Область изобретения
Изобретение относится к областям молекулярной биологии и генетической инженерии микроорганизмов, в особенности дрожжей.
Введение
Метаболизм аргинина является предметом интенсивных биохимических исследований. После открытия цикла мочевины для использования аргинина в качестве источника азота (Krebs and Henseleit, (1932) Wochenschrift 11: 757-759; Krebs, (1973) Biochemical Education 1: 19-23) внимание сфокусировалось на его роли в качестве предшественника для синтеза полиамина и сигнальных соединений γ-аминобутирата (ГАМК) и оксида азота (Knowles and Moncada (1994) Biochem J 298: 249-258; Pitkanen et al., (2001) Biochem Biophys Res Commun 287: 1051-1057). Наиболее распространенный путь деградации аргинина, встречающийся среди всех трех царств (Abdelal, (1979) Annu Rev Microbiol 33: 139-168), инициируется аргиназой (EC 3.5.3.1), уреогидролазой, превращающей аргинин в орнитин и мочевину. Ее активный центр, содержащий несколько участков связывания Mn2+, также является консервативным в других уреогидролазах, таких как агматиназа (EC 3.5.3.11), формиминоглутамаза (EC 3.5.3.8) и проклаваминат-амидиногидролаза (EC 3.5.3.22), гуанидинобутираза (EC 3.5.3.7) и гуанидинопропионаза (EC 3.5.3.17) (Ouzounis and Kyrpides, (1994) J Mol Evol 39: 101-104). Считают, что гены, кодирующие эти ферменты, возникли на ранних этапах эволюции (Hartman, (1975) Orig Life 6: 423-427), и их используют в качестве маркеров в филогенетических исследованиях (Ouzounis and Kyrpides (1994) J Mol Evol 39: 101-104; Sekowska et al., (2000) Microbiology 146: 1815-1828).
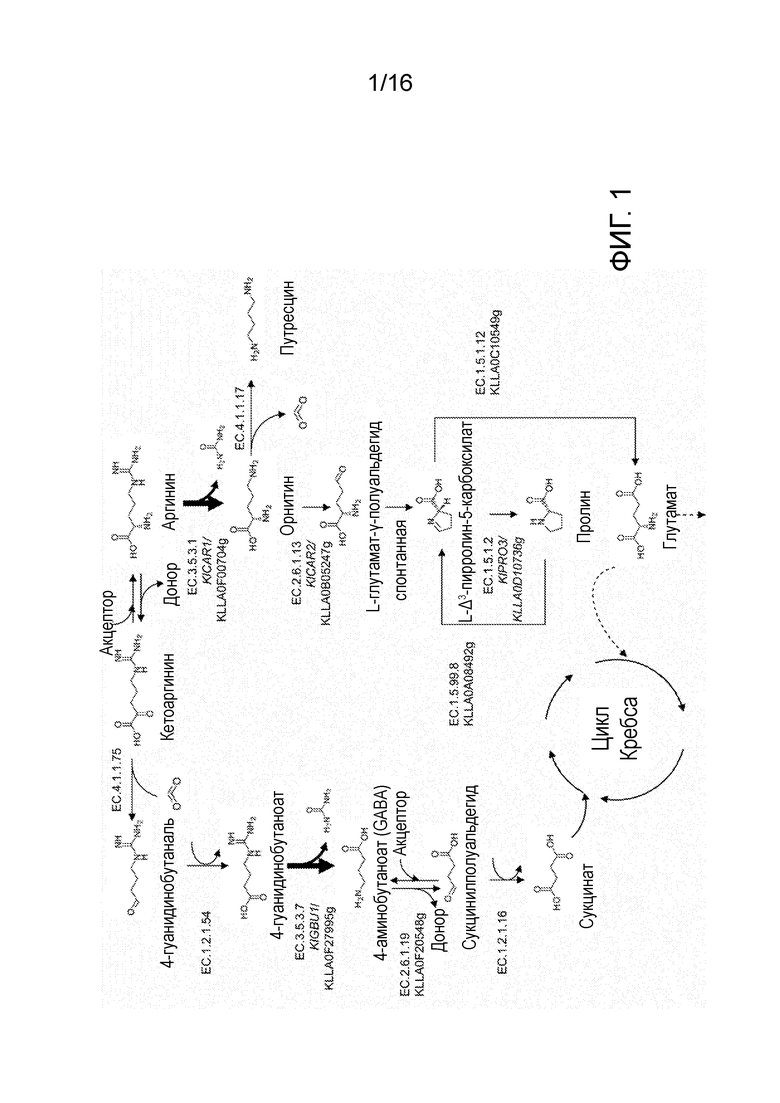
Ранее у эукариот описано только два типа уреогидролаз. В дополнение к аргиназе, высшие эукариоты экспрессируют агматиназу (Coleman et al., (2004) Biochem J 379: 849-855), участвующую в альтернативном пути катаболизма аргинина (фигура 1). В этом пути аргинин сначала декарбоксилируется до агматина, превращаемого агматиназой в путресцин и мочевину. Затем путресцин может превращаться в ГАМК или в полиамины спермин и спермидин (Pegg, (2009) IUBMB Life 61: 880-894). Быстрорастущее количество полных последовательностей геномов делает возможным предполагаемую идентификацию генов аргиназы и агматиназы у многих эукариот. Однако, т.к. такое аннотирование основано исключительно на гомологии последовательностей, это не позволяет сделать окончательные выводы о каталитической функции кодируемых белков.
Значительная часть знания о метаболизме аргинина у грибов основана на исследованиях с использованием модельного организма Saccharomyces cerevisiae. У S. cerevisiae аргинин транспортируется в клетку, а затем гидролизуется аргиназой (Car1) для получения орнитина и мочевины (Sumrada and Cooper, (1992) Yeast 8: 311-314; Cooper et al., (1992) J Bacteriol 700 174: 48-55; Shima 64 et al., (2003) Appl Environ Microbiol 69: 715-718). Затем АТФ-зависимая амидолиаза (Dur1.2) превращает мочевину в аммиак и диоксид углерода. Далее орнитин превращается орнитин-специфической трансаминазой (Car2) в глутамат-γ-полуальдегид (GSA), спонтанно образующий 1-пирролин-5-карбоксилат (P5C) (Martin et al., (2003) Appl Environ Microbiol 69: 1623-1628). В результате субклеточной компартментализации S. cerevisiae неспособны превращать цитозольный P5C напрямую в глутамат (Davis, (1986) Microbiol Rev 50: 280-313). Вместо этого, P5C восстанавливается до пролина с использованием пирролин-5-карбоксилатредуктазы (Pro3). Затем пролин транспортируется в митохондрии (Brandriss and Falvey, (1992) Bacteriol 174: 5176), превращается обратно в P5C оксидазой (Put1) и, в конечном итоге, превращается в глутамат митохондриальной P5C-дегидрогеназой (Put2) (Davis, (1986) Microbiol Rev 50: 280-313). Т.к. проведено очень мало физиологических исследований метаболизма аргинина в дрожжах, не относящихся к Saccharomyces, неизвестно, является ли путь аргиназы, важный для роста S. cerevisiae на аргинине в качестве единственного источника азота (Bossinger and Cooper, (1977) J Bacteriol 131: 163-173), единственным путем катаболизма аргинина у грибов.
S. cerevisiae и Kluyveromyces lactis принадлежат к семейству Saccharomycetaceae. Считают, что эти два родственных вида дрожжей генетически разделились до полногеномной дупликации (WGD), видоизменившей геном S. cerevisiae, кроме того K. lactis рассматривают в качестве подобия пред-WGD предка S. cerevisiae (Dujon, (2010) Nat Rev Genet 11: 512-524). Хотя осуществлено множество исследований, касающихся различий метаболизма сахаров у этих двух видов, различия метаболизма аминокислот не исследовали подробно. Тем не менее, получив полную последовательность генома K. lactis, выявили множество предполагаемых ортологов генов S. cerevisiae, участвующих в метаболизме аргинина (Dujon et al., (2004) Nature 430: 35-44; Souciet et al., (2009) BMC Genomics 13: 517; Dias et al., (2012) BMC Genomics 13: 517).
Селекцию микроорганизма, трансформированного с помощью рекомбинантной ДНК, значительно облегчает использование подходящего селективного маркера. Молекулярный биолог, работающий с Saccharomyces cerevisiae, имеет доступ к большому количеству селективных маркеров (Solis-Escalente et al., (2013) FEMS Yeast Research 13: 126-139). Однако ауксотрофные маркеры и маркеры устойчивости к антибиотикам иногда являются нежелательными, таким образом, количество подходящих маркеров значительно снижается. В конечном итоге, молекулярный генетик остается с ограниченным количеством, если он хочет получить штаммы дикого типа, аллополиплоидные и/или анеуплоидные прототрофные штаммы дрожжей.
Исследование метаболизма аргинина привело к идентификации двух новых доминантных маркеров с "приобретением функции", которые можно использовать, например, для встраивания геномных изменений в микроорганизмы, предпочтительно, в лаборатории, штаммов дрожжей дикого типа и промышленных штаммов дрожжей, включая штаммы S. cerevisiae.
Таким образом, изобретение относится к молекуле нуклеиновой кислоты, содержащей (a) нуклеотидную последовательность, кодирующую селективный маркер гуанидинобутиразу; и/или (b) нуклеотидную последовательность, кодирующую селективный маркер гуанидиноамидгидролазу, при этом нуклеотидная последовательность функционально связана с (гетерологичными) последовательностями промотора и терминатора. Эта молекула нуклеиновой кислоты, также обозначаемая как кассета доминантного маркера, представляет собой удобную систему доминантного селективного маркера, пригодную для использования в микроорганизмах, предпочтительно, в дрожжах.
Гуанидинобутираза (гуанидино-кислая гидролаза) и гуанидиноамидгидролаза принадлежат к более крупному семейству белков, уреогидролаз, содержащему ферменты, обладающие 3-слойной альфа-бета-альфа-структурой и играющие важные роли в метаболизме аргинина/агматина, цикле мочевины и деградации гистидина. Гуанидинобутираза катализирует гидролиз 4-гуанидинобутанoата в 4-аминобутанoат и мочевину. Гуанидиноамид-гидролаза гидролизует агматин в мочевину и путресцин, предшественник для биосинтеза полиамины, спермидин и спермин. Семейство уреогидролаз содержит три дополнительные подгруппы: i) гидролазу гуанидино-аминокислот (аргиназа, EC3.5.3.1), катализирующую превращение аргинина в мочевину и орнитин, ii) проклаваминатамидиногидролазу (EC 3.5.3.22), катализирующую превращение амидинопроклавамината в мочевину и проклаваминат, промежуточный продукт в биосинтезе клавулановой кислоты, и iii) формиминоглутамазу (EC 3.5.3.8), катализирующую четвертый этап деградации гистидина посредством гидролиза N-формимидоил-L-глутамата в L-глутамат и формамид.
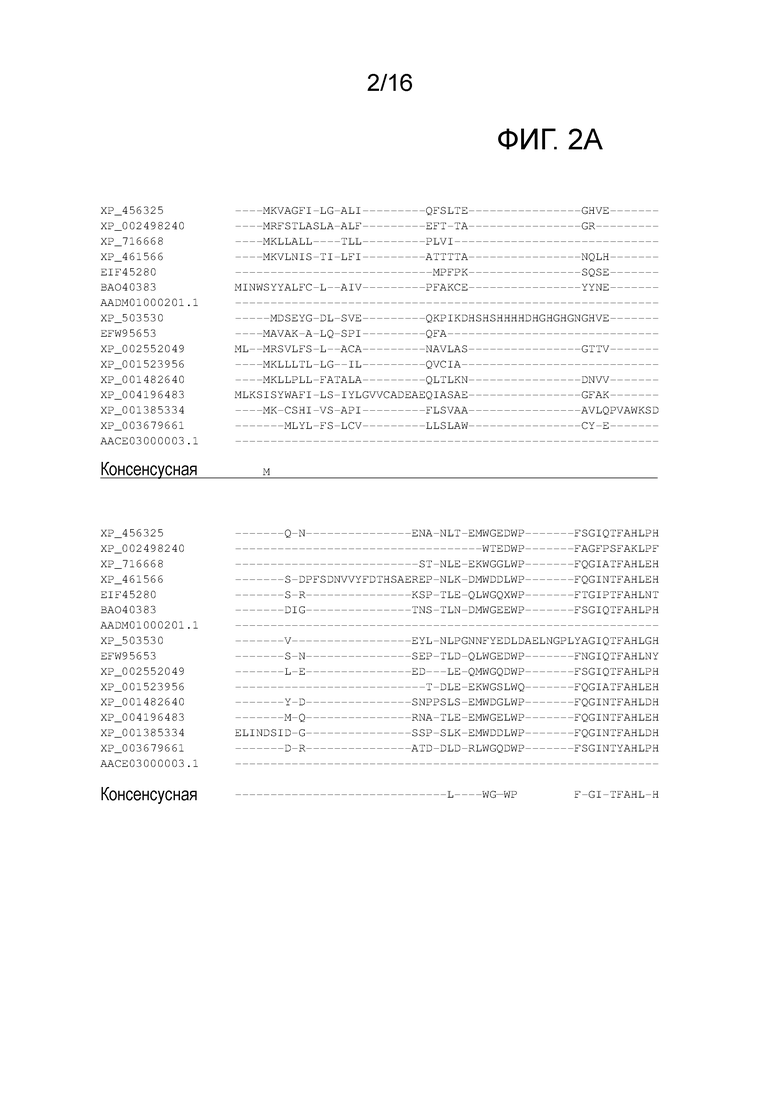
Указанная нуклеотидная последовательность, кодирующая гуанидинобутиразу, кодирует гуанидино-кислую гидролазу (EC.3,5.3,7). Нуклеотидная последовательность предпочтительно кодирует гуанидино-кислую гидролазу, имеющую один или оба консервативных домена, как показано на фигуре 2A и фигуре 2B, и/или кодирует консенсусную последовательность, как показано на фигуре 2A. Указанная гуанидино-кислая гидролаза предпочтительно содержит от 350 до 450 аминокислот, предпочтительно - приблизительно 400 аминокислот. Указанная нуклеотидная последовательность, кодирующая гуанидинобутиразу, предпочтительно кодирует гипотетический белок NRRL Y-1140 Kluyveromyces lactis, имеющий последовательность GenBank XP_456325.1, как показано на фигуре 3.
Указанная нуклеотидная последовательность, кодирующая гуанидиноамидгидролазу, кодирует агматинуреогидролазу (агматиназу) (EC.3.5.3.11). Нуклеотидная последовательность предпочтительно кодирует агматинуреогидролазу, имеющую консервативный домен, как показано на фигуре 4A и фигуре 4B, и/или кодирует консенсусную последовательность, как показано на фигуре 4A. Указанная агматинуреогидролаза предпочтительно содержит от 290 до 330 аминокислот, предпочтительно - приблизительно 306 аминокислот. Указанная агматинуреогидролаза предпочтительно кодирует последовательность GenBank AAC75974.1, как показано на фигуре 5.
Гуанидинобутираза и агматиназа катализируют образование мочевины, источника азота, повсеместно усваиваемого микроорганизмами, такими как S. cerevisiae. Таким образом, эти два гена уреогидролазы представляют собой важные характеристики потенциального доминантного селективного маркера с "приобретением функции" у микроорганизмов, таких как S. cerevisiae, при выращивании на гуанидинобутирате и/или агматине в качестве единственного источника азота.
Таким образом, изобретение относится к способу культивирования микроорганизма рода Saccharomycetaceae в присутствие гуанидинобутирата в качестве единственного источника азота, включающему (a) встраивание молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую гуанидинобутиразу, в микроорганизм, при этом нуклеотидная последовательность функционально связана с последовательностями промотора и терминатора, (b) культивирование микроорганизма таким образом, что молекула нуклеиновой кислоты, кодирующая гуанидинобутиразу, экспрессируется в микроорганизме, и (c) культивирование микроорганизма в присутствие гуанидинобутирата в качестве единственного источника азота.
Указанная нуклеотидная последовательность, кодирующая гуанидинобутиразу, предпочтительно кодирует гипотетический белок NRRL Y-1140 Kluyveromyces lactis.
Указанный промотор регулирует экспрессию селективного маркера в микроорганизме. Указанный терминатор опосредует эффективное образование 3'-конца мРНК. Указанный промотор предпочтительно является промотором дрожжей, более предпочтительно - промотором дрожжей, выбранным из гликолитического гена, такого как промотор PGI1 (фосфоглюкозоизомеразы 1), PFK1 (фосфофруктокиназы-1), PFK2 (фосфофруктокиназы-2), FBA1 (фруктозо-бисфосфатальдолазы-1), TPI1 (триозофосфатизомеразы-1), TDH1 (глицеральдегид-3-фосфат дегидрогеназы 1), TDH3 (глицеральдегид-3-фосфатдегидрогеназы 3), PGK1 (фосфоглицераткиназы 1), GPM1 (глицератфосфомутазы 1), PYK1 (пируваткиназы 1), ENO1 (альфа-енолазы) и/или ENO2 (енолазы 2), или выбранным из промотора ACT1 (актина 1), TEF1 (трансляционного фактора элонгации EF-1 альфа), TEF2 (трансляционного фактора элонгации EF-1 альфа 2), AgTEF2 (гена TEF2 Ashbya gossypii), PMA1 (H+-АТФазы плазматической мембраны типа P2). Предпочтительные последовательности промоторов выбраны из последовательностей промоторов генов PGI1, FBA1; TPI1; TDH3; PGK1; GPM1; ENO1; ENO2 и PYK1. Терминаторы ряда генов известны специалисту в этой области, и их используют, например, в экспрессирующих векторах, включающих терминаторы генов CYC1, TRP1, ADH1, MFl, FLP и D (Romanos et al., 1992. Yeast 8: 423-488).
Предпочтительные последовательности промоторов и последовательности терминаторов не содержат последовательности Saccharomyces. Предпочтительные последовательности промоторов и последовательности терминаторов получают из гена TEF Ashbya gossypii, кодирующего трансляционные факторы элонгации и/или регуляторные последовательности из генов URA3 и LEU2 K. lactis, кодирующих оротидин-5'-фосфат (OMP)-декарбоксилазу и бета-изопропилмалатдегидрогеназу, соответственно.
Молекулу нуклеиновой кислоты по изобретению, предпочтительно кодирующую гуанидинобутиразу, предпочтительно предоставляют в виде амплифицированного продукта или части амплифицированного продукта. Указанный амплифицированный продукт может дополнительно содержать последовательности, гомологичные первой части генома микроорганизма, предпочтительно - вышележащей части гена, подлежащего изменению (обозначаемого как ген-мишень), и последовательности, гомологичные второй части генома микроорганизма, предпочтительно - нижележащей части гена-мишени, смежной с молекулой нуклеиновой кислоты. Термин "смежный" используют для указания того, что последовательности, гомологичные первой части генома, локализованы на одной стороне молекулы нуклеиновой кислоты, в то время как последовательности, гомологичные второй части генома, локализованы на другой стороне молекулы нуклеиновой кислоты. Последовательности, гомологичные первой и второй части генома, предпочтительно содержат последовательности от 20 до 1000 п.н., более предпочтительно - от 30 до 500 п.н., более предпочтительно - от 40 до 250 п.н., более предпочтительно - от 50 до 80 п.н. Последовательности, гомологичные вышележащей части гена, предпочтительно включают инициаторный кодон. Последовательности, гомологичные нижележащей части гена, предпочтительно включают стоп-кодон. Способы получения амплифицированного продукта, содержащего молекулу нуклеиновой кислоты по изобретению, известны специалисту в этой области, включая, например, полимеразную цепную реакцию и амплификацию, основанную на последовательности нуклеиновой кислоты (NASBA).
Молекула нуклеиновой кислоты по изобретению предпочтительно содержится в векторе. Вектор содержит гены бактериальной устойчивости, например, делающие возможным рост бактерий в присутствие антибиотика. Наиболее предпочтительным вектором является плазмида, двухцепочечная молекула ДНК, способная реплицироваться в бактериях независимо от хромосомной ДНК.
Указанный вектор, предпочтительно - плазмида, предпочтительно дополнительно содержит последовательности, гомологичные первой части генома микроорганизма, предпочтительно - вышележащей части гена, подлежащего изменению (обозначаемого как ген-мишень), и последовательности, гомологичные второй части генома микроорганизма, предпочтительно - нижележащей части гена-мишени, смежные с молекулой нуклеиновой кислоты. Термин "смежный" используют для указания того, что последовательности, гомологичные первой части генома локализованы на одной стороне молекулы нуклеиновой кислоты, в то время как последовательности, гомологичные второй части генома, локализованы на другой стороне молекулы нуклеиновой кислоты.
Последовательности, гомологичные первой части генома, предпочтительно содержат от 20 до 1000 п.н., более предпочтительно - от 30 до 500 п.н., более предпочтительно - от 40 до 250 п.н., более предпочтительно - от 50 до 80 п.н. последовательностей, являющихся гомологичными первой части генома, предпочтительно - вышележащей части гена, подлежащего изменению или делеции. Последовательности, гомологичные вышележащей части гена, предпочтительно включают инициаторный кодон.
Последовательности, гомологичные второй части генома, предпочтительно содержат от 20 до 1000 п.н., более предпочтительно - от 30 до 500 п.н., более предпочтительно - от 40 до 250 п.н., более предпочтительно - от 50 до 80 п.н. последовательностей, являющихся гомологичными второй части генома, предпочтительно - нижележащей части гена, подлежащего изменению или делеции. Последовательности, гомологичные нижележащей части гена, предпочтительно включают стоп-кодон.
Указанные последовательности, гомологичные первой и/или второй части генома, предпочтительно - гену-мишени, могут содержать последовательности, являющиеся измененными по сравнению с последовательностями генома. Термины "изменение" и "измененный" относятся к замене одного или нескольких нуклеотидов, инсерции одного или нескольких нуклеотидов и/или делеции одного или нескольких нуклеотидов где-либо в гомологичных последовательностях.
Замену одного или нескольких нуклеотидов можно осуществлять посредством изменения одного или нескольких нуклеотидов в первой части и/или второй части, предпочтительно - в последовательностях, гомологичных вышележащей и/или нижележащей части гена. Если первая часть участка гомологии и вторая часть участка гомологии покрывают смежные области генома, предпочтительно - гена-мишени, встраивание направленного вектора будет приводить к изменению генома.
Указанный вектор, предпочтительно - плазмида, предпочтительно дополнительно содержит участок распознавания эндонуклеазы на одном конце молекулы нуклеиновой кислоты по изобретению и копию этого участка распознавания на другом конце молекулы нуклеиновой кислоты по изобретению. Указанные участки распознавания предпочтительно локализованы непосредственно смежно с молекулой нуклеиновой кислоты по изобретению. Указанная эндонуклеаза предпочтительно является редкощепящей эндонуклеазой, такой как, например, PacI (последовательность распознавания мишени 5'-TTAATTAA); AscI (последовательность распознавания мишени 5ʹ-GGCGCGCC), и AsiSI (последовательность распознавания мишени 5'-GCGATCGC). PacI, AscI и AsiSI доступны в New England Biolabs. Более предпочтительно, эндонуклеаза является хоминг-эндонуклеазой. Термин "хоминг-эндонуклеаза" относится к эндонуклеазам, кодируемым в виде самостоятельных генов в интронах, слитыми с белком хозяина или в виде самосплайсирующегося интеина. Предпочтительный список хоминг-эндонуклеаз представлен в таблице 1. Дополнительными примерами хоминг-нуклеаз являются I-DirI, I-NjaI, I-NanI, I-NitI, F-TevI, F-TevII, F-CphI, PI-MgaI, I-CsmI, все из которых известны специалисту в этой области. Дополнительные примеры хоминг-нуклеаз представлены в Benjamin K (патентной заявке США № 2012/052582), включенной в настоящее описание в качестве ссылки.
Указанный вектор, предпочтительно - плазмида, предпочтительно дополнительно содержит последовательность нуклеиновой кислоты от 20 до 200 п.н., предпочтительно - от 30 до 100 п.н., более предпочтительно - приблизительно 40-50 п.н., дуплицированную на любой стороне молекулы нуклеиновой кислоты по изобретению, предпочтительно - вне участков распознавания эндонуклеазы. Указанная последовательность нуклеиновой кислоты от 20 до 200 п.н. предпочтительно локализована между последовательностями, гомологичными вышележащей части гена-мишени, и участком распознавания эндонуклеазы на одном конце молекулы нуклеиновой кислоты по изобретению, и между последовательностями, гомологичными нижележащей части гена-мишени, и участком распознавания эндонуклеазы на втором конце молекулы нуклеиновой кислоты по изобретению. Дуплицированная последовательность нуклеиновой кислоты предпочтительно идентична области в геноме-мишени, предпочтительно - гена-мишени. Дуплицированная последовательность нуклеиновой кислоты от 20 до 200 п.н. обеспечивает бесшовное удаление маркера из генома-мишени посредством гомологичной рекомбинации.
Изобретение дополнительно относится к способу получения микроорганизма, предпочтительно - дрожжей, содержащего молекулу нуклеиновой кислоты по изобретению, включающему получение микроорганизма с использование молекулы нуклеиновой кислоты по изобретению, селекцию микроорганизма, имеющего указанную молекулу нуклеиновой кислоты, таким образом, получение микроорганизма, содержащего молекулу нуклеиновой кислоты по изобретению.
Способы селекции микроорганизм, предпочтительно - дрожжей, имеющих нуклеиновую кислоту по изобретению, известны в этой области и включают Саузерн-блоттинг и амплификацию продукта нуклеиновой кислоты, содержащего, по меньшей мере, часть молекулы нуклеиновой кислоты, с использованием по меньшей мере одного праймера, специфичного для молекулы нуклеиновой кислоты. Альтернативно, вектор, содержащий молекулу нуклеиновой кислоты по изобретению, предпочтительно дополнительно содержит селективный маркер, делающий возможной селекцию микроорганизма, содержащего молекулу нуклеиновой кислоты по изобретению.
Как применяют в настоящем описании, термин "специфичный" относится к праймеру или полинуклеотиду, который будет гибридизоваться только с его подпоследовательностью-мишенью, как правило, в сложной смеси нуклеиновых кислот, но, по существу, не с другими последовательностями при использовании строгих условий, как известно специалисту. Строгие условия зависят от последовательности и будут отличаться в разных обстоятельствах. Обширное руководство по гибридизации нуклеиновых кислот можно найти в Tijssen (Tijssen, (1993) Hybridization with Nucleic Acid Probes, vol. 2, Laboratory techniques in biochemistry and molecular biology, Volume 24. Elsevier, Amsterdam). Как правило, выбирают строгие условия, представляющие собой температуру на приблизительно 5-10°C ниже температуры плавления (Tm) для конкретной последовательности при определенной ионной силе и pH. Tm является температурой (при определенной ионной силе, pH и концентрации нуклеиновой кислоты), при которой 50% праймеров, комплементарных мишени, гибридизуются с последовательностью-мишенью при равновесии (т.к. последовательности-мишени присутствуют в избытке, при Tm 50% зондов заняты при равновесии). Строгие условия будут такими, при которых концентрация соли составляет менее приблизительно 1,0 M ионов натрия, как правило, приблизительно от 0,01 до 1,0 M ионов натрия (или других солей) при pH от 7,0 до 8,3, и температура составляет по меньшей мере приблизительно 30°C для коротких зондов (например, от 10 до 50 нуклеотидов) и по меньшей мере приблизительно 60°C для длинных зондов (например, более 50 нуклеотидов). В случае избирательной или специфичной гибридизации положительный сигнал является по меньшей мере в два раза более высоким, чем фон, предпочтительно - в 10 раз выше фона.
Предпочтительный способ по изобретению включает селекцию микроорганизма, предпочтительно - дрожжей, функционально экспрессирующего молекулу нуклеиновой кислоты, кодирующую селективный маркер. Например, можно подвергать селекции дрожжи, экспрессирующие гуанидинобутиразу и/или агматиназу, как определяют в анализе активности фермента. Способы определения активности гуанидинобутиразы или агматиназы известны в этой области. Количество продуцируемой мочевины можно количественно анализировать с использованием, например, способа Арчибальда (Archibald, (1945) J Biol Chem 157: 507-518). Можно получать калибровочную кривую в диапазоне от 0-1 мМ мочевины для количественного анализа количества мочевины.
В настоящем описании термин "функционально экспрессируют" используют для указания того, что с интересующего гена экспрессируется белок, кодируемый геном, в активной форме.
Указанный микроорганизм, предпочтительно - дрожжи, предпочтительно - рода Saccharomycetaceae, предпочтительно подвергают селекции, т.к. он способен расти на субстрате, предпочтительно синтетической среде, содержащей гуанидинобутират и/или агматин в качестве единственного источника азота.
Изобретение дополнительно относится к микроорганизму, предпочтительно - дрожжам, содержащему молекулу нуклеиновой кислоты по изобретению. Указанную молекулу нуклеиновой кислоты по изобретению предпочтительно встраивают в геном указанного микроорганизма.
Указанный микроорганизм предпочтительно принадлежит к роду Saccharomycetacea и содержит нуклеотидную последовательность, кодирующую гуанидинобутиразу, предпочтительно - нуклеотидную последовательность, кодирующую гуанидинобутиразу, кодирующую гипотетический белок Y-1140 Kluyveromyces lactis NRRL.
Способ по изобретению можно использовать по отношению ко всем микроорганизмам. Если микроорганизм имеет эндогенную активность гуанидинобутиразы и/или агматиназы, можно получать мутанты этого организма, в которых эта активность инактивирована. В этой области известны способы инактивации гена, кодирующего гуанидинобутиразу или агматиназу в микроорганизме. Подходящие способы описаны, например, в Akada et al., (2002). Yeast 19: 393-402; McNabb et al., (1997). Biotechniques, 22: 1134-1139; Storici et al., (1999). Yeast 15: 271-283; Gueldener et al., (2002). Nucleic Acids Res 30: e23; и Iwaki and Takegawa, (2004). Biosci Biotechnol Biochem 68: 545-550. Любой из этих способов можно использовать для получения микроорганизма, в котором эндогенный ген, кодирующий гуанидинобутиразу и/или агматиназу, можно функционально инактивировать.
В настоящем описании термин "функционально инактивированный" используют для указания сниженного функционального присутствия белкового продукта гена в микроорганизме, являющегося результатом сниженного уровня экспрессии или сниженного уровня активности белка. Указанное сниженное функциональное присутствие предпочтительно приводит к снижению количества и/или активности белка более чем на 90%, более предпочтительно - снижению количества и/или активности белка более чем на 95%, наиболее предпочтительно - снижению количества и/или активности белка более чем на 99% по сравнению с соответствующей активностью белка в родственном микроорганизме, не содержащем функционально инактивированный ген. В наиболее предпочтительном варианте осуществления функционально инактивированный ген не имеет остаточной активности и эквивалентен нокаутному гену. Термин "нокаутный ген" относится к гену, сделанному функционально неактивным посредством частичной или полной делеции кодирующей области из геномной ДНК, кодирующей указанный ген.
Микроорганизм, предпочтительно - дрожжи, по изобретению предпочтительно является дрожжами рода Saccharomycetaceae. Этот род включает Saccharomyces sensu stricto, Kazachstania, Naumovozyma, Nakaseomyces и Vanderwaltozyma. Предполагают, что эти рода, принадлежащие к семейству Saccharomycetaceae, возникли после полногеномной дупликации (после WGD), играющей основную роль в эволюции этого подвида.
При сравнении метаболизма азота Saccharomyces cerevisiae и Kluyveromyces lactis выявили значительные различия усвоения аргинина между двумя видами дрожжей. Хотя делеция единственного гена аргиназы CAR1 S. cerevisiae является достаточной для прекращения роста на аргинине в качестве единственного источника азота, соответствующий мутант Klcar1Δ в K. lactis все равно способен расти на аргинине. Это фенотипическое различие вызвано наличием в K. lactis гена (KLLA0F27995g/KlGBU1), кодирующего гуанидинобутиразу (EC.3.5.3.7), фермент, ранее не обнаруженный в грибах. Наличие этого фермента обеспечивает способность расти на гуанидинобутирате в качестве единственного источника азота.
Обнаружено, что ортологи гуанидинобутиразы не присутствуют у рода Saccharomycetaceae после WGD. Примечательно, что одна из
34 аннотированных последовательностей генома S. cerevisiae, доступных в базе данных SGD (www.yeastgenome.org/), принадлежащих к штамму EC1118, несет ген, прогнозируемая белковая последовательность которого обладает 62% идентичности последовательности по отношению к последовательности гуанидинобутиразы Kluyveromyces lactis. При дальнейшем исследовании структуры генома штамма EC1118 выявлено, что этот ортолог обнаружен в геномной области, приобретенной посредством горизонтального переноса, наиболее вероятно - из Torulaspora delbrueckii. Это генетическое происхождение соответствует филогенетическому распределению ортологов гуанидинобутиразы в пре-WGD родах Saccharomycetaceaee (Zygosaccharomyces, Lachancea, Torulaspora, Kluyveromyces и Eremothecium), и в родах, принадлежащих к группе CTG и Dipodascacae (Kurtzman, (2003) FEMS Yeast Res 4: 233-245; Dujon, (2010) Nat Rev Genet 11: 512-524).
Предпочтительными дрожжами являются лабораторные дрожжи, дрожжи дикого типа и промышленные лагерные дрожжи Saccharomyces pastorianus, лабораторный штамм, штамм дикого типа и промышленный штамм S. cerevisiae, предпочтительно - в буквальном смысле Saccharomyces (Saccharomyces paradoxus, S. mikatae, S. bayanus, S. eubayanus, S. kudriavzevii, S. paradoxus, S. arboricolus), или штамм Kazachstania, Naumovozyma, Nakaseomyces или Vanderwaltozyma, наиболее предпочтительно - лабораторные дрожжи, дрожжи дикого типа и промышленные лагерные дрожжи Saccharomyces pastorianus или штамм S. cerevisiae.
Изобретение дополнительно относится к способу изменения генома микроорганизма, предпочтительно - генома дрожжей, предпочтительно - дрожжей рода Saccharomycetaceae, включающему получение молекулы нуклеиновой кислоты по изобретению, предпочтительно - кодирующей гуанидинобутиразу, более предпочтительно - кодирующей гипотетический белок NRRL Y-1140 Kluyveromyces lactis, в указанном микроорганизме, и селекцию микроорганизма, в котором геном изменяют посредством инсерции в геном молекулы нуклеиновой кислоты по изобретению. Предпочтительный способ включает получение набора конструкций по изобретению в указанном микроорганизме и селекцию микроорганизма, в котором геном изменен, предпочтительно - посредством селекции микроорганизма, функционально экспрессирующего указанную гуанидинобутиразу. Предпочтительный способ включает селекцию микроорганизма посредством культивирования в присутствие гуанидинобутирата в качестве единственного источника азота.
Настоящее изобретение дополнительно относится к набору конструкций, содержащему первую конструкцию, содержащую первую часть нуклеотидной последовательности, кодирующей селективный маркер, как указано в п. 1 формулы изобретения, и вторую конструкцию, содержащую вторую часть нуклеотидной последовательности, кодирующую селективный маркер, как указано в п. 1 формулы изобретения, при этом фрагмент первой части селективного маркера перекрывается с фрагментом, присутствующим во второй части нуклеотидной последовательности, делая возможной рекомбинацию между первой и второй частью нуклеотидной последовательности.
С помощью этого набора конструкций преодолевают низкую эффективность направленного воздействия, получая набор направленных конструкций, в которых правильная экспрессия селективного маркера зависит от рекомбинации между направленными конструкциями. Обнаружено, что частота рекомбинации между направленными конструкциями значительно повышается после встраивания направленных конструкций в правильном локусе-мишени. Таким образом, направленная система по настоящему изобретению, содержащая набор направленных конструкций, значительно повышает процентную долю правильно встроенных конструкций у микроорганизмов, экспрессирующих селективный маркер, по сравнению с направленной системой с одним вектором. Разделение маркера на две отдельные конструкции ограничивает частоту ложноположительных результатов по причине отдельных кроссинговеров. С помощью подхода с раздельными маркерами улучшают соотношение положительных и ложноположительных результатов (Nielsen et al., 2006. Fungal Gen Biol 43: 54-64).
Как применяют в настоящем описании, термин "конструкция" или "направленная конструкция" относится к искусственно сконструированному сегменту нуклеиновой кислоты. Предпочтительной конструкцией является вектор, предпочтительно - вектор, содержащий гены бактериальной устойчивости для роста бактерий. Наиболее предпочтительной конструкцией является плазмида, линейная или кольцевая двухцепочечная ДНК, способная реплицироваться в бактериях независимо от хромосомной ДНК.
Как применяют в настоящем описании, термин "перекрывание" относится к дуплицированной области нуклеотидной последовательности, кодирующей селективный маркер, присутствующей в наборе конструкций. Дуплицированная область, по существу, является идентичной и предпочтительно составляет от 40 до 400 п.н., предпочтительно - приблизительно 200 п.н. Как применяют в настоящем описании, термин "по существу" используют для указания того, что область является по меньшей мере на 90% идентичной для обеспечения эффективной рекомбинации между направленными конструкциями, более предпочтительно - по меньшей мере на 95% идентичной, более предпочтительн - по меньшей мере на 99% идентичной, более предпочтительно - на 100% идентичной.
Первая конструкция в наборе направленных конструкций предпочтительно дополнительно содержит участок распознавания эндонуклеазы и первую область гомологии с геномом-мишенью микроорганизма, и вторая конструкция дополнительно содержит вторую область гомологии с геномом-мишенью микроорганизма и копию участка распознавания эндонуклеазы, при этом кодирующая последовательность, кодирующая эндонуклеазу и соединенная с индуцибельным промотором, присутствует в первой или второй конструкции; и часть первой области гомологии с геномом-мишенью в первой конструкции является дуплицированной между копией участка распознавания эндонуклеазы и второй областью гомологии с геномом-мишенью во второй конструкции; или часть второй области гомологии с геномом-мишенью во второй конструкции является дуплицированной между первой области гомологии с геномом-мишенью и участком распознавания эндонуклеазы в первой конструкции.
Указанная дуплицированная область гомологии с геномом-мишенью в первой и второй направленной конструкции предпочтительно составляет от 20 до 200 п.н., предпочтительно - от 40 до 100 п.н., предпочтительно - приблизительно 80 п.н. Указанная дуплицированная область гомологии с геномом-мишенью в первой и второй направленной конструкции делает возможным бесшовное удаление маркера из генома-мишени посредством гомологичной рекомбинации.
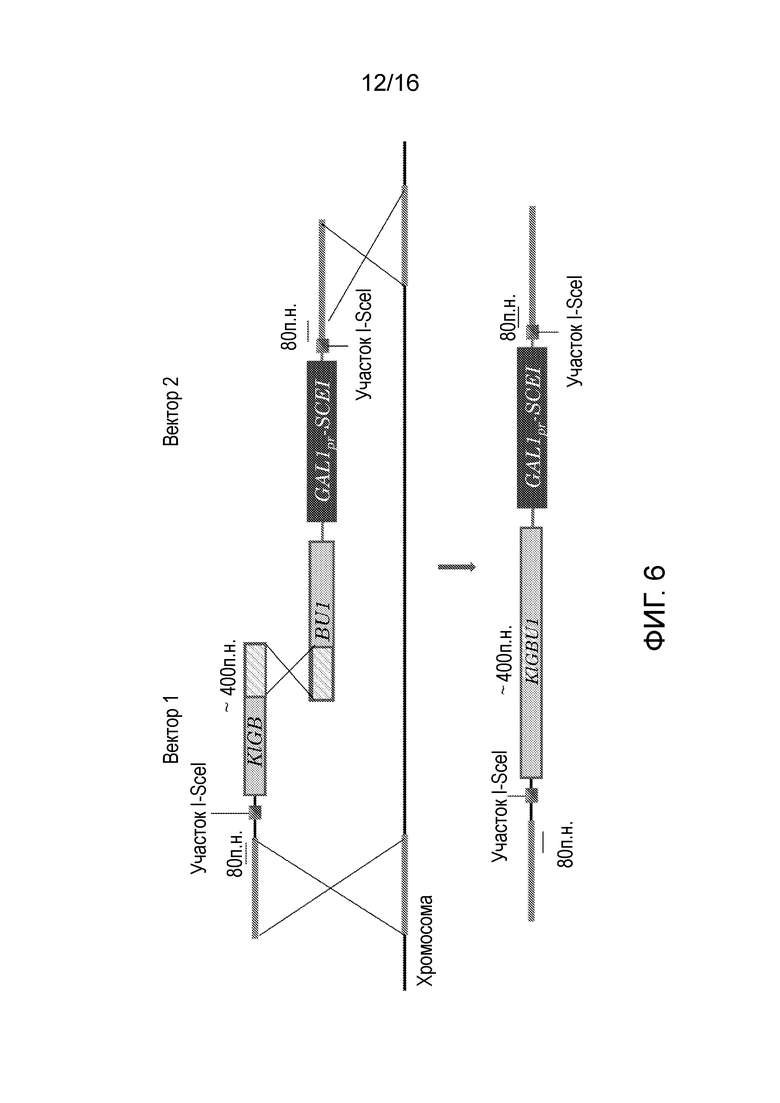
Первая конструкция предпочтительно содержит в этом порядке первую область гомологии с геномом-мишенью микроорганизма, участок распознавания эндонуклеазы и первую часть селективного маркера. Вторая конструкция предпочтительно содержит в этом порядке область перекрывания с первой частью нуклеотидной последовательности, кодирующей селективный маркер, затем вторую часть нуклеотидной последовательности, кодирующей селективный маркер, кодирующую последовательность, кодирующую эндонуклеазу и соединенную с индуцибельным промотором, копию участка распознавания эндонуклеазы, копию части первой области гомологии с геномом-мишенью, присутствующим в первой конструкции, и вторую область гомологии с геномом-мишенью микроорганизма. Эта конфигурация изображена на фигуре 6.
Геном-мишень может иметь любую локализацию, предпочтительно - ген, в геноме микроорганизма, предпочтительно - дрожжей, предпочтительно - дрожжей рода Saccharomytacea, геномная последовательность которого подлежит изменению. Как применяют в настоящем описании, термин "ген" относится к части генома микроорганизма, содержащей интронные и экзонные части гена, промоторную область указанного гена и геномные последовательности, опосредующие экспрессию указанного гена, такие как, например, энхансерные последовательности.
Специалисту в этой области будет понятно, что направленные конструкции предпочтительно используют для изменения гена микроорганизма. Таким образом, изобретение дополнительно относится к набору направленных конструкций, содержащему первую конструкцию, содержащую первую область гомологии с геном-мишенью микроорганизма, участок распознавания эндонуклеазы и первую часть нуклеотидной последовательности, кодирующей селективный маркер, и вторую конструкцию, содержащую область перекрывания с первой частью нуклеотидной последовательности, кодирующей селективный маркер, затем нуклеотидную последовательность, кодирующую вторую часть селективного маркера, копию участка распознавания эндонуклеазы и вторую область гомологии с геном-мишенью микроорганизма, при этом перекрывающиеся фрагменты делают возможной рекомбинацию между первой и второй частью нуклеотидной последовательности, кодирующей селективный маркер; при этом кодирующая последовательность, кодирующая эндонуклеазу и соединенная с индуцибельным промотором, присутствует в первой или второй конструкции; и при этом часть первой области гомологии с геном-мишенью в первой конструкции является дуплицированной между копией участка распознавания эндонуклеазы и второй областью гомологии с геном-мишенью во второй конструкции; или часть второй области гомологии с геном-мишенью во второй конструкции является дуплицированной между первой областью гомологии с геном-мишенью и участком распознавания эндонуклеазы в первой конструкции.
Указанная дуплицированная область гомологии с геном-мишенью в первой и второй направленной конструкцией предпочтительно составляет от 20 до 200 п.н., предпочтительно - от 40 до 100 п.н., предпочтительно - приблизительно 80 п.н.
Термин "изменение" геномной последовательности включает замену одного или нескольких нуклеотидов, инсерцию одного или нескольких нуклеотидов и/или делецию одного или нескольких нуклеотидов где-либо в геноме, предпочтительно - в гене.
Например, если первая и вторая области гомологии с геном-мишенью содержат смежные геномные последовательности гена, замена одного или нескольких нуклеотидов в первой области гомологии и/или второй области гомологии будет приводить к изменению гена после гомологичного таргетинга с использованием набора направленных конструкций по изобретению. Указанную замену одного или нескольких нуклеотидов предпочтительно осуществляют в области гомологии с геном-мишенью, присутствующей в первой и во второй конструкции.
Указанное изменение геномной последовательности предпочтительно является делецией одного или нескольких нуклеотидов, предпочтительно - где-либо в гене. Например, если первая и вторая области гомологии с геном-мишенью содержат геномные последовательности гена, разделенные в геноме организма, изменение гена после гомологичного таргетинга с использованием набора направленных конструкций по изобретению будет приводить к делеции области, локализованной между первой и второй областями гомологии на родительской хромосоме.
Указанная первая конструкция предпочтительно содержит первую часть, предпочтительно - первые две трети или первую половину, области, кодирующей селективный маркер. Например, белок гуанидинобутираза K. lactis содержит 410 аминокислот, кодируемые последовательностью нуклеиновой кислоты размером 1230 п.н. Указанная первая конструкция предпочтительно содержит от 400 до 800 п.н. кодирующей области этого белка, более предпочтительно - от 500 до 700 п.н. Вторая конструкция предпочтительно содержит от 400 до 800 п.н. кодирующей области этого белка, более предпочтительно - от 500 до 700 п.н.
Область перекрывания между первой и второй части селективного маркера предпочтительно составляет приблизительно от 50 п.н. до приблизительно 600 п.н., предпочтительно - приблизительно 200 п.н.
Первая или вторая направленная конструкция содержит кодирующую последовательность, кодирующую эндонуклеазу и соединенную с индуцибельным промотором. Эндонуклеаза предпочтительно является редкощепящей эндонуклеазой, такой как, например, PacI (последовательность распознавания мишени 5'-TTAATTAA); AscI (последовательность распознавания мишени 5ʹ-GGCGCGCC) и AsiSI (последовательность распознавания мишени 5'-GCGATCGC). PacI, AscI и AsiSI доступны в New England Biolabs. Более предпочтительно, эндонуклеаза является хоминг-эндонуклеазой. Термин "хоминг-эндонуклеаза" относится к эндонуклеазам, кодируемым в виде самостоятельных генов в интронах, слитыми с белком хозяина или в виде самосплайсирующегося интеина. Предпочтительный список хоминг-эндонуклеаз представлен в таблице 1. Дополнительными примерами хоминг-нуклеаз являются I-DirI, I-NjaI, I-NanI, I-NitI, F-TevI, F-TevII, F-CphI, PI-MgaI, I-CsmI, все из которых известны специалисту в этой области. Дополнительные примеры хоминг-нуклеаз представлены в патентной заявке США № 2012/052582, включенной в настоящее описание в качестве ссылки.
Предпочтительной хоминг-нуклеазой является PI-PspI (New England Biolabs; последовательность распознавания 5ʹ-TGGCAAACAGCTATTATGGGTATTATGGGT)) или PI-SceI (New England Biolabs; последовательность распознавания 5ʹ-ATCTATGTCGGGTGCGGAGAAAGAGGTAAT). Известны кодирующие последовательности большинства хоминг-эндонуклеаз. Например, кодирующие последовательности PI-SceI и PI-PspI доступны в общедоступных базах данных (инвентарный номер Genbank Z74233.1 и инвентарный номер Genbank U00707.1, соответственно). Специалисту в этой области будет понятно, что последовательность, отличающаяся от общедоступной последовательности нуклеазы, все равно может кодировать нуклеазу. Например, термин "кодирующая область PI-PspI" может включать последовательность, отличающуюся от общедоступной последовательности, например, оптимизацией кодонов, но все равно экспрессирующую активную эндонуклеазу, распознающую и расщепляющую указанную последовательность распознавания мишени.
Указанная эндонуклеаза предпочтительно находится под контролем индуцибельного промотора. Как применяют в настоящем описании, термин "индуцибельный промотор" относится к промотору, экспрессию которого можно регулировать. Индуцибельные промоторы известны специалисту в этой области. Примерами индуцибельных промоторов, используемых в дрожжах, являются промотор GAL1 и промотор GAL10, индуцируемые галактозой, промотор SUC2, индуцируемый сахарозой, промотор MAL12, индуцируемый мальтозой; промотор CUP1, индуцируемый медью, и промоторы tetO7 и tetO2, индуцируемые тетрациклином [Gari et al., (1997) Yeast 13: 837-48; Yen et al., (2003) Yeast 20 1255-62]. Предпочтительным индуцибельным промотором является промотор GAL1.
Один участок распознавания, содержащий последовательность распознавания мишени для эндонуклеазы, локализован смежно (позади) с первой областью гомологии с геном-мишенью микроорганизма в первой конструкции. Копия этого участка распознавания локализована смежно (перед) со второй областью гомологии с геном-мишенью микроорганизма во второй конструкции. Специалисту в этой области будет понятно, что, если часть первой области гомологии с геном-мишенью в первой конструкции является дуплицированной между копией участка распознавания эндонуклеазы и второй областью гомологии с геном-мишенью во второй конструкции, указанная копия участка распознавания локализована смежно (перед) с дупликацией первой области гомологии с геном-мишенью во второй конструкции. Альтернативно, участок распознавания локализован смежно (позади) с дуплицированной частью второй области гомологии с геном-мишенью в первой конструкции, если часть второй области гомологии с геном-мишенью во второй конструкции является дуплицированной в первой конструкции. Селективный маркер, включая последовательности промотора и терминатора, и кодирующая область эндонуклеазы, включая индуцибельный промотор, находятся между участком распознавания в первой конструкции и копией этого участка распознавания во второй конструкции.
Изобретение дополнительно относится к способу изменения генома, предпочтительно - гена-мишени, в микроорганизме, включающему снабжение набором направленных конструкций по изобретению указанного микроорганизма и селекцию микроорганизма, в котором изменяют геном. Указанную селекцию микроорганизма, в котором изменяют геном, предпочтительно осуществляют посредством селекции микроорганизма, функционально экспрессирующего рекомбинированный селективный маркер.
Как указано в настоящем описании выше, частота рекомбинации между направленными конструкциями значительно повышается после встраивания направленных конструкций в правильный локус таргетинга. Таким образом, наличие функционально рекомбинированного селективного маркера очень показательно для наличия правильно встроенных направленных конструкций в геноме-мишени и, таким образом, измененного генома в микроорганизме.
Как указано в настоящем описании выше, термины "изменение" и "измененный" относятся к замене одного или нескольких нуклеотидов, инсерции одного или нескольких нуклеотидов и/или делеции одного или нескольких нуклеотидов где-либо в геноме, предпочтительно - в гене-мишени.
Замену одного или нескольких нуклеотидов можно осуществлять, изменяя один или несколько нуклеотидов в первой области гомологии и/или во второй области гомологии. Если первая область гомологии и вторая область гомологии с геномом-мишенью покрывают смежные области генома, предпочтительно - гена-мишени, встраивание направленных векторов будет приводить к изменению генома. При наличии, указанную замену одного или нескольких нуклеотидов предпочтительно осуществляют, изменяя один или несколько нуклеотидов в перекрывающейся области гомологии с геномом, присутствующей в первой и во второй конструкции.
Указанное изменение геномной последовательности предпочтительно является делецией одного или нескольких нуклеотидов где-либо в геноме, предпочтительно - в гене. Например, если первая и вторая области гомологии с геномом-мишенью содержат геномные последовательности, разделенные в геноме организма, изменение генома после гомологичного таргетинга с использованием набора направленных конструкций по изобретению будет приводить к делеции области, локализованной между первой и второй областями гомологии на родительской хромосоме.
Изобретение дополнительно относится к способу получения микроорганизма, содержащего измененный геном, предпочтительно - измененный ген, включающему снабжение набором направленных конструкций по изобретению указанного микроорганизма и селекцию микроорганизма, в котором изменяют геном и который функционально экспрессирует рекомбинированный селективный маркер.
Способ получения микроорганизма, содержащего измененный геном, предпочтительно включает индуцирование индуцибельного промотора для экспрессии эндонуклеазы, таким образом, удаление селективного маркера и кодирующей области эндонуклеазы, включая индуцибельный промотор, из генома-мишени.
Изобретение дополнительно относится к микроорганизму, содержащему изменение генома, получаемое способами по изобретению. При наличии, дуплицированные области гомологии с геномом-мишенью в первой и второй направленных конструкциях обеспечивает бесшовное удаление маркера из генома-мишени посредством гомологичной рекомбинации. Получаемый микроорганизм содержит только изменение или изменения, присутствовавшие в первой и/или второй направленных конструкциях или индуцированные посредством рекомбинации направленных конструкций в подлежащий таргетингу геном, такое как инсерция в подлежащий таргетингу геном или делеция из подлежащего таргетингу генома.
Изобретение дополнительно относится к микроорганизму, содержащему геномное изменение, предпочтительно - изменение гена-мишени, изменение, содержащее инсерцию функционально рекомбинированного селективного маркера и кодирующую последовательность для эндонуклеазы, соединенную с индуцибельным промотором, при этом геном-мишень содержит одну копию последовательности распознавания для эндонуклеазы в обоих участках инсерции.
Изобретение дополнительно относится к набору, содержащему молекулу нуклеиновой кислоты по изобретению или набор конструкций по изобретению. Указанный набор может дополнительно включать способы и средства для выращивания микроорганизма, предпочтительно дрожжей, предпочтительно - из Saccharomycetaceae, в синтетической среде, содержащей гуанидинобутират и/или агматин, и/или идентификации фермента гуанидинобутиразы или агматиназы, такие как, например, гуанидиномасляная кислота и/или агматин.
Изобретение также относится к способу культивирования микроорганизма, предпочтительно - дрожжей, предпочтительно - из Saccharomycetaceae, в присутствие гуанидинобутирата или агматина в качестве единственного источника азота, включающему: (a) встраивание молекулы нуклеиновой кислоты по изобретению в микроорганизм и (b) культивирование микроорганизма таким образом, что молекула нуклеотида экспрессируется в микроорганизме.
Изобретение также относится к способу культивирования микроорганизма, предпочтительно - дрожжей, предпочтительно - из Saccharomycetaceae, в присутствие гуанидинобутирата или агматина в качестве единственного источника азота, включающему: (a) встраивание набора конструкций по изобретению в микроорганизм и (b) культивирование микроорганизма таким образом, что после рекомбинации направленных конструкций молекула нуклеотида, кодирующего селективный маркер, экспрессируется в микроорганизме.
Изобретение дополнительно относится к способу получения микроорганизма, содержащего измененный геном, включающему получение микроорганизма, содержащего изменение генома, предпочтительно - гена-мишени, изменение, включающее инсерцию функционально рекомбинированной нуклеотидной последовательности, кодирующей селективный маркер, и кодирующей последовательности эндонуклеазы, соединенной с индуцибельным промотором, при этом геном-мишень содержит одну копию последовательности распознавания для эндонуклеазы в обоих участках инсерции, и индуцирование индуцибельного промотора для удаления последовательности нуклеиновой кислоты между последовательностями распознавания эндонуклеазы. И снова, при наличии, дуплицированные области гомологии с геном-мишенью в первой и второй направленных конструкциях обеспечивают бесшовное удаление маркера из генома-мишени посредством гомологичной рекомбинации с помощью геномной ДНК с небольшим гомологичным фрагментом для эффективного повторного соединения разорванных цепей ДНК. Получаемый микроорганизм содержит только изменение или изменения, присутствовавшие в первой и/или второй направленных конструкциях или индуцированные посредством рекомбинации направленных конструкций в подлежащем таргетингу геноме, такое как инсерция в подлежащий таргетингу геном или делеция из генома, предпочтительно - инсерция в подлежащий таргетингу ген, или делеция подлежащего таргетингу гена, или делеция из подлежащего таргетингу гена.
В целях ясности и краткости описания признаки в настоящем описании описывают как часть одного или отдельных вариантов осуществления, однако следует понимать, что объем изобретения может включать варианты осуществления, включающие комбинации всех или некоторых из описываемых признаков.
Таблица 1
3ʹ AACTCCTCCAAAGAGACATTTATT 5ʹ
3' ---AACTCCTCC AAAGAGACATTTATT--- 5'
3' ATTGATATTGCCAGGATTCCATCGCT 5ʹ
3' ---ATTGATATTGCCAG GATTCCATCGCT--- 5'
3' CTTCCAAACCGTGGAGCTACAGCCGAGTAG 5ʹ
3' ---CTTCCAAACCGTG GAGCTACAGCCGAGTAG--- 5'
3' GCTAGGATTCCATCGCTTTAAGT 5ʹ
3' ---GCTAGGATTCCATC GCTTTAAGT--- 5'
3' GGGCCGATTGAGACACGGTC 5ʹ
5' ---GGGCCGAT TGAGACACGGTC--- 3'
3' GACCCAAGTTTTGCAGCACTCTGTCAAACC 5ʹ
3' ---GACCCAAGTTTTGCAG CACTCTGTCAAACC--- 5'
3' TACGGAACGGCCCATTCAAGGCCGCGCGTA 5ʹ
3' ---TACGGAACGGCC CATTCAAGGCCGCGCGTA--- 5'
3' GTTTTGCAGCATTCAAGGCCGCGC 5'
3' ---GTTTTGCAG CATTCAAGGCCGCGC--- 5'
3' TCATTACTCGGATTGCGAGTCGTT 5ʹ
3' ---TCATTACTCGGATTGC GAGTCGTT--- 5'
3'TCATTACTCGGATTGCGAGTTGTT 5ʹ
3' GTGTAGGTATTGGTATAGTAAAAA 5ʹ
3' ---GTGTAGGTATTGGTATAGTAA AAA--- 5'
3' GACCCAAGTTTTGCAGCACTCTGTCAAACC 5ʹ
3' ---GACCCAAGTTTTGCAG CACTCTGTCAAACC--- 5'
3' CTTCTACCCTCCTCCCTGGCCTGAGTTGAA 5ʹ
3' ---CTTCTACCCTCC TCCCTGGCCTGAGTTGAA--- 5'
3' GTCATGATGCCAATG5ʹ
3' ---GTCATG ATGCCAATG--- 5'
3' CGCTCGGGCATTCCCACACATGCCC
3' ---CGCTCGGGCATT CCCACACATGCCC--- 5'
3' ATTGATACTGAGAGAATTCCATCGGTTTA
3' ---ATTGATACTGAGAG AATTCCATCGGTTTA--- 5'
3' ACCGTTTGTCGATAATACCCATAATACCCA
3' ---ACCGTTTGTCGAT AATACCCATAATACCCA--- 5'
3' ACAGTGTAACTCCACGTGATCAATAATG
3' ---ACAGTGTAACTCCAC GTGATCAATAATG--- 5'
3' TCAATGCGATCCCTATTGTCCCATTATATC
3' ---TCAATGCGATCCC TATTGTCCCATTATATC--- 5'
TAGATACAGCCCACGCCTCTTTCTCCATTACTTTACCGT
3' ---TAGATACAGCC CACGCCTCTTTCTCCATTACTTTACCGT--- 5'
3' AAAACTAAGAAACCAGTGGGACTTCATAT
3' ---AAAACTAAGAAACCAG TGGGACTTCATAT--- 5'
3' TAACCTCCAAAACCATTGATAAATAATGG
3' ---TAACCTCCAAAACC ATTGATAAATAATGG--- 5'
3' AGAAAAGAGAACTAATCGGGATTAGATGC
3' ---AGAAAAGAGAAC TAATCGGGATTAGATGC--- 5'
3' TTATTAAAAGAAGAATCATTACGG
3' ---TTATTAAAAGAAGAATCATTA CGG--- 5'
3' CAATAAATTACAAAATCATCAACC
3' ---CAATAAATTACAAAATCATCA ACC--- 5'
3' ACAGTGTAACTCCACGTGATCAATAATG
3' CAGCCCGAGTATTGGGCTT
3' TCACCATAGT TGCGAGTCATCTAC
3' ---TCACCATAGT TGCGAGTCATCTAC--- 5'
3' CGAATACTCATACTTCACTTGTGCAATAAG
3' ---CGAATACTCATACTTCACTTGTG CAATAAG--- 5'
3' ATACATAGAAAACGCACATGGAAATTGAAG
3' ---AT ACATAGAAAACGCACATGGAAATTGAAG--- 5'
3' ATRCGNCTRTGNCTGCCTAARA
3' ---ATRCGNCTRTGNC TGCCTAARA--- 5'
3' TTTAACGAACGTTTGTCGATAATGCCGATA
3' GAAGTCATACGGGGCTTTG
3' ---GAAGT CATACGGGGCTTTG--- 5'
3' GGACTGAGAGAATTCCATCGGTTT
3' ---GGACTGAG AGAATTCCATCGGTTT--- 5'
Таблица 1: Обзор хоминг-эндонуклеаз и их последовательностей-мишеней.
Сокращения: SF Структурное семейство: HI: семейство LAGLIDADG; HII: семейство GIY-YIG; HIII: семейство H-N-H; HIV: семейство His-Cys-box.
D: Биологический домен источника: A: археи; B: бактерии; E: эукариоты.
SCL: Субклеточная локализация: chloro: хлоропласт; chrm: хромосома; mito: митохондрия; nuclear: экстрахромосомная ядерная локализация; phage: бактериофаг.
Таблица 2: Штаммы Saccharomyces cerevisiae, используемые в этом исследовании.
Ссылки:
Nijkamp et al., (2012) Microb Клетка Fact 11: 36
Entian & Kötter, (2007) Meth Microbiol 36: 629-666
Таблица 3: Праймеры
Таблица 4: Плазмиды, используемые в настоящем исследовании. a DR Прямой повтор.
Ссылка: Alberti et al., (2007) Yeast 24: 913-919
Таблица 5: Активности гуанидинобутиразы, измеряемые в экстрактах клеток штаммов IME215 и CEN.PK113-7D S. cerevisiae, выращенных в периодических культурах с аргинином в качестве единственного источника азота. Штаммы и IME215 S. cerevisiae предварительно выращивали на синтетической среде с глюкозой и аммонием в качестве единственного источника азота. B.D. означает ниже предела чувствительности, определяемого при 0,005 мкмоль мин-1 мг-1 белка. Данные представляют собой среднее ± среднее отклонение для независимых биологических параллельных структур.
Краткое описание чертежей
Фигура 1. Обзор ключевых реакций в метаболизме аргинина эукариот. Толстыми линиями указаны реакции уреогидролаз. EC 3.5.3.1: аргиназа, EC 4.1.1.17, орнитиндекарбоксилаза, EC 2.6.1.13: орнитинаминотрансфераза, EC 1.5.1.2: пирролин-5-карбоксилатредуктаза, EC 1.5.99.8: пролиндегидрогеназа, EC 1.5.1.12: 1-пирролин-5-карбоксилатдегидрогеназа, EC 2.6.1.-аминотрансфераза, EC 4.1.1.75 2-оксо-кислая декарбоксилаза, EC 1.2.1.54 гамма-гуанидинобутиральдегиддегидрогеназа, EC 3.5.3.7 гуанидинобутират, EC 2.6.1.19 ГАМК-трансаминаза, EC 1.2.1.16 сукцинат-полуальдегиддегидрогеназа.
Фигура 2.A Выравнивание аминокислотных последовательностей гуанидино-кислой гидролазы (EC.3.5.3.7) дрожжей Saccharomycotina. Аминокислотные последовательности выравнивали с использованием Clustal W (V1.7). Аминокислотные остатки, консервативные во всех последовательностях, указаны символом *. XP_456325: референсная GBU1 из NRRL Y-1140 Kluyveromyces lactis; XP_002498240: CBS 732 Zygosaccharomyces rouxii; XP_716668: SC5314 Candida albicans; XP_461566: CBS767 Debaryomyces hansenii; EIF45280: AWRI1499 Dekkera bruxellensis; BAO40383: DMKU3-1042 Kluyveromyces marxianus; AADM01000201.1: NCYC 2644 Lachancea waltii; XP_503530: CLIB122 Yarrowia lipolytica; EFW95653.1: DL-1 Ogataea parapolymorpha (Hansenula polymorpha); XP_002552049: CBS 6340 Lachancea thermotolerans (CBS6340 Kluyveromyces thermotolerans); XP_001523956: NRRL YB-4239 Lodderomyces elongisporus; XP_001482640: ATCC 6260 Meyerozyma guilliermondii (ATCC6260Pichia guilliermondii); XP_004196483: CBS 7064 Millerozyma farinosa (Pichia sorbitophila); XP_001385334: CBS 6054 Scheffersomyces stipitis (Pichia stipitis CBS 6054); XP_003679661: CBS 1146 Torulaspora delbrueckii; AACE03000003.1: NRRL Y-12651 Lachancea kluyveri. Консенсусная последовательность представлена под выравниванием последовательностей и аминокислотами, консервативными по меньшей мере в 50% последовательностей. Указанные полужирным шрифтом и подчеркнутые аминокислоты обозначают консервативные домены, обнаруженные в гуанидино-кислой гидролазе.
Посредством B-Weblogo (Schneider and Stephens (1990). Nucleic Acids Research 18, 6097-6100) представлены консервативные мотивы гуанидино-кислой гидролазы из выравнивания последовательностей.
Фигура 3. Запись белка K. lactis XP_456325.1 в GenBank.
Фигура 4. Выравнивание аминокислотных последовательностей бактериальной гуанидиноамидгидролазы (EC.3.5.3.11). A. Аминокислотные последовательности выравнивали с использованием Clustal W (V1.7). Аминокислотные остатки, консервативные во всех последовательностях, указаны символом *. NP_289508: Escherichia coli; WP_000105576: Shigella flexneri; YP_005016506: Klebsiella oxytoca KCTC 1686; YP_001337000: Klebsiella pneumoniae subsp. pneumoniae MGH 78578; YP_003614749: Enterobacter cloacae subsp. cloacae ATCC 13047; YP_001455807: Citrobacter koseri ATCC BAA-895; XP_004532666: Ceratitis capitata; WP_006734551: Salmonella enterica; AHE29794: Burkholderia pseudomallei NCTC 1317. B - Консенсусная последовательность, полученная при выравнивании последовательностей. Консенсусная последовательность представлена под выравниванием последовательностей и аминокислотами, консервативными по меньшей мере в 50% последовательностей. Указанные полужирным шрифтом и подчеркнутые аминокислоты обозначают консервативные домены, обнаруженные в гуанидиноамидгидролазе.
Посредством B-Weblogo представлены консервативные мотивы гуанидиноамидгидролазы из выравнивания последовательностей.
Фигура 5. Запись белка E. coli (агматиназы) AAC75974.1 в GenBank.
Фигура 6. Вектор 1 и 2 со всеми необходимыми частями для стандартной генной кассеты для делеции. Область перекрывания 400 оснований в селективном маркере KlGBU1 (указана крестом) сконструирована для рекомбинации по причине гомологии.
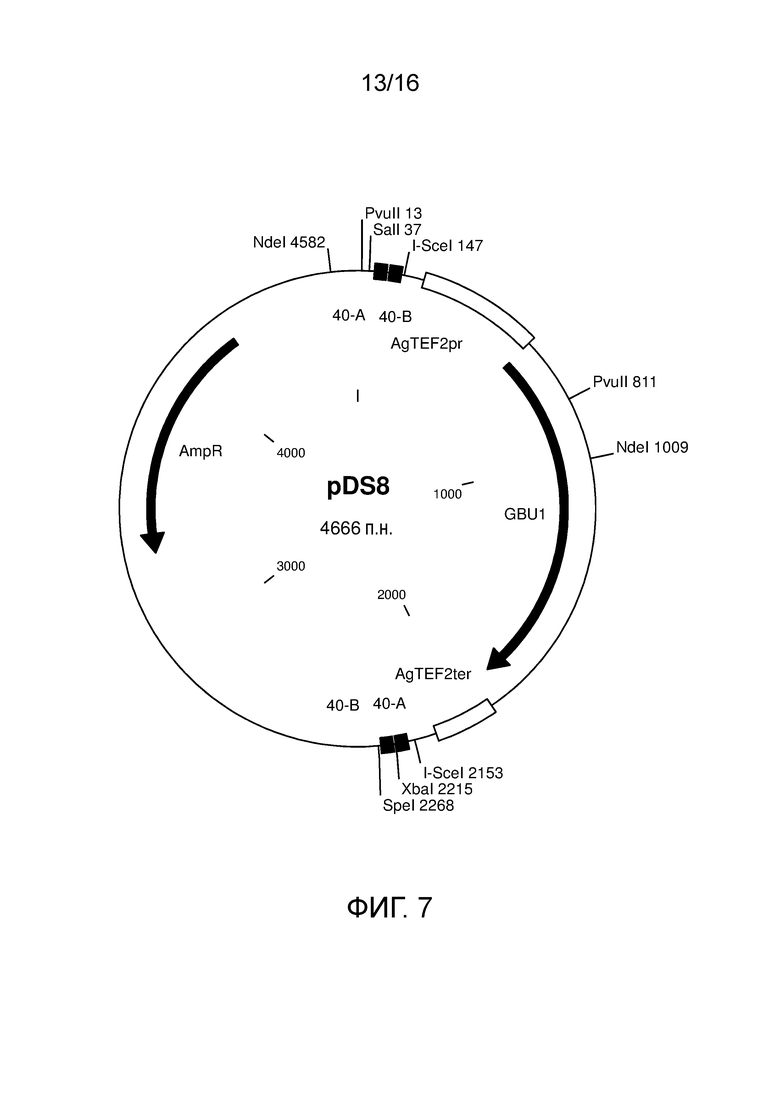
Фигура 7. Карта плазмиды pDS8, содержащая модуль маркера KlGBU1YM.
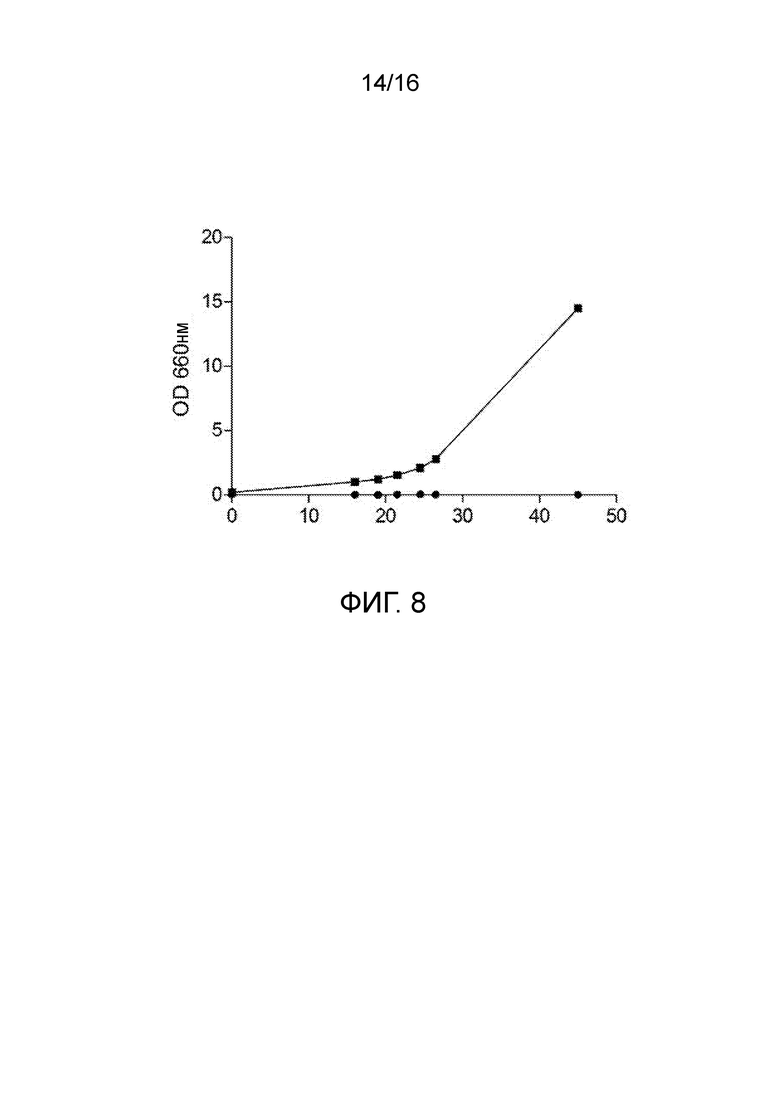
Фигура 8. Типичный профиль роста штаммов S. cerevisiae на гуанидинобутирате: штаммы CEN.PK113-7D (закрашенные круги) и IME215 (TDH3pr::KlGBU1::CYC1ter) (закрашенные квадраты) S. cerevisiae выращивали во встряхиваемых колбах на синтетической среде с глюкозой, содержащей гуанидинобутират в качестве единственного источника азота.
Фигура 9. Трансформация S. cerevisiae и S. pastorianus с использованием pUDE264. Штаммы CEN.PK113-7D S. cerevisiae и CBS1483 S. pastorianus трансформировали с использованием плазмиды pUDE264, несущей ген гуанидинобутиразы из K. lactis. Трансформированные клетки (50 мкл) высевали на SM с разными источниками азота ((NH4)2SO4, гуанидинобутиратом и агматином). Нетрансформированные штаммы (-ДНК) также высевали на схожие среды в качестве отрицательного контроля.
Фигура 10. Примеры генных кассет для делеции. Генные кассеты для делеции содержат несколько областей: A - (1) последовательность от 50 до 80 п.н., гомологичную вышележащей части гена, подлежащей делеции, включая инициаторный кодон, и последовательность от 50 до 80 п.н., гомологичную нижележащей части гена, подлежащей делеции, включая стоп-кодон, и (4) селектируемую кассету, включающую промотор TEF2 Ashbya gossypii, ген GBU1 K. lactis и терминатор TEF2A. gossypii. B - В дополнение к области (1) и (4) генная кассета для делеции включает (2) последовательность 40 п.н., фланкируемую (3) участком рестрикции I-SceI, локализованным выше и ниже модуля маркера.
Примеры
Пример 1
Материалы и способы
Штаммы и содержание
Штаммы Saccharomyces cerevisiae, используемые в настоящем исследовании, приведены в таблице 2. Штаммы S. cerevisiae конструировали в условиях CEN.PK (Nijkamp et al., (2012) Microb Cell Fact 11: 36; Entian & Kötter, (2007) Meth Microbiol 36: 629-666). Штаммы дрожжей, не несущие плазмиду, поддерживали на среде YPD (деминерализованная вода; 10 г/л дрожжевого экстракта; 20 г/л пептона; 20 г/л глюкозы). Штаммы дрожжей, несущие плазмиду, поддерживали на синтетической среде, содержащей соли, микроэлементы и витамины, полученной и стерилизованной, как описано ранее (Verduyn et al., (1992) Yeast 8: 501-517), в которой, по возможности, используют мочевину вместо сульфата аммония. Стоки культур получали из культур во встряхиваемых колбах, инкубируемых при 30°C и перемешиваемых при 200 об./мин., посредством добавления 20% (об./об.) глицерина и хранили при -80°C.
Среды и условия культивирования
Эксперименты по выращиванию осуществляли на синтетической среде, содержащей соли, микроэлементы и витамины, полученной и стерилизованной, как описано ранее (Verduyn et al., (1992) Yeast 8: 501-517). Добавляли глюкозу до конечной концентрации 20,0 г/л. Если сульфат аммония не являлся источником азота в синтетической среде, его заменяли гуанидинобутиратом, стерилизуемым посредством фильтрации и добавляемым в стерильную среду до концентрации 2,9 г/л. Кроме того, добавляли 3,3 г/л сульфата калия для компенсации удаления сульфата аммония.
При необходимости, добавляли 0,15 г/л урацила и/или 200 мг/л G418 (Geneticin) для завершения сред. Чашки с агаром для селекции получали посредством добавления 20,0 г/л агара в эти синтетические среды.
Культуры во встряхиваемых колбах получали во встряхиваемых колбах емкостью 500 мл или 250 мл, содержащих 100 мл или 20 мл жидкой среды, соответственно, и инкубировали на орбитальном шейкере (New Brunswick Scientific, Edison, NJ) при 200 об./мин. и 30°C.
Клонирование и гиперэкспрессия KLLA0F27995g K. lactis в S. cerevisiae.
Геномную ДНК прототрофного референсного штамма CEN.PK113-7D S. cerevisiae получали, как описано ранее (Burke et al., 2000. Cold Spring Harbor Laboratory. Methods in yeast genetics: a Cold Spring Harbor Laboratory course manual). ORF KLLA0F27995g (KlGBU1) клонировали из геномной ДНК с использованием полимеразы Phusion Hot-Start (Finnzymes) и прямого праймера GBU1/обратного праймера GBU1 из таблицы 3.
Продукты ПЦР расщепляли с использованием SpeI и XhoI, клонированных в pAG426GPDccdB (Alberti et al., (2007) Yeast 24: 913-919; Таблица 4), предварительно расщепленную теми же ферментами, с использованием лигазы T4 (Life Technologies, Breda, The Netherlands), получая плазмиду pUDE264.
С помощью плазмиды pUDE264 трансформировали CEN.PK113-5D S. cerevisiae (ura3-52) способом с использованием LiAc, как описано ранее (Gietz & Woods, (2002) Methods Enzymol 350: 87-96), получая штамм IME215 (MATa ura3-52 pUDE264 (TDH3pr-KlGBU1-CYC1ter URA3 2μ). С помощью плазмиды-основы pAG426GPD-ccdB трансформировали CEN.PK113-5D S. cerevisiae (ura3-52) способом с использованием LiAc, как описано ранее (Gietz & Woods, (2002) Methods Enzymol 350: 87-96), получая штамм IMZ312 (MATa ura3-52 pAG426GPD-ccdB (TDH3pr-CYC1ter URA3 2μ).
Конструирование плазмиды pDS8
ORF KLLA0F27995g (KlGBU1) переносили из pUDE264 в pDS1 вместо гена kanR. Для конструирования плазмиды pDS1 использовали вектор pUG6 (Guldener et al. (1996) Nucleic Acids Res 24:2519-24) в качестве матрицы для ПЦР с использованием ДНК-полимеразы Phusion™ Hot Start II (Thermo Scientific, Waltham, MA) по инструкциям производителя и пары праймеров pUGfw/pUGrv для амплификации основы вектора и pDSfw/pDSrv для амплификации селективного маркера. Праймер pUGfw содержал участок рестрикции XbaI и синтетическую последовательность 40 п.н. (повтор B), используемую для конструирования прямого повтора 80 п.н. Праймер pUGrv содержал участок рестрикции BglII и другую синтетическую последовательность 40 п.н. (повтор A) для конструирования второго прямого повтора 80 п.н. Праймер pDSfw содержал участок рестрикции BglII, затем повтор B 40 п.н. Праймер pDSrv содержал участок рестрикции XbaI и повтор A 40 п.н. Все ПЦР визуализировали с использованием электрофореза в геле. Фрагменты выделяли из агарозного геля с использованием набора для выделения ДНК из геля Zymoclean™ (Zymo Research, Irvine, CA). Фрагменты, полученные посредством ПЦР, рестрицировали с использованием XbaI и BglII и лигировали с использованием ДНК-лигазы T4 (Thermo Scientific) по инструкциям производителя. Химически компетентные Escherichia coli, штамм DH5α, трансформировали с использованием смеси для лигирования и правильные колонии выбирали посредством ПЦР с использованием следующих праймеров FK140 pUDI065 fw, SLT1_control_rv, KANMX4 fw и FK105-MP1.
Перенос KlGBU1 вместо последовательности kanR осуществляли посредством сборки по Гибсону (Gibson et al., (2009) Nat Methods 6: 343-345). Фрагмент, несущий ген KlGBU1, амплифицировали посредством ПЦР с использованием полимеразы Phusion Hot-Start (Finnzymes) и праймеров GBU1-fw и GBU1-rv (таблица 3). Плазмиду pUDE264 использовали в качестве матрицы. Плазмиду-основу амплифицировали посредством ПЦР с использованием полимеразы Phusion Hot-Start (Finnzymes) и праймеров Backbone-fw и Backbone-rv. Плазмиду pDS1 использовали в качестве матрицы. Два фрагмента обладали идентичностью по меньшей мере 43 нуклеотидов на их фланкирующих участках, и их собирали in vitro с использованием набора для клонирования со сборкой по Гибсону от New England Biolabs (Ipswich, MA). С помощью собранной смеси трансформировали DH5α E. coli и высевали на чашки с LB, содержащие ампициллин (100 мг/л). Собранную плазмиду, содержащую модуль маркера KlGBU1YM, подтверждали посредством рестрикционного анализа и секвенирования и правильный клон обозначали как pDS8.
Делеция ADE2 в S. cerevisiae.
Делеции генов в S. cerevisiae осуществляли посредством встраивания кассет KlGBU1YM (участок SceI::участок AgTEF2pr-KlGBU1-AgTEF2ter::SceI) посредством ПЦР гомологии коротких фланкирующих последовательностей (Wach et al., (1994) Yeast 10: 1793-1808). Последовательности олигонуклеотидных праймеров представлены в таблице 3. Генную кассету для делеции для ADE2 амплифицировали с использованием полимеразы Phusion Hot-Start (Finnzymes, Landsmeer, The Netherlands) и плазмиды-матрицы pDS8 с использованием праймеров GBU1-ADE2ko-CENPK-fw/GBU1-ADE2ko-CENPK-rv. Трансформацию CEN.PK113-7D S. cerevisiae с использованием генных кассет для делеции ADE2 осуществляли способом с использованием LiAc, как описано ранее в (Gietz & Woods, (2002) Methods Enzymol 350: 87-96), получая штамм IMX598.
Правильное встраивание кассеты KlGBU1SY и замену интересующего гена подтверждали посредством диагностической ПЦР с использованием прямого праймера, специфичного для 5ʹ-UTR (нетранслируемой области) ADE2, и обратного праймера для генной кассеты для делеции (таблица 3).
Получение экстрактов клеток
Для получения экстрактов клеток образцы культуры собирали посредством центрифугирования, дважды промывали 10 мМ буфером фосфата калия (pH 7,5), содержащим 2 мМ ЭДТА, и хранили при -20°C. Перед разрушением клеток образцы размораживали при комнатной температуре, промывали и ресуспендировали в 100 мМ буфере фосфата калия (pH 7,5), содержащем 2 мМ MgCl2 и 2 мМ дитиотреитол. Экстракты получали посредством обработки ультразвуком со стеклянными бусами диаметром 0,7 мм при 0°C в течение 2 мин при интервалах 0,5 мин с использованием соникатора MSE (Wolf Laboratories Limited, Pocklington, United Kingdom) (мощность 150 Вт; двойная амплитуда 8 мкм) (Luttik et al., (2008) Metab Eng 10: 141-153). Неразрушенные клетки и дебрис удаляли посредством центрифугирования при 4°C (20 мин; 36000×g). Полученный экстракт клеток использовали для анализов ферментов.
Анализы ферментативной активности
Для ферментативного анализа аргиназы 50 мкл экстракта клеток активировали в 950 мкл буфера малеата марганца (50 мМ сульфат марганца, 50 мМ малеиновая кислота, pH 7) в течение 1 ч. при 37°C (Messenguy et al., (1971) Eur J Biochem 22: 277-286). Реакционная смесь для анализов аргиназы, полученная в темных пробирках Eppendorf, содержала 60 мкл активированного экстракта клеток, 400 мкл раствор 713 мМ аргинина (pH 9,5) и деминерализованную воду до 1 мл. Реакционную смесь инкубировали в течение 30 мин при 37°C. Для остановки реакции в реакционную смесь добавляли 0,7 мл смеси серной и фосфорной кислот (20% об./об. концентрированной серной кислоты и 60% об./об. сиропообразной фосфорной кислоты в деминерализованной воде). Количество образовавшейся мочевины определяли способом Арчибальда (Archibald, (1945) J Biol Chem 157: 507-518) с калибровочной кривой в диапазоне от 0 до 0,6 мМ мочевины. Добавляли 0,06 мл 4% об./об. α-изонитрозопропиофенона в растворе этанола и образцы тщательно смешивали перед кипячением в течением 1 ч. на водяной бане с температурой 100°C для появления окраски. Образцы охлаждали при комнатной температуре в течение 15 мин и измеряли поглощение при 540 нм с помощью спектрофотометра Libra S11 (Biochrom, Cambridge, United Kingdom).
Реакционную смесь для анализов фермента гуанидинобутиразы (GBU) получали в темных пробирках Eppendorf, содержащих в конечном объеме 1 мл: 50 мМ буфера глицина (pH 9), 5 мМ MnSO4 и от 50 мкл до 100 мкл экстракта клеток. Реакцию инициировали посредством добавления 50 мМ гуанидиномасляной кислоты. Через 30 мин инкубации при 37°C реакцию останавливали посредством добавления 700 мкл смеси серной и фосфорной кислот (20% об./об. концентрированной серной кислоты и 60% об./об. сиропообразной фосфорной кислоты в деминерализованной воде). Количество образовавшейся мочевины определяли способом Арчибальда (Archibald, 1945) с калибровочной кривой в диапазоне от 0 до 0,6 мМ мочевины. Добавляли 0,06 мл 4% об./об. α-изонитрозопропиофенона в растворе этанола и образцы тщательно смешивали перед кипячением в течение 1 ч. на водяной бане с температурой 100°C для появления окраски. Образцы охлаждали при комнатной температуре в течение 15 мин и измеряли поглощение при 540 нм с помощью спектрофотометра Libra S11 (Biochrom).
Результаты
Экспрессия KlGBU1 в S. cerevisiae придает способность расти на гуанидинобутирате в качестве единственного источника азота.
Для характеризации функции предполагаемого гена уреогидролазы K. lactis клонировали KLLA0F27995g под контролем сильного конститутивного промотора TDH3 (TDH3pr) в экспрессирующем векторе и с его помощью трансформировали S. cerevisiae (фигура 8).
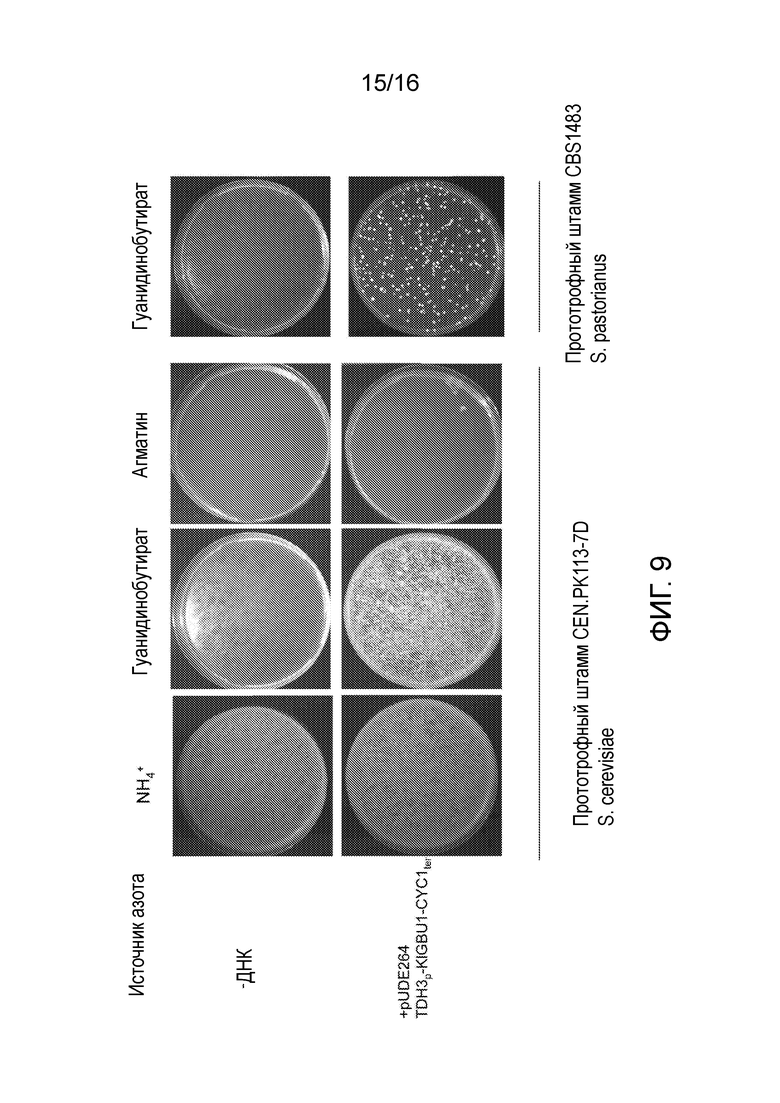
Посредством экспрессирующей плазмиды pUDE264 трансформировали лабораторный штамм CEN.PK113.7D (прототроф) и штамм CBS1483 Saccharomyces pastorianus дикого типа для пивоварения (www.cbs.knaw.nl/Collections/). Трансформированные клетки высевали на синтетической среде, содержащей гуанидинобутират или агматин или сульфат аммония. Как и следовало ожидать, в случае обоих штаммов все трансформанты могли расти на неселективных чашках, содержащих сульфат аммония. В соответствии с измерениями ферментов не получали трансформанты на чашках с агматином, в то время как более тысячи и сотни трансформантов определяли на чашках с гуанидинобутиратом в случае CEN.PK113-7D и CBS1483, соответственно (фигура 9). Это свидетельствует о том, что KlGBU1 можно эффективно использовать в качестве доминантного селективного маркера в прототрофном штамме S. pastorianus.
Конструирование плазмид и генных кассет для делеции.
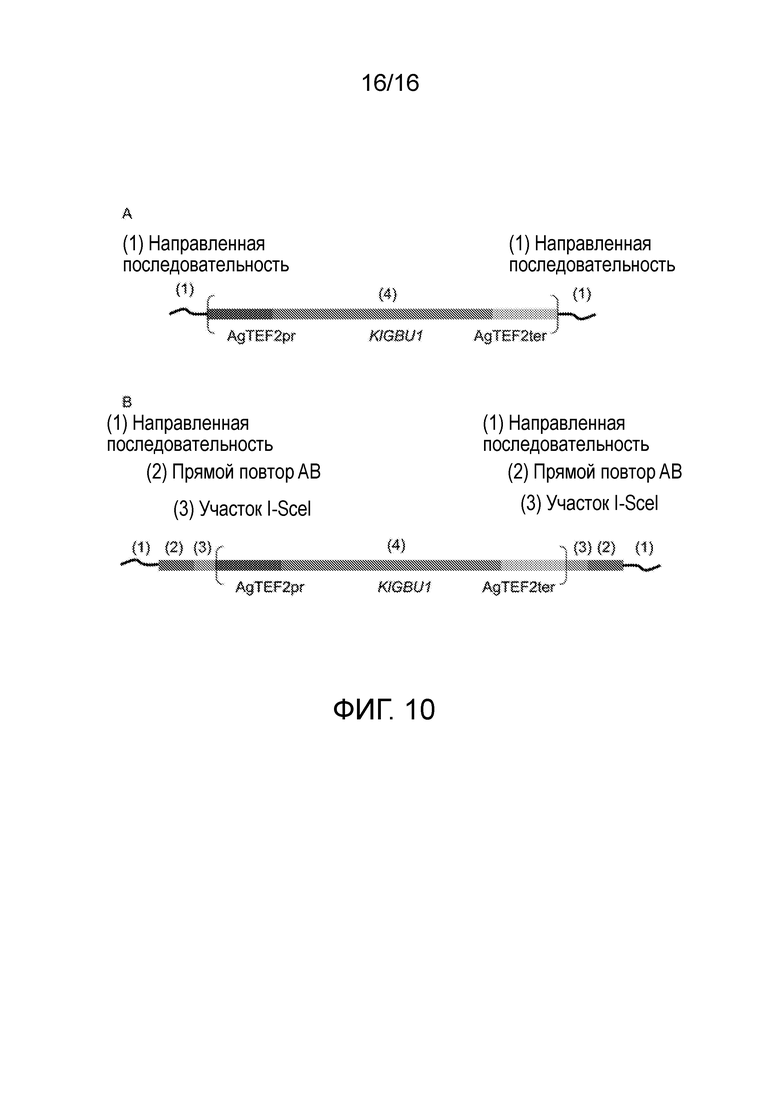
Кодирующую последовательность гена GBU1 K. lactis, фланкируемую промотором TEF2 и терминатором Ashbya. gossypii, клонировали в вектор pDS1, заменяя ген KanMX, получая плазмиду pDS8 (фигура 7). Получаемый модуль KlGBU1YM содержал лишь гетерологичные последовательности, таким образом, снижая вероятность неправильного встраивания (Wach et al., (1994) Yeast 10: 1793-1808). Плазмиду pDS8 легко можно использовать в качестве матрицы для генных кассет для делеции, содержащих новый модуль маркера KlGBU1YM, и ее использовали для конструирования всех генных кассет для делеции, используемых в настоящем исследовании.
Генные кассеты для делеции содержали три основные области (фиг. 10): (1) последовательность от 50 до 55 п.н., гомологичную вышележащей части гена, подлежащей делеции, включая инициаторный кодон, и последовательность от 50 до 55 п.н., гомологичную нижележащей части гена, подлежащей делеции, включая стоп-кодон. Эти области использовали для направленной гомологичной рекомбинации (Baudin et al., (1993) Nucleic Acids Res 21: 3329-3330), (2) последовательность 40 п.н., фланкируемую (3) участком рестрикции I-SceI, локализованным выше и ниже модуля маркера, и (4) маркер KlGBU1YM. После рестрикции эндонуклеазой гомологичная рекомбинация прямого повтора будет достаточной для удаления и рециркуляции модуля маркера.
Делеция гена в S. cerevisiae с использованием KlGBU1YM
Для оценки того, является ли новый маркер KlGBU1YM подходящим для нокаута гена у S. cerevisiae, пытались подвергнуть делеции ген в лабораторном штамме CEN.PK113-7D. Для этого контрольно-проверочного эксперимента ADE2 подвергали селекции, т.к. фенотип, вызываемый делецией ADE2, можно подвергать визуальному скринингу, осуществляя быструю предварительную оценку направленного встраивания. ADE2 кодирует фермент фосфорибозиламиноимидазолкарбоксилазу, участвующую в биосинтезе пуриновых нуклеотидов. Мутантам по ade2 необходим внешний источник аденина, и они накапливают предшественники пуриновых нуклеотидов в вакуолях, что придает колониям красный цвет (Zonneveld & van der Zanden, (1995) Yeast 11: 823-827).
Потенциал KlGBU1YM в качестве доминантного маркера тестировали посредством трансформации с использованием генной кассеты для делеции для разрушения ADE2 в CEN.PK113-7D. После трансформации клетки выращивали на чашках с агарной синтетической средой (SM), содержащей гуанидинобутират в качестве единственного источника азота. Направленную делецию гена подтверждали по неспособности отдельных колоний расти на SM (в отсутствие аденина) и посредством ПЦР. Средняя эффективность трансформации составляла 5 трансформантов на микрограмм ДНК, при этом 100% колоний имели правильное встраивание и были способны расти на гуанидинобутирате в качестве единственного источника азота.
























































































Изобретение относится к области молекулярной биологии. Предложен способ культивирования микроорганизма, выбранного из родов Saccharomyces sensu stricto, Kazachstania, Naumovozyma, Nakaseomyces и Vanderwaltozyma, в присутствии гуанидинобутирата в качестве единственного источника азота, включающий встраивание в указанный микроорганизм молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую гуанидинобутиразу c номером EC 3.5.3.7 и функционально связанную с последовательностями промотора и терминатора, последующее культивирование указанного микроорганизма таким образом, что молекула нуклеиновой кислоты, кодирующая гуанидинобутиразу, экспрессируется в указанном микроорганизме, и культивирование указанного микроорганизма в присутствии гуанидинобутирата в качестве единственного источника азота. Предложен также микроорганизм для роста на гуанидинобутирате в качестве единственного источника азота, выбранный из родов Saccharomyces sensu stricto, Kazachstania, Naumovozyma, Nakaseomyces и Vanderwaltozyma, содержащий нуклеотидную последовательность, кодирующую гуанидинобутиразу с номером EC 3.5.3.7. Предложен также способ изменения генома микроорганизма, выбранного из родов Saccharomyces sensu stricto, Kazachstania, Naumovozyma, Nakaseomyces и Vanderwaltozyma. Изобретение обеспечивает эффективное использование гуанидинобутиразы в качестве селективного маркера. 3 н. и 7 з.п. ф-лы, 10 ил., 5 табл., 1 пр.
1. Способ культивирования микроорганизма, выбранного из родов Saccharomyces sensu stricto, Kazachstania, Naumovozyma, Nakaseomyces и Vanderwaltozyma, в присутствии гуанидинобутирата в качестве единственного источника азота, включающий:
(a) встраивание молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую гуанидинобутиразу c номером EC 3.5.3.7, в указанный микроорганизм, при этом нуклеотидная последовательность функционально связана с последовательностями промотора и терминатора;
(b) культивирование указанного микроорганизма таким образом, что молекула нуклеиновой кислоты, кодирующая гуанидинобутиразу, экспрессируется в указанном микроорганизме; и
(c) культивирование указанного микроорганизма в присутствии гуанидинобутирата в качестве единственного источника азота.
2. Способ по п. 1, где указанная нуклеотидная последовательность, кодирующая гуанидинобутиразу, кодирует гипотетический белок NRRL Y-1140 Kluyveromyces lactis, имеющий последовательность, как показано на фигуре 3.
3. Способ по п. 1 или 2, где последовательности промотора и/или терминатора выбраны из гликолитического гена, выбранного из PGI1, PFK1, PFK2, FBA1, TPI1, TDH1, TDH3, PGK1, GPM1, PYK1, ENO1 и/или ENO2.
4. Микроорганизм для роста на гуанидинобутирате в качестве единственного источника азота, выбранный из родов Saccharomyces sensu stricto, Kazachstania, Naumovozyma, Nakaseomyces и Vanderwaltozyma, содержащий нуклеотидную последовательность, кодирующую гуанидинобутиразу с номером EC 3.5.3.7.
5. Микроорганизм, выбранный из родов Saccharomyces sensu stricto, Kazachstania, Naumovozyma, Nakaseomyces и Vanderwaltozyma, по п. 4, где указанная нуклеотидная последовательность, кодирующая гуанидинобутиразу, кодирует гипотетический белок NRRL Y-1140 Kluyveromyces lactis, имеющий последовательность, как показано на фигуре 3.
6. Способ изменения генома микроорганизма, выбранного из родов Saccharomyces sensu stricto, Kazachstania, Naumovozyma, Nakaseomyces и Vanderwaltozyma, включающий снабжение набором конструкций указанного микроорганизма, включающим первую конструкцию, содержащую первую часть нуклеотидной последовательности, кодирующей гуанидинобутиразу с номером ЕС 3.5.3.7, и вторую конструкцию, содержащую вторую часть нуклеотидной последовательности, кодирующей гуанидинобутиразу, при этом фрагмент первой части нуклеотидной последовательности перекрывается с фрагментом, присутствующим во второй части нуклеотидной последовательности, делая возможной рекомбинацию между первой и второй частью нуклеотидной последовательности и выбор микроорганизма, в котором изменяют геном посредством инсерции функциональной рекомбинированной гуанидинобутиразы с номером ЕС 3.5.3.7 в качестве селективного маркера.
7. Способ по п. 6, в котором перекрывающиеся фрагменты нуклеотидной последовательности, кодирующей гуанидинобутиразу с номером ЕС 3.5.3.7 в первой и второй конструкциях, составляют от 40 до 400 п.н.
8. Способ по п. 6 или 7, в котором перекрывающиеся фрагменты нуклеотидной последовательности, кодирующей гуанидинобутиразу с номером ЕС 3.5.3.7 в первой и второй конструкциях, составляют приблизительно 200 п.н.
9. Способ по любому из пп. 6-8, в котором ген-мишень изменен в геноме микроорганизма, выбранного из родов Saccharomyces sensu stricto, Kazachstania, Naumovozyma, Nakaseomyces и Vanderwaltozyma.
10. Способ по любому из пп. 6-9, в котором микроорганизм выбран посредством культивирования микроорганизма в присутствии гуанидинобутирата в качестве единственного источника азота.
| FAIRHEAD C | |||
| ET AL | |||
| New Vectors for Combinatorial Deletions in Yeast Chromosomes and for Gap-repair Cloning using "Split-marker" Recombination | |||
| YEAST, 1996, Vol | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| DAVE K | |||
| ET AL | |||
| A novel selectable marker based on Aspergillus niger arginase expression | |||
| Enzyme and Microbial Technology, 51, (2012), 53- 58 | |||
| NIELSEN M.L | |||
| ET AL | |||
| Efficient PCR-based gene targeting with a recyclable marker for Aspergillus nidulans | |||
| Зубчатое колесо со сменным зубчатым ободом | 1922 |
|
SU43A1 |
| МИХАЙЛОВА Ю.В | |||
| Сравнительная геномика дрожжей Saccharomyces | |||
| Автореферат дис | |||
| кандидата биологических наук.- Москва, 2009 | |||
| Пишущая машина для тюркско-арабского шрифта | 1922 |
|
SU24A1 |

