Изобретение относится к векторам доставки генов, содержащим молекулы нуклеиновых кислот, кодирующих индуцирующие апоптоз белки VP2 и/или апоптин (VP3) со схожей активностью.
Изобретение также относится к противоопухолевой терапии и к диагностике рака. Инфицирование клеток различных опухолей человека векторами доставки генов по изобретению приведет, в результате, к индукции апоптоза опухолевых клеток и значительно уменьшенному апоптозу, если не к полному его отсутствию, нормальных диплоидных нетрансформированных/незлокачественных клеток.
Экспрессия белка апоптина (VP3), происходящего от вируса анемии кур (CAV), in vitro в трансформированных куриных клетках индуцирует апоптоз (Noteborn et al., 1994, Noteborn and Koch, 1995). Апоптоз характеризуется сжатием клеток, сегментацией ядра, конденсацией и расщеплением ДНК на фрагменты, приблизительно соответствующие размеру домена, с последующей, в большинстве клеток, межнуклеосомной деградацией. Наконец, апоптозные клетки распадаются на заключенные в оболочку апоптозные тельца, которые быстро фагоцитируются соседними клетками. Следовательно, апоптоз вызывает значительно меньшее разрушение ткани, чем некроз, представляя нефизиологический тип гибели клеток (Wyllie et аl., 1980, Arenas and Wyllie, 1991, и White, 1996).
Апоптин представляет собой недлинный белок протяженностью только в 121 аминокислоту, являющийся скорее основанием, с большим содержанием пролина, серина и треонина (Noteborn et аl., 1991). В анализированных трансформированных куриных клетках, которые все претерпевают индуцированный апоптином апоптоз, апоптин локализуется строго в пределах ядра клетки, процессирование С-концевого основного участка апоптина приводит к пониженной локализации в ядре и существенно пониженной апоптозной активности (Noteborn et al., 1994).
Апоптин и другие белки с апоптиноподобной активностью также могут индуцировать апоптоз в линиях человеческих злокачественных и трансформированных клеток, но не в линиях нетрансформированных клеток человека. Авторы установили, что индуцированный апоптином апоптоз имеет место в отсутствие функционального р53 (Zhuang et al., 1995 a) и не может блокироваться Bcl-2, BCR-ABL (Zhuang et al., 1995), связывающим Bcl-2 белком BAG-1 и белком вируса коровьей оспы CrmA (Noteborn, 1996). In vitro апоптин не в состоянии индуцировать запрограммированную гибель клеток в нормальных лимфоидных, дермальных, эпидермальных, эндотелиальных и гладкомышечных клетках. Однако когда нормальные клетки становятся трансформированными, они становятся восприимчивыми к апоптозу с участием апоптина или других белков с апоптиноподобной активностью. Длительная экспрессия апоптина в нормальных человеческих фибробластах показывает, что апоптин не обладает токсической или трансформирующей активностью в этих клетках, в нормальных клетках апоптин обнаружен преимущественно в цитоплазме, в то время как в трансформированных или злокачественных клетках, т.е., характеризующихся гиперплазией, метаплазией или дисплазией, он локализуется в ядре, что наводит на мысль, что локализация апоптина связана с его активностью (Danen-Van Oorschot et al., 1997, Noteborn, 1996).
Апоптоз является запрограммированным физиологическим процессом для элиминации избыточных, изменившихся или злокачественных клеток (Earnshaw, 1995). Процесс апоптоза может быть инициирован различными регуляторными стимулами (Wyllie, 1995, и White, 1996). Изменения уровня выживаемости клеток играют важную роль в патогенезе человека, например, при развитии рака, вызванного усиленной пролиферацией клеток, но также пониженной гибелью клеток (Кеrr et al., 1994). Показано, что различные химиотерапевтические соединения и облучение индуцируют апоптоз в опухолевых клетках во многих случаях через дикий тип р53 (Thompson, 1995, Bellamy et al., 1995, Steller, 1995).
Однако многие опухоли приобретают мутацию в р53 во время своего развития, часто коррелирующую с плохой реакцией на противораковую терапию (Hooper, 1994). В случае некоторых (лейкозных) опухолей высокий уровень экспрессии протоонкогена Всl-2 ассоциируется с сильным сопротивлением различным индуцирующим апоптоз химиотерапевтическим средствам (Hockenberry, 1994, Kerr et al., 1994, и Sachs and Lotem, 1993).
Следовательно, апоптин может стать потенциальным кандидатом на роль средства для разрушения опухолевых клеток или других клеток, характеризующихся гиперплазией, метаплзией или дисплазией, которые стали устойчивыми к (химио)терапевтической индукции апоптоза из-за потери функционального р53 и повышенной экспрессии Всl-2 и других нарушающих апоптоз факторов. Тот факт, что апоптин не индуцирует апоптоз в нормальных нетрансформированных человеческих клетках, по меньшей мере in vitro, наводит на мысль, что токсическое действие обработки апоптином in vivo должно быть весьма слабым.
Однако до сих пор экспрессия апоптина в опухолевых клетках осуществляется путем использования временной трансфекции клеток культур тканей. Недостатком такого способа экспрессии является очень низкий процент клеток, которые могут экспрессировать апоптин в условиях in vitro. Используемые способы трансфекции in vivo будут громоздкими и неэффективными, если вообще будут возможны, и не внесут никакого вклада в эффективное лечение рака.
Можно получить аденовирус из аденовирусов человека (Ads), которые являются безоболочными двадцатигранными ДНК-вирусами. Геном состоит из линейной двухцепочечной молекулы ДНК примерно в 36 кб, содержащей инвертированные концевые повторы (Horvitz, 1990). Серотипы, использованные для развития вектора (Ad2 и Ad5), не ассоциируются с тяжелой патологией человека (Horvitz, 1990). Вирус исключительно эффективен при введении его ДНК в клетку-хозяина. Ads могут инфицировать множество делящихся и неделящихся клеток широкого ряда видов, и вирус можно получить в больших количествах с относительной легкостью. В противоположность ретровирусам, Ads не интегрируют в геном клетки-хозяина. Все используемые в настоящее время Ads имеют делецию в области Е1, куда можно ввести новую ДНК. В результате делеции Е1 рекомбинантный вирус становится дефектным по репликации (Stratford-Perricaudet and Perricaudet, 1991). С одной стороны, это обеспечивает существенное увеличение безопасности: rAdV не может реплицироваться в клетках человека в отсутствие белков Е1А. Таким образом, rAdV может доставить свою генетическую информацию в клетку человека в отсутствие литической или продуктивной инфекции. С другой стороны, появляется проблема получения этих векторов. Однако для функций Е1 нет необходимости в кодировании самим вектором. Они могут обеспечиться in trans специальными хелперными клетками, которые экспрессируют гены Е1. После инфицирования или трансфекции этих хелперных клеток Ad-вектором с делецией Е1 клеточные белки Е1 будут комплементировать репликацию rAdV, что приведет к продуцированию потомства rAdV. Ad-хелперные клетки должны быть человеческого происхождения и они должны содержать и экспрессировать область AdE1, т.е., Ad-трансформированные человеческие клетки, такие как клеточная линия 293 (Graham and Prevec, 1991), клеточная линия 911 (Fallaux et al., 1996) и клеточная линия PER.C6 (Fallaux, 1996).
Данное изобретение относится к вектору доставки генов, который обеспечивает возможность использования особенностей противоопухолевого средства апоптина или других белков с апоптиноподобной активностью для лечения рака с помощью применения генной терапии или для лечения злокачественных перерождений, характеризующихся гиперплазией, метаплазией или дисплазией. Такой вектор доставки генов, который является независимым инфекционным вектором, может представлять, например, вирус или липосому, или полимер, или нечто подобное, что само по себе может инфицировать или любым другим способом доставлять генетическую информацию, например, к опухолевым клеткам, которые можно обработать. Генетическая информация включает молекулу нуклеиновой кислоты, кодирующую апоптиноподобную активность. Изобретение также относится к вектору доставки генов, у которого существенно возросла способность экспрессировать апоптиноподобную активность. Неожиданно было обнаружено, что изменение некодирующих нуклеотидных последовательностей в обратном направлении, расположенных в пределах сайта инициации трансляции, которые предшествуют последовательностям, кодирующим апоптиноподобный белок, существенно увеличивает экспрессию указанного белка в опухолевых клетках. Изобретение также относится к вектору доставки генов, содержащему нуклеиновую кислоту, кодирующую VP-2-подобную активность. Неожиданно было показано, что VP-2-подобная активность действует синергитически с апоптиноподобной активностью в отношении индукции апоптоза в опухолевых клетках, VP-2-подобный белок сам также может действовать синергитически или аддитивно, например, в случае (химио) терапевтической индукции апоптоза. Изобретение также относится к вектору доставки генов, содержащему нуклеиновую кислоту, кодирующую VP-2-подобную активность, в дополнение к молекуле нуклеиновой кислоты, кодирующей апоптиноподобную активность. По изобретению, например, вектор доставки генов представляет собой вирус. Кроме того, изобретение относится к вектору доставки генов, который сам является дефектным по репликации вирусом, но который может реплицироваться в хелперных или упаковывающих клетках для получения самого вектора доставки генов. Так, вектор доставки генов по изобретению может представлять собой, например, аденовирус или ретровирус, или любые другие ДНК-овые или РНК-овые рекомбинантные вирусы, которые можно использовать в качестве вектора или плазмовируса. Кроме того, изобретение относится к вектору доставки генов, который дополнительно снабжен специфическим лигандом или молекулой-мишенью или молекулами-мишенями, с помощью которых вектор доставки генов может специфически направляться для доставки своей генетической информации к выбранной клетке-мишени. Такая молекула-мишень может представлять собой, например, вирусный спайковый белок или рецепторную молекулу или антитело, реакционно способное в отношении поверхностного рецептора или белка опухолевой клетки.
Изобретение также относится к вектору доставки генов, который можно использовать при диагностике, например, рака. Такой вектор доставки генов может использоваться для диагностики in vitro, когда у человека или животного берутся образцы тканей или клеток или биоптаты. Такие образцы затем оцениваются или испытываются путем их инфицирования, в культуре или непосредственно, указанным вектором доставки генов, способным к экспрессии, т.е. с апоптиноподобной активностью. Только трансформированные клетки или клетки, показывающие различные стадии гиперплазии, дисплазии или метаплазии, или опухолевые или раковые клетки экспрессируют белок с апоптиноподобной активностью в пределах ядра. Присутствие указанного белка можно продемонстрировать классическими (иммуно) гистохиммческими методами, т.е., методами световой микроскопии или методами автоматизированной клеточной сортировки. С другой стороны, указанные выше инфицированные клетки характеризуются апоптозом, и, таким образом, их можно выявить по известным характеристикам апоптоза.
Изобретение также относится к или описывает все стадии, необходимые для построения рекомбинантного дефектного по репликации аденовируса, эксперссирующего индуцирующий апоптоз фактор апоптин. Высокие титры рекомбинантного, содержащего апоптин, аденовируса можно получить с помощью линий аденовирусных упаковывающих клеток, таких как 293, 911 и PER.C6. Отрицательного воздействия апоптина на всех необходимых стадиях репликации аденовируса и других событий цикла жизни аденовируса в условиях выращивания клеток обнаружено не было.
Кроме того, в изобретении описывается построение контрольного рекомбинантного аденовируса, который содержит все последовательности, какие и у рекомбинантного, содержащего апоптин, аденовируса, но, из-за ориентации 3’-5’ кодирующей апоптин последовательности, под контролем регулирующих промоторных элементов, неспособного к экспрессии апоптина. С помощью этого контрольного аденовирусного вектора можно исследовать специфическое действие экспрессии апоптина рекомбинантным аденовирусом.
Рекомбинантный дефектный по репликации аденовирус экспрессирует апоптин в больших количествах в различных опухолевых и/или трансформированных клетках, результатом чего является индукция апоптоза. В противоположность этому, экспрессия апоптина при помощи рекомбинантного аденовируса в нормальных нетрансформированных человеческих клетках не приводит в результате к индукции индуцируемого апоптином апоптоза.
в частности, изобретение относится к противоопухолевой терапии. Лечение опухолевой клетки (клеток) будет происходить путем экспрессии апоптина при помощи инфицирования (опухолевых) клеток векторами доставки генов, такими как аденовирусные векторы, которые содержат кодирующую последовательность для белка с апоптиноподобной активностью. Следовательно, изобретение относится к векторам доставки генов, таким как аденовирус, эксперсиирующий апоптин, который является весьма мощным противоопухолевым средством. Аденовирусная регуляция апоптина не индуцирует апоптоз, или по меньшей мере заметно не индуцирует апоптоз, в нормальных клетках, что показывает, что токсичность обработки in vivo рекомбинантным, содержащим апоптин аденовирусом будет низкой. С помощью инфицирования рекомбинатным, содержащим апоптин аденовирусом значительно большее количество (опухолевых) клеток будет экспрессировать апоптин. Это открытие является основным улучшением в отношении экспрессии апоптина по сравнению с трансфекциями ДНК.
Изобретение также относится к построению элемента экспрессии VP2 без синтеза апоптина и/или части апоптина. Кроме того, авторы доказали, что экспрессия белка VP2 вируса анемии кур (CAV) усиливает индуцируемый апоптином апоптоз в клетках опухолей человека. Точнее, VP2 и апоптин действуют синергитически в отношении индукции апоптоза опухолевых клеток. Это открытие показывает, что коэкспрессия VP2 и апоптина даст, в результате, улучшение при лечении, основанном на апоптине.
В изобретении описывается существенное улучшение экспрессии апоптина посредством изменения направления его прямых последовательностей в противоположном направлении от инициирующего кодона ATG. Улучшение экспрессии не требует изменения аминокислот в апоптине, как это и предсказывает правило Козак. Улучшение последовательностей перед инициирующим ATG-кодоном других белков CAV также приведет в результате к усовершенствованию их синтеза.
Изобретение также относится к построению ретровирусных векторов, которые экспрессируют апоптин в клетках опухолей человека, результатом чего является индукция апоптоза. Данные по рекомбинантному, содержащему апоптин ретровирусу в сочетании с данными по рекомбинантному, содержащему апоптин аденовирусу показывают, что экспрессия апоптина не является токсическим фактором для репликации ДНК-овых и РНК-овых вирусов.
Экспрессия апоптина в (опухолевых) клетках также может происходить при инфицировании клеток другими ДНК-овыми и/или РНК-овыми вирусными векторами, кроме аденовирусных и ретровирусных векторов, содержащими кодирующую последовательность для апоптина. Кроме того, для индукции индуцируемого апоптином апоптоза опухолевых клеток можно использовать полученные на основе вирусов векторные системы, такие как плазмовирусы.
Изобретение будет поясняться подробнее с помощью описанной далее экспериментальной части. Это делается только с целью пояснения и не должно интерпретироваться как ограничение объема притязаний изобретения.
Экспериментальная часть
Клетки и условия их культивирования
Трансформированные E1 Ad5 линии клеток первичной почки человека (НЕК; 293) и первичной сетчатки человека (HER; 911 и PER. C6) выращивают в модифицированной по способу Дульбекко среде Игла (DMEM) с добавлением 10% фетальной телячьей сыворотки (FCS) в атмосфере с 5% СО2 при 37°С. Клеточная линия 293 получена из Американской коллекции типовых культур (АТСС CRL 1573). Клеточные линии 911 и PER. C6 получены по Fallaux et al. (1996). Среды для выращивания клеток, реагенты и сыворотки получены от GIBCO Laboratories (Гранд Исланд, Нью-Йорк). Культуральные планшеты получены от Greiner (Нюртинген, Германия).
Эпидермальные керациты человека извлекают из крайней плоти и выращивают в присутствии слоя мышиных фибробластов 3Т3, облученных летальной дозой 137-Сs. Первичные культуры кератиноцитов (FSK-1) инициируют в полной среде так, как описано ранее (Rheinwald and Green, 1975), с незначительными изменениями.
Онкогенные кератиноциты SCC-15 (Rheinwald and Beckett, 1981), полученные от плоскоклеточной карциномы, культивируют в смешанной среде DMEM и F12 (3:1), содержащей 5% фетальной телячьей сыворотки, 0,4 мкг гидрокортизона и 1 мкМ изопротеренола. Клетки HepG2, полученные от гепатомы человека (Aden et al., 1979), и U20S и Saos-2, полученные от остеосаркомы человека (Diller et al., 1990), выращивают в DMEM (GIBCO/BRL) с добавлением 10% фетальной телячьей сыворотки. Штамм спонтанно трансформированных кератиноцитов НаСаТ (Boukamp et al., 1988) подарен д-ром R. Fusenig, DKFZ, Гейдельберг, Германия. Клетки НаСаТ выращивают в DMEM с добавлением 10% фетальной телячьей сыворотки.
Муриновые клеточные линии выращивают в модифицированной по способу Дульбекко среде Игла с высоким содержанием глюкозы (4,5 г на литр) и 10% фетальной телячьей сыворотки в атмосфере с 5% СО2 при 37°С. Линия экоторопных упаковывающих клеток Psi-2 (Mann et al., 1983) и линия амфотропных упаковывающих клеток РА317 описаны ранее (Miller, 1990 a, b).
Методы работы с вирусами
Анализы бляшкообразования выполняются так, как описано ранее (Fallaux et al., 1996). Коротко, штаммы аденовирусов серийно разводят в 2 мл DMEM, содержащей 2% лошадиной сыворотки, и добавляют к почти слившимся клеткам 911 в 6-луночных планшетах. После инкубации в течение 2 час при 37°С среду заменяют минимальной поддерживающей средой F-15 (MEM), содержащей 0,85% агарозы (Sigma, США), 20 мM HEPES (рН 7,4), 12,3 мМ MgCl2, 0,0025% L-глутамина и 2% лошадиной сыворотки (теплоинактивированной при 56°С в течение 30 минут).
Получение небольших количеств аденовируса осуществляют так, как описано ранее (Fallaux et al., 1996). Коротко, монослои почти слившихся клеток 911 или PER. C6 инфицируют приблизительно 5 бляшкообразующими единицами (бое) на клетку в PBS, содержащем 1% лошадиной сыворотки. После инкубации в течение 1 часа при комнатной температуре инокулят заменяют свежей средой (DMEM/2% лошадиной сыворотки). Через 48 часов почти полностью отслоившиеся клетки собирают и помещают в 1 мл PBS, содержащего 1% лошадиной сыворотки. Вирус выделяют из клеток-продуцентов посредством 3 циклов быстрого замораживания и оттаивания. Лизаты очищают центрифугированием при 3000 об/мин в течение 10 минут и хранят при -20°С.
Штаммы rAdV, продуцированные 911 и PER. C6, скринируют на присутствие рекомбинат-компетентного аденовируса с помощью PCR с праймерами, полученными для области itr Ad5 (5_-GGGTGGAGTTTGTGACGTG-3_) И области, кодирующей Е1А (5_-TCGTGAAGGGTAGGTGGTTC-3_), как описано в Noteborn and De Boer (1995), с использованием аппарата для PCR Perkin Elmer. Присутствие амплифицированного фрагмента в 600 п.о. указывает, что в анализируемом штамме вируса имеется репликационно-компетентный (содержащий область Е1) аденовирус (Hoebon, неопубликованные результаты), или (это подтверждается) серией опытов с инфицированием клеток HepG2 rAdV. В течение по меньшей мере 10 дней клетки проверяются на возможное цитопатогенное действие и с помощью метода непрямой иммунофлуоресценции с использованием специфической моноклональной антисыворотки к белку Е1А.
Плазмиды и ДНК-трансфекции
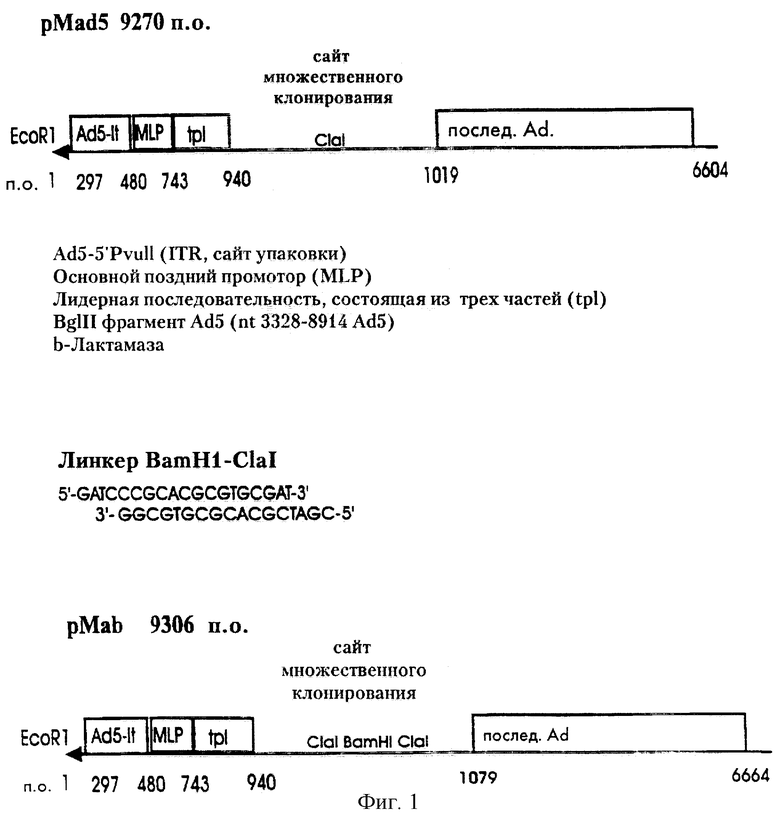
Адапторная плазмида pMad5 была сконструирована из pMLP10 (Levrerno et al., 1991) так, как описано ниже. Плазмиду pMLP10-lin получают из pMLP10 путем клонирования синтетического ДНК-фрагмента с уникальными сайтами для рестрикционных эндонуклеаз MluI, SplI, SnaBI, SpI, AsuII и MunI в сайт HindIII pMLP10. BglII фрагмент аденовируса от nt 3328 до 8914 генома Ad5 встраивают в сайт MunI pMLP-lin. Из образующейся в результате плазмиды удаляют SalI-BamHI фрагмент для инактивации гена устойчивости к тетрациклину. Полученная в результате плазмида была охарактеризована с помощью рестрикционного анализа и названа pMad5. Экспрессия генов, встроенных в сайт множественного клонирования, будет контролироваться основным поздним промотором аденовируса, который в этой конфигурации связан с энхансером немедленно-раннего гена 1 (E1) аденовируса.
Все ДНК-последовательности CAV первоначально получают из плазмиды pIc-20H/CAV-EcoRI (Noteborn and Boer, 1990). Экспрессирующая плазмида pCMV-fs, ранее названная pCMV-VP3 (Noteborn et al., 1994), содержит ДНК-последовательности CAV, кодирующие исключительно апоптин (nt 427-868).
Плазмида pCMV-VP2mu (Noteborn, неопубликованные результаты) содержит ДНК-последовательности CAV от 380 до 1512. Этот фрагмент ДНК CAV содержит кодирующую область для VP2, фланкированную 5’-некодирующей области участком в 25 п.о. и 3’-некодирующей области участком в 484 п.о. ДНК-последовательности CAV. На 106 п.о. в прямом направлении от стартового кодона для VP2 в другой рамке считывания располагается стартовый кодон для апоптина. Для предотвращения синтеза апоптина без изменения синтеза VP2 вводят мутацию в инициирующий кодон апоптина (ATG заменяется на ACG) и, кроме того, точечную мутацию в положении 549 (Т заменяется на А), в результате чего появляется дополнительный стоп-кодон в пределах гена, кодирующего апоптин. Поэтому клонирование последовательности CAV приведет к экспрессии только полноразмерного белка VP2. С помощью непрямой иммунофлуоресценции было показано, что VP2 может продуцироваться, но апоптин не синтезируется.
Для клонирования амплифицированных PCR ДНК-фрагментов авторы использовали плазмиду pCR-3.1, которую получили от InVitrogen (Карлсбад, Калифорния). Для создания рекомбинантного дефектного по репликации, содержащего апоптин ретровируса использовали ретровирусный вектор pLXSN (Miller, 1990 a, b).
Все стадии клонирования с плазмидными ДНК осуществляют, в принципе, методами, приведенными в Maniatis et al. (1992).
Все используемые ферменты получены от Boehringer Mannheim, Германия, и/или у BioLabs, США.
Плазмидную ДНК очищают путем центрифугирования в градиенте СsСl и колоночной хроматографией на сефакриле S500 (Pharmacia, Упсала, Швеция). Линии клеток человека НаСАТ, HepG2, SCC-15, 293, 911 и PER.C6 трансфецируют плазмидной ДНК методом осаждения с фосфатом кальция, как описано в Graham and Van der Eb (1973). Нормальные человеческие диплоидные кератиноциты (FSK-1; второй пассаж), клетки U20S и Saos-2 трансфецируют DOTAP (Fisher et al., 1996).
Анализ методом непрямой иммунофлуоресценции
Клетки фиксировали 80% ацетоном и использовали для иммунофлуоресцентных анализов со специфическими к CAV или специфическими к Е1А аденовируса моноклональными антителами и козьим антимышиным и/или козьим антикроличьим IgC, конъюгированными с флуоресцеином (Jackson Immunoresearch Laboratories Inc., Уэст-Гров, Пенсильвания), как описано в Noteborn et al. (1990). Ядерную ДНК окрашивали 2,4-диамино-2-фенилиндолом (DAPI) или пропидийиодидом (PI).
Результаты и обсуждение
Создание адапторного вектора pMab
Для того чтобы ввести сайт рестриктазы BamHI в адапторную плазмиду pMAd5, ее расщепляли ClaI и обрабатывали щелочной фосфатазой кишечника теленка. Линкер ClaI-BamHI обрабатывали Т4-киназой и лигировали с самим собой с использованием Т-4-ДНК-лигазы с последующим расщеплением ClaI. Линкер ClaI/BamHI/ClaI выделяли и лигировали с линейным вектором pMAd5. Бактериальный штамм JM109 трансформировали продуктами лигирования.
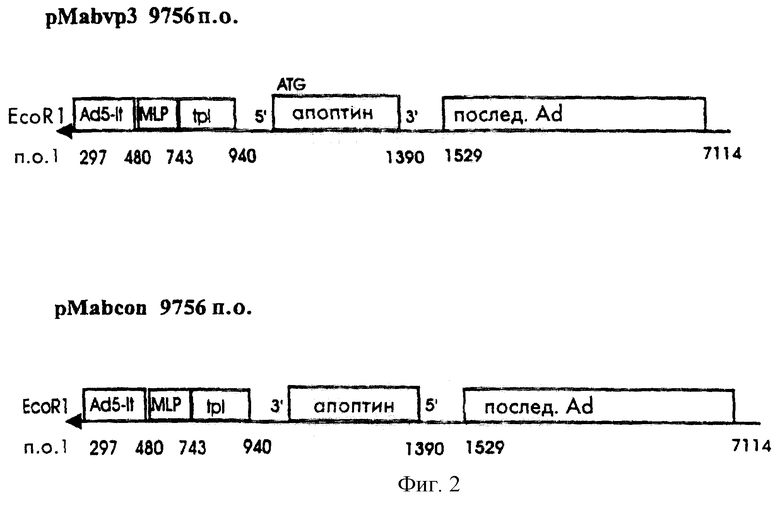
Полученный вектор рМАb характеризовали посредством расщепления рестрикционными ферментами. С помощью вектора рМАb чужеродные гены можно клонировать в уникальный сайт BamHI под контролем аденовирусного главного позднего промотора. Схема представление pMad5 и рМаb приведена на фиг.1.
Создание рекомбинантного, содержащего апоптин, и контрольного адапторного вектора
Для того чтобы создать адапторный вектор для введения гена апоптина в аденовирус, pMab обрабатывали BamHI, а затем фосфатазой кишечника теленка. Затем pCMV-fs обрабатывали BamHI и выделяли ДНК-фрагмент в 0,45 кб, содержащий кодирующие апоптин последовательности. Фрагмент ДНК апоптина лигировали в сайт BamHI линейного адапторного вектора pMab. Продукты лигирования клонировали в бактериальный штамм JM109. Ориентацию апоптина в pMab определяли с помощью рестрикционного анализа.
Конструкция pMab, содержащая ген апоптина в ориентации 5’-3’, под контролем аденовирусного главного позднего промотора будет экспрессировать ген апоптина. Этот адапторный вектор обозначили pMAb-VP3 и использовали для генерации аденовирусного вектора, экспрессирующего апоптин. Плазмида ДНК pMab, содержащая кодирующие апоптин последовательности в ориентации 3’-5’ в прямом направлении от аденовирусного главного позднего промотора, не обеспечивает экспрессии апоптина, и была использована для создания контрольного рекомбинантного аденовирусного вектора. Схема этих адапторных векторов приведена на фиг.2.
Индукция апоптоза плазмидой CMV в сравнении с рекомбинантным, содержащим апоптин адапторным вектором, экспрессирующим апоптин
Сначала авторы проверили, на самом ли деле ДНК-вектор pMAb-VP3 способен экспрессировать апоптин в трансфецированных клетках, в то время как рМАb-соn не обладает этой способностью. Для этого человеческие клетки 293 и 911, трансформированные аденовирусом, трансфецировали pMAb-VP3, рМАb-соn и, в качестве положительного контроля, pCMV-VP3. Приблизительно через двое суток после трансфекции клетки фиксировали и проверяли на экспрессию апоптина методом непрямой иммунофлуресценции. Клеточные культуры, трансфецированные pCMV-VP3 и pMAb-VP3, содержали примерно 1% клеток, реагирующих с апоптинспецифическими моноклональными антителами, в то время как клеточные культуры, трансфецированные рМАb-соn, их не содержали. Эти результаты подтверждают, что pMAb-VP3 экспрессирует апоптин, а рМАb-соn, как и ожидалось, его не экспрессирует.
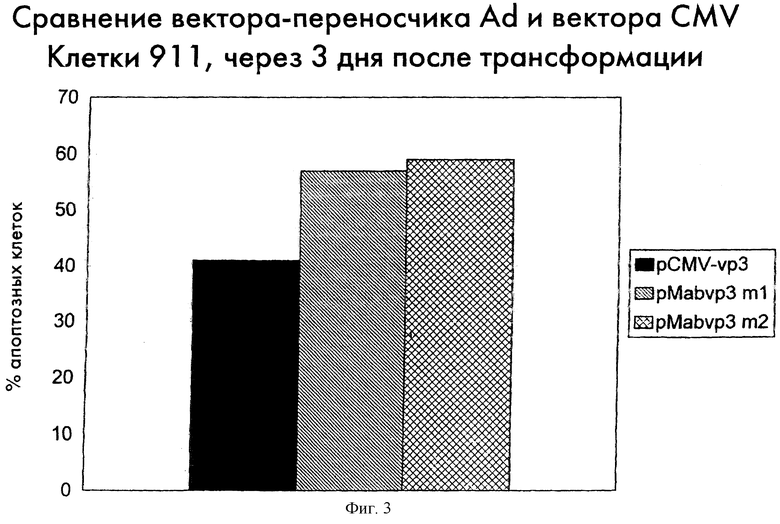
В другом эксперименте по трансфекции авторы анализировали индукцию апоптоза в клетках 911 после трансфекции pMAb-VP3 по сравнению с pCMV-VP3. Через три дня после трансфекции клетки 911 собирали и определяли экспрессию апоптина методом непрямой иммунофлуоресценции. Кроме этого, клетки окрашивали DAPI или PI; эти красители интенсивно окрашивают интактную ДНК, а апоптозную ДНК окрашивают слабо и/или неравномерно (Telford, 1992).
Приблизительно 60% апоптин-положительных клеток 911, трансфецированных pMAb-VP3, были апоптозными, в то время как из апоптин-положительных клеток 911, трансфецированных pCMV-VP3, апоптоз претерпевали примерно 40%. Эти результаты показывают, что экспрессия апоптина, регулируемая pMAb-VP3, приводит к такому же, а иногда - и к более высокому, уровню индукции апоптоза, по сравнению с экспрессией апоптина, регулируемой pCMV-VP3. Результаты приведены на фиг.3.
Более того, апоптин способен индуцировать апоптоз человеческих клеток, трансформированных аденовирусом, при этом Е1В не ингибирует апоптоз, индуцированный апоптином. В противоположность этому, Е1В способен блокировать апоптоз, индуцированный многими химиотерапевтическими средствами. Эти результаты показывают, что апоптин является очень сильным противоопухолевым средством.
Создание рекомбинантного аденовируса, содержащего апоптин
Рекомбинантные аденовирусные векторы, содержащие апоптин, получают посредством котрансфекции в хелперную клеточную линию 911 адапторных плазмид рДМb-VР3, несущих кодирующие последовательности для апоптина и также некоторые аденовирусные последовательности, и плазмиды ДНК JM17, содержащей полную ДНК аденовируса за исключением области Е1 и Е3 (McGrory et al., 1988). Котрансфекции проводили кальций-фосфатным методом согласно Graham and Van der Eb (19735. ДНК рекомбинантного аденовируса образуется путем гомологичной рекомбинации между гомологичными вирусными последовательностями, которые присутствуют в плазмиде pAMb-VP3, и аденовирусной ДНК из ДНК JМ17.
Подобным образом осуществляли котрансфекции клеток 911 с рМАb-соn и ДНК pJM17 для получения контрольного рекомбинантного аденовируса, который не может экспрессировать апоптин и который был использован в качестве контроля в случае апоптозных эффектов, индуцируемых апоптином.
Через несколько часов после трансфекции монослои клеток 911 покрывали верхним агарозным слоем и инкубировали при 37°С до появления отчетливо различимых бляшек, образованных рекомбинантным аденовирусом. Вирус собирали из бляшек в виде культуры, сохраняющейся в PBS с лошадиной сывороткой, как описано в Fallaux et al., (1996). Затем часть биомассы рекомбинантного вируса добавляли в 24-луночные планшеты, содержащие свежие клетки 911. Через несколько дней эти инфицированные клетки 911 лизировали и собирали рекомбинантные вирусы.
Затем исследовали экспрессию апоптина рекомбинантными, содержащими апоптин аденовирусами (rAd-VP3), или отсутствие такой экспрессии в случае контрольных рекомбинантных аденовирусов (rAd-con). Часть биомассы рекомбинантных вирусов, полученной из инфицированных 24-луночных планшет, использовали для инфицирования свежих клеток 911, которые выращивали в виде монослоев на покровных стеклах. На следующий день инфицированные клетки 911 фиксировали ацетоном и анализировали с помощью иммунофлуоресценции с использованием апоптинспецифических моноклональных антител 85.1. Пять из 5 анализируемых клеточных культур, инфицированных предполагаемым rAd-VP3, содержали клетки, экспрессирующие апоптин. Ни одна из культур клеток 911, инфицированных Ad-con, и неинфицированных клеток 911 не является апоптин-положительной.
Эти результаты показывают, что после котрансфекции линий аденовирусных упаковывающих клеток, таких как клетки 911, требуемым адаптером и аденовирусной ДНК можно получить жизнеспособный rAd-VP3, экспрессирующий апоптин.
Два штамма, полученных из бляшек rAd-VP3 или rAd-con, использовали для очистки rAd посредством осуществления трех последовательных пассажей бляшек на клетках 911 или, параллельно, в системе лимитирующих разведений на клетках PER.C6, как описано в Fallaux (1996).
На основе описанных выше способов, результатом которых является продуцирование rAd-VP3, экспрессирующего апоптин под контролем аденоверусного главного позднего промотора, авторы также получили аденовирусный вектор, экспрессирующий апоптин под контролем цитомегаловирусного (CMV) промотора. Эти результаты показывают, что можно получить различные типы рекомбинантных аденовирусов, которые контролируются одним из своих собственных или гетерологичных промоторных элементов.
Получение rAd-VP3 и rAd-con с использованием клеток PER.C6
Получение в небольшом масштабе партий rAd-VP3 и rAd-con с использованием клеток PER.C6 осуществляли так, как описано ранее (Fallaux, 1996). Коротко процедура описана в экспериментальном разделе.
С помощью анализа бляшкообразования определяются титры, составляющие приблизительно 1011-12 на мл осветленного лизата как для rAd-VP3, так и для rAd-con. Полученные титры не ниже тех, которые авторы наблюдали для других rAd.
С помощью PCR-анализа и инфицирования HepG2 rAd-VP3 и rAd-con исследовали, содержали ли полученные препараты вирусов комплементирующий репликацию аденовирус (см. также экспериментальный раздел). Как партии rAd-VP3, так и партии rAd-con не содержали RCA, что было подтверждено двумя методами.
Авторы приходят к заключению, что экспрессия апоптина не оказывает отрицательного влияния на все необходимые стадии цикла жизни аденовируса в условиях культивирования клеток. Следовательно, генная терапия на основе аденовирусных векторов, экспрессирующих апоптин, возможна.
Вследствие экспрессии антиапоптозных белков Ad5 El (White, 1996) апоптин оптимально индуцирует апоптоз после продуцирования рекомбинантного, содержащего апоптин аденовируса в больших количествах. Тот факт, что аденовирусный вектор, экспрессирующий апоптин, можно получить в человеческих клетках, трансформированных белками Е1 аденовируса типа 5 (Ad5), таких как клетки 293, 911 и PER.C6, показывает, что белок Е1 дает возможность этому ДНК-вирусу реплицироваться до высоких титров в присутствии индуцирующего апоптоз белка апоптина.
Эти результаты показывают, что возможно также сконструировать другие рекомбинантные ДНК-вирусные векторы, экспрессирующие апоптин в клеточных линиях, трансформированных белком Е1 аденовируса 5. Например, можно вырастить рекомбинантные парвовирусные векторы на основе парвовирусов Н-1 или MVM в клетках 293Т, трансформированных белком Е1 Ad5 (Dinsart et al., 1996).
Парвовирусы Н-1 или MVM специфически индуцируют гибель клеток в трансформированных клетках, но не во всех (Lopez-Guerrero, 1997). Парвовирусные векторы, экспрессирующие апоптин, будут более сильными при индукции опухолеспецифического апоптоза, чем парвовирус как таковой, из-за дополнительной опухолеспецифической индукции апоптоза апоптином (Dinsart et al., 1996, Danen-Van Oorschot, 1997).
Схема получения рекомбинантных, содержащих апоптин вирусных векторов на основе трансформированных белком Е1 Ad5 клеток также справедлива для видов РНК-вирусов, таких как ретровирусы.
Индукция апоптоза в линиях человеческих трансформированных и/или злокачественных клеток
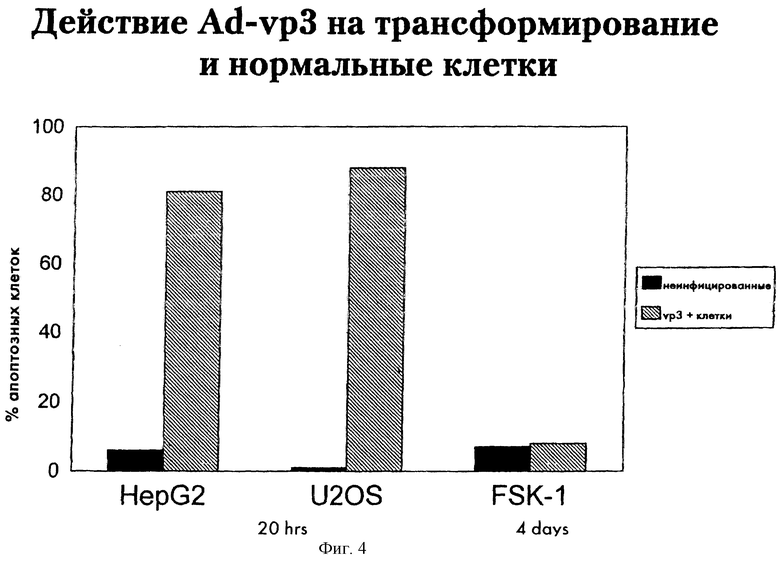
Авторы исследовали, приведет ли инфицирование человеческих опухолевых клеток rAd-VP3 к апоптозу, индуцированному апоптином. для этого клетки гепатомы человека HepG2, остеосаркомы U20S, клетки SCC-15, полученные из плоскоклеточной карциномы, и клетки из спонтанно трансформированной линии кератиноцитов НаСаТ инфицировались rAd-VP3. Через день после трансфекции клетки фиксировали и, при помощи иммунофлуоресценции и окрашивания DAPI, клетки исследовали на синтез апоптина и возникновение апотоза. Уже на другой день после инфицирования почти все проанализированные апоптин-положительные клетки опухолей человека были апоптозными. В неинфицированных культурах только небольшой процент клеток опухолей человека оказался апоптозным. Результаты для клеток HepG2 и U20S приведены на фиг.4.
Эти результаты демонстрируют, что апоптин, экспрессированный rAd-VP3, может индуцировать апоптоз в различных онкогенных и/или трансформированных клеточных линиях млекопитающего.
Экспрессия апоптина в нормальных клетках, инифицированных rAd-VP3
Для того чтобы проанализировать действие апоптина, экспрессированного rAd-VP3, в неинфицированных нормальных нетрансформированных клетках, rAd-VP3 инфицировали клетки FSK-1. Через четыре дня после трансфекции клетки анализируют при помощи непрямой иммунофлуоресценции с использованием моноклональных антител 85.1 и окрашивания DAPI. Самое большее 8% апоптин-положительных клеток показывают окрашивание DAPI с отклонением от нормы, что указывает, что они могли претерпеть индуцированный (апоптином) апоптоз. Однако 7% клеток, которые не были инфицированы, также имеют картину аберрантного окрашивания ДНК DAPI. Результаты приведены на фиг.4.
Данные о гепатотропном характере человеческого Ad5 после системной доставки также являются важными для исследования действия апоптина в нормальных диплоидных гепатоцитах. Для их получения выделенные гепатоциты крысы выращивали в среде Е Вильямса (Gibco/Life Technologies, Гран-Исланд, Нью-Йорк, США) с добавлением инсулина (2 мЕ/мл) и дексаметазона (1 нМ). Клетки выращивали на сенсибилизированных коллагеном предметных стеклах (Micronic).
Первичные крысиные гепатоциты инфицировали вектором на основе аденовируса Ad-VP3, экспрессирующим апоптин, контрольным аденовирусом, экспрессирующим LacZ, или псевдоинифицируют. Через два дня клетки фиксировали и, при помощи иммунофлуоресценции и окрашивания DAPI, проверяли процент апоптозных клеток. Не наблюдали различия в проценте погибших клеток между клетками, инфицированными для экспрессии апоптина, lacZ, или псевдоинфицированными. Эти наблюдения показывают, что (крысиные) гепатоциты не претерпевают индуцированного апоптином апоптоза, также и в том случае, когда апоптин синтезируется аденовирусным вектором.
Эти результаты показывают, что направляемая rAd-vp3 экспрессия апоптина не приводит к индуцируемому апоптином апоптозу в нормальных нетрансформированных человеческих клетках, в отличие от трансформированных или онкогенных человеческих клеток.
Увеличение синтеза апоптина
Для того чтобы исследовать действие прямых последовательностей перед инициирующим ATG-кодоном апоптина, авторы создали две конструкции pCR-3.1-апоптин. pCR-VP3ori содержит исходные прямые последовательности в обратном направлении (5_-ТТТСАА-3_) от ATG-кодона, в то время как другая конструкция pCR-VP3mu содержит прямую последовательность в обратном направлении 5_-GCCAAC-3_. С помощью бесклеточной системы (in vitro) транскрипции/трансляции из зародышей пшеницы было показано, что экспрессия апоптина pCR-VP3mu по меньшей мере в пять раз превышает экспрессию, наблюдаемую в случае pCR-Vp3ori. Эти данные показывают, что характер последовательностей непосредственно перед ATG-кодоном апоптина влияет на синтез апоптина. Создание (вирусных) векторов с прямой, в обратном направлении, последовательностью 5_-GCCAAC-3_ перед ATG-кодоном апоптина приведет к более высокому продуцированию апоптина и, косвенно, к повышенному индуцируемому апоптином апоптозу.
Важно также упомянуть, что аминокислотная последовательность апоптина не изменяется так, как это необходимо для повышенной эффективности трансляции согласно "правилу Козак" (Caventer and Stuart, 1991). В соответствии с этим правилом, авторы должны были бы заменить нуклеотид в позиции +4 с А на G, в результате чего должна была измениться вторая аминокислота апоптина, которая возможно могла изменить его активность.
Идентификация существенного фрагмента апоптина, содержащего апоптозную активность
Для того чтобы исследовать, является ли часть белка апоптина существенной для его апоптозной активности, была сконструирована плазмида, кодирующая химерные белки, состоящие из белка зеленой флуресценции (GFP; Rizzuto, 1995) и N-концевых 71 аминокислот апоптина (N-апоптин) или его с-концевых 50 аминокислот (С-апоптин). Человеческие трансформированные клетки, такие как клетки Saos-2 (Zhuang, 1995), трансфецировали плазмидами, экспрессирующими химерный GFP/N-апоптин или GFP/C-апоптин. Апоптоз претерпевают только клетки Saos-2, экспрессирующие GFP/C-апоптин. Это совпадает с тем фактом, что С-апоптин (связанный с GFP), может входить в ядро.
Эти результаты показывают, что для индукции апоптоза в (человеческих) онкогенных или трансформированных клетках также может быть достаточно части апоптина. Следовательно, можно также разработать эффективную (генную) терапию, основанную на (вирусных) векторах, экспрессирующих только часть апоптина. Кроме того, эти данные демонстрируют, что часть апоптина содержит его апоптозную активность, когда она ковалентно связана с чужеродным белком.
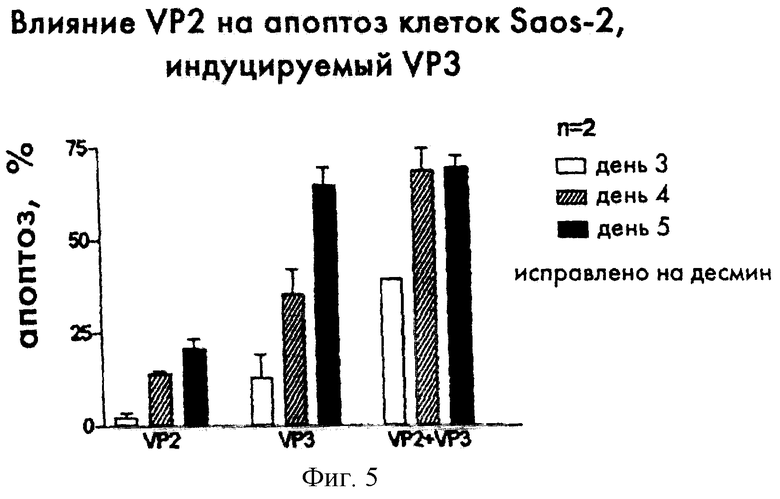
Коэкспрессия VP2 и апоптина в клетках опухолей человека синергитически увеличивает индукцию апоптоза
Для того чтобы исследовать влияние коэкспрессии VP2 и апоптина на индукцию апоптоза, клетки Saos-2 (ко)трансфецируют pCMV-fs, экспрессирующей апоптин, и/или pCMV-VP2mu, экспрессирующей VP2. Клетки фиксировали ацетоном через разные промежутки времени после трансфекции. Методом непрямой иммунофлуоресценции с моноклональными антителами CVI-CAV-111.1 (Noteborn and Koch, 1996) определяли УР2-положительные клетки, а с моноклональными антителами CVI-CAV-85.1 - апоптин-положительные клетки. На 3 день после трансфекции претерпевали апоптоз только примерно 3% экспрессирующих VP2 клеток и только примерно 10% клеток, экспрессирующих апоптин. В противоположность этому, примерно 40% клеток Saos-2, экспрессирующих как VP2, так и апоптин, уже были апоптозными. Также через 4 дня после трансфекции процент УР2/апоптин-положительных клеток, которые претерпели апоптоз, существенно выше, чем для клеток, экспрессирующих один апоптин или один VP2.
Эти результаты, которые приведены на фиг.5, показывают, что VP2 усиливает индуцируемый апоптином апоптоз.
Конструирование и получение rAD-VP2
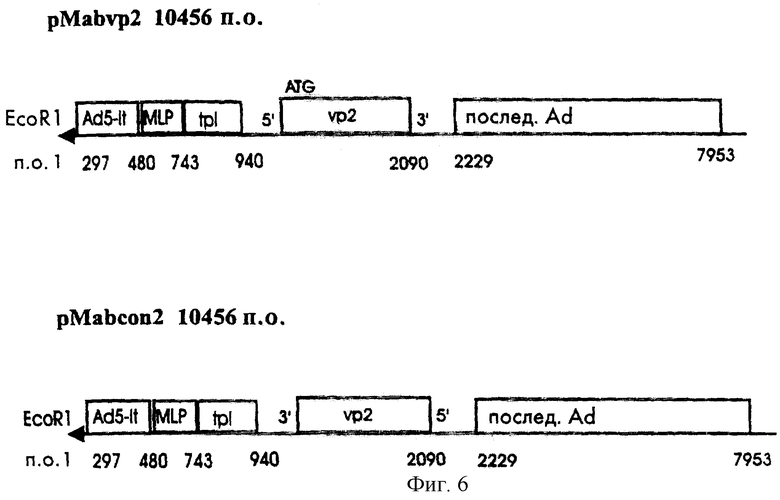
Для того чтобы сконструировать рекомбинантный аденовирус, экспрессирующий вирусный белок 2 (VP2) вируса анемии кур, была получена адапторная плазмида pMAb-VP2. Фрагмент BamHI в 1,1 кб выделяли из плазмиды pCMV-VP2mu, содержащей все кодирующие последовательности VP2, но с 2 точечными мутациями в пределах области, кодирующей апоптин (см. экспериментальный раздел), и лигировали с адапторным вектором рМАb, расщепленным BamHI и обработанным щелочной фосфатазой кишечника теленка. Полученную конструкцию pMAb-VP2, показанную на фиг.6, характеризовали с помощью рестрикционного анализа и анализа последовательностей.
rAD-VP2 создают посредством котрансфекции клеток 911 ДНК pMAb-VP2 и ДНК pJM17. Котрансфекцию и все другие требуемые стадии, необходимые для характеризации, очистки и получения rAD-VP2, осуществляли так же, как было описано в случае rAd-VP3 и rAd-con. Посредством непрямой иммунофлуоресценции с использованием моноклональных антител CVI-CAV-111.1 показали, что клетки 911 и PER. C6, инифицированные rAd-VP2, действительно могут экспрессировать белок VP2.
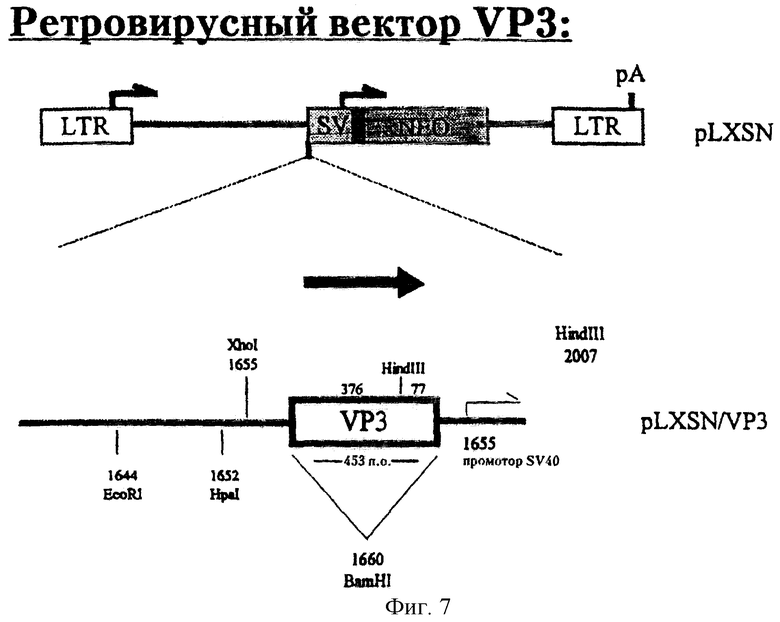
Создание ретровирусного вектора, экспрессирующего апоптин
Для того чтобы получить плазмиду pL-VP3-SN (см. фиг.7), фрагмент BamHi, несущий кодирующие апоптин последовательности, встраивали в уникальный сайт BamHI pLXSN. Правильную ориентацию вставки подтверждали рестрикционным анализом. Чтобы проверить целостность вставки, клетки COS-7 и HepG2 трансфецировали плазмидой pL-VP3-SN методом соосаждения с фосфатом кальция. Через четыре дня после трансфекции клетки фиксировали и анализировали с помощью моноклональных антител 85.1 на экспрессию белка апоптина. Экспрессия апоптина была обнаружена приблизительно в 1-2% клеток. Большинство клеток претерпевали апоптоз, что определяли с помощью окрашивания DAPI. Эти данные продемонстрировали, что провирусный LTR-промотор способен контролировать экспрессию белка апоптина, что его ген остается неповрежденным в ДНК-конструкции, и что в трансфецированных клетках HepG2 и COS-7 экспрессия апоптина индуцирует апоптоз.
Для того чтобы получить вирусы, плазмиду pL-VP3-SN трансфецировали в клетки Psi-2 и в клетки РА 317 методом соосаждения с фосфатом кальция. Супернатанты клеток собирали через сорок восемь часов после трансфекции, и их разведения использовали для инифицирования клеток HepG2 (супернатант РА 317) и клеток NIH3T3 (супернатант Psi-2) в присутствии 4 мкг/мл полибрена. Через четыре дня после инифицирования клетки фиксировали и анализировали на экспрессию апоптина с помощью моноклональных антител 85.1. Обнаружено, что приблизительно 1% клеток экспрессировали апоптин. Большинство апоптин-положительных клеток HepG2 были апоптозными. Эти данные подтверждают, что клетки были трансдуцированы ретровирусами, содержащими L-апоптин-SN. Кроме этого, полученные результаты демонстрируют, что одной копии провируса достаточно для экспрессии достаточного количества белка апоптина, которое можно обнаружить иммунофлуоресценцией, и этого количества достаточно для индукции апоптоза в клеточной линии опухоли человека, а именно клеточной линии гепатомы HepG2.
В совокупности эти данные показывают, что ретровирусные векторы, несущие ген апоптина, можно получать и можно использовать для индукции апоптоза в клетках опухолей человека. Это формально доказывает, что ни ген апоптина, ни его экспрессия не оказывают влияния на существенные стадии цикла жизни ретровируса. Они также показывают, что содержащие апоптин ретровирусы можно получать в количествах, достаточных для трансдукции клеток опухолей человека в культуре ткани.
Эти результаты также означают, что экспрессия апоптина и, следовательно, индуцируемый апоптином апоптоз (человеческих) опухолевых клеток также будет возможен с помощью векторных систем, полученных на основе (ретро)вирусов, таких как плазмовирусы (Noguiez-Hellin, 1996). Критический момент для такой системы на основе рекомбинатного, содержащего апоптин плазмовируса состоит в том, что не блокируется ли репликации ретровируса экспрессией апоптина. Авторы получили доказательства, что этого на самом деле не происходит в случае описанного выше рекомбинатного, содержащего апоптин ретровируса, что подразумевает успешное получение рекомбинатного, содержащего апоптин плазмовируса.
Диагностический анализ на раковые клетки на основе rAD-VPS
Клеточная локализация апоптина в онкогенных или трансформированных клетках иная по сравнению с локализацией в нормальных нетрансформированных клетках. Кроме того, другим показателем является специфическая способность апоптина индуцировать апоптоз в онкогенных или трансформированных клетках и не индуцировать апоптоз в нормальных клетках.
Посредством инфицирования клеток rAd-VP3 и анализа локации апоптина и/или индукции апоптоза в этих клетках можно проверить является ли клетка злокачественной или нет. Первичные клетки выделяют из (подозрительной) ткани и культивируют в требуемой среде. Клетки инфицируют rAd-VP3 и, параллельно, rAd-con, и затем анализируют. Например, применяют иммунофлуоресцентный анализ, основанный на моноклональных антителах 85.1, специфических для апоптина. Клетки контролируют на наличие апоптина в цитоплазме (нормальные клетки) или в ядре (трансформированные клетки). Кроме того, или вместо этого, можно оценить процент апоптозных клеток. Если процент апоптозных клеток в случае инифицированных rAd-VP3 клеток значительно выше, чем в случае клеток, инфицированных rAd-con, это означает, что первые клетки стали злокачественными.
Эксперименты по токсичности рекомбинатного, содержащего апоптин аденовируса на здоровых крысах
В условиях культивирования ткани апоптин, экспрессированный рекомбинантным аденовирусом rAd-VP3 в нормальных клетках, например, полученных от человека или грызуна, не индуцирует апоптоз. В описанном ниже эксперименте авторы исследовали, не приводит ли экспрессия апоптина с помощью рекомбинатного вирусного вектора rAd-VP3 у здоровых крыс к острой токсичности.
Оба вектора - используемый вектор rAd-VP3 и контрольный вектор rAd - выращивали на клетках РЕR.C6, и посредством PCR-анализа подтверждали, что препарат вируса не содержит аденовируса, комплементирующего репликацию (RCA). Векторы rAd очищали методом центрифугирования в градиенте СsСl.
Самцам крыс Wag/Rij (Harlan, Нидерланды) с массой тела примерно 200 граммов инъецировали рекомбинантный аденовирус, экспрессирующий апоптин (rAd-VP3; 2,5×109 бляшкообразующих единиц, бое) и контрольный рекомбинантный аденовирус, экспрессирующий генный продукт люциферазу (rAd-luc; 2,5×109). оба аденовирусных вектора ресуспендировали в PBS, содержащем 0,1% бычьего сывороточного альбумина и 10% глицерина (PBS+). Этот раствор без аденовирусного вектора также инъецировали крысам, и он служил в качестве дополнительного отрицательного контроля. rAd-VP3, rAd-luc или суспензии PBS+ вводили внутривенно, интраперитонеально или подкожно - двум крысам каждым способом.
Макроскопический патологический анализ крыс, обработанных rAd-VP3
Первым способом исследования возможного токсического действия экспрессированного rAd-VP3 апоптина является определение общего состояния здоровья и, в частности, массы тела обработанных крыс, которое проводят после инъекций ежедневно. В продолжение эксперимента у всех крыс наблюдалось хорошее состояние здоровья. Масса тела в различных группах существенно не различалась. Через 1 неделю все проверяемые крысы, включая инъецированных rAd-VP3, прибавили в весе, что указывает, что ни одно животное не страдало от острой токсичности вследствие одной из обработок rAd-VP3. Для дополнительного доказательства отсутствия острой токсичности осуществляли следующие определения. За два часа до умервщления всем крысам инъецируют BrdU. После умервщления некоторые ткани (печень, почки, кишки, сердце, легкое, селезенка, гонады и пенис) исследовали на патологию непосредственно и/или собирали также для гистологического анализа (см. ниже).
Макроскопический анализ показывает, что ни у одной из обработанных rAd-VP3 крыс нет органов с существенными паталогическими изменениями.
Определение ДНК Ad-VP3 в печени
Основной мишенью инъецированного внутривенно (рекомбинантного) аденовируса (вектора) является печень и, до некоторой степени, селезенка. Следовательно, любое токсическое действие апоптина будет наблюдаться в печени.
Проводят ряд экспериментов для исследования присутствия ДНК Ad-VP3, экспрессии апоптина Ad-VP3 и возможного цитопаталогического действия в печени.
Сначала методом Саузерн-блоттинга авторы исследовали содержит ли ДНК, выделенная в день умервщления, т.е., через 8 дней после инъекции, из печени крыс, обработанных Ad-VP3, ДНК апоптина. В качестве отрицательного контроля параллельно исследовали ДНК из печени крыс, обработанных Ad-luc. Перед нанесением выделенной ДНК на агарозный гель ДНК расщепляли BamHI, в результате чего наблюдали фрагмент ДНК апоптина примерно в 0,5 т.п.о.. Саузерн-блот гибридизировали с меченным 32Р зондом ДНК апоптина.
BamHI фрагмент ДНК апоптина отчетливо виден на Саузерн-блоте в случае ДНК, полученной от животных, обработанных Ad-VP3, и отсутствует, как и ожидалось, на полосах, содержащих ДНК, выделенную из печени крыс, обработанных контрольным rAd-luc. Для исследования количества копий Ad-VP3 в печени различные количества ДНК апоптина наносили параллельно на Саузерн-блот и гибридизировали с меченным зондом ДНК апоптина. Даже через восемь дней после внутривенного инфицирования можно определить 0,25 копий Ad-VP3 на клетку, что указывает на весьма существенную трансдукцию Ad-VP3 в печени.
Экспрессия апоптина и его токсическое действие на клетки печени
Посредством иммунного окрашивания парафиновых срезов печени, обработанной Ad-VP3, или контрольной печени, с использованием моноклональных антител CVI-CAV-85.1 или CVI-CAV-111.3, авторы показали, что примерно 20-30% клеток печени животных, обработанных Ad-VP3, экспрессировали апоптин. Срезы печени контрольных крыс были отрицательными в отношении апоптина.
Для того чтобы исследовать возможное цитотоксическое действие апоптина, экспрессированного Ad-VP3, на клетки печени, применяли два различных метода. В первом методе срезы печени всех крыс, обработанных Ad-VP3, и срезы от контрольных животных обоих типов окрашивали гематоксилином и эозином (НЕ). Во всех проверенных срезах печени не наблюдали морфологических паталогических изменений, что указывает, что экспрессия апоптина не является токсичной для клеток печени крысы.
Возможное вредное действие можно было бы обнаружить при мечении BrdU, которое выявляет вновь разделившиеся клетки. В случае печени, содержащей Ad-VP3, количество меченных BrdU клеток примерно такое же (примерно 2%), как в печени контрольных, обработанных Ad-luc крыс. Следовательно, экспрессия апоптина, как таковая, не оказывает существенного токсического действия in vivo.
Апоптин не оказывает токсического действия на модели in vivo
Как макроскопический, так и гистологический анализ в сочетании с биохимическими и иммунологическими данными показывают, что экспрессия апоптина не оказывает (острого) токсического действия на модели in vivo.
Эти результаты демонстрируют, что лечение, основанное на экспрессии апоптина посредством применения вектора доставки генов или с помощью других способов, будет иметь ограниченное отрицательное побочное действие.
Исследования противоопухолевого действия на модели гепатомы человека
Регулируемая Ad-VP3 экспрессия апоптина приводит к индукции апоптоза трансформированных человеческих клеток в условиях культивирования тканей. Например, регулируемая Ad-VP3 экспрессия апоптина имеет результатом индукцию апоптоза клеток HepG2, полученных из гепатомы человека. До сих пор противоопухолевая активность апоптина (например, экспрессированного Ad-VP3) in vivo не исследовалась.
Поэтому авторы определяли на модели in vivo, будет ли результатом регулируемой Ad-VP3 экспрессии апоптина противоопухолевая активность. Для этого самцам голых мышей Balb/C/nu/nu инъецировали подкожно 1×106 человеческих клеток HepG2 на инъекцию. Клетки гепатомы человека инъецировали каждой мыши по меньшей мере в два или три места. Через три недели после инъекции развиваются отчетливые гепатомные опухоли, видимые под кожей, имеющие средний размер по меньшей мере 5×5 мм.
В опухоли инъецировали векторы rAd-VP3 и контрольный rAd-conl суспендированные в PBS, содержащем 5% сахарозы и 0,1% бычьего сывороточного альбумина.
Вектор rAd-VP3, экспрессирующий апоптин, и контрольный вектор rAd-conl, содержащий ген апоптина в противоположной ориентации 3’-5’ (обратной или антисмысловой) по отношению к промотору Ad MLP, выращивали на клетках PER.С6. Методом PCR-анализа подтверждали, что оба препарата рекомбинантного аденовируса свободны от RCA. rAd очищали с применением центрифугирования в градиенте CsCl.
Инъецировали 7×109 бое частиц rAd в 40 мкл суспензии на опухоль. Каждым типом rAd-вектора обрабатывали по 6 мышей с 2-3 опухолями HepG2. В качестве дополнительного контроля группе из 4 голых мышей, имевших опухоли HepG2, в опухоли инъецировали PBS, содержащий 5% сахарозы и 0,1% бычьего сывороточного альбумина (группа PBS+-).
Апоптин обладает противоопухолевым действием на модели in vivo
Для того чтобы проверить возможное противоопухолевое действие экспрессированного Ad-VP3 апоптина на опухоли от человеческих HepG2, измеряли размер подкожных опухолей во время эксперимента, продолжающегося еще в течение 7 дней после инъекции rAd-VP3 и контрольных суспензий.
Как группа PBS+, так и группа, обработанная контрольным rAd-conl, показывают постепенное увеличение размера опухолей. В противоположность этому, группа, которой в опухоли инъецировали rAd-VP3, показывает уменьшение размера опухолей. На седьмые сутки после инъекции голых мышей умервщляли. Опухоли извлекали и исследовали макроскопически. Отчетливо видно, что гепатомные опухоли, обработанные контрольным вектором rAd-conl или PBS+, представляют сильно васкуляризованную ткань опухоли HepG2 и стали крупнее после обработки. Совершенно иная картина наблюдалась у HерG2-опухолей, обработанных rAd-VP3. Масса оставшейся опухоли бледная на вид из-за уменьшенной васкуляризации опухоли. Опухоли после обработки rAd-VP3 уменьшались в размере. Опухоли также содержали белые пузырьковидные структуры, что является показателем погибших клеток.
Кроме индуцированной апоптином регрессии опухолей никакого другого отрицательного действия экспрессии апоптина на органы (проверялись, в частности, печень и селезенка) зарегистрировать не удалось.
Апоптин обладает противоопухолевой активностью в системе in vivo
Итак, опухоли, обработанные rAd-VP3, показали уменьшение размера, в то время как при контрольных обработках этого не происходит. Это означает, что экспрессия апоптина обладает противоопухолевым действием на модели in vivo.
Тот факт, что экспрессированный Ad-VP3 апоптин может индуцировать регрессию опухоли в быстрорастущей опухоли, такой как HepG2, доказывает сильное противоопухолевое действие апоптина. Тот факт, что апоптин замедляет рост опухолей у голых мышей, показывает, что сама экспрессия апоптина "убивает" опухолевые клетки без дополнительной иммунной реакции.
Описанные исследования токсичности и противоопухолевого действия показывают, что противоопухолевая терапия, основанная на апоптине, безопасна и осуществима.
Описание фигур
Фиг.1 дает схематическое представление о существенных областях аденовирусных адапторных векторов pMAd5 и pMab.
Фиг.2 дает схематическое представление о существенных областях рекомбинантных аденовирусных адапторных векторов pMab-VP3 и рМаb-соn.
Фиг.3 показывает индуцирующую апоптоз активность апоптина в клетках 911, трансфецированных pMab-VP3 или pCMV-VP3. Для трансфекции клеток 911 использовали два независимо клонированных и очищенных препарата ДНК pMab-VP3 (рМаb-VР3/m1 и pMab-VP3/m2). Клетки фиксировали через 3 дня после трансфекции и анализировали методом непрямой иммунофлуоресценции с использованием апоптин-специфических моноклональных антител CVI-CAV-85.1 (85.1; Noteborn et al., 1991). Процент клеток, аномально окрашиваемых DAPI, приводится как относительная мера апоптоза.
Фиг.4 показывает активность апоптина (названного VP3) при индукции апоптоза в человеческих онкогенных клетках гепатомы HepG2, клетках остеосаркомы U20S и нормальных нетрансформированных диплоидных кератиноцитах FSK-1, инфицированных рекомбинантным, содержащим апоптин дефектным по репликации аденовирусом Ad-VP3. Клетки анализировали методом непрямой иммунофлуоресценции с использованием моноклональных антител 85.1 и окрашивания DAPI. Клетки HepG2 и U20S фиксировали на следующий день после трансфекции, а клетки FSK-1 собирали и фиксировали через 4 дня после трансфекции. Процент апоптин-положительных клеток, которые аномально окрашиваются DAPI, приводится как мера индуцируемого апоптином апоптоза (черные прямоугольники). В качестве контроля приводится процент неинфицированных клеток, аномально окрашенных DAPI (незаштрихованные прямоугольники).
Фиг.5 показывает активность апоптоза при индукции апоптином и/или VP2 в клетках Saos-2, трансфецированных 2,5 мкг ДНК pCMV-fs, экспрессирующей апоптин (ранее названой pCMV-VP3; апоптин называется VP3) и 2,5 мкг ДНК pCMV-neoBam (Danen-Van Oorschot, 1997); или 2,5 мкг ДНК pCMV-Vp2, экспрессирующей белок CAV 2 (VP2), и 2,5 мкг ДНК pCMV-neoBam; или 2,5 мкг pCMV-fs и 2,5 мкг pCMV-Vp2, результатом чего является экспрессия как апоптина (VP3), так и VP2. Клетки фиксировали через 3, 4 и 5 дней после трансфекции и анализировали методом непрямой иммунофлуоресценции с использованием апоптин-специфических моноклональных антител CVI-CAV-85.1 (85.1; Noteborn et al., 1991) или моноклональных антител CVI-CAV-111.1 (Noteborn and Koch., 1996). Процент клеток, которые аномально окрашиваются DAPI, дается в качестве относительной меры апоптоза.
Фиг.6 дает схематическое представление о существенных областях рекомбинантных аденовирусных адапторных векторов pMab-VP2.
Фиг.7 дает схематическое представление о существенных областях рекомбинантного ретровирусного вектора-переносчика PLS-VP3-N.





| название | год | авторы | номер документа |
|---|---|---|---|
| ОПРЕДЕЛЕНИЕ ТРАНСФОРМИРУЮЩЕЙ СПОСОБНОСТИ АГЕНТОВ | 1998 |
|
RU2214598C2 |
| СПОСОБ ПОЛУЧЕНИЯ СИСТЕМЫ НАПРАВЛЕННОЙ ДОСТАВКИ БЕЛКОВЫХ МОЛЕКУЛ (ОНКОЛИТИЧЕСКИХ БЕЛКОВ) В ОПУХОЛЕВЫЕ ТКАНИ НА ОСНОВЕ АКТИВИРОВАННЫХ ЛИМФОЦИТОВ | 2013 |
|
RU2552609C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pcDNA4-Apo-2NLS2, НЕСУЩАЯ СИНТЕТИЧЕСКИЙ ГЕН БЕЛКА АПОПТИНА, ИНДУЦИРУЮЩЕГО p53 НЕЗАВИСИМЫЙ АПОПТОЗ ОПУХОЛЕВЫХ КЛЕТОК | 2013 |
|
RU2541777C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pGEM-Puro-DS-Apo, СОДЕРЖАЩАЯ СИНТЕТИЧЕСКИЙ ГЕН АПОПТИНА, ФЛАНКИРОВАННЫЙ ПОСЛЕДОВАТЕЛЬНОСТЯМИ ГЕНОМА ВИРУСА ОСПОВАКЦИНЫ ИЗ РАЙОНА C10L-C12L, И РЕКОМБИНАНТНЫЙ ШТАММ VVdGF-ApoS24/2 ВИРУСА ОСПОВАКЦИНЫ, ПРОДУЦИРУЮЩИЙ АПОПТИН | 2012 |
|
RU2492238C1 |
| АДЕНОВИРУСНЫЙ/АЛЬФА-ВИРУСНЫЙ ГИБРИДНЫЙ ВЕКТОР ДЛЯ ЭФФЕКТИВНОГО ВВЕДЕНИЯ И ЭКСПРЕССИИ ТЕРАПЕВТИЧЕСКИХ ГЕНОВ В ОПУХОЛЕВЫЕ КЛЕТКИ | 2005 |
|
RU2394104C2 |
| УНИВЕРСАЛЬНЫЕ РАКОВОСПЕЦИФИЧНЫЕ ПРОМОТОРЫ И ИХ ИСПОЛЬЗОВАНИЕ В ПРОТИВООПУХОЛЕВОЙ ТЕРАПИИ | 2013 |
|
RU2539764C2 |
| СЛИТЫЙ БЕЛОК ТЕЛОМЕРАЗНОЙ ОБРАТНОЙ ТРАНСКРИПТАЗЫ, КОДИРУЮЩИЕ ЕГО НУКЛЕОТИДЫ И ИХ ПРИМЕНЕНИЕ | 2007 |
|
RU2473691C2 |
| ГЕМОПОЭТИЧЕСКАЯ КЛЕТКА CD34+ И ЕЕ ПРИМЕНЕНИЕ | 2005 |
|
RU2390558C2 |
| ПРИМЕНЕНИЕ РЕЦЕПТОРА EDG2 НА МОДЕЛИ СЕРДЕЧНО-СОСУДИСТОЙ НЕДОСТАТОЧНОСТИ У ЖИВОТНЫХ | 2003 |
|
RU2337963C2 |
| ТЕРАПЕВТИЧЕСКИЕ СОЕДИНЕНИЯ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2015 |
|
RU2711147C2 |
Изобретение относится к генной инженерии, конкретно к векторам доставки генов, индуцирующих апоптоз, и может быть использовано для лечения и диагностики рака, гиперплазии, метаплазии и дисплазии. Рекомбинантные аденовирусные вектора, содержащие апоптин, получают путем котрансфекции в хелперную клеточную линию 911 адапторных плазмид p.AMb-VP3 (в случае экспрессии белка VP3) или плазмид pMAb-VP2 (в случае экспрессии белка VP2) и плазмидной ДНК JM17. Плазмиды pAMb-VP3 несут ген апоптина в ориентации 5’-3’, экспрессия которого осуществляется под контролем аденовирусного главного позднего промотора. Плазмидная ДНК JM17 содержит полную ДНК аденовируса за исключением области Е1 и Е3. Плазмиды pMAb-VP2 несут ген апоптина, с 2 точечными мутациями в пределах кодирующей области. Котрансфекции проводили кальций-фосфатным методом. ДНК рекомбинантного аденовируса образуется путем гомологичной рекомбинации между гомологичными вирусными последовательностями, которые присутствуют в плазмиде pAmb-Vp3 (или pMAb-VP2), и аденовирусной ДНК из плазмидной ДНК JM17. Инфицирование клеток различных опухолей человека векторами доставки генов приводит к индукции апоптоза опухолевых клеток и значительно уменьшенному апоптозу нормальных диплоидных нетрасформированных или незлокачественных клеток. 7 з.п. ф-лы, 7 ил.
| WO 9503414 А, 02.02.1995 | |||
| WO 9630512 А, 03.10.1996 | |||
| ZHUANG S.M | |||
| et al | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Carcinogenesis | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |
| Рекомбинантная плазмидная ДНК pLT21, кодирующая полипептид со свойствами лимфотоксина человека, и штамм бактерий ESCHERICHIACOL - продуцент полипептида со свойствами лимфотоксина человека | 1990 |
|
SU1709731A1 |

