Изобретение относится, главным образом, к области медицины и конкретно к способам и композициям, предназначенным для ингибирования ангиогенеза тканей с использованием антагонистов витронектинового рецептора αvβ3.
Интегрины представляют собой класс клеточных рецепторов, которые, как известно, связывают внеклеточные матричные белки и, в связи с этим, регулируют взаимодействия клетка-клетка и клетка-внеклеточная матрица, которые обычно обозначаются как события адгезии клеток. Однако, хотя многие интегрины и лиганды, связывающие интегрин, описаны в литературе, биологическая функция многих интегринов остается неопределенной. Интегриновые рецепторы составляют семейство белков с разделенными структурными характеристиками нековалентных гетеродимерных гликопротеиновых комплексов, образованных из α и β субъединиц.
Витронектиновый рецептор, названный за его первоначальную характеристику преимущественного связывания с витронектином, как известно в настоящее время, имеет отношение к трем различным интегринам, обозначенным как αvβ1 αvβ3 и αvβ5, см. Horton Int. J. Exp. Pathol. 71:741-759 /1990/. αvβ1 Связывает фибронектин и витронектин. αvβ3 Связывает большое число лигандов, включая фибрин, фибриноген, ламинин, тромбоспондин, витронектин, фактор Виллебранда, остеоспонтин и костный сиалопротеин I. αvβ5 Связывает витронектин. Специфические роли в адгезии клеток эти три интегрина играют во многих клеточных взаимодействиях в тканях, исследуемых в настоящее время, однако очевидно, что имеются различные интегрины с различными биологическими функциями.
Одним из важных сайтов узнавания в лиганде для многих интегринов является аргинин-глицин-аспаргиновая кислота /RGD/ трипептидная последовательность. RGD обнаружена во всех лигандах, идентифицированных выше для витронектиновых рецепторных интегринов. Такой RGD - распознающий сайт может быть мимикрирован с помощью полипептидов /"пептиды"/, которые содержат RGD последовательность, и такие RGD-пептиды являются известными ингибиторами интегриновой функции. Однако важно отметить, что в зависимости от последовательности и структуры RGD-пептида специфичность ингибирования может изменяться в отношении мишень-специфичных интегринов.
Обсуждение RGD распознающего сайта можно найти в статье Pierschbacher с сотр. , в журнале Nature 309:30-33 /1984/ и Pierschbacher с сотр., в журнале Proc. Natl. Acad. Sci. США, 81:5985-5988 /1984/. Различные RGD-полипептиды различной интегриновой специфичности были также описаны Grant с сотр., в Cell, 58:933-943 /1989/ Cheresh с сотр. Cell, 58:945-953 /1989/, Aumailley с сотр. , FEBS Zetts. 291: 50-54 /1991/ и Pfaff с сотр. J.Biol. Chem., 269: 20233-20238 /1994/, а также в патентах США NN 4.517.686, 4.578.079, 4.589.881, 4.614.517, 4.661.111, 4.792.525, 4.683.291, 4.879.237, 4.988.621, 5.041.380 и 5.061.693.
Ангиогенез представляет собой процесс васкуляризации ткани, включающий рост вновь развивающихся в ткани кровеносных сосудов, и называется также неоваскуляризацией. Этот процесс опосредован инфильтрацией эндотелиальных клеток и клеток гладкой мышцы. Предполагается, что такой процесс реализуется одним из трех путей: сосуды могут прорастать из уже существующих сосудов, может иметь место de novo развитие сосудов из клеток предшественников /васкулогенез/, либо существующие мелкие сосуды могут увеличиваться в диаметре. См. Blood с сотр. Bioch. Biophys. Acta, 1032:89-118 /1990/. Как известно, сосудистые эндотелиальные клетки содержат, по крайней мере, пять RGD - зависимых интегринов, включая витронектиновый рецептор / αvβ3 или αvβ5 /, рецепторы коллагенового типа I и IV / α1β1 /, ламининовый рецептор / α2β1 /, фибронектин /ламинин/, коллагеновый рецептор / α3β1 / и фибронектиновый рецептор / α5β1 /. См. статью Davis с сотр., в J. Cell. Biochem. 52:206-218. /1993/. Известно, что клетка гладкой мышцы содержит, по крайней мере, шесть RGD-зависимых интегринов, включая α5β1, αvβ3 и αvβ5 .
Ангиогенез является важным процессом неонатального роста, но он столь же важен в заживлении ран и в патогенезе большого числа клинических заболеваний, таких как воспаление тканей, артриты, рост опухолей, диабетическая ретинопатия, пятнистая дегенерация в результате неоваскуляризации сетчатки глаза и при родственных состояниях. Такие клинические состояния связанны с ангиогенезом и на них ссылаются, как на ангиогенные заболевания. См. Folkman с сотр., Science 235:442-447 /1987/. Ангиогенез обычно отсутствует во взрослых или зрелых тканях, однако он обычно присутствует при лечении ран и в цикле роста corpeus leuteum. См. , например, Moses с сотр. Science, 248: 1408-1410 /1990/.
Было предположено, что ингибирование ангиогенеза может служить полезной терапией ограничения роста опухолей. Ингибирование ангиогенеза может осуществляться
/1/ ингибированием выделения таких "ангиогенных молекул", как βFGF /фактор роста фибробласта/,
/2/ нейтрализацией ангиогенных молекул, например путем использования анти-βFGF антител, и
/3/ ингибированием реакции эндотелиальной клетки на ангиогенную стимуляцию. Последнему варианту было уделено внимание и Folkman с сотр., (Cancer Biology, 3:89-96, 1992) описал несколько ингибиторов эндотеллиального клеточного ответа, включая коллагеназный ингибитор, базовый мембранный циклический ингибитор, ангиостатические стероиды, ингибиторы ангиогенеза грибкового происхождения, тромбоцитный фактор 4, тромбоспондин, такие артритные лекарства, как D-пеницилламин и тиомолат золота, аналоги витамина D3, альфа-интерферон и многие другие препараты, которые могут использоваться для ингибирования ангиогенеза. Другие предложенные ингибиторы ангиогенеза описаны Blood с сотр. , Bioch. Bioph. Acta, 1032-89-118 /1990/, Moses с сотр. Science 248: 1408-1410 /1990/, Ingber с сотр. Zab Invest. 59:44:51 /1988/ и также в патентах США NN 5.092.885, 5,112.946, 5.192.744, и 5.202.352. Ни один из ингибиторов ангиогенеза, описанный ранее, не обладал способностью ингибировать αvβ3 RGD-содержащие пептиды, ингибирующие витронектиновый рецептор αvβ3 также был описан. Aumailley с сотр. FEBS Zetts 291:50:54 /1991/., Choi с сотр. J. Vasc. Surg 19: 125-134 /1994/, Smith с сотр., J. Biol. Chem. 265: 12267-12271 /1990/ и Pfaff с сотр., J.Biol. Chem. 269:20233-20238 /1994/. Однако, до момента создания настоящего изобретения роль интегрина αvβ3 в развитии ангиогенеза не была предположена или установлена.
Ингибирование клеточной адгезии in vitro с использованием моноклональных антител, иммуноспецифичных для различных интегриновых субъединиц α или β, вовлекает αvβ3 в адгезию клеток большого числа типов, включая микрососудистые эндотелиальные клетки. См. Davis с сотр. J. Cell Biol: 51:206-218 /1993/. Кроме этого, Nicosia с сотр., Am. J. Pathol. 138:829-833 /1991/, описали использование RGD-пептида GRGDS для in vitro ингибирования образования "микрососудов" крысиной аорты, культурированной в коллагеновом геле. Однако, ингибирование образования "микрососудов" in vitro в коллагеновых гелевых культурах не является моделью ингибирования ангиогенеза в ткани, поскольку не показано, что микрососудистые структуры аналогичны капиллярным наростам, или что образование микрососудов в коллагеновой гелевой культуре аналогично росту новых сосудов в интактной ткани, например в артритной ткани, опухолевой ткани или вольной ткани, где желательно ингибирование ангиогенеза.
Поэтому, помимо сообщенных выше исследований, заявители неожиданно обнаружили другую демонстрацию того факта, что ангиогенез может ингибироваться в тканях с использованием ингибиторов клеточной адгезии. Главным образом, никогда ранее не было показано, что αvβ3 функция требуется для ангиогенеза в тканях, или что αvβ3-антагонисты могут ингибировать ангиогенез в тканях.
Краткое изложение сущности изобретения
В настоящем изобретении продемонстрирован тот факт, что ангиогенез в тканях требует наличия интегрина αvβ3 и что ингибиторы αvβ3 могут ингибировать ангиогенез.
Обнаружение этого факта также демонстрирует то, что антагонисты других интегринов, например, αvβ5 или αvβ1 не ингибируют ангиогенез, возможно по причине их несущественности для развития ангиогенеза.
В этой связи, в изобретении описываются способы ингибирования ангиогенеза в тканях, включающие применение на ткани композиции, включающей ангиогенез-ингибирующее количество αvβ3-антагониста.
Лечению может подвергаться любая ткань, для которой желательно ингибирование ангиогенеза, такая как "больная" ткань, где происходит неоваскуляризация. Примерами таких тканей могут служить воспаленная ткань, твердые опухоли, метастазы, ткани, подверженные ретенозу, и другие ткани.
Используемый в настоящем изобретении αvβ3-антагонист способен связывать αvβ3 и полностью ингибировать способность αvβ3 связываться с природным лигандом. Предпочтительно, чтобы такой антагонист обладал специфичностью в отношении αvβ3 по сравнению с другими интегринами. Согласно особенно предпочтительному воплощению αvβ3-антагонист ингибирует связывание фибриногена или другого RGD-содержащего лиганда с αvβ3, но несущественно ингибирует связывание фибриногена с α11bβ3. Предпочтительным αvβ3-антагонистом может служить полипептид, или моноклональное тело, или их функциональный фрагмент, который иммунореагирует с αvβ3 .
Краткое описание чертежей.
На чертежах, составляющих часть настоящего описания, показано следующее.
На фиг. 1A-1Г проиллюстрировано распределение в ткани интегриновых субъединиц β3 и β1 в случае нормальной кожи и кожи в процессе заживления ран, обозначенной как гранулированная ткань. Иммуногистохимия с использованием антител к β3 и β1 осуществлялась, как описано в примере 3А. На фиг. 1A и 1Б соответственно проиллюстрирована иммунореактивность анти-β3 в нормальной коже и в гранулированной ткани. На фиг. 1В и 1Г соответственно проиллюстрирована иммунореактивность анти- β1 в нормальной коже и в гранулированной ткани.
На фиг. 2A-2Г соответственно показано тканевое распределение фактора фон Виллебранда и ламининовых лигандов, которые соответственно связывают интегриновые субъединицы β3 и β1 в нормальной коже и в коже в процессе заживления ран, обозначенной как гранулированная ткань. Иммуногистохимию с использованием антител на фактор фон Виллебранда /анти-vWF /и ламинин /анти-ламинин/ осуществляли, как описано в Примере 3B. На фиг. 2A и 2Б соответственно проиллюстрирована иммунореактивность анти-vWF в нормальной коже и в гранулированной ткани. Фиг. 2В и 2Г соответственно показывают иммунореактивность анти-ламинина в нормальной коже и в гранулированной ткани.
На фиг. 3A-3Г продемонстрировано распределение витронектинового интегринового рецептора, αvβ3 в биопсии ткани рака мочевого пузыря, рака толстой кишки, рака молочной железы и рака легких соответственно.
Иммуногистохимию с использованием LM609 антитела к αvβ3 проводили, как описано в примере 3C.
На фиг. 4 показана типичная микрофотография CAM настоящего изобретения, лишенной кровеносных сосудов в необработанном 10-дневном курином эмбрионе. Препарат описан в Примере 5B.
На фиг. 5A-5В проиллюстрировано распределение в ткани интегринов β1 и αvβ3 в препарате CAM настоящего изобретения. На фиг. 5A показано распределение β1 субъединицы в необработанном 10-дневном препарате CAM, как детектировано с помощью иммунофлуоресцентной иммунореактивности с использованием CSAT, анти- β1-антителом. На фиг. 5Б показано распределение αvβ3 рецептора в необработанном 10-дневном препарате CAM, как детектировано с помощью иммунофлуоресцентной иммунореактивности с LM609, анти- αvβ3-антителом. На фиг. 5В показано распределение рецептора αvβ3 в обработанном β FGF 10-дневном препарате CAM, как детектировано с помощью иммунофлуоресцентной иммунореактивности с LM609, анти- αvβ3-антителом. Методы лечения и полученные результаты описаны в примере 5C.
На фиг. 6 проиллюстрировано количественное определение в столбце диаграммы относительной экспрессии αvβ3 и β1 в необработанных и обработанных βFGF 10-дневных препаратах CAM в соответствии с описанным в Примере 6A. Среднее значение интенсивности флуоресценции отложено по оси-Y, а интегриновые профили отложены по оси X.
На фиг. 7A-7В проиллюстрирован внешний вид необработанного 10-дневного CAM, образца CAM, обработанного βFGF, и образца CAM, обработанного TNFα , соответственно, методики и результаты описаны в примере 6A.
Фиг. 8A-8Д иллюстрируют влияние актуальной обработки антителом на FGF-индуцированный ангиогенез на 10 день CAM, как это описано в Примере 7A1/. На фиг. 8A показан необработанный CAM препарат, лишенный кровеносных сосудов. На фиг. 8Б продемонстрирована инфильтрация новой сосудистой сети в область, предварительно лишенную сосудистой системы, индуцированную βFGF обработкой. Фиг. 8В, 8Г и 8Д соответственно демонстрируют эффекты антител против β1 /антитело-β, CSAT/, αvβ5 /анти-αvβ5 P3C2/ и αvβ3 /анти- αvβ3 ; LM609/.
На фиг. 9A-9В проиллюстрировано влияние внутривенной инъекции синтетического пептида 66203 на ангиогенез, индуцированный опухолью, как описано в примере 7Д2. На фиг. 9A показано отсутствие ингибиторного эффекта внутривенной обработки контрольным пептидом /контрольная пептидная опухоль/ на ангиогенез, вызванный индукцией опухоли. Ингибирование такого ангиогенеза путем внутривенной инъекции пептида 66203 /циклическая RGD опухоль/ представлено на фиг. 9Б. Отсутствие ингибирующего эффекта или цитотоксичности на зрелые пресуществующие сосуды после внутривенного вливания пептида 66203 в область, примыкающую к опухоль-обработанной области, показано на фиг. 9В /циклический RGD, соседствующий с CAM/.
На фиг. 10A-10В проиллюстрирован эффект внутривенного применения моноклональных антител на ангиогенез, индуцируемый фактором роста, как это описано в примере 7B1/. Фиг. 10A демонстрирует индуцированный с помощью βFGF ангиогенез без обработки антителом /контрольный эксперимент/. Ингибирование ангиогенеза не имеет место, когда аналогичный препарат был обработан анти- αvβ5 антителом P3G2, как это показано на фиг. 10Б.
Ингибирование ангиогенеза реализуется при обработке анти- αvβ3 антителом LM609, как это показано на фиг. 10В.
Фиг. 11A-11В иллюстрируют эффект на эмбрионный ангиогенез после местного применения анти-интегриновых антител, как описано в примере 7C. Ангиогенез не ингибируется в результате обработки 6-дневного CAM анти- β1 и анти αvβ5 антителами соответственно, что показано на фиг. 11A и 11Б. Напротив, обработка анти- αvβ3 антителом LM609 приводит к ингибированию образования кровеносных сосудов, как это показано на фиг. 11В.
На фиг. 12 произведено количественное определение числа сосудов, входящих в опухоль в препарате CAM, как это описано в примере 7Д1/. На этом графике число сосудов нанесено на ось Y, как результат местного применения или CSAT /анти- β1 / LM609 /анти- αvβ3 или P3G2/, анти- αvβ5 /.
Фиг. 13A-13Г иллюстрируют сравнение между весами мокрых опухолей через 7 дней после обработки и первоначальными весами, как описано в примере 9A1/а. Каждая полоса представляет среднее ± S.E. 5-10 опухолей на группу. Используемые опухоли были производными человеческой меланомы /М21-L/ /фиг. 13A/, рака поджелудочной железы /Fg/ /фиг. 13Б/, легочной карциномы /YCLAP-3/ /фиг. 13В/ и ларингеальной карциномы /Hep3/ /фиг. 13Г/, CAM-препараты и внутривенно обработанные PBS, CSAT /aнти- β1 / или LM609 /анти- αvβ3 /. Графики показывают вес опухоли, значение которого нанесено на ось Y в результате внутривенного применения или CSAT /анти- β1 /, LM609 /анти- αvβ3 / или PBS как показано на оси X.
На фиг. 14A и 14Б показаны гистологические разрезы опухолей, обработанных P3G2 /aнти- αvβ5 / /фиг. 14A/ и LM609 /анти- αvβ3/ фиг. 14Б, и окрашенных гематоксилином и эозином, как это описано в примере 9A1/а. Как показано на фиг. 14A опухоли, обработанные контрольным антителом /P3G2/, демонстрировали большое число жизнеспособных и активно делящихся опухолевых клеток, о чем свидетельствовали митотические значения /показаны стрелками сверху/, а также множественные кровеносные сосуды /стрелки/ по всей строме опухоли. Напротив, очень небольшое количество, если оно вообще определялось, жизнеспособных опухолевых клеток или кровяных сосудов детектировалось в клетках, обработанных LM609 /анти- αvβ3 /, как показано на фиг. 14Б.
Фиг. 15A-15Д соответствуют М21L опухолям, обработанным пептидами как описано в Примере 9A1/b, и они представляют собой следующее:
фиг. 15A - контрольный циклический RAD пептид /69601/; фиг. 15Б - циклический RGD пептид /66203/; фиг. 15В - соседняя CAM ткань, взятая из того же эмбриона, обработанного циклическим RGD пептидом /66203/, и на фиг. 15Г представлено сильное увеличение /13х/ опухолей, обработанных контрольным RAD /69601/, а на фиг. 15Д показано увеличение при обработке циклическим RGD пептидом /66203/. Фиг. 15Г показывает нормальные сосуды из опухоли, обработанной RAD контрольным пептидом 69601/. На фиг. 15Д приведены примеры разрушенных кровеносных сосудов из опухолей, обработанных циклическим RGD пептидом /66203/ /стрелки/.
На фиг. 16А-16Д показано ингибирование ангиогенеза под действием антагонистов ангиогенеза в модельном анализе in vivo на кроличьем глазу в соответствии с описанным в примере 10. На фиг. 16A и 16Д изображен ангиогенез кроличьего глаза в присутствии βFGF и mabPIF6 /анти- αvβ5 /. На фиг. 16Б, 16В и 16Д показано ингибирование ангиогенеза кроличьего глаза в присутствии βFGF и mAbLM609 /анти- αvβ3 /.
На фиг. 17 представлена диаграмма, показывающая как можно создать in vivo мышь : человек химерную мышиную модель, что описано в примере 11A. Часть шкуры мыши SC1D заменяли на человеческую неонатальную крайнюю плоть и давали системе залечиваться в течение 4 недель. После залечивания прививки человеческую крайнюю плоть инокулировали клетками человеческой опухоли. В течение следующих 4 недель проявлялась измеримая опухоль, которая состояла из человеческой опухоли с человеческой сосудистой системой, проходящей из человеческой плоти в человеческую опухоль.
На фиг. 18 проиллюстрирован процент единичных клеток, полученных от CAM обработанной mAB и пептидом и окрашенных Apop Tag, как установлено FACS анализом и описано в примере 12A и 12B соответственно. Черные и полосатые столбцы представляют собой клетки эмбрионов, обработанный за 24 и 48 часов до анализа соответственно. Каждый столбец представляет среднее ±S.E. трех измерений. CAM обрабатывали mAb LМ609 /анти- αvβ3/ или CSAT /анти- β1/ или PBS, как описано в Примере 12A2. CAM также обрабатывали циклическим пептидом 69203 /цикло-RGD f Y, обозначаемым как пептид 203/ или контрольным циклическим пептидом 69601 /цикло-RAD fY, обозначаемым как пептид 601/, как это описано в примере 12B.
На фиг. 19A и 19Б проиллюстрированы объединенные результаты суспензий отдельных клеток CAM из эмбриона, обработанного либо CSAT /анти- β1/ /фиг. 19A/ или LM609 /анти- αvβ3/ /фиг. 19Б/, окрашенных Apop Tag и иодистым пропидием и проанализированных методом FACS, как описано в Примере 12C. По оси Y отложены результаты окрашивания Apop Tag в числе клеток /Аполтоз/, по оси X отложены результаты, соответствующие окрашиванию иодистым пропидием /содержание ДНК/. Горизонтальные линии представляют собой отрицательный вход для окрашивания Apop Tag. Левая и правая панели показывают на клетки CAM из эмбрионов, обработанных CSAT /анти- β1/ /фиг. 19A/ и LM609 /анти- αvβ3/ /фиг. 19Б/. Осуществляли анализ клеточного цикла, определяя примерно 8.000 событий на состояние.
Фиг. 20A-20C представляют собой изображения CAM ткани из CSAT /анти- β1/ обработанных эмбрионов, а фиг. 20D-20F отображают CAM ткани из LM609 /анти- αvβ3/ обработанных эмбрионов, полученных как описано в примере 12C. Фиг. 20A и 20D демонстрируют ткани, окрашенные Apop Tag и визуализированные методом флуоресценции /FITC/ суперналоженным на D.I.C. изображением. На фиг. 20B и 20E показаны те же ткани, окрашенные mAb LM609 /анти- αvβ3/ и визуализированные с помощью флуоресценции /родамин/. На фиг. 20C и 20F представлены изображения тех же тканей, окрашенных как Apop Tag, так и LM609, причем желтое окрашивание свидетельствует о солокализации. Полосы в левой и правой панели составляют 15 и 50 мкм соответственно.
Детальное описание изобретения
A. Обозначения:
Аминокислотные остатки: Аминокислота образуется в ходе химического переваривания /гидролиз/ полипептида по его пептидной связи. Описанные в тексте аминокислотные остатки предпочтительно находятся в "L" изомерной форме. Однако, остатки, находящиеся в "D" изомерной форме, могут быть заменены на любой L-аминокислотный остаток, если полипептид сохраняет желаемое функциональное свойство. NH2 относится к свободной аминогруппе на амино-конце полипептида. COOH относится к свободной карбоксигруппе, присутствующей на C-конце полипептида. Придерживаясь стандартной полипептидной номенклатуры /описанной в J. Biol. Chem. 243:3552-59 /1969/ и принятой 37 CFP 1.822/b/2/, сокращенные названия аминокислотных остатков представлены в Таблице Соответствия (см. в конце описания).
Следует отметить, что все аминокислотные последовательности представлены в тексте формулами, левая и правая ориентация которых представлены в традиционном направлении от амино-конца к карбокси-концу. Кроме этого следует также отметить, что штрих в начале или конце аминокислотной последовательности указывает на пептидную связь со следующей последовательностью одного или более аминокислотных остатков.
Полипептид: относится к линейному ряду аминокислотных остатков, связанных друг с другом пептидными связями между алфа-аминогруппой и карбоксигруппой непрерывной аминокислотной последовательностью.
Пептид: этот термин в контексте изобретения относится к линейным рядам не более примерно 50 аминокислотных остатков, связанных друг с другом как в полипептиде.
Циклический пептид: является производным соответствующего линейного пептида, относится к пептиду, в котором не существуют N- или C-концы и в котором соответствующие линейные пептидные N-терминальные формы и амидная связь с C-терминальным карбоксилатом существуют, как часть указанного линейного пептида.
Термин белок /протеин/: относится к линейным сериям из более, чем 50 аминокислотных остатков, связанных друг с другом как в полипептиде.
Синтетический пептид: относится к химически полученной цепи аминокислотных остатков, связанных друг с другом пептидными связями, которые не содержат встречающиеся в природе белки и их фрагменты.
B. Общие соображения
Настоящее изобретение относится главным образом к тому открытию, что ангиогенез опосредован специфическим витронектиновым рецептором αvβ3 и что ингибирование αvβ3 функции ингибирует ангиогенез. Такое открытие является важным из-за роли, которую играет ангиогенез в большом числе заболеваний. В результате ингибирования ангиогенеза можно вмешиваться в ход заболевания, улучшать симптомы и, в ряде случаев, излечивать заболевание.
В том случае, когда рост новых кровеносных сосудов вызывается или вносит вклад в патологию, связанную с заболеванием, ингибирование ангиогенеза будет уменьшать вредные эффекты такого заболевания. Примеры таких заболеваний включают ревматоидный артрит, диабетическую ретинопатию, воспалительные заболевания, рестеноз и т.п. В том случае, когда рост новых кровеносных сосудов требуется для поддержки роста вредной ткани, ингибирование ангиогенеза приведет к понижению подачи крови в ткань и тем самым внесет вклад в уменьшение образовавшейся тканевой массы, основанной на условиях подачи крови. Примеры такого рода включают рост опухолей, в которых неоваскуляризация является постоянным требованием с тем, чтобы размер опухоли достиг нескольких миллиметров в толщину и для образования твердых опухолевых метастазов.
Способы настоящего изобретения эффективны частично, поскольку такая терапия высоко селективна в отношении ангиогенеза, а не для других биологических процессов. Как показано в примерах, лишь рост новых сосудов содержит достаточное количество αvβ3 и поэтому терапевтические методы не оказывают вредного влияния на зрелые сосуды. Кроме этого, αvβ3 не является широко распространенным в нормальных тканях и в основном обнаруживается селективно на новых сосудах, что застраховывает тот факт, что такая терапия может селективно избирать мишенью рост новых сосудов.
Открытие того факта, что ингибирование лишь αvβ3 эффективно ингибирует ангиогенез, позволяет создавать терапевтические композиции с потенциально высокой специфичностью и поэтому с относительно низкой токсичностью. Таким образом, хотя настоящее изобретение раскрывает использование реагентов на базе пептидов, обладающих способностью ингибировать один или более интегринов, можно применять и другие реагенты, которые более селективно ингибируют αvβ3 , и поэтому не оказывают побочных эффектов ингибирования других биологических процессов, кроме опосредуемых αvβ3 .
Так например, как показано в настоящем изобретении, существует возможность получения моноклональных антител, высоко селективных в отношении иммунной реакции с αvβ3, которые аналогичным образом селективны в ингибировании αvβ3-функции. Кроме этого, RGD-содержащие пептиды могут применяться в качестве селективных агентов в ингибировании αvβ3, как будет описано ниже.
Перед открытием настоящего изобретения не было известно, что ангиогенез и любые процессы, зависящие от ангиогенеза, могут ингибироваться in vivo с применением реагентов, являющихся антагонистами биологической функции αvβ3
C. Способы ингибирования ангиогенеза.
Настоящее изобретение обеспечивает способ ингибирования ангиогенеза в ткани и в результате ингибируются события в ткани, которые зависят от ангиогенеза. Обычно такой способ включает применение на ткани композиции, включающей ангиогенез-ингибирующее количество αvβ3-антагониста.
Как указывалось ранее, ангиогенез ответственен за большое число процессов, включающих неоваскуляризацию ткани, включая "прорастание", васкулогенез или расширение сосудов, причем все такие ангиогенезные процессы опосредованы и зависят от экспрессии αvβ3. За исключением случаев заживления травматической раны, образования corpus leuteum и эмбриогенеза полагают, что большая часть ангиогенезных процессов связана с заболеваниями и поэтому использование терапевтических методов настоящего изобретения является селективным для таких заболеваний и не дает вредных побочных эффектов.
Имеется большое число заболеваний, в которых предполагается, что важным их фактором является ангиогенез и в этом отношении можно сослаться на ангиогенные заболевания, не ограничиваясь только ими, такие как воспалительные расстройства, например иммунные и не иммунные воспаления, хронический артикулярный ревматизм и псориаз, нарушения, связанные с неправильной и несвоевременной инвазией сосудов, такие как диабетическая ретинопатия, неоваскулярная глаукома, рестеноз, капиллярная пролиферация в атеросклеротических бляшках и остеопороз, а также нарушения связанные с развитием рака, такие как твердые опухоли, метастазы твердых опухолей, ангиофибромы, ретролентальная фиброплазия, гемангиома, саркома Капоши и подобные раковые заболевания, которые требуют неоваскуляризации для поддержки роста опухоли.
Таким образом, способы, ингибирующие ангиогенез в заболевших тканях, облегчают симптомы заболевания и, в зависимости от типа заболевания, могут способствовать излечению заболевания. Согласно одному из воплощений, настоящее изобретение охватывает ингибирование ангиогенеза, per se, в тканях. Степень ангиогенеза в ткани и поэтому степень ингибирования, достигаемая настоящими методами, может быть оценена большим числом способов, например теми, что описаны в Примерах детекции αvβ3-иммуноположительной зрелости и структуры зародившихся сосудов с помощью иммуногистохимии.
Следует иметь в виду, что большое число тканей или органов, состоящих из организованной ткани, могут поддерживать ангиогенез в болезненном состоянии, причем к таким тканям и органам могут относиться кожа, мышца, кишки, соединительная ткань, суставы, кости и т.д., то есть те случаи, когда кровеносные сосуды способны вторгаться в систему в результате ангиогенного стимулирования.
Таким образом, в одном из случаев ткань, подвергаемая лечению, представляет собой воспаленную ткань и ангиогенез, подвергаемый ингибированию, представляет собой ангиогенез воспаленной ткани, когда реализуется неоваскуляризация воспаленной ткани. В случае таких нарушений способ изобретения предусматривает ингибирование ангиогенеза в артритных тканях, как это имеет место у пациентов с хроническим артикулярным ревматизмом, в иммунных или неиммунных воспаленных тканях, в псориатических тканях и т.п.
Пациент, подвергаемый лечению с помощью метода настоящего изобретения, согласно большинству решений, желательно представляет собой человека, хотя следует иметь в виду, что принципы изобретения показывают, что оно эффективно в отношении всех млекопитающих, которые подпадают под термин "пациент". В этом контексте под млекопитающим понимается любая его разновидность, для которого желательно лечение заболевания, связанного с ангиогенезом, особенно это относится к разновидностям сельскохозяйственных и домашних млекопитающих.
Согласно другому родственному решению ткань, подлежащая лечению, представляет собой ткань сетчатки пациента больного диабетической ретинопатией, пятнистой дегенерацией или неоваскулярной глаукомой и ангиогенез, подлежащий ингибированию, представляет ангиогенез ткани сетчатки, когда реализуется неоваскуляризация ткани сетчатки.
Согласно другому воплощению ткань, подлежащая лечению, представляет опухолевую ткань пациента с твердой опухолью, метастазами, раком кожи, раком молочной железы, больного гемангиомой или ангиофибромой и другими видами рака, и ангиогенез, подлежащий ингибированию, представляет собой ангиогенез опухолевой ткани, в которой реализуется неоваскуляризация опухолевой ткани. Типичными твердыми опухолевыми тканями, обрабатываемыми методами изобретения, могут быть ткани легких, поджелудочной железы, молочной железы, толстой кишки, гортани, яичников и аналогичные ткани. Примеры ангиогенеза опухолевых тканей и его ингибирование описано в Примерах.
Ингибирование ангиогенеза опухолевой ткани является особенно предпочтительным воплощением изобретения в связи с важной ролью неоваскуляризации в росте опухоли. В отсутствие неоваскуляризации опухолевой ткани такая ткань не получает требуемых питательных веществ, снижается ее рост, прекращается дополнительный рост, она регрессирует и в конечном счете становится некротической тканью, приводя к гибели опухоли.
Говоря другими словами, настоящее изобретение обеспечивает способ ингибирования опухолевой неоваскуляризации путем ингибирования ангиогенеза опухоли в соответствии с методом настоящего изобретения. Аналогичным образом, изобретение обеспечивает способ ингибирования роста опухоли в результате практической реализации методов ингибирования ангиогенеза.
Способы изобретения особенно эффективны при их применении против развития метастазов поскольку
/1/ их образование требует васкуляризации первичной опухоли так, что метастазные клетки рака могли бы входить в первичную опухоль и
/2/ их образование на второй стадии требует неоваскуляризации для поддержки роста метастазов.
Согласно родственному воплощению настоящее изобретение охватывает практическую реализацию способа, связанного с другими терапиями, например с традиционной химиотерапией, направленной против твердых опухолей и для контроля образования метастазов. Применение ингибитора ангиогенеза обычно проводится в ходе или после химиотерапии, хотя иногда предпочтительно ингибировать ангиогенез после режима химиотерапии, когда опухолевая ткань будет ответственна за токсичный "штурм" индукцией ангиогенеза для регенерации при условии подачи к опухолевой ткани крови и питательных веществ. Кроме этого, предпочтительно применять методы ингибирования ангиогенеза после хирургических операций, в которых твердые опухоли были удалены в качестве профилактики метастазов.
Несмотря на то, что настоящие способы направлены на ингибирование опухолевой неоваскуляризации такие методы могут также применяться для ингибирования роста опухолевой ткани, ингибирования образования опухолевых метастазов и регрессии образовавшихся опухолей. Представленные примеры демонстрируют регрессию образовавшихся опухолей после единичного внутривенного применения αvβ3-антагониста настоящего изобретения.
Рестеноз представляет собой процесс миграции и пролиферации клеток гладкой мышцы /SMC/ в место подкожной транслюминальной коронарной ангиопластии, что затрудняет успех ангиопластики. Миграция и пролиферация SMC в ходе рестеноза может рассматриваться, как процесс ангиогенеза, который ингибируется методами настоящего изобретения. Поэтому настоящее изобретение также предусматривает ингибирование рестеноза путем ингибирования ангиогенеза, согласно методу изобретения, у пациента после применения методов ангиопластики. Для ингибирования рестеноза αvβ3-антагонист обычно применяют после ангиопластики через 2-28 дней и как правило в течение 14 дней после операции.
Настоящий метод ингибирования ангиогенеза в тканях и поэтому также предназначенный для практической реализации методов лечения, связанных с ангиогенезом заболеваний, заключается в контактировании ткани с развившимся аутогенезом, или при риске его появления с композицией, включающей терапевтически эффективное количество антагониста, способного ингибировать αvβ3, осуществляя связывание с его природным лигандом. Таким образом, такой способ заключается в применении на пациенте терапевтически эффективного количества физиологически переносимой композиции, содержащей αvβ3-антагонист изобретения.
Дозированные интервалы для введения αvβ3-антагониста зависят от формы антагониста и его мощности, как это описано далее в тексте, и составляют достаточно большие количества для обеспечения желаемого эффекта, согласно которому ангиогенез и симптомы заболевания, опосредованные ангиогенезом, подвергаются улучшению. Дозировка не должна быть столь высокой, чтобы вызывать нежелательные побочные эффекты, например синдромы гипервязкости, легочную эдему, сердечные нарушения, связанные с перегрузкой, и т.п. Обычно, дозировка будет меняться в зависимости от возраста, состояния больного, пола и степени заболевания пациента и она может быть определена специалистом. Такая дозировка также может регулироваться индивидуальным терапевтом в случае осложнений.
Терапевтически эффективное количество представляет собой такое количество αvβ3-антагониста, которое достаточно для обеспечения измеримого ингибирования ангиогенеза в обрабатываемой ткани, например ангиогенез-ингибирующее количество. Ингибирование ангиогенеза может быть измерено in situ методом иммуногистохимии, как описано в настоящем документе, или другими методами, известными специалистам.
Поскольку αvβ3-антагонист может принимать вид мимикрирующего αvβ3 и RGD -содержащего пептида, анти- αvβ3 моноклонального антитела или его фрагмента, следует убедиться, что мощность и поэтому экспрессия "терапевтически эффективного" количества могут изменяться. Однако, как показано методами анализа изобретения, специалист в данной области может легко оценить мощность αvβ3 -антагониста /кандидата настоящего изобретения.
Мощность αvβ3 -антагониста может быть измерена множеством способов, включая ингибирование ангиогенеза в анализе CAM, в in vivo анализе на глазах кролика, в in vivo анализе с использованием химерного мышь-человека и путем измерения ингибирования связывания природного лиганда с αvβ3 , причем все эти анализы описаны в тексте или с помощью аналогичных анализов.
Предпочтительный αvβ3 антагонист обладает способностью существенно ингибировать связывание такого природного лиганда, как фибриноген или витронектин, с αvβ3 в растворе при концентрациях антагониста менее 0,5 микромоля /мкм/, предпочтительно менее 0,1 мкм и более предпочтительно менее 0,05 мкм. Под термином "существенно" подразумевается, что по крайней мере 50% снижение связывания фибриногена наблюдается в результате ингибирования в присутствии антагониста и на 50% ингибирования ссылаются, как на значение IC50.
Более предпочтительный αvβ3 -антагонист проявляет селективность в отношении αvβ3 сравнению с другими интегринами. Так например, предпочтительный αvβ3 антагонист существенно ингибирует связывание фибриногена с αvβ3 но не существенно ингибирует связывание фибриногена с такими интегринами, как αvβ1, αvβ5 или 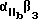 . Особенно предпочтительным является αvβ3 антагонист, проявляющий в 10-100 раз более низкую активность по значению IC50 при ингибировании связывания фибриногена с αvβ3, по сравнению с IC50 активностью при ингибировании связывания фибриногена с другим интегрином. Примеры анализов измерения IC50 активности при ингибировании связывания фибриногена с интегрином описаны в Примерах изобретения.
. Особенно предпочтительным является αvβ3 антагонист, проявляющий в 10-100 раз более низкую активность по значению IC50 при ингибировании связывания фибриногена с αvβ3, по сравнению с IC50 активностью при ингибировании связывания фибриногена с другим интегрином. Примеры анализов измерения IC50 активности при ингибировании связывания фибриногена с интегрином описаны в Примерах изобретения.
Терапевтически эффективное количество αvβ3 антагониста изобретения в виде моноклонального антитела обычно составляет такое количество, которое при применении в физиологически переносимой композиции достаточно для достижения концентрации плазмы в диапазоне от 0,01 микрограмм /μг/ на миллилитр /мл/ до 1000 мкг/мл, предпочтительно 1 мкг/мл - 5 мкг/мл и обычно около 5 мкг/мл. Иначе говоря, дозировка может изменяться от 0,1 мг/кг до 300 мг/кг, предпочтительно от 0,2 мг/кг до 200 мг/кг, наиболее предпочтительно 0,5 мг/кг до 20 мг/кг при применении одной или более дозировки ежедневно в течение одного или нескольких дней.
В том случае, когда антагонист находится в виде фрагмента моноклонального антитела, такое количество может легко регулироваться на основании отношения массы фрагмента относительно массы всего антитела. Предпочтительная концентрация плазмы в молярном выражении составляет от приблизительно 2 микромолярной /μМ/ до 5 миллимолярной /mM/ и предпочтительно от около 100 мкм до 1 mM антительного антагониста.
Терапевтически эффективное количество αvβ3-антагониста изобретения в виде полипептида или другого молекулярного αvβ3 миметика аналогичного малого размера обычно составляет такое количество полипептида, которое при введении в физиологически толерантной композиции достаточно для достижения концентрации плазмы от около 0,1 микрограмма /μг/ на миллилитр /мл/ до приблизительно 200 мкг/мл, предпочтительно от около 1 мкг/мл до приблизительно 150 мкг/мл. В расчете на полипептид с массой около 500 г/моль предпочтительная концентрация плазмы в молярном выражении составляет от около 2 микромолярной /μM/ до около 5 миллимолярной /mM/ и предпочтительно около 100 мкМ - 1 mM полипептидного антагониста. Иначе говоря дозировка в расчете на вес тела может изменяться в интервале от около 0,1 мг/кг до около 300 мг/кг, предпочтительно от около 0,2 мг/кг до около 200 мг/кг при применении одной или более доз ежедневно в течение от одного до нескольких дней.
Моноклональные антитела или полипептиды изобретения могут быть введены парентерально путем инъекции или постепенного вливания в течение времени. Хотя ткань, подлежащая лечению, обычно доступна в организме в результате системного применения и поэтому лечение, как правило, проводят путем внутривенных инъекций терапевтических композиций, подразумеваются и другие ткани и распределяющие устройства в том случае, когда существует вероятность того, что целевая ткань содержит молекулу-мишень. Так например, моноклональные антитела или полипептиды изобретения могут применяться внутривенно, внутрибрюшинно, внутримышечно, подкожно, внутрь полости, трансдермально и могут вводиться с помощью перистальтических средств.
Терапевтические композиции, содержащие моноклональное антитело, или полипептид изобретения обычно применяются внутривенно, например путем инъекции единичной дозировки. Термин "единичная дозировка", в том случае, когда он используется в отношении терапевтической композиции настоящего изобретения, относится к физически дискретным единицам, используемым в качестве единичных дозировок для субъекта, причем каждая единичная дозировка содержит определенное количество активного материала, рассчитанное с целью достижения желаемого терапевтического эффекта совместно с требуемым разбавителем; например носителем или связующим. Согласно одному из предпочтительных решений изобретения, как показано в Примерах, αvβ3-антагонист применяют внутривенно в виде единичной дозы.
Композиции изобретения применяют способом, совместимым с формой дозировки, и в терапевтически эффективном количестве. Количество, подлежащее применению, и длительность применения зависят от объекта применения, емкости системы субъекта, способной утилизировать активный ингредиент, и степени желаемого терапевтического эффекта. Точные количества активного ингредиента, требуемые для применения, зависят от мнения практикующего врача и различны для каждого индивидуума. Однако, подходящие дозировочные интервалы для систематического применения указаны в описании и они зависят от типа применения. Подходящие режимы применения тоже могут изменяться, но как правило, производят начальное применение с последующими повторяющимися дозами с интервалами в один или более часов с последующей инъекцией или другими применениями. Однако с другой стороны предусматриваются также непрерывные внутривенные вливания, достаточные для поддержания концентрации в крови в интервалах, специфичных для in vivo терапии.
Как продемонстрировано настоящими примерами, ингибирование ангиогеназа и регрессия опухоли происходят уже на 7 день после начального контактирования с антагонистом. Дополнительное или пролонгированное воздействие антагониста предпочтительно в период от 7 дней до 6 недель, предпочтительно в период 14-28 дней.
В соответствии с родственным решением приведенные Примеры демонстрируют взаимосвязь между ингибированием αvβ3 и индуцированием апоптоза в неоваскуляторных клетках, несущих αvβ3 . Таким образом, настоящее изобретение также охватывает способы ингибирования апоптоза в неоваскуляторной ткани. На практике осуществлен способ изобретения, включающий ингибирование ангиогенеза во всех тканях и при условиях, описанных в тексте. Заметным различием оказалось лишь начало эффекта, который в случае апоптоза проявляется быстро, обычно через 48 часов после контактирования с антагонистом, тогда как ингибирование ангиогенеза и регрессия опухоли проявляются более медленно, как это было указано выше. Такое отличие оказывает эффект на терапевтический режим в плане времени применения и желаемого эффекта. Обычно, применение в случае апоптоза неоваскулятора может проводиться в течение времени от 24 часов до 4 недель, хотя период от 48 часов до 7 дней является предпочтительным.
D. Терапевтические композиции.
Настоящее изобретения охватывает терапевтические композиции, используемые для практической реализации описанных терапевтических способов. Терапевтические композиции настоящего изобретения содержат физиологически переносимый носитель совместно с αvβ3-анатагонистом, растворенный или диспергированный в нем в качестве активного ингредиента. Согласно предпочтительному воплощению композиция αvβ3-антагониста не является иммуногенной при применении на млекопитающих или людях в терапевтических целях.
Используемые в тексте термины "фармацевтически применимый", "физиологически переносимый" и их грамматические варианты в той мере, в которой они относятся к композициям, носителям, разбавителям и реагентам, являются взаимозаменяемыми и означают, что такие материалы способны применяться на млекопитающих без нежелательных физиологических эффектов, таких как тошнота, головокружение, желудочные расстройства и т.п.
Приготовление фармакологической композиции, содержащей активные ингредиенты, растворенные или диспергированные в ней, хорошо известно из литературы и не ограничивает типы рецептур. Обычно такие композиции готовят в виде растворов для инъекций или жидких растворов, либо суспензий, однако могут также готовиться твердые формы, пригодные для раствора или суспензии непосредственно в жидкости перед применением. Такие препараты могут также представлять собой эмульсии.
Активный ингредиент может смешиваться с эксципиентами, являющимися фармацевтически приемлемыми и совместимыми с активным ингредиентом, в количествах, подходящих для использования в описанных терапевтических методах. Подходящими эксципиентами могут служить, например, вода, физиологический раствор, декстроза, глицерин, этанол и т.п. или их комбинации. Кроме этого, если желательно, композиция может содержать небольшие количества вспомогательных веществ, таких как смачивающие или эмульгирующие агенты, изменяющие pH буфферные агенты и т.п., которые повышают эффективность активного ингредиента.
Терапевтическая композиция настоящего изобретения может включать фармацевтически приемлемые соли компонентов. Фармацевтически приемлемые соли включают соли присоединения кислот /образованные с участием свободных аминогрупп полипептида/, которые готовят с помощью таких неорганических солей, как, например, хлористоводородная или фосфорная кислоты, или таких органических кислот, как уксусная, винная, миндальная и т.п. Соли, приготовленные с участием свободных карбоксильных групп, также могут быть производными таких неорганических оснований, как гидроксиды натрия, калия, аммония, кальция или железа, или таких органических оснований, как изопропиламин, триметиламин, 2-этиламиноэтанол, гистидин, прокаин и т.п.
Особенно предпочтительными при получении циклических полипептидных αvβ3 -антагонистов являются соли TFA и HCl. Представители солей пептидов описаны в Примерах.
Физиологически переносимые носители хорошо известны в данной области. Примерами жидких носителей являются стерильные водные растворы, несодержащие никаких материалов, кроме активных ингредиентов и воды, или содержащие такой буфер, как натрий фосфатный при физиологическом значении pH, физиологический раствор или то и другое, т.е. фосфатно-буфферный физиологический раствор. Кроме этого, водные носители могут содержать более одной буфферной соли или такие соли, как смешанные хлориды натрия и калия, декстрозу, полиэтиленгликоль и другие растворенные вещества.
Жидкие композиции также могут содержать жидкие фазы в присутствии или в отсутствии воды. Примерами таких дополнительных жидких фаз являются глицерин, такие растительные масла, как хлопковое масло и эмульсии типа вода в масле.
Терапевтическая композиция содержит ангиогенез-ингибирующее количество αvβ3-антагониста настоящего изобретения, причем она обычно формируется так, что содержит, по крайней мере, 0,1% вес. антагониста в расчете на общий вес терапевтической композиции. Весовой процент представляет собой весовое соотношение ингибитора к общему весу композиции. Так например, 0,1% вес. представляет собой 0,1 г ингибитора на 100 г всей композиции.
E. Антагонисты Интегрина αvβ3
αvβ3 -антагонисты используются в настоящих методах для ингибирования ангиогенеза в тканях и можно упомянуть большое число форм, включающих соединения, взаимодействующие с αvβ3 таким образом, что они препятствуют функциональному взаимодействию с природными αvβ3-лигандами. Примерами антагонистов могут служить аналоги αvβ3, являющиеся производными сайта связывания лиганда на αvβ3, миметики αvβ3 или природного лиганда αvβ3 , который мимикрирует структурную область, принимающую участие во взаимодействиях связывания αvβ3 -лиганда, полипептиды, имеющие последовательность, соответствующую домену функционального связывания природного лиганда, специфичного к αvβ3, особенно соответствующую RGD-содержащему домену природного лиганда αvβ3, а также антитела, которые иммунореагируют с αvβ3 или с природным лигандом, причем все эти вещества проявляют антагонистическую активность.
1. Полипептиды
В соответствии с одним из технических решений настоящее изобретение охватывает αvβ3-антагонисты в виде полипептидов. Полипептидный /пептидный/ αvβ3-антагонист может иметь характеристики последовательности природного лиганда αvβ3 или самого αvβ3 на участке, принимающем участие во взаимодействии αvβ3 -лиганда и проявляет активность αvβ3 -антагониста. Предпочтительный антагонист -пептид содержит RGD трипептид и соответствует по последовательности природному лиганду в RGD-содержащей области.
Предпочтительные RGD-содержащие полипептиды имеют последовательность, соответствующую последовательности аминокислотных остатков RGD-содержащей области природного лиганда αvβ3, такого как фибриноген, витронектин, фактор фон Виллебранда, ламинин, тромбоспондин и подобные лиганды. Последовательность таких αvβ3 лигандов хорошо известна.
Так например, αvβ3 -антагонист-пептид может быть производным любого из природных лигандов, хотя предпочтительными являются фибриноген и витронектин.
Особенно предпочтительные αvβ3 -антагонисты-пептиды преимущественно ингибируют αvβ3 связывание с их природными лигандами по сравнению с другими интегринами, описанными выше. Эти αvβ3 -специфичные пептиды особенно предпочтительны, по крайней мере, по причине того, что специфичность на αvβ3 уменьшает возможность нежелательных побочных эффектов, таких как ингибирование других интегринов. Идентификация предпочтительных αvβ3 антагонистов-пептидов, обладающих селективностью на αvβ3 может быть легко осуществлена типичным анализом на ингибирование связывания, например, с помощью анализа ELISA, описанного в Примерах.
Согласно одному из воплощений, полипептид настоящего изобретения содержит не более 100 аминокислотных остатков, предпочтительно не более 60 остатков, более предпочтительно не более 30 остатков. Пептиды могут быть линейными или циклическими, хотя особенно предпочтительными являются циклические пептиды.
Предпочтительные циклические и линейные пептиды и их обозначения приведены в Таблице 1 и в Примерах.
Целевой полипептид включает аналог, фрагмент или химическое производное полипептида, чья последовательность остатков аминокислот, представленная в описании свидетельствует о том, что такой полипептид представляет собой αvβ3-антагонист. Поэтому полипептид изобретения может быть объектом различных изменений, замещений, вставок и делеций, если такие изменения обеспечивают некоторые улучшения его применения. В этом отношении следует отметить, что αvβ3-антагонист-полипептид изобретения соответствует, а не идентичен последовательности описанного пептида, в котором произведено одно или более изменений и он сохраняет способность функционировать как αvβ3-антагонист в одном или более анализов, как это показано в тексте.
Таким образом, полипептид может присутствовать в любой из большого числа форм пептидных производных, включающих амиды, конъюгаты с белками, циклические пептиды, полимеризованные пептиды, аналоги, фрагменты, химически модифицированные пептиды, и аналогичные производные.
Термин "аналог" включает любой полипептид, имеющий последовательность аминокислотных остатков, идентичную последовательности, специально показанной там, где один или более остатков консервативно заменены на функционально аналогичный остаток и где проявляется αvβ3-антагонистическая активность. Примеры консервативных замен включают замену одного /неполярного/ гидрофобного остатка, такого как изолейцин, валин, лейцин или метионин на другой, замену одного полярного /гидрофильного/ остатка на другой, как это имеет место при замене аргинина на лизин, глутамина на аспарагин, глицина на серин, замену одного основного остатка, такого как лизин, аргинин или гистидин на другой, или замену одного кислотного остатка, такого как аспарагиновая кислота или глутаминовая кислота на другой.
Фраза "консервативное замещение" также подразумевает использование химически дериватизированных остатков вместо недериватизированного остатка при условии, что такой полипептид обладает достаточной ингибирующей активностью.
Термин "химическое производное" относится к целевому полипептиду, содержащему один или более остатков, химически дериватизированных по реакции функциональных боковых групп. Такие дериватизированные молекулы включают, например, те молекулы, в которых свободные аминогруппы были преобразованы с образованием амингидрохлоридов, n-толуол сульфонильных групп, карбобензокси групп, трет.-бутилоксикарбонильных групп, хлорацетильных групп или формильных групп. Свободные карбоксильные группы могут быть преобразованы с образованием солей, метиловых или этиловых эфиров, эфиров других типов или гидразидов. Свободные гидроксильные группы могут быть преобразованы с образованием О-ацил или О-алкильных производных. Имидазольный азот гистидина может быть дериватизирован с образованием N-имбензилгистидина. Термином "химические производные" охватываются те пептиды, которые содержат одно или более встречающихся в природе аминокислотных производных двадцати стандартных аминокислот. Так например, пролин может быть заменен на 4-гидроксипролин; лизин - на 5-гидроксилизин; гистидин может быть заменен на 3-метилгистидин; серин может быть заменен на гомосерин; а лизин может быть заменен на орнитин.
Полипептиды настоящего изобретения также включают любой полипептид, имеющий одну или более вставок и/или делеций или остатков с родственной полипептиду последовательностью, показанной в тексте при условии, что сохраняется необходимая активность.
Термин "фрагмент" относится к любому целевому полипептиду, имеющему последовательность аминокислотных остатков более короткую, чем у полипептида, аминокислотная последовательность которого показана в тексте.
В том случае, когда полипептид настоящего изобретения имеет последовательность, не идентичную последовательности αvβ3 природного лиганда, это означает, что были произведены одна или более консервативных или неконсервативных замещений, обычно в этом случае заменяют не более 30% и, предпочтительно не более 10% аминокислотных остатков. Дополнительные остатки также могут добавляться к любому окончанию полипептида в целях обеспечения "линкера", с помощью которого полипептиды изобретения могли бы соответствующим образом прикрепляться к метке твердой матрицы или носителя.
Метки, твердые матрицы и носители, которые могут использоваться с полипептидами настоящего изобретения, описаны ниже.
Линкеры аминокислотных остатков обычно представляют собой, по крайней мере, один остаток и могут содержать 40 или более остатков, чаще 1-10 остатков, но они не образуют эпитопов αvβ3-лиганда. Типичными аминокислотными остатками, используемыми для связывания являются тирозин, цистеин, лизин, глутаминовая и аспаргиновая кислоты или другие остатки. Кроме этого, целевой полипептид может отличаться, если не указано особо, от природной последовательности αvβ3 -лиганда последовательностью, которая модифицирована ацилированием терминальной NH2-группы, например, путем ацетилирования или амидированием тиогликолевой кислоты в результате терминального карбоксиламидирования, например, с помощью аммиака, метиламина, и аналогичными терминальными модификациями. Терминальные модификации, как хорошо известно, используются для снижения восприимчивости к протеиназному перевариванию и поэтому они служат для пролонгирования периода полураспада полипептидов в растворе, особенно в биологических жидкостях, где могут присутствовать протеазы. В этом отношении циклизация полипептида также является ценным методом терминальной модификации и она особенно предпочтительна из-за устойчивости структур, полученных в результате циклизации, и из-за биологических активностей, наблюдаемых для таких циклических пептидов, как это отмечается в описании.
Любой пептид настоящего изобретения может использоваться в виде фармацевтически приемлемой соли. Подходящие кислоты, способные образовывать соли пептидов настоящего изобретения, включают такие неорганические кислоты, как трифторуксусную кислоту /TFA/, хлористоводородную кислоту /HCl/, бромистоводородную кислоту, перхлорную кислоту, азотную кислоту, тиоциановую кислоту, серную кислоту, фосфоруксусную кислоту, пропионовую кислоту, гликолевую кислоту, молочную кислоту, пировиноградную кислоту, оксалиновую кислоту, малоновую кислоту, янтарную кислоту, малеиновую кислоту, фумаровую кислоту, антраниловую кислоту, коричную кислоту, нафталинсульфокислоту, сульфанилиновую кислоту и т.п. Особенно предпочтительными являются соли HCl и TFA.
Подходящие основания, способные образовывать соли с пептидами настоящего изобретения, включают такие неорганические основания, как гидроксид натрия, гидроксид аммония, гидроксид калия и т.п.; и такие органические основания, как моно-, ди- и триалкил и ариламины /например, триэтиламин, диизопропиламин, метиламин, диметиламин и т.п./ и, необязательно, замещенные этаноламины /например, этаноламин, диэтаноламин и т.п./.
Пептид настоящего изобретения, на который также ссылаются, как на целевой пептид, может быть синтезирован любым методом, известным специалисту в области полипептидов, например методами рекомбинантной ДНК. Синтетические химические методы, например, твердофазные синтезы типа Меррифилда являются предпочтительными по причинам чистоты продукты, антигенной специфичности, отсутствия нежелательных побочных продуктов, простоты выполнения и т.п. Отличное резюме большинства доступных способов содержится в книге Steward с сотр. "Твердофазные пептидные синтезы", W.H. Freeman, Сан-Франциско, 1969; работе Bodansky с сотр. , "Пептидные синтезы" Джон Вилей энд санс, второе изд. , 1976; J. Meienhofer "Гормональные протеины и пептиды", т. 2, стр. 46, Академик Пресс /Нью-Йорк/, 1983; в статье /Merrifield Adv. Enzymol, 32: 221-96, 1969; Fields с сотр., Int. J.Peptide Protein Res. 35:161-214, 1990 и патенте США N 4.244.946, в том, что касается твердофазных пептидных синтезов, и в книге Schroder с сотр., "Пептиды", т. 1, Академик Пресс /Нью-Йорк, 1965, в том, что касается классических синтезов в растворе, причем на все эти работы ссылаются в данном описании. Подходящие защитные группы, используемые в таких синтезах, описаны в приведенных выше работах и в работе J.F. M. McOmie "Защитные группы в органической химии". Пленум Пресс, Нью-Йорк, 1973, причем на эти работы также ссылаются в настоящем описании.
Как правило используемые методы твердофазного синтеза включают последовательное добавление одного или более аминокислотных остатков или подходящим образом защищенных аминокислотных остатков к растущей пептидной цепи. Обычно, каждая амино или карбоксильная группа первого аминокислотного остатка защищены подходящей, селективно удаляемой защитной группой.
Различные, селективно удаляемые защитные группы используются для аминокислот, содержащих реакционноспособную боковую группу, например для лизина.
С использованием твердофазного синтеза в качестве примера защищенная или дериватизированная аминокислота присоединяется к инертному твердому носителю через незащищенные карбоксильную или аминогруппу. Защитную группу амино- или карбоксильной группы затем селективно удаляют и следующую аминокислоту в последовательности с комплиментарной /амино или карбоксильной/ группой, соответствующим образом защищенной смешивают с системой и проводят реакцию в условиях, подходящих для образования амидной связи с остатком, уже присоединенным к твердой подложке. Затем защитную группу амино или карбоксильной группы удаляют из вновь добавленного аминокислотного остатка и следующую аминокислоту /соответствующим образом защищенную/ добавляют к системе и т.д. После того, как все желаемые аминокислоты связаны в надлежащей последовательности, любые оставшиеся терминальные и побочные группы, а также защитные группы /и твердый носитель/ последовательно или параллельно удаляют с образованием целевого линейного полипептида.
Полученные в результате линейные полипептиды /получены, например, как описано выше/ могут реагировать с образованием соответствующих циклических пептидов. Один из примеров методов циклизации пептидов описан Zimmer с сотр. , Пептиды 1992, стр. 393-394, ESCOM Сайенс Паблишерс, В.У. 1993. Обычно защищенный тетрабутоксикарбонилом пептидный метиловый эфир растворяют в метаноле и добавляют раствор гидроксида натрия и смеси дают реагировать при 20oC с целью гидролитического удаления метилэфирной защитной группы. После выпаривания растворителя тетрабутоксикарбонил-защищенный пептид экстрагируют этилацетатом из подкисленного водного растворителя. Затем тетрабутоксикарбонильную защитную группу удаляют в мягких кислых условиях в диоксановом корастворителе. Полученный таким образом незащищенный линейный пептид со свободными амино- и карбокси-концами превращают в соответствующий циклический пептид в результате реакции разбавленного раствора линейного пептида в смеси дихлорметана и диметилформамида, с дициклогексилкарбодиимидом в присутствии 1-гидроксибензотриазола и N-метилморфолина. Полученный в результате циклический пептид затем очищают хроматографией.
Особенно предпочтительный способ синтеза циклического пептида описан Gurrath с сотр., Eur. J.Biochem. 210:911-921 /1922/ и упомянут в примерах. Особенно предпочтительными пептидами для способов изобретения являются c-/Gr GDFY/ /последовательность SEQ 1D N 4/, с-/RGD fY/ /последовательность SEQ 1D N 5/, с-/RAD f V/ /последовательность SEQ 1D N 6/, с-/RGDFv/ /последовательность SEQ 1D N 7/ и линейный пептид YTAECKPQYTRCDYF /последовательность SЕQ 1D N 8/, где символ "c-" обозначает циклический пептид, заглавные буквы в скобках соответствуют однобуквенному коду для L-аминокислоты, а строчные буквы в скобках соответствуют однобуквенному коду для D-аминокислот. Последовательности аминокислотных остатков таких пептидов также показаны последовательностями SEQ 1D NN 4, 5, 6, 7 и 8 соответственно.
2. Моноклональные антитела
Согласно одному из воплощений, в настоящем изобретении описывается αvβ3-антагонист в виде моноклональных антител, которые иммунореагируют с αvβ3 и ингибируют αvβ3 связывание с его природным лигандом, как это показано в тексте. В настоящем изобретении также описываются линии клеток, продуцирующие такие антитела, способы продуцирования клеточных линий и способы получения моноклональных антител.
Моноклональное антитело настоящего изобретения включает молекулы антитела, которые 1/ иммунореагируют с выделенным αvβ3 и 2/ ингибируют связывание фибриногена с αvβ3. Предпочтительные моноклональные антитела, которые преимущественно связываются с αvβ3, включают моноклональное антитело, обладающее иммунореакционноспособными характеристиками mAB LM609, секретированные гибридомной клеточной линией АТСС HB 9537. Гибридомная клеточная линия АТСС HB 9537 депонирована в соответствии с требованиями Будапештского договора в Американской коллегии типовых культур /АТСС/, 1301 Парклаун Драйв, Рокквилл, МД, США, 15 сентября 1987 г.
Термин "антитело" или "молекула антитела" в различных грамматических формах используется в тексте в качестве коллективного значения, относящегося к популяции иммуноглобулиновых молекул и/или иммунологических активных частей иммуноглобулиновых молекул, т.е. молекул, содержащих сайт, объединяющий антитело или паратоп.
Под термином "сайт объединяющий антитело" подразумевается, что структурная часть молекулы антитела состоит из вариабельных или гипервариабельных участков тяжелых и легких цепей, специфически связывающих антиген.
Примерами антител, используемых в настоящем изобретении, могут служить интактные молекулы иммуноглобулина, частично интактные молекулы иммуноглобулина и те части молекулы иммуноглобулина, которые содержат паратоп, включая части, известные в литературе, как Fab, Fab', f/ab'/ и F/y/ и также относящиеся к фрагментам антитела.
Согласно другому предпочтительному воплощению, настоящее изобретение охватывает укороченную молекулу иммуноглобулина, включающую Fab фрагмент, являющийся производным моноклонального антитела изобретения. Fab фрагмент, в котором отсутствует Fc рецептор, является растворимым и обладает терапевтическими преимуществами в отношении полураспада сыворотки и диагностическими преимуществами в отношении способа применения растворимого Fab фрагмента. Приготовление растворимого Fab фрагмента обычно известно в области иммунологии и может осуществляться большим числом методов.
Так, например, Fab и F/ab'/ части /фрагменты/ антител получают протеолитической реакцией папаина и пепсина соответственно на существенно интактных антителах с помощью способов, хорошо известных из литературы. См. например, патент США N 4342566, выданный на имя Theofilopolus и Dixon. Части Fab' антитела также хорошо известны и могут быть получены из фрагментов F/ab'/2 с последующим восстановлением дисульфидных связей, связывающих участки тяжелых цепей, меркаптоэтанолом, после чего проводят алкилирование полученного в результате протеин-меркаптана таким реагентом, как иодоацетамид. Антитела, содержащие целые иммуноглобулиновые молекулы, являются предпочтительными и их используют, как проиллюстрировано в тексте.
Фраза "моноклональное антитело" в различных грамматических формах относится к популяции молекул антител, которые содержат только одну разновидность сайта, объединяющего антитело, способную иммунореагировать с конкретным эпитопом.
Таким образом, моноклональное антитело обычно проявляет единое связывающее сродство к любому эпитопу, с которым оно иммунореагирует. Поэтому моноклональное антитело может содержать молекулу антитела, имеющую множество антительных комбинирующих сайтов, каждый из которых иммуноспецифичен в отношении различного эпитопа, например биспецифичное моноклональное антитело.
Моноклональное антитело обычно состоит из антител, продуцированных клонами одной клетки, называемой гибридомой, которая секретирует /производит/ лишь один вид молекулы антитела. Гибридомная клетка формируется слиянием антитело-продуцирующей клетки и миеломы или другой бессмертной клеточной линии. Получение таких антител вначале было описано Kohler и Milstein, Nature 256: 495-497 /1975/, причем на эту работу ссылаются в описании. Дополнительные способы описаны Zola, моноклональные антитела: справочник по методам, CRC Пресс, Инк. /1987/. Гибридомный супернатант, полученный таким образом, может быть подвергнут отбору на присутствие молекул антител, иммунореагирующих с αvβ3 и на ингибирование αvβ3 связывания с природными лигандами.
Короче говоря, для получения гибридомы, из которой продуцируют композицию моноклонального антитела, миелому или другую бессмертную клеточную линию сливают с лимфоцитами, полученными из селезенки млекопитающего, гипериммунизированного источника αvβ3 , например αvβ3 выделенным из М21 человеческих мелономных клеток, как это описано Cheresh с сотр., J. Biol. Chem. 262: 17703-17711 /1987/.
Предпочтительно, чтобы миеломная клеточная линия, используемая для получения гибридомы, была той же разновидности, что лимфоциты. Обычно предпочтительным млекопитающим являются мыши штамма 129 GIX*. Подходящие для целей изобретения мышиные миеломы включают гипоксантин-аминоптерин-тимидин-чувствительные /HAT/ клеточные линии P3X63-Ag8.653 и Sp2/0-Ag14, которые получены из Американской коллекции типовых культур, Роквилл, МД под обозначениями CRL 1580 и CRL 1581 соответственно.
Спленоциты обычно сливают с миеломными клетками с использованием полиэтиленгликоля /ПЭГ/ 1500. Слитые гибриды подвергают селекции на их чувствительность к HAT. Гибридомы, производящие моноклональное антитело изобретения, идентифицируют с использованием энзим-связанного иммуносорбентного анализа /ELISA/, описанного в примерах изобретения.
Моноклональное антитело настоящего изобретения также может быть получено инициированием моноклональной гидридомной культуры, включающей питательную среду, содержащую гибридому, секретирующую молекулы антитела соответствующей специфичности. Культуру выдерживают в таких условиях и в течение такого периода времени, которые достаточны для того, чтобы гибридома секретировала молекулы антитела в среду. Затем собирают антитело-содержащую среду. Далее, молекулы антитела могут быть дополнительно выделены хорошо известными способами.
Среды, используемые для получения таких композиций, хорошо известны из литературы, выпускаются промышленностью и включают синтетические культурные среды инбредных мышей и т.п. Примерами синтетических сред являются минимально необходимая среда Дулбекко /ДМЕМ; Dulbecco с сотр., Virol, 8:396, 1959/, дополненная 4,5 мг/л глюкозы, 20 мМ глутамина, и 20% сыворотки телячьего плода. Примером инбредного мышиного штамма может служить Ва1b/с.
Другие способы продуцирования моноклонального антитела, гибридомной клетки или гибридомной клеточной культуры также хорошо известны. См., например, способ выделения моноклональных антител из иммунологического репертуара, описанный Sastry с сотр. , Proc. Natl. Acad. Sci. USA 86:5728-5732 /1989/; и Huse. с сотр. Science, 246:1275-1281 /1989/.
Настоящее изобретение также охватывает гибридомную клетку и культуры, содержащие гибридомную клетку, вырабатывающую моноклональное тело настоящего изобретения. Особенно предпочтительной является гибридомная клеточная линия, секретирующая моноклональное антитело mAb LM609, обозначенное как АТСС HB 9537. mAb LM609 было получено как описано Cheresh с сотр., J, Biol. Chem. 262: 17703-17711 /1987/ и это получение также отражено в Примерах изобретения.
Согласно одному из воплощений, настоящее изобретение охватывает моноклональное антитело, имеющее иммунореакционные характеристики mAb LM609.
Можно также определить без ненужного экспериментирования обладает ли моноклональное антитело такой же /т.е. эквивалентной/ специфичностью /иммунореакционными характеристиками/, как моноклональное антитело изобретения путем выяснения того факта, имеет ли место предотвращение первым объектом последнего объекта от связывания с предварительно выбранной молекулой-мишенью. Если испытуемое моноклональное антитело конкурирует с моноклональным антителом изобретения, о чем может свидетельствовать уменьшение степени связывания моноклональным антителом изобретения в стандартных условиях сравнительного анализа на связывание с мишенью-молекулой при реализации твердофазного варианта, то существует вероятность того, что два этих моноклональных антитела связаны с одним или близко родственным ему эпитопом.
Еще один путь определения того, обладает ли моноклональное антитело специфичностью моноклонального антитела изобретения, состоит в предварительной инкубации моноклонального антитела изобретения с молекулой-мишенью, с которой оно реагирует в обычных условиях с последующим добавлением испытуемого моноклонального антитела для определения факта ингибирования способности испытуемого антитела связывания молекулы-мишени. Если такое ингибирование имеет место, то по всей вероятности оно идентично, или функционально эквивалентно эпитопной специфичности моноклонального антитела изобретения.
Еще одним путем определения одинаковой специфичности моноклонального антитела и моноклонального антитела изобретения является определение последовательности аминокислотных остатков CDR-участков сравниваемых антител. Молекулы антитела, обладающие идентичной функционально-эквивалентной последовательностью аминокислотных остатков в их CDR областях, обладают одинаковой связующей специфичностью. Способы установления последовательностей полипептидов хорошо известны из литературы.
Иммуноспецифичность антитела, его связующая емкость в отношении молекулы-мишени, аффинность антитела в отношении эпитопа определяются эпитопом, с которым иммунореагирует антитело. Эпитопная специфичность определяется, по крайней мере, частично последовательностью аминокислотных остатков вариабельного участка тяжелой цепи иммуноглобулинового антитела и частично вариабельным участком легкой цепи последовательности аминокислотных остатков.
Использование термина "обладающий специфичностью связывания" указывает на то, что эквивалентные моноклональные антитела обладают одинаковыми или похожими иммунореакционными /связующими/ характеристиками и конкурируют за связывание с предварительно отобранной молекулой мишенью.
Гуманизированные моноклональные антитела предполагают особые преимущества над мышиными моноклональными антителами особенно в том плане, что они могут терапевтически применяться на людях. Говоря конкретно, человеческие антитела не так быстро подвергаются циркуляции, как "инородные" антигены и не активируют иммунную систему тем же образом, что инородные антигены и инородные антитела. Способы получения "гуманизированных" антител обычно хорошо известных из литературы и могут быть легко применимы к антителам настоящим изобретением.
Таким образом, изобретение предусматривает, согласно одному из воплощений, моноклональное антитело, которое гуманизируется трансплантацией с целью введения компонентов человеческой иммунной системы без существенных препятствий в отношении способности антитела связывать антиген.
3. αvβ3-Специфичные миметики.
Настоящее изобретение демонстрирует тот факт, что αvβ3 -антагонисты обычно могут использоваться в изобретении, причем такие антагонисты могут включать полипептиды, антитела и другие молекулы, обозначенные как "миметики", которые обладают способностью препятствовать αvβ3-функции. Особенно предпочтительными являются антагонисты, которые специфически препятствуют αvβ3 функционированию и не являются помехой функции других интегринов.
В этом контексте следует отметить, что большое количество реагентов может подходить для использования в способах изобретения при условии, что такие реагенты обладают требуемой биологической активностью. На такие реагенты ссылаются как на миметики, поскольку они обладают способностью "подражать" связыванию домена на αvβ3 или αvβ3-лиганде, принимающем участие в функциональном взаимодействии рецептора и лиганда и вследствие этого препятствовать /например, ингибировать/ нормальное функционирование.
αvβ3-миметик представляет собой любую молекулу, отличную от антитела или лиганд-производного пептида, которая обладает описанными выше свойствами. Это может быть синтетический аналог пептида, соединение, имеющее форму связующего кармана, указанного выше домена, или другая молекула.
Создание αvβ3-миметиков может осуществляться любым из большого числа структурно-аналитических методов создания лекарственных препаратов, известных из литературы, включающих молекулярное моделирование, двумерный ядерный магнитный резонанс /2-Д ЯМР/, рентгеновскую кристаллографию, случайный скрининг пептида, пептидного аналога или других химических полимерных библиотек и другие методы создания лекарств.
На основании широких структурных данных, представленных в настоящем описании, которое показывает, что αvβ3 -антагонист может быть небольшим полипептидом или моноклональным антителом, был сделан вывод о наличии двух взаимно различных химических структурах, которые разделяют функциональные свойства селективного ингибирования αvβ3, причем структура αvβ3 -антагониста, используемого в настоящем способе, не требует такого ограничения, а включает любой αvβ3 миметик, упомянутый в описании.
F. Способы идентификации антагонистов αvβ3
В изобретении также описываются аналитические способы идентификации кандидатов в αvβ3-антагонисты, предназначенные для использования в методах изобретения. В таких способах анализа молекулы-кандидаты оцениваются на их мощность в ингибировании αvβ3 связывания с нейтральными лигандами и, кроме этого, оцениваются на мощность в ингибировании ангиогенеза в тканях.
В первом анализе измеряется ингибирование прямого связывания природного лиганда с αvβ3 , а предпочтительное техническое решение подробно описано в Примерах.
В анализе обычно измеряется степень ингибирования связывания природного лиганда, например, фибриногена с изолированным αvβ3 в твердой фазе методом ELISA.
Такой анализ может также использоваться для идентификации соединений, обладающих специфичностью на αvβ3 и не ингибирующих природные лиганды от связывания с другими интегринами. Специфичность анализа проверяется параллельным проведением анализов ELISA, когда оба αvβ3 и другие интегрины отбираются конкурентно в отдельные аналитические камеры с целью определения соответствующих способностей к связыванию с природным лигандом, а соединение-кандидат оценивается на способность ингибировать соответствующие способности интегринов к соединению с предварительно отобранным лигандом. Предпочтительные форматы скринирующего анализа описаны в Примерах.
Согласно второму методу анализа измеряется ангиогенез в куриной хориоаллантоисной мембране /CAM/ и на этот анализ ссылаются как на анализ CAM. CAM анализ был описан подробно в других источниках и позже этот анализ был использован для измерения ангиогенеза и неоваскуляризации опухолевых тканей. Cм. Ausprunk с сотр. Am.J. Pathol. 79:597-618 /1975/ и Ossonski с сотр., Canser Res, 40:2300-2309 /1980/.
Анализ CAM является хорошо известным методом анализа для in vivo ангиогенеза, поскольку неоваскуляризация ткани имеет место в этом случае и действительные кровеносные сосуды куриного эмбриона прорастают в CAM либо в ткань, растущую на CAM.
Как продемонстрировано в описании, CAM - анализ иллюстрирует ингибирование неоваскуляризации, основанной на количестве и степени роста новых сосудов. Кроме этого, легко следить за ростом любой ткани, трансплантированной на CAM, например опухолевой ткани. Наконец, такой анализ особенно ценен, поскольку осуществляется внутренний контроль токсичности анализируемой системы. Куриный эмбрион подвергается воздействию любого испытуемого реагента и поэтому состояние здоровья эмбриона является указанием токсичности.
Третий анализ измерения ангиогенеза представляет собой in vivo модель на глазах кроликов и на него ссылаются, как на анализ на глазах кроликов. Такой анализ на глазах кролика подробно описан в других источниках и он дополнительно используется для измерения как ангиогенеза, так и неоваскуляризации в присутствии таких ангиогенных ингибиторов, как талидомид. См. D'Amato с сотр. Proc. Natl. Acad. Sci., 91:4082-4085 /1994/.
Кролично-глазной анализ представляет собой хорошо известную аналитическую модель in vivo ангиогенеза, поскольку процесс неоваскуляризации, примером которого является рост кровеносных сосудов кролика от края роговой оболочки глаза в роговую оболочку, легко визуализируется через обычно прозрачную роговую оболочку глаза.
Кроме этого, степень и количество стимуляции или ингибирования неоваскуляризации или регрессии неоваскуляризации легко может регистрироваться во времени.
Кроме этого, кролик может быть подвержен действию любого испытуемого реагента, и поэтому состояние здоровья кролика является свидетельством токсичности испытуемого реагента.
В четвертом анализе измеряется ангиогенез в химерной модели мышь : человек и на этот анализ ссылаются, как на химерный мышиный анализ. Этот анализ подробно описан в других источниках и этот анализ дополнительно описывается в тексте, как средство для измерения ангиогенеза, неоваскуляризации и регрессии опухолевой ткани. См. Yan с сотр. J. Clin. Intest.91:986-996 /1993/. Химерный мышиный анализ является ценной аналитической моделью для in vivo ангиогенеза, поскольку трансплантированная кожа, плотно привитая в ходе операции, напоминает нормальную человеческую кожу в гистологическом отношении, и неоваскуляризация ткани, происходящая в системе, включает проращивание действительных человеческих кровеносных сосудов от привитой человеческой кожи в человеческую опухолевую ткань на поверхности привитой человеческой кожи. Происхождение неоваскуляризации в человеческую прививку может быть продемонстрировано иммуногистохимическим окрашиванием неоваскулятуры человеко-специфичными эндотелиальными клеточными маркерами.
Как продемонстрировано в тексте, химерный мышиный анализ демонстрирует регрессию неоваскуляризации на основе количества и степени регрессии роста новых сосудов. Кроме этого, легко осуществлять наблюдение за эффектами в ходе роста любой ткани, трансплантированной на привитую кожу, например опухолевой ткани. Наконец, такой анализ является ценным средством, поскольку позволяет осуществлять внешний контроль токсичности аналитической системы. Химерная мышь подвергается любому испытуемому реагенту и поэтому состояние ее здоровья является индикатором токсичности.
Примеры
Следующие ниже примеры, относящиеся к изобретению носят иллюстративный характер и, разумеется, не ограничивают изобретение. Кроме этого, варианты изобретения, известные в настоящее время или те, что будут разработаны позже в рамках компетенции специалиста в данной области, рассматриваются как входящие в сферу настоящего изобретения.
1. Получение синтетических пептидов.
Линейные и циклические полипептиды, перечисленные в Таблице 1, синтезировали с использованием стандартных методов твердофазного синтеза, как это, например, описано Mirrifield Adv. Enzymol. 32:221-96, /1969/ и Pields G.B. и Noble R.Z. Int. J. Peptido Protein Res. 35:161-214 /1990/.
Два грамма /г/ BOG-Cly-D-Arg-Gly-Asp-Phe-Val-OMe /последовательность SEQ 1D N 1/ вначале растворяли в 60 миллилитрах /мл/ метанола, к которому добавляли 1,5 мл 2N раствор гидроксида натрия с образованием смеси. Такую смесь затем перемешивали в течение 3 часов при 20oC. После выпаривания остаток переносили в воду, подкисляли до pH 3 разбавленной HCl и экстрагировали этилацетатом. Экстракт сушили над Na2SO4, снова выпаривали и полученную в результате последовательность BOC-Gly-D-Arg-Gly-Asр-Phe-Val-OH /SEQ 1D N 2/ перемешивали при 20oC в течение 2 часов в присутствии 20 мл 2N HCl в диоксане. Полученную смесь выпаривали с получением последовательности H-Gly-D-Arg-Gly-Asp-Phe-Val-OH /SEQ 1D N3/, которую далее растворяли в смеси 1800 мл дихлорметана и 200 мл диметилформамида /DMF/ с последующим охлаждением до 0oC. После этого, последовательно при перемешивании добавляли 0,5 г дициклогексилкарбодиимида /DCCI/, 0,3 г 1-гидроксибензотриазола /HOBt/ и 0,23 мл N-метилморфолина.
Полученную в результате смесь перемешивали в течение 24 часов при 0oC и затем при 20oC в течение еще 48 часов. Раствор концентрировали и обрабатывали смешанным слоем ионообменника для освобождения от солей. После удаления полученной в результате смолы фильтрацией, осветленный раствор выпаривали и остаток очищали методом хроматографии, получая цикло/Gly-D-Arg-Gly-Asp-Phe-Val /последовательность /SEQ 1D N 4/. Следующие пептиды, перечисленные в Таблице 1 с использованием однобуквенного кода сокращений аминокислотных остатков и идентифицированные обозначениями номера пептида, получали аналогичным образом: цикло/Arg-Gly-Asp-D-Phe-Val/ /SEQ 1D N 5/; цикло/Arg-Ala-Asp-D-Phe-Val/ /SEQ 1D N6/; цикло/Arg-D-Ala-Asp-Phe-Val/ /SEQ 1D N9 /, цикло/Arg-Gly-Asp-Phe-D-Val/ /SEQ 1D N 7/. Пептид, обозначенный как 66203, имеющий последовательность идентичную последовательности пептида 62184, отличается от последнего лишь тем, что содержит соль HCl, а не соль TFA, присутствующую в 62184. В анализе по ингибированию ангиогенеза в соответствии с примером 7, в котором использовали синтетические пептиды, 66203 пептид, содержащий HCl, оказался несколько более эффективным в ингибировании ангиогенеза, чем идентичный пептид в TFA.
Пептид, обозначенный номером 69601, имеющий идентичную последовательность последовательности пептида 62185, отличается от последнего тем, что содержит соль HCl, а не соль TFA, присутствующую в 62184.
Циклический пептид с-RADfv /69601/, как было показано, ингибирует связывание фибриногена с интегрином αvβ3 и не ингибирует связывание фибриногена с интегринами 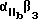 или α5β1, / Pfaff с
или α5β1, / Pfaff с
сотр. J. Biol. Chem. 269:20233-20238 /1994/. Таким образом, пептид c-RADFV специфичен на αvβ3.
2. Моноклональные антитела
Моноклональное антитело LM609, секретированное гибридомой АТСС HB 9537, было получено с использованием стандартных гибридомных методов в результате иммунизации выделенной αvβ3 адсорбированного на Сефарозалентиллектиновых шариках. αvβ3 Выделяли из человеческих меланомных клеток, обозначены как М21, и антитело получали в соответствии со способом Cheresh с сотр. J. Biol. Chem. 262: 17703-17711 /1987/. М21 клетки были получены д-ром D.L. Morton /Калифорнийский университет Лос-Анжелеса, Ка./ и выращены в суспензионной культуре в RPMI 1640 культуральной среде, содержащей 2 mM L-глутамина, 50 мг/мл гентамицинсульфата и 10% фетальной телячьей сыворотки.
Было показано, что моноклональное антитело LM609 специфически иммунореагирует с αvβ3 комплексом и не иммунореагирует с αv субъединицей, с β3 субъединицей или с другими интегринами.
3. Характеристика тканевого распределения αvβ3 экспрессии.
А. Иммунофлуоресцения с анти-интегрин рецепторными антителами.
В ходе заживления ран основные мембраны кровеносных сосудов экспрессируют несколько адгезивных протеинов, включающих фактор фон Виллебранда, фибронектин и фибрин.
Кроме этого, некоторые члены интегринового семейства адгезионных рецепторов экспрессируются на поверхности культивированных гладких мышц и эндотелиальных клеток. см. Cheresh Proc. Natl. Acad. Sci. USA 84:6471 /1987/; Janat с сотр. J. Cell Physiol, 151:588 /1992/; а также Cheng J. Cell Physiol. 139: 275 /1989/. Среди этих интегринов присутствуют αvβ3 эндотелиальный клеточный рецептор фактора фон Виллебранда, фибриноген /фибрин/ и фибронектин согласно Cheresh Proc. Natl. Acad. Sci. USA 84:6471 /1987/. Эти интегрины инициируют кальций-зависимый сигнальный путь, ведущий к миграции эндотелиальных клеток, и поэтому по-видимому они играют фундаментальную роль в биологии сосудисных клеток в соответствии с Zeavelsey с сотр., J. Cell Biol. 121:163 /1993/.
Для исследования экспрессии αvβ3 в ходе ангиогенеза человеческую раневую грануляционную ткань или соседнюю нормальную кожу получали от пациентов-добровольцев, промывали 1 мл фосфатного буфферного раствора и вводили в О.Т. С. среду /Ткань Tek/. Внедренные ткани замораживали в жидком азоте на 30-45 секунд. Секции толщиной шесть микрон вырезали из замороженных блоков на криостатном микротоме для последующего иммунопероксидазного окрашивания с антителами, специфичными к β3 интегринам /αvβ3 или 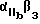 / или β1 подсемейству интегринов.
/ или β1 подсемейству интегринов.
Результаты окрашивания нормальной человеческой кожи и раневой грануляционной ткани представлены на фиг. 1A-1Г. Моноклональные антитела AP3 и LM534, направленные к β3 и β1 интегринам, соответственно, использовали для иммуногистохимического анализа замороженных секций. Эксперименты с тканью от четырех различных человеческих доноров дали идентичные результаты. Фотомикрографии приведены при увеличении в 300 раз.
αvβ3 интегрин обильно экспрессировался на кровеносных сосудах в грануляционной ткани /фиг. 1Б/, но не детектировался в дермисе и эпителии нормальной кожи того же донора /фиг. 1A/. В отличие от этого αvβ3 интегрины обильно экспрессируются на кровеносных сосудах и стромальных клетках как на нормальной коже /фиг. 1В/, так и на грануляционной ткани /фиг. 1Г/ и, как предварительно показано и описано Adams с сотр., Cell, 63:425 /1991/, на базальных клетках внутри эпителия.
B. Иммунофлуоресценция в присутствии анти-лигандных антител
Дополнительные участки человеческой нормальной кожи и грануляционных тканей, полученных выше, подвергали также исследованию на присутствие лигандов β3 и β1 интегринов, фактора фон Виллебранда и ламинина, соответственно. Фактор фон Виллебранда оказался локализованным на кровеносных сосудах нормальной кожи /фиг. 2A/ и грануляционной кожи /фиг. 2Б/, тогда как ламинин был локализован на всех кровеносных сосудах, а также эпителиальной основной мембраны в обоих препаратах ткани /фиг. 2В и 2Г/.
C. Распределение анти αvβ3 антител на раковых тканях.
Помимо проведенных выше анализов биопсии раковой ткани человека также исследовали на присутствие и распределение αvβ3 . Ткани получали, как описано Примере 1A за исключением того, что их окрашивали моноклональным телом LM609, полученным в Примере 2, специфичным на интегриновый рецепторный комплекс αvβ3 . Кроме этого также получали опухоли для микроскопического гистологического анализа путем фиксации представителей опухолей в Bulins Fixative в течение 8 часов и серийные секции вырезали и окрашивали H & E.
Результаты иммунопероксидазного окрашивания пузырьков, а также раковых тканей толстой кишки, грудной железы и легких представлены на фиг. 3A-3Г, соответственно. αvβ3 обильно экспрессировались лишь на кровеносных сосудах, присутствующих в четырех раковых биопсиях, подвергнутых анализу, и ни на каких-либо других клетках, присутствующих в ткани.
Таким образом, описанные результаты показывают, что αvβ3 интегриновый рецептор селективно экспрессируется на тканях специфических типов, а именно грануляционных, метастатических тканях и других тканях, в которых имеет место ангиогенез, и в нормальных тканях, в которых образование новых кровеносных сосудов прекращено. Поэтому такие ткани представляют собой идеальную мишень для терапевтических аспектов изобретения.
4. Идентификация αvβ3 -специфических синтетических пептидов, детектируемых лиганд-рецепторным связыванием.
Синтетические пептиды, полученные в Примере 1, подвергали скринингу, измеряя их способность антагонизировать αvβ3 и 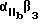 рецепторную связующую активность в анализах на связывание между очищенным лигандом и рецептором. Метод такого изучения связывания описан Barbas с сотр., в Proc. Natl. Acad. Sci. , USA 90:10003-10007 /1993/., Smith с сотр. J.Biol, Chem. 265:11008-11013 /1990/ и Pfaff с сотр., J. Biol. Chem. 269:20233-20238 /1994/, причем на содержание этих работ ссылаются в настоящем изобретении.
рецепторную связующую активность в анализах на связывание между очищенным лигандом и рецептором. Метод такого изучения связывания описан Barbas с сотр., в Proc. Natl. Acad. Sci. , USA 90:10003-10007 /1993/., Smith с сотр. J.Biol, Chem. 265:11008-11013 /1990/ и Pfaff с сотр., J. Biol. Chem. 269:20233-20238 /1994/, причем на содержание этих работ ссылаются в настоящем изобретении.
В настоящем описании описывается способ идентификации антагонистов в анализе лиганд-рецепторного связывания, в котором рецептор иммобилизован на твердой подложке, а лиганд и антагонист находятся в растворенном состоянии. Описывается также анализ на лиганд-рецепторное связывание, в котором лиганд иммобилизован на твердой подложке, а рецептор и антагонисты находятся в растворенном состоянии.
Вкратце, отобранные очищенные интегрины по отдельности иммобилизуют в лунках для микротитрования Titertek, при концентрации покрытия 50 нанограммов /ηг/ на ампулу. Метод очистки рецепторов, используемых в анализах на лиганд-рецепторное связывание, хорошо известен из литературы и может быть легко осуществлен специалистами в данной области. После инкубирования в течение 18 часов при 4oC неспецифичные связующие сайты на пластине блокируются 10 мг/мл альбумина бычьей сыворотки /BSA/ в Трис-буфферном растворе. Для изучения ингибирования различные концентрации пептидов, указанных в Таблице 1, испытывали на способность блокировать связывание 125I-витронектина или 125I-фибриногена с интегриновыми рецепторами αvβ3 и 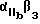 . Хотя такие лиганды демонстрируют оптимальное связывание с конкретным интегрином, витронектином в случае αvβ3 и фибриногеном в случае
. Хотя такие лиганды демонстрируют оптимальное связывание с конкретным интегрином, витронектином в случае αvβ3 и фибриногеном в случае 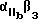 , исследования ингибирования связывания с использованием пептидов с целью блокировки связывания фибриногена с любым рецептором позволяют точно определить количество микромолей пептида, необходимое для половинного ингибирования связывания рецептора с лигандом. Радиомеченные лиганды использовали в концентрациях порядка 1 нМ и связывание вызывалось отдельно немеченными синтетическими пептидами.
, исследования ингибирования связывания с использованием пептидов с целью блокировки связывания фибриногена с любым рецептором позволяют точно определить количество микромолей пептида, необходимое для половинного ингибирования связывания рецептора с лигандом. Радиомеченные лиганды использовали в концентрациях порядка 1 нМ и связывание вызывалось отдельно немеченными синтетическими пептидами.
Через три часа инкубации свободный лиганд удаляли промывкой, а связанный лиганд детектировали гамма подсчетом. Данные анализов, в которых отобранные циклические пептиды, перечисленные в Таблице 1, использовали для ингибирования связывания рецепторов и радиомеченного фибриногена с отдельно иммобилизованными αvβ3 и 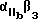 рецепторами были высоко воспроизводимыми при ошибке между полученными значениями обычно ниже 11%. Данные IC50 в микромолях /IC50 мкМ/ выражали как среднее значение из двух измерений ± стандартное отклонение, как это показано в Таблице 2.
рецепторами были высоко воспроизводимыми при ошибке между полученными значениями обычно ниже 11%. Данные IC50 в микромолях /IC50 мкМ/ выражали как среднее значение из двух измерений ± стандартное отклонение, как это показано в Таблице 2.
Таким образом, RGD-содержащие или RGD-дереватизированные циклические пептиды 62181, 62184, 62185 и 62187, каждый из которых содержит один D-аминокислотный остаток, демонстрируют преимущественное ингибирование фибриногенного связывания с αvβ3 рецептором, как измерено низкой концентрацией пептида, требуемой для полу максимального ингибирования, по сравнению с этой величиной для 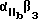 рецептора. В отличие от этого RGD-содержащие или RGD-дериватизированные пептиды 62186, 62175 и 62179 либо не столь эффективны в блокировании фибриногенного связывания с αvβ3 или демонстрируют преимущественное ингибирование фибриногенного связывания с
рецептора. В отличие от этого RGD-содержащие или RGD-дериватизированные пептиды 62186, 62175 и 62179 либо не столь эффективны в блокировании фибриногенного связывания с αvβ3 или демонстрируют преимущественное ингибирование фибриногенного связывания с  по сравнению c αvβ3 . Эти результаты находятся в соответствии с недавно опубликованными Pfaff с сотр. J. Biol. Chem, 2690233-20238 /1994/, в которых циклический пептид
по сравнению c αvβ3 . Эти результаты находятся в соответствии с недавно опубликованными Pfaff с сотр. J. Biol. Chem, 2690233-20238 /1994/, в которых циклический пептид 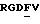 / где
/ где 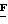 обозначает D-аминокислотный остаток/ специфически ингибировали связывание фибриногена с интегрином αvβ3 , а не с
обозначает D-аминокислотный остаток/ специфически ингибировали связывание фибриногена с интегрином αvβ3 , а не с 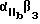 или α5β1 интегринами. Аналогичные анализы по ингибированию связывания проводились с линеаризованными пептидами, имеющими или не имеющими RGD мотив, последовательности которых являются производными αv рецепторной субъединицы,
или α5β1 интегринами. Аналогичные анализы по ингибированию связывания проводились с линеаризованными пептидами, имеющими или не имеющими RGD мотив, последовательности которых являются производными αv рецепторной субъединицы, 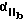 рецепторной субъединицы или последовательностей аминокислотных остатков витронектинового лиганда. Последовательности линейных пептидов 62880 /VN-производные аминокислотные остатки 35-49/, 62411 / αv -производные аминокислотные остатки 676-687/; 62503 / αv -производные аминокислотные остатки 655-667/ и 62502 / αv -производные аминокислотные остатки 296-306/ перечислены в Таблице 1. Каждый из таких пептидов использовали в отдельных анализах для ингибирования связывания либо витронектина /VN/ или фибриногена /FG/ с
рецепторной субъединицы или последовательностей аминокислотных остатков витронектинового лиганда. Последовательности линейных пептидов 62880 /VN-производные аминокислотные остатки 35-49/, 62411 / αv -производные аминокислотные остатки 676-687/; 62503 / αv -производные аминокислотные остатки 655-667/ и 62502 / αv -производные аминокислотные остатки 296-306/ перечислены в Таблице 1. Каждый из таких пептидов использовали в отдельных анализах для ингибирования связывания либо витронектина /VN/ или фибриногена /FG/ с 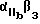 или αvβ3 . Значения IC50, выраженные в микромолях /IC50 мкМ/, индивидуального анализа каждого эксперимента представлены в Таблице 3.
или αvβ3 . Значения IC50, выраженные в микромолях /IC50 мкМ/, индивидуального анализа каждого эксперимента представлены в Таблице 3.
Результаты анализов по ингибированию лигандного связывания выбранных интегриновых рецепторов с линеаризованными пептидами показывают, что лишь пептид 62880 эффективен при ингибировании полу-максимального связывания FG или VN с αvβ3, что измерено низкой концентрацией пептида, требуемого для полу-максимального ингибирования по сравнению c 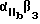 рецептором. Никакой другой из линеаризованных пептидов не был эффективен в блокировке лигандного связывания с αvβ3, хотя пептид 62502 оказался эффективным в блокировке VN связывания с
рецептором. Никакой другой из линеаризованных пептидов не был эффективен в блокировке лигандного связывания с αvβ3, хотя пептид 62502 оказался эффективным в блокировке VN связывания с 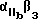 .
.
Таким образом, описанный в тексте лиганд-рецепторный анализ может использоваться для скрининга как кольцевых, так и линеаризованных синтетических пептидов, проявляющих селективную специфичность в отношении конкретного интегринового рецептора, особенно αvβ3, при их использовании в качестве витронектиновых рецепторных / αvβ3 / антагонистов при практической реализации изобретения.
5. Характеристики необработанной хориоаллантоисной мембраны цыпленка /CAM/
A. Получение CAM
Ангиогенез может индуцироваться на хориоаллантоисной мембране цыпленка /CAM/ после нормального эмбрионального ангиогенеза, приводящего к образованию зрелых кровеносных сосудов. Было показано, что ангиогенез индуцируется в ответ на специфический цитокинез фрагментов опухоли, как описано Zeibovich с сотр. Nature, 329:630 /1987/ и Ausprunk с сотр. Am. J.Pathol. 79:597 /1975/. CAM получали из цыплячьих эмбрионов с целью последующего индуцирования ангиогенеза и его ингибирования, как описано в примерах 6 и 7. Десятидневные куриные эмбрионы получали с птицеводческой фермы МакИнтайр /Лэйксайд, КА/ и инкубировали при 37oC при 60% влажности. Небольшое отверстие проделывали в оболочке на конце яйца непосредственно над воздушным мешочком с использованием небольшой дрели /Дремел, Отделение Эмерсон электрик Ко., Расайн WI/. Второе отверстие просверливали на широкой стороне яйца в области, лишенной эмбрионных кровеносных сосудов, что было установлено предварительной проверкой яйца на свет. К исходному отверстию прилагали отрицательное давление, что приводило к вытягиванию CAM /хориоаллантоисная мембрана/ из мембраны оболочки и созданию ложного воздушного мешочка над CAM. Квадратное окно размером 1,0х1,0 см вырезали в оболочке над CAM с использованием размалывающего колеса небольшой модели /Дремел/. Небольшое окошко позволяло осуществлять непосредственный доступ к находящемуся внизу CAM.
Полученный в результате препарат CAM использовали либо через 6 дней эмбриогенеза, стадии отличающейся активной неоваскуляризацией без дополнительной обработки с образованием CAM, отражающей модель, используемую для оценки эффектов эмбрионной неоваскуляризации, или использовали через 10 дней эмбриогенезиса, когда ангиогенез начинал убывать. Последний препарат таким образом использовали в изобретении для индуцирования нового ангиогенеза в ответ на цитокиновую обработку или контакт с опухолью, как это описано в Примере 6.
B. Гистология CAM.
Для анализа микроскопической структуры CAM куриных эмбрионов и/или человеческих опухолей, которые подвергали резекции из куриного эмбриона, как описано в примере 8, CAM и опухоли получали для изготовления срезов в замороженном состоянии, как описано в Примере 3A. Шестимикроновые срезы /по толщине/ вырезали из замороженных блоков на криостатном микротоме для иммунофлуоресцентного анализа.
На фиг. 4 представлена типичная фотомикрография площади, лишенной кровеносных сосудов, в необработанном CAM через 10 дней. Ангиогенез в CAM-системе уменьшался на этой стадии эмбриогенеза, систему использовали в изобретении для стимуляции продуцирования новой сосудистой системы из существующих сосудов от соседних участков в участки, где CAM лишен каких-либо сосудов.
C. Профили интегринов в CAM, детектируемой методом иммунофлуоресценции.
Для определения распределения ткани интегриновых рецепторов, присутствующих в CAM тканях, 6-микронные замороженные срезы опухолевой ткани и тканей CAM куриного эмбриона фиксировали в течение 30 секунд в ацетоне и окрашивали методом иммунофлуоресценции 10 микрограмм/миллилитр /мкг/мл/ mAB CSAT, моноклональным антителом, специфичным к β1 интегриновой субъединице, в соответствии с описанным Buck с сотр., J. Chell. Biol., 107:2351 /1988/ и таким образом использовали для контрольных образцов или LM609 в соответствии с методикой примера 2. Первичное окрашивание сопровождалось окрашиванием 1: 250 разбавлением меченного вторичного антитела козьего анти-мышиного родамина /Tago/ с целью детекции первичного иммунореактивного продукта. Затем такие срезы анализировали на микроскопе Цейса в присутствии иммунофлуоресцирующего соединения.
Результаты иммунофлуоресцентного анализа показывают, что зрелые кровеносные сосуды присутствуют в необработанном 10-дневном курином эмбрионе, экспрессирующем субъединицу интегрина β1 /фиг. 5A/. В отличие от этого, в серийном срезе ткани, показанном на фиг. 5A, не обнаружено иммунореактивности в присутствии LM609 /фиг. 5Б/. Таким образом, интегрин αvβ3 детектируемый LM609 антителом активно не экспрессируется зрелыми кровеносными сосудами, присутствующими в 10-дневном необработанном курином эмбрионе. Как показано в модели CAM и следующих примерах, хотя кровеносные сосуды подвергаются новому росту в ходе нормального эмбриогенеза или индуцируются цитокинами или опухолью, кровеносные сосуды экспрессируют αvβ3 . Однако после активного процесса неоваскуляризации, развитие сосудов прекращается и экспрессия αvβ3 уменьшается до уровней, не детектируемых иммунофлуоресцентным анализом. Такая регуляция αvβ3 экспрессии в кровеносных сосудах, подвергаемых ангиогенезу, контрастирующая с отсутствием экспрессии в зрелых сосудах, обеспечивает уникальную способность настоящего изобретения контролировать и ингибировать ангиогенез, как это показано в следующих ниже примерах в качестве модели, использующей систему анализа CAM-ангиогенеза.
6. Анализ CAM ангиогенеза.
А. Ангиогенез, индуцированный факторами роста
Было показано, что ангиогенез может индуцироваться цитокинами или факторами роста, на что ссылаются в Примере 5A. В описанных экспериментах ангиогенез в препарате CAM, описанном в Примере 5, индуцировали факторами роста, которые применяли местно на кровеносных сосудах CAM.
Ангиогенез индуцировали путем помещения фильтровального диска Ватмана /размером 5 х 5 см/ Ватман, фильтровальная бумага N 1/, насыщенного сбалансированным солевым раствором Ханкса /HBSS GIBCO Гранд айлэнд, Нью-Йорк/ или HBSS, содержащим 150 нанограммов/ миллилитр /нг/мг/ рекомбинантного основного фибробластного фактора роста / βFGF/ /Гензим, Кэмбридж, МА/ на CAM 10-дневного куриного эмбриона в области, лишенной кровеносных сосудов, и окошко далее закрывали лентой. В других анализах 125 нг/мл βFGF также оказались эффективными в индуцировании роста кровеносных сосудов. За развитием ангиогенеза следили через 72 часа с помощью фотомикроскопии. CAM резко замораживали и 6 мкм криостатные срезы фиксировали ацетоном и окрашивали методом иммунофлуоресценции, как описано в примере 5C, 10 мкг/мл анти- β⊥ моноклонального антитела CSAT или LM609.
Иммунофлуоресцентная фотомикрография на фиг. 5В показывает усиленную экспрессию αvβ3 в ходе βFGF индуцированного ангиогенеза на куриной CAM в контрасте с отсутствием αvβ3 экспрессии в необработанных куриных CAM, как это показано на фиг. 5Б. αvβ3 легко детектируется на большинстве /75-80%/ сосудов βFGF - обработанных CAM. Кроме этого, экспрессия интегрина β1 не изменяется относительно экспрессии, наблюдаемой в необработанных CAM, поскольку β1 также легко детектируется на стимулированных кровеносных сосудах.
Относительную экспрессию αvβ3 и β1 интегринов далее определяли количественно в ходе βFGF-индуцированного ангиогенеза с помощью анализа лазерного конфокального изображения CAM криостатных срезов. Затем окрашенные срезы анализировали на лазерном конфокальном микроскопе Цейса. Двадцать пять сосудов, окрашенных LM609, и 15, окрашенных CSAT, /средний размер - 1200 мм2, интервал размеров - 350-3500 мм2/ выбирали из случайной партии и среднюю родаминовую флуоресценцию для каждого сосуда в расчете на единицу площади измеряли в произвольных единицах методом анализа лазерного конфокального изображения. Полученные данные выражали как среднюю интенсивность флуоресценции в произвольных единицах сосудов ± стандартная ошибка /SE/.
Результаты, представленные на фиг. 6, показывают, что окрашивание αvβ3 существенно усилено /примерно в 4 раза/ на CAM, обработанных βFGF в соответствии с определением методом Wilcoxon Rank Sum test /P < 0.0001/, тогда как β1 окрашивание существенно не менялось при обработке βFGF.
Далее CAM-анализ использовали для изучения эффекта другого мощного индуктора ангиогенеза альфа-фактора некроза опухоли /TNFa/ на экспрессию β1 и β3 интегринов. Фильтровальные диски, пропитанные либо βFGF или TNFa и помещенные на CAM 10-дневных эмбрионов, как было обнаружено, промотируют локальный ангиогенез через 72 часа.
Полученные результаты представлены на фотомикрографиях CAM в необработанном состоянии /фиг. 7A/, обработанных βFGF /фиг. 7Б/ или обработанных TNFa /фиг. 7В/. Кровеносные сосуды легко различимы на препаратах, обработанных как βFGF, так и TNFa, но они отсутствуют на необработанном CAM. Таким образом, местное применение системы фактор роста/цитокин приводит в результате к индуцированию ангиогенеза от зрелых сосудов на площади, соседней с участком, на котором первоначально отсутствовали кровеносные сосуды. На основании βFGF-индуцированных кровеносных сосудов и сопутствующей экспрессии как показано на фиг. 5В, можно сделать вывод, что обработка TNFa обеспечивает сравнимую активность.
Эти результаты указывают на то, что как у человека, так и у курицы кровеносные сосуды, принимающие участие в ангиогенезе, демонстрируют повышенную экспрессию αvβ3 . Аналогичным образом, экспрессия αvβ3 на культивированных эндотелиальных клетках может вызываться различными цитокинами in vitro, как это описано Janat с сотр., в J. Cell Physiol, 151:588 /1992/, Enenstein с сотр., Exp. Cell Res., 203:499 /1992/ и Swerlicr с сотр., в J.Invest. Derm., 99:715 /1993/.
Влияние на ангиогенез, индуцированный фактором роста, антитела и пептидного ингибитора продемонстрировано в Примерах 7A и 7B.
B. Эмбриональный ангиогенез.
Получение CAM для оценки эффекта ингибиторов ангиогенеза на естественное образование эмбрионной сосудистой системы осуществлялось с использованием 6-дневных эмбрионных куриных эмбрионов, как было описано ранее. На этой стадии развития кровеносные сосуды подвергают de novo росту и таким образом обеспечивают полезную систему для определения вероятности участия αvβ3 в эмбрионном ангиогенезе. CAM-систему готовили как описано выше за исключением того, что анализ проводили на 6-дневных эмбрионах, а не на 10-й день. Влияние на эмбрионный ангиогенез с помощью обработки антителами и пептидами настоящего изобретения продемонстрировано в примере 7C.
C. Ангиогенез, индуцированный опухолями.
С целью исследования роли αvβ3 в ангиогенезе, индуцированном опухолью, различные αvβ3-отрицательные человеческие меланомные и карциномные фрагменты использовали в CAM-анализе, при этом их предварительно выращивали и выделяли из CAM 17-дневного куриного эмбриона, как это описано Brooks с сотр., J.Cell Biol. , 122: 1351 /1993/ и в настоящем тексте. Такие фрагменты вызывали обширную неоваскуляризацию в присутствии лишь буфера.
Ангиогенез индуцировали в CAM-аналитической системе непосредственным присоединением фрагмента опухоли к CAM. Приготовление CAM куриного эмбриона идентично методике, описанной выше. Вместо фильтровального бумажного диска 50 мг - 55 мг фрагмент меланомной опухоли человека М211, человеческой опухоли легочной карциномы UCLAP-3, клеточной линии FG-карциномы поджелудочной железы человека /Cheresh с сотр., Cell, 58:945-953, 1989/, или человеческой ларингеальной карциномной клеточной линии Нер3, все из которых являются αvβ3 отрицательными опухолями, помещали на CAM на площади, которая первоначально была лишена кровеносных сосудов.
M21L человеческая меланомная клеточная линия, UCLAP-3 человеческая легочная карциномная клеточная линия, FG клеточная линия карциномы поджелудочной железы или Нep3 человеческая ларингеальная карциномная клеточная линия, все αvβ3 отрицательные, использовали для выращивания твердых человеческих опухолей на CAM куриного эмбриона. Суспензию в виде отдельных клеток 8 · 106 М21-1, UCLAP-3 и FB или 5 · 105 Нeр3 клеток вначале применяли на CAM в общем объеме 30 микролитров стерильной HBSS. Окошки закрывали лентой и эмбрионы инкубировали в течение 7 дней для обеспечения роста человеческих опухолевых повреждений. По истечении 7 дней, теперь уже 17-дневные эмбрионные опухоли подвергали резекции из CAM и проводили операцию очистки от окружающей CAM ткани. Опухоли нарезали на фрагменты весом 50-55 мг для использования в анализе на рост опухоли или ангиогенез. Фрагменты опухоли помещали на новый ряд 10-дневных куриных эмбрионных CAM в соответствии с описанным в примере 6A на площади, лишенной кровеносных сосудов.
Опухоли, растущие in vivo на куриных эмбрионных CAM, окрашивали для определения αvβ3 экспрессии mAB LM609, как это описано в примере 3A. Специфического окрашивания опухолевых клеток не наблюдалось, что свидетельствует об отсутствии αvβ3 экспрессии.
Такие CAM опухолевые препараты далее обрабатывали в соответствии с 7D и 7E с целью измерения эффектов антител и пептидов на ангиогенез, индуцированный опухолью. CAM опухолевые препараты также обрабатывали, как описано в Примерах 8, 9 и 12 для измерения влияния антител и пептидов на регрессию опухолей и апоптоза ангиогенных кровеносных сосудов и сосудистых клеток.
7. Ингибирование ангиогенеза, измеренное в CAM анализе.
A. Ингибирование ангиогенеза, индуцированного фактором роста путем местного применения ингибиторов.
1/ Обработка моноклональными антителами.
Для определения того, играет ли αvβ3 активную роль в ангиогенезе, фильтровальные диски, насыщенные βFGF или TNFa помещали на CAM, затем моноклональные антитела /обозначаемые также, как mAb/, LM609 /специфичные к αvβ3/, CSAT /специфичные к β1/, или P3G2 /специфичные к αvβ3/ добавляли к препарату.
Ангиогенез индуцировали на CAM 10-дневных куриных эмбрионах с помощью фильтровальных дисков, насыщенных βFGF . Затем такие диски обрабатывали 50 мл HBSS, содержащего 25 мг mAb в общем объеме 25 мкл стерильного HBSS через 0,24 и 48 часов. Через 72 часа CAM собирали и помещали в 35 мм чаши Петри и однократно промывали 1 мл фосфатного буфферного физиологического раствора. Донную сторону фильтровальной бумаги и CAM ткань далее анализировали с помощью стереомикроскопа Олимпус с использованием двух наблюдателей, работающих двойным слепым /double-blind/ методом. Ингибирование ангиогенеза считалось значительным, когда CAM проявляла > 50% - уменьшение инфильтрации кровеносных сосудов CAM непосредственно под диском. Эксперименты повторяли четыре раза в расчете на антитело в присутствии 6 - 7 эмбрионов в зависимости от условий.
Результаты влияния mAB на βFGF-индуцированный ангиогенез представлены на фиг. 8A-8Б. Необработанный CAM-препарат, лишенный кровеносных сосудов, показан на фиг. 8A с целью сравнения с βFGF-кровеносно-сосудистой индукцией, показанной на фиг. 8Б, а на фиг. 8В-8Д показано влияние действия mAb. Около 75% таких CAM, обработанных mAb LM609, проявляли > 50% ингибирование ангиогенеза, как это показано на фиг. 8Д, и во многих образцах не наблюдалась инфильтрация сосудов. В отличие от этого буферный контроль / фиг. 8A/ и диски, обработанные mAb CSAT /фиг. 8В/ и P3G2 /фиг. 8Г/, показали обширную васкуляризацию.
Идентичные результаты были получены при индуцировании ангиогеназа с помощью TNFa. Для изучения эффектов тех же антител на предварительно существующие зрелые кровеносные сосуды, присутствующие в результате нормального их развития по соседству с участками, лишенными сосудов, фильтровальные диски, насыщенные mAb, помещали на сосудистые участки CAM 10-дневных эмбрионов, на которых не проводили местное применение цитокина. Ни один из трех mAb не воздействовал на предварительно существующие сосуды, о чем свидетельствовало визуальное наблюдение с помощью стереомикроскопа. Так mAb LM609 селективно ингибирует только рост новых кровеносных сосудов и не влияет на зрелые кровеносные сосуды, присутствующие на соседних площадях. Тот же эффект наблюдался при применении синтетических пептидов локально или внутривенно, как это описано в примерах 7A2/ и 7E2/ соответственно.
2/ Обработка синтетическими пептидами.
CAM-анализы также осуществлялись с помощью синтетических пептидов изобретения с целью определения влияния циклических и линеаризованных пептидов на ангиогенез, индуцированный фактором роста. Такие пептиды готовили как описано в Примере 1 и 80 мкг пептида помещали в общий объем в 25 мкл стерильного HBSS. Пептидный раствор применяли на CAM-препарате немедленно и затем снова через 24 и 48 часов. Через 72 часа фильтровальную бумагу и окружающую CAM ткань отсекали и наблюдали, как описано выше.
Результаты такого анализа были аналогичны тем, что показаны на фиг. 9A-9В, как описано в Примере 7E2/, где синтетические пептиды внутривенно вводили в кровеносные сосуды, индуцированные опухолью. В присутствии контрольного пептида 62186 βFGF-индуцированные кровеносные сосуды оставались незатронутыми, как показано на фиг. 9A. В отличие от этого, при применении на фильтре циклического RGD пептида, 62814, образование кровеносных сосудов ингибировалось, оставляя площадь, лишенную новой сосудистой системы. Этот эффект по внешним признакам аналогичен эффекту, показанному на фиг. 9Б, как это описано в примере 7E2/ ниже. Кроме этого, также, как показано на фиг. 9В, для случая внутривенно инъецированных пептидов на площадях, где зрелые кровеносные сосуды находились достаточно далеко от расположения фильтра, насыщенного фактором роста, эффект от местного применения синтетических пептидов не наблюдался на таких основных сосудах. Таким образом, ингибиторная активность пептидов в отношении ангиогенеза ограничена участками ангиогенеза, индуцированными факторами роста, и при этом не затрагиваются соседние предварительно существующие зрелые сосуды и не наблюдается вредной цитотоксичности в отношении окружающей площади.
Аналогичные анализы были осуществлены с использованием других синтетических пептидов, полученных в Примере 1 и перечисленных в Таблице 1.
B. Ингибирование ангиогенеза, индуцированного фактором роста в результате внутривенного применения ингибиторов.
1/ Обработка моноклональными антителами
Эффект на ангиогенез, индуцированный фактором роста, моноклональных антител, внутривенно инъектированных в CAM препараты, также оценивался в отношении использования в настоящем изобретении.
Получение куриных эмбрионных CAM для внутривенных инъекций проводилось практически аналогично методике Примера 7A при некоторых модификациях. В ходе проверки яиц на свет рельефные кровеносные сосуды подвергали отбору и на оболочке яйца ставили метки для указания их положения. В скорлупе просверливали отверстия и в них загружали CAM, а βFGF-насыщенные фильтровальные бумаги помещали на CAM, как описано выше. Окна закрывали стерильной лентой и эмбрионы помещали в инкубатор. Через 24 часа второе небольшое окошко тщательно вырезали в боковой стороне яичной скорлупы непосредственно над выступающими кровеносными сосудами, отобранными ранее.
Тщательно удаляли внешнюю яичную скорлупу, оставляя незатронутыми эмбрионные мембраны. Мембранную оболочку делали прозрачной с помощью небольшой капли минерального масла /Перкин-Элмер Корп., Норволк, СТ/, что позволяет легко визуализировать кровеносные сосуды. Очищенные стерильные mAb или синтетические пептиды, последние из которых описаны ниже, инокулировали непосредственно в кровеносные сосуды с помощью иглы 30 калибра при дозе 200 мкг IgG на эмбрион в общем объеме в 100 мкл стерильной PBS. Окна заклеивали лентой и эмбрионам давали инкубироваться в течение 72 часов. Фильтровальные диски и окружающие CAM ткани анализировали, как описано выше.
Для определения локализации LM609 в CAM тканях или в опухолевых тканях, как показано в тексте и следующих примерах, которые предварительно инокулировали внутривенно LM609, фиксированные срезы блокировали 2,5% BSA в HBSS в течение 1 часа при комнатной температуре с последующим окрашиванием 1:250 разбавлением козьего анти-мышиного родамин-меченого вторичного антитела /Tago/. Затем срезы анализировали с помощью иммунофлуоресцентного микроскопа Цейса.
Результаты внутривенной обработки антителом индуцированных βFGF кровеносных сосудов CAM препаратов представлены на фиг. 10A-10В. На фиг. 10A показан ангиогенез, индуцированный в результате обработки βFGF. Как видно из фиг. 10Б не наблюдается изменений относительно присутствия βFGF индуцированной сосудистой системы при внутривенном воздействии на mAb P3G2, анти- αvβ3-антитело. В отличие от этого, обработка CAM препарата, ангиогенезиндуцированного βFGF с помощью LM609, анти- αvβ3-антителом, приводит в результате к полному ингибированию роста новых сосудов в фильтровальную область, как это показано на фиг. 10В. Ингибирующее влияние на ангиогенез, таким образом, является результатом ингибирования αvβ3 рецепторной активности под воздействием LM609 анти- αvβ3-специфичного антитела. Поскольку блокировка αvβ3 не ингибирует образование сосудистой системы в CAM фильтрующем сайте, αvβ3 не является существенным компонентом по сравнению с αvβ3 в отношении роста новых сосудов.
2/ Обработка синтетическими пептидами
Синтетические пептиды, полученные в Примере 1, по отдельности внутривенно инъецировали в кровеносные сосуды, индуцированные фактором роста в CAM препаратах, как описано выше. Влияние пептидов на жизнеспособность сосудов оценивали аналогичным образом.
C. Ингибирование эмбрионного ангиогенеза путем местного применения.
1/ Обработка моноклональными антителами.
Для определения факта участия αvβ3 в эмбрионном ангиогенезе влияния LM609 на de novo рост кровеносных сосудов на CAM изучали на 6-дневных эмбрионах, т.е. на стадии, отмеченной активной неоваскуляризацией, в соответствии с описанным в Примере 5A. CAM анализ проводили в соответствии с методикой примера 6C с последующим местным применением дисков, насыщенных mAb, помещенных на CAM 6-дневных эмбрионов в отсутствие цитокинов. Через 3 дня CAM подвергали резекции и фотографировали. Каждый эксперимент включал 6 эмбрионов на группу и повторяли 2 раза.
В таких условиях антитело LM609 /фиг. 11В/, но не CSAT /фиг. 11A/ или P3G-2 /фиг. 11Б/, предотвращали сосудистый рост; это указывает на то, что αvβ3 играет существенную роль в эмбрионной неоваскуляризации, которая не зависит от добавленного фактора роста для индукции ангиогенеза.
2/ Обработка синтетическими пептидами
Синтетические пептиды, полученные в примере 1, по отдельности добавляли в эмбрионный CAM препарат, полученный выше, в соответствии с описанным в Примере 5A2/, путем местного применения на CAM или внутривенного применения на кровеносных сосудах.
Влияние пептидов на жизнеспособность сосудов оценивали аналогичным образом.
D. Ингибирование ангиогенеза, индуцированного опухолью в результате местного применения.
1/ Обработка моноклональными антителами
Помимо описанного выше анализа на ангиогенез, в котором оценивали влияние анти- αvβ3 -антагонистов, LM609 и пептидов 62181, 62184, 62185, 62187 и 62880 на эмбрионный ангиогенез, исследовали также роль αvβ3 в ангиогенезе, индуцированном опухолью. В качестве индуктора использовали αvβ3 негативные человеческие M21-L меланомные фрагменты, предварительно выращенные и выделенные из CAM 17-дневного куриного эмбриона. Эти фрагменты получали в соответствии с методикой примера 6C.
Как описано выше в Примере 7A1, mAb раздельно локально применяли на опухолевых фрагментах при концентрациях 25 мкг в 25 мкл HBSS и затем окна заклеивали лентой. Снова добавляли mAb тем же способом через 24 и 48 часов. Через 72 часа опухоли и окружающие CAM ткани анализировали в соответствии с методикой примера 7A1.
Как описано в примере 6C, опухоли начально получали трансплантацией культивированных M21-L клеток, которые не экспрессировали интегрин αvβ3, как это описано Felding-Habermann с сотр., J.Clin. Invest. 89:2018 /1992/ на CAM 10-дневного куриного эмбриона. Такие αvβ3 негативные фрагменты индуцировали обширную неоваскуляризацию в присутствии только буфера, или mAb CSAT /анти- β/, либо P3G2 /анти- αvβ5/. В отличие от этого mAb LM609 / /анти- αvβ3/ снимали инфильтрацию большинства сосудов в опухолевой массе и окружающей CAM.
С целью количественного определения влияния mAb на ангиогенез, индуцированный опухолью, кровеносные сосуды, входящие в опухоль в рамках фокусной плоскости CAM подсчитывали с помощью стереомикроскопа двумя наблюдателями двойным слепым /double-blind/ методом. Каждая полоса данных на фиг. 12 представляет собой среднее число сосудов ±SE от 12 CAM в каждой группе дублированных экспериментов.
Такой количественный анализ обнаруживает трехкратное понижение числа сосудов, входящих в опухоль, обработанную mAb LM609 по сравнению с опухолями, обработанными буфером или другими mAb, P3G2 или CSAT /P < 0,0001/ в соответствии с разрядным суммарным тестом Вилкоксона. Тот факт, что М21-L опухоли не экспрессируют αvβ3 указывает на то, что mAB LМ609 ингибирует ангиогенез непосредственным воздействием на кровеносные сосуды, а не на опухолевые клетки. Эти результаты согласуются с гистологическим распределением αvβ3 раковой тканевой биопсии, представленной на фиг. 3A-3Г, где распределение αvβ3 ограничено кровеносными сосудами в опухоли, а не самими клетками опухоли.
2/ Обработка синтетическими пептидами
Синтетические пептиды, полученные в примере 1, локально применяли на опухоль-индуцированной ангиогенной CAM аналитической системе в соответствии с описанным выше. Аналогичным образом оценивали влияние пептидов на жизнеспособность сосудов.
E. Ингибирование ангиогенеза, индуцированного опухолью с помощью внутривенного применения
1/ Обработка моноклональными антителами
Опухоль-индуцированные кровеносные сосуды, полученные согласно методике примера 7D1, также обрабатывали mAb, применяемой путем внутривенной инъекции. Опухоли помещали на CAM, как описано в Примере 7D1, и окна заклеивали лентой и спустя 24 часа 200 мкг очищенного mAb инокулировали внутривенно в кровеносные сосуды куриного эмбриона, как это описано выше. Затем куриные эмбрионы инкубировали в течение 7 дней. Затем степень ангиогенеза наблюдали, как описано выше. Как описано в примере 8 ниже, после этого периода времени опухоли подвергали резекции и анализировали их вес для определения влияния воздействия антитела на рост или подавление опухоли.
2/ Обработка синтетическими пептидами
Оценивали также влияние воздействия пептида на сосудистую систему, индуцированную опухолью в CAM аналитической системе. Опухолево-CAM препарат использовали, как описано выше за исключением того, что вместо внутривенной инъекции mAb синтетические пептиды, полученные в соответствии с методиками примера 1 и примера 7A2, по отдельности внутривенно инъецировали в видимые кровеносные сосуды.
Результаты CAM анализа в присутствии циклического пептида 66203, содержащего HCl-соль, и контрольного пептида 62186 представлены на фиг. 9A-9В. Из фиг. 9A видно, что обработка контрольным пептидом не оказывает влияния на обильное образование крупных кровеносных сосудов, которые индуцируются обработкой опухолью с ростом в области первоначально лишенной кровеносных сосудов CAM. В отличие от этого в том случае, когда циклический RGD-пептид, 66203, антагонист на αvβ3 применяли на фильтре, образование кровеносных сосудов ингибировалось, оставляя площадь, лишенную новой сосудистой системы, как это показано на фиг. 9Б. Ингибирующий эффект RGD-содержащего пептида был специфичным и локализованным, о чем свидетельствует отсутствие каких-либо нежелательных эффектов, касающихся сосудов, расположенных по соседству с расположением опухоли. Так, согласно фиг. 9В, в том случае, когда ингибиторные пептиды внутривенно инъецируются в CAM аналитическую систему, не наблюдается эффекта на ранее существовавшие зрелые сосуды, присутствующие в CAM на площадях дистанцированных от положения опухоли. Предварительно существующие сосуды при таком расположении не подвергают влиянию ингибирующего пептида, который протекает внутри таких сосудов, хотя генерация новых сосудов из предварительно существующих сосудов в опухолевую массу подвергается ингибированию. Так, синтетические пептиды, включающие пептиды 66203 и 62184, предварительно описанные в лиганд-рецепторном анализе в Примере 4, являющиеся антагонистами, как было продемонстрировано в настоящее время ингибируют ангиогенез, который ограничен сосудами, подверженными развитию, а не зрелыми и предварительно существующими сосудами. Кроме этого, внутривенное вливание пептидов не приводит в результате к какому-либо вредному цитотоксическому воздействию на окружающую область о чем свидетельствует интактная сосудистая система, показанная на фиг. 9В.
Аналогичные анализы осуществляли с другими синтетическими пептидами, полученными в примере 1 и перечисленными в Таблице 1.
8. Ингибирование роста опухолевой ткани с помощью αvβ3 -антагонистов, измеренное в CAM анализе
Как показано в примере 7D1, помимо визуальной оценки влияния анти-αvβ3 -антагонистов на ангиогенез, индуцированный фактором роста или опухолью, эффект -антагонистов также оценивали измерением любых изменений опухолевой массы после воздействия. Для такого анализа CAM аналитическая система ангиогенеза, индуцированного опухолью, была получена как описано в Примере 6C и 7D. К концу 7-го дня инкубационного периода полученные в результате опухоли подвергали резекции от CAM и обрезали от любой оставшейся CAM ткани, промывали 1 мл фосфатного буферного физиологического раствора и для каждой опухоли определяли вес во влажном состоянии.
Кроме этого, получение опухоли для микроскопического гистологического анализа включает фиксацию представителей опухолей в Bulins Fixative в течение 8 часов и погружение в парафин. Серийные секции вырезали и окрашивали гематоксилином и эозином /H & E/ для микроскопического анализа. Гладсон с сотр. , J. Clin. Invest. 88: 1924 /1991/. Секции фотографировали с помощью микроскопа Олимпус при 250-кратном увеличении.
A. Местное применение
Результаты, полученные при взвешивании типичной человеческой меланомной опухоли /M21L/ в результате местного применения контрольного буфера /HBSS/, P3G2 /анти- αvβ5 /или LМ609 /анти- αvβ3 /, представлены в Таблице 4. Число эмбрионов оценивали для каждой обработки средним весом опухоли в миллиграммах /мг/, причем каждое значение рассчитывалось с учетом стандартного отклонения среднего значения.
Обработка mAb - Средний вес опухоли /мг/
HBSS контроль - 172 ± 26
P3G2 - 171 ± 36
LM609 - 52 ± 13
Экспозиция αvβ3 негативной человеческой меланомной опухолевой массы в CAM аналитической системе к LM609 вызывает уменьшение среднего веса необработанной опухоли от значения 172 мг ± 26 до 52 мг ± 13.
P3G2 антитело не оказывает влияния на массу опухоли. Так, блокировка αvβ3 рецептора местным применением αvβ3-специфичного LM609 антитела приводит в результате к регрессии опухолевой массы, а также к ингибированию ангиогенеза, как это показано в предыдущих примерах. Измеренный диаметр опухолевой массы, полученной в результате воздействия P3G2, составлял в среднем величину от 8 миллиметров до 1 сантиметра. В отличие от этого, LM609-обработанные опухоли имели диаметр 2-3 мм.
Замороженные срезы таких опухолей обнаруживали незатронутую цитоархитектуру опухоли, подвергшейся воздействию P3G2 в отличие от потери организованной клеточной структуры в опухоли, подвергшейся воздействию LM609. Поэтому αvβ3 рецепторная активность существенна для того, чтобы αvβ3 негативная опухоль сохраняла свое питание для поддержания массы в результате развития αvβ3 -экспрессирующей сосудистой системы. Блокировка αvβ3 с помощью αvβ3-антагонистов изобретения приводит в результате к ингибированию ангиогенеза в опухоль, что в конечном счете приводит к уменьшению массы опухоли.
B. Внутривенное применение
Результаты взвешивания типичной карциномной опухоли /UCLAP-3/ в результате внутривенного применения контрольного буфера /PBS, фосфатный буферный физиологический раствор/, CSAT /анти- β1/ или LM609 /анти- αvβ3/ представлены в таблице 5. Число эмбрионов оценивали для каждой обработки средним весом опухоли с учетом стандартного отклонения среднего значения.
Обработка mAb - Средний вес опухоли /мг/
PBS контроль - 85 ± 7
CSAT - 108 ± 22
LM609 - 30 ± 6
Экспозиция αvβ3-негативной человеческой карциномной опухолевой массы в CAM аналитической системе к LM609 вызывало уменьшение среднего веса необработанной опухоли от значения 85 мг ± 7 до 30 мг ± 6. CSAT антитело не оказывает существенного влияния на вес опухолевой массы. Таким образом, блокировка αvβ3-рецептора в результате внутривенного применения αvβ3-специфичного LM609 антитела приводит к регрессии карциномы, как это имеет место и в случае меланомной опухолевой массы, описанной выше, а также к ингибированию ангиогенеза, как это продемонстрировано в предыдущих примерах. Кроме этого, рост человеческой меланомной опухоли аналогичным образом ингибируется внутривенной инъекцией LM609.
9. Регрессия роста опухолевой ткани в присутствии αvβ3-антагонистов, измеренная в CAM анализе.
Для оценки влияния αvβ3-антагонистов на рост опухоли и ее выживание фрагменты человеческой меланомы и фрагменты карциномы легких, поджелудочной железы и гортани помешали на CAM 10-дневных эмбрионов в соответствии с описанным в примере 5A.
A. Внутривенное применение.
1/ Обработка моноклональными антителами
а. Обработка LM609 /анти- αvβ3/ и CSAT /анти- β1/.
Через 24 часа после имплантации CAM фрагментами карциномы αvβ3-негативной человеческой меланомы М21-L, карциномы поджелудочной железы FG, человеческой легкочной карциномы UCLAP-3 или человеческой ларингеальной карциномы НЕр3 эмбрионы внутривенно инъецировали только PBS или однократной дозой /300 мкг/100 мкл/ mAb LM609 /анти- αvβ3/ или CSAT /анти- β1/. Опухолям давали возможность размножаться еще в течение 6 дней. К концу периода инкубации опухоли подвергали тщательной резекции и освобождали от окружающей CAM ткани. Резекцию опухоли проводили два независимых исследователя, удаляющих только легко определяемую твердую опухолевую массу. Опухоли имели хорошо обозначенные края и таким образом тонкая полупрозрачная мембрана /CAM/, которая легко различима с твердой опухолевой массой удалялась без нарушения самой опухолевой массы. Подвергнутые резекции опухоли взвешивали и подвергали морфологическому и гистологическому изучению.
Как показано на фиг. 13 веса влажных опухолей к концу 7-го дня определяли и сравнивали с исходным весом опухоли до обработки. Каждая полоса представляет собой среднее значение ± SE. для 5-10 опухолей на группу. mAb LM609 значительно ингибирует рост опухоли /p < 0,001/ по сравнению с контрольными образцами для всех испытуемых опухолей. Во всех случаях имела место пролиферация опухолей, обработанных PBS или CSAT. В отличие от этого, mAb LM609 не только предотвращает рост таких опухолей, но индуцирует обширную регрессию в большинстве случаев. Важно отметить, что клетки таких опухолей не экспрессируют интегрин αvβ3, демонстрируя тот факт, что ингибирование роста обязано анти-ангиогенным эффектам такого анти-тела на сосудистую систему, а не на непосредственно опухолевые клетки.
b. Обработка LM609 /анти- αvβ3/ P3G2 /анти- αvβ5/.
Человеческие M21-L меланомные опухолевые фрагменты /50 мг/ имплантировали на CAM 10-дневных эмбрионах, как описано в примере 5A. Через 24 часа эмбрионы внутривенно инъецировали только PBS или однократной дозой /300 мкг/100 мкл / mAb LM609 /анти- αvβ3/ или P3G2 /анти- αvβ5/. Опухолям давали размножаться, как описано в примере 9A1/а, и подвергали морфологическому и гистологическому изучению.
Представители М21-L опухолей, обработанных mAb P3G2 /анти- αvβ5/ или LM609 /анти- αvβ3/ подвергались морфологическому исследованию. Обработанные P3G2 опухоли были крупными /8 мм в диаметре/ и имели хорошо выраженную сосудистую систему, тогда как обработанные mAb LM609 были значительно мельче /3 мм в диаметре/ и не содержали детектируемых кровеносных сосудов.
Далее опухоли дополнительно исследовали приготовлением гистологических срезов и окрашиванием гематоксилином и эозином, как описано в примере 9A1/а. Как показано на фиг. 14 / верхняя часть/, опухоли, обработанные mAb P3G2 /анти- αvβ5/ демонстрировали многочисленные жизнеспособные и активно делящиеся опухолевые клетки, как показано митотическими значениями /острие стрелки/ и множеством кровеносных сосудов /стрелки/ по всех опухолевой строме. В отличие от этого, небольшое количество или отсутствие жизнеспособных опухолевых клеток или кровеносных сосудов детектировалось в опухолях, обработанных mAb LM609 /анти- αvβ3/ /фиг. 14, нижняя часть/. Эти результаты демонстрируют тот факт, что антагонисты интегрина αvβ3 ингибируют индукцированный опухолью ангиогенез, что приводит к остановке роста и регрессии большого числа человеческих опухолей in vivo. Важно отметить, что эмбрионы, изученные через 7 дней роста опухоли /эмбрионный 17-ый день/, оказались нормальными в ходе тщательного изучения в независимости от того были ли они обработаны αvβ3-антагонистом. Эти открытия указывают на то, что антагонисты такого интегрина нетоксичны в отношении развивающихся эмбрионов.
2/ Обработка синтетическими пептидами.
Человеческие M21-L меланомные опухолевые фрагменты /50 мг/ имплантировали на CAM 10-дневных эмбрионов, как описано в Примере 5A. Через 24 часа эмбрионам делали однократную внутривенную инъекцию 300 мкг/100 мкл цикло RADfV /69601/ или цикло RGDfV /66203/. Через 72 часа общего времени опухоли удаляли, подвергали морфологическому изучению и фотографировали стереомикроскопом, как описано в примере 9A1/.
Увеличенные фотографии, показанные на фиг. 15A-15Д, соответствуют следующему: на фиг. 15A изображены дубликатные образцы, обработанные цикло-RADfV пептидом /69601/; на фиг. 15Б - дубликатные образцы, обработанные цикло-RGDfV пептидом /66203/; на фиг. 15В показана соседняя ткань, взятая с некоторых эмбрионов, обработанных цикло-RGDfV пептидом /66203/; на фиг. 15Г и 15Д изображено сильное увеличение /13-кратное/ опухоли, обработанной пептидом. На фиг. 15Г изображены нормальные кровеносные сосуды опухоли, обработанной контрольным пептидом /69601/. На фиг. 15Д изображены примеры разрушенных кровеносных сосудов опухолей, обработанных цикло-RGDfV пептидом /66203/ /стрелки/.
Полученные результаты иллюстрируют тот факт, что лишь пептид 66203 ингибирует образование сосудов, а также что сосуды в CAM ткани, соседней с опухолью, не затрагиваются.
10. Регрессия роста опухолевой ткани в присутствии αvβ3-антагонистов, измеренная in vivo модельным анализом на кроличьем глазу.
Влияние анти- αvβ3-антагонистов на ангиогенез, индуцированный фактором роста, может наблюдаться в природных прозрачных структурах, примерами которых может служить роговая оболочка глаза. Новые кровеносные сосуды растут от края роговой оболочки глаза, который хорошо снабжается кровью, в направлении центра роговой оболочки, которая в нормальном состоянии не снабжается кровью. Такие стимуляторы ангиогенеза, как βFGF при применении на роговой оболочке глаза индуцируют рост новых кровеносных сосудов от края оболочки. Антагонисты ангиогенеза, применяемые на роговой оболочке глаза, ингибируют рост новых кровеносных сосудов от края оболочки. Так, роговая оболочка подвергается ангиогенезу в результате инвазии эндотелиальных клеток от края роговой оболочки в жесткую коллаген-упакованную ткань роговой оболочки, что можно легко наблюдать визуально. Поэтому модельный анализ на кроличьем глазу обеспечивает in vivo модель для непосредственного наблюдения стимуляции и ингибирования ангиогенеза после имплантации соединений непосредственно в роговую оболочку глаза.
A. In vivo модельный анализ на кроличьем глазу.
1/ Ангиогенез, индуцированный факторами роста.
Ангиогенез, индуцированный в in vivo модельном анализе на кроличьем глазу в присутствии фактора роста βFGF, причем подробности такого метода описаны ниже.
а. Приготовление гидроновых гранул, содержащих фактор роста и моноклональные антитела.
Гидроновые полимерные гранулы, содержащие фактор роста и mAb, получали как описано D'Amato с сотр., в Proc. Natl. Acad. Sci. 91:4082-4085 /1994/. Индивидуальные гранулы, содержащие 650 нг фактора роста βFGF, связывали с сукралфатом /Карафет, Марион Меррелл Доу корпорейшн/ с целью стабилизации βFGF и обеспечения его медленного выделения в окружающую ткань. Кроме этого, гидроновые гранулы готовили таким образом, чтобы они содержали 40 мкг mAb LМ609 /анти- αvβ3/или mAb P1F6 /анти- αvβ5/ в PBS. Гранулы отливали в специально приготовленные Тефлоновые втулки, которые имели 2,5 мм сердцевину, просверленную в их поверхностях. Примерно 12 мкл отлитого материала помещали в каждую втулку и полимеризовали в течение ночи в стерильном колпаке. Затем гранулы стерилизовали облучением ультрафиолетовым светом.
b. Обработка моноклональными антителами.
В каждом эксперименте использовали по три кролика, в один глаз которых помещали гранулу, содержащую βFGM и LM609, а в другой глаз загружали гранулу, содержащую βFGF и мышиный mAb P1F6 /анти- αvβ5/. Использование спаренного глазного тестирования для сравнения LM609 /анти- αvβ3/ с другими mAb и PBS контрольными образцами обеспечивает средство точного тестирования для демонстрации значительных различий между испытанными mAb.
P1F6 mAb иммунореагирует с интегрином αvβ5 , который обнаружен на поверхности сосудистых эндотелиальных клеток но, по-видимому, не участвует в ангиогенезе. Для определения факта участия mAb P1F6 в ангиогенезе готовили гранулы, содержащие только mAb, и анализировали их, как описано ниже, на факт того, что mAb не индуцирует ангиогенез.
Все испытуемые mAb очищали от асцитной жидкости с использованием Протеин-A Сефароза C1-4B аффинной колоночной хроматографии согласно хорошо известным способам. Элюированный иммуноглобулин затем диализировали против PBS и обрабатывали Детокси-гелем /Пьерсе Кэмикалс/ с целью удаления эндотоксина. Как было показано, эндотоксин является мощным ангиогенным и противовоспалительным стимулянтом. Поэтому mAb тестировали на присутствие эндотоксина с помощью Хромогенного лимулус амебоцитного лизатного анализа (Сhromogenic Zimulus Amebocyte Zysate Assay) /Био-Виттакер/ и лишь те mAb, которые не содержали детектируемых количеств эндотоксина использовали в модельном анализе на кроличьем глазу.
Гидроновую гранулу, содержащую βFGF и mAb LM609 /анти- αvβ3/ или P1F6 /анти- αvβ5/, вставляли в карман роговой оболочки глаза, образованной на глазах кролика. Гидроновая гранула также содержала сукралфат для стабилизации βFGF в ходе анализа. Индивидуальные гранулы имплантировали в созданные хирургическим путем "карманы", сформированные в мид-строме роговой оболочки глаз кролика. Хирургические операции проводили в стерильных условиях с использованием операционного микроскопа Вилд модели М691, снабженного пучковым дробителем, с которым была смонтирована камера для фотографической регистрации индивидуальных роговых оболочек глаза. "Карман" размером 3х5 мм создавали в строме роговой оболочки, делая 3 мм разрез на половину толщины роговой оболочки лезвием 69 Биавер. Строму рассекали периферически с использованием шпателя для радужной оболочки глаза и гранулу имплантировали по ее периферическим краям на 2 мм от лимбуса.
В течение последующих 14 дней βFGF и mAb диффундировали из имплантированной гранулы в окружающую ткань и в результате этого оказывали влияние на ангиогенез от края роговой оболочки глаза.
Представительные результаты каждой обработки изображены на фиг. 16A-16Д. Число присутствующих сосудов рассчитывали количественно и выражали в терминах часов, которые определяли следующим образом. Глаз делили на 12 равных секторов также, как это имеет место на циферблате часов. "Один циферблатный час сосудов" относится к такому количеству сосудов, которое заполняет площадь глаза, эквивалентную одному часу на циферблате. Пять кроликов, получивших лишь βFGF, демонстрировали свежий ангиогенез, в котором новые кровеносные сосуды росли от края роговой оболочки в направлении ее центра, который в нормальном состоянии не имеет кровеносных сосудов. Один из таких кроликов имел лишь 1 циферблатный час сосудов в расчете на гранулу. Два кролики, получивших как βFGF, так и LM609, абсолютно не проявляли ангиогенеза вплоть до 14 дня после хирургического вмешательства. Один из таких кроликов имел 3 фокуса кровоизлияния и почечные сосуды к 14 дню. Два кролика, получивших βFGF и mAb P3G2 /анти- αvβ5/, демонстрировали обширную васкуляризацию, в которой новые кровеносные сосуды росли от края роговой оболочки глаза в нее. Один из таких кроликов имел 1-2 часа сосудов в расчете на гранулу.
Как следует из модельного анализа на кроличьем глазу, ангиогенный эффект не наблюдается на нормальных паралимбальных сосудах в присутствии фактора роста βFGF у кроликов, получивших mAb LM609 /анти- αvβ3/. В отличие от этого, ангиогенез наблюдался на паралимбальных сосудах в присутствии фактора роста βFGF у кроликов, получивших mAb P3G2 /анти- αvβ5/. Полное ингибирование ангиогенеза роговой оболочки глаза при воздействии mAb LM609 значительно выше, чем в присутствии анти-ангиогенных реагентов, о которых сообщалось ранее.
11. In vivo регрессия роста опухолевой ткани в присутствии антагонистов, измеренная химерным анализом мышь-человек.
In vivo химерную модель человек-мышь генерировали заменой части кожи мыши линии SC1D человеческой неонатальной крайней плотью /фиг. 17/. После укоренения кожного трансплантанта человеческую крайнюю плоть инокулировали клетками карциномы. После образования способной к изменению опухоли mAb LM609 /анти- αvβ3/ или PBS инъецировали в хвостовую вену мыши. Через 2-3 недели опухоль вырезали и анализировали путем взвешивания и гистологии.
A. In vivo анализ химерной системы мышь-человек.
In vivo химерную модель мышь-человек готовили в соответствии с описанным Yan с сотр., J. Clin. Invest., 91:986-996 /1993/. Если говорить вкратце, то 2 см2 квадратной площади кожи хирургически удаляли у мышей разновидности SC1D /в возрасте 6-8 недель/ и заменяли человеческой крайней плотью. Мышей анестезировали, с площади 5 см2 удаляли волосы с каждой стороны латерального абдоминального участка путем выбривания. Два круглых трансплантата площадью 2 см2 готовили путем удаления полной толщины кожи ниже фасции. Полную толщину человеческих кожных трансплантантов того же размера, производных человеческой неонатальной крайней плоти, помещали на раневые слои и зашивали на место. Трансплантант покрывали Band-Aid, который пришивали к коже. Для покрытия раны также применяли микропористую тканевую ленту.
M21L человеческую клеточную линию или МДА 23.1 клеточную линию карциномы грудной железы /АТСС НТВ 26; αvβ3-негативная в соответствии с иммунореактивностью тканевых срезов в присутствии mAb LM609/ использовали для образования твердых человеческих опухолей на трансплантатах человеческой кожи на мышах линии SC1D. Одноклеточную суспензию 5 · 106 M21-L или МДА 23.1 клеток инъецировали интрадермально в трансплантант человеческой кожи. Затем мышей наблюдали в течение 2-4 недель с целью оценки роста измеряемой человеческой опухоли.
B. Внутривенное применение
1/ Обработка моноклональными антителами.
После роста измеряемых опухолей мышей линии SC1D, которым делали инъекции M21L опухолевыми клетками, внутривенно инъецировали в хвостовую вену 250 мкг mAb LM609 /анти- αvβ3/ или PBS дважды в неделю в течение 2-3 недель (результаты см. в таблице 6). После этого опухоли подвергали резекции с кожи и обрезали окружающую ткань. Нескольких мышей оценивали на каждую из обработок, определяя средний вес опухоли при каждой обработке.
Обработка - Средний вес опухоли /мг/
PBS - 198
LM609 - 113
Воздействие М21L αvβ3-негативной человеческой карциномной опухолевой массы в химерной аналитической / системе мышь-человек на LM609 /анти- αvβ3/ вызывало уменьшение среднего веса опухоли, обработанной PBS от 198 мг до 213 мг.
Представители опухолей M21L, обработанных mAb LM609 /анти- αvβ3/ и PBS были подвергнуты морфологическому исследованию. PBS -обработанные опухоли были крупными /с диаметром 8-10 мм/ и хорошо васкуляризованными, тогда как опухоли, обработанные mAb LM609 /анти- αvβ3 были значительно более мелкими /с диаметром 3-4 мм/ и не содержали детектируемых кровеносных сосудов.
Опухоли, полученные в кожных трансплантантах, которые инъецировали МДА 23.1 клетками, были детектируемыми и измеряемыми. Морфологическое исследование полученных опухолей обнаружило, что имеет место неоваскуляризация от трансплантированных человеческих тканей в МДА 23.1 опухолевые клетки.
Таким образом, блокировка αvβ3 рецептора внутривенным введением αvβ3 специфичного LM609 антитела приводит в результате к регрессии карциномы в такой модельной системе тем же образом, что и в случае CAM и модельных систем на кроличьих глазах, как это описано в примерах 9 и 10 соответственно.
12. Стимуляция сосудистых клеток с целью вхождения в клеточный цикл и их вовлечения в апоптоз в присутствии антагонистов интегрина αvβ3 при измерении с помощью CAM-анализа.
Ангиогенный процесс явно зависит от способности цитокинов, таких как βFGF и VEGF, стимулировать пролиферацию сосудистых клеток. Mignatti с сотр. J. Cell Biochem., 471:201 /1991/; Jakeshita с сотр. J.Clin. Invest., 93:6 62 /1994/; и Koyama с сотр. J. Cell Physiol., 158:1 /1994/. Однако, также следует отметить, что передача сигнала может регулировать дифферециацию таких сосудистых клеток в зрелые кровеносные сосуды. Так, возможно, что интерференция с сигналами, относящимися к росту или дифферециации сосудистых клеток, подвергающихся новому росту или ангиогенезу, может в результате приводить к расстройству ангиогенеза.
Лигирование интегрина, как было показано, принимает участие как в пролиферации клеток, так и в апоптозе или программированной клеточной смерти in vitro. См. Schwartz, Cancer Res. 51:1503 /1993/; Meredith с сотр., Mol. Biol. Cell. , 4:953 /1993/; Frisch с сотр., J. Cell Biol., 124:619 /1994/; и Ruoslahti с сотр. Cell, 77: 477 /1994/. Аналогичное исследование влияния αvβ3-антагонистов на ангиогенез обнаружило присутствие прерванных и разрушенных связанных с опухолью кровеносных сосудов. Поэтому, возможно, что потеря непрерывности кровеносных сосудов может быть следствием селективного некроза или апоптоза сосудистых клеток.
Для исследования такой возможности CAM изучали после индуцирования ангиогенеза в присутствии фактора роста βFGF и обработки mAb и циклическими пептидами настоящего изобретения.
A. Обработка моноклональными антителами
Апоптоз может детектироваться большим числом методов, которые включают непосредственное изучение ДНК, выделенной из ткани для определения фрагментации ДНК, и детекцию
3'OH в интактной ткани с помощью антитела, которое специфически определяет свободную 3'ОН группу фрагментированной ДНК.
1/ Анализ фрагментации ДНК
Ангиогенез индуцировали помещением фильтровальных дисков, насыщенных βFGF на CAM 10-дневных эмбрионов, как это описано в Примере 6A. Иммуногистологический анализ CAM в присутствии LM609 /анти- αvβ3/ позволил обнаружить пик экспрессии αvβ3 на кровеносных сосудах через 12-24 часа после инициирования ангиогенеза с помощью βFGF. Так, через 24 часа после стимуляции βFGF эмбрионы инокулировали внутривенно 100 мкл только PBS или PBS, содержащим 300 мкг CSAT /анти- β1/ или LM609 /анти- αvβ3/.
Фрагментацию ДНК детектировали путем резекции CAM ткани непосредственно ниже βFGF насыщенных фильтровальных дисков через 24 или 48 часов после внутривенной инокуляции с помощью mAb LM609 /анти- αvβ3 /, CSAT /анти- β1 / или PBS. Подвергнутые резекции ткани CAM трижды промывали стерильной PBS и мелко крошили, ресуспендировали в 0,25% бактериальной коллагеназе /Ворсингтон Биокемикал; Фрихолд, Нью-Йорк/ и инкубировали 90 минут при 37oC при периодическом вихревом перемешивании. ДНК экстрагировали из равного числа CAM клеток из одноклеточной суспензии, как описано выше см. Bissonette с сотр., Nature, 359:552 /1992/. Вкратце, равное число клеток CAM лизировали в 10 мМ трис-Cl, pH 8.0, 10 мМ ЕДТА в 0,5% /об./об./ Тритона Х-100 /Сигма, Сент-Льюис, МО/. Клеточный лизат центрифугировали со скоростью 16,000 xg в течение 15 минут при 4oC для отделения растворимых фрагментов ДНК от интактной хроматиновой гранулы. Фрагментированную ДНК промывали, осаждали и анализировали на 1,2% /вес. об./ агароза геле.
Растворимую фрагментированную ДНК выделяли из равного числа клеток CAM при каждой обработке, разделяли с помощью электрофореза на агарозном геле и визуализировали путем окрашивания этидиум бромидом. Никаких отличий не наблюдалось в относительной степени ДНК фрагментации в результате трех различных обработок через 24 часа после обработки. Однако, через 48 часов после обработки mAb LM609 /анти αvβ3/, наблюдалось значительное повышение фрагментации ДНК по сравнению с эмбрионами, обработанными либо mAb CSAT /анти- β1/ или только PBS.
2/ Стимуляция сосудистых клеток для вхождения в клеточный цикл
С целью экспериментального изучения роли αvβ3 в этих процессах клетки, произведенные из CAM, обработанных или не обработанных βFGF, окрашивали йодистым пропидиумом и проводили иммунную реакцию с mAb LM609 /анти- αvβ3/.
CAM, изолированные из эмбрионов через 24 и 48 часов после обработки mAb LM609 /анти- αvβ3/ CSAT /анти β1/ или PBS, диссоциировали в одноклеточную суспензию путем инкубации в присутствии бактериальной коллагеназы, как описано выше. Единичные клетки затем подвергались условиям проницаемости и окрашивались с помощью детекционного набора Apop Tag in situ согласно инструкциям производителя /Онкор, Гайтерсбург, МД/. Apop Tag представляет собой антитело, которое специфически определяет 3'OH группы фрагментированной ДНК. Детекция таких свободных 3'OH групп представляет собой установленный способ определения апоптотических клеток. См. Gavrieli. с сотр., J. Cell. Biol., 19:493 /1992/.
Окрашенные Apop Tag клетки затем промывали в 0,1% /об./об./ Тритона X-100 в PBS и ресуспендировали в FACS буфере, содержащем 0,5% /вес./об./ BSA, 0,02% /вес./об./ азида натрия и 200 мкг/мл RNазы A в PBS. Клетки инкубировали в течение 1,5 часа, промывали и анализировали флуоресцентной сортировкой активированных клеток. Флуоресценцию клеток измеряли с использованием PACScan проточного цитометра и полученные данные анализировали, как описано ниже.
Клеточную флуоресценцию измеряли с помощью PACScan /сканирующего клеточного сортера с возбуждением флуоресценции прим. пер. /проточного цитометра/ /Бектон Дикинсон, Маунтэйн Вью. Боковое рассеяние //SSC/ и переднее рассеяние. /FSC/ определяли одновременно и все полученные данные собирали в компьютере Хьюлет-Пакард /НР9000, снабженным FACScan исследовательским программным обеспечением /Бектон Дикинсон, Маунтэйн Вью, КА/. Данные анализировали с помощью программы Р.С. Лизис, версия 1 /Бектон Дикинсон, Маунтэйн Вью, КА/. Негативные контрольные входы устанавливали с использованием клеточной суспензии без добавления первичных антител из набора Apop Tag. Аналогичное открытие мембранного канала применяли на обеих клеточных популяциях в результате анализа примерно 8.000 клеток в расчете на различные обработки клеток.
Процент одиночных клеток, производных от CAM, обработанных mAb и окрашенных Apop Tag в соответствии с определением методом FACS анализа, изображен на фиг. 18. Черная полоса представляет собой клетки из эмбрионов, обработанных за 24 часа до анализа. Пунктирная полоса представляет собой клетки эмбрионов обработанные за 48 часов до анализа. Каждая полоса представляет собой среднее значение ± SE. при трех параллельных измерениях.
Как следует из фиг. 18 CAM, обработанные за 2 дня до воздействия mAb LM609 /анти- αvβ3/, демонстрируют 3-4-кратное увеличение окрашивания Apop Tag по сравнению с CAM, обработанными только PBS или CSAT /анти- β1/.
B. Обработка синтетическими пептидами.
CAM-анализы с использованием ангиогенеза, индуцированного фактором роста, как это описано в примере 6A, проводили также с использованием синтетических пептидов изобретения с целью определения влияния циклических пептидов на апоптоз. Пептиды цикло-RGDfV /66203/ и цикло-RADfV /69601/ получали как описано в примере 1.
Пептидные растворы или PBS инъектировали в CAM препарат с концентрацией 300 мкг/мл. Через 24 и 48 часов фильтровальную бумагу и окружающую CAM ткань вскрывали и окрашивали Apop Tag для детекции апоптоза, как описано выше в примере 12A2/.
Как показано на фиг. 18 CAM, обработанные за два дня до взаимодействия с пептидом 69203 /цикло-RGDfV /, демонстрировали 3-4-кратное увеличение окрашивания Apop Tag по сравнению с CAM, обработанными или только PBS или контрольным циклическим пептидом 69601 /цикло- RADfV /.
C. Влияние обработки моноклональными телами на апоптоз и клеточный цикл
Одноклеточные суспензии также изучали с целью определения числа копий хромосомных ДНК путем окрашивания иодистым пропидиумом с целью определения эффекта обработки моноклональными телами на клеточный цикл и на апоптоз путем окрашивания с помощью Apop Tag.
Одноклеточные суспензии CAM, обработанные за 24 или 48 часов до взаимодействия с mAb LM609 /анти- αvβ3 / или CSAT /анти- β1 / либо PBS, готовили, как описано в примере 12A1/.
Для окрашивания клеток с помощью Apop Tag клеточные суспензии трижды промывали буфером, содержащим 2,5% /вес.об./ BSA и 0,25% /вес.об./ азида натрия в PBS. Затем клетки фиксировали в 1% /вес./об./ параформальдегиде в PBS в течение 15 минут с последующим тройным промыванием, как описано выше. Для предотвращения неспецифического связывания одноклеточные суспензии блокировали 5% /вес.об./ BSA в PBS в течение ночи при 4oC. Затем клетки промывали как описано выше, окрашивали с помощью Apop Tag и клеточную флуоресценцию измеряли с помощью FACScan как описано выше в примере 12A.
Клетки при каждых экспериментальных условиях окрашивали йодистым пропидиумом /Сигма, Сент-Льюис, МО/ при концентрации 10 мкг/мл в PBS в течение 1 часа, дважды промывали PBS и анализировали на ядерные характеристики типичные для апоптоза, включая хроматиновую конденсацию и сегментацию. Процентное количество апоптотических клеток оценивали морфологическим анализом в произвольно выбранных микроскопических полях, имеющих значения по крайней мере 10-15.
Объединенные результаты исследования одноклеточной суспензии CAM из эмбрионов, обработанных либо CSAT /анти- β1/ или LM609 /анти- αvβ3/ , окрашенных Apop Tag и иодистым пропидиумом и проанализированных методом FACS, представлены на фиг. 19. На оси Y отображено окрашивание с помощью Apop Tag /апоптоз/, на оси X отображено окрашивание иодистым пропидиумом /содержание ДНК/. Горизонтальная линия отображает негативный мембранный канал для окрашивания Apop Tag. Левые и правые панели указывают CAM клетки из CSAT и LM609-обработанных эмбрионов соответственно. Анализ клеточного цикла осуществляли анализом примерно 8.000 событий в расчете на состояние и полученные данные представлены на контурном графике.
Образцы единичных клеток, окрашенных красителем ДНК иодистым пропидиумом, позволили установить, что 25-30% LM609 /анти- αvβ3/ обработанных CAM клеток через 48 часов после обработки демонстрируют ядерную конденсацию и/или сегментацию. Такие процессы характерны для клеток, подвергаемых апоптозу. Этот факт отличается от CAM, обработанных CSAT /анти-β1/, когда 90-95% клеток демонстрируют нормальное клеточное окрашивание.
Как показано на фиг. 19 в соответствии с индукцией апоптоза под действием LM609, наблюдается значительное число клеток в пике, содержащем менее одной копии ДНК /AO/. Этот пик был показан ранее для отражения фрагментированной ДНК на поздней стадии развития апоптотических клеток. См. Telford с сотр., Cytometry, 13:137 /1992/.
Кроме этого, такие AO клетки легко окрашиваются с помощью Apop Tag, что подтверждает способность такого реагента у детектировать апоптототические клетки. Однако, в дополнение к окрашиванию клеток в AO, значительное число клеток, содержащих более одной копии ДНК, также окрашивается с помощью Apop Tag /фиг. 19/. Эти результаты демонстрируют тот факт, что LM609 обладает способностью промотировать апоптоз среди сосудистых клеток, которые уже вошли в клеточный цикл. В отличие от этого клетки, производные от контрольных CAM, которые вошли в клеточный цикл, демонстрируют минимальное окрашивание Apop Tag, что соответствует нескольким апоптотическим клеткам, определяемым в контрольных обработанных CAM.
Среди тех клеток в CAM, стимулированных βFGF, которые вошли в клеточный цикл /S и G2/M фаза/, 70% демонстрируют положительное окрашивание LM609 /анти- αvβ3/. Это значение можно сравнить с 10% LM609 окрашивания, наблюдаемого в циклических клетках из CAM, необработанных β FGF. Эти открытия указывают на то, что после β FGF стимуляции большая часть αvβ3 -несущих клеток демонстрируют активную пролиферацию.
Совместное рассмотрение этих фактов указывает на то, что внутривенное инъецирование mAb LM609 или циклического пептида - антагониста αvβ3 промотирует апоптоз в курином CAM с последующей индукцией ангиогенеза.
CAM уже изучали гистологически на экспрессию αvβ3 путем исследования иммунореактивности с LM609 и на наличие клеток, подвергающихся апоптозу в результате иммунной реакции с Apop Tag. Срезы CAM, подвергнутые резекции из эмбрионов, обработанных за 48 часов LM609 /анти- αvβ3 /, CSAT /анти- β1 / или PBS, полученными в примере 5A, промывали, погружали в OTC /Бакстер/ и мгновенно замораживали в жидком азоте. Шестимикронные срезы CAM тканей вырезали, фиксировали в ацетоне в течение 30 секунд и хранили при -70oC до дальнейшего использования. Тканевые срезы готовили для окрашивания быстрым промыванием в 70% /об./об./ этаноле /EtOH/ с последующим тройным промыванием в PBS. Далее, срезы блокировали 5% /вес./об./ BSA в PBS в течение 2 часов, после чего проводили инкубацию в присутствии 10 мкг/мл mAb LM609 в течение 2 часов. Затем срезы промывали и инкубировали в присутствии 1:50 разбавления родаминового конъюгированного козьего анти-мышиного IgG /Фишер Сайнтифик, Питсбург, РА/ в течение 2 часов. Наконец эти срезы промывали и окрашивали с помощью Apop Tag, как описано в Примере 12A2/. Окрашенные тканевые срезы превращали в препараты и анализировали методом конфокальной иммунофлуоресцентной микроскопии.
На фиг. 20 панели A-C показывают CAM-ткань от эмбрионов, обработанных CSAT / анти- β1/, а панели D-F представляют CAM-ткань из LM609 /анти- αvβ3/ обработанных эмбрионов. Панели A и D показывают ткани, окрашенные Apop Tag и визуализированные флуоресценцией /FITC/, наложенной на D.I.C. изображение. Панели B и E содержат изображение тех же тканей, окрашенных mAb LM609 /анти- αvβ3 / и визуализированных с помощью флуоресценции /родамин/. Панели C и F представляют собой слитые изображения тех же тканей, окрашенных Apop Tag и LM609, где желтое окрашивание относится к солокализации. Полосы имеют размеры 15 и 50 мкм в левой и правой панелях соответственно.
Как показано на фиг. 20 /A-C/, после внутривенного инъецирования CSAT или PBS контрольных образцов окрашивание Aрор Tag было минимальным и случайным, что указывает на минимальный уровень апоптоза внутри ткани.
В отличие от этого, CAM из эмбрионов, предварительно обработанных LM609 или циклическим пептидом 203, демонстрировали преимущественное количество сосудов, интенсивно окрашенных Apop Tag, тогда как минимальная реактивность наблюдалась у окружающих несосудистых клеток /фиг. 20 D-F/. Кроме этого, в том случае, когда Apop Tag и LM609 использовали для окрашивания таких тканей /19C и 19F/ значительная солокализация наблюдалась лишь между этими маркерами в CAM, являющихся производными эмбрионов, обработанных αvβ3 антагонистами /фиг. 20F/. Эти открытия демонстрируют тот факт, что после индукции ангиогенеза in vivo ингибиторы интегрина αvβ3 селективно промотируют aпoптoз αvβ3 -несущих кровеносных сосудов.
Хотя ангиогенез представляет собой сложный процесс, включающий большое число молекулярных и клеточных биологических событий, некоторые черты доказательств позволяют предположить, что сосудистые клеточные интегрины αvβ3 играют относительно позднюю роль в таком процессе. Во-первых, иммуногистологический анализ позволил обнаружить, что экспрессия αvβ3 на сосудистых клетках достигает максимума через 12-24 часа после индукции ангиогенеза с помощью βFGF. Во-вторых, антагонисты αvβ3 нарушают ангиогенез, индуцированный множеством активаторов, что позволяет предположить, что такой рецептор принимает участие в общем каскаде реакций, протекающих вниз от возможно всех первичных событий по передаче сигнала, приводящих к ангиогенезу. В-третьих, CAM, обработанные mAb ML609 или циклическим пептидом, не проявляют значительного усиления апоптозиса, как было измерено лестничным анализом ДНК через 48 часов после обработки такими антагонистами. Наконец, антагонисты промотируют апоптоз сосудистых клеток уже подвергнутых индуцированию для вхождения в клеточный цикл.
Представленные в тексте результаты дают первое прямое доказательство того, что лигирование интегринов может регулировать выживание клетки in vivo. Поэтому можно высказать гипотезу, что после начала ангиогенеза индивидуальные сосудистые клетки начинают делиться и двигаться в направлении источника ангиогенеза, после чего αvβ3-лигирование обеспечивает сигнал непрерывному выживанию клетки, что приводит к дифференциации и образованию зрелых кровеносных сосудов. Однако, если αvβ3-лигирование предотвращается, то клетки не могут получать такой молекулярный сигнал и вовлекаются в апоптоз в недостаточном количестве. Такая гипотеза также предсказывает, что после дифференциации зрелые кровеносные сосуды больше не требуют αvβ3 передачи сигнала для выживания и они становятся устойчивыми в отношении антагонистов такого интегрина.
Наконец, представленные в тексте результаты дают доказательство того, что антагонисты интегрина αvβ3 могут обеспечивать мощный терапевтический подход к лечению неоплазии других заболеваний, характеризующихся ангиогенезом. Во-первых, антагонисты αvβ3 нарушают вновь образовавшиеся кровеносные сосуды, не оказывая влияния на уже существующую сосудистую систему. Во-вторых, такие антагонисты не оказывают значительного влияния на жизнеспособность куриных эмбрионов, что предполагает их нетоксичность. В-третьих, ангиогенез в значительной мере блокируется в независимости от природных ангиогенных стимулов. Наконец, систематическое применение αvβ3-антагонистов вызывает драматическую регрессию различных гистологических отличающихся человеческих опухолей.
Таким образом, приведенные выше Примеры демонстрируют тот факт, что интегрин αvβ3 играет ключевую роль в ангиогенезе, индуцируемом множеством стимулов, и такой αvβ3 представляет собой ценную терапевтическую мишень в присутствии αvβ3-антагонистов изобретения для заболеваний, характеризующихся неоваскуляризацией.
Приведенное выше описание можно считать достаточным для того, чтобы специалист в данной области мог реализовать изобретение на практике. Сфера настоящего изобретения не ограничивается депонированными клеточными линиями, поскольку депонированное воплощение рассматривается как одна из иллюстраций одного аспекта изобретения и любая клеточная линия, функционально эквивалентная описанной выше, входит в сферу изобретения. Депонирование материала не означает, что приведенное выше описание неадекватно для практической реализации любого аспекта изобретения, включая наилучшее его решение, также не имеется в виду, что этим ограничивается сфера формулы изобретения в отношении специфической иллюстрации того, что она представляет. Разумеется, что различные модификации изобретения помимо тех, что показаны и описаны в тексте станут ясными для специалистов в данной области из приведенного описания и они входят в сферу прилагаемой формулы изобретения.










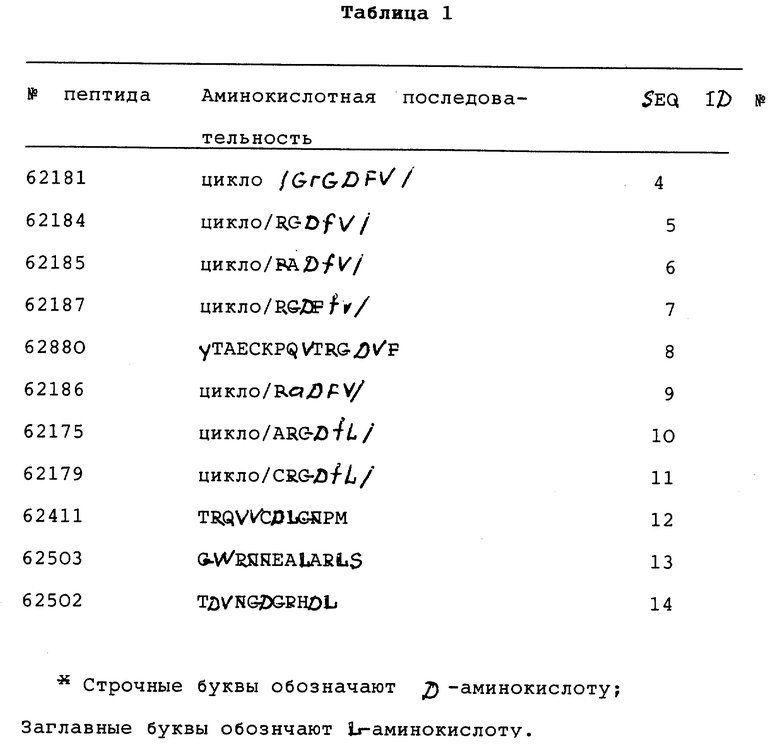
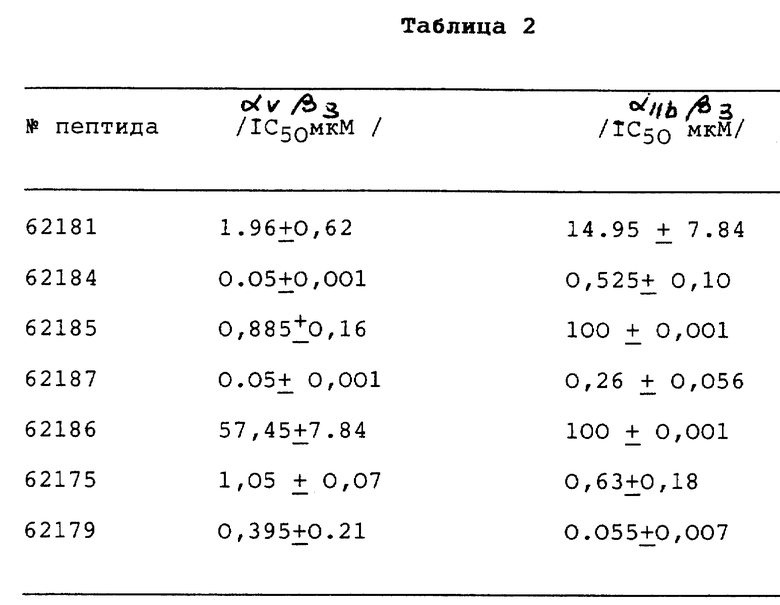
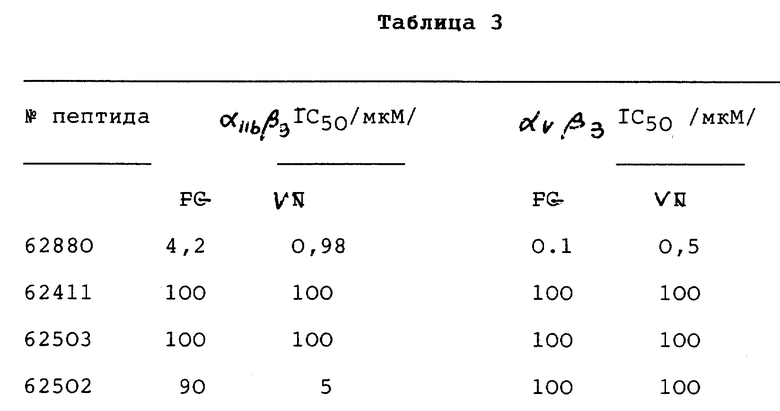


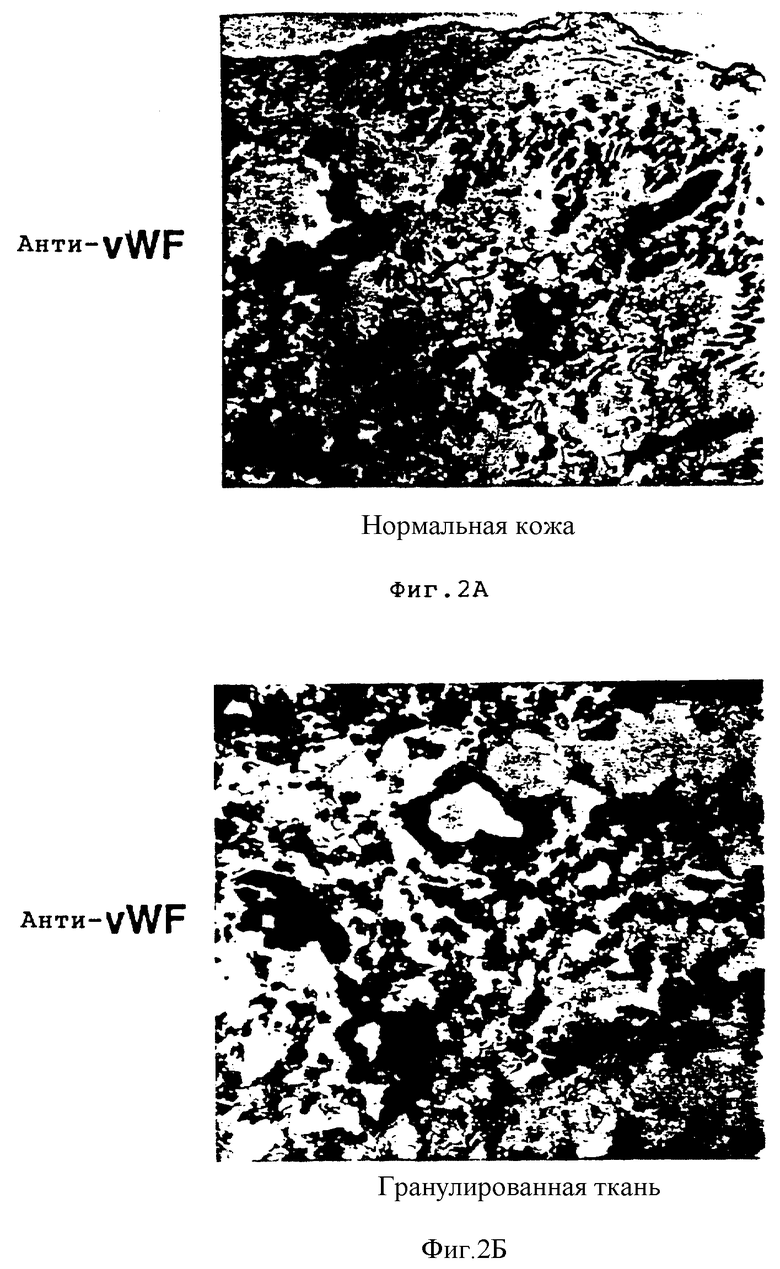
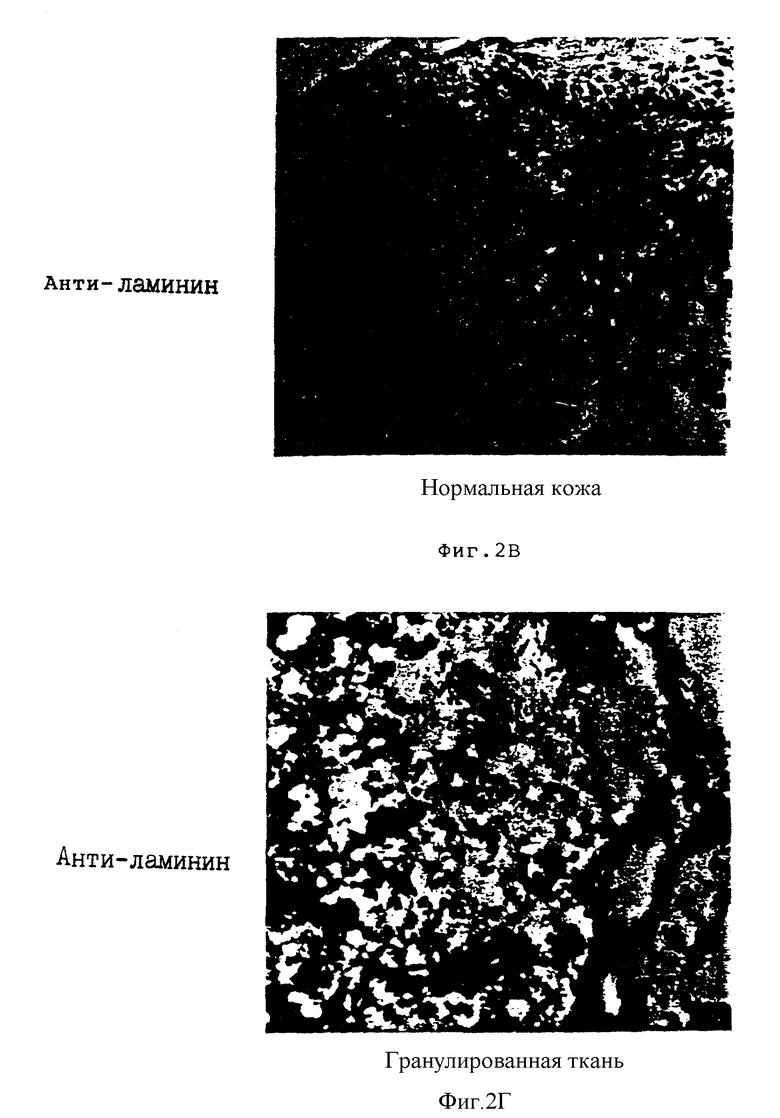



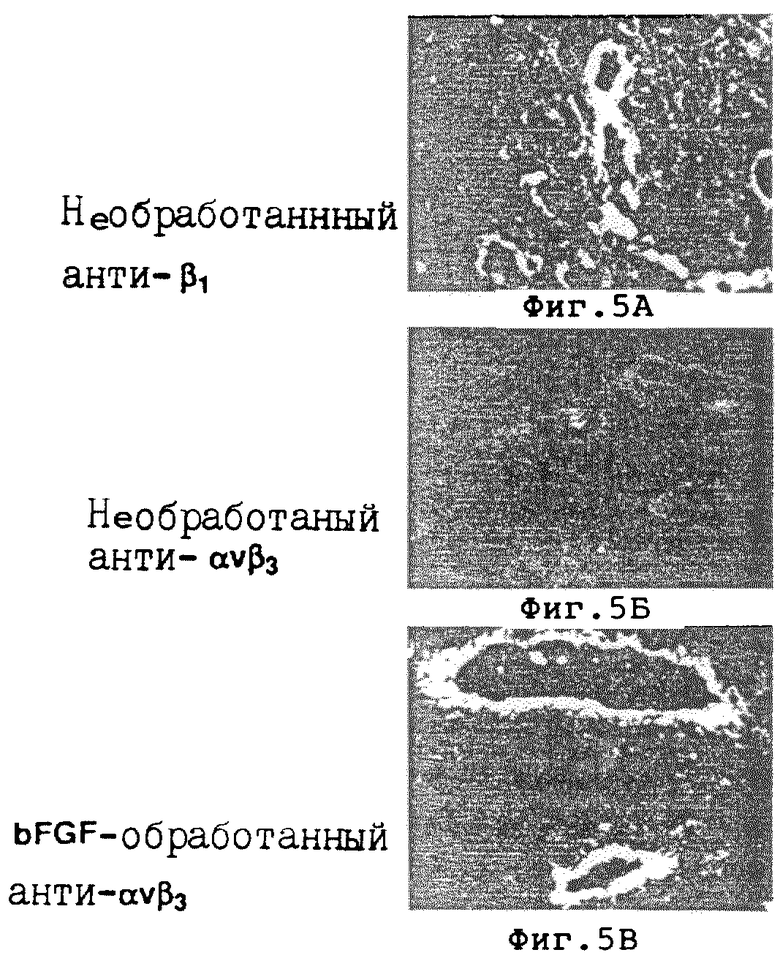
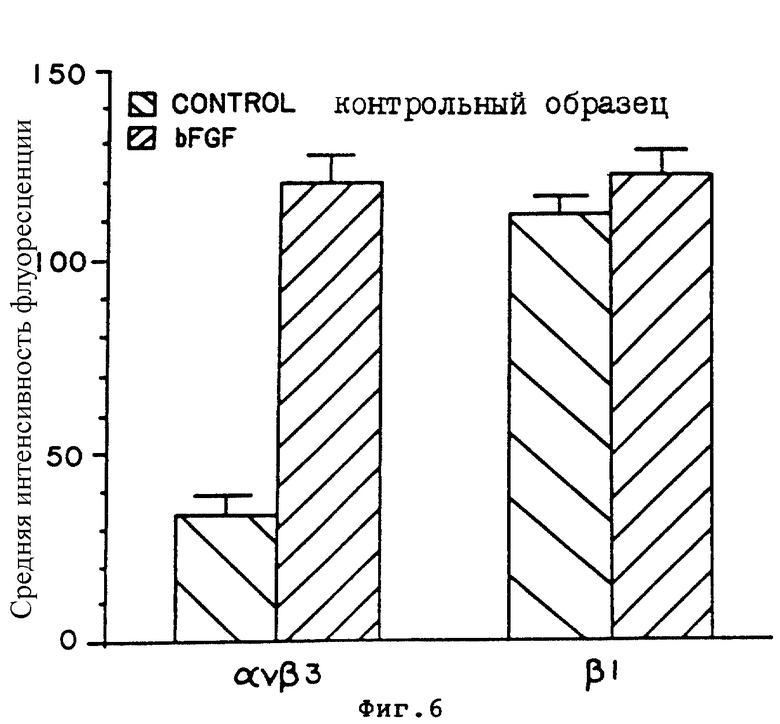

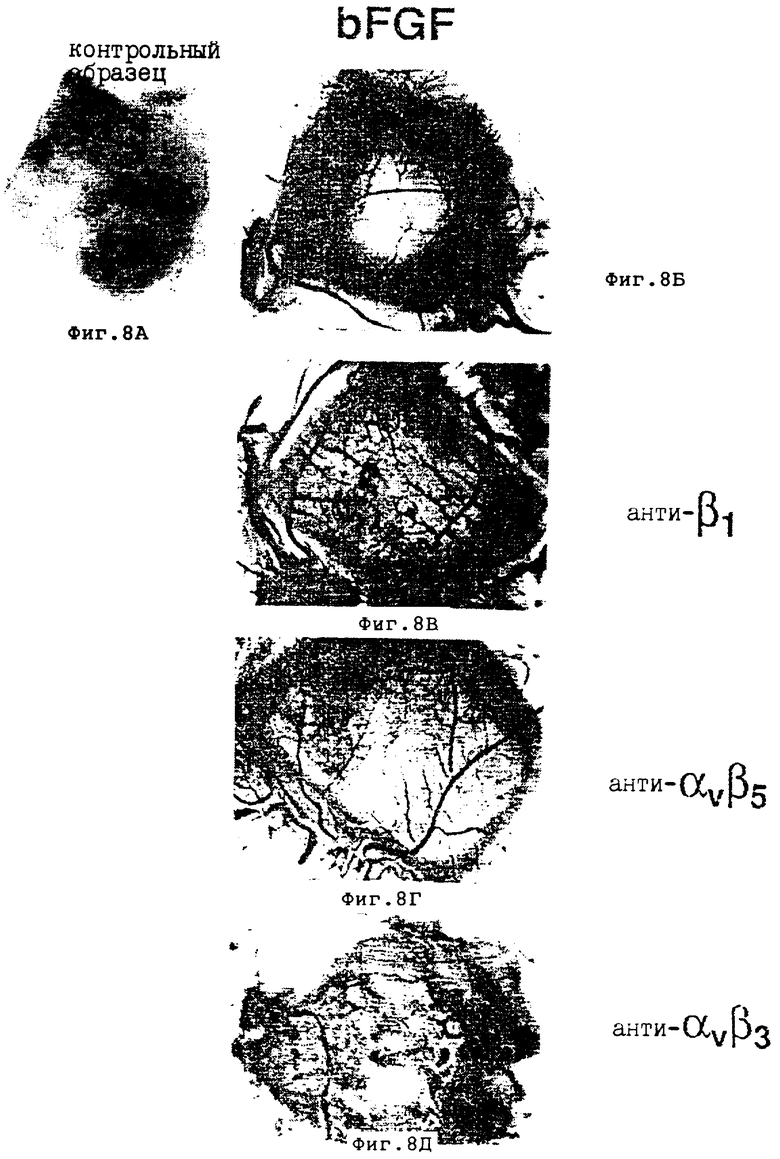

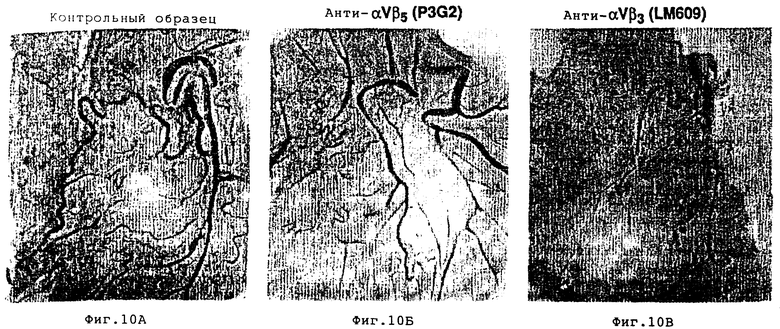
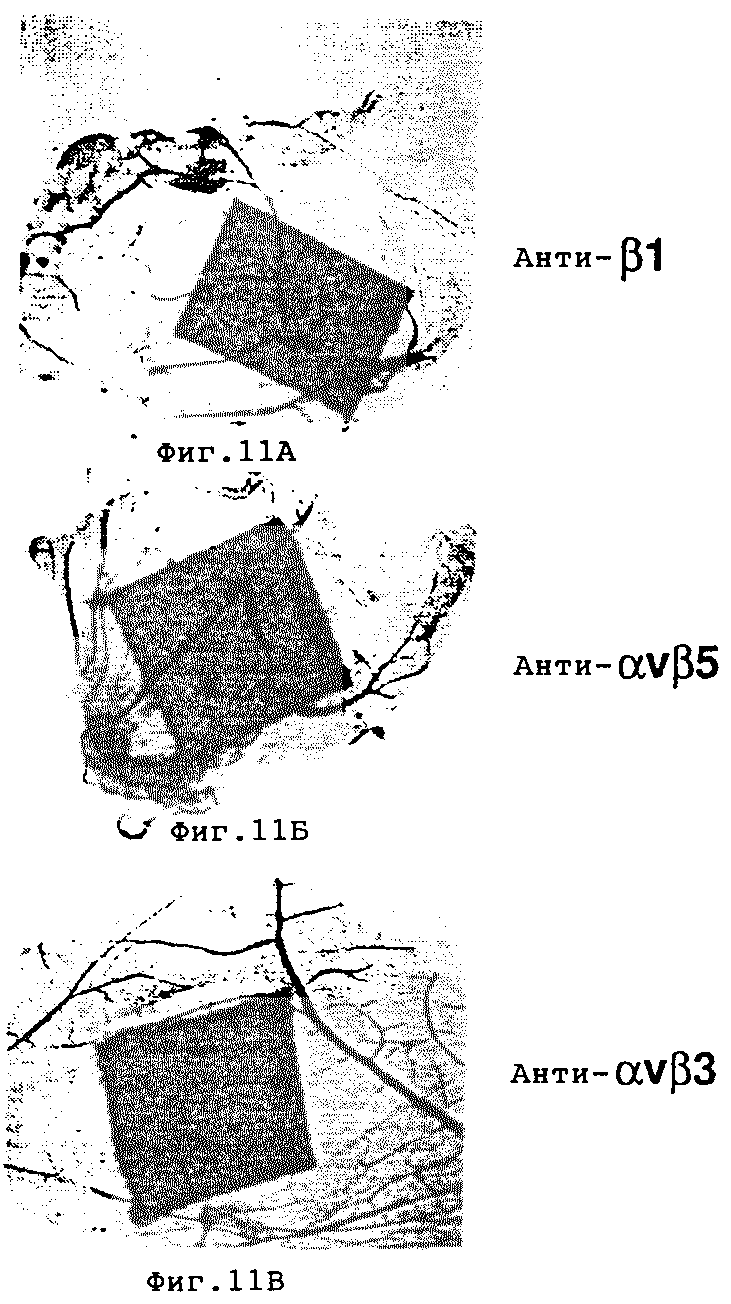
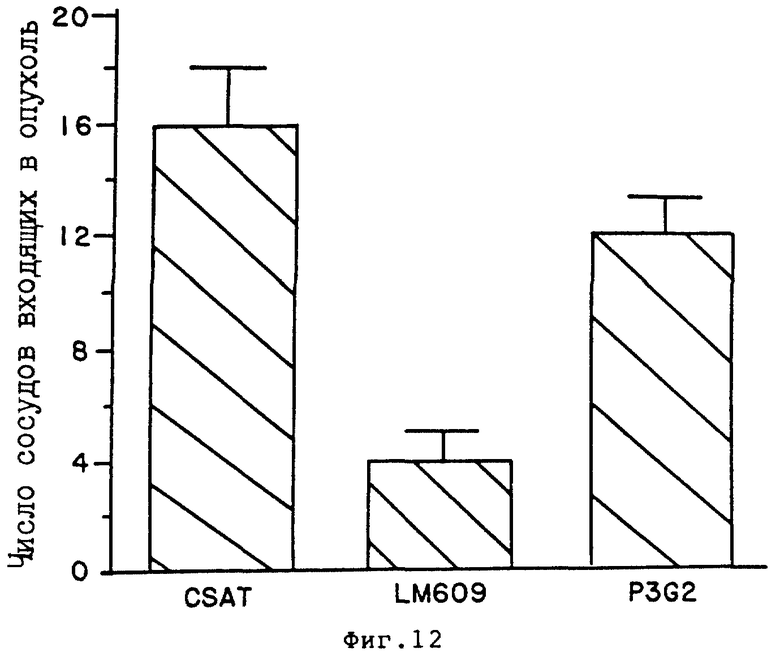
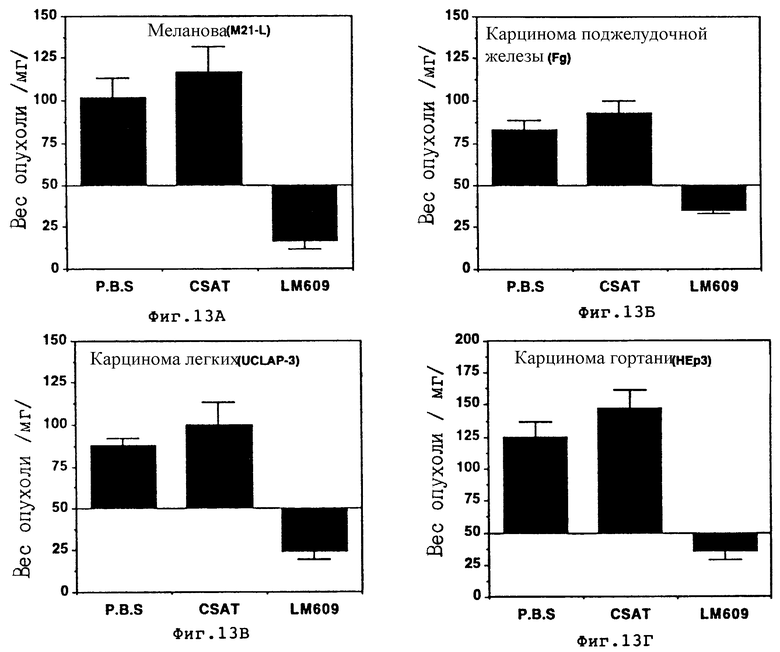
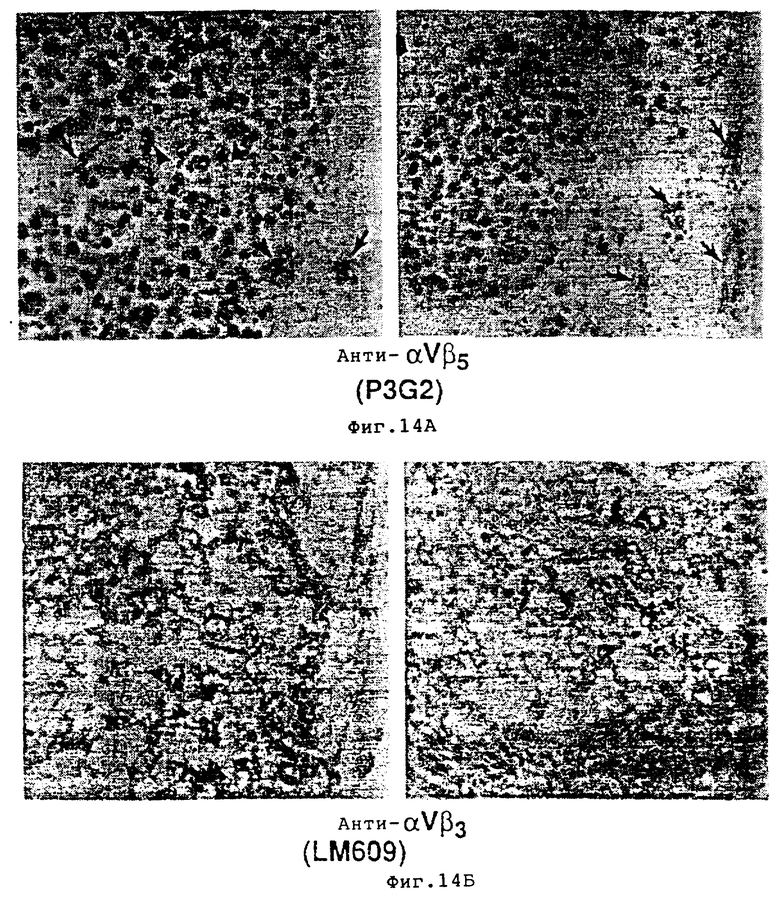
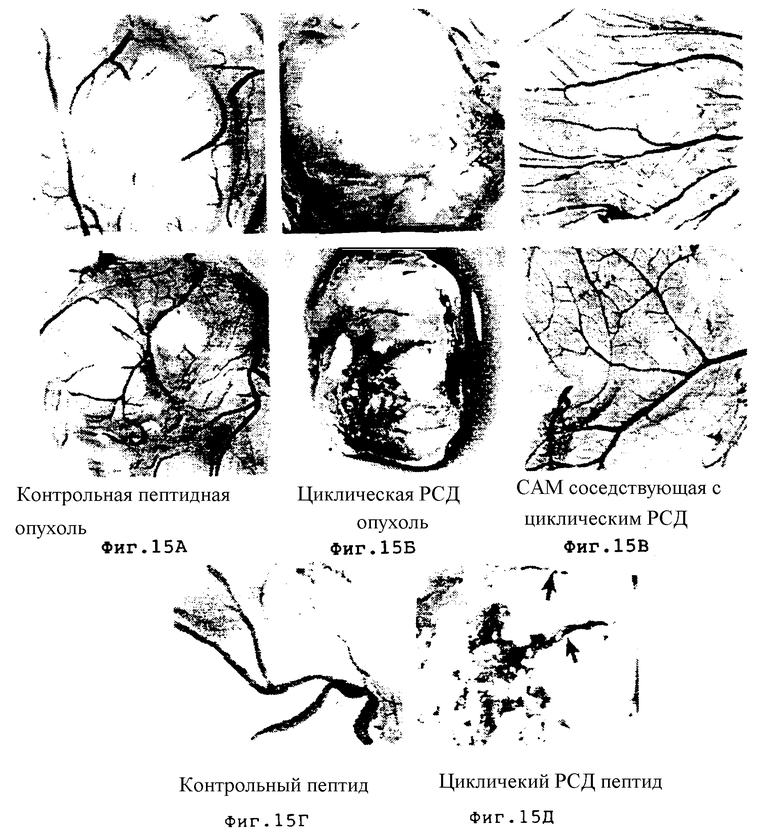
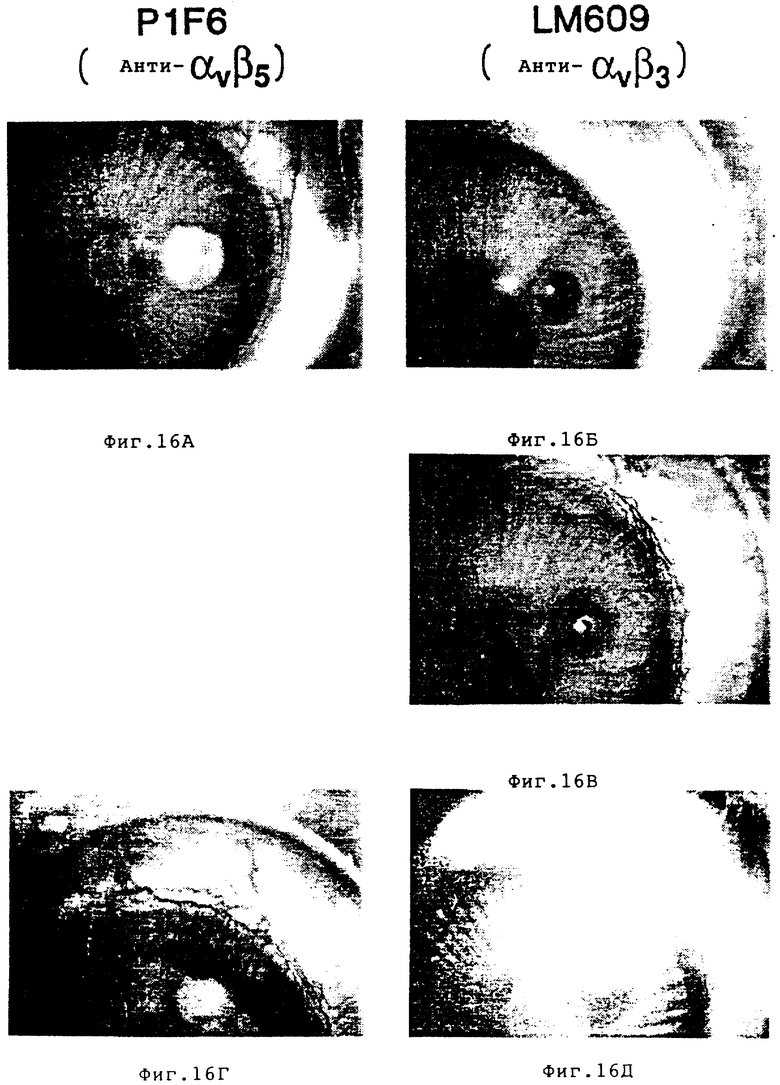
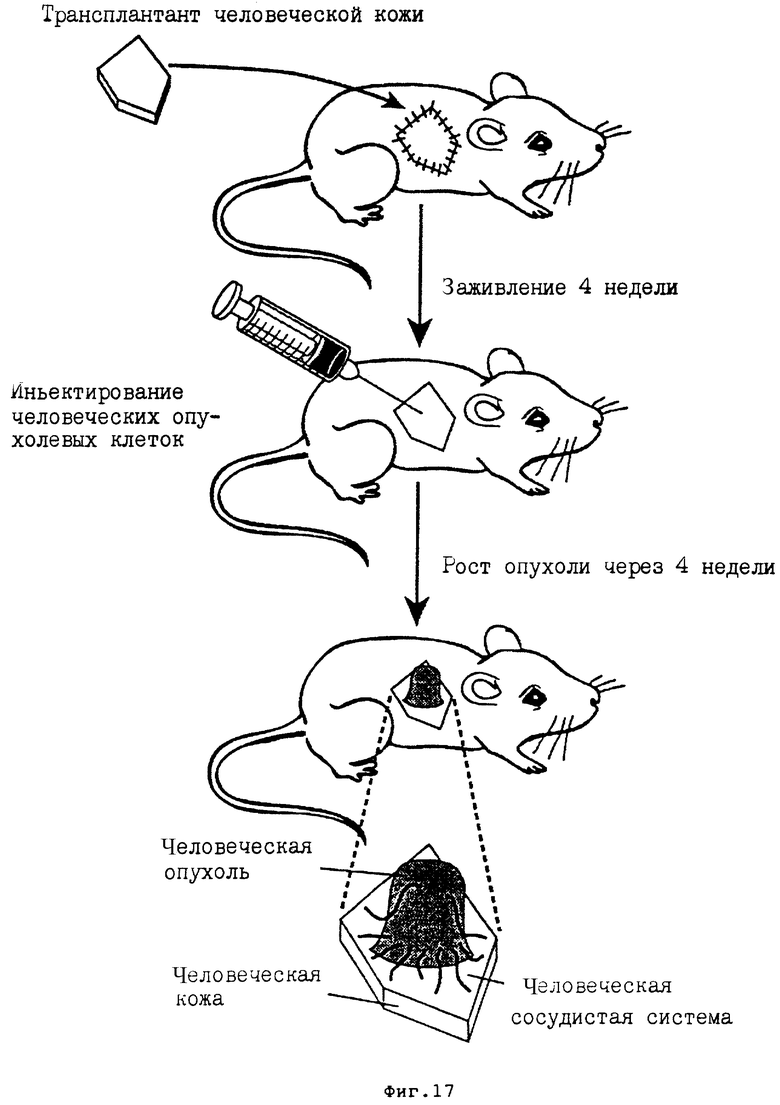
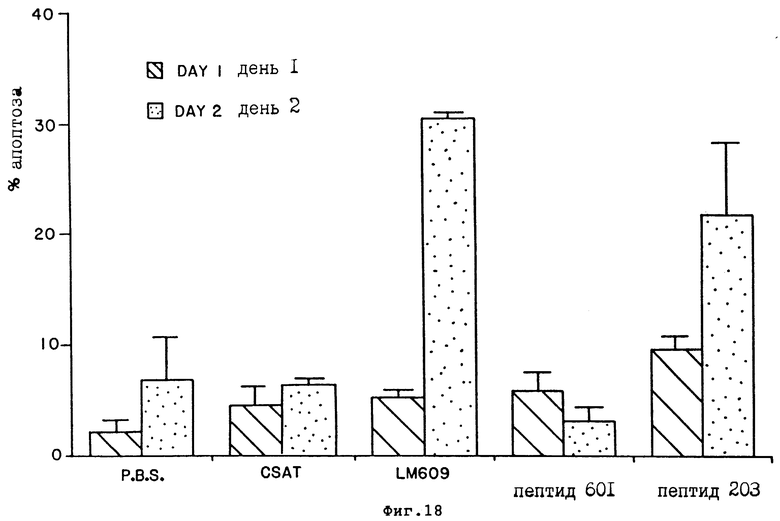
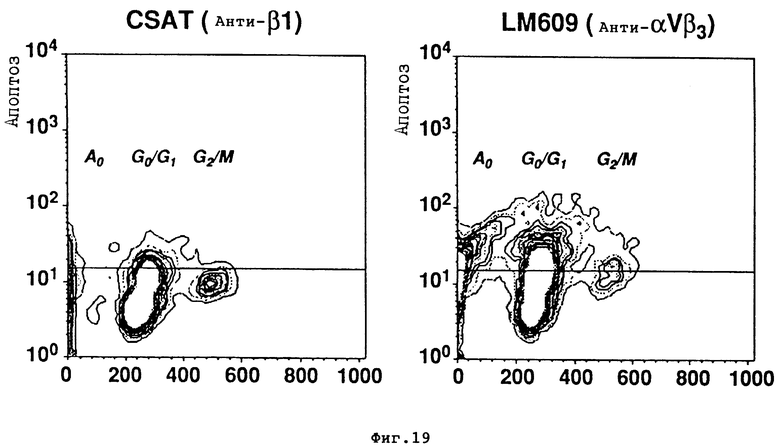


Изобретение относится к медицине и описывает способы ингибирования ангиогенеза в тканях с использованием витронектиновых антагонистов и, главным образом, ингибирование антиогенеза в воспаленных тканях и опухолевых тканях и метастазе с использованием терапевтических композиций, содержащих αvβ3-антагонисты, в частности анти-αvβ3 моноклональное антитело или RGD-содержащий полипептид. Способы позволяют ингибировать ангиогенез в тканях, где происходит неоваскуляризация, например в воспаленной ткани, твердой опухоли, матастазах, ткани, подверженной рестенозу. 6 с. и 30 з.п.ф-лы, 6 табл., 61 ил.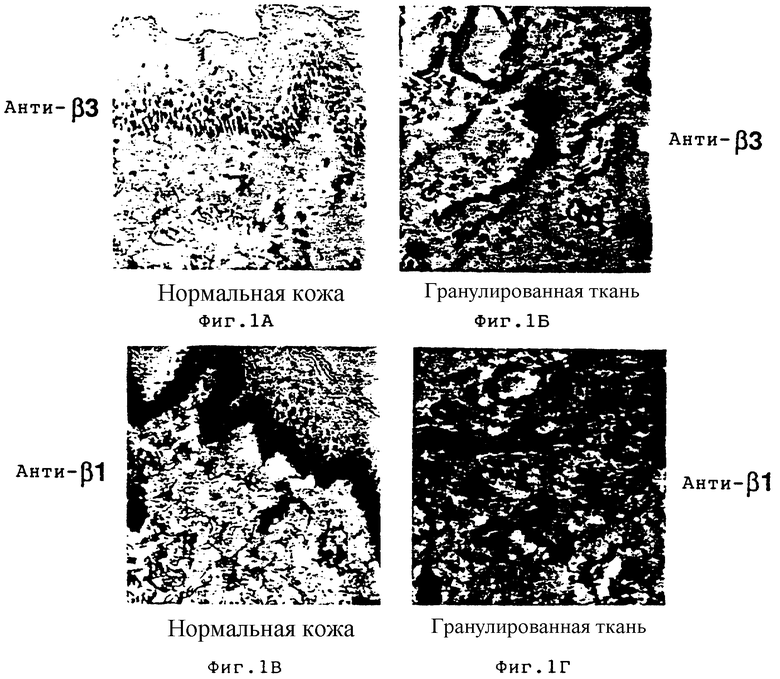
Приоритет по пунктам:
30.12.1994 по пп.7, 28, 34 и 36;
18.03.1994 - остальные пункты.
| Folkman M.et.al | |||
| Cancer Biology, 1992, 3:89-96 | |||
| Способ размножения копий рисунков, текста и т.п. | 1921 |
|
SU89A1 |

