Область техники
Настоящее изобретение относится к области генной инженерии и биотехнологии, в частности к созданию вектора-хозяина для генетической трансформации дрожжей Candida utilis, что обеспечивает экспрессию и секрецию гетерологичных белков в этих дрожжах, которые в дальнейшем можно использовать для разных целей.
Предпосылки изобретения
Генная инженерия и биотехнология открывают широкие возможности для получения многих интересных белков, применение которых в медицинских, пищевых или промышленных целях является весьма перспективным.
Компании, работающие в области биотехнологии, особенно широко используют бактерии Escherichia coli благодаря знанию их генетики, простоте обращения и возможности их культивирования с высокой плотностью.
Однако на получение требуемых белков в этих микроорганизмах влияют разные факторы. Прежде всего следует отметить, что наличие пирогенных и токсичных соединения в клеточной стенке Escherichia coli ограничивает применение этих микроорганизмов для получения веществ, используемых для изготовления лекарственных средств или продуктов питания для людей. Кроме того, белки, которые в избытке экспрессированы в Escherichia coli, обычно имеют нерастворимую форму и поэтому не могут быть секретированы. С другой стороны, механизмы транскрипционных, трансляционных и посттрансляционных модификаций отличаются от эукариотных систем, что ведет к образованию рекомбинантных белков, которые некоторым образом отличаются от белков, получаемых из природных источников.
Возможность продуцирования гетерологичных белков в эукариотных системах, таких как дрожжи, характеризуется рядом преимуществ по сравнению с прокариотными системами. К таким преимуществам относится получение указанных клеток с большой плотностью и возможность их культивирования в системах непрерывного действия. Кроме того, эти дрожжи способны секретировать белки в культуральную среду в гораздо больших количествах, чем Escherichia coli, и среды для выращивания указанных дрожжей являются более экономичными по сравнению со средами для выращивания бактерий (Lemoine, Y., 1988. Heteroiogous expression in yeast. 8th International Biotechnology Symposium, Париж, 17-22 июля).
Эти системы могут также включать другие посттрансдукционные модификации, имеющие место в случае гликозилирования, которое отсутствует в бактериальных системах (Fiers, W., 1988. Engineering Maximal Expression of Heterologous Gene in Microorganism, 8th International Biotechnology Symposium, Париж, 17-22 июля). Кроме того, эти системы обычно предпочтительно используют тот же кодон, что и высшие эукариотные системы (Kigsman, S.M. et al., 1990. Heterologous Gene Expression in Saccharomyces cerevisiae. Biotechnology & Genetic Engineering Reviews, 3, Ed. G.E. Russell).
Все эти факторы легли в основу создания новых эукариотных систем трансформации, особенно полезных для дрожжей, т.к. такие системы первоначально были описаны для рода Saccharomyces, причем особое внимание было уделено виду Saccharomyces cerevisiae. Однако экспрессия белков в Saccharomyces вызывает проблемы, связанные с уровнями экспрессии, достигаемыми при использовании гомологичных промоторов, а также с гипергликозилированием белков, секретируемых в среду. Именно это заставило обратиться к исследованию менее известных дрожжей с целью их использования для экспрессии гетерологичных белков.
Создание системы трансформации дрожжей, не относящихся к роду Saccharomyces, в частности Hansenula polymorpha, Pichia pastoris, и дрожжей рода Kluyveromyces (Sudbery, P., 1994, Yeast 10: 1707-1726), позволило всесторонне изучить и исследовать эти системы, а также увеличить количество чужеродных белков, экспрессируемых в этих системах, предназначенных для вакцинации, диагностики и промышленного применения.
Кроме того, в научной литературе было описано несколько систем трансформации и экспрессии дрожжей рода Candida, в том числе Candida tropicalis, Candida boidiini, Candida glabrata, Candida parapsilosis, Candida maltosa и Candida albicans, которые представляют интерес с медицинской точки зрения, так как многие эти виды являются причиной возникновения у людей заболеваний, вызываемых условно-патогенными микроорганизмами.
Candida utilis, относящийся к роду Candida, представляет особый интерес благодаря своим уникальным характеристикам. Прежде всего, Candida utilis использует целый ряд недорогих источников углерода, таких как ксилоза, сахароза и мальтоза. Другой интересной особенностью является то, что большое количество клеток можно эффективно продуцировать в непрерывных культурах. Candida utilis, также как Saccharomyces cerevisiae и Kluyveromyces lactis, сертифицированы Управлением по контролю за продуктами и лекарствами (FDA) как безопасные источники для производства продуктов питания. Кроме того, вид Candida utilis используют в промышленности для производства L-глутамина, этилацетата, инвертазы и других продуктов.
Первоначальная система трансформации Candida utilis была описана Но I. и др. (Но I. et al., 1984, Biotechnology and Bioengineering Symp. 14: 295-301). Это описание является неполным, так как в нем не рассмотрены устойчивые к лекарственным средствам маркеры и не приведены данные, относящиеся к процессу трансформации. Недавно была разработана новая стратегия создания системы трансформации Candida utilis (Kondo, К. et al., 1995, J. Bacteriol. 177:7171-7177). Авторы этой статьи получили устойчивые к циклогексимиду (СНУ) трансформанты, используя ген-маркер, который содержит мутированную форму рибосомного белка L41, сообщающую требуемую устойчивость, и фрагмент рибосомной ДНК (рДНК) в качестве мультикопийной мишени для интеграции плазмиды, так как маркер должен присутствовать в нескольких копиях для выбора CHY-устойчивых трансформантов.
Было предпринято много попыток использовать Candida utilis в качестве хозяина для экспрессии гетерологичного гена, и тем не менее до сих пор не разработан метод трансформации Candida utilis с использованием ауксотрофных мутантов.
С учетом данных, полученных в результате промышленного применения Candida utilis, и новизны генетики этого вида указанные микроорганизмы можно считать весьма привлекательными для коммерческого применения в качестве системы, экспрессирующей гетерологичные белки.
Краткое изложение существа изобретения
Объектом настоящего изобретения является создание системы трансформации дрожжей Candida utilis с целью экспрессии гетерологичных белков, в основе которой лежит получение ауксотрофных мутантов этого вида и выделение из геномной библиотеки разных генов, комплементирующих указанные ауксотрофные мутанты.
Процесс трансформации, рассматриваемый в этом описании изобретения, обеспечивает введение фрагментов или последовательностей ДНК в клетки-хозяева Candida utilis и использование Candida utilis в качестве системы-хозяина для экспрессии гена и продуцирования белка.
Кроме того, с помощью методов по настоящему изобретению можно идентифицировать и отбирать трансформированные дрожжевые клетки. В объем настоящего изобретения входят также новые штаммы Candida utilis, векторы и субклоны. Новые штаммы дрожжей используют в качестве хозяев для введения фрагментов рекомбинантной ДНК.
Настоящее изобретение далее относится к стабильной трансформации и сохранению ДНК в клетках-хозяевах, где маркер гомологично интегрирован в геном дрожжей.
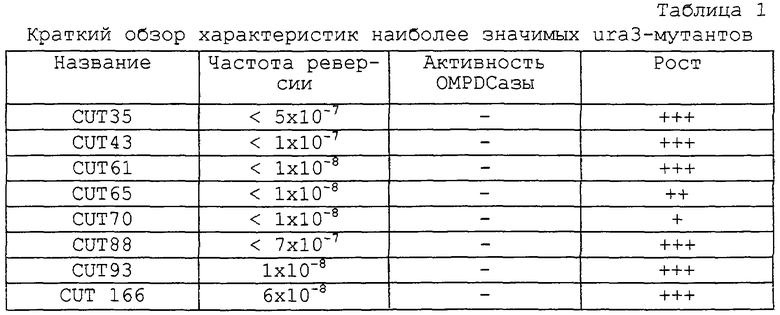
Настоящее изобретение, в частности, относится к системе трансформации дрожжей Candida utilis, где в качестве хозяев использованы новые ауксотрофные мутанты, выделенные из штамма NRRL Y-1084 указанных дрожжей. У этих мутантов отсутствует фермент оротидин-5’-фосфат-декарбоксилаза, биосинтетического пути метаболизма урацила или биосинтетического пути гистидина, и эти мутанты получают посредством классического мутагенеза под действием ультрафиолета и нефротоксического глобулина (NTG), которые являются хорошо известными мутагенными средствами (Sherman, F. et al., 1986.Laboratory course: Manual for methods in yeast genetics. Cold. Spring Harbor Laboratory Press, NY). Указанные мутанты характеризуются высокой стабильностью (частота реверсии равна примерно 10-8) и могут быть эффективно трансформированы в соответствии со способом по настоящему изобретению.
Кроме того, получены селективные маркеры для мутантов Candida utilis, а именно ген URA3, кодирующий фермент оротидин-5’-фосфат-декарбоксилаза, и ген HIS3, кодирующий фермент имидазолглицеролфосфат-дегидратаза, которые выделены из библиотеки генов Candida utilis в pUC19 и идентифицированы путем комплементации мутаций pyrF и hisb463 в штамме Escherichia coli MC1066. Мутацию ura3 штамма SEY 2202 Saccharomyces cerevisiae аналогичным образом используют для идентификации этого гена, выделенного из Candida utilis. Определена полная последовательность этих генов, при этом прогнозируемые аминокислотные последовательности обладают большим сходством с аналогичными последовательностями того же гена, выделенного из других дрожжей и грибов.
В качестве векторов в этой системе трансформации использованы плазмиды pURA5 и pUREC3, содержащие ген URA3, который можно интегрировать в гомологичный локус мутанта-хозяина Candida utilis в результате гомологичной рекомбинации.
Настоящее изобретение относится также к набору плазмид, получаемых на основании вышеописанных плазмид, которые используют для трансформации мутантов, выделенных из Candida utilis, с целью получения гетерологичных белков.
В системе трансформации по настоящему изобретению в качестве хозяев использованы новые ауксотрофные мутанты, полученные из штамма NRRL Y-1084 Candida utilis, в которых отсутствуют пути метаболизма урацила и гистидина. Среди этих мутантов наиболее предпочтительными благодаря своим характеристикам являются мутант CUT-35 (ura-) и мутант TMN-3 (his-).
Пример 1
Мутагенез Candida utilis
Для развития системы трансформации микроорганизма обычно необходимы три элемента:
(1) маркер для селекции трансформантов, который может быть ауксотрофным или доминантным маркером;
(2) мутант или приемлемый хозяин для такой селекции и
(3) метод воспроизводимого введения внеклеточной ДНК в клетку-хозяина в эффективном виде.
Для достижения второй цели выполняют классический мутагенез дрожжей Candida utilis. Культуры выбранного штамма дрожжей (NRRL Y-1084) инокулируют в 100 мл YPG-среды (1% дрожжевого экстракта, 2% пептона, 2% глюкозы) и инкубируют в шейкере при температуре 30°С в течение 10-20 часов. Полученную культуру (50 мл) центрифугируют со скоростью 3000 об/мин в течение 5 мин. Клетки дважды промывают 0,1 М раствором стерильного цитратного буфера (рН 5,5) и вновь суспендируют в 50 мл того же буфера. Затем 10 мл этой суспензии инкубируют с раствором нефротоксического глобулина (NTG) до достижения конечной концентрации, равной 50 мг/мл. Эту суспензию инкубируют в состоянии покоя при температуре 30°С в течение 30 мин.
Нефротоксический глобулин удаляют из суспензии и дважды промывают дистиллированной водой. Полученные клетки вновь суспендируют в 50 мл YPG-среды, после чего их переносят в аппарат Эрленмейера, содержащий 100 мл YPG-среды. Эту культуру мутантных клеток инкубируют при температуре 30°С в течение 48 часов.
Обогащением нистатином
Примерно 5 мл культуры, экспрессированной в YPG-среде в течение 48 часов, используют для инокуляции 100 мл минимальной среды. Минимальная среда (YNB, Yeast Nitrogen Base), используемая для обогащения антибиотиком, не должна содержать метаболит, который образуется в соответствии с биосинтетическим путем метаболизма, имеющим искомый дефект. Например, в среду не добавляют урацил, если нужно выделить ауксотрофные мутанты для этого вещества.
Культуру инкубируют до достижения оптической плотности, равной 20-30% исходного показателя. После достижения требуемой оптической плотности суспензию клеток обрабатывают 25 единицами нистатина на 1 мл раствора. Содержащий антибиотик раствор инкубируют, не перемешивая, при температуре 30°С в течение 30 мин. Нистатин удаляют из среды, дважды промывая суспензию клеток дистиллированной водой, после чего клетки вновь суспендируют в требуемом объеме, чтобы получить 150-200 колоний на одном планшете.
Скрининг и селекция
Планшеты, содержащие мутагенные колонии по примеру 1, культивируют в YNB-средах с урацилом или без него. Для дальнейшего анализа отбирают колонии, не растущие без урацила.
В частности, чтобы идентифицировать ura3- или ura5-мутанты, клетки выращивают в присутствии 5-фтороротовой кислоты (5FOA). Устойчивые колонии отбирают в качестве ura3-или urа5-подобных мутантов.
Пример 2
Выделение иrа3-мутантов
Обогащенную нистатином культуру дважды промывают дистиллированной водой и культивируют на планшетах с YNB-средой, содержащей 0,75 мкг/мл 5-FOA (5-фтороротовая кислота, Fluka) и 40 мкг/мл урацила. Планшеты инкубируют в течение четырех дней, после чего выросшие колонии анализируют с целью обнаружения фенотипа urа". Из 4×104 жизнеспособных клеток, полученных после обогащения нистатином, 79 колоний характеризуются устойчивостью к 5-FOA. Эти колонии могут быть ura3, ura5 или просто устойчивыми к 5-FOA. Чтобы подтвердить ауксотрофность урацила, предполагаемые мутанты помещают в YPG-среду, инкубируют в течение 48 часов при температуре 30°С и реплицируют на планшетах, содержащих YNB-среду с урацилом или без него. В общей сложности 67 колоний неспособны расти в YNB-среде без урацила, что свидетельствует о наличии фенотипа urа-.
Частоту реверсии всех этих мутантов определяют, исследуя группу из 23 мутантов с частотой реверсии порядка 10-8, которая сообщает им стабильность, необходимую для использования в качестве хозяина для системы трансформации.
Активность оротидин-5’-монофосфат-декарбоксилазы (ОДК-аза) всех урацилауксотрофных мутантов определяют по методу Йошимото и др. (Yoshimoto et al., 1978. Methods Enzymol. 52:74-79), который используют также для определения условий их роста. Эти результаты приведены в табл.1.
Пример 3
Выделение других мутантов, не относящихся к фенотипу ura-
Чтобы создать ряд ауксотрофных мутантов, отличающихся от урациловых, суспензию клеток, полученную в результате обогащения нистатином, помещают в YPG-среду и инкубируют при температуре 30°С в течение 50 часов. Затем колонии, находящиеся на планшетах с YPG-средой, реплицируют на планшетах, содержащих YNB-среду, и инкубируют при температуре 30°С в течение 48 часов. Для дальнейшего анализа отбирают колонии, не растущие на планшетах с YNB-средой.
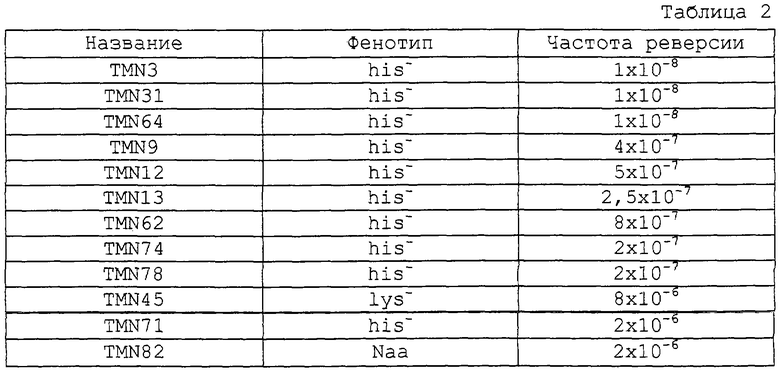
В результате исследования 2411 колоний было получено 2% ауксотрофных мутантов. Эти мутанты анализируют в соответствии с критериями Холлидея и Финчана. Результаты показывают, что 90% его мутантов имеют фенотип his-, 2% соответствуют фенотипу lys-, 1% соответствует фенотипу leu-, 1% соответствует фенотипу met-, 1% соответствует фенотипу ade- и 5% не относятся к простому ауксотрофному фенотипу (Naa).
Для дальнейшего анализа отбирают мутанты с частотой реверсии в диапазоне от 10-7 до 10-8 (табл.2).
Пример 4
Конструкция геномной библиотеки Candida utilis
Хромосомную ДНК, экстрагированную из штамма NRRL Y-1084 Candida utilis, частично расщепляют ферментом Sаu3А и с помощью электрофореза в низкотемпературном агарозном геле (LGT) выделяют фрагменты длиной от 6 до 9 т.п.о. Эти фрагменты лигируют в вектор pUC19, который предварительно расщепляют BamHI и обрабатывают щелочной фосфатазой. Это лигирование трансформируют в штамме МС 1066 Escherichia coli (F’, D Lacx74, hsr, hsm, rpsl, galU, galK, trip С 9030F, ieuB, pyrF::tn5). Геномная библиотека включает примерно 95% рекомбинантов.
Пример 5
Выделение гена URA3 из Candida utilis
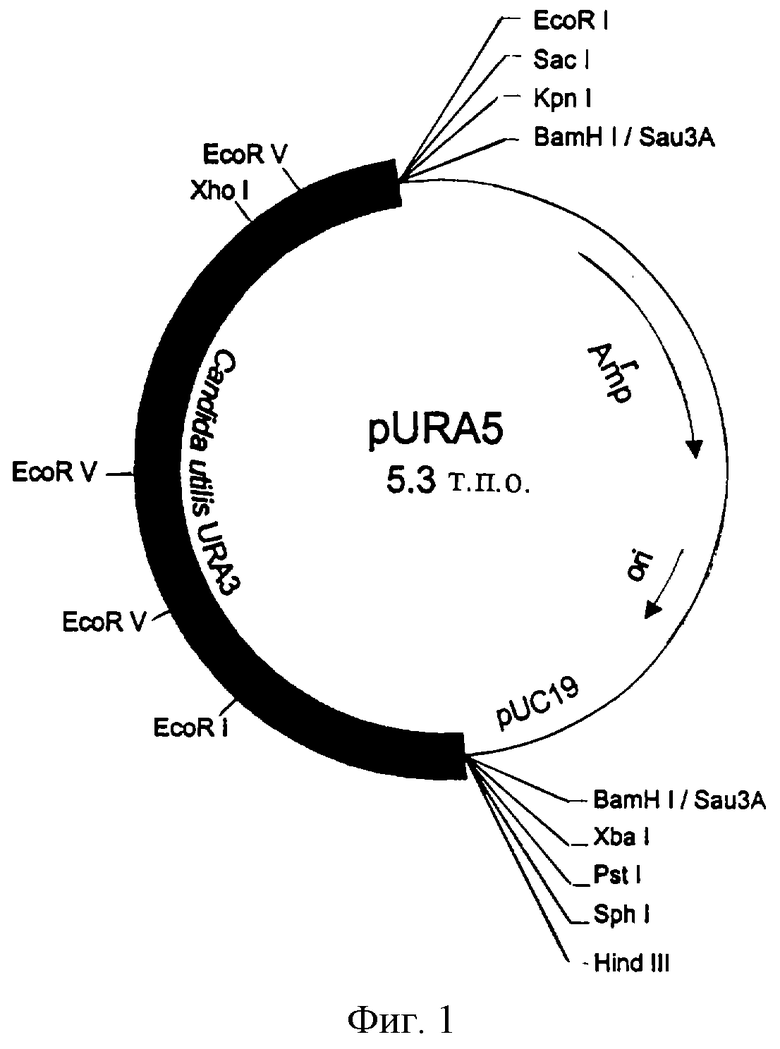
Ген URA3 выделяют из Candida utilis и анализируют с целью его использования в качестве маркера для трансформации клетки-хозяина ura3 Candida utilis. Фрагменты ДНК, содержащие ген URA3 вида Candida utilis, выделяют из геномной библиотеки pUC19 Candida utilis на основании его способности комплементировать мутацию pyrF Escherichia coli, принимая во внимание, что ген URA3, выделенный из Saccharomyces cerevisiae, комплементирует мутацию pyrF рода Е.coli, используя случайную промоторную активность в Escherichia coli. При переносе этой библиотеки в среду без урацила выделяют 12 независимых колоний pyrF+. Два из этих клонов (pURA-2 и pURA-5) имеют одинаковую геномную вставку ДНК длиной 2,6 т.п.о. вида Candida utilis на векторе pUC19, используя рестрикционное расщепление HindIII и EcoRI. ДНК, выделенная из обеих плазмид, трансформирует штамм МС1066 Escherichia coli в Ura+ с высокой частотой. Карта одной рекомбинантной плазмиды (pURA-5) с геном - рUС19 URA3 Candida utilis показано на фиг.1. Эту плазмиду используют для дальнейшей комплементации и секвенирования.
Пример 6
Разграничение и секвенирование гена URA3 Candida utilis
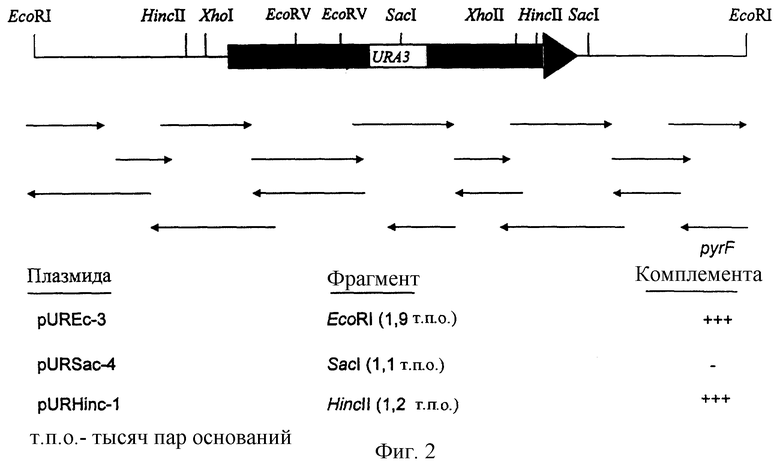
Плазмиду pURA5 расщепляют несколькими рестрикционными ферментами. Фрагменты, соответствующие расщеплению EcoRI (1,9 т.п.о.), HincII (1,3 т.п.о.), SacI (1,1 т.п.о.), субклонируют в векторе pBluescript SK (+), что дает соответственно плазмиды pUREc-3, pURHinc-1, pURSac-4. Фрагмент, соответствующий плазмиде pURSac-4, не способен комплементировать мутацию pyrF Escherichia coli (фиг.2).
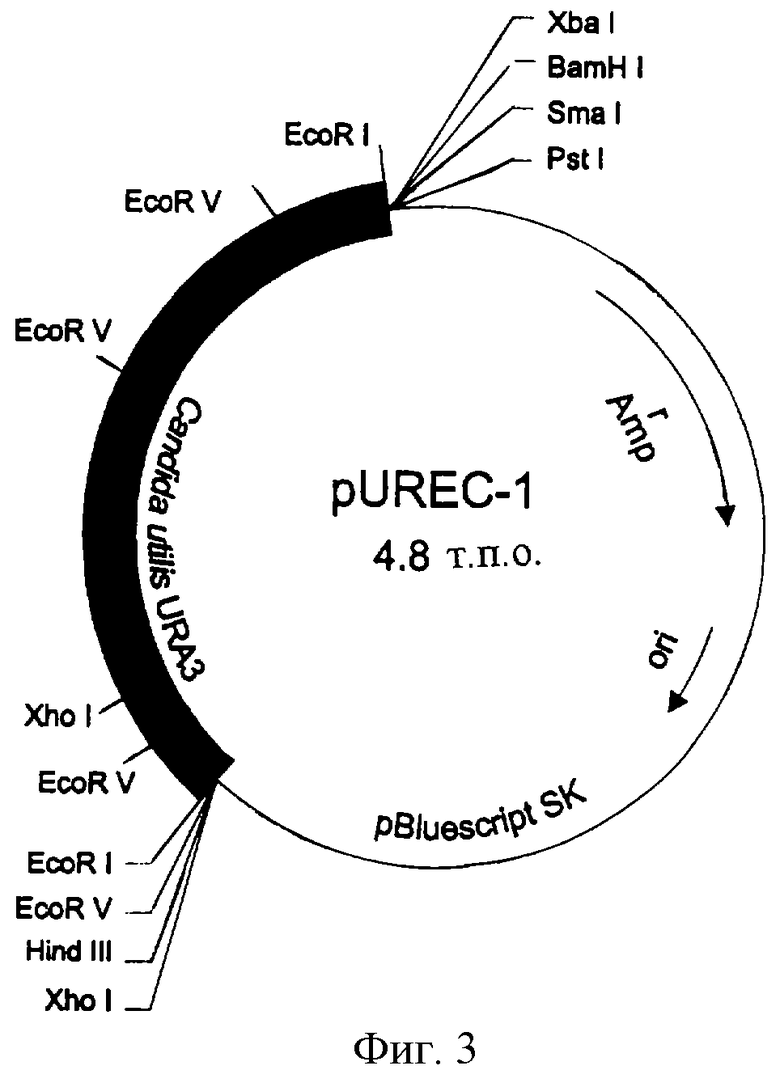
У фрагмента EcoRI длиной 1,9 т.п.о. (pUREc-3, фиг.3), содержащего ген URA3 Candida utilis, полностью секвенируют двухцепочечную молекулу по методу Сангера и др. (Sanger et al., 1977, Proc. Natl. Acad. Sci USA 74:5463-5467).
На этой стадии используют универсальные олигонуклеотиды серии M13mp/pUC и внутренние олигонуклеотиды, выделенные из этой последовательности. Полная последовательность длиной 1179 пар оснований фрагмента EcoRI показана на фиг.4 (SEQ ID №1, 2). Этот фрагмент содержит открытую рамку считывания длиной 800 пар основаной (266 кодонов). Ген URA3 Candida utilis кодирует белок с теоретической молекулярной массой 29436 Да. Нуклеотидная последовательность, фланкирующая к инициирующему кодону ATG (GAAAATG), соответствует согласованной последовательности, обнаруженной в дрожжах (A/YAA/YAATG) Чиганом и Донехью (Cigan and Donehue, 1987 (Gene, 59:1-18)).
3’-Нетранслированная область содержит мнимый сайт полиаденилирования (ТАТАААА, согласованная последовательность ААТАААА), находящийся в 3’-концевой области большинства эукариотных генов (Guo, Z. and Sherman, F., 1995, Mol. Cell. Biol. 15: 5983-5990).
Пример 7
Анализ комплементации Saccharomyces cerevisiae
С целью проверки соответствия клонированного фрагмента, соответствующего гену URA3 Candida utilis и не соответствующего фрагменту ДНК с супрессорной активностью, фрагмент KpnI/XbaI длиной 2,8 т.п.о. плазмиды pURA5 клонируют в производном векторе pBR322 (pBSARTR-3). Вектор pBSARTR-3 имеет автономную реплицирующую последовательность (ARS1) и ген TRP1 в качестве селективного маркера, полученные из Saccharomyces cerevisiae. Затем получают плазмиду pUT64 (фиг.5) и используют ее для трансформации штамма SEY2202 (ura3-52-, leu2-112, his3) Saccharomyces cerevisiae по методу на основе ацетата лития, описанному Ито и др. (Ito et al., 1983, J. Bacteriol, 153: 163-168).
Трансформанты получают через 48 часов после трансформации. Наличие репликативной плазмиды проверяют методами гибридизации колоний и саузерн-блоттинга.
Полученная частота трансформации (2-5×102 трансформаций/мг) соответствует значению, приведенному в научной литературе для ауксотрофных маркеров, выделенных из других дрожжей.
Из вышесказанного следует, что ген URA3, выделенный из Candida utilis, способен комплементировать мутацию ura3 Saccharomyces cerevisiae.
Пример 8
Трансформация CUT35 штамма Candida utilis с помощью плазмид pURA5 и pUCURA3 по методу на основе LiAc
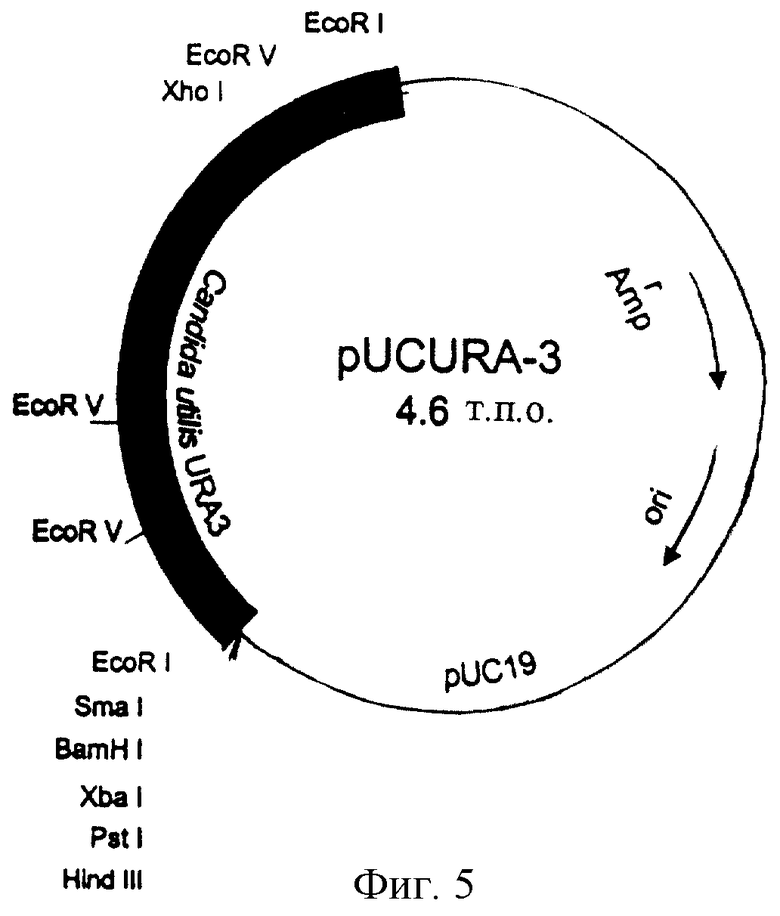
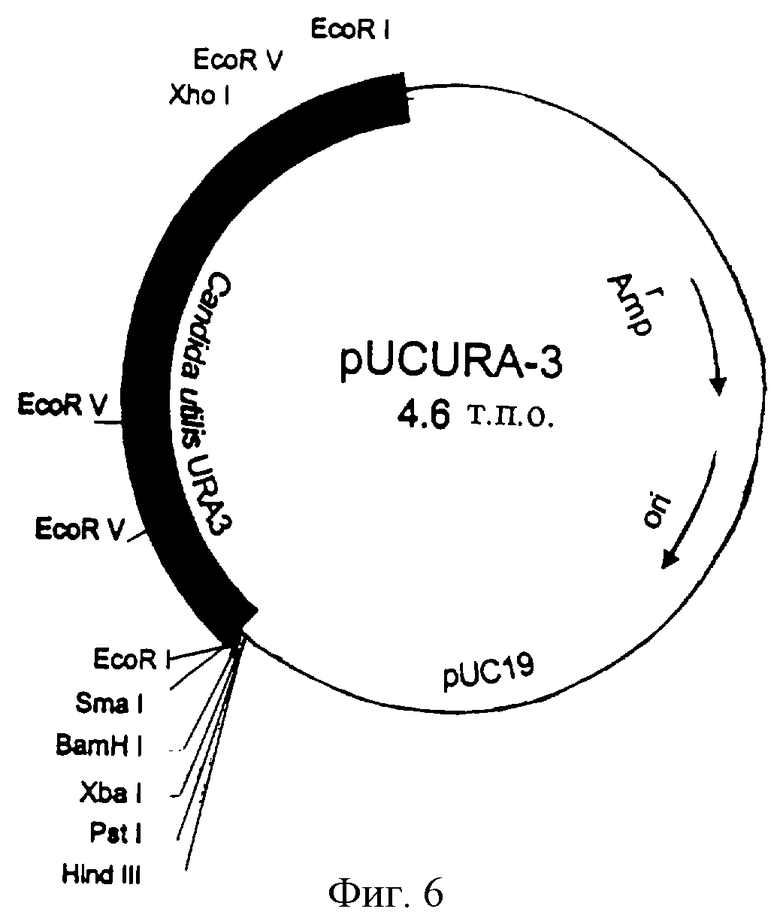
Мутантный штамм ura3 CUT35 Candida utilis, фенотип urа-, который включен под номером доступа CBS 100085 в коллекцию типовых культур Centralbureau voor Schimmelcultures 1 октября 1997 г., трансформируют по способу на основе ацетата лития, описанному Ито и др. (Ito et аl., 1983), с использованием гена URA3, предварительно выделенного из Candida utilis в качестве селективного маркера. Векторы (pURA5 и pUCURA3), используемые в этой системе трансформации, сконструированы с возможностью прямой интеграции в гомологичный локус мутантного штамма Candida utilis, в результате гомологичной рекомбинации. Плазмиду pUCURA3 получают путем клонирования фрагмента EcoRI длиной 1,8 т.п.о. гена URA3 Candida utilis в соответствующем сайте вектора pUC19 (фиг.6). До трансформации обе плазмиды расщепляют XhoI, который находится в 5’-концевой затравке структурного гена. Перевод плазмид в линейную форму способствует гомологичной интеграции в геномный локус.
Трансформацию выполняют аналогично процедуре на основе ацетата лития, описанной Ито и др (Ito et ai., 1983), за исключением того, что в данном случае концентрация LiAc составляет 50 мМ. В соответствии с этим методом колонию культуры выращивают на планшете с YPD-средой, выбранный штамм культивируют, встряхивая, в 5 мл жидкой YPD-среды при температуре 30°С в течение примерно 8 часов при концентрации, соответствующей оптической плотности OD600=0,003, после чего культуру выращивают, встряхивая, при температуре 30°С в течение примерно 16 часов. Когда клетки вырастают до логарифмической фазы (ОD600=0,6), их собирают центрифугированием со скоростью 3000 об/мин в течение 5 мин. Клетки промывают один раз 3 мл дистиллированной воды. Затем клетки суспендируют в 1 мл 50 мМ LiAc. После этого клетки инкубируют, перемешивая, при температуре 30°С в течение 1 часа. Аликвоту (100 мкл) клеток вводят в микроцентрифужную пробирку и добавляют 5 мкг ДНК. Смесь клетки ДНК инкубируют при температуре 30°С в течение 30 мин. Далее клетки ДНК обрабатывают 0,7 мл 40% PEG 4000 и 50 мМ LiAC и инкубируют при температуре 30°С в течение 1 часа. Затем полученную смесь в течение 5 мин подвергают температурному шоку в водяной бане при температуре 42°С. Сразу же после этого смесь клетки ДНК центрифугируют в течение 30 с и дважды промывают в 10 мМ трис-буфера, 1 мМ этилендиаминтетрауксусной кислоты (EDTA), рН 8. И наконец, удаляют супернатант и клетки суспендируют в 200 мкл 10 мМ трис-буфера, рН 8, 1 мМ EDTA, рН 8, до достижения конечного объема.
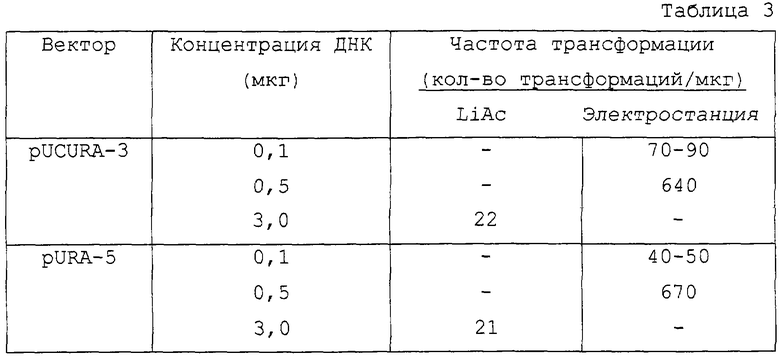
Трансформанты отбирают в минимальной YNB-среде, не содержащей урацила. Механизм гомологичной интеграции обеспечивает высокую митотическую устойчивость этих трансформантов. Частота трансформации соответствует значению, приведенному для Saccharomyces cerevisiae и других нестандартных дрожжей при использовании интеграционных векторов (табл.2).
Пример 9
Трансформация мутанта CUT35 Candida utilis с помощью плазмид pURA5 и pUCURA3 по методу электропорации
Мутантный штамм ura3 CUT35 Candida utilis трансформируют по методу электропорации, описанному Кондо и др. (Kondo, К. et al., 1995, J. Bacteriol. 177:7171-7177), используя ген URA3, предварительно выделенный из Candida utilis в качестве селективного маркера. Векторы (pURA5 и pUCURA3), используемые в этой системе трансформации, сконструированы с возможностью прямой интеграции в гомологичный локус мутантного штамма Candida utilis в результате гомологичной рекомбинации.
В основе используемой процедуры лежит обработка интактных дрожжевых клеток электрическим полем. Обработку производят в следующих условиях: подаваемый импульс 0,7 кВ (3,5 кВ/см), сопротивление 800 Ом и электрическая емкость 25 мкФ.
До трансформации обе плазмиды расщепляют XhoI, который находится в 5’-концевой затравке структурного гена, что облегчает гомологичную интеграцию в геномный локус.
Трансформанты отбирают в минимальной YNB-среде без урацила.
Частота трансформации при использовании pURA5 и pUCURAS зависит от концентрации плазмиды. Результаты сравнения обоих методов (LiAc и электропорация) приведены в табл,3.
Данный механизм интеграции обеспечивает высокую митотическую устойчивость этих трансформантов.
Частоты трансформации соответствуют значениям, приведенным для Saccharomyces cerevisiae и других нестандартных дрожжей при использовании интеграционных векторов.
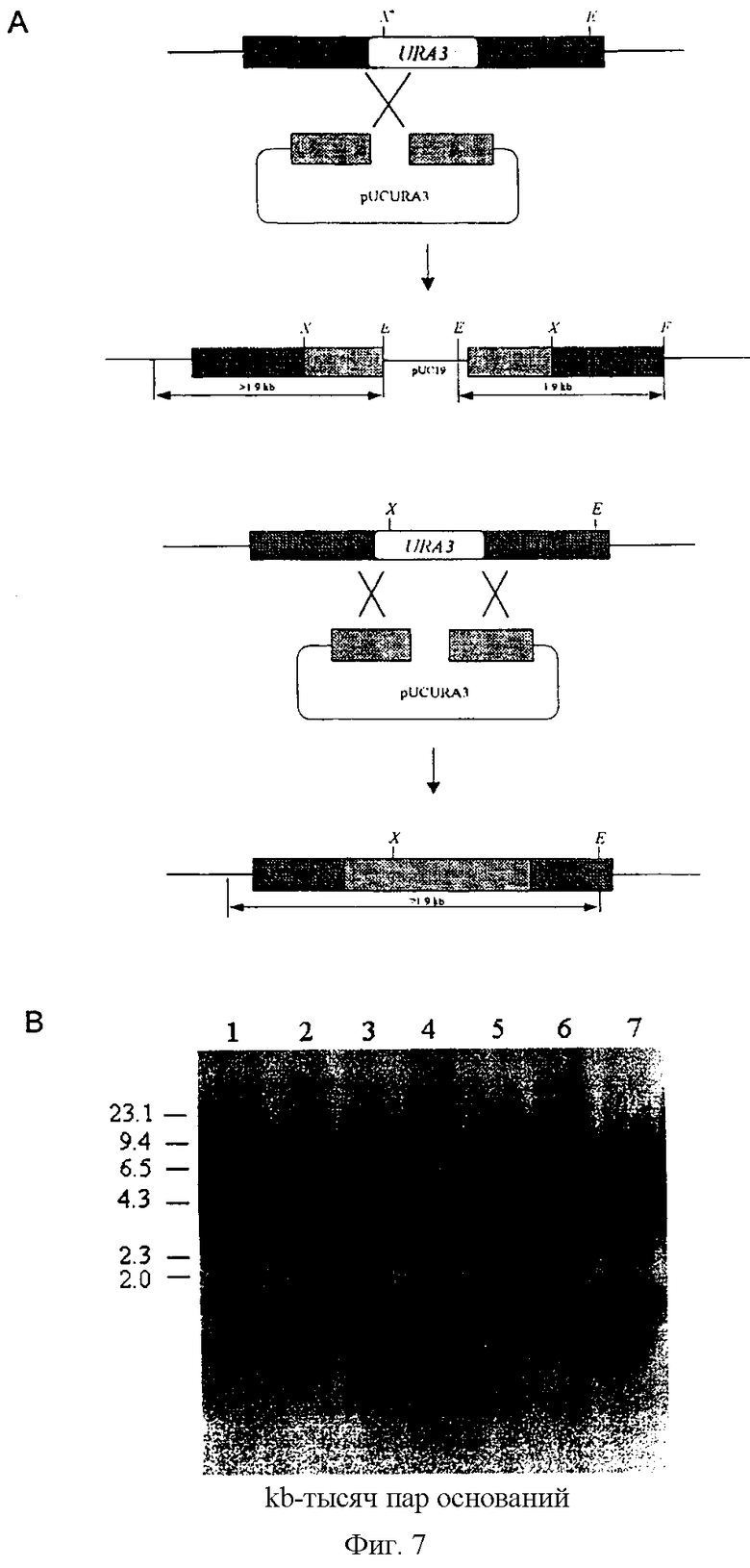
На фиг,7 показаны все возможные варианты интеграции в геном Candida utilis, а также результаты саузерн-блоттинга некоторых трансформантов.
Пример 10
Выделение гена HIS3 Candida utilis
Ген HIS3 выделяют из Candida utilis и исследуют с использованием библиотеки, описанной в примере 4. Фрагменты ДНК, содержащие ген HIS3 Candida utilis, выделяют из геномной библиотеки Candida utilis с учетом его способности комплементировать мутацию hisb463 в штамме КС8 Escherichia coli (hsd, hisB463, leuB6, pyrF::Tn5 Kmr, trp (9830 (lact YA), stm, galU, gal), принимая во внимание, что ген HIS3, выделенный из Saccharomyces cerevisiae, комплементирует мутацию hisb463 Escherichia coli, используя случайный промотор в Escherichia coli.
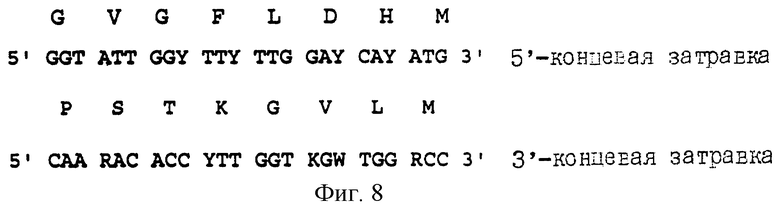
Чтобы выделить ген HIS3, клетки в количестве 105 культивируют на минимальной среде (М9), дополненной урацилом, триптофаном и лейцином. Плазмидную ДНК экстрагируют из колоний, способных расти в этой среде и комплементировать мутацию hisb463 в мутантном штамме КС8 Escherichia coli. Выделенные плазмидные ДНК используют для повторной трансформации мутантного штамма КС8 Escherichia coli. Все плазмиды, способные удовлетворять потребностям мутантного штамма в гистидине, получили название pHCU. Чтобы подтвердить, что колонии his+ содержат ген HIS3 Candida utilis, а не фрагмент ADN с супрессорной активностью, две плазмиды, полученные из трансформантов his+ (pHCU37 и pHCU40), подвергают полимеразной реакции синтеза цепи. Для этой цели используют два вырожденных олигонуклеотида, полученных из двух высококонсервативных областей в пяти последовательностях IGPDasas дрожжей и грибов. Олигонуклеотид и аминокислотные последовательности вырожденных олигонуклеотидов показаны на фиг.8.
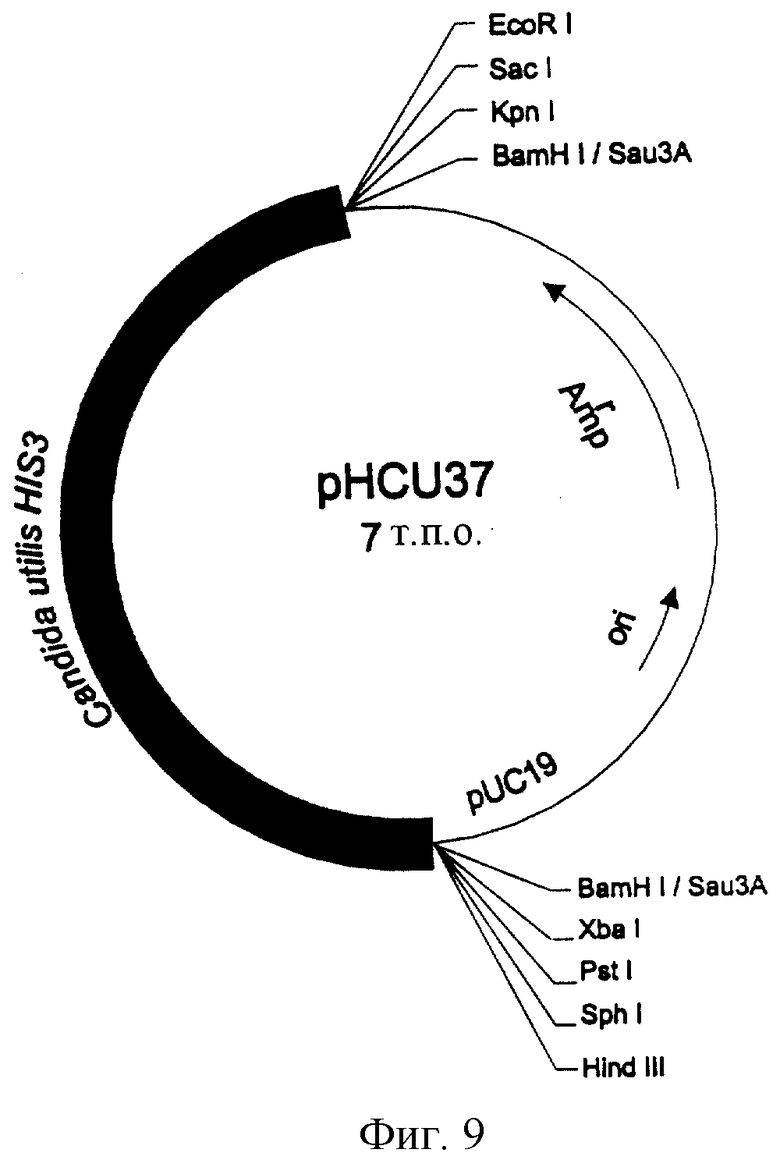
В результате полимеразной реакции синтеза цепи амплифицирована полоса длиной 500 пар оснований, соответствующая кодирующей последовательности гена HIS3 из Candida utilis. Фрагмент длиной примерно 500 пар оснований, полученный с помощью полимеразной реакции синтеза цепи, который, как показали результаты саузерн-блоттинга, способен гибридизировать с геномной ДНК Candida utilis, клонируют в Т-векторе (pMOSBLUE, Amershan) и прогнозируемая аминокислотная трансляцией ее последовательности в значительной степени идентична аналогичному показателю для гена His3p, выделенного из других дрожжей и грибов. Плазмиду pHCU37 (фиг.9) используют для определения всей последовательности гена HIS3 из Candida utilis.
Пример 11
Секвенирование гена HIS3 Candida utilis
У гена HIS3, выделенного из Candida utilis, полностью секвенируют двухцепочечную молекулу по методу Сангера и др. (Sanger et al., 1977). С этой целью используют олигонуклеотиды универсальной серии M13mp/pUC. Для инициации секвенирования всего гена используют затравки, выделенные из фрагмента, полученного в результате полимеразной реакции синтеза цепи. Секвенировано в общей сложности 1190 пар оснований плазмиды pHCU37. Вся последовательность HIS3, выделенная из Candida utilis, показана на фиг.10 (SEQ ID №5, 6).
Этот фрагмент содержит открытую рамку считывания, состоящую из 210 кодонов. Ген HIS3 Candida utilis кодирует белок с теоретической молекулярной массой, равной 24518 Да.
Пример 12
Выделение гена INV1, кодирующего инвертазу Candida utilis
Для того чтобы выделить ген INV1, кодирующий фермент инвертаза в Candida utilis, было использовано такое преимущественное свойство, что аминокислотная последовательность этого фермента имеет области, с высокой сходностью характерные для разных видов, таким образом, сопоставляют последовательности β-фруктофуранозидазы из разных дрожжей. Два вырожденных олигонуклеотида, используемых при осуществлении указанной полимеразной реакции синтеза цепи, сконструированы в соответствии с использованием кодонов в Candida utilis. Полипептидные последовательности и вырожденные олигонуклеотиды показаны на фиг.11.
В результате выполнения полимеразной реакции синтеза цепи получают полосу длиной 417 пар оснований, которую субклонируют в Т-векторе (pMOBlue, Amersham). Данную полосу полностью секвенируют, и трансляция указанного фрагмента ДНК подтверждает наличие согласованных областей и большой гомологии между ферментами инвертаза, приведенными в научной литературе. Полученные результаты показывают, что выделенный фрагмент относится к гену INV1, который кодирует этот фермент в Candida utilis. Этот фрагмент используют в качестве зонда для выделения гена INV1 из Candida utilis.
Обследование библиотеки Candida utilis позволяет выделить в общей сложности 6 клонов, имеющих ген INV1. Два из этих клонов выбирают с учетом их размера для секвенирования (рСI-6 и pCI-12) по методу полимеразной реакции синтеза цепи, используя олигонуклеотиды, представленные на предыдущей стадии. Эти олигонуклеотиды инициируют полное секвенирование гена из обеих цепей, относящихся к плазмиде рСI-6.
Пример 13
Секвенирование гена INV1 Candida utilis
В общей сложности 2607 пар оснований клона pCI-6, содержащего ген INV1, кодирующий инвертазу Candida utilis, полностью секвенируют по методу Сангера и др. (1977) и с этой целью используют универсальные олигонуклеотиды, относящиеся к серии M13mp/pUC, и внутренние олигонуклеотиды, выделенные из этой последовательности. Результаты полного секвенирования фрагмента длиной 2607 пар оснований показаны на фиг.12 (SEQ ID №5, 6). Указанный фрагмент имеет открытую рамку считывания длиной 1602 пар оснований (534 кодона). Ген INV1 Candida utilis кодирует белок с теоретической молекулярной массой, равной 60703 Да.
Принимая во внимание, что инвертаза в Candida utilis является периплазматическим ферментом, он должен иметь сигнальный пептид в N-конце. В результате анализа последовательности до 5’-конца этого гена обнаружены два кодона ATG (показаны на фиг.12 как ATG1 и ATG2), где находится открытая рамка считывания для белков, которые отличаются только размером N-концов. С помощью алгоритма фон Хейне (Heijne, 1986, Nucl. Acids Res. 14:4683-4690), используемого для прогнозирования сайта рестрикции сигнальной пептидазы зрелого белка, выделенного из обоих кодонов ATG, установлено, что сайты рестрикции в обоих случаях расположены между остатками S39 и S40 для ATG1 и между остатками S26 и S27 для ATG2. Это свидетельствует о наличии сигнальных пептидов, состоящих соответственно из 39 и 26 аминокислот. С учетом среднего размера сигнальных последовательностей в дрожжах (примерно 20 остатков) можно предположить, что инициирующим кодоном гена INV1 является второй кодон ATG.
В аспарагинах, занимающих положения 40, 88, 141, 187, 245, 277, 344, 348, 365, 373, 379 и 399 в последовательности зрелого белка, в соответствии с общим правилом N-X-T/S обнаружено одиннадцать вероятных сайтов N-гликозилирования.
В 5’-концевой нетранслированной области обнаружено два предполагаемых ТАТА-блока (согласованная последовательность ТАТАА), которые соответствуют областям от -18 до -14 и от -212 до -208, а также разные вероятные объединенные сайты репрессора Migl (согласованная последовательность SYGGRG).
Краткое описание фигур
Фиг.1. Плазмида pURA5, полученная в геномной библиотеке Candida utilis путем комплементации мутации pyrF штамма МС1066 Escherichia coli и мутации ura3 штамма SEY 2202 Saccharomyces cerevisiae.
Фиг.2. Карта рестрикционных ферментов, стратегия секвенирования и анализ комплементации гена URA3, выделенного из Candida utilis.
Фиг.3. Плазмида pUREC3, полученная путем клонирования фрагмента EcoRI длиной 1,9 т.п.о. плазмиды pURA5 в pBLUESCRIPT SK(+).
Фиг.4. Аминокислотная последовательность, выделенная из последовательности ДНК гена URA3, и последовательность ДНК, кодирующая этот ген.
Фиг.5. Плазмида pUT64, полученная для эксперимента по комплементации в мутантном штамме ura3 Saccharomyces cerevisiae.
Фиг.6. Плазмида pUCURA3, используемая в экспериментах по трансформации Candida utilis.
Фиг.7.
(A) Предполагаемое расположение векторной ДНК, интегрированной в локус URA3 путем гомологичной рекомбинации.
(B) Гибридизация по методу ДНК-блоттирования геномной ДНК, выделенной из некоторых трансформантов.
Фиг.8. Последовательность ДНК затравок, используемых для выделения гена HIS3, и выделенная аминокислотная последовательность, кодирующая этот ген.
Фиг.9. Плазмида pHCU37, полученная в геномной библиотеке Candida utilis путем комплементации мутации hisb463 штамма КС8 Escherichia coli.
Фиг.10. Аминокислотная последовательность, выделенная из последовательности ДНК гена HIS3, и последовательность ДНК, кодирующая этот ген.
Фиг.11. Аминокислотная последовательность и соответствующая последовательность ДНК олигонуклеотидов, используемых при осуществлении полимеразной реакции синтеза цепи с целью выделения гена INV1 Candida utilis.
Фиг.12. Последовательность ДНК, соответствующая фрагменту, содержащему ген INVl Candida utilis.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ФИБРОБЛАСТНОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА КЛЕТКАМИ ДРОЖЖЕЙ, СПОСОБ ЕЕ КОНСТРУИРОВАНИЯ И ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE-ПРОДУЦЕНТ ФИБРОБЛАСТНОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА | 1998 |
|
RU2180003C2 |
| ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE 1-60-Д578 (MSIL), - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ИНТЕРЛЕЙКИНА-2 ЧЕЛОВЕКА И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2002 |
|
RU2230781C1 |
| СПОСОБ КОНСТРУИРОВАНИЯ ШТАММОВ ДРОЖЖЕЙ Saccharomyces cerevisiae - СТАБИЛЬНЫХ ПРОДУЦЕНТОВ СОМАТОТРОПИНА ЧЕЛОВЕКА, ШТАММ ДРОЖЖЕЙ Saccharomyces cerevisiae - СТАБИЛЬНЫЙ ПРОДУЦЕНТ СОМАТОТРОПИНА ЧЕЛОВЕКА (ВАРИАНТЫ) | 2009 |
|
RU2420567C1 |
| ШТАММ ДРОЖЖЕЙ Yarrowia lipolytica - ПРОДУЦЕНТ ЛИПАЗЫ | 2007 |
|
RU2355754C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pPBS-St9, КОДИРУЮЩАЯ ПОЛИПЕПТИД СОМАТОТРОПИНА, И ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE ДЛЯ ПРОДУКЦИИ РЕКОМБИНАНТНОГО СОМАТОТРОПИНА | 2011 |
|
RU2465315C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ YARROWIA-LIPOLYTICA - ПРОДУЦЕНТ ЛИПАЗЫ | 2011 |
|
RU2451075C1 |
| Рекомбинантная плазмидная ДНК pJDB (MSIL), обеспечивающая синтез интерлейкина-2 человека в клетках дрожжей SасснаRомUсеS ceReUISIaL, способ ее получения и штамм дрожжей SасснаRомYсеS ceReUISIaL - продуцент интерлейкина-2 человека | 1988 |
|
SU1770359A1 |
| ПРИМЕНЕНИЕ ШТАММА ДРОЖЖЕЙ Komagataella pastoris В КАЧЕСТВЕ РЕЦИПИЕНТА ДЛЯ КОНСТРУИРОВАНИЯ ПРОДУЦЕНТОВ ЦЕЛЕВОГО БЕЛКА | 2013 |
|
RU2522479C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА PFS 19, ОПРЕДЕЛЯЮЩАЯ СИНТЕЗ ПОВЕРХНОСТНОГО АНТИГЕНА S ВИРУСА ГЕПАТИТА B ЧЕЛОВЕКА | 1992 |
|
RU2115730C1 |
| СПОСОБ ПОЛУЧЕНИЯ КСИЛИТА | 1993 |
|
RU2142999C1 |















Изобретение относится к способу трансформации, предназначенному для экспрессии гетерологичных белков в дрожжах Candida utilis путем получения ауксотрофных мутантов указанных видов и выделение из геномной библиотеки разных генов, комплементирующих указанные ауксотрофные мутанты. В качестве клеток-хозяев в указанной системе трансформации использованы новые ауксотрофные штаммы, полученные из дрожжей штамма NRRL Y-1084 Candida utilis (с фенотипом ura и с фенотипом his, обозначенным как TMN3), в которых отсутствуют главным образом биосинтетические пути метаболизма урацила и гистидина и которые могут быть трансформированы плазмидами, содержащими в качестве селективных маркеров гены URA3 и HIS3 Candida utilis. Другими объектами настоящего изобретения является рекомбинантный ДНК материал для трансформации указанных штаммов и кодирующая последовательность ДНК гена HIS3 указанных штаммов дрожжей; изобретение позволяет получать гетерологичные белки в дрожжах, которые в дальнейшем можно использовать в различных целях. 5 н. и 15 з.п.ф-лы, 11 ил., 3 табл.
| OHI R | |||
| et | |||
| al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Gene | |||
| Предохранительное устройство для паровых котлов, работающих на нефти | 1922 |
|
SU1996A1 |
| Композиция на основе высокомолекулярного силоксанового каучука | 1976 |
|
SU717107A1 |

