Изобретение относится к области биотехнологии, в частности к антителам, связывающим компонент С3 комплемента человека и подавляющим активацию системы комплемента по альтернативному пути.
Система комплемента играет важную роль в иммунной защите организма, в частности при аутоиммунных заболеваниях и заболеваниях, связанных с воспалением тканей. Список заболеваний, связанных с активацией комплемента, включает такие аутоиммунные заболевания, как ревматоидный артрит, системная красная волчанка, васкулиты, рассеянный склероз и др. [Ballanti. Immunol Res 56(2-3):477-91 (2013)], а также повреждения тканей после ишемии и реперфузии, хронический легочный дистресссиндром, тиреоидит, сахарный диабет, ряд глазных заболеваний и многие другие заболевания [Walport. N Engl J Med 3441058-1066 (2001)]. Система включает более 30 белков, выполняющих регуляторные и иные функции. Так, повышенные концентрации его компонентов С3а, С5а, C5b-9, циркулирующих в крови и других жидкостях организма в условиях острого или хронического воспаления, вызывает и поддерживает активацию нейтрофилов, моноцитов, тромбоцитов, клеток эндотелия сосудов.
Вместе с тем при нарушении баланса между белками активации и регуляторными белками происходит гиперактивация комплемента, которая приводит к анаке на собственные клетки и ткани, что становится причиной возникновения и развития тяжелых болезней, включая аутоиммунные болезни, опреативные вмешательства, обширные травмы, нейродегенеративные процессы, трансплантацию органов и тканей.
Список заболеваний, связанных с активацией комплемента, включает такие аутоиммунные заболевания, как ревматоидный артрит, системная красная волчанка, васкулиты, рассеянный склероз и др. [Ballanti. Immunol Res 56(2-3): 477-91 (2013)], а также повреждения тканей после ишемии и реперфузии, хронический легочный дистресссиндром, тиреоидит, сахарный диабет, ряд глазных заболеваний и многие другие заболевания [Walport. N Engl J Med 3441058-1066 (2001)]. Система включает более 30 белков, выполняющих регуляторные и иные функции. Так, повышенные концентрации его компонентов C3a, С5а, C5b-9, циркулирующих в крови и других жидкостях организма в условиях острого или хронического воспаления, вызывает и поддерживает активацию нейтрофилов, моноцитов, тромбоцитов, клеток эндотелия сосудов.
Система комплемента активируется тремя различными путями: классическим, альтернативным и лектиновым. [https://ru.wikipedia.org/wiki/Система_комплемента; Pyz et al., Ann Med 38, 242-251 (2006)]. Классический путь активируется комплексами антиген-антитело, лектиновый - узнаванием углеводных компонентов лектинами, альтернативный - связыванием С3 с распознаваемыми мотивами на поверхности патогена.
Все три пути активации приводят к протеолитическому расщеплению С3 с образованием фрагментов C3b и C3a, при этом в молекулах С3 и C3b экспонируется сложный тиоэфир, нуклеофильная атака которого приводит к ковалентному связыванию C3b с антигеном (обычно - с поверхностными структурами микробной клетки). Дальнейшее расщепление связанного C3b специфическими протеиназами приводит к образованию фрагментов C3b, C3c и C3dg, которые, наряду с C3b и C3a, распознаются различными рецепторами фагоцитов. Расположенный на поверхности микробных клеток C3b связывает фактор В комплемента, в результате чего фактор В становится субстратом для сериновой протеиназы - фактора D. В результате протеолиза фактором D фактор В распадается на фрагменты Ва и Bb. Bb также является активной протеазой, которая остается связанной с C3b на поверхности микробной клетки. Комплекс C3bBb является С3-конвертазой, атакующей C3, концентрация которого в сыворотке крови человека составляет 1,2 мг/мл. В результате количество C3b еще более увеличивается, поскольку одна молекула С3-конвертазы нарабатывает около 1000 молекул C3b, ковалентно связанных с микробной клеткой и опсонизирующих ее для последующего фагоцитоза. Кроме того, С3 конвертаза, присоединяя дополнительно молекулу C3b, становится С5 конвертазой, (C3b)2Bb, расщепляющей С5 на С5а и C5b. Анафилотоксины С5а и C3a являются активаторами нейтрофилов воспалительных фагоцитов и хемоаттрактантами, притягивающими тучные клетки к очагу заражения, что приводит к локальному выделению гистамина, вазодилятации, накоплению в очаге воспаления фагоцитирующих клеток и к другим воспалительным эффектам. Дальнейшие каскадные реакции в системе комплемента приводят также к образованию мембраноатакующего комплекса, образующего поры в мембранах микробных клеток.
Альтернативный путь (АП) активации относится к одному из факторов врожденного иммунитета и направлен на распознавание и уничтожение патогенов, а также на клиренс клеток, погибших вследствие апоптоза, некроза, и их дериватов [Merle NS, Church SE, Fremeaux-Bacchi V, Roumenina LT. Complement System Part I - Molecular Mechanisms of Activation and Regulation. Front Immunol. 2015; V. 6: Article 262]. Активация комплемента по альтернативному пути происходит непрерывно за счет медленного спонтанного гидролиза тиоэфирной связи в молекуле C3, в результате которой происходит изменение ее конформации и переход из неактивной формы в биоактивную форму C3(H2O). Время существования активной формы C3(H2O) исчисляется миллисекундами, после чего молекула превращается в неактивную форму, обозначаемую C3i, и быстро выводится из организма. Количество измененного С3 в форме C3(H20) и/или C3i составляет 1-2% от всего циркулирующего С3 и равно приблизительно 1 мкг.
Далее каскад реакций активации разворачивается в определенной последовательности. С3 в форме C3(H2O) связывается с фактором В (FB). Образовавшийся комплекс C3(H2O)В подвергается атаке уникальной сериновой протеазой фактором D (FD) с отщеплением части молекулы фактора В - Ва. Другая часть FB - Bb остается связанной с C3(H2O). Вновь образовавшийся комплекс C3(H2O)Bb представляет собой жидкофазную С3 конвертазу, субстратом которой является нативный C3. В результате действия вновь образованного фермента постоянно циркулирующий в крови С3 расщепляется на два фрагмента - C3b и C3a. Последний остается в растворе и выполняет собственные функции анафилатоксина, в то время как реакционноспособный C3b может ковалентно связаться через амидную или эфирную связи с подходящими акцепторными сайтами на поверхности находящегося в микроокружении активатора (А), образуя комплекс А-C3b. Далее происходит цикл превращений, приводящий к образованию С3 и С5 конвертаз и генерации дополнительных количеств C3b.
Эффективность данного процесса невелика, однако она может быть усилена через механизм положительной обратной связи. В этом случае А-C3b взаимодействует с теми же факторами В и D, образуя более стабильную твердофазную С3 конвертазу А-(C3b)nBb, которая в дальнейшем стабилизируется еще одним уникальным белком пропердином или фактором Р (FP). Образовавшийся комплекс А-(C3b)nBbP уже представляет собой фермент с новыми свойствами - C3/С5 конвертазу, которая эффективно атакует свои субстраты С3 и С5.
С5 компонент, подобно C3, расщепляется на большой фрагмент C5b и малый С5а (высокопотентный анафилатоксин). C5b взаимодействует с подходящей мишенью, становясь первым звеном при формировании мембраноатакующего комплекса (МАК), который вызывает гибель клетки-мишени, перфорируя ее мембрану [Merle NS, Church SE, Fremeaux-Bacchi V, Roumenina LT. Complement System Part I - Molecular Mechanisms of Activation and Regulation. Front Immunol. 2015; V. 6: Article 262]. В качестве активаторов (A) альтернативного пути выступают не только попавшие в микроокружение микроорганизмы, но и собственные клетки. Однако все клетки нашего организма надежно защищены ингибиторами, называемыми регуляторными белками, которые прерывают каскад активации комплемента на стадиях формирования C3/С5 конвертаз и на терминальной стадии формирования МАК, не допуская поражения собственных клеток. В то же время микроорганизмы, не имеющие такого защитного аппарата, подвергаются лизису и последующей элиминации комплементом.
Ингибирующими регуляторными молекулами служат, в частности, растворимый кофакторный белок, называемый фактором Н (FH), мембранный кофакторный белок МСР (CD46), рецептор комплемента типа 1 CR1 (CD35) и фактор, ускоряющий распад конвертаз DAF (CD55) [Merle NS, Church SE, Fremeaux-Bacchi V, Roumenina LT. Complement System Part I - Molecular Mechanisms of Activation and Regulation. Front Immunol. 2015; V. 6: Article 262; Merle NS, Noe R, Halbwachs-Mecarelli L, Fremeaux-Bacchi V, Roumenina LT. Complement System Part II: Role in Immunity. Front Immunol. 2015; V. 6: Article 257.]. Упомянутые кофакторы, к которым относится и CRI, взаимодействуют со связанным с активирующей поверхностью C3b и изменяют его конформацию, делая доступным для атаки сериновой протеазой комплемента фактором I (FI), которая отщепляет фрагмент и переводит его в неактивную форму iC3b. Функции регуляторных белков модулируются рядом гликопротеинов на поверхности клеток [Merle NS, Noe R, Halbwachs-Mecarelli L, Fremeaux-Bacchi V, Roumenina LT. Complement System Part II: Role in Immunity. Front Immunol. 2015; V. 6: Article 257].
При физиологических условиях в нормальной сыворотке крови существует баланс между белками, участвующими в активации по альтернативному пути: C3, FB, FD, FP - и регуляторными белками: FH, FI, CD35, CD46 и CD55. Однако, есть множество причин, приводящих к нарушению этого баланса. В частности, в случае генетических нарушений, связанных с мутациями в генах, ответственных за продукцию хотя бы одного из регуляторных белков, возникает нерегулируемая активация, которая приводит к атаке комплементом собственных клеток, что становится причиной серьезных болезней. Среди них хорошо изучены пароксизмальная ночная гемоглобинурия, атипический гемолитический синдром, возрастная дегенерация сетчатки глаза. Такие болезни относят к комплемент-ассоциированным болезням.
Кроме того, в результате внешних воздействий, таких как травмы, оперативные вмешательства, пересадка органов и тканей, гемодиализ, гемосорбция, а также вследствие внутренних нарушений: ишемические повреждения (инфаркты, инсульты, отек мозга), аутоиммунные процессы, сепсис - происходят усиление активации комплемента и повышенная генерация провоспалительных молекул C3a, С5а и макромолекулярного комплекса C5b-9, которые являются хемоатрактантами для нейтрофилов, моноцитов, тучных клеток и базофилов, мигрирующих в очаг воспаления, индукторами цитокинов и адгезионных молекул, а также стимулируют апоптоз. Таким образом, нерегулируемая активация комплемента ведет к патологическому воспалению и тяжелым болезням [Holers V.М. Complement and Its Receptors: New Insights Into Human Disease. Annu. Rev. Immunol. 2014. 32: 433-59; He S, Atkinson C, Qiao F, Cianflone K, Chen C, Tomlinson S. 2009. A complement-dependent balance between hepatic ischemia/reperfusion injury and liver regeneration in mice. J. Clin. Investig. 119: 2304-16; Chen M, Daha MR, Kallenberg CG. The complement system in systemic autoimmune disease. J Autoimmun. 2010; 34: J276-86; Parkes M, Cortes A, van Heel DA, Brown MA. 2013. Genetic insights into common pathways and complex relationships among immune-mediated diseases. Nat. Rev. Genet. 14: 661-73; Holers V.M. The spectrum of complement alternative pathway-mediated disease. Immunol. Rew. 2008, V. 223, P. 300-316; Mastellos DC, Ricklin D, Hajishengallis E, Hajishengallis G, Lambris JD. Complement therapeutics in inflammatory diseases: promising drug candidates for C3-targeted intervention. Mol Oral Microbiol. 2016; 31(1): 3-17].
Для решения вышеупомянутых проблем в качестве терапевтических средств целесообразно использование искусственных блокаторов комплемента, которые способны прерывать каскад реакций активации на каком-либо из его этапов. Наиболее эффективным представляется блокирование активации альтернативного пути за счет связывания или блокирования активации активного сайта на молекуле C3, который вовлечен во взаимодействие с другими компонентами реакционной цепи на стадии инициации активации, но не экспонирован на нативной молекуле, в организме больного, например, с помощью химерных или гуманизированных антител. При этом прямая нейтрализация С3 компонента является нецелесообразной из-за его высокой концентрации в сыворотке крови.
Известно, что все последовательные превращения С3 и его дочерних компонентов, происходящие в процессе активации альтернативного пути комплемента, сопровождаются конформационными перестройками молекул, что приводит к появлению новых антигенных детерминант, так называемых неоантигенов. Были получены моноклональные антитела к некоторым неоантигенам, способные подавлять активацию альтернативного пути комплемента, в частности, мышиные, химерные и гуманизированные антитела к С3 и C3b и их функциональные фрагменты (WO 2004031240, WO 2013152024, RU 2473563, ЕА 2011101593, WO 2006012621), которые избирательно подавляют альтернативный путь активации комплемента. Однако ни одно из заявленных ранее антител до сих пор не применяется в медицинской практике в связи с недостаточной эффективностью технологии их получения.
Наиболее близкими по технической сущности к заявляемому изобретению является разработанная ранее авторами технология получения мышиного моноклонального антитела СС3-4, распознающее конформационный эпитоп (неоантиген), который обычно отсутствует в молекуле С3 и появляется при его активации по альтернативному пути, антитела СС3-4, которое специфично и с высокой аффинностью связывается с молекулой С3 компонента комплемента человека и блокирует альтернативный путь активации комплемента [RU 2584582, 2016] на основе использования штамма гибридомы мыши, депонированного в коллекции РККК под номером РККК(П)764Д и продуцирующий данное антитело.
Недостатком данной технологии является продуцирование негуманизированных антител, что снижает их практическую значимость. В частности, анализ последовательностей аминокислот в легкой и тяжелой цепях антитела СС3-4 показал наличие участков CDR, отличающихся от участков CDR других известных антител к С3 компоненту комплемента человека. Было установлено, что константа диссоциации СС3-4 по С3 компоненту комплемента человека с экспонированной тиофирной связью, определенная с помощью иммуноферментного анализа и вычисленная по Скэтчарду, составляет только 2,92×10-10 М.
Технической задачей, решаемой авторами, являлось расширение круга антител к компоненту С3 человека, способных к ингибированию его биологической активности, путем создания штамма-продуцента нового гуманизированного антитела к конформационному эпитопу С3 компонента комплемента человека, которое может быть испытано в качестве терапевтического антитела для блокирования нерегулируемой активации системы комплемента по альтернативному пути.
Технический результат достигается созданием группы изобретений, включающих в себя моноклональное гуманизированное антитело hC34, характеризующееся наличием измененных (гуманизированных) последовательностей легкой и тяжелой цепей, которые содержат участки, связывающие конформационный эпитоп С3 компонента комплемента человека (CDR-участки) мышиного моноклонального антитела СС3-4, окруженные каркасными участками IgG человека, а также константные области IgG человека, которое специфично и с высокой аффинностью связывается с молекулой C3 компонента комплемента человека и блокирует альтернативный путь активации комплемента, и штамма клеток яичников китайского хомячка СНО-humC34-продуцент рекомбинантных гуманизированных антител и гуманизированных антител к конформационному эпитопу (неоантигену) активированной формы компонента С3 комплемента человека.
При этом используются аминокислотные последовательности CDRH-1, CDRH-2, CDRH-3 мышиного антитела СС3-4, которые участвуют в формировании экспрессионного вектора pTVK4/humC34-H и аминокислотные последовательности CDRL-1, CDRL-2, CDRL-3 мышиного антитела СС3-4, которые участвуют в формировании экспрессионного вектора pTVK4d/humC34-L.
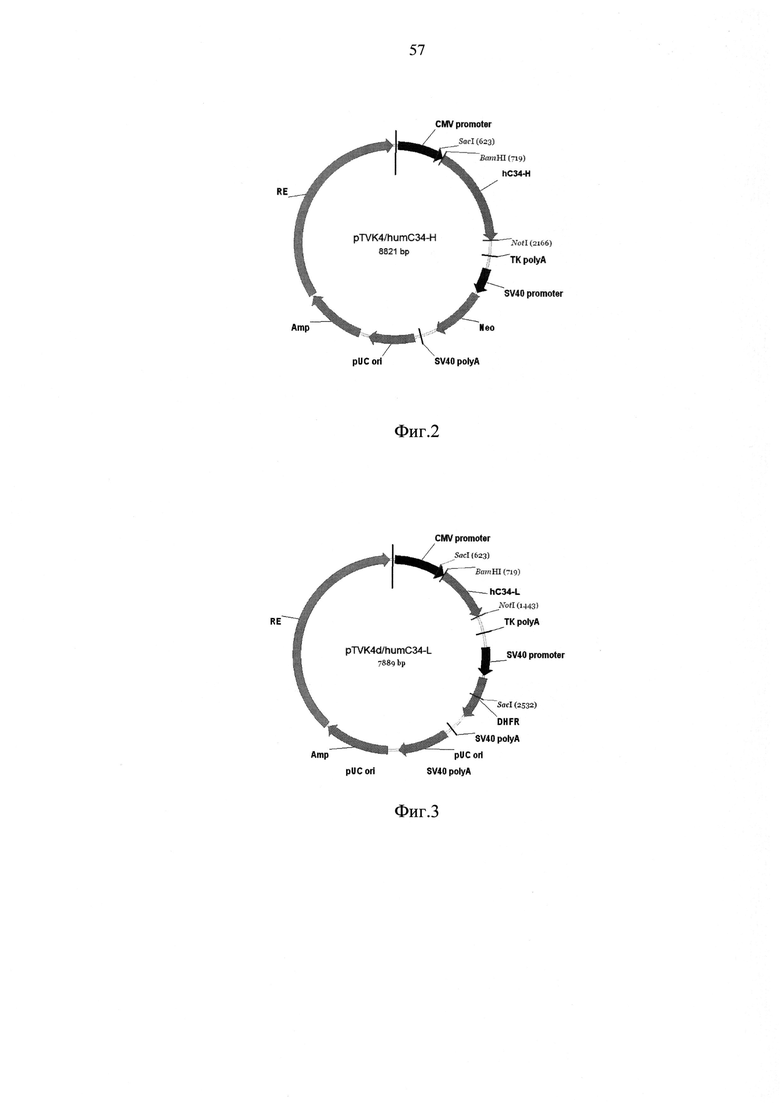
Экспрессионный вектор pTVK4/humC34-H включает в себя промотор предранних белков цитомегаловируса; ген тяжелой цепи гуманизированного антитела к конформационному эпитопу С3 человека; сайт полиаденилирования РНК из гена тимидинкиназы HSV; промотор ранних белков вируса SV40; ген устойчивости к неомицину; сайт полиаденилирования ранних белков вируса SV40; ген устойчивости к ампициллину и регуляторный элемент, усиливающий экспрессию целевого гена.
Экспрессионный вектор pTVK4d/humC34-L включает в себя промотор предранних белков цитомегаловируса; ген легкой цепи гуманизированного антитела к конформационному эпитопу С3 человека; сайт полиаденилирования РНК из гена тимидинкиназы HSV; промотор ранних белков вируса SV40; ген дигидрофолатредуктазы; сайт полиаденилирования ранних белков вируса SV40; ген устойчивости к ампициллину и регуляторный элемент, усиливающий экспрессию целевого гена.
Полученный штамм клеток яичников китайского хомячка CHO-humC34 характеризовался следующими свойствами.
Родословная штамма: Родительская клеточная линия CHOdhfr-. Котрансфекция плазмидами pTVK4/humC34-H и pTVK4d/humC34-L, отбор стабильного высокопродуктивного клона на среде без гипоксантина/тимидина с 200 мкг/мл G-418 и постепенным увеличением концентрации метотрексата. Число пассажей к моменту паспортизации и депонирования: 20.
Стандартные условия выращивания: Среда CDM4CHO (HyClone) без гипоксантина/тимидина с 6 мМ глутамина, 10 нМ метотрексата, 200 мкг/мл G-418, 37°C, 5% CO2.
Культуральные свойства: Суспензионное культивирование в пробирках, колбах или 6-луночных платах на орбитальном шейкере с частотой вращения 130-180 об/мин. Посевная доза 0,3-0,5×106/мл, пересев каждые 3-4 суток. Время удвоения 25-27 часов.
Данные по видовой принадлежности: Cricetulus griseus (ПЦР-анализ с видоспецифичными праймерами).
Маркерные признаки штамма и методы их оценки. Культура устойчива к росту в среде без гипоксантина и тимидина в присутствии 10 нМ метотрексата и 200 мкг/мл G-418
Контроль контаминации бактериями, грибами, микоплазмами и вирусами: контаминации нет.
Характеристики полезного вещества, продуцируемого штаммом. Рекомбинантные гуманизированные антитела hC34 (изотип IgG1, каппа) к неоантигену активированной формы компонента С3 комплемента человека, блокирующие активацию системы комплемента по альтернативному пути (оценка с помощью гемолитического теста с использованием эритроцитов кролика, теста ингибирования образования анафилотоксинов C3a и С5а при активации зимозаном).
Характеристика биосинтеза полезных продуктов (выход продукта в среду, уровень активности и способ ее определения). Рекомбинантные гуманизированные антитела секретируются в культуральную среду в количестве около 20 пг на клетку в сутки. Стабильность культивирования - не менее 30 пассажей.
Способ криоконсервирования: 45% свежей среды CDM4CHO, 45% кондиционной среды, 10% DMSO, 3-5×106 клеток/мл, заморозка до -70°C со скоростью 1°C/мин, далее помещение в жидкий азот, жизнеспособность после размораживания и отмывки от криоконсерванта 90% (с трипановым синим).
Другие особенности штамма: Возможно культивирование клеток в среде DMEM/F12 с 10% фетальной сыворотки (адгезионный тип роста) без дополнительной адаптации.
Штамм депонирован в коллекцию ИНЦ РАН под № РККК(П)776Д 31.03.2016 Полученное моноклональное антитело узнает конформационный эпитоп (неоантиген), который обычно отсутствует в молекуле С3 и появляется при его активации по альтернативному пути. Моноклональное антитело по настоящему изобретению не связывается с нативным C3, у которой тиоэфирная связь находится внутри молекулы, изолированная от ее поверхности петлей, образованной α-цепью фрагмента молекулы и не блокирует активацию комплемента по классическому пути.
Характеристика созданных антител и последовательностей приведена в предлагаемом списке последовательностей.
На фиг. 1 приведено сравнение последовательностей вариабельных областей Н- и L-цепей антитела СС3-4 и вариабельных областей цепей ААС09074 и АВА26038, где СС3-4-Н - последовательность аминокислот в вариабельной области тяжелой цепи антитела СС3-4; CC3-4-L - последовательность аминокислот в вариабельной области легкой цепи антитела СС3-4. CDR-участки подчеркнуты. Аминокислоты в каркасных участках цепей мышиного антитела СС3-4, подвергнутые замене, выделены жирным курсивом.
Список последовательностей:
Seq ID No 1 Последовательность аминокислот CDRH-1 мышиного антитела СС3-4
Seq ID No 2 Последовательность аминокислот CDRH-2 мышиного антитела СС3-4
Seq ID No 3 Последовательность аминокислот CDRH-3 мышиного антитела СС3-4
Seq ID No 4 Последовательность аминокислот CDRL-1 мышиного антитела СС3-4
Seq ID No 5 Последовательность аминокислот CDRL-2 мышиного антитела СС3-4
Seq ID No 6 Последовательность аминокислот CDRL-3 мышиного антитела СС3-4
Seq ID No 7 Последовательность аминокислот вариабельной области Н-цепи гуманизированного антитела hC34.
Seq ID No 8 Последовательность аминокислот вариабельной области L-цепи гуманизированного антитела hC34, вариант L-0
Seq ID No 9 Последовательность аминокислот вариабельной области L-цепи гуманизированного антитела hC34, вариант L-2
Seq ID No 10 Последовательность аминокислот вариабельной области L-цепи гуманизированного антитела hC34, вариант L-4
Seq ID No 11 Последовательность аминокислот вариабельной области L-цепи гуманизированного антитела hC34, вариант L-5
Seq ID No 12 Последовательность аминокислот вариабельной области L-цепи гуманизированного антитела hC34, вариант L-7
Seq ID No 13 Последовательность ДНК, кодирующая Н-цепь гуманизированного антитела hC34 (с сигнальным пептидом).
Seq ID No 14 Последовательность ДНК, кодирующая вариант L-0 L-цепи гуманизированного антитела hC34 (с сигнальным пептидом).
Seq ID No 15 Последовательность ДНК, кодирующая вариант L-2 L-цепи гуманизированного антитела hC34 (с сигнальным пептидом).
Seq ID No 16 Последовательность ДНК, кодирующая вариант L-4 L-цепи гуманизированного антитела hC34 (с сигнальным пептидом).
Seq ID No 17 Последовательность ДНК, кодирующая вариант L-5 L-цепи гуманизированного антитела hC34 (с сигнальным пептидом).
Seq ID No 18 Последовательность ДНК, кодирующая вариант L-7 L-цепи гуманизированного антитела hC34 (с сигнальным пептидом).
Особенности группы изобретений дополнительно поясняется следующими чертежами:
На фиг. 2 и 3 показаны схемы экспрессионных векторов, использованных для создания высокопродуктивного штамма-продуцента гуманизированного антитела hC34 - на фиг. 2 pTVK4/humC34-H, на фиг. 3 - pTVK4d/humC34-L.
При этом используются следующие обозначения:
- CMV promoter - промотор предранних белков цитомегаловируса;
- hC34-H - ген тяжелой цепи гуманизированного антитела к конформационному эпитопу С3 человека;
- hC34-L - ген легкой цепи гуманизированного антитела к конформационному эпитопу С3 человека;
- TK polyA - сайт полиаденилирования РНК из гена тимидинкиназы HSV;
- SV40 promoter - промотор ранних белков вируса SV40;
- DHFR - ген дигидрофолатредуктазы;
- Neo - ген устойчивости к неомицину;
- SV40 poly А - сайт полиаденилирования ранних белков вируса SV40;
- pUC ori - точка начала репликации;
- Amp - ген устойчивости к ампициллину;
- RE - регуляторный элемент, усиливающий экспрессию целевого гена.
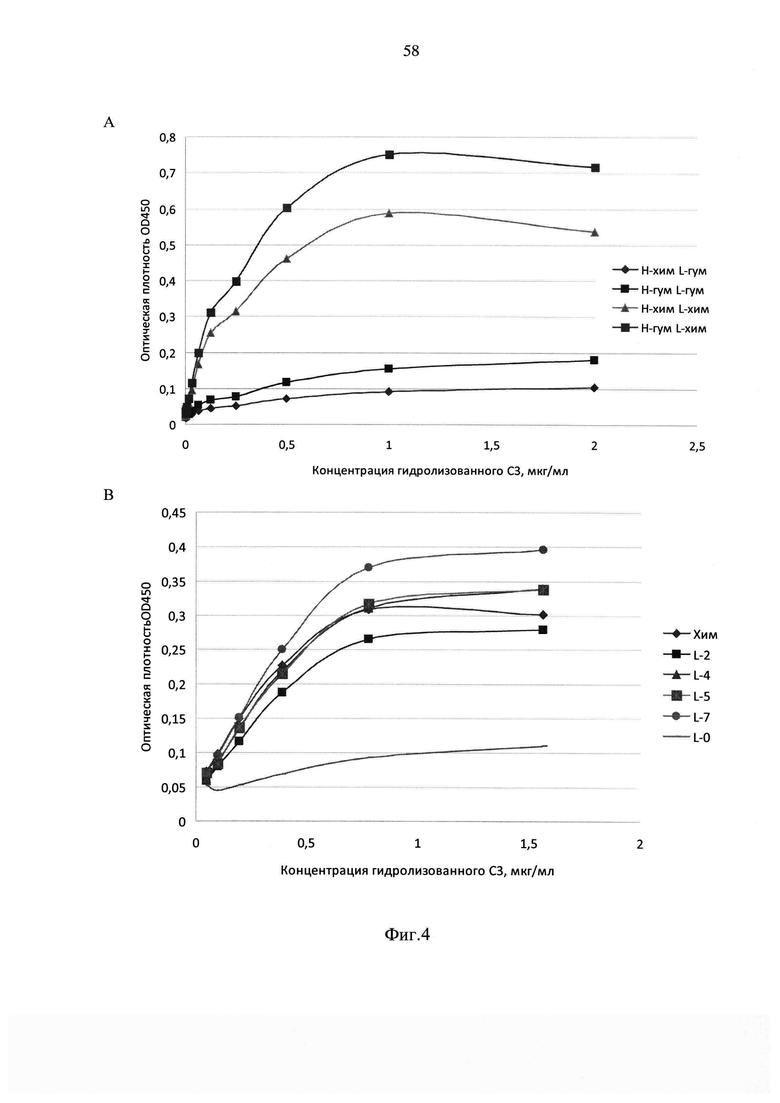
На фиг. 4 приведены результаты анализа аффинности гуманизированных
вариантов антитела hC34, где А - анализ аффинности антител, полученных при коэкспрессии химерных (хим) и гуманизированных (гум), цепей антитела к конформационному эпитопу С3 компонента комплемента (для легкой цепи использовался вариант L-0); В - анализ аффинности антител, полученных при коэкспрессии гуманизированной тяжелой цепи и различных вариантов легкой цепи (Хим - химерный вариант, L-0, L-2, L-4, L-5, L-7 - соответствующие варианты гуманизированной легкой цепи) антитела к конформационному эпитопу С3 компонента комплемента.
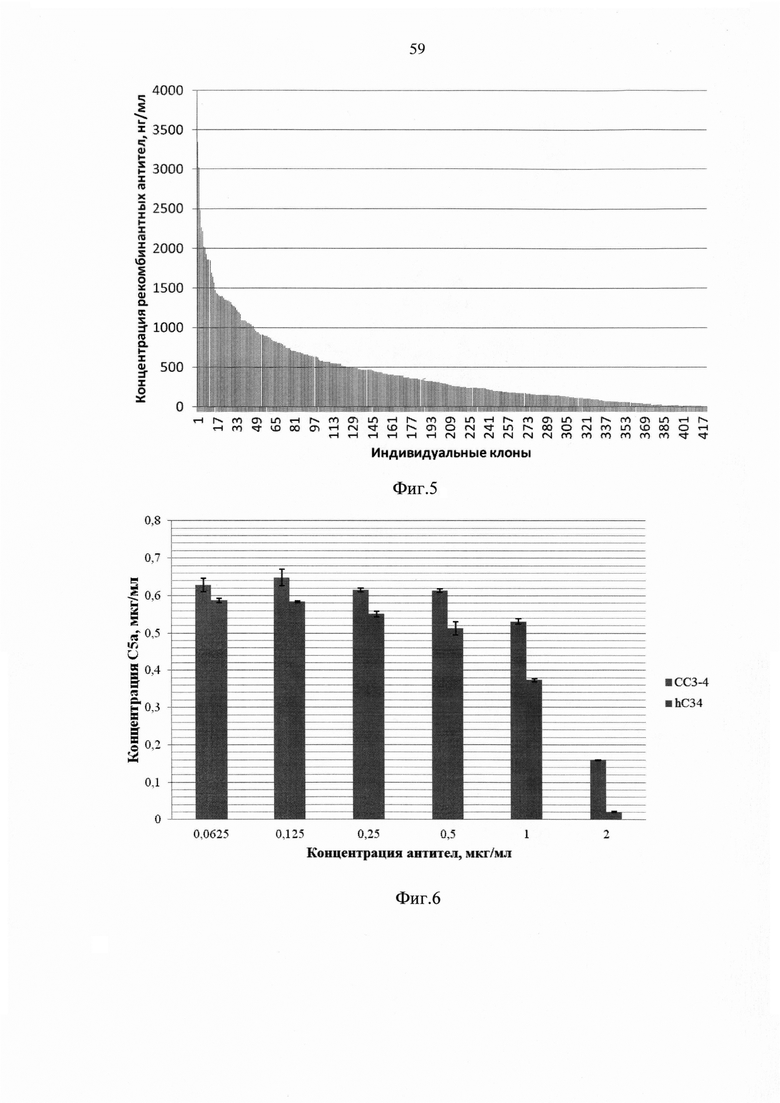
На фиг. 5 показана концентрация человеческих иммуноглобулинов в супернатантах единичных клонов-продуцентов антитела hC34, ранжированных по данному показателю.
На фиг. 6 продемонстрировано блокирование альтернативного пути активации комплемента (АПК) антителами СС3-4 и hC34.
Изобретение поясняется следующими примерами:
Пример 1. Гуманизация вариабельных областей моноклонального антитела СС3-4.
1.1. Анализ структуры антител и конструирование гуманизированного антитела к конформационному эпитопу С3-компонента комплемента человека.
Исходя их определенных нами ранее последовательностей аминокислот в вариабельных областях тяжелой (Н) и легкой (L) цепей мышиного антитела СС3-4, с помощью метода анализа по Кэбэту (2) были вычислены последовательности участков, определяющих комплементарное взаимодействие антитела с антигеном (CDR-участков) и окружающих каркасных участков (Фиг. 1). Далее, с помощью программы Ig-BLAST были найдены легкая и тяжелая цепи известных IgG человека, последовательности которых обладают наибольшим сходством с последовательностями аналогичных цепей антитела СС3-4. Таковыми антителами человека оказались ААС09074 для тяжелой цепи и АВА26038 для легкой цепи, в последовательностях которых также были выделены CDR-участки и каркасные участки. Сравнение последовательностей каркасных участков ААС09074 и АВА26038 с последовательностями соответствующих участков антитела СС3-4 показало, что «мышиные» каркасные участки отличаются от «человеческих» заменами аминокислот, которых в тяжелой цепи было обнаружено всего 31 и в легкой цепи - 37 (Фиг. 1).
Гуманизация вариабельных областей цепей мышиного антитела СС3-4 была проведена путем замены каркасных участков на соответствующие каркасные участки человеческих антител ААС09074 и АВА26038 в тяжелой и легкой цепях, соответственно. Спроектированное таким способом гуманизированное антитело было обозначено hC34. Аминокислотные последовательности вариабельных фрагментов легкой (L) и тяжелой (Н) цепей антитела hC34 представлены в Seq. ID No 7 и Seq. ID No 8. Для легкой цепи, помимо варианта, спроектированного описанным способом и обозначенного как L-0, было дополнительно спроектировано еще 4 варианта, отличающихся от L-0 заменами двух, четырех, пяти или семи аминокислот. Эти варианты были обозначены L-2, L-4, L-5 и L-7, соответственно. Их аминокислотные последовательности представлены в Seq. ID No 9-12.
1.2. Получение генов, кодирующих вариабельные участки химерного и вариантов гуманизированного антитела к конформационному эпитопу С3-компонента комплемента человека, и содержащих их экспрессионных плазмид.
Последовательности ДНК, кодирующие вариабельные фрагменты тяжелой и легкой цепей мышиного моноклонального антитела СС3-4 были получены с помощью ПНР на матрице кДНК, полученной из гибридомы СС3-4 (описано нами ранее в RU 2584582, 2016).
Последовательности ДНК, кодирующие вариабельные фрагменты тяжелой и L-0 варианта легкой цепей гуманизированного антитела были получены синтетическим путем. При проектировании последовательностей триплеты нуклеотидов, кодирующие необходимые аминокислоты выбирались с учетом частоты использования кодонов в геноме китайского хомячка (Cricetulus griseus), представленной в базе codon usage database (kazusa.or.jp).
Последовательность ДНК, кодирующую сигнальный пептид легкой цепи иммуноглобулина мыши - MDFQVQIFSFLLISASVIISRG и последовательность Козак, собирали из синтетических олигонуклеотидов. В качестве источника генов константной части тяжелой и легкой цепей использовали векторы pAL-TA/IGG1const и pAL-TA/IGKconst, созданные нами ранее (RU 2550262). Дальнейшая сборка генов проводилась с помощью перекрывающей ПЦР. Для легкой цепи независимо амплифицировали фрагмент, содержащий сигнальный пептид и последовательность Козак, фрагмент вариабельного участка и фрагмент константного участка, при этом использованные праймеры обеспечивали необходимое перекрывание концов соседних последовательностей. Далее цельную конструкцию амплифицировали из полученных фрагментов за два этапа перекрывающего ПЦР, после чего конечный продукт амплификации клонировали в экспрессионный вектор pIRES-DHFR [RU 2550262, 2015] по сайтам рестрикции BamHI и NotI и подтверждали соответствие полученной нуклеотидной последовательности запланированной с помощью секвенирования. Сборку тяжелой цепи проводили аналогичным способом. В первой реакции ПЦР объединяли последовательности Козак, сигнального пептида и вариабельной области, а затем присоединяли константную часть тяжелой цепи и клонировали в идентичный экспрессионный вектор так же, как легкую цепь. В результате были получены экспрессионные плазмиды pID/CC3-4-H и pID/CC3-4-L, кодирующие тяжелую и легкую цепи химерного антитела, содержащего вариабельные фрагменты мышиного антитела СС3-4 и константные фрагменты человеческого IgG, а также экспрессионные плазмиды pID/hC34-H и pID/hC34-L0, кодирующие тяжелую и легкую цепи гуманизированного антитела hC34.
Для получения генов, кодирующих варианты легкой цепи гуманизированного антитела L-2, L-4, L-5 и L-7, был проведен мутагенез с помощью ПЦР. В качестве матрицы использовали плазмиду pID/hC34-L0. Изменения в последовательности нуклеотидов вносили с помощью двух комплементарных праймеров длиной не менее 24 и.о., содержащих необходимые мутации. После проведения ПЦР и удаления исходной плазмиды обработкой рестриктазой DpnI реакционную смесь использовали для трансформации компетентных клеток. Из полученных колоний выделяли плазмиды и определяли наличие необходимых мутаций с помощью секвенирования. Полученные плазмиды обозначили pID/hC34-L2, pID/hC34-L4, pID/hC34-L5 и pID/hC34-L7, соответственно.
Нуклеотидные последовательности вариабельных участков тяжелой и легкой цепей (варианты L-0, L-2, L-4, L-5 и L-7) гуманизированного антитела hC34 представлены в Seq ID No 13-18.
Пример 2. Анализ связывания С3 компонента комплемента различными вариантами гуманизированного антитела hC34 в составе сыворотки крови человека.
Для получения аналитических образцов вариантов гуманизированного антитела hC34 культуру клеток HEK-293 котрансфецировали смесью плазмид, кодирующими тяжелую и легкую цепь антител. На первом этапе в качестве плазмид, кодирующих тяжелую цепь, использовали pID/hC34-H или pID/CC3-4-H, в качестве плазмид, кодирующих легкую цепь - pID/hC34-L0 или pID/CC3-4-L. Трансфекцию выполняли в 24-луночных платах с помощью коммерческого липосомального реагента. Супернатанты для анализа собирали через 5 суток после трансфекции, затем в них определяли концентрацию иммуноглобулинов человека, а также анализировали связывание С3 компонента комплемента из сыворотки человека с помощью иммуноферментного анализа (ИФА). Для этого в лунки планшета для ИФА сорбировали моноклональные антитела И-4 к IgG человека в концентрации 1,5 мкг/мл в объеме 100 мкл на ячейку. Планшет инкубировали при комнатной температуре 12-14 часов, после чего отмывали промывочным буфером. Затем в плату вносили исследуемые образцы антител, предварительно разведенные до 100 нг/мл в промывочном буфере с 0,1% бычьего сывороточного альбумина. Каждый исследуемый образец вносили не менее чем в 2 стрипа (2×8 лунок). После инкубации в течение 1 часа и отмывки вносили последовательные разведения сыворотки крови человека: от 1/640 до 1/20480 (ряд серийных двухкратных разведений) (по 12 лунок каждой концентрации). Плату инкубировали еще 2 часа и после отмывки добавляли конъюгат моноклональных антител CC3a-1 к C3a фрагменту С3 компонента комплемента человека с пероксидазой хрена в концентрации 0,5 мкг/мл. После часовой инкубации и отмывки плату окрашивали ТМБ.
Об аффинности тестируемых антител судили по углу наклона экспериментальных кривых, полученных в лунках, содержавших образцы исследуемых антител. Результаты представлены на фиг. 4А, из которого видно, что при экспрессии тяжелой цепи гуманизированного антитела hC34 совместно с легкой цепью химерного антитела аффинность полученных антител не снижается по сравнению с химерным антителом. При этом антитела, полученные при использовании варианта L-0 легкой цепи гуманизированного антитела, демонстрируют меньшую аффинность, чем химерные иммуноглобулины соответствующей специфичности.
В связи с этим были спроектированы дополнительные варианты легкой цепи гуманизированного антитела, в которых несколько аминокислот в позициях, которые по данным литературы с наибольшей вероятностью могут повлиять на фолдинг легкой цепи антитела, были заменены вновь на «мышиные», т.е. содержащиеся на соответствующих позициях в исходной последовательности легкой цепи антитела СС3-4. Анализ связывания рекомбинантных антител, полученных путем котрансфекции плазмиды pID/hC34-H и плазмиды, кодирующей один из вариантов легкой цепи (pID/hC34-L0, pID/hC34-L2, pID/hC34-L4, pID/hC34-L5 или pID/hC34-L7) показал (фиг. 4В), что все четыре спроектированных варианта легкой цепи гуманизированного антитела в сочетании с гуманизированной тяжелой цепью имеют сопоставимую с химерным вариантом аффинность, в частности, вариант L-7 показывает в данном тесте наилучший результат.
Пример 3. Получение штамма клеток CHO-humC34 - продуцента гуманизированного антитела к конформационному эпитопу С3 человека.
3.1. Получение экспрессионных плазмид, содержащих гены тяжелой и легкой цепей антитела hC34.
Для создания штамма-продуцента антитела hC34 были использованы вектора pTVK4 и pTVK4d, содержащие, помимо экспрессионных элементов, указанных на фиг. 4 и 5, специализированный регуляторный элемент UCOE (3), обозначенный как RE, при этом pTVK4 несет ген устойчивости к неомицину, a pTVK4d - ген дигидрофо-латредуктазы. Исходные плазмиды pID/hC34-H и pID/hC34-L7 обрабатывали рестриктазами BamIII и NotI и выделяли фрагмент ДНК, содержащий, помимо последовательностей тяжелой и легкой цепей антитела, последовательность Козак, обеспечивающую инициацию трансляции, и последовательность, кодирующую сигнальный пептид из иммуноглобулина мыши (MDFQVQIFSFLLISASVIISRG). Далее фрагменты ДНК, кодирующие легкую и тяжелую цепи гуманизированного антитела, были переклонированы в pTVK4 и pTVK4d, рестрицированные аналогичным образом. В результате было получено два экспрессионных вектора, pTVK4/humC34-H и pTVK4d/humC34-L, схемы которых приведены на Фиг. 2 и 3. Рестрикционный анализ и сиквенирование подтвердили правильность сборки плазмид и отсутствие мутаций в генах цепей гуманизированного антитела.
3.2. Получение штамма-продуцента гуманизированного антитела hC34
Для постоянной трансфекции использовали клетки линии яичников китайского хомячка, дефицитные по гену дигидрофолатредуктазы (CHOdhfr-), ранее адаптированные к суспензионному росту в бессывороточных условиях. Клетки культивировали в среде CDM4CHO («HyClone», США) с добавлением 4 mM аланилглугамина, 0,1 мМ гипоксантина, 0,016 мМ тимидина и 0,1% Pluronic F-68 («Sigma», США) в микробиореакторах типа TubeSpin 50 («ТРР», Швейцария) на орбитальном шейкере Sanyo MIR-S100C (200 об/мин), который был помещен в CO2-инкубатор (5% CO2, 37°C).
Трансфекцию проводили с помощью липосомального реагента «Freestyle» («Life Technologies», США) по инструкции производителя, используя смесь 2,4 мкг плазмиды pTVK4/humC34-H и и 3,6 мкг плазмиды pTVK4d/humC34-L на 5×106 клеток. Через 48 часов после трансфекции клетки переводили на среду без гипоксантина/тимидина с добавлением 200 мкг/мл генитицина G-418 («Life Technologies»). В последующие дни среду меняли с интервалом 3 суток. Через 15 суток клетки переносили в колбы Эрленмейера для последующего наращивания, после чего было проведено клонирование методом серийных разведений с целью отбора индивидуальных клонов с высокой продукцией. Клонирование проводили путем высева клеток в плотности 1 клетка на лунку 96-луночной платы. Всего таким образом было засеяно 20 96-луночных плат. Через 10 суток визуально отбирали лунки, содержащие единственный клон. Было идентифицировано 420 клонов. Через 3 суток с помощью иммуноферментного анализа (ИФА) оценивали содержание иммуноглобулинов человека в супернатантах индивидуальных клонов. Результат представлен на фиг. 5.
Для дальнейшей работы было отобрано 40 клонов, в супернатантах которых концентрация иммуноглобулинов была более 1000 нг/мл. Отобранные клоны были целиком перенесены в 48-луночные платы. Через 48 часов был проведен повторный анализ концентраций иммуноглобулинов в супернатантах. По результатам второго скрининга было отобрано 12 клонов с максимальной волюметрической продукцией рекомбинантных антител, концентрация иммуноглобулинов в супернатантах которых была более 4800 нг/мл. Отобранные клоны переносили в 6-луночные для последующего культивирования. Далее клетки каждого клона высевали в 12-луночные платы по 0,5×106 клеток на лунку в 2 мл среды для культивирования без G-418. Продукцию антител оценивали на 3 сутки после посева. В результате были отобраны 8 клонов с максимальной продукцией на 3 сутки (более 5000 нг/мл).
Данные клоны культивировали на шейкере в микробиореакторах типа TubeSpin 50 в объеме 5 мл. Начальная плотность культуры составляла 0,5×106 клеток/мл, пересев проводили с интервалом 3 суток. После третьего пассажа на шейкере клетки высевали в плотности 0,5×106 клеток/мл в 5 мл среды без G-418 и анализировали продукцию антител и динамику роста клеток. На основании полученных данных рассчитывали удельную продуктивность клеток клонов, которая оказалась наилучшей у клона N3 (4,5 пг на клетку в сутки).
Далее клетки клона N3 адаптировали к росту на среде с 10 нМ метотрексата (МТХ). После 7 пассажей в среде с МТХ клетки субклонировали в 96-луночные платы в среде с 10 нМ МТХ и 200 мкг/мл G-418, при этом был получен 80 субклонов. В результате последовательного отбора субклонов с наибольшей продуктивностью, как это описано выше, было установлено, что субклон N3/8 обладает наилучшими ростовыми характеристиками и продуктивностью: на 7 сутки культивирования концентрация жизнеспособных клеток составила 4,6×106 клеток/мл при жизнеспособности 90%, а максимальная волюметрическая продукция рекомбинантных антител составила 220 мкг/мл.
Данный субклон был использован для создания Главного клеточного банка штамма-продуцента, а также депонирован в Российской коллекции клеточных культур под номером РККК(П) 776Д и рабочим названием «СНО-humC34».
3.3. Анализ нейтрализующей активности гуманизированных антител к конформационному эпитопу С3 человека, секретируемы штаммом-продуцентом CHO-humC34.
Клетки штамма-продуцента CHO-humC34 культивировали на шейкере в колбах Эрленмейера, в результате чего было получено 150 мл культуральной жидкости, из которой выделяли антитела с помощью аффинной хроматографии на иммобилизованном белке А. В результате был получен лабораторный образец рекомбинантнного гуманизированного антитела hC34 в количестве 20 мг. Способность антител hC34 блокировать активацию комплемента по альтернативному пути оценивали в сравнении с мышиным моноклональным антителом СС3-4.
Активацию комплемента по альтернативному пути проводили в 20 мМ буфере HEPES (pH 7,2-7,4), содержащем 150 мМ NaCl, 10 мМ ЭГТА, 5 мМ MgCl2, 1 мг/мл БСА (буфер для АПК) путем инкубации сыворотки с активатором АПК зимозаном в 96-луночном планшете для ИФА. В ряды ячеек планшета вносили по 50 мкл раствора гуманизированных антител hC34 или мышиных антител СС3-4 в различных концентрациях и 100 мкл сыворотки крови здоровых доноров, содержащей 1 М ε-аминокапроновой кислоты, разведенной в 10 раз в буфере для АПК. Плату инкубировали при 37°C при постоянном перемешивании в течение 10 мин., после чего в ячейки вносили по 10 мкл суспензии зимозана (с концентрацией 10 мг/мл), приготовленной в буфере для АПК. Для отрицательного контроля вместо зимозана вносили буфер АПК. Плату инкубировали 1 час при температуре 37°C при постоянном перемешивании на шейкере. Активацию комплемента останавливали внесением в ячейки 10 мкл раствора, содержащего 0,2 М ЭДТА, 10 мМ PMSF. Концентрации антител в инкубационных пробах варьировали от эквимолярного значения по отношению к концентрации антигена C3, которую предварительно определяли в сывороточных пробах с помощью ИФА, в сторону понижения. Установленные отношения молярных концентраций МАТ: С3 в экспериментальных пробах составили 1:1, 1:2, 1:4, 1:8, 1:16. Степень активации комплемента определяли по содержанию анафилатоксина С5а в пробах после инкубации. О нейтрализующей (блокирующей) способности судили по изменению уровней анафилактосина С5а в сывороточных пробах в зависимости от концентрации антител.
Результаты приведены на фиг. 6, из которого видно, что гуманизированные антитела hC34 обладают способностью блокировать активацию альтернативного пути комплемента даже несколько более эффективно, чем мышиные антитела СС3-4, которые подвергали гуманизации. Это делает гуманизированные антитела hC34, производимые штаммом-продуцентом СНО-humC34, перспективными для терапевтического применения с целью блокады АПК.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО К КОНФОРМАЦИОННОМУ ЭПИТОПУ СЗ КОМПОНЕНТА КОМПЛЕМЕНТА ЧЕЛОВЕКА | 2018 |
|
RU2687609C1 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО СС3-4 К КОНФОРМАЦИОННОМУ ЭПИТОПУ С3 ЧЕЛОВЕКА, ШТАММ ГИБРИДНОЙ ДНК МЫШИ РККК(П)764Д - ПРОДУЦЕНТ МОНОКЛОНАЛЬНОГО АНТИТЕЛА СС3-4 | 2015 |
|
RU2584582C1 |
| ГУМАНИЗИРОВАННОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СПЕЦИФИЧНОЕ К СИНДЕКАНУ-1 | 2013 |
|
RU2551235C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО 6H8HU, СВЯЗЫВАЮЩЕЕСЯ С ОПУХОЛЕВЫМ АНТИГЕНОМ PRAME, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ АНТИТЕЛА | 2020 |
|
RU2768737C1 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО К АМИЛОИДУ БЕТА | 2013 |
|
RU2668161C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО | 2008 |
|
RU2538709C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА D И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2799044C2 |
| ГУМАНИЗИРОВАННОЕ ТАУ-АНТИТЕЛО | 2013 |
|
RU2644242C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К АМИЛОИДУ БЕТА | 2008 |
|
RU2567151C2 |
| АНТИТЕЛА К Р-СЕЛЕКТИНУ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ И ИДЕНТИФИКАЦИИ | 2011 |
|
RU2603097C2 |































Изобретение относится к области биохимии, в частности к гуманизированному антителу, которое связывается с эпитопом С3 компонента комплемента человека. Также раскрыты ДНК, кодирующая указанное антитело, экспрессионный вектор, для экспрессии указанного антитела, а также штамм клеток яичников китайского хомячка, который продуцирует указанное антитело. Изобретение обладает способностью специфически связывается с эпитопом С3 компонента комплемента человека, что позволяет эффективно лечить заболевания, ассоциированные с С3 компонентом комплемента человека. 8 н. и 4 з.п. ф-лы, 6 ил., 3 пр.
1. Гуманизированное антитело к конформационному эпитопу С3 компонента комплемента человека, содержащее фрагмент, выбранный из установленной по Кэбэту группы, включающей в себя CDRH-1 участок по Seq ID No: 1; CDRH-2 участок по Seq ID No: 2; CDRH-3 участок по Seq ID No: 3; CDRL-1 участок по Seq ID No: 4; CDRL-2 участок по Seq ID No: 5; CDRL-3 участок по Seq ID No: 6 для связывания с конформационным эпитопом С3 компонента комплемента человека.
2. Гуманизированное антитело к конформационному эпитопу С3 компонента комплемента человека по п. 1, имеющее последовательность вариабельной области тяжелой цепи по Seq ID No 7.
3. Гуманизированное антитело к конформационному эпитопу С3 компонента комплемента человека по п. 1, имеющее последовательность вариабельной области легкой цепи по Seq ID No 8.
4. Гуманизированное антитело к конформационному эпитопу С3 компонента комплемента человека по п. 1, имеющее последовательность вариабельной области легкой цепи по Seq ID No 9.
5. Гуманизированное антитело к конформационному эпитопу С3 компонента комплемента человека по п. 1, имеющее последовательность вариабельной области легкой цепи по Seq ID No 10.
6. Гуманизированное антитело к конформационному эпитопу С3 компонента комплемента человека по п. 1, имеющее последовательность вариабельной области легкой цепи по Seq ID No 11.
7. Гуманизированное антитело к конформационному эпитопу С3 компонента комплемента человека по п. 1, имеющее последовательность вариабельной области легкой цепи по Seq ID No 12.
8. ДНК, кодирующая тяжелую цепь гуманизированного антитела к конформационному эпитопу С3 компонента комплемента человека по п. 1 или 2, представленная на Seq ID No 13 для связывания конформационного эпитопа С3 компонента комплемента человека ДНК, кодирующая вариант L-0 легкой цепи гуманизированного антитела к конформационному эпитопу С3 компонента комплемента человека по пп. 1 и 3, представленная на Seq ID No 14 для связывания конформационного эпитопа С3 компонента комплемента человека.
9. ДНК, кодирующая вариант L-2 легкой цепи гуманизированного антитела к конформационному эпитопу С3 компонента комплемента человека по пп. 1 и 4, представленная на Seq ID No 15 для связывания конформационного эпитопа С3 компонента комплемента человека.
10. ДНК, кодирующая вариант L-4 легкой цепи гуманизированного антитела к конформационному эпитопу С3 компонента комплемента человека по пп. 1 и 5, представленная на Seq ID No 16 для связывания конформационного эпитопа С3 компонента комплемента человека.
11. ДНК, кодирующая вариант L-5 легкой цепи гуманизированного антитела к конформационному эпитопу С3 компонента комплемента человека по пп. 1 и 6, представленная на Seq ID No 17 для связывания конформационного эпитопа С3 компонента комплемента человека ДНК, кодирующая вариант L-7 легкой цепи гуманизированного антитела к к конформационному эпитопу С3 компонента комплемента человека по пп. 1 и 7, представленная на Seq ID No 18 для связывания конформационного эпитопа С3 компонента комплемента человека.
12. Экспрессионный вектор pTVK4/humC34-H, включающий промотор предранних белков цитомегаловируса; ген тяжелой цепи гуманизированного антитела к конформационному эпитопу С3 человека; сайт полиаденилирования РНК из гена тимидинкиназы HSV; промотор ранних белков вируса SV40; ген устойчивости к неомицину; сайт полиаденилирования ранних белков вируса SV40; ген устойчивости к ампициллину и регуляторный элемент, усиливающий экспрессию целевого гена а также ДНК, выбранную из группы ДНК, представленных в пп. 8-13, для экспрессии гуманизированного антитела для связывания конформационного эпитопа С3 компонента комплемента человека в клетках млекопитающих.
13. Экспрессионный вектор pTVK4d/humC34-L, включающий в себя промотор предранних белков цитомегаловируса; ген легкой цепи гуманизированного антитела к конформационному эпитопу С3 человека; сайт полиаденилирования РНК из гена тимидинкиназы HSV; промотор ранних белков вируса SV40; ген дигидрофолатредуктазы; сайт полиаденилирования ранних белков вируса SV40; ген устойчивости к ампициллину и регуляторный элемент, усиливающий экспрессию целевого гена, а также последовательность ДНК, выбранную из группы последовательностей ДНК, представленных в пп. 8-13, для экспрессии гуманизированного антитела для связывания конформационного эпитопа С3 компонента комплемента человека в клетках млекопитающих.
14. Штамм клеток яичников китайского хомячка CHO-humC34, трансфецированных векторами по пп. 8 и 13, депонированный в РККК под номером РККК (П) 776Д - продуцент гуманизированного антитела для связывания конформационного эпитопа С3 компонента комплемента человека.
| US 0008377437 B2 от 19.02.2013 | |||
| WO 2009114585 A1 от 17.09.2009 | |||
| US 20090280116 A1 от 12.11.2009 | |||
| Трость для обнаружения опасного нагревания материалов, сложенных в кучу | 1929 |
|
SU13876A1 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО СС3-4 К КОНФОРМАЦИОННОМУ ЭПИТОПУ С3 ЧЕЛОВЕКА, ШТАММ ГИБРИДНОЙ ДНК МЫШИ РККК(П)764Д - ПРОДУЦЕНТ МОНОКЛОНАЛЬНОГО АНТИТЕЛА СС3-4 | 2015 |
|
RU2584582C1 |

