Настоящее изобретение относится к способам скрининга свиней для выявления наличия или отсутствия аллелей остеопонтина (OPN), связанного с повышенной плодовитостью, к применению таких способов в предсказании плодовитости свиней и к наборам для осуществления таких способов.
Эффективность производства мяса и разведения соответствующих пород животных могла бы быть повышена, если бы было возможным повысить плодовитость животных. Одно и то же поголовье домашнего скота могло бы быть получено от меньшего числа родительских пар животных, что приводит к уменьшению расходов. Кроме того, организации, занимающиеся выведением пород домашнего скота, будут извлекать выгоду из возможности оценивать большее число животных с целью отбора особей с улучшенными генетическими показателями. Однако плодовитость обычно является трудным критерием для селекции, т.к. она ограничена одним полом и на нее почти не влияют негенетические факторы (способность наследования, мера фенотипического изменения, являющегося следствием генетических различий, составляет приблизительно 0,1 в расчете на объем приплода свиньи).
Один подход для увеличения объема приплода может быть в введении нужных генов в производство пород скота, обладающих значительно повышенной плодовитостью. Однако количественная генетика наводит на мысль о том, что комплексные признаки, такие как объем приплода, контролируются большим количеством генов, каждый из которых сам по себе обладает небольшим влиянием на этот признак. Если это так, то похоже, что генетическое улучшение посредством выведения комплексных признаков будет очень медленным. Альтернативная точка зрения заключается в том, что хотя много генов формируют комплексные признаки, значительные эффекты оказывают некоторые гены (главные гены). Если этот альтернативный взгляд верен, тогда генетическое улучшение таких свойств могло бы быть быстрым при условии, что возможно идентифицировать и отобрать для нужных аллелей соответствующие главные гены.
С появлением геномного картирования стало возможным идентифицировать гены, воздействующие на наследственные признаки (локусы наследуемых признаков, QTL) поиском связей между признаком и молекулярными маркерами, распределенными равномерно в геноме животных, в отношении которых проведено картирование. Для селекционных целей важно, что наследуемость таких маркерных фенотипов составляет 1,0.
Известно, что китайская Мейшанская (Meishan) порода свиней дает около 4 дополнительных поросят в каждом помете в сравнении с наиболее плодовитыми европейскими породами. Гены плодовитости (объема приплода) от этой породы должны иметь огромное значение в программах, направленных на повышение плодовитости коммерческих западных пород свиней. Действительно, было показано, что генетический маркер, связанный с геном рецептора эстрогена (ESR) Мейшанской свиньи, показывает благотворный эффект на объем приплода, что описано в WO 92/18651.
Порода овцы Booroola Merino является исключительно плодовитой. Обычная численность приплода составляет три или более. Было показано, что существенно повышенная плодовитость этой овцы является следствием действия единичного гена, FECB (см. GW Montgomery, et. al, Endocrine Reviews, 13: 309-328 (1992)). Генетическое картирование с использованием ДНК маркеров человека показал, что аналог FECB человека локализован на хромосоме 4 (GW Montgomery, et. al, Nature Genetics, 4: 410-114 (1993)) и тесно связан с геном, кодирующим секретируемый фосфопротеин-1 (SPP-I), известный также как остеопонтин (OPN), 2ar, сиалопротеин -1 кости, 44 кДа фосфопротеин кости и фосфопротеин, выделенный из опухоли. Сравнительное картирование (H. Ellegren, et. al, Genomics, 17: 599-603 (1993)) показало, что человеческая хромосома 4 и хромосома 8 свиньи в высшей степени подобны (синтеничны). SPP-1 ген свиньи также локализован на хромосоме 8.
Совсем недавно было показано, что FECB-связанный маркер крупного рогатого скота не действует как маркер повышенной плодовитости в стаде животных, отобранных по повышенной скорости овуляции (Blattman et al, Mid-West Animal Science Meeting, 18:43 (1995)).
Однако нами неожиданно обнаружено, что у свиней определенные ДНК маркеры для OPN связаны с объемом приплода и могут быть таким образом использованы для скрининга свиней, обладающих лучшей возможностью приносить более многочисленный приплод, и выбраковывать свиней с аллелями, дающими меньшую численность приплода. Приводимое здесь выражение "увеличенный объем приплода" обозначает значительное увеличение численности приплода по сравнению с данной величиной у взятой популяции.
Интересно отметить, что наблюдается явная точка нарушения в хромосомной синтении в области OPN между овцой, крупным рогатым скотом и человеком, с одной стороны, и мышью и свиньей, с другой (Montgomery et al, J. Reproduction and Fertility supplement, 49: 113-121 (1995)). Это наводит на мысль, что структура хромосомы может изменяться в этом участке между животными, имеющими большие приплоды (мышь и свинья) и животными, дающими небольшие приплоды (человек, овца, корова), так что эффект главного гена плодовитости изменяется. Возможные объяснения заключаются в том, что экспрессия главного гена может быть увеличена или уменьшена включением в более транскрипционно активный или неактивный участок; главный ген может быть включен непосредственно под контролем измененного промоторного элемента; положение главного гена относительно OPN может быть изменено таким образом, что OPN становится более удобным для использования в качестве маркера при оценке потенциала плодовитости свиньи, чем овцы или коровы.
Таким образом, данное изобретение относится, во-первых, к способу скрининга свиней для определения особей с более вероятной повышенной плодовитостью и/или особей с менее вероятной повышенной плодовитостью, причем этот способ включает следующие стадии;
(i) взятие пробы геномной ДНК от свиньи; и
(ii) исследование геномной ДНК, полученной на стадии (i), для определения, какая OPN аллель (аллели) присутствует/присутствуют.
Соответственно, стадии (ii), а именно - определение OPN аллелей, осуществляются путем поиска особых ДНК маркеров, непосредственно или косвенно связанных с OPN.
Связь между генетическими маркерами и генами, ответственными за особый признак, может быть разрушена генетической рекомбинацией. Таким образом, чем меньше физическое расстояние между маркером и рассматриваемым геном, тем менее вероятно, что при рекомбинации произойдет их разделение.
Также возможно установить связь между специфичными аллелями альтернативных ДНК маркеров и аллелями ДНК маркеров, связь которых с особым геном известна (например, упомянутый здесь OPN ген), относительно которого предварительно было установлено его влияние на особый признак. Таким образом, в данной ситуации, рассматривая OPN ген, может быть возможным, по крайней мере, в короткий период времени отобрать свиней с более вероятной повышенной плодовитостью или, напротив, более низкой плодовитостью опосредованно путем селекции определенных аллелей OPN-ассоциированного маркера в результате отбора специфических аллелей альтернативных маркеров хромосомы 8. Примерами таких маркеров, в отношении которых известно, что они связаны с OPN на хромосоме 8 свиньи, являются Sw 61, Sw 1085, Sw 194, Sw 16, Sw 790 и SO 178, все маркеры являются микросателлитами.
В частном варианте осуществления изобретения используется ряд таких маркеров. Например, пары маркеров можно использовать для установления пределов диапазона главного гена, чтобы уменьшить какие-либо возможные эффекты рекомбинации. Примеры таких комбинаций маркеров включают SO 178 и Sw 61, а также SO 178 и Sw 790.
Так как эффект может быть отнесен к различию в структуре гена свиней (или мышей) и овцы (и человека и крупного рогатого скота), это дает возможность предположить, что наиболее полезный второй маркер будет в негомологичном (несинтеничном) участке 8-й хромосомы свиньи. Примером известной подходящей комбинации маркеров, устанавливающей пределы этого участка, является OPN и SO 178. Однако квалифицированный специалист поймет, что легко могут быть найдены другие полезные маркеры.
Специфический генетический маркер, ассоциированный с ОРN, является микросателлитом. Они являются простыми повторами последовательностей из 4,3 или, более часто, 2 нуклеотидов, которые располагаются в основном в пространстве вокруг генома и в целом включают приблизительно 50000 оснований (около 60000 микросателлитов на гаплоидный геном). По-видимому, сбои ДНК-полимеразы во время репликации и неравный кроссинговер во время рекомбинации приводит к утрате или увеличению повторяющихся единиц. Это означает, что микросателлиты обычно полиморфны и могут иметь несколько повторяющихся протяженных аллелей. Примером микросателлита, ассоциированного с данным геном является (CA)n, приводящий к возможному повторению структурных фрагментов аллелей, например (CA)2, (CA)9, (CA)10, (CA)11 и (CA)12.
С использованием праймеров, способных к гибридизации (например, в строгих условиях) с участками, фланкирующими микросателлит, ассоциированный с рассматриваемым геном, в сочетании со стандартными PCR методиками можно получить PCR продукты различной длины, которая будет зависеть от конкретной повторяющейся структурной протяженностью аллели микросателлита.
Исследование связи таких PCR продуктов, полученных с использованием микросателлита, связанного с OPN геном, с объемом приплода, позволило определить аллели маркерной длины, связанные с увеличенной или пониженной плодовитостью у свиней.
Подходящие праймерные пары, которые будут гибридизоваться с фланкирующими участками таких микросателлитов, включают пары, имеющие следующие последовательности:
(GCTAGTTAATGACATTGTACATAA; или
CCAATCCTATTCACGAAAAAGC; и
GTGTCATGAGGTTTTTTCCACTGC; или
CAACCCACTTGCTCCCAC.
В частности, было установлено, что аллели с повторяющейся фрагментарной последовательностью вышеуказанного маркера микросателлита, обозначенные как 132 и 136, связаны с повышенной плодовитостью свиней. Кроме того, установлено, что аллель, обозначенный как 112, связан с пониженной плодовитостью свиней.
Фактически аллель, связанный с повышенной плодовитостью, преимущественно выведен из европейского поголовья скота. Это противоречит ожиданиям по той причине, что, как обсуждалось ранее, Мейшанская порода приносит на четыре поросенка больше в одном опоросе, чем порода Landrace или порода Duroc, и в связи с этим можно предположить, что нужные маркеры должны быть связаны с генами, не наследуемыми от породы Мейшан.
Вторым аспектом настоящего изобретения является способ скрининга свиней для определения особей с более вероятной повышенной плодовитостью и/или особей с менее вероятной повышенной плодовитостью, включающий стадии:
(i) взятие пробы геномной ДНК от свиньи;
(ii) гибридизация геномной ДНК, полученной на стадии (i) с одним или более подходящими праймерами;
(iii) осуществление одного или более циклов PCR с использованием гибридной нуклеиновой кислоты со стадии (ii); и
(iv) анализ длины PCR продуктов, полученных на стадии (iii).
Соответственно, способы настоящего изобретения выполняют с использованием реагентов и инструкций, присутствующих в форме набора.
Таким образом, третий аспект настоящего изобретения касается набора для скрининга свиней для определения особей с более вероятной повышенной плодовитостью и/или особей с менее вероятной повышенной плодовитостью, который включает один или несколько реагентов или материалов, пригодных для идентифицирования OPN аллелей в пробе геномной ДНК свиньи.
Предпочтительный набор по изобретению будет включать реагенты или материалы, пригодные для идентифицирования аллелей, связанных с ДНК маркерами, присоединенными к OPN гену, например, микросателлитным маркером. Такой набор будет предпочтительно содержать один или более ДНК праймеров, произвольно объединенных со стандартными PCR реагентами.
В конце концов специалисту будет понятно, что описанные способы и наборы могут быть использованы в сочетании с другими уже описанными методами и наборами для отбора свиней с более вероятной повышенной плодовитостью (или свиней с менее вероятной повышенной плодовитостью). Примеры других способов и наборов описаны в WO 92/18651. Конечно, предполагается, что возможно создание комбинированных наборов, которые могли бы быть использованы для оценки ДНК свиньи с использованием обоих способов.
В WO-A-92/18651 и USSN 08/312312 раскрыты способы определения свиней с более вероятной повышенной плодовитостью, основанные на связи с ESR геном. Специалисту будет понятно поэтому, что оценочные способы данного изобретения могут сочетаться с ранее раскрытыми ESR - оценочными способами, что обеспечивает еще более мощное средство для таких исследований. Таким образом, дальнейший аспект настоящего изобретения касается способа скрининга свиней для определения особей с более вероятной повышенной плодовитостью и/или особей с менее вероятной повышенной плодовитостью, который включает следующие стадии:
(i) взятие пробы геномной ДНК от свиньи;
(ii) исследование геномной ДНК, полученной на стадии (i) для определения, какая OPN аллель (аллели) присутствует/присутствуют; и
(iii) исследование геномной ДНК, полученной на стадии
(i) для определения, какой аллель (аллели), по крайней мере, какого-либо иного гена, связанного с повышенной плодовитостью свиней, присутствует/присутствуют.
В предпочтительном варианте осуществления данного способа аспекта изобретения "какой-либо другой ген" является, по крайней мере, ESR геном, как описано в WO-A-9218651 и USSN 08/312312.
Последний аспект настоящего изобретения касается набора для скрининга свиней для определения особей с более вероятной повышенной плодовитостью и/или менее вероятной повышенной плодовитостью, который включает один или более реагентов или материалов, пригодных для идентифицирования OPN аллелей в пробе геномной ДНК свиньи, вместе с одним или несколькими реагентами или материалами, пригодными для идентифицирования аллелей, по крайней мере, какого-либо другого гена, связанного с повышенной плодовитостью свиней, в пробе геномной ДНК свиньи.
Предпочтительные особенности каждого аспекта изобретения являются подходящими для mutatis mutandis.
Далее будет описано изобретение с приведением нижеследующих примеров, которые не должны истолковываться как ограничивающие объем изобретения.
Пример 1
Получение ДНК
ДНК может быть получена из любой исходной ткани, содержащей клеточные ядра, например, лейкоцитов, волосяных фолликулов, ткани уха и мышечной ткани. Процедура, кратко описанная здесь, касается подготовки клеток крови; другие ткани могут быть также обработаны непосредственным суспендированием материала в K буфере с последующим продолжением от той же самой стадии подготовки крови. Описанный здесь метод позволяет получать клеточный лизат, содержащий ДНК, которая пригодна для PCR амплификации. Однако любой метод приготовления очищенной или грубой ДНК должен быть в равной степени эффективным.
Кровь следует поместить в 50 мМ ЭДТА pH 8,0 для предотвращения коагуляции. 50 μл крови распределяют в маленькую пробирку для микроцентрифуги (0,5 мл Eppendorf или эквивалентная). Добавляют 450 μл TE буфера, чтобы лизировать красные кровяные тельца (гемовые группы ингибируют PCR) и смесь вращают в течение 2 секунд. Затем интактные белые и остаточные красные кровяные тельца были отцентрифугированы в течение 12 сек при 13000 q в микроцентрифуге. Супернатант был удален мягким отсасыванием с использованием вакуумного насоса с пониженным давлением. Другие 450 μл TE буфера добавляют, чтобы лизировать оставшиеся красные кровяные тельца и белые кровяные тельца, собранные центрифугированием, как и прежде. Если в осадке осталось красное окрашивание, процесс повторяют до тех пор, пока осадок не будет белым. После удаления последней капли супернатанта из осажденных белых кровяных телец, добавляют 100 μл K буфера, содержащего протеиназу К, и смесь инкубируют при 55oC в течение 2 часов. Затем смесь нагревали до 95-100oC в течение 8 минут и ДНК-содержащие лизаты хранили при -20oC до востребования.
Реагенты
ТЕ буфер:
10 мМ Трис-HCl pH 8,0
1 мМ ЭДТА
К буфер:
50 мМ KCl
10 мМ Трис-HCl pH 8,0
2,5 мM MgCl2
0,5% Tween 20
Перед использованием лизатов на 1,0 мл буфера К добавляли 10 μл раствора протеиназы К (Boehringer Mannheim) с концентрацией 20 мг/мл.
PCR
Реакции проводили как описано выше, в тонкостенных 0,25 мл пробирках (Perkin Elmer):
1,5 μл 10 х буфер;
1,5 μл 15 мМ MgCl2
1,5 μл 2 мМ dNTPs (Pharmacia);
0,5 μл каждого праймера при 5 мМ (Genosys);
9 μл стерильной деионизированной воды;
0,1 μл (0,5 единицы) (Ampli Tag ДНК-полимеразы Perkin Elmer);
1 μл ДНК-лизат.
Реакционные пробирки помещали затем на Perkin Elmer 9600 термальный циклер и проводили PCR в соответствии с показанным ниже режимом:
94o в течение 4 минут;
30 циклов 94oC в течение 30 секунд;
58oC в течение 1 минуты;
и 72oC в течение 1 минуты;
72oC в течение 4 минут;
4oC пока не востребуется.
Реагенты
10 х PCR буфер
100 мМ Трис-HCl pH 8,3 (250, 500 мМ KCl)
Прямой праймер
GCTAGTTAATGACATTGTACATAA или CCAATCCTATTCACGAAAAAGC
Обратный праймер
GTGTCATGAGGTTTGTGCCACTGC или CAACCCACTTGCTCCCAC
Если один из праймеров помечен флуоресцентным маркером, полученные продукты могут быть подвергнуты анализу на автоматическом секвенаторе ДНК, таком как Applied Biosystem 373 DNA Sequencer, с помощью систем Genescan и Genotyper с мягким носителем.
Пример 2
Электрофорез в полиакриламидном геле.
5 μл PCR продуктов смешивали с 2 μл рабочего буфера и разделяли на неденатурирующем полиакриламидном вязком геле в 1 х ТВЕ буфера при 100 V в течение 4 часов. Затем гель окрашивали в 50 нг/мл этидиум-бромида в течение 30 минут и PCR продукты рассматривали и фотографировали в ультрафиолетовом световом трансиллюминаторе. Размеры PCR продуктов в основных парах рассчитывали по их относительной подвижности в сравнении с аналогичными показателями маркеров с известной молекулярной массой в том же самом геле. Рассчитанный размер PCR продуктов отражает длину аллели микросателлита.
PCR продукты были также исследованы на Applied Biosystems DNA Секвенаторе при использовании в PCR флуоресцентно меченного праймера.
Результаты
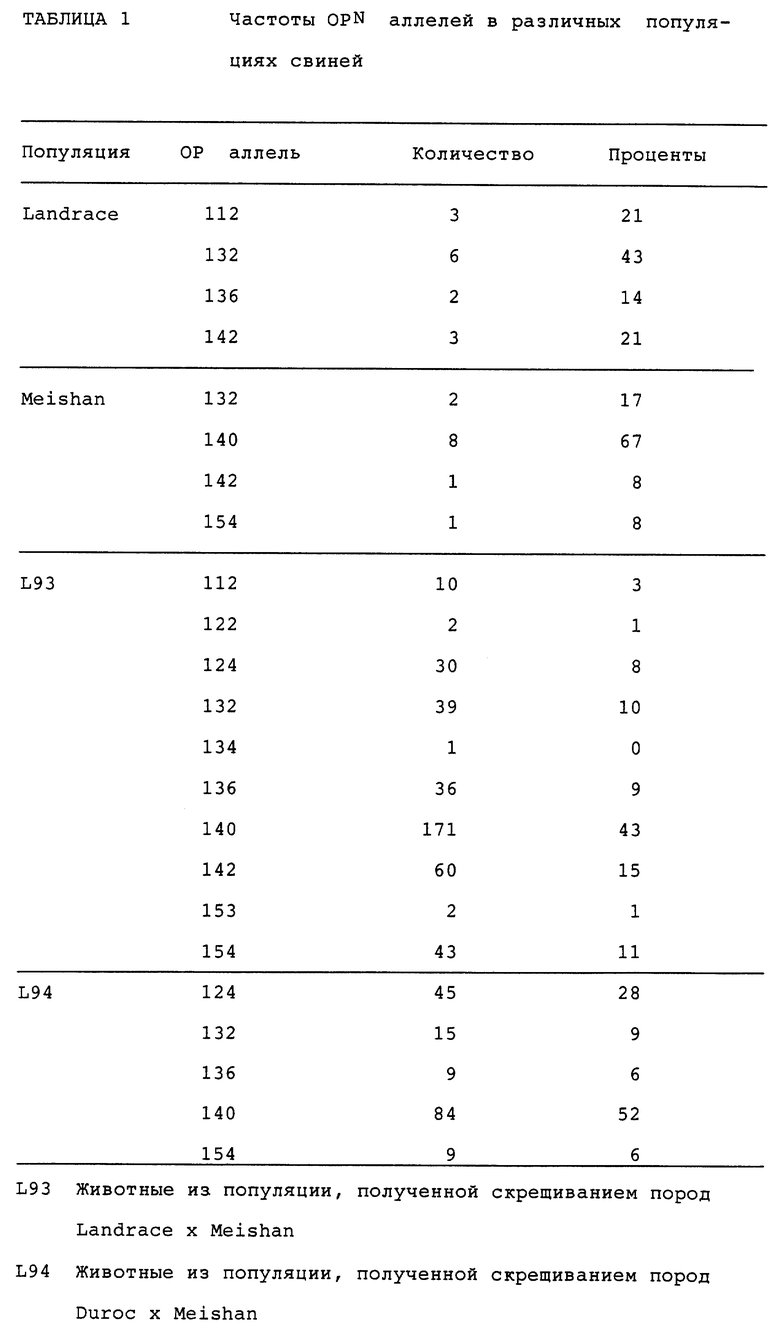
Частоты OPN аллелей
В таблице 1 приводится повторяемость (частота) результатов OPN аллелей в различных популяциях свиней.
Статистический анализ
Самки животных, полученные от L93 и L94, обсчитывались по плодовитости (как по общему количеству родившихся в приплоде (TNB), так и по количеству родившихся живыми (NBA)), в нескольких повторностях, если это возможно, и эти данные сравнивали с генотипами OPN микросателлитов для того же набора животных. Статистические связи между плодовитостью и OPN генотипами были исследованы с помощью метода наименьших квадратов для построения главной линейной модели. Для каждого OPN генотипа были рассчитаны значения наименьших квадратов по объему приплода (LSM-ы). LSM-ы представляют собой значения, установленные для других эффектов в модели, которые могли влиять на плодовитость.
Влияние индивидуальных OPN аллелей далее разбирали с использованием модели аллельного замещения, в которой животные были классифицированы по группам в зависимости от того, несли ли они 0, 1 или 2 копии специальной аллели. Для каждой группы вычисляли LSM-ы плодовитости. Результаты для L93 показаны в таблице 2.
Исходя из приведенных данных видно, что аллель 112, по-видимому, связан с негативным эффектом в плане плодовитости, в то время, как позитивные тенденции наблюдаются для аллелей 132 (NBA) и 136 (TNB). Результаты, представленные в таблице 1, наводят на мысль, что, хотя аллели 112 и 136 были, вероятно, получены от породы Landrace, аллель 132 мог быть получен от предков или породы Landrace, или породы Meishan. Однако, так как аллель 132 более чем вдвое превышает общие данные Landrace и Meishan, по всей видимости, значительная доля аллелей 132 в L93 наследована от породы Landrace.
Для того, чтобы исследовать возможность действия этих аллелей в качестве предсказателей объема приплода, анализу были подвергнуты дополнительные данные от L94. В этой линии аллель 112 не обнаруживался (по-видимому, этот аллель отсутствовал у породы Duroc). Объединенные результаты по аллелям 132 и 136 показаны в таблице 3.
Эти данные показывают, что только аллель 132 оказывает существенное положительное влияние на объем приплода как по TNB, так и по NBA. Хотя аллель 136 был близок по значениям TNB, возможно, что данный эффект здесь обусловлен небольшим количеством животных (3) из 136/136 находящихся под тщательным наблюдением.
Ассоциация между OPN аллелем 132 и большим объемом приплода продемонстрирована на двух различных линиях свиней (L93 и L94). Это показывает, что QTL, касающийся объема приплода, тесно связан со свиным OPN геном. Тем не менее, возможно, что в других семействах, линиях и породах свиней с повышенным объемом приплода будет связан другой OPN аллель.
Результаты еще одного анализа данных, полученных на основе L93 и L94 и дополнительной линии L07 (большая белая линия) показаны в таблице 4 с использованием альтернативной модели. Она включает значения соответствий каждого OPN аллеля как вариабельного и кодирующего каждое животное с 0, 1 и 2 для каждого аллеля (т.е. 0, 1 или 2 копии каждого аллеля).
Фиксируемыми эффектами были тип "стадо-сезон-обслуживание" и способность приносить потомство. Производитель включался как случайный эффект. ESR и OPN подгонялись для совариабельности. Включались все данные линии, не только единокровные и полуединокровные семейства. OPN аллели с менее чем 10 приплодами второго генотипа исключались из анализа.
Анализируемыми признаками были общее число родившихся (TNB) и число родившихся живыми (NBA).
Для каждой линии испытывались три модели, включая фиксируемый, случайный и ESR эффекты, как указано выше.
1. Модель с исключением OPN.
2. Модель, включающая все OPN аллели.
3. Модель, включающая OPN аллели индивидуально.
- 2 log вероятность (2 log liklihood) была получена для каждой модели. Значение для модели вычисляли путем вычитания log вероятности (log liklihood) от модели 2 или 3 из модели 1 и сравнивая полученный результат с распределением Chi-квадратов. Использованные степени свободны (df)-разность между двумя моделями.
Уровни значимости на линию для модели 2 и какие-либо значимые аллели в модели 3 приводятся в таблице 4.
Из приведенных данных могут быть сделаны следующие выводы:
1. OPN обусловила значительную степень изменения плодовитости (после включения ESR) для L07 (P < 0,10); L93 (TNB: P < 0,10; NBA: P < 0,05) и L94 (TNB; p < 0,01; NBA: p < 0,05).
2. OPN аллель 132 показала значительный положительный эффект в отношении плодовитости для L93 и L94.
3. Другие аллели OPN 122 (L07), OPN 112 и OPN 154 (L93) и OPN 124 (L94) показали значительные отрицательные эффекты.
Пример 3
Были получены и исследованы пробы геномной ДНК от другой линии L03 (другая линия, основанная на большой белой). Результаты показаны в таблице 5. Было исследовано 416 животных с рекордным приплодом 1010.
Испытывали несколько различных моделей. Для всех моделей учитывали эффект, обусловленный местом и временем, где и когда происходил опорос ("ферма-месяц"), способность приносить потомство и эффект производителя. ESR подгоняли как совариабельный во всех анализах.
Анализируемыми признаками были общее число родившихся (TNB) и число родившихся живыми (NBA).
Используемые модели:
1. Общее число рожденных = яловость - месяц + производитель + OPN аллель
2. TNB или NBA = яловость - месяц + производитель +ESR + OPN 112 + OPN 122 etc.
Эти данные показывают, что OPN 124 показывает значимый (p < 0,01) положительный эффект в плане TNB = 0,7 для каждой копии аллели. Кроме того, OPN 142 показала тенденцию к негативному эффекту в отношении объема потомства в L03, подобный эффект наблюдался и для L93.
Как обсуждалось выше, уже показано, что другой ген ESR оказывает влияние на плодовитость свиней и, вероятно, в будущем будут найдены и другие гены, связанные с плодовитостью. Мы анализировали, оказывают ли определенные полезные комбинации аллелей двух отдельных генов, OPN и ESR, добавочный эффект на плодовитость.
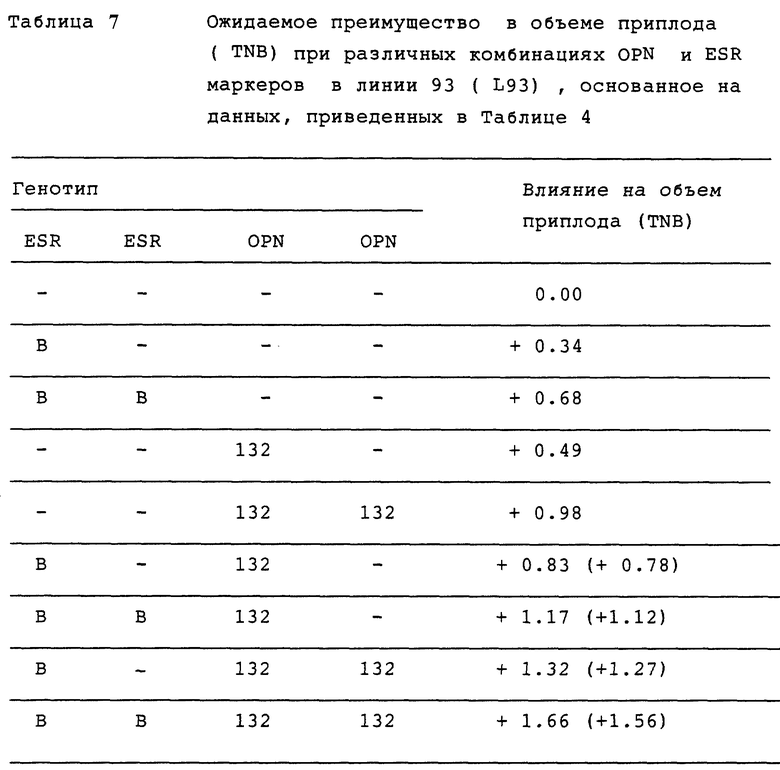
Для того, чтобы проверить эту возможность, мы наблюдали за связью между объемом приплода и различными комбинациями ESR и OPN аллелей. Результаты, приведенные в таблицах 6 и 7, показывают, что, действительно, полезные аллели OPN могут позитивно сочетаться с полезными аллелями ESR, так что может быть успешно получен даже больший приплод, чем полученный в результате использования полезных аллелей OPN и ESR по отдельности.
Эффекты плодовитости предполагают абсолютное увеличение (OPN 132 = + 0,49; ESR В = + 0.39) за исключением значений в скобках, которые допускают эффект OPN 132 или ESR В = + 0,39л
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ селекции на повышение воспроизводительных качеств свиноматок | 2022 |
|
RU2809200C1 |
| Способ ускорения селекционного процесса при создании специализированных линий свиней | 2022 |
|
RU2827487C2 |
| СПОСОБ СКРИНИНГА ЖИВОТНЫХ С ПОВЫШЕННЫМ УРОВНЕМ ЧИСЛЕННОСТИ ПРИПЛОДА И/ИЛИ ПРЕДПОЧТИТЕЛЬНЫМИ ПРИЗНАКАМИ КАЧЕСТВА МЯСА, БЕЛОК PRKAG3 (ВАРИАНТЫ) И КОДИРУЮЩАЯ ЕГО НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ (ВАРИАНТЫ) | 2001 |
|
RU2267538C2 |
| Разработка способа ускоренной селекции для создания специализированных линий свиней | 2022 |
|
RU2806803C1 |
| Способ селекции свиней на высокую воспроизводительную способность | 2021 |
|
RU2756315C1 |
| СПОСОБ ОТБОРА ЖИВОТНЫХ ПО ПРИЗНАКАМ, НАСЛЕДУЕМЫМ ПО МЕХАНИЗМУ РОДИТЕЛЬСКОГО ИМПРИНТИНГА | 1999 |
|
RU2262229C2 |
| СПОСОБ ОЦЕНКИ ЭФФЕКТИВНОСТИ СЕЛЕКЦИОННОЙ РАБОТЫ НА ОСНОВЕ ПОЛИЛОКУСНОГО ГЕНОТИПИРОВАНИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ | 2020 |
|
RU2756922C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ДОСТОВЕРНОСТИ ПРОИСХОЖДЕНИЯ И ЧИСТОПОРОДНОСТИ СВИНЕЙ | 2019 |
|
RU2744733C1 |
| ГЕНЕТИЧЕСКИЙ ТЕСТ ДЛЯ ИДЕНТИФИКАЦИИ НОСИТЕЛЕЙ РЕЦЕССИВНОГО ГЕНА КОМПЛЕКСНЫХ ВЕРТЕБРАЛЬНЫХ МАЛЬФОРМАЦИЙ У КРУПНОГО РОГАТОГО СКОТА | 2001 |
|
RU2276690C2 |
| НУКЛЕОТИДНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ, УЧАСТВУЮЩИЕ В УВЕЛИЧЕНИИ ИЛИ УМЕНЬШЕНИИ СКОРОСТИ ОВУЛЯЦИИ У МЛЕКОПИТАЮЩИХ | 2001 |
|
RU2283866C2 |






Изобретение относится к области животноводства. Предложенные способы скрининга свиней для определения особей с более вероятной повышенной плодовитостью и/или особей с менее вероятной повышенной плодовитостью основаны на идентификации OPN аллелей, присутствующих в пробе геномной ДНК свиньи. Предложенные наборы для использования в таких способах включают один или более реагентов или материалов, способных идентифицировать OPN аллелей в образце геномной ДНК свиньи. Изобретение позволяет значительно увеличить численность приплода по сравнению с данной величиной у взятой популяции. 6 с. и 16 з.п. ф-лы, 7 табл.
GCTAGTTAATGACATTGTACATAA,
CCAATCCTATTCACGAAAAAGC,
GTGTCATGAGGTTTTTTCCACTGC или
CAACCCACTTGCTCCCAC.
GCTAGTTAATGACATTGTACATAA,
CCAATCCTATTCACGAAAAAGC,
GTGTCATGAGGTTTTTTCCACTGC или
CAACCCACTTGCTCCCAC.
GCTAGTTAATGACATTGTACATAA,
CCAATCCTATTCACGAAAAAGC,
GTGTCATGAGGTTTTTTCCACTGC или
CAACCCACTTGCTCCCAC.
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| Science, 25 March 1994, v | |||
| Железнодорожный снегоочиститель на глубину до трех сажен | 1920 |
|
SU263A1 |
| Центрифуга, приводимая во вращение переменным током | 1924 |
|
SU1771A1 |

