г.). Полученная двунитчатая ДНК была фракционирована по размерам на колонке сефароза CL-4B; элюированный материал примерно со средними размерами 400, 300, 200 и 100 пар оснований депонировали в пулах кольцевой ДНК 1, 2, 3 и 4 соответственно. Пул лямбда-gt11 кольцевой ДНК был генерирован из пула N 3 кольцевой ДНК.
Пул кольцевой ДНК лямбда-gt11, генерированный из пула N 3, исследовали на эпитопы, которые могли бы специфически связываться с сывороткой больного, который ранее перенес гепатит ни A, ни B. Было исследовано около 106 фагов с сыворотками больных, используя методы Huynh и соавт. (1985 г.), за исключением того, что связанное антитело человека определяли иммуноглобулином человека в антисыворотке овцы, меченным радиоактивным йодом-125.
Было идентифицировано пять положительных фагов, которые были очищены. Затем пять положительных фагов испытывали на специфичность связывания с сывороткой 8 различных лиц, ранее инфицированных возбудителем гепатита ни A, ни B, используя тот же метод. Четыре фага кодировали полипептид, который взаимодействовал иммунологически только с сывороткой одного человека, т.е. с одной из сывороток, использованных для первичного скрининга пула фагов. Пятый фаг кодировал полипептид, который реагировал иммунологически с 5 из 8 испытанных сывороток. Кроме того, этот полипептид не неарировал иммунологически с сыворотками от 7 нормальных доноров крови. Поэтому кажется, что клон 5-1-1 кодирует полипептид, который специфически распознается иммунологически сыворотками от больных гепатитом ни A, ни B.
IV. A. 2. Последовательности кольцевой ДНК вируса гепатита C в рекомбинантном фаге 5-1-1 и полипептида, закодированного в последовательности
Кольцевая ДНК в рекомбинантном фаге 5-1-1 была подвергнута определению последовательностей по методу Sanger и соавт. (1977 г.). В сущности кольцевая ДНК была фрагментирована с помощью EcoRI, выделена фракционированием по размерам с использованием гель-электрофореза. Фрагменты ограничения EcoRI были субклонированы в M13 векторы, mp18 и mpl9 (Messing, 1983 г.) и исследованы их последовательности с использованием метода прекращения двойной дезоксицепи по методу Sanger и соавт. (1977). Полученная последовательность показана на фиг. 1.
Полипептид закодированный на фиг. 1, т.е. закодированный в кольцевой ДНК вируса гепатита C, находится в той же самой структуре, что и N-концевая часть молекулы бета-галактозидазы, с которой она сплавлена. Как показано в разделе IV. A, открыто читаемая трансляционная структура клона 5-1-1 кодирует эпитоп(ы), специфически распознанные сыворотками больных людей и шимпанзе, с инфекциями гепатита ни A, ни B.
IV.A.3. Выделение перекрытия генетического кода кольцевой ДНК вируса гепатита C с кольцевой ДНК в клоне 5-1-1
Перекрытие кольцевой ДНК вируса гепатита C с кольцевой ДНК в клоне 5-1-1 было получено исследованием того же пула лямбда-gt11, созданного, как описано в разделе IV.A.1, с синтетическим полинуклеотидом, полученным от последовательности кольцевой ДНК вируса гепатита C в клоне 5-1-1, как показано на фиг. 1. Последовательность полинуклеотида, использованного для проведения скрининга, была:
5'-TCC CTT GCT CGA TGT ACG GTA AGT GCT GAG AGC ACT CTT CCA TCT CAT CGA ACT CTC GGT AGA GGA CTT CCC TGT CAG GT-3'.
Пул лямбда-gt11 был исследован этой пробой с использованием метода, описанного Huynh (1985). Приблизительно один из 50000 клонов скрещивается с пробой. Три клона, гибридизирующиеся с синтетической пробой, были занумерованы 81, 1-2 и 91.
IV. A.4. Нуклеотидные последовательности в перекрывающихся кольцевых ДНК вируса гепатита C кольцевой ДНК в клоне 5-1-1
Последовательности нуклеотидов в трех клонах: 81, 1-2 и 91 определяли, по существу, так же, как и в разделе IV.A.2. Последовательности этих клонов относительно последовательности кольцевой ДНК вируса гепатита C в фаге 5-1-1 представлены на фиг. 2, которая показывает нить, кодирующую определенный эпитоп вируса гепатита C, и где гомологи в нуклеотидных последовательностях указаны вертикальными линиями между последовательностями.
Последовательности клонированных кольцевых ДНК вируса гепатита C высокогомологичны в перекрываемых участках (см. фиг. 2). Однако имеются различия в двух участках. Нуклеотид 67 в клоне 1-2 является тимидином, тогда как другие три клона содержат цитидиновый остаток в этом положении. Следует заметить, что та же самая аминокислота закодирована, если это положение занимает либо C, либо T.
Второе различие заключается в том, что клон 5-1-1 содержит 28 пар оснований, которые не присутствуют в трех других клонах. Эти пары оснований занимают начало последовательности кольцевой ДНК в 5-1-1 и обозначены строчными буквами. Основываясь на данных радиоиммунного анализа, который обсуждается ниже в разделе IV.D, можно допустить, что эпитоп вируса гепатита C может быть закодирован в этом 28 п.о. (пар оснований) участке.
Отсутствие 28 пар оснований в клонах 31, 1-2 и 91, присутствующих в клоне 5-1-1, может означать, что кольцевая ДНК в этих трех клонах получена от дефектных геномов; альтернативно 28 п.о. участок может быть концевым артефактом в клоне 5-1-1.
Последовательность маленьких букв в нуклеотидной последовательности клонов 81 и 91 указывает на то, что эти последовательности не были найдены в других кольцевых ДНК, потому что кольцевые ДНК, перекрывающие эти участки, до сего времени еще не выделены.
Композиционная последовательность кольцевой ДНК вируса гепатита C, полученная от перекрывающихся ДНК в клонах 5-1-1, 81, 1-2 и 91, показана на фиг. 3. Однако на этой фигуре уникальные 28 пар оснований клона 5-1-1 пропущены. На фигуре показана также последовательность полипептида, закодированного в открыто читаемой структуре композиционной кольцевой ДНК вируса гепатита C.
IV. A.5. Выделение перекрывающихся кольцевых ДНК вируса гепатита C с кольцевой ДНК в клоне 81
Выделение последовательностей кольцевой ДНК вируса гепатита C, перекрывающих кольцевую ДНК в клоне 81 в верхней части, осуществляли следующим образом. Лямбда-gt11 пул кольцевой ДНК, полученный как описано в разделе IV. A.1, был проверен гибридизацией с синтетической полинуклеотидной пробой, гомологичной 5'-концевой последовательности клона 81. Последовательность клона 81 представлена на фиг. 4. Эта последовательность синтетического полинуклеотида, использованная для скрининга, представляла собой:
5' CTG TCA GGT ATG ATT GCC CGC TTC CCG GAG 3'.
Методы выделения в основном соответствовали описанным Huynh (1985), за исключением того, что фильтры пулов дважды промывали в строгих условиях, т. е. пять раз промывали SSC с 0,1% додецилсульфата натрия при 55oC в течение 30 минут каждая промывка. Примерно один из 50000 клонов образовывал гибрид с пробой. Положительный рекомбинантный фаг, который содержал кольцевую ДНК, дающую гибрид с последовательностью, выделяли и очищали. Этот фаг был занумерован как клон 36.
Нисходящие последовательности кольцевой ДНК, которая перекрывает оканчивающиеся карбоксилом последовательности кольцевой ДНК в клоне 81, были выделены с использованием метода, аналогичного таковому для выделения восходящих последовательностей кольцевой ДНК, за исключением того, что пробу синтетического олигонуклеотида получали гомологичную 3'-концевой последовательности в клоне 81. Последовательность синтетического полинуклеотида, использованного для отбора последовательностей кольцевой ДНК, была следующей:
5' TTT GGC TAG TGG TTA GTG GGC TGG TGA CAG 3'.
Положительный рекомбинантный фаг, который содержал кольцевую ДНК, образующую гибрид с этой последней последовательностью, был выделен, очищен и получил нумерацию клона 38.
IV. A.6. Нуклеотидная последовательность кольцевой ДНК вируса гепатита C в клоне 36
Последовательность нуклеотидов кольцевой ДНК в клоне 36 определяли по существу методом, описанным в разделе IV.A.2. Двунитчатая последовательность этой кольцевой ДНК, участок ее перекрытия с кольцевой ДНК в клоне 81 и полипептид, закодированный открыто читаемой структурой, представлены на фиг. 5.
Открыто читаемая структура в клоне 36 находится в том же участке, что и антиген вируса гепатита C, закодированный в клоне 81. Таким образом в комбинации открыто читаемые структуры в клонах 36 и 81 кодируют полипептид, который представляет часть крупного антигена вируса гепатита C. Последовательность этого предполагаемого полипептида вируса гепатита C и кодирующая его последовательность двунитчатой ДНК, которая получена от сочетания открыто читаемых структур кольцевых ДНК вируса гепатита C клонов 36 и 81, представлены на фиг. 6.
IV.A.7. Последовательности нуклеотидов кольцевой ДНК вируса гепатита C в клоне 32
Нуклеотидную последовательность кольцевой ДНК в клоне 32 определяли в основном так, как описано в разделе IV.A.2 для последовательности клона 5-1-1. Данные о последовательности указывают на то, что кольцевая ДНК в рекомбинантном фаге клона 32 получена из двух различных источников. Один фрагмент кольцевой ДНК включал 418 нуклеотидов, производных от генома вируса гепатита C; другой фрагмент содержал 172 нуклеотида, полученных от генома бактериофага MS2, который был использован в качестве носителя во время приготовления пула кольцевой ДНК плазмы лямбда-gt11.
Последовательность кольцевой ДНК в клоне 32, соответствующая последовательности генома вируса гепатита C, представлена на фиг. 7. Участок последовательности, перекрывающий таковую клона 81, и полипептид, закодированный открыто читаемой структурой, также показан на этой фигуре. Эта последовательность содержит одну непрерывную открыто читаемую структуру, находящуюся в той же самой трансляционной структуре, что и антиген вируса гепатита C, закодированный клоном 81.
IV.A.8. Выделения перекрытия кольцевой ДНК вируса гепатита C с кольцевой ДНК в клоне 36
Выделение восходящих последовательностей кольцевой ДНК вируса гепатита C, которая перекрывает последовательность кольцевой ДНК в клоне 36, осуществляли, как описано в разделе IV.A.5, для последовательности, которая перекрывает кольцевую ДНК клона 81, за исключением того, что синтетический полинуклеотид базировался на 5'-участке клона 36. Последовательность синтетического полинуклеотида, использованного для скрининга, была:
5' AAG CCA CCG TGT GCG CTA GGG CTC AAG CCC 3'.
Примерно в одном из 50000 клонов происходила гибридизация с пробой. Выделенный, очищенный клон рекомбинантного фага, содержащего кольцевую ДНК, которая гибридизировала с этой последовательностью, был назван клоном 35.
IV.A.9. Последовательность нуклеотидов кольцевой ДНК вируса гепатита C в клоне 35
Нуклеотидную последовательность кольцевой ДНК в клоне 35 определяли по существу так, как описано в разделе IV.A.2. Последовательность, ее участок перекрытия с последовательностью кольцевой ДНК в клоне 36 и предполагаемый полипептид, закодированный в клоне 36, клоне 81 и клоне 32. На фиг. 9 показана последовательность длинной непрерывной открыто читаемой структуры, которая простирается через клоны 35, 36, 81 и 32 вместе с предполагаемым полипептидом, закодированным в ней. Эта объединенная последовательность была подтверждена с использованием других самостоятельных клонов кольцевой ДНК, полученных из того же пула кольцевой ДНК лямбда-gt11.
IV.A.10. Выделение перекрытия кольцевой ДНК вируса гепатита C с кольцевой ДНК в клоне 35
Выделение восходящих последовательностей кольцевой ДНК вируса гепатита C, которые перекрывают последовательности кольцевой ДНК в клоне 35, осуществляли, как описано в разделе IV.A.8, для последовательностей, которые перекрывают кольцевую ДНК клона 36, за исключением того, что синтетический полинуклеотид базировался на 5'-участке клона 35. Последовательность синтетического полинуклеотида, использованного для скрининга, была следующей:
5' CAG GAT GCT GTC TCC CGC ACT CAA CGT 3'.
Примерно один из 50000 клонов гибридизировался с пробой. Выделенный, очищенный клон рекомбинантного фага, содержащего кольцевую ДНК, которая гибридизируется в эту последовательность, была названа клоном 37b.
IV.A.11. Нуклеотидная последовательность вируса гепатита C в клоне 37b
Последовательность нуклеотидов кольцевой ДНК в клоне 37b определяли в основном так, как описано в разделе IV.A.2. Последовательность, участок ее перекрытия с последовательностью кольцевой ДНК в клоне 35 и предполагаемый полипептид, закодированный в ней, представлены на фиг. 10.
5'-концевой нуклеотид клона 35 является T (тимин), в то время как соответствующим нуклеотидом в клоне 37b является аденин (A). Кольцевые ДНК из трех других самостоятельных клонов, которые были выделены во время процедуры выделения клона 37b, описанного в разделе IV.A.10, подвергали определению последовательностей. Кольцевые ДНК этих клонов также содержали аденин в этом положении. Таким образом, концевой 5' тимин в клоне 35 может быть артефактом процедуры клонирования. Известно, что артефакты часто появляются в 5'-концах молекул кольцевой ДНК.
Клон 37b очевидно содержит одну непрерывную открыто читаемую структуру, которая кодирует полипептид, находящийся в продолжении полипептида, закодированного в открыто читаемой структуре, которая простирается через перекрытие клонов 35, 36, 81 и 32.
IV. A. 12. Выделение перекрытия кольцевой ДНК вируса гепатита C к кольцевой ДНК в клоне 32
Выделение нисходящих последовательностей кольцевой ДНК вируса гепатита C к клону 32 проводили следующим образом. Сначала выделяли клон cla, используя синтетическую гибридизационную пробу, которая основывалась на нуклеотидной последовательности кольцевой ДНК вируса гепатита C и последовательности в клоне 32. Метод выделения существенно не отличался от описанного в разделе IV. A. 5, за исключением того, что синтетическая проба была такой последовательности:
5' AGT GCA GTG GAT GAA CCG GCT GAT AGC CTT 3'.
Используя нуклеотидную последовательность из клона cla, синтезировали иную нуклеотидную последовательность такого вида:
5' TCC TGA GGC GAC TGC ACC AGT GGA TAA GCT 3'.
Отбор из пула лямбда-gt11 с использованием полученной из клона cla последовательности в качестве пробы давал примерно один клон на 50000 положительных колоний. Выделенный очищенный клон, который образовывал гибрид с этой пробой, был назван клоном 33b.
IV.A.13. Последовательность нуклеотидов кольцевой ДНК в клоне 33b
Нуклеотидную последовательность в клоне 33b определяли по существу так, как описано в разделе IV.A.2. Последовательность, участок ее перекрытия с последовательностью кольцевой ДНК в клоне 32 и предполагаемый полипептид, закодированный в ней, представлены на фиг. 11.
Клон 33b очевидно содержит одну непрерывную открыто читаемую структуру, которая является протяжением открыто читаемых структур в перекрывающихся клонах 37b, 35, 36, 81 и 32. Полипептид, закодированный в клоне 33b, находится в той же самой трансляционной структуре, что и полипептид, закодированный в протяженной открыто читаемой структуре этих перекрывающихся клонов.
IV.A.14. Выделение перекрывающихся кольцевых ДНК вируса гепатита C с кольцевой ДНК клона 37b и с кольцевой ДНК в клоне 33b
Для выделения кольцевых ДНК вируса гепатита C, которые перекрывают кольцевые ДНК в клонах 37b и 33b, были использованы следующие синтетические пробы, полученные из кольцевых ДНК в тех клонах, которые использовали для проверки пула лямбда-gt11, используя по существу метод, описанный в разделе IV.A.3. Использованные пробы были:
5' CAG GAT GCT GTC TCC CGC ACT CAA CGT C 3'
и
5' TCC TGA GGC GAC TGC ACC AGT GGA TAA GCT 3',
для определения колоний, содержащих последовательности кольцевой ДНК вируса гепатита C, которые перекрывают последовательности кольцевых ДНК в клонах 37b и 33b соответственно. Примерно по одной из 50000 колоний было определено с каждой пробой. Клон, содержащий кольцевую ДНК, восходящую и перекрывающую кольцевую ДНК в клоне 37b был назван клоном 40b. Клон, который содержал нисходящую кольцевую ДНК, которая перекрывала кольцевую ДНК в клоне 33b, был назван клоном 25с.
IV. A. 15. Нуклеотидная последовательность кольцевой ДНК в клоне 40b и в клоне 25c
Последовательности нуклеотидов в кольцевой ДНК в клоне 40b и в клоне 25c определяли по существу так, как описано в разделе IV.A.2. Последовательности 40b и 25c, их участки перекрытия с кольцевыми ДНК в клонах 37b и 33b и предполагаемые полипептиды, закодированные в них, представлены на фиг. 12 (клон 40b) и на фиг. 13 (клон 25c).
5'-концевой нуклеотид клона 40b является C. Однако кольцевые ДНК от пяти других независимых клонов, которые были выделены в процессе выделения клона 40b, описанного в разделе IV.A.14, были также подвергнуты исследованиям на определение последовательностей. Кольцевые ДНК от этих клонов также содержали нуклеотид T в этом положении. Таким образом, G может представлять артефакт клонирования (см. обсуждение в разделе IV.A.11).
5'-терминирующим кодоном клона 25c является ACT, но последовательность этого участка в клоне cla (последовательность не показана) и в клоне 33b является TCA. Это различие может также представлять артефакт клонирования, как могут представлять таковой 28 лишних 5'-терминальных нуклеотидов в клоне 5-1-1.
Клоны 40b и 25c каждый очевидно содержат открыто читаемую структуру, которая является протяженностью непрерывной открыто читаемой структуры в клонах с' ранее определенными последовательностями. Нуклеотидная последовательность открыто читаемой структуры, простирающейся через клоны 40b, 37b, 35, 36, 81, 32, 33b и 25c, и последовательность аминокислот предполагаемого полипептида, закодированного в ней, представлены на фиг. 14. На этой фигуре потенциальные артефакты исключены из последовательности и вместо этого показаны соответствующие последовательности в неконцевых 5'-участках многократного перекрытия клонов.
IV. A.16. Получение композиционной кольцевой ДНК из кольцевых ДНК в клонах 36, 81 и 32
Композиционная кольцевая ДНК вируса гепатита C, C100, была создана следующим образом. Сначала вырезали кольцевые ДНК из клонов 36, 81 и 32 с EcoRI. Фрагмент EcoRI кольцевой ДНК из каждого клона индивидуально клонировали в локализацию EcoRI вектора pGEM3-синего (Promega Biotec). Полученные рекомбинантные векторы, которые содержали кольцевые ДНК от клонов 36, 81 и 32, были названы как pGEM3-синий/36, pGEM3-синий/81 и pGEM3-синий/32 соответственно. Подходяще ориентированный рекомбинант pGEM3-синего/81 сбраживали NaeI и NarI, и крупный (≈2850 п.о.) фрагмент был очищен и лигирован небольшим (≈570 п.о.) очищенным смесью NaeI-NarI ограничивающим фрагментом из вектора pGEM3-синего/36. Этот композит кольцевых ДНК из клонов 36 и 81 был использован для генерирования другого pGEM3-синего вектора, содержащего непрерывную открыто читаемую структуру вируса гепатита C, заключенную в перекрывающихся кольцевых ДНК в этих клонах. Полая плазмида была затем сброжена с PvuII и EcoRI для освобождения фрагмента размером примерно 680 п.о., который затем лигировали небольшим (580 п.о.) фрагментом PvuII-EcoRI, выделенным из соответственно ориентированной плазмиды pGEM3-синяя/32? и композиционная кольцевая ДНК из клонов 36, 81 и 32 была лигирована в вытянутый в линию EcoRI вектор pSODcf1, который описан в разделе IV.B.1 и который использовали для экспрессии клона 5-1-1 в бактериях. Рекомбинанты, содержащие 1270 п. о. EcoRI фрагмент композиционной ДНК вируса гепатита C (C100), были отобраны и кольцевая ДНК из плазмид была выращена с EcoRI и очищена.
IV.A.17. Выделение нуклеотидных последовательностей кольцевых ДНК вируса гепатита C в клонах 14i, 11b, 7f, 7e, 8h, 33c, 14c, 8f, 33f, 33g и 39c
Кольцевые ДНК вируса гепатита C в клонах 14i, 11b, 7f, 7e, 8h, 33c, 14c, 8f, 33f, 33g и 39c выделяли методами выделения фрагментов перекрывающихся кольцевых ДНК, из пула кольцевых ДНК вируса гепатита C лямбда-gt11, описанных в разделе IV. A.1. Использованный метод по существу был аналогичен описанному в разделе IV.A.3, за исключением того, что использованные пробы были получены от нуклеотидной последовательности последних выделенных клонов от 5'- и 3'-окончаний объединенных последовательностей вируса гепатита C. Частота клонов, гибридизирующихся с пробами, описана ниже и составляла примерно один клон на 50000 в каждом случае.
Последовательности нуклеотидов кольцевых ДНК вируса гепатита C в клонах 14i, 7f, 7e, 8h, 33c, 14c, 8f, 33f, 33g и 39c определяли по существу так, как описано в разделе IV.A.2, за исключением того, что кольцевую ДНК, выделенную из клона 5-1-1, заменяли на кольцевые ДНК, вырезанные из этих фагов.
Клон 33c выделяли, используя гибридизационную пробу, основанную на последовательности нуклеотидов в клоне 40b. Нуклеотидная последовательность клона 40b представлена на фиг. 12. Нуклеотидная последовательность пробы, использованной для выделения клона 33c, была следующей:
5' ATC ACG ACC GGG GTG ACA ACA ATT ACC ACT 3'.
Последовательность кольцевой ДНК вируса гепатита C в клоне 33c и перекрытие с последовательностью в клоне 40b показано на фиг. 15, на которой также представлена последовательность аминокислот, закодированных в ней.
Клон 8h был выделен с использованием пробы, основанной на последовательности нуклеотидов в клоне 33c. Нуклеотидная последовательность пробы была:
5' AGA GAC AAC CAT GAG GTC CCC GGT GTT C 3'.
Последовательность кольцевой ДНК вируса гепатита C в клоне 8h, перехлест с последовательностью в клоне 33c и аминокислоты, закодированные в ней, представлены на фиг. 16.
Клон 7e был изолирован с использованием пробы, основанной на последовательности нуклеотидов в клоне 8h. Последовательность нуклеотидов пробы была:
5' TCG GAC CTT TAC CTG GTC ACG AGG CAC 3'.
Последовательность кольцевой ДНК вируса гепатита C в клоне 7е, перекрытие с клоном 8h и аминокислоты, закодированные в ней, представлены на фиг. 17.
Клон 14C был выделен с пробой, основанной на последовательности нуклеотидов в клоне 25c. Последовательность клона 25C показана на фиг. 13. Проба для выделения клона 14C имела последовательность:
5' ACC TTC CCC ATT AAT GCC TAC ACC ACG GGC 3'.
Последовательность кольцевой ДНК вируса гепатита C в клоне 14c, ее перекрытие с последовательностью в клоне 25c и закодированные в ней аминокислоты представлены на фиг. 18.
Клон 8f был выделен с использованием пробы, основанной на последовательности нуклеотидов в клоне 14с. Последовательность нуклеотидов пробы была:
5' TCC ATC TCT CAA GGC AAC TTG CAC CGC TAA 3'.
Последовательность кольцевой ДНК вируса гепатита C в клоне 8f, ее перекрытие с последовательностью в клоне 14с и аминокислоты, закодированные в ней, представлены на фиг. 19.
Клон 33f был выделен с использованием пробы, основанной на последовательности, присутствующей в клоне 8f. Последовательность нуклеотидов пробы была:
5' TCC ATG GCT GTC CGC TTC CAC CTC CAA AGT 3'.
Последовательность кольцевой ДНК вируса гепатита C в клоне 33f, ее перекрытие с последовательностью в клоне 8f и аминокислоты, закодированные в ней, представлены на фиг. 20.
Клон 33g был выделен с использованием пробы, основывающейся на последовательности нуклеотидов в клоне 33f. Последовательность нуклеотидов пробы была:
5' GCG ACA ATA CGA CAA CAT CCT CTG AGC CCG 3'.
Последовательность кольцевой ДНК вируса гепатита C в клоне 33g, ее перекрытие с последовательностью в клоне 33f и закодированная в ней последовательность аминокислот представлены на фиг. 21.
Клон 7f выделяли, используя пробу, основанную на последовательности нуклеотидов в клоне 7e. Нуклеотидная последовательность пробы была:
5' AGC AGA CAA GGG GCC TCC TAG GGT GCA TAA T 3'.
Последовательность кольцевой ДНК вируса гепатита C в клоне 7f, ее перекрытие с клоном 7e и аминокислоты, закодированные в ней, представлены на фиг. 22.
Клон 11b был выделен с использованием пробы, основанной на последовательности клона 7f. Нуклеотидная последовательность пробы была:
5' CAC CTA TGT TTA TAA CCA TCT CAC TCC TCC 3'.
Последовательность кольцевой ДНК вируса гепатита C в клоне 11b, ее перекрытие с клоном 7f и аминокислоты, закодированные в ней, представлены на фиг. 23.
Клон 14i выделяли с использованием пробы, основанной на последовательности нуклеотидов в клоне 11b. Последовательность нуклеотидов в пробе была:
5' CTC TGT CAC CAT ATT ACA AGC GCT ATA TCA 3'.
Последовательность кольцевой ДНК вируса гепатита C в клоне 14i, ее перекрытие с 11b и закодированные в ней аминокислоты представлены на фиг. 24.
Клон 39c был выделен с использованием пробы, основанной на последовательности нуклеотидов в клоне 33g. Последовательность нуклеотидов пробы была:
5' CTC GTT GCT ACG TCA CCA CAA TTT GGT GTA 3'.
Последовательность кольцевой ДНК вируса гепатита C в клоне 39c, ее перекрытие с клоном 33g и закодированные в ней аминокислоты, представлены на фиг. 25.
IV.A.18. Последовательность композиционной кольцевой ДНК вируса гепатита C, полученной от выделенных клонов, содержащих кольцевую ДНК вируса гепатита C
Последовательности кольцевой ДНК вируса гепатита C, описанные выше, выстраивали в линию, чтобы создать последовательность композиционной кольцевой ДНК вируса гепатита C. Выделенными клонами, упорядоченными в линию в направлении 5' - 3', были: 14i, 7f, 7e, 8h, 33c, 40b, 37b, 35, 36, 81, 32, 33b, 25c, 14c, 8f, 33f, 33g и 39c.
Последовательность композиционной кольцевой ДНК вируса гепатита C, полученная от выделенных клонов, и закодированные в ней аминокислоты показаны на фиг. 26.
При создании композиционной последовательности были приняты во внимание следующие гетерогенности последовательности. Клон 33c содержит кольцевую ДНК вируса гепатита C с 800 парами оснований, которая перекрывает кольцевую ДНК в клонах 40b и 37c. В клоне 33c, а также и в пяти других перекрывающихся клонах нуклеотидом N 789 является гуанин. Однако в клоне 37b (см. раздел IV. A. 11), соответствующим нуклеотидом является аланин. Различие последовательностей создает очевидную гетерогенность в аминокислотах, закодированных в них, которые будут либо цис, или тир для гуанина или аденина соответственно. Эта гетерогенность может иметь важные разветвления с точки зрения белкового синтеза.
Нуклеотидный остаток N 2 в клоне 8h кольцевой ДНК вируса гепатита C является тимином. Однако, как показано ниже, соответствующим остатком в клоне 7e является аденин; более того, аденин в этом положении найден также в трех других изолированных перекрывающихся клонах. Таким образом, тиминовый остаток в клоне 8h может представлять собой артефакт клонирования. Поэтому на фиг. 26 остаток в этом положении обозначен как A (аденин).
3'-концевой нуклеотид в клоне 8f кольцевой ДНК вируса гепатита C является гуанином. Однако соответствующим остатком в клоне 33f и в двух других перекрывающихся клонах является тимин (T). Поэтому на фиг. 26 остаток в этом положении обозначен как T.
3'-терминирующая последовательность в клоне 33f кольцевой ДНК вируса гепатита C является TTGC. Однако соответствующей последовательностью в клоне 33g и в двух других перекрывающихся клонах является АТТС. Поэтому на фиг. 26 соответствующий участок представлен как АТТС.
Нуклеотидный остаток N 4 в клоне 33g кольцевой ДНК вируса гепатита C является T. Однако в клоне 33f и в двух других перекрывающихся клонах соответствующим остатком является A. Поэтому на фиг. 26 соответствующий участок обозначен как A.
3'-терминирующий кодон клона 14i является AA, тогда как соответствующий динуклеотид в клоне 11b и в трех других клонах является TA. Следовательно, на фиг. 26 указан остаток TA.
Разделение других гетерогенных последовательностей рассмотрено выше.
Испытания композиционной кольцевой ДНК вируса гепатита C показывают, что она содержит одну большую открыто читаемую структуру. Это предполагает, что вирусный геном транслирован в большой полипептид, который подвергается обработке сопутствующе или последовательно с трансляцией.
IV.A.19. Выделение нуклеотидных последовательностей кольцевых ДНК вируса гепатита C в клонах 12f, 35f, 19g, 26g и 15e
Кольцевые ДНК вируса гепатита C в клонах 12f, 35f, 19g, 26g и 15e выделяли по существу методом, описанным в разделе IV.A.17, за исключением того, что использовали указанные ниже пробы. Частота клонов, которые гибридизировались с пробами, составляла примерно один клон из 50000 в каждом случае. Нуклеотидные последовательности кольцевых ДНК в этих клонах определяли в основном методом, описанным в разделе IV.A.2, за исключением того, что кольцевую ДНК, выделенную из клона 5-1-1, заменяли кольцевой ДНК указанных клонов.
Выделение клона 12f, который содержит кольцевую ДНК восходяще к кольцевой ДНК вируса гепатита C на фиг. 26, проводили с использованием гибридизационной пробы, основанной на последовательности нуклеотидов в клоне 14i. Нуклеотидная последовательность пробы была:
5' TGC TTG TGG ATG ATG CTA CTC ATA TCC CAA 3'.
Последовательность кольцевой ДНК вируса гепатита C в клоне 12f, ее перехлест с клоном 14i и аминокислоты, закодированные в ней, представлены на фиг. 27.
Выделение клона 35f, который содержит кольцевую ДНК нисходяще к кольцевой ДНК вируса гепатита C на фиг. 26, осуществляли с использованием гибридизационной пробы, основанной на последовательности нуклеотидов в клоне 39c. Нуклеотидная последовательность пробы была:
5' AGC AGC GGC GTC AAA AGT GAA GGC TAA CTT 3'.
Последовательность клона 35f, ее перехлест с последовательностью в клоне 39c и аминокислоты, закодированные в ней, показаны на фиг. 28.
Выделение клона 19g проводили с использованием гибридизационной пробы, основанной на 3'-последовательности клона 35f. Нуклеотидная последовательность пробы была:
5' TTC TCG TAT GAT ACC CGC TGC TTT GAC TCC 3'.
Последовательность кольцевой ДНК вируса гепатита C в клоне 19g, ее перекрытие с последовательностью в клоне 35f и аминокислоты, закодированные в ней, представлены на фиг. 29.
Выделение клона 26g проводили с использованием гибридизационной пробы, основанной на 3'-последовательности клона 19g. Нуклеотидная последовательность пробы была:
5' TGT GTG GCG ACG ACT TAG TCG TTA TCT GTG 3'.
Последовательность кольцевой ДНК вируса гепатита C клона 26g, ее перекрытие с последовательностью в клоне 19g и аминокислоты, закодированные в ней, представлены на фиг. 30.
Клон 15e выделяли с использованием гибридизационной пробы, основанной на 3'-последовательности клона 26g. Нуклеотидная последовательность пробы была:
5' CAС ACT CСA GTC AAT TCC TGG CTA GGC AAC 3'.
Последовательность кольцевой ДНК вируса гепатита C в клоне 15e, ее перекрытие с последовательностью в клоне 26g и аминокислоты, закодированные в ней, представлены на фиг. 31.
Клоны, описанные в этом разделе, депонированы в Американской коллекции культур клеток под названиями и при условиях, описанных в разделе II.A, и зарегистрированы под номерами, представлеными в табл. Б.
Последовательности кольцевой ДНК вируса гепатита C в выделенных клонах описаны выше и упорядочены в линию для создания последовательности композиционной кольцевой ДНК вируса гепатита C. Выделенные клоны, упорядоченные в линию в направлении 5'-3' являются: 12f, 14i, 7f, 7e, 8h, 33c, 40b, 37b, 35, 36, 81, 32, 33b, 25c, 14c, 8f, 33f, 33g, 39c, 35f, 19g, 26g и 15e.
Последовательность композиционной кольцевой ДНК вируса гепатита C, полученная из выделенных клонов, и закодированные в ней аминокислоты, представлены на фиг. 32.
IV. A. 20. Альтернативный метод выделения последовательностей кольцевой ДНК, восходящей к последовательности кольцевой ДНК вируса гепатита C в клоне 12f
Основываясь большей частью на 5'-последовательности вируса гепатита C на фиг. 32, полученной от кольцевой ДНК вируса гепатита C в клоне 12f, синтезировали небольшие синтетические олигонуклеотидные затравки обратной транскриптазы и использовали их для связывания с соответствующей последовательностью в геномной РНК вируса гепатита C, чтобы инициировать обратную транскрипцию восходящих последовательностей. Инициирующие последовательности близко родственны известной 5'-терминирующей последовательности клона 12f, но значительно ниже, чтобы допустить конструирование последовательностей проб в восходящем направлении к последовательностям затравки. Использовали известные методы затравки и клонирования. Полученные пулы кольцевой ДНК исследовали последовательностями восходяще к инициирующим локализациям (как выведено из разъясненной последовательности в клоне 12f). Геномную РНК вируса гепатита C получали либо из плазмы, либо из образцов печени шимпанзе, с гепатитом ни A, ни B, или из аналогичных образцов человека с гепатитом ни A, ни B.
IV. A. 21. Альтернативный метод использования отростка вириона для выделения последовательностей из 5'-терминирующего участка генома вируса гепатита C
Чтобы выделить дополнительные 5'-концевые последовательности РНК генома вируса гепатита C, продукт кольцевой ДНК первой стадии обратной транскрипции, который удвоен матрицей РНК, образует отросток с олигоцистином. Это достигается инкубированием продукта с терминирующей трансферазой в присутствии СТР. Второй раунд синтеза кольцевой ДНК, который дает комплемент первой нити кольцевой ДНК, осуществляется с использованием олигогуанина в качестве затравки для реакции обратной транскриптазы. Источники геномной РНК вируса гепатита C описаны в разделе IV.A.20. Методы формирования отростков с терминирующей трансферазой и для реакции обратной транскриптазы описаны Maniatis и соавт. (1982). Затем продукты кольцевой ДНК анализировали и определяли их последовательности.
IV. A.22. Альтернативные методы выращивания отростков для выделения последовательностей из 3'-концевого участка генома вируса гепатита C
Этот метод основан на ранее использовавшихся методах клонирования кольцевых ДНК РНК содержащего флавивируса. В этом методе РНК помещают в условиях денатурирования для удаления вторичных структур при 3'-терминирующем кодоне, и затем выращивают отросток с поли-A-полимеразой, используя r-аденозинтрифосфат в качестве субстрата.
Обратную транскрипцию поли-A-отростковой РНК катализируют, используя олиго dT в качестве затравки. Затем синтезируют вторую нить кольцевой ДНК и продукты клонируют, отсеивают и определяют последовательности.
IV. A. 23. Создание пулов лямбда-gt11 кольцевой ДНК вируса гепатита C, содержащих крупные вставки кольцевой ДНК
Метод, использованный для создания и проверки пулов лямбда-gt11, по существу, тот же, что описан в разделе IV.A.1, за исключением того, что этот пул создавали из более крупных размеров кольцевых ДНК, элюированных из колонки сефароза CL-4B.
IV. A.24. Создание пулов кольцевой ДНК вируса гепатита C, используя синтетические олигомеры в качестве инициаторов реакции
Новые пулы кольцевой ДНК вируса гепатита C создавали из РНК, полученной из пула плазмы инфекционного шимпанзе, описанного в разделе IV.A.1, и из фракции поли-A+ РНК, полученной из печени инфицированного животного. Кольцевую ДНК конструировали в основном как описано Gubler и Hoffman (1983), за исключением того, что затравками для синтеза первой нити кольцевой ДНК служили два синтетических олигомера, основанных на описанной выше последовательности генома вируса гепатита C. Затравки, основанные на последовательности клона 11b и 7e, были соответственно следующими:
5' CTG GCT TGA AGA ATC 3' и
5' AGT TAG GCT GGT GAT TAT GC 3'.
Полученные кольцевые ДНК клонировали в векторы лямбда-бактериофага, испытывали с различными другими синтетическими олигомерами, последовательность которых базировалась на последовательности, представленной на фиг. 32.
IV. B. Экспрессия полипептидов, закодированных в кольцевых ДНК вируса гепатита C, и идентификация воспроизведенных продуктов как индуцированных антигенов вируса гепатита C
IV.B.1. Экспрессия полипептида, закодированного в клоне 5-1-1
Полипептид, закодированный в клоне 5-1-1 (см. раздел IV.A.2 выше), воспроизводили как полипептид слияния с пероксиддисмутазой. Это осуществлялось субклонированием кольцевой ДНК клона 5-1-1, вставкой в вектор экспрессии pSODcfl (Steimer и соавт., 1986 г.), следующим образом.
Сначала ДНК, выделенную из вектора pSODcfl обрабатывали BamHI и EcoRI и следующее присоединение было лигировано в линейную ДНК, созданную ограничивающими ферментами:
5' GAT CCT GGA ATT CTG ATA A 3',
3' GA CCT TAA GAC TAT TTT AA 5'.
После клонирования плазмида, содержащая вставку, изолировалась.
Плазмида, содержащая вставку, была ограничена EcoRI. Вставка кольцевой ДНК вируса гепатита C в клон 5-1-1 была вырезана EcoRI и, лигированная в этой фермент EcoRI, упорядочивала в линию плазмидную ДНК. Смесь ДНК использовали для преобразования штамма E.coli D1210 (Sadler и соавт., 1980 г.). Рекомбинанты с кольцевой ДНК 5-1-1, правильно ориентированные на экспрессию открыто читаемой структуры, показаны на фиг. 1. Они были идентифицированы ограничительным картированием и определением нуклеотидной последовательности.
Рекомбинантные бактерии одного клона были индуцированы для экспрессии полипептида пероксиддисмутаза-ни A, ни B5-1-1 выращиванием бактерий в присутствии IPTG.
IV.B.2. Экспрессия полипептида, закодированного в клоне 81
Кольцевая ДНК вируса гепатита C, содержавшаяся в клоне 81, была воспроизведена как полипептид слияния пероксиддисмутаза-ни A, ни B81. Метод получения вектора, кодирующего этот полипептид слияния, был аналогичен методу, использованному для создания вектора, кодирующего пероксиддисмутазы-ни A, ни B5-1-1, за исключением того, что источником кольцевой ДНК вируса гепатита C был клон 81, который был выделен, как описано в разделе IV.A.3, и для которого последовательность кольцевой ДНК была определена, как описано в разделе IV. A. 4. Последовательность нуклеотидов кольцевой ДНК вируса гепатита C в клоне 81 и предполагаемая последовательность аминокислот полипептида, закодированного в ней, представлена на фиг. 4.
Вставка кольцевой ДНК вируса гепатита C в клон 81 была вырезана посредством EcoRI и лигирована в pSODcf1, который содержал присоединение (см. раздел IV. B. 1.) и который был упорядочен в линию обработкой EcoRI. Вмесь ДНК использовали для преобразования штамма E.coli D1210. Рекомбинанты с кольцевой ДНК вируса гепатита C клона 81 в правильной ориентации на экспрессию открыто читаемой структуры, представленные на фиг. 4, были идентифицированы ограничивающим картированием и определением последовательностей нуклеотидов.
Рекомбинантные бактерии одного клона были стимулированы на экспрессию полипептида пероксиддисмутаза-ни A, ни B81 выращиванием бактерий в присутствии IPTG.
IV.B.3. Идентификациия полипептида, закодированного клоне 5-1-1 как антигена, связанного с гепатитом C и гепатитом ни A, ни B
Полипептид, закодированный в кольцевой ДНК вируса гепатита C клона 5-1-1, был идентифицирован как антиген, связанный с гепатитом ни A, ни B путем демонстрации того, что сыворотки шимпанзе и людей, инфицированных гепатитом ни A, ни B, иммунологически реагировали с полипептидом слияния пероксиддисмутаза-ни A, ни B5-1-1, который содержал пероксиддисмутазу в своем N-окончании и структурный 5-1-1 антиген в своем C-концевом кодоне. Это осуществляли "западным" пятнением (Towbin и соавт., 1979 г.) следующим образом.
Рекомбинантный штамм бактерий, трансформированный вектором экспрессии, кодирующим полипептид пероксиддисмутаза-ни A, ни B5-1-1, описанный в разделе IV. B. 1, был стимулирован на экспрессию полипептида слияния выращиванием в присутствии IPTG. Общий бактериальный лизат подвергали электрофорезу через полиакриламидные гели в присутствии додецилсульфата натрия по методу Laemmli (1970). Разделенные полипептиды переносили на нитроцеллюлозные фильтры (Towbin и соавт., 1979 г.). Затем фильтры разрезали на узкие полоски и полоски инкубировали отдельно с различными сыворотками шимпанзе и человека. Связанные антитела определяли дальнейшим инкубированием с меченной йодом-125 сывороткой овцы против Ig человека, как описано в разделе IV.A.1.
Характеристики сывороток шимпанзе, использованных для "западных" пятен, и результаты, показанные на фотографии ауторадиографированных полосок, представлены на фиг. 33. Нитроцеллюлозные полосы, содержащие полипептиды, инкубировали с сыворотками, полученными от шимпанзе в различные периоды времени в процессе острой инфекции гепатита на A, ни B (линия Hutchinson) (строки 1-16), инфекции гепатита A (строки 17-24 и 26-33) и инфекции гепатита B (строки 34-44). Строки 25 и 45 показывают положительные контроли, в которых иммунные пятна инкубировали с сывороткой больного, использованной для идентификации рекомбинантного клона 5-1-1 в первоначальном скрининге пула кольцевой ДНК лямбда-gt11 (см. раздел IV.A.1).
Полоса, видимая на контрольных строках 25 и 45 на фиг. 23, отражает связывание антител в части ни A, ни B5-1-1 полипептида слияния с пероксиддисмутазой. Эти антитела не показывают связывания только с пероксиддисмутазой, поскольку она была включена также как отрицательный контроль в эти образцы и появилась как полоса, мигрирующая значительно быстрее, чем полипептид слияния пероксиддисмутаза-ни A, ни B5-1-1.
Строки 1-16 на фиг. 33 показывают связывание антител в образцах сывороток четырех шимпанзе: сыворотки были получены как раз перед инфекцией гепатитом ни A, ни B и последовательно в процессе острой инфекции. Как видно на фигуре, в то время как антитела, которые реагировали иммунологически с полипептидом пероксиддисмутаза-ни A, ни B5-1-1, отсутствовали в образцах сывороток, полученных до введения инокулята инфекции вируса гепатита C и в ранней острой фазе инфекции, все четыре животных в конечном счете индуцировали циркулирующие антитела к этому полипептиду на последней части острой фазы или в последующей за острой фазой. Дополнительные полосы, наблюдаемые на иммунологических пятнах в случае шимпанзе номер 3 и 4, были обусловлены фоном связывания с белками бактериального хозяина.
В противоположность этим результатам, полученным с сывороткой от шимпанзе, инфицированных гепатитом ни A, ни B, образования антител к части молекулы ни A, ни B5-1-1 полипептида слияния не наблюдали у 4 шимпанзе, инфицированных вирусом гепатита A, или у трех шимпанзе, инфицированных вирусом гепатита B. Единственным связыванием в этих случаях было фоновое связывание с белками бактериального хозяина, которое появляется также в инфицированных вирусом гепатита C образцах.
Характеристики сывороток человека, использованных для получения "западных" пятен, и результаты, которые показаны на фотографии ауторадиографированных полос, представлены на фиг. 34. Нитроцеллюлозные полосы, содержащие полипептиды, были инкубированы с сывороткой, полученной от лиц в различные периоды времени инфекции гепатитом ни A, ни B (строки 1-21), вирусом гепатита A (строки 33-40) и вирусом гепатита B (строки 41-49). Строки 25 и 50 показывают положительные контроли, в которых иммунологические пятна были инкубированы с сывороткой больного, использованной для первоначального отбора в пуле лямбда-gt11, описанного выше. Строки 22-24 и 26-32 показывают "неинфицированные" контроли, в которых сыворотка получена от "нормальных" доноров крови.
Как видно на фиг. 34, сыворотка от девяти больных гепатитом ни A, ни B, включая сыворотку, использованную для скрининга пула лямбда-gt11, содержали антитела к части ни A, ни B5-1-1 полипептида слияния. Сыворотки трех больных гепатитом ни A, ни B не содержали этих антител. Возможно, что антитела против ни A, ни B5-1-1 будут развиваться в будущем у этих больных. Возможно также, что отсутствие реакции проистекает от различий возбудителя - вируса ни A, ни B, ответственного за вызывание болезни у лиц, у которых была взята нереагирующая сыворотка.
На фиг. 34 показано также, что сыворотки многих больных, инфицированных вирусом гепатита A и вирусом гепатита B, не содержат антител против ни A, ни B5-1-1 и что эти антитела не присутствуют также в сыворотках от "нормальных" контролей. Хотя один больной вирусом гепатита A (строка 36) очевидно содержит антитела против ни A, ни B5-1-1, возможно, что этот больной был прежде инфицирован вирусом гепатита C, поскольку распространенность заболеваний гепатитом ни A, ни B очень высока и поскольку он часто бессимптомный.
Эти серологические исследования показывают, что кольцевая ДНК в клоне 5-1-1 кодирует эпитопы, которые распознаются специфически сыворотками больных людей и животных, инфицированных вирусом BB-ни A, ни B. В дополнение кольцевая ДНК не кажется явно полученной от генома приматов. Гибридизационная проба, приготовленная из клона 5-1-1 или из клона 81, не гибридизирует до "южных" пятен геномную ДНК и контрольного человека и шимпанзе от неинфицированных индивидов в условиях, при которых определялись уникальные однокопийные гены. Эти пробы не гибридизировали также до южных пятен контрольную геномную ДНК бычьей сыворотки.
IV. B. 4. Экспрессия полипептида, закодированного в композите кольцевых ДНК вируса гепатита C в клонах 36, 81 и 32
Полипептид вируса гепатита C, который закодирован в открыто читаемой структуре, протянувшейся через клоны 36, 81 и 32, был воспроизведен как полипептид слияния с пероксиддисмутазой. Это осуществляли встраиванием композиционной кольцевой ДНК, C100, в вектор экспрессии и экспрессией полипептида в дрожжах.
Кассету для проведения экспрессии, содержащую композит C100 кольцевой ДНК, полученный из клонов 36, 81 и 32, конструировали встраиванием фрагмента ≈1270 п. о. EcoRI в локализацию EcoRI вектора р33-56 (называемого также pS356), дающего плазмиду pS3-56C100. Конструирование C100 описано в разделе IV.A.16 выше.
Вектор рS3-56, производный от pBP322, содержит экспрессионную кассету, которая содержит гибрид ADH2/GAPDH дрожжевого промотора восходяще к гену пероксиддисмутазы человека и нисходящей GAPDH окончания транскрипции. Аналогичная кассета, которая содержит эти контрольные элементы и ген проксиддисмутазы, была описана Cousens и соавт. (1987) и в продолжающейся заявке на Европейский патент N 196056, опубликованной 1 октября 1986 года, принадлежащей настоящим заявителям. Кассета в векторе рS3-56 отличается, однако, от кассеты у Cousens и соавт. (1987) тем, что гетерологичный ген проинсулина и иммуноглобулиновая петля утрачены, и тем, что ген пероксиддисмутазы сопровождается последовательностью адаптера, который содержит локализацию EcoRI. Последовательность адаптера такова:
5'-AAT TTG GGA ATT CCA TAA TGA G-3'
AC CCT TAA GGT ATT ACT CAG CT
Локализация EcoRI позволяет встраивание гетерологичных последовательностей, которые, будучи воспроизведены из вектора, содержащего кассету, дают полипептиды, слитые с пероксиддисмутазой через олигопептидную связку, содержащую последовательность аминокислот:
-ans-leu-gly-ile-arg-
(-аспарагин-лейцин-глицин -изолейцин-аргинин-).
Образец вектора pS356 депонирован 29 апреля 1988 года в Американском собрании типов культур (АТСС). 12301 Parklawn Dr., Rockville, Maryland 20853 в терминах Budapest/Treaty и ему присвоили номер 67683. Сроки и условия доступа к депонированному вектору такие же, что и для штаммов, содержащих кольцевые ДНК вируса гепатита ни A, ни B, определенные в разделе II.A. Депозит имеется в виду только для удобства и не требуется для осуществления настоящего изобретения при наличии настоящего описания. Депонированный материал приведен в библиографической справке.
После того как рекомбинанты, содержащие вставку C100 кольцевой ДНК в точной ориентации, были выделены, экспрессионную кассету, содержащую C100 кольцевой ДНК, вырезали из вектора pS3-56C100 с помощью BamHI, и фрагмент ≈3400 п.о., который содержит кассету, выделяли и очищали. Этот фрагмент был затем встроен в локализацию BamHI дрожжевого вектора рAB24.
Плазмида рAB24, существенные признаки которой показаны на фиг. 35, является дрожжевым челночным вектором, который содержит полную двухмикронную последовательность для репликации (Broach, 1981 г.) и последовательность pBR322. Она содержит также дрожжевой URA3 ген, производный от плазмиды YEp24 (Botstein и соавт., 1979 г.) и дрожжевой LEU2d ген, полученный из плазмиды pCl/1 (публикация о Европейском патенте N 116201). Плазмида рAB24 была синтезирована ферментацией плазмиды YEp24 с EcoRI и религацией вектора для удаления частичных двухмикронных последовательностей. Полученная плазмида, YEP24deltaRI была упорядочена линейно ферментацией с ClaI и лигирована полной двухмикронной плазмидой, которая была упорядочена в линию с ClaI. Полученная плазмида pCBou была затем сброжена с XbaI и фрагмент вектора 8605 п. о. был выделен гелевой фильтрацией. Этот выделенный фрагмент XbaI был лигирован фрагментом 4460 п. о. XbaI, содержащим ген LEU2d, выделенный из pCl/1; ориентация LEU2d гена в том же направлении, что и гена URA3. Встраивание экспрессии осуществлено в уникальную локализацию BamHI последовательности pBR322, прерывая таким образом ген для бактериальной резистентности к тетрациклину.
Рекомбинантная плазмида, содержащая пероксиддисмутаза-C100 экспрессионную кассету рAB24C100-3, была трансформирована в дрожжевой штамм JSC 308, а также в другие дрожжевые штаммы. Клетки были видоизменены, как описано Hinnen и соавт. (1978), и выращены на урацилселективных пластинках. Отдельные колонии были инокулированы в лейцинселективную среду для выращивания. Культуру стимулировали на экспрессию полипептида пероксиддисмутаза-C100 (назван C100-3) выращиванием в YEP, содержащем 1% глюкозы.
Штамм JSC 308 принадлежит к генотипу MAT @ leu2, ura3 (del) DM15 (GAP/ADR1), интегрированных в локусе ADR1. В штамме JSC 308 сверхэкспрессия положительного активатора генного продукта ADR1 приводит к гипердепрессии (относительно контроля дикого типа ADR1) и к значительно более высоким выходам экспрессивных гетерологичных белков, если такие белки синтезируют через ADH2 UAS регуляторную систему. Конструирование дрожжевого штамма JSC 308 описано в продолжающейся заявке США серийный номер (квитанция проверенного N 2300-0229), зарегистрированной одновременно, и которая приведена в библиографической ссылке. Образец JSC 308 был депонирован 5 мая 1988 года в АТСС по условиям Будапештского договора и получил порядковый номер 20879. Сроки хранения и условия доступа к депозиту такие же, что и описанные в разделе II.A для штаммов, содержащих кольцевые ДНК вируса гепатита C.
Полный C100-2 полипептид слияния, закодированный в pAB24C100-3, должен содержать 154 аминокислоты пероксиддисмутазы человека при аминотерминале, остаток пяти аминокислот от синтетического адаптера, содержащего локализацию EcoRI, остатки 363 аминокислот, производных от кольцевой ДНК C100 и 5 карбокситерминирующих аминокислот, производных от последовательности нуклеотида MS2, соединенной с последовательностью кольцевой ДНК в клоне 32 (см. раздел IV. A. 7). Предполагаемая последовательность аминокислот карбоксиокончания этого полипептида, начинающаяся при полуконечном Ala-остатке пероксиддисмутазы, показана на фиг. 36; показана также нуклеотидная последовательность, кодирующая эту часть полипептида.
IV.B.5. Идентификация полипептида, закодированного в C100 как связанного с гепатитом ни A, ни B антигеном
Полипептид слияния C100-3, полученный экспрессией из плазмиды рAB24C100-3 в дрожжевом штамме JSC 308, был охарактеризован с учетом размера и полипептид, закодированный в C100, был идентифицирован как связанный с гепатитом ни A, ни B антиген по его иммунологической реактивности с сывороткой человека, больного хроническим гепатитом ни A, ни B.
Полипептид C100-3, полученный экспрессией, как описано в разделе IV.B.4, подвергали анализу следующим образом. Дрожжевые клетки штамма JSC 308 были трансформированы дрожжевым вектором рAB24 или плазмидой рAB24C100-3 и стимулированы на экспрессию гетерологичной плазмиды закодированного полипептида. Стимулированные дрожжевые клетки в 1 мл культуры (OD650 нанометров ≈ 20) превращали в шарики центрифугированием при 10000 оборотов в минуту в течение одной минуты и затем подвергали лизису энергичным завихрением из (10 раз по одной минуте) с 2 объемами раствора и одним объемом стеклянных шариков (0,2 миллимикронного диаметра). Раствор содержал 50 мМ трис-HCl, pH 8,0, 1 мМ этилендиаминтетрауксусной кислоты, 1 мМ фтористого фенилметилсульфонила и микрограмм/мл пенстатина. Нерастворимый материал в лизате, который включает полипептид C100-3, собирали центрифугированием (при 10000 оборотов/мин в течение пяти минут) и растворяли кипячением в течение пяти минут в образце буфера додецилсульфата натрия (см. Laermmli, 1970 г.). Количество полипептидов, эквивалентное 0,3 мл стимулированной дрожжевой культуры, подвергали электрофорезу через 10% полиакриламидный гель в присутствии додецилсульфата натрия по методу Laemmli (1970). Стандартные белки подвергали параллельному электрофорезу на гелях. Гели, содержащие полипептиды, подвергнутые депрессии, или окрашивали бриллиантовым синим Coomassie, или подвергали "западному" пятнению, как описано в разделе IV.B.2, используя сыворотку больного хроническим гепатитом на A, ни B для определения иммунологической реактивности полипептидов, полученных экспрессией от рAB24 и рAB24C100-3.
Результаты представлены на фиг. 37. На фиг. 37A полипептиды окрашены бриллиантовым синим Coomassie. Нерастворимые полипептид(ы) от штамма JCS 308, трансформированные рAB24, и от двух различных колоний штамма JSC 308, трансформированного рAB24C100-3, показаны на строке 1 (рAB24) и на строках 2 и 3 соответственно. Сравнение строки 2 и строки 3 со строкой 1 показывает, что имеет место стимулированная экспрессия полипептида, соответствующего молекулярному весу ≈ 54000 дальтон от штамма JSC 308, трансформированного рAB24C100-3, которая не стимулирована в штамме JSC 308, трансформированном плазмидой рAB24. Этот полипептид указан стрелкой.
На фиг. 37B представлены результаты "западных" пятен нерастворимых полипептидов, полученных экспрессией в трансформированном плазмидой рAB24 штамме JSC 308 (строка 1) или плазмидой рAB24C100-3 (строка 2). Полипептиды, полученные экспрессией от рAB24, были иммунологически нереакционноспособными с сывороткой человека, больного гепатитом ни A, ни B. Однако, как показано стрелкой, штамм JSC 308, трансформированный плазмидой рAB24C100-3, синтезирует полипептид с молекулярным весом примерно 54000 дальтон, который реагирует иммунологически с сывороткой человека, больного гепатитом ни A, ни B. Другие иммунологически реакционноспособные полипептиды в строке 2 могут быть продуктами распада и/или агрегации этого полипептида с молекулярным весом около 54000 дальтон.
IV.B.6. Очистка полипептида слияния C100-3
Полипептид слияния C100-3, содержащий пероксиддисмутазу при N-окончании и структурный полипептид вируса гепатита C C100 при C-окончании, был очищен дифференциальной экстракцией нерастворимой фракции экстрагированных дрожжевых клеток хозяина, в которых был синтезирован полипептид.
Полипептид слияния C100-3 был синтезирован в штамме C308, трансформированном рAB24C100-3, как описано в разделе IV.B.4. Дрожжевые клетки затем подвергали лизису гомогенизацией, нерастворимые материалы в лизате экстрагировали при pH 12,0 и C100-3 в оставшейся нерастворимой фракции переводили в растворенное состояние в буфере, содержащем додецилсульфат натрия.
Дрожжевой лизат готовили в основном по методу Nagahuma и соавт. (1984). Получали суспензию дрожжевых клеток, содержащую 33% клеток, суспендированных в растворе (буфер A), состоящем из 20 мМ трис-HCl, pH 8,0, 1 мМ дитиотрейтола и 1 мМ фтористого фенилметилсульфонила. Аликвоту суспензии (15 мл) смешивали с равным объемом стеклянных сфер (диаметром 0,45-0,50 мм), завихряли с максимальной скоростью на супермиксере (Lab Line Instruments, Inc.) в течение восьми минут. Гомогенат и стеклянные шарики разделяли и стеклянные шарики три раза промывали таким же объемом буфера A, что и первоначально упакованные клетки. После объединения промывок и гомогената нерастворимый материал в лизате получали центрифугированием гомогената при 7000 g в течение 15 минут при 4oC, ресуспендированием полученных шариков в буфере A, равном двойному объему первоначально упакованных клеток, и повторным получением шариков центрифугированием материала при 700 g в течение 15 минут. Процесс промывки повторяли три раза.
Нерастворимый материал из лизата был экстрагирован при pH 12,0 следующим образом. Осадок суспендировали в буфере, содержащем 0,5 М хлористого натрия, 1 мМ этилендиаминтетрауксусной кислоты, в котором объем суспензии был равен 1,8-кратному объему первоначально упакованных клеток. В суспензии поддерживали pH добавлением 0,2 объема 0,4 М натрийфосфатного буфера, pH 12,0. После перемешивания суспензию центрифугировали при 7000 g в течение 15 минут при 4oC и надосадочный слой удаляли. Экстрагирование повторяли два раза. Экстрагированные шарики промывали суспендированием их в 0,5 М хлористого натрия, 1 мМ этилендиаминтетрауксусной кислоты, используя объем суспензии, равный двум объемам первоначально консервированных клеток, с последующим центрифугированием при 7000 g в течение 15 минут при 4oC.
Полипептид C100-3 в экстрагированном осадке переводили в растворимое состояние обработкой додецилсульфатом натрия. Шарики суспендировали в объеме буфера A, равном 0,9 объема первоначально консервированных клеток, и 0,1 объема 2% додецилсульфата натрия добавляли в суспензию. После перемешивания суспензии ее центрифугировали при 7000 g 15 минут при 4oC. Полученный осадок три раза экстрагировали додецилсульфатом натрия. Полученные надосадочные слои, которые содержали C100-3, объединяли в общий фонд.
Эта процедура очищает C100-3 более чем в 10 раз из нерастворимой фракции дрожжевого гомогената и выделение полипептида составляло более 50%.
Очищенный препарат полипептида слияния подвергали анализу методом электрофореза на полиакриламидном геле согласно Laemmli (1970). Основываясь на данных анализа, полипептид имел чистоту более 80% и имел средний молекулярный вес примерно 54000 дальтон.
IV. C. Идентификация РНК инфицированных индивидуумов, которая образует гибриды с кольцевой ДНК вируса гепатита C
IV. C.1. Идентификация РНК в печени шимпанзе с гепатитом ни A, ни B, который гибридизует с кольцевой ДНК вируса гепатита C
РНК из печени шимпанзе, больного гепатитом ни A, ни B, как было установлено, содержит виды РНК, образующие гибриды с кольцевой ДНК, содержащейся в клоне 81, при "северном" пятнении следующим образом.
РНК выделяли из печеночной биопсии шимпанзе, от которой был получен высокий титр плазмы (см. раздел IV.A.1), используя метод, описанный Maniatis и соавт. (1982), для выделения общей РНК из клеток млекопитающих и для ее разделения на поли-A+- и поли-A--фракции. Эти фракции РНК подвергнуты электрофорезу на формальдегид-агарозном геле (1% от веса по объему) и перенесены на нитроцеллюлозу. (Maniatis и соавт., 1982). Нитроцеллюлозные фильтры гибридизировали с кольцевой ДНК вируса гепатита C из клона 81, снабженной радиоактивной меткой. (См. на фиг. 4 нуклеотидную последовательность вставки). Чтобы получить радиоактивную метку, вставку кольцевой ДНК вируса гепатита C выделяли из клона 81 и метили радиоактивным фосфором-32 точной трансляцией, используя ДНК-полимеразу 1 (Maniatis и соавт., 1982 г.). Гибридизацию проводили 18 часов при 42oC в растворе, содержащем 10% (от веса по объему) сульфата декстрина, 50% (вес, объем) деионизированного формамида, 750 мМ хлористого натрия, 75 мМ цитрата натрия, 20 мМ кислого фосфорнокислого натрия, pH 6,5, 0,1% додецилсульфата натрия, 0,02% (вес/объем) альбумина бычьей сыворотки, 0,02% (вес/объем) Ficoll-400, 0,02% (вес/объем) поливинилпирролидона, 100 микрограмм/мл ДНК спермы лосося, которую гидродинамически фрагментировали ультразвуком и денатурировали, и пробу точно меченной транслированной кольцевой ДНК активностью 106 регистраций импульсов на миллиметр.
Авторадиография исследованных фильтров показана на фиг. 38. Полоса 1 содержит меченные фосфором-3 ограничения фрагмента. Полосы 2-4 содержат РНК печени шимпанзе в следующем объеме: полоса 2 содержит 30 микрограммов общей РНК; полоса 3 содержит 30 микрограммов поли-A--РНК и полоса 4 содержит 20 микрограммов поли-A+-РНК. Как показано на фиг. 38, печень шимпанзе, больной гепатитом ни A, ни B, содержит гетерогенную популяцию родственных молекул поли-A+-РНК, которая гибридизирует пробу кольцевой ДНК вируса гепатита C и которая, по-видимому, насчитывает примерно от 5000 до 11000 нуклеотидов по размеру. Эта РНК, которая образует гибрид с кольцевой ДНК вируса гепатита C, может представлять собой вирусные геномы и/или специфические транскрипты вирусного генома.
Эксперимент, описанный в разделе IV.C.2 ниже, согласуется с предположением, что вирус гепатита C содержит геном РНК.
IV.C.2. Идентификация РНК, производной от вируса гепатита C, в сыворотке инфицированнных индивидов
Нуклеиновые кислоты экстрагировали из частиц, выделенных из высокого титра плазмы шимпанзе, больной гепатитом ни A, ни B, как описано в разделе IV. A. 1. Аликвоты (эквивалентные 1 мл исходной плазмы) выделенных нуклеиновых кислот суспендировали в 20 микролитах 50 мМ Hepes, pH 7,5, 1 мМ этилендиаминтетрауксусной кислоты и 16 микрограмм/мл дрожжевой растворимой РНК. Пробы денатурировали кипячением в течение пяти минут с последующим быстрым замораживанием и обрабатывали рибонуклеазой A (5 микролитров, содержащих 0,1 мг/мл рибонуклеазы A в 25 мМ этилендиаминтетрауксусной кислоты, 40 мМ Hepes pH 7,5), или дезоксирибонуклеазой 1 (5 микролитров, содержащих одну единицу дезоксирибонуклеазы/ в 10 мМ хлористого магния, 25 мМ Hepes, pH 7,5); контрольные образцы инкубировали без фермента. После инкубации добавляли 230 микролитров охлажденного льдом 2 x SSC содержащего 2 микрограмма/мл растворимой дрожжевой РНК, и образцы фильтровали на нитроцеллюлозном фильтре. Фильтры гибридизировали с пробой кольцевой ДНК, полученной из клона 81, которая была помечена фосфором-32, трансляцией метки. На фиг. 39 представлена ауторадиография фильтра. Сигналы гибридизации определялись в обработанных дезоксирибонуклеазой образцах и в контрольных образцах (полосы 2 и 1 соответственно), но не определялись в обработанном рибонуклеазой образце (полоса 3). Таким образом, поскольку обработка рибонуклеазой 1 разрушает нуклеиновые кислоты, выделенные из частиц, а обработка дезоксирибонуклеазой 1 не вызывает такого действия, то это свидетельствует настоятельно о том, что геном вируса гепатита C состоит из РНК.
IV. C. 3. Определение увеличенных последовательностей нуклеиновых кислот вируса гепатита C, полученных из последовательностей нуклеиновых кислот вируса гепатита C в печени и образцах плазмы шимпанзе, больной гепатитом ни A, ни B
Нуклеиновые кислоты вируса гепатита C, присутствующие в печени и плазме шимпанзе с гепатитом ни A, ни B и у контрольных шимпанзе, были увеличены с использованием в основном реакций полимеразной цепи по методу, описанному Saiki и соавт. (1986). Затравочные олигонуклеотиды были получены из последовательностей кольцевой ДНК вируса гепатита C в клоне 81 или в клонах 36 и 37. Увеличенные последовательности были определены электрофорезом через гель и "южным" пятнением, используя в качестве проб подходящий олигомер кольцевой ДНК с последовательностью от участка между двумя затравками, но не включая его.
Образцы последовательностей РНК содержащего вируса гепатита C, подлежащие испытаниям системой увеличения, были выделены из биопсий печени трех шимпанзе, больных гепатитом ни A, ни B, и от двух контрольных шимпанзе. Выделение фракции РНК проводили с помощью тиоцианата гуанидиния, методом, описанным в разделе IV.C.1.
Образцы РНК, которые подлежали исследованию системой увеличения, были также выделены из плазмы двух шимпанзе, больных гепатитом ни A, ни B, и одной контрольной шимпанзе, а также из пула плазмы от контрольных шимпанзе. Один инфицированный шимпанзе имел инфекционную дозу шимпанзе на 1 мл, равную или больше чем 106, а другой инфицированный шимпанзе имел инфекционную дозу на 1 мл, равную или больше чем 105.
Нуклеиновые кислоты экстрагировали из плазмы следующим образом. Либо 0,1 мл, либо 0,01 мл плазмы разбавляли до конечного объема 1,0 мл раствором TENB-протеиназа K-додецилсульфат натрия (0,05 М трис-HCl, pH 8,0, 0,001 М этилендиаминтетрауксусной кислоты, 0,1 М хлористого натрия, 1 мг/мл протеиназы K и 0,5% додецилсульфата натрия), содержащим 10 микрограмм на 1 мл полиадениловой кислоты, и инкубировали при 37oC в течение 60 минут. После этой ферментации протеиназы K полученные фракции плазмы были освобождены от белка экстракцией смесью TE [трис-HCl-этилендиаминтетрауксусной кислоты (10,0 мМ трис-HCl, pH 8,0, 1 мМ ЭДТA)], насыщенной фенолом. Фенольную фазу отделяли центрифугированием и повторно экстрагировали смесью трис-HCl ЭДТА-NaCl, содержащей 0,1% додецилсульфата натрия. Полученные водные фазы от каждой экстракции были объединены и дважды экстрагированы равными объемами смеси фенол-хлороформ-изоамиловый спирт в отношении 1:1 (99:2) и два раза равным объемом смеси хлороформ-изоамиловый спирт в отношении 99:1. После фазы разделения центрифугированием водную фазу концентрировали до конечной концентрации 0,2 М ацетата натрия и нуклеиновые кислоты осаждали добавлением двух объемов этанола. Выпавшие в осадок нуклеиновые кислоты были выделены ультрацентрифугированием в роторе SW 41 при 38 K в течение 60 минут при 4oC.
B дополнение к сказанному выше, экстрагировали высокий титр плазмы шимпанзе и накопленной контрольной плазмы альтернативно по методу Chomcyzski and Sacchi (1987) с 50 микрограммами поли A-носителя. В этом процессе использовали экстракцию кислым тиоцианатом гуанидиния. РНК выделяли центрифугированием при 10000 оборотов/мин в течение 10 минут при 4oC в микроцентрифуге Eppendorf.
В двух случаях до проведения синтеза кольцевой ДНК в полимеразной цепной реакции нуклеиновые кислоты, экстрагированные из плазмы протеиназа K-додецилсульфат натрия-фенольным методом, были далее очищены связыванием и элюированием из S и S Elutip-R-колонок. Процесс проводили по инструкциям поставщиков оборудования.
Кольцевую ДНК, использованную в качестве матрицы для цепной реакции полимеразы, получали из нуклеиновых кислот (либо из общих нуклеиновых кислот, или из РНК), приготовленных как описано выше. После преципитации этанолом осажденные аминокислоты сушили и ресуспендировали в обработанной DEPC дистиллированной воде. Вторичные структуры в нуклеиновых кислотах разрывали нагреванием при 65oC в течение 10 минут и пробы немедленно охлаждали льдом. Кольцевую ДНК синтезировали с использованием от 1 до 3 микрограммов общей РНК из печени шимпанзе, или из нуклеиновых кислот (или РНК), экстрагированных из 10-100 микролитров плазмы. В синтезе использовали обратную транскриптазу и его проводили в объеме 25 микролитров, используя инструкции, определенные изготовителем, BRL. Затравками для синтеза кольцевой ДНК служили также затравки, использованные в цепной реакции полимеразы, описанной ниже. Все реакционные смеси для синтеза кольцевой дНК содержали 23 единицы ингибитора рибонуклеазы RNASINm (Fisher/Promega). После синтеза кольцевой ДНК реакционную смесь разбавляли водой, кипятили 10 минут и быстро охлаждали льдом.
Полимеразные цепные реакции проводили в основном согласно предписаниям изготовителей препаратов (Cetus-Perkin-Elmer), за исключением добавления одного микрограмма рибонуклеазы A. Реакции проводили в конечном объеме 100 микролитров. Цепную полимеразную реакцию осуществляли за 35 циклов, используя режимы 37oC, 72oC и 94oC.
Инициаторы синтеза кольцевой ДНК и цепных полимеразных реакций получали из последовательностей кольцевой ДНК вируса гепатита C либо в клоне 81, клоне 36 или в клоне 37b. (Последовательности кольцевых ДНК вируса гепатита C в клонах 81, 36 и 37b представлены на фиг. 4, 5 и 10 соответственно). Последовательности двух 16-членных инициаторов, полученных из клона 81, были следующими:
5' CAA TCA TAG CTG ACA G 3' и
5' GAT AAC CTC TGC CTG A 3'.
Последовательность инициатора из клона 36 была:
5' GCA TGT CAT GAT GTA T 3'.
Последовательность инициатора из клона 37b была:
5' ACA ATA CGT GTG TCA C 3'.
В полимеразных цепных реакциях совмещаются пары инициаторов либо двух 16-членных последовательностей из клона 81, или 16-членная последовательность из клона 36 и 16-членная последовательность из клона 37b.
Продукты цепной реакции полимеразы анализировали разделением продуктов электрофорезом на щелочном геле с последующим "южным" пятнением и определением увеличенных последовательностей кольцевой ДНК вируса гепатита C пробой меченного фосфором-32 внутреннего олигонуклеотида, полученного из участка кольцевой ДНК вируса гепатита C, который не перекрывает последовательностей инициаторов. Смеси цепных реакций полимеразы экстрагировали смесью фенол-хлороформ, и нуклеиновые кислоты осаждали из водной фазы солью или этанолом. Выпавшие в осадок нуклеиновые кислоты собирали центрифугированием и растворяли в дистиллированной воде. Аликвоты образцов подвергали электрофорезу на гелях, содержащих 1,8% щелочной агарозы. Однонитчатые ДНК длинами в 60, 108 и 161 нуклеотид подвергали параллельному электрофорезу на гелях в качестве маркеров молекулярного веса. После электрофореза кольцевые ДНК в геле переносили на бумагу Biorad Zeta Probem. Условия предварительной гибридизации и гибридизации и промывки определялись спецификациями изготовителя (Biorad).
Пробы, использованные для гибридизации-определения увеличенных последовательностей кольцевой ДНК вируса гепатита C, были следующие. Если пара инициаторов цепной реакции полимеразы были получены из клона 81, то пробой служили 108 нуклеотидов с последовательностью, соответствующей последовательности, которая локализована на участке между последовательностями двух инициаторов. Если пара инициаторов цепной реакции полимеразы получена из клона 36 и 37b, то пробой служила nick-транслированная вставка кольцевой ДНК вируса гепатита C, полученная из клона 35. Инициаторы были получены из нуклеотидов 155-170 вставки клона 37b и 206-268 нуклеотидов вставки клона 36. 3'-конец вставки кольцевой ДНК вируса гепатита C в клоне 35 перекрывает нуклеотиды 1-186 вставки в клоне 36; и 5'-конец вставки клона 35 перекрывает нуклеотиды 207-269 вставки в клоне 37b. (Сравните фиг. 5, 8 и 10). Таким образом, вставка кольцевой ДНК в клоне 35 перекрывает часть участка между последовательностями инициатора, полученными из клона 36 и 37b, и является полезной как проба на увеличенные последовательности, которые включают эти инициаторы.
Анализ РНК из образцов печени был проведен согласно вышеописанной процедуре с использованием обоих наборов инициаторов и проб. РНК из печени трех шимпанзе с гепатитом ни A, ни В давали положительные результаты гибридизации для увеличенных последовательностей ожидаемого размера (161 и 586 нуклеотидов для клона 81 и клонов 36 и 37b соответственно), тогда как контрольные шимпанзе дали отрицательные результаты гибридизации. Те же результаты были достигнуты при повторении эксперимента три раза.
Анализ нуклеиновых кислот и РНК из плазмы был проведен также в соответствии с описанной выше процедурой, используя инициаторы и пробу из клона 81. Плазму получали от двух шимпанзе с гепатитом ни A, ни B, у контрольного шимпанзе и из пула плазмы контрольных шимпанзе. Оба образца плазмы шимпанзе, больных гепатитом ни A, ни B, содержали смесь нуклеиновые кислоты - РНК, которая давала положительные результаты анализа в реакции роста полимеразной цепи, тогда как обе контрольные плазмы давали отрицательные результаты. Эти результаты воспроизводились в повторных экспериментах.
IV. D. Радиоиммунный анализ для определения антител вируса гепатита C в сыворотке инцифированных индивидов
Твердофазный радиоммунологический анализ для определения антител к антигенам вируса гепатита C был усовершенствован, основываясь на исследованиях Tsu and Herzenberg (1980). Пластинки для микротитров (иммулон 2, Removawell полоски) покрывали очищенными полипептидами, содержащими эпитопы вируса гепатита C. Покрытые пластинки инкубировали либо с образцами сыворотки человека, подозрительными на содержание антител к эпитопам вируса гепатита C, или с соответствующими контрольными образцами. В процессе инкубации антитела, если таковые присутствовали, связывались с твердофазным антигеном. После удаления несвязанного материала и промывания микротитровых пластинок определяли комплексы антиген вируса ни A, ни B - антитело человека инкубированнием с меченным йодом-125 человеческим иммуноглобулином антисыворотки овцы. Несвязанное меченое антитело удаляли отсасыванием и пластинки промывали. Определяли радиоактивность в индивидуальных ячейках: количество связанных антител к антигену вируса гепатита C человека пропорционально радиоактивности в источниках.
IV.D.1. Очистка полипептида слияния пероксиддисмутаза-ни A, ни B5-1-1
Полипептид слияния пероксиддисмутаза-ни A, ни B5-1-1, полученный экспрессией в рекомбинантных бактериях, как описано в разделе IV.B.1, очищали от рекомбинантныой E.coli дифференциальной экстракцией экстрактов клеток мочевиной с последующей хроматографией на анионо- и катионообменных колонках следующим образом.
Оттаявшие клетки от одного литра культуры суспендировали в 10 мл 20% (вес/объем) сахарозы, содержащей 0,01 М трис-HCl, pH 8,0 и 0,4 мл добавляли 0,5 М этилендиаминтетрауксусной кислоты, pH 8,0. Через 5 минут при 0oC смесь центрифугировали при 4000 g в течение 10 минут. Полученный осадок суспендировали в 10 мл 25% (вес/объем) сахарозы, содержащей 0,05 М трис-HCl, pH 8,0, 1 мМ фтористого
фенилметилсульфонила и 1 микрограмм/мл пепстатина A, с последующим добавлением 0,5 мл лизоцима (10 мг/мл) и инкубировали при 0oC в течение 10 минут. После добавления 10 мл 1,0% (по объему) тритона-Х-100 в 0,05 М трис-HCl, pH 8,0, и 1,0 мМ этилендиаминтетрауксусной кислоты смесь инкубировали дополнительно 10 минут при 0oC с возможным встряхиванием. Полученный вязкий раствор гомогенизировали шестикратным пропусканием через гиподермальную иглу 20 размера и центрифугировали при 13000 g 25 минут. Выпавший в осадок материал суспендировали в 5 мл 0,01 трис-HCl, pH 8,0 и суспензию центрифугировали при 4000 g 10 минут. Осадок, содержащий белок слияния пероксиддисмутаза ни A, ни B5-1-1, растворяли в 5 мл 6 М мочевины в 0,02 М трис-HCl, pH 8,0, 1 мМ дитиотрейтола (буфер A) и помещали в колонку с быстро пропускающей Q-сефарозой, уравновешенной буфером A. Полипептиды элюировали линейным градиентом от 0,0 до 0,3 М хлористого натрия в буфере A. После элюирования фракции анализировали электрофорезом через полиакриламидный гель в присутствии додецилсульфата натрия для определения содержания пероксиддисмутаза-ни A, ни B5-1-1. Фракции, содержащие этот полипептид, собирали и подвергали диализу с 6 М мочевины в 0,02 М буфера фосфата натрия, pH 6,0, 1 мМ дитиотрейтола (буфер B). Диализованный образец помещали в колонку с S-сефарозой, уравновешенной буфером B, и полипептиды элюировали линейным градиентом от 0,0 до 0,3 М хлористого натрия в буфере B. Фракции анализировали электрофорезом через полиакриламидный гель на присутствие полипептида пероксиддисмутаза-ни A, ни B5-1-1, и соответствующие фракции собирали.
Окончательное получение пероксиддисмутаза-ни A, ни B5-1-1 испытывали электрофорезом на полиакриламидных гелях, в которых присутствовал додецилсульфат натрия. По результатам этого анализа препарат имел чистоту более 80%.
IV.D.2. Очистка полипептида слияния пероксиддисмутаза-ни A, ни B81
Полипептид слияния пероксиддисмутаза-ни A, ни B81, полученный экспрессией в рекомбинантных бактериях, как описано в разделе IV.B.2, был очищен от рекомбинантной E.coli дифференциальным экстрагированием клеточных экстрактов мочевиной с последующей хроматографией на анионо- и катионообменных колонках с использованием процесса, описанного в разделе IV.D.1, при выделении полипептида слияния пероксиддисмутаза-ни A, ни B5-1-1.
Конечное получение полипептида пероксиддисмутаза-ни A, ни B81 проверяли электрофорезом на полиакриламидных гелях в присутствии додецилсульфата натрия. На основании этого анализа препарат имел чистоту более 50%.
IV. D. 3. Определение антител к эпитопам вируса гепатита C твердофазным радиоиммунным методом
Образцы сыворотки 32 пациентов, у которых диагностировали гепатит ни A, ни B, подвергали радиоиммунному анализу на определение того, присутствуют ли антитела к эпитопам вируса C в полипептидах слияния пероксиддисмутаза-ни A, ни B5-1-1 или в пероксиддисмутаза-ни A, ни B81 и их регистрацию.
Микротитровые пластинки покрывали полипептидом пероксиддисмутаза-ни A, ни B5-1-1 или пероксиддисмутаза-ни A, ни B81, которые были частично очищены методами, описанными в разделах IV. D.1 и IV.D.2 соответственно. Aнализы проводили следующим образом.
100-микролитровые аликвотные пробы, содержащие 0,1-0,5 микрограммов пероксиддисмутаза-ни A, ни B5-1-1 или пероксиддисмутаза-ни A, ни B81 в 0,125 M боратно-натриевого буфера, pH 8,3, 0,075 М хлористого натрия (BBS) добавляли к каждой ячейке микротитровой пластинки (Dynatech Immulon 2 Removawell Strips). Пластинку инкубировали при 4oC, оставляя на ночь во влажной камере, после чего белковый раствор удаляли и ячейки промывали три раза BBS, содержащим 0,02% тритона X-100 (BBST). Чтобы предотвратить неспецифическое связывание, ячейки покрывали альбумином бычьей сыворотки добавлением 100 микролитров раствора альбумина бычьей сыворотки в BBS при концентрации 5 мг/мл с последующей инкубацией при комнатной температуре в течение часа; после этого инкубирования раствор альбумина бычьей сыворотки удаляли. Полипептиды в покрытых ячейках взаимодействовали с сывороткой при добавлении 100 микролитровых образцов плазмы, разбавленных в отношении 1:100 в 0,01 М фосфатного буфера, pH 7,2, 0,15 М хлористого натрия (PBS), содержащего 10 мг/мл альбумина бычьей сыворотки, и содержащие сыворотку ячейки инкубировали один час при 37oC. После инкубирования образцы сыворотки удаляли отсасыванием и ячейки пять раз промывали боратным буфером BBST. Антитела против ни A, ни B5-1-1 и ни A, ни B81 определяли связыванием меченных йодом-125 F'(ab)2 овцы против IgG человека с покрытыми ячейками. Аликвоты в 100 микролитров меченой пробы (удельная активность 5-20 микрокюри/микрограмм) добавляли к каждой ячейке и пластинки инкубировали при 37oC один час с последующим удалением избыточной пробы отсасыванием и пятикратным промыванием боратным буфером с тритоном Х-100 (BBST). Количество радиоактивности, связанной в каждой ячейке, определяли подсчетом числа распадов в счетчике, детектирующем гамма-излучение.
Результаты определения анти-ни A, ни B5-1-1 и анти-ни A, ни B81 у индивидов с гепатитом ни A, ни B представлены в табл. 1.
Как видно из таблицы, сыворотки больных 1, 19 и 32, у которых диагностировался вирусный гепатит ни A, ни B, были положительны в отношении антител, направленных против эпитопов вируса C, присутствующих в пероксиддисмутаза-ни A, ни B5-1-1 и пероксиддисмутаза-ни A, ни B81.
Однако образцы сыворотки, которые были положительными, неодинаково взаимодействовали иммунологически с пероксиддисмутаза-ни A, ни B5-1-1 и пероксиддисмутаза-ни A, ни B81. Образцы сыворотки больного N 1 были положительными к полипептиду пероксиддисмутаза-ни A, ни B81, но не были положительными к пероксиддисмутаза-ни A, ни B5-1-1. Образцы сыворотки больных N 10, 15 и 17 были положительными к пероксиддисмутаза-ни A, ни B5-1-1, но не были положительными к пероксиддисмутаза-ни A, ни B81. Образцы сыворотки больных N 3, 8, 11 и 12 реагировали одинаково с обоими полипептидами слияния, тогда как образцы сыворотки больных N 2, 4, 7 и 9 были в 2-3 раза сильнее в реакции с пероксиддисмутаза-ни A, ни B5-1-1, чем с пероксиддисмутаза-ни A, ни B81. Эти результаты наводят на мысль о том, что ни A, ни B5-1-1 и ни A, ни B81 могут содержать по меньшей мере три различных эпитопа, т.е. возможно, что каждый полипептид содержит по меньшей мере один уникальный эпитоп и что два полипептида делят между собой по меньшей мере один эпитоп.
IV. D. 4. Специфичность твердофазного радиоиммунологического анализа для ни A, ни B
Специфичность твердофазного радиоиммунного анализа для вируса ни A, ни B испытывали с использованием исследований сыворотки больных, инфицированных вирусом гепатита A или вирусом гепатита B, и на сыворотке контрольных индивидов. Анализы, в которых использовали частично очищенные полипептиды пероксиддисмутаза-ни A, ни B5-1-1 и пероксиддисмутаза-ни A, ни B81, проводили в основном так, как описано в разделе IV.D.3, за исключением того, что сыворотки были получены от больных, у которых ранее определяли гепатит вируса A или вирусный гепатит B, или у индивидов, являющихся донорами банка крови. Результаты, полученные на сыворотках инфицированных вирусами гепатита A и B, представлены в табл. 1. Радиоиммунный анализ проверяли на 11 видах сыворотки больных, инфицированных вирусом гепатита A, и на 20 видах сыворотки больных, инфицированных вирусом гепатита B. Как показано в табл. 1, ни одна из этих сывороток не давала положительной иммунологической реакции с полипептидами слияния, содержащими эпитопы вируса BB-ни A, ни B.
Радиоиммунологический анализ с использованием антигена ни A, ни B5-1-1 был применен для определения иммунологической реактогенности сыворотки контрольных индивидов. Из 230 образцов сыворотки, полученных от нормальной популяции донорской крови, только две показали положительные реакции радиоиммунного анализа (данные не показаны). Возможно, что два донора крови, кому принадлежали эти два образца сыворотки, ранее подвергались инфекции вируса гепатита C.
IV. D.5. Реактогенность ни A, ни B5-1-1 в процессе инфекции вируса гепатита ни A, ни B
Присутствие антител против ни A, ни B5-1-1 при течении инфекции вируса гепатита ни A, ни B у двух больных людей и 4 шимпанзе определяли с использованием радиоиммунного анализа, как описано в разделе IV.D.3. В дополнение радиоиммунный анализ был использован для определения присутствия или отсутствия антител против ни A, ни B5-1-1 при течении инфекций вируса гепатита A и вируса гепатита B у инфицированных шимпанзе.
Результаты, представленные в табл. 2, показывают, что антитела против ни A, ни B5-1-1 обнаруживались после начала острой фазы инфекции гепатита ни A, ни B. Антитела против ни A, ни B5-1-1 не определялись в образце сыворотки от шимпанзе, инфицированных либо вирусом гепатита A, или вирусом гепатита B. Таким образом, антитела против пептида ни A, ни B5-1-1 служат маркером для индивидов, подвергшихся инфекции вируса гепатита C.
IV.Е. Очистка поликлональных антител сыворотки к ни A, ни B5-1-1
На основе специфической иммунологической реактогенности полипептида пероксиддисмутаза-ни A, ни B5-1-1 в отношении антител образцов сыворотки больных гепатитом ни A, ни B был разработан метод очистки антител сыворотки, которые реагируют иммунологически с эпитопом(ами) в ни A, ни B5-1-1. Этот метод использует аффинную хроматографию. Очищенный полипептид пероксиддисмутаза-ни A, ни B5-1-1 (см. раздел IV.D.1) прикрепляли на нерастворимой подложке; присоединение осуществляли так, что фиксированный полипептид сохранял свое сродство с антителом к ни A, ни B5-1-1. Антитело в образцах сыворотки абсорбировано к матично-связанному полипептиду. После удаления неспецифически связанных материалов и несвязанных материалов связанное антитело освобождали от связанного полипептида пероксиддисмутаза - вирус гепатита C изменением pH и/или применением хаотропного реагента, например мочевины.
Нитроцеллюлозные мембраны, содержащие связанный полипептид пероксиддисмутаза-ни A, ни B5-1-1, готовили следующим образом. 2,1 см Sartorius с размером пор 0,2 микрона промывали по три минуты три раза буфером BBS (боратный буфер). Полипептид пероксиддисмутаза-ни A, ни B5-1-1 связывали с мембраной инкубированием очищенного препарата в буфере BBS при комнатной температуре в течение двух часов; альтернативно его инкубировали при 4oC, оставляя на ночь. Раствор, содержащий несвязанный антиген, удаляли, а фильтр три раза промывали боратным буфером (BBS) по три минуты каждая промывка. Оставшиеся активные локализации на мембране были блокированы альбумином бычьей сыворотки инкубированием с 5 мг/мл раствором альбумина бычьей сыворотки в течение 30 минут. Избыток альбумина бычьей сыворотки удаляли промыванием мембран пять раз боратным буфером (BBS) и три раза дистиллированной водой. Мембрану, содержащую вирусный антиген и альбумин бычьей сыворотки, обрабатывали 0,05 М гидрохлорида глицина, pH 2,5, 0,10 М хлористого натрия (Cly-HCI) в течение 15 минут с последующим трехкратным промыванием фосфатно-солевым буферным раствором, продолжительностью три минуты.
Поликлональные антитела против ни A, ни B5-1-1 выделяли инкубированием мембран, содержащих полипептид связывания с сывороткой индивидов, больных гепатитом ни A, ни B, в течение двух часов. После инкубации фильтры пятикратно промывали боратным буферным раствором (BBS) и два раза дистиллированной водой. Связанные антитела элюировали затем из каждого фильтра пятью вымываниями гидрохлоридом по три минуты каждое элюирование. Значение pH элюатов поддерживали равным 8,0 сбором каждого элюата в экспериментальную пробирку, содержащую 2,0 М трис-HCl, pH 8,0. Выход антител против ни A, ни B5-1-1 после аффинной хроматографии составил приблизительно 50%.
Эти нитроцеллюлозные мембраны, содержащие связанный вирусный антиген, могут быть использованы несколько раз без видимого уменьшения связывающей способности. Чтобы повторно использовать мембраны после элюирования антител, мембраны промывали боратным буферным раствором три раза по три минуты каждый. Затем их хранили в боратном буферном растворе при 4oC.
IV. F. Захват частиц вируса гепатита C из инфицированной плазмы, используя очищенные поликлональные антитела человека против вируса гепатита C; гибридизация нуклеиновой кислоты в захваченных частицах с кольцевой ДНК вируса гепатита C
IV. F. 1. Захват частиц вируса гепатита C из инфицированной плазмы, используя поликлональные антитела человека против вируса гепатита C
Комплексы белок - нуклеиновая кислота, присутствующие в инфекционной плазме шимпанзе с гепатитом ни A, ни B, были выделены с использованием очищенных поликлональных антител человека против вируса гепатита C, которые были связаны со слоями полистирола.
Поликлональные антитела против ни A, ни B5-1-1 очищали из сыворотки больного человека гепатитом ни A, ни B, используя полипептид пероксиддисмутаза - вирус гепатита C, закодированный в клоне 5-1-1. Метод очистки соответствовал таковому, описанному в разделе IV.Е.
Очищенные антитела против ни A, ни B5-1-1 были связаны с полистироловыми слоями (около 6,5 мм диаметром, зеркальная обработка. Precision Plastic Ball Co. , Chicago, Illinois) инкубированием каждого при комнатной температуре, оставляя их на ночь с 1 мл антител (1 микрограмм/мл боратного буфера с физиологическим раствором, pH 8,5). После ночной инкубации подложки один раз промывали с помощью TBST [50 мМ трис-HCl, pH 8,0, 0,150 мМ хлористого натрия, 0,05% (по объему) твина 20] и затем физиологическим раствором, стабилизированным фосфатным буфером (PBS), содержащим 10 мг/мл альбумина бычьей сыворотки.
Контрольные подложки готовили идентичным образом, за исключением того, что очищенные антитела против ни A, ни B5-1-1 были заменены общим иммуноглобулином человека.
Улавливание вируса гепатита C из инфицированной плазмы шимпанзе с гепатитом ни A, ни В связанными с подложкой антителами против ни A, ни B5-1-1 осуществляли следующим образом. Плазму шимпанзе с гепатитом ни A, ни B использовали как описано в разделе IV.A.1. Аликвоту (1 мл) инфицированной вирусом гепатита ни A, ни B плазмы шимпанзе инкубировали три часа при 37oC с каждой из пяти подложек, покрытых либо антителами против ни A, ни B5-1-1, либо контрольными иммуноглобулинами. Подложки промывали три раза трис-буферным раствором (TBST).
IV. F.2. Гибридизация нуклеиновой кислоты в захваченных частицах с кольцевой ДНК вируса ни A, ни B
Компонент нуклеиновой кислоты, освобожденный из частиц, захваченных антителами против ни A, ни B5-1-1, анализировали гибридизацией с кольцевой ДНК вируса гепатита C, полученной из клона 81.
Частицы вируса гепатита C захватывали из плазмы шимпанзе, инфицированной гепатитом ни A, ни B, как описано в разделе IV.F.1. Чтобы освободить нуклеиновые кислоты из частиц, промытые подложки инкубировали 60 минут при 37oC с 0,2 мл на подложку раствора, содержащего протеиназу k (1,0 мг/мл), 10 мМ трис-HCl, pH 7,5, 10 мМ этилендиаминтетрауксусной кислоты, 0,25% (от веса по объему) додецилсульфата натрия, 10 микрограммов/мл растворимой дрожжевой РНК и надосадочный слой удаляли. Надосадочный раствор экстрагировали фенолом и хлороформом и нуклеиновые кислоты осаждали этанолом, оставляя раствор на ночь при -20oC. Преципитат нуклеиновой кислоты собирали центрифугированием, сушили и растворяли в 50 мМ гепеса, pH 7,5. Удвоенные аликвоты растворимых нуклеиновых кислот из образцов, полученных на подложках, покрытых антителами против ни A, ни B5-1-1 и с контрольными подложками, содержащими общий иммуноглобулин человека, фильтровали на нитроцеллюлозных фильтрах. Фильтры гибридизировали с меченной фосфором-32, nick-транслированной пробой, полученной из очищенного фрагмента кольцевой ДНК вируса гепатита C из клона 81. Методы получения пробы и проведения гибридизации описаны в разделе IV.C.1.
Ауторадиография исследуемых фильтров, содержащих нуклеиновые кислоты из частиц, уловленных слоями, содержащими антитела против ни A, ни B5-1-1, показаны на фиг. 40. Экстракт, полученный с использованием антитела против ни A, ни B5-1-1 (A1, A2), дает ясные сигналы гибридизации относительно контрольного экстракта (A3, A4) и контрольной дрожжевой РНК (B1, B2). Стандарты, содержащие 1 пг, 5 пг и 10 пг (пикограмм) очищенного фрагмента кольцевой ДНК клона 81 показаны в Cl-3 соответственно.
Эти результаты показывают, что частицы, захваченные из плазмы, инфицированной гепатитом ни A, ни B, антителами против ни A, ни B5-1-1, содержат нуклеиновые кислоты, которые гибридизируют с кольцевой ДНК вируса гепатита C в клоне 81 и представляют дальнейшее свидетельство того, что кольцевые ДНК в этих клонах получены из этиологического агента для гепатита ни A, ни B.
IV. G. Имунологическая реактогенность C100-3 с очищенными антителами против ни A, ни B5-1-1
Иммунологическая реактогенность C100-3 полипептида слияния с антителами против ни A, ни B5-1-1 определяли радиоиммунным анализом, в котором антигены, которые были связаны с твердой фазой, провоцировали очищенными антителами против ни A, ни B5-1-1 и определяли комплекс антиген - антитело меченными йодом-125 антителами овцы против антигена человека. Иммунологическую реактивность полипептида C100-3 сравнивали с реактивностью антигена пероксиддисмутаза-ни A, ни B5-1-1.
Полипептид слияния C100-3 синтезировали и очищали, как описано в разделе IV. B. 5 и в разделе IV.B.6 соответственно. Полипептид слияния пероксиддисмутаза-ни A, ни B5-1-1 синтезировали и очищали, как описано в разделе IV.B.1 и в разделе IV.D.1 соответственно. Очищенные антитела против ни A, ни B5-1-1 получали, как описано в разделе IV.Е.
Стомикролитровые аликвоты, содержащие различные количества очищенного антигена C100-3 в 0,125 М буфера бората натрия, pH 8,3, 0,075 М хлористого натрия (BBS), добавляли к каждой ячейке микротитровой пластины (Dynatech Immulon 2 Removawell Strips). Пластину инкубировали при 4oC, оставляя на ночь во влажной камере, после чего белковый раствор удаляли и ячейки промывали три раза боратным буферным раствором, содержащим 0,02% Тритона Х-100 (BBST). Чтобы предотвратить неспецифическое связывание, ячейки покрывали альбумином бычьей сыворотки добавлением 100 микролитров 5 мг/мл раствора альбумина бычьей сыворотки в боратном буферном растворе с последующим инкубированием при комнатной температуре в течение одного часа, после чего избыток раствора альбумина бычьей сыворотки удаляли. Полипептиды в покрытых ячейках подвергали взаимодействию с очищенными антителами против ни A, ни B5-1-1 добавлением одного микрограмма антител на ячейку и инкубируя пробы образцов один час при 37oC. После инкубации избыток раствора удаляли отсасыванием и ячейки промывали пятикратно BBST. Антитела против ни A, ни B5-1-1, связанные с полипептидами слияния, определяли связыванием меченных йодом-125 F'(ab)2 овцы против IgG человека к покрытым альбумином бычьей сыворотки ячейкам. Аликвоты по сто микролитров меченой пробы (удельная активность 5-20 микрокюри/микрограмм) добавляли к каждой ячейке и пластинки инкубировали при 37oC один час с последующим удалением избыточной пробы отсасыванием и пятикратной промывкой буферным раствором BBST. Количество радиоактивности в каждой ячейке определяли подсчетом импульсов в счетчике, который детектирует гамма-излучение.
Результаты иммунологической реактогенности C100 с очищенными антителами против ни A, ни B5-1-1 с очищенными антителами представлены в табл. 3.
Результаты, представленные в табл. 3, показывают, что антитела против ни A, ни B5-1-1 распознают эпитоп(ы) в части C100 полипептида C100-3. Таким образом, ни A, ни B5-1-1 и C100 делят один общий эпитоп(ы). Результат наводит на мысль о том, что последовательность кольцевой ДНК, кодирующая этот ни A, ни B-эпитоп(ы), является последовательностью, которая присутствует как в клоне 5-1-1, так и в клоне 81.
IV.Н. Характеристика вируса гепатита C
IV.H.1. Характеристика нитчатости генома вируса гепатита C
Геном вируса гепатита C характеризовали в отношении его нитчатости изолированием фракции нуклеиновой кислоты из частиц, захваченных антителом против ни A, ни B5-1-1 на покрытых полистироловых слоях, и определяли, гибридизирует ли выделенная нуклеиновая кислота с плюс- и/или минус-нитями кольцевой ДНК вируса гепатита C.
Частицы были захвачены из плазмы шимпанзе, инфицированной вирусом гепатита C, с использованием полистироловых слоев, покрытых иммуноочищенными антителами против ни A, ни B5-1-1, как описано в разделе IV.F.1. Компонент нуклеиновой кислоты частиц освобождали, используя метод, описанный в разделе IV.F.2. Аликвоты выделенной геномной нуклеиновой кислоты, эквивалентные 3 мл плазмы высокого титра, наносили пятнами на нитроцеллюлозные фильтры. Контролем служили аликвоты денатурированной кольцевой ДНК вируса гепатита C из клона 81 (два пикограмма), которые также наносили пятнами на те же фильтры. Фильтры испытывали меченной фосфором-32 смесью плюс- или смесью минус-нити однонитчатой ДНК, клонированной из кольцевых ДНК вируса гепатита C; кольцевые ДНК вырезали из клонов 40b, 81 и 25c.
Однонитчатые пробы получали вырезанием кольцевых ДНК вируса гепатита C из клонов 81, 40b и 25c с помощью EcoRI и клонированием фрагментов кольцевой ДНК в М13 векторы, mp18 и mp19 (Messing, 1983 г.). Клоны М13 подвергали определению последовательностей на содержание плюс- или минус-нитей ДНК, производных от кольцевых ДНК вируса гепатита C. Определение последовательностей проводили методом окончания дидезоксицепи по Sanger и соавт. (1977).
Каждый комплект двойных фильтров, содержащих аликвоты генома вируса гепатита C, выделенного из уловленных частиц, гибридизировали либо с плюс-, либо с минус-нитчатыми пробами из кольцевых ДНК вируса гепатита C. Ауторадиография, полученная от испытываемого генома вируса гепатита C со смесью проб, полученных из клонов 81, 40b и 25c, показана на фиг. 41. Эта смесь были использована для увеличения чувствительности гибридизационного анализа. Образцы в перечне I гибридизировали смесью пробы плюс-нити. Образцы в перечне II испытывали гибридизацией со смесью пробы минус-нити. Состав образцов в перечнях иммунных пятен представлен в табл. 4.
Как видно из результатов на фиг. 41, только пробы минус-нити ДНК гибридизируют с выделенным геномом вируса гепатита C. Этот результат в сочетании с результатом, показывающим, что геном чувствителен к рибонуклеазе и нечувствителен к дезоксирибонуклеазе (см. раздел IV.C.2) наводит на мысль о том, что геном вируса ни A, ни В является положительной нитчатой РНК.
Эти данные и данные, полученные в других лабораториях относительно физико-химических свойств предполагаемого вируса ни A, ни B, согласуются с возможностью того, что вирус гепатита C представляет новый класс вирусного агента, и не исключают такую возможность.
IV. H. 2. Определение последовательности в уловленных частицах, которые, будучи увеличены посредством цепной реакции полимеразы, гибридизировали с кольцевой ДНК вируса гепатита A, полученного из клона 81
РНК в уловленных частицах получали как описано в разделе IV.Н.1. Анализы на определение последовательностей, которые гибридизируют с кольцевой ДНК вируса гепатита C, полученной из клона 81, осуществляли, используя процесс увеличения, лежащий в основе цепной реакции полимеразы, описанный в разделе IV. C. 3, за исключением того, что гибридизационной пробой служил киназированный олигонуклеотид, полученный из последовательности кольцевой ДНК слона 81. Результаты показывают, что увеличенные последовательности гибридизируют с пробой кольцевой ДНК вируса гепатита C, полученной из клона 81.
IV. Н. 3. Гомология между неструктурным белком Dengue-флавивируса (MNWWVD1) и полипептидами вируса гепатита C, закодированными объединенными открыто читаемыми структурами в клонах 14i до 39c
Объединенные кольцевые ДНК вируса гепатита C в клонах 14i до 39c содержат одну непрерывную открыто читаемую структуру, как показано на фиг. 26. Закодированный в ней полипептид был проанализирован на гомологию последовательности с участком неструктурного полипептида(ов) у Dengue-флавивируса (MNWWVD1). В анализе использовали базу данных о белках Dayhoff и обработали на компьютере. Результаты представлены на фиг. 42, на которой символ (:) означает точную гомологию и символ (.) указывает на консервативное замещение в последовательности; пунктирные линии показывают пространства, введенные в последовательность для достижения наибольшей гомологии. Как видно на фигуре, имеется значительная гомология между последовательностью, закодированной в кольцевой ДНК вируса гепатита C, и неструктурным полипептидом(ами) Денгье-вируса. В дополнение к гомологии, показанной на фиг. 42, анализ полипептидного сегмента, закодированного на участке в направлении 3'-конца кольцевой ДНК, также содержит последовательности, гомологичные последовательностям в Денгье-полимеразе. Следствием этой находки является то, что каноническая Cly-Asp-Asp (GDD) последовательность, хотя и существенная для РНК зависимых РНК полимераз, содержится в полипептиде, закодированном в кольцевой ДНК вируса гепатита C в локализации, которая согласуется с локализацией в Денгье-2 вирусе (данные не приводятся).
IV.H.4. ДНК вируса гепатита C не определяется в инфицированных гепатитом ни A, ни B тканях
Два вида исследований представляют данные, свидетельствующие о том, что ДНК вируса гепатита C не определяется в тканях индивидов с гепатитом ни A, ни B. Эти результаты в сочетании с результатами, описанными в разделах IV.C, IV. H. 1 и IV.H.2, служат подтверждением того, что вирус гепатита C не является ДНК-содержащим вирусом и что его репликация не вовлекает кольцевую ДНК.
IV.Н.4.a. Процесс "южного" пятнения
Чтобы определить, содержит ли печень инфицированной гепатитом ни A, ни B шимпанзе определяемую ДНК вируса гепатита C (или кольцевую ДНК вируса гепатита C), фрагменты ограничивающего фермента ДНК были выделены из этого источника и подвергнуты "южному" пятнению и пятна исследованы меченной фосфором-32 кольцевой ДНК вируса гепатита C. Результаты свидетельствуют о том, что меченая кольцевая ДНК вируса гепатита C не гибридизирует с исследуемой ДНК в пятнах из печени инфицированного шимпанзе. Она также не гибридизирует с контрольной ДНК в пятнах из печени нормального шимпанзе. И напротив, в положительном контроле меченая проба гена бета-интерферона сильно гибридизировала южные пятна ограничивающего фермента сброженной плацентарной ДНК человека. Эти системы были созданы для определения отдельной копии гена, который подлежал определению меченой пробой.
ДНК были выделены из печени двух шимпанзе с гепатитом ни A, ни B. Контрольные ДНК были выделены из печени неинфицированной шимпанзе и из плаценты человека. Процесс экстрагирования ДНК был осуществлен, в основном, по методу Maniatis и соавт. (1982 г.), и пробы ДНК были обработаны рибонуклеазой в процессе выделения.
Каждый образец ДНК был обработан либо EcoRI, MboI или HincII (12 микрограммов) согласно предписаниям изготовителя. Сброженные ДНК подвергали электрофорезу на 1% нейтральном агарозном геле, наносили "южными" пятнами на нитроцеллюлозу и пятнистый материал гибридизировали с подходяще nick-транслированной пробой кольцевой ДНК (3·106 регистраций распадов в минуту на миллилитр гибридизационной смеси). ДНК печени инфицированных шимпанзе и из нормальной печени гибридизировали с меченной фосфором-32 кольцевой ДНК вируса гепатита C из клонов 36 плюс 81; ДНК из плаценты человека гибридизировали с меченной фосфором-32 ДНК из гена бета-интерферона. После гибридизации пятна промывали в строгих условиях, т.е. раствором, содержащим 0,1 x SSC, 0,1% додецилсульфата натрия при 65oC.
ДНК гена бета-интерферона была получена по методу Honghton и соавторов (1981).
IV.Н.4.b. Увеличение методом цепной реакции полимеразы
Чтобы выяснить, будет ли система ДНК-вируса гепатита C определяться в печени шимпанзе, больных гепатитом ни A, ни B, ДНК выделяли из ткани и подвергали цепной реакции полимеразного увеличения, используя инициаторы и пробы полинуклеотидов, производных из кольцевой ДНК вируса гепатита C в клоне 81. Отрицательными контролями были образцы ДНК, выделенные из неинфицированных клеток культуры ткани HepG2 и из вероятно неинфицированной плаценты человека. Положительными контролями были образцы отрицательного контроля ДНК, к которым добавляли известное относительно небольшое количество (250 молекул) кольцевой ДНК вируса гепатита C в виде вставки из клона 81.
В дополнение, чтобы подтвердить, что фракции РНК, выделенные из тех же печеней шимпанзе с гепатитом ни A, ни B, содержали последовательности, комплементарные пробе ДНК-вируса гепатита C, использовали также систему увеличения-детектирования реакции роста цепи полимеразы на выделенных образцах РНК.
В этих исследованиях ДНК были выделены способом, описанным в разделе IV. Н. 4. a, и РНК экстрагировали, в основном, как описано Chirgwin и соавт. (1981).
Образцы ДНК были выделены из двух печеней инфицированных шимпанзе, из неинфицированных клеток HepG2 и из плаценты человека. Один микрограмм каждой ДНК сбраживали с HindIII согласно инструкции изготовителей. Сброженные образцы были подвергнуты увеличению по реакции роста полимеразной цепи и определению увеличенной кольцевой ДНК вируса гепатита C, в основном, как описано в разделе IV.C.3, за исключением того, что исключали операцию с обратной транскриптазой.
Инициаторы реакции роста полимеразной цепи и проба были получены из кольцевой ДНК вируса гепатита C клона 81 и описаны в разделе IV.C.3. До увеличения для положительного контроля один микрограмм образца каждой ДНК был "заострен" добавлением 250 молекул вставки кольцевой ДНК вируса гепатита C, выделенной из клона 81.
Для определения присутствия последовательности вируса гепатита C в РНК, выделенных из печени шимпанзе с гепатитом ни A, ни B, образцы, содержащие 0,4 микрограмма общей РНК, подвергали процессу увеличения, в основном, как описано в разделе IV. C.3, за исключением того, что пренебрегали обратной транскриптазой в некоторых образцах, как отрицательный контроль. Инициаторы реакции роста полимеразной цепи и проба были получены из кольцевой ДНК вируса гепатита C клона 81, как описано выше.
Результаты показывают, что увеличенные последовательности, комплементарные пробе кольцевой ДНК вируса гепатита C, не определялись в ДНК из печени инфицированных шимпанзе, не определялись они и в отрицательных контролях. И, напротив, если образцы, включающие ДНК из печени инфицированных шимпанзе, были "заострены" кольцевой ДНК вируса гепатита C до увеличения, последовательности клона 81 определялись во всех положительных контрольных образцах. В дополнение в исследованиях РНК увеличенные последовательности кольцевой ДНК вируса гепатита C клона 81 определяли, только если использовали обратную транскриптазу, поэтому можно с уверенностью заключить, что результаты не обусловлены загрязнением ДНК.
Эти результаты показывают, что гепатоциты от шимпанзе с гепатитом ни A, ни B не содержат или содержат нерегистрируемые уровни ДНК вируса гепатита C. Основываясь на исследовании "заострения", если ДНК вируса гепатита C присутствует, то ее уровень гораздо ниже 0,06 копий на гепатоцит, и, напротив, последовательности вируса гепатита C в общей РНК от одних и тех же образцов легко определялись реакцией роста полимеразной цепи.
IV.I. Определение иммуноферментным твердофазным анализом инфекции вируса гепатита C, используя C100-3 как экспериментальный антиген
Все образцы анализировали, используя иммуноферментный твердофазный анализ (ELISA) антигена C100-3 вируса гепатита C (который синтезировали и очищали как описано в разделе IV.B.5), и конъюгат пероксидазы хрена и моноклонального антитела мыши против IgG человека.
Пластинки, покрытые антигеном C100-3 вируса гепатита C, были приготовлены следующим образом. Раствор, содержащий покровный буфер (50 мМ бората натрия, pH 9,0), 21 мл/пластинка, альбумин бычьей сыворотки (25 микрограмм/мл), C100-3 (2,50 микрограмм/мл), готовили непосредственно перед добавлением к пластинкам (в съемные ячейки Immulon 1 Dynatech Corp.). После перемешивания в течение пяти минут добавляли по 0,2 мл раствора на ячейку и их покрывали и инкубировали два часа при 37oC, после чего раствор удаляли отсасыванием. Ячейки промывали один раз 400 микролитрами промывочного буфера (100 мМ фосфата натрия, pH 7,4, 140 мМ хлористого натрия, 0,1% (от веса по объему) казеина, 1% (по весу от объема) тритона Х-100, 0,01% (вес/объем) тимерозала). После удаления промывочного раствора добавляли 200 микролитров/ячейку послепокровного раствора (10 мМ фосфата натрия, pH 7,2, 150 мМ хлористого натрия, 0,1% (вес/объем) казеина и 2 мМ фтористого фенилметилсульфонила), пластинки свободно накрывали для предотвращения испарения и выдерживали при комнатной температуре 30 минут. Затем ячейки отсасывали для удаления раствора и лиофилизировали сушкой, оставляя на ночь без нагревания оболочек. Полученные пластинки хранили при 2-8oC в запаянных алюминиевых пакетах.
Чтобы провести иммуноферментный твердофазный анализ, 20 микролитров образца сыворотки или контрольный образец добавляли к ячейке, содержащей 200 микролитров образца, разбавителя (100 мМ фосфата натрия, pH 7,4, 500 мМ хлористого натрия, 1 мМ этилендиаминтетрауксусной кислоты, 0,1% (вес/объем) казеина, 0,015 (вес/объем) терозала, 1% (вес/объем) тритона Х-100, 100 микрограмм/мл дрожжевого экстракта). Пластинки закупоривали и инкубировали при 37oC два часа, после чего раствор удаляли отсасыванием и ячейки промывали 400 микролитрами промывочного буфера (фосфатно-солевой буфер), содержащего 0,05% твина 20. Промытые ячейки обрабатывали 200 микролитрами сыворотки мыши против конъюгата IgG-HRP человека, содержащейся в растворе ортоконъюгата разбавителя (10 мМ фосфата натрия, pH 7,2, 150 мМ хлорида натрия, 50% (объем/объем) фетальной бычьей сыворотки, 1% по объему обработанной нагреванием сыворотки лошади, 1 мМ K3Fe(CN)6, 0,05% (вес/объем) твина 20, 0,02% (вес/объем) тимерозала). Обработку продолжали один час при 37oC, раствор удаляли отсасыванием и ячейки промывали промывочным буфером, который также удаляли отсасыванием. Чтобы определить количество связанного ферментного конъюгата, добавляли 200 микролитров субстратного раствора (10 мг ортофенилендиамина дигидрохлорида на 5 мл проявляющего раствора). Проявляющий раствор содержит 50 мМ цитрата натрия с pH 5,1, устанавливаемого фосфорной кислотой, и 0,6 микролитров/мл 30% перекиси водорода. Пластинки, содержащие субстратный раствор, инкубировали в темноте в течение 30 минут при комнатной температуре. Реакцию останавливали добавлением 50 микролитров/мл 4 н. серной кислоты и определяли оптические плотности.
Примеры, представленные ниже, показывают, что исследование микротитровых панелей иммуноферментным твердофазным анализом, в котором используется C100-3 антиген вируса гепатита C, имеет высокую степень специфичности, о чем свидетельствует начальная скорость реактивности около 1% с повторной скоростью реактивности около 0,5% на случайных донорах. Анализ способен определять иммунную реакцию как в после острой стадии течения инфекции, так и в хронической стадии болезни. В дополнение анализ может определять некоторые образцы, которые расцениваются как отрицательные в тестах-заменителях на гепатит ни A, ни B; эти образцы исходят от индивидов с анамнезом гепатита ни A, ни B, или от доноров, вовлеченных в перенос гепатита ни A, ни B.
В примерах, описанных ниже, использованы следующие сокращения:
ALT - аланинаминотрансфераза
Anti-HBc - антитело против HBc
Anti-HBsAg - антитело против HBsAg
HBc - сердцевинный антиген гепатита B
HBsAg - поверхностный антиген гепатита B
IgG - иммуноглобулин G
IgM - иммуноглобулин М
IU/L - международные единицы/литр
NA - не оценивалось
NT - не испытывалось
N - размер образца
Neg - отрицательный
OD - оптическая плотность
Pos - положительный
S/CO - сигнал/порог
SD - стандартное отклонение
x - среднее значение
WNL - в пределах нормы
IV.I.1. Инфекция гепатита C в популяции случайных доноров крови
Группа образцов в количестве 1056 (свежая сыворотка) была получена от случайных доноров крови из мемориального банка крови Ирвина, Сан-Франциско, Калифорния. Полученные результаты теста на этих образцах суммированы в гистограмме, показывающей распределение значений оптической плотности (фиг. 43). Как видно на фиг. 43, 4 образца показывают больше 3, один образец показывает между 1 и 3, пять образцов показывают между 0,4 и 1, а остальные 1046 образцов показывают менее 0,4, при этом свыше 90% этих образцов показывают оптическую плотность менее 0,1 нанометра.
Результаты проверки на реактивность случайных образцов представлены в табл. 5. Используя пороговое значение, равное среднему, плюс 5 стандартных отклонений, 10 образцов из 1056 (0,95%) были изначально реакционноспособными. Из них пять образцов (0,47%) подтвердили свою реактивность при повторном анализе иммуноферментным твердофазным методом. В табл. 5 показаны значения аланинаминотрансферазы и статус антител против сердцевинного антигена вируса гепатита B для каждого из образцов, повторно проявляющих реактивность. Особенный интерес вызывает тот факт, что все пять повторно реактивных образцов были отрицательными в тестах-заменителях на гепатит ни A, ни B, в то время как вели себя как положительные при иммуноферментном твердофазном анализе.
IV.I.2. Образцы сывороток шимпанзе
Образцы сыворотки одиннадцати шимпанзе были испытаны на C100-3 вируса гепатита C иммуноферментным твердофазным анализом. Четыре из этих шимпанзе были инфицированы гепатитом ни A, ни B от загрязненной пробы фактора VIII (предположительно штамм Hatchinson'a), следуя установленной процедуре в сотрудничестве с доктором Daniel Bradley в Центре контроля заболеваний. Для контроля 4 других шимпанзе заразили вирусом гепатита A и три-вирусом гепатита B. Образцы сывороток получали в различные периоды течения инфекции.
Результаты, которые обобщены в табл. 6, показывают документированное появление в сыворотке специфических антител во всех случаях заражения шимпанзе штаммом Hutchinson гепатита ни A, ни B. После острой фазы инфекции (как свидетельствует значительное возрастание и последующее возвращение к нормальным уровням аланинаминотрансферазы) антитела к C100-3 вируса гепатита С становились определяемыми в сыворотке четырех из четырех зараженных гепатитом ни A, ни B шимпанзе. Эти образцы, как обсуждалось в разделе IV.B.3, определялись ранее как положительные при анализе "западного" пятнения и при радиоиммунном анализе. В противоположность этому, ни один из контрольных шимпанзе, зараженных вирусом гепатита A или вирусом гепатита B, не проявлял свидетельства своей реактивности при иммуноферментном твердофазном анализе.
IV.I.3. Панель 1: достоверно подтвержденные инфекционные сыворотки носителей хронического гепатита ни A, ни B
Закодированная панель содержала 22 единичных образца, каждый из которых был представлен в двух экземплярах, всего 44 образца. Образцы взяты из сыворотки носителей гепатита ни A, ни B, инфекционность которой подтверждена диагностически, из инфекционной сыворотки больных гепатитом ни A, ни B в острой фазе. В дополнение были представлены образцы близкородственных отрицательных контрольных и других контрольных заболеваний. Эта панель разработана доктором H.Alter из департамента здоровья и обслуживания Национального института здоровья, Бетезда, Мериленд. Панель была сконструирована доктором Alter несколько лет назад и была им использована как квалифицированная панель для предполагаемых анализов гепатита ни A, ни B.
Всю панель подвергали анализу ELISA (иммуноферментный твердофазный анализ) два раза и результаты были посланы д-ру Alter на расшифровку. Результаты оценки представлены в табл. 7. Хотя в таблице представлены результаты одного набора образцов, те же результаты были получены от каждого двойного образца.
Как представлено в табл. 7, шесть сывороток, которые были достоверно инфекционными на модели шимпанзе, были строго положительными. Семь инфекционных сывороток, соответствующих образцу с острым гепатитом ни A, ни B, оказались нереакционноспособными при этом анализе ELISA. Образец от вовлеченного в процесс донора как с нормальными уровнями аланинаминотрансферазы, так и с сомнительными результатами при изучении на шимпанзе дал отрицательный анализ. Три других серийных образца от одного лица с острым гепатитом ни A, ни B были также нереакционноспособными. Все образцы от близкородственных отрицательных контролей, полученных от доноров, имевших по меньшей мере 10 сдач крови, без вовлечения гепатита, были нереактивными в иммуноферментном твердофазном анализе. В заключение, четыре испытанных образца, ранее расценивавшиеся как положительные в анализах предполагаемого гепатита ни A, ни B, разработанных другими исследователями, настоящими анализами не подтвердились. Эти образцы расценены отрицательно иммуноферментным твердофазным анализом гепатита C.
IV.I.4. Панель 2: донорно-реципиентный гепатит ни A, ни B
Закодированная панель содержала 10 достоверных случаев связанного с трансфузией гепатита ни A, ни B, в общей сложности 188 образцов. Каждый случай содержал образцы некоторых или всех доноров реципиенту и серийные образцы (взятые 3, 6 и 12 месяцев после трансфузии) от реципиента. Включены также образцы от лиц, которым предстояло переливание крови до проведения трансфузии. Закодированная панель была предложена д-ром Альтером из Национального института здоровья и полученные результаты были посланы ему для оценки.
Результаты, которые обобщены в табл. 8, показывают, что иммуноферментный твердофазный анализ определяет гепатит ни A, ни B, связанный с трансфузией, в 9 случаях из десяти. Образцы от случая 4 (в которых не определялась сероконверсия) постоянно слабо реагировали при проведении иммуноферментного твердофазного анализа. Два из десяти образцов сыворотки реципиентов были реактивны через три месяца после трансфузии. Через 6 месяцев 8 образцов сыворотки реципиентов были реактивными и через 12 месяцев, за исключением случая 4, все образцы были реактивными. В дополнение по меньшей мере положительный на антитело донор был найден в 7 из 10 случаев, при этом в случае 10 имелось два положительных донора. Итак, в случае 10 реципиент до переливания крови был положительным на антитела против вируса гепатита C. Через месяц кровь этого реципиента упала до граничного значения реактивных уровней, тогда как оно повышалось до положительного через 4 и 10 месяцев после переливания. Таким образом, этот случай может представлять собой первичную инфекцию индивида вирусом гепатита C.
Состояние аланинаминотрансферазы и сердцевинного антигена вируса гепатита B для всех реактивных, т.е. положительных образцов, обобщены в табл. 9. Как видно из таблицы, образцы сыворотки доноров 1-8 были отрицательны для маркеров-заменителей и реактивны в иммуноферментном твердофазном анализе антител к вирусу гепатита C. С другой стороны, образцы сыворотки реципиентов (взятые через 12 месяцев после трансфузии) имели повышенное содержание аланинаминотрансферазы, положительные антитела против сердцевинного антитела вируса B или то и другое вместе.
IV.I.5. Определение инфекции вируса гепатита C в группе высокого риска
Образцы сыворотки из группы повышенного риска обсчитывали, используя иммуноферментный твердофазный анализ для определения реактивности к антигену C100-3 вируса гепатита C. Эти образцы были получены от д-ра Gary Tegtmeier, Community Blood Bank, Kansas City. Результаты обобщены в табл. 10.
Как видно из таблицы, образцы с наивысшей реактивностью были получены у больных гемофилией (76%). В дополнение, образцы от индивидов с повышенным содержанием аланинаминотрансферазы и положительные на антитела против сердцевинного антигена вируса гепатита B оценивались на 51% реактивности, значение, которое согласуется со значением из клинических данных, и преобладание гепатита ни A, ни B в этой группе. Появление антител к вирусу гепатита C было также выше в крови доноров с повышенным уровнем аланинаминотрансферазы только в крови доноров, положительной на антитела к антигену сердцевины вируса гепатита B и в крови доноров, отклоненной по причинам иным, чем высокое содержание аланинаминотрансферазы или наличия антител против сердцевинного антигена вируса гепатита B в сравнении со случайными добровольцами донорами.
IV.I.6. Сравнительные исследования моноклональных антител против иммуноглобулина G или иммуноглобулина М, или поликлональных антител как второго антитела в иммуноферментном твердофазном анализе антигена C100-3 вируса гепатита C
Чувствительность метода ELISA с использованием конъюгата моноклональных антител с антииммуноглобулином G сравнивали с чувствительностью, полученной с использованием конъюгата моноклональных антител к антииммуноглобулину М или при замещении моноклональных антител поликлональной антисывороткой, которая, как сообщалось, специфична с тяжелой и легкой цепью. Были проведены следующие исследования.
IV.I.6.a. Серийные образцы от сероконвертеров
Серийные образцы от трех случаев сероконвертеров вируса ни A, ни B были исследованы иммуноферментным анализом в твердой фазе антигена C100-3 вируса гепатита C с использованием ферментного конъюгата либо только с моноклональным антителом к иммуноглобулину G, или в сочетании с моноклональным антителом против иммуноглобулина М, или используя поликлональную антисыворотку. Образцы были предоставлены д-ром Cladd Stevens, N.Y. Blood Center, N. Y.C., N.Y. Истории образцов показаны в табл. 11.
Результаты, полученные с ферментным конъюгатом с моноклональным антителом против иммуноглобулина G, представлены в табл. 12. Эти данные свидетельствуют о том, что сильную реактивность сначала определяли в образцах 1-4, 2-8 и 3-5 случаев 1, 2 и 3 соответственно.
Результаты, полученные с использованием сочетания моноклонального конъюгата против иммуноглобулина G и конъюгата против иммуноглобулина М, показаны в табл. 13. Исследовали три различных отношения антител против иммуноглобулина G к антителам иммуноглобулина М: разбавления 1:10000 были постоянными во всех исследованиях. Испытанные разведения для моноклонального конъюгата против иммуноглобулина М были: 1:30000, 1:60000 и 1:120000. Данные свидетельствуют, что, в согласии с изучением только с одним антииммуноглобулином G, начальная сильная реактивность определялась в образцах 1-4, 2-8 и 3-5.
Результаты, полученные анализом ELISA с использованием моноклонального конъюгата антител против иммуноглобулина G (разбавление 1:10000), или с поликлональным конъюгатом Tago (разведение 1:80000), или поликлональным конъюгатом Jackson (разбавление 1:80000), показаны в табл. 14. Эти данные показывают, что начальная сильная реактивность определялась в образцах 1-4, 2-8 и 3-5 с использованием всех трех конфигураций: поликлональные антитела Tago дали самые низкие сигналы.
Представленные выше результаты показывают, что все три конфигурации определяют реактивные образцы в одно и то же время после острой фазы болезни (как свидетельствует повышение уровня аланинаминотрансферазы). Кроме того, результаты показывают, что чувствительность иммуноферментного твердофазного анализа антигена с 100-3 вируса гепатита C с использованием ферментного конъюгата с моноклональным антителом против иммуноглобулина G равна или выше чувствительности, полученной с использованием других испытываемых конфигураций ферментных конъюгатов.
IV.I.6.b. Образцы от случайных доноров крови
Образцы от случайных доноров крови были исследованы на инфекцию вируса гепатита C (см. раздел IV.I.1), используя иммуноферментный твердофазный анализ C100-3 вируса гепатита C, в котором конъюгатом антитело - фермент являлся либо моноклональный конъюгат антииммуноглобулина G, или поликлональный конъюгат. Общее число проверенных образцов составляло 1077 и 1056 для поликлонального конъюгата и моноклонального конъюгата соответственно. Суммарный результат скрининга представлен в табл. 15 и распределение образцов показано на гистограмме на фиг. 44.
Расчеты среднего значения и стандартного отклонения были проведены, исключая образцы, которые давали сигнал выше 1,5, т.е. 1073 значения оптической плотности были использованы в расчетах при применении поликлонального конъюгата и 1051 значение для моноклонального конъюгата антииммуноглобулина G. Как видно из табл. 15, если использовали поликлональный конъюгат, то среднее значение отклонялось от 0,0493 до 0,0931, а стандартное отклонение увеличивалось от 0,074 до 0,0933. Кроме того, результаты показывают также, что если применяют критерий x +5 стандартных отклонений для определения порога анализа, то конфигурация поликлональное антитело - фермент конъюгата требует при ELISA более высокого порогового значения. Это указывает на пониженную специфичность анализа в сравнении с моноклональной системой. В дополнение, как показано на гистограмме на фиг. 44, большее разделение результатов между положительными и отрицательными распределениями появляется, если случайные доноры крови проверяются иммуноферментным твердофазным анализом с использованием моноклонального конъюгата антииммуноглобулина G в сравнении с анализом, использующим коммерческую поликлональную метку.
IV.J. Определение сероконверсии вируса гепатита C у больных гепатитом ни A, ни B из различных географических локализаций
Сыворотки больных, подозрительных на гепатит ни A, ни B на основании повышенных уровней аланинаминотрансферазы и у которых были отрицательные пробы на вирус гепатита A и вирус гепатита B, проверяли с использованием радиоиммунного анализа в основном так, как описано в разделе IV.D, за исключением того, что антиген C100-3 вируса гепатита C использовали как отсеивающий антиген в микротитровых панелях. Как видно из результатов, представленных в табл. 16, радиоиммунный анализ определял положительно образцы в высоком проценте случаев.
IV. K. Определение сероконверсий вируса гепатита C у больных "бытовым" (коммунальным) гепатитом ни A, ни B
Сыворотки, полученные от 100 больных гепатитом ни A, ни B, у которых не было очевидного пути передачи (т.е. не было переливания крови, внутривенного введения лекарств, беспорядочных связей и так далее, которые идентифицированы как факторы риска), были предоставлены д-ром М.Альтером из Центра по контролю заболеваний и д-ром J.Dienstag из Гарвардского университета. Эти образцы были проверены с использованием радиоиммунного анализа в основном так, как описано в разделе IV.D, за исключением того, что использовали антиген C100-3 вируса гепатита C как исключающий антиген, связанный с микротитровыми панелями. Результаты показывают, что из 100 образцов сыворотки 55 образцов содержали антитела, реагирующие иммунологически с антигеном C100-3 вируса гепатита C.
Описанные выше результаты наводят на мысль о том, что "коммунальный" гепатит ни A, ни B также вызывается вирусом гепатита C. Кроме того, поскольку это было продемонстрировано здесь, что вирус гепатита C относится к флавивирусам, большинство из которых переносятся членистоногими, то можно полагать, что передача вируса гепатита C в "бытовых" случаях является также результатом переноса членистоногими.
IV. L. Сравнение частоты появления антител против вируса гепатита C и маркеров-заменителей у доноров, вовлеченных в перенос гепатита ни А, ни B
Перспективное исследование было проведено для определения того, являются ли реципиенты крови доноров, подозрительных на гепатит ни A, ни B, которые проявляют гепатит ни A, ни B, сероконвертированы положительно к антителам против вируса гепатита С. Донорскую кровь испытывали другими маркерами анормальностей, т. е. повышенные уровни аланинаминотрансферазы и присутствие антител против сердцевинного антигена. В дополнение доноры были проверены также на присутствие антител против вируса гепатита C, проводили с использованием радиоиммунного анализа, как описано в разделе IV.K. Результаты исследования представлены в табл. 17, которая показывает: количество больных (колонка 1); присутствие антител против вируса гепатита C в сыворотке больных (колонка 2); количество вливаний, полученных больным с каждым переливанием, полученным от различного донора (колонка 3); присутствие антител в сыворотке донора (колонка 4) и отклонения от нормы, определенные другими маркерами в крови донора (колонка 5) (NT или -- означает "не испытывали") (ALT означает "повышенная трансминаза и ANTI-HBc означает антитело против сердцевинного антигена вируса гепатита B).
Результаты, представленные в табл. 17, показывают, что тест на антитела к вирусу гепатита C является более точным в определении инфицированных доноров крови, чем тесты маркерами-заменителями. Девять из десяти больных, у которых проявлялись симптомы гепатита ни A, ни B, дали положительный тест на сероконверсию антител против вируса гепатита C. Из 11 подозрительных доноров (пациент 6 получал кровь от двух различных индивидов, подозрительных на носителей вируса гепатита ни A, ни B) девять были положительными на антитела против вируса гепатита C и один больной был на нижнем пределе положительного и поэтому сомнителен (донор для больного 1). В противовес этому, используя тест на повышенную трансминазу шесть из десяти доноров дали отрицательный результат теста, а используя тест антителосердцевинный антиген пять из десяти доноров показали отрицательный результат теста. Более последователен тест на антитела против сердцевинного антигена вируса гепатита B, дали непоследовательные результаты.
IV. М. Увеличение клонирования последовательностей кольцевой ДНК вируса гепатита C, используя реакцию роста полимеразной цепи и инициаторы, полученные из законсервированных участков геномных последовательностей флавивируса
Результаты, представленные выше, говорят в пользу того, что вирус гепатита C является флавивирусом или флавиподобным вирусом, и позволяют стратегию клонирования неохарактеризованных последовательностей кольцевой ДНК вируса гепатита C, используя реакцию роста полимеразной цепи и инициаторы, полученные из участков, кодирующих последовательности законсервированных аминокислот флавивирусов. Вообще один из инициаторов получен из определенной геномной последовательности вируса гепатита C, а другой инициатор, который примыкает к участку непоследовательного полинуклеотида вируса гепатита C, получен из законсервированного участка генома флавивируса. Геномы флавивируса известны как содержащие консервированные последовательности в NS1 и E полипептиды, которые закодированы в 5'-участке генома флавивируса. Соответствующие последовательности, кодирующие эти участки, лежат на восходящей от последовательности кольцевой ДНК вируса гепатита C, показанной на фиг. 26. Таким образом, чтобы выделить последовательность кольцевой ДНК, производную из этого участка генома вируса гепатита C, конструировали восходящие инициаторы, которые получены из законсервированных последовательностей в этих полипептидах флавивирусов. Восходящие инициаторы получали от восходящего конца известной части кольцевой ДНК вируса гепатита C.
Из-за нарушений кода вероятны несоответствия между пробами флавивируса и соответствующей геномной последовательностью вируса гепатита C. Поэтому используют подход, аналогичный описанному Lee (1988). Метод Lee использует смешанные олигонуклеотидные инициаторы, комплементарные продуктам обратной трансляции аминокислотной последовательности; последовательности в смешанных инициаторах принимают во внимание каждое нарушение кодона для консервированной последовательности аминокислот.
Были генерированы три состава инициирующих смесей, основываясь на аминокислотных гомологах, найденных у различных флавивирусов, включая Dengue-2,4 (D-2,4), японский энцефалитный вирус, вирус желтой лихорадки и вирус Западного Нила. Инициирующая смесь, полученная из главным образом восходящей законсервированной последовательности (5'-1), основанной на аминокислотной последовательности глицин-триптофан-глицин, которая является частью законсервированной последовательности аспарагин-аргинин-глицин-триптофан-глицин-аспарагин, найденной в E белке вируса Денгье, японского энцефалитного вируса, вируса желтой лихорадки и вируса Западного Нила. Следующая инициирующая смесь (5'-2) основана на нисходяще законсервированной последовательности в E белке, т.е. фенилаланин- аспарагин-глицин-аспарагин-серин-тиронин-изолейцин-фенилаланин- глицин-аспарагин-серин-тиронин-изолейцин, и получена из фенилаланина-глицина-аспарагина; консервированная последовательность присутствует в вирусах Денгье, японском энцефалитном, желтой лихорадки и Западного Нила. Третья инициирующая смесь (5'-3) основана на аминокислотной последовательности аргинин-серин-цистин, которая является частью консервированной последовательности цистин-цистин-аргинин-серин-цистин в белке NS 1 вирусов Денгье-2, D-4, японского энцефалитного, желтой лихорадки и Западного Нила. Отдельные инициаторы, которые образуют смесь в 5'-3, показаны на фиг. 45. В дополнение к различным последовательностям, полученным от консервированного участка, каждый инициатор в каждой смеси также содержит постоянный участок в 5'-конце, который содержит последовательность, кодирующую локализации для ограничивающих ферментов: HindIII, MboI и EcoRI.
Нисходящий инициатор ssc5h20A получен из нуклеотидной последовательности в клоне 5h, который содержит последовательности кольцевой ДНК вируса гепатита C, перекрывающие последовательности в клонах 14i и 11b. Последовательность ssc5h20A является:
5' GTA ATA TGG TGA CAG AGT CA 3'.
Может быть получен также альтернативный инициатор ssc5h34A. Этот инициатор получают из последовательности в клоне 5h, в дополнение содержит нуклеотиды в 5'-конце, которые образуют локализацию ограничивающего фермента, облегчая таким образом клонирование. Последовательность ssc5h34A следующая:
5' GAT CTC TAG AGA AAT CAA TAT GGT GAC AGA GTC A 3'.
Реакцию полимеразной цепи, которую первоначально описали Saiki и соавт. (1986), проводили, в основном, по методу Lee и соавт. (1988), за исключением того, что матрицей для кольцевой ДНК является РНК, выделенная из печени шимпанзе, инфицированного вирусом гепатита C, как описано в разделе IV.C.2, или из вирусных частиц, выделенных из сыворотки инфицированных вирусом гепатита C шимпанзе, как описано в разделе IV.A.1. B дополнение условия ренатурации менее строгие в первом раунде увеличения (0,6 М хлористого натрия и 25oC), поскольку часть инициатора, которому предстоит ренатурировать до последовательности вируса гепатита C, является последовательностью только 9 нуклеотидов и здесь могут возникать несоответствия. Кроме того, если используют ssc5h34A, то дополнительные последовательности, не полученные из генома вируса гепатита C, имеют тенденцию дестабилизировать гибрид матрица - инициатор. После первой стадии увеличения условия ренатурации могут быть, более строгими (0,066 М хлористого натрия и 32-37oC), поскольку увеличенные последовательности содержат теперь участки, которые комплементарны инициаторам или дублируют их. В дополнение первые 10 циклов увеличения протекают с ферментом 1 Кленова при подходящих условиях для этого фермента. После завершения этих циклов образцы экстрагировали и разгоняли Tag полимеразой в соответствии с инструкциями, выработанными Cetus/Perkin-Elmer.
После увеличения увеличенные последовательности кольцевой ДНК вируса гепатита C определяли гибридизацией с использованием пробы, полученной из клона 5h. Эта проба получена из последовательностей, восходяще направленных к последовательностям, использованным для получения инициатора, и не перекрывает последовательности инициатора, производных из клона 5h. Последовательность пробы такова:
5' CCC AGC GGC GTA CGC GCT GGA CAC GGA GGT GGC CGC GTC GTG TGG CGG TGT TGT TCT CGT CGG GTT GAT GGC GC 3'.
IV.N.1. Создание пула кольцевой ДНК вируса гепатита C из печени шимпанзе с инфекционным гепатитом ни A, ни B
Пул кольцевой ДНК вируса гепатита C был создан из печени шимпанзе, из которой был создан пул кольцевой ДНК вируса гепатита C в разделе IV.A.1. Методика создания пула была аналогична таковой, описанной в разделе IV.A.24, за исключением этого отличного источника РНК и того, что был использован инициатор, основанный на последовательности кольцевой ДНК вируса гепатита C в клоне 11b. Последовательность инициатора была следующей:
5' CTG GCT TGA AGA ATC 3'.
IV. N. 2. Выделение нуклеотидной последовательности кольцевой ДНК вируса гепатита C в клоне k9-1, перекрывающей кольцевую ДНК в клоне 11b
Клон k9-1 был выделен из пула кольцевой ДНК вируса гепатита C, созданного из печени инфицированного гепатитом ни A, ни B шимпанзе, как описано в разделе IV. A. 25. Пул исследовали на клоны, которые перекрывают последовательность в клоне 11b, путем использования клона, который перекрывает клон 11b в 5'-терминале, т.е. клона 11e. Последовательность клона 11b показана на фиг. 23. Положительные клоны были выделены с частотой один клон на 500000. Один изолированный клон, k9-1, подвергали дальнейшему изучению. Природа перекрытия кольцевой ДНК вируса гепатита C в клоне k9-1 в 5'-конце последовательности кольцевая ДНК-вируса гепатита C на фиг. 26 была подтверждена испытанием клона с клоном Alех46; этот последний клон содержит последовательность кольцевой ДНК вируса гепатита C в 30 пар оснований, которая соответствует последовательности пар оснований при 5' терминале кольцевой ДНК вируса гепатита C в клоне 14i, описанном выше.
Нуклеотидная последовательность кольцевой ДНК вируса гепатита C из клона k9-1 была определена с использованием метода, описанного выше.
Последовательность кольцевой ДНК вируса гепатита C в клоне k9-1, перекрытие с кольцевой ДНК вируса гепатита C на фиг. 26 и закодированные в ней аминокислоты показаны на фиг. 46.
Последовательность кольцевой ДНК вируса гепатита C в клоне k9-1 была упорядочена в линию с последовательностями клонов, описанных в разделе IV.A. 19, чтобы создать композиционную последовательность кольцевой ДНК вируса гепатита C с последовательностью k9-1, помещенной вверху в направлении последовательности, показанной на фиг. 32. Композиционная кольцевая ДНК вируса гепатита C, которая включает последовательность клона k9-1 и закодированные в ней аминокислоты, представлены на фиг. 47.
Последовательность аминокислот, закодированных в 5'-участке кольцевой ДНК вируса гепатита C, показанной на фиг. 47, сравнивали с соответствующим участком одного из штаммов вируса Dengue, описанного ввыше, с учетом профиля участков гидрофобности и гидрофильности. Это сравнение показало, что полипептиды от вируса гепатита C и вируса Dengue, закодированные в этом участке, который соответствует участку кодирования белка NS1 (или его части), имеют аналогичный гидрофобный/гидрофильный профиль.
Представленная ниже информация позволяет идентифицировать штаммы вируса гепатита C. Выделение и характеристики других штаммов вируса гепатита C могут быть осуществлены путем выделения нуклеиновых кислот из компонентов тела, содержащего вирусные частицы, создания пулом кольцевой ДНК, используя полинуклеотидные пробы, основанные на пробах кольцевой ДНК вируса гепатита C, описанных выше, исследования пулов на отсеивание клонов, содержащих последовательности кольцевой ДНК вируса гепатита C, описанные ниже, и сравнения кольцевых ДНК вируса гепатита C из вновь выделенных образцов с кольцевыми ДНК, описанными ниже. Полипептиды, закодированные в них или в вирусном геноме, могут быть зарегистрированы и проверены на иммунологическую перекрестную реактивность, используя описанные выше полипептиды и антитела, описанные выше. Штаммы, которые отвечают параметрам вируса гепатита C, как описано в разделе определений выше, легко идентифицируемы. Другие методы идентификации штаммов вируса гепатита C будут очевидны специалистам в данной области на основании представленной здесь информации.
Промышленная применимость
Настоящее изобретение, раскрытое в различных проявлениях в описании, имеет различное промышленное применение, к которому можно отнести следующее. Кольцевые ДНК вируса гепатита C могут быть использованы для конструирования проб для определения нуклеиновых кислот вируса гепатита C в образцах. Пробы, полученные из кольцевых ДНК, могут быть использованы для определения нуклеиновых кислот вируса гепатита C, например, в реакциях химического синтеза. Они могут быть также использованы в программах скрининга антивирусных агентов, для определения действия агентов в ингибировании вирусной репликации и системах клеточных культур и на модельных системах животных. Полинуклеотидные пробы вируса гепатита C также полезны при определении вирусных нуклеиновых кислот у человека, и таким образом, могут служить основанием для диагностики инфекции вируса гепатита C у людей.
В дополнение к сказанному выше, описанные здесь кольцевые ДНК дают информацию о средстве для синтезирования полипептидов, содержащих эпитопы вируса гепатита C. Эти полипептиды полезны при определении антител к антигенам вируса гепатита C. Ряд иммунологических анализов на инфекцию вируса гепатита C, основанных на рекомбинантных полипептидах, содержащих эпитопы вируса гепатита C, приведены в описании и могут найти коммерческое использование в диагностике вируса гепатита C, вызванного гепатитом ни A, ни B, и исследовании (скрининге) банков крови доноров на выявление вызванных вирусом гепатита C гепатитов, а также для определения заражения крови от инфекционных доноров крови. Вирусные антигены могут быть также полезны при оценке эффективности антивирусных агентов в модельных системах животных. В дополнение полипептиды, полученные из кольцевых ДНК вируса гепатита C, описанные в заявке, могут найти применение в качестве вакцин для лечения инфекций вируса гепатита C.
Полипептиды, полученные из кольцевых ДНК, кроме уже упоминавшегося применения, могут быть также полезны для стимулирования появления антител против вируса гепатита C. Таким образом, они могут быть использованы в вакцинах против вируса гепатита C. Разумеется, что и антитела, образующиеся в результате иммунизации полипептидами вируса гепатита C, могут быть также полезны для определения наличия вирусных антигенов в образцах. Следовательно, они могут быть использованы также для исследования продуцирования полипептидов вируса гепатита C в химических системах. Антитела против вируса гепатита C могут быть также использованы для оценки эффективности противовирусных агентов при реализации скрининговых программ, в которых эти агенты испытывают в системах культур тканей. Они могут быть использованы также для пассивной иммунотерапии и для диагностики гепатита ни A, ни B, вызванного вирусом гепатита C, позволяющей определение вирусного антигена(ов) как в крови доноров, так и реципиентов. Другое важное применение находят антитела против вируса гепатита C в аффинной хроматографии для очистки вируса и вирусных полипептидов. Эти очищенные препараты вируса и вирусных полипептидов могут быть использованы в вакцинах. Разумеется, что очищенные вирусы могут быть полезны для развития систем клеточных культур, в которых реплицирует вирус гепатита C.
Системы клеточных культур, содержащие зараженные вирусом гепатита C клетки, могут найти различное применение. Они могут быть использованы для крупномасштабного продуцирования вируса гепатита C, который нормально является вирусом низкого титра. Эти системы могут быть полезны для разъяснения молекулярной биологии вируса и привести к разработке противовирусного агента(ов). Системы культур клеток будут также полезны в исследованиях эффективности антивирусных агентов. В дополнение системы клеточных культур, прошиваемых для вируса гепатита C, полезны для продуцирования ослабленных штаммов вируса гепатита C.
Для удобства антитела против вируса гепатита C и полипептида вируса гепатита C, природные или рекомбинантные, могут быть упакованы в наборы (комплекты).
Метод, использованный для выделения кольцевой ДНК вируса гепатита C, который включает получение пула кольцевой ДНК, полученной из инфицированной ткани индивидов в векторе экспрессии, и выбор клонов, которые продуцируют продукты экспрессии, реагирующие иммунологически с антителами в содержащих антитела компонентах тела от другого(их) инфицированных индивидов, но не от незараженных индивидов, может также найти применение для выделения кольцевых ДНК, полученных от других неохарактеризованных связанных с заболеванием агентов, которые содержат геномный компонент. Это, в свою очередь, может привести к выделению и характеристике этих агентов и к диагностическим реагентам и вакцинам против этих других связанных с болезнью агентов.
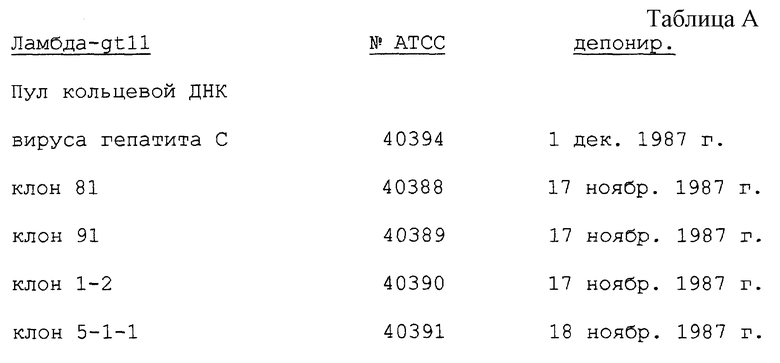


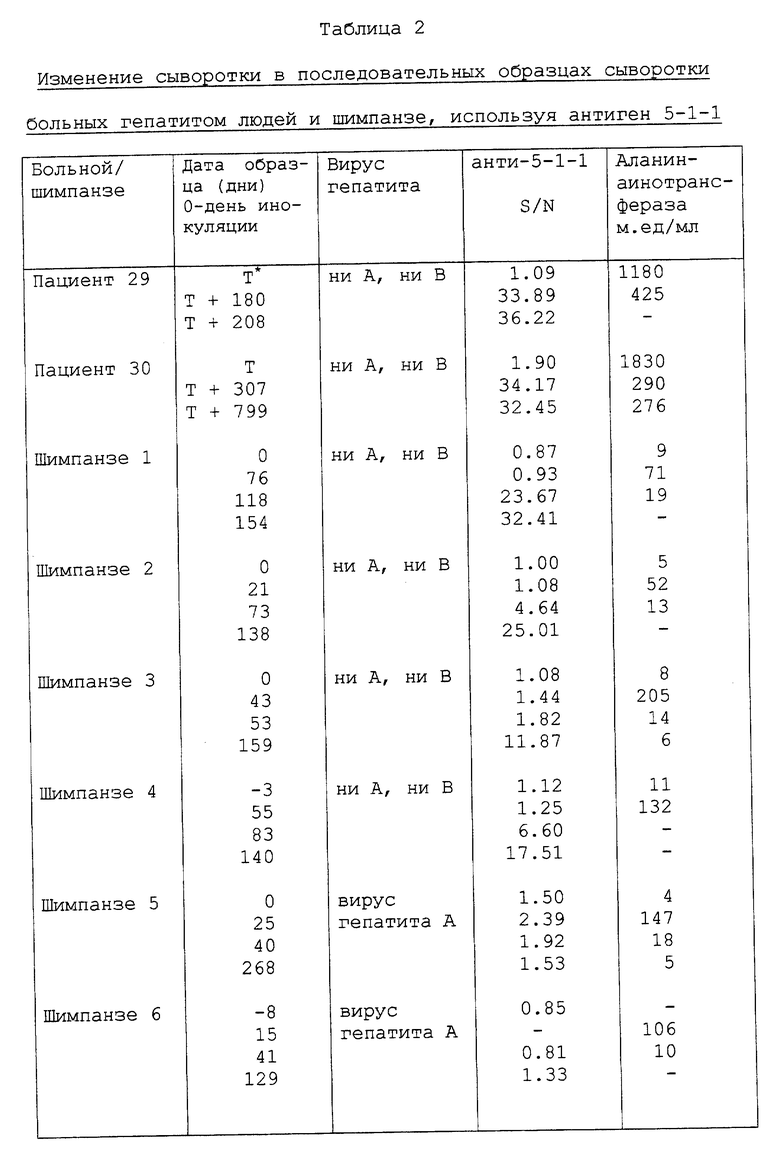
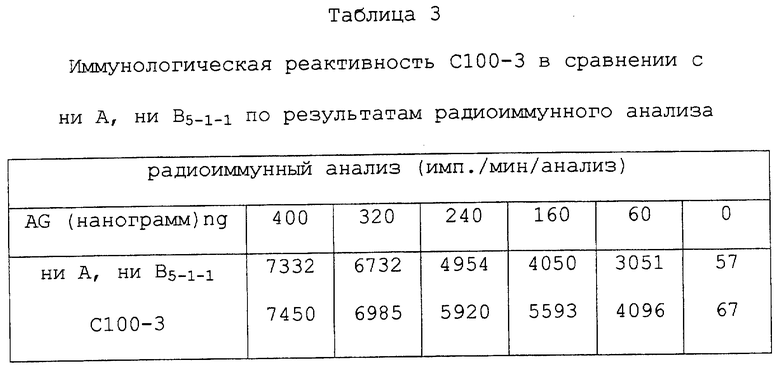

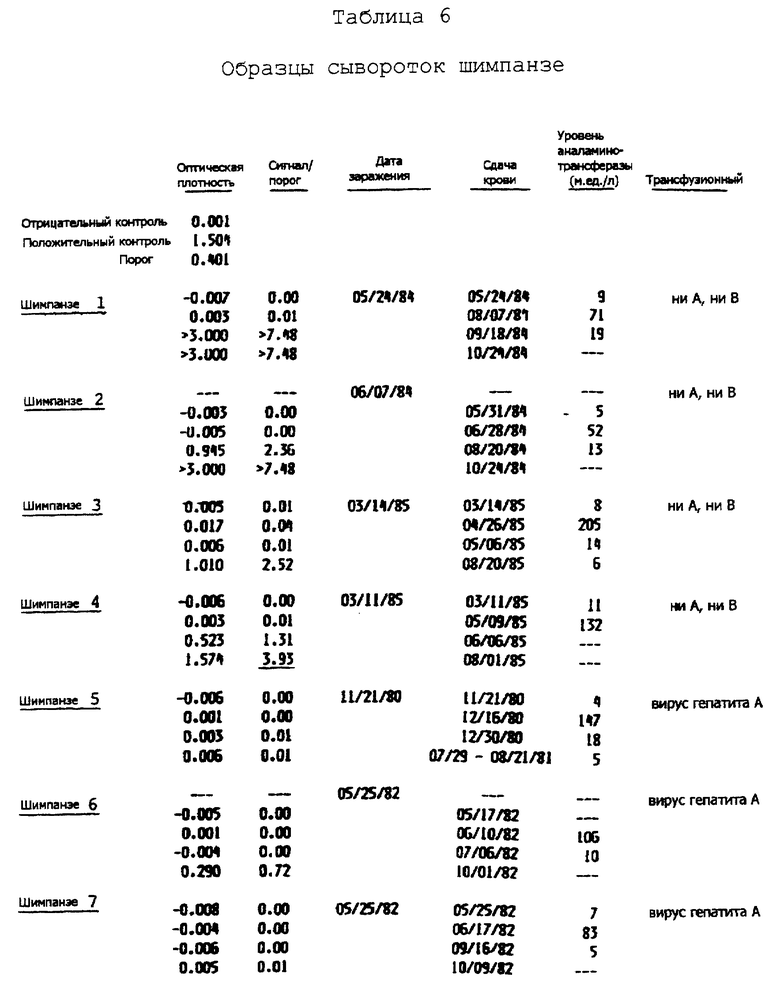
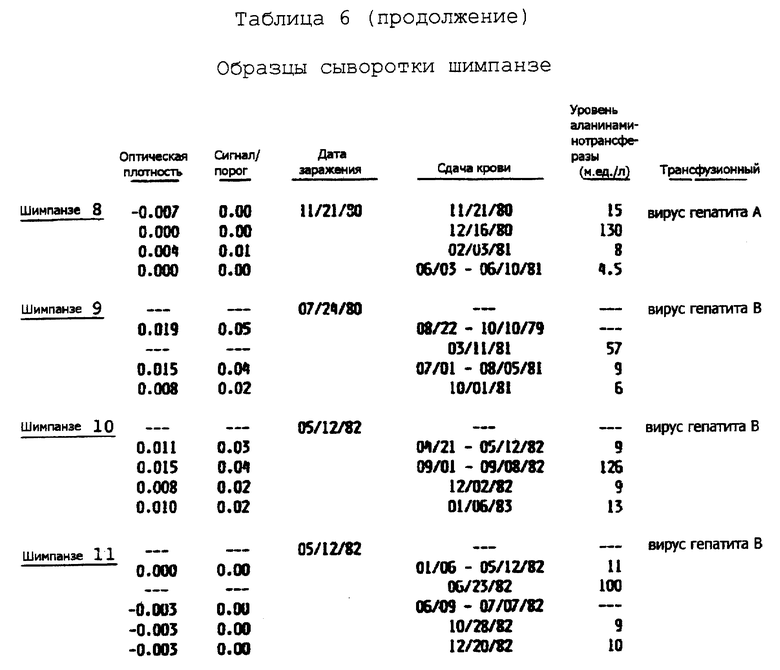

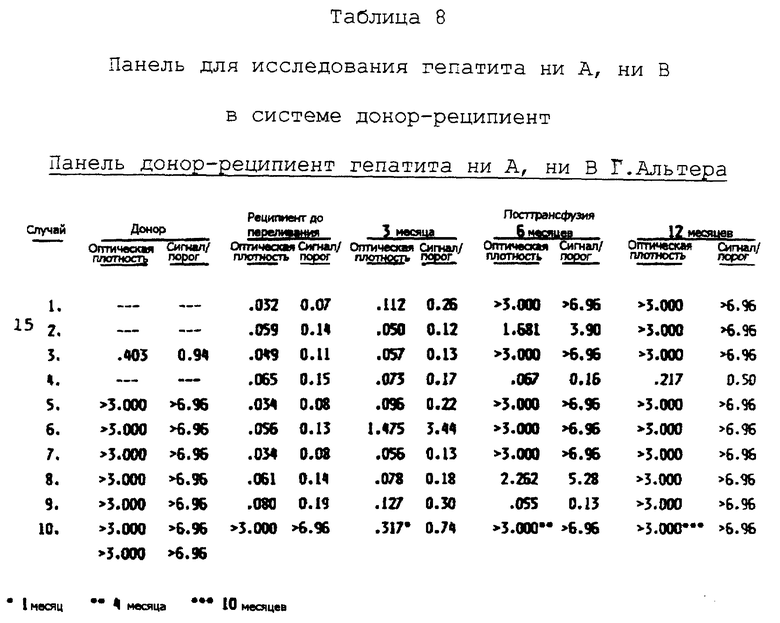
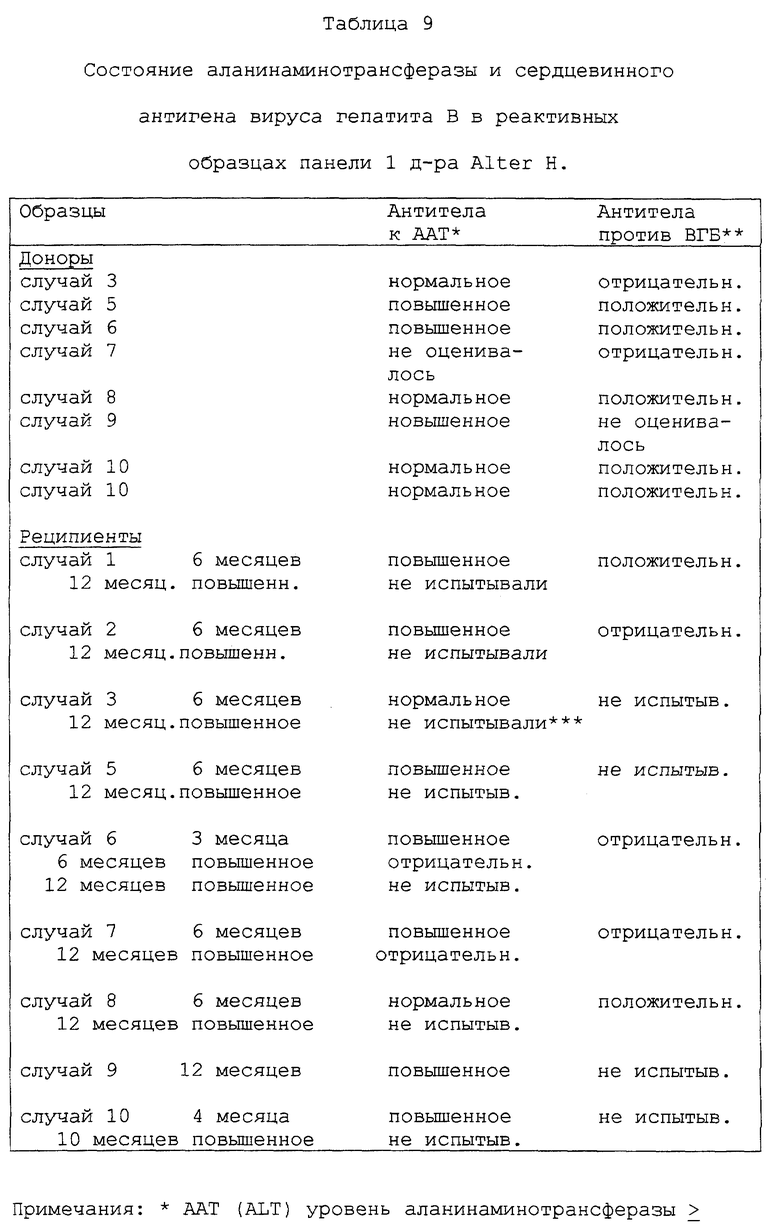

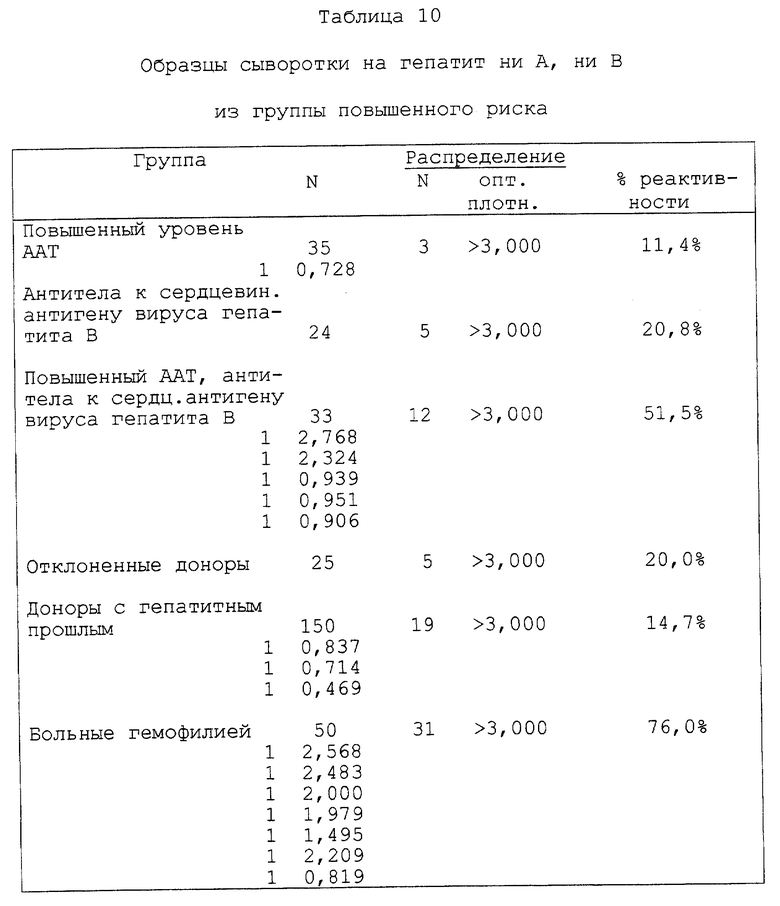
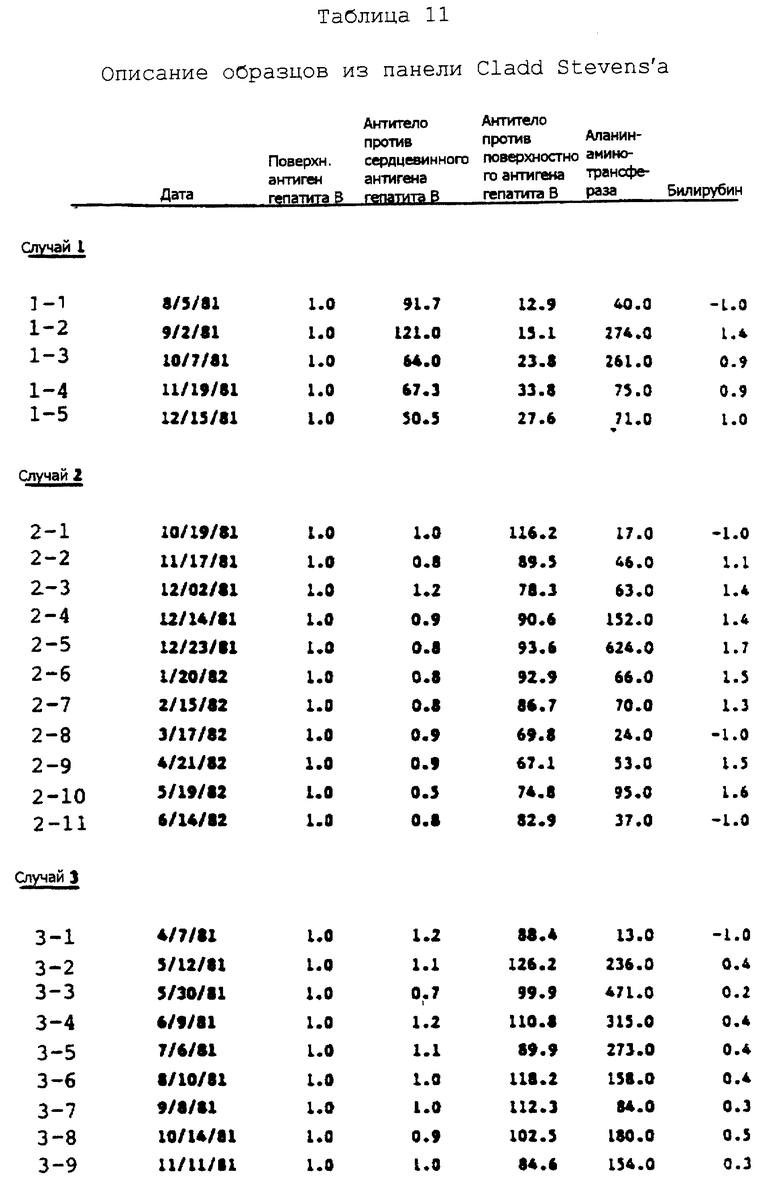
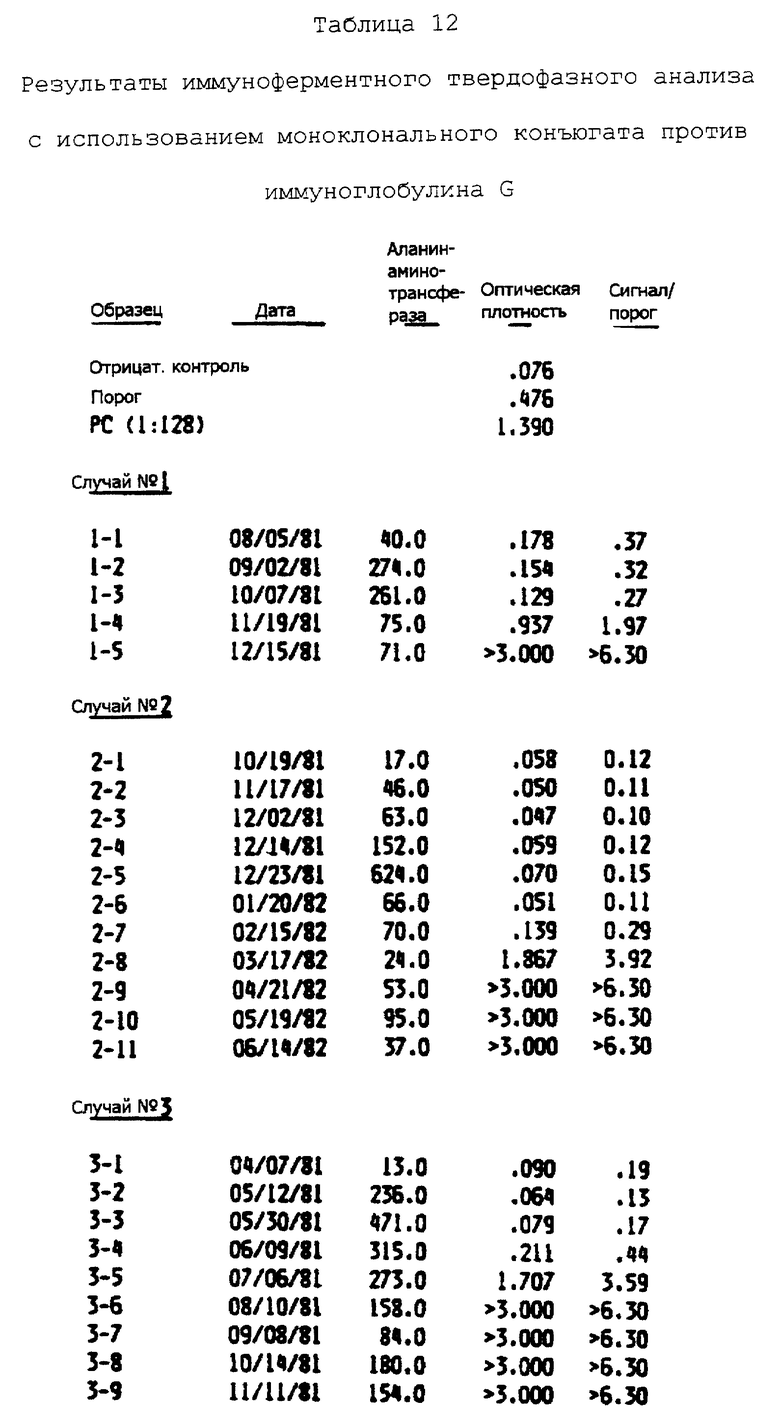
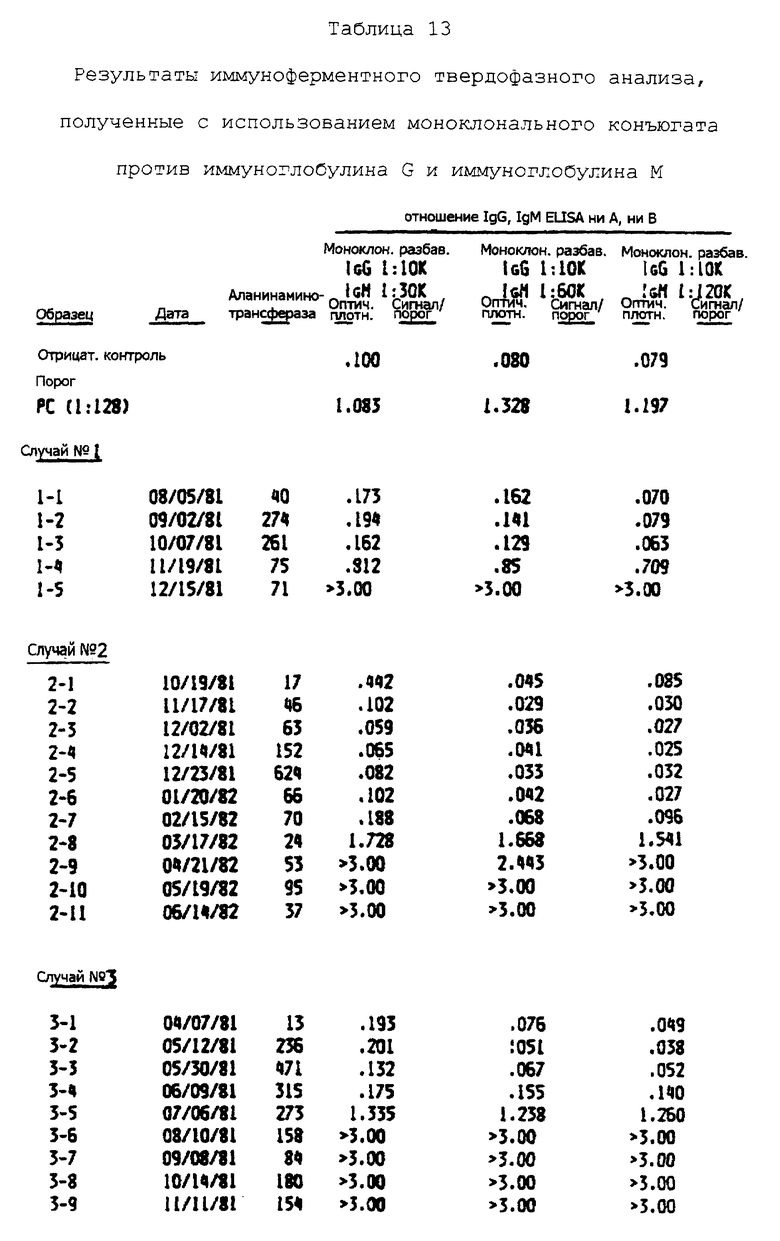
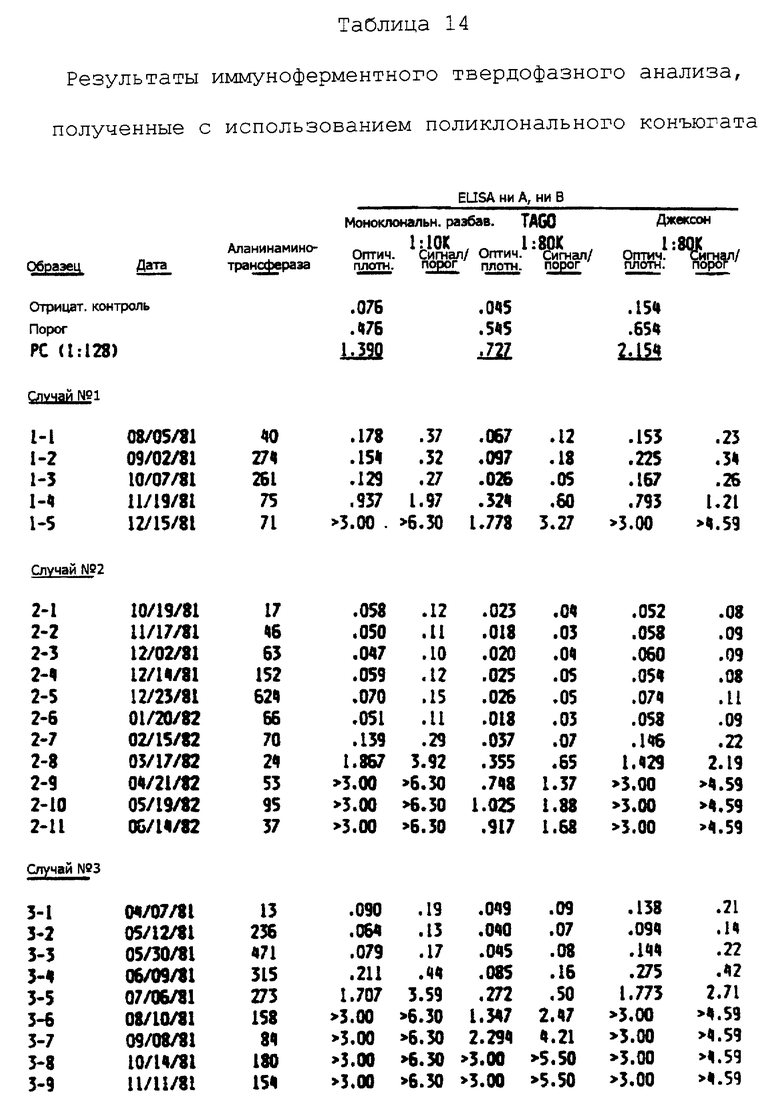
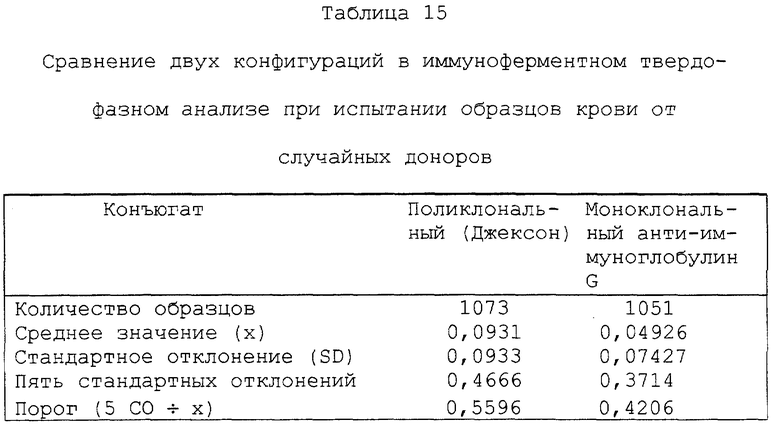

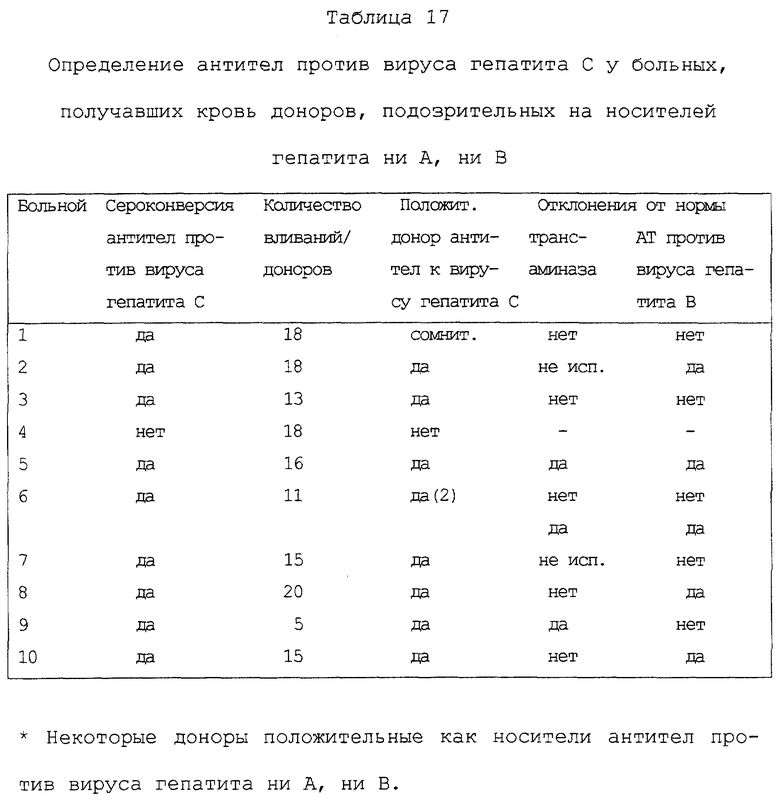
















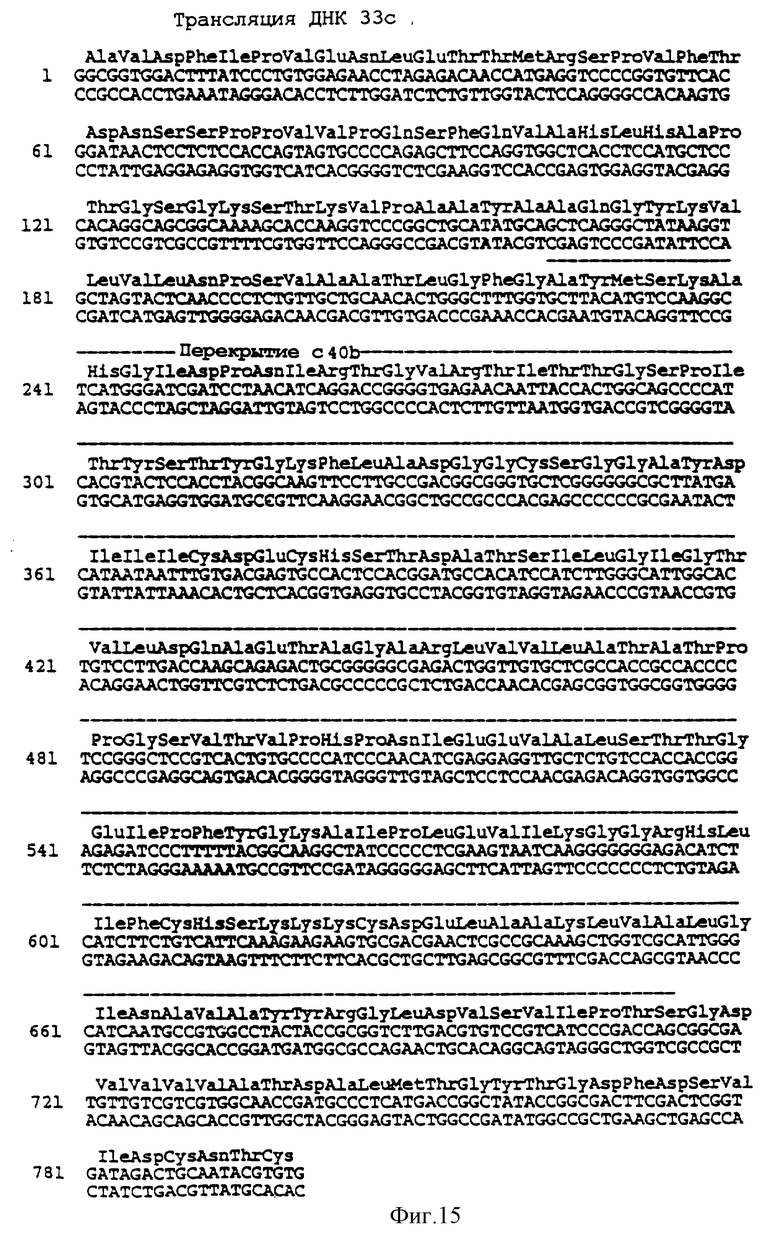































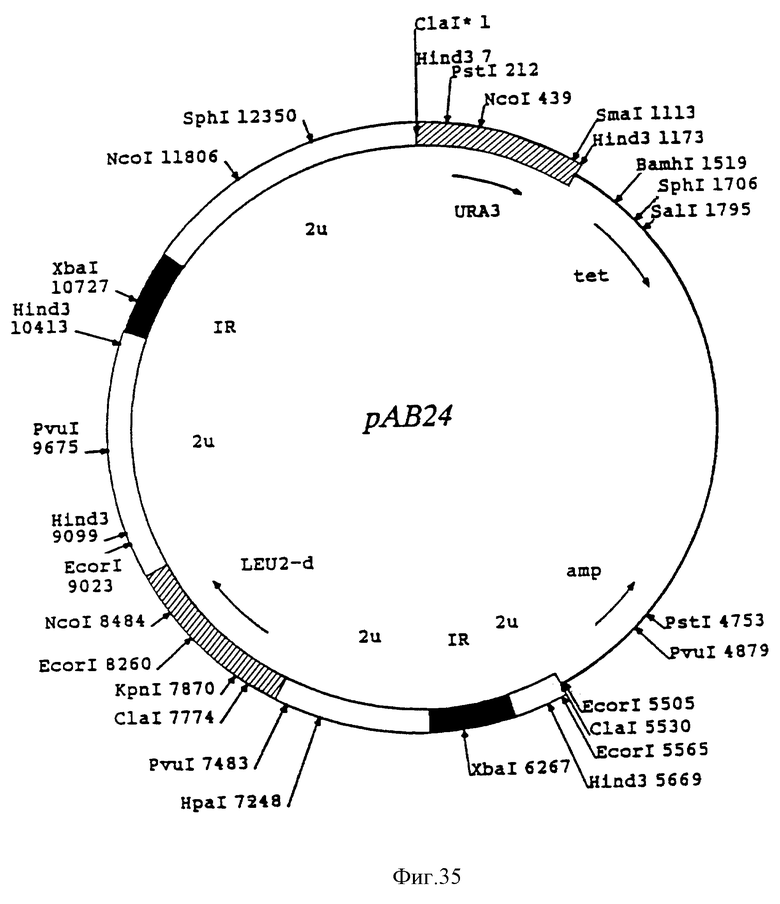







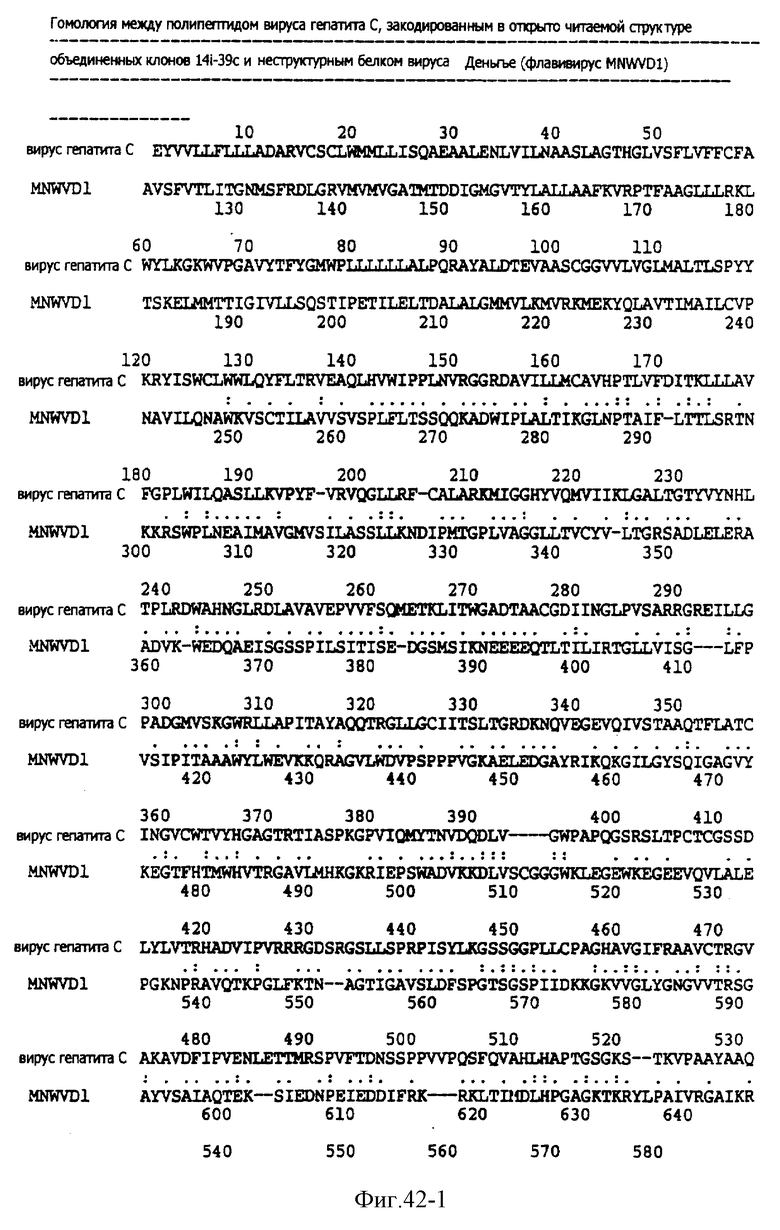

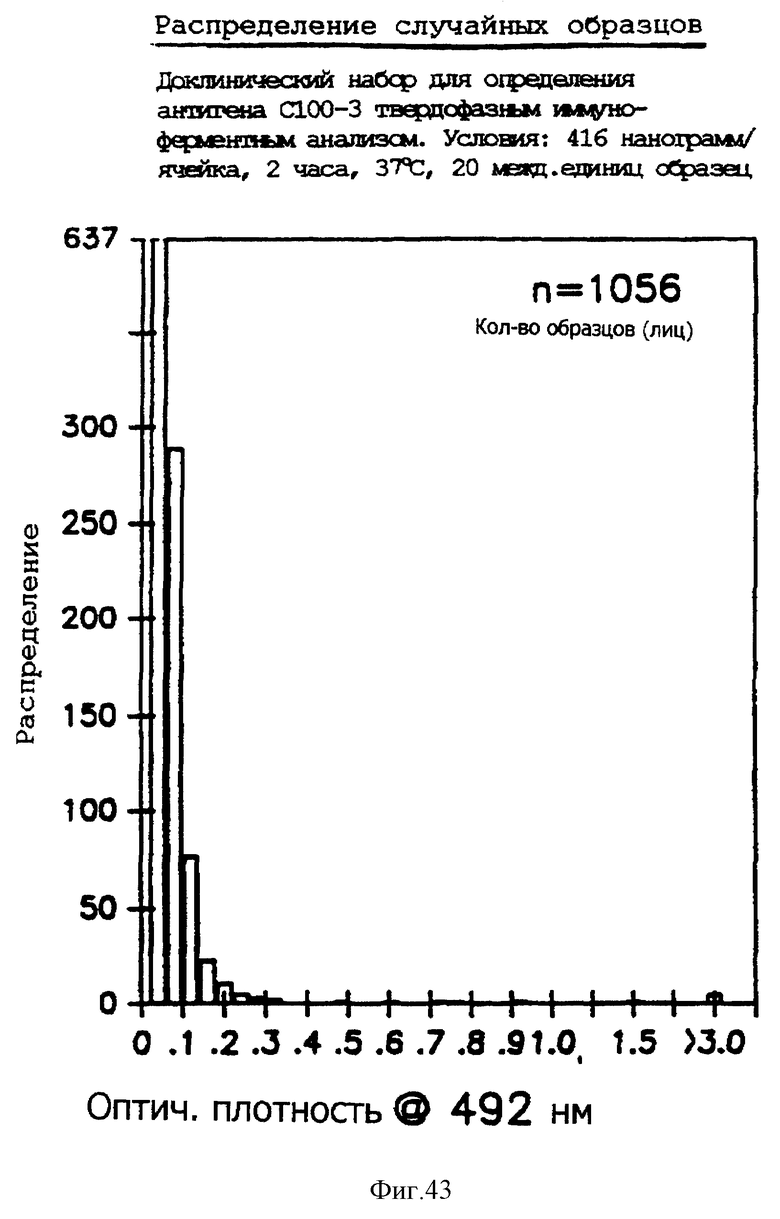
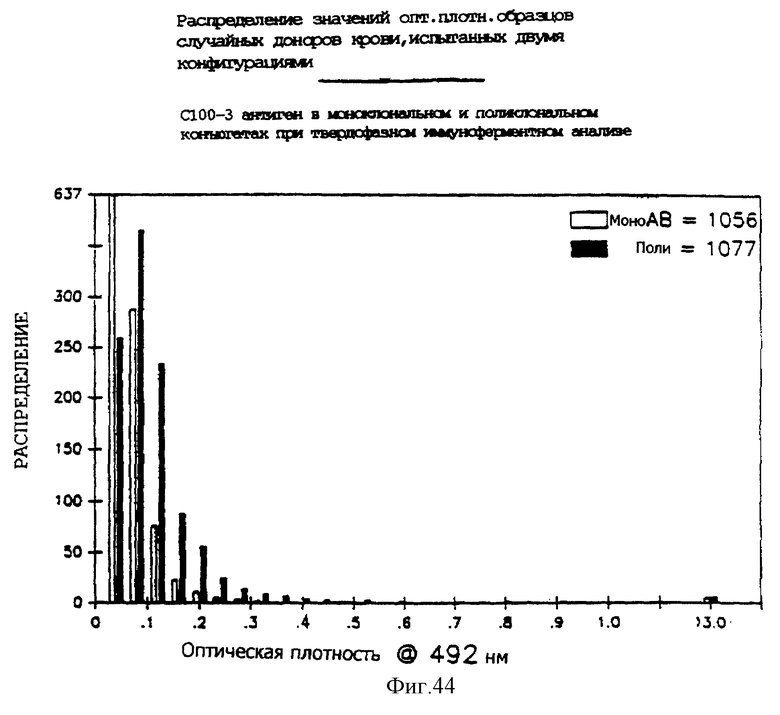











Изобретение относится к материалам и методам сдерживания распространения инфекции вируса гепатита ни А, ни В (вируса гепатита С). Предложены полипептиды, обладающие антигенными свойствами вируса гепатита С и их фрагменты. Предложен диагностический реагент для обнаружения антител против вируса гепатита С, набор для обнаружения антител против вируса гепатита С, включающий диагностический реагент, и способ обнаружения антител против вируса гепатита С. Изобретение позволяет получить надежные диагностические и прогностические средства для определения нуклеиновых кислот, антигенов и антител, связанных с вирусом гепатита С. 17 с. и 13 з.п. ф-лы, 47 ил., 19 табл.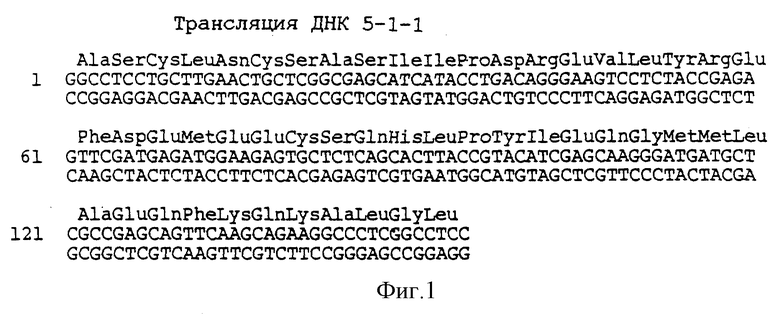
Приоритет по пунктам:
18.11.87 - по пп.6-9, 16, 17, 22, 26, 27;
06.05.88 - по пп.10-13, 18, 19, 20, 23, 28, 29;
18.11.88 - по пп.1-5, 14, 15, 21, 24, 25.
| УСТРОЙСТВО для АВТОМАТИЧЕСКОЙ УСТАНОВКИ ЭКСПОЗИЦИИ в ФОТОАППАРАТЕ | 0 |
|
SU218474A1 |
| ЦИЛИНДРИЧЕСКИЙ СОСУД ДЛЯ ВЫСОКИХ ВНУТРЕННИХ ДАВЛЕНИЙ | 1934 |
|
SU38642A1 |
| РАБОЧИЙ ОРГАН РОТОРНОГО ЭКСКАВАТОРА | 0 |
|
SU279460A1 |

