Изобретение относится в общем к молекулярной биологии растений и, более конкретно, к новому классу глифосат толерантных 5-енолпирувилшикимат-3-фосфат синтаз.
Последние достижения генетической инженерии обеспечили необходимые средства для трансформации растений для введения чужеродных генов. Теперь возможно получать растения, которые обладают уникальными характеристиками агрономической важности. Несомненно одним таким выгодным путем является более эффективный, совместный с окружающей средой контроль сорняков через толерантность к гербицидам. Гирбицидтолерантные растения могут уменьшить необходимость в обработке земли для контроля сорняков, в результате чего эффективно снижается эрозия почвы.
Гербицидом, который является объектом многих исследований в этом отношении, является N-фосфонометилглицин, обычно называемый глифосат. Глифосат ингибирует путь шикимовой кислоты, который приводит к биосинтезу ароматических соединений, включая аминокислоты, гормоны растений и витамины. Особенно глифосат ограничивает конверсию фосфоенолпировиноградной кислоты (РЕР) и 3-фосфошикимовой кислоты в 5-енолпирувил-3-фосфошикимовую кислоту при ингибировании фермента 5-енол-пирувилшикимат-3-фосфат синтазы (здесь далее упоминается как EPSP синтаза или EPSPS).
Уже было показано, что глифосат толерантные растения могут быть получены путем внесения в геном растения способности продуцировать более высокий уровень EPSP синтазы в хлоропласте клетки (Shah et al. ., 1986), этот фермент является предпочтительно толерантным к глифосату (Kishore et al., 1988). Были выделены варианты дикого типа EPSPS фермента, которые являются глифосат толерантными в результате изменений в кодирующей аминокислотной последовательности EPSPS (Kishore and Shah 1988; Schulz et al., 1984; Sost et al. 1984; Kishore et al., 1986). Эти варианты обычно имеют более высокий Ki для глифосата, чем фермент EPSPS дикого типа, который придает фенотипу толерантность к глифосату, но эти варианты также характеризуются высоким Km для PEP, который делает фермент кинетически менее эффективным (Kishore and Shah 1988; Sost et al., 1984; Schulz et al., 1984; Kishore et al., 1986; Sost and Amrhein, 1990). Например, кажущийся Km для PEP и кажущийся Ki для глифосата для нативного EPSPS из E.coli равны 10 мкМ и 0,5 мкМ, тогда как для толерантного к глифосату изолята, имеющего единственное аминокислотное замещение аланина на глицин в положении 96, эти величины равны 220 мкМ и 4,0 мМ соответственно. С помощью мутагенеза был сконструирован ряд глифосаттолерантных вариантов EPSPS генов растений. Снова глифосат толерантный EPSPS снижается благодаря увеличению Km для PEP и незначительного снижения Vmax нативного фермента растения (Kishore and Shah, 1988), в результате чего снижается каталитическая эффективность (Vmax/Km) фермента. Поскольку кинетические константы вариантов ферментов ухудшились в отношении PEP, было предположено, что высокие уровни сверхпродукции варианта фермента, 40-80-кратные, будут требоваться для поддержания нормальной каталитической активности в растениях в присутствии глифосата (Kishire et al., 1988).
Хотя было показано, что такие варианты EPSP-синтаз являются полезными для получения трансгенных растений, толерантных к глифосату, было бы крайне интересно получить EPSP-синтазу, которая является высоко толерантной к глифосату, в то время как еще кинетически эффективной, чтобы снизилось то количество толерантных к глифосату EPSPS, которое должно быть продуцировано для сохранения нормальной каталитической активности, или чтобы была получена улучшенная толерантность с тем же самым уровнем экспрессии.
Предшествующие исследования показали, что EPSPS-ферменты из различных источников сильно меняются в отношении степени их чувствительности к ингибированию глифосата. Изучение активности растительного и бактериального EPSPS-фермента как функции концентрации глифосата показало, что имеется очень широкий интервал степени чувствительности к глифосату. Степень чувствительности не показывает корреляции с какими-либо генами или испытанными видами (Schulz et al., 1985). Уже сообщалось о нечувствительности к ингибированию глифосата активности EPSPS из Pseudomonas sp.PG2982, но не проводились детали исследования (Fitzgibbon, 1988). В общем, хотя известна такая природная толерантность, не сообщалось предположений о кинематическом превосходстве EPSPS- ферментов природного бактериального происхождения, толерантных к глифосату, по сравнению с мутированными EPSPS-ферментами, не были охарактеризованы какие-либо гены. Подобным образом, не имеется сообщений об экспрессии природно толерантных к глифосату EPSPS-ферментов в растениях для придания толерантности к глифосату.
Имеется молекула ДНК, представляющая собой ДНК, кодирующую кинетически эффективную толерантную к глифосату EPSP-синтазу. EPSP-синтазы настоящего изобретения снижают количество сверхпродукции EPSPS-фермента в трансгенном растении, необходимое для фермента, чтобы поддерживать каталитическую активность при придании к тому же толерантности к глифосату. Эта и другие EPSP-синтазы, описанные здесь, представляют собой новый класс EPSPS-ферментов, упоминаемых здесь далее как класс II EPSPS-ферментов. Класс II EPSPS-ферментов обладает малой гомологией к известным бактериальным или растительным EPSPS-ферментам и показывает толерантность к глифосату при сохранении подходящих интервалов Km(PEP). Подходящие интервалы Km (PEP) для EPSPS-ферментов настоящего изобретения находятся между 1-150 μM при более предпочтительном интервале между 1-35 μM и наиболее предпочтительном интервале между 2-25 μM. Эти кинематические константы определены в опытных условиях, определенных здесь далее. Vmax фермента должна быть предпочтительно по крайней мере 15% неингибированного фермента растения, а более предпочтительно более 25%. EPSPS настоящего изобретения предпочтительно имеет Ki для глифосатного интервала между 25-10000 μM. Отношение Ki/Km может быть между 3-500, а более предпочтительно между 6-250. Vmax предпочтительно может быть в интервале 2-100 единиц/кг/ μмоль/мин.мг при 25oC/ и Km для шикимат-3-фосфата может быть предпочтительно в интервале от 0,1 до 50 μМ.
Гены, кодирующие ферменты класса II EPSPS, были выделены из трех (3) различных бактерий: Agrobacterium tumefaciens sp. штамм CP4, Achromobacter sp. штамм LBAA и Pseudomonas sp. штамм PG2982. Гены LBAA и PG2982 класса II EPSPS были определены для идентификации, а белки, кодируемые этими двумя генами, очень похожи на CP4 белок и имеют примерно 84% аминокислот, идентичных им. Класс II EPSPS-ферментов может быть легко разграничен от класса I EPSPS по их неспособности к взаимодействию с поликлональными антителами, приготовленными из ферментов класса I EPSPS в условиях, когда другие ферменты класса I EPSPS будут легко реагировать с антителами класса I.
Другие ферменты класса II EPSPS могут быть легко выделены и идентифицированы при использовании зонда нуклеиновой кислоты из одного из генов класса II EPSPS, описанного здесь с использованием стандартной методики гибридизации. Такой зонд из CP4 штамма был приготовлен и использован для выделения генов класса II EPSPS из штаммов LBAA и PG2982. Эти гены также могут быть приспособлены для повышения экспрессии в растениях по известной методологии. Такой зонд также был использован для идентификации гомологичных генов в бактериях, изолированных de novo из почвы.
Класса II EPSPS-ферменты предпочтительно сливают с хлоропластным транзитным пептидом (СТР), имея в виду получить белок в хлоропластах растений, в которые он может быть введен. Химерные гены, кодирующие этот СТР-класс II EPSPS слитый белок, могут быть приготовлены с подходящим промотором и 3'-сайтом полиаденилирования для введения в желаемое растение по стандартным методикам.
Следовательно, в одном аспекте, настоящее изобретение обеспечивает новый класс EPSP-синтаз, который показывает низкий Km для фосфоенолпирувата (PEP), высокое отношение Vmax/Km и высокое Ki для глифосата, таких, что при введении в растение, растение становится толерантным к глифосату, так что сохраняются по существу в нормальном состоянии каталитическая активность фермента и метаболизм растения. При настоящем обсуждении высокая эффективность EPSPS относится к его эффективности в присутствии глифосата.
В другом аспекте настоящего изобретения обсуждается двунитевая молекула ДНК, представляющая собой ДНК, кодирующую фермент EPSPS класса II. Последовательность ДНК ферментов EPSPS класса II раскрыта из трех источников: штамм Agrobacterium sp. , обозначенный CP4, штамм LBAA Achromobacter sp. и штамм PG2982 Pseudomonas sp.
В дополнительном аспекте настоящего изобретения представлен нуклеиново-кислотный зонд из гена EPSPS класса II, пригодный для применения в скрининге генов EPSPS класса II в других источниках посредством анализа на способность последовательности ДНК из другого источника гибридизоваться с зондом.
В еще одном аспекте настоящего изобретения описаны трансгенные растения и трансформированные клетки растений, которые обеспечивают толерантность к глифосату за счет введения гена EPSPS класса II в геном растения. Генетически трансформированные растения, толерантные к гербициду глифосату, могут быть получены способом, включающим стадии:
а) введение в геном клетки растения рекомбинантной двунитевой молекулы ДНК, включающей:
i) промотор, функционирующий в растительной клетке, обеспечивающий продукцию последовательности РНК;
ii) структурную последовательность ДНК, обеспечивающую продукцию последовательности РНК, которая кодирует слитый полипептид, включающий аминоконцевой транзитный пептид хлоропласта и EPSPS-фермент класса II;
iii) 3'-нетранслируемую последовательность ДНК, функционирующую в растительной клетке, обеспечивающую добавление тяжа поли-A к 3'-концу последовательности РНК;
причем промотор является гетерологичным по отношению к структурной последовательности ДНК и адаптирован таким образом, чтобы обеспечить достаточную экпрессию слитого пептида для повышения толерантности к глифосату клеток растений, трансформированных указанным геном;
b) получение трансформированной растительной клетки;
c) регенерацию из трансформированной растительности клетки генетически трансформированного растения, которое обладает повышенной толерантностью к глифосату.
Структурная последовательность ДНК, кодирующая фермент EPSPS класса II, может быть выбрана из группы последовательностей, включающих SEQ ID NO:2, SEQ ID NO: 4 и SEQ ID NO: 6.
Промотор может быть получен из ДНК вируса растения и может быть промотором CaMV 35D или FMV 35S.
В еще одном аспекте настоящего изобретения рекомбинантная двунитевая молекула ДНК включает:
a) промотор, функционирующий в растительных клетках и обеспечивающий продукцию последовательности РНК;
b) структурную последовательность ДНК, обеспечивающую продукцию последовательности РНК, кодирующей фермент EPSPS класса II; и
c) 3'-нетранслируемую область, функционирующую в растительных клетках, обеспечивающую добавление тяжа поли-A к 3'-концу последовательности РНК,
причем промотор является гетерологичным по отношению к структурной последовательности ДНК и адаптирован таким образом, чтобы обеспечить достаточную экспрессию слитого пептида для повышения толерантности к глифосату растительных клеток, трансформированных указанной молекулой ДНК.
В еще одном аспекте настоящего изобретения представлен способ селективного контроля за сорняками путем посадки семян растений или растений, трансформированных геном EPSPS класса II, для придания толерантности к глифосату растениям, что позволяет наносить глифосатсодержащие гербициды на растения для уничтожения чувствительных к глифосату сорняков, но не культурных растений.
Способ селективного контроля за сорняками в полевых условиях, предусматривающий посадку семян или растений, может включать следующие стадии:
a) посадку указанных семян растений или растений, толерантных к глифосату, вследствие введения в указанные семена или растения рекомбинантной двунитевой молекулы ДНК, включающей:
i) промотор, функционирующий в растительных клетках и обеспечивающий продукцию последовательности РНК;
ii) структурную последовательность ДНК, обеспечивающую продукцию последовательности РНК, кодирующей полипептид, который включает аминоконцевой транзитный пептид хлоропласта и EPSPS фермент класса II;
iii) 3'-нетранслируемую последовательность ДНК, функционирующую в растительных клетках, обеспечивающую добавление тяжа поли-A к 3'-концу последовательности РНК;
причем промотор является гетерологичным по отношению к структурной последовательности ДНК и адаптирован таким образом, чтобы обеспечить достаточную экспрессию слитого полипептида для повышения толерантности к глифосату растительных клеток, трансформированных указанным геном; и
b) нанесение на указанные растения и сорняки достаточного количества глифосата для обеспечения контроля за сорняками, но без существенного влияния на культурное растение.
Структурная последовательность ДНК, кодирующая SPSPS фермент класса II, может быть выбрана из последовательностей, представленных в SEQ ID NO:2, SEQ IN NO:4 или SNQ ID NO:6.
Молекула ДНК может дополнительно включать промотор, выбранный из группы, состоящей из промоторов CAMV 35S и FMV 35S. Культурное растение может быть выбрано из группы, состоящей из кукурузы, пшеницы, риса, сои, хлопка, сахарной свеклы, масличного рапса, канолы, льна, подсолнечника, картофеля, табака, томатов, люцерны, тополя, сосны, яблони и винограда.
Другие и дополнительные объекты, преимущества и аспекты изобретения будут очевидны из приложенных чертежей и описания изобретения.
На фиг. 1 показана последовательность ДНК (SEQ ID N 1) для полной длины промотора мозаичного вируса норичника (FMV 35S).
На фиг. 2 показан космидный вектор клонирования pMON 17020.
На фиг. 3 показана последовательность структурной ДНК (SEQ ID N 2) для гена класса 11 EPSPS из бактериального изолята Agrobacterium sp. штамм CP4 и происходящая из нее аминокислотная последовательность (SEQ ID N 3).
На фиг. 4 показана последовательность структурной ДНК (SEQ ID N 4) для гена класса II EPSPS из бактериального изолята Achromobacter sp. штамм LBAA и происходящая из нее аминокислотная последовательность (SEQ ID N 5).
На фиг. 5 показана последовательность структурной ДНК (SEQ ID N 6) для гена класса II EPSPS из бактериального изолята Pseudomonas sp. штамм PG2982 и происходящая из нее аминокислотная последовательность (SEQ ID N 7).
На фиг. 6 показано сравнение Бетфита аминокислотной последовательности E.coli EPSPS (SEQ ID N 8) с последовательностью CP4 EPSPS (SEQ ID N 3).
На фиг. 7 показано сравнение Бестфита аминокислотной последовательности CP4 EPSPS (SEQ ID N 3) с последовательностью LBAA EPSPS (SEQ ID N 5).
На фиг. 8 показана структурная последовательность ДНК (SEQ ID N 9) для синтетического гена CP4 класса II EPSPS.
На фиг. 9 показана последовательность ДНК (SEQ ID N 10) хлоропластного транзитного пептида (CTP) и кодируемая аминокислотная последовательность (SEQ ID N 11), происходящая из Arabidopsis thaliana EPSPS CTP и содержащая SphI сайт рестрикции в сайте переработки хлоропласта, здесь далее упоминаемая как CTP2.
На фиг. 10 показана последовательность ДНК (SEQ ID N 12) хлоропластного транзитного пептида и кодируемая аминокислотная последовательность (SEQ ID N 13), происходящая из Arabidopsis thaliana EPSPS-гена и содержащая Sco RI сайт рестрикции в зрелой области EPSPS, здесь далее упоминаемая как CTP3.
На фиг. 11 показана последовательность ДНК (SEQ ID N 14) хлоропластного транзитного пептида и кодируемая аминокислотная последовательность (SEQ ID N 15), происходящая из Petunia hybrida EPSPS CTP и содержащая SphI-сайт рестрикции в сайте переработки хлоропласта и в которой аминокислоты в сайте переработки заменены на -Cys-Met, здесь далее упоминаемая как CTP4.
На фиг. 12 показана последовательность ДНК (SEQ ID N 16) хлоропластного транзитного пептида и кодируемая аминокислотная последовательность (SEQ ID N 17), происходящая из Petunia hybrida EPSPS-гена с сайтом ScoRI природного происхождения в зрелой области EPSPS-гена, здесь далее упоминаемая как CTP5.
На фиг. 13 показана плазмидная карта CP4 трансформации растения/вектор экспрессии pMON 17110.
На фиг. 14 показана плазмидная карта CP4 EPSPS синтетического гена трансформации растения/вектор экспрессии pMON 17131.
На фиг. 15 показана плазмидная карта CP4 EPSPS свободная ДНК трансформации растения/вектор экспрессии pMON 13640.
На фиг. 16 показана плазмидная карта CP4 трансформации растения/вектор прямой селекции pMON 17227.
На фиг. 17 показана плазмидная карта CP4 трансформации растения/вектор экспрессии pMON 19653.
Экспрессия растительного гена, который существует в форме двуниточной ДНК, включает синтез матричной РНК (мРНК) из одной нити ДНК с помощью фермента РНК полимеразы, и последующий прессинг мРНК продукта транскрипции внутрь ядра. Этот процессинг включает 3'-нетранслируемую область, которая дополняет полиаденилированные нуклеотиды к 3'-концу РНК.
Транскрипция ДНК в мРНК регулируется областью ДНК, обычно упоминаемой как "промотор". Область промотора содержит последовательность оснований, которая сигнализирует РНК полимеразе ассоциировать с ДНК и инициировать транскрипцию в мРНК, используя одну нить ДНК в качестве матрицы, чтобы получить комплементарно соответствующую нить РНК.
В литературе был описан ряд промоторов, которые активны в клетках растений. Они включают промоторы нопалин синтазу (NOS) и октопин синтазу (OCS) (которые переносятся индуцирующими опухоль плазмидами Agrobacterium tumetaciens), промоторы вируса мозаики цветной капусты (CaMV) 19S и 35S, индуцируемый светом промотор из малой субъединицы рибулоза бис-фосфат карбоксилазы (ss RUBISCO, очень часто встречающийся полипептид растений) и промотор транскрипции полной длины из вируса мозаики норичника (FMV35S). Все эти промоторы были использованы для создания различных типов конструкций ДНК, которые были экспрессированы в растениях, смотри, например, PCT публикацию WO 84/02913 (Роджерс с сотр., Монсанто).
Промоторы, которые являются известными или вызывающими транскрипцию ДНК в клетках растений, могут быть использованы в настоящем изобретении. Такие промоторы могут быть получены из множества источников, таких как растения и ДНК вирусов растений, и включают, но не ограничиваются промоторами CaMV35S и FMV35S и промоторы, выделенные из генов растений, таких, как ssRUBISCO-гены. Как описано ниже, предпочтительно, чтобы конкретный выбранный промотор мог быть способен вызывать достаточную экспрессию, чтобы получить в результате эффективное количество класса II EPSPS, чтобы придать растению значительную толерантность к глифосатным гербицидам. Количество класса II EPSPS, необходимое для индукции желаемой толерантности, может варьировать с видом растения. Предпочтительно, чтобы использованные промоторы имели относительно высокую экспрессию во всех меристематических тканях в дополнение к другим тканям, так как теперь известно, что глифосфат транслокирован и аккумулирован в тканях растений этого типа. Альтернативно может быть использовано сочетание химерных генов для совокупного результата при необходимом общем уровне экспрессии выбранного фермента класса II EPSPS, чтобы получить в результате толерантный к глифосату фенотип.
Продуцированная конструкцией ДНК настоящего изобретения мРНК также содержит 5'-нетранслируемую лидерную последовательность. Эта последовательность может происходить из промотора, выбранного для экспрессии гена, и может быть специально модифицирована, чтобы увеличить трансляцию мРНК. 5'-нетранслируемые области также могут быть получены из вирусных РНК, из подходящих эукариотных генов, или из синтетических генных последовательностей. Настоящее изобретение не ограничивается конструкциями, которые представлены в следующих примерах, где нетранслируемая область происходит из обеих 5'-нетранслируемой последовательности, которая сопровождает промоторную последовательность, и части 5'-нетранслируемой области гена белка оболочки вируса. Скорее нетранслируемая лидерная последовательность может происходить из неродственного промотора или кодирующей последовательности, как обсуждено выше.
Предпочтительным промотором для использования в настоящем изобретении является матричный промотор полной длины (SEQ ID N 1) из вируса мозаики норичника (FMV35S), который функционирует как сильный и равномерный промотор с особенно хорошей экспрессией в меристематической ткани для химерных генов, вставленных в растения, особенно двудольные. Полученное в результате трансгенное растение в общем экспрессирует белок, кодируемый вставленным геном, на более высоком и однородном уровне в тканях и клетках трансформированного растения, чем тот же самый ген под действием усиленного CaMV35S - промотора. Сссылаясь на фиг. 1, последовательность ДНК (SEQ ID N 1) FMV35S промотора локализована между 6368 и 6930 нуклеотидами FMV генома. 5'-нетранслируемая лидерная последовательность предпочтительно связана с промотором. Лидерная последовательность может быть из самого генома FMV35S или может быть из другого источника, чем FMV35S.
3'-Нетранслируемая область химерного гена содержит сигнал полиаденилирования, который функционирует в растениях, вызывая присоединение полиаденилированных нуклеотидов к 3'-концу вирусной РНК. Примерами подходящих 3'-областей являются (1) 3'-транскрибированные нетранслируемые области, содержащие сигнал полиаденилирования генов, вызывающих опухоль, плазмиды (Ti) Agrobacterium, такие, как ген нопалин синтазы (NOS), и (2) растительные гены, подобные генам запасных белков сои и маленькой субъединицы гена рибулоза-1,5-бисфосфат карбоксилазы (ssRUBISCO). Примером предпочтительной 3'-области является таковая из ssRUBISCO гена гороха (E9), более детально описанная ниже.
Конструкция ДНК настоящего изобретения также содержит структурную кодирующую последовательность в виде двуниточной ДНК, которая кодирует толерантный к глифосату высоко эффективный фермент класса II EPSPS.
Идентификация толерантных к глифосату, высоко эффективных EPSPS ферментов.
При попытке идентифицировать и выделить толерантные к глифосату, высоко эффективные ферменты EPSPS провели кинетический анализ EPSPS ферментов из ряда бактерий, обладающих толерантностью к глифосату или выделенных из подходящих источников. Было обнаружено, что в некоторых случаях ферменты EPSPS не показывают толерантность для ингибирования глифосата, и был сделан вывод, что толерантность фенотипа бактерии возникает благодаря непроницаемости глифосата или других факторов. Однако в ряде случаев были идентифицированы микроорганизмы, у которых фермент EPSPS показал более высокую степень толерантности к ингибированию глифосата и которые показывают низкий Km для PEP при сравнении с ранее приведенным для других микробных и растительных источников. EPSPS ферменты из этих микроорганизмов затем были подвергнуты дополнительному изучению и анализу.
В таблице 1 приведены данные, полученные для EPSPS ферментов, идентифицированных и выделенных в результате описанного выше анализа. В таблице 1 включены данные для трех идентифицированных ферментов класса II EPSPS, для которых наблюдалось, что они имеют высокую толерантность к ингибированию глифосфата и низкую Km для PEP, а также данные для нативного EPSPS петунии и толерантного к глифосату варианта EPSPS петунии, упоминаемого как GA101. Вариант GA101 был так назван потому, что в нем имеется замещение аланинового остатка на остаток глицина в положении 101 (в отношении петунии) в инвариантной области. При внесении изменения EPSPS петунии (GA101) в ряд других ферментов EPSPS наблюдались подобные изменения в кинетике, повышение Ki для глифосата и Km для PEP.
Первоначально идентифицируют Agrobacterium sp. штамм CP4 по его способности расти на глифосате как источнике углерода (10 мМ) в присутствии 1 мМ фосфата. Штамм CP4 был идентифицирован из коллекции, полученной из колонки со стационарным слоем иммобилизованных клеток, в которой применены шарики диатомитовой земли Маннвилл K-635. Колонку гоняли три месяца на сбросовой воде с установки по производству глифосата. Колонка содержала 50 мг/мл глифосата и NH3 и NH4Cl. Общий органический углерод составляет 300 мг/мл и БПК (биологическое потребление кислорода - мера доступности "мягкого" углерода) составляет менее 30 мг/мл. Обработанная таким образом колонка была описана (Heitkamp et al., 1990). Минимальна солевая среда Дворкина-Фостера, содержащая глифосат в количестве 10 мМ и 1 мМ фосфата, была использована для отбора микробов из промывки этой колонки, которые способны расти на глифосате в качестве единственного источника углерода. Минимальную среду Дворкина-Фостера готовят объединением в 1 л (с автоклавной водой) по 1 мл каждого из A, B и C и 10 мл D (см. ниже) и 5 мг тиамина HCl.
A. Д-Ф соли (1000Х stock; на 100 мл; автоклавированная):
H3BO3 - 1 мг
MnSO4 • 7 H2O - 1 мг
ZnSO4 • 7 H2O - 12,5 мг
CuSO4 • 5 H2O - 8 мг
NaMoO3 • 3 H2O - 1,7 мг
B. FeSO4 • 7 H2O (1000Х stock; на 100 мл; автоклавированная):
0,1 г
C. MgSO4 • 7 H2O (1000Х stock; на 100 мл; автоклавированная):
20 г
D. (NH4)2SO4 (1000Х stock; на 100 мл; автоклавированная):
20 г
Дрожжевой экстракт (УЕ, Дифко) прибавляют до конечной концентрации 0,01 или 0,001%. Штамм CP4 также выращивают на среде, составленной из Д-Ф солей, исправленной, как описано выше, содержащей глюкозу, глюконат и нитрат (каждый по 0,1%) в качестве источников углерода и неорганический фосфат (0,2-1,0 мМ) в качестве источника фосфора.
Другие микроорганизмы, содержащие класс II EPSPS, были идентифицированы как Achromobacter sp. штамм LBAA, который был взят из коллекции бактерий, ранее описанной (Hallas et al., 1988), и Pseudomonas sp. штамм PG2982, который был описан в литературе (Moore et al., 1983; Fitzgibbon, 1983). Ранее уже сообщалось, из измерений в сырых лизатах, что EPSPS фермент из штамма PG2982 был менее чувствительным к ингибированию глифосатом, чем штамм из E. coli, но не сообщались детали этой потери чувствительности и не приводились данные о Km для PEP для этого фермента или о последовательности ДНК для гена этого фермента (Fitzgibbon, 1988; Fitzgibbon and Braymer, 1990).
Отношение класс II EPSPS к ранее изученным ферментам
Все EPSPS белки, изученные до сегодняшнего дня, показали замечательную степень гомологии. Например, бактериальные и растительные EPSPS примерно на 54% являются идентичными и при таком высоком подобии, как 80%. Внутри самих бактериальных EPSPS и растительных EPSPS степень идентичности и подобие являются очень высокими (смотри таблицу 2).
При зондировании сырых экстрактов CP4 и LBAA бактерий (50 μг белка) с использованием анти-EPSPS антител кролика (Padgette et al., 1987) белков EPSPS петунии с помощью анализа Вестерна, не смогли определить положительный сигнал даже при увеличенном времени экспозиции (Белок A - 125I проявляющая система) и в условиях, когда контроль EPSPS (20 нг EPSPS петунии, класс I EPSPS) легко осуществим. Наличие EPSPS-активности в этих экстрактах подтверждают ферментным анализом. Этот неожиданный результат, указывающий на потерю подобия между EPSPS из этих бактериальных изолятов и ранее изученными EPSPS, ассоциируемый с сочетанием низкой Km для PEP и высокой Ki для глифосата, иллюстрирует, что эти новые EPSPS-ферменты отличаются от известных EPSPS-ферментов, теперь упоминаемых как класс I EPSPS.
Толерантные к глифосату ферменты в микробных изолятах
Для ясности и краткости изложения последующее описание выделения генов, кодирующих класс II EPSPS-ферментов, относится к выделению такого гена из бактериального изолята. Специалисты в данной области должны понимать, что такая же или подобная стратегия может быть использована для выделения таких генов из других микробных изолятов, из растительных и грибных источников.
Клонирование Agrobacterium sp. штамм CP4 EPSPS гена(ов) в E.coli.
После установления существования подходящего EPSPS в Agrobacterium sp. штамм CP4, были сделаны две параллельные попытки проклонировать ген: клонирование, основанное на ожидаемом фенотипе толерантного к глифосату EPSPS; и очистка фермента, чтобы получить материал для извлечения антител и получения аминокислотных последовательностей из белка для облегчения подтверждения клонов. Клонирование и генетические методики, если нет других указаний, в общем являются такими, как описано у Маниатиса с сотр., 1982 или у Самбрука с сотр., 1987. Стратегия клонирования является следующей: введение космидного банка штамма Agrobacterium sp. штамм CP4 в E.coli и селекция EPSPS гена с помощью селекции роста на ингибирующих концентрациях глифосата.
Готовят хромосомальную ДНК из штамма Agrobacterium sp. штамм CP4 следующим образом: Повторно суспендируют клеточный шарик из 200 мл L-бульона (Miller, 1972) последней лог-фазы культуры Agrobacterium sp. штамм CP4 в 10 мл раствора 1; 50 мМ глюкозы, 10 мМ ЕДТА, 25 мМ Трис-Cl pH 8,0 (Birnboim and Doly, 1979). Прибавляют SDS до конечной концентрации 1% и суспензию подвергают трем циклам замораживание-оттаивание, каждый состоит в погружении в сухой лед на 15 минут и в воду при 70oC на 10 минут. Затем лизат четыре раза экстрагиурют равными объемами фенола : хлороформа (1:1, фенол, насыщенный TE, TE = 10 мМ Трис, pH 8,0; 1 мМ ЕДТА) и разделяют фазы центрифугированием (15000 g; 10 минут). Осажденный эталоном материал собирают в шарик, отделяя от супернатанта краткосрочным центрифугированием (8000 g, 5 минут) с последующим добавлением двух объемов этанола. Шарик повторно суспендируют в 5 мл TE и диализуют в течение 16 часов при 4oC против 2 л TE. Этот препарат представляет собой 5 мл раствора ДНК 552 мкг/мл.
Следующим образом готовят частично рестриктированную ДНК. Обрабатывают три аликвотных образца 100 мкг CP4 ДНК в течение 1 часа при 37oC рестрикционной эндонуклеаной Hind III при дозах 4, 2 и 1 единица фермента/мкг ДНК соответственно. Собирают образцы ДНК, устанавливая 0,25 мМ ЕДТА, и экстрагируют равным объемом фенола:хлороформа. После добавления ацетата натрия и этанола осаждают ДНК двумя объемами этанола и центрифугируют для получения шарика (12000 g, 10 минут). Сухой шарик ДНК повторно суспендируют в 500 мкл TE и наслаивают на градиент 10-40% сахарозы (с 5% инкрементами 5,5 мл каждого) в 0,5 М NaCl, 50 мМ Трис pH 8,0, 5,0 мМ ЕДТА. После центрифугирования в течение 20 часов при 26000 об/мин в SW 28 роторе трубки прокалывают и собирают фракции примерно 1,5 мл. Образцы (20 мкл) каждой второй фракции наносят на 0,7% агарозный гель и определяют размер ДНК путем сравнения с линеаризованной лямбда ДНК и Hind III-переваренной лямбда ДНК в качестве стандартов. Собирают фракции, содержащие фрагменты ДНК 25-35 ко, обессоливают на колонках AMICON10 (7000 об/мин, 20oC, 45 минут) и концентрируют осаждением. Эта процедура дает 15 мкг CP4 ДНК требуемого размера. Составляют банк космид, используя вектор pMON17020. Этот вектор, карта которого представлена на фиг. 2, основан на pBR327-репликоне и содержит спектиномицин/стрептомицин (Spr, spc) ген устойчивости из Тп7 (Fling et al., 1985), ген устойчивости к хлорамфениколу (Cmr, cat) из Тп9 (Alton et al., 1985), промоторную область гена 10 из фага Т7 (Dunn et al., 1983) и cos-фрагмент 1,6 ко Bgl II фага лямбда из pHC79 (Hohn and Collins, 1980). Ряд сайтов клонирования локализованы ниже cat-гена. Поскольку преобладающий блок экспрессии генов из других микробных источников должен появиться в E.coli на уровне транскрипции, применение T7 промотора и подача T7 полимеразы в in trans от pGP1-2 плазмиды (Tabor and Richardson, 1985) делают возможной экспрессию больших сегментов ДНК от чужеродной ДНК, даже тех, что содержат терминальные последовательности транскрипции РНК полимеразы. Экспрессия spc гена ослабляется транскрипцией от Т7 промотора, так что только Cmr может быть селектирован в нитях, содержащих pGP1-2. Использование устойчивости к антибиотикам, такой как Cmr устойчивость, которая не использует мембранный компонент, является предпочтительным благодаря наблюдению, что высокий уровень устойчивости генов, которые включают мембранный компонент, а именно устойчивость к β-лактамазе и Amp-устойчивость, приводит к увеличению толерантного к глифосату фенотипа. По-видимому это происходит благодаря исключению глифосата из клетки с помощью мембраны, локализующей устойчивость белка. Также предпочтительно, чтобы отбираемый маркер был ориентирован в том же самом направлении, что и T7 промотор. Затем вектор отрезают Hind III и обрабатывают щелочной фосфатазой теленка (CAP) в препарате для клонирования. Последовательности вектора и мишени лигируют путем сочетания следующего:
Вектор ДНК (Hind III/CAP) - 3 мкг
Фракционированные по размерам фрагменты СР4 Hind III - 1,5 мкг
1OX буфер лигирования - 2,2 мкл
T4 ДНК лигаза (Нью Инглэнд Биолабс) (400 E/мкл) - 1,0 мкл
и добавления воды до - 22,0 мкл
Эту смесь инкубируют в течение 18 часов при 16oC. 10X буфер лигации представляет собой 250 мМ Трис-HCl, pH 8,0; 100 мМ MgCl2; 100 мМ дитиотрейтола; 2 мМ спермидина. Упаковывают 5 мкл лигированной ДНК в частицы лямбда фага (Стратаген, Джигапакк Голд), используя процедуру производителя.
Образец 200 мкл E.coli HB101 (Boyer and Rolland-Dussoix, 1973), содержащий плазмиду экспрессии pGP1-2 T7 полимеразы (Tabor and Richardson, 1985) и выращенную в течение ночи в L-бульоне (с 0,2% мальтозы и 50 мкг/мл канамицина), инфицируют 50 мкл упакованной ДНК. Трансформанты отбирают при 30oC на M9 (Miller, 1972) агаре, содержащем 50 мкг/мл канамицина. 25 мкг/мл хлорамфеникола, 50 мкг/мл L-пролина, 50 мкг/мл L-лейцина и 5 мкг/мл B1 и 3,0 мМ глифосата. Также высевают на ту же среду, лишенную глифосата, аликвотные образцы, чтобы оттитровать упакованные космиды. Изолируют космидные трансформанты на этой последней среде в количестве 5•105 на мкг CP4 Hind III ДНК после 3 дней при 30oC. Колонии выращивают на глифосатном агаре от 3 до 15 дней при конечном количестве от ≈ 1 на 200 космид. ДНК получают из 14 толерантных к глифосату клонов и после подтверждения этого фенотипа трансформируют в E.coli GB100 (pGP1-2), E.coli GB100 является aro A производным MM294 (Talmadge and Gilbert, 1980) и испытывают на комплементарность для роста в отсутствие добавленных ароматических аминокислот и аминобензойных кислот. Могут быть использованы другие aro A штаммы, такие как SR481 (Bachman et al. , 1980; Padgette et al., 1987), и они будут пригодными для этого эксперимента. Использование GB 100 приведено только в качестве примера и не должно рассматриваться в ограничивающем смысле. Такой aro A штамм обычно требует, чтобы среда роста была дополнена L-фенилаланином, L-тирозином и L-триптофаном, каждой по 100 мкг/мл, и п-оксибензойной кислотой, 2,3-диоксибензойной кислотой и п-аминобензойной кислотой, по 5 мкг/мл каждой, для роста на минимальной среде. Из четырнадцати испытанных космид только одна показала комплементарность aro-A-фенотипу. Трансформанты этой космиды, pMON17076, показали слабый, но равномерный рост на недополненной минимальной среде после 10 дней.
Белки, кодируемые космидами, были определены in vivo с использованием T7 системы экспрессии (Tabor and Richardson, 1985), а тестовые и контрольные космиды были выращены при 30oC на 2 мл L-бульона с 25 и 50 мкг/мл соответственно хлорамфеникола и канамицина для считывания Клетта ≈ 50. Извлекают аликвот и собирают клетки центрифугированием, промывают M9 солями (Miller, 1972) и повторно суспендируют в 1 мл среды M9, содержащей 0,2% глюкозы, 20 мкг/мл тиамина и содержащей по 0,01% 18 аминокислот (минут цистеин и метионин). После инкубации при 30oC в течение 90 минут культуры переносят на водяную баню при 42oC и оставляют на 15 минут. Прибавляют рифампицин (Сигма) до 200 мкг/мл и оставляют культуры при 42oC на дополнительные 10 минут, а затем переносят на 20 минут при 30oC. Клетки собирают центрифугированием и суспендируют в 60-120 мкл дробящего буфера 60 мМ Трис-HCl, pH 6,8, 1% SDS, 1% 2-меркаптоэтанола, 10% глицерина, 0,01% бромфенольного голубого. Аликвотные образцы подвергали электрофорезу на 12,5% SDS-PAGE и смачивают потом в течение 60 минут 10 объемами уксусной кислоты-метанола-воды (10:30:60), гель смачивают ENLIGHTNINGTM (Дюпон), следуя инструкциям производителя, сушат и выдерживают при -70oC на рентгеновской пленке. Белки около 45 кд размером, меченные 35S-метионином, были детектированы в ряде космид, включая pMON17076.
Очистка EPSPS из Agrobacterium sp. штамм CP4
Все процедуры по очистке белков проводили при 3-5oC. Анализ ферментов EPSPS осуществляют с использованием или выделения фосфата или метода радиоактивной высоко эффективной жидкостной хроматографии (ВЭЖХ), как ранее было описано у Padgette et al., 1987, используя 1 мМ фосфоенолпирувата (PEP, Боерингер) и 2 мМ шикимат-3-фосфата в субстрате. Для радиоактивного анализа ВЭЖХ используют 14C-PEP (Амстердам). S3P синтезируют, как описано ранее у Wibbenmeyer et al., 1988. Осуществляют секвенирование N-концевых аминокислот путем загрузки образцов на полибреновый прецикличный фильтр в аликвотах при сушке. Используют автоматизированную химию деградации для определения N-концевой последовательности белка, применяя газофазный секвенсер Апплайд Биосистемс Мордель 470A (Hunkapiller et al., 1983) с анализатором Апплайд Биосистемс 120A PTH.
Проводят пять 10 л ферментаций со спонтанным "гладким" изолятом штамма CP4, который показывает меньшую агглютинацию при росте в жидкой культуре. Такая пониженная агглютинация и гладкая морфология колоний может быть благодаря сниженной продукции полисахарида этим изолятом. В следующем разделе, относящемся к очистке EPSPS фермента, CP4 упоминается как "гладкий" изолят - CP4-SI. Собирают клетки из трех партий, показывающие самую высокую специфическую активность. Пасту клеток Agrobacterium sp. CP4 (300 г) дважды промывают 0,5 л 0,9% солевого раствора и собирают центрифугированием (30 минут, 8000 об/мин в G53 Сорволл роторе). Шарик клеток суспендируют в 0,9 л экстракционного буфера (100 мМ Трис-HCl, 1 мМ ЕДТА, 1 мМ ВАМ (Бензамидина), 5 мМ ДТТ, 10% глицерина, pH 7,5) и лизируют за 2 прохода через Manton Gaulin cell. Полученный в результате раствор центрифугируют (30 минут, 8000 об/мин) и супернатант обрабатывают 0,21 л 1,5% протамин сульфата (в 100 мМ Трис-C1, pH 7,5, 0,2% масс./объем конечная концентрация протамин сульфата). После 1 часа перемешивания смесь центрифугируют (50 минут, 8000 об/мин) и полученный в результате супернатант обрабатывают твердым сульфатом аммония до 40% насыщения и перемешивают 1 час. После центрифугирования (50 минут, 8000 об/мин) полученный в результате супернатант обрабатывают твердым сульфатом аммония до 70% насыщения, перемешивают 50 минут и собирают нерастворимый белок центрифугированием (1 час, 8000 об/мин). Эту 40-70% фракцию сульфата аммония затем растворяют в экстракционном буфере, чтобы получить конечный объем 0,2 л, и дважды диализуют (Spectrum 10000 MW cutoff dialysis tubing) против 2 л экстракционного буфера всего 12 часов.
К полученной в результате диализа 40-70% фракции сульфата аммония (0,29 л) прибавляют твердый сульфат аммония до получения конечной концентрации 1 М. Этот материал загружают (2 мл/мин) в колонку (5 см х 15 см, 295 мл), заполненную фенилсефарозной CL-4B смолой (Фармасиа), уравновешенной экстракционным буфером, содержащим 1М сульфата аммония, и промывают тем же самым буфером (1,5 л, 2 мл/мин). EPSPS элюируют при линейном градиенте экстракционного буфера от 1 М до 0,00 М сульфата аммония (общий объем 1,5 л, 2 мл/мин). Собирают фракции по 20 мл и анализируют на EPSPS-активность по методу выделения фосфата. Собирают фракции с наиболее высокой активностью (фракции 36-50) и диализуют против 3х2 л (18 часов) 10 мМ Трис-C1, 25 мМ KCl, 1 мМ ЕДТА, 5 мМ ДТТ, 10% глицерина, pH 7,8.
Загружают 350 мл диализованного EPSPS-экстракта со скоростью 5 мл/мин в колонку 2,4 см х 30 см, 136 мл, заполненную смолой Q-Сефароза Фаст Флоу (Фармасиа), уравновешенной 10 мМ Трис-C1, 25 мМ KCl, 5 мМ ДТТ, 10% глицерина, pH 7,8 (Q-Сефарозный буфер) и промывают 1 л того же буфера. EPSPS элюируют с линейным градиентом сефарозного буфера от 0,025 М до 0,40 М KCl (общий объем 1,4 л, 5 мл/мин). Собирают фракции по 15 мл и анализируют на EPSPS-активность методом выделения фосфата. Объединяют фракции с самой высокой EPSPS-активностью (фракции 47-60) и осаждают белок добавлением твердого сульфата аммония до 80% насыщения и перемешивают 1 час. Собирают осажденный белок центрифугированием (20 минут, 12000 об/мин, в роторе GSA Сорволл), растворяют в Q-Сефарозном буфере (общий объем 14 мл) и диазилизуют против того же буфера (2 х 1 л, 18 часов).
Загружают 19 мл полученного в результате диализа частично очищенного EPSPS-экстракта со скоростью 1,7 мл/мин в Моно Q 10/10 колонку (Фармациа), уравновешенную Q-Сефарозным буфером, и промывают 35 мл того же буфера. Элюируют EPSPS при линейном градиенте от 0,025 М до 0,35 М KCl (общий объем 119 мл, 1,7 мл/мин). Собирают фракции по 1,7 мл и анализируют на EPSPS-активность методом выделения фосфата. Объединяют фракции с наиболее высокой активностью (фракции 30-37) (6 мл).
В пул Моно Q добавляют твердый сульфат аммония до 1 М по сульфату аммония и аликвоты 2 мл хроматографируют на колонке с фенилсуперозой 5/5 (Фармасиа), уравновешенной 10 мМ Трис-Cl, 5 мМ ДТТ, 1 М сульфата аммония, 10% глицерина, pH 7,5 (фенилсуперозный буфер). Образцы загружают со скоростью 1 мл/мин, промывают фенилсуперозным буфером (10 мл) и элюируют с линейным градиентом фенилсуперозного буфера от 1 М до 0,00 М сульфата аммония (общий объем 60 мл, 1 мл/мин). Собирают фракции по 1 мл и анализируют на EPSPS-активность методом выделения фосфата. Объединяют вместе фракции от каждого опыта с наиболее высокой EPSPS-активностью (фракции 35-40) (10 мл, 2,5 мг белка). Для определения N-концевой аминокислотной последовательности часть одной фракции (# 39 из опыта 1) диализуют против 50 мМ NaHCO3 (2 х 1 л). В полученном в результате чистом образце EPSPS (0,9 мл, 77 мкг белка) было найдено, что имеется единственная аминокислотная последовательность: XH(G) ASSRPATARKSS (G)LX(G)(T)V(R) IPG(D)(K)(M) (SEQ ID NO:18).
В этой и во всех последующих аминокислотных последовательностях использована стандартная однобуквенная номенклатура. Все пептидные структуры, представленные в последующем описании, показаны в удобном формате, где аминогруппа у N-конца появляется слева, а карбоксильная группа у C-конца - справа. Подобным образом, номенклатура аминокислот для встречающихся в природе аминокислот, найденных в белке, является следующей: аланин (Ala, A), аспарагин (Asn, N), аспаргиновая кислота (Asp, D), аргинин (Arg, R), цистеин (Cys, C), глютаминовая кислота (Glu, E), глютамин (Gln, Q), глицин (Gly, G), гистидин (His, H), изолейцин (Ile, I), лейцин (Leu, L), лизин (Lys, K), метионин (Met, M), фенилаланин (Phe, F), пролин (Pro, P), серин (Ser, S), треонин (Thr, T), триптофан (Trp, W), тирозин (Tyr, Y) и валин (Val, V). "X" используют, когда аминокислотный остаток неизвестен, а скобка означает, что недвусмысленное обозначение невозможно и обозначение аминокислоты в скобках является наиболее вероятным, исходя из известной информации.
Оставшийся фенилсуперозный EPSPS пул диализуют против 50 мМ Трис-Cl, 2 мМ ДТТ, 10 мМ KCl, 10% глицерина, pH 7,5 (2х 1 л). Аликвот (0,55 мл, 0,61 мг белка) загружают со скоростью 1 мл/мин в Моно Q 5/5 колонку (фармасиа), уравновешенную Q-сефарозным буфером, промывают тем же буфером (5 мл) и элюируют с линейным градиентом Q-сефарозного буфера 0-0,14 М KCl в течение 10 минут, затем выдерживают при 0,14 М KCl (1 мл/мин). Собирают фракции (1 мл) и анализируют на EPSPS активность методом выделения фосфата и подвергают SDS-PAGE (10-15%, Фаст Систем, Фармасиа, с окрашиванием серебром) для определения чистоты белка. Объединяют фракции, показывающие одну полосу белка при SDS-PAGE (22-25, 222 мкг) и диализуют против 100 мМ бикарбоната аммония, pH 8,1 (2 х 1 л, 9 часов).
Трипсинолиз и секвенирование белка Agrobacterium sp. штамм CP4 EPSPS.
К полученным в результате 111 мкг чистого EPSPS Agrobacterium sp. штамм CP4 прибавляют 3 мкг трипсина (Кальбиохем) и позволяют пройти реакции трипсинолиза в течение 16 часов при 37oC. Трипсиновый перевар затем хроматографируют (1 мл/мин) на колонке с C18 обратимой фазой ВЭЖХ (Видак), как было описано ранее Padgette et al., 1988 для E.coli EPSPS. Для всех пептидных очисток 0,1% трифторуксусная кислота (TFA, Пьерс) обозначена как "RP-A" буфер, а 0,1% TFA в ацетонитриле является буфером "RP-B". Градиент, использованный для элюирования трипсинизированного EPSPS из Agrobacterium sp. CP4, составил: 0-8 минут, 0% RP-B, 8-28 минут 0-15% RP-B, 28-40 минут, 15-21% RP-B, 40-68 минут, 21-49% RP-B, 68-72 минут, 49-75% RP-B, 72-74 минуты, 75-100% RP-B. Собирают фракции по 1 мл и, основываясь на профиле элюции при 210 нм, получают по крайней мере 70 отдельных пептидов из трипсинизированного EPSPS. Фракции 40-70 выпаривают досуха и повторно растворяют в 150 мкл 10% ацетонитрила, 0,1% тирфтороуксусной кислоте.
Фракцию 61 пептида дополнительно очищают на C18 колонке при градиенте: 0-5 минут, 0% RP-B, 5-10 минут, 0-38% RP-B, 10-30 минут, 38-45% B. Фракции собирают на основе УФ-сигнала при 210 нм. Большой пептидный пик во фракции 24 элюируют при 42% RP-B и сушат снова, повторно суспендируют, как описано выше, и повторно хроматографируют на C18 колонке с градиентом: 0-5 минут, 0% RP-B, 5-12 минут, 0-38% RP-B, 12-15 минут, 38-39% RP-B, 15-18 минут, 39% RP-B, 18-20 минут, 39-41% RP-B, 20-24 минуты, 41% RP-B, 24-28 минут, 42% RP-B. Пептид из фракции 25, элюированный при 41% RP-B и обозначенный как пептид 61-24-25, подвергают N-концевому аминокислотному секвенированию и определяют следующую последовательность:
APSM(1)(D)EYPILAV (SEQ ID N 19)
Фракцию 53 CP4 EPSPS трипсинизированного белка дополнительно очищают на C18 ВЭЖХ при градиенте 0% B (5 минут), 0-30% B (5-17 минут), 30-40% B (17-37 минут). Пептид из фракции 28, элюированный при 34% B и обозначенный как пептид 53-28, подвергают N-концевому аминокислотному секвенированию и определяют следующую последовательность:
ITGLLEGEDVINTGK (SEQ ID N 20)
Для подтверждения космидного клона CP4 EPSPS был сконструирован ряд олигонуклеотидных зондов на основе последовательности из двух трипсинизированных последовательностей из СP4 фермента (таблица 3). Зонд, идентифицированный как MID, имел очень низкое вырождение и был использован для первоначального скринирования. Зонды, идентифицированные как EDV-C и EDV-T, были основаны на тех же самых аминокислотных последовательностях и различаются в одном положении (подчеркнуто в таблице 3 ниже) и были использованы, как подтверждающие зонды с положительным результатом только от одного из этих зондов. В олигонуклеотидах ниже альтернативно приемлемые нуклеотиды в конкретном положении обозначены через "/" так, как A/C/T.
Зонды были помечены, используя 32P-ATP и нуклеотид киназу. ДНК из четырнадцати космид, описанных выше, рестриктировали Eco RI, переносили на мембрану и зондировали нуклеотидными зондами. Использованными условиями были следующие: проводят предгибридизацию в 6X SSC, 10X Денхарте в течение 2-18 часов при 60oC, а гибридизацию проводят в течение 48-72 часов в 6X SSC, 10X Денхарте, 100 мкг/мл мРНК при 10oC ниже Td для зонда. Td зонда определяют по формуле 2oC • (A+T) + 4oC • (G+C). Затем фильтры промывают три раза 6X SSC в течение 10 минут каждый раз при комнатной температуре, сушат и ауторадиографируют. При использовании MID зонда фрагмент ≈ 9,9 ко в pMON17076 космиде дает единственный положительный сигнал. Эту космидную ДНК затем зондируют EDV-C (SEQ ID N 22) и EDV-T (SEQ ID N 23) зондами отдельно и снова эта полоса ≈ 9,9 ко дает сигнал и только с EDV-T-зондом.
Объединенные данные по толерантному к глифосату фенотипу, комплементарности E.coli aro-A фенотипа, экспрессии ≈ 45 Кд белка, и гибридизации двумя зондами, происходящими из CP4 EPSPS аминокислотной последовательности, четко подтверждают, что pMON17076 космида содержит EPSPS ген.
Локализация и субклонирование CP4 EPSPS гена
CP4 EPSPS ген далее был локализован следующим образом: был проведен ряд дополнительных анализов по Саузерну различных рестрикционных переваров pMON17076 с использованием MID (SEQ ID N 21) и EDV-T (SEQ ID N 23) зондов отдельно. Исходя из этих анализов и поледующего рестрикционного картирования pBluescript (Стратаген) субклонов фрагмента ≈ 9,9 ко из pMON17076, был идентифицирован фрагмент 3,0 ко EcoRI-SalI, с которым гибридизируются оба зонда. Этот анализ также показывает, что MID (SEQ ID N 21) и EDV-T (SEQ ID N 23) зонды гибридизуются с различными сторонами Bam HI, Cla I и Sac II сайтов. Этот фрагмент 3,8 кв клонируют в обеих ориентациях в pBluescript с образованием pMON17081 и pMON17082. Затем были определены фенотипы E.coli, ухудшенные этими клонами. Толерантность к глифосату была определена после трансформации в E. coli MM294, содержащий pGP1-2 (pBluescript также содержит T7-промотор), на M9 агаровой среде, содержащей 3 мМ глифосата. Как pMON17081, так и pMON17082 показали толерантные к глифосату колонии через 3 дня при 30oC при примерно половине размера контроля на той же среде, лишенной глифосата. Этот результат подтверждает, что фрагмент 3,8 ко содержит интактный EPSPS-ген. Кажущаяся потеря ориентационной зависимости этого фенотипа может быть объяснена наличием T7-промотора на одной стороне сайтов клонирования и lac-промотора на другой. Фенотип aro-A был определен в трансформантах E.coli GB100 на M9 агаровой среде, лишенной ароматических добавок. В этом эксперименте, проведенном с и без Plac-индуктора IPTG, pMON17082 показывает намного больший рост, чем pMON17081, подтверждая, что EPSPS ген был экспрессирован от Sal I-сайта к EcoRI-сайту.
Нуклеотидное секвенирование было начато от ряда концов рестрикционных сайтов, включая Bam HI-сайт, обсужденный выше. Последовательности, кодирующие белковые последовательности, которые близко соответствуют N-концевой белковой последовательности и последовательности трипсинированного фрагмента 53-28 (SEQ ID N 20) (основа EDV-T зонда) (SEQ ID N 23), были локализованы с SalI стороны этого Bam HI сайта. Эти данные обеспечивают убедительное доказательство для клонирования CP4 EPSPS гена и для направления транскрипции этого гена. Эти данные сочетаются с данными по рестрикционному картированию и также указывают, что полный ген был локализован у фрагмента ≈ 2,3 ко XhoI, и этот фрагмент был субклонирован в pBlueScript. Нуклеотидная последовательность почти 2 ко этого фрагмента была определена при сочетании секвенирования клонированных фрагментов рестрикции и при использовании специфических праймеров для расширения последовательности. Нуклеотидная последовательность CP4 EPSPS гена и фланкирующих областей показана на фиг. 3 (SEQ ID N 2). Также была локализована последовательность, соответствующая пептиду 61-24-25 (SEQ ID N 19). Последовательность была определена при использовании как Секвеназного набора из IBI (Интернейшнл Биотекнолоджис Инк.), так и T7 секвенирования (Деаза Кит из Фармасиа).
То, что клонированный ген кодируют EPSPS активность, очищенную от Agrobacterium sp. штамм CP4, было подтверждено следующим образом. С помощью серии сайт-направленных мутагенезов Bgl II и NcoI сайты помещают с N-конца с fMet содержащей внутри NcoI распознаваемой последовательности, первый внутренний NcoI сайт был удален (второй внутренний NcoI сайт был удален позже), а SacI сайт был помещен после стоп колонов. На последней стадии внутренний NcoI сайт был также удален путем сайт-направленного мутагенеза. Следующий список включает праймеры для сайт-направленного мутагенеза (дополнение или удаление рестрикционных сайтов) CP4 EPSPS гена. Мутагенез проводят с помощью методик Кункеля с сотр. (1987), в основном описанных в Sambrook et al. (1989).
Праймер BgNc (дополнение BglI и NcoI сайтов к N-концу)
CGTGGATAGATCTAGGAAGACAACCATGGCTCACGGTC (SEQ ID N 24)
Праймер Sph 2 (дополнение SphI сайта к N-концу)
GGATAGATTAAGGAAGACGCGCATGCTTCACGGTGCAAGCAGCC (SEQ ID N 25)
Праймер SI (дополнение SacI сайта непосредственно после стоп кодонов)
GGCTGCCTGATGAGCTCCACAATCGCCATCGATGG (SEQ ID N 26)
Праймер N 1 (удаление внутреннего NotI сайта распознавания)
CGTCGCTCGTCGTGGGTGGCCGCCCTGACGGC (SEQ ID N 27)
Праймер NcoI (удаление первого внутреннего NcoI сайта распознавания)
CGGGCAAGGCCATGCAGGCTATGGGCGCC (SEQ ID N 28)
Праймер Nco2 (удаление второго внутреннего NcoI сайта распознавания)
CGGGCTGCCGCCTGACTATGGGCCTCGTCGG (SEQ ID N 29)
Этот CP4 EPSPS ген затем клонируют в виде NcoI-BamHI N-концевого фрагмента плюс BamHI-SacI C-концевой фрагмент в PrecA-gene 10L-вектор экспрессии, подобный вектору, описанному (Wong et al., 1988; Olins et al., 1988) с образованием pMON17101. Km для PEP и Ki для глифосата были определены для EPSPS активности в сырых лизатах трансформантов pMON17101/GB100 с последующей индукцией налидиксиновой кислотой (Wong et al., 1988) и было найдено, что они являются одинаковыми с теми, что были определены для препаратов очищенного и сырого фермента из Agrobacterium sp. штамма CP4.
Характеристика EPSPS гена из Achromobacter sp. штамм LBAA и из Pseudomonas sp. штамм PG292.
Космидный банк из частично Hind III-рестриктированной LBAA ДНК был сконструирован в E.coli MM294 в векторе pHC79 (Hohn and Collins, 1980). Этот банк зондируют CP4 EPSPS генным зондом полной длины путем гибридизации колоний и идентифицируют положительные клоны в количестве ≈ 1 на 400 космид. Далее LBAA EPSPS ген был локализован в этих космидах с помощью анализа по Саузерну. Ген локализован на ≈ 2,8 ко XhoI фрагменте и путем серии стадий секвенирования как из рестрикционных концов фрагмента, так и при использовании олигонуклеотидных праймеров из секвенирования CP4 EPSPS гена нуклеотидная последовательность LBAA EPSPS гена была закончена и представлена на фиг. 4 (SEQ ID N 4).
Также был клонирован EPSPS ген из PG2982. EPSPS белок был очищен, в основном как описано для CP4 фермента, со следующими отличиями: После колонки с Сефарозой CL-4B собирают фракции с наиболее высокой активностью и осаждают белок при добавлении твердого сульфата аммония до 85% насыщения и перемешивании в течение 1 часа. Осажденный белок собирают центрифугированием, повторно суспендируют в Q-Сефарозном буфере и подвергают диализу против того же буфера, а затем загружают в колонку (как для CP4 фермента). После очистки на колонке с Q-Сефарозой примерно 40 мг белка в 100 мМ Трис, pH 7,8, 10% глицерина, 1 мМ ЕДТА, 1 мМ ДТТ и 1 М сульфата аммония загружают в колонку с Фенил Суперозой (Фермасиа). Колонку элюируют со скоростью 1,0 мл/мин 40 мл градиента от 1,0 М до 0,00 М сульфата аммония в приведенном выше буфере.
Приблизительно 1,0 мг белка из активных фракций с колонки с Фенил Суперозой 10/10 загружают в Фармасиа Моно P 5/10 Хроматофокусирующую колонку со скоростью потока 0,75 мл/мин. Исходный буфер представляет собой 25 мМ бис-Трис при pH 6,3 и элюируют колонку 39 мл Полибуфера 74, pH 4,0. Приблизительно 50 мкг пиковой фракции из хроматографирующей колонки диализуют в 25 мМ бикарбонате аммония. Затем этот образец используют для определения N-концевой аминокислотной последовательности.
Полученная N-концевая последовательность представляет собой XHSASPKPATARRSE (где X = неидентифицированный остаток) (SEQ ID N 30). Ряд дегенеративных олигонуклеотидных зондов был сконструирован на основе этой последовательности и использован для зондирования библиотеки PG2982 частично Hind III ДНК в космиде pHC79 (Hohn and Collins, 1980) при гибридизации колоний в нестрогих условиях. Условия конечной промывки были 15 минут с IX SSC, 0,1% SDS при 55oC. Один зонд с последовательностью GCGGTBGCSGGYTTSGG (где B=C, G или T; S = C или G, а Y = C или T) (SEQ ID N 31) идентифицировал группу космидных клонов.
Космидная группа, идентифицированная этим путем, была составлена из космид с различными Hind III фрагментами. Однако при зондировании этой группы CP4 EPSPS генным зондом была идентифицирована космида, содержащая PG2982 EPSPS ген (обозначена как космида 9C1 первоначально, а позже как pMON20107). С помощью серий рестрикционного картирования и анализа по Саузерну этот ген был локализован в XhoI фрагмент ≈ 2,8 ко и была определена нуклеотидная последовательность этого гена. Эта последовательность ДНК (SEQ ID N 6) показана на фиг. 5. Не имеется различий в нуклеотидах между последовательностями EPSPS-генов из LBAA (SEQ ID N 4) и PG2982 (SEQ ID N 6). Кинетические параметры обоих ферменетов находятся внутри интервала экспериментальной ошибки.
Ген из PG2982, который придает толерантность к глифосату в E.coli, был секвенирован (Fitzgibbon, 1988; Fitzgibbon and Braymer, 1990). Последовательность PG2982 EPSPS класс II гена непоказывает гомологии к ранее приведенной последовательности, подтверждая, что фенотип толерантности к глифосату в более ранней работе не относится к EPSPS.
Протоколы альтернативного выделения для других структуральных генов EPSPS класса II.
Ряд генов класса II был изолирован и описан здесь. Ясно, что первоначальное клонирование гена, который является геном из CP4, было затруднено из-за низкой степени подобия между ферментами и генами класса I и класса II. Однако идентификация других генов была сильно облегчена при использовании этого первого гена в качестве зонда. При клонировании LBAA EPSPS-гена CP4 генный зонд позволяет быструю идентификацию космидных клонов и локализацию интактного гена в маленький фрагмент рестрикции, а некоторые из CP4 секвенирующих праймеров также были использованы для секвенирования LBAA (и PG2982) EPSPS-гена(ов). CP4-генный зонд также был использован для подтверждения клона PG2982 гена. Высокая степень подобия класса II EPSPS генов может быть использована для идентификации и клонирования дополнительных генов почти таким же путем, как были использованы зонды из EPSPS-генов класса I для клонирования других генов класса I. Примером последнего было клонирование A. thaliana EPSPS-гена с использованием P.hybrida гена в качестве зонда (Klee et al., 1987).
Толерантная к глифосату EPSPS активность указывалась ранее для EPSPS-синтаз из ряда источников. Эти ферменты не были охарактеризованы по какой-либо степени в большинстве случаев. Применение генных зондов EPSPS-класса I и класса II или антител-зондов обеспечивает быстрые средства начального скрининга природных EPSPS и обеспечивает средства для быстрого клонирования и характеристики генов таких ферментов.
Два из трех описанных генов были изолированы из бактерий, которые были выделены из обработанных глифосатом установок (штамм CP4 и LBAA). Третий (PG2982) был из бактерии, которая была выделена из культуры коллекционного штамма. Эта последняя изоляция подтверждает, что экспонирование глифосатом может быть необязательным для выделения высоко толерантных к лифосату EPSPS-ферментов и что скринирование коллекций бактерий может дать дополнительные изоляты. Можно обогатить разлагающие глифосат или устойчивые к глифосату микробные популяции (Quinn et al. 1988; Talbot et al., 1984) в случаях, когда получают, что обогащение для таких микроорганизмов будет увеличивать частоту выделения микроорганизмов EPSPS класса II. Также были идентифицированы дополнительные бактерии, содержащие EPSPS-ген класса II. Бактерия, названная C12, изолированная из шариков той же обработанной колонки как CP4 (см. выше), но в среде, в которую был введен глифосат как источник углерода и фосфора, было показано с помощью анализа по Саузерну, что она гибридизуется с зондом, состоящим из CP4 EPSPS кодирующей последовательности. Этот результат в сочетании с результатом для штамма LBAA подтверждает, что этот метод обогащения облегчает идентификацию изолятов EPSPS класса II. Новые бактериальные изоляты, содержащие гены EPSPS класса II, также были идентифицированы из окружающих сред, других чем установки, обработанные глифосатными сбросами. Был приготовлен инокулюм при экстракции почвы (из недавно убранных соевых полей в Джерсивилле, Иллинойс) и популяция бактерий, отобранных по росту при 28oC на среде Дворкина-Фостера, содержащей 10 мМ глифосата в качестве источника углерода (и 100 мкг/мл циклогексилимида для предотвращения роста грибов). При выращивании на L-агаровой среде было идентифицировано пять типов колоний. Готовят хромосомальную ДНК из 2 мл L-бульонных культур этих изолятов и было прозондировано наличие EPSPS-гена класса II с использованием зонда - кодирующей последовательности CP4 EPSPS анализом по Саузерну при жестких условиях гибридизации и промывки. Один из почвенных изолятов, S2, явился положительным при таком скрининге.
Соотношение между различными EPSPS-генами
Сравнивали уменьшенные аминокислотные последовательности ряда EPSPS ферментов класса I и класса II, используя компьютерную программу Бестфита, поданную в UWGCG упаковке (Devereux et al., 1984). Приведена степень подобия и идентичность как определены с использованием этой программы. Степень подобия (идентичность, определенные внутри последовательностей белков класса I и класса II, является очень высокой, например, при сравнении E.coli с S. typhimurium (подобие) идентичность = 93% (88%) и даже при сравнении E.coli с растительным EPSPS (Petunia hybrida; 72%) (55%). Эти данные приведены в таблице 4. Сравнение последовательностей между классом I и классом II, однако, показывает только очень низкую степень родства между классами (подобие) идентичность = 50-53% (23-30%). Данные анализа по Бестфиту для E.coli (SEQ ID N 8) и CP4 (SEQ ID N 3) последовательностей показывают положения сохраненных остатков и представлены на фиг. 6. Предыдущие анализы EPSPS последовательностей отметили высокую степень сохранения последовательностей ферментов и почти инвариантность последовательностей в двух областях - областях "20-35" и "95-107" (Gasser et al., 1988; пронумерованные в соответствии с EPSPS последовательностью петунии) - и эти области являются менее сохраненными в случае CP4 и LBAA при сравнении последовательностей бактериального EPSPS класса I и растительного EPSPS (см. фиг. 6 для сравнения последовательностей EPSPS E. coli и CP4 с последовательностью E.coli, приведенной в виде верхней последовательности на фиг. Соответствующими последовательностями в CP4 EPSPS класса II являются:
PGDKSISHRSFMFGGL (SEQ ID N 32) и LDFGNAATGCRLT (SEQ ID N 33).
Эти сравнения показывают, что общее родство белков EPSPS класса I и класса II является низким и что последовательности в мнимо сохраненных областях также значительно расходятся.
В CP4 EPSPS аланиновый остаток находится в положении "глицин 101". Замещение сохраненного глицина (из области "95-107") на остаток аланина приводит в результате к повышенному Ki для глифосата и к повышению Km для PEP в EPSPS класса I. В случае CP4 EPSPS, который содержит аланин в этом положении, Km для PEP находится в нижней области, указывая, что ферменты класса II отличаются во многих аспектах от ферментов EPSPS, охарактеризованных ранее.
Внутри изолятов класса II степень подобия/идентичности является такой же высокой, как отмечено для этого соотношения внутри класса I (таблица 4). На фиг. 7 показано регулирование компьютерной программы Бестфита CP4 (SEQ ID N 3) и LBAA (SEQ ID N 5) EPSPS уменьшенных аминокислотных последовательностей с CP4 последовательностью, приведенной как верхняя последовательность на фиг. Символы, использованные на фиг. 6 и 7, являются стандартными символами, использованными в компьютерной программе Бестфита для обозначения степени подобия и идентичности.
Однако различие, которое может быть отмечено между уменьшенными аминокислотными последовательностями CP4 и LBAA EPSPS белков, заключается в положении 100, где находят аланин в случае фермента CP4 и глицин в случае LBAA фермента. В ферментах EPSPS класса I глицин обычно находят в эквивалентном положении, т.е. глицин 96 в E.coli и K.pneumoniae и глицин 101 в Petunia. В случае этих трех ферментов сообщалось, что превращение этого глицина в аланин приводит в результате к повышению app Ki для глифосата и сопутствующего увеличения app Km для PEP (Kishore et al., 1986; Kishore and Shan, 1988; Sost and Amrhein, 1990), что, как обсуждалось выше, делает ферменты менее эффективными, особенно в условиях более низких концентраций PEP. Глицин 100 в EPSPS LBAA превращают в аланин и как app Km для PEP, так и app Ki для глифосата были определены для варианта. Замена глицина 100 на аланин была вызвана мутагенезом с использованием следующего праймера:
CGGCAATGCCGCCACCGGCGCGCGCC (SEQ ID N 34) и гены как дикого типа, так и варианта были экспрессированы в E.coli в RecA-промоторе вектора экспрессии (pMON17201 и pMON17264 соответственно) и в сырых лизатах были определены app Km и app Ki. Данные указывают, что app Ki (глифосат) для G100A варианта повышается примерно в 16 раз (таблица 5). Этот результат находится в согласии с наблюдением о важности этой G-A замены при возрастании app Ki (глифосат) в EPSPS ферментах класса I. Однако в противоположность результатам для G-A вариантов класса I app Km (PEP) для варианта G-A класса II (LBAA) остается неизменным. Это обеспечивает еще одно различие между EPSPS ферментами класса II и класса I.
LBAA G100A-вариант благодаря его превосходным кинетическим свойствам способен к приданию улучшения глифосату в растении.
Модификация и повторный синтез последовательности EPSPS-гена из штамма CP4 Agrobacterium sp.
EPSPS-ген из Agrobacterium sp. штамм CP4 содержит последовательности, которые могут быть inimical к высокой экспрессии гена в растениях. Эти последовательности включают сайты потенциального аденилирования, которые часто являются богатыми A + T, и имеют более высокую G+C%, чем часто находимую в генах растений (63% по сравнению с 50%), концентрированные протяженности G- и C-остатков, и кодоны, которые часто не используются в генах растений. Высокая G + C% в CP4 EPSPS гене имеет ряд потенциальных следствий, включая следующие: более высокое использование G или C, чем было найдено в генах растений в третьей позиции в кодонах, и потенциал для образования прочной hair-pin структуры, которая может воздействовать на экспрессию или стабильность РНК. Снижение содержания G + C в CP4 EPSPS-гене, перерыв в протяженности G или C, удаление потенциальных последовательностей полиаденилирования и улучшения в применении кодона, что более часто используется в растительных генах, может в результате привести к более высокой экспрессии CP4 EPSPS-гена в растениях.
Синтетический CP4 ген предназначен для замены по возможности полностью этих inimical последовательностей, обсужденных выше. В общем, была снова сконструирована генная последовательность для удаления по возможности больше следующих последовательностей или характерных признаков последовательности (избегая при этом введения необходимых сайтов рестрикции): протяженности G и C от 5 или более; и богатых A+T областей (преимущественно), которые могут функционировать в качестве сайтов полиаденилирования или область потенциальной дестабилизации РНК. Последовательность этого гена показана на фиг. 8 (SEQ ID N 9). Эта кодирующая последовательность экспрессирована в E.coli от RecA-промотора и проанализирована на EPSPS-активность и ее сравнили с последовательностью нативного CP4 EPSPS-гена. Кажущаяся Km для PEP для нативного и синтетического генов составила 11,8 и 12,7 соответственно, указывая, что фермент, экспрессированный из синтетического гена, является неизменным. N-конец кодирующей последовательности был мутагенезирован, чтобы поместить SphI сайт при ATG, чтобы смочь сконструировать CTP-2-CP4 синтетическое слияние для введения хлоропласта. Был использован следующий праймер для осуществления этого мугагенеза:
GGACGGCTGCTTGCACCGTGAAGCATGCTTAAGCTTGGCGTAATCATGG (SEQ ID N 35)
Экспрессия хлоропласт-направленного CP4 EPSPS
Глифосатная мишень в растениях, 5-енолпирувилшикимат-3-фосфатсинтаза (EPSPS) фермент, локализована в протопласте. Многие хлоропласт-локализованные белки, включая EPSPS, экспрессируются из ядерных генов как предшественники и направляются в хлоропласт с помощью хлоропласт транзитного белка (CTP), который удаляется во время стадий введения. Примеры других таких хлоропластных белков включают малую субъединицу Рубилоза-1,5-бисфосфат карбоксилазы (KuBISCO), ферродоксин, ферродоксин оксидоредуктазу, созревающий на свету комплекс белка I и белка II и тиоредоксин F. Было показано in vivo и in vitro, что нехлоропластные белки могут быть нацелены на хлоропласт при использовании слияния белков с CTP и что последовательности CTP достаточно, чтобы направить белок в хлоропласт.
Было сконструировано слияние CTP-CP4 EPSPS между Arabidopsis thaliana EPSPS CTP (Klee et al., 1987) и CP4 EPSPS кодирующими последовательностями. Arabidopsis CTP был сконструирован путем сайт-направленного мутагенеза, чтобы поместить SphI сайт рестрикции на сайт процессинга CTP. Этот мутагенез заменяется Glu-Lys в этом месте на Cys-Met. Последовательность этого CTP, обозначенная как CTP2 (SEQ ID N 10), показана на фиг. 9. N-конец CP4 EPSPS-гена модифицирован для размещения SphI - сайта, который охватывает Met-кодон. Второй кодон на этой стадии также превращен в кодон для лейцина. Эта замена не оказывает заметного влияния на in vivo активность CP4 EPSPS в E. coli, как оценено по скорости комплементации aro A аллеля. Этот модифицированный N-конец затем объединяют с SacI C-концом и клонируют ниже CTP2-последовательностей. CTP2-CP4 EPSPS-слияние клонируют в p Blue Script KS(+). Вектор может быть транскрибирован in vitro с использованием T7-полимеразы и трансляцией PHK 35s-Метионина, чтобы получить материал, который может быть оценен на внесение в хлоропласты, изолированные из Lactuca sativa, с использованием описанных здесь далее методов (della-Cioppa et al. 1986, 1987). Эту матрицу транскрибируют in vitro, используя T7-полимеразу и 35s-метионин-меченый CTP2-CP4 EPSPS-материал, как было показано, входит в хлоропласты с эффективностью, сравнимой с активностью контрольного EPSPS петуньи. (Контроль = 35s-меченый Pre EPSPS /pMON 6140; della-Cioppa et al. 1986)).
В другом примере Arabidopsis EPSPS CTP, обозначенный как CTP3, сливают с CP4 EPSPS через EcoRI-сайт. Последовательность этого CTP3 (SEQ ID N 12) показана на фиг. 10. EcoRI-сайт был введен в зрелую область Arabidopsis EPSPS вокруг аминокислоты 27, заменив последовательность Arg-Ala-Leu-Leu на Arg-Ile-Leu-Leu в процессе. Праймер следующей последовательности был использован для модификации N-конца CP4 EPSPS-гена, чтобы добавить EcoRI-сайт для воздействия на слияние с CTP3:
GGAAGACGCCCAGAATTCACGGTGCAAGCAGCCGG (SEQ IД и 36)
(EcoRI-сайт подчеркнут).
Это CTP3-CP4 EPSPS-слияние также было клонировано в вектор BlueScript и T7-экспрессированное слияние, как было найдено, также введено в хлоропласты с эффективностью, сравнимой с эффективностью контрольного EPSPS петунии (pMON 6140).
Родственные серии CTP, обозначенные как CTP (SphI) и CTP5 (EcoRI), основанные на EPSPS CTP петунии, и ген также были слиты с SphI- и EcoRI модифицированными последовательностями CP4 EPSPS-гена. SphI-сайт был добавлен при сайт-направленном мутагенезе, чтобы поместить этот сайт рестрикции (и заменить аминокислотную последовательность на -Cys-Met-) в сайте процессинга хлоропласта. Было показано, что все CTP-CP4 EPSPS-слияния вносят в хлоропласты примерно равную эффективность. Последовательности CTP4 (SEQ ID N 14) и CTP5 (SEQ ID N 15) показаны на фиг. 11 и 12.
Слияние CTP2-LBAA EPSPS также было сконструировано после модификации N-конца LBAA EPSPS-гена при введении SphI-сайта. Также было найдено, что это слияние входит эффективно в хлоропласты. С помощью подобных попыток также было показано, что слияния CTP2-CT4 EPSPS и CTP4-CP4 EPSPS эффективно входят в хлоропласты, приготовленные из листовых оболочек кукурузы. Эти результаты показывают, что эти слияния CTP-CP4 также могут обеспечить полезные гены, придавая толерантность к глифосату однодольным видам.
Специалистам в данной области должно быть понятно, что могут быть сделаны различные химерические конструкции, которые используют функциональность конкретного CTP для внесения EPSPS-фермента класса II в хлоропласт клетки растения. Хлоропласт с внесенным EPSPS класса II может быть определен с использованием следующего анализа.
Быстрый анализ хлоропластов
Изолируют интактные хлоропласты из латука (Latuca sativa, сорт длиннолистый) центрифугированием в градиенте Перколл/фиколл, модифицированном по Барлетту с сотр. (1982). Конечный шарик интактных хлоропластов суспендируют в 0,5 мл стерильного 330 мМ сорбита в 50 мМ Гепес-КОН, pH 7,7, анализируют на хлорофилл (Arnon, 1949) и устанавливают конечную концентрацию хлорофилла 4 мг/мл (используя сорбит/Гепес). Выход интактных хлоропластов из одной головки латука составляет 3-6 мг хлорофилла.
Типичный образец для анализа 300 мкл содержит 5 мМ ATP, 8,3 мМ немеченного метионина, 322 мМ сорбита, 58,3 мМ Гепес-КОН (pH 8,0), 50 мкл продуктов трансляции ретикулоцитного лизата и интактные хлоропласты из L. sativa (200 мкг хлорофилла). Поглощающую смесь осторожно встряхивают при комнатной температуре (в стеклянных трубках 10х75 мм) непосредственно перед иллюминатором из оптического волокна, настроенным на максимум интенсивности света (лампа 150 Вт). Аликвотные образцы поглощающей смеси (около 50 мкл) отбирают через различные промежутки времени и фракционируют на 100 мкл градиентах силикон-масло (в полиэтиленовых трубках на 150 мкл) центрифугированием при 11000g в течение 30 секунд. В этих условиях интактные хлоропласты образуют шарик на слое масло-силикон и инкубационная среда (содержащая ретикулоцитный лизат) всплывает на поверхность. После центрифугирования градиенты силикон-масло немедленно замораживают на сухом льду. Хлоропластный шарик затем повторно суспендируют в 50-100 мкл лизисного буфера (10 мМ Гепес-КОН, pH 7,5, 1 мМ PMSF, 1 мМ бензамидина, 5 мМ е-амино-н-капроновой кислоты и 30 мкг/мл апротинина) и центрифугируют при 15000g в течение 20 минут, чтобы получить шарик тилакоидных мембран. Смешивают в равных объемах чистый супернатант (стромные белки) из этого опыта и аликвот ретикулоцитлизатной инкубационной среды из каждого эксперимента и образец буфера 2X SDS-PAGE для электрофореза (Леммли, 1970).
SDS-PAGE проводят согласно Леммли (1970) в 3-17% (мас./об) акриламидного плоского геля (60 мм х 1,5 мм) с 3% (мас./об.) акриламидного кускового геля (5 мм х 1,5 мм). Гель фиксируют в течение 20-30 мин в растворе с 40% метанола и 10% уксусной кислоты. Затем гель замачивают в EN3HANCEТМ (Дюпон) на 20-30 минут, затем сушат гель в сушилке для геля. Создают изображение на геле ауторадиографией, используя интенсифицирующий экран и экспозицию в течение ночи, чтобы определить, вошел ли CP4 EPSPS в изолированные хлоропласты.
Трансформация растений
Растения, которые могут быть сделаны толерантными к глифосату в практике настоящего изобретения, включают, но не ограничиваются, сою, хлопок, кукурузу, canola, масличный рапс, лен, сахарную свеклу, подсолнечник, картофель, табак, томаты, пшеницу, рис, люцерну и латук, а также различные виды деревьев, орехов и винограда.
Двуниточная молекула ДНК настоящего изобретения ("химерный ген") может быть вставлена в геном растения любым подходящим методом. Подходящие векторы трансформации растений включают векторы, происходящие из Ti-плазмиды Agrobacterium tumefaciens, а также те, что описаны, например Herrera-Estrella (1983), Bevan (1984), Klee (1985) и в европейской заявке 120516 (Schilperoort et al.). В дополнение к векторам трансформации, происходящим из Ti- или корнеиндуцирующих (Ri) плазмид Agrobacterium, могут быть использованы альтернативные способы вставки конструкции ДНК изобретения в клетки растений. Такие методы могут включать, например, использование липосом, электропорацию, химикаты, которые увеличивают поглощение свободной ДНК, выделение свободной ДНК через микроотверстия, и трансформацию с использованием вирусов или пыльцы.
EPSPS класса II векторы трансформации растений
Последовательности ДНК EPSPS класса II могут быть сконструированы в векторы, способные трансформировать растения при использовании известных методик. Последующее описание является только иллюстративным и не рассматривается в ограничивающем смысле. Средний специалист в данной области должен понимать, что другие плазмиды, векторы, маркеры, промоторы и т.п. могут быть использованы с подходящими результатами. Слияние CTP2-CP4 EPSPS клонируют в виде BglII-EcoRI фрагмента в растительной вектор pMON 979 (описан ниже) с образованием pMON 17110, карта которого представлена на фиг. 13. В этом векторе CP4 ген экспрессирует из усиленного CaMV35S-промотора (E35S,Kay et al, 1987). Конструкцию промотора FMV35S (pMON 17116) заканчивают следующим путем: Лигируют SalI-NotI и NotI-BglII фрагменты из pMON 979, содержащей Spc /AAC/ 3/-III/ oriV и pBR 322 /правый край /NOS3'/ CP4 EPSPS-генный сегмент из pMON 17110 с фрагментом промотора XhoI-BglII FMV35S из pMON 981. Эти векторы вводят в табак, хлопок и канолу (canola).
Также комплектуют серии векторов в векторе pMON 977, в который клонируют CP4 EPSPS ген, CTP2-CP4 EPSPS-слияние и CTP3-CP4-слияние в виде фрагментов BglII-SacI с образованием pMON 17124, pMON 17119 и pMON 17120 соответственно. Эти плазмиды вводят в табак. Также комплектуют производное pMON 977, содержащее CTP2-LBAA EPSPS-ген (pMON 17206) и вводят в табак.
Вектор трансформации/экспрессии в растениях происходит из pMON 886 (описан ниже) при замене гена неомицин фосфотрансферазы тип II (KAN) в pMON 886 на фрагмент 0,89 ко, содержащий ген бактериальный гентамицин-3-N-ацетилтрансферазы тип III /AAC/3/-III/ (Hayford et al. 1988). Химерный P-35S /AA/3/-III-NOS 3'-ген кодирует устойчивость к гентамицину, что позволяет провести селекцию трансформированных клеток растений, pMON 979 также содержит 0,95 ко полигенный экспрессирующий кластер, состоящий из усиленного CaMV35S промотора (Kay et al., 1987), несколько уникальных сайтов рестрикции и NOS 3'-конец (P-En-CaMV35S/ NOS 3'). Остальные pMON979 ДНК-сегменты являются точно такими же, как в pMON 886.
Плазмида pMON 886 составлена из следующих сегментов ДНК. Первый представляет собой 0,93 ко AvaI инженерный EcoRV-фрагмент, изолированный из транспозона Тп7, который кодирует бактериальную спектиномицин/стрептомициновую устойчивость (Spc /Str), которая является определяющей для селекции в E. Coli и Agrobacterium tumefaciens. Они объединены в 1,61 ко сегмент ДНК, кодирующий химерную устойчивость к канамицину, что позволяет провести селекцию трансформированных клеток растений. Химерный ген (P-35S/KAN/NOS 3'/ состоит из (CaMV) 35S-промотора вируса мозаики подсолнечника, гена неомицин фосфотрансферазы тип II (KAN), и 3'-нетранслируемой области гена нопалин синтазы (NOS3') (Fraley et al., 1983). Следующий сегмент представляет собой 0,75 ко oriV, содержащий с источником репликона из RK2 плазмиды. Он соединен в 3,1 ко SalI до PvuI сегмент pBR322 (ori 322), который обеспечивает источник репликона для сохранения в E.coli и bom сайт для конъюгационного переноса в клетки Agrobacterium tumefaciens. Следующий сегмент является 0,36 ко Pvu I до Bcl I от Ti T37, который несет правый край нопалин-тип T-ДНК (Fraley et al., 1985).
Вектор pMON977 является таким же, как pMON981, за исключением наличия P-En-CAMV35S-промотора вместо FMV35S-промотора (смотри ниже).
Плазмида pMON981 содержит следующие сегменты ДНК: 0,93 кв фрагмент, изолированный из транспозона Tn7, кодирующий бактериальную спектиномицин/стрептомициновую устойчивость (Spc) Str, определяющий для селекции в E. coli и Agrobacterium tumefaciens (Fling et al., 1985)), химерный ген устойчивости к канамицину инженерный для экспрессии в растении, чтобы можно было провести селекцию трансформированной ткани, состоящей из 0,35 ко 35S промотора вируса мозаики подсолнечника (P-35S) (Odell et al. 1985), 0,83 ко ген неомицин фосфотрансферазы тип II-КАN/ и 0,26 ко 3'-нетранслируемая область гена нопалин синтазы (NOS 3') (Fraley et al., 1983). 0,75 ко ориджин репликации из RK2-плазмиды (oriV/stalker et al., 3,1 ко SalI до PvuI сегмента pBR 322), который обеспечивает ориджин репликации для сохранения в E.coli (ori-322) и bom-сайт для конъюгированного переноса в клетки Agrobacterium tumefaciens и 0,36 ко фрагмента из T T37 плазмиды, содержащей область правого края нопалин-типа T-ДНК (Fraley et al., 1985). Полигенный экспрессирующий кластер состоит из 0,6 ко 35S промотора из вируса мозаики норичника (P-FMV35S) (Gowda et al. , 1989) и 0,7 ко 3'-нетранслируемой области гена гороха rbcS-E9 (E9 3') (Coruzzi et al., 1984, и Morelli et al., 1985). Фрагмент 0,6 ко SspI, содержащий FMV35S промотор (фиг. 1) был подстроен к подходящему месту сайтов клонирования вниз от стартового сайта транскрипции. Слитый CTP2-CP4 syn ген был введен в растительные векторы экспрессии (включая pMON981, для образования pMON17131; фиг. 14) и трансформированы в табак, канолу, картофель, томаты, сахарную свеклу, хлопок, латук, огурцы, масличный рапс, тополь и Arabidopsis.
Растительный вектор, содержащий EPSPS ген класса II, может быть мобилизован в любой подходящий штамм Agrobacterium для трансформации желаемых видов растений. Растительный вектор может быть мобилизован в штамм ABI Agrobacterium. Подходящим штаммом ABI является A208 Agrobacterium tumefaciens, несущий обезоруженную Ti-плазмиду pTiC58 (pMP90RK) Konez and Schell, 1986). Ti-плазмида не должна вести T-ДНК фитогормональных генов и, следовательно, штамм не способен вызвать заболевание корневым раком. Скрещивание растительного вектора в ABI было сделано с помощью трипарентеральной системы конъюгации с использованием хелперной плазмиды pRK2013 (Ditta et al., 1980). При инкубации растительной ткани с ABI: конъюгатом растительного вектора вектор переносится в клетки растений с помощью vir функций, кодируемых обезвреженной pTiC58-плазмидой. Вектор открыт с области прямого края T-ДНК и полная векторная последовательность может быть вставлена в хромосому растения-хозяина. pTiC58 Ti-плазмида не переносится в клетки растения, а остается в Agrobacterium.
Векторы EPSPS класса II свободной ДНК
Гены EPSPS класса II также могут быть введены в растения с помощью методов прямого высвобождения. Для CP EPSPS-гена было составлено несколько векторов прямого высвобождения. Вектор pMON13640, карта которого представлена на фиг. 15, описан здесь. Плазмидный вектор основан на pUC-плазмиде (Vieira and Messing, 1987), содержащей в этом случае nptII ген (устойчивость к канамицину, KAN) из Tn903, чтобы обеспечить селектирующий маркер в E. coli. Слитый CTP4-EPSPS ген экспрессируется из p-FMV35S промотора и содержит NOS3'-фрагмент последовательности полиаденилирования и из второго полигенного экспрессирующего кластера, состоящей из E35S-промотора, слитого CTP4-CP4 гена и NOS3'-последовательностей. Апробированный GUS маркерный ген (Jefferson et al. , 1987) экспрессируется из маннопин синтазного промотора (P-MAS, Velten et al. , 1984) и 3'-последовательностей гена 7S запасного белка сои (Schuler et al., 1982). Также могут быть получены подобные плазмиды, в которых слитые CTP-CP4 EPSPS были экспрессированы из усиленного CaMV35S-промотора или других растительных промоторов. Могут быть получены другие векторы, которые являются пригодными для высвобождения свободной ДНК в растениях, и они известны специалистам в этой области и считается, что они входят в область настоящего описания.
Регенерация растений
При достижении экспрессии EPSPS-гена класса II в трансформированных клетках (или протопластах), клетки (или протопласты) регенерируют в целое растение. Выбор методологии для стадии регенерации не является критическим, подходящие протоколы доступны для хозяев из Leguminosae (люцерна, соя, клевер и т. п. ), Umbelliferae (морковь, сельдерей, пастернак), Cruciferae (капуста, редис, рапс и т. п.), Cucurbitaceae (дыня и огурец), Gramineae (пшеница, рис, кукуруза и т.п.), Solanaceae (картофель, табак, томаты, перец), различные цветочные культуры, а также различные деревья, такие как тополь и яблоня, ореховые культуры или виноградные растения, такие как виноград. Смотри, например, Ammirato, 1984, Shimamoto, 1989, Fromm 1990, Vasil, 1990.
Следующие примеры приведены для лучшего понимания практики настоящего изобретения и не должны интерпретироваться каким-либо образом для ограничения области настоящего изобретения. Специалисты в данной области должны понимать, что различные модификации, сокращения и т.п. могут быть сделаны в способах и генах, описанных здесь, не выходя при этом из духа и области настоящего изобретения.
В приведенных ниже примерах EPSPS-активность в растениях была проанализирована следующим способом. Образцы тканей собирают и немедленно замораживают в жидком азоте. Замораживают в ступке 1 г молодой листовой ткани и измельчают ее пестиком в тонкий порошок. Затем порошок переносят во вторую ступку, добавляют 1 мл/г экстракционного буфера и измельчают порошок еще 45 секунд. Экстракционный буфер для канолы состоит из 100 мМ Трис, 1 мМ EDTA, 10% глицерина, 5 мМ DTT, 1 мМ BAM, 5 мМ аскорбата, 1,0 мг/мл BSA, pH 7,5 (4oC). Экстракционный буфер для табака состоит из 100 мМ Трис, 10 мМ EDTA, 35 мМ KCl, 20% глицерина, 5 мМ DTT, 1 мМ BAM, 5 мМ аскорбата; 1,0 мг/мл BSA, pH 7,5 (4oC). Смесь переносят в микрофужную трубку и центрифугируют 5 минут. Обессоливают полученные в результате супернатанты на спин G-50 колонках (Фармасиа), заранее уравновешенных экстракционным буфером (без BSA), в аликвотах 0,25 мл. Обессоленные экстракты анализируют на EPSP синтазную активность с помощью радиактивного анализа ВЭЖХ. Концентрацию белка в образцах определяют с помощью БиоРад микробелкового анализа, используя в качестве стандарта BSA.
Концентрации белка определяли, используя БиоРад микробелковый метод. Используют BSA для получения стандартной кривой в интервале 2-24 мкг. Смешивают 800 мкл стандартного или разбавленного образца с 200 мкл концентрированного реагента БиоРад Бредфорд. Образцы интенсивно перемешивают и считывают при A(595) после 5 минут и сравнивают со стандартной кривой.
Аналитический набор для EPSPS фермента содержит 50 мМ ГЕПЕС, 2 мМ шикимат-3-фосфата, 0,1 мМ молибдата аммония и 5 мМ KF с добавкой 0,5 или 1,0 мМ глифосата или без него. Предварительно инкубируют 30 мкл аналитической смеси и 10 мкл растительного экстракта в течение 1 минуты при 25oC и инициируют реакции добавлением 1 мМ 14C-PEP. Через 3 минуты реакции закаливают 50 мкл 90% EtOH/0,1 М HOAc, pH 4,5. Образцы центрифугируют при 6000 об/мин и анализируют полученные в результате супернатанты на 14C-EPSP продукцию с помощью ВЭЖХ. Процент устойчивого EPSPS вычисляют из EPSPS активностей с и без глифосата.
Определяют процент конверсии 14C-меченого PEP в 14C EPSP с помощью радиоанализа ВЭЖХ с C18 предохранительной колонкой (Броунли) и AX100 ВЭЖХ колонкой (0,4 х 25 см, Синхропак), элюентом является 0,28 М изократический фосфат калия, pH 6,5 при
1 мл/мин. Рассчитывают начальные скорости путем умножения фракционных оборотов в единицу времени при начальной концентрации меченого субстрата (1 мМ). Анализ был линейный при времени до 3 минут и 30% превращения в EPSPS. Образцы разбавляют 10 мМ Трис 10% глицерина, 10 мМ DTT, pH 7,5 (4oC), если необходимо для получения результатов внутри линейного интервала.
Для этих анализов DL-дитиотейтол (DTT), бензамидин (BAM) и бычий сывороточный альбумин (BSA), практически чистые от глобулина, были получены от Сигмы. Фосфоенолпируват (PEP) был получен от Боерингер Маннхейм, а фосфоенол-(1-14C)пируват (28 мКю/ммоль) был получен из Амерсхайма.
Пример 1. Трансформированные растения табака были генерированы с рядом векторов гена EPSPS класс II, содержащим CP4 EPSPS последовательность ДНК, как описано выше, с подходящей экспрессией EPSPS. Эти трансформированные растения показывают толерантность к глифосату, придаваемую CP4 EPSPS класса II.
Для трансформации табака применяют протокол трансформации листового диска табака, в котором использована здоровая листовая ткань примерно месячного возраста. После 15-20 минут стерилизации поверхности 10% хлороксом плюс поверхностно-активное вещество листья промывают 3 раза стерильной водой. Используя стерильные бумажные штампы, листовые диски прижимают и помещают верхней стороной вниз на среду MS104 (MS соли 4,3 г/л, сахароза 30 г/л, B5 витамины 500X 2 мл/л, NAA 0,1 мг/л и BA 1,0 мг/л) на один день прекультуры.
Затем диски инокулируют ночной культурой обезвреженного ABI штамма Agrobacterium, содержащего целевой вектор, которая была разбавлена 1/5 (т.е. примерно 0,6 ОД). Инокулирование проводят при помещении дисков в центрифужные трубки с культурой. После 30-60 секунд жидкость сцеживают и диски промокают между стерильной фильтровальной бумагой. Затем эти диски помещают верхней стороной вниз на пластины с MS104 питательной средой с фильтровальным диском для сокультуры.
После 2-3 дней сокультуры диски переносят, еще верхней стороной вниз, для селекции пластин с MS104-средой. Через 2-3 недели образуется каллюсная ткань и индивидуальные скопления отделяют от листового диска. Ростки стерильно отрезают от каллюса, когда они становятся достаточно большими, чтобы отличаться от стеблей. Ростки помещают на не содержащую гормонов среду для укоренения (MSO: MS соли 4,3 г/л, сахароза 30 г/л и B5 витамины 500X 2 мл/л/ с отбором на устойчивость к соответствующему антибиотику. Образование корней происходит через 1-2 недели. Любой анализ листового каллюса предпочтительно проводить на укорененных ростках, пока еще стерильных. Укорененные ростки затем помещают в почву и выдерживают в окружающей среде с высокой влажностью (например, пластиковых контейнерах или мешочках). Ростки затвердевают при постепенной выдержке их в условиях комнатной влажности.
Экспрессия CP4 EPSPS-белка в трансформированных растениях
Клетки табака трансформировали рядом растительных векторов, содержащих нативный CP4 EPSPS-ген, и используя различные промоторы и/или CTP. Предварительным доказательством экспрессии гена является способность листовой ткани из отобранных на антибиотике трансформированных ростков снова давать каллюс на глифосате. В некоторых случаях толерантный к глифосату каллюс отбирают непосредственно после трансформации. Определяют уровень экспрессии CP EPSPS по уровню EPSPS-активности, толерантной к глифосату (анализируют в присутствии 0,5 мМ глифосата) или с помощью блот-анализа по Вестерну при использовании козьих анти-CP4 EPSPS-антител. Делают количественную оценку пятен по Вестерну с помощью денситометрического копирования и сравнения ее стандартной кривой, установленной для очищенного CP4 EPSPS. Эти данные представлены как % растворимого белка листа. Данные от ряда трансформированных линий растений и векторов трансформации представлены в таблице 6 ниже.
Толерантность к глифосату также была показана на уровне целого растения в трансформированных растениях табака. Для табака R0-трансформанты CTP2-CP4 EPSPS опрыскивали 0,4 фунт/акр (0,448 кг/га) дозой, достаточной для уничтожения контрольных нетрансформированных растений табака, соответствующей оценкам 3, 1 и 0 в день 7, 14 и 28 соответственно, и проанализированы вегетативно и продуктивно (таблица 7).
Пример 2.
Растения канолы трансформировали векторами pMON17110, pMON17116 и pMON17131, и был получен ряд линий растений трансформированной канолы, которые обладают толерантностью к глифосату.
Растительный материал
Ростки Brassica napus сорт Вестар помещали в 2-дюймовые (≈ 5 см) горшочки, содержащие Метро Микс 350. Их выращивали в камере роста при 24oС и фотопериоде 16/8 часов, интенсивности света 400 uEm-2 сек-1 (HID лампы). Они были удобрены Петерс 20-10-20 для общеспециальной цели. Через 2,5 недели их переносили в 6-дюймовые (≈ 15 см) горшочки и выращивали в ростовой камере при 15oC (10oC) температурах день/ночь, фотопериоде 16/8 часов, интенсивности света 800 uEm-2 сек-1 (HID лампы). Их удобряли Петерс 15-30-15 Хи-Фос-Спесиал.
Трансформация/Селекция/Регенерация
Четыре концевых междоузлия от растений почти перед выбросом цветоноса или в процессе выброса цветоноса, но перед цветением удаляют и поверхностно стерилизуют в 70%-м объем/объем этаноле в течение 1 минуты, 2% масс./объем гипохлорита натрия в течение 20 минут и 3 раза промывают стерильной деионизированной водой. Стебли с прикрепленными к ним листьями могут быть помещены в холодильник на 72 часа перед стерилизацией. Шесть-семь сегментов стебля нарезают на диски 5 мм с Редко Вежетабль Слайсер 200, сохраняя ориентацию базального конца.
Agrobacterium выращивают в течение ночи на качалке при 24oC в 2 мл Луриа бульоне, содержащем 50 мг/л канамицина, 24 мг/л хлорамфеникола и 100 мг/л спектиномицина. Делают разбавление 1:10 в среде MS (Мурашиге и Скуг), получая примерно 9•108 клеток/мл. Это подтверждается показаниями оптической плотности при 660 нм. Диски стебля (экспланты) инокулировали 1 мл Agrobacterium и избыток отсасывали от эксплантов.
Экспланты помещают нижней стороной вниз в пластины петри, содержащие 1/10X стандартных MS-солей, B5-витамины, 3% сахарозы, 0,8% агара, pH 5,7, 1,0 мг/л 6-бензиладенина (BA). На пластины наслаивают 1,5 мл среды, содержащей MS-соли, B5-витамины, 3% сахарозы, pH 5,7, 4,0 мг/л п-хлорфеноксиуксусной кислоты, 0,005 мг/л кинетина и покрывают стерильной фильтровальной бумагой.
После 2-3 дней сокультуры экспланты переносят в глубокие плоские чашки петри, содержащие MS-соли, B5-витамины, 3% сахарозы, 0,8% агара, pH 5,7, 1 мг/л BA, 500 мг/л карбенициллина, 50 мг/л цефотаксима, 200 мл/г канамицина или 175 мг/л гентамицина, для селекции. На каждую пластину помещают семь эксплантов. Через 3 недели их переносят на свежую среду по 5 эксплантов на пластину. Экспланты культивируют в ростовой камере при 25oC при непрерывном освещении (Кул Вайт).
Анализ экспрессии
Через 3 недели ростки отрезают от эксплантов. Инициируют повторный анализ каллюса листьев для подтверждения модификации R0-ростков. Три крошечных кусочка листовой ткани помещают на среду для рекаллюзации, содержащей MS-соли, B5-витамины, 3% сахарозы, 0,8% агара, pH 5,7, 5,0 мг/л BA, 0,5 мг/л нафталинуксусной кислоты (NAA), 500 мг/л карбенициллина, 50 мг/л цефотаксима и 200 мг/л канамицина или гентамицина или 0,5 мМ глифосата. Анализируемый лист инкубируют в ростовой камере в тех же условиях, что и культуру экспланта. Через 3 недели оценивают опыт по рекаллюзации листа на толерантность к гербициду (каллюс или зеленая ткань листа) или чувствительность (отбеленный).
Трансплантация
Во время процедуры боковые побеги погружают в Рутон® и помещают в 2-дюймовые (≈ 5 см) горшочки, содержащие Метро Микс 350, и вносят в замкнутую влажную среду. Их помещают в ростовую камеру при 24oC, 16/8 часах фотопериода, 400 uEm-1 сек-2 (HID лампы) в течение периода отверждения примерно 3 недели.
Семена, собранные от растений R0, представляют собой семена R1, из которых выращивают растения R1. Для оценки толерантности к глифосату растений R0 оценивают их потомство. Поскольку растение R0 предполагается должно быть гемизиготным в каждом вставленном участке, самоопыление приведет в результате к максимальному разделению генотипа в R1. Поскольку каждая вставка действует как доминантный аллель, в отсутствие сцепления и предположив, что только одна гемизиготная вставка требуется для экспрессии толерантности, одна вставка должна разделять 3:1, две вставки 15:1, три вставки 63:1 и т.д. Следовательно, относительно маленькое количество растений R1 нужно вырастить, чтобы найти по крайней мере один устойчивый фенотип.
Семена от растения R0 собирают, обмолачивают и сушат перед выращиванием в опыте с опрыскиванием глифосатом. Были использованы различные методики для выращивания растений для оценки опрыскивания R1. Тесты проводили как в теплице, так и в ростовой камере. Использовали две системы выращивания: 10 см горшки или ящики для выращивания, содержащие 32 или 36 ячеек. Для выращивания использовали почву или Метро 350 плюс три типа медленно выделяющегося удобрения или плант Метро 350. Полив был или сверху в теплицах или снизу (подпочвенный) в ростовых камерах. Удобрение вносят по потребности с поливной водой. Поддерживают температурный режим, подходящий для канолы. Поддерживают 16-часовой фотопериод. В начале цветения растения пересаживают в 15 см горшки для получения семян.
Опрыскивание "партии" заключается в опрыскивании в один и тот же день нескольких групп потомства R1. Некоторые группы так же могут включать оценки других, чем R1 растений. Каждая группа также включает опрысканные и неопрысканные нетрансгенные генотипы, представляющие генотипы в конкретной группе, которые были предполагаемо трансформированы. Также включенными в группу являются один или более несегрегированных трансформированных генотипов, ранее идентифицированных, как имеющие некоторую устойчивость.
Не опрыскивают два-шесть растений из каждого индивидуального потомства R0 и оставляют в качестве контроля для сравнения и измерения толерантности к глифосату, а также для оценки любой вариабельности, не индуцированной глифосатом. Когда другие растения достигают стадии 2-4 листьев, обычно через 10-20 дней после посадки наносят глифосат в дозах, меняющихся от 0,28 до 1,12 кг/га, в зависимости от целей исследования. Технологии с низкими дозами, использующие малые объемы, были приемлемы. Лабораторный гусеничный опрыскиватель был откалиброван для выдачи дозы, эквивалентной полевым условиям.
Для оценки опрысканных растений на вегетативную устойчивость используют шкалу от 0 до 10. Шкала относится к неопрысканным растением. Более высокие числа между 0 и 10 означают прогрессивно меньшее повреждение по сравнению с неопрысканным растением. Растения оценивают на 7, 14 и 28 день после обработки (ДАТ) или до тех пор, пока не произойдет стрелкование, и линия дает среднюю оценку опрысканных растений внутри семейства растений R0.
Используют шесть целых чисел для количественного описания степени репродуктивного повреждения для глифосата:
0 : Не развиваются цветочные почки
2 : Цветочные почки имеются, но абортируются перед раскрытием
4 : Цветки раскрыты, но нет пыльников, или пыльники не дают впоследствии выйти лепесткам
6 : Стерильные пыльники
8 : Частично стерильные пыльники
10 : Полностью фертильные цветки
Растения оценивают, используя эту шкалу во время или сразу после инициации цветения, в зависимости от степени развития цветочной структуры.
Экспрессия EPSPS в каноле
После 3-недельного периода трансформированные растения канолы оценивают на наличие EPSPS-активности толерантности к глифосату (оценка в присутствии 0,05 мМ глифосата). Результаты представлены в таблице 8.
R1 - трансформанты канолы затем были выращены в ростовой камере и опрысканы глифосатом в дозе 0,56 кг/га (килограмм/гектар) и оценены вегетативно. Эти результаты представлены в таблице 9A-9C. Следует отметить, что экспрессия устойчивого к глифосату EPSPS во всех тканях является предпочтительной, чтобы наблюдать оптимальный толерантный к глифосату фенотип в этих трансгенных растениях.
В таблицах ниже приведены только результаты экспрессии, полученные из листовой ткани.
Данные, полученные для трансформантов EPSPS класса II, могут быть сравнены с толерантными к глифосату трансформантами EPSPS класса I, в которых использован один и тот же промотор для экспрессии EPSPS генов и в которых уровень толерантной к глифосату EPSPS-активности сравним для двух типов трансформантов. Сравнение данных для pMON17110 (в таблице 9A) и pMON17131 (таблица 9B) с данными для pMON899 (в таблице 9C; ген класса I в pMON 899 является геном из A.thaliana (Klee et al., 1987), в котором глицин в положении 101 заменен на аланин), показывает, что EPSPS класса II является по крайней мере таким же хорошим, как и EPSPS класса I. Улучшение в вегетативной толерантности EPSPS класса II появляется, когда принимают во внимание, что растения класса II опрыскивают дважды дозой и испытывают, как растения R1.
Пример 3.
Растения сои трансформируют вектором pMON13640 (фиг.15) и получают ряд линий трансформированных растений сои, которые проявляют толерантность к глифосату.
Растения сои трансформируют методом микроинъекций, используя технологию отдельных капель, как описано Christou et al. (1988). Семена, собранные от растений R0, являются R1 - семенами, которые дают растения R1. Для оценки толерантности к глифосату растений R0 оценивают его потомство. Поскольку предполагается, что растение R0 должно быть гемизиготным в месте каждой вставки, самоопыление приводит в результате к максимуму генотипической сегрегации в R1. Поскольку каждая вставка действует как доминантный аллель, в отсутствие связывания и предполагая, что только одна гемизиготная вставка требуется для экспрессии толерантности, одна вставка будет давать сегрегацию 3: 1, две вставки 15:1, три вставки 63:1 и т.д. Следовательно, нужно относительно небольшое количество растений R1, чтобы найти по крайней мере один устойчивый фенотип.
Собирают семена от растений сои R0 и сушат перед посадкой для теста с опрыскиванием глифосатом. Семена высевают в 4-дюймовые (≈ 5 см) квадратные горшочки, содержащие Метро 350. Двадцать ростков из каждого растения R0 считается адекватным для испытания. Регулируют 12,5 - 14-часовой фотопериод и температуру 30oC днем и 24oC ночью. При необходимости применяют водорастворимое удобрение Петерс Пет Лайт.
Опрыскивание "группы" заключается в опрыскивании в один и тот же день нескольких партий потомства R1. Некоторые группы также могут включать опрысканные и неопрысканные нетрансгенные генотипы, представляющие генотипы в конкретной группе, которые вероятно трансформированы. Также включены в группу один или более несегрегированных трансформированных генотипов, ранее идентифицированных как имеющие некоторую устойчивость.
Одно-два растения из каждого индивидуального R0 - потомства не опрыскивают и они служат в качестве контроля для сравнения и измерения толерантности к глифосату, а также для оценки любой вариабельности, не индуцированной глифосатом. Когда другие растения достигают стадии первого трилистника, обычно 2-3 недели после посева, глифосат наносят в дозе, эквивалентной 128 фунт/акр/ 8,895 кг/га Раундир®. Лабораторный гусеничный опрыскиватель был откалиброван для выдачи дозы, эквивалентной этим условиям.
Используют вегетативную оценку от 0 до 10. Шкала относится к неопрысканному потомству от одного и того же R0 - растения. 0 означает мертвое растение, а 10 означает отсутствие видимой разницы с неопрысканным растением. Более высокое число между 0 и 10 означает постепенное снижение повреждения при сравнении с неопрысканным растением. Растения оценивают на 7, 14 и 28 день после обработки (ДАТ). Данные от анализа одной группы трансформированных и контрольных растений сои представлены в таблице 10 и показывают, что CP4 EPSPS ген придает толерантность к глифосату также и сое.
Пример 4
CP4EPSPS - ген может быть использован для отбора трансформированного растительного материала непосредственно на среде, содержащей глифосат. Способность к отбору и идентификации трансформированного растительного материала зависит в большинстве случаев от использования доминантного селективного маркерного гена, способного к предпочтительному и непрерывному росту трансформированных тканей в присутствии нормально ингибирующих веществ. Устойчивость к антибиотикам и толерантность к гербицидам генов была использована почти исключительно как такие доминантные селективные маркерные гены в присутствии соответствующих антибиотиков или гербицидов. Схема селекции npt II/канамицин возможно используется чаще всего. Было показано, что CP4EPSPS также является полезным и по-видимому превосходит схему селективный маркер/отбор для получения и идентификации трансформированных растений.
Вектором трансформации растений, который может быть использован в этой схеме, является pMON17227 (фиг. 16). Эта плазмида объединяет многие другие плазмиды, описанные ниже, и по существу составлена из ранее описанной бактериальной системы репликона, которая придает этой плазмиде способность к репликации в E.coli, и она может быть введена и реплицировать в Agrobacterium, бактериального селективного маркерного гена (Spc/Str), и локализованным между правым краем T-ДНК и левым краем является CTP2-CP4 синтетический ген в FMV35S промоторе-E93'-полигенного экспрессирующего кластера. Эта плазма также имеет уникальные сайты для ряда рестрикционных ферментов, расположенные внутри краев и вне полигенного экспрессирующего кластера. Это делает возможным легко добавлять другие гены и генетические элементы в вектор для введения в растения.
Протокол прямой селекции трансформированных растений на глифосате приведен для табака. Экспланты готовят в пред-культуре, как в стандартной процедуре, описанной в примере 1: поверхностная стерилизация листьев месячного возраста растений табака (15 минут в 10% клороксе + поверхностно-активное вещество; 3X dH2O промывки); экспланты нарезают на квадратики 0,5 х 0,5 см, удаляя листовую кромку, среднюю жилку, кончик и конец черешка для ткани однородного типа; экспланты помещают на однослойные, верхней стороной вниз, MS104 пластин + 2 мл 4C005K среды для увлажнения поверхности; пре-культура 1-2 дня. Экспланты инокулируют, используя ночную культуру Agrobacterium, содержащую плазмиду трансформации растений, которую регулируют до титра 1,2•109 бактерий/мл средой 4C005K. Экспланты помещают в центрифужные трубки и прибавляют суспензию Agrobacterium и смесь бактерий и эксплантов интенсивно перемешивают до максимального загущения в течение 25 секунд, чтобы обеспечить проникновение бактерий. Бактерии сливают и экспланты промокают между слоями сухой стерильной фильтровальной бумаги для удаления избытка бактерий. После этого экспланты помещают верхней стороной вниз на MS104 пластины + 2 мл среды 4C005K + фильтровальный диск. Со-культура проводится 2-3 дня. Экспланты переносят на MS104 + Карбенициллин 1000 мг/л + цефотаксим 100 мг/л на 3 дня (замедленная фаза). Затем экспланты переносят на MS104 + глифосат 0,05 мМ + Карбенициллин 1000 мг/л + цефотаксим 100 мг/л для селективной фазы. Через 4-6 недель ростки отрезают от каллюса и помещают на MSO + Карбенициллин 500 мг/л среду укоренения. Корни образуются на 3-5 день, за это время кусочки листа могут быть взяты с пластин для укоренения для подтверждения толерантности к глифосату и что этот материал трансформирован.
Наличие CP4 EPSPS-белка в этих трансформированных тканях было подтверждено иммуноблотинговым анализом листовых дисков. Данные из одного эксперимента с pMON17227 представляют собой следующее: 139 побегов образовалось на глифосате из 400 эксплантов, инокулированных Agrobacterium ABI/pMON17227; 97 из них были положительными на повторное каллюсообразование на глифосате. Эти данные указывают, что степень трансформации составляет 24 на 100 эксплантов, что делает ее очень эффективной и сокращает время процедуры трансформации растений. Подобные частоты трансформаций были получены с pMON17131, а прямая селекция трансформантов на глифосате с CP4 EPSPS генами также была показана на других видах растений, включая Arabidopsis, картофель, томаты, хлопок, латук и сахарную свеклу.
pMON17227 плазмида содержит уникальные сайты распознавания расщепляющими ферментами (NotI, XhoI и BstXI) между областью CP4 селекции на глифосат и левым краем вектора клонирования дополнительных генов и облегчения введения этих генов в растения.
Пример 5.
Ген CP4 EPSPS также был введен в клетки кукурузы Черная мексиканская сладкая (BMS) эккспрессией белка и устойчивости к глифосату, детектирована в каллюсе.
Основой для этой плазмиды явилось производное высококопирующей плазмиды puC119 (Viera and Messing, 1987). Фрагмент 1,3 ко FspI-DraI puC119, содержащей ориджин репликации, сливают с наполненным фрагментом 1,3 ко SmaI-HindIII из pKC7 (Rao and Rogers, 1979), который содержит ген неомицин фосфотрансферазы тип II для придания бактериальной устойчивости к канамицину. Эту плазмиду используют для конструирования вектора монокот полигенного экспрессирующего кластера, содержащего 0,6 ко промотор вируса мозаики подсолнечника (CaMV) 35S РНК с удвоением области от -90 до -300 (Kay et al., 1987), 0,8 ко фрагмента, содержащего интрон из гена кукурузы в 5'-нетранслируемой лидерной области, после полилинкера и 3'-концевых последовательностей из нопалин синтазного (NOS) гена (Fraley et al., 1983). Фрагмент 1,7 ко, содержащий 300 во хлоропластного транзитного белка из Arabidopsis EPSP синтазы, слитый в рамке с 1,4 ко кодирующей последовательностью для бактериальной CP4 EPSP-синтазы, вставлен в монокот полигенный экспрессирующий кластер в полилинкер между интроном и NOS-концевой последовательностью для образования плазмиды pMON19653 (фиг. 17).
pMON19653 ДНК была вставлена в клетки Черной мексиканской сладкой кукурузы (BMS) со-бомбардировкой EC9, плазмидой, содержащей сульфонилмочевина-устойчивую форму ацетолактат синтазного гена кукурузы. 2,5 мг каждой плазмиды покрывают частицы вольфрама и интродуцированы в log-фазу BMS клеток, используя пушки PDS-1000 частиц, по существу как описано (Klein et al., 1989). Отбирают трансформанты на MS среде, содержащей 20 ppb хлорсульфурона. После начальной селекции на хлорсульфуроне каллюс может быть проанализирован непосредственно блот-анализом по Вестерну. Толерантность к глифосату может быть оценена путем переноса каллюса на среду, содержащую 5 мМ глифосата. Как показано в таблице 11, CP4 EPSPS придает толерантность к глифосату каллюсу кукурузы.
Для измерения экспрессии CP4 EPSPS в каллюсе кукурузы была использована следующая процедура: BMS каллюс (3 г влажного веса) сушат на фильтровальной бумаге (Ватман # 1) в вакууме, повторно взвешивают и прибавляют экстракционный буфер (500 мкл/г сухого веса; 100 мМ Трис, 1 мМ EDTA, 10% глицерина). Ткань гомогенизируют с помощью мешалки с головкой Витона в течение 30 секунд при снятой мощности 2,8. После центрифугирования (3 минуты, микрофуга Эппендорфа) удаляют супернатант и количественно определяют белок (БиоРад белковый анализ). Загружают образцы 50 мкг/ячейку в SDS-PAGE гель (Жюль, 3-17%) рядом со стандартом CP4 EPSPS (10 нг), проводят электрофорез и переносят на нитроцеллюлозу подобно ранее описанному методу (Padgelle, 1987). Нитроцеллюлозный блот зондируют козьим анти-CP4 EPSPS IgG и проявляют 1-125 Протеином G. Радиоактивный блот был визуализирован ауторедиографией. Результаты были представлены в количественной форме с помощью денситометрии на лазерном денситометре LKB UltraScan XL и сведены в таблицу 12 ниже.
Улучшения в экспрессии EPSPS класса I также могут быть достигнуты путем экспрессии гена с использованием более сильных растительных промоторов, использованием лучших сигнальных последовательностей 3'-полиаденилирования, оптимизации последовательностей вокруг кодона инициации для рибосомной загрузки и инициации трансляции, или путем сочетания этих или других регуляторных или экспрессирующих последовательностей или факторов. Также должна быть благоприятной трансформация желаемых растений геном EPSPS класса I в сочетании с другим геном EPSPS толерантности к глифосату, или геном, способным к деградации глифосата, чтобы повысить толерантность к глифосату трансформированных растений.
Из вышесказанного видно, что настоящее изобретение хорошо приспособлено к достижению всех вышеприведенных целей вместе с преимуществами, которые являются очевидными и которые присущи изобретению.
Должно быть понятно, что некоторые признаки и сочетания являются полезными и могут быть применены без ссылок на другие признаки и сочетания. Это является ожидаемым и входит в область формулы изобретения.
Поскольку может быть сделано много возможных вариантов изобретения без выхода за его область, должно быть понятно, что все приведенное здесь существо или показанное на приложенных чертежах должно быть интерпретировано как иллюстративное и не ограничивающее.
Пример 6
Ген LBAA EPSPS класса II был введен в растения и он также придает толерантность к глифосату. Данные о табаке, трансформированном pMON17206 (ниже), представлены в таблице 13.
Список литературы
Alton, N.K. and Vapnek, D. (1979) Nature 282:864-869.
Ammirato, P. V. , et al. Handbook of Plant Cell Culture - Crop Species. Macmillan Publ. Co. (1984).
Arnon, D.I. Plant Physiol. 24:1-15 (1949).
Bachmann, В.J. et al., Microb. Rev. 44:1-56 (1980).
Bartlett, S. G. , Grossman, A.R., and Chua, N.H. (1982) in Methods in Chloroplast Molecular Biology. pp. 1081-1091. M. Edelman, R.B., Hallick, and Chua, N.H.,eds.
Bevan, M. (1984) Nucleic Acids Res. 12 (22): 8711-8721.
Birnboim, H.C. and Doly, J. (1979) A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucl. Acids. Res. 7:1513-1525.
Boyer, H.W. and Rolland-Dussoix, D. (1969) A complementation analysis of the restriction and modification of DNA in Escherichia coli. J. Mol. Biol. 41:459.
Christou, P. , D. E.McCabe, and W.F. Swain (1988) Stable transformation of Soybean Callus by DNA-Coated Gold Particles. Plant Phvsiol. 87:671-674.
Coruzzi, G. , Broglie, R., Edwards, C., and Chua, N.H. (1984). Tissue-specific and light-regulated expression of a pea nuclear gene encoding the small subunit of ribulose-1,5-bisphosphate carboxylase. EMBO J 3:1671.
della-Cioppa, G., Bauer, S.C., Klein, B.K., Shah, D.M., Fraley, R.T. and Kishore G. K. (1986) Translocation of the precursor of 5-enolpyruvylshikimate-3-phosphate synthase into chloroplasts of higher plants in vitro. Proc. Natl. Acad Sci. USA 83: 6873-6877.
della-Cioppa, G., Bauer, S.C., Taylor, M.Т., Rochester, D. E., Klein, B. K. , Shah, D.M., Fraley, R.T. and Kishore G.M. (1987) Targeting a herbicide-resistant enzyme from Escherichia coli to chloroplasts of higher plants. Bio/Technology 5: 579-584.
Devereux, J. , Haeberli, P. and Smithies, O. (1984) A comprehensive set of sequence analysis programs for the VAX. Nucl. Acids. Res. 12:387-395.
Ditta, G. , Stanfield, S., Corbin, D., and Helinski, D.R. (1980) Broad host range DNA cloning system for Gram-Negative bacteria: construction of a gene bank of Rhizobium meliloti. Proc Natl Acad Sci USA 77,7347-7351.
Duncan, K., Edwards, R.M., Coggins, J.R. (1987) The pentafunctional aroM enzyme of Saccharomyces cerevisiae is a mosaic of monofunctional domains. Biochem. J. 246: 375-386.
Dunn, J.J. and Studier, F.W., (1983) J. Mol. Biol. 166:477- 535.
Fitzgibbon, J. E. (1988) Pseudomonas sp. strain PG2982: uptake of glyphosate and cloning of a gene which confers increased resistance to glyphosate. Ph. D. Dissertation, Louisiana State University.
Fitzgibbon, E. F. and Braymer, H.D. (1990) Cloning of a gene from Pseudomonas sp. PG2982 conferring, increased glyphosate resistance Appl. Environ. Microbiol. 56:3382-3388.
Fling, M. E., Kopf, J., and Richards, C. (1985). Nucleotide sequence of the transposon Tn7 gene encoding an aminoglycoside- modifying enzyme, 3"(9)-O-nucleotidyltransferase. Nucleic Acids Research 13 no. 19, 7095-7106.
Fraley, R. T. , Rogers, S.G., Horsch, R.B., Sanders, P.R. Flick, J.S., Adams, S.P., Bittner, M.L., Brand, L.A., Fink, C.L., Fry, J.S., Galluppi, G. R. , Goldberg, S.B., Hoffman, N.L., and Woo, S.C. 1983. Expression of bacterial genes in plant cells. Proc. Natl. Acad. Sci. USA 80:4803-4807.
Fraley, R. Т., Rogers, S.G., Horsch, R.B., Eichholtz D.A., Flick, J.S., Fink, C. L. , Hoffmann, N.L. and Sanders, P.R. (1985) The SEV system: a new disarmed Ti plasmid vector system for plant transformation.
Fromm, M. , (1990) UCLA Symposium on Molecular Strategies for Crop Improvement, April 16-22, 1990. Keystone, CO.
Gasser, C. S. , Winter, J. A. , Hironaka, C.M. and Shah, D. M. (1988) Structure, expression, and evolution of the 5- enolpyruvyishikimate 3-phosphate synthase genes of petunia and tomato. J. Biol. Chem. 263: 4280-4289.
Gowda, S. , Wu, F.C., and Shepard, R.J. (1989). Identification of promoter sequences for the major RNA transcripts of figwort mosaic and peanut chlorotic streak viruses (caulimovirus group). Journal of Cellular Biochemistry supplement 13D, 301 (Abstract).
Hallas, L. E., Hahn, E.M. and Korndorfer, C. (1988) Characterization of microbial traits associated with glyphosate biodegradation in industrial activated sludge. J. Industrial Microbiol. 3: 377-385.
Hayford, M.B., Medford, J.I., Hoffmann, N.L., Rogers, S. G. and Klee, H. J. (1988) Development of a plant transformation selection system based on expression of genes encoding gentamicin acetyltransferases. Plant Physiol. 86: 1216-1222.
Herrera-Estrella, L. , et al. (1983) Nature 303:209 Horsch. R.B. and H. Klee. (1986) Proc. Natl. Acad. Sci. U.S.A. 83:4428-32.
Heitkamp, M.A., Hallas, L. and Adams, W.J. (1990) Biotreatment of industrial wastewater with immobilized microorganisms - Presented in Session 11, Paper S40, Society for Industrial Microbiology Annual Meeting, Orlando, Florida, July 29- August 3, 1990.
Hohn, В. and Collins J. (1980) A small cosmid for efficient cloning of large DNA fragments. Gene 11: 291-298.
Hunkapiller, M.W., Hewick, R.M., Dreyer, R.J., and Hood, L. (1983) Methods Enzymol. 91, 399-413.
Jefferson, R. A. , Kavanaugh, T.A. and Bevan, M.W., EMBO J. 6:3901-3907 (1987).
Kay, R. , Chan, A., Daly, M. and McPherson, J. 1987. Duplication of the CaMV 35S promoter sequence creates a strong enhancer for plants. Science 236,1299-1302.
Kishore, G. , Shah, D., Padgette, S., della-Cioppa, G., Gasser, C., Re, D. , Hironaka, C., Taylor, M., Wibbenmeyer, J., Eichholtz, D., Hayford, M., Hoffman, N., Delannay, X., Horsch, R., Klee, H., Rogers, S., Rochester, D., Brundage, L. , Sanders, P. and Fraley, R.T. (1988) 5-Enolpyruvylshikimate 3-phosphate synthase: From Biochemistry to genetic engineering of glyphosate tolerance, in Biotechnology for Crop Protection ACS Symposium series N. 379. Eds. Hedlin P. A., Menn, J. J. and Hollingsworth, R. M. pp. 37-48.
Kishore, G. and Shah, D. (1988) Ann. Rev. Biochem. 57:627- 663.
Kishore, G. M. , Brundage, L., Kolk, K., Padgette, S.R., Rochester, D., Huynh, Q.K. and della-Cioppa, G. (1986) Fed. Proc. 45:1506.
Klee, H.J., et al. (1985) Bio/Technology 3:637-42.
Klee H. J. , Muskopf, Y.M. and Gasser, C.S. (1987) Cloning of an Arabidopsis thaliana gene encoding 5-enolpyruvylshikimate- 3-phosphate synthase: sequence analysis and manipulation to obtain glyphosate tolerant plants. Mol. Gen. Genet. 210:437-442.
Klein, T.M., Kornstein, L., Sanford, J.C., and Fromm, M.E. 1989. Genetic transformation of maize cells by particle bombardment. Plant Phys. 91:440-444.
Koncz, C. and Schell, J. (1986) The promoter of TL-DNA gene 5 controls the tissue-specific expression of chimeric genes carried by a novel type of Agrobacterium binary vector. Mol. Gen. Genet. 204:383-396.
Kunkel, T.A., Roberts, J.D. and Zakour, R.A. (1987) Rapid and efficient site-specific mutagenesis without phenotypic selection. Methods Enzymol. 154:367.
Laemmli, U. K., "Cleavage of structural proteins during the assembly of the head of the bacteriophage T4" Nature. 227:680 (1970).
Maniatis, T., Fritsch, E.F. and Sambrook, J. (1982) Molecular Cloning: a laboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
Maskell, D. J., Morrissey, P. and Dougan, G. (1988) Cloning and nucleotide sequence of the aroA gene of Bordetella pertussis. J. Bacteriol. 170: 2467-2471.
Miller, J.H. (1972). Experiments in Molecular Genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
Moore, J.K., Braymer, H.D. and Larson, A.D. (1983) Isolation of a Pseudomonas sp. which utilizes the phosphonate herbicide glyphosate. Appl. Environ. Microbiol. 46: 316-320.
Morelli, G. , Nagy, F. , Fraley, R.T., Rogers, S.G., and Chua, N.H. (1985). A short conserved sequence is involved in the light- inducibility of a gene encoding ribulose 1,5-bisphosphate carboxylase small subunit of pea. Nature 315, 200-204.
Odell, J. T. , Nagy, F., and Chua, N.H. (1985). Identification of DNA sequences required for activity of the cauliflower mosaic virus 35S promoter. Nature 313, 810-812.
Olins, P. O. , Devine, C.S., Rangwala, S.H. and Kavka, K.S. (1988) Gene 73:227-235.
Padgette, S.R., Huynh, Q.K., Borgmeyer, J., Shah, D.M., Brand, L.A., Re, D. B., Bishop, B.F., Rogers, S.G., Fraley, R.Т., and Kishore, G. (1987) Bacterial expression and isolation of Petunia hybrida 5-enol-pyruvylshikimate-3-phosphate synthase. Arch. Biochem. Biophys. 258, 564-573.
Padgette, S.R., Huynh, Q.K., Aykent, S., Sammons, R.D., Sikorski, J.A., and Kishore, G.M. (1988) J. Biol. Chem. 263, 1798-1802.
Quinn, J.P., Peden, J.M.M. and Dick, E. (1988) Glyphosate tolerance and utilization by the microflora of soils treated with the herbicide. Appl. Microbiol. Biotechnol. 29: 511-516.
Rao, R. N. and Rogers, S.G. 1979. Plasmid pKC7: A vector containing ten restriction endonuclease sites suitable for cloning DNA segments. Gene 7:79.
Rogers, S. G. , Brand, L.A. Holder, S.B. Sharps, E.S. and Brackin, M.J. (1983) Amplification of the aroA gene from E. coli results in tolerance to the herbicide glyphosate. Appl. Environ. Microbiol. 46:37-43.
Sambrook, J., Fritsch, E.F. and Maniatis, Т., Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York (1989).
Schuler, M.A., Schmitt, E.S. and Beachy, R.N. (1982) Nucleic Acids Res. 10:8225-8244.
Schuiz, A. , Kruper, A. and Amrhein, N. (1985) Differential sensitivity of bacterial 5-enolpyruvylshikimate-3-phosphate synthases to the herbicide glyphosate. FEMS Microbiol. Lett. 28: 297-301.
Schuiz, A., Sost, D. and Amrhein, D. (1984) Arch. Microbiol. 137: 121-123.
Shah, D. , Horsch, R. , Klee, H., Kishore, G., Winter, J., Tumer, N., Hironaka, C. , Sanders, P., Gasser, C., Aykent, S., Siegal, N., Rogers, S., and Fraley, R. (1986). Engineering herbicide tolerance in transgenic plants. Science 233, 478-481.
Shimamoto, K. et al. (1989) Nature 338:274-276.
Sost, D., Schuiz, A. and Amrhein, N. (1984) FEBS Lett. 173: 238-241.
Sost, D. and Amrhein, N. (1990) Substitution of Gly-96 to Ala in the 5-enolpyruvylshikimate 3-phosphate synthase of Klebsiella pneumoniae results in greatly reduced affinity for the herbicide glyphosate. Arch. Biochem. Biophys. 282: 433-436.
Stalker, D. M., Thomas, C.M., and Helinski, D.R. (1981). Nucleotide sequence of the region of the origin of replication of the broad host range plasmid RK2. Mol Gen Genet 181: 8-12.
Stalker, D. M. , Hiatt, W. R. and Comai, L. (1985) A single amino acid substitution in the enzyme 5-enolpyruvylshikimate 3- phosphate synthase confers resistance to glyphosate. J. Biol. Chem. 260:4724-4728.
Tabor, S. and Richardson, C.C. (1985) A bacteriophage T7 RNA polymerase/promoter system for controlled exclusive expression of specific genes. Proc. Natl. Acad. Sci. USA 82: 1074-1078.
Talbot, H.W., Johnson, L.M. and Munnecke, D.M. (1984) Glyphosate utilization by Pseudomonas sp. and Alcaligenes sp. isolated from environmental sources. Current Microbiol. 10: 255-260.
Talmadge, K. , and Gilbert, W., "Construction of plasmid vectors with unique PstI cloning sites in the signal sequence coding region" Gene. (12) 235-241 (1980).
Vasil, V., F. Redway and I. Vasil., Bio/Technology 8:429-434 (1990).
Velten, J., Velten, R., Hain, R. and Schell, J. (1984) EMBO J. 3:2723-2730.
Viera, J. and Messing, J. 1987. Production of single-stranded plasmid DNA. Methods Enzym. 153:3-11.
Wibbenmeyer, J., Brundage, L., Padgette, S.R., Likos, J.J., and Kishore, G.M. (1988) Biochem. Biophys. Res. Comm. 153, 760- 766.
Wong, E. Y. , Seetharam, R. , Kotts, C.E., Heeren, R.A., Klein, B.K., Braford, S.R., Mathis, K.J., Bishop, B.F., Siegel, N.R., Smith, C.E. and Tacon, W. C. (1988) Expression of excreted insulin-like growth factor-1 in Escherichia coli. Gene 68: 193-203.6




























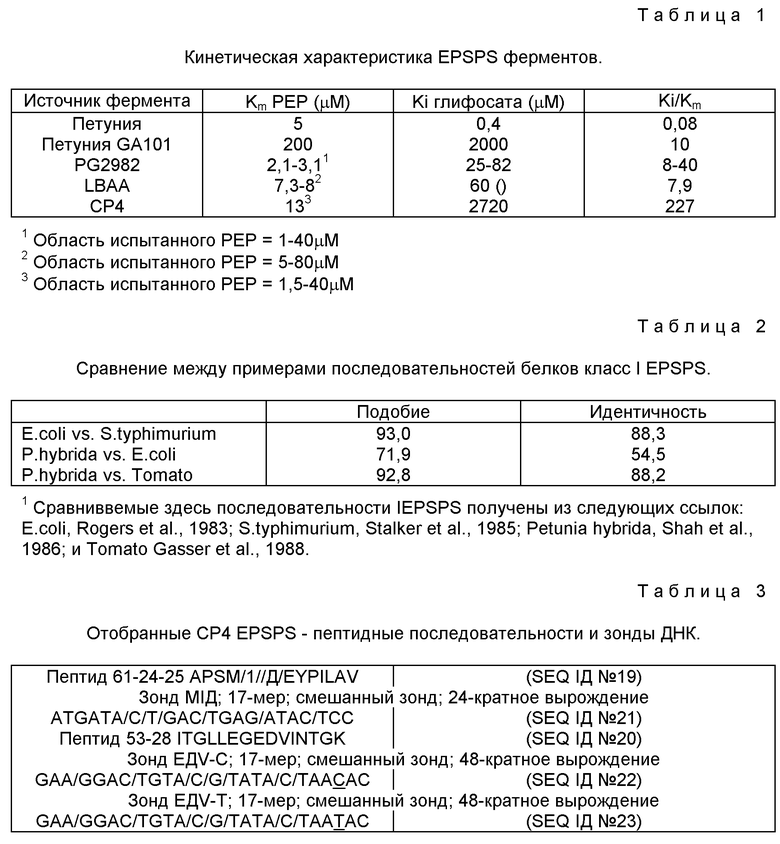
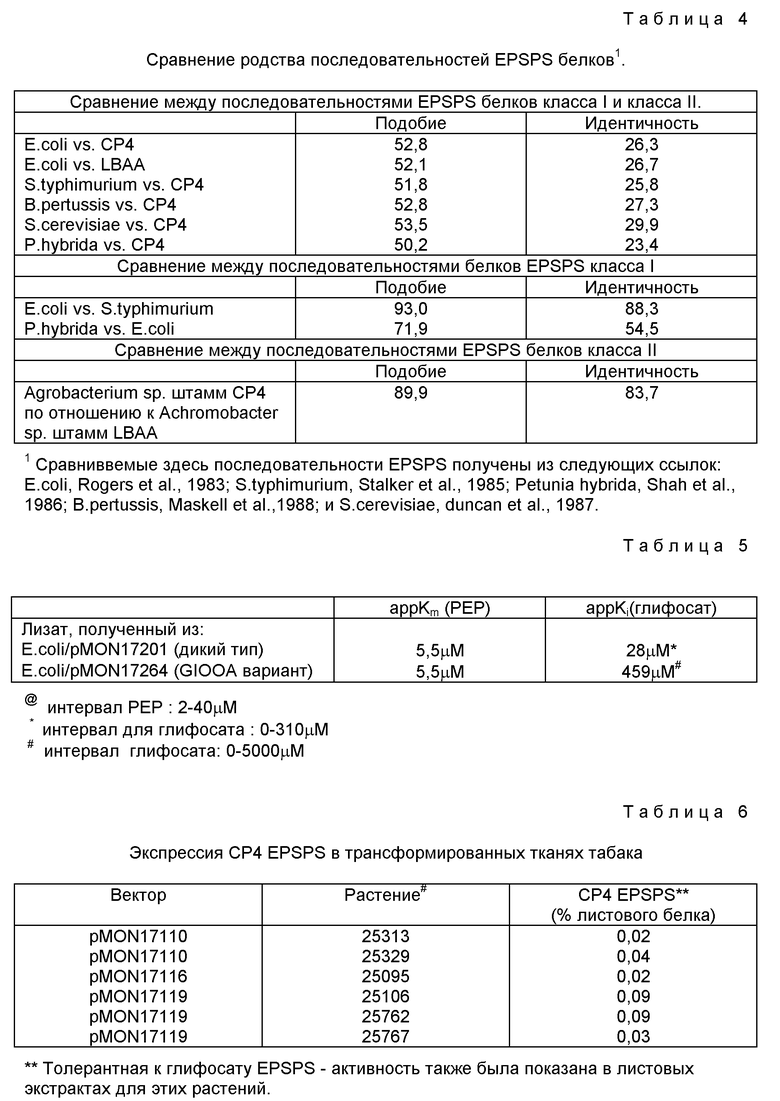

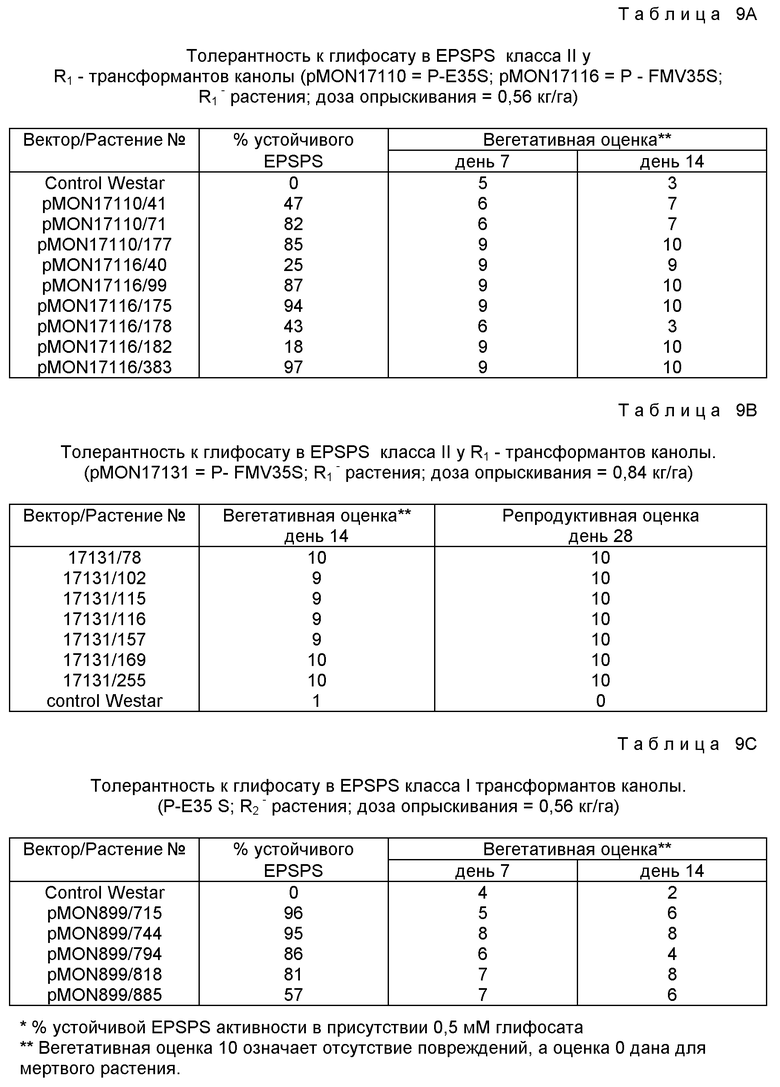
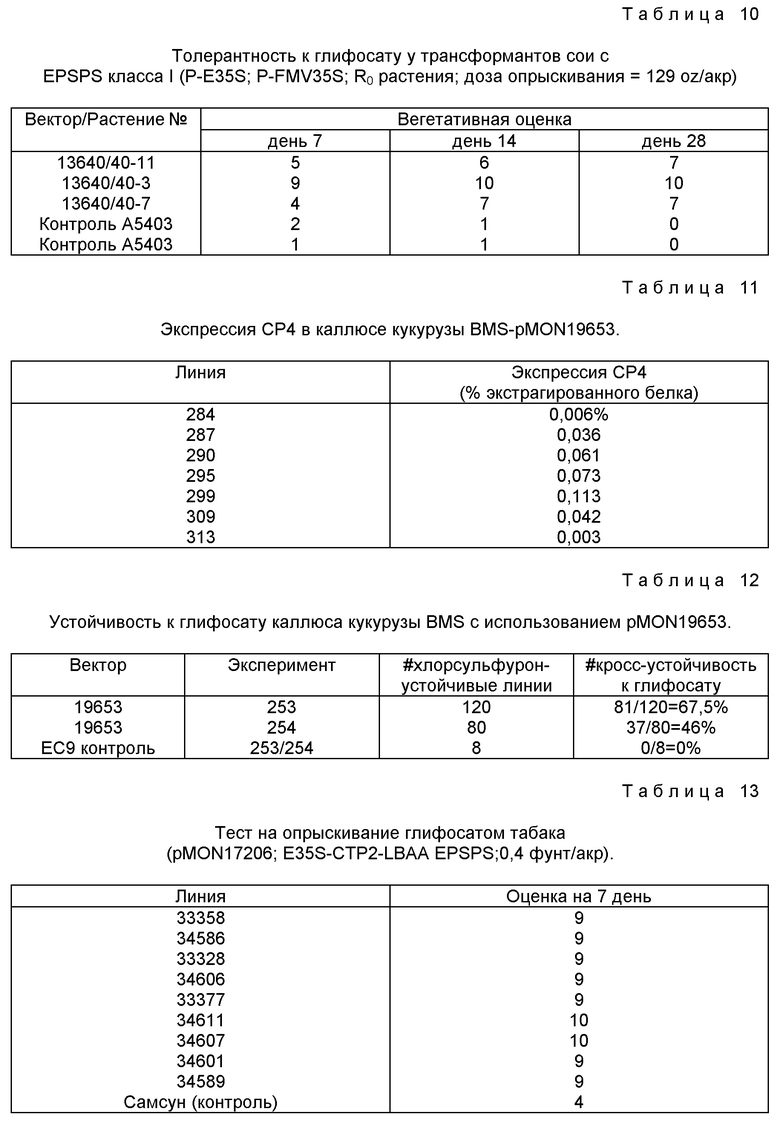


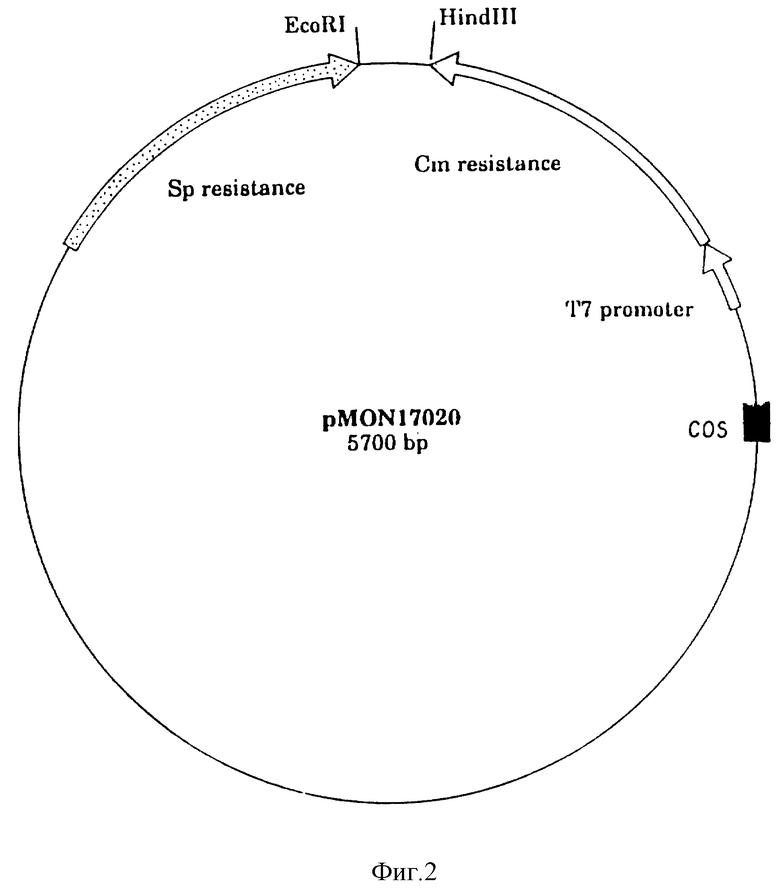
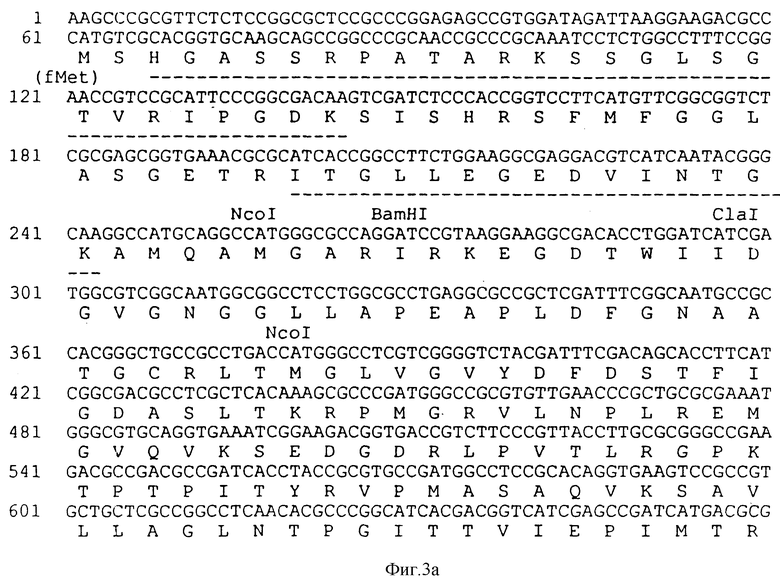


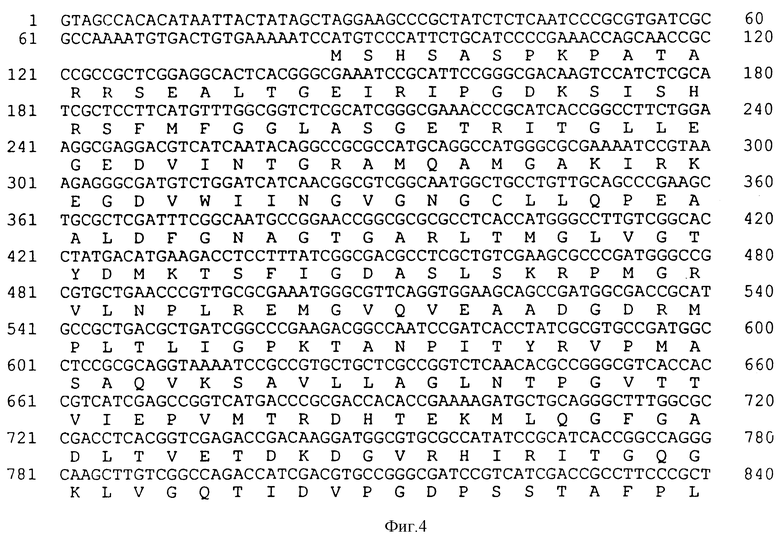
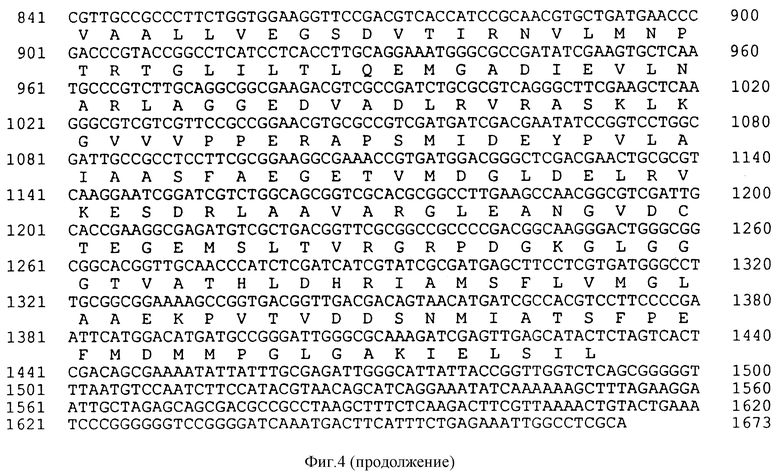







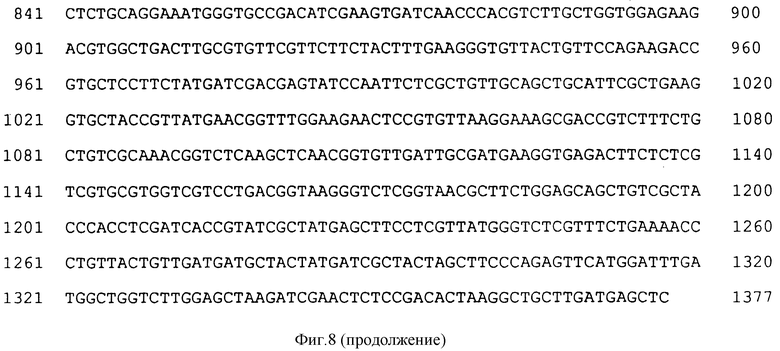







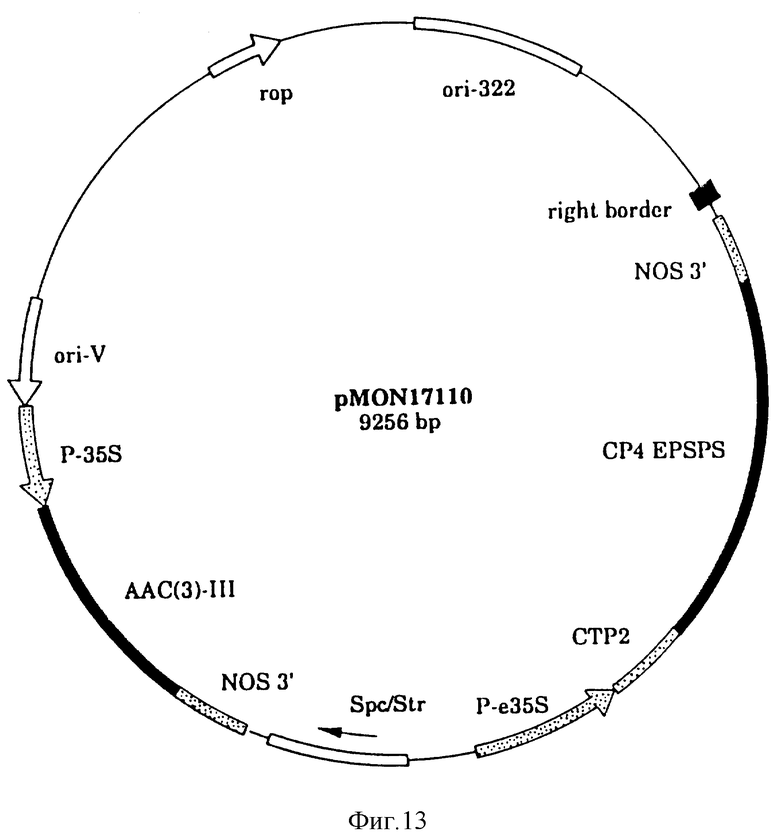
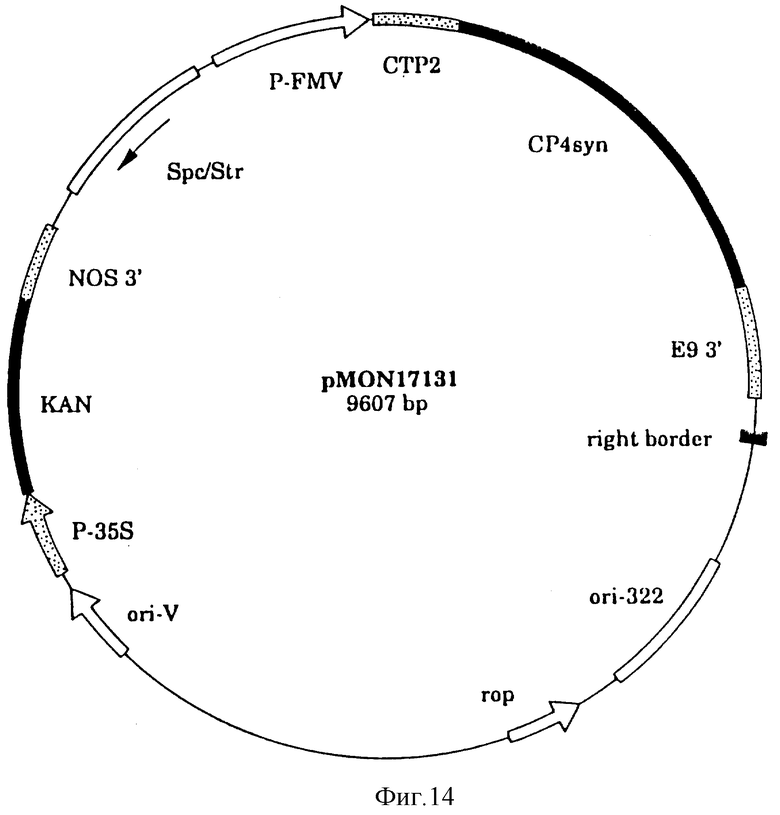
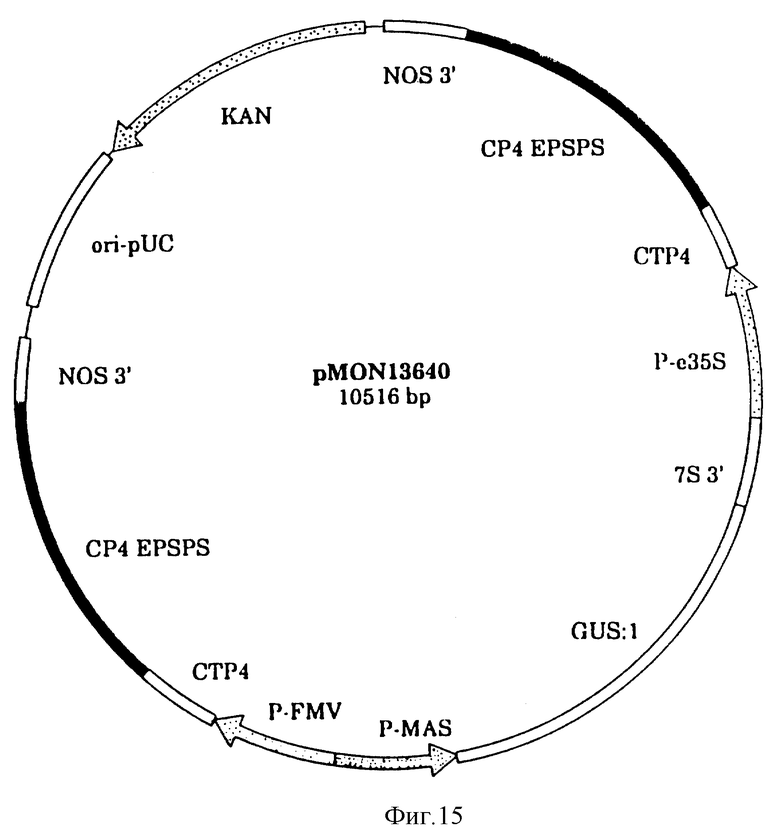
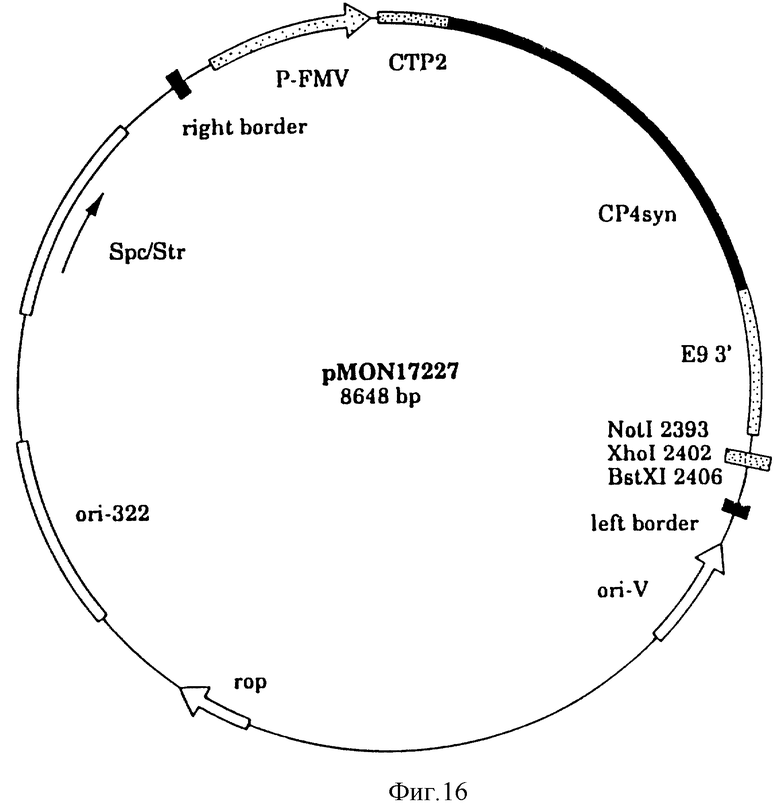
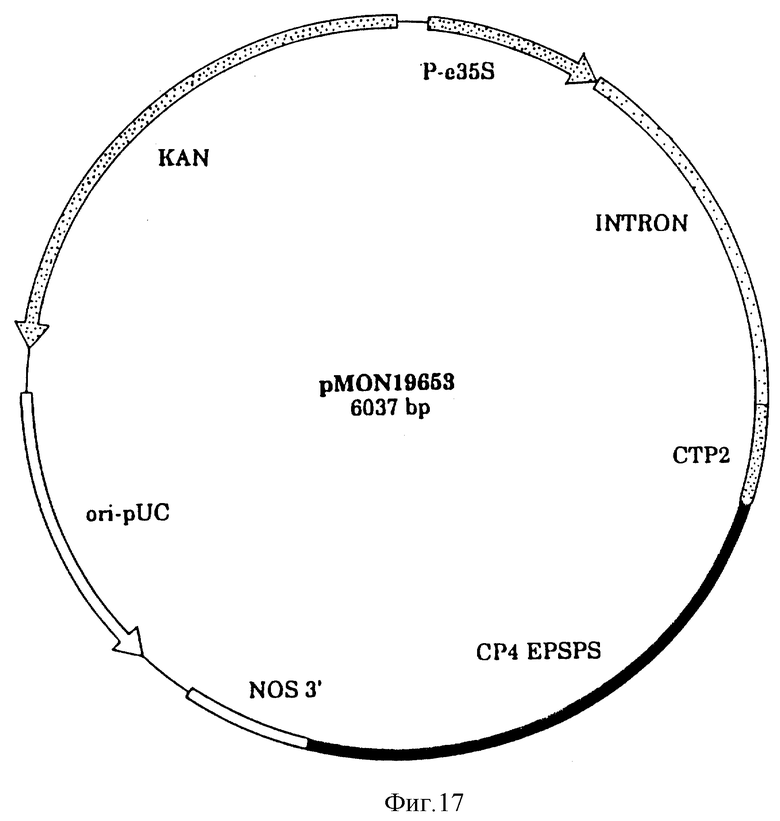
Последовательности ДНК предназначены для экспрессии ферментов ЕРSРS нового класса и могут быть использованы для получения растений, устойчивых к гербициду глифосату. Ферменты EPSPS этого класса (II) характеризуются большей кинетической эффективностью, чем известного класса (I), в присутствии глифосата. 4 с. и 13 з.п. ф-лы, 17 ил, 13 табл.
| БОЛТОВОЕ СОЕДИНЕНИЕ | 0 |
|
SU218571A1 |
| 0 |
|
SU293358A1 | |
| US 4769061 A1, 06.09.88 | |||
| Способ отбора устойчивых к 2,4- @ форм ячменя | 1983 |
|
SU1165314A1 |
| Способ селекции пшеницы,устойчивой к неблагоприятным факторам | 1982 |
|
SU1170991A1 |
