ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к композициям антигенов папилломавируса человека (HPV) и полипептидов, происходящих из них, используемым по отдельности или в комбинациях и также в качестве химерных молекул, слитых с бактериальными или вирусными антигенами, для профилактики и/или медикаментозного лечения инфекции HPV, а также вирусом гепатита B, у млекопитающих, в частности у человека.
Настоящее изобретение относится к фармацевтическому составу, способному вызывать защитный ответ антител и цитотоксических T-клеток против инфекции папилломавирусом человека (HPV). Иммуногенный состав содержит очищенные рекомбинантные антигены HPV в стабильном составе. Обсуждается способ индукции защитных ответов антител и цитотоксических T-клеток с использованием одновалентных, двухвалентных и поливалентных химерных вакцин. Иммунногенную композицию изготавливают для введения человеку in vivo.
УРОВЕНЬ ТЕХНИКИ
Папилломавирусы человека (HPV) представляют собой двухцепочечные ДНК-вирусы, которые инфицируют плоский и слизистый эпителий. Злокачественные эпителиальные новообразования, вызываемые типами HPV "высокого риска" или онкогенными типами HPV, могут прогрессировать в злокачественную карциному шейки матки (Bosch and de Sanjose, 2003). Персистирующая инфекция HPV в значительной степени коррелирует с инвазивной карциномой. Генотипы "высокого риска" могут постепенно прогрессировать от LSIL (низкозлокачественных внутриэпителиальных повреждений плоского эпителия) до злокачественной опухоли. (Schlecht et al. 2003). HPV является главным этиологическим фактором для рака шейки матки, поскольку выявлено, что практически в 99,7% случаев рака шейки матки имеются персистирующие инфекции HPV (Walboomers et al. 1999). Встречаемость инфекции HPV является частой среди молодых женщин с половой активностью, и оказалось, что свыше 95% встречающихся инфекций самопроизвольно разрешаются с течением времени. Инвазивная карцинома шейки матки развивается менее чем у 5% женщин, инфицированных HPV16 (Schlecht et al. 2003).
Среди почти 100 типов HPV, молекулярно идентифицированных до настоящего времени, и более 35 типов HPV, идентифицированных в половых путях, HPV16 составляет 50-60% случаев рака шейки матки в большинстве стран, за которым следуют HPV18 (10-20%), и HPV31 и 45 (4-5% каждый). Таким образом, HPV16 является наиболее распространенным типом во всех географических областях, рядом с которым следует HPV18, в частности, являясь более распространенным в Юго-Восточной Азии. Хотя эта картина является общей для плоскоклеточного рака (SCC) по всему миру, HPV18 является наиболее преобладающим в случае аденокарцином (ADC), а за ним следует HPV16 (Clifford et al. 2003). Эпидемиологические исследования выявили смешанную инфекцию HPV нескольких генотипов.
Данные других исследований показывают, что HPV16 является более персистирующим (Richardson et al. 2003; Londesborough et al. 1996), а также с большей вероятностью прогрессирует в CIN3, чем другие генотипы HPV высокого риска. Таким образом, в программах скрининга шейки матки на основе HPV, генотипирование может иметь диагностическую пользу при отделении случаев LSIL, положительных по HPV16 (и, возможно, HPV18 и другим генотипам), для которых требуется наиболее строгое наблюдение, от случаев LSIL, инфицированных другими генотипами высокого риска (Clifford et al. 2005).
Стратегии вакцинации, основанные на доступных эпидемиологических данных о характерном распределении инфекции HPV по возрасту и типу, могут формировать основу для разработки эффективной вакцины для контроля и устранения инфекции папилломавирусом человека. Для эффективной вакцинации должна рассматриваться стратегия вакцин-кандидатов, которая обеспечивает широкую защитную эффективность против множества генотипов. Доступные вакцины с использованием капсидных белков L1 являются типоспецифическими для HPV16, HPV18, HPV6 и HPV11 и обеспечивают недостаточную защиту от инфекции, вызываемой другими генотипами, такими как HPV31, HPV35, HPV52, HPV58 и т.д., которые также ассоциированы с прогрессией в карциному шейки матки. В настоящее время отсутствуют доступные вакцины против HPV, которые обеспечивают перекрестную защиту против нескольких генотипов HPV.
Были выданы лицензии на две профилактические вакцины, содержащие рекомбинантные вирусоподобные частицы L1 HPV (VLP). Они нацелены только на два из приблизительно 15 известных онкогенных типов HPV. Хотя приблизительно 70% случаев рака шейки матки свойственны этим двум типам, существуют данные о некоторой степени защиты "Ciption of tross" против других близкородственных типов. Однако текущая стоимость этих вакцин исключает непрерывную доставку по всему миру, в частности, в треть стран мира, где встречаемость инфекции HPV является высокой.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На фиг.1 представлена амплификация посредством ПЦР гена малого антигена гепатита B (HepBsAg). Дорожка 1:~700 п.н. гена HbsAg; дорожка 2: лестничная ДНК по 100 п.н.
На фиг.2 представлена амплификация посредством ПЦР мультиэпитопных последовательностей синтетического фрагмента гена E6 и E7 для конструирования химеры HbsAg-HPV. Дорожка 1:мультиэпитопный фрагмент гена размером ~150 п.н., содержащий мультиэпитопную последовательность размером 129 п.н.; дорожка 2:лестничная ДНК по 100 п.н.
На фиг.3 представлена амплификация посредством ПЦР фрагмента гена HPV16L2 размером ~530 п.н., содержащего ген HPV16L2 размером 510 п.н., для получения химерной последовательности HbsAg-HPV; дорожка 2: лестничная ДНК по 100 п.н.
На фиг.4 представлена амплификация посредством ПЦР фрагмента HPV16L2, соответствующего аминокислотам 100-220 гена L2, для получения фрагмента ПЦР размером ~380 п.н., который содержит конкретную последовательность размером 360 п.н.
На фиг.5 проиллюстрирована амплификация посредством ПЦР фрагмента гена HPV16L1 размером 99 п.н., амплифицированного посредством ПЦР, с получением фрагмента размером ~120 п.н., соответствующего аминокислотам 100-220 гена L2, для получения фрагмента ПЦР размером ~380 п.н., который содержит конкретную последовательность размером 360 п.н.
На фиг.6 проиллюстрировано, что амплификация посредством ПЦР гена HPV18L1 приводит к фрагменту размером ~140 п.н., содержащему последовательность HPV18L1 размером 117 п.н.
На фиг.7 показано, что амплификация посредством ПЦР гена HPV18L1 приводит к фрагменту размером ~500 п.н., содержащему фрагмент HPV18L1 размером 480 п.н.
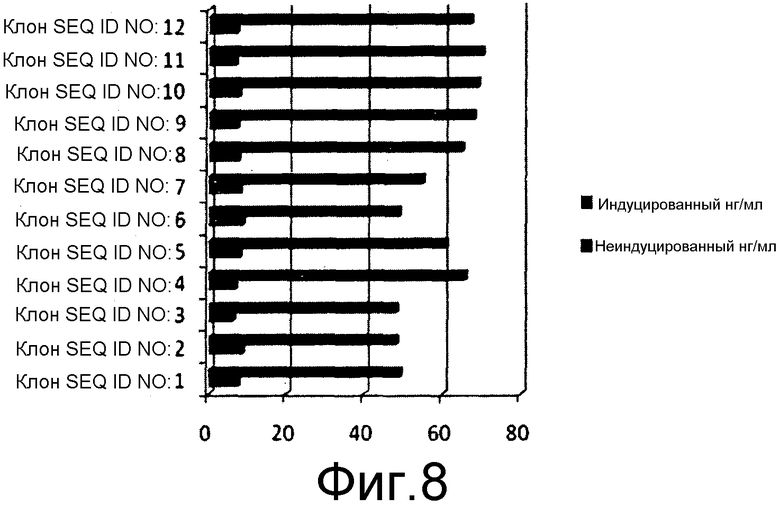
На фиг.8 представлен уровень экспрессии химерного белка HbsAg-HPV, выявленный посредством ELISA по содержанию HbsAg и взятый в качестве показателя экспрессии химерного белка в индуцируемых культурах Pichia pastoris.
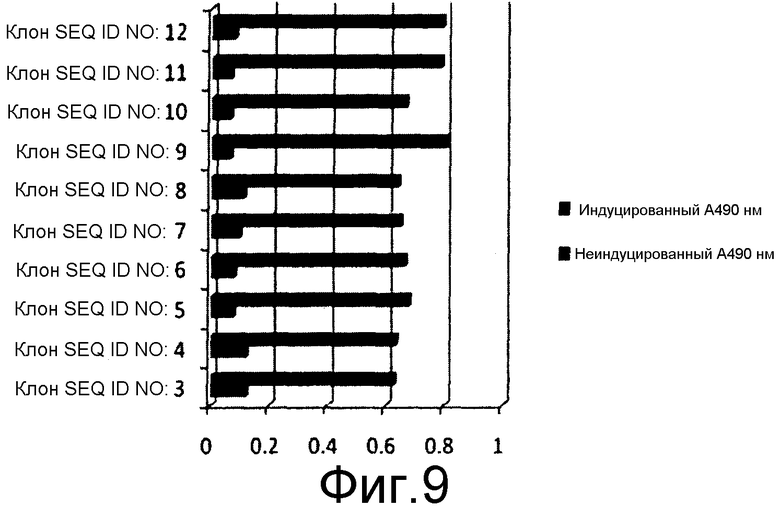
На фиг.9 представлен уровень экспрессии химерного белка HbsAg-HPV, выявленный посредством ELISA по содержанию антигена HPV в индуцируемых культурах Pichia pastoris.
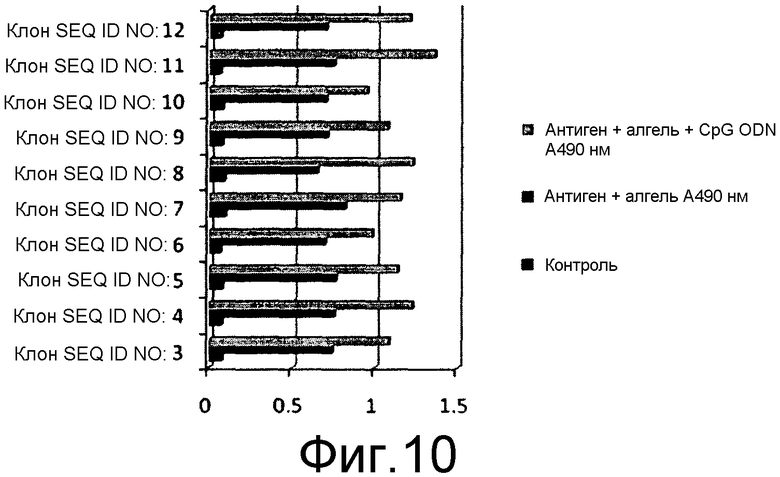
На фиг.10 представлена иммуногенность химерных белков, выявленных с использованием специфичного к HPV ELISA с использованием первичного антитела, индуцированного против коммерчески доступной четырехвалентной вакцины, и антисыворотки кролика против HPV16L2, индуцированной против очищенного белка.
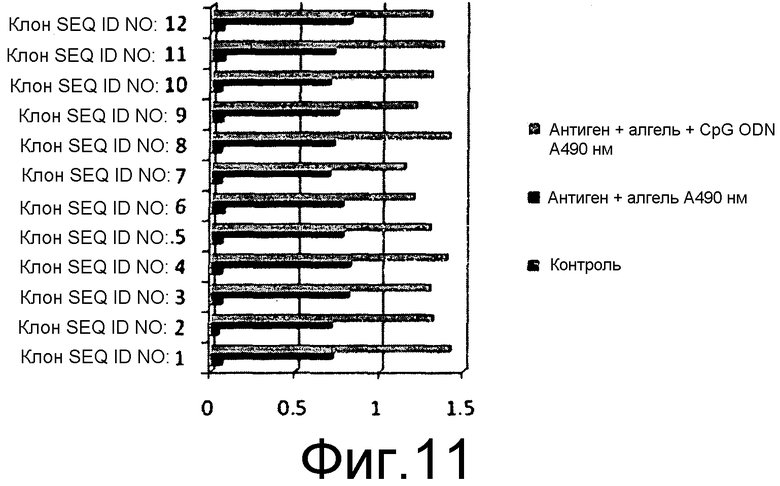
На фиг.11 представлена иммуногенность химерных белков, выявленных с использованием специфичного к HbsAg ELISA с использованием коммерчески доступного набора. Титр антител против конструкции, содержащей белок HPV16L2, оценивали с использованием антисыворотки кролика, индуцированной против очищенного белка L2.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Таким образом, настоящее изобретение относится к вакцинным композициям, содержащим химерные белки антигенов HPV, слитые с вирусными или бактериальными белками, обеспечивающим усиленную иммуногенность, полезную при инфекциях вирусом гепатита B, а также при инфекциях папилломавирусом человека (HPV).
Антигены HPV могут быть выбраны из группы, включающей, но не ограничивающейся ими, HPV16, HPV18, HPV6, HPV11, HPV31, HPV33, HPV35, HPV39, HPV45, HPV51, HPV52, HPV56, HPV58, HPV59, HPV68.
Антигенная композиция, содержащая полные/неполные последовательности/мутанты/варианты антигенов HPV, может быть выбрана из списка, включающего антигены L1, L2, E2, E5, E6 и E7, или конкретные T-клеточные или B-клеточные эпитопы указанных антигенов, либо в качестве отдельной последовательности, либо в тандемных повторах в химерных молекулах, слитых с бактериальными/вирусными белками, способными индуцировать сильный антительный ответ при введении хозяину.
Вирусные и бактериальные белки могут быть выбраны из списка, включающего, но не ограничивающегося ими, антигены вируса гепатита B, такие как ядерный антиген вируса гепатита B, малый антиген вируса гепатита B, инактивированный столбнячный токсоид, дифтерийный токсоид, термолабильный токсин E.coli, холерный токсин, экзотоксин A Pseudomonas, шига-токсин, листериолизин, либо отдельно, либо в комбинациях, или эпитопы, полученные из указанных выше белков.
Антигенная композиция состоит из химерных молекул антигенов HPV, слитых с малым антигеном гепатита B (HbsAg).
Антиген HPV содержит либо полные, либо неполные последовательности/эпитопы капсидных белков, таких как L1 и L2, или полипептидов, полученных из L1 и/или L2, слитые с HbsAg, такие как в с SEQ ID NO: 3 по SEQ ID NO: 13.
Антигены HPV содержат полную/неполную последовательность/ мутанты/варианты/эпитопы ранних белков HPV, таких как E2, E5, E6 и E7, либо отдельно, либо в их комбинациях, таких как в SEQ ID NO: 1 и 2.
Антигены HPV подвергают слиянию с N-концом, с C-концом или с любой областью в открытой рамке считывания малого антигена гепатита B с добавлением линкерной последовательности между двумя последовательностями или без ее добавления.
Согласно другому аспекту этого изобретения предусматривается способ экспрессии указанных выше химерных антигенов HPV и HPV-HBsAg по п.1, включающий культивирование клетки-хозяина, трансформированной антигеном/слитым антигеном по п.1, выделения и очистки из них рекомбинантных белков.
Рекомбинантная экспрессия указанных выше антигенов HPV может происходить в хозяине либо в качестве внутриклеточных белков, либо в качестве секреторных белков, причем хозяином является либо прокариотическая, либо эукариотическая экспрессирующая система, предпочтительно дрожжи.
Фармацевтическая композиция содержит антигенные композиции, описанные в настоящем документе выше, в эффективном количестве, подлежащие применению в вакцине, и фармацевтически приемлемый носитель.
Антигены используют в диапазоне 10 мкг - 1000 мкг белка на дозу, предпочтительно от 10 мкг до 200 мкг и наиболее предпочтительно от 10 мкг до 100 мкг.
Кроме того, фармацевтическая композиция содержит пригодный адъювант, причем адъювант выбран по меньшей мере из одного из следующих: гидроксид алюминия, фосфат алюминия, монофосфориллипид A, другие органические липофильные соединения, содержащие CpG и не содержащие CpG олигонуклеотиды, цитокины.
Способ индукции защитного ответа антител на антигенную композицию и/или ответа цитотоксических T-клеток согласно любому из предшествующих пунктов, включает введение указанной композиции субъекту, нуждающемуся в таком лечении, по меньшей мере одним из путей, таким как пероральный путь, путь через слизистую оболочку или парентеральный путь введения. Указанные выше антигенные композиции заключают в фармацевтически приемлемые биодеградируемые полимеры.
Указанные выше антигенные составы применяют для профилактики или медикаментозного лечения инфекций папилломавирусом и ассоциированного с ними риска злокачественной опухоли у млекопитающих, в частности у человека.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Изобретение относится к разработке профилактических или терапевтических вакцин-кандидатов для человека от инфекций папилломавирусом человека. Из уровня техники известно, что основной капсидный белок HPV, белок L1, может собираться в вирусоподобную частицу (VLP), либо отдельно, либо в комбинации с минорным капсидным белком L2, при экспрессии в качестве рекомбинантных белков in vitro. Эти вирусоподобные частицы являются иммуногенными и их используют в качестве вакцин для профилактики инфекций HPV у людей. Карцинома шейки матки вызывается несколькими "онкогенными" типами HPV, включающими HPV16, HPV18, HPV31, HPV33, HPV35, HPV39, HPV45, HPV51, HPV52, HPV56, HPV58, HPV59, HPV68 и т.д. VLP, содержащие капсидные белки L1, являются хорошими вакцинами-кандидатами, однако антитела, которые распознают конкретные структурные эпитопы на VLP, являются специфичными к генотипу, и антитела против одного типа VLP демонстрируют небольшую перекрестную реактивность в отношении других генотипов HPV, или демонстрируют ее отсутствие. Однако является распространенной смешанная инфекция HPV нескольких генотипов.
В объем настоящего изобретения входит разработка универсальных вакцин, которые могут обеспечить защитный иммунитет против основных типов HPV, которые ассоциированы с риском карциномы шейки матки. Минорный капсидный белок HPV L2 обеспечивает потенциально широкую нейтрализующую активность против нескольких генотипов вируса вследствие более высокой консервативности последовательностей в белке L2. Однако в отличие от структурных эпитопов L1, которые являются высоко антигенными, белок L2 является относительно слабоиммуногенным. Его низкое содержание в природных капсидах - приблизительно 12-72 молекул на 360 копий L1, ограничивает его иммуногенность (Pereira et al. 2009). Возможным подходом для расширения иммунитета при более низких затратах является рассмотрение вакцинации против L2. Вакцины против L2 можно получать при относительно низких затратах и они также имеют перспективу обеспечения значительно более широкого защитного иммунитета перекрестного типа, чем в случае, наблюдаемом при иммунизации VLP L1 (Kanda and Kondo 2009). Минорный капсидный белок L2 HPV, который играет важную роль в проникновении вируса в клетки, локализации вирусных компонентов в ядре, в связывании ДНК, образовании и стабильности капсида, индуцирует антитела, которые являются более перекрестно-реактивными среди типов HPV, чем в случае основного капсидного белка L1, делая его привлекательной потенциальной мишенью для субъединичных вакцин против инфекций HPV нового поколения с более широкой защитой.
Задачей изобретения является повышение иммуногенности вакцины-кандидата(вакцин-кандидатов), позволяя представление множества копий антигенных эпитопов для усиления иммуногенности, посредством химерного слияния с вирусными и бактериальными белками. До настоящего времени отсутствует коммерческое производство мультиэпитопной вакцины против инфекции HPV. Мультиэпитопные вакцины обеспечивают преимущество широкой защитной активности вследствие высокой консервативности последовательностей антигенных эпитопов капсидных и ранних белков среди различных генотипов вируса.
Вирусоподобные частицы, образованные малым антигеном гепатита B (HbsAg), представляют собой мультимерную структуру, которая является высоко иммуногенной при введении в качестве вакцины против инфекции вирусом гепатита B. Структура VLP позволяет встраивание гетерологичных последовательностей из других белков в его открытую рамку считывания в различных областях, позволяя применение химерных слитых белков с HbsAg в качестве потенциальных вакцин-кандидатов. Антигены HPV включают капсидные белки L1 и L2, и ранние белки, такие как E1, E2, E5, E6 и E7. Эти белки могут состоять из полной/неполной последовательности/мутантов/вариантов/эпитопов белков L1, L2, E1, E2, E5, E6 и E7 HPV, либо по отдельности, либо в сочетаниях. T-клеточные или B-клеточные эпитопы этих белков-кандидатов используют либо по отдельности, либо в тандемных повторах, и подвергают слиянию с HbsAg для применения в качестве вакцин-кандидатов для вакцинации против HPV. Антигены HPV подвергают слиянию либо с N-концом или C-концом, либо с любой областью в открытой рамке считывания малого антигена гепатита B с добавлением линкерной последовательности между двумя антигенами, или без ее добавления.
Химерная молекула капсидных белков HPV, слитая с HbsAg, не была описана до настоящего времени. Химера минорного капсидного белка L2 HPV с HbsAg является привлекательной в качестве кандидата для вакцины, поскольку белок L2, хотя и обеспечивает широкую нейтрализующую активность против множества генотипов вируса, сам по себе является слабым антигеном, как и его линейный полипептид. Предоставление областей L2, ответственных за его широкую нейтрализующую активность против множества генотипов вируса HPV в качестве мультимерных копий на VLP HbsAg, повышает его иммуногенность посредством предоставления множества копий VLP, по сравнению с тем, когда они представлены в виде одной копии. Области белков L1 HPV16 и HPV18, выбранные для создания химерных конструкций с HbsAg, также представляют собой области, которые обладают широкой нейтрализующей активностью против других генотипов, и потенциально могут обеспечить перекрестную защиту. Авторы настоящего изобретения исследовали универсальность HbsAg, самосборка которого в вирусоподобные частицы все еще позволяет химерное слияние с гетерологичными последовательностями других белков. Авторы настоящего изобретения изготовили химеру из HbsAg с B-клеточными и T-клеточными эпитопами белков E6 и E7 HPV, которые предоставлены в тандеме, для повышения их иммуногенного потенциала, а также с белками L1 и L2 HPV16 и HPV18. Была описана химерная конструкция различных областей E7 HPV с белком HbsAg. Авторы настоящего изобретения использовали множество T- и B-клеточных эпитопов, происходящих из белков E6 и E7 HPV, поскольку присутствие этих последовательностей в тандеме приводит к более высокой иммуногенности, чем когда они присутствуют в качестве единичной копии. Их предоставление на мультимерной VLP из HbsAg в свою очередь увеличивает количество копий, которые оказываются на VLP, что дополнительно повышает их иммуногенность и обеспечивает превосходный кандидат для вакцины. Известно, что B- и T-клеточные эпитопы вызывают сильные цитотоксические T-клеточные ответы, обеспечивающие перспективу хорошей терапевтической вакцины против инфекций HPV.
Специалисты в данной области смогли установить, что также в сходной стратегии в качестве химерных последовательностей с антигенами HPV для повышения вакцинного потенциала можно использовать следующие кандидаты. Список кандидатов включает, но не ограничивается ими, ядерный антиген вируса гепатита B, инактивированный столбнячный токсоид, дифтерийный токсоид, термолабильный токсин E.coli, холерный токсин, экзотоксин A Pseudomonas, шига-токсин, листериолизин, белки теплового шока, кальретикулин или эпитопы, происходящие из указанных выше эпитопов.
Указанные выше белки можно экспрессировать либо в прокариотических, либо в эукариотических экспрессирующих системах, в качестве внутриклеточных или секреторных белков. Pichia pastoris в качестве экспрессирующей системы являются преимущественными в промышленном масштабе, поскольку они являются экономически выгодными, и несколько белков, которые экспрессированы и очищены из Pichia pastoris успешно выпустили в серийное производство, поскольку было определено, что они являются безопасными для применения у человека. Эта система может быть применима к дрожжам любого рода или вида. Химерная слитая молекула антигенов HPV с секреторной сигнальной последовательностью HSA (сывороточного альбумина человека) для повышения синтеза и секреции белка в культуральную среду, обеспечивает не только высокий уровень экспрессии, но также простоту очистки белка в масштабе промышленной ферментации. Последовательность HSA на его N-конце совместима с высоким уровнем синтеза и эффективной секрецией в Pichia pastoris. Очистка вирусных антигенов с использованием запатентованной технологии HimaxTM упрощает выделение химерных антигенов HbsAg-HPV, в частности, VLP, в экономически выгодном способе, который является безопасным для применения у человека и экономичным в промышленном масштабе, по сравнению с общепринятым ультрацентрифугированием в градиенте плотности на хлориде цезия. Это позволит продукцию недорогой вакцины, которая является безвредной для окружающей среды вследствие избегания применения тяжелых металлов, таких как хлорид цезия.
Белки экспрессируются в качестве VLP, капсомеров, полипептидов или химерных полипептидов, содержащих HPV и бактериальные/вирусные антигены. Указанные антигены включены либо в качестве единичной последовательности, либо в виде тандемных повторов, поскольку химерные слитые молекулы с бактериальными/вирусными белками способны индуцировать сильный ответ антител или ответ цитотоксических T-клеток при введении хозяину. Субъединичные иммуногены, содержащие копии T- и B-клеточных эпитопов в тандеме, потенциально являются более иммуногенными, чем когда они встречаются в виде единичной копии. Химерные конструкции с эпитопами E6 и E7, а также с белками L2 и L1, индуцируют сильный ответ антител как против эпитопов HbsAg, так и против эпитопов HPV, при определении посредством ELISA. Это обеспечивает вакцину для двойного применения, как против гепатита B, так и против множества генотипов вируса HPV.
Антигенные композиции указанных выше беков изготавливают в фармацевтически приемлемом носителе для иммунизации у человека. Антигенные составы можно вводить любо отдельно, либо с адъювантом, выбранным из списка, который включает, но не ограничивается ими: гидроксид алюминия, фосфат алюминия, монофосфориллипид A, другие органические липофильные соединения, содержащие CpG и не содержащие CpG олигонуклеотиды, цитокины и т.д. Альтернативно антигенный состав может содержать два или более очищенных антигена, которые смешаны in vitro в пригодном составе, и он может использоваться в качестве вакцины. Антигенные составы можно доставлять одним из нескольких способов пероральным путем, путем через слизистую оболочку или парентеральным путем у млекопитающих, предпочтительно у человека, для того, чтобы вызвать защитный ответ антител и/или цитотоксических T-клеток. Также антигены можно доставлять в фармацевтически приемлемых биодеградируемых полимерах. Указанные выше составы можно использовать для профилактики или медикаментозного лечения инфекций папилломавирусом и связанного с ними риска злокачественной опухоли у млекопитающих.
HPV проникает в организм через слизистые клетки и не распространяется системно. Таким образом, вакцина против HPV возможно будет индуцировать сильный и длительный иммунный ответ в слизистых оболочках половых путей. В связи с этим, исследования назальной и пероральной иммунизации у животных показали, что VLP индуцирует антитела в слизистой половых путей. Вакцины, обеспечивающие иммунитет слизистых оболочек против антигенов HPV, являются преимущественными для профилактики инфекции HPV. Составы, пригодные для доставки вакцин через слизистую оболочку, могут быть протестированы. Для применения на слизистую оболочку не существует таких коммерчески доступных вакцин против HPV.
Кроме того, изобретение иллюстрируется следующими примерами. Однако их можно использовать только для цели иллюстрации, и они не подразумевают объем охраны.
Примеры:
201. Пример 1: Стратегия клонирования
a) Стратегия 1: Химерная молекула мультиэпитопов, происходящих из ранних белков, слитых с HbsAg:
Эта последовательность состоит из малого антигена вируса гепатита B, слитого с мультиэпитопом, происходящим из белков E7 HPV16 и HPV18, которые расположены тандемно, с последующим химерным слиянием молекулы мультиэпитопа с C-концом и N-концом белка HbsAg. (последовательность белка HbsAg:

Следующие эпитопы, происходящие из белков E7 и/или E6, располагают тандемно для получения: 5'-EYMLDLQPETTEEDEIDGPAGQAEPDRAHYNIDEIDGVNHQHL-3'-43 аминокислоты (129 нуклеотидов). Этот главный эпитоп представляет собой следующую комбинацию и он выбран из списка T- и B-клеточных эпитопов: T-клеточный эпитоп E7 HPV16: а.к. 20-29: TDLYCYEQLN; а.к. 45-54 AEPDRAHYNI; B-клеточные эпитопы: а.к. 10-20: EYMLDLQPETT; а.к. 35-49 EEDEIDGPAGQAEPDR; слитых с B-клеточным эпитопом E7 HPV18 DEIDGVNHQHL, который представляет собой иммунодоминантный эпитоп E7 HPV18, с получением следующей химерной слитой молекулы, обозначенной как SEQ ID NO: 1. Сходная химерная слитая молекула мультиэпитопа, слитого на N-конце с HbsAg, представляет собой последовательность, предоставленную в SEQ ID NO: 2. Для инициации трансляции на 5'-конец мультиэпитопа помещали инициирующий метионин для трансляции.

b) Стратегия 2: Химерная слитая молекула минорного капсидного белка L2 и HbsAg.
N-концевую область минорного капсидного белка L2 подвергали слиянию с HbsAg на C-конце HbsAg. Амплифицированные области L2 представляют собой 171 аминокислоту со следующей последовательностью (инициирующий метионин встраивали для обеспечения трансляции), слитой с C-концом HbsAg

SEO ID NO: 4:
Последовательность минорного белка капсида L2, содержащую 171 аминокислоту со следующей последовательностью, подвергают слиянию на ее C-конце с N-концом HbsAg с получением SEQ ID NO: 4. Для трансляции добавляют инициирующий метионин.

c) Стратегия No.3:
Химерная слитая молекула фрагмента L2 с гнездовыми делециями с HbsAg.
N-концевую область минорного капсидного белка L2 подвергали слиянию с HbsAg на C-конце HbsAg. Амплифицированная область L2 составляет приблизительно 120 а.к. (360 п.н.) со следующей последовательностью:

Ее лигируют с C-концом HbsAg с получением SEQ ID No: 5:

SEO ID No: 6:
Последовательность минорного капсидного белка L2, содержащую следующую последовательность из 120 аминокислот:
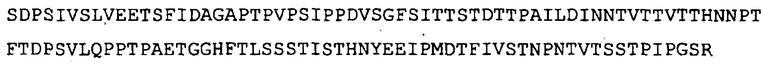
подвергают химерному слиянию на ее C-конце с N-концом HbsAg с получением SEQ ID NO: 6. Для трансляции добавляли инициирующий метионин.


d) Стратегия No. 4: Химерная слитая молекула области L1 HPV16 с HbsAg:
Получали химерную слитую молекулу области L1 белка HPV16 на C-конце HbsAg, состоящей из 33 аминокислот, происходящих из области L1 белка HPV16 (99 п.н.) с последовательностью: NTVIQDGDMVDTGFGAMDFTLQANKSEVPLDI

Химерную слитую молекулу области L1 белка HPV16, состоящей из 33 аминокислот, происходящих из области L1 HPV16 последовательности: NTVIQDGDMVDTGFGAMDFTTLQANKSEVPLDI с N-концом HbsAg для получения химерной последовательности SEQ ID NO: 8. Для трансляции добавляли инициирующий метионин.

e) Стратегия No. 5:
Химерная слитая молекула области HPV18 L1 с HbsAg:
Химерную слитую молекулу области белка L1 HPV18 (39 аминокислот; 117 п.н.) со следующей последовательностью
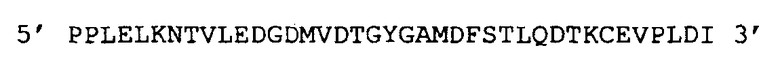
подвергали слиянию с C-концом последовательности HbsAg для получения следующей химерной последовательности:

Ее подвергают слиянию с N-концом белка HbsAg с получением SEQ ID NO: 10, где для трансляции добавляли инициирующий метионин.

f) Стратегия No. 6: Химерное слияние более крупного полипептида области L1 HPV18 с HbsAg:
Химерная слитая молекула более крупного фрагмента HPV18 состоит из следующей последовательности

и подвергают слиянию на ее N-конце с C-концом HbsAg с получением химерной последовательности:

Химерная слитая молекула более крупного фрагмента HPV18 состояла из следующей последовательности:

и ее подвергали слиянию на ее C-конце с N-концом HbsAg с получением химерной последовательности SEQ ID 12:

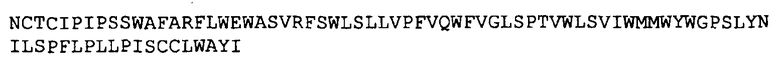
Стратегия No. 7:
Химерная слитая молекула полноразмерной последовательности белка L1 HPV16 на ее N-конце с сигнальной последовательностью из 24 аминокислот сывороточного альбумина человека для секреторной экспрессии в pichia pastoris с получением SEQ ID NO: 13

SEQ ID NO: 14:
Химерная слитая молекула полноразмерной последовательности белка L2 HPV16 на ее N-конце с сигнальной последовательностью из 24 аминокислот сывороточного альбумина человека для секреторной экспрессии в pichia pastoris с получением SEQ ID NO: 14

SEQ ID NO: 15:
Химерная слитая полноразмерная последовательность белка L1 HPV18 на его N-конце с сигнальной последовательностью из 24 аминокислот сывороточного альбумина человека для секреторной экспрессии в pichia pastoris с получением SEQ ID NO: 15:

2. Пример 2: Клонирование гена и конструирование химер антигена HbsAg-HPV:
Ген гепатита B, кодирующий полноразмерный белок HbsAg, амплифицировали посредством ПЦР с N-концевыми и C-концевыми праймерами, содержащими участки рестрикции EcoRI и BamHI, соответственно, с получением фрагмента ПЦР размером ~700 п.н. (См. фиг. 1). Праймеры, использованные для амплификации посредством ПЦР гена HbsAg, представляют собой:
HBNTERM: 5' CACGAATTCACCATGGAGAACACAACATCAGG 3'
HBCTERM: 5' TCAGGATCCAATGTATGCCCAAAGACAACAGG 3'
Условия амплификации посредством ПЦР представляли собой денатурацию при 94°С в течение 45 секунд, отжиг при 65°С в течение 40 секунд и удлинение при 72°С в течение 1,5 минут в течение 30 циклов с последующим конечным удлинением при 72°С в течение 10 минут. 10 нг плазмидной ДНК, содержащей ген HbsAg, клонированный в pBluescript SKII+, использовали в качестве матрицы для реакции ПЦР. Реакция состояла из 1x буфера для ДНК-полимеразы Taq, 0,25 мМ dNTPS, 20 пикомоль каждого праймера и 1,5 Ед ДНК-полимеразы Taq. Все реагенты были от Bangalore Genei. Фрагмент ПЦР очищали из геля и расщепляли BamHI. Расщепленный BamHI фрагмент ПЦР HbsAg использовали для лигирования с различными антигенами HPV для получения химерных конструкций. Для конструирования химерных конструкций, которые несли ген HbsAg на C-конце и антигены HPV на N-конце, HbsAg амплифицировали с помощью ПЦР с C-концевым праймером, содержащим участок NotI, и N-концевым праймером, который содержал участок BamHI. В конструкциях, использованных для амплификации посредством ПЦР, где антигены HPV находятся на N-конце и HbsAg находится на C-конце, праймеры ПЦР для антигенов HPV содержали участок EcoRI на их N-конце и участок BamHI на их C-конце. В обоих случаях, получали химерные слитые молекулы гена HbsAg и гена HPV посредством лигирования на BamHI-конце обеих последовательностей. Участок BamHI вносил две аминокислоты "GS" (глицин-серин) в область соединения последовательностей HbsAg-HPV. Все описанные в данном описании химерные слияния (с SEQ ID No: 1 по SEQ ID No: 15) проводили на нуклеотидном уровне с использованием полимеразной цепной реакции (ПЦР) и лигирования амплифицированных посредством ПЦР генов.
Следующие фрагменты генов HPV амплифицировали посредством ПЦР для лигирования.
Клонирование мультиэпитопной последовательности
EYMLDLQPETTEEDEIDGPAGQAEPDRAHYNIDEIDGVNHQHL - 43 аминокислоты (129 нуклеотидов). Синтетический фрагмент гена, кодирующий указанную выше последовательность, синтезировали в GenScript Corporation. Фрагмент гена амплифицировали с помощью следующих праймеров, которые вносили участок BamHI на N-конце и участок NotI на C-конце. Реакция ПЦР представляла собой денатурацию при 94°С в течение 45 секунд, отжиг при 62°С в течение 45 секунд и удлинение при 72°С в течение 30 секунд в 33 циклах с получением фрагмента ПЦР размером ~140 п.н., содержащего эпитопную последовательность размером 129 п.н.
Праймер MENTERM: 5'-ATAGGATCCGAGTACATGTTGGATTTG-3'
Праймер MECTERM 5'-ATCGCGGCCGCTTATCACAAATGTTGGTGGTTG-3'
Фрагмент ПЦР размером 129 п.н. (см. фиг. 2) с использованием указанных выше праймеров очищали из геля и расщепляли BamHI. Расщепленный BamHI фрагмент ПЦР лигировали в расщепленный BamHI ген HbsAg в течение ночи с использованием ДНК-лигазы T4 для получения химерного фрагмента размером ~840 п.н., транслированная последовательность которого обозначена как SEQ ID NO: 1. Лигированный фрагмент правильного размера (~840 п.н.) очищали из геля, и расщепляли EcoRI и NotI для клонирования в дрожжевой вектор pPIC3.5K. Для получения SEQ ID NO: 2, где мультиэпитопная последовательность лигирована на 5'-конце гена HbsAg, изготавливали праймеры со сходными последовательностями, за исключением того, что C-концевой праймер содержал участок BamHI вместо участка NotI, и в N-концевом праймере участок BamHI был заменен участком EcoRI. Продукт ПЦР размером ~140 п.н. расщепляли BamHI и лигировали с расщепленным BamHI HbsAg в течение ночи с ДНК-лигазой T4. Лигированный фрагмент размером ~840 п.н., обозначенный как SEQ ID NO: 2, очищали из геля и расщепляли EcoRI и NotI для клонировали в дрожжевой экспрессирующий вектор pPIC3.5K.
Химерное слияние фрагмента L2 HPV16 на C-конце HbsAg
N-концевую область минорного капсидного белка L2 подвергали слиянию с HbsAg на C-конце HbsAg. Ген L2 HPV16 синтезировали в GenScript Corp. Амплифицированные области L2 соответствовали аминокислотам 50-220 (510 п.н.) и их лигировали на C-конце HbsAg с получением SEQ ID NO: 3. Для лигирования фрагмента гена L2 с C-концом гена HbsAg, ген, кодирующий белок L2, амплифицировали посредством ПЦР со сходным N-концевым праймером, содержащим участок BamHI, и C-концевым праймером, содержащим участок NotI. Амплифицированный посредством ПЦР фрагмент размером ~530 п.н. расщепляли BamHI и лигировали с расщепленным BamHI HbsAg, который содержал участок BamHI на его C-конце. Лигированный фрагмент размером ~1230 п.н. очищали из геля и расщепляли EcoRI и NotI для клонирования в дрожжевой вектор pPIC3.5K.
Для получения химерной конструкции HPV-HbsAg, ген L2 (положение аминокислот 50-220 п.н.) амплифицировали с праймерами со сходной последовательностью, за исключением того, что N-концевой праймер содержал участок EcoRI вместо участка BamHI и C-концевой праймер содержал участок BamHI вместо участка NotI. Амплификация посредством ПЦР и расщепления были такими, как описано выше, с получением SEQ ED NO: 4.
Химерное слияние фрагмента L2 HPV16 с гнездовыми делециями на C-конце HbsAg
N-концевую область минорного капсидного белка L2 подвергали слиянию с HbsAg на C-конце HbsAg. Амплифицированные области L2 представляли собой 100-220 аминокислот (120 а.к.; 360 п.н.). Праймеры, использованные для амплификации посредством ПЦР фрагмента гена L2, представляют собой:
6P2CFNTERM: 5'-ACCGGATCCGATCCATCTATCGTTTC-3'
6P2CFCTERM: 5'-ATCGCGGCCGCTTATCATCTTGAACCTGGAATTG-3'
Реакцию ПЦР проводили в объеме 50 мкл, содержавшем 1x буфер для ДНК-полимеразы Taq, 0,25 мМ dNTP и 20 пикомоль каждого праймера и 1,5 Е ДНК-полимеразы Taq. Условия ПЦР представляли собой денатурацию при 94°С в течение 45 секунд, 64°С в течение 40 секунд и удлинение при 72°С в течение 1 мин в 30 циклах.
Фрагмент ПЦР размером ~380 п.н., содержавший 360 п.н. последовательности L2, очищали из геля, расщепляли BamHI и лигировали в расщепленный BamHI ген HbsAg с получением химеры HbsAg-HPV размером ~1060 п.н., обозначенной как SEQ ID NO: 5. Для получения химерной последовательности HPV-HbsAg, где фрагмент гена L2 лигирован с N-концом гена HbsAg, N-концевые и C-концевые праймеры для L2 содержали участки рестрикции EcoRI и BamHI. Лигирование проводили между расщепленным BamHI фрагментом гена L2 и расщепленным BamHI HbsAg с получением химерной последовательности из 1060 п.н., обозначенной как SEQ ID NO: 6.
Получение химерной последовательности с областью L1 HPV16 и HbsAg
Получение химерной последовательности, состоящей из 33 аминокислот, происходящих из области L1 белка HPV16 (99 п.н.) последовательности: NTVIQDGDMVDTGFGAMDFTTLQANKSEVPLDI с геном HbsAg. Указанную область гена L1 HPV16 (ген, синтезированный в GenScript Corporation), амплифицированную посредством ПЦР с получением фрагмента ПЦР размером ~120 п.н., содержащего 99 п.н. указанной выше последовательности, амплифицировали с использованием следующих праймеров для ПЦР:
6P1CFNTERM: 5'-ATCGGATCCAACACCGTTATCCAAGACGG-3'
6P1CFCTERM: 5'-ACTGCGGCCGCTTATCAAATATCCAATGGAACTTC-3'
Реакцию ПЦР проводили в объеме 50 мкл, содержавшем 1x буфер для ДНК-полимеразы Taq, 0,25 мМ dNTP и 20 пикомоль каждого праймера и 1,5 Ед ДНК-полимеразы Taq (все реагенты были получены от Bangalore Genei). Цикл ПЦР состоял из денатурации при 94°С в течение 45 секунд, отжиг при 65°C в течение 40 секунд и удлинение при 72°C в течение 45 секунд в 30 циклах. Фрагмент ПЦР размером ~120 п.н. очищали из геля, расщепляли BamHI и лигировали в расщепленный BamHI ген HbsAg в течение ночи с ДНК-лигазой T4 с получением химерной последовательности размером ~810 п.н., обозначенной как SEQ ID NO: 7, в которой фрагмент L1 HPV16 является C-концевым для гена HbsAg. Для получения HPV-HbsAg, где указанный выше фрагмент лигирован с N-концом гена HbsAg, фрагмент L1 HPV16 амплифицировали посредством ПЦР с праймерами со сходной последовательностью, содержавшей участок EcoRI в N-концевом праймере и последовательность BamHI в С-концевом праймере вместо NotI. Фрагмент ПЦР со сходным фрагментом расщепляли BamHI и лигировали в расщепленный BamHI HbsAg, как описано выше, с получением последовательности, обозначенной как SEQ ID NO: 8.
Получение химерной последовательности с областью L1 HPV18 и HbsAg
Получение химерной последовательности белка L1 HPV18 (39 аминокислот; 117 п.н.) со следующей последовательностью PPLELKNTVLEDGDMVDTGYGAMDFSTLQDTKCEVPLDI, лигированной с C-концом последовательности HbsAg посредством ПЦР. Синтетический ген L1 HPV18 подвергали оптимизации кодонов для экспрессии в Pichia pastoris и синтезировали в GenScript Corporation. Последовательность L1 HPV18, указанную выше, подвергали амплификации посредством ПЦР со следующими праймерами для ПЦР:
8PICFNTERM: 5'-GATGGATCCCCTCCTTTGGAATTGAAG-3'
8PICFCTERM: 5'-AGTGCGGCCGCTTATCAGTAATCTGGATATTTGC-3'
Реакцию ПЦР проводили в объеме 50 мкл, содержавшем 1x буфер для ДНК-полимеразы Taq, 0,25 мМ dNTP и 20 пикомоль каждого праймера и 1,5 Ед ДНК-полимеразы Taq. Реакция ПЦР состояла из денатурации при 94°C в течение 45 секунд, отжига 65°C в течение 30 секунд и удлинения при 72°C в течение 40 секунд в 30 циклах. Фрагмент ПЦР размером ~135 п.н. очищали из геля, расщепляли BamHI и лигировали в расщепленный BamHI ген с получением химерной последовательности HbsAg-HPV, где последовательность L1 HPV18 лигирована с C-концом HbsAg для получения SEQ ID NO: 9. Для получения химерной последовательности HPV-HbsAg праймеры для ПЦР содержали участок EcoRI в N-концевом праймере и участок BamHI в C-концевом праймере. После расщепления BamHI, фрагмент лигировали в HbsAg, который содержал BamHI на N'-конце с получением SEQ ID NO: 10. Оба лигированных фрагмента ПЦР расщепляли EcoRI и NotI для клонирования в дрожжевой экспрессирующий вектор pPIC3.5K.
Получение химерной последовательности более крупного фрагмента L1 HPV18 с HbsAg
Химерную последовательность более крупного фрагмента L1 HPV18, состоящего из следующей последовательности:

лигировали с C-концом HbsAg посредством ПЦР. Фрагмент L1 амплифицировали посредством ПЦР с помощью следующих праймеров для ПЦР:
8P1CFLFNTERM:
5'-TCTGGATCCGCTACATCTAATGTCTC-3'
8P1CFLFCTERM:
5'-ATTGCGGCCGCTTATCAGGATGGACTGTAAACG-3'
Реакционная смесь объемом 50 мкл содержала 1x буфер для ДНК-полимеразы Taq, 0,25 мМ dNTP и 20 пикомоль каждого праймера и 1,5 Ед ДНК-полимеразы Taq. Реакция ПЦР состояла из денатурации при 94°C в течение 30 секунд, отжига 64°C в течение 40 секунд и удлинения при 72°C в течение 1 мин в 30 циклах. Фрагмент ПЦР очищали из геля, расщепляли BamHI и лигировали с С-концом расщепленного BamHI HbsAg с получением химерного фрагмента SEQ ID NO: 11. Для лигирования указанного выше фрагмента гена L1 HPV18 с N'-концом последовательности HbsAg, праймеры для ПЦР содержали участок EcoRI в N-концевом праймере и участок BamHI в C-концевом праймере. Расщепленный BamHI фрагмент лигировали в расщепленный BamHI HbsAg с получением химерной последовательности, в которой фрагмент L1 HPV18 является N-концевым для гена HbsAg с получением SEQ ID NO: 12. Обе указанных выше химерных последовательности расщепляли посредством EcoRI и NotI и лигировали с расщепленным EcoRI и NotI дрожжевым экспрессирующим вектором pPIC3.5K для клонирования в Pichia pastoris.
Получение химерных последовательностей антигенов HPV c сигнальной последовательностью гена сывороточного альбумина человека для секреторной экспрессии антигенов HPV в Pichia pastoris
Сигнальную последовательность гена сывороточного альбумина человека из 24 аминокислот амплифицировали посредством ПЦР со следующими праймерами:
HASSNTERM: 5'-CCTGAATTCACCATGAAGTGGGTAACCTTTATTTC-3'
HASSCTERM: 5'-ATTGGATCCTCGACGAAACACACCCCTG-3'
с получением фрагмента размером 72 п.н., который очищали из геля, расщепляли BamHI и лигировали с 5' концом полноразмерных генов L1 HPV16, L2 HPV16 и L1 HPV18, которые содержали участок BamHI на 5'-конце с получением последовательностей SEQ ID NO: 13, SEQ ID NO: 14 и SEQ ID NO: 15, соответственно.
Пример 3: Клонирование и экспрессия в Pichia Pastoris
Все химерные последовательности проверяли секвенированием обеих цепей ДНК в отношении правильности химерной последовательности и рамки считывания перед проведением трансформации дрожжей. Все химерные последовательности, упомянутые в предшествующих разделах, после расщепления EcoRI и NotI очищали из геля и лигировали в расщепленный EcoRI и NotI вектор pPIC3.5K (Invitrogen Corporation, Carlsbad, USA) и лигировали в течение ночи с ДНК-лигазой T4. Смесь для лигирования трансформировали в компетентные клетки E.coli DH5a и высевали на планшет с агаром LB, содержащим 100 мкг/мл ампициллина. Плазмидную ДНК выделяли из трансформированных клонов для каждой из конструкций в крупном масштабе, очищали и расщепляли BglII для линеаризации плазмиды для трансформации дрожжей.
В основном линеаризованную плазмидную ДНК, кодирующую химерные последовательности из с SEQ ID NO: 1 по SEQ ID NO: 15, трансформировали в Pichia Pastoris GS115 согласно протоколам изготовителей (Invitrogen) и как описано в их руководстве. Все химерные последовательности клонировали в локус AOX1 и экспрессировали под промотором AOX1 посредством индукции метанолом. Клонирование, скрининг, выделение рекомбинантных штаммов Pichia и индукцию клонированного гена метанолом проводили согласно руководству пользователя "A Manual of Methods for Expression of Recombinant Proteins in Pichia pastoris" Version M Jan 2002, набора для экспрессии в Pichia, каталожный # K1710-01, Invitrogen Corporation, Carlsbad, USA).
Пример 4: очистка химерных антигенов HbsAg-HPV
Клетки после инкубации с метанолом собирали и лизировали обработкой ультразвуком и/или с помощью стеклянных гранул. Лизат обрабатывали PEG6000 в концентрации 7,5% и NaCl в концентрации вплоть до 3%. Суспензию клеток центрифугировали при 8000 × g в течение 10 мин. Супернатант преципитировали обработкой солью (запатентованная смесь) и преципитировали. Белок элюировали из преципитата с солью буфером Tris-HCl, pH 8,5. Элюат концентрировали и подвергали диафильтрации 50 мМ Tris-HCl, pH 8,5 и наносили на катионную колонку, уравновешенную тем же буфером. Белок элюировали NaCl в различной концентрации для различных антигенных белков. Чистоту препарата проверяли на 12% SDS-PAGE с окрашиванием серебром. Затем точный молекулярный размер различных химерных белков подтверждали вестерн-блоттингом.
Пример 5: ELISA химерных белков HPV-HbsAg
Экспрессию химерных белков выявляли посредством ELISA для определения содержания малого антигена гепатита B с помощью набора MONOLISA Anti-Hbs3.0 (продукт # 72400; Bio-Rad) в точности согласно протоколу, описанному в руководстве набора, и выражали в нг/мл. 100 мкл клеточного лизата после индукции использовали для оценки содержания HbsAg. Оценку антигенов HPV в химерных белках в клеточном лизате после индукции также проводили посредством ELISA. 1 мкг очищенных белков в 200 мкл наносили на 96-луночный микропланшет для титрования в присутствии буфера для нанесения (карбонатный буфер, pH 9,6) и инкубировали в течение ночи при 4°C. Содержание лунок удаляли, а затем планшетом постукивали по бумажному полотенцу для удаления остаточного буфера в лунках. Лунки блокировали 225 мкл/лунка блокирующего раствора (1 x PBST, содержащий 0,5% BSA) и инкубировали при 37°C в течение 1 часа.
Лунки промывали три раза буфером для промывания (200 мкл/лунка, 30 секунд на промывание). После каждого промывания планшеты постукивали до сухого состояния по бумажному полотенцу для удаления из лунок остаточного буфера. Лунки промывали в 1x PBST (фосфатный буфер, содержащий 0,5% tween 20) пять раз для удаления не связавшихся белков. Антитело к полноразмерному L1 HPV6, L2 HPV16 и L1 HPV18, индуцированное у кролика, использовали в качестве первичного антитела. После добавления первичного антитела в разведении 1:1000, планшеты инкубировали при комнатной температуре в течение одного часа при встряхивании. Содержимое лунок удаляли и промывали пять раз буфером для промывания, содержавшим 1x PBST. Добавляли конъюгат вторичного антитела против IgG кролика с HRP в разведении 1:2000 в 10 мМ фосфатно-солевом буфере и инкубировали при комнатной температуре в течение одного часа при встряхивании. Содержимое лунок удаляли и планшеты постукивали по сухому бумажному полотенцу для удаления остаточного буфера из лунок. 200 мкл свежеполученного раствора для субстрата OPD (дигидрохлорид ортофенилендиамина) и пероксида водорода добавляли на лунку и инкубировали при комнатной температуре в течение 30 мин. Реакцию останавливали добавлением 75 мкл буфера для остановки и поглощение при 490 нм считывали в устройстве для считывания планшетов ELISA.
Пример 6: вестерн-блоттинг
Химерные белки, как описано в предшествующих разделах, разделяли на 12% геле для SDS-PAGE, подвергали электроблоттингу на мембране PVDF (Hybond-P, GE Healthcare) и блокировали в растворе, содержавшем 10 мМ фосфатный буфер, pH 7,4, содержавший 3% порошок обезжиренного молока. После блокирования в течение ночи блокирующим раствором, блокирующий раствор удаляли, и проводили промывание 3×5 минут в PBST. Блот инкубировали при 37°C с первичной антисывороткой кролика в разведении 1:500, индуцированной коммерчески доступной четырехвалентной вакциной. После инкубации с первичным антителом в течение одного часа, блот промывали пять раз и инкубировали с вторичным антителом в разведении 1:2000 в PBST, т.е. конъюгатом IgG против антител кролика с HRPO, и инкубировали при RT в течение одного часа при встряхивании. Добавляли субстрат диаминобензиден (DAB) и пероксид водорода и цвет проявляли до достижения требуемой интенсивности цвета. Реакцию останавливали удалением раствора субстрата и промыванием WFI.
Вестерн-блоттинг выявил в химерных последовательностях L1 HPV16 и L1 HPV18 с помощью антисыворотки, индуцированной против коммерчески доступной четырехвалентной вакцины против HPV.
Пример 7: Иммуногенность химерных белков
10 мышей BALB/C (15-20 г) в каждой тестируемой группе иммунизировали 50 мкг очищенного белка каждого из химерных антигенов, заключенных в геле из гидроксида алюминия, и вводили внутримышечно. Вспомогательную дозу давали через 21 сутки после первой дозы 1, и вторую дозу вводили через 45 суток после 1 дозы. Титр антител измеряли посредством непрямого ELISA для всех клонов, содержавших SEQ ID NO: 3-12, как описано в предыдущем разделе. Титр антител оценивали как для антигена HbsAg, так и для антигена HPV, у мышей, иммунизированных химерными антигенами. Первичное антитело для оценки L1 HPV16 и L1 HPV18 представляло собой антисыворотку кролика, индуцированную против коммерчески доступной четырехвалентной вакцины, за исключением клонов, экспрессирующих белок L2 HPV16, который измеряли с помощью антисыворотки кролика против L2 HPV16, индуцированной против очищенного белка L2 HPV16. Титр антител против антигенов HPV измеряли посредством непрямого ELISA в объединенной сыворотке от каждой группы мышей. Вторичное антитело представляло собой конъюгат антител кролика против IgG с HRPO. Эксперимент повторяли, где эквивалентные количества антигенов в алгеле вводили с 50 мкг олигонуклеотидов CpG на дозу. Титр антител у всех мышей оценивали на 60-65 сутки после введения первоначальной дозы антигена. Для оценки титра антител против HbsAg у иммунизированных мышей, оценку проводили с помощью набора Monolisa Anti HBs 3.0 Kit от Bio-Rad laboratories, в точности согласно инструкциям изготовителя. Значения регистрировали при 490 нм и наносили на график.


| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ПАПИЛЛОМАВИРУСНОЙ ИНФЕКЦИИ ЧЕЛОВЕКА И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2014 |
|
RU2546243C1 |
| Способ получения рекомбинантной вакцины для профилактики папилломавирусной инфекции человека, рекомбинантная вакцина | 2018 |
|
RU2681174C1 |
| ПАПИЛЛОМАВИРУСНЫЕ ВАКЦИНЫ | 1995 |
|
RU2206608C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ PICHIA ANGUSTA - ПРОДУЦЕНТ КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 18 | 2011 |
|
RU2445358C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ PICHIA ANGUSTA - ПРОДУЦЕНТ КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 16 | 2011 |
|
RU2445357C1 |
| ОПТИМИЗИРОВАННАЯ ЭКСПРЕССИЯ L1 HPV45 В ДРОЖЖАХ | 2004 |
|
RU2360001C2 |
| ОПТИМИЗИРОВАННАЯ ЭКСПРЕССИЯ HPV 58 L1 В ДРОЖЖАХ | 2004 |
|
RU2370538C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ ГЛАВНОГО КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 18 | 2014 |
|
RU2546242C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ ГЛАВНОГО КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 16 | 2014 |
|
RU2546241C1 |
| МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, КОДИРУЮЩАЯ HPV31 L1, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР, КЛЕТКА-ХОЗЯИН, ВИРУСОПОДОБНАЯ ЧАСТИЦА И СПОСОБ ЕЕ ПОЛУЧЕНИЯ, ВАКЦИНА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И СПОСОБЫ С ИХ ИСПОЛЬЗОВАНИЕМ | 2004 |
|
RU2356943C2 |











Изобретение относится к области медицины, а именно к биофармацевтики, и может быть использовано для приготовления химерной вакцины для профилактики папилломавируса человека (HPV) и инфекции, вызываемой гепатитом B у человека. Для этого вакцина содержит антигенную композицию, которая состоит из химерных слитых молекул полипептидов или эпитопов капсидных белков-антигенов L1 и L2 HPV с поверхностным антигеном гепатита B (HbsAg), как раскрыто в SEQ ID NO:3 по SEQ ID NO:13. Разработка химерной вакцины позволяет увеличить плотность L1 или L2 нейтрализующих эпитопов на поверхности вирусоподобной частицы HbsAg, при этом химерная слитая конструкция экспрессируется в виде вирусоподобной частицы, обеспечивая защиту от любого известного серотипа HPV и от гепатита В посредством введения одного белка. 9 з.п. ф-лы, 7 пр., 11 ил.
1. Вакцинная композиция для профилактики и лечения любого известного серотипа папилломавируса человека (HPV) и инфекции, вызываемой вирусом гепатита B у человека, содержащая антигенную композицию, которая состоит из химерных слитых молекул полипептидов или эпитопов капсидных белков-антигенов L1 и L2 HPV с поверхностным антигеном гепатита В (HbsAg), как раскрыто в SEQ ID NO:3 по SEQ ID NO:13.
2. Вакцинная композиция по п.1, где антигены HPV содержат полипептиды или эпитопы ранних белков HPV, таких как E2, E5, E6 и E7, либо отдельно, либо в комбинациях, таких как раскрыты в SEQ ID NO:1 и 2.
3. Вакцинная композиция по п.1, где антигенная композиция содержит полные или неполные последовательности или мутанты или варианты антигенов HPV, выбранных из антигенов L1, L2, E2, E5, E6 и E7, или специфичные T-клеточные или B-клеточные эпитопы указанных антигенов, либо в качестве отдельной последовательности, либо в тандемных повторах в химерных слитых молекулах с поверхностным антигеном гепатита В, способные индуцировать достаточный титр антител, требующийся для профилактики инфекции, вызываемой любым известным серотипом папилломавируса человека (HPV) и инфекции, вызываемой вирусом гепатита В, при введении человеку.
4. Вакцинная композиция по п.1, где антигены HPV выбраны из группы, включающей HPV16, HPV18, HPV6, HPV11, HPV31, HPV33, HPV35, HPV39, HPV45, HPV51, HPV52, HPV56, HPV58, HPV59, HPV68.
5. Вакцинная композиция по п.1, где антигены HPV подвергнуты слиянию с N-концом или с C-концом или с любой областью в открытой рамке считывания поверхностного антигена гепатита B с добавлением линкерной последовательности между двумя последовательностями, или без ее добавления.
6. Вакцинная композиция по п.1, где химерные слитые молекулы полипептидов или эпитопов капсидных белков-антигенов L1 и L2 HPV с поверхностным антигеном гепатита В (HbsAg), как раскрыто в SEQ ID NO:3 по SEQ ID NO:13, получены посредством рекомбинантной экспрессии в хозяине, включающей следующие этапы: культивирование клетки-хозяина, трансформированной химерными слитыми молекулами антигенной композиции по п.1, с применением вектора pPIC3.5K, выделение и очистку из них рекомбинантных белков.
7. Вакцинная композиция по п.6, где рекомбинантная экспрессия указанных выше антигенов HPV осуществляется либо в качестве внутриклеточных белков, либо в качестве секреторных белков в хозяине, причем хозяин представляет собой дрожжевую экспрессирующую систему Pichia pastoris.
8. Вакцинная композиция, содержащая антигенную композицию по любому из пп.1-4, дополнительно содержащая фармацевтически приемлемый носитель.
9. Вакцинная композиция по п.1, где антигены используются в диапазоне 10-1000 мкг белка на дозу вакцинной композиции.
10. Вакцинная композиция по п.8 или 9, дополнительно содержащая пригодный адъювант, причем адъювант выбран по меньшей мере из одного из следующих: гидроксид алюминия, фосфат алюминия, монофосфориллипид A, содержащие CpG и не содержащие CpG олигонуклеотиды, цитокины.
| TINDLE RW et al | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| Приспособление к основовязальной машине для ее выключения при обрыве нити | 1958 |
|
SU117550A1 |
| SCHODEL F | |||
| Hybrid hepatitis В virus core antigen as a vaccine carrier moiety: I. | |||

