Данное изобретение было по крайней мере частично поддержано проектом под номером IOWO 3600 (Hatch Funds, USDA). Правительству Соединенных Штатов могут принадлежать определенные права на данное изобретение.
Перекрестные ссылки на связанные заявки
Данная заявка является продолжением следующих одновременно находящихся на рассмотрении предварительных заявок на выдачу патента США, принадлежащих настоящему заявителю: 60/231045, поданной 8 сентября 2000 г.; 60/260239, поданной 8 января 2001 г.; и 60/299111, поданной 18 июня 2001 г. Притязания на приоритет осуществляются согласно 35 U.S.C. Section 120.
Область изобретения
Данное изобретение в общем относится к выявлению генетических различий среди животных. Более конкретно, изобретение относится к генетическим маркерам, указывающим на наследуемые фенотипы с улучшенным качеством мяса, увеличенной численностью приплода, и другими экономически выгодными свойствами животных. Также описаны способы и композиции для применения данных маркеров при генотипировании и селекции животных.
Предпосылки изобретения
Между отдельными особями животных, а также между их породами существуют генетические различия, которые могут использоваться в способах разведения для получения животных с требуемыми характеристиками. Например, известно, что животные китайских пород достигают половой зрелости в раннем возрасте и характеризуются большой численностью приплода, тогда как животные американских пород быстрее растут и бывают менее упитанными. Часто, однако, требуемые свойства слабо передаются по наследству, и стандартные способы разведения, в которых особей выбирают на основе вариаций фенотипа, не полностью берут в расчет существующую генетическую изменчивость или комплексные взаимодействия генов.
Для исследования свиной ДНК несколькими группами применялся анализ полиморфизма длин рестрикционных фрагментов (RFLP). В работе Jung et al., Theor. Appl. Genet., 77: 271-274 (1989), включенной сюда в качестве ссылки, описано применение способов RFLP для выявления генетической изменчивости между двумя породами свиней. Для данных пород демонстрировали полиморфизм генов свиного лейкоцитарного антигена (SLA) класса I. В работе Hoganson et al., Abstract for Annual Meeting of Midwestern Section of the American Society of Animal Science, March 26-28, 1990, включенной сюда в качестве ссылки, сообщается о полиморфизме генов свиного главного комплекса гистосовместимости (MHC) для свиней китайских пород, что также продемонстрировано путем PFLP-анализа. В работе Jung et al., Theor. Appl. Genet., 77: 271-274 (1989), включенной сюда в качестве ссылки, сообщается о RFLP-анализе генов SLA класса I некоторых свиней. Данные авторы констатируют, что по их результатам можно предположить, что могут иметь место ассоциации между генами свиного SLA/MHC класса I и свойствами продукции и производительности. Кроме того, они констатируют, что применение рестрикционных фрагментов SLA класса I в качестве генетических маркеров может в будущем потенциально служить для улучшения производительности выращивания свиней.
Способность выявлять конкретный выбранный генный аллель приводит к осуществлению нового и продолжительного процесса идентификации молекулярного ДНК-маркера для главного действующего гена. Данный маркер может быть связан с одним геном, или с некоторым числом генов с дополнительным действием. ДНК-маркеры имеют несколько преимуществ; отличие легко оценивать, и оно однозначно, и ДНК-маркеры являются кодоминантными, т.е. могут быть идентифицированы отличия между гетерозиготными и гомозиготными животными. Если система маркеров установлена, решения по селекции могут быть вынесены очень легко, поскольку ДНК-маркеры можно анализировать каждый раз после того, как может быть взят образец ткани или крови от отдельной молодой особи, или даже от эмбриона.
Применение генетических различий в генах рецепторов стало ценной системой маркеров для селекции. Например, в патентах США 5550024 и 5374526, выданных Rothschild et al., описан полиморфизм в свином гене рецептора эстрогена, который ассоциирован с большей численностью приплода, причем описание данных патентов включено сюда в качестве ссылки. В патенте США №5935784 описаны полиморфные маркеры в свином гене рецептора пролактина, которые ассоциированы с большей численностью приплода и общей репродуктивной эффективностью.
Численность приплода, конечно, имеет большое экономическое значение для животновода, а также для мясных пород животных важно качество мяса. Качество мяса является характеристикой, с трудом поддающейся оценке, так как этот общий параметр составляется из множества различных аспектов, как объективных, так и субъективных. Список факторов, определяющих качество мяса, как и для других продуктов, является довольно длинным (Wood et al., Proceedings of The Nutrition Society (1999) 58: 363-70). Он включает отсутствие микробиологических загрязнений (пищевая безопасность) и предотвращение эксплуатации животных (забота о животных). Он также включает сенсорную привлекательность мяса, т.е. его вкус или пищевое качество, и полезность для здоровья при потреблении, особенно в отношении количества и типа жира.
На качество сырой свинины влияет большое число генетических и негенетических факторов. Последние включают условия содержания на ферме, условия транспортировки, забоя и переработки. Специалисты по мясу провели значительное количество исследований данных факторов, что привело к заметному улучшению качества. Некоторая часть исследований была также посвящена генетическим характеристикам животных, и в некоторых исследованиях была выявлена значимость генетических факторов. За счет этого в промышленности стало известно, что селекционное разведение животных и применение генной технологии может играть важную роль для улучшения качества свинины.
Информация на уровне ДНК может помочь установить специфический основной ген, но также она может способствовать выбору количественной характеристики уже выбранного признака. Молекулярная информация в дополнение к фенотипическим данным может повышать точность селекции и, поэтому, реакции на селекцию. Степень дополнительной реакции при проведении такой программы селекции с помощью маркеров (Marker Assisted Selection; MAS) рассматривалась многими работниками с теоретической точки зрения. В общих терминах MAS является более выгодной для слабо наследуемых признаков, и для признаков, фенотипическая оценка которых является дорогостоящей. В частности, качество мяса расценивается как отличная возможность применения MAS. Например, в работе Meuwissen, T. H. E. and Goddard, M. E. (1996) "The use of Marker Haplotypes in Animal Breeding Schemes", Genet. Sel. Evol., 28 161-176 рассматривается применение селекции с помощью маркеров для таких признаков, как репродукция и качество мяса, которые трудно улучшить с использованием традиционных способов. Данные результаты являются крайне обнадеживающими, и они показывают, что для таких признаков, как качество мяса, при оценке признака после забоя могла достигаться дополнительная реакция до 64%.
Конечно, лучшим подходом генетического улучшения экономически значимых свойств, таких как качество мяса или численность приплода, является непосредственное обнаружение в популяции значимых ДНК-маркеров во время селекции. Измерения качества мяса могут выполняться непрерывно для некоторых животных из племенных популяций в животноводческих организациях. Поскольку полная оценка качества мяса может выполняться только после забоя, эти данные могут собираться только для выбракованных животных и не могут быть получены для потенциально племенных животных. Сходным образом, численность приплода маток может определяться только после размножения. Идентификация генетической предрасположенности для данных признаков может сделать возможной селекцию на генетическом уровне.
Эти фенотипические данные собирают для подтверждения обнаружения значимых ДНК-маркеров, и для подтверждения маркеров, идентифицированных с использованием экспериментальных популяций, или для тестирования генов-кандидатов. Значимые маркеры или гены могут затем непосредственно включаться в процесс селекции. Преимущество данной молекулярной информации состоит в том, что ее можно получить уже в очень раннем возрасте разводимых животных, и это значит, что животные могут подвергаться предварительной селекции, основываясь на ДНК-маркерах, до выполнения ими ростового производительного теста. Это является большим преимуществом для всей системы тестирования и селекции.
Из следующего выше можно видеть, что существует потребность в идентификации маркеров, которые могут использоваться для улучшения качества мяса, а также репродуктивных характеристик животных путем идентификации и селекции животных с улучшенными характеристиками на генетическом уровне.
Целью настоящего изобретения является предоставление генетического маркера, основанного на гене PRKAG3 или последовательности внутри него, который является индикатором предпочтительных характеристик мяса, таких как определяемые по pH, по мраморности, по цвету и по потере жидкости вследствие вытекания сока, и/или большей численности приплода.
Другой целью изобретении является предоставление анализа для определения данного генетического маркера.
Дальнейшей целью изобретения является предоставление способа оценки животных, повышающего точность способов селекции и разведения для требуемых признаков.
Еще одной целью изобретения является предоставление теста амплификации путем PCR, который значительно ускорит определение присутствия данного маркера.
Другие цели и преимущества данного изобретения будут частично указаны в последующем описании, и часть их будет очевидно вытекать из описания, или они могут быть освоены при практическом осуществлении изобретения. Другие цели и преимущества данного изобретения могут достигаться при помощи средств и комбинаций, подробно указанных в прилагаемой формуле изобретения.
Сущность изобретения
Данное изобретение относится к обнаружению альтернативных генных форм гена PRKAG3, которые могут использоваться в качестве генетических маркеров, ассоциированных с признаками качества мяса и репродуктивными признаками животных. Ген PRKAG3 является высоко консервативным среди видов животных, и ожидается, что описанные здесь различные аллели также будут коррелировать с изменчивостью в данном гене у других экономически значимых или мясных животных, таких как крупный рогатый скот, овцы, куры, и т.д.
Для достижения цели и по замыслу изобретения, что осуществлено и развернуто описано здесь, настоящее изобретение относится к обнаружению альтернативных генотипов, что предоставляет способ скрининга животных с целью определения тех, которые с большей вероятностью характеризуются предпочтительными признаками качества мяса, или для их отбора от тех свиней, что имеют аллели, свидетельствующие о менее предпочтительных признаках качества мяса. Применяемое здесь выражение «предпочтительный признак качества мяса» означает значимое улучшение (повышение или снижение) одного из многих измеряемых признаков качества мяса по сравнению со средним значением для данной популяции, так что данная информация может использоваться для разведения с достижением однородной популяции с оптимизированным качеством мяса, причем это может включать повышение некоторых признаков или снижение других в зависимости от требуемых характеристик мяса. Данные факторы, которые могут приниматься во внимание, включают в качестве неограничивающих примеров следующие:
Яркость спинной части туши Minolta (L*): приемлемым является интервал в пределах 43-47 единиц (от более темного до более светлого), но лучшим является значение L*, равное 43; т.е. оно имеет большую экономическую ценность вообще в данном интервале (это может зависеть от рынка, например, в Японии предпочитают свинину более темного цвета).
Японский цветовой коэффициент (JCS) спинной части туши: приемлемым является интервал в пределах 2,5-5,0 единиц (от более темного до более светлого), но лучшим является значение JCS, равное 3-4.
Мраморность спинной части туши (уровень внутримышечного жира): в общем, лучшей является высокая мраморность, поскольку она ассоциирована с улучшенными характеристиками пищевого качества мяса.
pH спинной части туши: (окончательная кислотность мяса, измеряемая через 24 часа после забоя; данное свойство является наиболее важным признаком качества свинины) требуемым интервалом является 5,50-5,80, но лучшим является значение 5,80, поскольку оно положительно влияет на цвет и (слабо) на очистку (purge) мяса.
Яркость окорока Minolta (L*): приемлемым является интервал в пределах 43-52 единиц, но лучшим является более низкое значение (43).
pH окорока: лучшим является более высокое значение, т.е. 5,80.
Потеря жидкости вследствие вытекания сока или усыхание (purge): приемлемым является интервал в пределах 1%-3%, но лучшим является более низкое значение.
Данные способы оценки качества мяса служат примерами тех, что в основном применяются специалистами в данной области. Для обзора признаков качества мяса можно обратиться к следующему источнику: Sosnicki, A.A., E.R.Wilson, E.B.Sheiss, A. deVries, 1998 "Is there a cost effective way to produce high quality pork?", Reciprocal Meat Conference Proceedings, Vol.51.
Таким образом, настоящее изобретение относится к способу скрининга свиней для идентификации тех особей, что с большей вероятностью дадут мясо предпочтительного качества, и/или тех особей, что с меньшей вероятностью дадут мясо предпочтительного качества, с целью оптимизации способов разведения и селекции на предмет лучшего качества мяса.
Данное изобретение также включает способ скрининга свиней для определения тех особей, что с большей вероятностью дадут больший приплод при разведении, или для их отбора от тех свиней, что имеют аллели, указывающие на малую численность приплода. Применяемое здесь выражение «больший приплод» означает значимое повышение численности приплода относительно среднего значения в данной популяции. Таким образом, настоящее изобретение относится к способу скрининга свиней для определения тех особей, что с большей вероятностью дадут больший приплод, и/или тех, что с меньшей вероятностью дадут больший приплод.
Способы анализа данных признаков в общем включают стадии: 1) получения от свиньи биологического образца; и 2) анализа геномной ДНК или белка, полученных на стадии 1) с целью определения, какой аллель(и) PRKAG3 присутствует(ют). Также сюда включаются данные по гаплотипу, которые позволяют комбинировать при выполнении протокола селекции или идентификации серию полиморфизмов гена PRKAG3 для максимизации преимуществ каждого из данных маркеров.
Поскольку некоторые из полиморфизмов влекут изменения аминокислотного состава белка PRKAG3, способы анализа могут даже включать определение аминокислотного состава белка PRKAG3. Способы данного типа очистки или анализа обычно используют выделение белка с помощью средств, включающих флуоресцентное мечение антителами, разделение и очистку белка (например, в системе обращенно-фазовой ВЭЖХ), и применение автоматического белкового секвенатора для идентификации присутствующей последовательности. Протоколы данного анализа являются стандартными и известными в данной области и описаны Ausubel et al. (eds.), Short Protocols in Molecular Biology Fourth ed. John Wiley and Sons, 1999.
В предпочтительном осуществлении анализируют генетический образец. В кратком изложении, образец генетического материала получают от животного, и данный образец анализируют для определения наличия или отсутствия полиморфизма в гене регуляторной гамма-субъединицы активируемой АМФ протеинкиназы (PRKAG3), который коррелирует с повышенной численностью приплода или с улучшенным качеством мяса, или с обоими признаками, в зависимости от формы гена.
Как хорошо известно специалистам в данной области, при сравнении молекул нуклеиновой кислоты на предмет различий в последовательности могут использоваться разнообразные способы. Например, они включают анализ полиморфизма длин рестрикционных фрагментов, анализ гетеродуплексов, анализ полиморфизма однонитевой конформации, электрофорез с денатурирующим градиентом и электрофорез с температурным градиентом.
В предпочтительном осуществлении полиморфизм представляет собой полиморфизм длин рестрикционных фрагментов, и анализ включает идентификацию свиного гена PRKAG3 в выделенном генетическом материале; воздействие на данный ген ферментом рестрикции, что приводит к образованию рестрикционных фрагментов гена различной длины; разделение рестрикционных фрагментов с образованием профиля рестрикции, например, путем электрофореза или разделения в ВЭЖХ; и сравнения полученных в результате профилей рестрикционных фрагментов из гена PRKAG3, с теми, о которых известно, что они содержат или не содержат требуемый маркер. Если при тестировании животное является позитивным в отношении данных маркеров, оно может считаться подходящим для включения в программу разведения. Если при тестировании животное не признано позитивным в отношении маркерного генотипа, оно может выбраковываться из группы и использоваться иным образом. В скрининг множественных аллелей для исследования качества мяса и/или численности приплода также может включаться применение данных по гаплотипу.
В наиболее предпочтительном осуществлении ген выделяют путем использования праймеров и ДНК-полимеразы для амплификации конкретного региона гена, который содержит полиморфизм. Затем амплифицированный регион расщепляют ферментом рестрикции, и фрагменты снова разделяют. Визуализация паттерна RFLP осуществляется путем простой окраски фрагментов или путем мечения праймеров или нуклеозидтрифосфатов, применяемых при амплификации.
В другом осуществлении данное изобретение относится к способу идентификации генетического маркера качества мяса и/или численности приплода в конкретной популяции. Скрещивают хряков и свиноматок одной и той же породы, или разных пород, или одной генетической линии, и определяют число поросят в помете (для свиноматок) и/или качество мяса, продуцированного каждой свиньей. Идентифицируют полиморфизм гена PRKAG3 каждой свиньи, и ассоциируют его с числом поросят в помете или качеством мяса. Предпочтительно, для определения полиморфизма применяют анализ RFLP.
В другом осуществлении данное изобретение относится к способу идентификации генетического маркера качества мяса и/или численности приплода (числа рожденных детенышей) для любого конкретного экономически значимого животного, отличного от свиньи. По причине высоко консервативной природы данного гена среди различных животных, ожидается, что посредством не более чем описанного выше рутинного тестирования данный маркер может применяться на других видах животных для отбора качества мяса или численности приплода (число рожденных детенышей), основываясь на приведенных здесь указаниях. Скрещивают самцов и самок одной и той же породы, или разных пород, или одной генетической линии, и определяют и соотносят число детенышей в помете и/или качество мяса, продуцированные каждым животным. Для других животных, последовательности которых доступны, может проводиться сравнение последовательностей посредством BLAST для того, чтоб удостовериться, является ли конкретный аллель аналогичным описанному здесь. Аналогичный полиморфизм будет присутствовать у других животных и в других близкородственных генах. Термин «аналогичный полиморфизм» относится к полиморфизму, который совпадает с любым из описанных здесь, что определяется путем сравнений посредством BLAST.
Для описания отношений последовательности между двумя или более нуклеиновыми кислотами или полинуклеотидами используются следующие термины: (a) «последовательность сравнения», (b) «окно сравнения», (c) «идентичность по последовательности», (d) «процентная идентичность по последовательности» и (e) «существенная идентичность».
(a) Применяемый здесь термин «последовательность сравнения» относится к определенной последовательности, применяемой как основа для сравнения последовательностей. В данном случае это последовательность сравнения PRKAG3. Последовательность может представлять собой часть определенной последовательности или всю ее; как, например, сегмент полноразмерной последовательности кДНК или гена, или целая последовательность кДНК или гена.
(b) Применяемый здесь термин «окно сравнения» включает ссылку на непрерывный и определенный сегмент полинуклеотидной последовательности, где полинуклеотидная последовательность может сравниваться с последовательностью сравнения, и где часть полинуклеотидной последовательности, находящаяся в окне сравнения, может включать добавления или делеции (т.е. вставки) по отношению к последовательности сравнения (которая не включает добавления или делеции) для оптимального выравнивания двух последовательностей. В основном, окно сравнения составляет в длину по крайней мере 20 следующих друг за другом нуклеотидов, и необязательно может быть длиной 30, 40, 50, 100 нуклеотидов, или длиннее. Специалисты в данной области понимают, что во избежание решения о высокой степени сходства после включения вставок в нуклеотидную последовательность, обычно вводят штраф за вставку и его вычитают из числа совпадений.
Способы выравнивания последовательностей для их сравнения хорошо известны в данной области. Оптимальное выравнивание последовательностей для сравнения может проводиться по алгоритму локальной гомологии Smith and Waterman, Adv. Appl. Math. 2: 482 (1981); по алгоритму гомологичного выравнивания Needleman and Wunsch, J. Mol. Biol. 48: 443 (1970); по способу поиска сходства Pearson and Lipman, Proc. Natl. Acad. Sci. 85: 2444 (1988); с помощью компьютерных реализаций данных алгоритмов, включая в качестве неограничивающих примеров CLUSTAL в составе программы PC/Gene от Intelligenetics, Mountain View, Калифорния; GAP, BESTFIT, BLAST, FASTA, и TFASTA в составе Wisconsin Genetics Software Package, Genetics Computer Group (GCG), 575 Science Dr., Madison, Висконсин, США; программа CLUSTAL подробно описана Higgins and Sharp, Gene 73: 237-244 (1988); Higgins and Sharp, CABIOS 5: 151-153 (1989); Corpet, et al., Nucleic Acids Research 16: 10881-90 (1988); Huang, et al., Computer Applications in the Biosciences 8: 155-65 (1992), и Pearson, et al., Methods in Molecular Biology 24: 307-331 (1994). Семейство программ BLAST, которое может использоваться для поиска сходства в базах данных, включает: BLASTN для поиска запросов по нуклеотидным последовательностям в базах данных нуклеотидных последовательностей; BLASTX для поиска запросов по нуклеотидным последовательностям в базах данных белковых последовательностей; BLASTP для поиска запросов по белковым последовательностям в базах данных белковых последовательностей; TBLASTN для поиска запросов по белковым последовательностям в базах данных нуклеотидных последовательностей; и TBLASTX для поиска запросов по нуклеотидным последовательностям в базах данных нуклеотидных последовательностей. См. Current Protocols in Molecular Biology, Chapter 19, Ausubel, et al., Eds., Greene Publishing and Wiley-Interscience, New York (1995).
Кроме указанных по другому случаев, приведенные здесь значения идентичности/сходства относятся к значению, полученному с использованием пакета программ BLAST 2.0 с использованием параметров по умолчанию. Altschul et al., Nucleic Acids Res. 25: 3389-3402 (1997). Программное обеспечение для выполнения анализов BLAST является общедоступным, например, через National Center for Biotechnology-Information (http://www.ncbi.nlm.nih. gov/).
Данный алгоритм вначале включает выявление пар последовательностей с высоким коэффициентом (HSP) путем идентификации коротких слов длиной W в последовательности запроса, которые либо совпадают, либо удовлетворяют некоторому коэффициенту позитивного пограничного значения T при выравнивании со словом той же длины в последовательности из базы данных. T обозначается как граничное значение коэффициента соседних слов (Altschul et al., supra). Данные первоначальные совпадения соседних слов действуют как основа для возникновения инициирующих поисков с целью поиска содержащих их более длинных HSP. Концы слов затем продлеваются в обоих направлениях по каждой последовательности до тех пор, пока может повышаться кумулятивный коэффициент выравнивания. Кумулятивные коэффициенты вычисляют с использованием для нуклеотидных последовательностей параметров M (коэффициент вознаграждения за пару совпадающих остатков; всегда > 0) и N (коэффициент штрафа за несовпадающие остатки; всегда < 0). Для аминокислотных последовательностей для вычисления кумулятивного коэффициента применяют матрицу коэффициентов. Продление концов слов в каждом направлении прекращается в случае, когда кумулятивный коэффициент выравнивания снижается на число Х от максимально достигнутого им значения; кумулятивный коэффициент становится равен нулю или ниже вследствие накопления при выравнивании одного или нескольких остатков с отрицательным коэффициентом; или при достижении конца одной из последовательностей. Параметры алгоритма BLAST W, T и X определяют чувствительность и скорость выравнивания. В программе BLASTN (для нуклеотидных последовательностей) по умолчанию применяются длина слова (W), равная 11, ожидание (E), равное 10, уровень отсечения, равный 100, M=5, N=-4, и сравнение обеих цепей. Для аминокислотных последовательностей в программе BLASTP по умолчанию применяются длина слова (W), равная 3, ожидание (E), равное 10, и матрица коэффициентов BLOSUM62 (см. Henikoff & Henikoff (1989) Proc. Natl. Acad. Sci. USA 89: 10915).
В дополнение к вычислению процентной идентичности последовательностей алгоритм BLAST также проводит статистический анализ сходства между двумя последовательностями (см., например, Karlin & Altschul, Proc. Natl. Acad. Sci. USA 90: 5873-5787 (1993)). Одно измерение сходства, обеспечиваемое алгоритмом BLAST, представляет собой вероятность наименьшей суммы (P (N)), которая представляет оценку вероятности, с которой совпадение двух нуклеотидов или аминокислотных последовательностей произойдет случайным образом.
Поиски BLAST предполагают, что белки могут моделироваться как случайные последовательности. Однако, многие реальные белки включают регионы неслучайных последовательностей, которые могут являться гомополимерными участками, повторами с коротким периодом или регионами, обогащенными одной или несколькими аминокислотами. Такие регионы с низкой комплексностью могут выравниваться при выравнивании двух неродственных белков даже несмотря на то, что другие регионы белков являются абсолютно не сходными. Для снижения такого выравнивания регионов низкой комплексности может применяться некоторое количество программ-фильтров низкой комплексности. Например, фильтры низкой комплексности SEG (Wooten and Federhen, Comput. Chem., 17: 149-163 (1993)) и XNU (Claverie and States, Comput. Chem., 17: 191-201 (1993)) могут использоваться отдельно или в комбинации.
(c) Применяемый здесь термин «идентичность последовательности» или «идентичность» в контексте двух последовательностей нуклеиновой кислоты или полипептидных последовательностей включает ссылку на остатки двух последовательностей, которые совпадают при выравнивании с максимальным соответствием посредством конкретного окна сравнения. Когда по отношению к белкам используется процентная доля идентичности последовательности, подразумевается, что позиции остатков, которые не идентичны, часто отличаются консервативными аминокислотными заменами, в которых аминокислотные остатки замещены другими аминокислотными остатками со сходными химическими свойствами (например, зарядом или гидрофобностью), и поэтому не меняют функциональных свойств молекулы. Когда последовательности отличаются консервативными заменами, процентная идентичность последовательности может повышаться для поправки на консервативную природу замены. О последовательностях, отличающихся такими консервативными заменами, говорят, что они имеют «сходство последовательности» или «сходство». Средства для осуществления такой поправки хорошо известны специалистам в данной области. Обычно они включают учет консервативной замены как частичной, в отличие от полного несовпадения, с повышением таким образом процентной идентичности последовательности. Так, например, в случае придания идентичной аминокислоте счета 1, и придания неконсервативной замене нулевого счета, консервативной замене придают счет между нулем и 1. Коэффициенты консервативных замен вычисляют, например, по алгоритму Meyers and Miller, Computer Applic. Biol. Sci., 4: 11-17 (1988), что, например, реализовано в программе PC/GENE (Intelligenetics, Mountain View, Калифорния, США).
(d) Применяемый здесь термин «процентная доля идентичности последовательности» означает значение, определенное путем сравнения двух оптимально выровненных последовательностей посредством окна сравнения, где часть полинуклеотидной последовательности в окне сравнения может включать добавления или делеции (т.е. вставки) по отношению к последовательности сравнения (которая не включает добавления или делеции) с целью оптимального выравнивания двух последовательностей. Процентную долю вычисляют путем определения числа позиций, где в обеих последовательностях имеет место идентичное основание нуклеиновой кислоты или аминокислотный остаток, с получением числа совпадающих позиций, деления числа совпадающих позиций на общее число позиций в окне сравнения и умножения результата на 100, с получением процентной доли идентичности последовательности.
(e) (I) Термин «существенная идентичность» полинуклеотидной последовательности означает, что полинуклеотид включает последовательность, которая характеризуется по крайней мере 70%-ной идентичностью последовательности, предпочтительно, по крайней мере 80%-ной идентичностью, более предпочтительно, по крайней мере 90%-ной идентичностью, и наиболее предпочтительно, по крайней мере 95%-ной идентичностью, в отношении последовательности сравнения, что вычисляется с использованием одной из программ для выравнивания, описанных с применением стандартных параметров. Специалисту в данной области ясно, что данные значения могут быть подобраны подходящим образом для определения соответствующей идентичности белков, кодируемых двумя нуклеотидными последовательностями, с учетом вырожденности кодонов, сходства аминокислот, позиционирования рамки считывания и тому подобного. Существенная идентичность аминокислотных последовательностей для данных целей обычно означает идентичность последовательности, равную по крайней мере, 60%, более предпочтительно, по крайней мере 70%, 80%, 90%, и наиболее предпочтительно, по крайней мере 95%.
Данные программы и алгоритмы могут выявлять аналогию конкретного полиморфизма в целевом гене с тем, что описан здесь. Как указывалось раньше, на основе высоко консервативной природы гена PRKAG3 (Jeon T.J., V. Armeger, C. Rogel-Gaillard, A. Robic, E. Bongcam-Rudloff et al., 2001 Genomics 72: 297-303), ожидается, что данный полиморфизм существует и у других животных, и его применение у отличных от описанных здесь животных включает не более чем рутинную оптимизацию параметров, которые используют приведенные здесь указания. Последовательность свиного PRKAG3 показана на фигуре 1.
Также возможно установить связь между специфическими аллелями альтернативных ДНК-маркеров и аллелями ДНК-маркеров, о которых известно, что они ассоциированы с конкретными генами (например, с обсуждаемым здесь геном PRKAG3), которые, как ранее показано, ассоциированы с конкретным признаком. Так, в настоящей ситуации, для гена PRKAG3, по крайней мере, в краткие сроки возможна селекция свиней, вероятно, способных дать приплод большей численности и/или мясо лучшего качества, или, альтернативно, отбраковка тех свиней, которые, вероятно, дадут приплод меньшей численности и/или мясо менее предпочтительного качества, осуществляемая непрямым образом, путем выбора особых аллелей ассоциированного с PRKAG3 маркера путем выбора специфичных аллелей альтернативных маркеров хромосомы 15. Примеры таких маркеров, о которых известно, что они сцеплены с PRKAG3 на свиной хромосоме 15, включают SW1683 и SW1983. Применяемый здесь термин «генетический маркер» включает не только полиморфизм, открытый любыми способами анализа изменений белка, ассоциированных с данным полиморфизмом, но также сцепленные маркеры, применение микросателлитов, или даже другие способы анализа причинных изменений белка, на которые указывает маркер, и их применение для воздействия на качество мяса животного.
При использовании здесь обозначение конкретного полиморфизма часто делается по названию конкретного фермента рестрикции. Это не предполагает того, что единственным способом, которым может идентифицироваться данный участок, является применение данного фермента рестрикции. Имеется множество доступных специалистам баз данных и ресурсов, с помощью которых можно определить другие ферменты рестрикции, которые могут использоваться для идентификации конкретного полиморфизма, например, http://darwin.bio.genese.edu, на котором после анализа последовательности и подлежащего идентификации полиморфизма можно получить список подходящих ферментов рестрикции. В действительности, как описано в приведенных здесь указаниях, существуют многие пути идентификации конкретного полиморфизма или аллеля альтернативными способами, которые даже могут не включать фермент рестрикции, но которые анализируют ту же генетическую или протеомную альтернативную форму.
Сопутствующие фигуры, которые включены сюда и которые составляют часть данной спецификации, иллюстрируют одно из осуществлений данного изобретения и, вместе с данным описанием, служат для объяснения принципов данного изобретения.
Описание фигур
Фигура 1 представляет собой изображение нуклеотидной последовательности свиного PRKAG3, включая аминокислоты, идентифицированные альтернативные полиморфные локусы и изменения соответствующих им аминокислот.
На фигурах 2A и 2B изображена последовательность 5'-концевого фланкирующего региона гена PRKAG3, включая экзон 1, экзон 2 и новую последовательность интрона между ними.
На фигуре 2A присутствует SINE (11), а на фигуре 2B SINE отсутствует (22). Данная последовательность может использоваться для формирования дополнительных праймеров. Жирный шрифт обозначает прямые повторы между SINE; жирным шрифтом и курсивом обозначены экзоны гена PRKAG3 (экзон 1 и экзон 2).
Фигуры 3a и 3b представляют собой графики, на которых изображены кривые отношения дисперсий (критерий Фишера; F-ratio) доказательства QTL, ассоциированного с качеством мяса для SSC 15. Ось х означает относительные позиции на карте сцепления. Ось y представляет отношение дисперсий. Стрелки на оси x обозначают позиции, в которых присутствует маркер. Три линии представляют собой 5%-ный хромосомный (-----), 5%-ный геномный ( ) и 1%-ный геномный (- - -) уровень значимости. A, признаки среднего содержания гликогена, среднего содержания лактата и среднего гликолитического потенциала. На фиг.3b показаны признаки pH.
) и 1%-ный геномный (- - -) уровень значимости. A, признаки среднего содержания гликогена, среднего содержания лактата и среднего гликолитического потенциала. На фиг.3b показаны признаки pH.
На фигуре 4 продемонстрировано действие замены гаплотипа PRKAG3 на коэффициент pH и цветовой коэффициент окорока и спинной части туши. Действие замены гаплотипа оценивали для 5 пород (ALL) и для каждой породы. Породы основывались на породе ландрас (LR), большой белой (Large White; LW) или породе дюрок (Duroc; DU), синтетической породе, основанной на породе дюрок (DS), и породе, основанной на беркширской породе (BE). Для породы BE использовали отдельную шкалу. Оценки в колонке, которые имеют одинаковый верхний индекс, значимо отличаются при p<0,005 для оценок по всем породам и при p<0,005 для оценок по отдельным породам.
Подробное описание изобретения
Здесь будут сделаны подробные ссылки для указанных здесь осуществлений изобретения, которые вместе с последующими примерами служат для объяснения принципов изобретения.
Активируемая АМФ протеинкиназа участвует во включении продуцирующих АТФ метаболических путей. Также она может посредством фосфорилирования инактивировать гликогенсинтазу. АМРК состоит из трех субъединиц: каталитической α цепи и двух регуляторных субъединиц β и γ.
В опубликованной заявке WO/01/20003, выданной Institut National De Le Recherche Agronomique, описаны варианты гена PRKAG3. Они включают замену R41Q (которая в данном случае соответствует аминокислоте 200), замену V40I (аминокислота 199), что связано с известным RN-аллелем PRKAG3. В заявке сообщается об обнаружении мутации в кодоне 200 гена PRKAG3, связанной (в гомозиготном и гетерозиготном состоянии) с высоким гликолитическим потенциалом у гемпширской свиньи, называемой RN-(200Q) фенотипом. Свиньи с таким фенотипом имеют предельно низкое значение рН, уменьшенную способность удерживать воду и дают уменьшенную продукцию консервированного готового окорока. Однако анализ различных линий свиней указывает на то, что эта мутация в кодоне 200 возникла у гемпширской породы и встречается с очень низкой частотой или полностью отсутствует у других пород свиней. Кроме того, как описано в примере 5 публикации PCT, было показано, что 200Q всегда присутствовала совместно с 199V, и это подразумевает, что у маркера в позиции 199 отсутствуют вариации или независимое значение в качестве генетического маркера в отличие от маркера 200Q.
В заявке WO/01/20003 было определено, что маркер 200Q ассоциирован с неблагоприятной RN-мутацией. В данной заявке показано, что этот маркер всегда обнаруживается совместно с 199V, однако, 199V также присутствует вместе с 200R, который имеется в мясе лучшего качества. Заявители неожиданно обнаружили, что третья комбинация 199I/200R имеет в среднем лучшее качество мяса, чем 199V/200R. Кроме того, заявители обнаружили, что полиморфизм V199I неожиданным образом связан с изменением численности потомства. Данная информация позволяет использовать маркер 199 в качестве средства для селекции.
Кроме того, настоящие заявители еще идентифицировали новый полиморфный локус G52S, который связан с улучшенным качеством мяса. В заключение, для того, чтобы оценить взаимодействие между 199I-52G- и известным полиморфизмом 30T (описанным в работе Milan et. al., 2000), проводился анализ гаплотипа. Согласно этому осуществлению гаплотип 30T-52G-199I (в дальнейшем гаплотип 3) был самым благоприятным для признаков качества мяса.
На фигуре 1 изображен ген PRKAG3 и все обсуждаемые здесь полиморфизмы. (SEQ ID NO:1 является диким типом). До работы, описанной в данной заявке, не существовало никаких доказательств влияния этого гена на экономически значимые свойства других пород. В настоящее время неожиданно было обнаружено, что новые маркеры в гене PRKAG3 PRKAG3-199, PRKAG3-30 и PRKAG3-52 коррелируют с изменениями признаков качества мяса, а также с репродуктивными свойствами, такими как численность потомства, во многих отличных от гемпширской породы породах свиней. Эти новые маркеры, как было показано, коррелируют с мясом самого высокого технического качества по показателям цвета, уровня рН, мраморности и потери вследствие вытекания сока и также с признаками численности потомства. Согласно данному изобретению, связь этих полиморфизмов с такими признаками позволяет использовать генетические маркеры для идентификации определенных пород или генетических линий для того, чтобы определить животных с благоприятными характеристиками мяса и/или численностью потомства в начальный период жизни животного.
Различные генотипы маркеров PRKAG3-199 являются результатом полиморфизма гена PRKAG3, который приводит к преобразованию гуанина в аденин в 595 нуклеотидной позиции, (SEQ ID NO:7) приводящему к изменению аминокислоты валина на изолейцин (номер аминокислоты 199) (SEQ ID NO:8). Данное преобразование в свою очередь вызывает появление сайта рестрикции в аллеле 1, связанного с более низким содержанием гликогена, лактата и гликолитическим потенциалом. Также было обнаружено, что этот сайт коррелирует с увеличенной численностью потомства, если присутствует по крайней мере одна копия.
Различные генотипы маркеров PRKAG3-52 являются результатом полиморфизма гена PRKAG3, который приводит к преобразованию гуанина в аденин в 154 нуклеотидной позиции, (SEQ ID NO:5 (аминокислота 52), приводящему к изменению аминокислоты глицина на серин (SEQ ID NO:6). Данное преобразование в свою очередь вызывает появление сайта рестрикции в аллеле 2, связанного с более низким содержанием гликогена, лактата и гликолитическим потенциалом.
Различные генотипы маркеров PRKAG3-30 являются результатом полиморфизма гена PRKAG3, который приводит к преобразованию аденина в цитозин в 89 нуклеотидной позиции SEQ ID NO:3 (аминокислота 30), приводящему к изменению аминокислоты аспарагина на треонин SEQ ID NO:4. Ранее уже сообщалось о данном полиморфизме, но не было обнаружено, что он коррелирует с каким-нибудь фенотипом качества мяса. Треонин в значительной степени был ассоциирован с улучшенным качеством мяса.
Данное изобретение относится к генетическим маркерам экономически значимых признаков у животных. Данные маркеры представляют собой аллели, которые в значительной степени связаны с признаками качества мяса и/или численности потомства, репродуктивными признаками и, таким образом, относятся к способу скрининга животных для того, чтобы определить те особи, которые, наиболее вероятно, произведут при размножении больше потомства или лучшее качество мяса (или и то, и другое), посредством идентификации наличия или отсутствия полиморфизма в гене PRKAG3, который с ними таким образом коррелирует.
Таким образом, изобретение относится к генетическим маркерам и способам идентификации этих маркеров у животного определенной породы, рода, популяции, или группы, посредством чего особь женского пола, более вероятно, произведет потомство, которое в значительной степени увеличится выше среднего в численности для данной определенной породы, рода, популяции, или группы. Подобным образом данный способ можно использовать для идентификации животных, которые, наиболее вероятно, будут давать мясо с предпочтительным его качеством.
Может быть использован любой способ идентификации наличия или отсутствия такого маркера, включая, например, анализ полиморфизма одноцепочечной конформации (SSCP), сканирование последовательности на выпадение оснований (BESS), анализ RFLP, гетеродуплексный анализ, электрофорез в денатурирующем градиентном геле, аллельная PCR, непосредственное определение последовательности при помощи лигазной цепной реакции, минисеквенирование, гибридизация нуклеиновых кислот, определение гена PRKAG3 по типу микрополей, или других последовательностей, сцепленных с геном PRKAG3. Также объем изобретения включает анализ конформационных изменений или изменений последовательности белка, которые встречаются при наличии этого полиморфизма. Данный полиморфизм может быть вызван мутацией или этого может не быть, но он будет указывать на наличие такого изменения и можно проводить анализ генетических или белковых основ фенотипических различий.
Ниже представлен общий обзор способов, которые могут быть использованы для исследования полиморфизмов данного изобретения.
Образец генетического материала в настоящем изобретении получают из животных. Образцы могут быть получены из крови, ткани, спермы и т.д. В общем случае, в качестве источника используют клетки периферической крови, а генетическим материалом является ДНК. Для того чтобы предоставить достаточное количество ДНК для анализа, получают достаточное количество клеток. Это количество будет известно или легко определимо специалистами в данной области. ДНК выделяют из клеток крови посредством способов, известных специалистам в данной области.
Выделение и амплификация нуклеиновых кислот
Образцы геномной ДНК выделяют из любого подходящего источника, включая слюну, клетки щек, корни волос, кровь, пуповинную кровь, амниотическую жидкость, интерстициальную жидкость, перитонеальную жидкость, ворсинки хориона и любой другой подходящий образец клетки или ткани с интактными интерфазными ядрами или метафазными клетками. Клетки могут быть получены из цельной ткани из свежего или законсервированного органа, или из образца ткани, или биопсии. Образец может содержать такие соединения, как консерванты, антикоагулянты, буферы, фиксаторы, питательные вещества, антибиотики и подобные им вещества, которые в природе не перемешиваются с биологическим материалом.
Способы выделения геномной ДНК из этих различных источников описаны, например, в работе Kirby, DNA Fingerprinting, An Introduction, W.H. Freeman & Co. New York (1992). Геномную ДНК также можно выделять из культивируемых первичных или вторичных клеточных культур или из трансформированных клеточных линий, происходящих из любого из вышеперечисленных образцов тканей.
Также можно использовать образцы РНК животных. РНК можно выделять из экспрессирующих ген PRKAG3 тканей, как описано в работе Sambrook et al., указанной выше. РНК может быть общей клеточной РНК, мРНК, поли-А+ РНК, или любой их комбинацией. Для достижения наилучшего результата РНК очищают, но можно использовать также неочищенную цитоплазматическую РНК. РНК можно обратно транскрибировать для получения ДНК, которую затем используют в качестве матрицы амплификации, таким образом, что PCR опосредованно амплифицирует определенную популяцию транскриптов РНК. См., например, работы Sambrook, указанной выше, Kawasaki et al., главу 8 в PCR Technology (1992, указанной выше, и Berg et al., Hum.Genet. 85: 655-658 (1990).
PCR-амплификация
Самым распространенным средством амплификации является полимеразная цепная реакция (PCR), как описано в патентах США №4683195, 4683202, 4965188, каждый из которых включен сюда в качестве ссылки. В случае, когда PCR используется для амплификации целевой области клеток крови, гепаринизированную цельную кровь следует собирать в герметичную вакуумную пробирку, держать отдельно от других образцов и обрабатывать в чистых перчатках. Для достижения наилучшего результата, кровь следует обрабатывать непосредственно после сбора; если это невозможно, ее следует хранить до использования в герметичном контейнере при температуре 4°С. Также можно анализировать клетки других физиологических жидкостей. При использовании любой из таких жидкостей, клетки жидкости следует отделять от компонентов жидкости посредством центрифугирования.
Ткани следует грубо измельчить с использованием стерильного одноразового скальпеля и стерильной иглы (или двух скальпелей) в чашке Петри размером 5 мм. Процедура удаления парафина со срезов ткани описана во множестве специализированных руководств, хорошо известных специалистам в данной области.
Для того чтобы амплифицировать целевую последовательность нуклеиновой кислоты образца посредством PCR, последовательность должна быть доступна для компонентов системы амплификации. Одним способом выделения целевой ДНК является грубая экстракция, которая применима для относительно больших образцов. Короче говоря, мононуклеарные клетки из образцов крови, амниоциты из амниотической жидкости, культивируемые клетки ворсинок хориона, или им подобные клетки выделяют посредством расслоения в стерильном градиенте Ficoll-Hypaque по стандартной процедуре. Перед экстракцией ДНК, интерфазные клетки собирают и отмывают три раза в стерильном фосфатно-солевом буфере. В случае исследования ДНК из лимфоцитов периферической крови, предлагается применение осмотического шока (обработка осадка дистиллированной водой в течение 10 секунд), который сопровождается двумя дополнительными отмывками, если после первоначальных отмывок видны остаточные красные клетки крови. Такая процедура предотвращает при реакции PCR ингибиторный эффект гемовой группы, которую несет гемоглобин. В случае, если исследование PCR выполняется не сразу же после сбора образца, аликвоты клеток количеством 106 можно осаждать в стерильных пробирках типа Эппендорф и замораживать сухой осадок при температуре -20°С до использования.
Клетки ресуспендируют (106 ядерных клеток на 100 мкл) в буфере 50 мМ Tris-HCl (рН 8,3), 50 мМ KCl, 1,5 мМ MgCl2, 0,5% Tween 20, 0,5% NP40, с добавлением 100 мкг/мл протеиназы К. После инкубации в течение 2 часов при температуре 56°С, для инактивации протеиназы К клетки нагревают в течение 10 минут до 95°С и немедленно переносят на влажный лед (резкое охлаждение). В случае, если присутствуют крупные агрегаты, следует выполнить еще один цикл расщепления в таком же буфере. Для амплификации используется десять мкл данного экстракта.
При экстракции ДНК из тканей, например, из клеток ворсинок хориона или объединенных культивируемых клеток, количество выше указанного буфера с протеиназой К может изменяться в зависимости от размера образца ткани. Данный экстракт инкубируют в течение 4-10 часов при температуре 50-60°С и затем для инактивации протеиназы в течение 10 минут при температуре 95°С. Во время более длительной инкубации следует после примерно 4 часов добавлять свежую протеиназу К в первоначальной концентрации.
Если образец содержит небольшое количество клеток, то экстракцию можно выполнять при помощи способов, описанных в работе Higuchi, "Simple and Rapid Preparation of Samples for PCR", in PCR Technology, Ehrlich, H. A. (ed.),Stockton Press, New York, которая включена сюда в качестве ссылки. PCR можно применять для амплификации целевых областей в очень малом количестве клеток (1000-5000), полученных из индивидуальных колоний костного мозга и культур периферической крови. Клетки образца суспендируют в 20 мкл лизирующего буфера для PCR (10 мМ Tris-HCl (рН 8,3), 50 мМ KCl, 2,5 мМ MgCl2, 0,1 мг/мл желатина, 0,45% NP40, 0,45% Tween 20) и замораживают до использования. В случае, когда должна быть выполнена PCR, к клеткам в лизирующий буфер для PCR добавляют 0,6 мкл протеиназы К (2 мг/мл). Затем данный образец нагревают примерно до 60°С и инкубируют в течение 1 часа. Расщепление останавливают посредством инактивации протеиназы К путем нагревания в течение 10 минут до температуры 95°С и затем охлаждают на льду.
Относительно простой процедурой выделения ДНК для PCR является процедура высаливания, адаптированная из способа, описанного в работе Miller et al., Nucleic Acids Res. 16:1215 (1988), которая включена здесь в качестве ссылки. Мононуклеарные клетки разделяются в градиенте Ficoll-Hypaque. Клетки ресуспендируют в 3 мл лизирующего буфера (10 мМ Tris-HCl, 400 мМ NaCl, 2 мМ Na2EDTA, pH 8,2). К клеткам добавляют пятьдесят мкл раствора протеиназы К в концентрации 20 мг/мл и 150 мкл 20% раствора SDS и затем инкубируют всю ночь при температуре 37°С. Покачивание пробирки во время инкубации улучшает расщепление образца. Если после ночной инкубации расщепление протеиназой К остается неполным (фрагменты еще видны), дополнительные 50 мкл раствора протеиназы К в концентрации 20 мг/мл смешивают с раствором и инкубируют еще одну ночь при температуре 37°С на слабо покачивающейся или вращающейся платформе. После адекватного расщепления один мл 6М раствора NaCl добавляют к образцу и энергично перемешивают. Полученный в результате раствор центрифугируют в течение 15 минут при 3000 об./мин. Осадок содержит осажденные клеточные белки, в то время как супернатант содержит ДНК. Супернатант удаляют в пробирку на 15 мл, в которой находится 4 мл изопропанола. Содержимое пробирки аккуратно перемешивают до тех пор, пока смешаются водная и спиртовая фазы и образуется белый осадок ДНК. Осадок ДНК удаляют и помещают в раствор 70% этанола и аккуратно перемешивают. Осадок ДНК удаляют из этанола и высушивают на воздухе. Осадок помещают в дистиллированную воду и растворяют.
Наборы для экстракции высокомолекулярной ДНК для PCR включают Genomic Isolation Kit A.S.A.P. (Boehringer Mannheim, Indianapolis, Ind.), Genomic DNA Isolation System (GIBCO BRL, Gaithersburg, Md.), Elu-Quik DNA Purification Kit (Schleicher & Schuell, Keene, N. H.), DNA Extraction Kit (Stratagene, LaJolla, Calif.), TurboGen Isolation Kit (Invitrogen, San Diego, Calif.) и тому подобное. Использование этих наборов согласно инструкциям производителей, как правило, приемлемо для очистки ДНК перед осуществлением способов настоящего изобретения.
Концентрацию и чистоту экстрагированной ДНК можно определять при помощи спектрофотометрического анализа абсорбции разбавленной аликвоты при 260 нм и 280 нм. После экстракции ДНК можно проводить PCR-амплификацию. Первый этап каждого цикла PCR включает в себя разделение двойной нуклеиновой кислоты, образованной при наращивании праймера. Как только цепи разделяются, следующий этап PCR включает в себя гибридизацию разделенных цепей с праймерами, которые ограничивают целевую последовательность. Затем праймеры наращиваются с образованием комплементарных копий целевых цепей. Для успешной PCR-амплификации праймеры конструируют таким образом, что положение, в котором каждый праймер гибридизуется вдоль двойной последовательности, таково, что синтезированный продукт наращивания с одного праймера после разделения от матрицы (комплемента) служит в качестве матрицы для наращивания другого праймера. Цикл денатурации, гибридизации и наращивания повторяется так много раз, сколько это необходимо для получения желаемого количества амплифицированных нуклеиновых кислот.
В особенно полезном осуществлении PCR-амплификации, разделение цепочек достигается путем нагревания реакционной смеси до достаточно высокой температуры в течение значительного времени, для того чтобы вызвать денатурацию двойной цепи, но не привести к необратимой денатурации полимеразы (см. патент США №4965188, включенный сюда в качестве ссылки). Типичная тепловая денатурация включает в себя диапазон температур приблизительно от 80°С до 105°С в диапазоне времени от секунд до минут. Однако, разделение цепей можно выполнять при помощи любого подходящего денатурирующего способа, включая физические, химические или ферментативные средства. Разделение цепей может быть вызвано геликазой или, например, ферментом, способным проявлять геликазную активность. Например, фермент RecA имеет геликазную активность в присутствии АТФ. Подходящие условия реакций для разделения цепей при помощи геликаз известны в данной области (см. в работах Kuhn Hoffman-Berling, 1978, CSH-Quantitative Biology, 43: 63-67; и Radding, 1982, Ann.Rev. Genetics 16: 405-436, каждая из которых включена сюда в качестве ссылки).
Зависимое от матрицы наращивание праймеров в PCR катализируется посредством полимеризации в присутствии адекватных количеств четырех дезоксирибонуклеотидтрифосфатов (типично дАТФ, дГТФ, дЦТФ и дТТФ) в реакционной среде, включающей соответствующие соли, катионы металлов и системы рН-буферов. Приемлемыми агентами полимеризации являются ферменты, которые, как известно, катализируют зависимый от матрицы синтез ДНК. В некоторых случаях целевые области могут кодировать по крайней мере часть экспрессируемого клеткой белка. В этом случае для амплификации целевой области можно использовать мРНК. Альтернативно, PCR можно использовать для создания с РНК библиотеки кДНК для дальнейшей амплификации, причем первоначальной матрицей для наращивания праймера является РНК. Агентами полимеризации, приемлемыми для синтезирования комплементарной, копии ДНК (кДНК), последовательности с матрицы РНК являются такие обратные транскриптазы (RT), как RT вируса птичьего миелобластоза, RT вируса мышиной лейкемии Moloney, или ДНК-полимераза из Thermus thermophilus (Tth), термостабильная ДНК-полимераза с активностью обратной транскриптазы, предоставляемые Perkin Elmer Cetus, Inc. Типично, геномная матрица РНК деградирует при нагревании во время первого этапа денатурации, оставляя после первоначального этапа обратной транскрипции только матрицу ДНК. Приемлемые полимеразы для использования с матрицей ДНК включают в себя, например, ДНК-полимеразу I из E.coli или ее фрагмент Кленова, Т4 ДНК-полимеразу, Tth полимеразу, и Taq полимеразу, стабильную при нагревании ДНК-полимеразу, выделенную из Thermus aquaticus и коммерчески доступную от Perkin Elmer Cetus, Inc. Последний фермент широко используют при амплификации и определении последовательности нуклеиновых кислот. Условия реакции для использования Taq полимеразы известны в данной области и описаны в работе Gelfand, 1989, PCR Technology выше.
Аллель-специфическая PCR
Аллель-специфическая PCR проводит различия между целевыми областями, отличающимися по наличию отсутствия изменений или полиморфизма. Выбирают те праймеры PCR-амплификации, которые связывают только определенные аллели целевой последовательности. Этот способ описан Gibbs, Nucleic Acid Res. 17: 12427-2448 (1989).
Способы скрининга аллель-специфических олигонуклеотидов
Кроме того, диагностические способы скрининга используют способы скрининга аллель-специфических олигонуклеотидов (ASO), как описано в работе Saiki et al., Nature 324: 163-166 (1986). Для любого определенного аллеля создают олигонуклеотиды с одним или несколькими несовпадениями пар оснований. Способ скрининга ASO определяет несовпадения между вариантами целевой геномной или амплифицированной при PCR ДНК и немутантными олигонуклеотидами, показывая повышенное связывание олигонуклеотида относительно мутантного олигонуклеотида. Олигонуклеотидные зонды можно конструировать таким образом, что при низкой жесткости они будут связывать обе полиморфные формы аллеля, а при высокой жесткости будут связывать тот аллель, которому они соответствуют. Альтернативно, можно разработать условия жесткости, при которых по существу получается двойной ответ, то есть, ASO, соответствующий вариантной форме целевой последовательности, будет гибридизоваться к данному аллелю, а не к аллелю дикого типа.
Способ опосредованного лигазой определения аллелей
Целевые области ДНК тестируемого субъекта можно сравнивать с целевыми областями здоровых и пораженных болезнью членов семьи при помощи опосредованного лигазой определения аллелей. См. работу Landegren et al., Science 241: 107-1080 (1988). Лигазу также можно использовать для определения точечных мутаций в реакции амплификации лигирования, описанной в работе Wu et al., Genomics 4: 560-569 (1989). В реакции амплификации лигирования (LAR) используется амплификация специфичной последовательности ДНК, с применением последовательных циклов зависимых от матрицы лигирований, как описано в работах Wu, указанной выше, и Barany, Proc. Nat. Acad. Sci. 88: 189-193 (1990).
Электрофорез в денатурирующем градиентном геле
Продукты амплификации, полученные с использованием полимеразной цепной реакции, можно анализировать посредством использования электрофореза в денатурирующем градиентном геле. Основываясь на различных зависящих от последовательности нуклеотидов свойствах плавления и электрофоретической подвижности ДНК в растворе, можно идентифицировать различные аллели. В условиях повышенной температуры или денатурации молекулы ДНК плавятся в сегментах, называемых доменами плавления. Каждый домен плавления кооперативно плавится при различной, специфичной для оснований температуре плавления (ТМ). Доменами плавления являются по крайней мере 20 пар оснований в длину и это количество может доходить вплоть до нескольких сотен пар оснований.
Дифференциация между аллелями, основанная на различиях, определяемых последовательностью доменов плавления, можно оценить, используя электрофорез в полиакриламидном геле, как описано в главе 7 из Erlich, ed., PCR Technology, Principles and Applications for DNA Amplification, W.H.Freeman and Co., New York (1992), содержание которой включено сюда в качестве ссылки.
В общем случае, целевую последовательность, которую необходимо проанализировать при помощи электрофореза в денатурирующем градиентном геле, амплифицируют с использованием ограничивающих целевую область праймеров PCR. Амплифицированный продукт PCR вносят в полиакриламидный гель с линейным денатурирующим градиентом, как описано в работах Myers et al., Meth. Enzymol. 155: 501-527 (1986), и Myers et al., in Genomic Analysis, A Practical Approach, K. Davies Ed. IRL Press Limited, Oxford, pp.95-139 (1988), содержание которых включено сюда в качестве ссылки. В системе электрофореза поддерживают температуру чуть ниже Tm доменов плавления целевых последовательностей.
В альтернативном способе электрофореза в денатурирующем градиентном геле целевые последовательности можно первоначально присоединять к участку с нуклеотидами GC, называемому GC-скобка, как описано в главе 7 из Erlich, указанной выше. Предпочтительно, чтобы по крайней мере 80% нуклеотидов в GC-скобке являлись либо гуанинами, либо цитозинами. Предпочтительно, чтобы GC-скобка содержала по крайней мере 30 оснований в длину. Данный способ особенно подходит для целевых последовательностей с высокими Tm.
В общем случае, целевую последовательность амплифицируют посредством полимеразной цепной реакции, как описано выше. Один из олигонуклеотидных праймеров PCR несет на своем 5' конце область GC-скобки, по крайней мере 30 оснований, богатой GC последовательности, которая во время амплификации включается в 5' конец целевой области. Полученная в результате целевая область разгоняется в электрофорезном геле в денатурирующих градиентных условиях, как описано выше. Фрагменты ДНК, отличающиеся при одиночной замене нуклеотида, будут мигрировать до разных положений в геле, которые можно визуализировать при помощи окраски бромидом этидия.
Электрофорез в температурном градиентном геле
Электрофорез в температурном градиентном геле (TGGE) основан на тех же самых основополагающих принципах, что и электрофорез в денатурирующем градиентном геле, за исключением того, что денатурирующий градиент получают при помощи разницы температуры вместо использования разницы концентраций химических денатурантов. Для стандартного TGGE используют аппарат для электрофореза с градиентом температур, идущим вдоль электрофоретической дорожки. Поскольку образцы перемещаются в геле с однородной концентрацией химического денатуранта, они претерпевают увеличение температур. В альтернативном способе TGGE, электрофорезе во временном температурном градиентном геле (TTGE или tTGGE), для достижения такого же результата используют постоянно увеличивающуюся температуру для всего электрофорезного геля. Пока образцы перемещаются в геле, температура всего геля увеличивается, приводя к тому, что образцы во время их перемещения в геле претерпевают увеличение температуры. Приготовление образцов, включая амплификацию при PCR с включением GC-скобки и визуализация продуктов, такое же, что и при электрофорезе в денатурирующем градиентном геле.
Анализ полиморфизма одноцепочечной конформации
Целевые последовательности или аллели локуса PRKAG3 можно различать с использованием анализа полиморфизма одноцепочечной конформации, который идентифицирует различия оснований по изменению электрофоретической подвижности одноцепочечных продуктов PCR, как описано в работе Orita et al., Proc. Nat. Acad. Sci. 85: 2766-2770 (1989). Амплифицированные продукты PCR можно получать, как описано выше, и нагревать или денатурировать иным способом для того, чтобы образовались одноцепочечные продукты амплификации. Одноцепочечные нуклеиновые кислоты могут повторно сворачиваться или формировать вторичные структуры, которые частично зависят от последовательности оснований. Таким образом, по электрофоретической подвижности одноцепочечных продуктов амплификации можно определить изменение в последовательности оснований между аллелями или целевыми последовательностями.
Химическое или ферментативное расщепление несовпадающих оснований.
Различия между целевыми последовательностями можно также определять при помощи дифференциального химического расщепления несовпадающих пар оснований, как описано в работе Grompe et al., Am. J. Hum. Genet. 48: 212-222 (1991), в другом способе различия между целевыми последовательностями можно также определять при помощи ферментативного расщепления несовпадающих пар оснований, как описано в работе Nelson et al., Nature Genetics 4: 11-18 (1993). Короче говоря, генетический материал животного и пораженного болезнью члена семьи можно использовать для создания свободных от несовпадений гетерогибридных дуплексов ДНК. Используемый здесь термин, «гетерогибридный» означает двойную цепочку ДНК, состоящую из одной цепочки одного животного, и второй цепочки от другого животного, обычно отличающегося по фенотипу интересующего признака животного. Положительная селекция свободных от несовпадений гетерогибридов позволяет определить небольшие вставки, делеции или другие полиморфизмы, которые могут быть связаны с полиморфизмами PRKAG3.
Безгелевые системы
Другие возможные способы включают такие безгелевые системы, как TaqMan™ (Perkin Elmer). В этой системе олигонуклеотидные праймеры PCR конструируют таким образом, чтобы ограничить рассматриваемую мутацию и позволить проведение PCR-амплификации данной области. Затем конструируют третий олигонуклеотидный зонд для гибридизации области, содержащей основание, подвергшееся изменению между различными аллелями гена. Этот зонд помечают флуоресцентными красителями как с 5', так и 3' конца. Данные красители выбирают таким образом, чтобы при данной близости друг к другу, флуоресценция одного из них тушилась другой и не могла определяться. Наращивание при помощи Taq полимеразы праймера PCR, расположенного на 5' конце матрицы относительно зонда, приводит к расщеплению красителя, присоединенного к 5' концу отожженного зонда вследствие 5' нуклеазной активности Taq ДНК-полимеразы. Это удаляет эффект тушения, позволяя определять флуоресценцию красителя на 3' конца зонда. Установление различий между разными последовательностями ДНК возникает благодаря тому факту, что если гибридизация зонда к молекуле матрицы не полная, то есть существует несовпадение некоторых форм, расщепление красителя не имеет места. Таким образом, удаление эффекта тушения будет происходить только в том случае, если нуклеотидная последовательность олигонуклеотидного зонда полностью комплементарна к молекуле матрицы, с которой он связан. Реакционная смесь может содержать две различные последовательности зонда, каждая из которых сконструирована для различных аллелей, которые могли бы присутствовать, позволяя таким образом определять оба аллеля в одной реакции.
Другой способ все же включает Invader Assay, который содержит изотермическую амплификацию, которая основана на каталитическом испускании флуоресценции. См. Third Wave Technology на сайте www.twt.com.
Диагностика ДНК, не основанная на PCR
Идентификацию сцепленной с PRKAG3 последовательности ДНК можно выполнить без основанного на полиморфизмах этапа амплификации, включая полиморфизмы длин рестрикционных фрагментов животного и члена семьи. В общем случае зондами гибридизации являются олигонуклеотиды, которые связываются посредством комплементарного спаривания оснований со всей или с частью целевой нуклеиновой кислоты. Зонды типично связываются с целевыми последовательностями, имеющими недостаток полной комплементарности с последовательностью зонда, зависящей от жесткости условий гибридизации. Зонды предпочтительно помечают прямым или непрямым способом так, что, анализируя наличие или отсутствие зонда, можно определять наличие или отсутствие целевой последовательности. Способы прямого мечения включают в себя мечение такими радиоизотопами, как 32P или 35S. Способы непрямого мечения включают в себя флуоресцентные метки, комплексы с биотином, которые могут связываться с авидином или стрептавидином, или пептидные или белковые метки. Способы визуального определения включают средства фотолюминесценции, техасский красный, родамин и его производные, краситель red leuco и 3,3',5,5'тетраметилбензидин (TMB), флуоресцеин и его производные, дансил, умбиллиферон и ему подобные средства или пероксидазу хрена, алкалинфосфатазу и ей подобные средства.
Зонды гибридизации включают любую нуклеотидную последовательность, обеспечивающую гибридизацию свиной хромосомы, где располагается PRKAG3, и, таким образом, определение сцепленного с PRKAG3 генетического маркера, включая полиморфизм длины рестрикционного фрагмента, гипервариабельную область, повторяющиеся элементы или тандемные повторы различного количества. Зонды гибридизации могут находиться в любом гене или подходящем аналоге. Кроме того, подходящие зонды гибридизации включают фрагменты экзонов или участков кДНК или генов, которые, как известно, картированы в подходящую область хромосомы.
Предпочтительными зондами гибридизации тандемных повторов, для использования согласно настоящему изобретению, являются зонды, которые распознают небольшое количество фрагментов в определенном локусе при условиях гибридизации высокой жесткости, или которые распознают большое количество фрагментов в определенном локусе в случае, если условия жесткости занижены.
Можно использовать один или несколько дополнительных ферментов рестрикции и/или зондов и/или праймеров. Дополнительные ферменты, сконструированные зонды и праймеры можно определять посредством обычного экспериментирования обычным специалистом в данной области, и они предназначены для включения в объем данного изобретения.
Несмотря на то, что способы, описанные здесь, могут относиться к использованию одного рестрикционного фермента или одного набора праймеров, способы не ограничены таким образом. При желании можно использовать один или несколько дополнительных ферментов рестрикции и/или зондов и/или праймеров. На самом деле в некоторых ситуациях можно предпочтительно использовать комбинации маркеров, дающих определенный гаплотип. Дополнительные ферменты, сконструированные зонды и праймеры можно определять посредством обычного экспериментирования, в сочетании с предоставленными и включенными сюда указаниями.
Согласно данному изобретению были идентифицированы полиморфизмы гена PRKAG3, которые связаны с качеством мяса и численностью потомства. В одном осуществлении наличие или отсутствие маркеров можно анализировать посредством анализа RFLP PCR с использованием рестрикционных эндонуклеаз, и праймеры амплификации можно конструировать с использованием аналогичных генов человека, свиньи или других родственных PRKAG3 генов вследствие высокой гомологии в окружающей полиморфизмы области, или можно конструировать их с использованием известных данных последовательности гена PRKAG3, как проиллюстрировано в GenBank, или даже можно конструировать из последовательностей, полученных из данных о сцеплении генов близкого окружения, основанных на имеющихся здесь указаниях и ссылках. Последовательности, окружающие полиморфизм, облегчают разработку альтернативных тестов PCR, в которых праймер из приблизительно 4-30 смежных оснований, взятый из непосредственно прилегающей к полиморфизму последовательности, используется в полимеразной цепной реакции для значительной амплификации области перед обработкой желаемым рестрикционным ферментом. Не обязательно, чтобы праймеры были точно комплементарны; существенно, чтобы были приемлемы эквивалентные последовательности. Конструирование праймеров для амплификации путем PCR, известно специалистам в данной области и подробно обсуждается в Ausubel (ed.),"Short Protocols in Molecular Biology, Fourth Edition" John Wiley and Sons, 1999. Далее следует короткое описание конструирования праймера.
Стратегия конструирования праймера
Увеличенное использование способов полимеразной цепной реакции (PCR) стимулировало разработку большого количества программ для помощи при конструировании и выборе используемых в качестве праймеров для PCR олигонуклеотидов. Четырьмя примерами таких программ, которые свободно доступны через Интернет, являются: PRIMER, созданная Mark Daly и Steve Lincoln из Whitehead Institute (для UNIX, VMS, DOS, и Macintosh), Oligonucleotide Selection Program (OSP), созданная Phil Green и LaDeana Hiller из Washington University в St. Louis (для UNIX, VMS, DOS, и Macintosh), PGEN, созданная Yoshi (только для DOS), и Amplify, созданная Bill Engels из University of Wisconsin (только для Macintosh). В общем случае эти программы помогают конструировать праймеры PCR, путем поиска участков известных элементов повторяющейся последовательности и затем оптимизации Tm, анализируя длину и содержание GC предполагаемого праймера. Также доступна коммерческая программная продукция, и процедура выбора праймера быстро включается в большинство пакетов анализа последовательности.
Определение последовательности и PCR праймеров
Конструирование олигонуклеотидов для использования либо в качестве праймеров секвенирования, либо PCR праймеров требует выбора соответствующей последовательности, которая специфически распознает целевые участки, и затем проверки последовательности, для того, чтобы устранить возможность того, что у олигонуклеотида будет стабильная вторичная структура. Инвертированные повторы последовательности можно идентифицировать с использованием программы для идентификации повторов или сворачивания РНК, такие, как описано выше (см. предсказание структуры нуклеиновых кислот). Если замечают возможную исходную структуру, то в последовательности праймера можно сдвинуть несколько нуклеотидов в любом направлении для минимизации предсказанной вторичной структуры. Последовательность олигонуклеотида следует также сравнивать с последовательностями обеих цепей соответствующего вектора и ДНК вставки. Очевидно, что у последовательности праймера должно быть только одно совпадение с целевой ДНК. Также целесообразно исключать праймеры, которые имеют только одно несовпадение с нежелательной целевой последовательностью ДНК. Для используемых при амплификации геномной ДНК праймеров PCR, последовательность праймера следует сравнивать с последовательностями из базы данных GenBank для того, чтобы определить, имеют ли место какие-либо значительные совпадения. В случае, если последовательность олигонуклеотида присутствует в какой-либо известной последовательности ДНК или, что более важно, в каком-либо известном повторяющемся элементе, последовательность праймера следует изменить.
Способы и материалы изобретения можно также использовать в более общих случаях для оценки ДНК свиньи, генетического типирования индивидуальных свиней и определения генетических различий у свиней. В частности, образец геномной ДНК свиньи можно оценить по соотношению с одним или несколькими контролями для того, чтобы определить, присутствует ли полиморфизм гена PRKAG3. Предпочтительно, чтобы относительно гена PRKAG3 проводился анализ RFLP, и результаты его сравнивались с контролем. Контролем является результат анализа RFLP гена PRKAG3 свиньи у другой свиньи, для которой известен полиморфизм гена PRKAG3. Точно так же генотип PRKAG3 свиньи можно определить, получая образец ее геномной ДНК, проводя анализ RFLP гена PRKAG3 и сравнивая результаты с контролем. Опять же, контролем является результат анализа RFLP гена PRKAG3 другой свиньи. По данным результатам типируют свинью, определяя полиморфизм в ее гене PRKAG3. В конечном счете, генетические различия среди свиней можно определять, получая образец геномной ДНК по крайней мере двух свиней, идентифицируя наличие или отсутствие полиморфизма гена PRKAG3 и сравнивая результаты.
Эти анализы пригодны для идентификации генетических маркеров, связанных с качеством мяса, как обсуждалось выше, для идентификации других полиморфизмов гена PRKAG3, которые могут коррелировать с другими характеристиками, такими как численность потомства, и для общего анализа генотипов и фенотипов свиней.
Генетические маркеры, способы и новые аллели данного изобретения также пригодны в селекционной программе для улучшения качества мяса и/или репродуктивной эффективности (численности потомства) породы, линии или популяции свиней. В некоторых ситуациях продолжительная селекция и разведение свиноматок, которые по крайней мере гетерозиготны и предпочтительно гомозиготны по полиморфизму, связанному с предпочтительным качеством мяса, должны также привести к увеличению численности потомства. Это следует применять в популяционных исследованиях в примере 2.
В примерах и методах здесь описаны некоторые гены, в которых, как было идентифицировано, имеется полиморфизм, который связан либо положительно, либо отрицательно с выгодным признаком, который имеет влияние на качество мяса/численность потомства животных, имеющих данный полиморфизм. Идентификацию наличия полиморфизма гена часто проводят при помощи выбора единичного основания, что приводит к появлению сайта рестрикции в определенных аллельных формах. Однако, определенный аллель, как показано и как обсуждалось здесь, может иметь в себе множество изменений оснований, которые можно было бы проанализировать, которые указывают на тот же самый полиморфизм (аллель). Кроме того, другие генетические маркеры или гены могут быть связаны с описанным здесь полиморфизмом таким образом, что анализы могут включать в себя идентификацию других генов или фрагментов генов, но от которых, в конечном счете, зависит генетическая характеристика животных по тому же самому полиморфизму. Любой анализ, который сортирует и идентифицирует животных, основываясь на описанных здесь аллельных различиях, предназначен для включения в объем данного изобретения.
Специалист в данной области поймет, что существует множество способов генотипирования животных по данному полиморфизму, когда он был идентифицирован и установлена корреляция с определенным признаком. Конструирование таких альтернативных тестов просто представляет оптимизацию параметров, известных специалистам в данной области, и предназначено для включения в объем данного изобретения, как полностью описано здесь.
Материал и способы
Картирование родословных, сцепления и QTL. Авторами изобретения было проведено скрещивание беркширской и йоркширской пород свиней (BxY), что дало 525 потомка F2, и данное потомство использовали для картирования QTL в отношении качества мяса (Malek et al., 2001) с использованием интервального способа картирования (Haley et al., 1994). Для данного скрещивания выбрали беркширскую породу, поскольку она рассматривается как характеризующаяся очень хорошим качеством мяса, в частности, в терминах pH, цвета, способности удерживать влагу и нежности. Ген PRKAG3 картировали на карте сцепления семейства BxY с использованием программы картирования CRI-MAP (версия 2.4) (Green et al., 1990). Интервальный способ картирования (Haley et al., 1994), включающий информацию об участке PRKAG3, использовали для картирования QTL в отношении качества мяса на свиной хромосоме 15 (SSC 15) (фигура 1). Эффекты QTL были оценены и представляли собой средний аллельный эффект Беркширской породы и средний аллельный эффект Йоркширской породы.
Образцы ткани и выделение ДНК/РНК. Образцы крови и фенотипы собирали и записывали для животных F0, F1 и F2 скрещивающегося семейства (Malek et al., 2001) вместе с образцами крови и мышечной ткани из области окорока и спинной части от некоторых животных F3. Авторами изобретения также была собрана большая коллекция образцов крови от пяти различных коммерческих линий свиней (ландрас, большая белая, дюрок, дюрок-синтетическая и беркширская). Геномную ДНК выделяли из цельной крови путем стандартных процедур высаливания, и экстрагировали общую РНК из мышечной ткани окорока и спинной части с использованием способа, основанного на реагенте TRIzol по инструкциям производителя (GIBCO/BRL, Rockville, Мэриленд).
PCR, RT-PCR, RACE и обнаружение полиморфизма. Основываясь на последовательности свиного гена PRKAG3, доступной в GenBank (AF214521), авторы изобретения сконструировали праймеры для амплификации всех кодирующих регионов гена PRKAG3. Реакции PCR выполняли с использованием 12,5 нг свиной геномной ДНК, 1,5 мМ MgCl2, 0,125 мМ dNTP, 0,3 мкМ каждого праймера и 0,35 ед. ДНК-полимеразы Taq (Promega, Мэдисон, Висконсин) и буфера для PCR (10 мМ Tris-HCl, 50 мМ KCl и 0,1% Triton®X-100) в конечном объеме 10 мкл. Обратную транскрипцию общей РНК (3,5 мкг) проводили путем случайной гексануклеотидной затравки и Superscript II (GIBCO/BRL, Rockville, Мэриленд) по протоколу производителя (праймеры: набор A, прямой 5'ATGAGCTTCCTAGAGCAAGGAG 3' и обратный 5'CAGGTCTCAATCTTATGTTCTTC 3'; набор B, прямой 5'CGTCCGAGCGGCACCTTTGT 3', и обратный 5'AAGGTTCCAAGGTTCTCAGGC 3'). Эксперименты 5'-концевой быстрой амплификации концов кДНК (RACE) проводили с использованием набора FirstChoice RLM RACE (Ambion, Austin, Техас) по инструкциям производителя, после определения последовательности продуктов PCR (геноспецифические праймеры: внешний 5'CCCACGAAGCTCTGCTTCTT 3', и внутренний 5'TCCTTGCTCTAGGAAGCTCAT 3'). Последовательность ампликонов определяли с использованием концевых меток (PE Applied Biosystems, Foster City, Калифорния) на автоматическом секвенаторе ABI 377. Авторы изобретения использовали программное обеспечение Sequencher (Gene Codes Corporation, версия 4.0.5, Ann Arbor, Мичиган) для объединения последовательностей и идентификации полиморфизмов.
Генотипирование и анализ PCR-RFLP. Регион, фланкирующий каждую из анализированных бессмысленных мутаций, амплифицировали с использованием одной пары праймеров для замен T30N и G52S (прямой 5'ATGAGCTTCCTAGAGCAAGGAG 3' и обратный 5'GGCTGCATGATGTTATGTGCCT 3') и другой пары для I199V (прямой 5'GGAGCAAATGTGCAGACAAG 3' и обратной 5'CCCACGAAGCTCTGCTTCTT 3'). После расщепления ферментами рестрикции BsaHI (для I199V), HphI (для G52S) и StyI (для T30N) расщепленные продукты PCR разделяли на гелях 4%-ной агарозы NuSieve (FMC, Rockland, Мэн) и окрашивали этидий-бромидом. Для полиморфизма SINE после амплификации путем PCR (праймеры: прямой 5'GAAACTCTTCTCCCCACAGAC 3' и обратный 5'GGCTGCATGATGTTATGTGCCT 3') осуществляли разделение продуктов на 1%-ном агарозном геле (AMRESCO, Solon, Огайо). После генотипирования на предмет данных полиморфизмов всех животных с гаплотипом 2 (таблица 6) также генотипировали на предмет замены R200Q для повышения шанса на обнаружение аллеля RN- или 200Q аллеля (см. Milan et al., 2000). Были обнаружены две гомозиготные особи по аллелю 200Q и четыре носителя, и их удаляли для дальнейшего статистического анализа, так что мутация RN- не влияла на проведенный авторами изобретения анализ других замен. Для замены R200Q авторы использовали те же праймеры, что для мутации I199V, и расщепление проводили ферментом рестрикции BsrBI. В качестве окончательной проверки также анализировали случайную выборку примерно из 100 животных с различными гаплотипами на предмет замены R200Q, но ни одно из животных не несло аллеля 200Q.
Измерение фенотипических признаков. Измерения фенотипов в семействе BxY проводили с использованием обычных промышленных способов (Malek et al., 2001), и включали оценку pH, цвета и гликолитического потенциала. Данные, относящиеся к пяти коммерческим породам, собирали на коммерческих мясокомбинатах, и получали индивидуальные значения цвета мяса (отражательная способность спинной части туши и окорока - предпочтительными являются более низкие значения) и индивидуальные значения рН спинной части туши и окорока через 24 часа после забоя (предпочтительными являются более высокие значения). Данные, полученные на мясокомбинате, не включали измерений гликогена и гликолитического потенциала. Измерения фенотипических признаков цвета и рН представляют собой промышленные способы измерения качества мяса, которые опосредованно коррелируют с содержанием гликогена и гликолитическим потенциалом.
Статистический анализ популяции гибридов F2 беркширской x йоркширской породы. Ассоциации между заменой PRKAG3 I199V и признаками содержания гликогена, лактата, гликолитического потенциала и качества мяса в популяции F2 BxY тестировали с использованием процедур общих линейных моделей (SAS®, процедура GLM, SAS Institute Inc., Cary, Северная Каролина), с моделью, которая учитывала свиноматку-производительницу, дату забоя, пол и генотип I199V. При замене I199V получали наименьшие квадратичные средние для всех трех генотипов.
Анализ коммерческих пород. Ассоциации между полиморфизмами PRKAG3 и признаками качества мяса тестировали с использованием процедур смешанных моделей (процедура SAS® MIXED, SAS Institute Inc., Cary, Северная Каролина) с моделью, которая всегда учитывала хряка-производителя в качестве случайного эффекта, и дату забоя и маркерный(ые) генотип(ы) в качестве фиксированных эффектов. Породу добавляли в качестве фиксированного эффекта при анализе животных всех пород. Признаки пола и фермы не включали, поскольку все признаки измеряли только на самках, и каждой дате забоя соответствовало не более одной фермы. Поскольку самцов не включали в данную часть анализа, результаты авторов изобретения по BxY не подразумевают действия пола на генотип. Полную модель, включающую эффекты различных генотипов, для каждого из трех заменяющих сайтов подгоняли для пяти коммерческих пород. Незначимые генотипические эффекты удаляли путем обратной элиминации (p для удаления > 0,10) с целью идентификации того, какие замены ассоциированы с действием на признаки качества мяса.
Средние значения наименьших квадратов (LS) трех классов генотипов получали по каждой из коммерческих пород для каждой из индивидуально анализируемых замен. Не было обнаружено породы с взаимодействиями генотипов, и поэтому для увеличения надежности оценок аллельных эффектов данные от пяти пород объединяли для межпородного анализа.
Комбинированные эффекты трех замен оценивали как эффекты замен в гаплотипе. Отличия между гаплотипами оценивали посредством модели, учитывающей хряка-производителя (случайно), сутки забоя и одну переменную для каждого гаплотипа, принимающую значения -1, 0 и 1, соответствующие животному, имеющему 0, 1 или 2 копии данного гаплотипа. Эффекты замен в гаплотипе были представлены как отклонения от среднего из гаплотипов и отражали различия от наихудшего до наилучшего гаплотипа. Число животных, использованное в анализах ассоциации, изменялось в зависимости от измеряемого признака, и эти данные перечислены в таблицах 3, 4 и 5.
Результаты
Разработка маркеров и картирование сцепления. Некоторые значимые QTL детектировали на SSC15 (Malek et al., 2001) в регионе, где локализуется ген PRKAG3 (Milan et al., 2000), между маркерами SW1683 и SW1983 (фигура 1). Они включали QTL для среднего содержания гликогена и гликолитического потенциала, на которые, как сообщалось (Milan et al., 2000), влияет аллель PRKAG3 200Q, также как на признаки рН 24-часовых окорока и спинной части туши и на значения L коэффициента Hunter 24-часовой спинной части туши (отражение света). Предпочтительный аллель для данного QTL, который, что интересно, имеет дополнительный эффект (доминантной является мутация RN-), был выделен преимущественно у беркширской породы (которая, в общем, считается характеризующейся мясом очень хорошего качества), как и ожидалось (таблица 1). Ген PRKAG3 являлся уникальным геном-кандидатом в данной области, что основано на недавней разработке BAC-контига свиного RN-региона (Milan et al., 2000), на высокой степени сохранения порядка сцепления свиной карты данной области по сравнению с картой человеческого транскрипта (Jeon et al., 2001) и на недавно разработанной карте генома человека (Lander et al., 2001). Авторы изобретения вначале тестировали племенных животных, двух хряков-производителей беркширской породы и девять свиноматок йоркширской породы, на предмет опубликованной замены RN- (R200Q). Все племенные животные имели аллель rn+ (200R). Путем определения всего кодирующего региона гена PRKAG3 животных-родоначальников семейства BxY и четырех особей F3 с наивысшими значениями качества мяса, авторы изобретения идентифицировали три бессмысленных мутации. Они представляли собой замены T30N и I199V, описанные ранее (Milan et al., 2000) и новую бессмысленную мутацию (G52S). Другая несинонимичная замена (P53L), обнаруженная Milan et al. (2000), не имела вариантов среди животных-родоначальников семейства BxY, которые все характеризовались 53P. По причине недостатка информации по 5'UTR авторы изобретения применяли RACE для обнаружения полной 5'-концевой фланкирующей последовательности и генной организации данного региона. Интронный полиморфизм SINE был обнаружен начиная от 79 н.п. выше старт-кодона, но он присутствовал только у трех свиноматок-родоначальниц йоркширской породы. Основываясь на различиях между животными-родоначальниками гибридного семейства по аллельной частоте каждого участка, авторы изобретения сочли замены G52S и I199V наиболее вероятными кандидатами на QTL качества мяса, о котором сообщалось ранее. С использованием замены I199V авторы изобретения картировали ген PRKAG3 в карте сцепления BxY до позиции ниже широкого(их) пика(ов) QTL для содержания гликогена, лактата и гликолитического потенциала, а также рН мяса после 24 ч хранения (фигура 1). После добавления информации по I199V PRKAG3 длина карты и порядок маркеров на SSC 15 был тем же самым, что в работе Malek et al. (2001). Повторный анализ QTL, включающий I199V PRKAG3 (фигура 1) привел к малым изменениям значения отношения дисперсий и расположения пиков QTL на SSC 15 (от 0 до 3 сантиморган) по сравнению с результатами Malek et al. (2001).
Исследования ассоциации в F2. С использованием анализа ассоциации авторы изобретения обнаружили значимое действие всех трех замен (T30N, G52S и I199V) на среднее содержание гликогена и лактата и также на гликолитический потенциал в популяции F2 BxY (данные показаны только для замены I199V - таблица 2). Наиболее значительное действие, как было выявлено, оказывала замена I199V на большинство анализируемых признаков, включая содержание лактата и гликогена и измерения гликолитического потенциала, но также и на некоторые признаки качества мяса, связанные с этими показателями. Из данных по F2 аллели 30T, 52G и 199I были предпочтительны в смысле качества мяса. При наличии большой предсказанной неравновесности сцепления при скрещивании было необходимо исследовать и подтвердить действие данных мутаций в некоторых коммерческих породах свиней, относящихся к ауткроссингу, для того, чтобы определить, высока ли вероятность того, что данный ген непосредственно участвует в наблюдаемой изменчивости по качеству мяса.
Анализ коммерческих популяций. Генотипические частоты для анализируемых замен представлены в таблице 3. Для всех трех участков замены беркширская порода характеризуется более высокой частотой присутствия генотипов, ассоциированных с низким содержанием гликогена (и более высоким качеством мяса) в скелетных мышцах, основываясь на данных по F2 BxY. Другие коммерческие популяции характеризовались более низкими частотами предпочтительных аллелей, что особенно отмечалось для замены I199V по сравнению с беркширской популяцией. Мутации в PRKAG3 и их ассоциации с качеством мяса тестировали для каждой из пяти коммерческих пород и также по всем породам.
При обратной элиминации участков замены в случае анализа по всем породам I199V сохранялась в модели для всех шести признаков, G52S сохранялась для рН окорока, pH спинной части туши, коэффициента Minolta L спинной части туши и коэффициента Minolta b спинной части туши, а T30N сохранялась для коэффициента Minolta L окорока и коэффициента Minolta b окорока.
Поскольку каждая из замен продемонстрировала различные ассоциации по крайней мере с тремя признаками, эффекты каждой замены оценивали независимо. Среднеквадратические оценки генотипических средних по всем породам (таблица 4) и в пределах породы (таблица 5) показали значимое взаимодействие между анализируемыми заменами и мерами качества мяса, указывая на то, что могут существовать некоторые дополнительные (новые) аллели rn+.
При исследовании ассоциаций было выявлено, что наибольшие эффекты по всем породам (таблица 4) и также в пределах пород (таблица 5, данные показаны только для I199V) были получены для замены I199V, что относилось ко всем проанализированным признакам. Для данной замены ассоциации были высоко значимыми (p<0,0005) в отношении всех признаков качества мяса, использованных в данном исследовании при анализе по всем породам. Значимые ассоциации по крайней мере с одним признаком были выявлены для той же замены в пределах каждой из исследованных пород, при высоко значимом действии на коэффициент Minolta b окорока для пород дюрок и дюрок-синтетической и на pH спинной части туши для дюрок-синтетической. Данные две породы (дюрок-синтетическая, дюрок) характеризовались наилучшим распределением часто для анализа ассоциации при наличии существенного числа животных каждого генотипа (таблица 5). При межпородном анализе и в большинстве результатов по индивидуальным породам действие оказывалось в одном направлении для всех признаков, при том что аллель 199I являлся предпочтительным для высокого качества мяса.
Значимые эффекты, но меньшие, чем для I199V, были выявлены для замены T30N по пяти из признаков при межпородном анализе (таблица 4). При внутрипородном анализе T30N эффекты выявляли почти исключительно в популяциях дюрок и дюрок-синтетической (данные не показаны). В большинстве ситуаций действие оказывалось в одном и том же направлении, причем аллель 30T ассоциировался с лучшим качеством мяса.
Для замены G52S значимые (p<0,05) эффекты выявляли только для двух признаков (pH окорока и коэффициент Minolta L спинной части туши) при межпородном анализе, и для данных признаков как предпочтительный был идентифицирован другой аллель. При внутрипородном анализе были выявлены значимые ассоциации лишь в популяции дюрок-синтетической с цветовыми коэффициентами Minolta (данные не показаны).
В пяти тестируемых авторами изобретения коммерческих популяциях ими обнаружено лишь четыре гаплотипа (таблица 6). Беркширская популяция наименее полиморфна, при наличии в ней гаплотипа 3 (30T-52G-199I) с высокой частотой (0,87). В породе большая белая наиболее часто встречается гаплотип 2 (30T-52S-199V), а гаплотип 1 (30N-52G-199V) характеризовался максимальной частотой в популяциях ландрас, дюрок и дюрок-синтетической. Гаплотип 4 (30T-52G-199V) встречался с наименьшей частотой во всех популяциях.
Эффекты замены гаплотипа для каждой породы и по всем породам вычисляли как отклонение от среднего для четырех гаплотипов (Figure 2).При межпородном и внутрипородном анализе были показаны наибольшие различия между гаплотипами для pH окорока и цветовых измерений, в отличие от признаков спинной части туши. Для pH окорока межпородный и внутрипородный анализ показал, что гаплотип 3 оказывал наибольшее действие, которое значимо отличалось от такового для других гаплотипов при межпородном анализе (p < 0,0005) и по крайней мере от одного гаплотипа при анализе каждой индивидуальной породы (p<0,05). Гаплотип 2 был следующим лучшим для большинства признаков, и породы с гаплотипами 1 и 4 имели тенденцию к наихудшему качеству мяса. Данная иерархия неочевидна для беркширской популяции, где наблюдались значимые различия только для гаплотипа 4, характеризовавшегося самым низким значением, соответствующим результатам межпородного анализа. Незначимые результаты, полученные для беркширской породы, вероятно, частично являются следствием низкого уровня полиморфизма у данной породы и сопровождающим его очень низким числом наблюдений гаплотипов 1 и 4. Оценка гаплотипа 4 в популяции дюрок-синтетической оказалась отличной от таковой для других пород (особенно для pH окорока, где она существенно выше, чем для гаплотипа 2 (p<0,05) и гаплотипа 1 (p<0,01), но частота гаплотипа 4 в данной популяции была очень низкой (0,07). Синтетическая природа данной породы (хотя ее создание произошло шесть поколений назад) также обеспечивает благоприятную возможность для присутствия протяженной неравновесности сцепления, повышая шанс сцепленных локусов вносить вклад в эффекты замены гаплотипа.
Результаты по гаплотипам для коэффициентов Minolta соответствовали результатам для pH. Гаплотип 3, как было обнаружено, в основном оказывал предпочтительное действие (низкие значения цветовых коэффициентов). Имело место небольшое количество исключений из результатов для индивидуальных пород, что могло быть следствием выборки. Единственным значимым отклонением является гаплотип 2, который ассоциирован с более низким коэффициентом Minolta b в беркширской породе (p<0,05). При межпородном анализе гаплотип 2 был в большинстве случаев вторым после гаплотипа 3.
Обсуждение
Результаты, приведенные в данной работе, предоставляют убедительные доказательства преимуществ наличия новых аллелей гена PRKAG3, воздействующих на признаки качества мяса. Данное заключение основано на следующих трех пунктах. 1) Известно действие аллелей PRKAG3 rn+ и RN- на качество мяса. 2) Факт наблюдения некоторых QTL для связанных признаков качества мяса, обнаруженных на SSC15 в регионе местонахождения PRKAG3 в семействе BxY. Данные QTL были обнаружены в данном гибриде свиньи, где первоначальная замена R200Q не разделялась; и 3) представленные здесь результаты по ассоциации между заменами в PRKAG3, и содержанием гликогена и лактата, гликолитическим потенциалом и признаками качества мяса в популяции F2 BxY и с признаками качества мяса в некоторых неродственных коммерческих породах свиней.
При анализе ассоциаций отдельных замен было выявлено, что из трех исследованных здесь замен замена I199V характеризовалась наиболее значимыми и большими различиями признаков качества мяса. Например, анализ F2 BxY показал значимые различия между генотипическими классами I199V в плане гликолитического потенциала, но также по содержанию гликогена и лактата (таблица 2). Важные эффекты также были выявлены для большинства анализированных признаков качества мяса. Было обнаружено, что аллель 199I ассоциирован с более низким уровнем гликогена, лактата и гликолитического потенциала, с более высокими значениями pH окорока и спинной части туши, и с лучшими цветовыми коэффициентами. Данный маркер в F2 BxY по существу был информативным в плане обеспечения хороших оценок аллельных эффектов.
При анализе коммерческих популяций замена I199V была ассоциирована со значимыми различиями LS (наименьших квадратичных) средних между гомозиготными классами, составлявшими до 0,14 в породе ландрас и 0,1 по всем породам для pH окорока (таблицы 4 и 5). Для одного из коэффициентов цвета мяса, Minolta L окорока, были обнаружены значимые различия LS (наименьших квадратичных) средних между гомозиготными генотипами, составлявшие до 3,5 единиц отражательной способности (в породе ландрас) и 2,0 по всем породам. Данные эффекты находятся в интервале от 0,5 до 1 фенотипического стандартного отклонения. Значительные отличия также были выявлены для других признаков и пород. Эффекты данного множества признаков, важных для общего качества мяса, очень интересны для животноводческой промышленности.
Кроме I199V большие эффекты также были выявлены при анализе единственной замены T30N. Однако, при включении в данный анализ также I199V сохранялись лишь умеренные эффекты замены T30N. Следует считать, что за эффекты, выявленные для участка 30, большей частью несет ответственность сильная неравновесность сцепления между участками 30 и 199. Малые эффекты, которые по большей части были незначимыми, наблюдались при анализе единственного участка G52S.
Анализ гаплотипов способствовал разделению эффектов несинонимичных замен и предоставлял дополнительные доказательства эффекта позиции 199, а также 52. Гаплотип 3, представлявший собой единственный гаплотип, содержащий 199I, являлся наиболее предпочтительным гаплотипов в отношении pH и измерений цвета мяса. В большинстве тестируемых ситуаций гаплотип 2, представлявший собой единственный гаплотип, содержащий 52S, характеризовался промежуточно ценностью, особенно в отношении признаков качества окорока, где различия в эффектах были более значимыми и более выраженными, чем для других признаков. Ценность гаплотипов 1 и 4 близка к наименьшей, и в большинстве случаев они значимо не отличаются друг от друга.
По наблюдениям того, что значения, соответствующие двум этим гаплотипам (1 и 4), относительно сходны для большинства оценок, позволяет авторам настоящего изобретения сделать вывод, что замена T30N характеризуется минимальным вкладом в изменчивость качества мяса. При межпородном анализе и при анализе пород ландрас и большой белой, в которых частота гаплотипа 4 составляет более 0,10, авторами изобретения обнаружены предпочтительные эффекты гаплотипа 1 на коэффициенты Minolta окорока (данный гаплотип ассоциирован с вариантом 30N) по сравнению с гаплотипом. В других популяциях различия между эффектами данных гаплотипов слабо поддаются оценке вследствие очень низкой частоты присутствия гаплотипа 4.
Различие между гаплотипом 4 и гаплотипом 2 состоит только в участке G52S. Эффекты гаплотипа 4 и 2 значимо различаются для рН и коэффициентов Minolta L и в окороке, и в спинной части туши, при межпородном анализе и для некоторых признаков индивидуальных пород, наиболее заметно, большой белой. Гаплотип 2 (содержащий 52S и кодирующий серин) является предпочтительным по сравнению с гаплотипом 4 (который содержит 52G и кодирует глицин). Это явление противоположно тому, что обнаружено при исследовании BxY, где в качестве предпочтительного аллеля предсказан 52G. Сильная неравновесность сцепления с участком I199V, вследствие ограниченного числа родоначальников F2, может маскировать истинный эффект замены G52S в данной популяции. Интересно, что индивидуальный анализ G52S не показывает какого-либо эффекта для большинства признаков и пород. При таком анализе сравнивается гаплотип 2 с объединенными другими тремя. Из фигуры 2 можно видеть, что смесь трех других гаплотипов может, в зависимости от частот гаплотипов, приводить к получению среднего значения, близкого к таковому для гаплотипа 2, так что разница не может детектироваться при индивидуальном анализе G52S, что указывает на ценность анализа на основе гаплотипов.
Вариант 30T, присутствующий в гаплотипе 4, как было обнаружено, основываясь на анализе единственного участка, является предпочтительным в смысле качества мяса, причем данный вариант ассоциирован со значимыми эффектами в породах дюрок и дюрок-синтетической для большинства признаков. В данных двух популяциях гаплотип 3 характеризуется умеренной частотой (таблица 6) и содержит как 30T, так и предпочтительный вариант 199I. Таким образом, вариант 199I вносит вклад в более выраженное действие варианта с участком 30T за счет неравновесности сцепления.
Авторы настоящего изобретения заключают, что совместный анализ замен и анализ гаплотипов демонстрируют наличие трех несинонимичных замен в гене PRKAG3 с действием различной выраженности на меры качества мяса у свиней. Данная интересная модель «один ген - несколько полиморфизмов - различные фенотипы» основана на наличии различимых добавочных эффектов на комплексный фенотипический признак и может служить в качестве модели для будущих исследований других признаков. Также недавно было сделано предположение о наличии у свиней множественных аллелей как следствия последовательных мутаций в гене при селекции (Jeon et al., 1999; Nezer et al., 1999).
Замена I199V происходит в домене цистатионин-бета-синтетазы (CBS), очень консервативном регионе генов данного семейства (Milan et al., 2000). Роль домена CBS пока неясна, но предполагается, что он участвует в передаче сигнала на цитоплазматические мишени (Pontig, 1997), белок-белковом взаимодействии (Bateman, 1997) и/или регуляции (Bateman, 1997) активности белков. В гене PRKAG3 содержится четыре домена CBS (Milan et al., 2000), и замена I199V локализована в первом и самом консервативном домене. Выравнивание домена CBS и пептида γ3, проведенное с использованием программного обеспечения Pfam, показало, что предпочтительной аминокислотой в данной позиции является изолейцин (результаты не показаны). В данном исследовании интересно то, что была обнаружена ассоциация аллеля 199I (кодирующего изолейцин в участке 199) с лучшим качеством мяса в коммерческих популяциях и семействе F2 BxY и также с низкими уровнями гликогена, лактата и гликолитического потенциала в последнем.
Milan et al. (2000) показали, что вариант 200Q (RN-) всегда обнаруживается совместно с 199V. Однако, 199V обнаруживается как с 200R, так и с 200Q, а 199I всегда обнаруживается с 200R. Так как только три нуклеотида разделяют участки данных замен, вероятность рекомбинации между ними крайне мала. По этой причине авторы данного изобретения считают R200Q заменой, произошедшей наиболее поздно, причем данная гипотеза также поддерживается наличием данной мутации только у гемпширской породы свиней. Оба гаплотипа 199V-200R и 199I-200R могут быть предковыми, поскольку каждый из них был идентифицирован в большинстве проанализированных к настоящему моменту пород (Milan et al., 2000), включая дикого кабана и некоторые другие виды подотряда Suisformes (Ciobanu et al., неопубликованные результаты).
Гаплотип 199V-200R ассоциирован с более высоким содержанием гликогена и более низким значением рН окорока/спинной части туши после забоя по сравнению с гаплотипом 199I-200R (данные по F2 BxY). Замена в кодоне 199 преимущественно приводит к действию на метаболизм глюкозы, и, следовательно, на содержание в мышцах гликогена. Третий гаплотип 199V-200Q дает фенотип RN-. Действие на содержание гликогена, ассоциированное с 199V-200Q, более выражено, чем действие других гаплотипов, и гаплотип 199V-200Q является доминантным по отношению к другим. По этим причинам авторы изобретения предполагают, что фенотип RN- может представлять собой комбинированный эффект гаплотипа 199V-200Q, а не является результатом только замены R200Q. Данный эффект может быть вызван модификацией данными заменами домена CBS.
Конкретные функции регуляторных субъединиц AMPK β и γ пока неясны. Однако, известно, что обе они необходимы для киназной активности (Hardie and Carling, 1997). Эксперименты in vitro показывают, что β-субъединица может играть важную роль в образовании гетеротримерной структуры AMPK, тогда как β взаимодействует и с γ-, и с α-субъединицами, которые непосредственно не взаимодействуют друг с другом (Woods et al., 1996). В последнее время доказано, что в аллостерическом АМФ-связывающем участке могут участвовать как γ-, так и α-субъединицы комплекса AMPK (Cheung et al., 2000). Cheung et al. (2000) предложили элегантную модель, согласно которой в отсутствие AMP гетеротримерный комплекс может быть преимущественно неактивным без взаимодействия γ- и α-субъединиц. В такой ситуации фосфорилирование участка Thr172 в α-субъединице и взаимодействие с субстратами блокируется аутоингибиторным регионом α-субъединицы. В активной форме AMPK аутоингибирование предотвращается за счет взаимодействия α-аутоингибиторного региона с одним или несколькими CBS-доменами γ-субъединицы, и АМФ связывается с со обеими субъединицами, стабилизируя комплекс (Cheung et al., 2000). Информация по выравниванию, предложенная модель регуляции комплекса AMPK и также присутствие рядом участка R200Q поддерживает гипотезу возможного участия замены I199V в активности AMPK. Даже, несмотря на отсутствие разрешенной молекулярной структуры комплекса AMPK, авторы изобретения выдвигают гипотезу о том, что замена аминокислоты также может влиять на структуру и активность фермента, что приводит к наблюдаемому эффекту замены G52S.
Хотя γ3-субъединица выраженно экспрессирована в скелетных мышцах, активность AMPK оказывается в большей степени ассоциированной с изоформами γ1 и γ2 (Cheung et al., 2000). Однако, замена R200Q (или комбинация I199V-R200Q) в гене PRKAG3 по еще неясному механизму вызывает значимые различия в активности AMPK у свиней гемпширской породы (Milan et al., 2000), что указывает на важную роль изоформы γ3 в метаболизме глюкозы скелетной мышцы. Для прояснения ситуации необходимы подробные функциональные исследования различных сочетаний субъединиц. Физиологический смысл роли AMPK в метаболизме глюкозы основан на сравнениях с родственным комплексом SNF1 из дрожжей. Некоторые исследования также показывают, что AMPK участвует в метаболизме гликогена путем инактивации гликогенсинтазы (Carling and Hardie, 1989; Poulter et al., 1988; Zhang et al., 1993), путем активации нитроксидсинтазы (Fryer et al., 2000) и путем усиления перемещения транспортера глюкозы-4 на плазматические мембраны (Hayashi et al., 1998; Kurth Kraczek et al., 1999; Bergeron et al., 1999; Holmes et al., 1999).
Несмотря на то, что эффекты описанных здесь замен на меры качества мяса менее выражены, чем таковые эффекты доминантной мутации RN-, они важны и в биологическом, и в экономическом смысле. В частности, данные аллели сегрегируют во всех коммерческих линиях и породах, проанализированных к настоящему моменту, в отличие от мутации RN-, которая ассоциирована только с гемпширской породой и имеет ограниченное применение в большинстве программ продукции свинины. Сообщаемые здесь результаты по PRKAG3 также указывают на то, что генетикам следует искать дополнительные экономически значимые мутации в генах, известных более сильным действием внутри одного вида и среди нескольких видов. Данное замечание поддерживается сообщениями о больших эффектах, ассоциированных с другими генами, не относящимися к целевому виду или его породам, например, большие эффекты мутаций MC4R у мышей (Huszar et al., 1997) и людей (Yeo et al., 1998) и, в меньшей степени, у свиней (Kim et al., 2000).
Идентификация у видов животных новых биохимически значимых генов также предоставляет полезную информацию в плане новых биохимических мишеней. Данная информация становится еще более значимой при обнаружении новых и интересных аллелей. В случае PRKAG3, возникло предположение (Milan et al., 2000), что данный ген и другие гены, связанные с AMPK у человека, являются интересными кандидатами для участия в диабете II типа, что основано на их функции и локализации QTL, по этой причине эффект данных новых аллелей может обеспечивать новое понимание потенциальных факторов, воздействующих на метаболизм глюкозы, и их следует иметь в виду при дальнейших исследованиях данного заболевания.
Все цитируемые здесь ссылки включены сюда полностью в качестве ссылок, включая в качестве неограничивающих примеров следующие:
Bateman, A., 1997 The structure of a domain common to archaebacteria and the homocystinuria disease protein. Trends Biochem. Sci. 22: 12-13.
Bergeron, R., R.R. Russell 3rd, L.H. Young, J.M.A. Ren, M. Marcucci et al., 1999 Effect of AMPK activation on muscle glucose metabolism in conscious rats. Am. J. Physiol. 276: E938-E944.
Carling, D., and D.G. Hardie, 1989 The substrate and sequence specificity of the AMP-activated protein kinase. Phosphorylation of glycogen synthase and phosphorylase kinase. Biochim. Biophys. Acta 1012: 81-86.
Cheung, P.C., I.P. Salt, S.P. Davies, D.G. Hardie and D. Carling, 2000 Characterization of AMP-activated protein kinase gamma-subunit isoforms and their role in AMP binding. Biochem J. 346: 659-669.
Fryer, L.G., E. Hajduch, F. Rencurel, I.P. Salt, H.S. Hundal et al., 2000 Activation of glucose transport by AMP-activated protein kinase via stimulation of nitric oxide synthase. Diabetes 49: 1978-1985.
Gao, G., S. Fernandez, D. Stapleton, A.S. Auster, J. Widmer, J.R.B. Dyck et al., 1996 Non-catalytic beta- and gamma-subunit isoforms of the 5'-AMP-activated protein kinase. J. Biol. Chem. 271: 8675-8681.
Green, P., K. Falls and S. Crooks, 1990 Documentation for CRIMAP, version 2.4., Washington University, School of Medicine, St. Louis, MO.
Haley, C.S., S.A. Knott and J.M. Elsen, 1994 Mapping quantitative trait loci in crosses between outbred lines using least squares. Genetics 136: 1195-1207.
Hardie, D.G., and D. Carling, 1997 The AMP-activated protein kinase - fuel gauge of the mammalian cell? Eur. J. Biochem. 246: 259-273.
Hardie, D.G., D. Carling and M. Carlson 1998 The AMP-activated/SNFl protein kinase subfamily: metabolic sensors of the eukaryotic cell? Annu. Rev. Biochem. 67: 821-855.
Hayashi, T., M.F. Hirshman, E.J. Kurth, W.W. Winder and L.J. Goodyear, 1998 Evidence for 5' AMP-activated protein kinase mediation of the effect of muscle contraction on glucose transport. Diabetes 47: 1369-1373.
Holmes, B.F., E.J. Kurth-Kraczek and W.W. Winder, 1999 Chronic activation of 5'-AMP-activated protein kinase increases GLUT-4, hexokinase, and glycogen in muscle. J. Appl. Physiol. 87: 1990-1995.
Huszar, D., C.A. Lynch, V. Fairchild-Huntress, J.H. Dunmore, Q. Fang et al., 1997 Targeted disruption of the melanocortin-4 receptor results in obesity in mice. Cell 88: 131-141.
Jeon, J.T., О. Carlborg, A. Tornsten, E. Giuffra, V. Amarger, P. Chardon, L. Andersson-Eklund, K. Andersson, I. Hansson, K. Lundstrom, and L. Andersson, 1999. A paternally expresssed QTL affecting skeletal and cardiac muscle mass in pigs maps to the IGF2 locus. Nat. Genet. 21:157-158.
Jeon, T.J., V. Amarger, C. Rogel-Gaillard, A. Robic, E. Bongcam-Rudloff et al., 2001 Comparative analysis of a BAC contig of the porcine RN region and the human transcript map: implications for the cloning of trait loci. Genomics 72: 297-303.
Kim, K.S., N. Larsen, T. Short, G. Plastow and M.F. Rothschild, 2000 A missense variant of the porcine melanocortin-4 receptor (MC4R) gene is associated with fatness, growth, and feed intake traits. Mamm. Genome 11: 131-135.
Kluijtmans, L.A., G.H. Boers, E.M. Stevens, W.O. Renter, J.P. Kraus et al., 1996 Defective cystathionine beta-synthase regulation by S-adenosylmethionine in a partially pyridoxine responsive homocystinuria patient. J.Clin. Invest. 98: 285-289.
Kurth-Kraczek, E.J., M.F.Hirshman, L.J.Goodyear and W.W.Winder, 1999 5' AMP-activated protein kinase activation causes GLUT4 translocation in skeletal muscle. Diabetes 48: 1667-1671.
Lander, E.S., L.M.Linton, B.Birren, C.Nusbaum, M.C.Zody et al., 2001 Initial sequencing and analysis of the human genome. Nature 409: 860-921.
LeRoy, P., J.Naveau, J.M.Elsen and P.Sellier, 1990 Evidence for a new major gene influencing meat quality in pigs. Genet. Res. 55: 33-40.
Malek, M., J.C.M. Dekkers, H.K.Lee, T.J.Baas, K.Prusa et al., 2001 A molecular genome scan analysis to identify chromosomal regions influencing economic traits in the pig. II. Meat and muscle composition. Mamm. Genome (in press).
Milan, D., J.T.Jeon, C.Looft, V.Amarger, A.Robic et al., 2000 A mutation in PRKAG3 associated with excess glycogen content in pig skeletal muscle. Science 288: 1248-1251.
Monin, G. and P. Sellier, 1985 Pork of low technological quality with a normal rate of muscle pH fall in the immediate postmortem period: The case of Hampshire breed. Meat Sci. 3: 49-63.
Nezer, C., I.Moreau, L.Karim, B.Brouwers, W.Coppieters, J.Detilleux, R.Hanset, A.Kvasz, P.Leroy, and M. Georges 1999. An imprinted QTL with major effects on muscle mass and fat deposition maps to the IGF2 locus in pigs. Nat. Genet., 21: 155-156.
Ponting, C.P., 1997 CBS domains in CIC chloride channels implicated in myotonia and nephrolithiasis (kidney stones). J. Mol. Med. 75: 160-163.
Poulter, L., S.G.Ang, B.W.Gibson, D.H.Williams, C.F.Holmes et al, 1988 Analysis of the in vivo phosphorylation state of rabbit skeletal muscle glycogen synthase by fast-atom-bombardment mass spectrometry. J. Biol. Chem. 175: 497-510.
Sellier, P., 1998. Genetics of meat and carcass traits, pp.463-510 in The Genetics of the Pig, edited by M.F. Rothschild, and A. Ruvinsky. CABI, Wallingford, UK.
Stapleton, D., E.Woollatt, K.I.Mitchelhill, J.K.Nicholl, C.S.Fernandez et al., 1997 AMP-activated protein kinase isoenzyme family: subunit structure and chromosomal location. FEBS Lett. 409: 452-456.
Stapleton, D., K.I. Mitchelhill, G. Gao, J. Widmer, B.J. Michell et al., 1996 Mammalian AMP-activated protein kinase subfamily, J. Biol. Chem. 271: 611-614.
Thornton, C., M.A. Snowden and D. Carling, 1998 Identification of a novel AMP-activated protein kinase beta subunit isoform that is highly expressed in skeletal muscle. J. Biol. Chem. 273: 12443-12450.
Woods, A., P.C.F. Cheung, F.C. Smith, M.D. Davison, J. Scott et al., 1996 Characterization of AMP-activated protein kinase beta and gamma subunits. Assembly of the heterotrimeric complex in vitro. J. Biol. Chem. 271: 10282-10290.
Yeo, G.S., I.S.Farooqi, S.Aminian, D.J.Halsall, R.G.Stanhope et al., 1998 A frameshift mutation in MC4R associated with dominantly inherited human obesity. Nat. Genet. 20: 111-112.
Zhang, W., A.A DePaoli Roach and P.J. Roach, 1993 Mechanisms of multisite phosphorylation and inactivation of rabbit muscle glycogen synthase. Arch. Biochem. Biophys. 304: 219-225.
1
bСуммарные (а) и доминантные (d) эффекты QTL соответствовали генотипическим значениям +а, d и -а, соответственно, для особей, унаследовавших два беркширских аллеля, гетерозигот и особей с двумя йоркширскими аллелями. Положительные суммарные эффекты означают, что беркширские аллели усиливали проявление признака, отрицательные - что беркширские аллели снижали его. Эффекты доминантности приведены относительно среднего двух гомозигот.
c% отклонения = генетическое отклонение в QTL, рассчитанное на основе оцененных суммарных и доминантных эффектов и аллельных частот, равных 1/2, как процент отклонения, оставшегося в F2.
dединицы измерения - мкмоль/г
*значимое на 5% геномном уровне (F>8,22)
** значимое на 1% геномном уровне (F>9,96).
Следующие признаки были незначимыми при p<0,05: коэффициент Minolta окорока, измеренный в лаборатории, коэффициент Hunter окорока, измеренный в лаборатории, и коэффициент Minolta спинной части туши, измеренный на мясоперерабатывающем заводе. Наименьшие квадратичные средние значения оценивались для каждого признака и представлены со стандартными ошибками оценок, приведенными в скобках.
Число животных в каждом генотипическом классе составляло: n=131 (II), 260-265 (IV), и 111-113 (VV). Значимые отличия между среднеквадратичными оценками обозначены 2-буквенными верхними индексами: a-b p<0,05, c-d p<0,005, e-f p<0,0005. Оценка с верхним индексом «a» значимо отличается от оценки(нок) с верхним индексом «b», то же для c-d и e-f и соответствующих им уровней значимости.
Таблица 3. Генотипические частоты замен T30N, G52S и I199V в гене PRKAG3 для пяти коммерческих пород свиней

Наименьшие квадратичные средние значения оценивались отдельно для каждого участка замены и представлены со стандартными ошибками оценок, приведенными в скобках. Значимые отличия между среднеквадратичными оценками (в пределах одного признака и участка замены) обозначены 2-буквенными верхними индексами: a-b p<0,05, c-d p<0,005, e-f p<0,0005. Оценка с верхним индексом «a» значимо отличается от оценки(нок) с верхним индексом «b», то же для c-d и e-f и соответствующих им уровней значимости.
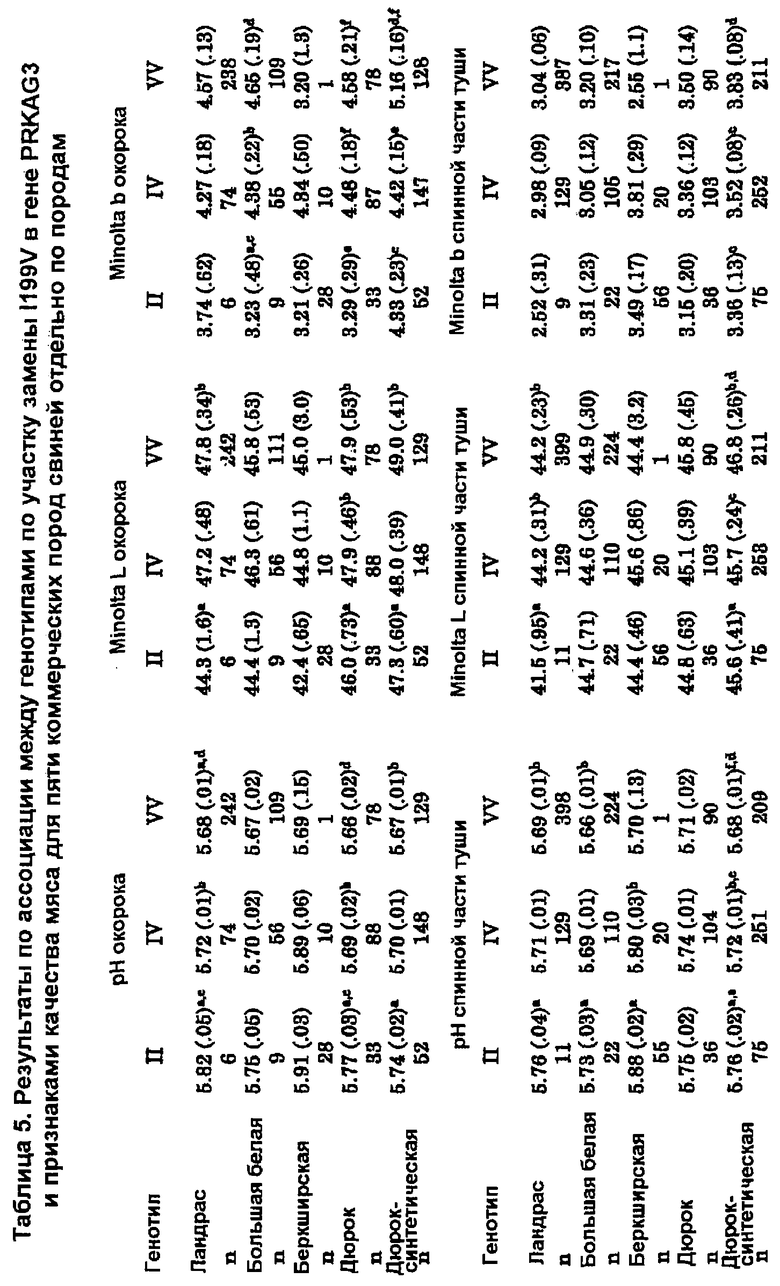
Наименьшие квадратичные средние значения оценивались отдельно для участка замены I119V и представлены со стандартными ошибками оценок, приведенными в скобках. Значимые отличия между среднеквадратичными оценками (в пределах одной породы и участка замены) обозначены 2-буквенными верхними индексами: a-b p<0,05, c-d p<0,005, e-f p<0,0005. Оценка с верхним индексом «a» значимо отличается от оценки(нок) с верхним индексом «b», то же для c-d и e-f и соответствующих им уровней значимости.
спинной
части туши
части туши
части туши
bN - число животных, использованных при анализе ассоциаций гаплотипов.
Пример 2
По данному изобретению было довольно неожиданно показано, что аллели PRKAG3 характеризовались значимой ассоциацией с численностью приплода у животных. Изобретение в данном конкретном осуществлении относится к генетическим маркерам численности приплода у животных. Оно относится к способу скрининга животных с целью определения тех особей, что с большей вероятностью произведут при разведении приплод большей численности, путем идентификации присутствия или отсутствия полиморфизма в гене PRKAG3, который коррелирует с повышенной численностью размера приплода. Применяемый здесь термин «повышенная численность приплода» означает биологически значимое повышение численности приплода по отношению к средней для данной популяции.
Ассоциация между генотипом PRKAG3 и численностью приплода
Полиморфизм кодона 199 PRKAG3 использовали для генотипирования свиноматок с данными о численности приплода. Задействовали две породы, соответствующие породе ландрас (A) и породе дюрок-синтетической (B), в которых ранее обнаруживали ассоциацию между данным полиморфизмом и признаками качества мяса.
Данные анализировали по записям о первом опоросе и по всем опоросам (таблица 7).
Статистически значимая ассоциация (p<0,05) была обнаружена между данным генотипом и признаками численности приплода (общее число рожденных, число рожденных живыми) для породы B в первом опоросе (таблица 8). Было обнаружено, что гетерозигота характеризуется большей численностью приплода, кроме того, генотип 11 характеризовался большими приплодами, чем гомозигота 22 (p<0,3), что указывает на то, что для свиноматок предпочтительно быть носителями по крайней мере одной копии аллеля 1. Интересно то, что сходный эффект наблюдался для породы A, хотя различия у этой породы не достигали статистической значимости, возможно, вследствие меньшего числа наблюдений. Однако в случае большего числа наблюдений разница между гетерозиготой и генотипом 22 по количеству рожденных живыми детенышей приближалась к статистически значимой (p<0,1).
Сходные эффекты наблюдались для набора данных по всем опоросам у двух пород. В данном случае эффект у породы A для общего числа рожденных характеризовался генотипической значимостью, составляющей p<0,08.
Уровни значимости средних LS:
a-b p<0,30; c-d p<0,10; e-f p<0,05; g-h p<0,01; i-j p<0,005.
Протоколы PCR-тестов:
Тест PCR-RFLP для PRKAG3-30
Полиморфизм StyI
Праймеры
RF1 - 5'ATG AGC TTC CTA GAG CAA GGA G 3'
RN52R2 - 5'GGC TGC ATG ATG TTA TGT GCC T 3'
Условия PCR
Смесь 1
Объедините смесь 1 и ДНК в реакционной пробирке. Наслоите минеральное масло. Запустите выполнение следующей программы PCR: 94°C в течение 4 мин; 35 циклов, включающих 94°C в течение 45 с, 59°C в течение 45 с и 72°C в течение 45 с; с последующим конечным продлением при 72°C в течение 12 мин.
Проверьте 3 мкл реакционной смеси PCR на 2% агарозном геле для подтверждения успешной амплификации и чистоты отрицательного контроля. Размер продукта составляет 270 н.п.
Расщепление может проводиться по следующей процедуре:
Реакция расщепления StyI
Получите коктейль из продукта PCR, буфера, фермента и воды. Инкубируйте в течение 2 часов при 37°C. Смешайте расщепленный продукт с красителем для загрузки геля (1:6) и проводите электрофорез в 4% агарозном геле.
Генотипы:
Тест полиморфизма PRKAG3-SINE (короткий рассеянный элемент)
Праймеры
RP1F -5' GAA ACT CTT CTC CCC ACA GAC3'
RN52R2-5'GGC TGC ATG ATG TTA TGT GCC T 3'
Условия PCR
Смесь 1
Объедините смесь 1 и ДНК в реакционной пробирке. Наслоите минеральное масло. Запустите выполнение следующей программы PCR:
1 цикл при 95°C в течение 4 мин;
15 циклов, включающих: 95°C в течение 1'20'', 64°C в течение 1' и 74°C в течение 1'40'';
30 циклов, включающих: 95°C в течение 1'20", 58°C в течение 1' и 73°C в течение 1'40'';
с последующим конечным продлением при 73°C в течение 12 мин.
Тест PCR-RFLP для PRKAG3-52
Полиморфизм HphI
Праймеры
RF1 - 5'ATG AGC TTC CTA GAG CAA GGA G 3'
RN52R2 - 5'GGC TGC ATG ATG TTA TGT GCC T 3'
Условия PCR
Смесь 1
Объедините смесь 1 и ДНК в реакционной пробирке. Наслоите минеральное масло. Запустите выполнение следующей программы PCR: 94°C в течение 4 мин; 35 циклов, включающих 94°C в течение 45 с, 59°C в течение 45 с и 72°C в течение 45 с; с последующим конечным продлением при 72°C в течение 12 мин.
Проверьте 3 мкл реакционной смеси PCR на 2% агарозном геле для подтверждения успешной амплификации и чистоты отрицательного контроля. Размер продукта составляет 270 п.н.
Расщепление может проводиться по следующей процедуре:
Реакция расщепления HphI
Получите коктейль из продукта PCR, буфера, фермента и воды. Инкубируйте в течение 2 часов при 37°C. Смешайте расщепленный продукт с красителем для загрузки геля (1:6) и проводите электрофорез в 4% агарозном геле.
Генотипы:
11 - 270 п.н.
12 - 270 п.н., 158 п.н. и 112 п.н.
22 - 158 п.н. и 112 п.н.
Тест PCR-RFLP для PRKAG3-199
Полиморфизм BsaHI
Праймеры
RNF - 5'GGA GCA AAT GTG CAG ACA AG 3'
RNR - 5'CCC ACG AAG CTC TGC TTC TT 3'
Условия PCR
Смесь 1
Объедините смесь 1 и ДНК в реакционной пробирке. Наслоите минеральное масло. Запустите выполнение следующей программы PCR: 94°C в течение 4 мин; 35 циклов, включающих 94°C в течение 45 с, 61°C в течение 45 с и 72°C в течение 1 мин; с последующим конечным продлением при 72°C в течение 12 мин.
Проверьте 3 мкл реакционной смеси PCR на 2% агарозном геле для подтверждения успешной амплификации и чистоты отрицательного контроля. Размер продукта составляет 258 п.н.
Расщепление может проводиться по следующей процедуре:
Реакция расщепления BsaHI
Получите коктейль из продукта PCR, буфера, фермента и воды. Инкубируйте в течение 2 часов при 37°C. Смешайте расщепленный продукт с красителем для загрузки геля (1:6) и проводите электрофорез в 4% агарозном геле.
Генотипы:
11 - 167 п.н. и 91 п.н.
12 - 167 п.н., 119 п.н. и 91 п.н.
22 - 119 п.н. и 91 п.н.


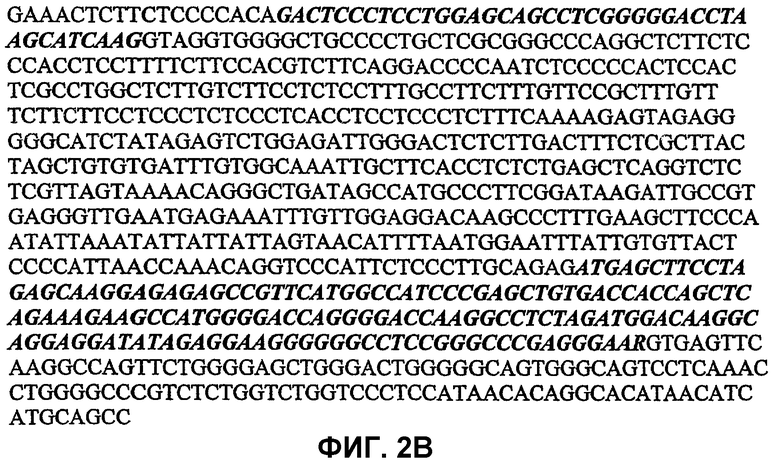


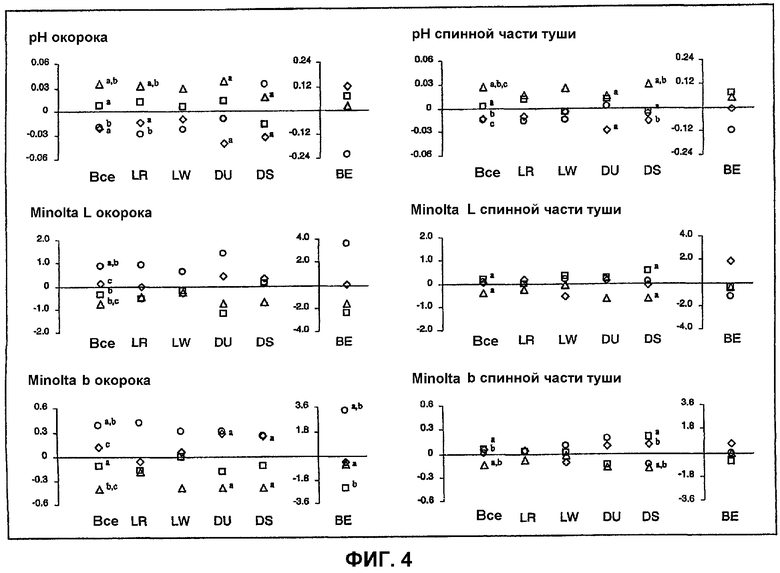
Изобретение относится к области генетической инженерии. Предложено несколько способов скрининга животных с повышенным уровнем численности приплода и/или предпочтительными признаками качества мяса. Способы основаны на детекции генетических маркеров, которые ассоциированы с вышеуказанными положительными признаками. Генетические маркеры основаны на наличии или отсутствии определенных полиморфизмов в гене PRKAG3. Предложены также различные варианты гена PRKAG3 и кодируемого им белка PRKAG3. Изобретение позволяет повысить эффективность проводимой селекции на повышение числа приплода и/или качества мяса. Изобретение может быть использовано в разведении животных. 15 н. и 31 з.п. ф-лы, 6 ил., 8 табл.
| MILAN D | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Science | |||
| ЩИТОВОЙ ДЛЯ ВОДОЕМОВ ЗАТВОР | 1922 |
|
SU2000A1 |
| WO 9735878, 02.10.1997. | |||

