Область техники
Изобретение относится к биотехнологии, генной инженерии, микробиологической, медицинской и фармацевтической промышленности и представляет собой сконструированную in vitro рекомбинантную плазмидную ДНК, которая обеспечивает синтез соматотропина человека в клетках дрожжей Saccharomyces cerevisiae, способ конструирования этой плазмидной ДНК и дрожжевой штамм-продуцент соматотропина человека, содержащий эту плазмиду.
Уровень техники
Соматотропин (синонимы: соматотропин, гормон роста человека, соматотропный гормон, соматотропный гормон роста) относится к наиболее изученным гормонам гипофиза и принадлежит к белковому семейству, включающему гормон роста, пролактин, ХС. Соматотропин синтезируется в передней доле гипофиза и у всех видов млекопитающих представляет собой одиночный пептид с молекулярной массой около 22000 Да. Несмотря на высокую степень гомологии последовательностей гормонов роста различных млекопитающих, в клетках человека активен только собственный гормон роста человека или гормон роста высших приматов. Молекула соматотропина состоит из 191 аминокислотного остатка и содержит два внутримолекулярных дисульфидных мостика [Roskam W.G., Rougeon F. // Molecular cloning and nucleotide sequence of the human growth hormone structural gene. Nucl.Acids Res., 1979, v.7, p.305-320.; Martial J.A., Hallewell R.A., Baxter J.D., Goodman H.M. // Human growth hormone: complementary DNA cloning and expression in bacteria. Science, 1979, v.205, No4406, p.602-607].
Соматотропин оказывает анаболическое действие, положительно влияет на минеральный обмен, вызывает увеличение роста и массы тела при карликовости, связанной с недостаточностью гормона роста. Соматотропин вырабатывается и секретируется в кровь специализированными клетками главным образом передней доли гипофиза - соматотрофами. Содержание соматотропина в гипофизе человека более чем на порядок превышает содержание других гормонов этой эндокринной железы. Соматотропин - полифункциональный гормон. Являясь специфическим стимулятором роста тела (скелета и мягких тканей), он участвует также в регуляции всех видов обмена веществ. Основной дефект развития организма человека и животных в условиях недостаточности соматотропина - задержка роста костей. Избыток соматотропина в растущем организме может приводить к гигантизму, а у взрослых - к аномальному увеличению отдельных органов и тканей. Действие соматотропина на рост костей опосредовано через соматомедины - инсулиноподобные ростовые факторы полипептидной природы. Биосинтез и секреция соматотропина находятся под сложным контролем, включающим регуляцию гормонами гипоталамуса - соматостатином и соматолиберином, а также некоторыми другими гормонами и продуктами обмена веществ.
Область применения соматотропина достаточно широка: нарушение процесса роста у детей при недостаточности эндогенного гормона роста, хроническая почечная недостаточность у детей, сопровождающаяся задержкой роста, синдром Шерешевского-Тернера, остеопороз, синдромы иммунодефицита, сопровождающиеся потерей массы тела.
Лекарственные формы соматотропина (Генотропин (PFIZER MFG. BELGIUM, N.V., производитель Vetter Pharma-Fertigung, GmbH & Со. KG; Хуматроп (LILLY FRANCE, (Франция), Сайзен MERCK SERONO, S.p.A. (Италия), Нордитропин (NOVO NORDISK, A/S (Дания), Биосома, производитель BIOFA, российский препарат Растан (ФАРМСТАНДАРТ-УфаВИТА, Россия), известны также нутропин или протропин (Genentech), Jintropin (GenSci, Китай), Dynatrope (Dynamic Development Laboratories Co., Ltd), Ansomone (Ankebio, Китай), Getropin (Getropin, Китай), Hygetropin (Китай), Somatohorm (Biomed, Польша) и другие.
Известен способ получения соматотропина, состоящий в его выделении из тканей гипофиза человека [Simionescu L., Dimitriu V., Zamfir-Grigorescu D., Aman E., Terbancea M. // The simultaneous isolation of human pituitary hormones. I. Human growth hormone. Endocrinologie 1982 Oct.-Dec.; 20 (4): 273-83]. Недостатками способа являются сложность с получением исходного материала и риск заражения инфекцией [Rappaport E.B. // latrogenic Creutzfeldt-Jakob disease. Neurology, 1987, Sep.; 37 (9): 1520-2].
Известен способ, основанный на получении рекомбинантного соматотропина в трансформированных клетках мыши линии С 127 [Carter M.J., Facklam T.J., Long P.C., Scotland R.A. // Are continuous cell lines safe as substrates for human drugs and biologies? A case study with human growth hormone. Dev Biol Stand 1989; 70: 101-7]. При таком подходе удается получить гормон роста человека в правильной конформации с хорошей физиологической активностью. Недостатками этого способа являются чрезвычайно низкий выход и длительное время культивации продуцирующих клеток.
Перспективным подходом для получения соматотропина человека в значительных количествах является использование микроорганизмов в качестве продуцентов этого препарата [Seeburg, et al. // Synthesis of growth hormone by bacteria. Nature, 1978, 276, 795-798; Seeburg et al. // Nucleotide sequence and amplification in bacteria of structural gene for rat growth hormone, 1978 Nature, 270, 486-494; Martial et al. // Human growth hormone: complementary DNA cloning and expression in bacteria, Science, 1979, 205, 602-607; Seeburg et al. // Efficient Bacterial Expression of Bovine and Porcine Growth Hormones, 1983, DNA 2: 37-45].
В настоящее время из уровня техники известен штамм бактерии Escherichia coli - продуцент свиного соматотропина (Патенты РФ 2072393). Известен способ получения соматотропина, включающий экспрессию в клетках Escherichia coli, заключающийся в достаточно быстром биосинтезе клетками бактерий рекомбинантного соматотропина в виде нерастворимых "телец включения" [Olson K.C., Fenno J., Lin N., Harkins R.N., Snider C., Kohr W.H., Ross M.J., Fodge D., Prender G., Stebbing N. // Purified human growth hormone from E.coli is biologically active. Nature 1981 Oct. 1; 293 (5831): 408-11].
Из патента США №5795745 известен продуцент соматотропина на основе плазмиды pHGH 107, несущей под контролем двух последовательных Lac-промоторов ген соматотропина, состоящий из синтетической части (соответствующей аминокислотам 1-24) и фрагмента природного гена (соответствующего аминокислотам 25-191). Использование Lac-промоторов, а также почти полностью (на 88%) природной кДНК, включающей редко встречающиеся в генах Escherichia coli (минорные) кодоны, приводит к существенному снижению уровня биосинтеза белка относительно теоретически возможного.
В российском патенте 2233879 разработана рекомбинантная плазмидная ДНК pES1-6, обеспечивающая высокий выход рекомбинантного соматотропина из бактерий E.coli. Недостатком может являться использование бактерии E.coli в качестве продуцента, так как это организм является условным патогеном.
Использование эукариотических систем экспрессии, в частности дрожжевых, позволяет осуществлять корректную посттрансляционную модификацию рекомбинантных белков, препараты, полученные из дрожжей, не содержат бактериальных пирогенов и эндотоксинов, что позволяет снизить спектр побочных эффектов и значительно улучшить качество жизни пациентов в период длительного лечения. Наиболее близким по технической сущности является патент США 4443539, в котором запатентован процесс выделения бычьего соматотропина из дрожжей Saccharomyces cerevisiae.
Для производства рекомбинантного соматотропина человека мы предлагаем использовать штамм-продуцент дрожжей Saccharomyces cerevisiae, несущий плазмидную ДНК с геном соматотропина.
Осуществление изобретения
Изобретение решает задачу конструирования плазмиды, детерминирующей синтез рекомбинантного белка, и создания высокопродуктивного бактериального штамма-продуцента, позволяющего получать рекомбинантный соматотропин с высоким выходом и по упрощенной технологии.
Стабильное поддержание экстрахромосомных генетических элементов (плазмид) в дрожжевой клетке зависит от наличия в таких элементах последовательностей, обеспечивающих репликацию ДНК синхронно с репликацией дрожжевых хромосом и равномерное распределение плазмид по дочерним клеткам, а также возможности селекции клеток, содержащих данную плазмиду.
Поставленная задача решается за счет конструирования рекомбинантной плазмидной ДНК pPBS-St9, кодирующей полипептид с последовательностью соматотропина, имеющей молекулярную массу около 4,1 МДа (6266 п.о.). Плазмида сконструирована на основе плазмиды pRS316 [Sikorski and Hieter P. // A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae, 1989, Genetics 122 (1): 19-27] стандартного центромерного челночного вектора, включающего бактериальные маркер ампициллин-резистентности и ориджин репликации, полилинкер в составе участка гена LacZ, ген URA3 S. cerevisiae, последовательности ARS и CEN Saccharomyces cerevisiae. Исходная плазмида pRS316 коммерчески доступна, например, в АТСС Number 77145 и может быть заказана по ссылке http://www.atcc.org/ATCCAdvancedCatalogSearch/ProductDetails/tabid/452/Default.aspx?ATC CNum=77145&Template=vectors. Рекомбинантная плазмидная ДНК pPBS-St9 включает в себя промотор гена PHO5 (вставлен по сайтам SacI/SacII положение 1998-2533, размер вставки 536 п.о.), фрагменты ДНК, кодирующие 24 N-терминальных аминокислоты интерлейкина человека hIL-1beta (Ser5-Ala28), вставленные по сайтам SacII/Xba1 (положение 2540-2611, размер вставки 72 п.о.), сайт для расщепления эндопептидазой Кех2 (Lys-Arg), 191 а.к. соматотропина, вставленные по сайтам Xba/BamHI (положение 2617-3192, размер вставки 576 п.о.), после которых находится сигнал терминации транскрипции гена CYC1 дрожжей, вставленный по сайтам HindIII/BamHI (положение 3198-3460, размер вставки 262 п.о.).
Для регуляции экспрессии гена соматотропина человека мы использовали промотор гена PHO5, содержащий области, обеспечивающие активацию транскрипции при отсутствии неорганического фосфата в культуральной среде, а также область инициации транскрипции. Промотор гена PHO5 относится к числу наиболее сильных дрожжевых промоторов. Уровень экспрессии генов, находящихся под контролем промотора PHO5, эффективно регулируется экзогенным неорганическим фосфатом. При концентрации неорганического фосфата в культуральной среде 1 г/л наблюдается только базальный уровень экспрессии. Это позволяет регулировать синтез соматотропина в клетках дрожжей. Таким образом, предлагаемая система позволяет нарабатывать большое количество дрожжевой биомассы в оптимальных условиях (на среде, содержащей неорганический фосфат) а затем, после смены условий культивирования (замены среды, содержащей неорганический фосфат, на среду без неорганического фосфата), за короткое время достигать высокого выхода конечного продукта - белка соматотропина.
Конструирование рекомбинантного гена, кодирующего соматотропин человека, осуществлено на основе плазмиды pRS316 4887 п.о. [Sikorski and Hieter P. // A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae, 1989, Genetics 122 (1): 19-27]. Синтезированный рекомбинантный ген встроен в плазмиду pRS316 по сайтам XbaI/BamHI. Искусственный ген, кодирующий соматотропин, фланкированный сайтами рестриктаз XbaI и BamHI, получают химико-ферментативным синтезом набора олигонуклеотидных фрагментов с последующей их сборкой и амплификацией при помощи полимеразной цепной реакции (ПЦР). Перед лигированием для генерации липких концов амплификат и векторную плазмиду обрабатывают рестриктазами XbaI и BamHI. Лигазную смесь используют для трансформации компетентных клеток E.coli DH5α. Отбор положительных клонов проводят при помощи ПЦР с использованием специфических праймеров, с последующим рестриктным анализом выделенной плазмидной ДНК рестриктазами PstI и XbaI. Структуру гена, кодирующего рекомбинантный соматотропин, определяют секвенированием по методу Сенгера на приборе ABI 377. Она должна полностью соответствовать нуклеотидной последовательности исходного искусственного гена соматотропина (фиг.1).
Ввиду высокой вариабельности индекса использования кодонов у дрожжей возможны отклонения в представленной нуклеотидной последовательности гена соматотропина. Изобретательский уровень распространяется на последовательности с гомологией от 90% до 100%, представленной в данном изобретении.
Рекомбинантная плазмидная ДНК pPBS-St9, кодирующая полипептид соматотропин, характеризуется следующими признаками:
- имеет молекулярную массу около 4,1 Мда (6266 п.о.);
- кодирует полипептид соматотропин;
- состоит из: XbaI/BamHI-фрагмента ДНК плазмиды pRS316-St, содержащей синтетический ген соматотропина, плазмиды pRS316, включающего бактериальные маркер ампициллин-резистентности и ориджин репликации, полилинкер в составе участка гена LacZ, ген URA3 S. cerevisiae, последовательности ARS и CEN S. cerevisiae; последовательность, фланкированная сайтами SacI/XbaI интерлейкина человека hIL-1beta из плазмиды YEpsec1-hI1; сигнал терминации транскрипции гена CYC1 дрожжей из плазмиды рСМ251 встроен по сайтам HindIII-BamHI, промотор гена PHO5 амплифицирован с хромосомной ДНК штамма 74-Д694 и затем встроен по сайтам SacII-SacI в вектор. Уникальные сайты узнавания PstI и XbaI дают с фрагментами 2240 и 6022 п.о. при обработке соответствующими рестриктазами.
Плазмида содержит маркерный ген для селекции - ген URA3. В качестве реципиента для данной плазмиды выступает штамм, несущий полную делецию гена ura3. Способность синтезировать урацил будет характерна только для клеток, несущих плазмиду, поэтому на синтетической среде без экзогенных аминокислот и азотистых оснований будут расти только стабильные трансформанты. В качестве штамма использован штамм дрожжей Saccharomyces cerevisiae BY4739 [leu2 ura3 lys2 prc1::LEU2].
Для получения штамма-продуцента рекомбинантного соматотропина плазмидную ДНК pPBS-St9 используют для трансформации компетентных клеток дрожжей BY4739 prc1::LEU2 и проводят отбор клонов, сохраняющих уровень биосинтеза рекомбинантного полипептида не ниже 20% от суммарного клеточного белка в течение, по крайней мере, шести последовательных пассирований. Для этого клоны трансформированных плазмидной ДНК выращивают в богатой среде YPD с добавлением KH2PO4 в концентрации 1 г/л в течение 12-14 часов, инокулируют новую порцию питательной среды в соотношении 1:100, инкубируют культуру до достижения оптической плотности А600=1, отмывают от питательной среды, добавляют богатую среду YEPD и инкубируют еще 3-6 часов. Получение из клеток продуцента рекомбинантного соматотропина включает следующие стадии: отделение культуральной жидкости; солюбилизацию и восстановление целевого белка, его рефолдинг и окончательную очистку.
Исходный штамм BY4739 генотипа МАТа leu2 lys2 ura3 свободно продается фирмой Gentaur, см. ссылку http://www.clonagen.com/clonagen/c1106c02-bbb5-45a3-a8bf-b011aabfle4c/yeast_parental_strains_by4742_product.aspx, и может быть использован для осуществления данного изобретения.
Полученный штамм-продуцент Saccharomyces cerevisiae BY4739 prcl::LEU2/pPBS-St9 характеризуется следующими признаками.
Морфологические признаки: клетки округлой формы.
Культуральные признаки: при росте на агаризованной среде YEPD колонии круглые, гладкие, белые, край ровный. При росте на жидкой среде YEPD образуют интенсивную ровную муть.
Физико-биологические признаки: клетки растут при температуре от 4°С до 40°С при оптимуме рН 5,5-6,0. В качестве источника азота используют как минеральные соли в аммонийной форме, так и органические соединения в виде пептона, триптона, дрожжевого экстракта, аминокислот и т.д. В качестве источника углерода используют глюкозу, глицерин, сахарозу.
Преимущества предлагаемого изобретения заключаются, во-первых, в использовании при химико-ферментативном синтезе гена соматотропина максимально широкого набора кодонов, являющихся оптимальными для продукции белка в Saccharomyces cerevisiae, расположение которых в синтетическом гене устраняет возможность образования на синтезируемой мРНК протяженных "шпилек", потенциально ингибирующих трансляцию. На фиг.1 приведена нуклеотидная последовательность XbaI/BamHI-фрагмента плазмиды pPBS-St9, содержащая рекомбинантный ген соматотропина человека, оптимизированный для экспрессии в дрожжах S. cerevisiae, объединенный с фрагментом гена интерлейкина и сайтом эндопептидазы Kех2; соответствующая ей аминокислотная последовательность представлена на фиг.2.
Во-вторых, применение для биосинтеза рекомбинантного белка оптимальных регуляторных элементов, контролирующих его экспрессию: регулируемый промотор PHO5, высокоэффективный терминатор транскрипции CYC1, последовательность гена интерлейкина человека hIL-1beta для выброса (экскреции) белка в межклеточное пространство, что значительно упрощает очистку белка, и позволяет добиться более высокой чистоты в сравнении с белком, имеющим внутриклеточную локализацию.
В третьих, преимущество предлагаемого штамма-продуцента заключается в использовании дрожжей с делецией вакуолярной пептидазы С, что обеспечивает стабильность чужеродного белка в клетках дрожжей за счет подавления протеолитического расщепления синтезируемого de novo рекомбинантного гормона роста (соматотропина).
Клетки штамма S. cerevisiae BY4739 prc1::LEU2/pPBS-St9 являются суперпродуцентом. При индукции промотора PHO5 происходит эффективный биосинтез рекомбинантного соматотропина, который накапливается в клетках в количестве не менее 20% суммарного белка дрожжей.
На фиг.1 представлена нуклеотидная последовательность XbaI/BamHI-фрагмента плазмиды pPBS-St9, содержащая рекомбинантный ген соматотропина человека, оптимизированный для экспрессии в дрожжах S. cerevisiae, объединенный с фрагментом гена интерлейкина (отмечен жирным шрифтом) и сайтом эндопептидазы Kех2 (отмечен подчеркиванием); на фиг.2 представлена аминоксилотная последовательность XbaI/BamHI-фрагмента плазмиды pPBS-St9, содержащая рекомбинантный ген соматотропина человека, оптимизированный для экспрессии в дрожжах S. cerevisiae, объединенный с фрагментом гена интерлейкина (отмечен жирным шрифтом) и сайтом эндопептидазы Kех2 (отмечен подчеркиванием); на фиг.3 представлена физическая карта плазмиды pPBS-St9; на фиг.4 - нуклеотидная последовательность промотора гена PHO5; на фиг.5 - нуклеотидная последовательность терминатора транскрипции CYC1.
Следует понимать, что специалист в данной области техники может без дополнительных экспериментов, с учетом настоящего описания, применить настоящее изобретение на практике в полном объеме. Следовательно, приведенные в данном контексте примеры представлены только для иллюстрации изобретения, не уменьшающие объем притязаний.
ПРИМЕР 1.
Конструирование рекомбинантной плазмидной ДНК pPBS-St9. Плазмида pPBS-St9 для экспрессии соматотропина включает в себя промотор гена PHO5, фрагменты ДНК, кодирующие 24 N-терминальных аминокислоты интерлейкина человека hIL-1beta (Ser5-Ala28), сайт для расщепления эндопептидазой Kех2 (Lys-Arg), 191 а.к. соматотропина, после которых находится сигнал терминации транскрипции гена CYC1 дрожжей.
Рекомбинантная плазмидная ДНК pPBS-St9 сконструирована на основе плазмиды pRS316 [Sikorski and Hieter P. // A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae, 1989, Genetics 122 (1): 19-27] - стандартного центромерного челночного вектора, включающего бактериальные маркер ампициллин-резистентности и ориджин репликации, полилинкер в составе участка гена LacZ, ген URA3 S. cerevisiae, последовательности ARS и CEN Saccharomyces cerevisiae. В качестве источника последовательности интерлейкина человека hIL-1beta была использована плазмида YEpsec1-hI1 [Baldari С, Murray JAH, Ghiara P, Cesareni G, Galeotti CL // A novel leader peptide which allows efficient secretion of a fragment of human interleukin 1β in Saccharomyces cerevisiae, 1987, EMBO J 6: 229-234]. Последовательность была амплифицирована с использованием пары праймеров:
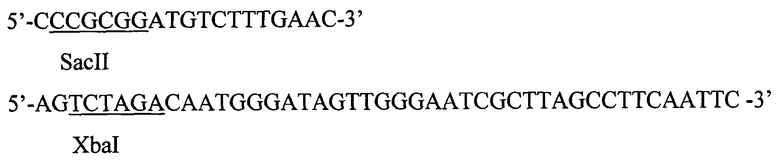
Полученный фрагмент затем был встроен в вектор по сайтам рестрикции SacII-XbaI.
Сигнал терминации транскрипции гена CYC1 дрожжей был вырезан из плазмиды рСМ251 [Belli G, Gari E, Piedrafita L, Aldea M, Herrero E. // An activator/repressor dual system allows tight tetracycline-regulated gene expression in budding yeast, 1998, Nucleic Acids Res. 26 (4): 942-7.] по сайтам рестрикции HindIII-BamHI и встроен в вектор по этим же сайтам.
Промотор гена PHO5 был получен путем ПЦР с парой праймеров
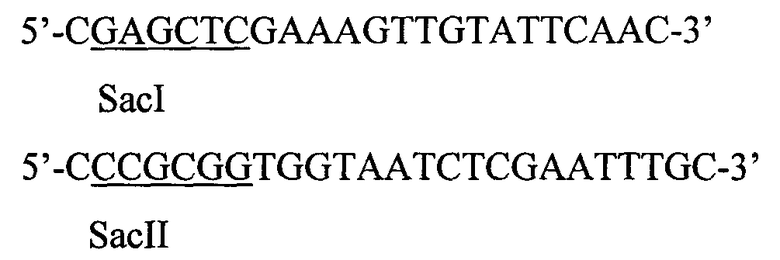
с хромосомной ДНК дрожжей (штамм BY4739 и затем встроен по сайтам SacII-SacI в вектор).
Рекомбинатный гена соматотропина человека получен методом химико-ферментативного синтеза и клонирован в рекомбинантную плазмидную ДНК pRS316 по сайтам XbaI-BamHI.
Конструирование рекомбинантной плазмидной ДНК pRS316-St9.
Нуклеотидную последовательность, соответствующую гену соматотропина, получают химико-ферментативным синтезом. Для этого теоретически рассчитанную последовательность ДНК разбивают на перекрывающиеся фрагменты размером около 50 п.о. Химический синтез олигонуклеотидов, соответствующих этим фрагментам, выполняют твердофазным фосфоамидитным методом при помощи, например, ДНК-синтезатора ASM-900 (БИОССЕТ, Новосибирск) с наращиванием олигонуклеотидной цепи в направлении от 3'-конца к 5'-концу с помощью защищенных фосфамидитов - 5'-диметокситритил-N-ацил-2'-дезоксинуклеозид-3'-O-(β-цианэтил-диизопропиламино)-фосфитов, активированных тетразолом. Синтез проводят в масштабе 0,5-0,7 мкмоль, используя в качестве носителя пористое стекло (размер пор 500 А), к которому через 3'-сукцинатную связь присоединяют первое нуклеозидное звено (нагрузка 20-30 мкмоль/г). Полученные олигонуклеотиды подвергают 5'-концевому фосфорилированию с использованием Т4 полинуклеотидкиназы (Fermentas, Литва). Для этого олигонуклеотиды в количестве 20 пмоль смешивают с ферментом в количестве 10 ед. в буферном растворе, содержащем 50 мМ Tris-HCl (pH 7,6 при 25°С), 10 мМ MgCl2, 5 мМ дитиотреита, 1 мМ спермидина, 0,1 мМ АТФ и 0,1 мМ ЭДТА. Реакцию ведут 30 минут, полинуклеотид киназу инактивируют нагреванием до 65°С в течение 10 мин.
Фосфорилированные олигонуклеотиды смешивают в эквимолярном соотношении в 50 мкл буфера, содержащего 20 мМ трис-HCl, pH 7,56, 10 мМ MgCl2, 0,2 мМ rATP, 10 мМ дитиотреита, прогревают до 65°С, медленно охлаждают до 37°С в течение часа и добавляют 10 ед. ак. Т4-ДНК-лигазы. Реакцию лигирования ДНК проводят 4 ч при 37°С. 0,1 мкл полученного раствора используют в качестве матрицы в полимеразной цепной реакции (ПЦР) в присутствии термостабильной ДНК-полимеразы Pfu и специфических праймеров
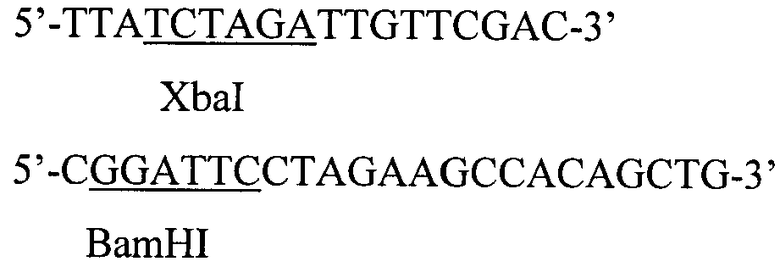
Проводят 25 циклов амплификации (95°С, 20 с; 62°С, 40 с; 72°С, 60 с) для синтеза полноразмерного фрагмента ДНК, содержащего последовательность гена соматотропина, фланкированного сайтами узнавания рестриктаз BamHI и XbaI. Продукт амплификации гидролизуют рестриктазами BamHI и XbaI, очищают электрофорезом в 5% акриламидном геле, полосу ДНК величиной около 600 п.о. выделяют из геля методом электроэлюции и осаждают ДНК из раствора этанолом.
Для приготовления плазмидной ДНК pRS316-St с рекомбинантным геном соматотропина исходный векторную плазмидную ДНК pRS316 (3 мкг, 1 пмоль) обрабатывают в 40 мкл буфера 2xY (33 мМ трис-ацетат, pH 7,9, 10 мМ Mg-ацетат, 66 мМ К-ацетат 1, 0,5 мМ DTT, 0,1 мг/мл BSA) рестриктазами BamHI и XbaI (по 10 ед. акт) в течение 1 ч при 37°С. Полученный фрагмент ДНК величиной около 4,8 т.п.о. после электрофоретического разделения в 1% агарозном геле выделяют из геля методом электроэлюции и осаждают ДНК из раствора этанолом. 1 мкг полученного векторного фрагмента лигируют с 2 пмоль BamHI/XbaI-фрагмента размером 576 п.о., содержащего синтетический ген рекомбинантного соматотропина и сайт эндопептидазы, в 10 мкл буфера (20 мМ трис-HCl, рН 7,56, 10 мМ MgCl2, 0,2 мМ rАТР, 10 мМ дитиотреита) с помощью 10 ед. акт. Т4-ДНК-лигазы в течение 12 ч при 10°С.
На завершающем этапе BamHI/XbaI-фрагмент плазмиды pRS316-St, содержащий рекомбинантный ген соматотропина и сайт эндопептидазы, лигировался в итоговую плазмиду. На фиг 3. представлена физическая карта плазмидной ДНК pPBS-St9.
Полученной лигазной смесью трансформируют клетки штамма DH5a Escherichia coli. Для получения компетентных клеток Escherichia coli, бактерии выращивают в 100 мл среды LB при 37°С до достижения культурой густоты клеточной суспензии, соответствующей 0,4-0,6 ед. оптической плотности при длине волны 550 нм. Клеточную суспензию охлаждают в ледяной бане, центрифугируют при 5000 об/мин в течение 10 минут при 4°С. Клетки суспендируют в 100 мл 10 мМ хлористого натрия, собирают центрифугированием в тех же условиях. Далее клетки суспендируют в 50 мл 75 мМ хлористого кальция, выдерживают в ледяной бане в течение 40 минут, осаждают центрифугированием в тех же условиях и суспендируют в 1 мл 75 мМ хлористого кальция. К суспензии компетентных клеток добавляют глицерин до конечной концентрации 15%, разделяют на аликвоты и хранят при -70°С.
На этапе трансформации компетентных клеток плазмидой pPBS-St9 суспензию компетентных клеток размораживают в ледяной бане, добавляют 1 мкл лигазной смеси и инкубируют в ледяной бане в течение 40 минут. Далее клетки подвергают действию теплового шока при 42°С в течение 2 минут, после чего инкубируют в 1,5 мл среды LB при 37°С в течение 1 часа. Клетки собирают центрифугированием при 5000 об/мин в течение 10 минут и высевают на чашки Петри со средой LB, содержащей 2% агара и 50 мг/л ампициллина. Чашки инкубируют при 37°С в течение 12-16 часов. Первичный отбор клонов, содержащих нужную плазмиду, проводят методом "ПЦР с клонов" с использованием специфических праймеров:
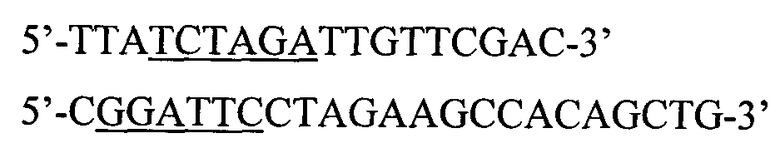
Из выросших отдельных клонов трансформантов выделяют плазмидную ДНК при помощи методики (микрометод), описанной ниже, за исключением того, что клетки Escherichia coli выращивают в 10 мл LB, и соответственно объемы всех растворов уменьшают в 100 раз. Кроме того, вместо стадии центрифугирования в градиенте плотности хлористого цезия проводят обработку ДНК панкреатической РНКазой. Для этого нуклеиновые кислоты, осажденные изо-пропиловым спиртом, растворяют в 100 мкл буфера ТЕ, добавляют 10 мкл раствора РНКазы (1 мг/мл) и инкубируют 30 минут при 37°С.
Для получения препаративных количеств плазмиды используют максиметод. Для этого клетки бактерий Escherichia coli, содержащие плазмиду pPBS-St9, выращивают в течение ночи в 1 л питательной среды LB (1% пептона, 0,5% дрожжевого экстракта, 1% хлористого натрия), содержащей ампициллин в концентрации 50 мг/л. Клетки собирают центрифугированием при 5000 об/мин в течение 10 минут при 4°С, осадок суспендируют в 20 мл 25 мМ трис-хлоридного буфера (рН 8,0), содержащего 10 мМ ЭДТА и 50 мМ глюкозы. К суспензии добавляют 30 мг лизоцима и инкубируют 10 минут при комнатной температуре. Далее добавляют 40 мл 0,2 М раствора NaOH, содержащего 1% додецилсульфата натрия, осторожно перемешивают и инкубируют в течение 10 минут при комнатной температуре. Раствор нейтрализуют добавлением 30 мл 3 М ацетата натрия (рН 5,0) и выдерживают в течение 10 минут при 4°С. После этого центрифугируют при 14000 об/мин в течение 40 минут при 4°С. К супернатанту добавляют 0,6 объема изо-пропилового спирта, выдерживают 20 минут при комнатной температуре и центрифугируют при 14000 об/мин в течение 20 минут при 20 С. Полученный осадок промывают 70% этиловым спиртом, высушивают в вакууме и растворяют в 4 мл дистиллированной воды. Далее добавляют 4,2 г хлористого цезия и 0,36 мл раствора бромистого этидия (10 мг/мл). Полученный раствор выдерживают в течение 1 часа при 4°С, затем центрифугируют при 15000 об/мин в течение 15 минут. Супернатант центрифугируют при 70000 об/мин в течение 16 часов в центрифуге TL-100 ("Beckman"). После центрифугирования отбирают полосу плазмидной ДНК (нижнюю из двух флюоресцирующих в ультрафиолетовом свете полос), дважды экстрагируют бромистый этидий равным объемом изо-амилового спирта, разбавляют в два раза дистиллированной водой и осаждают плазмидную ДНК двумя объемами этилового спирта и 1/15 объема 3 М ацетата натрия (рН 5,0). Осадок собирают центрифугированием при 10000 об/мин в течение 10 минут, промывают 70% этиловьм спиртом и растворяют в 0,5-1 мл буфера ТЕ (10 мМ трис-хлоридный буфер, рН 8,0, содержащий 1 мМ ЭДТА). Концентрацию плазмидной ДНК определяют по поглощению раствора при длине волны 260 нм. Чистоту препарата контролируют при помощи электрофореза в 0,7% агарозном геле в буфере ТВЕ (0,1 М трис-боратный буфер, рН 8,3, содержащий 1 мМ ЭДТА).
Из клеток идентифицированного клона препаративно выделяют плазмидную ДНК по приведенным ранее методикам и используют ее для трансформации клеток дрожжей, как описано в примере 2.
ПРИМЕР 2.
Штамм-продуцент рекомбинантного соматотропина.
Для получения штамма дрожжей Saccharomyces cerevisiae продуцента соматотропина нами использован штамм BY4739 генотипа МАТа leu2 lys2 ura3, с инактивированной вакуолярной пептидазой С и двумя маркерами ауксотрофности, трансформированный плазмидой pPBS-St9.
Для получения делеции вакуолярной пептидазы С мы проводили ПЦР хромосомной ДНК штамма BY4739 с праймерами

Первые 40 нуклеотидов этих праймеров гомологичны последовательностям, фланкирующим ген PRC1 на хромосоме, оставшиеся 20-кодирующей последовательности гена LEU2. В результате получается фрагмент, содержащий полноразмерный ген LEU2, фланкированный 3' и 5' последовательностями гена PRC1. Трансформация штамма [ura3 leu2 lys2] таким фрагментом приводит к гомологичной рекомбинации по фланкирующим последовательностям, появлению Leu+ клонов, в которых ген PRC1 замещен геном LEU2.
В дальнейшем мы трансформировали штамм [leu2 ura3 lys2 prc1::LEU2] сконструированной нами плазмидой с геном соматотропина и отбирали Ura+ клоны, несущие плазмиду.
Для этого клетки дрожжей выращивают в 100 мл среды YEPD до достижения культурой оптической плотности, соответствующей 2-4 ед. поглощения при длине волны 600 нм. Клетки дважды промывают стерильной водой, после чего суспендируют в 0,3 мл 100 мМ раствора ацетата лития и инкубируют при 30°С в течение 30 минут. К 50 мкл полученной суспензии клеток добавляют 0,1-1 мкг плазмидной ДНК, 50 мкг ДНК спермы лосося, предварительно денатурированной нагреванием (10 минут при 100°С) и 0,3 мл раствора 100 мМ ацетата лития, содержащего 40% полиэтиленгликоля PEG-4000. Далее пробу инкубируют 30 минут при 30°С и 20 минут при 42°С, помещают на 15 секунд в ледяную баню и центрифугируют 10 секунд при 10000 об/мин. Клетки суспендируют в 1 мл стерильной воды и высевают на твердую среду MD. Клоны трансформантов вырастают через 4-6 суток.
Для анализа продукции соматотропина клетками трансформантов их выращивают в 50 мл жидкой среды ПЕП до стационарной фазы роста. Клетки собирают центрифугированием при 3000 об/мин в течение 10 минут, промывают водой, суспендируют в 1 мл 50 мМ трис-хлоридного буфера (рН 7,5), содержащего 1 мМ ФМСФ (фенилметилсульфонилфторид), добавляют 1 г стеклянных бус (0,5 мм) и разрушают в дезинтеграторе "Braun" в течение 1 минуты при температуре 4°С. Полученный гомогенат центрифугируют при 10000 об/мин в течение 10 минут, осадок промывают 10-кратным объемом того же буфера и суспендируют в 1 мл 50 мМ натрий-фосфатного буфера (рН 7,0), содержащего 2% додецилсульфата натрия и 5% 2-меркаптоэтанола, и инкубируют 5 минут в кипящей водяной бане. По окончании инкубации пробу центрифугируют 15 минут при 12000 об/мин и в супернатанте определяют содержание соматотропина при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия и последующей гибридизации со специфическими антителами к соматотропину человека. Разделение белков проводят в 15% полиакриламидном геле в стандартной системе буферов (электродный буфер: 25 мМ трис, 192 мМ глицин, 0,1% додецилсульфат натрия, рН 8,3; буфер для геля: 375 мМ трис-хлоридный буфер, рН 8,8).
Параллельно проводят разделение белков контрольного (не трансформированного плазмидой) штамма BY4739 prcl::LEU2, выращенного и гомогенизированного в идентичных условиях.
В качестве стандартов молекулярной массы используют карбоангидразу, ингибитор трипсина, миоглобин, лизоцим фирмы "Sigma". По окончании электрофореза белки ренатурируют, выдерживая гели 15 минут в 10 мМ трис-хлоридном буфере (рН 7,5), содержащем 4 М мочевину, 20 мМ ЭДТА. Далее белки переносят на нитроцеллюлозную мембрану в 25 мМ трис-192 мМ глициновом буфере (рН 8,3), содержащем 20% метилового спирта, при 30-40 В, в течение 1,5 часов на приборе фирмы "Bio-Rad". Далее мембрану выдерживают в буфере TBST (10 мМ трис-хлоридный буфер (рН 8,0), 150 мМ хлористого натрия, 0,05% твин-20, 1% бычьего сывороточного альбумина) в течение 2 часов при 26°С. Затем помещают мембрану в тот же буфер, содержащий разведенные в 1000 раз кроличьи поликлональные антитела к соматотропину человека ("Abeam"), и инкубируют 2 часа при 26°С. Далее трижды промывают мембрану буфером TBST и инкубируют 1 час при 26°С с разбавленным в 7000 раз конъюгатом видоспецифических антител к иммуноглобулинам кролика и пероксидазы хрена ("Abcam"). После отмывки мембраны буфером PBST (58 мМ двузамещенного фосфата натрия, 17 мМ однозамещенного фосфата натрия, 68 мМ хлористого натрия, 0,1% Твин-20) ее помещают в раствор субстратов для пероксидазы: 0,02% DAB (3,'3-диаминобензидин тетрагидрохлорид), 0,006% перекись водорода в 10 мМ трис-хлоридном буфере, рН 7,5. Параллельно окрашивают гели 0,15% раствором кумасси G250 в 25% изопропаноле и 10% уксусной кислоте и отмывают в 10% уксусной кислоте. При сравнении спектра белков двух штаммов у штамма BY4739 prc1::LEU2/pPBS-St9 обнаруживают появление дополнительной белковой полосы, дающей положительную реакцию с антителами к соматотропину человека, с молекулярной массой 22 кДа, что соответствует молекулярной массе соматотропина. Уровень синтеза рекомбинантного соматотропина определяют, сравнивая интенсивность окрашивания полосы этого белка с полосой стандартного соматотропина. Согласно полученным данным клетки дрожжей штамма BY4739 prc1::LEU2/pPBS-St9 синтезируют внутриклеточно около 50 мг соматотропина на литр культуры дрожжей. При проведении анализа на подавление цитопатической активности вируса везикулярного стоматита в культуре человеческих фибробластов стандартными методами [М.G.Tovey et al. // Antiviral Activity of Bovine Interferons on Primate Cells, 1977, J. Gen. Virol., 36, 341-344] обнаруживается, что препарат соматотропина, синтезированного штаммом дрожжей Saccharomyces cerevisiae BY4739 prc1::LEU2/pPBS-St9 биологически активен. Активность препарата составляет 20-40 млн ед./л.
Суммируя вышесказанное, можно заключить, что полученный штамм дрожжей Saccharomyces cerevisiae BY4739 prc1::LEU2/pVBS-St9 синтезирует соматотропин человека в количестве, достаточном для его очистки в лабораторном и промышленном масштабе. В результате такой очистки могут быть получены препараты соматотропина, имеющие терапевтическую ценность, пригодные для применения в медицине.
Штамм дрожжей Saccharomyces cerevisiae BY4739 [leu2 ura3 lys2 prc1::LEU]/pPBS-St9, депонирован в Петергофской генетической коллекции микроорганизмов под номером С-016 - продуцент рекомбинантного соматотропина человека.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ КОНСТРУИРОВАНИЯ ШТАММОВ ДРОЖЖЕЙ Saccharomyces cerevisiae - СТАБИЛЬНЫХ ПРОДУЦЕНТОВ СОМАТОТРОПИНА ЧЕЛОВЕКА, ШТАММ ДРОЖЖЕЙ Saccharomyces cerevisiae - СТАБИЛЬНЫЙ ПРОДУЦЕНТ СОМАТОТРОПИНА ЧЕЛОВЕКА (ВАРИАНТЫ) | 2009 |
|
RU2420567C1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА СЕКРЕТИРУЕМОГО СОМАТОТРОПИНА ЧЕЛОВЕКА И ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE - ПРОДУЦЕНТ СЕКРЕТИРУЕМОГО СОМАТОТРОПИНА ЧЕЛОВЕКА | 2011 |
|
RU2460795C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PES1-6, КОДИРУЮЩАЯ ПОЛИПЕПТИД СОМАТОТРОПИН, И ШТАММ ESCHERICHIA COLI BL 21(DE3)/PES1-6-ПРОДУЦЕНТ РЕКОМБИНАНТНОГО СОМАТОТРОПИНА | 2002 |
|
RU2233879C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, КОДИРУЮЩАЯ ПОВЕРХНОСТНЫЙ АНТИГЕН (HBSAG) ВИРУСА ГЕПАТИТА B, ЕЕ ПОЛУЧЕНИЕ И ШТАММ ДРОЖЖЕЙ HANSENULA POLYMORPHA - ПРОДУЦЕНТ ПОВЕРХНОСТНОГО АНТИГЕНА (HBSAG) ВИРУСА ГЕПАТИТА B | 2002 |
|
RU2207374C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PSTH 2191, КОДИРУЮЩАЯ СИНТЕЗ СОМАТОТРОПИНА ЧЕЛОВЕКА, И ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ СОМАТОТРОПИНА ЧЕЛОВЕКА | 1986 |
|
SU1387414A1 |
| ШТАММ ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAL, СОДЕРЖАЩИЙ РЕКОМБИНАНТНУЮ ПЛАЗМИДУ YEP 63/AB, - ПРОДУЦЕНТ ПРОИЗВОДНОГО М-БЕЛКА ВИРУСА ГЕПАТИТА В ЧЕЛОВЕКА | 1994 |
|
RU2082759C1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА, ШТАММ Saccharomyces cerevisiae - ПРОДУЦЕНТ ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА (ВАРИАНТЫ) | 2010 |
|
RU2427645C1 |
| Рекомбинантная плазмидная ДНК pJDB (MSIL), обеспечивающая синтез интерлейкина-2 человека в клетках дрожжей SасснаRомUсеS ceReUISIaL, способ ее получения и штамм дрожжей SасснаRомYсеS ceReUISIaL - продуцент интерлейкина-2 человека | 1988 |
|
SU1770359A1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА ПАУТИНЫ, СЛИТЫЙ БЕЛОК, РЕКОМБИНАНТНАЯ ДНК, ВЕКТОР ЭКСПРЕССИИ, КЛЕТКА-ХОЗЯИН И ШТАММЫ-ПРОДУЦЕНТЫ | 2010 |
|
RU2451023C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА PFS 19, ОПРЕДЕЛЯЮЩАЯ СИНТЕЗ ПОВЕРХНОСТНОГО АНТИГЕНА S ВИРУСА ГЕПАТИТА B ЧЕЛОВЕКА | 1992 |
|
RU2115730C1 |



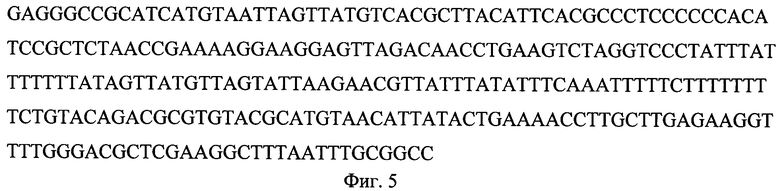
\Изобретение относится к области биотехнологии и генной инженерии. Предложена рекомбинантная плазмидная ДНК pPBS-St9, кодирующая полипептид с последовательностью гормона роста человека соматотропина, имеющая молекулярную массу 4,1 Мда (6266 п.о.), а также штамм Saccharomyces cerevisiae BY4739 [leu2 ura3 lys2 prc1::LEU]/pPBS-St9, содержащий рекомбинантную плазмидную ДНК pPBS-St9, - продуцент рекомбинантного соматотропина. Изобретение может быть использовано для получения рекомбинантного гормона роста человека при лечении карликовости. 2 н.п. ф-лы, 5 ил., 2 пр.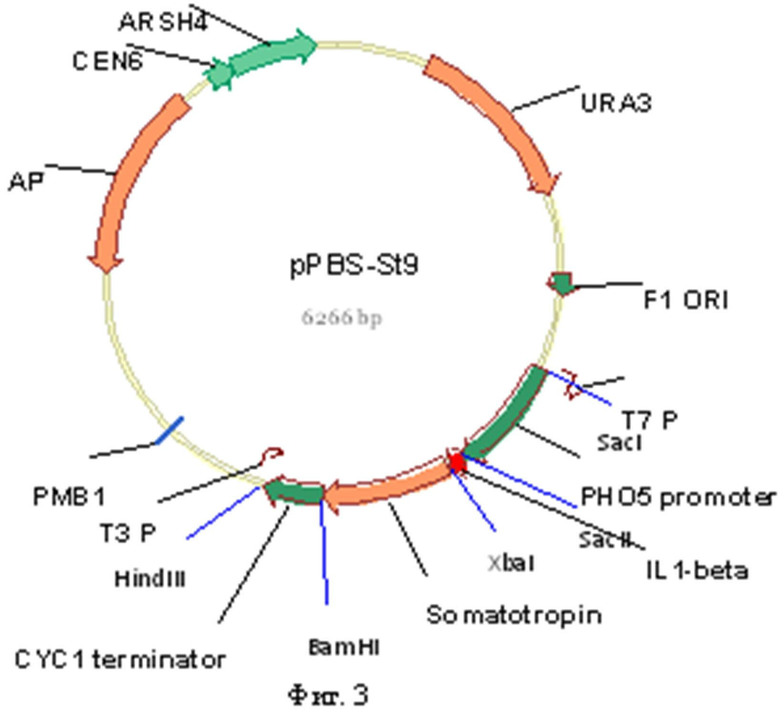
1. Рекомбинантная плазмидная ДНК pPBS-St9, кодирующая полипептид с последовательностью гормона роста человека соматотропина, имеющая молекулярную массу 4,1 МДа (6266 п.о.); состоящая из SacI/HindIII-фрагмента ДНК плазмиды pRS316, содержащего бактериальные маркер ампициллин-резистентности и ориджин репликации, полилинкер в составе участка гена LacZ, уникальные сайты узнавания эндонуклеаз рестрикции, расположенные на следующем расстоянии вправо от сайта PstI: XbaI - 2236 п.о., ген URA3, последовательности ARS и CEN Saccharomyces cerevisiae, a также вставленные в исходную плазмидную ДНК последовательность промотора гена РНO5, введенную по сайтам SacI/SacII в положении 1998-2533 (размер вставки 536 п.о.), фрагменты ДНК, кодирующие 24 N-терминальных аминокислоты интерлейкина человека hIL-1beta (Ser5-Аlа28), вставленные по сайтам SacII/XbaI (положение 2540-2611, размер вставки 72 п.о.), сайт для расщепления эндопептидазой Kех2 (Lys-Arg), 191 а.к. соматотропина, вставленные по сайтам Xba/BamHI (положение 2617-3192, размер вставки 576 п.о.), после которых встроена сигнальная последовательность терминации транскрипции гена CYC1 дрожжей, вставленная по сайтам HindIII/BamHI (положение 3198-3460, размер вставки 262 п.о.).
2. Штамм Saccharomyces cerevisiae BY4739 [leu2 ura3 lys2 prc1::LEU]/pPBS-St9, содержащий рекомбинантную плазмидную ДНК pPBS-St9 по п.1, - продуцент рекомбинантного соматотропина.
| http://sequence-www.stanford.edu/group/yeast-deletion-project/downloads.html | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| CECILIA J | |||
| et al | |||
| Genomic Screen for Vacuolar Protein Sorting Genes in Saccharomyces cerevisiae | |||
| Molecular Biology of the Cell | |||
| Насос | 1917 |
|
SU13A1 |
| Водоподъемное устройство | 1924 |
|
SU2487A1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PES1-6, КОДИРУЮЩАЯ ПОЛИПЕПТИД СОМАТОТРОПИН, И ШТАММ ESCHERICHIA COLI BL 21(DE3)/PES1-6-ПРОДУЦЕНТ РЕКОМБИНАНТНОГО СОМАТОТРОПИНА | 2002 |
|
RU2233879C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PSTH 2191, КОДИРУЮЩАЯ СИНТЕЗ СОМАТОТРОПИНА ЧЕЛОВЕКА, И ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ СОМАТОТРОПИНА ЧЕЛОВЕКА | 1986 |
|
SU1387414A1 |

