Настоящая заявка является частично продолженной заявкой относительно заявки США с серийным номером 08/136,783 от 14 октября 1993 г., которая является продолжением заявки США с серийным номером 08/117.366 от 7 сентября 1993 г., причем на обе эти заявки ссылаются в настоящем изобретении.
Настоящее изобретение, главным образом, относится к области слитых белков, а также к белкам, используемым для лечения и диагностики болезненных состояний, связанных с действием IL 4 и избытком продуцирования IgE, более конкретно настоящее изобретение относится к химерным и гуманизированным IL 4 антителам.
Область атопических аллергических заболеваний охватывает как относительно незначительные болезни, например, такие как сезонные риниты и коньюктивиты, так и более серьезные заболевания, такие как атопический дерматит и атопическая астма, а также такие болезни, угрожающие жизни, как анафилактический шок. Общим фактором таких болезненных состояний является иммунный ответ организма на аллергены, причем такая реакция заключается в продуцировании иммуноглобулиновых E (IgE) антител у генетически предрасположенных пациентов (атопия). Ингибирование продукции IgE в течение длительного времени составляло цель специфической иммунотерапии аллергических заболеваний с использованием десенсибилизирующих вакцин. Однако в последние годы безопасность и эффективность вакционной терапии стала вызывать сомнения, однако, потребность понижения уровня IgE не отпала.
Интерлейкин 4 (IL 4) представляет собой протеиновый медиатор в лимфоидной системе. Исследования лимфоцитов, взятых у атопических пациентов, позволило обнаружить повышенное количество Т-лимфоцитов, обладающих способностью секретировать IL 4 в ответ на стимуляцию и повышенные количества IL 4, секретированного после стимуляции.
Было обнаружено, что анти-IL 4 антитело ингибирует IgE, но не ингибирует IgG1 или IgG2a /Finkelman с сотр., Ann. Rev. Immunol. 8:303(1990)/, а также продукцию IL 5 секретирующих Т-клеток /Maggi с сотр., J. Immunol. 148:2142 (1992)/. Кроме этого, последние данные подтвердили, что IL 4 может оказывать влияние на аккумуляцию эозинофила в тканях. См., например, Tepper с сотр. Cell, 62:457(1990); Tepper с сотр. Cell, 57; 503(1989).
В данной области техники в настоящее время сохраняется необходимость в создании высокоаффинного IL 4 антагониста, способного снижать уровень зозинофильного воспаления в результате уменьшения пролиферации IL 5 секретирующих клеток и ингибирования механизма адгезии, в результате чего зозинофилы могли бы аккумулироваться в тканях и использоваться для лечения, профилактики и диагностики аллергических реакций.
В соответствии с первым аспектом настоящее изобретение обеспечивает слитый белок, обладающий сродством к связыванию интерлейкина-4 человека, который содержит участки, определяющие комплементарность (CDR) нечеловеческого нейтрализующего моноклонального антитела (Mab), характеризующегося константой диссоциации, равной или меньшей 2·10-10 М для человеческого IL 4, и первый партнер слияния, в котором, по крайней мере, один и предпочтительно все участки, определяющие комплементарность (CDR) замещены CDR моноклонального антитела нечеловеческого происхождения (Mab). Такое нейтрализующее моноклональное антитело нечеловеческого происхождения может быть выбрано из группы, состоящей из 3В9 и 6A1, в соответствии с тем, что более полно описано в подробном описании изобретения. Предпочтительно, когда слитый белок оперативно связан со вторым слитым белком, включающим всю или часть иммуноглобулиновой константной цепи.
В соответствии с родственным аспектом настоящее изобретение обеспечивает CDR нейтрализующего моноклонального антитела нечеловеческого происхождения (Mab), характеризующегося константой диссоциации, равной или меньшей 2·10-10 М для человеческого IL 4 и молекулы нуклеиновой кислоты, кодирующие такие CDR.
Согласно другому аспекту настоящее изобретение предусматривает гуманизированные (humanized) антитела, содержащие, по крайней мере, один и предпочтительно шесть участков, определяющих комплементарность (CDR), являющихся производными нейтрализующих моноклональных антител нечеловеческого происхождения (Mab), характеризующихся константой диссоциации, равной или меньшей 2·10-10 М для человеческого IL 4.
Согласно еще одному аспекту предусматривается химерное антитело, содержащее человеческие константные области тяжелой и легкой цепей и вариабельные области тяжелой и легкой цепей, произведенные из нечеловеческих нейтрализующих моноклональных антител (Mab), характеризующихся константой диссоциации, равной или меньшей 2·10-10 М для человеческого IL 4.
Еще один аспект настоящего изобретения относится к фармацевтической композиции, которая содержит один (или более) из указанных выше слитых протеинов или Mab (например, гуманизированных, химерных и т.п.) и фармацевтически применимый носитель.
Следующий аспект настоящего изобретения предусматривает метод лечения и/или профилактики аллергических состояний людей в результате применения эффективного количества фармацевтической композиции изобретения.
Другой аспект настоящего изобретения предусматривает способы и компоненты, применяемые для рекомбинантного продуцирования слитых белков, Mab (например, гуманизированных, химерных и т.п.), их CDR, Fab или F (ab')2 или их аналогов, являющихся производными нечеловеческих нейтрализующих моноклональных антител (Mab), характеризующихся константой диссоциации, равной или меньшей 2·10-10 М для IL 4. Такие компоненты включают изолированные кодирующие нуклеиново-кислотные последовательности, рекомбинантные плазмиды, содержащие нуклеиново-кислотные последовательности, контролируемые выбранными регуляторными последовательностями, способными направлять их экспрессию в клетках-хозяевах, а также клетки-хозяева (предпочтительно млекопитающих), трансфицированные рекомбинантными плазмидами. Способ получения включает культивирование трансфицированной линии клетки-хозяина настоящего изобретения в таких условиях, что антитело, предпочтительно гуманизированное (humanized) антитело, экспрессируется в указанных клетках, и выделение продукта экспрессии.
Еще один аспект настоящего изобретения охватывает способ диагностики аллергий и других болезненных состояний, связанных с избытком образования иммуноглобулина E в организме человека, заключающийся в контактировании образца биологической жидкости со слитыми протеинами, Mab's (например, гуманизированными, химерными и т.п.) и Fab's настоящего изобретения и анализе реализации связывания между указанными слитым протеином, Mab или Fab и человеческим интерлейкином 4.
Еще один аспект предусматривает способ скрининга моноклональных антител, обладающих высоким титром по отношению к человеческому интерлейкину 4, который включает:
a) получение гибридомной клеточной линии, отличающейся секретированием моноклонального антитела по отношению к человеческому интерлейкину 4; и (b) скрининг указанной гибридомной клеточной линии с альдегид-коньюгированным интерлейкином-4 или биотинилированным человеческим интерлейкином-4. Предпочтительно проводят скрининг гибридомной клеточной линии с биотинилированным человеческим интерлейкином-4.
Настоящее изобретение также предусматривает нейтрализующие Mab, обладающие высоким сродством к IL 4, их Fab фрагменты или F(ab)2 фрагменты, полученные скринингом библиотеки гибридомных продуктов в присутствии альдегид-коньюгированного человеческого интерлейкина-4 или биотинилированного человеческого IL 4.
Согласно другому аспекту настоящее изобретение предусматривает нейтрализующие моноклональные антитела грызунов, специфичные к человеческому интерлейкину-4 и обладающие сродством к связыванию, характеризующиеся константой диссоциации, равной или меньшей 2·10-10 М. Примерами таких моноклональных антител могут служить мышиные Mab, 3В9, крысиные Mab, 6A1 и другие Mab с идентичными характеристиками (т.е. способностью связываться с эпитопами 3В9 или 6A1 при специфичности к человеческому IL 4 и константой диссоциации, равной или меньшей 2·10-10 М). Другим аспектом настоящего изобретения является гибридома 3426A11C1B9.
Другие аспекты настоящего изобретения представлены ниже в подробном описании его предпочтительных воплощений.
На фиг. 1 (последовательности SEQ ID N: 1,2) проиллюстрирован вариабельный участок легкой цепи (аминокислоты 21-132) мышиного 114 антитела 3В9, человек/мышь 3В9 химерное антитело, а также нативная сигнальная последовательность (аминокислоты 1-20). Подчеркнутые участки обозначают CDR (Последовательности SEQ ID N: 15, 16; SEQ ID N: 17 и 18; и SEQ ID N: 19 и 20).
На фиг. 2 (SEQ ID N: 3 и 4) проиллюстрированы вариабельный участок тяжелой цепи (аминокислоты 20-140) мышиного 3В9, а также нативная сигнальная последовательность (аминокислоты 1-19). Подчеркнутые участки обозначают CDR (SEQ ID N:21, 22; SEQ ID N: 23 и 24; и SEQ ID N: 25,26).
На фиг. 3 (SEQ ID N: 9, 10) проиллюстрирован вариабельный участок тяжелой цепи (аминокислоты 21-141) человек/мышь 3В9 химерного антитела и его сигнальная последовательность (аминокислоты 1-19; SEQ ID N: 5 и 6). Подчеркнутые части обозначают CDR, являющиеся производными 3В9 (SEQ ID N: 21, 22; SEQ ID N: 23, 24 и SEQ ID N: 25, 26).
На фиг. 4 проиллюстрированы (SEQ ID N: 11, 12) вариабельный участок тяжелой цепи (аминокислоты 20-141) гуманизированного 3В9 антитела с сигнальной последовательностью (аминокислоты 1-19; SEQ ID N: 5, 6). Подчеркнутые участки обозначают CDR, являющиеся производными 3В9 (SEQ ID N: 54, 22; SEQ ID N: 55, 24; и SEQ ID N: 56 и 26).
На фиг. 5 (SEQ ID N: 13, 14) проиллюстрирован вариабельный участок легкой цепи (аминокислоты 21-131) гуманизированного 3В9 антитела и сигнальная последовательность (аминокислоты 1-20; SEQ ID N: 7, 8). Подчеркнутые участки обозначают CDR, являющиеся производными 3В9 (SEQ ID N: 53, 16; SEQ ID N: 17, 18 и SEQ ID N: 27, 28).
На фиг. 6A (SEQ ID N: 5, 6) показана сигнальная последовательность тяжелой цепи, используемая в приведенном ниже примере 4.
На фиг. 6B (SEQ ID N: 7, 8) показана сигнальная последовательность легкой цепи, используемая в представленном ниже примере 4.
На фиг. 7 дано схематическое изображение плазмиды plL4chhc3-pcd, используемой для экспрессии химерной IL 4 тяжелой цепи в клетках млекопитающих. Такая плазмида содержит бета-лактамазный ген (BETA LAC), точку начала репликации SV40 (SV40), цитомегаловирусную промоторную последовательность (CMV), сигнальную последовательность, химерную вариабельную тяжелую цепь от SEQ ID N 9, 10, константный участок человеческой тяжелой цепи, поли A сигнал бычьего гормона роста (BGH), бетаглобиновый промотор (beto glopro), дигидрофолатредуктазный ген (DHFR) и другая BGH последовательность поли A сигнала в среде pVC19.
На фиг. 8 представлена схема плазмиды pIL 4 chlc-pcdn, используемой для экспрессирования химерного IL 4 легкоцепного вариабельного участка SEQ ID N 1 и 2 в клетках млекопитающего. Эта плазмида отличается от изображенной на фиг. 7 тем, что она содержит химерный легкоцепной вариабельный участок, отличный от химерной тяжелой цепи, постоянный участок человеческой легкой цепи и неомициновый ген (Neo) помимо DHFP.
На фиг. 9 представлена схема плазмиды pIL 4 hzhc-1-pcd, используемой для экспрессии синтетического IL 4 вариабельного участка тяжелой цепи SEQ ID N 11 и 12 в клетках млекопитающего. Такая плазмида отличается от изображенной на фиг. 7 тем, что содержит вариабельный участок гуманизированной тяжелой цепи, отличный от того, который присутствует в химерной тяжелой цепи.
На фиг. 10 представлена схема плазмиды pIL 4hzlc-O-Pcn, используемой для экспрессии вариабельного участка гуманизированной IL 4 легкой цепи SEQ ID N 13 и 14 в клетках млекопитающего. Такая плазмида отличается от той, что изображена на фиг. 8 тем, что содержит вариабельный участок гуманизированной легкой цепи, отличающейся от той, что присутствует в химерной легкой цепи и не кодирует DHFP ген.
Настоящее изобретение обеспечивает ряд антител, их фрагментов и слитых белков, главным образом гуманизированных антител, которые характеризуются связующей специфичностью в отношении человеческого IL 4, нейтрализующей активностью и высоким сродством к человеческому IL 4, как это показано в примерах, относящихся к мышиным Mab или крысиным Mab 6A1. Такие продукты находят применение в терапевтических и фармацевтических композициях для лечения реакций, вызванных действием IL 4 и IgE. Такие продукты также используются для диагностики вызванного действием IL 4 болезненного состояния в результате измерений (например, методом твердофазного иммуноферментного анализа (EL1SA) циркуляционного эндогенного уровня IL 4 у людей.
1. Принятые определения.
Термин "слитый белок" относится к белку, закодированному слитой молекулой, который может быть получен экспрессией в выбранной клетке-хозяине. Такие слитые белки представляют собой генно-инженерные антитела, например химерные или гуманизированные антитела, или фрагменты антитела, в которых полностью или частично отсутствует константный участок иммуноглобулина, например Fy, Fab или F(ab)2 и т.д.
Термин "слитые молекулы" относится к нуклеиново-кислотной последовательности, кодирующей участки, определяющие комплементарность (CDR) нечеловеческого иммуноглобулина, которые вставлены в первый партнер слияния, включающий человеческие вариабельные каркасные последовательности. Иногда первый партнер слияния оперативно связан со вторым партнером слияния.
Термин "первый партнер слияния" относится к нуклеиново-кислотной последовательности, кодирующей человеческий каркас или вариабельный участок человеческого иммуноглобулина, где нативные (или встречающиеся в природе) CDRS замещены CDR донорского антитела. Человеческая вариабельная область может представлять собой иммуноглобулиновую тяжелую цепь, легкую цепь (или обе такие цепи), а также аналог их функциональных фрагментов. Такие CDR или CDR области, находящиеся внутри вариабельных участков антител (иммуноглобулинов), могут детерминироваться (определяться) известными способами. Так, например, Kabat с сотр. /Sequences of Protein of Immunological Interest, 4-е изд. Департамент здравоохранения США, Национальный Институт здоровья (1987)/ описывает правила расположения CDR. Кроме этого известны компьютерные программы идентификации участков или структур CDR.
Термин "высокий титр" относится к антителу с высокой аффиностью связывания, характеризующемуся Kd, равной или меньшей 2·10-10 М для человеческого IL 4.
Под термином "специфичность связывания относительно человеческого IL 4" подразумевается высокий титр (или сродство) в отношении человеческого, но не в отношении бычьего или мышиного IL 4.
Термин "второй партнер слияния" относится к другим нуклеотидным последовательностям, кодирующим протеин или пептид, с которыми коньюгирован первый партнер слияния в рамке или с использованием необязательной традиционной линкерной последовательности (например, путем оперативного связывания). Предпочтительно это вещество представляет собой иммуноглобулин. Второй партнер слияния может включать нуклеиново-кислотную последовательность, кодирующую полный константный район того же самого (например, вариант гомологии, когда первый и второй белки слияния имеют одинаковое происхождение) или дополнительного (например, гетерологичного) нужного антитела. Это вещество может представлять собой иммуноглобулиновую тяжелую или легкую цепи (либо обе такие цепи как часть единого полипептида). Второй партнер слияния не ограничивается конкретным классом иммуноглобулина или изотипом. Кроме этого, второй партнер слияния может включать часть иммуноглобулинового константного участка, такого как Fab или F(ab)2 (например, дискретную часть соответствующего человеческого константного участка или участка каркаса). Такой второй партнер слияния может также включать последовательность, кодирующую интегрально-мембранный протеин, экспонированный на внешней поверхности клетки-хозяина, например, как часть фаговой библиотеки, или последовательность, кодирующую белок в целях диагностики, например пероксидазу хрена, β-галактозидазу и т.п.
Термины Fy, Fc, Fab или F(ab)2 используются в стандартных значениях (см. , например, Harlow с сотр., Antibodies Alaboratore Manual, лаборатория Колд Спринг Харбор (1988).
Используемый в тексте термин "инженерное антитело" относится к типу слитого белка, т.е. синтетического антитела (например, химерного или гуманизированного антитела), в которых часть вариабельных доменов легкой и/или тяжелой цепей выбранного акцепторного антитела замещена на аналогичные части одного или более донорских антител, обладающих специфичностью в отношении выбранного эпитопа. Так, например, указанные молекулы могут включать антитела, характеризующиеся наличием гуманизированной тяжелой цепи, связанной с немодифицированной легкой цепью (либо химерной легкой цепью) или наоборот. Инженерные антитела могут также характеризоваться изменением нуклеиново-кислотных последовательностей, кодирующих вариабельные доменные каркасные участки легкой и/или тяжелой цепей акцепторного антитела с тем, чтобы сохранить специфичность к связыванию донорского антитела. Такие антитела могут включать замену одного или более CDR (предпочтительно всех) от акцепторного антитела на CDR от донорского антитела, описанного в настоящем документе.
Термин "химерное антитело", используемый в данном тексте, относится к типу инженерных антител, которые содержат природный вариабельный район (легкая цепь и тяжелые цепи), произведенный из донорского антитела, ассоциированного с константными районами легкой и тяжелой цепей, произведенных из акцепторного антитела.
Термин "гуманизированное антитело" относится к антителу инженерного типа с его CDR от нечеловеческого донорского иммуноглобулина, причем другие оставшиеся иммуноглобулиновые части молекулы являются производными одного (или более) человеческого иммуноглобулина. Следует отметить, что каркасные опорные остатки могут подвергаться изменению с тем, чтобы сохранить связующую аффинность (см. , например, Queen с сотр., Proc. Natl. Acad. Sci. UCA 86, 10029-10032 (1989), Hodgson с сотр., Bio /Technology, 9:421 (1991).
Используемый в тексте термин "донорское антитело" относится к антителу (поликлональному, моноклональному или рекомбинантному), которое способствует передаче нуклеиново-кислотным последовательностям его вариабельных участков, CDR или других его функциональных фрагментов или аналогов к первому партнеру слияния, в результате чего образуется слитая молекула и экспрессированный слитый протеин с антигенной специфичностью и нейтрализующей способностью, характерными для донорского антитела. Примером донорского антитела, подходящего для использования в настоящем изобретении, может служить нечеловеческое нейтрализующее моноклональное антитело (например, мышиное), обозначаемое как 3В9. Антитело 3В9 определяется, как обладающее высоким титром, человеческий-IL 4 специфичное (т.е. не распознающее бычий или мышиный IL 4), нейтрализующее антитело изотипа IgG1, имеющее ДНК вариабельной легкой цепи и аминокислотные последовательности SEQ ID N 1 и 2, а также ДНК вариабельной тяжелой цепи и аминокислотные последовательности SEQ ID N 3 и 4 в константной области подходящего мышиного IgG.
Термин "акцепторное антитело" относится к антителу (поликлональному, моноклональному или рекомбинантному), гетерологичному донорскому антителу, которое способствует передаче всех (или любой их части, но предпочтительно всех) нуклеиново-кислотных последовательностей, кодирующих каркасные районы его тяжелой и/или легкой цепи и/или константные районы его тяжелой и/или легкой цепи второму партнеру слияния. Предпочтительно человеческое антитело представляет собой акцепторное антитело.
Термин "CDRS" относится к определяющему комплементарность участку аминокислотных последовательностей антитела, которые представляют собой гипервариабельный участок иммуноглобулиновых легких и тяжелых цепей. См., например, Kabat с сотр. Последовательности протеинов, имеющих иммунологическое значение, 4-е изд., Департамент здравоохранения США, Национальный институт здоровья (1987). Имеется по три CDR (или CDR районов) в тяжелой и легкой цепях в вариабельной части иммуноглобулина. В связи с этим, используемый в тексте термин "CDR" относится ко всем трем CDR тяжелой цепи или всем трем CDR легкой цепи (либо ко всем CDR как легкой, так и тяжелой цепей).
CDR обеспечивают множество константных остатков для связывания антитела с антигеном или эпитопом. CDR настоящего изобретения являются производными последовательностей вариабельных тяжелой и легкой цепей донорского антитела и содержат аналоги встречающихся в природе CDR, причем такие аналоги обладают частью или сохраняют ту же антигенсвязующую специфичность и/или нейтрализующую способность, что и донорское антитело, из которого они произошли.
Под термином "распределение антигенсвязующей специфичности или нейтрализующей способности" предполагается, например, что хотя Mab 3В9 может характеризоваться некоторым уровнем антигенного сродства, а CDR, закодированный нуклеиново-кислотной последовательностью 3В9 в подходящем структурном окружении, может обладать более низким или более высоким сродством, предполагается, что CDR 3В9 в таких условиях все-таки будут распознавать тот же эгритоп, что и 3В9. Примерами тяжелоцепных CDR 3В9 могут служить последовательности ID N 22; последовательность ID N 24; последовательность ID N 26; а примерами легкоцепных CDR 3В9 могут служить последовательность ID N 16; последовательность ID N 18 и последовательность ID N 20.
Термин "функциональный фрагмент" относится к неполной вариабельной последовательности тяжелой или легкой цепи (например, небольшие делеции на амино- или карбоксиконцах иммуноглобулинового вариабельного участка), сохраняющей ту же антигенсвязующую специфичность и/или нейтрализующую способность, что и антитело, из которого был получен данный фрагмент.
Термин "аналог" относится к аминокислотной последовательности, модифицированной, по меньшей мере, одной аминокислотой, причем указанная модификация может быть осуществлена химически, представлять собой замещение или перегруппировку нескольких аминокислот (как правило не более 10) и такая модификация позволяет аминокислотной последовательности сохранять биологические характеристики, например специфичность к антигену, высокий титр или аффинность, присущие немодифицированной последовательности. Так, например, молчащие мутации могут реализоваться путем замещений с целью создания рестрикционных эндонуклеазных сайтов внутри или в пределах окружающих CDR области.
Аналоги могут также возникать в ходе аллельных вариаций. Под термином "аллельная вариация или модификация" подразумевается изменение нуклеиново-кислотной последовательности, кодирующей аминокислотную или пептидную последовательности изобретения. Такие вариации или модификации могут быть связаны с дегенерацией генетического кода или достигнуты в результате обдуманного конструирования с целью придания желаемых характеристик. Такие вариации или модификации могут в результате приводить или не приводить к изменению в любой кодирующей аминокислотной последовательности. Так, например, аминокислотные последовательности CDR легкой цепи, последовательность ID N 16 идентична нативной мышиной последовательности и гуманизированного 3В9 антитела. Однако такая CDR последовательность закодирована как SEQ ID N: 15, так и SEQ ID N: 53. Аналогичным образом, CDR SEQ ID N: 22 закодирована как SEQ ID N: 21, так и SEQ ID N:54, CDR SEQ ID N: 24 закодирована как SEQ ID N: 23, так и SEQ ID N: 55; и CDR SEQ ID N: 26 закодирована как SEQ ID N: 25, так и SEQ ID N:56.
Термин "эффекторные агенты" относится к непротеиновым молекулам-носителям, с которыми обычными методами могут быть связаны слитые протеины и/или природная или синтетическая, легкая или тяжелая цепь донорского антитела или других фрагментов донорского антитела. Такие небелковые носители могут включать традиционные носители, используемые в области диагностики, например полистирол или другие пластиковые шарики, полисахариды, например те, что используются в системе BlAcope /Pharmacia / или другие небелковые вещества, используемые в медицине и безопасные при применении на людях и животных. Другие агенты-эффекторы могут включать макроцикл для хелатирования атома тяжелого металла или радиоизотопы. Такие эффекторы могут также использоваться для повышения времени полураспада слитых протеинов; к таким агентам относится полиэтиленгликоль.
II. Высокоаффинные IL 4-моноклональные антитела.
Для осуществления конструирования антител, фрагментов и слитых протеинов изобретения могут применяться нечеловеческие разновидности (например, материалы, полученные от коров, овец, приматов, грызунов (например, мышей и крыс) и т.д.) и они используются для генерации желаемого иммуноглобулина путем представления с помощью нативного человеческого IL 4 или его пептидного эпитопа. Традиционные гибридомные методы используются для обеспечения гибридомной клеточной линии, секретирующей нечеловеческий Mab к IL 4. Затем такие гибридомы подвергают скринингу с использованием IL 4, ковалентно связанного с 96-луночными пластинами, или с использованием биотинилированного IL 4, предназначенного для применения в скрининге в соответствии с методикой, подробно описанной в следующем ниже примере 2. Таким образом, одним из отличительных признаков настоящего изобретения является способ детекции Mab на человеческий IL 4, в котором используемые аналитические средства позволяют избежать денатурацию IL 4. Согласно такому способу было обнаружено, что могут детектироваться Mab с высоким титром (или высоким сродством) к человеческому IL 4.
В качестве одного из примеров вначале будет описано получение нейтрализующего Mab с высоким титром из мышиного донора. Mab 3В9, представляющие собой желательное мышиное (донорское) антитело для использования в выработке химерного или гуманизированного антитела, подробно описано в следующем ниже примере 1. 3В9 Mab характеризуется антигенсвязующей специфичностью на человеческий IL 4 с Kd < 2 · 10-1 М (около 1,8 · 10-10 М) в отношении IL 4. Kd для 114 фрагмента Fab такого 3В9 имеет значение менее 3·10-10 М. Эпитоп такого антитела не может картироваться с помощью IL 4 линейных пептидов и, следовательно, такой эпитоп рассматривается как связывающийся с несоприкасающимся эпитопом. Картина связывания предполагает наличие связующего сайта в области B-C петля (остатки 60-69) ---> C-спираль (остатки 70-93). В отношении картированного обозначения таких участков можно сослаться на работы Cook с сотр. J.Mol. Biol. 218:675-678 (1991), Walter с сотр., J.Biol. Chem. 267: 20371-20376 (1992), Wlodaver с сотр., FEBS Lett. 309: 59-64(1992), Redfield с сотр. Biochem 30: 11029-11035 (1991), Smith с сотр., J.Mol. Biol. 224: 899-904(1992), Garrett с сотр., (1992) и Rowers c coтр. Biochem. 31: 4334-4346 (1992) и Science 256:1673-1677 (1992).
Другим желаемым донорским антителом является крысиный Mab, 6A1. Получение такого Mab описано ниже в примере 7. Такой Mab характеризуется тем, что является изотипом IgG2a и имеет константу диссоции в отношении IL 4 менее 2 · 10-10 М (около 1,6·10-10 М). Как и в случае 3В9 эпитоп-мишень такого 6A1 не картируется с IL 4 линейными пептидами и поэтому такой эпитоп рассматривается как несоприкасающийся и трехмерный. Картина связывания с IL 4 мутеинами и его биологическая активность указывают на связывание в области D-спирали человеческого IL 4 (аминокислотные остатки 109-127), по-видимому в области тирозина на аминокислотном остатке # 124.
Настоящее изобретение не ограничивается использованием 3В9 Mab, 6A1 Mab или их гипервариабельных (т.е. CDR) последовательностей. Любые другие подходящие IL 4 антитела с высоким титром, характеризующиеся константой диссоциации, равной или меньшей 2 · 10-10 М в отношении человеческого IL 4, и соответствующие анти-IL 4 CDR могут служить заменой указанным материалам. В следующем ниже описании донорское антитело идентифицируется как 3В9 или 6A1, причем такое обозначение дается лишь в целях иллюстрации и упрощения описания.
Ill. Фрагменты антител
Настоящее изобретение также охватывает использованием Fab фрагментов или F(ab)2 фрагментов, являющихся производными Mab, направленных против человеческого IL 4. Такие фрагменты используются в качестве агентов, защищающих in vivo от IL 4 - IgE-медиаторных состояний или in vitro в качестве части IL 4 диагностического средства. Фрагмент Fab содержит полную легкую цепь и аминотерминальную часть тяжелой цепи; а фрагмент F(ab')2 представляет собой фрагмент, образованный двумя фрагментами Fab, связанными посредством дисульфидной связи. Mab 3В9, 6A1 и другие аналогичные высокоаффинные, IL 4 связывающие антитела представляют собой источники фрагментов Fab и F(ab'), которые могут быть получены традиционными методами, например расщеплением Mab с помощью соответствующих протеолитических энзимов, папаина и/или пепсина, или рекомбинантными методами. Такие фрагменты Fab и F(ab')2 сами по себе используются в качестве терапевтических, профилактических или диагностических агентов, а также в качестве доноров последовательностей, включающих вариабельные участки и CDR последовательностей, используемых для образования рекомбинантных или гуманизированных антител, как это описано в настоящем документе.
IV. Анти-114 аминокислотная и нуклеотидные последовательности.
Mab 3В9 или другие описанные выше антитела могут обеспечивать последовательности, например вариабельные пептидные последовательности тяжелой и легкой цепей, рамочные последовательности, CDR последовательности, их функциональные фрагменты и аналоги, а также иодирующие их нуклеиново-кислотные последовательности, применяемые для конструирования и получения различных слитых протеинов (включая генно-инженерные антитела), которые характеризуются антигенсвязующей специфичностью донорского антитела.
Таким образом, в качестве одного из примеров настоящее изобретение предусматривает последовательности вариабельной легкой цепи и вариабельной тяжелой цепи из IL 4 мышиного антитела 3В9 и последовательности, являющиеся их производными. Вариабельный участок тяжелой цепи 3В9 характеризуется аминокислотными остатками 20-140 SEQ ID N: 4. CDR области указаны путем подчеркивания на фиг. 2 и представлены в SEQ ID N: 22; SEQ ID N: 24 и SEQ ID N: 26. Вариабельная область клона легкой цепи характеризуется аминокислотными остатками 21-132 на фиг. 1 /SEQ ID N: 2/. CDR области даны аминокислотными остатками 44-58 /SEQ ID N: 16/; 74-80/ SEQ ID N: 18/ и 113-121 / SEQ ID N: 20/.
Также предусмотрены вариабельная область химерной тяжелой цепи и сигнальная нуклеотидная и аминокислотная последовательности. Эти последовательности идентичны тяжелой цепи 3В9 за исключением сигнальной последовательности. Сигнальная последовательность химерной тяжелой цепи показана в SEQ ID N: 5 и 6. Области CDR подчеркнуты на фиг. 3 и они идентичны по аминокислотной последовательности нативным мышиным CDR /SEQ ID N: 21-26/. Нуклеотидные и аминокислотные последовательности вариабельного района химерной легкой цепи идентичны немодифицированным 3В9 последовательностям (аминокислотные остатки 21-132 SEQ ID N: 2), что делает возможным использование природных мышиных сигнальных последовательностей (аминокислотные остатки 1-20 SEQ ID N: 2).
Вариабельная область гуманизированной тяжелой цепи и сигнальные последовательности проиллюстрированы на фиг. 4 /SEQ ID N: 11 и 12/. Сигнальная последовательность также показана в SEQ ID N: 5 и 6. Другие подходящие сигнальные последовательности, известные специалистам в данной области, могут служить заменой сигнальным последовательностям, представленным в примерах. CDR аминокислотные последовательности такой конструкции идентичны CDR нативной мышиной и химерной тяжелой цепи и они показаны в SEQ ID N: 22 (закодированной SEQ ID N: 54), SEQ ID N: 24 (закодированной SEQ ID N: 55) и SEQ ID N: 56 (закодированной SEQ ID N: 26).
Приведенная в качестве примера (синтетическая) вариабельная последовательность гуманизированной легкой цепи проиллюстрирована на фиг. 5 /SEQ ID N: 13 и 14/. Такая сигнальная последовательность включает аминокислотные остатки 1-19 последовательности ID N: 8. CDR последовательности на этом чертеже обозначены путем подчеркивания и они отличаются от CDR нативной мышиной CDR по сигнальной аминокислотной последовательности SEQ ID N: 20. Таким образом, CDR гуманизированной легкой цепи представлены SEQ ID N: 53 и 16, SEQ ID N: 17 и 18 и SEQ ID N 27 и 28. Имеющееся отличие подробно описано в примере 3.
Нуклеиново-кислотные последовательности настоящего изобретения или их фрагменты, кодирующие пептидные последовательности вариабельной легкой цепи и тяжелой цепи, используются в немодифицированной форме или могут быть синтезированы с целью введения желаемых модификаций, например рестрикционных сайтов. Выделенные нуклеиново-кислотные последовательности природного или синтетического происхождения, являющиеся производными Mab 3В9 или других желательных IL 4 антител с высоким титром, могут необязательно содержать рестрикционные сайты, облегчающие инсерцию или лигирование в подходящую нуклеиново-кислотную последовательность, например ту, что кодирует каркасную область желаемого антитела, лигирование с мутантными CDR или слияние с нуклеиново-кислотной последовательностью, кодирующей выбранный второй партнер слияния.
Принимая во внимание вырожденности генетического кода, могут быть сконструированы различные кодирующие последовательности, которые способны кодировать вариабельные аминокислотные последовательности тяжелой и легкой цепи, а также CDR последовательности изобретения и их функциональные фрагменты и аналоги, которые делят антигенную специфичность донорского антитела. Выделенные нуклеиново-кислотные последовательности изобретения или их фрагменты, кодирующие пептидные последовательности вариабельной цепи или CDR, могут использоваться для получения слитых протеинов, химерных или гуманизированных антител или других инженерных антител настоящего изобретения при оперативном соединении со вторым партнером слияния.
Эти последовательности также используются для мутагенного введения специфических изменений в нуклеиново-кислотные последовательности, кодирующие CDR или каркасные области, а также для введения полученной в результате модифицированной или слитой нуклеиново-кислотной последовательности в плазмиду для экспрессии. Так, например, молчащие замещения в нуклеотидной последовательности каркасной и CDR-кодирующей областей использовали для создания рестрикционных сайтов, которые облегчают вставку мутагенизированных CDR (и/или каркасных) областей. Такие CDR области использовали для конструирования гуманизированного антитела изобретения.
Следует отметить, что помимо выделенных нуклеиново-кислотных последовательностей, кодирующих части слитого белка и антител, описанных в настоящем документе, могут использоваться и другие нуклеиново-кислотные последовательности, например, комплементарные нативным последовательностям. Используемые ДНК последовательности включают такие последовательности, которые гибридизуются в строгих условиях /см. T.Maniatis с сотр. Молекулярное клонирование (Лабораторное руководство), лаборатория Колд Спринг Харбор (1982), стр. 387-389/ с ДНК-последовательностями. Одним из примеров такой гибридизации в строгих условиях является гибридизация при 4XSSC при 65oC с последующей промывкой в 0,1 XSSC при 65oC в течение часа. Другим примером гибридизации в строгих условиях может служить обработка в 50% формамиде, 4 XSSC при 42oC. Предпочтительно, чтобы такие гибридные ДНК-последовательности содержали, по крайней мере, 18 нуклеотидов, т.е. имели размер, близкий к размеру CDR.
V. Слитые молекулы и слитые протеины
Слитые молекулы способны кодировать слитые белки, которые включают такие инженерные антитела, как химерные антитела и гуманизированные антитела. Желательная слитая молекула содержит CDR, последовательности, кодирующие пептиды, обладающие антигенной специфичностью IL 4 антитела, предпочтительно высокоаффинного антитела, предусматриваемого настоящим изобретением, вставленного в первый партнер слияния (человеческая каркасная область или человеческая иммуноглобулиновая вариабельная область).
Предпочтительно, чтобы первый партнер слияния был оперативно связан со вторым партнером слияния. Второй партнер слияния определен выше и он может включать последовательность, кодирующую область второго антитела, например Fc область. Вторые партнеры слияния могут также включать последовательности, кодирующие другие иммуноглобулины, с которыми, с сохранением рамки считывания, слита константная область легкой или тяжелой цепи, или такое слияние осуществляет с помощью линкерной последовательности. Инженерные антитела, направленные против функциональных фрагментов или аналогов IL 4, могут быть сконструированы таким образом, чтобы обеспечивать усиленное связывание с тем же антителом.
Второй партнер слияния может быть также связан с описанным выше агентом-аффектором, включающим небелковые молекулы-носители, с которыми второй партнер слияния может быть оперативно связан традиционными средствами.
Слияние или сцепление между вторыми партнерами слияния, например последовательностями антитела, и агентом-аффектором может осуществляться любыми подходящими методами, например путем создания традиционных ковалентных или ионных связей, слияния протеинов или с использованием таких гетеро-бифункциональных сшивателей, как карбодиимид, глутаровый альдегид и т.п. Такие методики известны в данной области и легко доступны из традиционной химической и биохимической литературы. Кроме этого, традиционные линкерные последовательности, которые просто обеспечивают желаемое пространство между вторым партнером слияния и агентом-аффектором, также могут быть сконструированы в слитой молекуле. Конструкция таких линкеров хорошо известна специалистам в данной области.
Следует отметить, что сигнальные последовательности молекул изобретения могут быть модифицированы с целью усиления экспрессии. В качестве одного из примеров можно отметить слитый белок, имеющий аминокислотную последовательность последовательности мышиной тяжелой цепи, которая идентична химерной вариабельной тяжелой цепи (Yн) фиг. 2 / SEQ ID N: 4/, содержит оригинальный сигнальный пептид, замененный другой сигнальной последовательностью (аминокислотные остатки 1-20) / SEQ ID N: 6/.
Пример слитого белка содержит пептидную или протеиновую последовательность вариабельной тяжелой и/или легкой цепи, обладающую антигенной специфичностью Mab 3В9, например Yн /аминокислотные остатки 21-141 последовательности SEQ ID N: 9 и 10/ и YL цепи /аминокислотные остатки 21-132 последовательностей SEQ ID N: 1 и 2/. Еще один желательный слитый белок изобретения характеризуется аминокислотной последовательностью, содержащей по крайней мере одну, а предпочтительно все CDR вариабельной области тяжелых и/или легких цепей мышиного антитела 3В9, причем оставшиеся последовательности имеют человеческое происхождение, либо их аналоги или функциональные фрагменты. См. , например, гуманизированные YH и YL области SEQ ID N: 11 и 12 и SEQ ID N: 13 и 14 (фиг. 4 и 5).
Согласно еще одному воплощению изобретения инженерное антитело может содержать присоединенный к нему дополнительный агент. Так, например, метод рекомбинантной ДНК технологии может быть использован для получения инженерного антитела изобретения, в котором Fc фрагмент CH3 домена молекулы всего антитела заменен на энзим или другую детектируемую молекулу (например, полипептидный эффектор или молекулу-репортер).
Второй партнер слияния может быть также оперативно связан с неиммуноглобулиновым пептидом, протеином или их фрагментом, гетерологичным CDR-содержащей последовательности, имеющей антигенную специфичность мышиного 3В9. Полученный в результате белок может в ходе экспрессии проявлять как анти-IL 4 антигенную специфичность, так и характеристики неиммунноглобулина. Характеристики такого партнера слияния могут включать такие функциональные характеристики, как другой связующий или рецепторный домен, или терапевтические характеристики, если партнер слияния сам по себе является терапевтическим белком, или дополнительные антигенные характеристики.
Другой желательный белок настоящего изобретения может содержать молекулу полного антитела, имеющую легкие и тяжелые цепи полной длины, или любой ее дискретный фрагмент, например Fab или F(ab')2, димер тяжелой цепи, или любые их минимальные рекомбинантные фрагменты, например Fy или одноцепочечное антитело (SCA), либо любую другую молекулу с той же специфичностью, что выбранное донорское Mab, например Mab 3В9 или 6A1. Такой протеин может использоваться в виде слитого белка или может применяться в неслитой форме.
В каждом случае, когда второй партнер слияния происходит из другого антитела, например любого изотипа или класса иммуноглобулиновой каркасной или константной области, возникает генно-инженерное антитело. Инженерные антитела могут содержать иммуноглобулиновые (Ig) константные области и вариабельные каркасные области из одного источника, например акцепторного антитела, и одну или более (предпочтительно все) CDR донорского антитела, например, описанное в тексте анти-IL 4 антитело. Кроме этого, могут осуществляться изменения, например делеции, замещения или вставки вариабельного домена каркасной области тяжелой, и/или легкой, и/или тяжелоцепной вариабельной доменной остовной области акцепторной цепи акцепторного Mab на нуклеиново-кислотном или аминокислотном уровнях или участков донорского CDR с тем, чтобы сохранить антигенную связующую специфичность донорского антитела.
Такие инженерные антитела конструируются таким образом, чтобы использовать одну (или обе) вариабельные тяжелые и/или легкие цепи IL 4 Mab (необязательно модифицированные, как это описано) или одну или более нижеидентифицированные CDR тяжелой или легкой цепи (см. пример 3). Инженерные антитела настоящего изобретения являются нейтрализующими антителами, т.е. они могут блокировать связывание с рецептором IL 4 протеина. Так, например, антитело, происходящее из Mab 3В9, направлено против специфического третичного белкового эпитопа человеческого IL 4, находящегося, как полагают, в области B-C петля ----> C-спираль, как это было отмечено выше.
Такие инженерные антитела могут включать гуманизированное антитело, содержащее каркасные участки выбранного человеческого иммуноглобулина или субтипа, или химерное антитело, содержащее человеческие константные области тяжелой или легкой цепи, слитые с функциональными фрагментами IL 4 антитела. Подходящее человеческое (или другого животного происхождения) ацепторное антитело может быть выбрано из традиционной базы данных, например базы данных КАВАТ®, Лос-Аламоской базы данных и Швейцарской белковой базы данных, по гомологии с нуклеотидными и аминокислотными последовательностями донорского антитела. Человеческое антитело, характеризующееся гомологией с каркасными областями донорского антитела (на аминокислотном базисе), может быть пригодным для обеспечения константной области тяжелой цепи и/или вариабельной каркасной области тяжелой цепи для вставки донорских CDR. Подходящее акцепторное антитело, способное быть донором константных или вариабельных каркасных областей легкой цепи, может быть выбрано аналогичным образом. Следует отметить, что тяжелые и легкие цепи акцепторного антитела необязательно должны иметь происхождение от одного и того же акцепторного антитела.
Желательно, чтобы гетерологичные каркасные и константные области выбирались из человеческих иммуноглобулиновых классов и изотипов, таких как IgG (подтипы 1-4), IgM, IgA и IgE. Однако акцепторное антитело необязательно должно содержать лишь человеческие иммуноглобулиновые белковые последовательности. Так, например, может быть сконструирован ген, в котором ДНК последовательность, кодирующая часть цепи человеческого иммуноглобулина, слита с ДНК-последовательностью, кодирующей неиммуноглобулиновую аминокислотную последовательность, например полипептидную эффекторную или репортерскую молекулу.
Один из примеров особенно желательного гуманизированного антитела содержит CDR, 3В9, вставленные в каркасные области последовательности выбранного человеческого антитела. В случае нейтрализующих гуманизированных антител один, два или предпочтительно три CDR из вариабельных областей тяжелой и/или легкой цепи IL 4 антитела вставляются в каркасные области последовательности, выбранного человеческого антитела, заменяя нативные CDR последнего антитела.
Предпочтительно в гуманизированном антителе вариабельные домены как тяжелых, так и легких цепей человека подвергаются генно-инженерным операциям путем замены одного или более CDR. Можно использовать все шесть CDR или различные комбинации из менее чем шести CDR. Предпочтительно заменяют все шесть CDR. Можно заменять CDR только в человеческой тяжелой цепи, используя в качестве легкой цепи немодифицированную легкую цепь человеческого акцепторного антитела. С другой стороны, совместимая легкая цепь может выбираться из другого человеческого антитела, прибегая к помощи традиционной базы данных антител. Оставшаяся часть инженерного антитела может быть производным любого подходящего акцепторного человеческого иммуноглобулина.
Таким образом, генно-инженерное гуманизированное антитело предпочтительно имеет структуру природного человеческого антитела или его фрагмента и обладает комбинацией свойств, требуемых для эффективного терапевтического использования, например, в лечении связанных с IL 4 воспалительными заболеваниями людей или для диагностического применения.
В качестве другого примера можно привести инженерное антитело, которое содержит три CDR вариабельной области легкой цепи 3В9 /SEQ ID 1: 16, 18, 20 и 28/ и три CDR, вариабельной области тяжелой цепи 3В9 /SEQ ID N 22, 24 и 26/.
Полученное в результате гуманизированное антитело характеризуется антигенсвязующей специфичностью и высокой аффинностью Mab 3В9.
Специалистам в данной области должно быть понятно, что инженерное антитело может подвергаться дополнительным модификациям в результате изменений аминокислот вариабельных доменов, причем это необязательно должно оказывать влияние на специфичность и высокую аффинность донорского антитела (т.е. аналога). Так, например, были сконструированы гуманизированные моноклональные антитела, в которых аминокислотный остаток в положении 120 легкой цепи представлял собой аргинин / SEQ ID N: 13 и 14/ или треонин /SEQ ID N: 57 и 58/. Следует отметить, что аминокислоты тяжелой и легкой цепей могут быть замещены другими аминокислотами либо в вариабельном домене каркасов, либо в CDR или в обеих указанных областях.
Кроме этого, константная область может подвергаться изменению, направленному на усиление или снижение селективных свойств молекул настоящего изобретения. В качестве примеров таких изменений можно привести димеризацию, связывание с Fc рецепторами, или изменение способности связывать и активировать комплемент (см. , например, Angal с сотр. Mol. Immunol. 30: 105-108(1993), Xu с сотр., J. Biol. Chem. 299: 3469-3474(1994), Winter с сотр., EP 307,434-B).
Слитый протеин, представляющий собой химерное антитело, отличается от описанных выше гуманизированных антител тем, что имеет вариабельные области тяжелой и легкой цепей полностью нечеловеческого донорского антитела, включает каркасные области, ассоциированные с человеческими иммуноглобулиновыми константными областями обеих цепей. Следует отметить, что химерные антитела, сохраняющие дополнительные нечеловеческие последовательности относительно гуманизированных антител изобретения, могут вызывать значительную иммунную реакцию у людей.
Как обсуждается ниже, такие антитела полезны в профилактике и лечении связанных с действием IL 4 аллергических расстройств.
VI. Получение слитых протеинов и инженерных антител.
Предпочтительно вариабельные последовательности легкой и/или тяжелой цепей и CDR Mab 3В9 /SEQ ID N: 16, 18, 20, 22, 24 и 26/ или других подходящих донорских Mab (например, 6A1) и их кодирующие нуклеиново-кислотные последовательности используются для конструирования слитых белков и инженерных антител, предпочтительно гуманизированных антител изобретения, с использованием следующего способа. Такой же или похожие методы могут использоваться для реализации других технических решений настоящего изобретения.
Гибридому, продуцирующую выбранное донорское Mab, например мышиное антитело 3В9, подвергают общепринятому клонированию и ДНК ее вариабельных участков тяжелой и легкой цепей получают методами, известными специалистам в данной области, например методами, описанными Sambrook с сотр. Молекулярное клонирование (Лабораторное руководство), 2-е изд., Лаборатория Колд Спринг. Харбор (1989). Вариабельные области тяжелой и легкой цепей 3В9, содержащие, по крайней мере, CDR и те части вариабельного домена каркасной области легкой и/или тяжелой цепей акцепторного Mab, которые требуются для сохранения связующей специфичности в отношении донорского Mab, а также оставшиеся иммуноглобулиновые части цепи антитела, происходящей из человеческого иммуноглобулина, получают с использованием полинуклеотидных праймеров и обратной транскриптазы. CDR идентифицируют с использованием известной базы данных и сравнением с другими антителами.
Затем может быть получено мышь/человек химерное антитело и проведен его анализ на способность к связыванию. Такое химерное антитело содержит YH и Y1 области полностью нечеловеческого донорского антитела, связанные с константными участками человеческого Ig для обеих цепей.
Гомологичные каркасные области вариабельного участка тяжелой цепи человеческого антитела идентифицировали с использованием компьютеризированной базы данных, например КАВАТ®, и человеческое антитело, обладающее гомологией к 3В9, выбирали в качестве акцепторного антитела. Последовательности синтетических вариабельных участков тяжелой цепи, содержащих 3В9 CDR внутри каркасов человеческого антитела, конструировали с необязательными заменами нуклеотидов в каркасных областях с целью введения рестрикционных сайтов. Такую сконструированную последовательность затем синтезировали с помощью перекрывающихся олигонуклеотидов, амплифицировали с помощью полимеразной цепной реакции (PCR) и исправляли ошибки.
Аналогичным образом была сконструирована подходящая вариабельная каркасная область легкой цепи.
Гуманизированное антитело может происходить из химерного антитела или предпочтительно может быть получено синтетически, вставкой CDR донорского Mab из тяжелых и легких цепей в выбранный каркас тяжелой и легкой цепи. С другой стороны гуманизированное антитело изобретения может быть получено с использованием стандартных приемов мутагенеза. Таким образом, полученное в результате гуманизированное антитело содержит человеческие каркасные области и CDR донорского Mab. Могут быть осуществлены последующие манипуляции с остатками каркаса. Полученное гуманизированное антитело может быть экспрессировано в рекомбинантных клетках-хозяевах, например клетках COS или CHO. Другие подробности такого метода представлены в примере 4. Другие гуманизированные антитела могут быть получены с использованием такого метода на других подходящих IL 4-специфичных, нейтрализующих, нечеловеческих антителах с высоким титром.
Традиционный вектор экспрессии или рекомбинантную плазмиду получают, проводя оперативную ассоциацию таких кодирующих последовательностей для слитого белка с традиционными регуляторными контрольными последовательностями, способными контролировать репликацию и экспрессию в, и/или секрецию из, клетки-хозяина. Регуляторные последовательности включают промоторные последовательности, например CMV промотор и сигнальные последовательности, которые происходят из других известных антител. Аналогичным образом получают второй вектор экспрессии, содержащий ДНК последовательность, кодирующую легкую или тяжелую цепь комплементарного антитела. Предпочтительно такой второй вектор экспрессии идентичен первому, за исключением тех случаев, когда кодирующие последовательности и способные к селекции маркеры предназначаются для максимально возможной функциональной экспрессии каждой полипептидной цепи.
Выбранную клетку-хозяина подвергают котрансфекции общепринятыми методами как с первым, так и со вторым векторами или просто трансфицируют одним вектором с целью получения трансфицированной клетки-хозяина настоящего изобретения, содержащей как рекомбинантную, так и синтетическую легкую и тяжелую цепи. Трансфицированную клетку затем культивируют общепринятыми методами с получением инженерного антитела изобретения. Гуманизированное антитело, включающее ассоциацию рекомбинантной тяжелой цепи и/или легкой цепи, тестируют из культуры с помощью соответствующих анализов, таких как ELISA или RIA. Аналогичные традиционные приемы могут использоваться для конструирования других слитых белков и молекул настоящего изобретения.
Подходящие векторы для клонирования и субклонирования, а также конструкции композиций настоящего изобретения могут быть выбраны специалистами в данной области техники. Так, например, могут применяться общепринятые pYC серии клонирующих векторов. Один из используемых векторов представляет собой pYC19 и он выпускается такими поставщиками, как Amersham (Букингхамшир, Великобритания) или Pharmacia (Уппсала, Швеция). Кроме этого, любой вектор, способный легко реплицироваться, содержит множество клонирующих сайтов и маркерных генов и в целях клонирования может легко осуществляться любая манипуляция. Таким образом, выбор клонирующего вектора не является ограничивающим фактором настоящего изобретения.
Аналогичным образом векторы, используемые для экспрессии инженерных антител согласно изобретению, могут быть выбраны специалистом в данной области из любого традиционного вектора. Такие векторы также содержат выбранные регуляторные последовательности, находящиеся в оперативной ассоциации с ДНК-кодирующими последовательностями иммуноглобулиновых областей и способны направлять репликацию и экспрессию гетерологических ДНК-последовательностей в выбранных клетках-хозяевах, например CMV-промоторы. Такие векторы содержат описанные выше ДНК-последовательности, которые кодируют инженерное антитело слитой молекулы. С другой стороны, такие векторы могут содержать выбранные иммуноглобулиновые последовательности, модифицированные вставкой желаемых рестрикционных сайтов в целях облегчения манипуляции.
Векторы экспрессии могут также характеризоваться маркерными генами, подходящими для усиления экспрессии гетерологичных ДНК-последовательностей, например дигидрофолатредуктазный ген млекопитающего (DHFP) или ген неомицинустойчивости (neo®). Другие предпочтительные векторные последовательности включают поли-A-сигнальную последовательность, например бычьего гормона роста (BGH), и бета-глобинпромоторную последовательность (betaglopro). Используемые векторы экспрессии могут быть синтезированы методами, хорошо известными специалистам в данной области.
Компоненты таких векторов, например репликоны, гены селекции, энхансеры, промоторы, сигнальные последовательности и т.п., могут быть получены из природных источников или синтезированы известными методами с целью их использования для направления экспрессии и/или секреции продукта рекомбинантной ДНК в выбранном хозяине. Другие подходящие векторы экспрессии, многочисленные типы которых известны в данной области для экспрессии у млекопитающих, бактерий, насекомых, дрожжей и грибков, также могут быть выбраны для использования в указанных выше целях.
Настоящее изобретение также охватывает клеточную линию, трансфицированную рекомбинантной плазмидой, содержащей кодирующие последовательности инженерных антител или их слитых молекул. Клетки-хозяева, используемые для клонирования и других манипуляций с такими клонирующими векторами, также являются традиционными. Однако наиболее желательно использовать клетки различных штаммов E.coli для репликации клонирующих векторов и других стадий конструирования слитых белков настоящего изобретения.
Подходящими клетками-хозяевами или клеточными линиями для экспрессии инженерных антител или слитых белков изобретения предпочтительно являются такие эукариотные клетки, как CHO, COS, фибробластная клетка (например, 3Т3) и миелоидные клетки, причем наиболее предпочтительной является такая клетка млекопитающего, как CHO-клетка или миелоидная клетка. Могут использоваться человеческие клетки, что позволяет модифицировать молекулу по профилям человеческого гликозилирования. Могут использоваться и другие эукариотные клеточные линии. Выбор подходящих клеток-хозяев от млекопитающих, методов их трансформации, культивирования, амплификации, скрининга, а также получения продукта и его очистки известны из литературы. См., например, цитированную выше работу Sambrook с сотр.
Бактериальные клетки могут оказаться полезными в качестве клеток-хозяев, подходящих для экспрессии рекомбинантых Mab настоящего изобретения. Однако, в связи с тенденцией белков к экспрессии в бактериальных клетках в неуложенной или неправильно уложенной форме или в негликозилированной форме, любые рекомбинантные Mab, полученные в бактериальной клетке, должны быть подвергнуты скринингу на сохранение способности связывать антиген. Если молекула, экспрессированная бактериальной клеткой, получена в правильно уложенной форме, то такая бактериальная клетка может считаться желательным хозяином. Так, например, различные штаммы E.coli, используемые для экспрессии, как хорошо известно, могут применяться в качестве клеток-хозяев в области биотехнологии. В этом методе могут также использоваться различные штаммы B.subtilis, Streptomyces, другие бактерии и т.п.
Как известно специалистам, штаммы дрожжевых клеток также могут использоваться в качестве клеток-хозяев, как и клетки насекомых, например Drasophila и Lepidoptera, а также вирусные экспрессирующие системы. См., например, Miller с сотр. Генная инженерия, 8:277-298, Пленум Пресс (1986) и цитированную в этой работе литературу.
Основные методы конструирования векторов настоящего изобретения, методы трансфекции, требующиеся для получения клеток-хозяев изобретения, и методы культивирования, необходимые для получения слитого белка или инженерного антитела настоящего изобретения из такой клетки-хозяина, являются хорошо известными приемами. Аналогичным образом после получения слитые белки или инженерные антитела изобретения могут быть очищены от компонентов клеточной культуры с помощью стандартных методов, включающих осаждение сульфатом аммония, очистку на аффинных колонках, хроматографических колонках, гель-электрофорез и т.п. Эти методы известны специалистам и не ограничивают сферу изобретения.
Еще один способ экспрессии гуманизированных антител может состоять в экспрессии в трансгенном животном в соответствии с описанным в патенте США N 4873316. Этот метод относится к экспрессирующей системе, использующей казеиновый промотор животного, который при трансгенном введении в млекопитающее позволяет женской особи продуцировать желаемый рекомбинантный белок в ее молоке.
После экспрессии желательным методом инженерное антитело исследуют на in vitro активность с использованием соответствующего анализа. В настоящее время общепринятые схемы анализа ELISA используются для оценки качественного и количественного связывания инженерного антитела с IL 4-эпитопом. Кроме этого, другие in vitro анализы, например Blacore/Pharmacia/, могут также использоваться для проверки нейтрализующей эффективности перед последующим клиническом исследованием на людях, осуществляемом для оценки персистем инженерного антитела в организме человека вне зависимости от обычных механизмов очистки.
Следуя методам, описанным для получения гуманизированных антител из 3В9, специалист в данной области может также сконструировать гуманизированные антитела из других донорских IL 4-антител, последовательностей вариабельных областей и CDR пептидов, описанных в настоящей заявке. Инженерные антитела могут быть получены с такими вариабельными каркасными участками, которые потенциально распознаются как "свои" реципиентами инженерного антитела. В вариабельной каркасной области могут быть произведены небольшие модификации с целью осуществления сильного увеличения антигенного связывания без ощутимого увеличения иммуногенности для реципиента. Такие инженерные антитела могут эффективно использоваться для лечения людей при болезненных состояниях, связанных с IL 4. Эти антитела могут также использоваться для диагностики указанных состояний.
VII. Терапевтическое и профилактическое применения.
Настоящее изобретение также относится к способу лечения людей, испытывающих аллергические нарушения, который заключается в применении эффективной дозы антител, включающей одно или более инженерных антител или слитых белков, описанных в настоящей заявке, или их фрагментов.
Терапевтическая реакция, вызываемая применением молекул настоящего изобретения, возникает вследствие связывания с человеческим IL 4 и последующим блокированием выделения IgE. Таким образом, молекулы настоящего изобретения в виде препаратов и рецептур для терапевтического использования весьма желательны для применения на пациентах с аллергическими нарушениями, такими как аллергический ринит, коньюктивит, атопический дерматит, атопическая астма и анафилактический шок.
Слитые белки, антитела, инженерные антитела или их фрагменты настоящего изобретения могут также использоваться совместно с другими антителами, особенно человеческими Mab, реактивными в отношении других маркеров (эпитопов), ответственных за болезненное состояние, против которого направлено действие инженерных антител изобретения. Аналогичным образом Mab, реакционноспособные в отношении эпитопов, ответственных за болезненное состояние выбранного животного, против которого направлено действие антитела изобретения, также могут использоваться в ветеринарных композициях.
Предполагается, что терапевтические агенты настоящего изобретения окажутся полезными для лечения аллергических состояний в течение периода времени от 2 дней до 3 недель или по необходимости. Так, например, более длительные времена лечения могут оказаться желательными при лечении сезонных ринитов и т. п. Такой метод представляет собой значительный шаг вперед по сравнению с используемым в настоящее время методом вливания в соответствии с известными приемами лечения нарушений, вызванных действием IL 4. Дозировка и длительность лечения зависят от относительной длительности пребывания молекул изобретения в организме человека и они могут регулироваться специалистом в зависимости от типа заболевания и общего состояния здоровья пациента.
Тип применения терапевтического агента изобретения может быть любым, обеспечивающим доставку такого агента в организм хозяина. Слитые белки, антитела, инженерные антитела и их фрагменты, а также фармацевтические композиции изобретения особенно полезны при парентеральном применении, т.е. подкожно, внутримышечно, внутривенно или интраназально.
Терапевтические агенты настоящего изобретения могут быть приготовлены в виде фармацевтических композиций, содержащих эффективное количество инженерного (например, гуманизированного) антитела настоящего изобретения в качестве активного ингредиента в фармацевтически применимом носителе. При профилактическом применении агента изобретения предпочитают использовать водную суспензию или раствор, содержащий инженерное антитело, предпочтительно в среде буфера при физиологическом значении pH, в форме, готовой для инъекции. Композиции для парентерального применения обычно содержат раствор инженерного антитела изобретения или его смесь, растворенную в фармацевтически применимом носителе, предпочтительно в водном носителе. Может использоваться большое число водных носителей, например 0,4% солевой раствор, 0,31% глициновый раствор и т.п. Такие растворы должны быть стерильными и, как правило, они не содержат мелких частиц вещества. Эти растворы могут быть простерилизованы хорошо известными методами (например, фильтрацией). Такие композиции могут содержать фармацевтически применимые вспомогательные соединения, требующиеся для аппроксимации физиологических условий, и ими могут быть регуляторы pH, буферные агенты и т.п. Концентрация антитела изобретения в такой фармацевтической рецептуре может изменяться в широких пределах, например от значения менее 0,5%, обычно, по крайней мере, от 1% до 15-20% вес., и требуемую концентрацию выбирают, основываясь на требуемом объеме жидкости, вязкости и т.п., в соответствии с конкретным типом применения.
Так, например, фармацевтическую композицию настоящего изобретения для внутримышечного вливания готовят таким образом, чтобы она содержала 1 мл стерильной забуференной воды и 1 нг - 100 мг, как правило, 50 нг - 30 мг или более предпочтительно 5-25 мг инженерного антитела изобретения. Аналогичным образом фармацевтическая композиция изобретения для внутривенного вливания должна содержать 250 мл стерильного раствора Рингера и 1- 30 мг, предпочтительно 5-25 мг инженерного антитела настоящего изобретения. Методы приготовления парентерально применяемых композиций хорошо известны специалистам и более детально они описаны, например, в Remington's Pharmaceutical Science, 15-е изд. Мак Паблишинг Компани, Истон, Пенсильвания.
Предпочтительно, чтобы терапевтический агент изобретения присутствовал в фармацевтическом препарате в виде единичной дозы. Соответствующая терапевтически эффективная доза может быть легко определена специалистом в данной области. Для эффективного лечения воспалительного расстройства у людей и животных следует применять парентерально, предпочтительно внутримышечно, единичную дозу, включающую примерно 0,1-20 мг на 70 кг веса тела белка или антитела настоящего изобретения. Такая доза, если необходимо, может быть повторена через соответствующий период времени в ходе воспалительной реакции в соответствии с предписаниями терапевта.
Настоящее изобретение также охватывает введение IL 4 слитых белков настоящего изобретения одновременно или последовательно с другими антителами или слитыми белками, характеризующимися анти-IL 4 активностью, например фактором активности противоопухолевого некроза или другими фармацевтическими активностями, совместимыми с IL 4 рецепторной связующей способностью слитых белков изобретения. Такие другие антитела выпускаются промышленностью или могут быть сконструированы в соответствии со способом, аналогичным описанному в настоящей заявке.
Слитые белки и инженерные антитела настоящего изобретения могут также использоваться в диагностических целях, например для определения расстройств, связанных с действием IL 4, или слежения за ходом лечения таких нарушений. В качестве диагностических реагентов такие слитые белки могут быть традиционным образом помечены для использования в анализе ELISA и других общепринятых анализах, предназначенных для измерения уровней IL 4 в сыворотке, плазме или других соответствующих тканях. Такие анализы, где используются слитые белки, являются общепринятыми и не ограничивают сферу изобретения.
Антитела, инженерные антитела или их фрагменты, описанные в данной заявке, могут быть лиофилизированы для хранения и вновь составлены в подходящем носителе перед использованием. Было показано, что такой прием эффективен для обычных иммуноглобулинов и известные методики лиофилизации и реконституции могут быть использованы.
Следующие ниже примеры иллюстрируют различные аспекты настоящего изобретения, включая конструирование представителей инженерных антител и их экспрессию в подходящих векторах и клетках-хозяевах, и такие примеры не ограничивают сферу изобретения. Все аминокислоты обозначены традиционными трехбуквенным или одно- буквенным кодами. Все необходимые рестрикционные энзимы, плазмиды и другие реагенты и материалы получали из коммерческих источников, если это не оговорено особо. Все общие методы клонирующего лигирования и другие методы рекомбинантной ДНК-технологии осуществляли в соответствии с описанным в работе T. Maniatis с сотр., цитированной выше, или вторым изданием этой книги (1989), ред. Sombrook с сотр., тех же издателей (Sambrook с сотр.).
Пример 1.-Получение Mab 3В9
A. Методика иммунизации.
Четырех мышей (F1 гибриды BaIb/c и C57B1/6) подкожно иммунизировали 50 мкг рекомбинантного E.coli человеческого IL 4 в полном адьюванте Фрейнда и через 4 недели проводили повторную внутри брюшинную иммунизацию 50 мкг IL 4 в неполном адьюванте Фрейнда. Основываясь на хорошем значении титра сывороточного антитела по отношению к IL 4, одну из мышей дополнительно иммунизировали через 8 недель 200 мкг IL 4 (внутрибрюшинно, физиологический раствор), через два дня - 100 мкг IL 4 (внутрибрюшинно, в физиологическом растворе) и спустя два дня 50 мкг IL 4 (внутрибрюшинно, в физиологическом растворе). Через два дня после финальной иммунизации проводили спленектомию.
B. Методика слияния и система скрининга.
Клетки мышиной селезенки использовали для получения гибридом (по стандартной методике, например, описанной Kohler с сотр., Nature 256; 495(1975), из которых > 250 клонов клеток подвергали скринингу на секрецию антитела к IL 4, с использованием выпускаемой промышленностью системы B1Acore и анализа ELISA, как это описано ниже, в целях связывания IL 4. В пяти лунках был получен положительный ответ. Лишь 1 клон мышей, 3В9, был сильно положительным. Все вторичные клоны, являющиеся производными 3В9 были положительными.
Пример 2. Анализы ELISA и константы аффинности.
A.ELISA
Скрининговый анализ, осуществляемый в соответствии с описанным ниже, предназначался для измерения аффинности на нативный человеческий IL 4. В эксперименте 1 активированные альдегидом пластины с 96 лунками покрывали IL 4 в количестве 1 мкг/мл, 100 мкл/лунку в 0,1М боратном буфере, pH 8,5 и инкубировали в течение ночи при комнатной температуре. К пластине ковалентно присоединяли hIL 4. Раствор IL 4 удаляли и неспецифически связанные (NSB) сайты блокировали 1% бычьим сывороточным альбумином (BSA) в TBS буфере (50 мМ Трис, 150 мМ NaCl, 1мМ MgCl2, 0,02% NaN3, pH 7,4) в течение 60 минут при 37oC. После этой и каждой из последующих стадий пластину промывали 4 раза промывным буфером (10 мМ Трис, 150 мМ NaCl, 0,05% Твин 20, 0,02% NaN3, pH 7,4). После этого добавляли 50 мкл гибридомной среды (очищенный 3В9 или Fab фрагменты) и 50 мкл аналитического буфера (0,5% бычьего гамма-глобулина в TBS буфере) и пластины инкубировали в течение 60 минут при 37oC. 100 мкл биотинилированного антимышиного антитела добавляли в каждую лунку в среде аналитического буфера и проводили инкубацию, как описано выше. 100 мкл стрептавидина, коньюгированного со щелочной фосфатазой, добавляли в лунку и проводили инкубацию (30 мин при 37oC). Добавляли 100 мкл/лунку PNP и проводили инкубацию в течение 30 мин при 37oC. Показания снимали по оптической плотности при длине волны 405 нм. В эксперименте 2 покрытые стрептавидином пластины (100 мкл/лунку, 1 мкг/мл в фосфатном буферном растворе (PBS)) инкубировали в течение ночи при 4oC и анализировали, как описано ниже. Стрептавидиновый раствор удаляли, NSB сайты блокировали 1% BSA в NBS буфере (60 мин при 37oC). После этой стадии и каждой из последующих стадий пластины четыре раза промывали промывным буфером 50 мкл биотинилированного IL 4, добавляли совместно с 50 мкл аналитического буфера и систему инкубировали в течение 30 минут при 37oC. После этого добавляли 50 мкл очищенного 3В9 IgG или Fab фрагмент (или гибридомную среду) плюс 50 мкл аналитического буфера и проводили инкубацию в течение 60 минут при 37oC. Добавляли 100 мкл антимышиного IgG-щелочно-фосфатазного коньюгата и проводили инкубацию в течение 60 минут при 37oC. Добавляли 100 мкл PNP субстрата и проводили инкубацию в течение 30 минут при 37oC. Отсчет показаний проводили, как описано выше.
B. Расчет 3В9 аффинности к IL 4.
Используя результаты описанных выше экспериментов и суммируя их, как описано ниже, рассчитывали Kd для 3В9 в соответствии с методом Beadty с сотр. J. Immunol. Methods, 100 : 173-179 (1987): Kaff =1/2(2[Abx]-[Ab],
Abx = концентрация связанного Ab при 150 нг/мл биотинилированного hIL 4;
Ab = концентрация связанного Ab при 300 нг/мл биотинилированного hIL 4.
Константы диссоциации рассчитывали из следующего соотношения: Kd = 1/Kaff.
Эксперимент 1: анализ ELISA на пластине с 96 лунками, покрытой стрептавидином (100 нг/лунку). Kd = 2,2 · 10-10 М (3В9 Fab).
Эксперимент 2: анализ ELISA на пластине с 96 лунками, покрытой стрептавидином (100 нг/лунку). Kd = 1,4 · 10-10 М (3В9 IgG).
C. Специфичность.
Mab 3В9 распознает человеческий IL 4, но не распознает бычий или мышиный IL 4. Один из путей определения такого явления состоит в следующем. Анализ ELISA может быть осуществлен с использованием пластины с 96 лунками, покрытой антимышиным IgG и затем заблокированной бычьим сывороточным альбумином, на которой 50 мкл 3В9 (100 нг/мл), 25 мкл нечеловеческого IL 4 и 25 мкл биотин-IL 4 инкубировали в течение 60 мин при 37oC, после чего проводили промывку стрептавидином, коньюгированным с щелочной фосфатазой и PNP.
Аналогичным образом было установлено, что Mab 6A1 не распознает бычий или мышиный IL 4.
Пример 3. Гуманизированное антитело
Одно гуманизированное антитело конструировали таким образом, чтобы оно содержало мышиные CDR внутри каркаса человеческого антитела. Такую гуманизированную версию IL 4 специфичного мышиного антитела 3В9 готовили путем осуществления следующих манипуляций.
A. кДНК-клонирование.
Клоны кДНК получали из мРНК 3В9 тяжелых и легких цепей, экстрагированной из 3В9 гибридомной клеточной линии /пример 1/, с использованием набора Boehringer Mannheim. Праймеры, специфичные как на мышиный шарнирный участок, так и на каппа-константную область, использовали для синтеза первой нити.
Праймер каппа-цепи представляет собой /SEQ ID 1:29/:
5' - CTAACACTCATTCCTCTTCAACCTCTTCACAATCCC-3'
Праймер тяжелой гамма-цепи представляет собой /SEQ ID N: 30/:
5' - CTACATATCCAACCCTTACAACCACAATC3'.
Двухнитевую кДНК клонировали непосредственно в плазмиде pGEM7f +/Promega/, которыми затем трансформировали Е.coli ДН-5а /Bethesda Research Labs/.
B. Секвенирование ДНК.
кДНК клонов восьми мышиных тяжелых и одной легкой цепи мышиных из части A подвергали секвенированию. Результаты секвенирования вариабельных областей таких клонов показаны в SEQ ID N: 1, 2, 3 и 4. Каждый клон содержал аминокислоты, известные как консервативные среди вариабельных районов мышиных тяжелых цепей или легких цепей, и мышиные сигнальные последовательности. Аминокислотные последовательности CDR перечислены ниже.
CDR-области тяжелой цепи представляют собой SEQ ID N: 22, 24 и 26 (аминокислоты 50-56, 71-86 и 119-129 последовательности SEQ ID N: 4) См. фиг. 2. Такие последовательности закодированы последовательностями SEQ ID N: 21, SEQ ID N: 23 и SEQ ID N: 25 соответственно. CDR области легкой цепи представляют собой SEQ ID N: 16, 18 и 20 (аминокислоты 45-58, 74-80 и 113-121 последовательности SEQ ID N: 2. См. фиг. 1. Такие последовательности закодированы последовательностями SEQ ID N: 15, 17 и 19 соответственно.
C. Отбор человеческих каркасов.
После клонирования 3В9 аминокислотные последовательности вариабельной области (аминокислоты 21-132 SEQ ID N: 2 и аминокислоты 20-140 SEQ ID N: 4) сравнивали с базой данных последовательностей человеческого иммуноглобулина с использованием баз данных КАВАТ® и SWiSS с целью идентификации человеческого каркаса как для тяжелой, так и для легкой цепи, которые наиболее близко совместимы с мышиным источником по гомологии последовательности. Помимо этих исследований по гомологии последовательности легкие и тяжелые цепи также оценивали относительно позиционной базы данных, разработанной из структурных моделей Fab домена с целью определения потенциальных конфликтов, связанных с заменами аминокислот, которые могут оказывать влияние на CDR представление. В настоящем случае структурными исследованиями не было обнаружено очевидных конфликтов; поэтому использовали ДНК-кодирование, выведенное на основании исследований гомологии аминокислотной последовательности.
Использовали каркасные области тяжелой цепи антитела, полученного из человеческого миеломного иммуноглобулина (COR) /E.M.Press, N.M.Hogg Biochem. J. 117: 641-660 (1970)). Такая последовательность оказалась примерно на 77% гомологичной (69,4% идентичность) 3В9 вариабельной области цепи на аминокислотном уровне.
Для подходящей вариабельной каркасной области легкой цепи использовали вариабельную каркасную последовательность легкой цепи человеческого антитела, идентифицированную H. G.Klobeck с coтp., Nucl.Acid Res. 13:6515-6529 (1985). Было установлено, что последовательность человеческого антитела примерно на 80,2% гомологична (идентичность 72,0%) мышиной вариабельной области легкой цепи на аминокислотном уровне.
Используя мышиные 3В9 CDR /SEQ ID N: 15-26/ и последовательность человеческого антитела, получали синтетическую тяжелую цепь и осуществляли PCR для амплификации ДНК. Такие последовательности синтезировали с помощью следующих перекрывающихся олигонуклеотидов и амплифицировали с помощью PCR. SEQ ID N: 31-37 предусматривает пять перекрывающихся олиго и 2 PCR праймера. Олиго 1 /SEQ ID N: 31/, как установлено, содержит основания 5-121. Олиго 2 /SEQ ID N: 32/ содержит основания в диапазоне 122-241, а олиго 3 /SEQ ID N: 33/ содержит основания 242-361. Два нижних нитевых праймера SEQ ID N: 34 и SEQ ID N: 35 содержат основания 134-110, и основания 134-110, и основания 253-230. Все ошибки в картированной последовательности, которые были внесены PCR, были исправлены. PCR осуществляли снова с использованием в качестве 5' праймера нуклеотиды 1-25 SEQ ID N: 36, а в качестве 3' - праймера - нуклеотиды 361-341 последовательности SEQ ID N: 37.
Синтетический вариабельный участок лигировали в экспрессирующий вектор pCD совместно с синтетической сигнальной последовательностью SEQ ID N; 5 и 6 из химерной конструкции тяжелой цепи совместно с IgG1 человеческой константной областью. Синтетическая YH и нуклеотидная и аминокислотная последовательности сигнальной последовательности представлены на фиг.4 /SEQ ID N: 11 и 12/. Аминокислотные последовательности CDR /SEQ ID N: 22, 24 и 26/ идентичны мышиным 3В9 BDR. Однако, кодирующие последовательности для CDR /SEQ ID N: 54, 55 и 56/ отличаются от мышиных 3В9 кодирующих последовательностей /SEQ ID N: 21, 23 и 25/. Полученный в результате вектор экспрессии, 114hzhc 1-1-Pcd, показан на фиг. 9.
CDR генные области пресуществующего каркаса легкой цепи удаляли рестрикционным перевариванием и заменяли на следующие синтетические IL 4 CDR гены, которые получали синтетическим путем.
Для CDR, 1:
SEO IDNO: 38: 5'CTAGCTGTGTCTCTGGGCGAGAGGGCCACCATCAACTGCAAGG 3'
SEQ IDNO: 39: CCTTGCAGTTGATGGTGGCCCTCTGGCCCAGAGACACAG
SEQ IDNO: 40: TCGAGAGGCCTCCCAAAGTGTTGATTATGATGGTGATAG
TTATATGAACTGGTATCAGCAGAAACCC
SEQ IDNO: 41: GGGTTTCTGCTGATACCAGTTCATATAACTATCACCATCATA
ATCAACACTTTGGGAGGCCTC
для CDR2:
SEQ IDNO: 44:
GGGCAGCCTCCTAAGTTGCTCATTTACGCTCGATGCAATCTA
GAATCTGGGGTAC
SEQ 1DNO: 45;
CCCAGATTCTAGATTGGATGCAGCGTAAATGAGCAACTTAGG
AGGCTGCCC
Для CDR3:
SEQ 1DNО: 42:
ATACTACTGTCAGCAAAGTAATGAGGATGGTCCGAGGTTCGG
CGGAGGGAC
SEQ 1DNO: 43:
CTTGGTCCCTCCGCCGAACCTCGGAGGATCCTCATTACTTTG
CTGACAGTAGT
Синтетическая YL и нуклеотидная и аминокислотные последовательности сигнальной последовательности показаны на фиг. 5 /SEQ ID N: 13 и 14/. Аминокислотные последовательности двух первых CDR /SEQ ID N: 16 и 18/ идентичны соответствующим мышиным 3В9 CDR. Однако кодирующая последовательность для первого CDR /SEQ ID N: 53/ отличается от мышиной 3В9 кодирующей последовательности /SEQ ID N: 15/. Кроме этого, в последнем CDR были созданы две гуманизированные конструкции 3В9 аминокислотной последовательности. Одна из них /SEQ ID N: 28/ отличается по сигнальной аминокислотной /SEQ ID N: 20/ от нативной мышиной 3В9 последовательности. SEQ ID N: 28 закодирована последовательностью SEQ ID N: 27. Синтетические вариабельные легкие области лигировали в вектор экспрессии совместно с сигнальной последовательностью /SEQ ID N: 7 и 8/. Один из полученных векторов экспрессии, 114hzic1-О-Pcn, показан на фиг. 10.
Такие синтетические вариабельные последовательности легкой и/или тяжелой цепи используются при конструировании гуманизированного антитела.
Пример 4. Экспрессия гуманизированного Mab в клетках COS и CHO.
pUC18-субклоны для YH получали таким образом, чтобы добавить сигнальную последовательность, первоначально полученную от человеческого антитела SEQ ID N: 5. Для YL получали субклоны pUC18 с целью добавления сигнальной последовательности SEQ ID N: 7.
Гуманизированная тяжелая цепь, являющаяся производным IgG1 изотипа, проявляет 89: 3% гомологию (идентичность 84,3%) на аминокислотном уровне к мышиной тяжелой цепи от 3В9. Синтетическая YH обеспечена аминокислотами 20-141 последовательности SEQ ID N: 11 и 12. Гуманизированная легкая цепь, человеческая каппа-цепь, демонстрирует 92,0% гомологию (идентичность 86,6%) с 3В9 на аминокислотном уровне. Синтетическая YL /аминокислоты 21-131 последовательности SEQ ID N: 13 и 14/, содержащая 3В9 CDR, была сконструирована и синтезирована в соответствии с описанным для синтетических тяжелых цепей.
Фрагменты ДНК, соответствующие сигналы которых связаны с вариабельными участками как гуманизированной тяжелой, так и легкой цепи, вставляли pUC19 - основанные экспрессирующие плазмиды клеток млекопитающих, содержащие CMY промоторы и константные области человеческой тяжелой или легкой цепи химеры, полученной в примере 5, используя для этого традиционные методы /Maniatis, цитированная выше/ с получением плазмид 114 hzhc1 - 1Pcd (тяжелая цепь) /фиг. 9/ и 114 hz1c1 - oPcn) (легкая цепь) /фиг. 10/. Плазмиды HZHC и HZLC котрансфицировали в клетки COS и супернатанты анализировали методом ELISA на присутствие гуманизированного антитела через 3 и 5 дней. Другое гуманизированное антитело конструировали с использованием IgG4 изотипа.
В представленном примере описывается получение инженерного антитела. Другие методики могут использоваться для создания других инженерных антител с использованием других анти-IL 4 антител (например, 6A1 - пример 7), полученных традиционными методами.
Пример 5. Конструирование химерного антитела
A. Химерную тяжелую цепь конструировали путем выделения мышиного вариабельного участка тяжелой цепи из оригинального мышиного Mab 3В9 в виде EcoR1-BstE11 рестрикционного фрагмента. Конструировали и синтезировали небольшой олигомер ДНК для соединения мышиного вариабельного участка с человеческой IgG1 константной областью (Bst EII - Apa1):
5' праймер: SEQ ID N: 50: GTCACCGTCTCCTCAGCTAGCACCAAGGGGG
3' праймер: SEQ ID N: 51: CTTGGTGCTAGCTGAGGAGACG.
Два таких фрагмента лигировали в плазмиду pCD (см. фиг. 7) (расщепленную с EcoR1 и Apa1), которая уже кодирует человеческий IgG1 константный участок. Такой клон не экспрессировался, поэтому 5' UTP дикого типа и сигнальную последовательность делетировали и заменяли на SEQ ID N: 5 и 6.
Поскольку традиционный рестрикционный эндонуклеазный сайт не доступен на 3' окончании сигнальной последовательности, вводили Bst EII сайт (т.е. молчащая мутация) с помощью PCR. Использовали следующие PCR праймеры;
SEQ IDNO: 48: 5' праймер: 5'CACCTTACCCTCAAACACTC 3'
SEQ IDNO: 49: 3' праймер: 5'GAAGTAGTCCTTGACCAG 3'
Затем из такой плазмиды выделяли Bst EII-PstI рестрикционный фрагмент. Затем конструировали и синтезировали новую сигнальную последовательность и 5'ИТР, содержащие EcoRI и Bst ЕII1 концы.
SEQ IDNO: 46: 5 праймер: AATTCGAGGACGCCAGCAACATGGTGTTGCA GACCCAGGTCTTCATTTCTCTGTTGCTCTGGAATCTCTGGTGCCTACGGGC
AG
SEQ IDNO: 47: 3' праймер: GTAACCTGCCCGTAGGCACGAGAGATCCAGA
GCAACAGAGAAATGAAGACCTGGGTCTGCAACACCATGTTGCTCGCGTC
CTCG
Химерную легкую цепь конструировали с применением техники PCR к исходной мышиной 3В9 легкой цепи, которую клонировали в pGEM72f (+) /Progema/. Используемые праймеры представляли собой выпускаемые промышленностью p UC18 универсальный обратный праймер на 5' конце (EcoR1) и 3' праймер, вводящий сайт Nar1:
[5'CATCTAGATGGCGCCGCCACAGTACGTTTGATCTCCAGCTTGGTCCC3'
используемый для слияния мышиного вариабельного участка с человеческим константным участком. Затем такую вариабельную область лигировали в экспрессионный вектор PcDN (EcoR1 Nar1) (фиг. 8), который уже содержал человеческую каппа-область.
Через 3 и 5 дней собирали супернатанты и проводили анализ методом ELISA, описанным ниже: пластины ELISA покрывали 0,1 мкг козьего антитела, специфичного на Fc область человеческих антител. Супернатанты среды добавляли в течение часа. Добавляли пероксидазу хрена, коньюгированную с козьим антителом, специфичным на полное человеческое IgG антитело. После этого в течение часа добавляли ABTS пероксидазный субстрат (Киркегаард и Перри Лабораторис Инк. , Гайтерсбург, МД). Детектировали экспрессию химерного антитела. Во втором анализе ELISA COS клеточные супернанты, содержащие химерное антитело, специфически связывались с рекомбинантным человеческим IL 4 протеином. Этот результат подтверждает тот факт, что гены, кодирующие антитело, специфичное на IL 4 подвергались клонированию.
B. Гуманизированная тяжелая цепь может быть также получена из такой химерной тяжелой цепи. Гуманизированную тяжелую цепь конструировали вставкой мышиных CDR в человеческий каркас. Использовали тот же человеческий каркас, что описан выше, наиболее гомологичная протеиновая последовательность в Швейцарской базе данных протеинов на мышиную 3В9 YH (аминокислоты 20-140 последовательности SEQ ID N: 4). Такую гуманизированную последовательность тяжелой цепи (EcoRI ApaI) получали синтетическим путем и осуществляли PCR с целью амплификации ДНК в соответствии с описанным выше. Синтетическую вариабельную область лигировали в экспрессирующий вектор pCD (EcoR1 Apa1) совместно с синтетической сигнальной последовательностью SEQ ID N: 5 и 6 из химерной конструкции тяжелой цепи и IgG человеческого константного участка.
Аналогичным образом гуманизированная легкая цепь может быть получена из химерной легкой цепи в соответствии с описанным выше для тяжелой цепи. Этот ген (ЕсоRY Nar1) получали также синтетическим путем. Гуманизированный YL лигировали в экспрессирующий вектор pCN, переваренный в присутствии EcoR1 Nar1, совместно с сигнальной последовательностью (EcoR1 EcoRY). Вектор экспрессии содержал человеческую каппа константную область.
Пример 6. Очистка и термодинамика гуманизированного Mab.
Очистка CHO экспрессированного химерного и гуманизированного 3В9 может достигаться традиционными методами белковой A (или G) аффинной хроматографии с последующим применением ионообменной хроматографии и гель-фильтрации. Аналогичные методы с успехом использовались для очистки до чистоты > 95% других Mab (например, респираторного синцитиального вируса и малярийных циркумспорозоитных антигенов).
Аффинность и подробная термодинамика IL 4 связывания с гуманизированным Mab 3В9 и мышиным 3В9 (пример 1) определяли методом микрокалориметрического титрования. В этом методе измеряются реакции связывания по их внутренним теплотам реакции (см., например, Wiseman с сотр. Anal. Biochem. 179: 131-137 (1989). Аффинность обоих Mab оказалась настолько прочной, что не было возможности провести измерения при окружающей температуре. Тогда был применен термодинамический подход: I) аффинность измеряли при 60oC, когда она достаточно слаба и поддается прямому измерению; и II) снимали температурную зависимость энтальпии связывания при 30-60oC. Совместно полученные результаты позволили рассчитать аффинность в широком температурном интервале с использованием уравнения Гиббса-Гельмгольца.
Результаты термодинамики IL 4 связывания для гуманизированного и мышиного 3В9 антител представлены в таблице 1. На
основании различий в свободной энергии, энтальпии, энтропии и теплоемкости двух таких Mab можно сделать вывод о неразличимости их термодинамического поведения.
Пример 7. Получение характеристики крысиного Mab.
Mab 6A1, выбранное для высокоаффинного связывания, получали от иммунизированной крысы с использованием протокола иммунизации, описанного для мышей в примере 1. 6A1 выбирали из гибридом (конкретно, гибридома 3426A11C1В9), полученных от крыс, иммунизированных человеческим IL 4.
Kd для 6A1 рассчитывали в соответствии с описанным Beaty с сотр. J.Immunol. Methods. 100: 173-179(1987) и полученное значение было равно 2 · 10-10 М.
Гибридома 3426A11C1B9 депонирована 6 октября 1993 г. в Европейской коллекции культур клеток животных (ECACC), Лаборатория здравоохранения сервисного центра по прикладной микробиологии и исследованиям, Нортон Даун, Салисбури, Вильтшайр, SP4 O1G, Великобритания, под каталожным номером 93100620 и акцептирована как патентный депонент в соответствии с Будапештским договором 1977 г., регулирующим депонирование микроорганизмов в целях патентования.
Пример 8. Биологическая активность Mabs: 3В9 (гуманизированного) 3В9 (мышиного) и 6A1.
Следующие ниже анализы проводили с использованием описанных ниже методик.
A. Связывание с гликозилированным rhIL 4.
Указанные выше антитела индуцировали к негликозилированному рекомбинантному человеческому IL 4 (rh IL 4), которое продуцировали в E.coli. Поскольку нативный человеческий IL 4 является гликозилированным, важно подтвердить связыванием с материалом, секретированным клеточной линией млекопитающих. 3В9 в одинаковой степени хорошо присоединяется как к гликозилированному, так и к негликозилированному человеческому рекомбинантному IL 4 и поэтому на направлен к эпитопу, который может быть замаскирован на природном человеческом IL 4.
B. Ингибирование IL 4 связывания с рецептором.
Способность 3В9 ингибировать связывание IL 4 с его рецептором изучалась с использованием 125I -rhIL 4 связывания с клеточной линией гибона, M1a /АТСС T1B201/, которая несет примерно 6000 рецепторов в расчете на клетку. М1A клетки инкубировали с 125I-IL 4 в течение 30 минут при 37oC. Поглощение радиоактивности определяли в гамма-счетчике после отделения клеточно-связанного 125I-IL 4 центрифугированием через масляный градиент. Hеспецифическое связывание определяли инкубацией в присутствии 100-кратного молярного избытка немеченного IL 4/Park с сотр., J. Exp. Med. 166: 476-488 (1987)/. Значение IC50 для немеченного IL 4 в таком анализе составило 22пМ при количестве добавленного IL 4 83пМ. Для интактного мышиного (IgG) 3В9 значение IC50 составило 63пМ и 93пМ для Fab фрагмента. При другой концентрации IL 4 (218пМ) полученное в анализе количество для мышиного (IgG) 3В9 составило 109 пМ.
C. Ингибирование пролиферации лимфоцитов.
С использованием метода, описанного Spits с сотр., J. Immunol. 139: 1142-1147 (1987), человеческие периферические лимфоциты крови инкубировали в течение трех дней в присутствии фитогемаглютинина, T-клеточного митогена, с целью активации IL 4 рецептора. Полученные в результате бластоидные клетки затем стимулировали в течение трех дней с помощью IL 4. Пролиферацию измеряли внедрением 3H тимидина. Пролиферацию B-клеток измеряли методом анализа согласно Collard с сотр., Лимфокинез и интерфероны. Практический подход, гл. 19, стр. 345, при следующих модификациях. Очищенные B-клетки человеческих миндалин стимулировали в течение 3 дней с помощью IL 4 и иммобилизованного анти-IgM. Пролиферацию измеряли внедрением 3H тимидина.
Мышиный 3В9 ингибировал внедрение 3H-тимидина в человеческие перефирические T-лимфоциты крови, стимулированные 133пМ IL 4, и в B-лимфоциты человеческих миндалин, стимулированные 167пМ IL 4. IL 2-стимулированные T-лимфоциты не подвергались воздействию. Значение IC50 для ингибирования пролиферации T-клеток составило 30пМ, а для пролиферации B-клеток - 103пМ. Соответствующие значения для Fab фрагмента мышиного 3В9 составили 108 и 393 пМ.
D. Ингибирование CD23 индукции.
CD23 представляет собой низко-аффинный рецептор IgE (FcER11) и индуцируется на мембране B-лимфоцитов низкими концентрациями IL 4, что является необходимой предпосылкой для IgE продуцирования. Очищенные B-клетки человеческих миндалин стимулировали в течение 2 дней с помощью IL 4. Процентное количество клеток, экспрессирующих CD23 рецептор, определяли методом проточной цитометрии /Defrance с сотр., J. Exp. Med. 165:1459-1467 (1987)/. Мышиный 3В9 ингибировал CD23 экспрессию на B-лимфоцитах человеческих миндалин, стимулированных 8.3пМ IL 4 при значении IC50 136пМ.
E. Ингибирование IgE секреции.
В отличие от других анализов, в которых IL 4 добавляли при EC50 концентрациях /Pere с сотр., Proc. Natl. Acad, Sci. 85: 6880-6884(1988), IgE секрецию исследовали в присутствии концентраций IL 4, обеспечивающих максимальную секрецию с тем, чтобы уменьшить вариабельность, присущую такой системе. Пролиферацию T-клеток измеряли следующим образом. Человеческие лимфоциты периферической крови инкубировали с IL 4 в течение 10-18, предпочтительно 12 дней. Концентрацию IgE в супернатанте культуры определяли методом ELISA.
IgE-секреция ингибировалась мышиным 3В9 и Fab фрагментом 3В9 в присутствии 1,7 нМ IL 4, давая значение IC50 1,9 и 5,0 нМ соответственно. Эксперимент повторяли с использованием более низкой концентрации IL 4, 667 пМ, что понижало значение IC50 до 0,65 нМ в случае мышиного 3В9. В изученных концентрационных пределах молярное соотношение антитела (IgC) к IL 4 оставалось неизменным (1:1).
F. Резюме и интерпретация данных.
Молярные соотношения IL 4 к различным Mab, требуемые для 50% ингибирования функции в биоанализах, представлены в таблице 2.
Во всех анализах, за исключением IgE секреции, IL 4 добавляли с примерно ЕД50 концентрацией. Молярные соотношения между антителом и IL 4, требуемые для 50% ингибирования, близки для гуманизированного 3В9, мышиного 3В9 и 6A1 в двух анализах по пролиферации лимфоцитов, но выше для гуманизированного 3В9 в CD23 индукционном анализе. Последний анализ является особенно чувствительным, требующим очень низких (~5%) степеней занятости рецептора (Kruse с сотр., EMBO J, 12:5121, 1993) и, как видно из результатов, полученных при использовании мышиного 3В9 и в связи с этим подвержен аналитическим вариациям.
Сравнение активностей крысиного 6A1 и мышиного 3В9 демонстрирует аналогичную картину функциональных эффектов, а также неожиданную неспособность 6A1 полностью ингибировать связывание радиоиодированного IL 4 с его рецептором. Предполагается, что радиоиодированный IL 4, используемый в рецепторно-связующем анализе, иодирован по доступному тирозину, остатку 124. При сравнении способности 6A1 ингибировать CD23 экспрессию, индуцированную немеченным или иодированным IL 4, было обнаружено, что ингибирование менее эффективно в случае иодированного лиганда. Эти результаты показывают, что 6A1 связывается с IL 4 в области тирозина 124, но без специфичности к этому остатку.
Таким образом, полученные данные показывают, что 6A1 представляет собой нейтрализующее антитело высокой аффинности, способное связываться с совершенно другим участком IL 4, чем 3В9.
Пример 9. Фармакокинетика
Фармакокинетические свойства гуманизированного 3В9 изучали на самцах крыс разновидности Sprague Dawley, гуманизированное 3В9 применяли на четырех животных в виде iv шаровидной массы с дозировкой 1 мг/кг, причем отбор образцов крови продолжали в течение 5 недель после применения. Концентрацию гуманизированного 3В9 в плазме определяли с использованием IL 4/античеловеческой IgG сэндвичевой EILSA, предназначенного подтвердить не только наличие циркулирующего человеческого IgG, но также его способность связываться с рекомбинантным человеческим IL 4.
Полученные результаты суммированы в таблице 3.
Полученные данные показывают, что вариабельность между животными относительно мала и исчезновение гуманизированного 3В9 из плазмы, по-видимому, является двухфазным. Кажущееся очищение плазмы было низким (0,5 мл/час/кг). Период полураспада составлял 11 дней. Таким образом, фармакокинетические характеристики гуманизированного 3В9, являющегося производным CHO клеток, согласуется с другими гуманизированными моноклональными антителами у крыс. Длительный период циркуляционного полувыведения гуманизированного 3В9 у крыс также позволяет предположить, что при применении на людях гуманазированное 3В9, по-видимому, будет эффективным в течение длительного периода времени.
Многочисленные модификации и вариации настоящего изобретения охватываются приведенным выше описанием и они должны быть очевидны для специалистов в данной области. Так, например, человеческие каркасные области или их модификации, отличные от приведенных выше примеров антител, могут использоваться для конструирования гуманизированных антител. Такие модификации и изменения композиций и способов настоящего изобретения охватываются сферой прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ МИЕЛИН-АССОЦИИРОВАННОГО ГЛИКОПРОТЕИНА (MAG) | 2003 |
|
RU2303461C9 |
| АНТИТЕЛА ПРОТИВ CXCR4 ДЛЯ ЛЕЧЕНИЯ ИНФЕКЦИИ ВИЧ | 2010 |
|
RU2573902C2 |
| ИММУНОГЛОБУЛИНЫ | 2005 |
|
RU2404192C2 |
| НОВЫЕ АНТИ-IL 13 АНТИТЕЛА И ИХ ИСПОЛЬЗОВАНИЕ | 2004 |
|
RU2487887C2 |
| АНТИТЕЛО ПРОТИВ c-Met | 2009 |
|
RU2560257C2 |
| БЛОКИРУЮЩИЕ АНТИТЕЛА ПРОТИВ Dkk-1 И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2548817C2 |
| АНТИТЕЛА ПРОТИВ IL2 | 2006 |
|
RU2425054C2 |
| НОВОЕ АНТИТЕЛО ПРОТИВ с-МЕТ | 2011 |
|
RU2607377C2 |
| ИММУНОГЛОБУЛИНЫ K CD52 ЧЕЛОВЕКА | 2010 |
|
RU2603743C2 |
| АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ ИНТЕРЛЕЙКИНУ-2 И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2745451C1 |








































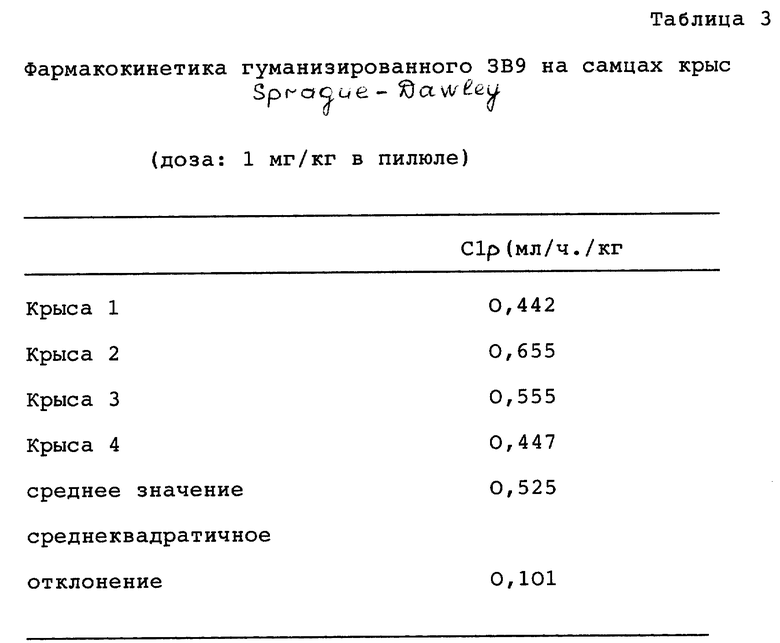







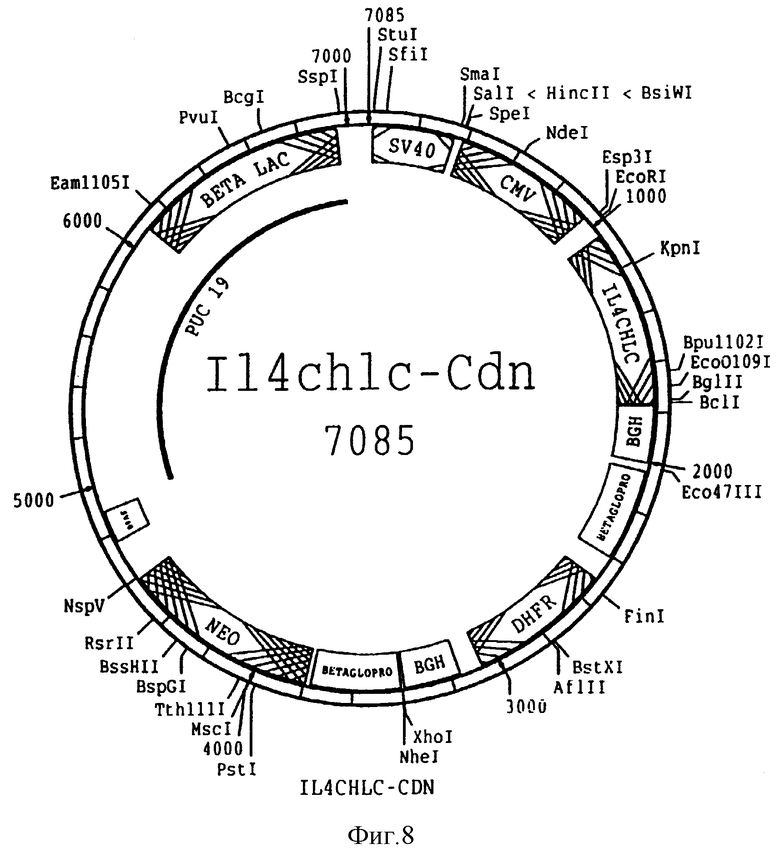
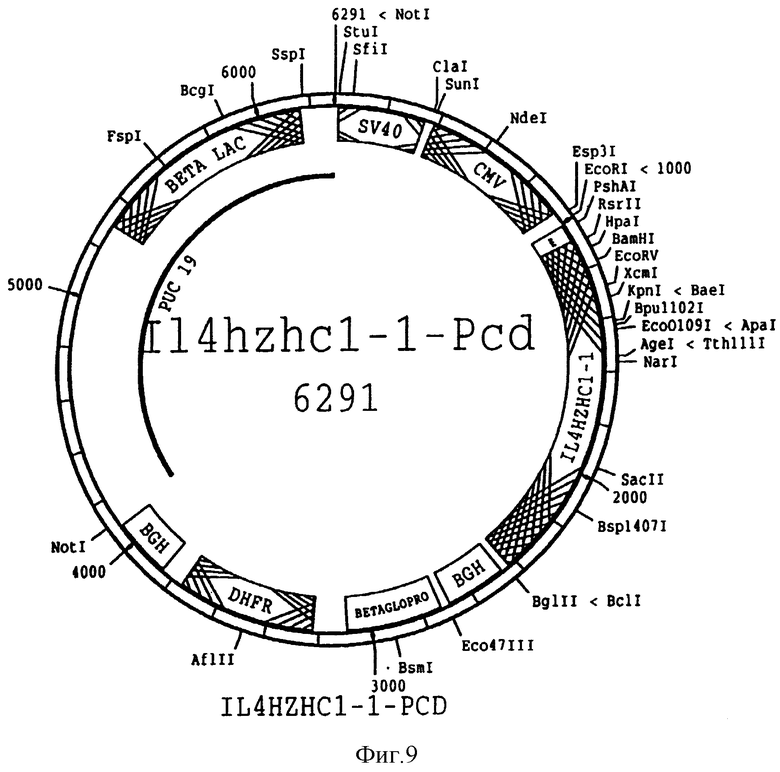
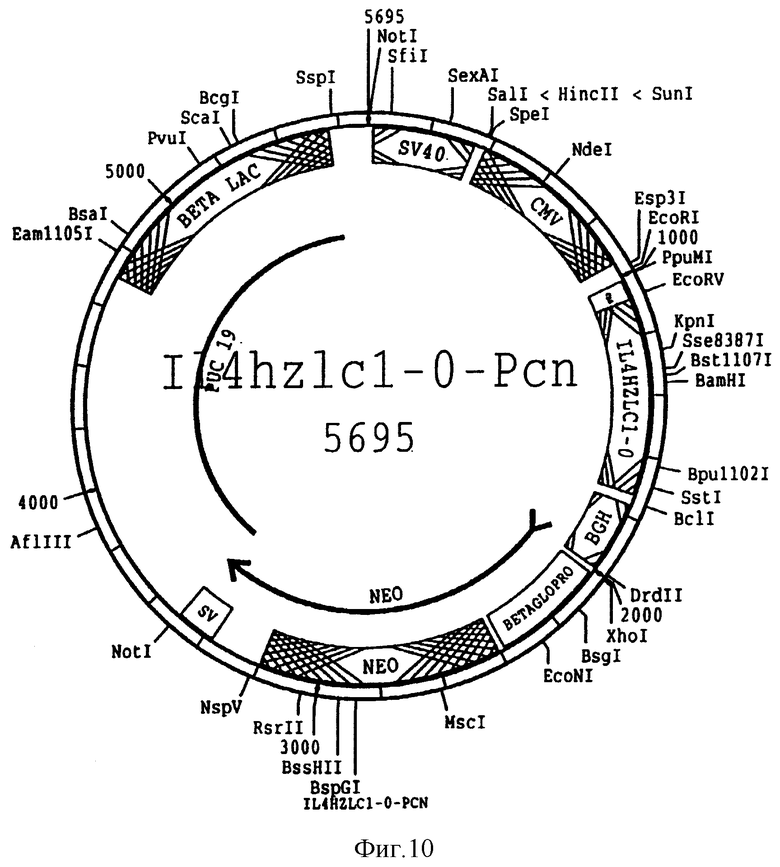
Изобретение относится к биотехнологии, а именно к рекомбинантным IL4-антителам, используемым для лечения нарушений, связанных с действием IL4. Сущность изобретения включает химерные и гуманизированные IL4 Mab-антитела, которые являются производными высокоэффективных Маb-антител, а также содержащие их фармацевтические композиции и способы диагностики и лечения. Преимущество изобретения заключается в создании высокоаффинного IL4-антагониста, способного снижать уровень эозинофильного воспаления. 22 с. и 3 з.п. ф-лы, 10 ил., 3 табл.
(a) ThrSerGlyMetGlyValSer: SEQ ID NO:22;
(b) HisIle TyrTrpAspAspLysArgTyrAsnProSerLeuLysSer: SEQ ID NO:24 или
(c) ArgGluThrVal PheTyrTrpPheAspVal: SEQ ID NO:26,
где аминокислотные последовательности областей, определяющих комплементарность легкой цепи, представляют собой
(a) LeuAlaSerGlnSerValAspTyrAspGlyAspSerTyrMetAsn: SEQ ID NO:16;
(b) AlaAlaSerAsnLeuGluSer: SEQ ID NO:18 или
(c) GlnGlnSerAsnGluAspProProArg: SEQ ID NO:28,
где аминокислотные последовательности областей, определяющих комплементарность легкой цепи, представляют собой
(a) LysAlaSerGlnSerValAspTyrAspGlyAspSerTyrMetAsn: SEQ ID NO:16;
(b) AlaAlaSerAsnLeuGluSer: SEQ ID NO:18 или
(c) GlnGlnSerAsnGluAspProProThr: SEQ ID NO:20,
происходящие из нечеловеческого моноклонального антитела, выбранного из группы, состоящей из 3B9 и 6А1, характеризуемого константой диссоциации, равной или меньшей 2 х 10-10 M относительно человеческого IL4, а также первый партнер слияния, где последовательность первого партнера слияния содержит последовательность тяжелой цепи: аминокислоты 21-50, 56-71, 88-119 и 131-141 последовательности SEQ ID NO: 12, где последовательность первого партнера слияния содержит последовательность легкой цепи: аминокислоты 20-42, 58-72, 80-111 и 121-131 и последовательности SEQ ID NO:14.
(a) ThrSerGlyMetGlyValSer: SEQ ID NO:22;
(b) HisIle TyrTrpAspAspAspLysArgTyrAsnProSerLeuLysSer: SEQ ID NO: 24, и
(c) ArgGluThrValPheTyrTrpPheAspVal SEQ ID NO:26.
(a) LeuAlaSerGlnSerValAspTyrAspGlyAspSerTyrMetAsn: SEQ ID NO:16;
(b) AlaAlaSerAsnLeuGluSer: SEQ ID NO: 18;
(c) GlnGlnSerAsnGluAspProProArg: SEQ ID NO:28 и
(c) GlnGlnSerAsnGluAspProProThr: SEQ ID NO:20.
(a) ACT TCT GGT ATG GGT GTG AGC: SEQ ID NO:21;
(b) CAC ATT TAC TGG GAT GAT GAC AAG CGC TAT; AACCCATCCCTGAAGAGC: SEQ ID NO:23;
(c)AGA GAG ACT GTG TTC TAC TGG TAC TTC GAT GTC: SEQ ID NO:25;
(d) ACC TCC GGT ATG GGT GTT TCC: SEQ ID NO:54;
(e) CAC ATC TAC TGG GAC GAC GAC AAA CGT TAC AAC CCG AGC CTG AAA TCC: SEQ ID NO: 55 и
(f) CGC GAA ACC GTT TTC TAC TGG TAC TTC GAC GTT: SEQ ID NO:56.
(a) AAG GCC AGC CAA AGT GTT GAT TAT GAT GCT GAT AGT TAT ATG AAC: SEQ ID NO:15;
(b) AAG GCC TCC CAA AGT GTT GAT TAT GAT GGT GAT AGT TAT ATG AAC: SEQ ID NO:53;
(c) GCT GCA TCC AAT CTA GAA TCT: SEQ ID NO:17;
(d) CAG CAA AGT AAT GAG GAT CCT CCG ACG: SEQ ID NO:19 и
(e) CAG CAA AGT AAT GAG GAT CCT CCG AGG: SEQ ID NO:27.
а) SEQ ID NO:21; SEQ ID NO:23;
SEQ ID NO:25; SEQ ID NO:54;
SEQ ID NO:55; SEQ ID NO:56;
SEQ ID NO:15; SEQ ID NO:53;
SEQ ID NO:17; SEQ ID NO:19 и
SEQ ID NO:27;
(b) нуклеино-кислотной последовательности, комплементарной а), с) нуклеиново-кислотной последовательности из 18 или более нуклеотидов, способной гибридизироваться в строгих условиях с (а) или (b), и
(d) фрагмента или аналога (а), (b) или (с), кодирующего белок, характеризующийся специфичностью в отношении человеческого интерлейкина-4.
| US 5317087 A, 31.05.1994 | |||
| EP 01626999 A2, 27.11.1985 | |||
| EP 0707065 A2, 17.04.1996 | |||
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| Огнетушитель | 0 |
|
SU91A1 |

