Настоящее изобретение относится к биосинтезу рибофлавина, более конкретно к последовательностям ДНК и штаммам Saccharomyces cerevisiae AJ 88, AJ 115, AJ 71, AJ 106, AJ 66 и AJ 121, трансформированным ими, для получения рибофлавина.
Известно получение рибофлавина путем ферментации таких грибов, как Eremothecium ashbyii или Ashbya gossypii (The Merck Index, Windholz и др., изд. Мерк и Ко., стр. 1183, 1983).
В заявке ЕР N 405370 описываются сверхпродуцирующие рибофлавин бактериальные штаммы, которые получают путем трансформации генов биосинтеза рибофлавина из Bacillus subtilis.
Так как генетика биосинтеза рибофлавина в бактериях и эукариотах различна, вышеупомянутые гены из Bacillus subtilis не подходят для разработки рекомбинантного способа получения рибофлавина с помощью эукариотических продуцирующих организмов, таких как Ashbya gossypii.
Задачей изобретения является выделение генов биосинтеза рибофлавина из эукариот с тем, чтобы осуществить рекомбинантный способ получения рибофлавина в продуцирующем эукариотическом организме.
Данная задача решается представленными в конце данного описания последовательностями ДНК, выбранными из группы, включающей последовательность ДНК, кодирующую полипептид с представленной в посл. N 2 аминокислотной последовательностью (ГТФ-циклогидролаза II) или аналог, или производное полипептида согласно поcл. N 2, где одна или несколько аминокислот были удалены, или добавлены, или заменены другими аминокислотами без существенного ослабления ферментативного действия полипептида, при обязательном условии, что последовательность ДНК для аналога или производного полипептида, по крайней мере, на 80% гомологична поcл. N 1, последовательность ДНК, кодирующую полипептид с представленной в поcл. N 4 аминокислотной последовательностью (ДРАПФ-дезаминаза) или аналог, или производное полипептида согласно поcл. N 4, где одна или несколько аминокислот были удалены, или добавлены, или заменены другими аминокислотами без существенного ослабления ферментативного действия полипептида, при обязательном условии, что последовательность ДНК для аналога или производного полипептида, по крайней мере, на 80% гомологична поcл. N 3, последовательность ДНК, кодирующую полипептид с представленной в поcл. N 6 аминокислотной последовательностью (ДБФ-синтетаза) или аналог, или производное полипептида согласно посл. N 6, где одна или несколько аминокислот были удалены, или добавлены, или заменены другими аминокислотами без существенного ослабления ферментативного действия полипептида, при обязательном условии, что последовательность ДНК для аналога или производного полипептида, по крайней мере, на 80% гомологична посл. N 5, последовательность ДНК, кодирующую полипептид с представленной в посл. N 8 аминокислотной последовательностью (ДМРЛ-синтетаза) или аналог, или производное полипептида согласно посл. N 8, где одна или несколько аминокислот были удалены, или добавлены, или заменены другими аминокислотами без существенного ослабления ферментативного действия полипептида, при обязательном условии, что последовательность ДНК для аналога или производного полипептида, по крайней мере, на 80% гомологична посл. N 7, последовательность ДНК, кодирующую полипептид с представленной в посл. N 10 аминокислотной последовательностью (рибофлавин - синтетаза) или аналог, или производное полипептида согласно посл. N 10, где одна или несколько аминокислот были удалены, или добавлены, или заменены другими аминокислотами без существенного ослабления ферментативного действия полипептида, при обязательном условии, что последовательность ДНК для аналога или производного полипептида, по крайней мере, на 80% гомологична посл. N 9 и последовательность ДНК, кодирующую полипептид с представленной в посл. N 12 аминокислотной последовательностью (5-гидрокси-триптофан-редуктаза) или аналог, или производное полипептида согласно посл. N 12, где одна или несколько аминокислот были удалены, или добавлены, или заменены другими аминокислотами без существенного ослабления ферментативного действия полипептида, при обязательном условии, что последовательность ДНК для аналога или производного полипептида, по крайней мере, на 80% гомологична посл. N 11.
Кроме того, данная задача решается штаммами Saccharomyces cerevisiae AJ 88, AJ 115, AJ 71, AJ 106, AJ 66 и AJ 121, трансформированными вышеприведенными последовательностями ДНК, для получения рибофлавина.
Гены и их генные продукты (полипептиды) вводятся в протокол соответствующей последовательности с их первичной структурой и расположены следующим образом:
Последовательность N 1: ген rib 1
Последовательность N2: продукт гена rib 1 (ГТФ-циклогидролаза II)
Последовательность N 3: ген rib 2
Последовательность N 4: продукт гена rib 2 (2,5-диамино-рибитиламино-2,4 (1H,3H)-пиримидин-5-фосфат/ДРАПФ/-дезаминаза)
Последовательность N 5: ген rib 3
Последовательность N 6: продукт гена rib 3 (L-3,4-дигидрокси-2-бутанон-4-фосфат/ДБФ/-синтетаза)
Последовательность N 7: ген rib 4
Последовательность N 8: продукт гена rib 4 (6,7-диметил-8-рибитиллюмазин/ДМРЛ/-синтетаза)
Последовательность N 9: ген rib 5
Последовательность N 10: продукт гена rib 5 (рибофлавин-синтетаза)
Последовательность N 11: ген rib 7
Последовательность N 12: продукт гена rib 7 (5-гидрокситриптофан-редуктаза)
Гуанозин-5'-трифосфат (ГГФ) превращается благодаря ГТФ-циклогидролазе II (продукту гена rib 1) в 2,5-диамино-6-рибозиламино-4-(3H)-пиримидинон-5-фосфат. Это соединение с помощью продукта гена rib 7 сразу восстанавливается в 2,5-диамино-рибитиламино-2,4(1H, 3H)-пиримидин-5-фосфат и затем дезаминируется посредством продукта гена rib 2 в 5-амино-6-рибитиламино-2,4(1H, 3H)-пиримидиндион. После этого в реакцию, катализируемую продуктом гена rib 4, вводится соединение с 4 атомами углерода (ДБФ) и образуется 6,7-диметил-3-рибитиллюмазин (ДМРЛ), из которого при реакции, катализируемой продуктом гена rib 5, получают рибофлавин. Соединение, содержащее 4 атома углерода, ДБФ (L-3,4-дигидрокси-2-бутанон-4-фосфат) образуется из D-рибулозо-5-фосфата при реакции, катализируемой продуктом гена rib 3.
Последовательности ДНК, описанные в посл. N 1, 3, 5, 7, 9, 11, кодируют полипептиды, которые описаны в посл. N 2, 4, 6, 8, 10, 12.
Помимо последовательностей ДНК, названных в протоколе последовательностей, годятся также те, которые вследствие дегенерации генетического кода обладают другой последовательностью ДНК, однако кодируют такой же полипептид.
Далее здесь упоминаются такие последовательности ДНК в соответствии с изобретением, которые кодируют генный продукт (полипептид) с другой, нежели представленная в протоколе последовательностей, первичной структурой, пока генный продукт в основном не будет обладать такими же биологическими свойствами, которыми обладает названный в протоколе последовательностей генный продукт. Под биологическими свойствами подразумеваются прежде всего те, которые способствуют ферментативной активности, обеспечивающей биосинтез рибофлавина.
Такие измененные генные продукты с похожими в основном биологическими свойствами получают при удалении или при добавлении одной или нескольких аминокислот или пептидов, или путем замены аминокислот другими аминокислотами, или могут быть выделены из других организмов, нежели Ashbya gossypii.
Последовательности ДНК, которые кодируют измененные генные продукты, гомологичны последовательностям ДНК согласно протоколу последовательностей, как правило, на 80% и более. Такие последовательности ДНК возможно выделить, исходя из описанных в посл. N 1, 3, 5, 7, 9, 11 последовательностей ДНК, например, с помощью обычных способов гибридизации или с использованием реакции удлинения цепи полимеразой из других, нежели Ashbya gossypii, эукариот. Эти последовательности ДНК гибридизируются в стандартных условиях с описанными в посл. N 1, 3, 5, 7, 9, 11 последовательностями ДНК.
Под стандартными условиями подразумеваются, например, температуры между 42 и 58oC в водном буферном растворе с концентрацией между 0,1 и 1 х SSC/SSC - раствор цитрата и хлорида натрия/(1 х SSC: 0,15 М хлорид натрия, 15 мМ цитрат натрия, pH 7,2). Экспериментальные условия для гибридизаций ДНК описываются в учебниках по генной инженерии, например в книге Sambrook и др. "Molecular Cloning", Cold Spring Harbor Laboratory, 1989.
Регуляторные последовательности, в частности промоторные последовательности, которые расположены в 5'-направлении от последовательностей ДНК, кодирующих соответствующие полипептиды, вводятся в протокол последовательностей и далее объясняются подробнее.
Регуляторная последовательность для гена rib 1:
Последовательность N 1: нуклеотид 1 - 242
Регуляторная последовательность для гена rib 2:
Последовательность N 3: нуклеотид 1 - 450
Регуляторная последовательность для гена rib 3:
Последовательность N 5: нуклеотид 1-314
Регуляторная последовательность для гена rib 4:
Последовательность N 7: нуклеотид 1 - 270
Регуляторная последовательность для гена rib 5:
Последовательность N 9: нуклеотид 1 - 524
Регуляторная последовательность для гена rib 7:
Последовательность N 11: нуклеотид 1 - 352
Регуляторные последовательности могут быть также укорочены еще в 5'- и/или 3'-направлении без существенного ослабления их функции.
Важными для регуляторного действия являются, как правило, фрагменты от 30 до 100, предпочтительно 40 - 70 нуклеотиды из вышеуказанных областей последовательностей.
Эти регуляторные последовательности могут быть также оптимизированы в своей функции, по сравнению с природными последовательностями, с помощью направленного мутагенеза.
Данное изобретение позволяет осуществить рекомбинантный способ получения рибофлавина, в котором трансформированные согласно изобретению штаммы Saccharomyces cerevisiae с вышеуказанным регистрационным номером культивируют известным способом путем ферментации, и образующийся при ферментации рибофлавин выделяют из ферментационной среды и в каждом случае подвергают очистке.
Гены rib и их генные продукты выделяются и охарактеризовываются, как описано в примере и в протоколе последовательностей.
Пример 1
Выделение генов биосинтеза рибофлавина гриба Ashbya gossypii (гены rib)
а) Строение банка комплементарной ДНК (кДНК) Ashbya gossypii
Совокупную РНК экстрагировали в поздней фазе логарифмического роста из мицелия сверхпродуцирующего рибофлавин штамма Ashbya gossypii АТСС 10195 после выращивания на среде YEPD (Sherman и др., "Methods in yeast genetics", Cold Spring Harbor, New-York, 1989).
Поли(A)+ РНК очищали на олиго(тимидин) - целлюлозе путем двухкратной адсорбции и элюции (Aviv и Leder, Proc. Natl. Acad. Sci. USA 69, 1972, 1408-1412). кДНК выделяли в соответствии с известным способом Гублера и Хоффманна (Gene 25, 1983, 263) и добавляли синтетические EcorI-адапторы к концам цепей двуцепочечных молекул кДНК, заканчивающихся парой соединенных комплементарных оснований. Расщепленные EcoRI фрагменты кДНК далее фосфорилировали с помощью полинуклеотидкиназы фага Т4 и клонировали в дефосфорилированном расщепленном EcoRI векторе pYEura3 (фиг. 1). pYEura3 (Clonetech Laboratories, Inc. , штат Калифорния) является экспрессирующим вектором дрожжей, который содержит индуцируемые галактозой промоторы GAL1 и GAL10 и URA, CEN4 и ARS1. Эти элементы дрожжей обеспечивают трансформацию и экспрессию клонированных фрагментов ДНК в клетках дрожжей.
Аликвоты реакции сшивания использовали с целью трансформирования в высококомпетентный (Hanahan, DNA Cloning, изд. D.M. Glover; IRL Press, Oxford 1985, 109) E. coli XL1-Blue (Bullock и др., Biotechniques 5 (1987) 376-378), и трансформанты были подвергнуты селекции на основе их устойчивости к ампициллину.
Около 3 х 105 устойчивых к ампициллину клеток объединяли и амплифицировали и оттуда выделяли плазмиду ДНК (Birnboim и Dolly, Nucleic Acids Res. 7, 1979, 1513).
б) Выделение клонов кДНК Ashbya gossypii, кодирующих образующие рибофлавин ферменты
Клоны кДНК Ashbya gossypii, кодирующие образующие рибофлавин ферменты, были выделены путем функциональной комплементации мутантов Saccharomyces cerevisiae, имеющих отношение к биосинтезу рибофлавина.
Штаммы AJ 88 (Mata leu2 his3 rib1:: URA3 ura3-52), AJ 115 (Matalpha leu2 inos1 rib2:: URA3 ura3-52), AJ 71 (Matalpha leu2 inos1 rib3:: URA3 ura3-52), AJ 106 (Matalpha leu2 inos1 rib4:: URA3 ura3-52), AJ 66 (Mata canR inos1 rib5: : URA3 ura3-52) и AJ 121 (Matalpha leu2 inos1 rib7:: URA ura3-52) являются мутантными штаммами, образующимися при изменении структуры одного из шести генов (от RIB1 до RIB5 и RIB7), которые вовлечены в биосинтез рибофлавина при участии Saccharomyces cerevisiae.
Эти штаммы в каждом случае трансформировались с помощью 25 мкг кДНК из банка кДНК Ashbya gossypii и выращивались на твердой содержащей галактозу среде без рибофлавина. После примерно одной недели роста из чашек с культурой выделяли трансформанты Rib+.
В каждом случае трансформант каждого трансформированного мутанта (Rib1+, Rib2+, Rib3+, Rib4+, Rib5+ и Rib7+) анализировали, и во всех случаях было найдено, что фенотип Rib+ был выражен только в среде с галактозой, а не с глюкозой.
Эти результаты подтверждают, что фенотип Rib+ выражался под контролем индуцируемых галактозой промоторов GAL10 с постоянной плазмидой.
ДНК-плазмида была выделена из трансформантов Rib1+, Rib2+, Rib3+, Rib4+, Rib5+ и Rib7+ в результате трансформации Е.coli и названа pJR715, pJR669, pJR788, pJR733, pJR681 и pJR827.
Частичное секвенирование содержащихся в этих плазмидах инсерций кДНК подтвердило, что они кодируют белки, аналогичные белкам продуктов генов Rib из Saccharomyces.
в) Выделение геномных клонов Ashbya gossypii, кодирующих ферменты, образующие рибофлавин
Для выделения геномных копий образующих рибофлавин генов гриба Ashbya gossypii геномный банк Ashbya gossypii АТСС 10195 упаковывали в космиде superCosl (Stratagene Cloning Systems, Калифорния) и проводили скрининг с помощью 32P-меченных зондов, полученных из копий кДНК генов RIB1, RIB2, RIB3, RIB4, RIB5 и RIB7 гриба Ashbya gossypii.
Космидные клоны с ДНК генов RIB1, RIB2, RIB3, RIB4, RIB5 и RIB7 выделяли путем гибридизации колоний (Grunstein и Hogness, Proc. Natl. Acad. Sci. USA 72, 1975, 3961-3965). Дальнейшие анализы по Саузерну ферментативно расщепленной космидной ДНК с использованием похожих RIB-специфичных зондов кДНК позволили идентифицировать определенные рестрикционные фрагменты, содержащие гены RIB1, RIB2, RIB3, RIB4, RIB5 и RIB7 гриба Ashbya gossypii.
Был найден фрагмент ДНК BamHI-ClaI длиной 3,1 Т.н., который содержал совокупный ген RIB1 гриба Ashbya gossypii, кодирующий ГТФ - циклогидролазу II. Этот фрагмент был выделен из агарозного геля и клонирован в рестрицированной BamHI и ClaI фагмиде pBluescript KS (+) (Stratagene Cloning Systems), при этом образовывалась плазмида pJR765 (фиг. 2).
Была получена последовательность ДНК длиной в 1329 П.н. (посл. N 1), которая содержала RIB1 с открытой рамкой считывания из 906 П.н., 242 П.н. 5'-незакодированного участка и 181 П.н. 3'-незакодированного участка.
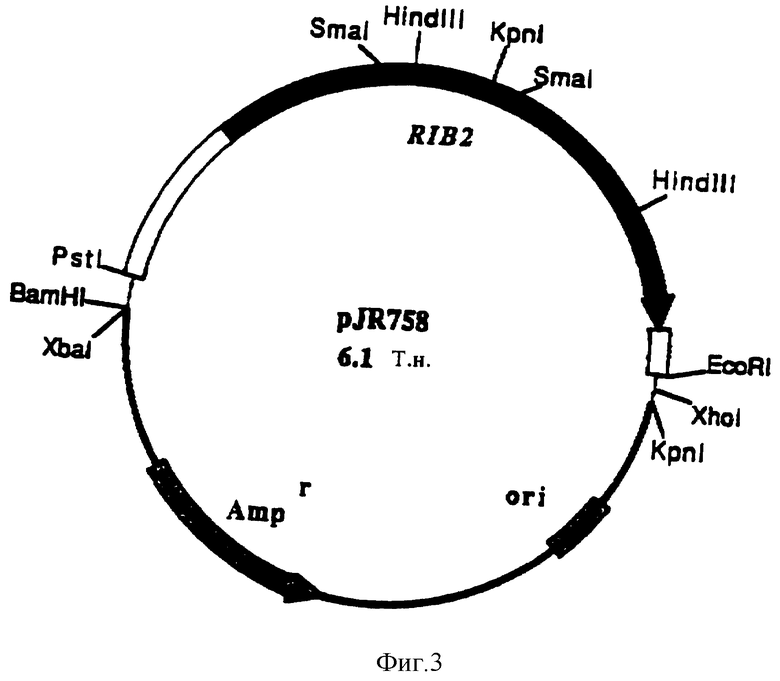
Совокупный ген RIB2 гриба Ashbya gossypii, кодирующий ДРАПФ-дезаминазу, был найден на фрагменте EcoRI-PstI длиной 3,0 Т.н., который клонируется в pBluescript KS (+), образуя плазмиду pJR758 (фиг. 3).
Было проведено определение последовательности участка длиной 2627 П.н. вставки EcoRI-PstI с открытой рамкой считывания гена RIB2 из 1830 П.н., 450 П. н. 5'-нетранслированного участка и 347 П.н. 3'-нетранслированного участка (посл. N 3).
Совокупный ген RIB3 гриба Ashbya gossypii, кодирующий ДБФ-синтетазу, был найден на фрагменте PstI-HindIII длиной 1,5 Т.н., который клонируется в pBluescript K.S (+), образуя плазмиду pJR790 (фиг. 4).
Было проведено определение последовательности участка длиной 1082 П.н. вставки PstI-HindIII с открытой рамкой считывания гена RIB3 из 639 П.н., 314 П. н. 5'-нетранслированного участка и 129 П.н. 3'-нетранслированного участка (посл. N 5).
Ген RIB4 гриба Ashbya gossypii, кодирующий ДМРЛ-синтетазу, был найден на фрагменте PstI-PstI длиной 3,2 Т.н., который клонируется в pBluescript KS (+), образуя плазмиду pJR762 (фиг. 5).
Было проведено определение последовательности участка длиной 996 П.н. вставки PstI-PstI с открытой рамкой считывания гена RIB4 из 519 П.н., 270 П. н. 5'-нетранслированного участка и 207 П.н. 3'-нетранслированного участка (посл. N 7).
Совокупный ген RIB5 гриба Ashbya gossypii, кодирующий рибофлавин-синтетазу, был найден на фрагменте PstI-PstI длиной 2,5 Т.н., который клонируется в pBluescript KS (+), образуя плазмиду pJR739 (фиг. 6).
Было проведено определение последовательности участка длиной 1511 П.н. вставки PstI-PstI с открытой рамкой считывания гена RIB5 из 708 П.н., 524 П. н. 5'-нетранслированного участка и 279 П.н. 3'- нетранслированного участка (посл. N 9).
Наконец, ген RIB7 гриба Ashbya gossypii, кодирующий 5-гидрокситриптофан-редуктазу, был найден на фрагменте EcoRI-EcoRI длиной 4,1 Т.н., который клонируется в pBluescript KLS (+), образуя плазмиду pJR845 (фиг.7).
Было проведено определение последовательности участка длиной 1596 П.н. вставки EcoRI-EcoRI с открытой рамкой считывания гена RIB7 из 741 П.н., 352 П. н. 5'-нетранслированного участка и 503 П.н. 3'-нетранслированного участка (посл. N 11).
Пример 2 Анализ мРНК генов RIB гриба Ashbya gossypii
Для идентификации RIB-специфичных транскриптов проводили Нозерн-блоттинг. Совокупную РНК выделяли из штамма АТСС 10195 гриба Ashbya gossypii, как описано в примере 1. Зонды РНК штамма (5 мкг) разделяли электрофоретически на 0,8%-ных агарозно-формальдегидных гелях вместе с маркерами величины РНК и переносили в вакууме на полиамидную мембрану (Thomas, Proc. Natl. Acad. Sci. USA, 77, 1980, 5201- 5205).
Полиамидные мембраны независимо друг от друга гибридизировали с 32P-меченными RIB-специфичными зондами ДНК при 42oC в 5 х SSC в присутствии 50%-ного формамида. Ген RIB1 гриба Ashbya gossypii выражается в виде единицы генетического кода из примерно 1150 нуклеотидов, что было показано для обоих штаммов с помощью зонда SmaI-SacI длиной 0,7 т.п.н. из плазмиды pJR765 (фиг. 2).
Аналогично были обнаружены в блотах единственные транскрипты RIB2, длиной в 1900 нуклеотидов, RIB3, длиной в 900 нуклеотидов, RIB4, длиной в 800 нуклеотидов, RIB5, длиной в 1050 нуклеотидов, и RIB7, длиной в 1000 нуклеотидов, с помощью фрагмента SmaI-SmaI длиной 0,5 т.п.н. из pJR758, фрагмента HindIII-KpnI длиной 0,6 т.п.н. из pJR790, фрагмента ScaI-HindIII длиной 0,5 т.п.н. из pJR739, фрагмента PstI-PstI длиной 0,3 т.п.н. из pJR845 в качестве специфических зондов.
Пример 3
Экспрессия гена RIB гриба Ashbya gossypii в Saccharomyces cerevisiae
Как описано в примере 1, хорошо изученные мутанты Saccharomyces cerevisiae, дефектные на ступени биосинтеза рибофлавина, могут расти на культуральных средах без рибофлавина, когда они несут плазмиду, кодирующую комплементарные ферменты Ashbya. Для проверки функции продуктов генов RIB гриба Ashbya gossypii, была измерена активность образующих флавины ферментов в бесклеточных экстрактах мутантов S. cerevisiae, которые несли одну из экспрессирующих плазмид pJR715, pJR669, pJR788, pJR733, pJR681 и pJR827.
Описанные в примере 1 и являющиеся производными pYEura3, эти плазмиды содержат RIB-специфичные фрагменты кДНК Ashbya gossypii под контролем индуцируемых галактозой промоторов GAL10.
Бесклеточные белковые экстракты S. cerevisiae получали из культур, выращенных в жидкой среде до оптической плотности около 2.
Клетки собирали, промывали холодным 20 мМ хлоргидратом трис(гидроксиметил)аминометана с pH 7,5 и заново суспендировали в таком же буфере, к которому добавляли 1 мМ фенилэтилсульфонилфторид.
Клеточные лизаты получали при встряхивании со стеклянными шариками и центрифугировании при 3000 g в течение 20 минут при 4oC.
Ферментативную активность ГТФ - циклогидролазы II, ДРАПФ - дезаминазы, ДБФ-синтетазы, ДМРЛ-синтетазы, рибофлавин-синтетазы и 5-гидрокситриптофан - редуктазы определяли, как описано в литературе (Shavlovsky и др., Arch. Microbiol. 124, 1980, 255-259; Richter и др., J. Bacteriol. 175, 1993, 4045-4051; Klein и Bacher, Z. Naturforsch. 35b, 1980, 482-484; Richter и др. , J. Bacteriol. 174, 1992, 4050-4056; Nielsen и др., J. Biol. Chem.261, 1986, 3661; Plaut и Harvey, Methods Enzymol. 18В, 1971, 515-538; Hollander и Brown, Biochem. Biophys. Res. Commun. 89, 1979, 759-763; Shavlovski и др., Biochim. Biophys. Acta, 428, 1976, 611-618).
Количественное определение белка осуществляли по методу Петерсона (Anal. Biochem. 83, 1977, 346-356). Как явствует из Таблицы 1, плазмида pJR715 способствует экспрессии активности ГТФ - циклогидролазы II в мутанте AJ88 S. cerevisiae. Далее эта активность наблюдается только в клетках, которые выращивают на среде с галактозой, что указывает на то, что экспрессия гена RIB1 кДНК гриба Ashbya gossypii происходит под контролем индуцируемых галактозой промоторов GAL10.
Эти результаты поэтому и подтверждают, что RIB1 кодирует ГТФ - циклогидролазу в Ashbya gossypii. Аналогичным способом было показано, что в этом грибе RIB2 кодирует ДРАПФ - дезаминазу, RIB3 кодирует ДБФ-синтетазу, RIB4 кодирует ДМРЛ - синтетазу, RIB5 кодирует рибофлавин-синтетазу, RIB7 кодирует 5-гидрокситриптофан - редуктазу (табл. 2 - 6).
| название | год | авторы | номер документа |
|---|---|---|---|
| УРЕОГИДРОЛАЗЫ В КАЧЕСТВЕ ДОМИНАНТНЫХ СЕЛЕКТИВНЫХ МАРКЕРОВ У ДРОЖЖЕЙ | 2015 |
|
RU2702766C2 |
| КЛЕТКА ДЛЯ ПОЛУЧЕНИЯ ГЕТЕРОПРОТЕИНОВ И СПОСОБ ПОЛУЧЕНИЯ НА ЕЕ ОСНОВЕ | 2008 |
|
RU2494148C2 |
| СПОСОБ ПОЛУЧЕНИЯ РИБОФЛАВИНА, ШТАММ BACILLUS SUBTILIS - ПРОДУЦЕНТ РИБОФЛАВИНА (ВАРИАНТЫ) | 2002 |
|
RU2261273C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pZEN16 ДЛЯ ПЕРЕНОСА И ЭКСПРЕССИИ ГЕНОВ В МИЦЕЛИАЛЬНОМ ГРИБЕ ACREMONIUM CHRYSOGENUM | 2009 |
|
RU2434944C2 |
| СПОСОБ ТРАНСФОРМАЦИИ ДРОЖЖЕЙ CANDIDA UTILIS, ШТАММ ДРОЖЖЕЙ CANDIDA UTILIS (ВАРИАНТЫ), РЕКОМБИНАНТНЫЙ ДНК МАТЕРИАЛ ДЛЯ ТРАНСФОРМАЦИИ ШТАММА ДРОЖЖЕЙ CANDIDA UTILIS, КОДИРУЮЩАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК ГЕНА HIS3 УКАЗАННЫХ ДРОЖЖЕЙ | 1997 |
|
RU2235127C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pPBS-St9, КОДИРУЮЩАЯ ПОЛИПЕПТИД СОМАТОТРОПИНА, И ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE ДЛЯ ПРОДУКЦИИ РЕКОМБИНАНТНОГО СОМАТОТРОПИНА | 2011 |
|
RU2465315C1 |
| СПОСОБ ПОЛУЧЕНИЯ ГЕТЕРОЛОГИЧНОГО БЕЛКА | 1994 |
|
RU2143495C1 |
| Фосфолипаза А2 для экспрессии в дрожжах (варианты) | 2018 |
|
RU2716087C1 |
| ВАРИАНТЫ СУБТИЛАЗ | 2010 |
|
RU2651525C2 |
| ДРОЖЖЕВОЙ ЭКСТРАКТ, СОДЕРЖАЩИЙ ГАММА-Glu-X ИЛИ ГАММА-Glu-X-Gly, И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2010 |
|
RU2496864C2 |


























Изобретение относится к генной инженерии. Установлена последовательность ДНК, кодирующая полипептид с аминокислотной последовательностью, выбранной из группы, включающей последовательности согласно номерам 2,4,6,8,10 и 12, их аналоги и производные, у которых одна или несколько аминокислот были удалены, или добавлены, или заменены другими аминокислотами без существенного ослабления ферментативного действия полипептида, при обязательном условии, что их гомологичность в отношении указанных последовательностей равна по меньшей мере 80%. Данными последовательностями трансформированы штаммы Saccharomyces cerevisiae AJ 88, AJ 115, AJ 71, AJ 106, AJ 66 и AJ 121, служащие для получения рибофлавина. Изобретение позволяет осуществить рекомбинантный способ получения рибофлавина в продуцирующем эукариотическом организме. 12 с.п.ф-лы, 7 ил., 6 табл.
| EP 405370 A, 02.01.91 | |||
| Система регулирования температурного режима прямоточного котла | 1974 |
|
SU569806A1 |
| Способ конструирования рекомбинантных векторов УСR р 2,УСR р 3 и УСR р 4 | 1985 |
|
SU1364241A3 |
