ОБЛАСТЬ ИЗОБРЕТЕНИЯ
В основном, настоящее изобретение относится к профилактике и/или лечению инфекции вирусом папилломы человека (HPV). Более конкретно, настоящее изобретение относится к синтетическим полинуклеотидам, кодирующим белок HPV 52 L1, и к содержащим указанные полинуклеотиды рекомбинантным векторам и организмам-хозяевам. Также это изобретение относится к подобным вирусу HPV 52 частицам (VLP), где VLP получают экспрессией рекомбинантных HPV 52 L1 или L1 + L2 в клетках дрожжей, и к их применению в вакцинах и фармацевтических композициях для профилактики и лечения инфекций HPV.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Существует более чем 80 типов вируса папилломы человека (HPV), многие из которых связывали с широким разнообразием биологических фенотипов, от доброкачественных пролиферирующих бородавок до злокачественных карцином (для обзора см. McMurray et al., Int. J. Exp. Pathol. 82(1): 15-33 (2001)). HPV6 и HPV11 представляют собой типы, наиболее часто связываемые с доброкачественными бородавками, доброкачественной остроконечной кондиломой и/или дисплазией низкой степени слизистой оболочки половой или дыхательной систем. HPV16 и HPV18 представляют собой типы высокого риска, наиболее часто связываемые с карциномами in situ и инвазивными карциномами шейки матки, влагалища, вульвы и анального канала. Более чем 90% карцином шейки матки связаны с инфекциями HPV16, HPV18 или менее распространенными онкогенными типами HPV31, -33, -45, -52 и -58 (Schiffman et al., J. Natl. Cancer Inst. 85(12): 958-64 (1993)). Наблюдение, что ДНК HPV выявляют в 90-100% раков шейки матки, обеспечивает убедительное эпидемиологическое доказательство того, что HPV вызывают карциному шейки матки (см. Bosch et al., J. Clin. Pathol. 55: 244-265 (2002)).
Вирусы папилломы представляют собой небольшие (50-60 нм), без оболочки, икосаэдрические ДНК-вирусы, кодирующие до восьми ранних и двух поздних генов. Открытые рамки считывания (ORF) вирусных геномов обозначают от E1 до E7, а также L1 и L2, где “E” означает ранний, а “L” означает поздний. L1 и L2 кодируют белки вирусного капсида, тогда как гены E связаны с функциями, такими как репликация вируса и клеточная трансформация.
Белок L1 представляет собой основной белок капсида и имеет молекулярную массу 55-60 кДа. Белок L2 представляет собой минорный белок капсида. Иммунологические данные позволяют предположить, что большая часть белка L2 является внутренней в отношении белка L1 в вирусном капсиде. Оба белка L1 и L2 являются высококонсервативными среди различных вирусов папилломы.
Экспрессия белка L1 или сочетания белков L1 и L2 в дрожжах, клетках насекомых, клетках млекопитающих или бактериях приводит к самосборке вирусоподобных частиц (VLP) (для обзора см. Schiller and Roden, в Papillomavirus Reviews: Current Research on Papillomaviruses; Lacey, ed. Leeds, UK: Leeds Medical Information, pp 101-12 (1996)). VLP морфологически сходны с аутентичными вирионами и способны индуцировать высокие титры нейтрализующих антител при введении животным или человеку. Поскольку VLP не содержат потенциально онкогенного вирусного генома, они представляют собой безопасную альтернативу использованию живого вируса в разработке вакцины HPV (для обзора см. Schiller and Hidesheim, J. Clin. Virol. 19: 67-74 (2000)). По этой причине гены L1 и L2 были определены как иммунологические мишени для разработки профилактических и терапевтических вакцин для инфекции и заболевания HPV.
Разработке вакцины HPV и осуществлению промышленного производства препятствовали проблемы, связанные с достижением высоких уровней экспрессии белков капсида в эффективно трансформированных организмах-хозяевах, что ограничивает продукцию очищенного белка. Поэтому, несмотря на выявление нуклеотидных последовательностей дикого типа, кодирующих белки HPV L1, такие как белок HPV 52 L1, очень желательно разработать легко возобновляемый источник неочищенного белка HPV L1, в котором используют кодирующие HPV 52 L1 нуклеотидные последовательности, оптимизированные для экспрессии в определенной клетке-хозяине. Кроме того, для использования в разработке вакцины целесообразно получать большие количества HPV 52 L1 VLP, которые обладают придающими иммунность свойствами нативных белков.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к композициям и способам для придания или усиления иммунности белковых продуктов, экспрессируемых генами HPV 52 L1. Конкретно, настоящее изобретение обеспечивает кодирующие белок HPV 52 L1 полинуклеотиды, где у полинуклеотидов были оптимизированы кодоны для высокого уровня экспрессии в клетке дрожжей. В альтернативных вариантах осуществления изобретения нуклеотидная последовательность полинуклеотида изменена для устранения распознаваемых дрожжами сигналов терминации транскрипции. Кроме того, настоящее изобретение обеспечивает подобные вирусу HPV 52 частицы (VLP), где указанные VLP получают экспрессией рекомбинантных HPV 52 L1 или L1+L2 в клетках дрожжей, а также в настоящем изобретении описано использование HPV 52 VLP в иммуногенных композициях и вакцинах для профилактики и/или лечения заболевания HPV и связанного с HPV рака.
Настоящее изобретение относится к кодирующим белок HPV 52 L1 синтетическим молекулам ДНК. Кодоны синтетических молекул сконструированы таким образом, чтобы использовать кодоны, предпочитаемые клеткой дрожжей. В альтернативном варианте осуществления изобретения нуклеотидная последовательность синтетической молекулы изменена для устранения распознаваемых дрожжами сигналов терминации транскрипции. Синтетические молекулы можно использовать в качестве источника белка HPV 52 L1, который может самособираться в VLP. Указанные VLP можно использовать в вакцине на основе VLP.
Характерный вариант осуществления настоящего изобретения включает синтетическую молекулу нуклеиновой кислоты, кодирующую белок HPV 52 L1, как указано ниже в SEQ ID №:2, где указанная молекула нуклеиновой кислоты содержит последовательность нуклеотидов, в которой кодоны оптимизированы для высокого уровня экспрессии в клетке дрожжей.
Также предоставляют рекомбинантные векторы и рекомбинантные прокариотические и эукариотические клетки-хозяева, которые содержат молекулы нуклеиновой кислоты, указанные на протяжении этого описания. В предпочтительном варианте осуществления настоящего изобретения клетка-хозяин представляет собой клетку дрожжей.
Также настоящее изобретение относится к процессу для экспрессии белка HPV 52 L1 в рекомбинантной клетке-хозяине, предусматривающему: (a) введение в дрожжевую клетку-хозяин вектора, содержащего нуклеиновую кислоту, которая кодирует белок HPV 52 L1; и (b) культивирование дрожжевой клетки-хозяина в условиях, позволяющих экспрессировать указанный белок HPV 52 L1.
Кроме того, настоящее изобретение относится к процессу для экспрессии белка HPV 52 L1 в рекомбинантной клетке-хозяине, предусматривающему: (a) введение в дрожжевую клетку-хозяин вектора, содержащего молекулу нуклеиновой кислоты, которая кодирует белок HPV 52 L1; где в молекуле нуклеиновой кислоты кодоны оптимизированы для оптимальной экспрессии в дрожжевой клетке-хозяине, и (b) культивирование дрожжевой клетки-хозяина в условиях, позволяющих экспрессировать указанный белок HPV 52 L1.
В предпочтительных вариантах осуществления молекула нуклеиновой кислоты содержит последовательность нуклеотидов, как указано ниже в SEQ ID №:1, (обозначенной здесь “последовательность 52 L1 R”).
Также это изобретение относится к подобным вирусу HPV 52 частицам (VLP), которые получают в клетках дрожжей, к способам получения HPV 52 VLP и способам применения HPV 52 VLP.
В предпочтительном варианте осуществления изобретения дрожжи выбирают из группы, состоящей из: Saccharomyces cerevisiae, Hansenula polymorpha, Pichia pastoris, Kluyveromyces fragilis, Kluyveromyces lactis и Schizosaccharomyces pombe.
В другом аспекте это изобретение относится к HPV 52 VLP, где VLP получают рекомбинантной экспрессией HPV 52 L1 или HPV 52 L1+L2 в клетке дрожжей.
В еще одном аспекте это изобретение относится к HPV 52 VLP, содержащей белок HPV 52 L1, который продуцируется геном HPV 52 L1 с оптимизированными кодонами. В характерном варианте осуществления этого аспекта изобретения ген HPV 52 L1 с оптимизированными кодонами содержит последовательность нуклеотидов, как указано ниже в SEQ ID №:1.
Также это изобретение обеспечивает способ индукции иммунной реакции у животного, предусматривающий введение животному частиц, подобных вирусу HPV 52. В предпочтительном варианте осуществления HPV 52 VLP получают посредством гена с оптимизированными кодонами.
В еще одном аспекте это изобретение относится к способу профилактики или лечения связанного с HPV рака шейки матки, предусматривающему введение млекопитающему вакцины, которая содержит HPV 52 VLP. В предпочтительном варианте осуществления этого аспекта изобретения HPV 52 VLP получают в дрожжах.
Также это изобретение относится к вакцине, содержащей подобные вирусу HPV 52 частицы (VLP), где HPV 52 VLP получают в дрожжах.
В альтернативном варианте осуществления этого аспекта изобретения вакцина дополнительно содержит VLP по меньшей мере одного дополнительного типа HPV. По меньшей мере один дополнительный тип HPV может представлять собой любой интересующий тип HPV, в том числе любой описанный в данной области тип HPV или типы, выявленные в последующем. В предпочтительном варианте осуществления тип HPV представляет собой тип, связанный с клиническим фенотипом, таким как бородавки или рак шейки матки. В еще одном предпочтительном варианте осуществления по меньшей мере один дополнительный тип HPV выбирают из группы, состоящей из: HPV6, HPV11, HPV16, HPV18, HPV31, HPV33, HPV35, HPV39, HPV45, HPV51, HPV55, HPV56, HPV58, HPV59 и HPV68.
Также настоящее изобретение относится к фармацевтическим композициям, содержащим подобные вирусу HPV 52 частицы, где HPV 52 VLP получают в дрожжах. Кроме того, это изобретение относится к фармацевтическим композициям, содержащим HPV 52 VLP и VLP по меньшей мере одного дополнительного типа HPV. В предпочтительном варианте осуществления по меньшей мере один дополнительный тип HPV выбирают из группы, состоящей из: HPV6, HPV11, HPV16, HPV18, HPV31, HPV33, HPV35, HPV39, HPV45, HPV51, HPV55, HPV56, HPV58, HPV59 и HPV68.
Как используют на протяжении всего описания и в прилагаемой формуле изобретения, формы единственного числа "некоторый" и "этот" включают в себя ссылку на множественное число до тех пор, пока в контексте явно не указано иначе.
Как используют на протяжении всего описания и в прилагаемой формуле изобретения, применены следующие определения и сокращения.
Термин “промотор” относится к участку распознавания на цепи ДНК, с которым связывается РНК-полимераза. Промотор формирует с РНК-полимеразой инициаторный комплекс для инициации и управления транскрипционной активностью. Комплекс можно изменять активирующими последовательностями, называемыми “энхансерами”, или “вышерасположенными активирующими последовательностями”, или ингибирующими последовательностями, называемыми “сайленсерами”.
Термин “вектор” относится к некоторым способам, которыми фрагменты ДНК можно вводить в организм-хозяин или ткань-хозяин. Существуют различные типы векторов, в том числе плазмиды, вирусы (включая аденовирус), бактериофаги и космиды.
Термин “кассета” относится к последовательности нуклеотидов или гена, подлежащей экспрессии из вектора, например последовательности нуклеотидов или гена, которая кодирует белок HPV 52 L1. Как правило, кассета содержит последовательность гена, вставленную в вектор, который в некоторых вариантах осуществления обеспечивает регуляторные последовательности для экспрессии последовательности нуклеотидов или гена. В других вариантах осуществления регуляторные последовательности для своей экспрессии обеспечивает последовательность нуклеотидов или гена. В еще одних вариантах осуществления некоторые регуляторные последовательности обеспечивает вектор, а другие регуляторные последовательности обеспечивает последовательность нуклеотидов или гена. Например, вектор может обеспечивать промотор для транскрипции последовательности нуклеотидов или гена, а последовательность нуклеотидов или гена обеспечивает последовательность терминации транскрипции. Регуляторные последовательности, которые может обеспечивать вектор, включают, но ими не ограничиваются, энхансеры, последовательности терминации транскрипции, акцепторные и донорные последовательности для сплайсинга, интроны, связывающие рибосомы последовательности и поли(A)-последовательности.
Обозначения “последовательность 52 L1 дикого типа” и “последовательность 52 L1 д.т.” относятся к последовательности HPV 52 L1, указанной здесь как SEQ ID №:3. Хотя последовательность HPV 52 L1 дикого типа была описана ранее, между получаемыми из клинических изолятов ДНК нередко находят незначительные отличия в последовательности. Поэтому характерную последовательность HPV 52 L1 дикого типа выделяли из клинических образцов, для которых ранее было показано содержание ДНК HPV 52 (см. пример 1). Последовательность HPV 52 L1 дикого типа использовали в качестве контрольной последовательности для сравнения описанных здесь последовательностей HPV 52 L1 с оптимизированными кодонами (см. фигуру 1).
Обозначения "HPV 52 L1 R" и "52 L1 R" относятся к описанной здесь характерной синтетической нуклеотидной последовательности HPV52 L1 (SEQ ID №:1), где последовательность реконструировали таким образом, что она содержит кодоны, предпочтительные для высокого уровня экспрессии клеткой дрожжей.
Термин “эффективное количество” означает количество вводимой композиции вакцины, достаточное для получения таких соответствующих уровней полипептида, чтобы возникла иммунная реакция. Специалист в данной области понимает, что этот уровень может различаться.
“Консервативная замена аминокислоты” относится к замене одного остатка аминокислоты другим, химически сходным аминокислотным остатком. Примерами таких консервативных замен являются: замена одного гидрофобного остатка (изолейцина, лейцина, валина или метионина) другим; замена одного полярного остатка другим полярным остатком с таким же зарядом (например, аргинина лизином; глутаминовой кислоты аспарагиновой кислотой).
Термин “млекопитающее” относится к любому млекопитающему, в том числе человеку.
"VLP" или "VLPs" означает(ют) вирусоподобную частицу или вирусоподобные частицы.
“Синтетический” означает то, что ген HPV 52 L1 сконструировали таким образом, что он содержит последовательность нуклеотидов, отличающуюся от нуклеотидной последовательности, которая представлена в указанном встречающемся в природе гене HPV 52 L1 дикого типа (52 L1 дикого типа, SEQ ID №:3). Как указано выше, предоставленные здесь синтетические молекулы содержат нуклеотидную последовательность, включающую в себя кодоны, предпочтительные для экспрессии клетками дрожжей. Предоставленные здесь синтетические молекулы кодируют те же последовательности аминокислот, что и ген HPV 52 L1 дикого типа (SEQ ID №:2).
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фигуре 1 представлено выравнивание последовательностей для сравнения нуклеотидов, измененных в синтетическом гене HPV 52 L1 по настоящему изобретению (SEQ ID №:1, обозначенная как "52 L1 R") (см. пример 2). Контрольная последовательность представляет собой последовательность 52 L1 дикого типа (SEQ ID №:3, обозначенная как "52 L1 дикого типа"; см. пример 1). Измененные нуклеотиды указаны в их соответствующем положении. Номер нуклеотида заключен в круглые скобки. Те же самые нуклеотиды в реконструированной последовательности 52 L1 указаны точками.
На фигуре 2 представлена реконструированная синтетическая двухцепочечная нуклеиновая кислота HPV 52 L1 (SEQ ID №№:1 и 7) и однобуквенная последовательность аминокислот (SEQ ID №:2). Номер нуклеотида указан слева.
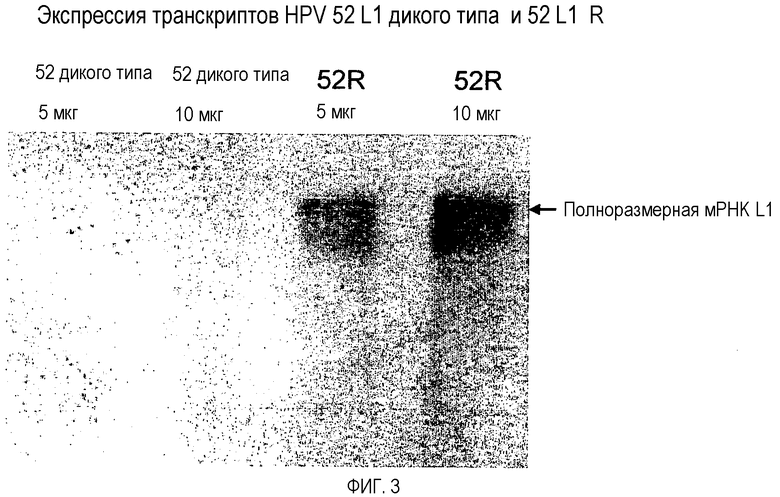
На фигуре 3 представлен нозерн-блот транскриптов HPV 52 L1 дикого типа и HPV 52 L1 R (см. пример 4). Блот анализируют с использованием смеси ДНК-зондов, полученных к обеим последовательностям 52 L1 дикого типа и 52 L1 R. Стрелка справа указывает ожидаемое положение полноразмерного транскрипта 52 L1. В случае РНК 52 L1 дикого типа в дорожках 5 и 10 мкг не было выявлено транскриптов какой-либо длины. В случае 52 L1 R полноразмерные транскрипты наблюдают в дорожках как 5, так и 10 мкг.
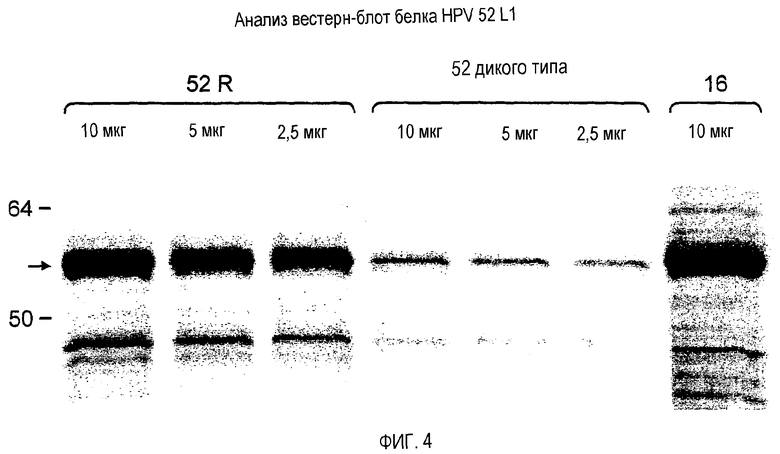
На фигуре 4 представлен вестерн-блот белков HPV 52 L1 дикого типа (52 дикого типа) и 52 L1 R (52R). HPV 16 L1 был включен в качестве контроля (16). Десять, пять и два, а также половину микрограмма общего белкового экстракта дрожжей денатурировали и наносили на 10% гель SDS-PAGE. Белок переносили по способу вестерн. Белок HPV 52 L1 на полученном блоте выявляли с использованием абсорбируемой дрожжами козьей поликлональной антисыворотки к trpE-HPV 31 L1, перекрестно реагирующей с HPV 52 L1 и HPV 16 L1. Слева указаны маркеры молекулярной массы в кДа. Стрелка указывает положение белка HPV 52 L1 ~55 кДа.
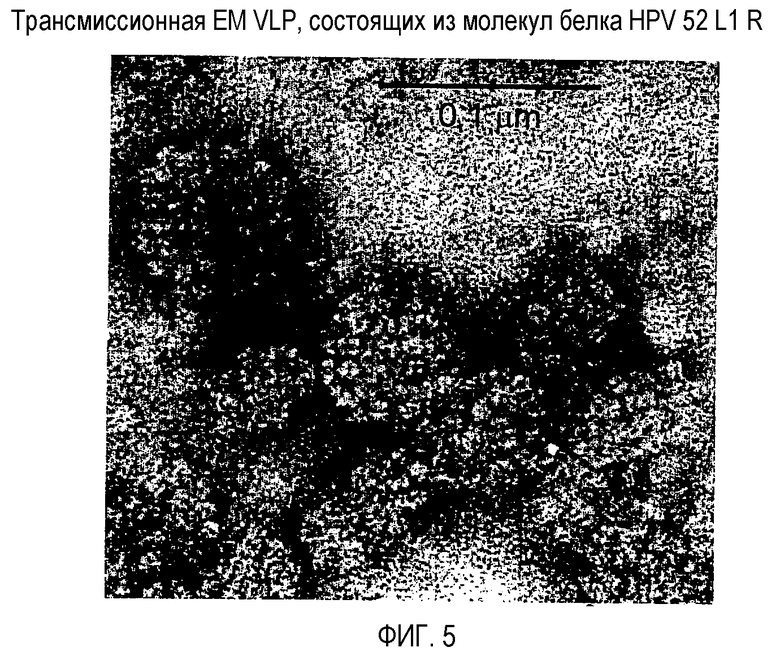
На фигуре 5 представлен характерный образец HPV 52 VLP, составленных из описанных здесь молекул белка HPV 52 L1 R, где изображение получено трансмиссионной электронной микроскопией (см. пример 7). Диаметр сферических частиц в этом неочищенном образце находится в диапазоне от 40 до 70 нм, причем у некоторых частиц наблюдают регулярное расположение капсомеров. Линейка представляет собой приблизительно 0,1 мкм.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Большинство карцином шейки матки связаны с инфекциями специфическими онкогенными типами вируса папилломы человека (HPV). Настоящее изобретение относится к композициям и способам для придания или усиления иммунности белковых продуктов, экспрессируемых генами онкогенных типов HPV. Конкретно, настоящее изобретение обеспечивает кодирующие HPV 52 L1 полинуклеотиды, где у полинуклеотидов кодоны оптимизированы для высокого уровня экспрессии в дрожжах. Также настоящее изобретение обеспечивает подобные вирусу HPV52 частицы (VLP), получаемые в дрожжах, а также в настоящем изобретении описано использование указанных полинуклеотидов и VLP в иммуногенных композициях и вакцинах для профилактики и/или лечения связанного с HPV рака.
Была опубликована нуклеотидная последовательность HPV52 L1 дикого типа (уникальный идентификатор банка генов №NC 001592). Настоящее изобретение обеспечивает синтетические молекулы ДНК, кодирующие белок HPV 52 L1. В одном из аспектов изобретения синтетические молекулы содержат последовательность кодонов, где по меньшей мере несколько кодонов были изменены с целью использования кодонов, которые предпочтительны для клетки дрожжей для высокого уровня экспрессии. В альтернативном аспекте изобретения нуклеотидная последовательность синтетической молекулы изменена для устранения распознаваемых дрожжами сигналов терминации транскрипции. Синтетические молекулы можно использовать в качестве кодирующей последовательности для экспрессии белка HPV 52 L1, который может самособираться в VLP. Указанные VLP можно использовать в вакцине на основе VLP для обеспечения эффективной иммунопрофилактики папилломавирусной инфекции посредством нейтрализующего антитела и опосредуемого клетками иммунитета. Также такие вакцины на основе VLP могут быть эффективны для лечения уже выявленных инфекций HPV.
Экспрессия HPV VLP в клетках дрожжей обеспечивает преимущества экономической эффективности и легкого приспособления к росту в значительных масштабах в ферментерах. Кроме того, геном дрожжей можно легко изменять для обеспечения отбора рекомбинантных трансформированных дрожжей с повышенной способностью роста и экспрессии. Однако многие белки HPV L1, в том числе HPV 52 L1, экспрессируются в клетках дрожжей на уровнях более низких, чем желаемые для коммерческого промышленного масштаба уровни (см. пример 2).
Соответственно настоящее изобретение относится к последовательностям гена HPV 52 L1, которые “оптимизированы” для высокого уровня экспрессии в среде клеток дрожжей.
“Триплетный” кодон из четырех возможных нуклеотидных оснований может существовать в более чем 60 различных формах. Поскольку эти кодоны обеспечивают информацию только для 20 различных аминокислот (а также инициации и терминации транскрипции), то несколько аминокислот могут кодироваться более чем одним кодоном, явление, известное как вырожденность генетического кода. По не совсем понятным причинам альтернативные кодоны неравномерно представлены в эндогенной ДНК различных типов клеток. Действительно, по-видимому, существует различная естественная иерархия или “предпочтительность” определенных кодонов в определенных типах клеток. В качестве одного из примеров аминокислота лейцин определяется любым из шести кодонов ДНК, включающих в себя CTA, CTC, CTG, CTT, TTA и TTG. В исчерпывающем анализе частот использования геномных кодонов у микроорганизмов было выявлено, что эндогенная ДНК E. coli наиболее часто содержит определяющий лейцин кодон CTG, тогда как ДНК дрожжей и слизистой плесени наиболее часто содержит определяющий лейцин кодон TTA. Учитывая эту иерархию, обычно предполагают, что вероятность достижения высоких уровней экспрессии богатого лейцином полипептида в E. coli-хозяине в некоторой степени зависит от частоты использования кодона. Например, вероятно, что богатый кодонами TTA ген плохо экспрессируется в E. coli, тогда как богатый CTG ген, возможно, экспрессируется в этом организме-хозяине на высоком уровне. Сходным образом, предпочтительный кодон для экспрессии богатого лейцином полипептида в дрожжевых клетках-хозяевах представляет собой TTA.
Последствия явления предпочтительности кодонов для способов рекомбинантной ДНК очевидны, а явление может служить для объяснения многих предыдущих неудач в достижении высоких уровней экспрессии экзогенных генов в эффективно трансформированных организмах-хозяевах - во вставляемом гене может быть многократно представлен менее “предпочтительный” кодон, и системы клетки-хозяина для экспрессии могут действовать не так эффективно. Это явление позволяет предположить, что синтетические гены, которые были сконструированы для включения в них предполагаемых предпочтительных для клетки-хозяина кодонов, обеспечивают оптимальную форму чужеродного генетического материала для практического осуществления экспрессии рекомбинантного белка. Таким образом, в одном из аспектов настоящее изобретение относится к гену HPV 52 L1 с оптимизированными кодонами для высокого уровня экспрессии в клетке дрожжей. В предпочтительном варианте осуществления настоящего изобретения было открыто, что использование альтернативных кодонов, кодирующих ту же самую последовательность белка, может устранять ограничения экспрессии белков HPV 52 L1 клетками дрожжей.
По этому изобретению участки гена HPV 52 L1 преобразовывали в последовательности, имеющие аналогичные транслированные последовательности, но с использованием альтернативного кодона, как описано Sharp and Cowe в (Synonymous Codon Usage in Saccharomyces cerevisiae. Yeast 7: 657-678 (1991)), приведенном здесь полностью в качестве ссылки. Как правило, способ состоит в выявлении в последовательности дикого типа кодонов, которые обычно не связаны с высокоэкспрессируемыми генами дрожжей, и в замене их оптимальными кодонами для высокой экспрессии в клетках дрожжей. Затем новую последовательность гена проверяют на предмет нежелательных последовательностей, получаемых этими заменами кодонов (например, последовательностей "ATTTA", случайного создания участков распознавания для сплайсинга интронов, нежелательных участков для ферментов рестрикции, высокого содержания GC, наличия распознаваемых дрожжами сигналов терминации транскрипции и т.д.). Нежелательные последовательности устраняют заменой существующих кодонов отличными кодонами, кодирующими ту же аминокислоту. Затем участки синтетического гена проверяют на предмет улучшенной экспрессии.
Указанные выше способы использовали для конструирования участков синтетического гена для HPV 52 L1, получая в результате ген, который содержит оптимизированные для высокого уровня экспрессии кодоны. Хотя указанная выше процедура обеспечивает краткое описание способа авторов изобретения для конструирования генов с оптимизированными кодонами для использования в вакцинах HPV, специалист в данной области понимает, что сходную эффективность вакцины или повышенную экспрессию генов можно достигать незначительными изменениями в процедуре или незначительными изменениями в последовательности.
Соответственно настоящее изобретение относится к синтетическому полинуклеотиду, содержащему последовательность нуклеотидов, которая кодирует белок HPV 52 L1, или биологически активный фрагмент, или мутантную форму белка HPV 52 L1, где полинуклеотидная последовательность содержит кодоны, оптимизированные для экспрессии в дрожжевой клетке-хозяине. Указанные мутантные формы белка HPV 52 L1 включают, но ими не ограничиваются: консервативные аминокислотные замены, усечения с аминоконцевой стороны, усечения с карбоксиконцевой стороны, делеции или добавления. Любой такой биологически активный фрагмент и/или мутант кодирует белок или фрагмент белка, который по меньшей мере в значительной степени воспроизводит иммунологические свойства белка HPV 52 L1, как указано ниже в SEQ ID №:2. Синтетические полинуклеотиды по настоящему изобретению кодируют молекулы мРНК, экспрессирующие функциональный белок HPV 52 L1 таким образом, чтобы быть эффективными в разработке терапевтической или профилактической вакцины HPV.
В одном из аспектов настоящее изобретение относится к молекуле нуклеиновой кислоты с оптимизированными кодонами, кодирующей белок HPV 52 L1, как указано ниже в SEQ ID №:2, где указанная молекула нуклеиновой кислоты содержит последовательность нуклеотидов с оптимизированными кодонами для высокого уровня экспрессии в клетке дрожжей. В предпочтительном варианте осуществления этого аспекта изобретения молекула нуклеиновой кислоты содержит последовательность нуклеотидов, как указано ниже в SEQ ID №:1.
Также настоящее изобретение относится к рекомбинантным векторам и рекомбинантным прокариотическим и эукариотическим клеткам-хозяевам, содержащим молекулы нуклеиновой кислоты, которые описаны на всем протяжении этого описания. В предпочтительном варианте осуществления этого изобретения клетка-хозяин представляет собой дрожжевую клетку-хозяин.
Синтетическую ДНК HPV 52 L1, ее функциональные эквиваленты и ее фрагменты, сконструированные описанными здесь способами, можно рекомбинантно экспрессировать посредством молекулярного клонирования в вектор экспрессии, который содержит приемлемый промотор и другие соответствующие элементы регуляции транскрипции. Указанный вектор экспрессии можно переносить в прокариотические или эукариотические клетки-хозяева для получения рекомбинантного белка HPV 52 L1. Способы для таких процедур подробно описаны в данной области (Sambrook et al. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, (1989); Current Protocols in Molecular Biology, Ausubel et al., Green Pub. Associates and Wiley-Interscience, New York (1988); Yeast Genetics: A Laboratory Course Manual, Rose et al., Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, (1990), которые приведены здесь полностью в качестве ссылки).
Таким образом, настоящее изобретение относится к процессу для экспрессии белка HPV 52 L1 в рекомбинантной клетке-хозяине, предусматривающему: (a) введение в дрожжевую клетку-хозяина вектора, содержащего нуклеиновую кислоту, которая кодирует белок HPV 52 L1; и (b) культивирование дрожжевой клетки-хозяина в условиях, позволяющих экспрессировать указанный белок HPV 52 L1.
Кроме того, настоящее изобретение относится к процессу для экспрессии белка HPV 52 L1 в рекомбинантной клетке-хозяине, предусматривающему: (a) введение в дрожжевую клетку-хозяин вектора, содержащего нуклеиновую кислоту, которая кодирует белок HPV 52 L1; где в молекуле нуклеиновой кислоты кодоны оптимизированы для оптимальной экспрессии в дрожжевой клетке-хозяине, и (b) культивирование дрожжевой клетки-хозяина в условиях, позволяющих экспрессировать указанный белок HPV 52 L1.
Кроме того, это изобретение относится к процессу для экспрессии белка HPV 52 L1 в рекомбинантной клетке-хозяине, предусматривающему: (a) введение в дрожжевую клетку-хозяин вектора, содержащего нуклеиновую кислоту, как указано ниже в SEQ ID №:1; и (b) культивирование дрожжевой клетки-хозяина в условиях, позволяющих экспрессировать указанный белок HPV 52 L1.
Синтетические гены по настоящему изобретению можно объединять в кассету экспрессии, содержащую последовательности, которые сконструированы для обеспечения эффективной экспрессии белка HPV 52 L1 в клетке-хозяине. Предпочтительно кассета содержит синтетический ген с функционально связанными с ним последовательностями, которые контролируют транскрипцию и трансляцию, такими как промотор и последовательности терминации. В предпочтительном варианте осуществления промотор представляет собой промотор GAL1 S. cerevisiae, хотя специалисты в данной области понимают, что можно использовать любые из множества других известных промоторов дрожжей, таких как промоторы GAL10, GAL7, ADH1, TDH3 или PGK, или промоторы других эукариотических генов. Предпочтительный терминатор транскрипции представляет собой терминатор ADH1 S. cerevisiae, хотя также можно использовать другие известные терминаторы транскрипции. Особенно предпочтительно сочетание промотор GAL1-терминатор ADH1.
В другом аспекте это изобретение относится к подобной вирусу HPV 52 частице (VLP), получаемой рекомбинантной экспрессией генов HPV 52 L1 или L1+L2 в клетке дрожжей, к способам получения HPV 52 VLP и способам применения HPV 52 VLP. VLP могут самособираться, если L1, основной белок капсида вирусов папилломы человека и животных, экспрессируется в дрожжах, клетках насекомых, клетках млекопитающих или бактериях (для обзора см. Schiller и Roden, в Papillomavirus Reviews: Current Research on Papillomaviruses; Lacey, ed. Leeds, UK: Leeds Medical Information, pp. 101-12 (1996)). Морфологически слабо отличимые HPV VLP также можно получать экспрессией сочетания белков капсида L1 и L2. VLP состоят из 72 пентамеров L1 в икосаэдрической структуре T=7 (Baker et al., Biophys. J. 60 (6): 1445-56(1991)).
VLP морфологически сходны с аутентичными вирионами и способны при введении животному индуцировать высокие титры нейтрализующих антител. Было показано, что иммунизация кроликов (Breitburd et al., J. Virol. 69 (6): 3959-63 (1995)) и собак (Suzich et al., Proc. Natl. Acad. Sci. USA 92 (25): 11553-57 (1995)) посредством VLP индуцирует нейтрализующие антитела и защищает от экспериментальной папилломавирусной инфекции. Кроме того, было показано, что иммунизация взрослых женщин посредством HPV 16 VLP защищает от инфекции HPV 16 и интраэпителиальной неоплазии шейки матки HPV 16 (Koutsky et al. N. Engl. J. Med. 347: 1645-51 (2002)). Поскольку VLP не содержат потенциально онкогенного вирусного генома и могут самособираться при экспрессии с одного гена, то они представляют собой безопасную альтернативу использованию живого вируса в разработке вакцины HPV (для обзора см. Schiller and Hidesheim, J. Clin. Virol. 19: 67-74 (2000)).
Таким образом, настоящее изобретение относится к вирусоподобным частицам, содержащим рекомбинантный белок L1 или рекомбинантные белки L1 + L2 HPV 52, где рекомбинантный белок экспрессируется в клетке дрожжей.
Как указано выше, в предпочтительном варианте осуществления изобретения HPV 52 VLP получают в дрожжах. В другом предпочтительном варианте осуществления дрожжи выбирают из группы, состоящей из: Saccharomyces cerevisiae, Hansenula polymorpha, Pichia pastoris, Kluyveromyces fragilis, Kluyveromyces lactis и Schizosaccharomyces pombe.
В другом аспекте это изобретение относится к HPV 52 VLP, которая содержит белок HPV 52 L1, продуцируемый геном HPV 52 L1 с оптимизированными кодонами. В предпочтительном варианте осуществления этого аспекта изобретения ген HPV 52 L1 с оптимизированными кодонами содержит последовательность нуклеотидов, как указано ниже в SEQ ID №:1.
В еще одном аспекте это изобретение относится к способу получения HPV 52 VLP, предусматривающему: (a) трансформацию дрожжей молекулой рекомбинантной ДНК, кодирующей белок HPV 52 L1 или белки HPV 52 L1+L2; (b) культивирование трансформированных дрожжей в условиях, позволяющих экспрессировать молекулу рекомбинантной ДНК для получения рекомбинантного белка HPV 52; и (c) выделение рекомбинантного белка HPV 52 для получения HPV 52 VLP.
В предпочтительном варианте осуществления этого аспекта изобретения дрожжи трансформируют геном HPV 52 L1 с оптимизированными кодонами для получения HPV 52 VLP. В особенно предпочтительном варианте осуществления ген HPV 52 L1 с оптимизированными кодонами содержит последовательность нуклеотидов, как указано ниже в SEQ ID №:1.
Также это изобретение обеспечивает способ для индукции иммунной реакции у животного, предусматривающий введение животному подобных вирусу HPV 52 частиц. В предпочтительном варианте осуществления HPV 52 VLP получают рекомбинантной экспрессией гена с оптимизированными кодонами, кодирующего HPV 52 L1 или HPV 52 L1+L2.
В еще одном аспекте это изобретение относится к способу профилактики и/или лечения связанного с HPV рака шейки матки, предусматривающему введение млекопитающему вакцины, которая содержит HPV 52 VLP. В предпочтительном варианте осуществления этого аспекта изобретения HPV 52 VLP получают в дрожжах.
Также это изобретение относится к вакцине, содержащей подобные вирусу HPV 52 частицы (VLP).
В альтернативном варианте осуществления этого аспекта изобретения вакцина дополнительно содержит VLP по меньшей мере одного дополнительного типа HPV. В предпочтительном варианте осуществления по меньшей мере один дополнительный тип HPV выбирают из группы, состоящей из: HPV6, HPV11, HPV16, HPV18, HPV31, HPV33, HPV35, HPV39, HPV45, HPV51, HPV55, HPV56, HPV58, HPV59 и HPV68.
В предпочтительном варианте осуществления этого аспекта изобретения вакцина дополнительно содержит HPV 16 VLP.
В другом предпочтительном варианте осуществления изобретения вакцина дополнительно содержит HPV 16 VLP и HPV 18 VLP.
В еще одном предпочтительном варианте осуществления изобретения вакцина дополнительно содержит HPV 6 VLP, HPV 11 VLP, HPV 16 VLP и HPV 18 VLP.
Также это изобретение относится к фармацевтическим композициям, содержащим подобные вирусу HPV 52 частицы. Кроме того, это изобретение относится к фармацевтическим композициям, содержащим HPV 52 VLP и VLP по меньшей мере одного дополнительного типа HPV. В предпочтительном варианте осуществления по меньшей мере один дополнительный тип HPV выбирают из группы, состоящей из: HPV6, HPV11, HPV16, HPV18, HPV31, HPV33, HPV35, HPV39, HPV45, HPV51, HPV55, HPV56, HPV58, HPV59 и HPV68.
Композиции вакцины по настоящему изобретению можно использовать по отдельности в соответствующих дозах, делающих возможным оптимальное ингибирование инфекции HPV 52 при минимальной возможной токсичности. Кроме того, может быть желательно совместное введение или последовательное введение других средств.
Количество вирусоподобных частиц, подлежащее введению в реципиента вакцины, зависит от иммуногенности экспрессируемого продукта гена. Как правило, иммунологически или профилактически эффективную дозу, составляющую приблизительно от 10 мкг до 100 мкг, а предпочтительно приблизительно от 20 мкг до 60 мкг VLP, вводят непосредственно в мышечную ткань. Также предусмотрены подкожная инъекция, внутрикожное введение, внедрение через кожу и другие способы введения, такие как внутрибрюшинный, внутривенный или доставка ингаляцией. Также предусмотрено, что могут быть обеспечены повторные вакцинации. Также эффективны парентеральное введение, такое как внутривенное, внутримышечное, подкожное или другие способы введения с адъювантами, такими как алюминиевые квасцы или адъювант из алюминиевых квасцов Merck, одновременно с или после парентерального введения вакцины по этому изобретению.
Все указанные здесь публикации приведены здесь полностью в качестве ссылки в целях изложения и описания способов и веществ, которые можно использовать в связи с настоящим изобретением. Ничто из указанного здесь не должно рассматриваться как признание, что изобретение не предоставляет право на датирование такого описания задним числом в силу предшествующего изобретения.
При наличии описанных предпочтительных вариантов осуществления изобретения со ссылкой на сопровождающие чертежи следует понимать, что изобретение не ограничивается этими точными вариантами осуществления и что специалист в данной области может осуществлять различные изменения и модификации без утраты объема или сущности изобретения, как определено в прилагаемой формуле изобретения.
Следующие примеры иллюстрируют, но не ограничивают изобретение.
ПРИМЕР 1
Определение характерной последовательности HPV 52 L1
Ранее была описана последовательность HPV 52 L1 (уникальный идентификатор банка генов №NC 001592). Однако между получаемыми из клинических изолятов ДНК нередко находят незначительные отличия в последовательности. Для определения характерной последовательности HPV 52 L1 дикого типа ДНК выделяли из трех клинических образцов, для которых ранее было показано содержание ДНК HPV 52. Последовательности HPV 52 L1 амплифицировали полимеразной цепной реакцией (PCR) с использованием ДНК-полимеразы Taq и следующих праймеров: 5'L1 5' - A T G T C C G T G T G G C G G C C T A G T - 3' (SEQ ID №:4) и 3' 52 Bgl II 5' - G A G A T C T C A A T T A C A C A A A G T G - 3' (SEQ ID №:5). Амплифицированные продукты подвергали электрофорезу в агарозных гелях и получали изображение посредством окрашивания бромидом этидия. Вырезали полосы L1 ~1500 п.о. и очищали ДНК с использованием набора Geneclean Spin Kit (Q-Bio Gene, Carlsbad, CA). Затем ДНК лигировали в вектор для клонирования TA, pCR2.1 (Invitrogen). Трансформировали клетки E. coli TOP10F', получаемой при лигировании смесью, и высевали на агар LB с канамицином и IPTG, а также X-gal для отбора синих/белых колоний. Планшеты переворачивали и инкубировали в течение 16 ч при 37°C.
PCR для колоний проводили для пяти белых колоний, происходящих из каждого из трех амплифицированных клинических изолятов. Праймеры 5' L1 и 3' 52 Bgl II использовали в двухстадийной PCR, в которой первая стадия содержала 10 циклов из 96°C в течение 15 с (денатурация), 55°C в течение 30 с (отжиг) и 68°C в течение 2 мин (удлинение), а вторая стадия содержала 35 циклов преимущественно сходной программы, за исключением того, что стадию отжига проводили при 50°C в течение 30 с. Продукты PCR подвергали электрофорезу в агарозных гелях и получали изображение посредством окрашивания бромидом этидия. Несколько колоний из каждого клинического изолята содержали амплифицированный продукт с полосами ~1500 п.о. Колонии культивировали в среде LB с канамицином, встряхивая при 37°C в течение 16 ч. Использовали способ выделения в малом объеме для выделения плазмидных ДНК, которые расщепляли эндонуклеазами рестрикции для выявления наличия в плазмиде гена L1. Получаемые фрагменты рестрикции наблюдали посредством электрофореза в агарозном геле и окрашивания бромидом этидия.
Для плазмид, содержащих вставки клонированного L1 из каждого из трех клинических изолятов, проводили секвенирование ДНК. ДНК и транслированные аминокислотные последовательности сравнивали друг с другом и с ранее опубликованными в банке генов последовательностями HPV 52 L1. В результате анализа последовательностей трех клинических изолятов было выявлено отсутствие последовательности, одинаковой с последовательностью из банка генов (уникальный идентификатор №NC 001592). В качестве характерной последовательности HPV 52 L1 был выбран клон pCR2.1 HPV 52L1 №2C и обозначен здесь как "последовательность 52 L1 дикого типа" (SEQ ID №:3, см. фигуру 1). В сравнении с последовательностью из банка генов выбранная в качестве 52 L1 дикого типа (д.т.) последовательность содержала одну точковую мутацию, состоящую в молчащей мутации нуклеотида 1308 (аденин → гуанин). Аминокислотная последовательность последовательности HPV 52 L1 дикого типа была аналогична последовательности 52 L1 из банка генов.
Для добавления удлиняющих участков Bgl II последовательность HPV 52 L1 дикого типа амплифицировали с использованием праймера 5' L1 Bgl II (5' - G A G A T C T C A C A A A A C A A A A T G T C C G T G T G G C - 3' (SEQ ID №:6)) и указанного выше праймера 3' 52 Bgl II. PCR выполняли с использованием полимеразы Taq. Продукт PCR подвергали электрофорезу в агарозном геле и получали изображение посредством окрашивания бромидом этидия. Вырезали полосу L1 ~1500 п.о. и очищали ДНК с использованием набора Geneclean Spin Kit (Q-Bio Gene, Carlsbad, CA). Затем продукт PCR лигировали в вектор pCR2.1 и получаемой при лигировании смесью трансформировали клетки TOP10F'. Белые колонии культивировали в среде LB с канамицином, встряхивая при 37°C в течение 16 ч. Для выделения плазмидной ДНК использовали способ выделения в малом объеме. Ген HPV 52 L1 выделяли из последовательностей вектора посредством расщеплений эндонуклеазой рестрикции Bgl II. Расщепленную ДНК подвергали электрофорезу в агарозном геле и наблюдали посредством окрашивания бромидом этидия. Полосу L1 очищали с использованием набора Geneclean и лигировали в дефосфорилированный, расщепленный BamHI вектор pGAL110. Получаемой при лигировании смесью трансформировали клетки E. coli TOP10F'. Для отбора вставки HPV 52 L1 в правильном направлении плазмидную ДНК из колоний подвергали амплификации PCR. Для подтверждения последовательности и направления вставок проводили секвенирование ДНК. Выбранный клон обозначили pGAL110-HPV 52L1 №5. Из выбранного клона способом выделения в большом объеме получали ДНК. Клетки Saccharomyces cerevisiae делали компетентными посредством образования сферопластов с использованием глузулазы и трансформировали посредством pGAL110-HPV 52L1 №5. Получаемую при трансформации дрожжей смесь высевали на покрытие из агара лейцин-сорбит на планшетах лейцин-сорбит и инкубировали в перевернутом положении в течение 3-5 сут при 30°C. Колонии отбирали и делали посевы штрихом для выделения на планшетах лейцин-сорбит. Затем выделенные колонии выращивали в 5 мл 5 X сорбита-лейцин-аденин с 1,6% глюкозой и 4% галактозой в культурах во вращающихся пробирках при 30°C для индукции транскрипции HPV 52 L1 и экспрессии белка.
ПРИМЕР 2
Оптимизация дрожжевого кодона
Были описаны предпочтительные для дрожжей кодоны (Sharp, Paul M and Cowe, Elizabeth. Synonymous Codon Usage in Saccharomyces cerevisiae YEAST 7:657-678 (1991)). Экспрессия белка HPV 52 L1 дикого типа поддавалась выявлению; однако уровень транскрипции был очень низким и нерегистрируемым посредством нозерн-блота. Было предположено, что за низкие уровни экспрессии гена HPV 52 L1 может быть ответственна преждевременная терминация транскрипции. Для повышения транскрипции этого гена и обеспечения получения полноразмерных транскриптов ген HPV 52 L1 реконструировали с использованием предпочтительных для дрожжей кодонов. Последовательность проверяли на наличие распознаваемых дрожжами сигналов терминации транскрипции для дрожжей, и эти последовательности устраняли заменой альтернативными кодонами при сохранении той же самой аминокислотной последовательности. Реконструированная последовательность HPV 52 L1, содержащая последовательности оптимизированных для дрожжей кодонов, включала в себя 379 изменений нуклеотидов в сравнении с последовательностью HPV 52 L1 дикого типа. Получаемая в результате последовательность обозначена здесь как "52 L1 R" (R=реконструированная, см. фигуру 1). Нуклеотидные изменения между последовательностями 52 L1 дикого типа (SEQ ID №:3) и 52 L1 R (SEQ ID №:1) представлены на фигуре 1. Транслированная аминокислотная последовательность 58 L1 R не была изменена (SEQ ID №:2, см. фигуру 2). Реконструированная последовательность обеспечивает повышенную экспрессию белка HPV 52 L1, что является значительным улучшением для использования в разработке вакцины в сравнении с диким типом.
Используемый для получения оптимизированного гена способ состоял в конструировании длинных перекрывающихся смысловых и антисмысловых олигомеров, которые перекрывают ген, заменяя нуклеотиды последовательностями предпочтительных для дрожжей кодонов при сохранении аминокислотной последовательности. Эти олигомеры использовали вместо образца ДНК в реакции PCR с ДНК-полимеразой Pfu. Для амплификации реконструированных последовательностей с образцов олигомеров конструировали и использовали дополнительные праймеры для амплификации.
Оптимальные условия для амплификации были специфичны для участков; однако для большинства реакций использовали программу, сходную с 94°C в течение 5 мин (денатурация) с последующими 25 циклами из 95°C в течение 30 с (денатурация), 50-55°C в течение 30 с (отжиг), 72°C в течение 1,5 мин (удлинение), с последующим конечным удлинением при 72°C в течение 7 мин и хранением при 4°C. Продукты PCR анализировали электрофорезом в агарозном геле. Вырезали полосы соответствующего размера и из пластины геля очищали ДНК. Затем амплифицированные фрагменты использовали в качестве образцов для сборки реконструированного гена HPV 52 L1 из 1512 нуклеотидов.
После реконструкции полосу из 1512 нуклеотидов очищали из геля и лигировали в вектор pCR4 Blunt (Invitrogen, Carlsbad, CA). После лигирования трансформировали получаемой при лигировании смесью компетентные клетки E. coli TOP10. Колонии выращивали в 4 мл LB с ампициллином, а плазмидную ДНК выделяли из колоний способами выделения в малом объеме. Для подтверждения наличия желаемых изменений в реконструированном HPV 52 L1 секвенировали плазмидную ДНК. Для добавления удлиняющих участков BamHI к обоим концам повторно амплифицировали 52 L1 R (реконструированный) из pCR4Blunt-52 L1 R. Амплифицированный фрагмент клонировали, как указано выше, и секвенировали получаемую плазмидную ДНК. Плазмиду pCR4 Blunt-52 L1 R (Bam) расщепляли BamHI и подвергали получаемые вставки фрагмента ДНК электрофорезу в агарозном геле. Фрагмент HPV 52 L1 R (Bam) ~1530 п.о. очищали из геля и лигировали в расщепленный BamHI pGAL110. Получаемой при лигировании смесью трансформировали клетки E. coli TOP10F' (Invitrogen).
Полученные колонии отбирали посредством PCR на предмет вставки HPV 52 L1 R в правильном направлении. Последовательность и направление подтверждали секвенированием ДНК. Плазмидную ДНК получали способом выделения в большом объеме. Клетки S. cerevisiae делали компетентными посредством образования сферопластов и трансформировали. Получаемые при трансформации дрожжи высевали на покрытие из агара лейцин-сорбит на планшетах с агаром лейцин-сорбит и инкубировали в перевернутом положении в течение 7 суток. Колонии отбирали и делали посевы штрихом для выделения клонов на планшетах с агаром лейцин-сорбит. Затем выделенные колонии выращивали в 5 мл 5 X сорбита лейцин-аденин с 1,6% глюкозой и 4% галактозой в культурах во вращающихся пробирках при 30°C для индукции транскрипции L1 и экспрессии белка. Через 48 и/или 72 часа объем культуры, эквивалентный OD600=10, осаждали центрифугированием, удаляли надосадочную жидкость, а осадки замораживали и хранили при -70°C.
ПРИМЕР 3
Получение РНК
Осадки клеток трансформированных дрожжей, которые индуцируют для экспрессии HPV 52 L1 посредством индукции галактозой, размораживали на льду, суспендировали в 0,8 мл реактива тризол (Life Technologies, Gibco BRL) и инкубировали при комнатной температуре в течение 5 мин. В сосуд добавляли одну пятую объема хлороформа. Затем осторожно встряхивали в течение 15 с для перемешивания и инкубировали при комнатной температуре в течение 3 мин. После центрифугирования в течение 5 мин при 13000 об/мин отбирали верхнюю фазу и переносили в новый сосуд. В сосуд добавляли 0,4 мл изопропанола. Смесь инкубировали при комнатной температуре в течение 10 мин. Для осаждения РНК проводили центрифугирование в течение 10 мин при 13000 об/мин. Сливали надосадочную жидкость, осадок РНК промывали 75% EtOH и повторяли стадию центрифугирования. Надосадочную жидкость сливали и позволяли осадку РНК сушиться на воздухе в течение 15 мин с последующим суспендированием в не содержащей РНКазу воде. Для определения концентрации РНК в образце проводили спектрофотометрию, используя допущение, что когда A260/280 составляет 1,7-2,0, то показание прибора при A260 1= 40 мкг/мл РНК.
ПРИМЕР 4
Анализ нозерн-блот
Заливали в форму 1,1% агарозно-формальдегидный гель. Сочетали пять и десять микрограмм РНК с денатурирующим буфером (конечные концентрации: 6% формальдегида, 50% формамида и 0,1 x MOPS) и нагревали до 65°C в течение 10 мин. Добавляли одну десятую объема буфера для загрузки геля и загружали образец в гель. Электрофорез проводили при 75 В в 1 x буфере MOPS в течение ~3 ч. Гель промывали в течение 60 мин в 10xSSC.
РНК переносили на нейлоновую мембрану Hybond-N+(Amersham Biosciences, Piscataway, NJ) посредством капиллярного действия в течение 16 ч в 10 x SSC. Затем РНК закрепляли на нейлоновой мембране перекрестными сшивками с использованием функции самосшивания Stratagene UV Stratalinker (Stratagene, LA Jolla, CA). После закрепления нейлоновой мембране позволяли сушиться на воздухе.
Для мечения последовательностей ДНК 52 L1 дикого типа и 52 L1 R применяли набор I для мечения и регистрации ДНК Roche DIG High Prime (Hoffmann-La Roche Ltd., Basel, Switzerland) с использованием DIG в качестве зондов для выявления РНК 52 L1 дикого типа и 52 L1 R на нозерн-блоте. Предварительную гибридизацию, гибридизацию и иммунологическую обработку с использованием конъюгированного с щелочной фосфатазой антитела к DIG проводили согласно рекомендациям производителей. Вкратце, проводили предварительную гибридизацию блота в течение 30 мин при 37°C при осторожном встряхивании. Зонд денатурировали нагреванием до 95°C в течение 5 мин и последующим охлаждением на льду. Зонд добавляли к раствору для гибридизации и наносили на мембрану в течение 4 ч при 44,6°C при осторожном встряхивании. Затем раствор для гибридизации удаляли и блот промывали 2 x в течение 5 мин в 2xSSC с 0,1% SDS при комнатной температуре с последующей дополнительной промывкой при 65°C с 0,5xSSC и 0,1% SDS. Затем блот блокировали в течение 30 мин и наносили конъюгированное с щелочной фосфатазой антитело к DIG при разведении 1:5000 в течение 30 мин. Блот промывали и определяли наличие связавшейся с зондом РНК посредством регистрации связавшегося, конъюгированного со щелочной фосфатазой антитела к DIG с использованием субстрата NBT/BCIP.
Первоначальный анализ экспрессирующих HPV 52 L1 дикого типа дрожжей позволил предположить, что белок HPV 52 L1 экспрессировался; однако уровень был низким. В результате анализа нозерн-блот РНК из дрожжевых экстрактов культур, индуцированных для экспрессии HPV 52 L1 дикого типа, не было обнаружено какой-либо поддающейся выявлению РНК HPV 52 L1. Было очевидно, что поскольку выявили некоторый белок соответствующего размера, то были получены некоторые полноразмерные транскрипты РНК. Ген HPV 52 L1 реконструировали с последовательностями предпочтительных для дрожжей кодонов и сконструировали для исключения любых возможных участков преждевременной терминации транскрипции для обеспечения устойчивой транскрипции. В анализе нозерн-блот транскрипта HPV 52 L1 R было выявлено, что образовывались полноразмерные транскрипты и что они поддавались выявлению анализом нозерн-блот (фигура 3).
ПРИМЕР 5
Экспрессия белка HPV 52 L1
Замороженные осадки клеток дрожжей из индуцированных галактозой культур, эквивалентные OD600=10, размораживали на льду и суспендировали в 300 мкл буфера PC (100 мМ Na2HPO4 и 0,5 М NaCl, pH 7,0) с 2 мМ PMSF. Добавляли промытые кислотой стеклянные шарики 0,5 мм в концентрации ~0,5 г/пробирка. Пробирки встряхивали в течение 3 циклов по 5 мин при 4°C с перерывом в 1 мин. Добавляли 7,5 мкл 20% TritonX100 и повторяли стадию встряхивания в течение 5 мин при 4°C. Пробирки помещали на 15 мин в лед с последующим центрифугированием в течение 10 мин при 4°C. Надосадочную жидкость переносили в стерильную пробирку для микроцентрифугирования, которую обозначали как общий белковый экстракт дрожжей, указывали дату и хранили при -70°C.
ПРИМЕР 6
Анализ вестерн-блот
Для подтверждения экспрессии белка HPV 52 L1 после индукции галактозой анализировали вестерн-блотом общий белковый экстракт дрожжей из двадцати отдельных колоний дрожжей для каждого конструкта HPV 52 L1.
Десять, пять и два, а также половину микрограмма общего белкового экстракта дрожжей сочетали с буфером для загрузки SDS-PAGE и нагревали до 95°C в течение 10 мин. В качестве положительного контроля включали белок HPV 16 L1, составляющий приблизительно 55 кДа, вместе с не содержащим HPV L1 общим белковым экстрактом дрожжей в качестве отрицательного контроля (данные не представлены). Белки загружали в 10% гель SDS-PAGE и проводили электрофорез в буфере трис-глицин. После разделения белков белки переносили способом вестерн из геля на нитроцеллюлозу и получаемый в результате блот блокировали в 1 x буфере для разбавления (Kirkegaard and Perry Laboratories, Gaithersburg, MD) в течение 1 ч при комнатной температуре при покачивании. Блот три раза промывали и наносили в течение 16 ч при комнатной температуре абсорбируемую дрожжами козью антисыворотку к trpE-HPV 31 L1, перекрестно реагирующую с белками HPV 16 и HPV 52 L1. Затем блот три раза промывали и инкубировали с конъюгированным с HRP антителом к козе в разведении 1:2500 в течение 1 ч. Блот снова промывали три раза и наносили субстрат NBT/BCIP для регистрации (Kirkegaard and Perry Laboratories). Иммунореактивные белки выявляли в виде фиолетовых полос на блоте.
Во всех случаях белок HPV 52 L1 выявляли в виде отдельной иммунореактивной полосы на нитроцеллюлозе, соответствующей приблизительно 55 кДа (фигура 4). Интенсивность полосы HPV 52 L1 R (дорожка 2,5 мкг) выглядела значительно более высокой, чем полосы HPV 52 L1 дикого типа (10 мкг). Было очевидно, что после реконструкции уровень экспрессии HPV 52 L1 R с оптимизированными кодонами увеличился более чем в четыре раза, что представляет собой предел непосредственного сравнения на вестерн-блоте.
ПРИМЕР 7
Трансмиссионная электронная микроскопия
Для выявления того, что белок 52 L1 действительно самособирается, образуя пентамерные капсомеры L1, которые, в свою очередь, самособираются в вирусоподобные частицы, подвергали частично очищенный экстракт белка HPV 52 L1 R трансмиссионной электронной микроскопии (TEM).
Дрожжи выращивали в условиях культивирования в незначительном масштабе и осаждали центрифугированием. Получаемые осадки подвергали очистным обработкам. Осадок и очищенные экстракты дрожжей анализировали иммуноблотом для выявления экспрессии белка HPV 52 L1 и задержания в ходе процедуры очистки. Затем очищенные экстракты дрожжей подвергали центрифугированию над подушкой 45%-сахарозы и получаемый осадок суспендировали в буфере для анализа HPV 52 L1 VLP посредством TEM.
Характерный образец полученных HPV 52 L1 R VLP представлен на фигуре 5. Диаметр сферических частиц в этом неочищенном образце находится в диапазоне от 40 до 70 нм, причем у некоторых частиц наблюдают регулярное расположение капсомеров.
| название | год | авторы | номер документа |
|---|---|---|---|
| ОПТИМИЗИРОВАННАЯ ЭКСПРЕССИЯ HPV 58 L1 В ДРОЖЖАХ | 2004 |
|
RU2370538C2 |
| МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, КОДИРУЮЩАЯ HPV31 L1, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР, КЛЕТКА-ХОЗЯИН, ВИРУСОПОДОБНАЯ ЧАСТИЦА И СПОСОБ ЕЕ ПОЛУЧЕНИЯ, ВАКЦИНА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И СПОСОБЫ С ИХ ИСПОЛЬЗОВАНИЕМ | 2004 |
|
RU2356943C2 |
| ОПТИМИЗИРОВАННАЯ ЭКСПРЕССИЯ L1 HPV45 В ДРОЖЖАХ | 2004 |
|
RU2360001C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ, ПРИГОДНАЯ ПРИ ИНФЕКЦИЯХ HPV И ВИРУСОМ ГЕПАТИТА В, И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2009 |
|
RU2509570C2 |
| ПАПИЛЛОМАВИРУСНЫЕ ВАКЦИНЫ | 1995 |
|
RU2206608C2 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ПАПИЛЛОМАВИРУСНОЙ ИНФЕКЦИИ ЧЕЛОВЕКА И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2014 |
|
RU2546243C1 |
| Способ получения рекомбинантной вакцины для профилактики папилломавирусной инфекции человека, рекомбинантная вакцина | 2018 |
|
RU2681174C1 |
| ХИМЕРНАЯ ЧАСТИЦА HPV | 2012 |
|
RU2642287C2 |
| ОЧИЩЕННЫЕ БЕЛКИ ВИРУСА ПАПИЛЛОМЫ | 1995 |
|
RU2161651C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ ГЛАВНОГО КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 56 | 2014 |
|
RU2546240C1 |











Изобретение относится к области молекулярной биологии и вирусологии. Предложена молекула нуклеиновой кислоты для экспрессирования белка HPV 52 L1 в клетках дрожжей, вектор, содержащий эту молекулу, дрожжевая клетка-хозяин, включающая вектор, и способ получения подобных вирусу HPV 52 частиц. Изобретение может быть использовано для получения вакцин и обеспечения эффективной иммунопрофилактики папилломавирусной инфекции. 4 н. и 4 з.п. ф-лы, 5 ил.
1. Молекула нуклеиновой кислоты для экспрессирования белка LI HPV52 в клетке дрожжей, содержащая последовательность нуклеотидов, по существу соответствующую последовательности нуклеотидов, которая представлена в SEQ ID №:1, где молекула нуклеиновой кислоты кодирует белок HPV52 L1, как представлено в SEQ ID №:2, и где последовательность нуклеиновой кислоты представляет собой последовательность с оптимизированными кодонами для высокого уровня экспрессии в клетке дрожжей.
2. Экспрессионный вектор для экспрессии в клетке дрожжей, содержащий молекулу нуклеиновой кислоты по п.1, функционально связанную с промотором.
3. Дрожжевая клетка-хозяин, содержащая экспрессионный вектор по п.2.
4. Дрожжевая клетка-хозяин по п.3, где клетку дрожжей выбирают из группы, состоящей из: Saccharomyces cerevisiae, Hansenula polymorpha, Pichia pastoris, Kluyveromyces fragilis, Kluyveromyces lactis и Schizosaccharomyces pombe.
5. Дрожжевая клетка-хозяин по п.3, где клетка-хозяин представляет собой Saccharomyces cerevisiae.
6. Способ получения подобных вирусу HPV52 частиц (VLP), причем способ включает:
(a) трансформацию клетки дрожжей молекулой нуклеиновой кислоты, кодирующей белок HPV52 L1, по п.1;
(b) культивирование трансформированной клетки дрожжей в условиях, позволяющих экспрессировать молекулу нуклеиновой кислоты для получения рекомбинантного белка вируса папилломы; и
(c) выделение рекомбинантного белка вируса папилломы для получения VLP HPV52.
7. Способ по п.6, где клетку дрожжей выбирают из группы, состоящей из: Saccharomyces cerevisiae, Hansenula polymorpha, Pichia pastoris, Kluyveromyces fragilis, Kluyveromyces lactis и Schizosaccharomyces pombe.
8. Способ по п.7, где клетка дрожжей представляет собой Saccharomyces cerevisiae.
| US 2003096259 A1, 22.05.2003 | |||
| CERTER J.J., WIPF G.C., BENKI S.F., CHRISTENSEN N.D., GALLOWAY D.A | |||
| - J Virol | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |

