Изобретение относится к фрагментам ДНК, кодирующим переносчики олигосахаридов, плазмидам, содержащим указанные фрагменты ДНК, способу модификации метаболизма углеводов в растении, способу снижения апикального доминирования в тканях растения, способу получения бактерии, способной поглощать сахарозу, способам выделения фрагмента ДНК, кодирующего переносчик олигосахаридов растений, дрожжевым штаммам.
Наиболее важным транспортным метаболитом для запасания энергии во многих растениях, например, картофеле, является сахароза. В других видах растений другие олигосахариды могут выполнять эту роль. В японских артишоках таким олигосахаридом является, например, стахиоза.
На трансгенных растениях было продемонстрировано центральное положение транспорта олигосахаридов при запасании энергии растениями, в которых за счет экспрессируемой инвертазы происходит расщепление сахарозы на моносахариды, что ведет, в свою очередь, к значительным изменениям габитуса растений (EP 442592). В связи со значимостью сахарозы в образовании запасных материалов было проведено множество исследований по биосинтезу и метаболизму дисахаридов. Из документа DE 4213444 известно, что можно достигнуть улучшения способностей к хранению у трансгенного картофеля, у которого через экспрессию апопластической инвертазы ингибируется транспорт богатых энергией материалов к гетеротрофным частям растущих побегов.
Несмотря на многочисленные попытки, до сих пор не удалось прояснить механизм распределения в растениях запасных материалов с целью воздействия на него таких, например, как олигонуклеотиды, так, все еще неизвестно, как в листьях после фотосинтеза образуется сахароза, как она достигает транспортных каналов флоэмы растений и как она отбирается из запасающих органов, таких, как картофельные клубни или семена. Было продемонстрировано на изолированных плазматических мембранах клеток листовой ткани сахарной свеклы (Beta vulgaris), что транспорт сахарозы через мембраны можно индуцировать созданием искусственного градиента pH и интенсифицировать посредством поддержания электрохимического потенциала (Lemoine & Delrot (1989) FEBS letters 249:129-133). Транспорт сахарозы через мембрану подчиняется кинетике уравнения Михаэлиса-Ментен, в котором значение Km для транспорта сахарозы находится в пределах 1 мМ (Slone & Buckhout 1991, Planta 183:484-589). Указанная форма кинетики предполагает участие в процессе транспортных белков. Эксперименты на плазматических мембранах сахарной свеклы, Ricinus communis и Cyclamen persicum показали, что транспорт сахарозы сопровождается сопутствующим транспортом протонов (Buckholet, 1989, Planta 178:393-399; Williams et al. , 1990, Planta 182: 480-485). Стехиометрия сопутствующего транспорта составляет 1: 1 (Bush, 1990, Plant Physiol. 93:1590-1596). Были предложены механизмы, объясняющие механизм транспорта сахарозы через плазмодий растительных клеток (Robards & Lucas, 1990, Ann Rev Plant Physiol 41:369-419).
Несмотря на доказанность существования активной транспортной системы, позволяющей клеткам доставлять сахарозу к транспортным каналам, белок с соответствующими этому процессу свойствами еще неизвестен. При окрашивании плазматических мембран сахарной свеклы N-этилмелеинимидом в присутствии и отсутствии сахарозы Галлет с соавт. (Gallet et al., 1989, Biochem. Biophys. Acfa 978:56-64) получили информацию о наличии белка размером 42 кДа, способного взаимодействовать с сахарозой. Антитела к фракции белка этого размера из плазматической мембраны могут ингибировать транспорт сахарозы через мембраны (Lemoine et al., 1989, Biochem. Biophys. Acfa 978:65-71).
В отличие от этого, была получена информация с применением фотоаффинного связывания белка из сои с аналогом сахарозы-дезоксиазидогидроксибензамидосахарозой - об участии белка 62 кДа в транспорте сахарозы через мембраны. Аминокислотная последовательность переносчика сахарозы неизвестна.
Описаны последовательности ДНК, которые содержат кодирующий регион для синтеза переносчика олигосахарида, при этом содержащаяся в нуклеотидной последовательности информация позволяет посредством интеграции последовательности в геном растения синтезировать РНК, с помощью которой транспортная активность в отношении олигосахаридов может быть введена в клетки растения, а также может экспрессироваться активность переносчика эндогенного олигосахарида. Под термином переносчик олигосахаридов подразумевается переносчик сахарозы в таких, например, растениях, как шпинат или картофель.
Идентификация кодирующего региона переносчика олигосахаридов производится с применением способа, позволяющего выделить последовательности растительной ДНК, кодирующие транспортные молекулы при экспрессии в специфических мутантах дрожжей Sacharemyces cerevisial. Для этой цели необходимы соответствующие дрожжевые мутанты, не способные утилизировать те вещества, которые определяются кодирующим регионом транспортной молекулы, выделяемой из генной библиотеки растений. Такой способ известен для случая применения при недостаточности транспорта калия в мутантах дрожжей с целью его комплементации (Anderson et al., 1992, Proc. Natl. Acad. Sci, USA 3736-3740).
Согласно этому способу, последовательности дрожжевой кДНК экспрессируются в мутантах дрожжей с использованием векторов экспрессии. Те дрожжи, которые способны усваивать транспортируемые в клетки вещества, содержат в векторе экспрессии кодирующий регион для транспортной молекулы. Однако, этот способ не пригоден для идентификации кодирующего региона переносчика сахарозы, поскольку дрожжи не содержат эндогенный переносчик сахарозы, который мог бы быть выключен из метаболизма в результате мутации. Дрожжи кодируют расщепляющую инвертазу, которая расщепляет внеклеточную сахарозу, так что получающиеся гексозы могут уже усваиваться из клеток с помощью переносчика гексозы.
Для получения и трансформации дрожжевых штаммов, которые можно использовать для целей идентификации переносчика олигосахаридов в растениях:
а) вначале с помощью гомологичной рекомбинации получают штамм дрожжей с дефектным suc 2 геном, который не расщепляется инвертазой, и затем из него
б) посредством трансформации с участием гена, кодирующего синтез активной синтазы сахарозы в растительных клетках, выделяют штамм дрожжей, способный расщеплять внутриклеточную сахарозу, и затем
в) через трансформацию этого штамма вектором экспрессии для дрожжевых клеток, содержащим библиотеку растительных кДНК, идентифицируют последовательности ДНК, кодирующие переносчик олигосахаридов. Полученные по этому способу для идентификации переносчиков олигосахаридов в растениях штаммы дрожжей включают, в частности, такие штаммы, как YSH 2.64-IA-SUSY (DSM 7106) и YSH-IA-INV (DSM 7105).
С использованием дрожжевого штамма YSH 2.64-IA-SUSY была идентифицирована последовательность ДНК, кодирующая синтез переносчика сахарозы в растениях, которая имеет вид, как представлено ниже. Последовательность N 1 для переносчика сахарозы в шпинате (Seq. ID N 1) и последовательность N 2 (Seq. ID N 2) для переносчика сахарозы из картофеля приведены в конце описания изобретения.
Идентифицированные последовательности ДНК могут быть введены в плазмиды и далее объединены с контролирующими элементами экспрессии в эукариотических клетках (см. Пример 5). Указанные контролирующие элементы являются, с одной стороны, промоторами транскрипции, а, с другой стороны, терминаторами транскрипции. С целью экспрессии транслируемой мРНК для синтеза в клетках переносчика сахарозы или с целью экспрессии нетранслируемой РНК, препятствующей синтезу в клетках эндогенного переносчика сахарозы, эукариотические клетки могут быть трансформированы с помощью последовательностей ДНК настоящего изобретения, содержащихся в плазмидах. Посредством экспрессии РНК, соответствующих последовательностям настоящего изобретения, которые определяют синтез переносчика олигосахаридов, представляется возможным модифицировать метаболизм углеводов таким образом, чтобы улучшить доставку запасных веществ в собираемые части растений и таким образом повысить продуктивность сельскохозяйственных культур растений. Возможность воздействовать на поглощение запасных веществ отдельными органами позволяет так изменить метаболизм всего растения, что рост определенных тканей, например листьев, можно замедлить, тогда как рост собираемых частей растений увеличить. Для этой цели представляется существенным удлинение вегетативной фазы роста культур растений, что должно привести к повышению уровня образуемых запасных веществ.
К настоящему времени имеется способ, позволяющий достигнуть генетической модификации двудольных и однодольных растений (см., например, Gasser, C.S., Fraley, R.T., 1989 г., Science 244:1293-1299; Potrykus, 1991, Ann Rev Plant Mol Physiol 42:205-225).
Для экспрессии в растениях кодирующие последовательности должны быть связаны с регуляторными элементами. Такие элементы, называемые промоторами, известны (EP 375091).
Далее для осуществления верной транскрипции кодирующие регионы должны быть обеспечены терминирующими сигналами транскрипции. Такие элементы также уже описаны (Gielen et al., 1989, EMBO J.8:22-29).
Стартовый регион транскрипции может быть как нативным и/или гомологичным, так и чужеродным и/или гетерологичным в отношении растения-хозяина. При необходимости, терминирующие кодоны могут заменять друг друга. Последовательности ДНК стартового и терминирующего регионов транскрипции могут быть синтезированы, получены из естественных источников или из смеси синтетических и натуральных составляющих ДНК. Для введения чужеродных генов в высшие растения должны быть доступны большое число векторов, которые включают, в том числе, сигнал репликации для E. coli и маркер для селекции трансформированных клеток. Примерами таких векторов являются pBR 322, pUC-Серии, M13 mp-Серии, pACYC 184 и т.д. В зависимости от способа введения желаемого гена в растения, могут быть приемлемы также другие последовательности ДНК. При использовании для трансформации растительных клеток Ti- или Ri-плазмид, по крайней мере, правый конец, а чаще и правый, и левый концы Т-ДНК Ti- и Ri-плазмид присоединяются, как фланкирующая область, к вводимому гену. Возможность использования Т-ДНК для трансформации клеток интенсивно изучалась и подробно описана в EP 120516 (Hoekama, In: The Binary Plant Vector System, Offset-drukkеrij Kanters B.V. Aеblasserdam (1985), Shapter V; Fraley, et al. , Crit. Rev Plant Sci, 4:1-46 and An et al., (1985) EMBO J.4:277-287.) Поскольку вводимая ДНК интегрируется в геном, она, как правило, стабильно сохраняется в нем и остается также в потомстве первоначально трансформированных клеток. В норме она содержит селективный маркер, который придает трансформируемым клеткам устойчивость к биоциду или антибиотикам, таким, как канамицин, G418, блеомицин, гигромицин или фосфинотрицин и т.д. Используемый селективный маркер должен позволять производить отбор трансформированных клеток от клеток, утративших введенную ДНК.
Для введения ДНК в хозяйскую клетку, помимо трансформации с использованием Agrobacteria, существует множество других доступных способов. Эти способы включают слияние протопластов, микроинъекцию ДНК и электропорацию, а также баллистические методы и вирусную инфекцию.
В подходящей среде, содержащей биоциды или антибиотики для селекции, может быть регенерировано целое растение. А полученные растения следует проверить на наличие введенной ДНК. Процессы инъекции плазмид или электропорации не требуют создания каких-либо специальных условий. Могут быть использованы простые плазмиды типа pUC-производных. Однако, при регенерации из таких трансформированных клеток целого растения необходимо присутствие селективного маркерного гена. Трансформированные клетки проращивают внутри растений с помощью известной процедуры (см. Mc Cormick et al. (1986) Plant Cell Reports 5:81-84).
Эти растения выращивают в нормальных условиях и затем скрещивают с другими растениями, обладающими теми же самыми или отличными от них трансформированными генами. Полученные гибридные экземпляры обладают соответствующими фенотипическими признаками.
Последовательности ДНК настоящего изобретения могут быть также введены в плазмиды и там объединены с контролирующими элементами для экспрессии в прокариотических клетках. Возможность получения из бактерий транслируемой РНК для последовательности, определяющей синтез переносчика сахарозы в эукариотических клетках, приводит, неожиданно, несмотря на существование значительных различий в мембранных структурах прокариотических и эукариотических клеток, к приобретению прокариотами способности усваивать сахарозу. Это явление создает основу для продукции интересных в техническом отношении бактериальных штаммов, способных расти на относительно дешевом субстрате, каким является сахароза (см. Пример 5). Так, к примеру, в литературе описано получение полиоксимасляной кислоты с использованием бактерий Alkaligenes eutrophus (Steinbuchel & Schubert, 1989, Arch Microbiol 153:101-104). Однако, используемый в данном методе вид бактерий в состоянии потреблять только очень ограниченный круг субстратов. Кроме прочих аспектов, в случае Alkaligenes eutrophus представляет большой интерес экспрессия гена переносчика сахарозы.
Последовательности ДНК настоящего изобретения могут быть также введены в плазмиды, обладающие мутагенным действием и способностью индуцировать модификацию нуклеотидной последовательности за счет рекомбинации последовательностей ДНК эукариотической и прокариотической систем. Таким способом может быть изменена специфичность переносчика сахарозы.
Используя стандартные методы (Sambrook et al., 1989, Molecular Cloning: A Laboratory Manual, 2 Edn, Cold Spring Harbor Press; NY, USA), производят замену оснований или добавляют естественные или синтезированные последовательности кДНК. Для связывания фрагментов ДНК друг с другом к их смеси добавляются адаптер или линкеры. Кроме того, необходимо обработать нуклеотидную последовательность, подготовив соответствующим образом сайты для рестрикционного расщепления, либо удалить избыток ДНК. Для осуществления инсерций, делеций или замещений нуклеотидных последовательностей, таких, как транзиции или трансверсии, может быть использована техника мутагенеза in vitro, репарации ДНК с помощью праймера, рестрикции или лигирования. Для анализа, в основном анализа нуклеотидной последовательности, может быть использован рестрикционный анализ, равно, как и другие биохимические, молекулярные и биологические методы. При каждой манипуляции используемая последовательность ДНК расщепляется и связывается с другой последовательностью ДНК. При этом каждая плазмидная последовательность может быть клонирована в тех же самых или отличных от них плазмидах.
Производные или отдельные части, в том числе плазмидные, последовательностей ДНК, составляющих предмет настоящего изобретения, могут быть также использованы для трансформации прокариотических или эукариотических клеток. Кроме того, последовательности ДНК настоящего изобретения могут быть использованы с применением стандартных методик для выделения сходных последовательностей из генома различного вида растений, также кодирующих синтез транспортных молекул для сахарозы или других олигосахаридов. С использованием этих последовательностей могут быть получены конструкции для трансформации клеток с целью модификации процесса транспортировки в трансгенных растениях.
Для определения родственных последовательностей ДНК, создаются, прежде всего, генные библиотеки, отражающие репрезентативно содержание генов в растениях данного типа или экспрессию генов в растениях данного типа. Первая коллекция представляет собой геномную библиотеку генов, тогда как вторая - библиотеку генов кДНК. На их основе далее могут быть получены родственные последовательности с использованием последовательностей настоящего изобретения в качестве проб. При этом, как только будет идентифицирован и выделен родственный ген, становятся возможными определение последовательности и анализ белков, кодируемых этими последовательностями.
Для лучшего понимания примеров, иллюстрирующих основы настоящего изобретения, ниже перечислены все известные к настоящему времени методы, применявшиеся для исследования:
1. Способ клонирования:
Для клонирования использовали фаг Ламбда ZAPII и вектор p BluescriptSK (Short et al., 1988, Nucl Acids Res 16:7583-7600). Для трансформации дрожжей применяли вектор YIplac 128 и YEplac 112 (Gietz & Sugio, 1988, Geue 74: 527-534). Для трансформации растений клонировали генные конструкции с помощью бинарного вектора p Bin AR HoRgen & Willnutzer, 1990, Plant Sci. 66: 221-230).
2. Бактериальные и дрожжевые штаммы
Для вектора p Bluescript SK и для p Bin AR конструкций использовался штамм E. coli XLI blue (Bullock et al., 1987, Biotechniques, 5, 376-378). В качестве стартового штамма для получения дрожжевого штамма YSH 2.64-IA-SUSY настоящего изобретения использовался штамм YSH 2.64-IA (Gozalbo & Hohmann, 1990, Curr Genet 17:77-79).
Трансформация плазмиды в растения картофеля выполнялась с использованием Agrobacterium tumefacilus штамм LBA4404 (Bevan, 1984, Nucl. Acids Res 12: 8711-8720).
3. Трансформация Agrobacterium tumefaciens.
Транспорт ДНК к Agrobacteria выполнялся посредством прямой трансформации по методу Хефгена и Вильмитцера (Hofgen & Wilmitzer 1988, Nucl. Acids Res 16: 9877). Плазмидная ДНК трансформированной Agrobacterium выделялась в соответствии с методом Бирнбоима и Доли (Birnboin & Doleg 1979, Nucl. Acids. Res 7:1513-1523) и анализировалась с помощью гель-электрофореза после соответствующего рестрикционного расщепления.
4. Трансформация растений
10 маленьких листьев, поврежденных скальпелем, из стерильной картофельной культуры помещали в 10 мл MS среды, содержащей 2% сахарозы и 30-50 мкл ночной культуры Agrobacterium tumеfaciens, росшей в условиях селекции. После 3-5 минут осторожного перемешивания листья вынимали и перекладывали в MS среду, содержащую 1,6% глюкозы, 2 мг/л зеантинрибозы, 0,62 мг/л нафтилуксусной кислоты, 0,02 мг/л гибереллиновой кислоты, 500 мг/л клафорана, 50 мг/л канамицина и 0,8% бактоагара. После инкубации в течение одной недели при температуре 25oC и освещении в 3000 люкс концентрация клафорана в среде была уменьшена наполовину.
Хранение
Приведенные ниже плазмиды и дрожжевые штаммы были депонированы в Германской Коллекции Микроорганизмов (DSM) в Брауншвейге, Германия,
12.06.92 - (депозитный номер):
Плазмида p SK-S21 - (DSM 7115)
Дрожжевой штамм YSH 2.64-1A-SUSY - (DSM 7106)
Дрожжевой штамм YSH 2.64-1A-INY - (DSM 7105)
Описание чертежей.
Фиг. 1. Временное распределение потребления сахарозы дрожжевыми клетками. "пмол/мг клеток" = количество 14C-меченной сахарозы в пмол, поглощенной на единицу влажного веса дрожжевых клеток в мг; "112"=стартовый штамм (без переносчика); "+глюкоза"=преинкубация клеток в среде, содержащей 10 мМ глюкозы; "-глюкоза"= отсутствие преинкубации клеток; "2,5 μM СССР"=коинкубация с ингибитором карбонилцианид-m -хлорфенилгидразоном (СССР) в концентрации 2,5 μM; "500 μMPCMBS″ =совместная инкубация с ингибитором п-хлормеркурийбензолсульфокислотой (PCMBS) в концентрации 500 μM; "10 μM Антимицина" = совместная инкубация с ингибитором антимицином в концентрации 10 μM.
Фиг. 2. Клонирование плазмиды pMA5-INY. Показывает инсерцию Hind III фрагмента размером 2,4 н.п. из pSEYC 306-1 в pMAS-10.
Фиг. 3 показывает плазмиду pBin AR-S21. На ней: CaMV 35S промотор - промотор гена для 35 S РНК вируса мозаики цветной капусты; A - кодирующий регион для транспортной молекулы, переносящей сахарозу в шпинате, с ориентацией в направлении считывания; OCS - терминатор гена октопинсинтазы из Agrobacterium tumefaciens; SmaI, NotI, Bam HI: расщепляемые рестрикционными ферментами позиции.
Фиг. 4 показывает плазмиду pBin AR-P62-анти. На ней CaMV 35S промотор - промотор гена для 35S РНК вируса мозаики цветной капусты; OCS - терминатор гена октопинсинтазы из Agrobacterium tumefaciens; SmaI, SacI, Bam HI, XbaI: позиции, расщепляемые рестрикционными ферментами.
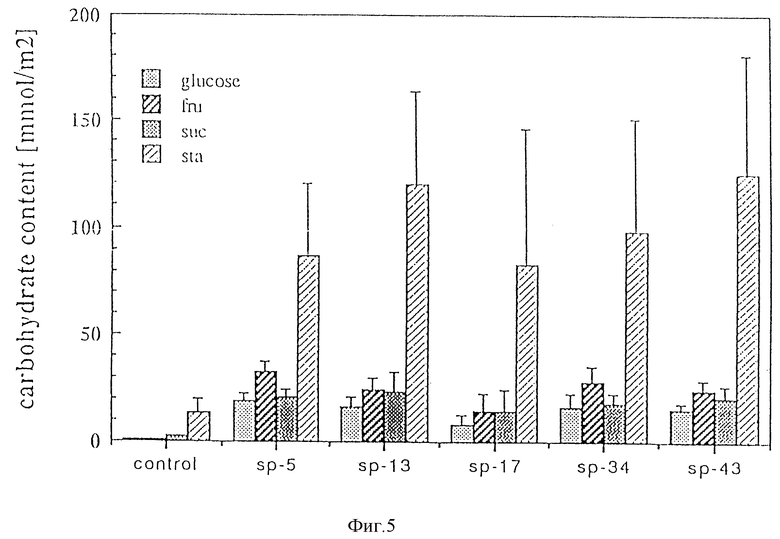
Фиг. 5. Содержание различных углеводов в листьях растений, трансформированных Bin AR-P62-анти. На ней fru - фруктоза; suc - сахароза; sna - крахмал; контроль - нетрансформированные исходные растения; sp-5 до sp-43 - трансформанты с индивидуальными номерами.
Фиг. 6. Выход углеводов из трансформантов Bin AR-P62-анти черешков; wt - дикий тип; sp-5 до sp-43 - трансформанты с индивидуальными номерами.
Приведенные ниже примеры описывают приготовление дрожжевых штаммов, идентификацию их и их функционирование, а также использование растительного переносчика сахарозы.
Пример 1
Приготовление дрожжевых штаммов YSH 2.64-IA-SUSY и YSH 2.64-IA-INY
Дрожжевой штамм YSH 2.64-IA обладает структурами suc2-, ma10, leu2, trp1 (Gozalbo & Hohmann, 1990, Curr Genet 17:77-79).
Для введения в данный штамм энзиматической активности, направленной на расщепление сахарозы, он был трансформирован с помощью интегрируемой плазмиды YipIac128A2-SUSY, которая содержит кодирующий регион для синтазы сахарозы из картофеля, слитой с промотором гена алкогольдегидрогеназы из дрожжей. Плазмида Yiplac128A2-SUSY приготавливалась следующим образом. Плазмида YIpIac 128 (Gietz & Sugine 1988, Geue 74:527-534) укорачивается путем расщепления с помощью PstI и EcoRI, дальнейшей деградации и/или введения в единичные нити с концевой избыточностью и лигирования в регионе полилинкера. В оставшуюся позицию расщепления SphI вводится кассета размеров в 728 н.п. с промотором алкогольдегидрогеназы из плазмиды pYT102U (Vernet et al., 1987, Geue 52:225-233).
Таким способом была получена плазмида VIpIac128A2. Кодирующий регион для синтазы сахарозы был амплифицирован с помощью полимеразной цепной реакции со следующими олигонуклеотидами SUSYI (ГАГАГАГГАТЦЦТГЦААТГТЦТГААЦГТГТТТТГАЦТЦГТГ) и SUSY2 (ГАГАГАГГАТЦЦТТЦАТТЦАЦТЦАГЦАГЦЦААТГГААЦАГЦТ)в ламбда клон синтазы сахарозы из картофеля (Salanoubad & Belliarol, 1987, Geue 60:47-56). Из этого продукта был получен в виде BamHI фрагмента кодирующий регион и вставлен в сайт расщепления BamHI полилинкера кассеты. Дрожжевой штамм YSH 2.64-IA был трансформирован с помощью приготовленной таким образом плазмиды VIpIac128A2-SUSY. Поскольку плазмида не содержит 2μ региона, она не может автономно реплицироваться в дрожжах. Поэтому такие трансформанты приобретают только признак ауксотрофности по лейцину, трансформанты, которые хотя бы частично интегрировали в свои хромосомы плазмидную ДНК.
Колонии, аутотрофные по лейцину, исследовались на экспрессию гена синтазы сахарозы. С этой целью клетки из 5 мл жидкой культуры были разрушены с помощью добавленных стеклянных шариков при интенсивном помешивании, затем, после центрифугирования, общий белок из супернанта был добавлен в систему для измерения энзиматической активности. Активность экспрессируемой синтазы сахарозы содержит 25 mU/мг белка.
Сходным образом вводилась инвертазная активность в дрожжевой штамм YSH 2.64-IA, в геном которого с помощью плазмиды YIpIac128AI-INY был интегрирован ген цитозольной, нерасщепляющей инвертазы. YIpIac128AI-INY содержит вместо кодирующего региона для синтазы сахарозы инвертазный ген, лишенный сигнальной последовательности для вывода генного продукта. Предшественником этой плазмиды является плазмида YIpIac128AI, которая отличается от YIpIac128A2 только ориентацией полилинкера в кассете относительно промотора алкогольдегидрогеназного гена. Кассета для этой плазмиды была получена из плазмиды pYT100U (Vernet et al., 1987, Geue 52; 225-233). Кодирующий регион для инвертазы был получен на ДНК suc2 гена с помощью полимеразной цепной реакции с олигонуклеотидами
INY3 (ГАГЦТГЦАГАТГГЦАААЦГАААЦТАГЦАТАГАЦЦТТТГГТЦАЦА)
и
INY4 (ГАГАЦТАГГТТАТААЦЦТЦТАТТТТАЦТТЦЦЦТТАЦТТГГАА), Кодирующий регион был лигирован в виде PstI/Spe1 фрагмента в линейном векторе YIpIac128AI den с использованием PstI и XbaI. Тест на наличие энзиматической активности экспрессируемой в дрожжевых клетках инвертазы выявил величину энзиматической активности, равную 68 mU/мг общего белка.
Пример 2. Клонирование кДНК переносчика сахарозы в растениях
На основе полиаденилированной РНК из ткани листьев растений шпината и картофеля была создана библиотека кДНК в фаге Ламбда ZAP II. Исходя из 5000000 Pfu, была приготовлена фаговая ДНК и очищена с использованием градиента цезия хлорида- саркозила. После расщепления фаговой ДНК с помощью фермента NotI инсерции в регионы размером выше и ниже 1,5 кило н.п. были приготовлены на 1% геле агарозы и лигированы на NotI расщепляемых сайтах вектора экспрессии YEpIac112AINE. Этот вектор является производным вектора YEpIac112 (Gozalbo & Hohmann 1990, Curr Geuet 17:77-79), с которым, как описано в Примере 1, полилинкер обменивался кассетой, содержащей промотор гена алкогольдегидрогеназы. Полилинкер кассеты был снова удален расщеплением с помощью фермента PstI и XbaI и замещен двунитевым олигонуклеотидом, который вводит расщепляемые сайты для NotI и EcoRI
(Последовательность: ГАТЦЦГЦГГЦЦГЦЦЦГТААТТЦТЦТАГАЦТГЦА).
Около 9000'00 клонов для региона размером менее 1,5 кило н.п. и около 40000 клонов для региона размером выше 1,5 кило н.п. были получены при трансформации E.coli. На их основе была получена плазмидная ДНК. 2 μг ДНК трансформировали четырнадцать раз в дрожжевой штамм YSH 2.64-IA suc-SUSY.
Трансформанты выращивали на среде, содержащей 2% глюкозы, затем от пяти до десяти тысяч трансформантов смыли жидкой агар-содержащей средой и высеяли на среду, содержащую сахарозу. Колонии, выявившие способность к более быстрому росту, проанализировали. Инсерции в вектор YEpIac112INH трансформантов S21 или P62 (YЕрIac112AINE-S21) были секвенированы. Последовательности представлены выше.
Пример 3
Анализ дрожжевых трансформантов, метаболизирующих сахарозу
Дрожжевые трансформанты YEpIac112AINE-S21, соответствующие таковым, полученным в Примере 2, выращивали в жидкой среде вплоть до достижения культурой логарифмической фазы. После центрифугирования культуры клетки преинкубировали в течение 5 минут в среде, содержащей глюкозу, и затем перенесли на среду, содержащую сахарозу, меченную 14C. Потребление меченной сахарозы измерялось способом, описанным Циримо (Cirillo, 1989, Meth Enzymol 174:617-622). Потребление меченной сахарозы без преинкубации с глюкозой сравнивалось с таковым в случае совместной инкубации с ингибиторами карбонилцианид-м- хлорфенилгидразоном (СССР), n-хлормеркурийбензолсульфокислотой (PCMBS) и антимицином. Временное распределение показано на фиг. 1. Вычисленное снижение потребления сахарозы в присутствии ингибиторов представлено в таблице I. Конкурентный эксперимент с применением различных сахаров в качестве конкурентов меченной сахарозы, на основе которого может быть получена информация о специфичности переносчика, показан в таблице II.
Аналогичные измерения были выполнены с использованием дрожжевого штамма YEpIac112AINE-P62. Эти исследования дали сходные результаты.
Пример 4
Трансформация бактериальных штаммов последовательностями ДНК для экспрессии переносчика сахарозы
Для приобретения способности метаболизировать потребляемую сахарозу бактериальные клетки нуждаются в энзиматической активности для расщепления моносахаридов. Для введения такой активности бактерии трансформировали с помощью плазмиды pMA5-INY и исследовали на наличие инвертазной активности. Плазмида pMA5-INY приготавливалась следующим образом. Плазмида pMA5-10 (Stanssens et al., 1989, Nucl. Acids Res 17:4441-4454) была превращена в линейную в расщепляемом сайте Hind III полилинкера. Hind III фрагмент размером 2,4 кв плазмиды pSEYC306-1 был клонирован в сайте расщепления III (Taussig & Carlson 1983, Nucl. Acids Res II:1943-1954). Соответствующий схематический разрез плазмиды показан на фиг. 2. Энзиматическая активность инвертазы в бактериальных клетках, трансформированных плазмидой pMA5-INY, определяли с помощью хорошо известного способа на основе гель-электрофореза Возможность образования функционально активного переносчика сахарозы при экспрессии растительной кДНК в бактериальных клетках исследовалась при трансформации E. coli плазмидой pSK-S21. Плазмида описана в Примере 3. После трансформации бактериальных клеток с помощью pSK-S21 было проведено исследование способности к потреблению сахарозы.
Пример 5
Трансформация растений с помощью конструкций для сверх-экспрессии кодирующего региона переносчика сахарозы
С помощью векторов YEpIac 112AINE-S21 и YEpIac 112AINE-P62, которые содержат в качестве инсерций кДНК для переносчика сахарозы из шпината и/или картофеля (см. Примеры 2 и 3), инсерции после расщепления NotI были выделены и лигированы в сайтах расщепления NotI на плазмидах pBluescript-SK (pSK-S21 и/или pSK-P62). Для pSK-S21 инсерция была приготовлена в виде SacI/XbaI фрагмента и клонирована после введения в однонитевую ДНК с избыточностью концов в "смысловой" ориентации на pBin AR (Hofgen & Wilmitzer, 1990, Plant Sci 66: 221-230), который предварительно был расщеплен ферментами SmaI и XbaI. Полученная плазмида pBin AR-S21 (см. фиг. 3) может быть использована для введения в растения с целью их трансформации в плане создания условий для сверх-экспрессии переносчика сахарозы. Для pSK-P62 инсерция была выделена в виде NotI фрагмента размером 1,7 кило н.п. и клонирована в "антисмысловой" ориентации на сайте расщепления SmaI бинарного вектора pBin AR-P62-анти (см. Рис.4).
Эта плазмида подходит для целей трансформации растений в плане достижения "антисмыслового" ингибирования экспрессии переносчика сахарозы. Далее с целью инфекции листовых сегментов табака или картофеля была применена трансформация с помощью Agrobacteria.
На десяти независимо полученных трансформантах, в которых с помощью Саузерн-блоттинг-анализа продемонстрировано наличие интактного неперестроенного химерного гена, были исследованы изменения в содержании сахарозы, гексозы и крахмала соответственно.
Трансформанты, содержащие Т-ДНК из pBin AR-P62-анти, показали значительное повышение концентрации крахмала, гексоз и сахарозы в листьях (см. фиг. 5). Выделение углеводов из черешков, взятых из растений, значительно уменьшается в водной среде (см. фиг. 6). Из этих данных хорошо видно значение переносчика сахарозы для транспорта фотоассимилированных веществ из фотосинтетически активных органов. Поскольку ингибирование активности переносчика ограничивает транспорт углеводов, это приводит к снижению транспорта фотоассимилятов к органам хранения в случае сверх- экспрессии, например, при использовании плазмиды pBin AR-S21. Растения табака, в которые Т-ДНК из pBin AR-S21 была интегрирована, демонстрируют снижение апикального доминирования, т. е. приобретают способность к кустовому росту. Такое фенотипическое изменение может быть в некоторых случаях весьма желательно, к примеру, для растений томата. Плазмида pBin AR-S21 с последовательностью ДНК настоящего изобретения (Seq. ID N 1) подходит в этой связи для модификации растений с целью улучшения важных для выращивания характеристик, таких, как кустообразный рост.

















Фрагменты ДНК, кодирующие переносчик олигосахаридов в растении, предназначены для трансформации растений. Введение фрагментов ДНК в геном модифицирует образование и транспорт запасных веществ как в трансгенных растениях, так и в плазмидах и бактериях. Способы модификации метаболизма углеводов, снижения апикального доминирования в тканях растения, получения бактерии и выделения фрагментов ДНК, кодирующих переносчик олигосахаридов, реализуются с использованием дрожжевых штаммов. 11 с. и 10 з.п. ф-лы, 6 ил., 2 табл.
| Способ получения спиртов и/или простых эфиров | 1972 |
|
SU442592A1 |
| БИОХИМИЯ РАСТЕНИЙ | |||
| Пер | |||
| с англ | |||
| / Под ред | |||
| В.А | |||
| Кретовича | |||
| - М.: Мир, 1968, с.129 - 136. | |||

