Данное изобретение относится, в основном, к комбинации технологий получения рекомбинантной ДНК и моноклонального антитела для получения новых терапевтических средств, в частности, для производства неиммуногенных антител и их использования.
Обоснование изобретения
Для млекопитающих иммунный ответ передается двумя типами клеток, специфически взаимодействующих с чужим материалом, т.е. антигенами. Один из типов этих клеток - B-клеток ответственны за производство антител. Второй класс клеток, T-клетки, включает большое разнообразие клеточных подклассов контролирующих in vivo функцию как B-клеток и широкий диапазон других кроветворных клеток, включая T-клетки.
Одним из способов осуществления этого контроля является производство лимфокина известного как интерлейкин-2-/IL-2/, первоначально названного фактором роста T-клеток. Основной функцией IL-2, по-видимому, является стимулирование и поддержка T-клеток. Некоторые иммунологи считают, что IL-2 может быть в центре полного иммунного ответа (см. Farrar, J., et. all. Immunol. Rev., 63:129-166 (1982) (в ссылках).
Для проявления своего биологического действия 1 - 2 взаимодействует со специфическими мембранными рецепторами, обладающими высоким сродством. /Green, IV. , et. all. Progress: n Hematology XIV., E.Brown, Ed. Gune and Statton, New York (1986), at pgs 283 tf/. У человека рецептором IL-2 является сложный многоцепной гликопротеин с одной цепью, известной как Тас-пептид, размером около 55 kD /Leonard:W, et. all., J. Biol. Chem., 260:1872 (1985)) включено в ссылку. Был выделен ген, кодирующий этот протеин и указывающий на пептид, состоящий из 272 аминокислот, включая и сигнальный пептид из 21 аминокислоты /Leonard: W, et. all. , Nature 311: 626 (1989)/. 219 NH2-терминальные аминокислоты p55 Tac протеина очевидно включают клеточные домены /Leonard W., et. all., Science, 230, 633-639 (1985)/.
В основном, объяснение структуры и функций рецепторов человеческого IL-2 связано с развитием специфически реактивных моноклональных антител. В особенности, одно моноклональное антитело мыши, известное как анти-Тас /Uchiyama, et al., H. Immunol. 126; 1393 (1981)/ показало, что IL-2 рецепторы могут быть обнаружены на T-клетках, также на клетках семейства моноцитов-макрофагов, на клетках Купфера в печени, клетках Лангерхана кожи, и, конечно, активированных T-клетках. Важно, что T-клетки в состоянии покоя, B-клетки или циркулирующие макрофаги, обычно не показывают IL-2-рецептор. /Hennmann, et. al., J Exp. Med., 162; 1111 (1985)/.
Анти-Тас моноклональное антитело также использовали для определения функции лимфоцитов, которая включает взаимодействие с IL-2, и показали, что оно замедляет различные функции T-клеток, включая выработку цитотоксичных и супрессорных T-лимфоцитов в культуре клеток. Также, основываясь на исследованиях с анти-Тас и другими антителами, сейчас связывают ряд заболеваний с неправильным выражением IL-2 рецептора T-клетками, в особенности T-клеточную лейкемию у взрослых.
Недавно показали, что IL-2 является идеальным объектом для новых терапевтических подходов к заболеваниям с участием T-клеток. Было предложено, что антитела, специфические к IL-2 рецептору, такие как анти-Тас моноклокальное антитело, можно использовать как само по себе, так и как иммуноконъюгат /напр. с Рицином A, изотопами и т.п./ для эффективного удаления клеток, несущих IL-2 рецептор. Данные агенты могут, например, теоретически исключить лейкемические клетки, несущие рецептор IL-2, некоторые B-клетки, или активированные T-клетки, участвующие в протекании болезни, но позволяют удерживать зрелые нормальные T-клетки и их предшественников для обеспечения возможности формирования нормального T-клеточного иммунного ответа при необходимости. В общем, большинство других специфических агентов для T-клеток могут существенно разрушать все периферические T-клетки, которые ограничивают терапевтическую эффективность агентов. В общем, использование соответствующих моноклональных антител, специфических для IL-2-рецептора, может иметь терапевтический эффект при автоиммунных заболеваниях, трансплантации органов и любых нежелательных реакциях активированных T-клеток. Действительно, были проведены клинические испытания с использованием, например, анти-Тас антител /см. в основном, Waedman, T., et al., Cancen. Res., 45, 625 (1985) and Waedman, T., Science, 232; 727-732 (1986), обе ссылки включены/.
К сожалению, использование анти-Тас и других нечеловеческих моноклональных антител имеет некоторые недостатки, особенно при повторяющихся терапевтических режимах, как объяснены ниже. Например, моноклональные антитела мыши недостаточно хорошо фиксируют комплемент человека и не обладают другими важными характеристиками функций иммуноглобулина при использовании для человека.
Возможно даже более важно то, что анти-Тас и другие нечеловеческие моноклональные антитела содержат существенные отрезки аминокислотных последовательностей, которые обладают иммуногенностью при введении человеку. Многочисленные исследования показали, что после введения чужого антитела человеку, иммунная реакция, вызываемая у пациента антителом, может быть достаточно сильной, существенно уменьшающей терапевтический эффект применения антител при первоначальном введении. Более того, поскольку увеличивающееся число различных мышиных или других антигенных /к человеку/ моноклональных антител можно ожидать для лечения различных заболеваний в дальнейшем, после первого и второго курса лечения любыми нечеловеческими антителами, последующее лечение даже и другими методами может быть неэффективным или даже опасным само по себе.
В то время, как производство так называемых "химерных антител" (т.е. изменяемых участков мыши, присоединенных к постоянным участкам человека) оказалось успешным, осталась существенной проблема иммуногенности. В общем, производство человеческих иммуноглобулинов, реактивных в IL-2 рецептору человека, было очень сложно при использовании обычных методов производства человеческих моноклональных антител. Аналогично использование технологии рекомбинантной ДНК для получения так называемых "гуманизированных" антител" /см. напр. EPO Publication N 0239400/ приводит к неопределенным результатам, особенно по отношению к непредсказуемой способности к связыванию.
Следовательно, необходимы улучшенные формы человекоподобных иммуноглобулинов, например, такие, которые специфичны для IL-2 рецептора человека, которые существенно неиммуногенны для человека, но которые можно легко и экономично получать методом, пригодным для целей терапии и других потребностей. Данное изобретение удовлетворяет всем этим условиям.
Краткое описание изобретения
Данное изобретение предусматривает новые композиции, полезные, например, при лечении расстройств человека, передаваемых T-клетками, композиции, содержащие человекоподобные иммуноглобулины, способные специфически блокировать связывание человеческого IL-2 со своим рецептором и/или способные к связыванию с протеином p55 Tac на человеческих рецепторах IL-2. У иммуноглобулинов может быть две пары комплексов - легкая цепь /тяжелая цепь, обычно, по крайней мере одна пара при этом имеет цепи, включающие участки, определяющие комлементарность для мыши, функционально присоединенные к сегментам, составляющим основу для человека. Например, участки, определяющие комплементарность для мыши, с или без дополнительных изначально существующих аминокислотных остатков, характерных для мыши, можно использовать для получения человекоподобных антител, способных к связыванию с человеческим рецептором IL-2, при уровне сродства более 108 М-1.
Иммуноглобулины, включающие связывающие участки и другие производные, приведенные здесь, можно легко получать согласно данному изобретению, используя многочисленные методы рекомбинантной ДНК, в основном реализуемые в трансфектных клетках, предварительно сделанных неумирающими, эукариотических клетках, таких как клетки миеломы или гибридомы. Полинуклеотиды, включающие первую последовательность, кодирующую основные участки человеческого иммуноглобулина, и вторую последовательность, кодирующую участки, определяющие желаемую комплементарность иммуноглобулина, можно получить синтетически или соответственно, комбинируя сегменты кДНК и ДНК генома.
Человекоподобные иммуноглобулины можно использовать сами по себе в достаточно чистой форме или в комплексе с цитотоксичным агентом, таким как радионуклид, рибосомальный ингибирующий протеин, или цитотоксичным агентом, активным на поверхности клетки. Все эти соединения будут особенно полезны при лечении расстройств, передаваемых T-клетками. Человекоподобные иммуноглобулины или их комплексы можно приготовить в фармацевтически удобной дозировке, которую можно изменять в зависимости от способа применения.
Данное изобретение также предусматривает новые способы конструирования цепей человекоподобного иммуноглобулина, имеющие один или более участков, отвечающих за комплементарность /CDR'S/ от донорского иммуноглобулина, и основной участок от человеческого иммуноглобулина, предпочтительные способы включают первоначально включение основы или аминокислотных последовательностей изменяемого участка донорского иммуноглобулина в соответствующие последовательности набора цепей человеческого иммуноглобулина и выбор одной наиболее гомологичной последовательности из набора, подобной человеческому иммуноглобулину. Человеческий иммуноглобулин или акцептирующую иммуноглобулиновую последовательность обычно выбирают из набора по крайней мере от 10 до 20 последовательностей цепей иммуноглобулина, и она обычно имеет наибольшую гомологичность к иммуноглобулиновой последовательности донора по сравнению с любой последовательностью в наборе. Основная последовательность человеческого иммуноглобулина обычно гомологична на 65-70% или больше к основным последовательностям донорского иммуноглобулина. Иммуноглобулин донора может быть как с легкой цепью, так и с тяжелой /или с обеими/ и набор человека будет содержать тот же тип цепи. Гуманизированные легкую и тяжелую цепи можно использовать для образования полного гуманизированного иммуноглобулина или антитела, имеющего две пары цепей типа легкая/тяжелая, с (или без) частичными или полными постоянными участками человека или с другими протеинами.
В другом варианте данного изобретения, как в соединении с вышеприведенным способом или отдельно, можно заместить дополнительные аминокислоты в акцепторной цепи иммуноглобулина на аминокислоты, образующие CDR иммуноглобулиновой цепи донора. Более специфично дальнейшее выборочное замещение аминокислоты из основы человека из акцептирующего иммуноглобулина соответствующей аминокислотой из донорского иммуноглобулина, оно может быть выполнено в таком положении на иммуноглобулинах, где:
/a/ аминокислота на основном "человеческом" участке акцептирующего иммуноглобулина редко встречается в этом положении и соответствующая аминокислота в донорском иммуноглобулине обычна для этого положения в последовательностях человеческого иммуноглобулина или
/b/ аминокислота сразу соседствует с одним из CDR'S или
/c/ предсказано, что аминокислота находится в пределах 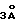 от CDR в трехмерной модели иммуноглобулина и способна взаимодействовать с антигеном или с CDR'S гуманизированного иммуноглобулина.
от CDR в трехмерной модели иммуноглобулина и способна взаимодействовать с антигеном или с CDR'S гуманизированного иммуноглобулина.
Гуманизированная цепь иммуноглобулина обычно включает по крайней мере около 3 аминокислот от донорского иммуноглобулина, кроме CDR'S, обычно, по крайней мере одна из которых сразу прилегает как к CDR в донорском иммуноглобулине. Можно конструировать как легкие? так и тяжелые цепи, используя любой или все критерии выбора положения.
При присоединении к интактному антителу гуманизированные легкие и тяжелые цепи по данному изобретению будут существенно неимунногенны у человека и сохранят, в основном, то же сродство к антигену, как и донорский иммуноглобулин /к такому как протеин или другое соединение, содержащее эпитоп/. Эти уровни сродства могут изменяться от примерно 108 M-1 или выше и могут примерно в четыре раза превышать сродство к антигену донорского иммуноглобулина.
Краткое описание рисунков
Фиг. 1. Сравнение последовательностей тяжелой цепи анти-Тас /верхние линии/ и тяжелой цепи Eu /нижние линии/. Использован однобуквенный код для аминокислот. Первая аминокислота на каждой линии пронумерована слева. Одинаковые аминокислоты в двух последовательностях соединены линиями. Три CDR'S подчеркнуты. Другие положения аминокислот, для которых больше использовали анти-Тас аминокислоту, чем Еu аминокислоту, отмечены.
Фиг. 2. Сравнение последовательностей легкой цепи анти-Тас /верхние линии/ и легкой цепи Eu /нижние линии/. Использован однобуквенный код для аминокислот. Первая аминокислота на каждой линии отмечена слева. Идентичные аминокислоты в двух последовательностях соединены линиями. Три CDR'S подчеркнуты. Другие положения аминокислот, для которых больше использовали анти-Тас аминокислоту, чем Eu аминокислоту в гуманизированной тяжелой цепи анти-Тас, отмечены *.
Фиг. 3 Нуклеотидная последовательность в гене для гуманизированного изменяемого участка тяжелой цепи анти-Тас гена. Транслируемая аминокислотная последовательность для части гена, кодирующего протеин, показана ниже нуклеодитной последовательности. Нуклеотиды ТCTAGA в начале и конце гена являются XbaI участками. Основная последовательность тяжелой цепи начинается с аминокислоты # 20 Q.
Фиг. 4 Нуклеотидная последовательность в гене для гуманизированного изменяемого участка легкой цепи анти-Тас гена. Транслируемая аминокислотная последовательность для части гена, кодирующего протеин, показана под нуклеодитной последовательностью. Нуклеотиды TCTAGA в начале и конце гена являются XbaI участками. Основная последовательность легкой цепи начинается с аминокислоты # 21D.
Фиг. 5 A. Последовательности из 4 олигонуклеотидов, используемых для синтеза гуманизированного анти-Тас тяжелоцепного гена, напечатаны от 5' до 3'. B. Относительное положение олигонуклеотидов. Стрелки указывают на положение 3' для каждого нуклеотида.
Фиг. 6 A. Последовательности из четырех олигонуклеотидов, используемых для синтеза гуманизированного анти-Тас легкоцепного гена, напечатаны от 5' до 3'. /B/. Относительные положения олигонуклеотидов. Стрелки указывают в направлении 3' для каждого олигонуклеотида. Показано положение участка Hind III при перекрывании JFD 2 и JFD 3.
Фиг. 7 Схематическая диаграмма плазмиды pHu GTACI, используемой для экспрессии гуманизированной анти-Тас тяжелой цепи. Показаны походящие области разрыва, кодирующие участки тяжелой цепи показаны прямоугольниками. Направление считывания с иммуноглобулинового (Iq) промотора стрелкой. E = ген ускоритель тяжелых цепей. Hyg = ген сопротивления гидромициновый.
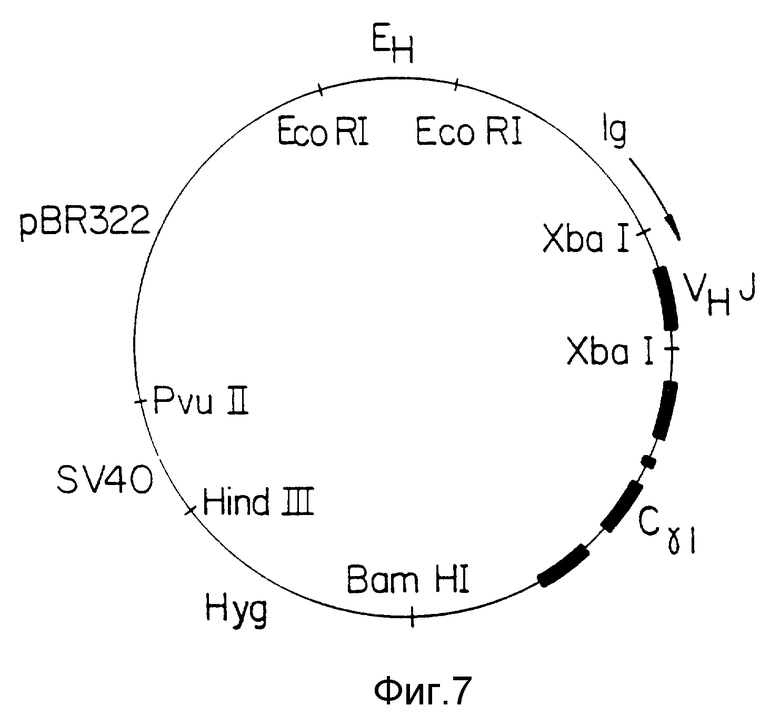
Фиг. 8 Схематическая диаграмма плазмиды pHuLTAC, используемой для выражения анти-Тас легкой цепи. Показаны подходящие области разрыва, кодирующие участки легкой цепи показаны прямоугольниками. Направление считывания с Iq промотора показано стрелкой.
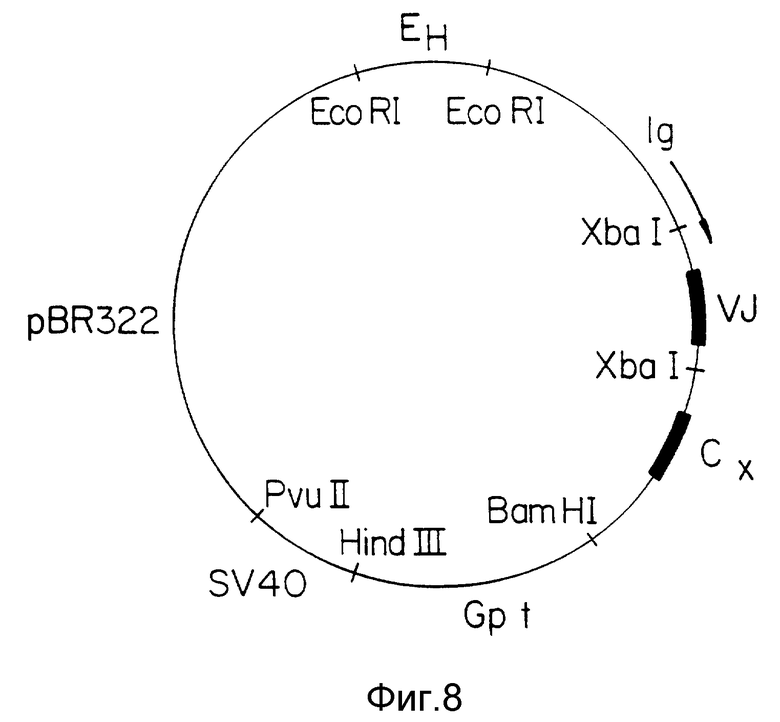
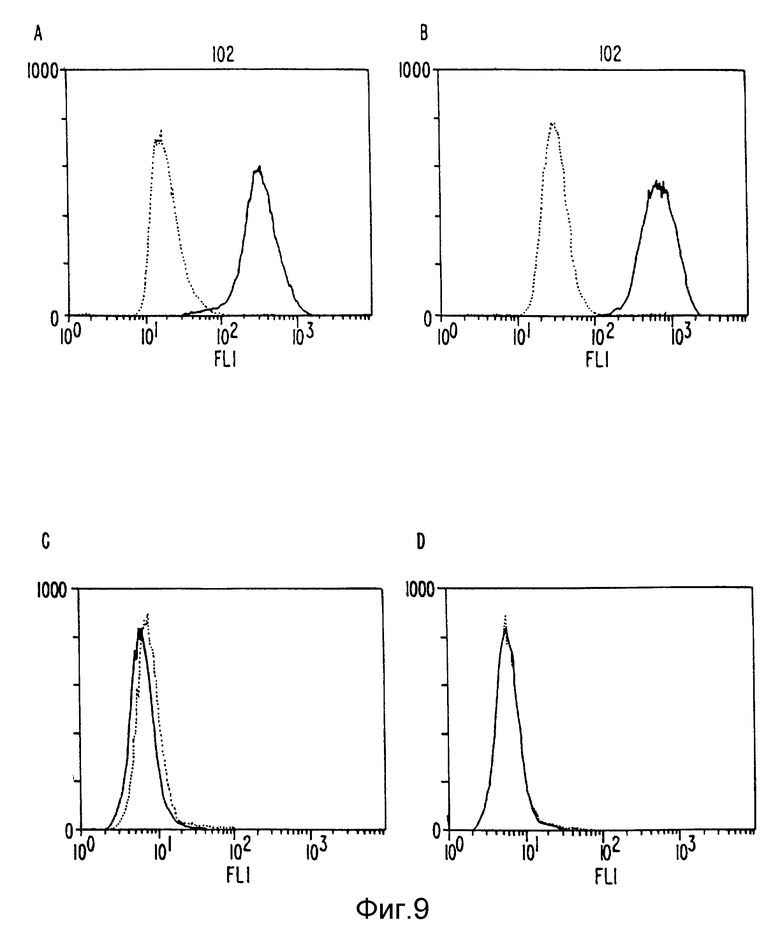
Фиг. 9 Флуороцитометрия клеток HUT-102 и Jurkut, покрытых анти-Тас антителом или гуманизированным анти-Тас антителом, вместе с соответственно сопряженными с флуоресцином Ig антителом козла анти-мыши или козла анти-человеческим Ig антителом, как отмечено. Прерывистая кривая на каждом графике показывает результат, полученный при исключении первого антитела, а сплошная кривая - результаты включения первого и второго /коньюгированного/ антител, как описано.
Фиг. 10
/A/ Флуороцитометрия клеток HUT-102 окрашенных о-40 нг анти-Тас, как показано, затем бионизированным анти-Тас и затем фикоэритрин-конъюгированным авидином.
/B/ Флуороцитометрия клеток HUT-102, окрашенных указанным антителом, затем бионизированным анти-Тас и затем фикоэритрин-конъюгированным авидином.
Подробное описание изобретения
В соответствии с одним из способов по данному изобретению предложены человеко-подобные иммуноглобулины, специфически реагирующие с желаемыми эпитопами, такими как на IL-2 рецепторе на T-клетках человека. Эти иммуноглобулины, имеющие связывающее сродство, по крайней мере около 108 M-1, предпочтительно 109 M-1 до 1010 M-1 или выше, способные, следовательно, к блокированию связывания IL-2 с рецепторами IL-2 человека. Человекоподобные иммуноглобулины будут иметь человекоподобную основу и могут иметь участки, определяющие комплементарность /CDR'S/ от иммуноглобулинов, обычно, мыши, специфически реагирующих с эпитопом на p55 Тас протеина. Иммуноглобулины по данному изобретению можно экономично получить в больших количествах и они находят применение, например, при лечение заболеваний, передаваемых T-клетками человека, многими способами.
Известно, что структурная единица базового антитела содержит тетрамер. Каждый тетрамер составлен из двух идентичных пар полипептидных цепей, при этом в каждой паре есть одна "легкая" /около 25 к/" и одна "тяжелая"/около 50-70 к/ цепь.
NH2-терминал каждой цепи начинает изменяемый участок из 100 до 110 или больше аминокислот, первично ответственных за распознавание антигена. COOH терминал каждой цепи определяет постоянный участок, первоначально отвечающий за функцию эффектора.
Легкие цепи классифицируют как каппа и лямбда. Тяжелые цепи классифицируют /и подразделяют/ как гамма, мю, альфа, дельта или эпсилон и определяют изотип антитела как IgG, IgM, IgA, IgD и IgE соответственно. Внутри легких и тяжелых цепей изменяемые и постоянные участки присоединены "J" участком, состоящим из 12 или более аминокислот, с тяжелой цепью также включающий "J" участок из 1 или больше аминокислот /см. в осн. Fundamental Immenology, Poul, W, Ed. Chapten 7, pgs. 131-166 Raven, Press, N.Y., (1984)/ включено в ссылки.
Изменяемый участок каждой пары легкой и тяжелой цепи образует связывающий антитело участок. Все цепи имеют одну и ту же общую структуру относительно устойчивых основных участков, соединенных тремя суперизменяющимися участками, также называемыми CDR'S /см. "Seguences of Proteins of Immunological Interest, "Kabat, E., et al., US. Department of Health and Human Services, (1983); and Cholthia and Lesk, J. Mol. Biol., 196; 901-917 (1987) включены/.
CDR'S от двух цепей каждой пары упорядочены основными участками, обуславливая связывание со специфическим эпитопом.
Употребляемый здесь термин "иммуноглобулин" относится к протеину, состоящему из одного или более полипептидов, кодируемых иммуноглобулиновыми генами. Изученные иммуноглобулиновые гены включают каппа, лямбда, альфа, гамма, дельта, эпсилон и мю гены постоянных участков наряду с огромным количеством генов изменяемых участков. Иммуноглобулины могут существовать в разнообразных формах помимо антител, включая, например, Fv, Fab, F(ab)2, а также в единичных цепях /см. например, Huston, et al., Proc. Nat., Acad. Sci, 85, 58795883 (1988), and Bird et al, Science, 242; 423-426 (1988) вкл. в ссылки), (см. в осн. Hood et al, "Immunilogy", Benjamin, W.J., 2 nd. ed (1984), and Hunkapillen and Hood, Nature, 323: 15-16 (1988), включены в ссылки).
Химерные антитела - это антитела, чьи легко и тяжело цепные гены были сконцентрированы, обычно путем генной инженерии, из участков иммуноглобулинового гена, принадлежащих к разным образцам, например, изменяемые /V/ участки генов из моноклонального антитела мыши можно присоединить к постоянным сегментам человека /C/, таким как γ1 и γ3. Типичное терапевтическое химерное антитело таким образом является гибридным протеином, состоящим из V или антиген-связывающего домена из человеческого антитела. /напр. A.T.C.C. доп. номер CRL 9688 выделяют анти-Тас химерное антитело/, хотя можно использовать и другие млекопитающие.
Используемый здесь термин "frame work" (основной) участок относится к тем частям вариабельных участков легких и тяжелых цепей иммуноглобулина, которые относительно консервативны /т.е. не такие как CDR'S среди различных иммуноглобулинов в единичных видах, как определил Кабат и др. (см. любую ст. ). Как указано здесь, основной (frame work) участок иммуноглобулина, подобного человеческому, это основной участок, который в каждой существующей цепи включает по крайней мере около 70 или больше аминокислотных остатков, типично от 75 до 85 или больше остатков, идентичных аналогичным в иммуноглобулине человека.
Используемый здесь термин "подобный человеческому" относится к иммуноглобулину, включающему основной участок иммуноглобулина, подобного человеческому и присутствующий в котором любой постоянный (constant) участок, по существу, гомологичен постоянному участку иммуноглобулина человека, т.е. по крайней мере идентичен на 85-90%, предпочтительно на 95%. Так все части человекоподобного иммуноглобулина, за исключением возможно CDR'S, существенно гомоголичны соответственным частям одной или более последовательностей природного иммуноглобулина человека. Например, человекоподобный иммуноглобулин не может включать химерное антитело на основе изменяемого участка мыши /постоянного участка человека/.
В соответствии с другим основным аспектом данного изобретения также включены критерии, по которым ограниченное число аминокислот в основе человекоподобной или гуманизированной иммуноглобулиновой цепи выбирают такими же как аминокислоты в тех же положениях в донорском Ig, а не в акцепторном Ig, для увеличения аффинности антитела, содержащего гуманизированную иммуноглобулиновую цепь.
Данный аспект изобретения частично основан на том, что существуют две причины потери аффинности в ранее известных способах получения гуманизированных антител/ используя в качестве примеров антитела мыши в качестве источника CDR'S/:
/1/ При соединении CDR'S мыши с человеческой основой аминокислоты, в основе близкие к CDR'S, становятся человеческими, а не мышиными. Не выдвигая специальной терапии, мы просто считаем, что эти измененные аминокислоты могут слегка искажать CDR'S, т.к. они создают различные электростатические и гидрофобные силы по сравнению донорским антителом мыши и искаженные CDR'S могут не обеспечить такие же эффективные контакты с антигеном, как CDR'S в донорском антителе.
/2/ Наряду с этим, аминокислоты в первоначальном антителе мыши, которые расположены близко к CDR'S, но не являются его частью /т.е. они все еще части основы/, могут контактировать с антигеном, что вносит вклад в аффинность. При гуманизации антитела эти аминокислоты теряются, т.к. все аминокислоты основы делаются человеческими.
Для того, чтобы избежать этого и получить гуманизированные антитела с высокой аффинностью к избранному антигену, в данном изобретении используют следующие 4 критерия для конструирования гуманизированных иммуноглобулинов. Эти критерии можно по одному или при необходимости в сочетании для достижения желаемой аффиности или других характеристик.
Критерий I. Используйте в качестве акцептора основу от особенного иммуноглобулина человека, который обладает необычной гомологичностью к донорскому иммуноглобулину, предназначенному для гуманизирования, или используйте совместную основу от многих человеческих антител. Например, сравнение последовательности тяжело /или легко/ цепного изменяемого участка мыши с тяжело /или легко/ цепными изменяемыми участками человека в банке данных /например, the National Biomedical Rescareh Foundation Protein Indentification Resource/ показывает, что степень гомологичности к различным участкам человека изменяется широко, типично от около 40% до около 60-70%. При выборе в качестве акцептора иммуноглобулина одного из человеческих тяжелых /относительно легких/ цепных изменяемых участков, наиболее гомологичных тяжело /относительно легко/ цепным изменяемым участкам донорского иммуноглобулина, необходимо сменить небольшое число аминокислот при переходе от донорского иммуноглобулина к гуманизированному. Не создавая специальной теории, можно считать, что существует меньший шанс изменения конформации CDR'S при смене аминокислот около них. Более того, точная общая форма гуманизированного антитела, включающая гуманизированную иммуноглобулиновую цепь, может более точно повторить форму донорского антитела, также уменьшая возможность искажения CDR'S.
Обычно выбирают одну из 3-5 наиболее гомологичных тяжелоцепных последовательностей изменяемого региона из набора по крайней мере от 10 до 20 известных тяжелых цепей человека в качестве акцептора для создания тяжелоцепной основы, и аналогично поступают с легкой цепью. Предпочтительно используют один из 1-3 наиболее гомологичных изменяемых участков. Выбранная акцепторная иммуноглобулиновая цепь должна обладать предпочтительно по крайней мере 65% гомологичности в основном участке по отношению к донорскому иммуноглобулину.
Независимо от способа выбора иммуноглобулина можно достичь высокого сродства, избирая небольшое количество аминокислот в основе гуманизированной иммуноглобулиновой цепи, таких же как аминокислоты в тех же положениях в доноре, а не в акцепторе. Следующие критерии определяют, какие аминокислоты можно выбрать таким образом. Предпочтительно можно выбрать большинство или все донорские аминокислоты, находящиеся в данном положении, удовлетворяющем один из данных критериев.
Критерий II. Если аминокислота в основе человеческого акцептора необычна/ т.е. "редкая", то есть встречается в данном положении не чаще, чем в 10% человеческих тяжелоцепных /относительно легких/ последовательностей V участка, находящихся в банке данных /и, если донорская аминокислота в данном положении типична для последовательностей человека/, т.е. "обычна", что в данном случае означает, что она встречается по крайней мере в 25% последовательностей в банке данных/, то скорее следует выбрать донорскую аминокислоту, а не акцепторную. Данный критерий позволяет быть уверенным, что атипичная аминокислота в человеческой основе не разрушит структуру антитела. Более того, замещая необычную аминокислоту на аминокислоту донорского антитела, которая типична для человеческих антител, можно сделать гуманизированное антитело менее иммуногенным.
Критерий III. В положении, непосредственно прилежащим к 3 CDR'S в гуманизированной цепи иммуноглобулина, лучше выбрать донорскую аминокислоту, а не акцепторскую. Эти аминокислоты особенно вероятно взаимодействуют с аминокислотами на CDR'S и выбранные с акцептора искажают донорские CDR'S и уменьшают аффинность. Более того, прилегающие аминокислоты могут непосредственно взаимодействовать с антигеном /Amitet all., Science, 233, 747-753 (1986)/, и выбрать эти аминокислоты от донора может быть желательно для поддержания всех контактов антитела, которые обеспечивают аффинность в исходном антителе.
Критерий IV. Трехмерная модель, типичная для исходного донорского антитела, показывает, что некоторые аминокислоты, находящиеся вне CDR'S тем не менее близки к ним и имеют большую вероятность взаимодействия с аминокислотами в CDR'S через водородную связь, силы Ван/дер-Ваальса, гидрофобное взаимодействие и т.п. В таких положениях лучше выбрать донорскую аминокислоту, а не акцепторную аминокислоту иммуноглобулина. Аминокислоты в соответствии с данным критерием должны иметь атом боковой цепи в пределах  от некоторого участка на CDR'S и должны содержать атомы, способные взаимодействовать с атомами CDR способами, указанными выше. Компьютерные программы для создания моделей протеинов, таких как антитела, в основном доступны и хорошо известны специалистам /см.Loen et al. Int.J.Quant.Chem.,Quant.Biol.Symp. 15: 55-56 (1988), Bruccoleni et.al., Nature, 335, 564-568 (1988); Ehothia et.al. , Science, 233: 755-758 (1986) (в кл. в ссылки). Это не является частью изобретения. В самом деле, поскольку у всех антител похожая структура, можно использовать известные структуры антител, которые доступны из банка данных протеинов Brookhaven при необходимости в качестве грубых моделей других антител. Можно использовать коммерчески доступные компьютерные программы для представления данных моделей на мониторе для вычисления расстояния между атомами и для оценки подобия различных аминокислотных взаимодействий /см. Fernin et.al., J.Mol.Graphics.b: 13-27, (1988)/.
от некоторого участка на CDR'S и должны содержать атомы, способные взаимодействовать с атомами CDR способами, указанными выше. Компьютерные программы для создания моделей протеинов, таких как антитела, в основном доступны и хорошо известны специалистам /см.Loen et al. Int.J.Quant.Chem.,Quant.Biol.Symp. 15: 55-56 (1988), Bruccoleni et.al., Nature, 335, 564-568 (1988); Ehothia et.al. , Science, 233: 755-758 (1986) (в кл. в ссылки). Это не является частью изобретения. В самом деле, поскольку у всех антител похожая структура, можно использовать известные структуры антител, которые доступны из банка данных протеинов Brookhaven при необходимости в качестве грубых моделей других антител. Можно использовать коммерчески доступные компьютерные программы для представления данных моделей на мониторе для вычисления расстояния между атомами и для оценки подобия различных аминокислотных взаимодействий /см. Fernin et.al., J.Mol.Graphics.b: 13-27, (1988)/.
Гуманизированные или подобные человеческим антитела имеют по крайней мере три потенциальных преимущества над мышинными или в некоторых случаях химерными антителами при лечении человека:
1. Поскольку в части эффектора оно человеческое, оно может лучше взаимодействовать с другими частями человеческой иммунной системы, например, разрушать клетки мишени более эффективным путем комплементарно-зависимой цитотоксичности /CDC/ или антитело зависимой клеточной цитотоксичности /ADCC/.
2. Иммунная система человека не сможет считать основу или постоянный участок гуманизированного антитела чужеродным, и следовательно, реакция антитела на такое введенное антитело будет слабее, чем по отношению к полностью чужеродному антителу мыши или частично чужеродному химерному антителу.
3. Известно, что у введенных антител мыши в человеческом организме период полураспада намного меньше, чем у нормальных антител /D.Shaw et.al.J.Immunol; 138: 4534-4538(1987)/. Введенные гуманизированные антитела будут, вероятно, иметь время полураспада более близким к настоящим антителам человека, что позволяет перейти к меньшим и менее часто даваемым дозам.
Данное изобретение имеет целью усовершенствованные гуманизированные иммуноглобулины /например, способные к связыванию IL-2 рецептора человека/ по отношению к описанным в EPA N 0239400. Эта заявка, существо которой не перекрывается настоящей заявкой, описывает для некоторых иммуноглобулинов замещение участков CDR'S в легко или тяжелоцепных изменяемых доменах акцептора антитела на аналогичные участки CDR'S/ обычно доступных для растворителя /от антитела другой специфичности. В данной заявке также обслуживается для некоторых иммуноглобулинов возможность только передающих остатков, которые доступны /для растворителя/ с участка связывания антигена, и данные остатки могут очевидно включать некоторые участки основы /в особенности, те остатки, про которые известно, что они участвуют в связывании антигена как описано у Amit et. al. Science 233:747-753 (1986), или возможно остатки, необходимые для межцепного взаимодействия, но для выбора которых в данной заявке дано неудовлетворительное объяснение/. Так, например, предпочтительный вариант осуществления данного изобретения заключается в замещении всех аминокислот CDR, S и основы непосредственно соседствующих /или предпочтительно каждой/ с CDR'S в общем, любой остаток основы, который также способен к контакту с CDR'S, т. е. поддерживает их конформацию /и обычно специфичность связывания их антигена/, специально включен в предпочтительный вариант осуществления данного изобретения, как описано подробно.
Один из аспектов данного изобретения связан с сегментами рекобинантной ДНК, кодирующей тяжело и/или легкоцепные CDR'S/ обычно с другими аминокислотными остатками, как описано выше, иммуноглобулина, способного к связыванию с желаемым эпитопом, таким как на IL-2 рецепторе человека /т.е. с анти-Тас моноклональным антителом/. Сегменты ДНК, кодирующие эти участки, обычно присоединяют к сегментам ДНК, кодирующим соответственные человекоподобные основные участки. Например, предпочтительные последовательности ДНК, которые содержат в коде для полипептидных цепей суперизменяемые участки тяжелых и легких цепей анти-Тас /с человекоподобными основными участками/, показаны на фиг. 3 и 4 соответственно. Из-за дегенерации кодонов и неточных замещений аминокислот другие последовательности ДНК можно легко заместить на эти последовательности, как описано ниже.
Сегменты ДНК обычно далее включают последовательность ДНК, осуществляющую контроль за кодированием, присоединенную к кодирующим последовательностям человекоподобного антитела, включая естественно существующие /ассоциированные/ или гетерологичные участки промотора. Предпочтительно, чтобы последовательностями, осуществляющими контроль за экспрессией, были эукариотические промоторные системы с векторами, способными к трансформации или трансфекции эукариотических клеток хозяина, но можно также использовать контрольные последовательности для прокариотических хозяев. Как только вектор включен в соответствующего хозяина, хозяина поддерживают в условиях, удобных для экспрессии нуклеотидных последовательностей на высоком уровне, и, как предусмотрено, следует сбор и очистка легких цепей, тяжелых цепей, димеров легких/тяжелых цепей или интактных антител, связывающих фрагментов или других форм иммуноглобулинов.
Последовательности ДНК человеческого постоянного региона можно выделить в соответствии с хорошо известными процедурами из многообразия клеток человека, но предварительно сделанных неумирающими B-клеток /см. Kabat, на выбор, и WP87/02671/. Например, гены и последовательности каппа иммуноглобулина постоянного участка и J участка описаны у Heiter et al., Cell. 22:197-207 /1980/, нуклеотидные последовательности гена человеческого иммуноглобулина C γ I описаны у Ellison et.al., Nucl. Acid.Res., 10: 4071/1982/ обе работы включены в ссылки. CDR'S для получения иммуноглобулинов по данному изобретению могут быть подобным же образом получены из моноклональных антител, способных к связыванию с нужным антигеном /т.е. IL-2-рецептором человека/, и произведены в любом подходящем млекопитающем, включая мышей, крыс, кроликов или других позвоночных, с помощью которых можно получать антитело хорошо известными способами. Подходящий источник клеток для последовательностей ДНК и клеток хозяина для экспрессии и секреции иммуноглобулина можно подобрать из ряда источников таких как American Type Culture Collection /"Catalogue of Cell Lines and Hybridomas", Fifth Edition (1985) Rockvill, Maryland, USA), включены в прилагаемые ссылки.
Кроме человекоподобных иммуноглобулинов, специально описанных здесь, другие "существенно гомологичные" измененные иммуноглобулины могут быть легко сконструированы и произведены с использованием методов рекомбинантных ДНК, хорошо известных специалистам. Например, для иммуноглобулинов 1 - 2 рецептора основные /каркасные/ участки могут отличаться от последовательностей на фиг. 3 и 4 на уровне первичной структуры несколькими замещенными аминокислотами, терминальными и промежуточными добавками и исключениями и тому подобным. Более того, ряд различных основных человеческих участков можно использовать отдельно или в комбинации в качестве основы для человекоподобных иммуноглобулинов данного изоберетения. В общем, модификации генов можно легко осуществить, используя большой набор известных способов, таких как направленный мутагенез /см. Gillman., Gene. 8-81-97 /1979/, Roberts, S., et. al., Nature, 328:731-734(1987) обе включены/.
С другой стороны, полипептидные фрагменты, включающие только часть первичной структуры антитела, можно получать и они будут обладать одной или более функциями иммуноглобулина /например, способностью к комплементарной фиксации/. Также, поскольку, как и многие гены, гены, связанные с иммуноглобулином содержат отдельные функциональные участки, каждый из которых имеет одну или больше определенную биологическую активность, гены можно слить с функциональными участками из других генов /например, ферментов, см. U. SS. N 132; 387, filed Dec 15, 1987, вкл. в ссылки/ с образованием слитных протеинов /например, иммунотоксинов/ с новыми свойствами.
Последовательности нуклеиновых кислот по данному изобретению, способные к полной экспрессии желаемых антител подобных человеческим, можно сформировать из разнообразия различных полинуклеотидов /генома, или к ДНА, РНК, синтетических олигонуклеотидов и т.п./ и компонентов /например, V, J. D- и C участков/ и с использованием разнообразных методов. Присоединение соответствующих последовательностей генома является в настоящее время наиболее обычным методом получения, но можно также использовать последовательности к ДНК /см. Eu. Paten Publication N 0239400 and Reichmann L. ey, al. Nature 332:323-327 /1988/ вкл. в ссылки.
Как показано выше, последовательности ДНК будут выражены в хозяине только после присоединения /т.е. поставлены на места, обеспечивающие функционирование/ к последовательностям, обеспечивающим контроль за экспрессией. Эти векторы экспрессии обычно воспроизводятся в организме хозяина или как эписомы или как интегральная часть хромосомной ДНК хозяина. Обычно векторы экспрессии содержат маркеры селекции, например тетрациклин или неомицин, для распознавания тех клеток, которые подвергались трансформации при участии нужных последовательностей ДНК. /см. например US Patent 4, 704, 362, вел. в ссылки/.
E. coli является одним из прокариотических хозяев, особенно полезных для клонирования последовательностей ДНК в данном изобретении. Другие микробы, пригодные для использования, включая бациллы, такие как Bacillus subtilus и другие энтеробактерии, такие как Salmonella, Serratia и разнообразные Pseudomonas. В данных прокариотических хозяевах можно также получать векторы экспрессии, которые обычно содержат контролирующие экспрессию последовательности, совместимые с клеткой хозяина/ например, происхождение репликации/. Кроме того, может быть представлено любое количество разнообразных промоторов, таких как система промотора лактозы, система промотора триптофана /Трп/ система промотора бета/лактамазы или система промотора для фага лямбда. Промоторы обычно контролируют экспрессию, альтернативно с последовательностью оператора, и имеют последовательности связывающего рибосомы участка и аналогичные для начала и окончания транскрипции и трансляции.
Для экспрессии можно использовать и другие микробы, такие как дрожжи. Saccharomyces является предпочтительным хозяином с подходящими векторами, имеющими последовательности контроля экспрессии, такими как промоторы, включая 3- фосфоглицерат киназу или другие гликолитические ферменты, и подходящим происхождением последовательностей репликации, обрыва и т.п.
Кроме микроорганизмов, можно также использовать клеточную культуру тканей млекопитающих для экспрессии и производства полипептидов по данному изобретению. /см. Winnacker "From benes to clones", VCH Publishers, N. Y. (1987), вкл. в ссылки. Предпочтительнее эукариотические клетки, так как в данной области известен ряд удобных клеточных линий-хозяев, способных к выделению интактных иммуноглобулинов, и включающий линии клеток CHO, разнообразные линии клеток COS, клетки HELa, линии клеток миеломы, и т.д., но предпочтительнее трансформированные B-клетки гидридом. Векторы экспрессии для этих клеток могут включать последовательности контроля экспрессии, такие как происхождение репликации, промотор, усилитель /Queen, C. et. al. Immund. Pev. 89:49-68, /1986/ и необходимо, чтобы они обладали информационными участками, такими как участок связывания рибосом, участок сращивания РНК, участки полиаденилирования, и транскрипционные последовательности терминатора. Предпочтительными последовательностями контроля экспрессии являются промоторы, полученные из SV40 с усилителем /см. Milligan and Beng, Science:209: 1422-1427 /1980//, ген иммуноглобулина, аденовирус, вирус Bonne Papilloma и т.д.
Векторы, содержащие интересующие нас сегменты ДНК /например, кодирующие последовательности тяжелых и легких цепей/, могут быть перенесены в клетку хозяина с помощью хорошо известных способов, которые зависят от клеточного типа хозяина. Например, для прокариотных клеток обычно используют трансфекцию CaCl2, в то время как для других клеточных хозяев можно применять обработку фосфатом кальция или электропорацию /см. в осн. Manuatis, et. al., Molecular cloning: a Laboratory Manual, Cold Spring Harbor Press, (1982), вкл. в ссылки/. После экспрессии, целые антитела, их димеры, индивидуальные тяжелые или легкие цепи можно очистить по стандартной методике, включающей осаждение сульфатом аммония, аффинные колонки, колончатую хроматографию, гель-электрофорез и т.п. /см. в осн. Scopes R., Protein Purifucation, Springer-Verlag, N.Y., /19823//.
Предпочитают существенно очищенные иммуноглобулины с гомогенностью по крайней мере около 90 до 95%, а лучше с гомогенностью около 98 до 99 или больше для фармацевтических целей. После очистки частичной или до гомогенного состояния, по желанию, полипептиды можно затем использовать в терапии /в т. ч. и для наружного применения/ или для развития и проведения анализов, флуоресцентного окрашивания и т.п. /см. в осн. Immunological Methods, Vols I and II, Lefcovits and Pernis, eds. Academic Press, New York, N.Y. (1979 and 1981)/.
Представленные в данном изобретении антитела, специфичные к IL-2 рецептору найдут индивидуальное применение при лечении расстройств, передаваемых Т-клетками. В общем, там, где были опознаны связанные с заболеванием клетки, несущие IL-2 рецептор, пригодны человекоподобные антитела, способные блокировать связывание IL-2 с IL-2 рецептором человека. /см. U.S.S.N 085, 707, заглавие: "Treatig Human Malignaneies and Disordens, вкл. в ссылки/. Например, типичные болезненные состояния, поддающиеся лечению, включают расстройства, вызванные взаимодействием трансплантата с хозяином и отторжение трансплантата у пациентов, перенесших трансплантацию органа, такого как сердце, легкие, почки, печень и т.д., другие заболевания, включая автоиммунные, такие как диабеты Типа I, рассеянный склероз, ревматоидный артрит, системная волчанка и миастения.
Антитела, подобные человеческим, по данному изобретению можно также использовать в сочетании с другими антителами, в особенности, с моноклональными антителами человека, реактивными по отношению к другим маркерам на клетках, ответственных за заболевания. Например, подходящие маркеры Т-клеток могут включать их, сгруппированных в так называемые "Кластеры дифференциации" /Clustens of Differentiation/, термин введен на First International Leukocyte Differentiation Workshop, Leuksocyte Typing Bernard et al., Eds., Springer - Verlag, N.Y. (1984), вкл. в ссылки/. Антитела можно также использовать как отдельные композиции, даваемые совместно с химиотерапевтическими или иммуносупрессивными агентами. Обычно эти агенты включают циклоспорин A или пуриновый аналог /метотрексат, 6-меркаптопурин или аналог/, но можно также использовать многочисленные добавочные агенты, хорошо известные специалистам /например, циклофосфамид, преднизолон и т.п./.
Предпочтительная фармацевтическая композиция по данному изобретению включает использование зависимых антител в иммунотоксинах. Иммунотоксины характеризуются двумя компонентами и особенно полезны для умерщвления выбранных клеток in vitro или in vivo. Один компонент - это цитотоксичный агент, который обычно убивает клетки при присоединении или поглощении его. Другой компонент, известный как "механизм доставки", обеспечивает способы для доставки токсичного агента к особому типу клеток, такому как клетки, включающие карциному. Два компонента обычно химически связаны вместе с помощью многочисленных хорошо известных химических способов. Например, когда цитотоксичный агент - протеин, а второй компонент - интактный иммуноглобулин, связывание можно провести с помощью гетеробифункиональных сшивателей, например SPDP, карбодиимида, альдегида глутаровой кислоты и т.п. Получение различных иммунотоксинов хорошо известно и способы его можно найти, например, в "Monoclonal Antibody - Toxin Conjugates: Aiming the Magu Bullet." Thorpe et al. , Monoclonal Antibodies in chinical Medicine, Academic Press. pp. 168 - 190 (1982), вкл. в ссылки/.
Для использования в иммунотоксинах пригоден целый ряд цитотоксичных агентов. Цитотоксичные агенты могут включать радионуклиды, такие как Иод-131, Иттрий-90, Рений-188 и Висмут-212, ряд химиотерапевтических средств, таких как виндесин, метотрексат, адриамицин и циспластин, и цитоксичные протеины, такие как ингибирующие протеины рибосом, подобные антивирусному протеину фитолакки, экзотоксин A Pseudomonas, рицин, токсин дифтерии, цепь рицина A и т.п. или агенты, активные на поверхности клетки, такие как ферменты фосфолипазы /например, фосфолипаза C/ /см. в основном U.S.S.N. 07/290, 968 пол. в Дек. 28, 1988//, "Chimeric Toxins", Olsnes and Phil., Pharmac. Ther.: 25:355 - 381 (1982), and "Monoclonal Antibodies for Cancer Detection and Therapy", eds. Baldink and Byers. DP. 159 - 179, 224 - 266, Academic Press (1985),
Доставочный компонент иммунотоксина будет включать иммуноглобулины, подобные человеческим по данному изобретению. Предпочтительно использованы интактные иммуноглобулины или их связывающие фрагменты, такие как Fab. Обычно антитела в иммунотоксинах будут относится к изотипам IgM или IgG человека, но при желании можно использовать постоянные участки других млекопитающих.
Антитела, подобные человеческим, и их фармацевтические композиции по данному изобретению особенно полезны для внутреннего применения, т.е. подкожно, внутримышечно или интравенозно. Композиции для внутреннего применения будут обычно включать раствор антитела или смесь их, растворенную в приемлемом носителе, предпочтительно в воде. Можно использовать ряд носителей, например воду, воду с буфером, 0,4% pp соли, 0,3% глицин и т.п. Эти растворы стерильны и свободны от примесей посторонних.
Данные составы можно стерилизовать с помощью хорошо известных методов. Составы могут содержать фармацевтически приемлемые дополнительные вещества в соответствии с необходимостью приближения к физиологическим условиям, такие как регуляторы pH и буферы, агенты, регулирующие токсичность и т.п., например ацетат натрия, хлорид натрия, хлорид калия, лактат натрия и т.д. Концентрация антитела в этих составах может широко изменяться, например, от менее чем около 0,5%, обычно от или по крайней мере 1% до не менее 15 или 20 весовых % и ее выбирают, основываясь на объеме жидкости, вязкостях и т.п. в соответствии с выбранным способом введения.
Так, обычная фармацевтическая композиция для внутримышечных инъекций может содержать 1 мл стерильной воды с буфером и 50 мг антитела. Типичная композиция для внутривенного вливания может состоять из 250 мл стерильного раствора Рингера и 150 мг антитела. Применяемые способы получения композиций для внутреннего употребления известны или знакомы специалистам и более подробно описаны, например, в Remington's Pharmaceutical Science, 15 th ed., Mack. Publishing Co., Eeaston, Pennsylania (1980) вкл. в ссылки/.
Антитела по данному изобретению можно подвергать лиофилизации и затем восстановить в необходимой среде перед использованием, можно использовать способы, хорошо известные для лиофилизации и восстановления, так как данный способ хорошо себя зарекомендовал для иммуноглобулинов. Специалисты должны учитывать, что лиофилизация и восстановление могут привести к изменению падения активности антитела в различной степени, например, с распространенными иммуноглобулинами, у IgM антител большее падение активности, чем у IgC антител, и поэтому используемые уровни нуждаются в регулировании с целью компенсации.
Композиции, содержащие данные антитела, подобные человеческим или их смесь, можно применять для профилактики и/или лечения. При лечении композиции применяют для пациента, уже страдающего от болезни, в количествах, достаточных для вылечивания или по крайней мере остановки, прогрессирования болезни и ее осложнений. Количество, необходимое для этого, определяют как терапевтически эффективную дозу". Количества, эффективные для данного применения, зависят от серьезности инфекции и от общего состояния собственной иммунной системы пациента, но обычно наиболее часто используют, в основном, от 1 до 200 мг антител на дозу с дозировкой от 5 до 25 мг на пациента. Необходимо учитывать, что материалы по данному изобретению можно, в основном, применять при серьезных заболеваниях, когда существует реальная или потенциальная угроза жизни. В таких случаях, ввиду сведения к минимуму периферических веществ и снижения вероятности "чужеродных веществ", что достигается применением антител, подобных человеческим по данному изобретению, вероятно и возможно желательно для лечащего врача применять существенный избыток этих антител.
В профилактических целях композиции, содержащие данные антитела или их смесь, вводят в пациента, еще не заболевшего, для усиления его сопротивляемости. Такое количество определяют как "профилактически эффективную дозу". В этом случае также точные количества зависят от состояния здоровья пациента и общего уровня его иммунитета, но обычно от 0,1 до 25 мг на дозу, от 0,5 до 2,5 мг на пациента. Предпочтительное использование в целях профилактики для предотвращения отторжения трансплантированной почки.
Единичное (или множественные) применение композиций должно быть выбрано лечащим врачом с учетом дозировки и способа, в любом случае фармацевтические препараты должны обеспечить количество антитела /антител/ по данному изобретению, достаточное для эффективного лечения пациента.
Антитела, подобные человеческим, по данному изобретению могут в дальнейшем найти широкое применение in vitro. Например, данные антитела можно использовать для типирования T-клеток, для выделения специфических, несущих IL-2 рецептора клеток или фрагментов рецептора, для получения вакцин и т.п.
Для целей диагностики антитела могут быть помечены или не помечены. Непомеченные антитела можно использовать в комбинации с помеченными антителами /вторыми антителами/, которые реактивны по отношению к антителу, подобному человеческому, такие как антитела, специфичные к постоянным участкам иммуноглобулина человека. С другой стороны, антитела можно пометить непосредственно. Можно использовать большое разнообразие меток, таких как радионуклиды, фтор, ферменты, субстраты энзимов, кофакторы энзимов, ингибиторы энзимов, лиганды /особенно гаптены/ и т.п. Специалистам известны разные типы доступных иммунных анализов.
В наборы также могут входить определенные антитела для защиты или распознавания клеточной активности или для присутствия выбранного антигена. Так определенная композиция антител по данному изобретению может быть получена, обычно в лиофилизированной форме в контейнере, как сама по себе, так и в соединении с дополнительными антителами, специфичными для нужного типа клеток. Антитела, которые можно соединить с меткой или токсином, или неприсоединенные, включены в наборы с буферами, такими как трис-, фосфат, карбонат и т. п., стабилизаторами, биоцидами, инертными протеинами, например сывороточным альбумином, и т.п., и с инструкциями к применению. В основном данные вещества составляют не более 5 вес.% по отношению к количеству активного антитела и обычно присутствуют в общем количестве, не превышающем 0,001% по весу по отношению к концентрации антитела. Часто желательно включать инертный расширитель или эксипиент для разбавления активных ингредиентов, где эксипиент может составлять от около 1 до 99 вес.% от всей композиции. Там, где применяют второе антитело, способное к связыванию с химерным антителом, для анализа, оно обычно находится отдельно. Второе антитело обычно связано с меткой и оформлено аналогично, как и описанное выше.
С целью иллюстрации ниже приведены примеры, которым не ограничивается изобретение.
Примеры
Экспериментальная часть
1. Конструирование генов для человекоподобных легких и тяжелых цепей
Последовательность человеческого антитела Eu /Seguences of Proteins of Immunological Interent, Kabet. E. , et al., US. Dept. of Health and Human Services, 1983) использовали для получения основы гуманизированного антитела, так как аминокислотная последовательность тяжелой цепи анти-Тас более гомоглогична к тяжелой цепи данного антитела, чем к любой другой тяжелоцепной последовательности из National Biomedical Foundation Ptotein Identification Recource.
Для выбора последовательности гуманизированной тяжелой цепи тяжелоцепную последовательность анти-Тас /см. обычно U.S.S.Ns 186.862 and 223.037; вкл. в ссылки/ сравнили с последовательностью Eu тяжелой цепью /фиг. 1/. Для каждого положения была выбрана Eu аминокислота для гуманизированной последовательности, если только эта позиция не попадает в одну из следующих категорий, в таком случае выбирали аминокислоту анти-Тас.
/1/ Позиция попала внутрь участка, определяющего комплементарность /CDR/, как опр. Kabat et al. op. cit /аминокислоты 31-55, 50-66, 99-106/.
/2/ Eu аминокислота необычна для тяжелых цепей человека в данном положении, в то время как анти-Тас аминокислота была типична для тяжелых цепей человека в данном положении /аминокислоты 27, 93, 95, 98, 107-109, 111/.
/3/ Положение было непосредственно около CDR в аминокислотной последовательности тяжелой цепи анти-Тас /аминокислоты 30 и 67/.
/4/ Трехмерное моделирование анти-Тас антитела позволило предположить, что аминокислота была физически вблизи участка связывания антигена /аминокислоты 48 и 68/.
Некоторые аминокислоты попали в более чем одну категорию, но отмечены только один раз.
Для выбора последовательности гуманизированной легкой цепи анти-Тас легкоцепные последовательности сравнили с последовательностями Eu легкой цепью /фиг. 2/. В каждом положении была выбрана e аминокислота, если только положение опять не попало в одну из категорий /1/ - /4/, /везде в категориях следует заменить термины на легкие цепи/:
/1/ CDR'S /аминокислоты 24-34, 50-56, 89-97/.
/2/ Анти-Тас аминокислота более типична, чем Eu аминокислоты 48 и 63.
/3/ Близость к CDR'S /нет аминокислот, Eu и анти-Taс уже не отличались во всех этих позициях/
/4/ Возможное приближение в объеме к связующему участку /аминокислота 60/.
Нуклеотидные последовательности тяжелой /фиг. 3/ и легкой /фиг. 4/ цепи генов выбраны следующим образом:
/1/ Код нуклеотидных последовательностей для аминокислотных последовательностей выбран как описано выше.
/2/ 5' этих кодирующих последовательностей, код нуклеотидной последовательности для первой /сигнальной последовательности, а именно лидера легкой цепи антитела МОРС 63 и лидера тяжелой цепи антитела PCH 108A /Kabat et al. Op. cit/. Эти лидерные последовательности выбраны как типичные у антитела.
/3/ 3' кодирующих последовательностей, нуклеотидные последовательности здесь это последовательности, которые следуют за сегментом J 5 легкой цепи мыши и сегментов J 2 тяжелой цепи мыши, которые являются частью последовательностей анти-Тас. Эти последовательности включены, так как они содержат сплетение донорских сигналов.
/4/ На каждом конце последовательности расположен участок Хва 1 для обрыва на участках Хва 1 клонирования в Хва 1 сайт вектора.
2. Конструирование гуманизированных генов тяжелой и легкой цепи
Для синтеза тяжелой цепи и были синтезированы четыре олигонуклеотида HES 12, HES 13, HES 14, HES 15 /фиг. 5A/, используя синтезатор ДНК для прикладных биосистем 380 В. Два из олигонуклеотидов являются частью каждого тяжа на тяжелой цепи, и каждый олигонуклеотид перекрывает следующий около 20 нуклеотидами, делая возможной ренатурацию /фиг. 5B/. Вместе олигонуклеотиды покрывают всю гуманизированную тяжелую цепь /фиг. 3/ с несколькими дополнительными нуклеотидами на каждом конце для обеспечения обрыва на сайтах Хва 1. Олигонуклеотиды были очищены на полиакриламидных гелях.
Каждый нуклеотид фосфорилировали АТФ и Т4 полинуклеотид киназу по стандартной методике /см. Maniatis, op. cit./. Для ренатурации фосфорилированных олигонуклеотидов их суспендировали вместе в 40 мкл ТА / 33 мМ трис-ацетата pH 7,9, 66 мМ ацетата калия 10 мМ ацетата магния/ при концентрации около 3,75 мкл для каждого, нагревали до 95oС за 4 минуты и медленно охлаждали до 4oС. Для синтеза полного гена из олигонуклеотидов путем синтеза противоположного тяжа каждого олигонуклеотида /фиг. 5B/, добавили следующие компоненты в конечный объем 100 мкл:
10 мкл ренатурированные олигонуклеотиды
0,16 мкл каждый дезоксирибонуклеотид
0,5 мкл АТФ
0,5 млк ДТТ
100 мкг/мл BSA
3,5 мкг/мл T4g43 протеин /ДНК полимераза/ 25 мкг/мл T4 g 44/ 62 протеин /добавочный протеин полимеразы/. 25 мкг/л 45 протеин /добавочный протеин полимеразы/.
Смесь выдерживали при 37oС 30 мин. Затем добавили 10 мкл Т4 ДНК лигазы и выдерживали при 37oС 30 мин. Полимераза и лигаза были инактивированны при инкубации при 70oС за 15 мин. Для усвоения геном Хва 1 в реак. смесь добавили 50 мкл 2 х ТА, содержащего BSA в конц. 200 мкг/мл и ДТТ 1 мМ, 43 мкл воды и 50 мкл Хва 1 в 5 мкл. Выдерживали 3 часа при 37oС и пропустили через гель. Фрагмент вр Хва 1 был очищен от геля и клонирован в сайт Хва 1 плазмиды pU C19 стандартным способом. Четыре изолята плазмид были очищены и разделены на последовательности с использованием дидеокси метода. Одна из них имела правильную последовательность /фиг. 3/.
Для синтеза легкой цепи были синтезированы четыре олигонуклеотида JFD1, JFD2, JFD3, JFD4 /фиг. 6a/. Два из олигонуклеотидов являются частью каждого тяжа на легкой цепи и каждый олигонуклеотид перекрывает следующий около 20 нуклеотидами для обеспечения ренатурации /фиг. 6B/. Все вместе олигонуклеотиды перекрывают всю гуманизированную легкую цепь /фиг. 4/ с несколькими дополнительными нуклеотидами на каждом конце для обеспечения обрыва на сайтах Хва 1. Олигонуклеотиды были очищены на полиакриламидных гелях.
Легкоцепной ген был синтезирован из данных олигонуклеотидов из двух частей. По 0,5 мкг каждого из JFD1 и JFD2 соединены в 20 мкл секвеназного буфера /40 мМ трис-РС, pH 7,5: 20 мМ хлорида магния, 50 мМ хлорида натрия/, нагреты до 70oС за три минуты и оставлены медленно остывать до 23oС для обеспечения ренатурации олигонуклеотидов. С JFD3 и JFD4 поступили таким же образом. Каждая реакция проводилась для 10 мМ в ДТТ и 0,5 мМ в каждом дезоксирибонуклеотиде и 6,5 мкл секвеназы /US Biochemicals/ в окончательном объеме 24 мкл и выдержке 1 час при 37oС для синтеза противоположных тяжей олигонуклеотидов. В каждую реакцию добавили Xвa I и Hind III для усвоения ДНК /т.к. существует сайт Hind III в области, где перекрываются JFD2 и JFD3 и, следовательно, в каждой из синтезированных ДНК, фиг. 6B/. Реакцию проводили на полиакриламидных гелях, фрагментом Хва I - Hind III были очищены и клонированы в pUC18 с помощью стандартных методов. Несколько изоляторов плазмид для каждого фрагмента были получены дидеокси методом и выбраны правильные.
3. Конструирование плазмид для экспрессии гуманизированных тяжелых и легких цепей
Тяжелоцепной фрагмент Хва I был извлечен из плазмиды pUC19, в которую он был внесен, и затем внесен в сайт Хва I вектора pVγ1 /см. обычно U.S.S.N 223, 037/ с правильной ориентацией с помощью стандартных методов для получения плазмиды pHii GTAСl /фиг. 7/. Эта плазмида будет выражать достаточно точно полную тяжелую цепь, будучи перенесенной в соответствующую клетку хозяина.
Два легкоцепных фрагмента Хва I - Hind III были выделены из плазмид pUC18, в которые они были внесены. Вектор плазмиды pVk1 /см. обычно U.S.S.N 223, 037/ был удален вместе с Хва I, дефосфорилирован и связан с двумя фрагментами по стандартной методике. Желаемый продукт реакции имел замкнутую форму: вектор - Хва-фрагмент I - Hind - III - фрагмент 2-Хва I - вектор. Несколько изолятов плазмид проанализировали с помощью рестрикции и секвенирования и выбрали один с данной формой. Следовательно, данная плазмида pHuLTAC /фиг. 8/ содержит полную гуманизированную легкую цепь /фиг. 4/ и будет выражать с высокой степенью точности легкую цепь послевнесения в соответствующую клетку хозяина.
4. Синтез и аффинность гуманизированного антитела
Плазмиды pHu и GTAC и pHuLTAC были перенесены в клетки мыши Sp2/0, и клетки, в которые были включены плазмиды, выбрали по их устойчивости к микрофенольной кислоте и/или гидромицину B, связанной с генами gpt и hyg на плазмиде /фиг. 7, 8/, по стандартным методикам. Для проверки, действительно ли эти клетки производят антитела, связывающиеся с IL-2 рецептором, клеточный экстракт инкубировали с клетками HUT-102, которые, как известно, производят IL-2 рецептор. После промывки клетки инкубировали с флуоресцентно связанным козьим античеловеческим антителом, промывали и смотрели флуоресценцию на флоуриметре FACSCAN. Результаты ясно показывают, что гуманизированное антитело связывается с этими клетками, но не с Т-клетками Jurkat, которые не выражают IL-2-рецептор /фиг. 3/. В качестве контроля также использовали настоящие анти-тас антитела мыши для окрашивания этих клеток /фиг. 2 9B, C/, дающие похожие результаты.
Для продолжения эксперимента клетки, производящие гуманизированное антитело, ввели в мышей и результаты обработали. Гуманизированное антитело очистили до существенной гомогенности, пропустив через колонку /аффинную/ с антителом с иммуноглобулином козы - античеловеческим, приготовленным на подложке Affigel-10 /Bio-Rad Laborayories, Inc., Richmond, CA/ в соответствии со стандартными способами. Для определения аффинности гуманизированного антитела по сравнению с исходным анти-Тас антителом провели сравнительный эксперимент по связыванию. Около 5•105 клеток HUT-102 инкубировали с известными количествами /10-40 нг/ анти-Тас антитела и гуманизированным анти-Тас антителом 10 мин при 4oС. Затем 100 нг биотинилированного анти-Тас добавили к клеткам и инкубировали 30 мин при 4oС. Это количество анти-Тас предварительно определили как достаточное для насыщения связывающих участков на клетках, но не являющееся значительным избытком. Затем клетки дважды промыли 2 мл буферного раствора соли /PBS/, содержащим 0,1% азида натрия. Затем клетки инкубировали 30 мин при 4oС с 250 нг авидина, конъюгированного с фикоэритрином, который связался с биотинилированным анти-Тас, уже присоединенным к клеткам. Клетки опять были промыты, как указано выше, зафиксированы в PBS, содержащем 1% параформальдегида, и проанализированы на флуоресценцию на цитофлуориметре FACSCAN.
Использование увеличивающихся количеств /10-40 нг/ анти-Тас антитела, как конкурента на первой стадии, уменьшило количество биотинизированного анти-Тас, который мог бы связаться с клетками на второй стадии, и следовательно, количество авидина, конъюгированного сфикоэритрином, который присоединился на последней стадии, уменьшая тем самым флуоресценцию (фиг. 10A). Эквивалентные количества /20 нг/ анти-Тас и гуманизированного анти-Тас, используемые как конкуренты, уменьшали флуоресценцию до приблизительно такого же уровня (фиг. 10B). Это показывает, что данные антитела имеют примерно ту же аффинность /в пределах 3-4 кратной/ так, как если бы одно из них имело гораздо большую аффинность, оно бы более успешно конкурировало с биотинилированным антителом, таким образом снижая флуоресценцию больше.
5. Биологические свойства гуманизированного антитела
Для оптимального использования при лечении болезней человека гуманизированное антитело должно быть способно разрушать T-клетки в теле, которые выражают рецептор IL-2. Один из механизмов, по которому антитела могут разрушать клетки/мишени, антителозависимая, передаваемая клетками цитотоксичность, сокращенно ADCC /Fundamental Immunology. Paul, W., Ed. Raven Press. N. J. (1984), pp. 681/, в котором антитело образует мостик между клеткой мишенью и клеткой-эффектором, таким как макрофаг, который может лизировать мишень. Для определения может ли гуманизированное антитело и исходное анти-Тас антитело мыши быть медиатором ADCC, был осуществлен по стандартным методикам анализ на высвобождение хрома. В частности, клетки лейкемии человека HUT-102, которые выражают IL-2 рецептор, были инкубированы с 51Cr для поглощения радионуклида. Затем клетки HUT-102 инкубировали с избытком как анти-Тас, так и с гуманизированным анти-Тас антителом. Клетки HUT-102 затем инкубировали 4 часа с клетками эффектора как в соотношении 30:1, так и 100:1, которые являлись нормальными очищенными периферическими моноклеарными клетками крови человека, которые были активированы инкубированием в течение около 20 часов с рекомбинантным IL-2 человека. Освобождение хрома, которое свидетельствовало о распаде клеток-мишеней HUT-102, было измерено и проанализировано. Результаты показывают (таблица), что при любом количестве клеток эффекторов анти-Тас не разрушает значительного количества клеток-мишеней /менее чем 5%/ в то время как гуманизированное антитело разрушает /более 20%/. Так гуманизированное антитело вероятно более эффективно, чем исходное антитело мыши при лечении T-клеточной лейкемии или других расстройств, передаваемых T-клетками.
Из изложенного ясно, что иммуноглобулины, подобные человеческим, по данному изобретению имеют многочисленные преимущества по сравнению с другими антителами. Например, по сравнению с анти-Тас моноклональными антителами мыши данные человекоподобные иммуноглобулины IL-2 рецептора можно получать более экономично и они содержат существенно меньше посторонних аминокислотных последовательностей. Эта уменьшенная вероятность антигенности при введении пациенту представляет существенное улучшение с точки зрения терапии для иммуноглобулинов, полученных согласно критериям, приведенным выше.
Хотя данное изобретение детально описано с помощью иллюстраций и примеров для более точного понимания, очевидно, что могут быть внесены различные изменения и модификации в объеме прилагаемой формулы изобретения.







Изобретение предназначено для получения реномбинантной ДНК и моноклональных антител, применяемых при изготовлении новых терапевтических средств. Реномбинантное антитело специфически взаимодействует с белком р 55 Тас рецептора Интерлейкина-2 человека (ИЛ-2). Указанное антитело ингибирует связывание ИЛ-2 с рецептором ИЛ-2 на Т-клетках человека и имеет сродство к рецептору ИЛ-2 более 108M-1. Оно состоит из легких и тяжелых цепей и содержит участки, определяющие комплементарность (CDRs). Предложены полинуклеотиды, включающие последовательность, кодирующую каркасные участки и участки, определяющие комплементарность связывания с антигеном (CDRs). Также предложены способы получения тяжелых и/или легких цепей рекомбинантного антитела. Изобретение позволяет получать новые терапевтические средства с использованием неиммуногенных антител. 5 c. и 6 з.п.ф-лы, 10 ил., 1 табл.
ингибирует связывание ИЛ-2 с рецептором ИЛ-2 на Т-клетках человека;
имеет сродство к рецептору ИЛ-2 человека более 108 М-1;
состоит из легких и тяжелых цепей, содержащих константные области акцепторного антитела человеческого происхождения, каркасные участки вариабельных областей акцепторного антитела человеческого происхождения и участки, определяющие комплементарность (CDRs), донорного антитела мышиного происхождения, при этом каркасные участки вариабельных областей акцепторного антитела человеческого происхождения представляют собой следующие последовательности:
а) последовательности, которые по крайней мере на 65% гомологичны таковым донорного антитела, и/или
б) последовательности, в которых по крайней мере около трех аминокислот замещены на аминокислоты донорного антитела в определенном положении, если аминокислоты каркасного участка вариабельной области акцепторного антитела человеческого происхождения являются редкими для данного положения, а соответствующие аминокислоты каркасного участка вариабельной области донорного антитела мышиного происхождения являются обычными для данного положения в антителе человеческого происхождения, и/или
в) последовательности, в которых по крайней мере около трех аминокислот замещены на соответствующие по положению аминокислоты донорного антитела, если указанные аминокислоты каркасного участка вариабельной области акцепторного антитела человеческого происхождения непосредственно прилегают к CDRs и/или
г) последовательности, в которых по крайней мере около трех аминокислот замещены на соответствующие по положению аминокислоты донорного антитела, если указанные аминокислоты каркасного участка вариабельной области акцепторного антитела человеческого происхождения находятся в пределах 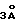 от CDRs в предсказанной трехмерной модели антитела и способны взаимодействовать с антигеном или с CDRs в рекомбинантном антителе.
от CDRs в предсказанной трехмерной модели антитела и способны взаимодействовать с антигеном или с CDRs в рекомбинантном антителе.
а) указанные аминокислоты каркасного участка вариабельной области акцепторного антитела человеческого происхождения являются редкими для данного положения, а соответствующие аминокислоты каркасного участка вариабельной области донорного антитела мышиного происхождения являются обычными для данного положения в антителе человеческого происхождения и/или
б) указанные аминокислоты непосредственно прилегают в CDRs и/или
в) указанные аминокислоты находятся в пределах  от CDRs в предсказанной трехмерной модели антитела и способны взаимодействовать с антигеном или с CDRs в рекомбинантном антителе.
от CDRs в предсказанной трехмерной модели антитела и способны взаимодействовать с антигеном или с CDRs в рекомбинантном антителе.

