Данное изобретение относится к способу трансформации C3-растения для обеспечения его C4-циклом путем введения двух или нескольких ферментов, которые участвуют в C4-фотосинтетическом пути.
Уровень техники
Известны три типа фотосинтетического пути в высших растениях, а именно типы C3, C4 и CAM. Ткани листьев растений, имеющих фотосинтетический путь C4-типа (далее иногда называемых C4-растениями), содержат мезофилльные клетки и клетки обкладки сосудистых пучков, существующие вокруг сосудисто-волокнистых пучков, образующие характерную структуру ткани листа, называемую анатомией Кранц-типа. C4-растения фиксируют диоксид углерода в C4- соединение действием фосфоенолпируваткарбоксилазы (далее иногда называемой ФЕПК (РЕРС)), локализованной в цитоплазме мезофилльных клеток. Фиксированный диоксид углерода высвобождается декарбоксилазой в клетки обкладки сосудистых пучков, что увеличивает уровень диоксида углерода вблизи рибулозо-1,5-бифосфаткарбоксилазы/оксигеназы (далее иногда называемой Rubisco), которая является ферментом основной фиксации диоксида углерода. Метаболит, образующийся в результате декарбоксилирования в клетках обкладки сосудистых пучков, переносится в мезофилльные клетки и превращается в фосфоенолпируват (далее иногда называемый ФЕП), субстрат для ФЕПК, под действием пируват-ортофосфатдикиназы (далее называемой иногда ПФДК), локализованной в мезофилльных клетках, с одновременным потреблением АТФ. Т.е. два типа клеток в зеленых листьях C4- растений являются функционально дифференцированными: мезофилльная клетка является местом образования C4-соединений при первоначальной фиксации углерода, а также местом регенерации субстрата ФЕПК, тогда как клетка обкладки сосудистого пучка является местом декарбоксилирования C4-соединения и основной фиксации диоксида углерода по пути цикла Кальвина-Бенсона.
Эти три стадии, т.е. фиксация диоксида углерода ФЕПК, высвобождение диоксида углерода вблизи Rubisco и регенерация субстрата ФЕПК, сопровождающаяся потреблением АТФ, составляют систему циклической реакции, которую называют C4-фотосинтетическим путем. Этот путь обеспечивает C4-растения с повышенной способностью аккумулировать диоксид углерода и избегать снижения эффективности фотосинтеза, которое в ином случае может иметь место при высокой интенсивности света вследствие сверхпродуцирования АТФ (избежание фотоингибирования). Эти свойства не обнаруживаются в C3-растениях, имеющих нормальный фотосинтетический путь (фотосинтез C3-типа). Таким образом, C4-растения не обнаруживают фотодыхания, как C3-растения, и, следовательно, первые обнаруживают меньшее ухудшение в эффективности фотосинтеза, чем последние, при помещении в атмосферу, которая является сухой, имеющей высокую интенсивность света или высокую температуру. Поэтому C4-растения превосходят C3-растения в их способности осуществлять фотосинтез.
Можно ожидать, что C4-фотосинтетический путь мог бы быть введен в C3-растение при помощи скрещивания и селекции. Однако большинство видов, имеющих C4-фотосинтетический путь, и большинство видов, имеющих нормальный C3-фотосинтетический путь, объединены в различные род или семейство, и скрещивание между ними является затруднительным. Кроме того, попытка ввести свойства C4-растения, в которой C3-растение скрещивали с C4-растением, выбранным из того же рода лебеды, была неудачной (Ohsugi, R. Nogyo-gijutsu (1995) Vol. 50, pp. 30-36).
Hudspeth, et. al. наблюдали, что зеленые листья трансгенного табака, в которые был введен ген ФЕПК под контролем промотора гена хлорофилл а/b-связывающего белка (Cab промотора), обнаружили удвоение активности ФЕПК и увеличение уровня малата (Hudspeth, et. al. Plant Physiol., (1992) 98:458-464). Kogami, et al. наблюдали, что зеленые листья трансгенного табака, в которые был введен ген ФЕПК под контролем промотора 35S вируса мозаики цветной капусты, имели приблизительно вдвое более высокую активность ФЕПК, чем нетрансформированный табак (Kogami, et al., Transgenic Research (1994) Vol. 3; 287-296).
Таким образом, Hudspeth, et. al. и Kogami, et al. просто наблюдали накопление C4-соединения, малата, без подтверждения какого-либо изменения в фотосинтетической способности, вызываемого введением ФЕПК в виде единственного гена в C3-растение табака. Клетки C3-растений неспособны быстро декарбоксилировать C4-соединение для подачи диоксида углерода в цикл Кальвина. Таким образом, было бы невозможно посредством простого введения гена ФЕПК в C3-растение, в попытке обеспечения растения способностью к C4-фотосинтетическому пути, концентрировать карбонат или избегать фотоингибирования, когда желательно улучшение фотосинтетической способности C3-растения.
В Japanese Patent Public Disclosure Hei 8-80197 сообщается, что фрагмент ДНК, кодирующий транзитный (транспортный) пептид, был соединен с геном фосфоенолпируваткарбоксикиназы (ФКК (РСК)). Химерный ген вводили в рис, который представляет собой C3-растение, и в результате обнаружили активность этого фермента в неочищенном экстракте зеленых листьев, а также локализацию белка ФКК в хлоропластах. Эти факты свидетельствуют о том, что возможно достичь локализации ФКК в хлоропластах. Однако ничего не сообщается об установлении C4-фотосинтетического пути или изменении фотосинтетической способности в трансгенных растениях.
Ichikawa, et al., Nihon Sakumotsu Gakkai Kiji, Vol. 63, Suppl.2, (1994), p. 247) сообщили, что при введении ПФДК в C3-растения, Arabidopsis и томат, этот белок накапливался в этих растениях. Однако не сообщается об установлении C4-фотосинтетического пути или изменении фотосинтетической способности в трансгенных растениях. В Japanese Patent Publication Hei 6-12990 описано изменение в эффективности фотосинтеза в протопластах семядолей Lycopersicon escu-lentum, когда был введен белок карбоангидраза (называемая здесь иногда КА (СА)). С другой стороны, Majeau et al., сообщают, что сверхэкспрессия КА in vivo не приводила к какому-либо изменению в фотосинтетической способности растения (Plant Mol. Biol. (1994) 25:337-385).
Как обсуждалось выше, прежние попытки введения гена из C4-фотосинтетического пути в C3-растение способом генной инженерии ограничивались введением гена КА, ФЕПК, ФКК или ПФДК в виде единственного гена. Эти попытки не смогли подтвердить присутствия какого-либо C4-фотосинтетического пути или изменения эффективности фотосинтеза, даже если в случае некоторых попыток наблюдали экспрессию введенного гена или ферментативную активность.
Сущность изобретения
Целью данного изобретения является обеспечение способа улучшения фотосинтетической способности C3-растений. Конкретно, данное изобретение обеспечивает способ трансформации C3-растения для обеспечения его C4-фотосинтетическим путем посредством введения двух или более ферментов, которые участвуют в C4-фотосинтетическом пути.
Другой целью данного изобретения является обеспечение растения, которое было трансформировано таким образом, что оно имеет C4-путь, в соответствии со способом данного изобретения.
Следующей целью данного изобретения является обеспечение вектора, который применим для проведения трансформации C3-растения.
Краткое объяснение рисунков
Фиг. 1 - иллюстрация цикла C4-фотосинтеза ФКК-типа.
Фиг. 2 - иллюстрация генной конструкции, используемой в трансформации C3-растения.
Фиг. 3 - график, на котором относительное количество меченого изотопом соединения углерода нанесено в зависимости от времени.
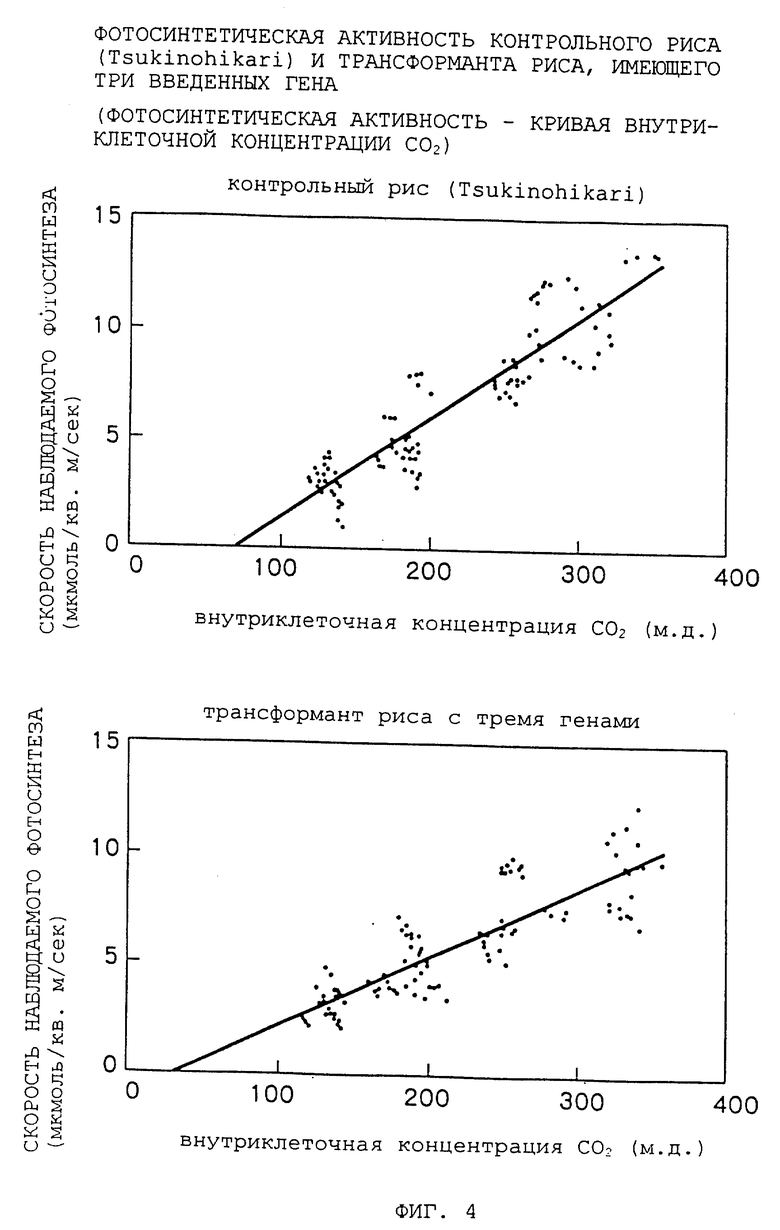
Фиг. 4 - график, показывающий фотосинтетическую активность трансформированного риса.
Подробное объяснение
Данное изобретение было выполнено как результат интенсивного исследования авторами изобретения путем введения гена ФЕПК в C3-растение вместе с геном ФФК, который был соединен с фрагментом ДНК, кодирующим транзитный пептид.
Авторы данного изобретения сконцентрировали внимание на том факте, что в прежних попытках соответствующий ген компонентов C4-фотосинтетического пути вводили в цитоплазму C3-растения в виде единственного гена. Поэтому авторы изобретения предположили, что этот факт может быть причиной, вследствие которой предшествующие попытки не смогли привести в действие C4-фотосинтетический путь или улучшить фотосинтетическую способность, хотя некоторые из трансгенных C3-растений приобретали активность введенных ферментов.
На основании этого предположения авторы данного изобретения сконструировали систему, в которой: гены двух или нескольких ферментов вводят в C3-растение таким образом, что каждый фермент будет экспрессироваться в определенном внутриклеточном местоположении: хлоропласты возьмут на себя роль клеток обкладки сосудистого пучка C4-растения; и два или не сколько ферментов, требующихся в C4-фотосинтетическом пути, будут экспрессироваться одновременно в зеленых мезофилльных клетках. В соответствии с этим планом можно не только обеспечить C3-растение соответствующей ферментативной активностью, но также заставить C3-растение использовать циклическую реакцию, которая имитирует C4-фотосинтетический путь C4-растений, повышая тем самым способность накапливать диоксид углерода в хлоропластах и избегать фотоингибирования, которое может быть вызвано избыточным потреблением АТФ. Ожидается, что растения, обеспеченные такими свойствами, будут обнаруживать улучшенную продуктивность, улучшенную засухоустойчивость, улучшенную устойчивость к высокой температуре (жаростойкость) и улучшенную фотосинтетическую способность в условиях низкой концентрации диоксида углерода, как результат улучшения фотосинтетической способности.
Способ данного изобретения для обеспечения C4-фотосинтетического пути предусматривает: обеспечение мезофилльных клеток зеленого листа C3-растения (например, риса); предоставление возможности ферменту (ФЕПК) первичного процесса фиксации диоксида углерода C4-растения действовать в цитоплазме этих клеток; предоставление возможности декарбоксилазе для C4-растений действовать в хлоропластах этих клеток; и одновременно предоставление возможности экспрессии фермента для регенерации ФЕП либо в цитоплазме, либо в хлоропластах этих клеток.
Декарбоксилазы, применимые для этой цели, включают ФФК. ФФК представляет собой фермент, который декарбоксилирует оксалоацетат с образованием ФЕП при потреблении АТФ. Таким образом, ФФК может быть выгодно использован в качестве декарбоксилазы, поскольку декарбоксилирование, потребление АТФ и регенерация ФЕП могут осуществляться единственным ферментом. Для того, чтобы сделать возможным проявление декарбоксилазой ее активности в хлоропластах, ген этого фермента соединяют с последовательностью транзитного пептида. Транзитный пептид перемещает полипептид декарбоксилазы в хлоропласты, чтобы позволить этому ферменту проявлять его функцию в хлоропластах.
Посредством трансформации, как указывалось выше, путь фиксации диоксида углерода, который имитирует C4-фотосинтетический путь, происходящий из дифференцированной структуры ткани листа C4-растений, учреждается в C3-растении, где цитоплазма мезофильных клеток зеленого листа C3-растения имитирует клетки мезофилла C4-растений, а хлоропласты C3-растений имитируют клетки обкладки сосудистого пучка C4-растений (Фиг. 1). Таким образом, можно обеспечить C3-растение со способностью концентрировать диоксид углерода и избегать фотоингибирования.
Сочетание ФЕПК, декарбоксилазы и фермента регенерации ФЕП будет достаточным для учреждения желательного C4-фотосинтетического пути. Однако в предпочтительном варианте, в цитоплазме может коэкспрессироваться КА для подачи иона бикарбоната в качестве непосредственного субстрата ФЕПК, так что C4-фотосинтетический путь будет функционировать более ровно. Кроме того, ПФДК, которая катализирует образование ФЕП из пирувата, может коэкспрессироваться в дополнение к ФЕПК, ФКК и КА, если желательна подача ФЕП в качестве субстрата для ФЕПК, вследствие чего C4-фотосинтетический цикл будет функционировать еще более гладко.
Далее данное изобретение будет описано более детально.
Данное изобретение относится к способу трансформации C3-растения для обеспечения его C4-фотосинтетическим путем посредством введения в это растение гена, кодирующего ФЕПК, гена, кодирующего ФКК, который был соединен с фрагментом ДНК, кодирующим транзитный пептид.
Гены ФЕПК, которые могут использоваться в данном изобретении, включают гены ФЕПК из бактерий, простейших, растений и т.д. В качестве характерного примера бактериального гена ФЕПК известен ген ФЕПК продуцирующего глутамат штамма Corinebacterium (Japanese Patent Publication Hei 7-83714).
Однако предпочтительными ФЕПК являются ФЕПК растительного происхождения, например ФЕПК, происходящие из кукурузы (Japanese Patent Publication Hei 6-30587), Amaranthus (Rydzik, E.and Berry, J.O., Plant physiol., (1995) 110: 713), Flaveria trinervia (Poetsch, W. , et al., FEBS Lett., (1991) 292: 133-136), табака (Koizumi, N., et al. Plant Mol. Biol., (1991) 17:535-539), сои (Japanese Patent Public Disclosure Hei 6-319567), рапса (Japanese Patent Public Disclosure Hei 6-90766), картофеля (Merkelbach, S., et al. Plant Mol. Biol. , (1993) 23: 881-888), люцерны (Pathariana, S.M., et al. Plant Mol. Biol., (1992) 20:437- 450), Mesembryanthenum crystallinum (Cushman, J.C. and Bohnart, H. J., Nuc. Acid Res., (1989) 6743-6744). Особенно предпочтительна ФЕПК из кукурузы.
Примерами кодирующих ФКК генов, используемых в данном изобретении, являются гены АТФ-зависимых ФКК, происходящих из растений и бактерий. Примеры ФКК растений включают ФКК из Urochioa panicoides (Japanese Patent Public Disclosure Hei 8-80197) и огурца (Kim, D.-J. and Smith, S.M., Plant Mol. Biol. , (1994) 26: 423-434), а примеры бактериальных ФКК включают ФКК из E.coli (Medina, V. et al. , J. Bacteriol. , (1990) 172: 7151-7156) и Rhizobium (Jsteras, M. et al., J. Bacteriol., (1995) 177: 1452-1460). Особенно предпочтителен ген ФКК, происходящий из растения, в частности из Urochioa panicoides.
Далее, требуется, чтобы ФКК проявляла свою функцию в хлоропластах, как отмечалось выше. Для гарантии этого с геном ФКК соединяют фрагмент ДНК, кодирующий транзитный пептид.
Ряд последовательностей транзитного пептида, гены которых могут быть соединены с геном ФКК, сообщался среди белков, локализованных в хлоропластах (Keegstra, K. et al., Annu. Rev. Plant Mol. Biol., (1989) 40:471-501). Для данного изобретения предпочтительно, чтобы последовательность транзитного пептида происходила из белков риса. Особенно предпочтительно, чтобы последовательность транзитного пептида была последовательностью малой субъединицы Rubisco (SEQ ID N 2), которая может быть получена в соответствии со способом, описанным в примерах ниже. Фрагмент ДНК, кодирующий последовательность транзитного пептида, соединен в рамке считывания со структурным геном ФКК против хода транскрипции и предпочтительно непосредственно против хода транскрипции от него.
В данном изобретении ген, кодирующий КА, также может быть введен в C3-растения в цитоплазму для обеспечения бикарбонатного иона в качестве субстрата для ФЕПК, как описано выше.
Ряд генов, кодирующих КА, которые могут быть использованы в данном изобретении, известны, например гены КА, происходящие из животных и растений. Однако гомологии последовательностей между КА высших растений и других организмов не являются высокими. Далее, на ферментативную активность КА из высших растений влияет неорганический фосфат (Sultemeyer, D. et al., Physiol. Plant., (1993) 88; 179-190). Таким образом, предпочтительно, чтобы этот ген происходил из растения, такого как шпинат (Burnell et al., Plant Physiol. , (1990) 92:37-40), горох (Roeske, C.A. and Ogren, W.L., Nuc Acid Res., (1990) 18: 3413), Arabidopsis (Raines, C.A., et al., Plant Mol. Biol., (1992) 20: 1143-1148), рис (WO 95/11979) и кукуруза (WO 95/11979). Особенно предпочтительна КА, происходящая из шпината. Поскольку КА шпината локализована в хлоропластах, ген этого фермента содержит район, кодирующий транзитный пептид. Таким образом, SEQ ID N 3, район, кодирующий транзитный пептид, элиминируют введением точковой мутации, как описано в Примере 1, и ген, имеющий последовательность, представленную в SEQ ID N 3, используют для генетического конструирования.
Гены, кодирующие ПФДК, используемые в данном изобретении, включают ген ПФДК кукурузы C4-типа (Matsuoka, М. et al., J. Biol. Chem., (1988) 263: 11080-11083), ген ПФДК риса (Japanese Patent Public Disclosure Hei 7-184657), ген ПФДК Flaveria pringlei (Rosche, E. et al. Plant Mol. Biol., (1994) 26: 763-769), Mesembryanthemum crystallinum (Fisslthaler, B. et al., Planta, (1995) 196: 492-500) и предпочтительным является ген ПФДК кукурузы C4-типа.
Ген ПФДК может экспрессироваться либо в хлоропластах, либо в цитоплазме. Если желательно экспрессировать ген ПФДК в хлоропластах, этот ген может быть соединен с фрагментом ДНК, кодирующим транзитный пептид.
Промоторные последовательности, применимые для экспрессии генов вышеупомянутых ферментов, не ограничиваются какими-либо специфическими промоторными последовательностями, хотя предпочтительны те, которые характерны для фотосинтетического органа. Например, предпочтительны промотор ПФДК кукурузы C4-типа (Glackin et al. , (1990) Proc. Natl. Acad. Sci. USA, 87: 3004-3008), промотор ФЕПК кукурузы C4-типа (Hudspeth, R.L. and Grula, J.M. Plant Mol. Biol. , (1989) 12: 579-589), промотор малой субъединицы Rubisco (Kyozuka, J. et al. Plant Physiol., (1993) 102:991-1000) и промотор светособирающего хлорофилл а/б-связывающего белка (Sa-kamoto, М. et al. Plant Cell Physiol. , (1991) 32:385-393). Особенно предпочтителен промотор ПФДК кукурузы C4-типа. В нижеследующих Примерах в качестве промотора используется SEQ ID N 1.
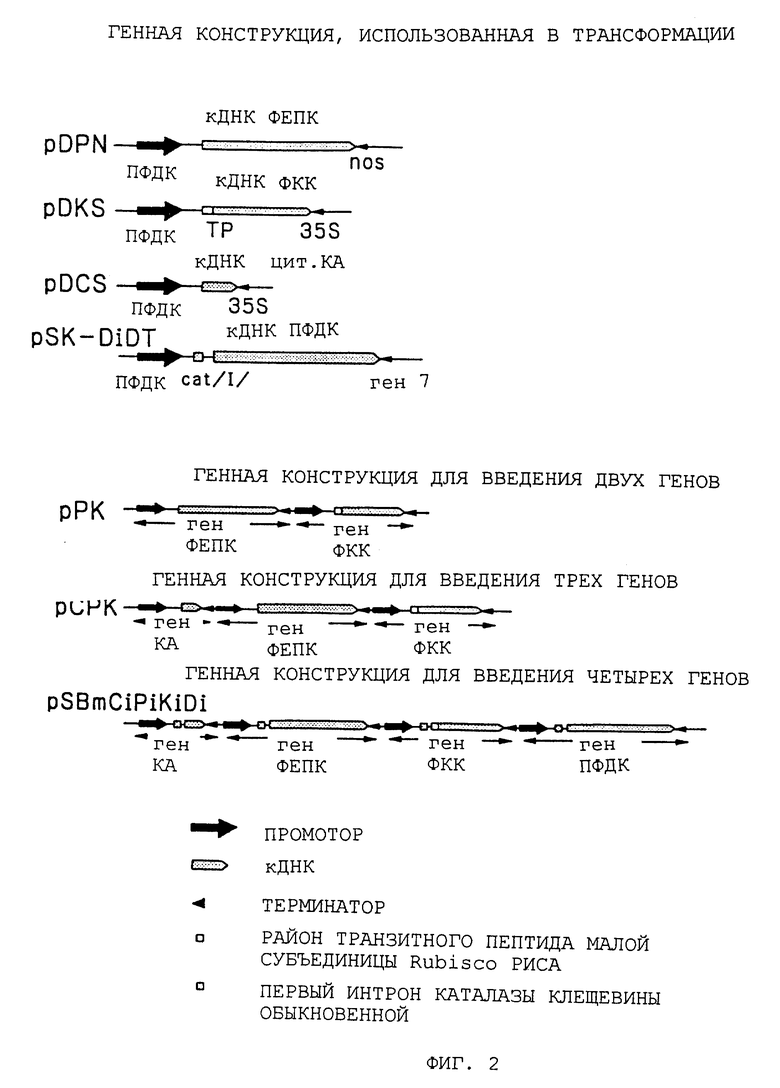
В соответствии с данным изобретением ген, который кодирует каждый из вышеупомянутых ферментов C4-пути, может переноситься на отдельной генной конструкции (вводящей ген конструкции), которую используют для трансформации C3-растения. Однако предпочтительно, чтобы два или более генов находились на одной вводящей ген конструкции, которую вводят в C3-растение для его трансформации. В этом случае нет особого ограничения в отношении порядка генов.
Трансформация клеток C3-растений такой вводящей гены конструкцией, содержащей полученные гены независимо или соединенные вместе, может выполняться в соответствии со стандартным способом введения вводящей гены конструкции в эти клетки или выбранное C3-растение. Основные способы введения генов известны в данной области, например, такие как электропорация, электроинъекция, химическая обработка, например, полиэтиленгликолем (ПЭГ) и генетическая бомбардировка. Среди них предпочтительно введение генов для трансформации клеток C3-растений при помощи способа с использованием Agrobacterium. Способ с Agrobacterium хорошо известен в данной области и способен трансформировать как двудольные (например, Japanese Patent Public Disclosure Hei 4-330234), так и однодольные растения (WO 94/00977). Успешно образованные трансформанты могут быть отобраны по способу, описанному ниже.
Фенотипы трансформантов могут закрепляться любым из подходящих общепринятых способов селекции и, следовательно, введенный ген может переноситься потомству этих трансформантов.
Способ данного изобретения может быть применен к любому из C3-растений и он особенно выгоден для культурных растений, таких как рис, пшеница, ячмень, соя, картофель, табак, рапс и т.п., принимая во внимание, что ожидается повышение продуктивности в расчете на сухой вес вследствие повышения фотосинтетической способности. Предпочтительно, данное изобретение применимо к однодольным растениям, наиболее предпочтительно к рису.
C4-фотосинтетический путь в данном изобретении состоит из трех процессов, т. е. фиксации диоксида углерода ФЕПК, описанной выше, высвобождения диоксида углерода вблизи Rubisco декарбоксилазой и регенерации субстратов для ФЕПК с использованием АТФ.
Способ подтверждения, что C4-фотосинтетический путь функционирует в трансформированном C3-растении, будет подробно описан в Примерах ниже. Вкратце, этот способ предусматривает:
(1) исследование, образуется ли C4-соединение в качестве первичного продукта под действием ФЕПК в трансформантах, когда отделенным листьям трансформированных и контрольных растений дают включать радиоактивный диоксид углерода (14CO2), или исследование, функционирует ли введенный C4-фотосинтетический путь в трансформанте, путем прослеживания соотношения соединений с меченым углеродом по прохождении времени;
(2) исследование, функционирует ли декарбоксилирование C4-соединения под действием ФКК в трансформантах, когда отделенным листьям трансформированных растений дают включать радиоактивный малат ([14C]-малат) и сравнивают относительное количество меченой сахарозы после определенного периода времени между трансформантами и контрольными растениями;
и/или
(3) исследование каких-либо изменений в фотосинтетической способности путем измерения фотосинтетической способности.
Примеры
Для дальнейшей более детальной иллюстрации данного изобретения, но не с целью ограничения, будут представлены следующие ниже примеры.
A. C4-фотосинтетический путь ФКК-типа
Пример 1. Конструирование трансгенов.
(1) Промоторная последовательность
Фрагмент ДНК района промотора ПФДК кукурузы C4-типа получали по способу ПЦР (Mcpherson, M. J. , Quirke, P. and Taylor, GR, ed.: PCR. A practical approach, Oxford Express Press, Oxford NY (1991)) с использованием следующих двух синтетических праймеров, изготовленных на основе известной нуклеотидной последовательности (Glackin, C.A. and Grula, J.W. (1990) Proc. Natl. Acad. Sci. USA 87: 3004-3008): 5' - CTAAAGACATGGAGGTGGAGG-3' (5'-сторона) (SEQ ID N 4) 5'- GTAGCTCGATGGGTGCACG-3' (3'-сторона) (SEQ ID N 5)
Амплификацию проводили с использованием в качестве матрицы геномной ДНК кукурузы, которую получали экстракцией общих нуклеиновых кислот из зеленого листа инбредной кукурузы В73 SDS-фенольным способом с последующей очисткой ультрацентрифугированием в хлориде цезия/бромиде этидия. Полученный таким образом фрагмент ДНК встраивали в клонирующий сайт плазмидного вектора pCR1000 (производимого Invitrogen, USA). Полученную таким образом плазмиду расщепляли SacI и концы затупляли, после чего к ней присоединяли линкер Ncol. После расщепления HindIII полученный ДНК-фрагмент 950 п.о., содержащий промоторный район ПФДК, использовали в конструировании генов. SEQ ID N 1 показывает используемую в нем последовательность ДНК.
(2) Ген ФЕПК
кДНК ФЕПК кукурузы C4-типа выделяли скринингом 20000 клонов из библиотеки кДНК, полученной с использованием мРНК, полученной гуанидин-HCl-методом из зеленых листьев проростков кукурузы (гибридного сорта "harvest queen"), и вектора λZAP (производимого Stratagene, U.S.A.) в соответствии с инструкцией, предоставляемой в прилагаемом руководстве, с использованием следующего синтетического олигонуклеотида в качестве зонда, приготовленного на основе известной нуклеотидной последовательности (Hudspeth, R.L. and Grula, J.W., Plant Mol. Biol. , (1989) 12: 579-589): 5'-GCCATGGCGCGGCGGGAAGCTAAGCACGGAAGCGA-3' SEQ ID N 6) общепринятым способом (Sambrook, J., Fritsch, E.F. and Maniatis, T. Ed. : Molecular Cloning: A Laboratory Manual, (1989) 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1989). Полученный таким образом клон расщепляли XhoI и затем частично расщепляли Ncol. Полученный таким образом ДНК-фрагмент ~ 3 т.п.о. использовали в конструировании генов.
(3) Ген ФКК
кДНК Urochloa panicoides использовали в генной конструкции, в которой район, кодирующий транзитный пептид малой субъединицы Rubisco риса, присоединяли, как описано ранее (Japanese Patent Public Disclosure Hei 8-80197). А именно, λPCK170204 и λPCK100101 лигировали вместе в сайтах KpnI, присутствующих в их вставках. С использованием полученного таким образом фрагмента ДНК в качестве матрицы проводили ПЦР с применением следующих синтетических праймеров:
PCK-f2: 5'-GCTCTAGATCTCTGGCACGTGAATATGGCCCCAACCTCG-3'(SEQ ID N 7);
PCK-r.2: 5'-CAGTGCATGCCGCCGAACAGGCATACAGATTTACACCAG-3' (SEQ ID N 8).
Отдельно, фрагмент ДНК, кодирующий транзитный пептид малой субъединицы Rubisco риса, выделяли по способу ПЦР с применением следующих праймеров, синтезированных на основе последовательности малой субъединицы Rubisco риса (Matsuoka et al. Plant Cell Physiol. 20: 1015-1022 (1988)):
TP-f1: 5'-GGAATTCCATGGTGCATCTCAAGAAGTAC-3' (SEQ ID N9) и
TP-r1: 5'-GCTCTAGACTGCATGCACCTGATCC-3' (SEQ ID N 10)
Матрицей была геномная ДНК риса, которая была получена из зеленых литьев японского сорта риса "Nihonbare" SDS-фенольным способом.
С использованием амплифицированного фрагмента ДНК в качестве матрицы проводили ПЦР опять с применением следующего синтетического праймера:
TP-f2: 5'-GGAATTCCATGGCCCCCTCCGTGATGG-3' (SEQ ID N 11) и вышеупомянутого праймера TP-r1. Затем фрагмент ДНК ~ 2 т.п.о., полученный частичным расщеплением фрагмента кДНК ФКК, амплифицированного ПЦР, рестриктазами XbaI и SphI, лигировали с фрагментом ~ 150 п.о. (представленным SEQ ID N 2), полученным расщеплением фрагмента ДНК последовательности транзитного пептида, амплифицированного второй ПЦР, рестриктазами Ncol и XbaI. Полученный фрагмент ДНК ~ 2,2 т.п.о. использовали для конструирования генов.
(4) Ген КА
Вектор получали встраиванием фрагмента 1,8 т.п.о., содержащего район кДНК КА шпината (который получали расщеплением клона фага λ (λLCA48), сообщенного ранее (Burnell, J.N. et al. (1990) Plant Physiol. 92:33-40) HindIII и KpnI), в сайт HindIII/KpnI) pBluescriptSK (производимой Stratagene, U.S.A.). Из этой кДНК делетировали район, кодирующий транзитный пептид, который участвует в транспорте КА в хлоропласты. Для выполнения этого делетирования концевой аминокислотный остаток в районе транзитного пептида изменяли с серина на метионин таким образом, что эта точковая мутация вводила сайт узнавания Ncol. Мутацию получали с применением следующего синтетического олигонуклеотида:
5'-GGTGGCACAGATAACCATGGATCCAGTTAGCCGACGGTGGC-3' (SEQ ID N 12) Mutan-kTM (производимого Takara Shuzo Co. , Ltd.). Полученную плазмиду, несущую эту мутацию, расщепляли Ncol для делетирования района, кодирующего транзитный пептид. Затем эту плазмиду расщепляли SphI и полученный фрагмент ДНК ~ 700 п. о. использовали в конструировании генов. SEQ ID N 3 показывает последовательность этого фрагмента.
(5) Терминаторные последовательности
Использованными терминаторными последовательностями были фрагмент ДНК района терминатора NOS, полученный расщеплением рВ1121 (Jefferson, R.A. (1987) Plant Mol. Biol. Reptr. 5: 387-405) Sall и EcoRI, и фрагмент ДНК района терминатора 35S, полученный расщеплением pGL2 (Bilang, R. Et al. (1991) Gene 100: 247-250) Sphlu EcoRI.
(6) Конструирование плазмид для введения
Фрагменты промотора, кДНК и терминатора, полученные таким образом, объединяли и лигировали, как показано ниже и затем встраивали между сайтами HindIII-EcoRI pBluescriptIISK - (производимой Stratagene, U.S.A.), для конструирования в результате этого плазмид (плазмид для введения отдельного гена, см. фиг. 2).
Промотор ПФДК::кДНК ФЕПК::терминатор NOS (pDPN).
Промотор ПФДК::кДНК КА::терминатор 35S (pDCS).
Промотор ПФДК::кДНК ФКК::терминатор 35S (pDKS).
Далее pDKS расщепляли ClaI, затупляли и затем расщепляли XbaI. Полученный таким образом фрагмент ДНК встраивали в pDPN в сайтах, расщепляемых SmaI и XbaI, с получением таким образом плазмиды pPK, несущей эти два гена, т. е. ФЕПК и ФКК (см. фиг. 2). Затем pDCS расщепляли SmaI и присоединяли к ней линкер HindIII. После расщепления HindIII полученный фрагмент ДНК встраивали в сайт HindIII pDPN с получением в результате плазмиды pCP. Кроме того, pDKS расщепляли ClaI, затупляли и затем расщепляли XbaI. Полученный таким образом фрагмент ДНК встраивали в pCP в сайте, расщепляемом ClaI и XbaI, с получением в результате плазмиды pCPK, несущей три гена, т.е. КА, ФЕПК и ФКК (см. Фиг. 2). Затем pPK расщепляли XbaI и затем частично расщепляли HindIII с получением в результате фрагмента ДНК ~ 7,5 т.п.о., тогда как pCPK расщепляли HindIII с получением другого фрагмента ДНК ~ 9,5 т.п.о. Эти фрагменты ДНК встраивали в pSBll (Komari, T. et al. Plant J. (1996) 10: 165-174), соответственно, в сайтах, расщепляемых XbaI и HindIII, с получением сконструированных супербинарных промежуточных плазмид pSPK и pSCHK.
Эти векторы, каждый, вводили в штамм Escherichia coli LE392. Затем проводили введение в Agrobacterium и гомологичную рекомбинацию (Komari, Т. Et al. Plant J. (1996) 10: 165-174) посредством трехродительского скрещивания с Agrobacterium LB4404/pSB4 и E.coli HB101/pRK2013 с получением плазмид pSB4PK и pSB4CPK.
(7) Конструирование векторов, несущих КА, ФЕПК, ФКК и/или ПФДК
Плазмидный вектор pCR1000 (производимый Invitrogen, U.S.A.), имеющий промоторный район ПФДК, который был получен по способу ПЦР, как описано выше, расщепляли HindIII и EcoRI и затем встраивали между сайтами HindIII и EcoRI pBluescript IISK-(производимой Stratagene, U.S.A.), из которой был делетирован сайт Sac1. Затем полученный продукт расщепляли Sac1 и концы затупляли. После этого фрагмент ~ 200 п.о., содержащий первый интрон каталазы Ricinus communis, (который был получен расщеплением plG221 (Ohta et al.: Construction and expression in tobacco of betagluronidase (GUS) reporter gene containing an intron within the coding sequence. Plant Cell Physiol. 31: 805-813 (1990) рестриктазами BamHI и Sall с последующим затуплением концов), встраивали в него с получением плазмиды pSK-Di, содержащей промотор ПФДК (catI). Затем эту плазмиду расщепляли Nde1 и концы затупляли с получением другой плазмиды pSK-Di2, имеющей вставленный в нее NcoI-линкер.
кДНК ПФДК кукурузы C4-типа выделяли скринингом библиотеки кДНК, полученной общепринятым способом (Sambrook et al., ibid.) с использованием вектора λZAP (производимого Stratagene, U.S.A.) с применением следующего синтетического олигонуклеотида; 5'-TAGCTCGATGGGTTGCACGATCATATGGAGCAAGG-3' (SEQ ID N 13), приготовленного на основе известной нуклеотидной последовательности (Matsuoka, М. , Ozeki, Y., Yamamoto, N., Hirano, H., Kano-Murakami, Y. and Tanaka, Y.: Primary Structure of maize pyruvate, orthophosphate dikinase as deduced from cDNA sequence, J. Biol. Chem. 163: 11080-11083 (1990)). Далее, выделенную кДНК ПФДК кукурузы подвергали амплификации по способу ПЦР (Mcpherson, M. J. et al., ibid) с применением следующих синтетических праймеров, приготовленных на основе известной последовательности (Sheen, J.: Molecular mechanisms underlying the differential expression of Maize Pyruvate, Opthophosphate dikinase genes. Plant Cell 3:225-245 (1991)):
5'-TTTCATATGGCGCCCGTTCAATGTGCGC GTTCGCAGAGGGTGGTTCCACTTCGGCAA-3' (5'-сторона) (SEQ ID N14) и
5'-GTACTCCTCCACCCACTGCA-3' (3'-сторона) (SEQ ID N15); с получением в результате фрагмента ДНК ~ 250 п.о. Этот фрагмент расщепляли NdeI и SacI и вводили вместо района между сайтами NdeI и SacI вышеупомянутой кДНК ПФДК. Затем ее расщепляли NdeI и SacI и полученный таким образом фрагмент ДНК ~ 2,9 т.п.о. использовали в качестве кДНК ПФДК в конструировании генов.
В качестве терминирующего района использовали терминатор гена 7, содержащийся в плазмиде pPGA643A (Gynheung AN, Paul R. Ebert, Amitava Minra and Sam B. HA: binary vectors, Plant Molecular Biology Manual A3: 1-19 (1988)). Терминатор гена 7 получали расщеплением pPGA643A (Gynheung AN et al. , ibid. ) ClaI и KpnI. Этот терминатор встраивали между сайтами ClaI и KpnI pBluescript IISK- ((производимой Stratagene, U. S.A.) с получением плазмиды. Эту плазмиду расщепляли KpnI, концы затупляли и присоединяли к ней XbaI-линкер. После расщепления ClaI и XbaI полученный фрагмент ДНК использовали в конструировании генов.
кДНК ПФДК и терминатор гена 7 встраивали между сайтами NdeI и XbaI pSK-Di с получением плазмиды pSK-DiDT, содержащей ген ПФДК.
Фрагмент ДНК ~ 3,4 т.п.о., который получали расщеплением вышеупомянутой плазмиды pDPN XbaI и затем частичным расщеплением NcoI, встраивали между сайтами NcoI и XbaI pSK-Di2 с получением плазмиды pSK-DiPN, содержащей ген ФЕПК.
Фрагмент ДНК ~ 2,4 т.п.о., который был получен расщеплением вышеупомянутой pDKS NcoI и XbaI, встраивали между сайтами NcoI и XbaI плазмиды pSK-Di2 с получением плазмиды pSK-DiKS, содержащей ген ФКК.
Фрагмент ДНК ~ 1 т.п.о., который был получен расщеплением вышеупомянутой плазмиды pDKS NcoI и XbaI, встраивали между сайтами NcoI и XbaI pSK-Di2 с получением плазмиды pSK-DiCS, содержащей ген КА.
Линкер SmaI встраивали в сайт Xho1 pSK-DiPN и расщепляли SmaI. Полученный таким образом фрагмент ДНК ~ 4,5 т.п.о. встраивали в pSK-DiCS в сайте, который был расщеплен PstI и имел затупленные концы, с получением плазмиды pSK-CiPi. После делеции сайта XbaI pSK-DiDT к сайту XhoI этой плазмиды присоединяли линкер XbaI и расщепляли XbaI и NotI. Полученный таким образом фрагмент ДНК ~ 4,8 т. п. о. встраивали между сайтами XbaI и NotI плазмиды pSK-CiPi с получением плазмиды pSK-CiPiDi. Линкер NotI присоединяли к сайту XhoI pSK- CiPiDi и расщепляли рестриктазой NotI с получением фрагмента ДНК ~ 12 т.п.о. pSB11 (Komari, T. Et al. Plant J. 10:165-174 (1996)) расщепляли HindIII и EcoRI и концы затупляли. После этого в нее добавляли линкер NotI. В этот сайт NotI встраивали вышеупомянутый фрагмент ДНК ~ 12 т.п.о. с получением плазмиды pSBmCiPiDi. Линкер XbaI присоединяли к сайту XhoI pSK-DiKS и расщепляли рестриктазой XbaI. Полученный таким образом фрагмент ДНК ~ 3,3 т. п.о. встраивали в сайт XbaI pSBmnCiPiDi с получением плазмиды pSBmCiPiKiDi.
Затем проводили введение в Agrobacterium и гомологичную рекомбинацию (Komari, T., ibid.) посредством трехродительского скрещивания между штаммом E. coli DH5α, несущим плазмиду pSBmCiPiKiDi, Agrobacterium LBA4404, несущим pSB4, и штаммом E. coli НВ101, несущим pRK2013, с получением в результате плазмиды pSB4CiPiKiDi.
Пример 2 Конструирование трансформантов
Во всем исследовании трансформации риса использовали Японский рис (культивар "Tsukinohikari").
Трансформанты риса, имеющие введенные в них pDPN, PDKS и pDCS, конструировали по способу электропорации, описанному ранее (Japanese Patent Public Disclosure Hei 8-80197).
Трансформанты риса, имеющие введенные в них pSB4PK, pSB4CPK и pSB4CiPiDiKi, конструировали по способу с Agrobacterium, описанному ранее (Hiel, Y. et al. (1994) Plant J. 6:271-282).
Эти трансформанты выращивали в оранжерее с кондиционированным воздухом (период дневного света: 16 часов, днем: 28oC, ночью: 23oC).
Пример 3. Обнаружение ферментных белков и измерение ферментативных активностей
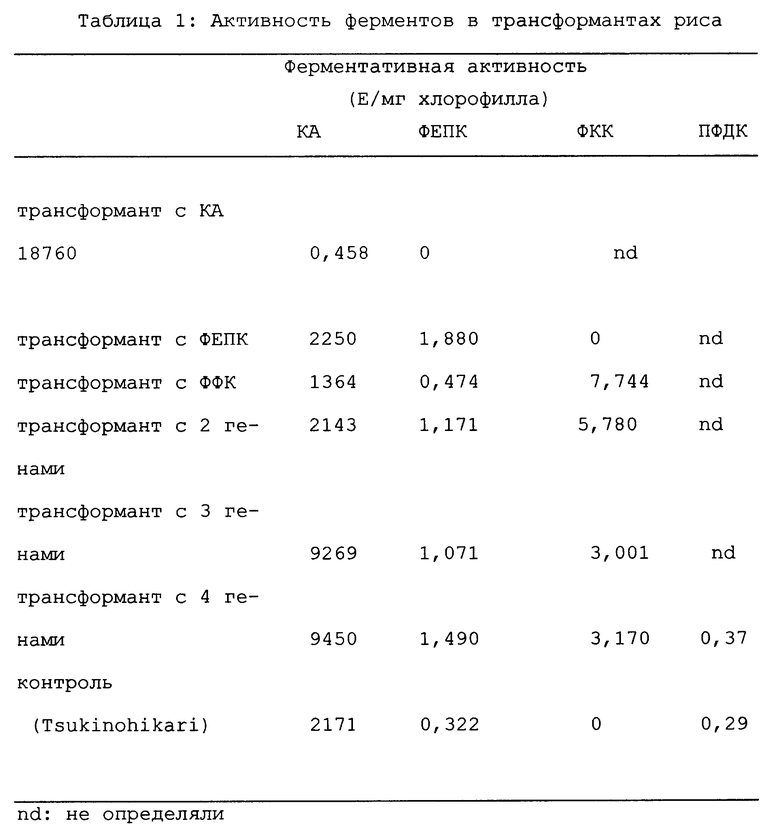
Приблизительно 0,1 г зеленых листьев трансформантов или контрольного риса ("Tsukinohikari") гомогенизировали в 1 мл ледяного буфера для экстракции (50 мМ HEPES-KOH pH 7,0, 10 мМ хлорид магния, 2 мМ хлорид марганца, 1 мМ пируват натрия, 1 мМ фосфорная кислота, 1 мМ ЭДТА, 0,1% 2-меркаптоэтанол, 20% глиферин, 1 мМ фенилметилсульфонилфторид, 1 мМ бензамидин, 1 мМ 6-амино-н-капроновая кислота, 0,2% (м/м изоаскорбиновая кислота и 2% (м/об) polyclar AT). Гомогенат центрифугировали при 15000 х q в течение 20 минут при 4oC. Затем полученный супернатант обессоливали пропусканием через колонку NAP5TM (производимую Pharmacia, Sweden), которая была уравновешена при комнатной температуре колоночным буфером (50 мМ HEPES-KOH pH 7,0, 10 мМ хлорид магния, 2 мМ хлорид марганца, 1 мМ ЭДТА, 0,1% 2-меркаптоэтанол и 2% глицерин) с получением неочищенного экстракта. Содержание хлорофилла в гомогенате определяли по методу, сообщенному ранее (Wintermans and deMots (1965) Biochem. Biophys. Acta 109: 448-453), тогда как содержание белка в неочищенном экстракте определяли с использованием набора Protein Assay KitTM (производимого BioRad, U.S.A.).
Экспрессию ферментов в трансформантах детектировали Вестерн-блоттингом следующим образом. Полученный, как описано выше, неочищенный экстракт подвергали электрофорезу в ПА-АГ-SDS таким образом, чтобы довести концентрацию белка до одного и того же уровня. Разделенные белки в геле электрически переносили на нитроцеллюлозную мембрану (производимую Schleicher and Schull, Germany) и экспрессию каждого из этих белков детектировали с применением кроличьей антисыворотки против белка ФЕПК кукурузы, белка ФКК Urochloa panicoides, белка КА шпината или белка ПФДК кукурузы, конъюгированной со щелочной фосфатазой козьей антисыворотки против кроличьего IgG (производимой Organon Teknika, U. S. A. ) и набора АР Immun-Blot Assay KITT (производимого BioRad, U.S.A.).
Активность ФЕПК определяли измерением скорости уменьшения поглощения НАДН при 340 нм с применением 1 мл реакционной смеси, содержащей 25 мМ HEPES-KOH (pH 8,0) 5 мМ сульфат магния, 4 мМ дитиотреитол, 5 мМ бикарбонат калия, 0,25 мМ НАДН, 1 мМ глюкозо-6-фосфат, 5 мМ фосфоенолпируват, 1 E малатдегидрогеназы (производимой Boeringer Mannheim, (Germany) и 25 мкл неочищенного экстракта.
Активность ФКК определяли измерением скорости уменьшения поглощения щавелевоуксусной кислоты при 280 нм с применением 1 мл реакционной смеси, содержащей 25 мМ HEPES-KOH (pH 8,0), 4 мМ дитиотректол, 0,2 мМ щавелевоуксусную кислоту, 1 E пируваткиназы (производимой Boeringer Mannheim, Germany), 0,2 мМ АТФ и 50 мкл неочищенного экстракта.
Активность КА определяли добавлением 0,5 мл ледяной насыщенной диоксидом углерода воды к 0,3 мл 50 мМ HEPES-KOH-буфера (pH 8,0), окрашенного Бромтимоловым синим, и 10 мкл неочищенного экстракта и измерением времени, необходимого для исчезновения окрашивания реакционной смеси на льду. Активность рассчитывали в соответствии с методом, сообщенным ранее (Burnell, J.N. and Hatch, M.D. (1988) Plant Physiol. 86:1252-1256).
Активность ПФДК определяли измерением скорости уменьшения поглощения НАДН при 340 нм с применением 1 мл реакционной смеси, содержащей 25 мМ HEPES-KOH (pH 8,0), 10 мМ дитиотреитол, 10 мМ бикарбонат калия, 2,5 мМ вторичный кислый фосфат натрия, 1 мМ АТФ, 1 мМ глюкозо-6-фосфат, 5 мМ пируват натрия, 0,2 мМ НАДН, 2 E малатдегидрогеназы, 2 E ФЕПК (производимой Wako Pure Chemical Industries, Japan) и 200 мкл неочищенного экстракта.
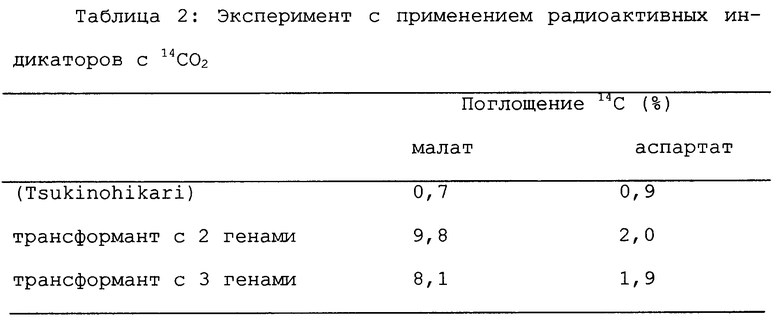
Пример 4. Эксперимент с применением радиоактивных индикаторов с 14CO2
Концы листьев (приблизительно 5 см) трансформанта и контроля ("Tsukinohikari"), которые росли в кондиционированной оранжерее, отделяли и каждый срезанный конец покрывали абсорбирующей ватой, пропитанной в воде. Эти пробы устанавливали в ассимиляционную камеру (емкостью 120 мл или 50 мл), изготовленную авторами данного изобретения. После пропускания атмосферного воздуха при скорости тока ~ 5 л/мин в течение 30 минут при освещении ~ 27000 люкс в эту замкнутую систему вводили радиоактивный газообразный диоксид углерода, который генерировали смешиванием 100-180 мкл 60% перхлорной кислоты с раствором NaH14CO3 50-70 мкКи (производимого Amersham, England) в газонепроницаемом шприце. После кратковременного введения (пульс-мечение-5 секунд) листья замораживали в жидком азоте для прекращения любых биологических активностей и затем помещали в 80% горячий этанол на ~ 30 минут для экстрагирования растворимого материала. После пульса 5 секунд в систему вводили атмосферный воздух. Через 10, 30 и 90 секунд после этого из камеры брали пробы листьев и погружали в жидкий азот для прекращения любых биологических активностей. Затем растворимые вещества экстрагировали в 80% горячем этаноле. Полученные таким образом экстракты выпаривали (каждый) на испарителе и подвергали двумерной тонкослойной хроматографии на пластинке Funaseru SF Cellulose Thin Layer PlateTМ (производимой Funakoshi Japan, 20 см x 20 см).
Проявление выполняли при комнатной температуре с применением смеси фенол-вода-ледяная уксусная кислота - 0,5 М ЭДТА (47:84:5,5:1,14, об/об) в качестве первичного проявляющего растворителя и изоволюметрической смеси раствора А (н-бутанол: вода 74:6 5, об/об) в качестве второго проявляющего растворителя. После завершения проявления пластинку сушили с последующей авторадиографией с системой Bioimage Analyzer Bas 1000 (Fuji Film Japan) для определения относительного количества каждого пятна. Таким образом определяли соотношение вещества, меченного радиоактивным изотопом.
Пример 5. Эксперимент с применением радиоактивных индикаторов с [14C]малатом
Листья трансформанта риса и контроля ("Tsukinohikari"), которые росли в кондиционированной оранжерее, отделяли, помещали в 10 мл фосфатный буфер (pH 6,4) и затем освещали при 27000 люкс в течение 1 часа. Затем эти листья помещали в 100 мкл раствора, к которому были добавлены 5 мкл 1 мкКи [14C]-малата (производимого Amersham, England). После определенного периода времени брали пробы листьев. Затем часть, погруженную в раствор, удаляли и погружали в кипящий 80% этанол для прекращения любых биологических активностей. Растворимым веществам давали элюироваться из них при кипячении в течение 30 минут. Элюат выпаривали на испарителе. Вещество, меченное радиоактивным изотопом, разделяли двумерной тонкослойной хроматографией с последующей авторадиографией с системой Bioimage Analyzer Bas 1000 (Fuji Film Japan) для определения радиоактивности каждого пятна. Таким образом определяли относительное количество вещества, меченного радиоактивным изотопом.
Пример 6. Измерение фотосинтетической активности
Трансформанты риса, имеющие введенные в них три гена, и контроль ("Tsukinohika-ri"), которые были выращены в кондиционированной оранжерее, переносили для роста в камеру для выращивания (период дневного света: 12 часов, освещение: ~ 35000 люкс, 25oС) для акклиматизации. Затем фотосинтетические активности полностью выросших листьев, не обнаруживающих симптомов старения, измеряли в системе для измерения фотосинтеза (LI-6200, производимой LI-COR, U.S.A.).
Пример 7. Получение трансгенного риса с C4-путем ФКК-типа и результаты определений
Были получены отдельные трансформанты для каждой генной конструкции, полученной, как описано выше, и экспрессию трансгенов исследовали при помощи Вестерн-блоттинга. Среди 19 трансформантов риса, имеющих введенную в них генную конструкцию pDPN (трансформант с ФЕПК), экспрессия белка ФЕПК была подтверждена в 15 трансформантах. Среди трансформантов (в количестве 31), имеющих введенную в них генную конструкцию pDKS (трансформант с ФКК), экспрессия белка ФКК была подтверждена в 20 трансформантах. Среди трансформантов (в количестве 41), имеющих введенную в них генную конструкцию pDCS (трансформант с КА), экспрессия белка КА была подтверждена в 3 трансформантах. Среди трансформантов (в количестве 21), имеющих введенную в них генную конструкцию pSB4PK (трансформант с двумя генами), экспрессия двух белков, т. е. ФЕПК и ФКК, была подтверждена в 12 трансформантах. Среди 40 трансформантов, имеющих введенную в них генную конструкцию pSB4CPK (трансформант с тремя генами), экспрессия трех белков, т.е. КА, ФЕПК и ФКК, была подтверждена в 15 трансформантах. Среди 72 трансформантов, имеющих введенную в них
генную конструкцию pSB4CiPiKiDi (трансформант с четырьмя генами), экспрессия четырех белков, т.е. КА, ФЕПК, ФКК и ПФДК, была подтверждена в 22 трансформантах.
Были получены R1-потомства из некоторых из этих трансформантов риса, обнаруживающие относительно высокие уровни экспрессии введенных в них ферментов. Затем испытывали активность каждого фермента в неочищенных экстрактах зеленых листьев. Эти результаты показаны в Таблице 1. Как ясно видно из Таблицы 1, неочищенные экстракты трансформантов генерации RI показали более высокие активности введенных в них ферментов, чем экстракты в контрольном рисе ("Tsukinohikari"). Эти факты указывают на то, что ферменты, экспрессируемые введенными генами в трансформантах, проявляют их ферментативные активности. Генные конструкции для введения ФКК, используемые в этом Примере, были химерными генами, к которым был присоединен район, кодирующий транзитный пептид малой субъединицы Rubisco риса, сходными с химерными генами, сообщенными ранее (Japanese Patent Public Disclosure Hei 8-80197). Таким образом, белок ФКК мог быть локализован в хлоропластах благодаря действию транзитного (транспортного) пептида.
14CO2 подавали к отрезкам листьев растений генерации R1 трансформантов, имеющих введенные в них два и три гена и контроля (Tsukinohikari). Спустя 5 секунд биологические активности в тканях прекращали и определяли относительное количество меченых C4-соединений. В результате было обнаружено, что содержания меченого малата и меченой аспарагиновой кислоты в трансформантах были соответственно приблизительно в 10 раз и приблизительно в 2 раза выше, чем в контрольной пробе (см. Таблицу 2). Эти факты свидетельствуют о том, что введенная ФЕПК функционировала в тканях зеленых листьев трансформантов таким образом, что в них осуществлялся процесс первичной фиксации диоксида углерода по C4-фотосинтетическому пути.
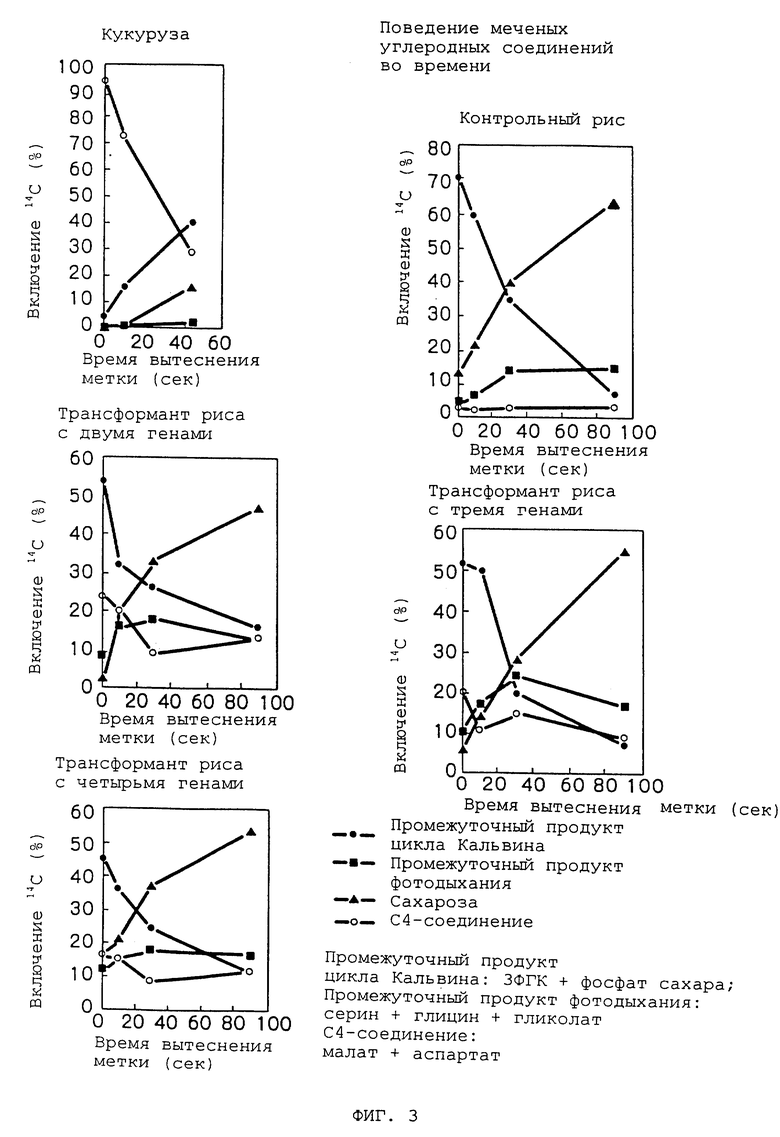
Также 14CO2 подавали в течение 5 секунд к отрезкам листьев генераций R1 трансформантов риса, имеющих введенные в них два, три или четыре гена, а также контрольного риса (Tsukinohikari) и контрольной кукурузы. Затем поведение меченых C4-соединений прослеживали методом "чейза" (слежения за меткой) во времени. В трансформантах риса меченые C4-соединения снижались (по метке) сходно с вариантом кукурузы (т.е. C4-растения). В противоположность этому, меченые C4-соединения в контрольном рисе не обнаруживали какого-либо значимого изменения (см. Фиг. 3). Эти факты указывают на то, что в трансформантах риса C4-соединения, образованные фиксацией диоксида углерода введенной ФЕПК, сразу метаболизировались в другие вещества, подобно феноменам, наблюдаемым в тканях зеленых листьев C4-растений.
[14C] -малат предоставляли отрезкам листьев R1-потомств трансформантов риса, имеющих введенные в них два и три гена, и контрольного риса (Tsukinohikari). После 15 минут биологические активности в тканях прекращали и определяли соотношение меченых соединений. В результате было обнаружено, что относительные количества меченой сахарозы в трансформантах были приблизительно в 3 раза выше, чем эти количества в контрольной пробе (см. Таблицу 3). Этот факт указывает на то, что введенная ФКК функционирует в тканях зеленых листьев трансформантов риса таким образом, что диоксид углерода переходит от C4-соединения в C4-фотосинтетическом пути к циклу Кальвина-Бенсона.
Фотосинтетические активности R1-потомств трансформанта риса, имеющего введенные в него три гена, и контрольного риса (Tsukinohikari) измеряли с вариированием концентрации диоксида углерода, подаваемого к листьям. Скорость фотосинтеза наносили на график в зависимости от внутриклеточной концентрации диоксида углерода (см. Фиг. 4). В результате, внутриклеточная концентрация диоксида углерода трансформанта в точке, когда наблюдаемая фотосинтетическая активность, по-видимому, исчезает (т.е. пересечение оси x линии на фиг. 4), была ниже, чем эта концентрация в контрольном рисе. Этот факт означает, что компенсационная точка CO2 трансформанта риса ниже, чем компенсационная точка контроля. Как правило, C4-растения обнаруживают компенсационные точки, более низкие, чем компенсационные точки C3-растений. Таким образом, можно сделать вывод, что сконструированные здесь трансформанты риса, сходны по их фотосинтетическим характеристикам с C4-растениями, в сравнении с контрольным рисом.
Описанные выше результаты свидетельствуют о том, что трансформанты риса, имеющие введенные в них два и три гена, содержат активные формы ферментов, относящихся к C4-фотосинтезу, которые представляют собой продукты экспрессии введенных генов, и, кроме того, что C4-фотосинтетический путь функционирует в этих растениях с усилением фотосинтетической способности. Таким образом, данное изобретение делает возможным функционирование C4-фотосинтетического пути ФКК-типа в C3-растениях с изменением тем самым фотосинтетической активности.
Выводы изобретения
Согласно данному изобретению, циклическая реакция, сходная с C4-фотосинтетическим путем C4-растений, может запускаться в мезофилльных клетках C3-растений таким образом, что C3-растения приобретают функцию повышения концентрации диоксида углерода в хлоропластах и другую функцию элиминации фотоингибирования, вызываемой потреблением АТФ. Ожидается, что вследствие повышенной фотосинтетической активности растения, приобретающие эти функции, будут иметь повышенную продуктивность на сухой вес, улучшенную засухоустойчивость, улучшенную устойчивость к повышенным температурам, улучшенную устойчивость к высокой интенсивности света и повышенную фотосинтетическую активность в условиях низких концентраций диоксида углерода.












Способ трансформации растения с помощью химерного гена предназначен для использования в селекции растений. Введение в растение гена, кодирующего фосфоенолпируваткарбоксилазу (ФЕПК), и гена, кодирующего фосфоенолпируваткарбоксикиназу (ФКК), который соединен с фрагментом ДНК, кодирующим транзитный пептид, обеспечивает получение растений с высокими агрохимическими показателями в условиях засухи, повышенной температуры и интенсивной освещенности, что достигается за счет придания растениям С4 фотосинтетического пути. 2 с. и 4 з.п.ф-лы, 4 ил., 3 табл.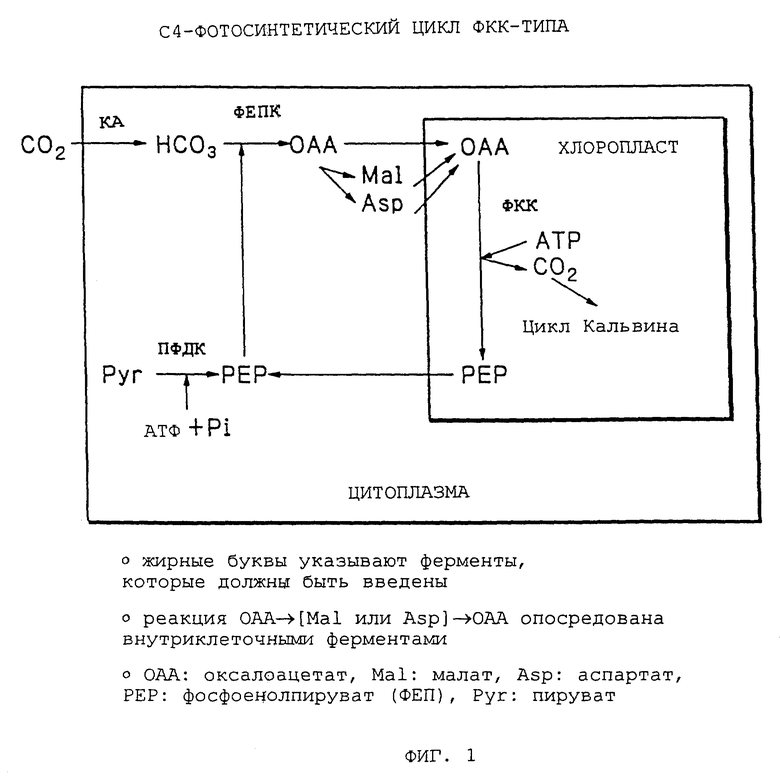
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |
| ГРИНН Н.Биология /Пер.с англ | |||
| - М.: Мир, 1993, т.1. | |||

