Изобретение касается использования ингибиторов пролиферации стволовой клетки для регулирования цикла стволовой клетки при лечении человека и животных, подверженных аутоиммунным заболеваниям, старению, раку, миелоэзу, прелейкозу, лейкозу, псориазу или другим болезням, включающим в себя условия гиперпролиферации. Настоящее изобретение также касается способа лечения человека и животных, имеющим симптомы или подвергающихся воздействию химиотерапевтических факторов, прочих факторов, пагубно воздействующих на цикличность стволовой клетки, или облученных радиацией. И, наконец, настоящее изобретение касается улучшения поддержания жизнедеятельности стволовой клетки или культур роста для процесса авто- и алло- трансплантации или переноса генов.
Большинство клеток на терминальной стадии дифференцировки в обновляющихся системах имеют короткий срок жизни и непрерывно заменяться на протяжении жизни человека. Например, клетки крови создаются в самообновляющейся популяции мультипотентных кроветворных стволовых клеток (КСК). Кроветворные стволовые клетки являются подпопуляцией кроветворных клеток. Кроветворные клетки могут быть получены, например, из костного мозга, умбиликальной пуповинной крови или периферической крови (не активированной или активированной таким фактором, как фактор роста - ликвор КСФ-Г; кроветворные клетки включают популяцию стволовой клетки, клетки-предшественника, дифференцированные клетки, придаточные клетки, клетки стромы и прочие клетки, которые вносят вклад в окружающую среду, необходимый для производства зрелых клеток в крови. Вследствие того, что кроветворные стволовые клетки необходимы для развития всех зрелых клеток кроветворной и иммунной систем, их выживаемость чрезвычайно важна для восстановления иммунной системы организма в полном объеме в людях, лечение которых производится с помощью химиотерапии или других агентов.
Воспроизводство кроветворных клеток регулируется серией факторов, которые стимулируют рост и дифференциацию кроветворных клеток, некоторые из которых, например, эритропоэтин и фактор роста - ликвер, сейчас используются в клинической практике. Одной из частей сети управления, которая однако не была широко рассмотрена, является механизм обратной связи, который образует отрицательный рычаг процесса регулирования (Ив и др. Кровь 78: 110-117, 1991).
Ранние исследования, проведенные Лордом и его коллегами, отразили существование фактора растворимого белка в обычных экстрактах костного мозга мышей и свиней, способного обратимо ингибировать цикл КСК (Лорд и др., Бр. Дж. Хаем. 34: 441-446, 1976). Эта ингибирующая деятельность (молекулярный вес 50-100 кД) была вызвана ингибитором стволовой клетки (ИСК).
Очистка этого фактора от первоначальных источников не была выполнена по причине трудностей, связанных с анализом in vivo, требующем большого количества облученных мышей. Пытаясь преодолеть эти проблемы, Прогнелл со своими коллегами разработал анализ с in vitro для примитивных кроветворных клеток (КОЕ-А, колониеобразующая единица) и протестировали клеточные, принятые в качестве источника ингибирующей деятельности ( см. Грэхэм и др. Природа 314: 442-444, 1990).
Как и в ранних исследованиях в качестве возможных источников для ИСК были выявлены макрофаги (Лорд и др. Клетки крови 6:581-593,1980), для исследования отобрали клеточную линию макрофагов мыши, J774.2 (Грэхэм и др. Природа 344: 442-444,1990). Для очистки Грэхэм и др. использовали обусловленную среду из этой клеточной линии, ингибирующий пептид был изолирован, что оказалось идентичным описанному выше возбуждающему белку макрофага цитокина 1 - альфа (ВБМ -1 альфа). Таким образом, ВБМ- 1 альфа был изолирован из клеточной линии, а не из первичного вещества. В то время, как Грэхэм и др. наблюдали за тем, как действие антитела к ВБМ - 1 альфа отменило действие необработанного экстракта костного мозга, другие сотрудники обратили внимание на важность другой ингибирующей деятельности. Например, Грэхэм и др. (Дж. Експ. Мед. 178: 925-32, 1993) выразили мысль, что главным ингибитором кроветворных стволовых клеток является TGF бетта, а не ВБМ-1 альфа. Далее, Ив и др. (PNAS 90:12015-19,1993) предложили, что как ВБМ-1 альфа, так и TGF бетта присутствуют на субоптимальных уровнях в обычном костном мозге, и ингибирование требует синергии (совместного действия) двух факторов.
Другие сотрудники описали дополнительные ингибирующие факторы стволовой клетки. Фриндель со своими коллегами изолировали тетраптид из эмбрионального экстракта костного мозга теленка и экстракта печени, в которой присутствует ингибирующая деятельность стволовой клетки (Lenfant и др. PNAS 86: 779-782, 1989). Paukovits и др. (Cancer res. 50: 328-332, 1990) дали описание пентапептида, который в своей изометрической форме является ингибитором, а в своей тетрагональной форме является стимулятором циклической деятельности стволовой клетки. В различных системах in vitro были признаны и другие факторы (см. Wright and Pragnell в Bailliere's Clinical Haematology, том 5, стр. 723-39, 1992(Bailliere's Tinadall)).
Цырлова и др. СССР 1561261 А1 рассмотрела процесс очистки для ингибитора пролифирации стволовой клетки.
Сегодня ни один из этих факторов не утвержден для использования в клинических целях. Однако необходимость в эффективных ингибиторах стволовой клетки существует. Главным источником токсичности, возникающей при химиотерапии и лучевой терапии, является разрушение обычных пролиферативных клеток, что может привести к супрессии костного мозга или к желудочно-кишечной токсичности. Эффективный ингибитор стволовой клетки предохранит эти клетки и позволит оптимизировать эти терапевтические режимы. Таким образом, при наличии обоснованной необходимости в разнообразных стимулирующих цитогенах ( таких как IL-1, IL-2, IL-3. IL-4, IL-5, IL-6, IL-7, IL-9, IL-11, IL-13, IL-14, IL-15, КОЕ-Г, КОЕ-ГМ, эритропоэтин, тромбопоэтин, фактор стволовой клетки, лигандов flk 2/flt 2 и т. д., которые стимулируют циркуляцию кроветворных клеток) в зависимости от клинической ситуации, таким образом, возможна ситуация, при которой видна необходимость в разнообразных ингибирующих факторах.
Гемоглобин представляет собой высоко консервированный тетрамерный белок с молекулярным весом 64000 Дальтон. Он состоит из двух альфа- и двух бета-цепей. Каждая цепь связывает одну молекулу гема (железопорфирина IX), железосодержащая протезная группа. Вертебральные альфа- и бета-цепи возможно были получены из одного наследственного гена, который удвоился и затем распространился; две цепи сохранили большую степень последовательной схожести, как между собой, так и между различными позвонками (см. рис.16 А). В человеческом организме скопление альфа-цепи на хромосоме 16 содержит два альфа-гена (альфа 1 и альфа 2), которые имеют следующий код для идентичных полипептидов, а также генов, кодирующих другие цепи, подобные альфа: зета, тета и несколько не транскрибированных псевдогенов (см. рис.16 В для кДНК и аминокислотной последовательности альфа-цепи человеческого организма). Скопление бета-цепи на хромосоме 11 состоит из одиночного гена бета-цепи и нескольких генов, похожих на бета: дельта, ипсилон, G-гамма и А-гамма, а также, по крайней мере, два невыраженных псевдогена (см. рис.16С для ДНК и аминокислотной последовательности бета-цепи человеческого организма). Экспрессия этих генов варьируется в период роста. В гемопоэзе человеческого организма, который был широко исследован, эмбриональные эритробласты успешно синтезировали тетрамеры двух зета- и двух ипсилон-цепей (Gower 1), двух альфа- и двух ипсилон-цепей (Gower II) или двух зета- и двух гамма-цепей (Hb Portland). В процессе развития эмбриогенеза преобладающая форма состоит из эмбрионального гемоглобина (Hb F), который состоит из двух альфа- и двух гамма-цепей. Гемоглобин зрелого организма (две альфа- и две бета-цепи) подвергается синтезированию во внутриутробный период; при рождении он составляет приблизительно 50% гемоглобина, а завершается переход в течение 6 месяцев. Значительная часть гемоглобина (приблизительно 97%) в зрелом организме наблюдается в виде следующих разновидностей: две альфа- и две бета-цепи (Hb А) с небольшим количеством Hb F или дельта-цепи (Hb А2).
Гем был широко изучен с точки зрения влияния на кроветворность (см. S. Sassa, Seminars Hemat. 25: 312-20, 1988 и N. Abraham и др., Int. J. Cell Cloning 9: 185-210, 1991 для анализа). Гем необходим для созревания эритробластов: in vitro, гемин Хлорожелезопорфирин IX , т.е. гем с дополнительным ионом хлора увеличивает пролиферацию КОЕ-ГЕММ, БОЕ-Е и КОЕ-Е. Таким же образом гемин увеличивает объем клеточного содержимого в долговременных культурах костного мозга.
1. Химиотерапия и лучевая терапия рака.
Произведенное исследование факторов, стимулирующих рост, явилось причиной клинического использования этих факторов (эритропоэтин, КОЕ-Г, КОЕ-ГМ и т.д.). Эти факторы снизили смертность и заболеваемость, связанные с химиотерапией и лучевой терапией. Дальнейшие клинические выгоды для пациентов, которые проходят курс химио- и лучевой терапии, могут быть обнаружены путем использования альтернативной стратегии блокировки доступа стволовой клетки в цикл клетки, таким образом предохраняя их от постороннего токсического воздействия.
II. Трансплантация костного мозга.
Трансплантация костного мозга (ТКМ) является хорошим средством для лечения разнообразных гематологических, аутоиммунных и злокачественных болезней; современные виды лечения включают кроветворные клетки, полученные в умбиликальной пуповинной крови или из периферической крови (немобилизованной или мобилизованной такими агентами как КОЕ-Г), а также из костного мозга.
Для расширения примитивной стволовой клетки до популяции, пригодной для трансплантации, в настоящее время используется обработка кроветворных клеток методом ex vivo. Оптимизация этой процедуры требует: (1) достаточного количества стволовых клеток, способных поддерживать долговременное воспроизведение гемопоэза; (2) снижение трансплантата по отношению к Т-лимфоциту и (3) отсутствие остаточных злокачественных клеток. Эта процедура может быть оптимизирована, если для расширения методом ех vivo включаются ингибиторы стволовой клетки.
Эффективность очищения кроветворных клеток цитотоксичными лекарственными средствами с целью уничтожения остаточных злокачественных клеток ограничено вследствие токсичности этих составов для нормальных кроветворных клеток, и особенно стволовых клеток. Необходимо наличие эффективной защиты нормальных клеток во время процесса очищения; защита может быть обеспечена путем извлечения стволовых клеток из цикла посредством эффективного ингибитора.
III. Сбор периферической стволовой клетки.
Периферические стволовые клетки крови (ПСКК) обеспечивают несколько потенциальных преимуществ по сравнению с костным мозгом при аутогенной трансплантации. Пациенты, которые не имеют удобных участков для сбора костного мозга вследствие вмешательства опухоли или предшествующей лучевой терапии, могут однако быть объектами сбора ПСКК. При использовании стволовых клеток крови отпадает необходимость общей анестезии и хирургического вмешательства для пациентов, которые плохо их переносят. Технология афереза, необходимая для сбора клеток крови, является эффективной и имеется в наличии большинства медицинских центров. Крупнейшими ограничениями метода являются как низкая частота стационарного состояния стволовых клеток в периферической крови и их высокий статус цикла после мобилизационного процесса с использованием лекарственных средств или факторов роста (т.е. циклофосфамид, КОЕ-Г, фактор стволовой клетки). Эффективный игибитор стволовой клетки был бы полезен для возвращения таких клеток в неподвижное состояние, таким образом предотвращая их потерю посредством дифференцировки.
IV. Лечение гиперпролиферационных нарушений.
Некоторые болезни характеризуются гиперпролиферационным состоянием, при котором потерявшие управление стволовые клетки стимулируют перепроизводство клеток на терминальной стадии дифференцировки. Такие состояния болезни включают без ограничений псориаз, при котором происходит перепроизводство эпидермальных клеток, предраковые состояния в желудочно-кишечном тракте, характеризующееся появлением кишечных полипов. Ингибитор стволовой клетки был бы полезен в лечении таких состояний.
V. Перенос генов.
Способность переноса генетической информации в кроветворные клетки применяется в данный момент в клинических целях. Кроветворные клетки являются полезным объектом генной терапии вследствие легкости доступа, обширного опыта обработки и лечения этой ткани ex vivo, и по причине способности клеток крови проникать через ткани. Более того, исправление некоторых человеческих дефектов могло бы быть возможным путем внедрения функционального гена в примитивные стволовые клетки кроветворной системы человека.
Для внедрения генов в кроветворные клетки человека с использованием либо инфекции ретро вируса, либо физических методов переноса генов:
(1) низкая частота стволовых клеток в кроветворных тканях потребовала разработки высокоэффективных способов переноса генов; (2) стволовые клетки, имеющие более быструю цикличность оказались более восприимчивы к инфекции переносчика, однако повышение частоты инфекции путем стимулирования пролиферации стволовой клетки факторами роста оказывает отрицательное воздействие на длительную экспрессию гена, так как клетки, содержащие гены-трансплантаты, принудительно необратимо дифференцированы и потеряли способность к самообновлению. Эти проблемы могут быть решены с помощью ингибитора стволовой клетки для того, чтобы предотвратить дифференцировку и потерю способности к самообновлению.
Настоящее изобретение касается полипептидов, которые являются ингибиторами пролиферации стволовой клетки (INPROL), и их использования.
Настоящее изобретение включает ингибитор пролиферации стволовой клетки, характеризующийся следующими свойствами:
(а) специфическая активность (IC50) менее или аналогична 20 нг/мл в анализе селезенки мышей (КОЕ-С) на колониеобразование (см. пример 4),
(б) молекулярный вес более 10000 и менее 100000 Дальтон (путем ультрафильтрации),
(в) активность чувствительна к деградации при использовании ципсина,
(г) более гидрофобен чем MIP- альфа или бета и может быть отделен от последних путем обращенной хроматографии (см. пример 12),
(д) биологическая активность сохраняется после нагревания в течение часа при температуре 37, 55 или 75oС в водном растворе и
(е) биологическая активность сохраняется при осаждении с добавлением 1% соляной кислоты в ацетон.
Далее настоящее изобретение характеризуется и отличается от прочих возможных ингибиторов стволовой клетки (например, MIP-1 альфа, TGF бета и различных олигопептидов) своей способностью достигать ингибирования при анализе in vitro после короткого преинкубационного периода ( см. пример 5).
Настоящее изобретение охватывает также фармацевтические составы, содержащие INPROL для лечения различных нарушений.
Настоящее изобретение охватывает способ лечения пациента, подвергающегося воздействию агента, способного уничтожить или повредить стволовые клетки путем введения этому пациенту действенной дозы ингибирующего состава стволовой клетки. Стволовые клетки, сохраненные таким способом, могут представлять собой кроветворные стволовые клетки, обычно содержащиеся и делящиеся в костном мозге. С другой стороны, стволовые клетки могут быть эпителиальными, расположенными, например, в кишках или на скальпе или в других частях тела, или терминальными, расположенными в органах размножения. Способом настоящего изобретения желательно пользоваться при лечении человека, однако, он также охватывает лечение животных. Термины "субъект" или "пациент" относятся к животным, таким как млекопитающие, включая человека.
С другой стороны, изобретение охватывает способ защиты и восстановления кроветворных, иммунных и прочих систем стволовых клеток пациента, подвергнутого химиотерапии, и предусматривает введение пациенту эффективной дозы INPROL.
Кроме того, настоящее изобретение включает способ вспомогательного лечения любых раковых заболеваний, включая те из них, которые характеризуются сплошной опухолью, путем введения пациенту, больному раком, эффективной дозы INPROL для защиты стволовых клеток костного мозга, желудочно-кишечного тракта или других органов от токсического воздействия химиотерапии и лучевой терапии.
Еще одной областью применения настоящего изобретения является лечение лейкоза, которое включает лечение кроветворных клеток, в состав которых входят пролиферативные лейкозные клетки, путем введения эффективной дозы INPROL для ингибирования пролиферации нормальных стволовых клеток, а также лечению костного мозга с помощью цитотоксического агента для уничтожения лейкозных клеток.
Можно усилить действие этого способа последующим лечением костного мозга другими агентами, стимулирующими его пролиферацию: то есть факторами, которые стимулируют колониеобразование. В сочетании эти виды лечения могут осуществляться in vivo. С другой стороны, этот способ также полезен для очищения ex vivo и экспансии кроветворных клеток для трансплантации.
Кроме того, этот способ включает лечение субъекта, имеющего любое нарушение, причиненное пролиферирующими стволовыми клетками. Такие нарушения как псориаз, миелодисплазия, некоторые аутоиммунные заболевания, иммунодепрессия при старении лечатся введением эффективной дозы INPROL для частичного ингибирования пролиферации сомнительной стволовой клетки.
Настоящее изобретение включает способ для обратной защиты стволовых клеток от повреждений, нанесенных цитотоксическим агентом, способным уничтожить или повредить стволовые клетки. Способ включает введение субъекту, которому надлежит быть объектом воздействия таким агентом, эффективной дозы INPROL.
Настоящее изобретение также включает:
Ингибитор пролиферации стволовой клетки, выделенной от костного мозга свиньи или другого животного путем следующей процедуры (см. пример 12):
(а) извлечение костного мозга и удаление частиц веществ посредством фильтрации;
(б) обработка нагреванием при 50oС в течение 40 минут с последующим охлаждением в ледяной бане;
(в) удаление осадка путем обработки в центрифуге при 10000 g в течение 30 минут при 4oС;
(г) кислотное осаждение посредством добавления супернатанта к 10 объемам перемешанного ацетона пониженной температуры, содержащего 1% по объему концентрированной соляной кислоты при инкубации при 4oС в течение 16 часов;
(д) выделение осадка путем обработки в центрифуге при 20000 g в течение 30 минут при 4oС и промывке холодным ацетоном после высушивания;
(е) выделение путем обращенной хромотографии и отслеживания активности путем ингибирования процесса колониеобразования клетками костного мозга, в котором были предварительно введены 5-флуороурацил и выдержанного в присутствии IL-3 мыши, а также путем абсорбции при 280 нг и SDS-PAGE.
Настоящее изобретение также включает:
Способы для очистки ингибитора пролиферации стволовой клетки, которые в значительной степени освобождены от прочих веществ, напоминающих белки, составляющие пункты, изложенные выше. Более подробно об этом изложено ниже.
Настоящее изобретение также включает:
Способ лечения людей и животных, при котором ингибитор пролиферации стволовой клетки содействует ослаблению иммунодепрессии, возникающей вследствие гиперпролиферации стволовой клетки.
Настоящее изобретение также включает:
Способ лечения людей и животных, при этом упомянутый ингибитор пролиферации стволовой клетки вводят после того, как стволовые клетки индуцированы для пролиферации путем воздействия цитотоксического лекарственного средства или облучения. Стволовые клетки обычно неактивны, однако их стимулируют внедряться в цикл клетки после химиотерапии. Это делает их более чувствительными ко второму приему химиотерапии; данный метод предохраняет их от такого лечения.
Настоящее изобретение также включает:
Способ лечения человека и животных, при этом упомянутый ингибитор пролиферации стволовой клетки вводится в качестве адъюванта перед или совместно с вакцинацией с целью улучшения иммунной реакции.
Настоящее изобретение также включает:
Способ лечения человека и животных, принимающих цитотоксическое лекарственное средство или подвергающихся лучевой терапии. Этот способ представляет собой введение эффективной дозы ингибитора пролиферации стволовой клетки для защиты стволовой клетки от повреждений.
Настоящее изобретение также включает фармацевтический состав, в который входят (а) полипептид, отобранный из группы, состоящей из альфа-цепи гемоглобина, бета-цепи гемоглобина, гамма-цепи гемоглобина, дельта-цепи гемоглобина, ипсилон-цепи гемоглобина и зета-цепи гемоглобина, и (б) фармацевтически приемлемый носитель. Такие фармацевтические составы могут состоять из одиночного полипептида, отобранного из упомянутой группы, смеси полипептидов, отобранных из упомянутой группы в форме димеров и мультимеров, с гемом или без него.
Настоящее изобретение также включает пептиды, имеющие следующую последовательность:
Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val,
Cys-Phe-Pro-His-Phe-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val-Cys
там, где две группы Cys образуют дисульфидную связь,
Cys-Phe-Pro-His-Phe-Phe-Asp-Leu-Ser- His-Gly-Ser-Ala-Gln-Val-Cys
там, где две группы Cys объединены углеродным мостиком, и
Asp-Ala-Leu-Thr-Asn-Ala-Val-Ala-His-Val-Asp-Asp-Met-Pro-Asn-Ala-Leu-Ser-Ala.
Настоящее изобретение также включает последовательности ДНК, кодирующие идентифицированные выше пептиды, инфекции, содержащие упомянутые последовательности ДНК и клетки-хозяева, содержащие упомянутые инфекции. Эти пептиды могут быть синтезированы с помощью стандартных химических приемов (например, синтез фазы твердого тела) или с помощью приема рекомбинации (например, системы синтеза, в которых применяется глютатион-S-трансфераз (D.B. Smith and K.S. Johnson, Gene 67:31-40, 1988), тиоредоксин (La Vallie и др., Biotechnology 11: 187-193, 1993) или убихитин (Butt и др., PNAS 86:2540-4, 1989; US Patent 5391490).
Настоящее изобретение довольно включает способ ингибирования пролиферации стволовой клетки, который предусматривает контактирование кроветворных клеток с составом, способным связывать рецепторы опиата, предпочтительно субкатегории мю рецепторов опиата. Пептиды, называемые геморфины, были выделены из гемоглобина, который демонстрировал активность, характерную для опиата (например, Branti и др., Eur. J. Pharm. 125: 309-10, 1986; Zhao и др. , Ann. N.Y. Acad. Sci 750:452-8,1995). На каждую из этих статей в изобретении имеется ссылка.
Изобретение также включает способ ингибирования пролиферации стволовой клетки, который предусматривает контактирование кроветворных клеток с пептидом, отобранным из группы пептидов геморфина, имеющих следующую последовательность:
Leu-Val-Val-Tyr-Pro-Trp-Thr-Gln-Arg- Phe,
Leu-Val-Val-Tyr-Pro-Trp-Thr-Gln-Arg,
Leu-Val-Val-Tyr-Pro-Trp-Thr-Gln,
Leu-Val-Val-Tyr-Pro-Trp-Thr,
Leu-Val-Val-Tyr-Pro-Trp,
Leu-Val-Val-Tyr-Pro,
Val-Val-Tyr-Pro-Trp-Thr-GIn,
Tyr-Pro-Trp-Thr-Gln-Arg-Phe,
Туr-Рrо-Тrp-Thr-Gln-Arg,
Tyr-Pro-Trp-Thr-GIn,
Туr-Рrо-Тrp-Thr.
Вышеупомянутые пептиды имеют последовательность, схожую с другими пептидами, подобными семейству Tyr-MIF-1 (см. Reed и др., Neurosci. Biobehav. Rev. 18:519-25, 1994 для обзора), казоморфины, полученные из казеина (Brantl и др. Норре-Seyler's Z. Physiol. Chem. 360:1211-16, 1979; Loukas и др. Biochem. 22: 4567-4573, 1983; Fiat and Jolles, Mol. Cell. Biochem.. 87:5-30, 1989), пептиды, полученные из цитохрома b, называемые цитохрофины (Brantl и др. Eur. J. Pharm. 111:293-4, 1985), а также пептиды, полученные из комбинаторных библиотек, отсортированные для связи с рецепторами опиата (см. Dooley и др., Ptptide Research 8:124-137, 1995 для обзора). На каждую из этих статей в изобретении имеется ссылка.
Изобретение также включает способ ингибирования пролиферации стволовой клетки, который предусматривает контактирование кроветворных клеток с пептидом, отобранным из группы, состоящей из пептидов, родственных Tyr-MIF-1, казоморфинов и цитохрофинов. Особо следует отметить пептиды Tyr-MIF-1, имеющие следующую последовательность:
Tyr-Pro-Try-Gly-NH2
Tyr-Pro-Lys-Gly-NH2
Tyr-Pro-Leu-Gly-NH2 и
Pro-Leu-Gly-NH2
Изобретение также включает способ проведения генотерапии млекопитающего, который состоит из:
а) удаления упомянутых кроветворных клеток из упомянутого млекопитающего,
б) дополнительного лечения упомянутых кроветворных клеток ex vivo используя, по крайней мере, один стимулирующий цитокин для индуцирования пролиферации стволовой клетки.
в) трансфекции упомянутых кроветворных клеток ех vivo заданным геном,
г) контактирования упомянутой зараженной кроветворной клетки ех vivo с INPROL,
д) трансплантации в млекопитающее кроветворной клетки, упомянутой в этапе (г),
е) дополнительного лечения упомянутого млекопитающего in vivo посредством INPROL.
Изобретение также включает способ проведения экспансии ех vivo стволовой клетки, предусматривающий лечение упомянутых стволовых клеток с помощью INPROL и, по крайней мере, одним стимулирующим цитокином. INPROL контактирует с кроветворными клетками до, в течение и после контакта со стимулирующим цитокином.
Изобретение также включает фармацевтическую структуру, в состав которой входят (a) INPROL и (б) по крайней мере, один ингибирующий состав, отобранный из группы, состоящей из MIP-1 альфа, TGF бета, TNF альфа, INF альфа, INF бета, INF гамма, пентапептид пиро Glu-Glu-Asp-Cys-Lys, тетрапептид N-Acetyl-Ser-Asp-Lys-Pro и трипептид глютатион (Glu-Cys-гамма Glu).
Изобретение также включает фармацевтическую структуру, в состав которой входят (a) INPROL и (б) по крайней мере, один стимулирующий состав, отобранный из группы, состоящей из IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-9, IL-11, IL-13, IL-14, IL-15, КОЕ-Г, КОЕ-ГМ. КОЕ-М, эритропоэтин, тромбопоэтин, фактор стволовой клетки и лиганд flk2/flk3.
В настоящем изобретении дается описание ингибитора стволовой клетки (INPROL), который отличается от тех, которые известны у специалистов, таких как MIP-1 альфа, TGF бета, тетрапептид Фринделя и его коллег или пентапептид Пауковица и его сотрудников (cf., Wright & Pragnell, 1992 (ор cit)). В естественных условиях INPROL имеет молекулярный вес, превышающий 10000 дальтон посредством ультрафильтрации, что выделяет его из тетрапептидов и пентапептидов. Он более гидрофобен, чем MIP-1 альфа и TGF бета в системах обращенной хромотографии, что выделяет его из этих цитокинов. Далее, режим его действия отличается от тех ингибиторов, описание которых приводится выше, и он остается активным при анализе in vitro при использовании только в течение преинкубационного периода. Например, MIP-1 альфа не является эффективным при использовании в течение преинкубационного периода (пример 5). Далее, в естественных условиях INPROL активен при анализе, проводимом для измерения "клеток с высоким пролиферативным потенциалом" (HPP-PFC), в то время как MIP-1 альфа не является таковым (пример 6).
Краткое описание чертежей
На рисунках 1-4 приведено расположение продукта геля полиакриламида SDS после каждого этапа очистки.
На рисунке 1: позиция 1 - химотрипсиноген, позиция 2 - овальбумин, позиция 3 -BSA, позиция 4 - фракции <30 kD, позиция 5 - фракции 30-50 kD и позиция 6 - фракции 50-100 kD.
На рисунке 2: позиция 1 - после осаждения сульфата аммония (40-80%) и позиции 2-5 - фракции DEAE (позиция 2 представляет собой активную фракцию).
На рисунке 3: позиция 1 - супернатант после осаждения сульфата аммония, позиция 2 - активная фракция DEAE, позиции 3-5 представляют собой фракции фильтрации геля (позиция 5 представляет собой активную фракцию).
Рисунок 4: позиция 2 представляет собой окончательный продукт.
На рисунке 5 приведена обращенная хроматография окончательной очистки.
На рисунке 6 приведено введение тритированного тимидина (cpm) в клетки линии смеси FDCP (контроль=0% ингибирования) и совместно с различными концентрациями очищенного INPROL, полученного от костного мозга свиньи (pINPROL). Данные нормализованы в соответствии с контрольной величиной.
На рисунке 7 приведен процент клеток в фазе S цикла клетки после лечения мышей с помощью пропионата тестостерона (TSP), TSP плюс pINPROL или раствор связующего (контроль). Каждая группа включала 25 животных (3-4 на временную точку).
На рисунке 8 приведена картина выживания мышей, которых лечили двумя дозами 5-ФУ, совместно с лечением посредством pINPROL или без него. В каждую группу входило 30 животных.
На рисунке 9 приведена картина выживания облученных мышей, прошедших или не прошедших лечения посредством pINPROL. В каждую группу входило 50 животных.
На рисунках 10А и 10В приведена картина восстановления обычных донорских клеток культуры декстеровского типа в течение 1 недели (10А) и 3 недель (10В) после лечения с помощью Аrа-с или Аrа-с плюс pINPROL.
На рисунке 11 приведена картина выживания мышей (75 на группу) после летального облучения и трансплантации 3•104 клеток костного мозга после преинкубации в питательной среде (контроль) или в pINPROL (25 нг/мл) в течение 4 часов. Выживание отслеживалось в течение 30 дней.
На рисунке указан показатель КОЕ-ГМ, полученный спустя 14 дней в культуре мышей из клеток костного мозга мышей после летальной радиации и восстановления посредством донорских клеток костного мозга, прошедших преинкубацию вместе с pINPROL или в питательной среде в течение 4 часов.
На рисунке 13 приведены клетки суспензии из лимфоидной культуры декстеровского типа, которые брали и каждую неделю промывали и высеивали на чашку Петри вместе с IL-7 (10 нг/мл) после преинкубации в питательной среде или с pINPROL в течение 4 часов.
На рисунке 14 зафиксировано улучшение репопуляционной способности лейкозных переферических клеток крови, прошедших лечение посредством pINPROL. Стимулирующие клетки культуры декстеровского типа (LTC-IC) измерялись высеиванием на чашку Петри сросшихся или несросшихся клеток LTC совместно с pINPROL или без него, при этом показатель - КОЕ-ГМ замеряется на седьмой день. Данные нормализуют согласно контрольным величинам.
На рисунке 15А приведена обращенная хроматограмма С4 очищенного pINPROL, эльюирующего при 53% ацетонитрила.
Позиция 1 - сырой материал, позиция 2 - маркеры молекулярного веса и позиция 3 - очищенный продукт. На рисунке 15В приведена обращенная хроматограмма С4 MIP-1 альфа, эльюирующего при 43,9% ацетонитрила. На рисунке 15 С приведена хроматограмма сырого препарата pINPROL и очищенного препарата после фазы обращения.
На рисунке 16 приведены последовательности гемоглобина: на рисунке 16А приведены последовательности кДНК и аминокислот альфа гемоглобина человека. На рисунке 16В приведены последовательности кДНК и аминокислоты бета гемоглобина человека. Исчисление ведется в соответствием с исчислением аминокислот. На рисунке 16С приведено сравнение последовательностей аминокислот альфа- и бета-цепей гемоглобина человека, мыши и свиньи.
На рисунке 17 приведено сравнение следов ЖХВР фазы обращения С 4 pINPROL (рисунок 17А) и кристаллизованный гемоглобин свиньи (рисунок 17В).
На рисунке 18 изображен гель SDS-PAGE фракций от разделения ЖХВР фазы обращения С4 кристаллизованного гемоглобина свиньи. Позиция 1 демонстрирует маркеры молекулярного веса, позиция 2 демонстрирует фракции 48-49, полученные при первом пике (при 47.11 мин), позиция 3 демонстрирует фракции 50-51, полученные при втором пике (при 49.153 мин), позиция 4 демонстрирует фракции 54-55, полученные при третьем пике (при 52.25 мин) и позиция 5 демонстрирует фракции 56-57, полученные при четвертом пике (при 56.613 мин).
На рисунке 19 приведены сравнения электрофореза геля в двух измерениях pINPROL (рисунок 19А) и очищенного бета гемоглобина свиньи (рисунок 19В).
На рисунке 20 приведено сравнение воздействия очищенного альфа гемоглобина свиньи, бета гемоглобина и pINPROL в анализе FDCP-MIX.
На рисунке 21 продемонстрирована обращенная сепарация гемоглобина свиньи с помощью пологого градиента элюции.
С целью более полного понимания настоящего изобретения ниже приведено его более подробное описание. Это описание, хотя и является иллюстративным, не должно быть истолковано в качестве ограничительной директивы для настоящего изобретения и такие вариации, которые входят в компетенцию опытного специалиста, считаются находящимися в диапазоне настоящего изобретения.
Подробное описание предпочтительного поля применения
INPROL обратимо ингибирует деление стволовых клеток. Особо следует отметить, что INPROL является действенным при временном ингибировании деления клетки кроветворных стволовых клеток. Таким образом, методика изобретения может использоваться для смягчения нежелательного побочного воздействия химиотерапии кроветворной и иммунной систем, системы костного мозга пациента путем защиты стволовых клеток от урона, причиненного химиотерапевтическими средствами и средствами лучевой терапии, используемыми для уничтожения клеток, зараженных раком или вирусом. В одном поле применения изобретения INPROL вводят пациенту в такой дозе, которая достаточна для ингибирования деления клеток, в то время как химиотерапевтические средства действуют на заболевшую клетку. После того, как химиотерапевтическое средство выполнило свою функцию, стволовая клетка, ингибированная INPROL, без дальнейшего лечения возвращается к делящимся клеткам. Если требуется усилить процесс регенерации гемопоэза, в дополнение к упомянутым выше процедурам могут быть использованы факторы, стимулирующие рост, и цитокин.
Используемый здесь термин INPROL включает белки млекопитающего, очищенные, как это показано в примерах, гемоглобин, альфа-цепь гемоглобина (вместе с группой гема или без нее), бета-цепь гемоглобина (вместе с группой гема или без нее), и фрагменты или аналоги этих белков, включая эмбрионные, зародышевые или зрелые формы (например, альфа-, бета-, гамма-, дельта-, ипсилон-цепей, либо одиночных, либо смешанных, димеров или мультимеров (вместе с группой гема или без нее), способные ингибировать пролиферацию стволовой клетки. Термин "INPROL" включает встречающееся в естественных условиях или в неестественных условиях (например, изготовленные методом рекомбинации) формы этих белков.
В типичной клинической ситуации INPROL вводят пациенту ежедневно путем внутривенной инъекции вливанием в форме единиц дозировки, например, от 0,01 до 100 мг/кг, желательно от 0,1 до 1,0 мг/кг, вводят INPROL, например, за от 4 до 60 часов до стандартной химиотерапии или лучевой терапии.
Другое поле применения данного изобретения предусматривает предварительное лечение с помощью INPROL, которое позволяет увеличить дозы химиотерапевтических средств и средств лучевой терапии и превысить дозы, обычно переносимые пациентами.
Большая часть кроветворных стволовых клеток обычно статичны (не циркулируют). Однако в виде компенсационного ответа на ущерб кроветворной системе, нанесенный химиотерапией, большая часть стволовых клеток после химиотерапии начинает циркулировать, что делает их особенно уязвимыми к последующим дозам цитотоксической химиотерапии или терапевтического облучения. Путем ингибирования цикла таких стволовых клеток лечение посредством INPROL позволяет осуществлять более раннее и более частое введение последующих доз цитотоксической химиотерапии обычных или увеличенных доз.
В одном поле применения данного изобретения INPROL (от 0,1 до 6 мг, предпочтительно от 1,0 до 60 мг) вводят от 24 часов до 10 дней после принятия первоначальной дозы химиотерапии. После периода от 4 до 60 часов, предпочтительно от 24 до 48 часов, вводят следующую дозу химиотерапии. Такой цикл сочетания химиотерапии и INPROL продолжается согласно лечебной необходимости. Средства химиотерапии и схемы их введения выбирается на основе пригодности для особых типов опухоли в стандартной клинической практике. Кроме того, для дальнейшего улучшения хода кроветворной реконструкции после химиотерапии и лучевой терапии используют стимулирующие факторы роста, такие как КОЕ-Г и фактор стволовой клетки.
Для применения ex vivo используют от 0,1 нг до 100 нг/106 клеток/мл, желательно 20-50 нг/106 клеток/мл INPROL.
В другом поле применения изобретения INPROL используется в методике приготовления аналогичных кроветворных клеток для трансплантации. Кроветворные клетки лечат ex vivo действенной дозой INPROL для ингибирования деления стволовой клетки, а затем происходит вытеснение раковых клеток путем введения в культуры костного мозга действенной дозы химиотерапевтического средства или средства лучевой терапии. Предпочтительно использовать химиотерапевтические средства, предназначенные специально для циркулирующих клеток. Костный мозг, подвергнутый такому лечению, повторно вводят в аналогичный донор. Кроме того, пациента лечат средством, которое стимулирует гемопоэз для улучшения кроветворной реконституции пациента.
В другом поле применения изобретения INPROL используют в качестве вспомогательного средства при лечении лейкоза. Например, в состояниях болезни, когда клетки, зараженные лейкозом, не отвечают на действие INPROL, лейкозные кроветворные клетки лечат ех vivo с помощью INPROL. Введение INPROL препятствует пролиферации нормальных стволовых клеток. Таким образом, популяция нормальных стволовых клеток предохранена от повреждений в течение периода лечения пролиферирующихся стволовых клеток посредством цитотоксического средства специфического цикла клетки. Кроме того, стимулирующий цитокин, такой как IL-3 или КОЕ-ГМ, вводится дополнительно для стимуляции циркулирования в лейкозной клетке в период лечения лекарством или облучением, в то время как нормальные стволовые клетки предохраняются посредством INPROL. Пациента лечат химиотерапевтическими средствами или средствами лучевой терапии для уничтожения лейкозных клеток, а десенсибилизованный костный мозг трансплантируется обратно пациенту для восстановления кроветворной реконституции.
Таким же образом, в другой области применения изобретения при лечении пациентов, имеющих серьезные вирусные инфекции, поразившие клетки крови или лимфоциты, такие как ВИЧ, кроветворные клетки лечат ex vivo совместно с INPROL с последующим введением антивирусных средств и лекарств, которые поражают пораженные инфекцией клетки или системы, основанные на антителах для удаления пораженных инфекцией клеток. После миелобластовой антивирусной или миелобластовой химиотерапии в целях эрадикации вирусных клеток-хозяев пациента, клетки костного мозга, прошедшие лечение INPROL, возвращаются к пациенту.
В другой области применения изобретения INPROL используется для лечения нарушений, связанных с гиперпролиферацией стволовых клеток. Например, псориаз является нарушением, вызванным гиперпролиферирующими эпителиальными стволовыми клетками кожи, и иногда лечится цитотоксическими лекарственными средствами. Другие предраковые поражения, в которых задействована пролиферация стволовой клетки, также поддаются лечению INPROL, который применяется для полного или частичного ингибирования пролиферации стволовых клеток. Для этих случаев, в качестве альтернативы парентеральному методу введения веществ, используются составы локальной или трансдермальной доставки (например, мази, примочки, гели или "пэтчи"), содержащие INPROL в соответствующем месте, в качестве альтернативы парентеральному введению. В большинстве случаев лейкоза, лейкозные клетки-предшественники представляют собой дифференцированные популяции клеток, которые не подвергаются воздействию INPROL и который, таким образом, лечат посредством методики использования INPROL, описанной выше. В случае, если лейкозные клетки-предшественники слишком примитивны и чувствительны к прямому ингибированию посредством INPROL, пролиферация лейкозных клеток снижается посредством действенных доз INPROL.
Антитела, моноклональные или поликлональные, превращаются путем стандартной методики в полипептиды INPROL. Эти антитела или полипептиды INPROL маркируют поддающимися обнаружению метками, многие типы которых хорошо известны в отрасли. Помеченные антитела INPROL или анти-INPROL используются затем в качестве маркеров стволовой клетки для индентификации и выделения стволовых клеток путем их прямого введения пациенту с диагностическими целями. С другой стороны, эти помеченные полипептиды или антитела используются ex vivo для опознания стволовых клеток в препарате кроветворной стволовой клетки для стимуляции их выведения перед десенсибилизацией раковых клеток в костном мозге. Таким же образом, такие помеченные полипептиды или антитела используются для выведения и идентификации эпителиальных или других стволовых клеток. Кроме того, такие тела, помеченные или непомеченные, используются в терапевтических целях путем нейтрализации активности INPROL или в диагностических целях через выявление уровней циркуляции INPROL.
INPROL может быть клонирован из человеческого гена или из библиотеки кДНК для экспрессии рекомбинантного INPROL человека посредством стандартной методики. Например, с помощью данных последовательностей, полученных из очищенного белка, были взяты олигонуклеотидные пробы, которые могут иметь метки, например фосфор-32, и использоваться для проведения скрининга соответствующей библиотеки кДНК (например, костного мозга). С другой стороны, экспрессия библиотеки соответствующего источника (например, костного мозга) подвергается скринингу на код кДНК для INPROL с помощью антител или функционального анализа (например, согласно описанию, приведенному в примере 2). Сам гемоглобин, также как индивидуальные альфа- и бета-цепи, был подвергнут клонированию и экспрессии на основе самой современной методики (см. Pagnier и др. , Rev. Fr. Transfus. Hemobion. 35:407-15, 1992; Looker и др. Nature 356: 258-60, 1992; Meathods in Enzymology, том 231, 1994). В настоящем труде имеется ссылка на каждую из этих статей.
Настоящее изобретение включает типы последовательностей ДНК, которые предусматривают в свою очередь включение ко донов, "предпочитаемых" для экспрессии путем отобранных клеток-хозяев, не принадлежащим млекопитающим, и предоставление участков для деления путем ограничения эндоядерных ферментов; а также предоставление дополнительных исходных, конечных или промежуточных последовательностей ДНК, которые способствуют либо образованию переносчиков, готовых подвергнутся экспрессии, либо производству или очистке альфа-, бета-, гамма-, дельта-, ипсилон- и/или зета-цепей гемоглобина.
Настоящее изобретение, кроме того, содержит собой последовательности ДНК, кодирующие аналоги полипептидов или производных гемоглобина альфа-, бета-, гамма-, дельта-, ипсилон- и/или зета-цепей, которые отличаются от образований, встречающихся в естественных условиях с точки зрения идентичности или расположения остатков одной или более аминокислот (т.е. аналоги делеции, содержащие менее всех указанных выше остатков; аналоги замены, при которых происходит замена одного или более указанных остатков другими остатками, и аналоги добавления, при которых остатки одной или более аминокислот добавляют к окончательной и промежуточной доли полипептида), и которые разделяют некоторые или все свойства образования, встречающегося в естественных условиях.
В лучшем случае, INPROL является продуктом экспрессии прокариотической или эукариотической клетки-хозяина (т.е. с помощью бактериальных, дрожжевых клеток, а также клеток высших растений, насекомых и млекопитающих) экзогенной последовательности ДНК, полученной посредством геномического клонирования, клонирования ДНК или путем генного синтеза. То есть в благоприятных условиях INPROL становится "рекомбинантным INPROL". Продукт экспрессии в типичных дрожжах (например, Saccharomyces cerevisiae) или прокариотической клетки-хозяина (например Е.соli) свободен от ассоциаций с белками млекопитающих. Продукты экспрессии вертебральных (например, млекопитающих, за исключением человека) (например, COS или СНО) клеток свободны от ассоциаций с любыми клетками человека. В зависимости от используемого хозяина полипептиды изобретения могут быть гликозилированы и негликолизированы. Кроме того, факультативно полипептиды настоящего изобретения включают исходный остаток метиониновой аминокислоты (в позиции 1).
Настоящее изобретение охватывает также другие продукты, такие, например, как аналоги полипептидов альфа-, бета-, гамма-, дельта-, ипсилон- и/или зета-цепей гемоглобина. Такие аналоги включают фрагменты альфа-, бета-, гамма-, дельта-, ипсилон- и/или зета-цепей гемоглобина. Следуя известным процедурам, можно без труда спроектировать и изготовить код генов для микробной экспрессии полипептидов, имеющих первичную последовательность, отличающуюся от упомянутых выше в отношении идентичности и расположения одного или более остатков (например, замены, окончательных и промежуточных добавлений и делений). С другой стороны, модификации кДНК и геномических генов могут быть весьма легко выполнены путем известной управляемой сайтом методики мутагенеза и используемых для генерации аналогов и производных альфа-, бета-, гамма-, дельта-, ипсилон- или зета-цепей гемоглобина. Такие продукты обладают, по крайней мере, одним из биологических свойств INPROL, но могут иметь различия в других. Например, продукты изобретения включают альфа-, бета-, гамма-, дельта-, ипсилон- или зета-цепи, которые сокращены, например, делециями, или те, которые более устойчивы к гидролизу (и поэтому могут носить более подчеркнутый характер или обладать более длительным сроком воздействия, чем продукты, встречающиеся в естественных условиях); или которые были изменены делецией или путем добавления одного или более потенциальных сайтов для О-гликозилирования и/или N-гликолизирования, или те, у которых один или более остатков цистеинов были подвергнуты делеции или замене с помощью, например, остатков аланина или серина. Кроме того, их легче выделить в активной форме из микробных систем; или те, у которых один или более остатков тирозина заменены фенилаланином и связываются очень просто с контрольными белками или рецепторами на контрольных клетках. Под ними также понимают фрагмент полипептида, дуплицирующие только часть непрерывной последовательности аминокислоты или вторичную конформацию в пределах альфа-, бета-, гамма-, дельта-, ипсилон- и зета-цепей, фрагменты которой могут обладать одним свойством INPROL (например, связка рецептора) и не обладать другими (например, активность по ингибированию стволовой клетки). Важно знать, что эта активность необязательна для одного или более продуктов изобретения, предназначенных для терапевтических целей (см. Weiland и др., Blut 44:173-5, 1982) или других целей, таких как анализ антагонизма ингибирующего фактора. Конкурирующие антагонисты полезны в случае перепроизводства ингибиторов стволовой клетки или их рецепторов.
Следует добавить, что пептиды, полученные из последовательности белка, которая сохраняет биологическую активность, могут быть химически синтезированы с помощью стандартной методики. Настоящее изобретение также содержит коды последовательностей для аналогов пептида или производных гемоглобина альфа-, бета-, гамма-, дельта-, ипсилон- и/или зета-цепей, которые отличаются от обычно встречающихся форм с точки зрения идентичности и расположения одного или более остатков аминокислот (например, аналоги делеции, содержащие менее всех упомянутых остатков; аналоги замены, при этом один или более упомянутых выше остатков заменяют другими остатками, встречающимися в естественных условиях, или другими аналогами, известными современной науке, например такими как аминокислоты-Д, и аналоги добавления, при этом один или более остатков аминокислот химически модифицируют для повышения устойчивости, растворимости и/или сопротивляемости к протеолизу) и которые обладают некоторыми или всеми свойствами форм, встречающимися в естественных условиях.
Последовательности пептида согласно вышеприведенному описанию могут быть идентифицированы различными способами. С помощью сравнения трехмерных структур цепей нативного гемоглобина, активных при анализе (например, альфа-цепи), со структурно родственными неактивными белками (например, миоглобин) можно иденцифицировать области, которые имеют различные конформации в трехмерном пространстве и которые поэтому являются потенциальными областями для активных пептидов. В качестве другого подхода используют протеолиз, при котором протеолитические ферменты используются в огранических переварах цепей гемоглобина; в результате получают пептиды, которые могут делится на части, например, посредством ЖХВР обращенной фазы, а затем подвергнутся анализу на ингибирование стволовой клетки. Пептиды могут быть также генерированы путем химического синтеза (например, твердофазного синтеза); ряд перекрывающихся пептидов (например, 15-меры), которые охватывают последовательность интересующей цепи гемоглобина (например, альфа-цепи) может быть легко генерирована и проверена путем анализа стволовой клетки. Могут быть генерированы комбинаторные библиотеки, в которых производятся сложные химические синтезы и где отобранные позиции аминокислот отличаются изменчивостью; следствием этого образуется большое количество пептидных аналогов для скрининга (например, Dooley и др. Peptide Research 8:124-137,1995). С другой стороны, могут применяться рекомбинантные методы. Для идентификации критических остатков, необходимых для активности конкретной цепи гемоглобина, может быть использован мутагенез направленных сайтов. Области цепи, которые считаются активными в качестве ингибитора стволовой клетки (например, альфа-цепь) могут быть заменены областями из родственного, но неактивного белка (например, миоглобин) и проверены анализом стволовой клетки, позволяя, таким образом, идентифицировать области, необходимые для активности. Такие идентифицированные области могут быть подвергнуты экспрессии в качестве пептидов и проверены на активность в анализах циркуляции стволовой клетки.
Гомологичные и аналогичные варианты INPROL из других образцов используются в разнообразных ветеринарных целях, подобных тем терапевтическим областям применения изобретения, которые были описаны выше.
INPROL действует на циркулирующие стволовые клетки путем их возвращения в неделимое состояние "покоя". При желании для стимуляции деления покоящейся стволовой клетки, например, после лечения пациента, больного раком, посредством средств химиотерапии или лучевой терапии, субъекту вводят факторы, стимулирующие колониеобразование, и другие кроветворные стимулирующие вещества. В качестве примеров таких факторов можно привести: КОЕ-М, (КСФ-1), ГМ-КСФ, Г-КСФ, мегакариоцит-КСФ, мегакариоцитопоэз, фактор стволовой клетки или другие цитокины, такие как IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-9, IL-11, IL-12, IL-13, IL-14 или эритропоэтин.
Полипептиды INPROL или активные фрагменты, стволовая клетка которых характеризуется ингибирующей активностью, очищаются или синтезируются путем обычных химических процессов в сочетании с соответствующими биоанализами ингибирующей активности стволовой клетки, что подтверждается примерами, которые приведены в приложении.
В соответствии с одной из областей применения изобретения терапевтически активная доза белка INPROL или его терапевтически активный фрагмент применяются в смеси с фармацевтически приемлимым носителем. Этот состав INPROL обычно вводят посредством парентеральной инъекции или вливанием. Подкожные, внутривенные или внутримышечные способы применения инъекции выбирают в соответствии с достигнутым лечебным эффектом.
При систематическом введении лечебный состав, используемый в соответствии с настоящим изобретением, имеет форму водного раствора, который принимают парентерально, свободного от парегена. Фармацевтически приемлемый стерильный раствор белка, в котором должное внимание уделено рН, изотоничности, устойчивости белков носителя и т.д., отвечает самым высоким требованиям.
Изобретение охватывает также фармацевтические составы, включающие эффективные дозы полипептидных продуктов настоящего изобретения, а также удобным растворителем (разжижающие средства), консерванты, солюбилизаторы, эмульгаторы, адъюванты и/или носители также полезны при лечении INPROL. Термин "терапевтически эффективная доза", используемый здесь, подразумевает дозы, которые обеспечивают лечебный эффект для данного режима кондиции и приема. К таким составам относятся жидкости, гели, мази или подвергнутые лиофилизации или другому виду сушки составы, и включают растворители с различным содержанием буферного раствора (например, Tris-НС1, ацетат, фосфат), рН и ионной активностью, добавки, такие как альбумин или желатин для того, чтобы предотвратить рассасывание поверхности, моющие вещества (например, Tween 20, Tween 80, Pluronic F16, соли желчных кислот), склонные к солюбилизации средства (например, глицерин, полиэтиленгликоль), антиоксиданты (например, аскорбиновая кислота, метабисульфит), консерванты (например, тимеросал, бензил, спирт, парабены), средства "ангро" или тонусные модификаторы (например, лактоза, маннит), ковалентное присоединение полимеров, таких как полиэтиленгликоль к белку, комплексообразование с ионами металла или введение материала в
макрочастичные препараты полимерных составов, таких как полимер молочной кислоты, полигликолевая кислота, гидрогели и т.д., или липосомы, ниосомы, микроэмульсии, мицеллы, однослойные или многослойные пузырьки, инжектируемые микрокапсулы или микросферы, построенные на основе биологического распада, или белковые матрицы, "тень" эритроцита, сферопласты, "пэтчи" кожи, или другие методы выделения или упаковки фармацевтических препаратов. Такие составы повлияют на физическое состояние, растворимость, устойчивость, интенсивность выделения in vivo, коэффициент очищения INPROL in vivo. Составы, контролирующие и поддерживающие выделение включают технологию приготовления лекарственных средств в липофильных депо (например, в жирных кислотах, парафине, растительных маслах). Настоящее изобретение охватывает также составы твердых веществ, покрытые полимерами (например, пологзамеры или пологзамины) и INPROL, связанные с антителами, направленными против рецепторов из особой ткани, лигандов или антител, или связанные с лигандами рецепторы из особой ткани. Другая область приминения составов изобретения включает формы твердых веществ предохранительного покрытия, протеазные ингибирующие факторы или стимуляторы проницаемости для различных способов введения лекарственных средств, включая парентеральный, легочный, назальный, локальный (кожный или через слизистую оболочку) и оральный. В других областях приминения состав, содержащий INPROL, вводят локально или через трансдермальный "пэтч".
В одной из областей применения, составы настоящего изобретения упаковывают в стерильные пробирки или ампулы в форме дозировочной единицы.
Изобретение также охватывает составы, включая один или более дополнительных факторов, таких как химиотерапевтические средства (например, 5-фторурацин, 5-ФУ), цитозин арабинозид, циклофосфамид, циплатин, карбоплатин, доксирубицин, этопозид, таксол, алкилирующие средства), антивирусные средства (например, AZT, ацикловир), TNF, цитокины (например, интерлейкины), противопролиферативные лекарственные средства, антиметаболиты и лекарственные средства, которые воздействуют на метаболизм ДНК.
Режим дозирования, используемый для лечения субъекта, подлежащего воздействию таких цитотоксических средств, или для лечения гиперпролифирирующих стволовых клеток определяется участковым врачом, учитывающим различные факторы, которые вносят изменения в действие лекарственных средств, например состояние, вес тела, пол и диета пациента, серьезность инфекции, время введения и прочие клинические факторы.
Следуя за воздействием на субъект цитотоксическим средством или средством лучевой терапии, терапевтический метод настоящего изобретения дополнительно предусматривает введение субъекту одного или более лимфокинов, колониеобразующие факторы или другие цитокины, гемапоэтины, интерлейкины или факторы роста для общей стимуляции роста и деления стволовых клеток (и их наследников), ингибированных предшествующим лечением посредством INPROL. Терапевтические средства, которые стимулируют гемопоэз, включают IL-I, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, Meg-CSF, MCSF (CSF-1), GM-CSF, G-CSF или эритропоэтин. Дозы этих средств определяют на основе знаний, полученных при их использовании в клинических испытаниях на эффективность в целях стимулирования кроветворной реконституции после химиотерапии или трансплантации кроветворных стволовых клеток. Эти дозы будут откорректированы для компенсации изменений в физическом состоянии пациента; следует также откорректировать дозу и тип химиотерапевтического средства или средства лучевой терапии, используемых для лечения субъекта. Прогресс в реверсировании ингибирования стволовых клеток, ставший следствием введения INPROL подвергнутого лечению пациента, отслеживают обычными способами.
При лечении лейкоза полезно вводить совместно INPROL для ингибирования циркуляции нормальных стволовых клеток и стимулятор роста лейкозной клетки, такой как IL-3 или ГМ-КОЕ, при этом следует параллельно проводить лечение посредством цитотоксических лекарственных средств или средств лучевой терапии. Согласно настоящему протоколу представляется возможным достичь значительной разницы между статусами циркулирования и чувствительности лекарственных средств в нормальных и лейкозных клетках.
Пример 1: Анализ in vivo ингибирования пролиферации стволовой клетки.
Для определения пролиферации стволовой клетки количество КОЕ-С в фазе-S стволового цикла измерялось с помощью метода "самоубийства" 3H-тимидин (Becker и др.. Blood 26:296-308,1965).
Незрелые кроветворные клетки-предшественники - колониеобразующие единицы в селезенке (КОЕ-С) могут быть обнаружены методом in vivo путем образования макроскопических колоний в селезенке смертельно облученных мышей 8-10 дней спустя после внутренней инъекции кроветворных клеток (Till&McCulloch, 1961).
Для стандартного анализа пролиферации КОЕ-С обычно используется метод "самоубийства" 3H-тимидин (Becker и др., 1965). Способ основан на введении тимидина, меченного радиоактивным изотопом,(3H-тимидин) предшественника ДНК, в клетки в течении синтеза ДНК. КОЕ-С, находящиеся в фазе-S цикла во время тестирования, убивают высокой радиоактивностью и, таким образом, не способны образовывать колонии в селезенке. Таким образом, разница между количеством КОЕ-С, образованного путем инъекции образца клетки, выращенного без 3H-тимидина, и те же самые клетки, выращенные вместе с 3H-тимидином, показывает процент пролиферирующего КОЕ-С в исходном образце.
Тестирование ингибитора не может быть проведено с популяцией стволовой клетки костного мозга, не стимулированных животных, поскольку ингибитор воздействует только на циркулирующие КОЕ-С, которые составляют всего 7-10% общей популяции КОЕ-С в костном мозге нормальной мыши.
Для стимуляции пролиферации КОЕ-С использовали фенилгидразин (ФГЗ) или сублетальное облучение (Lord, 1976).
Мы разработали способ применения тестостерон-пропионата (ТСП), основанный на моделирующем воздействии на КОЕ-С (Byron и др.. Nature 228: 1204,1970), что облегчает тестирование и не причиняет никаких побочных эффектов. Индуцированная ТСП стимуляция пролиферации КОЕ-С в течение 20-24 часов после инъекции, а эффект можно наблюдать, по крайней мере, 7 дней.
Процедура, используемая для скрининга фракций в период очистки ингибитора была следующая:
мыши: на протяжении всего периода тестирования использовались штаммы мышей BDF1 или CBF1.
Мышей-доноров лечили посредством дозы в 10 мл/100 г ТСП путем внутриперитонеальной инъекции объемом 0,2 мл /мышь для индуцирования 30-35% КОЕ-С в фазу-S.
Спустя 24 часа костный мозг берется из бедра для приготовления суспензии клетки. От пяти до десяти миллионов клеток на 1 мл выращивают с различными фракциями контроля и тестирования в течение 3,5 часов при 37oС в водяной бане, с двумя трубками для каждой группы (одна для горячей (радиоактивной) и одна для холодной (нерадиоактивной)).
Спустя 3,5 часа к каждой горячей трубке добавляют 3H-тимидин (1 MCi/мл, удельная активность 18-25 Ci/ммоль) в объеме 200 мкл на 1 мл клеточной суспензии; к холодным трубкам ничего не добавляют. Инкубация продолжается еще в течение 30 минут при 37oС.
После 30 минут инкубации реакция нейтрализации прекращается путем добавления 10 мл холодного (4oС) субстрата, содержащего 400 мкг/мл нерадиоактивного тимидина. Клетки экстенсивно промываются (3 раза).
Клетки ресуспендированы и растворяются до желательной концентрации для инъекции, обычно до 2-4•104 клеток на мышь в 0,3-0,5 мл.
Мышей-реципиентов, 8-10 на группу, облучают радиацией не позднее чем за 6 часов перед инъекцией.
Клетки селезенки реципиентов собирают на 9-12 день и фиксируют в растворе Теллесницкого; колонии подсчитывают на глаз. Процент клеток в фазе-S вычисляют по формуле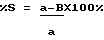 ,
,
где а - количество КОЕ-С без 3H-тимидина;
в - количество КОЕ-С с 3H-тимидином.
Данные тестирования INPROL, представленные в Таблице 1, показывают, что у циркулирующих клеток после лечения посредством INPROL усиливается сопротивляемость действию 3H-тимидина. Для этого и всех последующих примеров термин "pINPROL" касается очищенного белка из костного мозга свиньи. Такая же защита наблюдается для цитозина арабинозида гидроксимочевины особых цитотоксических лекарственных средств S-фазы. Если подвергнутые лечению стволовые клетки затем промываются холодным субстратом, содержащим нерадиоактивный тимидин, то выживающие стволовые клетки пролифелируют в селезенке мыши для образования колоний в обычном режиме.
Пример 2: Анализ ингибирования пролиферации стволовых клеток in vitro.
С помощью следующей тестовой системы (Lord и др. в The Inhibitors of Hematopoiesis, стр. 227-239, 1987) был продемонстрирован непосредственный эффект INPROL. Зависимая линия стволовой клетки фактора мультипроисхождения, смесь FDCP А4 (А4), содержалась в среде Дульбекко с добавлением 20% лошадиной сыворотки и 10% кондиционной Среды (КС) WEHI-3 в качестве колониестимулирующих средств для IL-3.
Анализ введения тритированного тимидина использовался для измерения пролиферации, клетки А4 (5•104 в среде 100 мкл с 20% лошадиной сыворотки и 50% WEHI-3 КС) выращивались при 37oС в 5% СО2 в течение 16 часов.
pINPROL или неочищенный ВМЕ (фракция IV) добавили вначале. Затем добавили тритированный тимидин ((3H-TdR) 3,7 Kbq в 50 мкл при 740 Gbq/ммоль) в каждую группу для последующих 3 часов инкубационного периода. Уровень пролиферации замерили путем сбора клеток, а процент ингибирования расчитали по формуле ,
,
где чим - число импульсов в минуту (прим. переводчика).
Включение тритированного тимидина (3H-TdR) посредством клеток смеси FDCP -А4, выращенных в присутствии дифференцированных доз экстракта нормального костного мозга или pINPROL изображено на рисунке 6. Можно заметить, что очищенный состав pINPROL, по крайней мере, в 1000 раз более активен, чем исходный материал. Время экспонирования (16 часов) является важным фактором для достижения эффективного ингибирования и является свидетельством прямого воздействия pINPROL на стволовые клетки клеточной линии А4.
Пример 3: Ингибирование пролиферации с помощью INPROL, инъектированного in vivo: дозы и продолжительность воздействия.
Изучение воздействия INPROL, введенного путем инъекции, показало, что INPROL может эффективно блокировать рекрутмент КОЕ-С в цикл, таким образом, предохраняя эти клетки от цитотоксического воздействия дальнейшего лечения, демонстрируя потенциальные возможности применения в клинических целях.
Эксперимент преследовал две задачи: проверить воздействие INPROL на КОЕ-С при инъецировании in vivo и определить эффективную продолжительность активности INPROL по отношению к циркулирующим стволовым клеткам.
В целях стимулирования пролиферации КОЕ-С, инъекция тестостерона пропионата использовалась на основе эффекта, упомянутого в примере 1.
Мышам BDF1 был инъецирован TSP ( 10 мг/100 г) в день 0, через 24 часа мышы каждой эксперементальной группы (4 мыши в группе) получили одноразовую инъекцию дозами 0, 5, 10 и 15 мкг одной мыши.
Спустя 24 часа после инъекции мышей умертвили, а процент циркуляции КОЕ-С измерили путем анализа, описание которого приведено в примере 1. Инъекция TSP индуцировала приблизительно 50% КОЕ-С для циркулирования по сравнению с 7% у мышей, не прошедших лечения. pINPROL в дозах менее 2 мкг был неспособен вызвать ингибирование пролиферации, индуцированной TSP, и довести ее уровень до нормального уровня.
Для продолжения оценки воздействия, одна группа мышей (21 на группу) была инъецирована только TSP, а другая группа была инъецирована как TSP, так и pINPROL (спустя 24 часа после TSP). Циркулирование КОЕ-С измеряли каждые 24 часа в течение недели - для этой цели брали по три донора из каждой группы и измеряли статус цикла КОЕ-С в их костном мозге посредством описанного выше способа (см. пример 1).
Для продолжения оценки воздействия одна группа мышей (21 на группу) была инъецирована только TSP, а другая группа была инъецирована как TSP, так и pINPROL (спустя 24 часа после TSP). Циркулирование КОЕ-С измеряли каждые 24 часа в течение недели . Для этой цели брали по 3 донора из каждой группы и измеряли статус цикла КОЕ-С в их костном мозге посредством описанного выше способа (см. пример 1).
Данные, предоставленные на рисунке 7, показывают, что воздействие TSP длится приблизительно семь дней, одноразовая инъекция INPROL может ввести КОЕ-С в состояние покоя и поддерживать их вне цикла в течение более 48-72 часов. Поскольку большая часть химиотерапевтических средств, используемых для лечения рака и лейкоза, имеет сравнительно короткое время полужизни in vivo, которое обычно составляет менее 24 часов, воздействие INPROL, согласно полученным данным, сохраняется в течение более длительного времени, чем время воздействия, в течение которого такие химиотерапевтические средства, как цитозин арабинозид или гидроксимочевина сохраняются активными in vivo. Для химиотерапии чрезвычайно важно сохранять длительные промежутки времени (более 24 и менее 96 часов) между первым (неповреждающим стволовые клетки) и вторым (повреждающим КОЕ-С) этапами лечения, одноразовая инъекция INPROL в течение интервалов между двумя приемами средства химиотерапии или лучевой терапии должно быть достаточным. Единая стратегия, основанная на продолжительности воздействия INPROL, применяется в течение нескольких повторяющихся циклов цитотоксической или лучевой терапии.
Пример 4: Наиболее примитивные кроветворные стволовые клетки, стимулированные на быстрое стимулирование после лечения посредством 5-ФУ, предохранены при содействии INPROL от второй экспозиции 5-ФУ.
Лекарственное средство 5-флуороурацил (5-ФУ) значительно снижает количество клеток в лимфатических узлах и секторах спинного мозга. Обычно при этом имеют в виду использование лекарственного средства для циркулирующей клетки, имеющего целью быструю пролиферацию клеток, так как введение аналога нуклеотида в ДНК во время фазы-S цикла клетки или до нее приводит к смерти клетки. Выживание в течение длительного периода времени и иммунокроветворная реконституция костного мозга мышей не поддаются воздействию после принятия одной дозы 5-ФУ; однако опыты показали (Harrison и др. Blood 78: 1237-1240, 1991), что плюрипотентные кроветворные стволовые клетки (ПКСК) становятся неуязвимыми ко второй дозе 5-ФУ в течение короткого периода времени в 3-5 дней после первоначальной фазы. Это можно объяснить тем, что нормальный цикл ПКСК слишком замедленен, чтобы одиночная доза 5-ФУ была эффективна и стимулировала быстрое циркулирование посредством стимула, вызванного первоначальным лечением посредством 5-ФУ. Мы предлагаем возвратить ПКСК в статус замедленного цикла посредством введения INPROL и, таким образом, предохранить от второго этапа лечения с использованием 5-ФУ.
Мыши, использованные в экспериментах, представляли собой особей мужского пола BDF1. Исходный раствор 5-ФУ (Сигма) приготовили в растворе физиологической соли с концентрацией 10 микрограмм/мл. Каждая мышь, подвергнутая лечению, получила 2 мг 5-ФУ на 10 г веса тела через хвостовую вену в день 0 эксперимента; через 24 часа мышей инъецировали интраперитонеально pINPROL (10 микрограмм/100 г веса тела), а в день 3 инъецировали вторую дозу 5-ФУ. Изучение процесса выживания проводили путем отслеживания смертности мышей в экспериментальной (лечение с использованием pINPROL) и контрольной круппах, по 30 мышей в каждой. Кривые выживаемости приведены на рисунке 8 оригинала.
Пример 5: Эффект преинкубационного периода совместно с INPROL по сравнению с MIP-lα в клетках косного мозга.
Целью эксперимента было сравнение in vitro ингибирующего воздействия преинкубации с pINPROL и MIP-lα на клетки костного мозга мышей.
Использовали следующую процедуру:
in vivo: мышей BDF1 в возрасте 6-15 недель инъецировали 200 мг/кг 5-ФУ при инкубационном периоде 48 часов до сбора костного мозга из бедренной кости;
in vitro: в суспензии одиночных, объединенных в пул клеток произвели подсчет и 5•106 клеток заключили в инкубатор, общий объем составил при этом 2 мл вместе или без pINPROL или MIP-lα, кроме этого, в состав вошли: 5% лошадиной сыворотки, среда Дульбекко, при этом добавили глютамин-Л, при 37oС и 5% СО2 вышеупомянутые компоненты оставили в инкубаторе в течение 4 часов. После этого клетки дважды промывают и пересчитывают. Их культивируют в метилцеллюлозе при следующих окончательных условиях:
0,8% метилцеллюлоза
25% лошадиная сыворотка
20 нг/мл рекомбинант мыши IL-3 добавили L-глютамин
5•105 клеток на 1 мл
Среда Дульбекко
Планшет проходил инкубацию в течение 11 дней при 37oС и 5% СО2 при 100% влажности. Были насчитаны колонии, состоящие из более 50 клеток (см. Таблицу 2).
Пример 6: INPROL ингибирует пролиферацию колониеобразующих клеток с высоким пролиферативным потенциалом (ККВПП).
Анализ колониеобразующих клеток с высоким пролиферативным потенциалом (ККВПП) является анализом in vitro, который используется для оценки реконституции стволовых клеток выше их ранних предшественников; другие схожие анализы, например КОЕ-А, КОЕ-ГМ, КОЕ-Е и КОЕ-ГЕММ, обнаруживают прогрессивно ограниченные популяции клеток-предшественников (М. Moore, Blood 177: 2122-2128, 1991). Этот пример показывает, что предварительное лечение клеток посредством pINPROL ингибирует их пролиферацию, в то время как применение MIP-lα не приводит к таким результатам при аналогичных условиях проведения эксперимента.
Мышей BDF1 лечили посредством 5-флуороурацила (200 мг/кг и.п.) перед тем, как их костный мозг был проанализирован на число ККВПП. Клетки были промыты посредством обработки в центрифуге и выдержаны в инкубаторе при плотности от 106 до 5•106 клеток/мл в среде вместе или без добавления агента (управления), pINPROL(25 нг/мл) или MIP-lα (200 нг/мл) в течение 4 часов. После инкубации клетки промывают и высеивают в агар (0,3%) с 30% ФТС (фетальной телячьей сыворотки) и смешанной кондиционной средой от 5637 и линией клеток WEHI-3B (7,5% каждой кондиционной Среды, согласно рекомендациям Sharp и др., 1991). Концентрация высеивания составила 5•10 4 клеток/мл в 60-миллиметровые планшеты. Колонии подсчитали на 14 день. Результаты приведены в Таблице 3.
Согласно этим результатам MIP-lα не ингибировал пролиферацию большинства незрелых предшественников, когда они присутствуют только в течение преинкубационного периода. pINPROL активно ингибировал пролиферацию при этих условиях, указывая, таким образом, на фундаментальные различия между pINPROL и MIP-lα в отношении биологической активности.
Пример 7: Лечебное воздействие INPROL на выздоровление от индуцированной радиацией аплазии костного мозга.
Аплазия костного мозга является основным видом ограничивающей токсичности лучевой терапии раковых заболеваний. Было продемонстрировано, что некоторые факторы роста (например, КСФ-Г, КСФ-ГМ, эритропоэтин) могут усилить выздоровление от аплазии костного мозга, вызванной радиацией. Концепция защиты путем использования ингибитора пролиферации стволовой клетки является другим дополнительным способом борьбы с гематологическим уроном. Следуя процедуре лечения, разработанной выше (примеры 3,4) была создана модель летального облучения мышей. Специалисты знают, что мышь, получившая 9 грей кобальта 60, начинает умирать через 10-14 дней, к 30 дню смертность составляет приблизительно 50%. Эта летальная доза использовалась в нашем моделировании путем разбивки этой дозы на две последовательно вводимые дозы по 4,5 грей каждая, с промежутком в 3 дня между ними. Предварительные данные показали, что кривая выживаемости при этой модели была очень близка к кривой, при которой облучение производилось один раз в 9 грей, более того, испытания на пролиферацию КОЕ-С показали, что спустя 24 часа после первого облучения 35-40% КОЕ-С индуцировано к пролиферации. Такие клетки могут быть предохранены с помощью ингибитора стволовой клетки, введенного до второй дозы.
Для проверки этой возможности мыши (50 мышей в группе) получили дозу в 4,5 грей в день 0. Спустя двадцать четыре часа одной группе был введен pINPROL (2 микрограмма на мышь и.п.), а другой контрольной группе инъецировали соль. Вторую дозу радиации (4,5 грей) мыши получили на третий день.
На рисунке 9 отражено улучшение показателя выживаемости после введения одной дозы pINPROL. Условия моделирования клинически схожи с лечением любого вида раковых заболеваний, включая те из них, которые характеризуются появлением твердых опухолей; такому лечению подвергают пациента, имеющего рак, посредством введения ему эффективных доз IMPROL в промежутке между двумя последовательными дозами радиации, таки образом, позволяя подвергать пациента более крупным дозам радиации, используемой при лечения рака. Кроме того, появляется возможность расширить пределы этого моделирования и на химиотерапевтические средства.
Пример 8: Использование INPROL при аутогенной трансплантации костного мозга.
Трансплантация костного мозга является единственным лечебным видом терапии для нескольких лейкозов (CML, AML и др.) Кондиционирование ex vivo аутогенных трансплантаций костного мозга для инфузии призвано обеспечить потенциальные аутогенные источники поступления нормальных стволовых клеток, не зараженных лейкозом и способных к репопуляции кроветворной системы реципиента, для обеспечения активного и эффективного лечения.
1. Модель лейкоза L1210 у культуры декстеровского типа для изучения воздействия INPROL, предохраняющего нормальный гемопоэз во время дс с помощью АrаС.
Культуры декстеровского типа (КДТ) были восстановлены согласно Toksoz и др. (Blood 55: 931-936, 1980), а линия лейкозных клеток L1210 была адаптирована к КДТ путем совместной культивации в течение 2 недель. Одновременный рост нормальных и лейкозных клеток-предшественников имел место в этих объединенных культурах КДТ/L1210, подобно ситуации в костном мозге пациента, зараженного лейкозом. Отделение нормальных колониеобразующих единиц (КОЕ) от лейкозных КОЕ стало возможным путем выращивания их как агарных колоний в присутствии или с отсутствием кондиционной среды из WEHI-3 (производительная линия клеток мыши IL-3). Нормальные клетки подвергают апоптозу при отсутствии IL-3, в то время как лейкозные клетки могут образовывать колонии в его отсутствие. Клетки суспензии из состава КДТ/1Л210 создают приблизительно 150 колоний в присутствии IL-3 (нормальные кроветворные клоны) и 70 колоний при выращивании без IL-3 (лейкозные клоны) на 50000 высеянных клеток.
Процедура дс проходила следующим образом: в день 0 все клетки суспензии и питательной среды (10 мл/колбу) были взяты из колб, содержащих КДТ-L1210 и заменены на 2 мл питательной среды, содержащей 200 микрограммов цитозина арабинозида (AraC) (Tsyrlova и др. в Leukemia: Advances in Biology and Therapy, т. 35, 1988); после 20 часов инкубации колбы были промыты и заменены 2 мл только свежей питательной среды (контрольная группа) или питательной Среды, содержащей pINPROL при 25 нг/мл в течение 4 часов. После такой предварительной инкубации клетки прошли инкубацию вместе со 100 микрограммами на колбу АrаС в течение 3 часов при 37oС. В каждой группе было 4 колбы. Культуры КДТ-L1210 промыли три раза и заполнили свежей питательной средой КДТ; их сохранили, как и в предыдущем примере, в целях изучения регенерации на 3-4 недели.
Данные предоставлены на рисунке 10. Рост клеток не был зафиксирован в контрольных культурах, подвергнутых воздействию только AraC, в то же время в колбах, предохраненных INPROL, регенерация гемопоэза происходила намного быстрее вследствие пролиферации клеток-предшественников из адгезивного слоя. Более того, клетки из экспериментальной группы при пересадке в агар вырастали только в присутствии IL-3, создавая около 100 КОЕ на 50000 клеток; в течение 4 недель не было замечено никакого роста лейкозных клеток. Таким образом, костный мозг, подвергнутый лечению ex vivo посредством введения эффективной дозы АrаС в сочетании с INPROL может быть от клеток, больных раком, в то время как стволовые клетки должны быть предохранены. Необходимо по возможности расширить такое моделирование до пределов применения других форм химиотерапии и лучевой терапии.
2. Возможность репопуляции костного мозга (ВРКМ) и тридцать дней радиозащиты увеличиваются посредством лечения INPROL in vitro.
ВРКМ, возможность клеток репопулировать костный мозг летально облученных радиацией мышей, вместе с возможностью предоставлять радиозащиту в течение 30 дней является прямым измерением in vivo потенциала спасения животных, имеющих пораженный костный мозг.
С целью изучения защиты от радиации мыши BDF1 были облучены силой в 9,5 грей и восстановлены путем трансплантации костного мозга от доноров, стимулированных тестостероном. Одну группу реципиентов лечили посредством пересадки клеток костного мозга, прошедших предварительную инкубацию в течение 4 часов в питательной среде (контрольная группа А) и другую (группа В) вместе с 25 нг/мл pINPROL. Клетки в обеих группах были промыты и пересажены в облученных животных из расчета 30000 на мышь. Данные выживаемости приведены на рисунке 11. Всего изображено 3 эксперимента, при этом контроль нормализован до 100%. Инкубация с pINPROL увеличила процент выживаемости мышей с 36,5% в контрольной группе до 61,8% к 30 дню.
Повышение ВРКМ, индуцированное преинкубацией совместно с INPROL, может быть одним из способов улучшения иммунитета в отношении воздействия на организм лучевой терапии. Для анализа этой гипотезы осуществили замер ВРКМ в соответствии с Visser и др. (ор cit). Короче говоря, доноры-мыши BDF1 получили предварительное лечение тестостероном, при этом их костный мозг прошел преинкубацию совместно с питательной средой или с INPROL в течение 4 часов и был инъекцирован в облученных радиацией животных. На 13 день клетки костного мозга бедренной кости реципиента были пересажены в агар в трех различных концентрациях (0,01; 0,05; 0,1 эквивалента бедренной кости) в присутствии 20% лошадиной сыворотки и 10% WEHI-CM. Количество колоний седьмого дня составило ВРКМ, так как колониеобразующие клетки в костном мозге реципиента в это время являлись предшественниками донорских незрелых стволовых клеток.
Как это видно из рисунка 12, ВРКМ популяции клеток, прошедших преинкубацию с INPROL, превышает ВРКМ контрольной группы (В).
Пример 9: Антипролиферационное воздействие INPROL на стволовые клетки может изменить их дифференциальные нарушения.
Гиперпролиферация КОЕ-С видна не только во время восстановления от цитотоксических лекарственных средств или облучения, но также как последствие обычного старения и считается главной чертой миелодиспластического синдрома (МДС). Она сопровождается дифференциальными нарушениями, такими как преобладание эритроидной дифференциации при одновременном снижении дифференциации гранулоцитного метаболического пути.
Клетки костного мозга подверглись инкубации в течение 4 часов при 37oС совместно с 25 нг/мл pINPROL или питательной средой (контрольная), промыты и высеяны в агар вместе с 20% лошадиной сыворотки, 24 ед./мл эритропоэтина и 10% WEHI-CH. Количество колоний БОЕ-Е и ГМ-КОЕ подсчитали на седьмой день. Данные, приведенные в Таблице 4, суммированы на основании трех экспериментов - взяли 4 животных на точку; высеяли 4 планшета.
Из Таблицы 4 становится очевидно, что инкубация нормального костного мозга (НКМ), взятого из нетронутых молодых животных (BDF1 8-12 недель), совместно с INPROL не изменила количество или соотношения колоний различных типов. Доноры BDF1, подвергнутые предварительной инкубации с тестостероном пропионатом (TSP), показали такое же увеличение пролиферации КОЕ-С, какое наблюдалось выше (Примеры 1, 3, 4), небольшое увеличение числа эритроидных предшественников (колонии БОЕ-Э и сокращение ГМ-КОЕ, которые были полностью нейтрализованы совместной инкубацией с INPROL. Кроме того, ненормально высокий уровень пролиферации КОЕ-С вернулся к 10% КОЕ-С в фазе-S цикла клетки. Гиперпролиферация КОЕ-С считается чертой некоторой нагрузки на мышь, индуцированной вирусным лейкозом, например мыши Balb/c (Таблица 4), и может также наблюдаться в более старых животных (Таблица 4). Аналогичная перегруппировка коммитированных предшественников, наблюдаемая в мышах BDF 1, лечение которых проводилось с помощью Т-клеточной сериновой протеазы (ТСП), наблюдается в Balb/c и в более старых (23-25 месяцев) особях BDF 1, которые также обладают ненормально высоким уровнем пролиферации КОЕ-С. Корректировка как пролиферации КОЕ-С, так и дифференциации была индуцирована посредством инкубации вместе с INPROL, что еще более ценно с клинической точки зрения. Анализ показал, что инъекция INPROL (2 микрограмма на мышь) воздействует как на пролиферацию КОЕ-С, так и на показатель эритроида (БОЕ-Э) и ГМ- колоний (Таблица 4).
Пример 10: Иммуностимулирующая активность INPROL.
Наблюдения показали, что инкубация клеток костного мозга, содержащих высокий процент пролиферирующих КОЕ-С, вместе с INPROL не только изменяет циркулирование КОЕ-С, но также и дифференциацию - происходит переход преобладания эритроидной дифференциации в пользу гранулоцитных и лимфоидных предшественников. Это свойство INPROL имеет важное значение вследствие иммуноподавляющих побочных эффектов цитотоксической химиотерапии и лучевой терапии, а также иммуноподавления, сопровождающее гиперпролиферативные нарушения стволовой клетки и старение.
Пример показывает прямое воздействие INPROL на дифференцирование незрелых предшественников из лимфоидной культуры декстеровского типа (ЛКДТ), образованной согласно Wittlock & Witte (Ann. Rev. Immun. 3:213-35, 1985) в пре-В предшественниках измерение производилось путем образования колоний в метилцеллюлозе, содержащей IL-7.
ЛКДТ были восстановлены, как это изложено, и прикормлены свежей питательной средой - ЛКДТ (Terry Fox Labs..Vancouver, Canada) дважды в неделю. Неадгезированные клетки собирали раз в неделю, вымывали факторы и подвергали инкубации в течение 4 часов вместе с 25 нг/мл pINPROL или с одной питательной средой для контроля. После инкубации клетки были промыты и высеяны с концентрацией 105 клеток на 1 мл в метилцеллюлозе, содержащей 30% ФТС (фетальная телячья сыворотка) и 10% нг/мл IL-7. Данные, начиная с третьей недели, показаны на рисунке 13. Количество крупных колоний пре-Б варьируется под контролем, повышаясь по времени, однако, преинкубация с INPROL всегда стимулирует рост колоний со складками 4-8 выше уровня контроля. Этот пример характеризует иммуностимулирующее свойство INPROL, которое используется при корректировке состояния иммунодефицита и при увеличении желаемых иммунных ответов, например, при прививке.
Пример 11: INPROL улучшает способность стволовых клеток к репопуляции - провоцирующие клетки декстеровского типа от пациента, больного хроническим костно-мозговым лейкозом.
Хронический костно-мозговой лейкоз (ХКЛ) - это летальная злокачественная болезнь кроветворной стволовой клетки. Лечение ХКЛ в хронической фазе путем применения одного химиотерапевтического средства, комбинации химиотерапевтических средств, спленэктомии или селезеночной лучевой терапией может контролировать клинические объективные и субъективные признаки болезни, однако жизнь при этом продлевается незначительно. По мере того как ХКЛ переходит из этапа хронического в обостренный, стандартная терапия перестает быть эффективной. Сейчас трансплантация костного мозга (ТКМ) является единственным видом лечебной терапии ХКЛ. Терапия посредством ТКМ не родственного донора представляет собой трудности по причине тканевой несовместимости. Менее 40% подходящих в других отношениях пациентов ХКЛ имеют подходящего родственного донора; таким образом, предпочтительно применять аутогенную трансплантацию. Кондиционирование аутогенной ТКМ с целью инфузии вместе со способностью отбора нелейкозных (Ф-отрицательных) костно-мозговых предшественников у пациентов, имеющих Ф-положительных предшественников, выращенных в культурах декстеровского типа (КДТ), предполагает возможность аутогенных источников нормальных стволовых клеток, позволяющую осуществлять активную и эффективную терапию ХКЛ.
В контексте ТКМ, кроветворная стволовая клетка может быть определена как клетка, обладающая способностью генерировать зрелые клетки крови в течение длительных периодов времени. Мы использовали систему КДТ человека, разработанную С. Eaves & A.Eaves как для анализа числа стволовых клеток, так и в качестве методики их обработки в терапевтических целях. Процедура включает высеивание клеток в предварительно восстановленном, подвергнутом облучению адгезивном слое костного мозга человека; эти культуры затем выдерживают в течение 5 недель. Предельной точкой считается общее клоногенное содержание клеток (адгезивных и неагдезивных) культур, собранных на заключительной стадии. Выход клоногенной клетки в этих условиях линейно связан с числом предшественников (провоцирующие клетки культур декстеровского типа (ПК КДТ)), добавленных первоначально; средняя производительность ПК КДТ человека составляет 4 клоногенных предшественника на ПК КДК. Выше было уже рассмотрено, что после того как костный мозг пациента, зараженный ХКЛ, содержат в таких условиях число лейкозных (Ф-положительные) клоногенных клеток резко сокращается. Путем подсчета нормальных ПК КДТ остатка у пациентов с ХКЛ представляется возможным отобрать те из них, которые могут принести пользу от интенсивной терапии, поддержанной трансплантацией выращенных трансплантантов (Phillips и др., Bone Marrow Transplantation 8:477-487, 1991).
Следующую процедуру использовали для изучения воздействия INPROL на число клогенных клеток (ПК КДТ) среди клеток трансплантанта костного мозга, восстановленных из периферической крови пациента с ХКЛ.
Культуры были инициированы как культуры декстеровского типа на строме предоперационного облучения. В качестве контрольной группы взяли периферическую кровь здорового донора. Периферические клетки крови пациента с ХКЛ прошли период предварительной инкубации совместно или без pINPROL (25 нг/мл) в течение 4 часов. были промыты и содержались в системе ПК КДТ в течение 5 недель для определения контрольного числа ПК КДТ. Для целей экспериментов другие, параллельные культуры проходили стадию восстановления в течение 10 дней. Смесь агдезивных и неагдезивных клеток, взятых из культур, выращиваемых в течение 10 дней, прошла период предварительной инкубации совместно и без pINPROL, далее ее инкубируют в предварительно восстановленных питательных фидерах в течение дополнительных 8 недель. Число ПК КДТ каждой экспериментальной культуры оценивалось путем высеивания как агдезивных, так и неагдезивных клеток в метилцеллюлозу вместе с соответствующими факторами роста (Terry Fox Laborotories, Vancouver, Canada) и подсчетом общего числа колониеобразующих клеток. Величины ПК КДТ, подсчитанные посредством этой процедуры, были получены путем оценки общего содержания клоногенных клеток (КОК - колониеобразующих клеток) по формуле
#ПК-КДТ=#КОК/4.
Данные, представленные на рисунке 14, свидетельствуют об отсутствии в течение первых 10 дней потерь ПК КДТ культуры, инициированной из костного мозга здорового донора, и приблизительно 30% количества индуцированных ПК КДТ еще продолжали присутствовать в культуре спустя 5 недель. Число ПК КДТ у пациента с ХКЛ значительно снизилось до приблизительно 8% в течение 10 дней и не восстанавливалось в течение последующей инкубации, в то время как после проведения предварительной инкубации клеток совместно с INPROL уровень ПК КДТ увеличился до 30% от исходного количества и сохранялось на этом уровне в течение 8 недель.
Клинически обоснованное использование INPROL, спрогнозированное этими предварительными данными, включает методику выборочного увеличения содержания нормальных стволовых клеток свежих или культивированных трансплантантов костного мозга, методику усиления рекрутмента остаточных нормальных стволовых клеток in vivo, а также протоколы для переноса нового генетического материала в стволовые клетки костного мозга человека для последующей трансплантации пациентам.
Пример 12А: Методика выделения иммуноактивного ингибитора пролиферации стволовых клеток из препаратов костного мозга.
Костный мозг был выделен из свиных ребер. Ребра свиной туши были очищены от мышечных волокон и жира и разрублены на кусочки. Костный мозг был выделен с помощью гидропресса, изготовленного фирмой "Биофизприбор". Клетки костного мозга выделили путем обработки в центрифуге К-70 при скорости вращения 2000 оборотов в минуту в течение 20 минут. Супернатант экстракта затем подвергли процедуре ультрафильтрации с помощью Amicon USA мембран ХМ-100, РМ-30, РМ-50. Согласно результатам анализа путем электрофореза главным продуктом является альбумин (см. рисунок 1).
Биохимическая очистка
Компоненты фракций экстракта костного мозга и белка были подвергнуты анализу на каждом этапе очистки посредством гелевого электрофореза в 10% полиакриламиде, содержащем 0,1% додецилсульфата натрия. До 7% додецилсульфата натрия и до 0,5-1% меркаптоэтанола добавили в образцы, которые подверглись инкубации в течение 5 минут при 70oС до закладки в гель.
Электрофорез проводили в 20-сантиметровом геле в течение пяти часов. Затем гель был окрашен в 0,25% Coomassie CBBC 250 в смеси этанол:вода:ацетиловая кислота 5:5:1 в течение одного часа при 20oС и промыты в нескольких переменах 7% ацетиловой кислоты. Активность продукта оценивалась методом ингибирования пролиферации стволовых кроветворных клеток (КОЕ-С). Способ подробно излагается далее.
Этап 1. Очистка путем образования осадка с сульфатом аммония.
Активность была очищена путем образования осадка с использованием сульфата аммония в 25% с насыщенностью 40-80%. Выбор насыщенности определяется на основе результатов, приведенных в Таблице 5.
Объем препарата, используемого для тестирования после каждого этапа очистки, определялся в соответствии с уровнем очистки и эквивалента дозе 2•10-2 мг исходного продукта. Активность определялась по формуле
%разницы=%Нa-Нв,
где % На представляет собой %Н в контрольной группе;
% Нв представляет собой %Н после инкубации с фракцией тестирования.
Фракция была опреснена с целью снижения концентрации сульфата аммония в 20 раз перед каждым тестированием активности и перед каждым последующим этапом очистки.
2 этап. Загрязненный ингибитор 1 этапа применяется после опреснения и фракционной утилизации ионообменной хроматографии, сюда же добавляют диэтиламиноэтилцеллюлозу (ДЭАЭ), целлюлозу 23, а затем элюируют в градиенте буфера ацетата натрия (рН 6,0).
Активные фракции элюируют в концентрации 3-5 мМ с получением ингибитора.
Объем колонны составил 1 мл, а скорость элюции 4 мл/час. Обнаружение проводилось с помощью хроматографа Милликром при 230 и 280 нм. Фракция 1 (смотри рисунок 2), которая демонстрирует наибольшую активность была выделена и элюирована в 5 мМ буфера ацетата натрия (см. Таблицу 6).
Данные электрофореза показывают, что главный загрязнитель белка - альбумин (см. рис.3) удаляется из этой фракции, что влечет за собой дополнительную четырехкратную очистку.
Этап 3. Частично очищенный ингибитор этапа 2 применяется непосредственно на колонну Сефедекс G-75.
Объем колонны составляет 20 мл (20 х1), норма элюирования составляет 2 мл/час. Буфер элюирования составляет 5 мМ NaCl, 10 mM Tris, pH 7,5. Обнаружение проводилось на хроматографе Миллихром при 230 и 280 нм. Была выделена фракция 5, обладающая наивысшей активностью.
Этап 4 обращенная хроматография (система Pharmacia FPLC), утилизирующая колонны PROREC, выполнялась на матрице Ультрасфера. Белок элюировали с помощью 0,1% трифторацетатной кислоты в градиенте ацетонитрил.
Однородность продукта с молекулярной массой 16-17 кД равна 90%, как это видно из анализа геля додецилсульфата акриламида натрия (см. рисунок 6). Результат показан на рисунке 4. Активность определяется по фракции 5. Окончательный выход продукта составил 5%. В результате объем белка с молекулярной массой 16 кД после очистки составил 650 нг/мл от исходного продукта. Во время процесса очистки продукт был подвергнут термической инкубации при 70oС в течение нескольких минут, однако потерь биологической активности зарегистрировано не было.
Пример 12В: Альтернативные способы выделения крупных объемов INPROL. Первоначальное выделение
Ребра свежей свиной туши очистили от мышечных волокон и жира, затем разрубили на куски и замочили в фосфат-буферном растворе соли в соотношении 1: 1 (вес/объем). Полученную смесь раздавили гидравлическим прессом с целью выделения костного мозга из твердого костного материала.
Суспензию клеток костного мозга собирают и фильтруют от твердых частиц через четыре слоя суровой марли. Фильтрат проходит инкубацию при 56oС в течение 40 минут, затем охлаждают в ледяной бане до 4oС. Полученный в результате процедуры осадок удаляют центрифугированием при 10000 g в течение 30 минут и затем сбрасывают.
Очищенный супернатант добавляют по капле в течение 30 минут к 10 объемам перемешанного холодного (ледяного) ацетата, содержащего 1% от объема концентрированной соляной кислоты. Образованную смесь хранят при 4oС в течение 16 часов для полного формирования осадка. Затем после центрифугирования при 20000 г в течение 30 минут при 4oС выпадает осадок. Этот осадок промывают ацетоном и высушивают.
Очистка ЖХВР (жидкостная хроматография высокого разрешения)
Осадок растворяют в А буфере элюэнта ЖХВР, содержащего 5% ацетонитрила (MeCN) и 0,1% трифлуорацетатной кислоты (ТФК), до окончательной концентрации белка 8-10 мг/мл. Этот раствор (0,5-0,6) загружают в колонну ЖХВР (250 х4,6 мм), упакованную с Полисил ODS-300 (10 мсм) и уравновешенную тем же самым А буфером.
Элюция завершается посредством градиента буфера В (90% MeCN, 0,1% TFA) в буфере А при скорости потока 1 мл/минуту в соответсвии со следующей программой:
Время, мин - %В
0 - 0
4 - 0
5 - 25
25 - 90
Дополнительный этап в 100% В в течение 5 минут используется для промывки колонны перед уравновешиванием. Затем колонну снова уравновешивают для возврата в исходное положение, после этого можно загрузить следующую порцию раствора белка. Типичная хроматограмма показана на рисунке 5.
В процессе выделения вытекающий поток отслеживается при 280 нм для обнаружения пиковых точек белка. Фракции, содержащие белковый материал, охлаждают, выделенные пики группируют в пулы и обрабатывают в испарителе роторного типа при 30oС до высушивания. Полученный остаток растворяют в дистиллированной воде и исследуют посредством теста на биоактивность и десенсибилизация - электрофорез в полиакриламидном геле (14% геля, восстанавливающие условия). Пик, содержащий активный материал, элюируют от 70 до 80% буфера В; он содержит бэнд белка 16 кД и следы более быстро двигающихся белков, как это видно из теста десенсибилизация - электрофорез в полиакриламидном геле. Анализ материала, полученного путем сбора только второго по величине пика жидкостной хроматографии высокого разрешения, показана на рисунке 15 (А и С). Материал, содержащий оба пика (например, рисунок 15), далее будет именоваться "Препарат pINPROL 1", а тот материал, который содержит только второй пик, далее будет именоваться "Препарат pINPROL 2". 500 г этого активного очищенного Препарата pINPROL 2 загрузили в колонну с обращенной фазой С4 (Vydac) и элюировали посредством линейного градиента 5,5% ацетонитрила в 0,1% трифлуороацетатной кислоты. Материал элюирован как единичный пик при 53% ацетонитрила (рисунок 15А). Когда, однако, в идентичные условия поместили 250 микрограмм MIP- альфа ( системы R&D), то он элюировал при 43,9% ацетонитрила (рисунок 15В), помните, что более ранние пики до 14% ацетонитрила являются артифактами и подвержены образованию воздущных пузырьков в датчике. Таким образом, INPROL, встречающийся в естественных условиях значительно более гидрофобен, чем MIP - альфа, при аналогичных условиях. Известно,что TGF-бета элюирует при более низких концентрациях чем те, при которых элюировал pINPROL при аналогичных условиях (Miyazono и др. J. Biol. Chem. 263: 6407-15,1988).
Гель элюированного материала pINPROL показан на рисунке 15С. Ряд 1 представляет собой сырой материал. Ряд 2 - маркеры молекулярной массы и Ряд 3 - очищенный материал. Можно заметить два крупных бэнда, один приблизительно в 14 кД и один приблизительно в 35 кД. Считают что бэнд в 35 кД является мультимерной формой бэнда в 14 кД.
Пример 13А: Активные Препараты INPROL содержат бета-цепь гемоглобина.
pINPROL был приготовлен, как показано на рисунке 5 (т.е. Препарат pINPROL 1, см. Пример 12В). Материал был подвергнут обработке в додецилсульфате натрия - электрофорез в полиакриламидном геле - и перенесен в нитроцеллюлозу посредством стандартной методики. Материал также подвергли анализу N-концевой последовательности посредством секвенатора белка ABI 477A с компьютерным анализатором РТН-АА 120А посредством стандартной методики. Получили следующую N-концевую последовательность:
VHLSAEEKEAVLGLWGKVNVVDEV....
Компьютерный поиск баз данных белка показал, что эта последовательность идентична N-концевой последовательности бета-цепи гемоглобина свиньи (смотри рисунок 16С).
Пример 13В: Активные Препараты INPROL содержат альфа-цепь гемоглобина.
Как показано на рисунке 15 С, белок, очищенный путем сбора второго по величине пика, показанного на рисунке 5 (например, Препарат pINPROL 2 ) имел следствием появление двух крупных бэндов, соответствующих молекулярной массе приблизительно 15 кД и 30 кД, а также нескольких мелких бэнда. Гели додецилсульфат натрия -электрофорез в полиакриламидном геле перенесли в нитроцеллюлозу посредством стандартной методики, а индивидуальные бэнды были подвергнуты эксцизии и анализу N-концевой последовательности, как это показано в примере 13А. Следующая N-концевая последовательность была получена для бэнда 15 кД:
VLSAADKANVKAAWGKVGGQ......
Бэнд 30 кД подвергли протеолитическому перевару и получили следующую внутреннюю последовательность:
**FPHFNLSHGSDQVK
Первая последовательность отражает идентичность с с N-концевой последовательностью альфа-цепи гемоглобина свиньи, в то время как вторая последовательность отражает идентичность с остатками 43-45 альфа-цепи гемоглобина (смотри на рисунке 16С последовательное сравнение альфа- и бета-цепей гемоглобина человека, мыши и свиньи). Повторяющаяся последовательность этих бэндов, также как некоторых других незначительных бэндов, постоянно вырабатывала порции последовательности альфа глобин. Таким образом, различные бэнды, видимые на рисунке 15 С, представляют собой фрагменты или совокупность альфа-цепи гемоглобина свиньи.
Пример 13 С: Продолжение характеристики INPROL свиньи.
Для дальнейшего сравнения гемоглобина pINPROL свиньи, у фирмы Сигма Кемикэл получили дважды кристаллизованный гемоглобин свиньи. Его подвергли жидкостной хроматографии высокого разрешения обращенной фазы, как это показано на рисунке 15. Как это видно на рисунке 17, хроматограмма жидкостной хроматографии высокого разрешения неповрежденного гемоглобина похожа на хроматограмму, составленную для Препарата pINPROL I. Далее при прямом сравнении Препарат pINPROL 2, показанный на рисунке 17А (то есть полученный из второго пика на рисунке 5) перекрывает два другие пика гемоглобина свиньи (рисунок 17В) со временем удерживания в 52.51 и 52.25 минут для наиболее значительных пиков соответственно. Следует отметить, что гем также перемещается с самым крупным пиком гемоглобина, в данном случае при 49.153 минутах; таким образом, гем является компонентом Препарата pINPROL 2. Это подтверждает отсутствие абсорбции этого Препарата pINPROL 1 при 575 нм при диагностике по длине волны на наличие гема.
Спрогнозированная молекулярная масса альфа- и бета-цепей гемоглобина составляет 15038 дальтон и 16034 дальтон. Как это видно из хроматограммы додецилсульфат натрия - электрофорез в полиакриламидном геле на рисунке 18, два первых пика состоят из цепи более высокой молекулярной массы, а вторые два составлены из цепи более низкой молекулярной массы. Таким образом, первые два пика представляют собой бета-цепи гемоглобина, а вторые два представляют собой альфа-цепи гемоглобина.
Дополнительное выделение гемоглобина свиньи было выполнено с помощью градиента поверхностной элюции (рисунок 21). N-Концевые анализы этих пиков продемонстрировали, что первый пик является альфа-цепью свиньи, а второй пик
является бета-цепью свиньи. Результаты биологического анализа подтвердили, что обе выделенных цепи гемоглобина биологически активны (например, это видно из примеров 14 и 15).
Для дальнейшего сравнения Препарата pINPROL 2 и бета-цепи гемоглобина был проведен двухмерный электрофорез (рисунок 19). В качестве первого измерения было проведено изоэлектрофокусирование в стеклянных трубках с помощью 2% (рН 4-8) амфолина в объеме 9600 вольт-часов. В качестве внутреннего стандарта использовали тропомиозин ( молекулярная масса 33 кД, pINPROL 5.2); его позиция помечена стрелкой на окончательном геле 2Д. Гель в трубке был уравновешен в буфере и герметизирован до верхней части концентрирующего геля в верхней части 12,5% акриламидного геля в присутствии додецилсульфата натрия. Электрофорез в гелевом блоке в присутствии додецилсульфата натрия проводили в течение 4 часов при 12,5 мА/гель. Гели были выкрашены в серебристый цвет и высушены.
Сравнение образцов электрофореза показало, что только одно или два пятна составляют разницу между очищенной бета-цепью гемоглобина жидкостной хроматографией высокого разрешения и pINPROL2. Анализы проводившиеся на Западе, проводившиеся посредством антисвиных антител гемоглобина и электрофорезов 1Д и 2Д, подтвердили присутствие бета-цепи гемоглобина в препарате. Таким образом, становится очевидным, что активный pINPROL2, приготовленный согласно процедуре приведенной в Примере 12В, является бета-цепью гемоглобина свиньи.
Пример 14: Альфа-цепь гемоглобина, бета-цепь гемоглобина или интактный гемоглобин обнаруживают ингибирующую деятельность стволовой клетки.
Для подтверждения факта, что бета-цепь гемоглобина обладает активностью INPROL, был проведен анализ по методике "самоубийства" используя костный мозг, взятый у мышей, подвергнутых лечению тестостероном, на основе методики, описание которой приведено в Примере 1, используя материал, очищенный таким же образом, как в Примере 12В. Как это видно из Таблицы 8, 15% клеток костного мозга нормальной мыши было нейтрализовано по сравнению с 36% животных, подвергнутых лечению тестостероном. Как и ожидалось, был подтвержден факт, что лечение тестостероном увеличивает процент клеток в цикле (таким образом, более подверженных нейтрализации, например, см. Пример 1). В противоположность этому, анализ клеток животных, подвергнутых лечению тестостероном, которые прошли инкубацию с pINPROL или с очищенной бета-цепью гемоглобина при 40 нг/мл, отразил снижение процента клеток в цикле с 36% до 0% и до 7% соответственно. Чем выше была доза 200 нг, тем менее эффективна она была для обоих белков. В качестве позитивной контрольной группы, охарактеризованный выше ингибитор стволовой клетки MIP-lα снизил цикл до 13%.
Похожий анализ был проведен in vitro, используя статус цикличности КОЕ-МИКС вместо КОЕ-С. Анализ был проведен согласно вышеприведенной процедуре для анализа КОЕ-С, за исключением того, что вместо тримидина, меченного тритием высокой дозировки (смотри B.I.Lord in HAEMOPOIESIS - F Practical approach. N.G. Testa and G. Molineux (Eds.), IRL Press 1993. Pragnell и др. In Culture of Hematopoietic Cells, R.I.Freshney, I.B. Pragnell и M.G.Freshney (Eds), Wiliy Liss 1994) и вместо мышей BDF1, подвергнутых воздействию тестостерона, используется штамм мыши с высокими эндогенными скоростями цикла Balb/c. Как это видно из Таблицы 9, бета-цепь свиньи высокой активности, как бета-цепь свиньи высокой очистки, так и альфа цепь свиньи высокой очистки отличаютя активностью при этом анализе. Следует отметить, что в этом анализе уровни цикла для клеток, подвергнутых лечению pINPROL, иногда имеют отрицательные числа, указывая, что в пуле, подвергнутом лечению Аrас, оказалось больше колоний, чем в пуле, лечение которого не проводилось.
Как это указано в Примере 2, pINPROL ингибирует пролиферацию линии стволовых клеток мыши FDCP-MIX в анализе поглощения тримидина, меченного тритием. На рисунке 20 видно, что как альфа-, так и бета-цепи гемоглобина проявляют активность при анализе, при ингибированиии, наблюдаемом при <2 нг/мл.
Вышеупомянутые данные свидетельствуют о том, что бета-цепь гемоглобина свиньи проявляет активность INPROL. Другие данные (например, Таблица 9, рисунок 20) показывают, что выделенная альфа-цепь, также как интактный гемоглобин, является активными ингибиторами стволовой клетки. Активные препараты также включают смешанные альфа- и бета-цепи (например, рисунок 5).
Наблюдения, в результате которых было определено, что выделенная цепь альфа глобин и/или выделенная цепь бета глобин отличаются активностью, указывают на то, что упомянутая здесь активность не требует интактной трехмерной структуры гемоглобина. Фрагменты альфа- и бета-цепи гемоглобина являются также активными в качестве ингибиторов стволовой клетки.
Пример 15: Очищенный INPROL, очищенная альфа-цепь гемоглобина свиньи или очищенная бета-цепь гемоглобина свиньи являются активными при методе in vivo.
Для того, чтобы проверить способность очищенных цепей гемоглобина свиньи действовать in vivo, мышей BDF1 инъецировали тестостероном пропионатом согласно процедуре, приведенной в Примере 1. Спустя двадцать четыре часа мыши получили 500 нг либо pINPROL, альфа-цепи гемоглобина (очищенного от периферических красных клеток крови, как это показано на рисунке 21), бета-цепи гемоглобина(очищенного от периферических красных клеток крови, как это показано на рисунке 21) или эквивалентный объем носителя внутривенно. Спустя сорок восемь часов собрали костный мозг каждой мыши и провели анализ КОЕ-МИКС согласно процедуре, приведенной в Примере 14. Как это видно из Таблицы 10, pINPROL, альфа-цепь гемоглобина свиньи, бета-цепь гемоглобина свиньи проявили активность in vivo, снижая процент КОЕ-МИКС в цикле до базальных уровней.
Пример 16: Очищенная альфа-цепь гемоглобина человека, биотинимированная альфа-цепь гемоглобина человека, биотинимированная бета-цепь гемоглобина человека, биотинимированная гамма-цепь гемоглобина человека, биотинимированная дельта-цепь гемоглобина человека демонстрируют in vitro ингибирующую активность стволовой клетки.
Гемоглобин человека был получен либо в химической корпорации "Сигма", либо был выделен посредством стандартной методики из периферической или умбиликальной пуповинной крови человека. Индивидуальные цепи были выделены посредством жидкостной хроматографии высокого разрешения обращенной фазы, таким же образом, как и в процедуре, описанной для альфа- и бета-цепей (смотри стр. 56) (смотри В. Masala и L.Manka, Methods in Enzymology том 231, стр. 21-44, 1994). Были получены очищенные альфа-, бета-, гамма- и дельта-цепи. Для биотинимированной альфа- и бета- цепей 1 мг гемоглобина взрослого человека подвергли воздействию 37 микрограммов NHS LC Биотин (проникающий) и цепями, выделенными посредством упомянутой выше хроматографии обращенной фазы.
Как это видно из Таблиц 11, 12 и 13, все очищенные альфа-цепи гемоглобина человека, биотинимированные альфа-цепи гемоглобина человека, биотинимированные бета-цепи гемоглобина человека, гамма-цепи гемоглобина человека и дельта-цепи гемоглобина человека являются активными в анализе цикла КОЕ-МИКС.
Пример 17: Очищенная альфа-цепь гемоглобина человека, димер или гемоглобин цепи альфа-бета являются активными in vivo.
Очищенная альфа-цепь гемоглобина человека, димер или гемоглобин цепи альфа-бета были исследованы при анализе in vivo согласно процедуре, приведенной в Примере 15. Каждый из них проявил активность при концентрации 500 нг/мышь.
Пример 18: INPROL свиньи активен в одноядерных клетках человека или в клетках CD 34+ пуповинной крови человека in vitro.
Для исследования способности очищенного INPROL костного мозга свиньи воздействовать на цикл клеток-предшественников человека были получены клетки умбиликальной пуповинной крови. Использовались либо общая фракция одноядерной клетки, полученной после выделения из фиколл, либо фракция CD 34+ (фирма "CellPro Inc."). Клетки подвергли инкубации в течение сорока восьми часов in vitro в присутствии интерлейкина 3 (IL-3) и фактора стволовой клетки (100 нг/мл каждый) для обеспечения присутствия ранних стволовых клеток в цикле. После такой предварительной инкубации провели анализы циклов согласно процедуре, приведенной в Примере 14 для мыши, за исключением того, что КОЕ-ГЕММ (вместо КОЕ-МИКС ) были подсчитаны на 18 день после высеивания. Как это видно из Таблицы 14, ингибирующий цикл INPROL свиньи КОЕ-ГЕММ находится в каждой из основных одноядерных клеток или во фракции CD 34+.
Пример 19: Гемоглобин очищенной альфа-цепи характеризуется активностью в КОЕ-ГЕММ человека.
Одноядерные клетки умбиликальной пуповинной крови человека были получены и прошли инкубацию в IL-3 и фактор стволовой клетки, и использовались в анализе цикла согласно процедуре, приведенной в Примере 18. Как это видно из Таблицы 15, как очищенный INPROL из костного мозга свиньи, так и гемоглобин альфа цепи человека, очищенный от переферической крови, проявили активность в ходе такого анализа.
Пример 20: Пептиды, полученные из последовательностей альфа-цепи и бета-цепи гемоглобина человека характеризуются активностью.
Для идентификации активных пептидных последовательностей трехмерная структура миеглобина (которая отличается активностью в настоящем анализе) была привнесена в естественную трехмерную структуру альфа-цепи, присутствующей в гемоглобине взрослого человека с помощью программы компьютерного моделирования. Два пептида (представляющие аминокислоты 43-55 и 64-82) которые являются участками, структурно отличающимися от миеглобина в трехмерном пространстве) были идентифицированы как активные в анализе цикла КОЕ-МИКС. Для того, чтобы приблизить звено, обнаруженное в естественной альфа-цепи, синтезировали циклический дериват пептида 43-45 (с43-55) (утилизирующий дисульфидную связь), который оказался активным.
Последовательность этих пептидов приведена ниже:
43-55 Phe- Pro-His-Phe-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val
c(43-55)Cys-Phe-Pro-His-Phe-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val-Cys
(при этом два остатка Cys связаны дисульфидно)
64-82 Asp-Ala-Leu-Thr-Asn-Ala-Val-Ala-His-Val-Asp-Asp-Met-Pro-Asn-Ala-Leu-Ser-Ala
Две последовательности геморфина, геморфин 10 (аминокислоты 32-41 последовательности бета-цепи) и геморфин 7 (аминокислоты 33-40) были исследованы и найдены активными.
Последовательности приводятся ниже:
Геморфин 10 Leu-Val-Val-Tyr-Pro-Trp-Thr-Gln-Arg-Phe
Геморфин 7 Val-Val-Tyr-Pro-Trp-Thr-Gln-Arg
Для тестирования активности этих последовательностей был проведен анализ цикла КОЕ-МИКС, как это видно из Примера 14. Как это видно из Таблицы 16-18, все эти пептиды характеризуются активностью в проведенном анализе.
Описание настоящего изобретения проводилось с точки зрения предпочтительных сфер применения, однако, вполне понятно, что специалисты могут вносить в него изменения и модификации. Поэтому мы предполагаем посредством приведенных ниже пунктов формулы охватить все такие эквивалентные изменения в пределах настоящего изобретения, то есть в том его виде, в котором оно было заявлено.
Относящиеся к изобретению документы
Категория. Наименование документа с указанием, где это уместно, номеров пунктов, относящихся к формуле изобретения
Y. "Природа", т. 344, издание от 29 марта 1990 года, Грэхэм и др. "Идентификация и характеристика ингибитора пролиферации кроветворной стволовой клетки", стр.442-444, смотри весь документ. 1-46
Y. WO, A, 91/04274 (фирма "Дженетикс Инститьют инк.", 4 апреля 1991 года, смотри весь документ. 1-46
A. Клетки крови, т.14, издание 1988 года, Ивз и др. "Поддержание жизнеспособности и контроль пролиферации примитивных кроветворных клеток-предшественников в культурах дексеровского типа костного мозга человека", стр. 355-368, смотри весь документ. 1-46
A. "Кровь", том 55, издание 1980 года, Токсоз и др. "Регулирование гемопоэза в культурах декстеровского типа костного мозга" II. Моделирование Ингибирования пролиферации стволовой клетки, стр. 931-936, смотри весь документ. 1-46
А. "Ингибиторы гемопоэза", том 162, издание 1987 года, Лорд и др., "Ингибитор пролиферации кроветворной КОЕ-С: анализы, производственные ресурсы и регулирующие механизмы", стр. 227-239, смотри весь документ. 1-46
A. J. Exp. Med., том 170, издание 1989 года, Броксмейер и др., "Результаты усиления миелопоэза воспаленных белках 1 и 2 макрофаги мыши на колониеобразование in vitro посредством клеток-предшественников гранулоцита/макрофаги костного мозга человека и мышей", стр.1583-1594, смотри весь документ. 1-46
A. Br.J.Paк, том 50, издание 1984 года, Теджеро и др. "Клеточная специфичность регуляторов пролиферации кроветворных стволовых клеток", стр. 335-341, смотри весь документ. 1-46
A. "Клетки крови", том 6, издание 1980 года, Лорд и др. "Источники пролиферации кроветворных стволовых клеток: стимуляторы и ингибиторы", стр. 581-593, смотри весь документ. 1-46т
| название | год | авторы | номер документа |
|---|---|---|---|
| ИНГИБИТОР И СТИМУЛЯТОР ПРОЛИФЕРАЦИИ СТВОЛОВОЙ КЛЕТКИ И ИХ ИСПОЛЬЗОВАНИЕ | 1997 |
|
RU2292352C9 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ЭКСПРЕССИРУЮЩИЕ EPO ЧЕЛОВЕКА | 2015 |
|
RU2799086C2 |
| УСТОЙЧИВЫЕ К РИТУКСИМАБУ ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2816370C2 |
| ИНГИБИТОР ДИФФЕРЕНЦИРОВКИ КРОВЕТВОРНЫХ КЛЕТОК-ПРЕДШЕСТВЕННИКОВ | 2006 |
|
RU2317074C1 |
| КОМБИНИРОВАННОЕ ПРИМЕНЕНИЕ ЛОВУШЕК GDF И АКТИВАТОРОВ РЕЦЕПТОРОВ ЭРИТРОПОЭТИНА ДЛЯ ПОВЫШЕНИЯ СОДЕРЖАНИЯ ЭРИТРОЦИТОВ | 2020 |
|
RU2814047C2 |
| ОПОСРЕДОВАННЫЙ ВИРУСОМ УСИЛЕННЫЙ ПЕРЕНОС ДНК | 1995 |
|
RU2174846C2 |
| ДОСТАВКА ПОЛЕЗНОЙ НАГРУЗКИ К СТВОЛОВЫМ КЛЕТКАМ | 2018 |
|
RU2795155C1 |
| ИММУНОМОДУЛИРУЮЩИЕ ОЛИГОПЕПТИДЫ | 2007 |
|
RU2503684C2 |
| ЛЕЧЕНИЕ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО РЕЦЕПТОРА АНТИГЕНА ПРОТИВ CD19 | 2015 |
|
RU2815417C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ СВЯЗАННЫХ С BCMA РАКОВЫХ ЗАБОЛЕВАНИЙ И АУТОИММУННЫХ РАССТРОЙСТВ | 2018 |
|
RU2799762C2 |











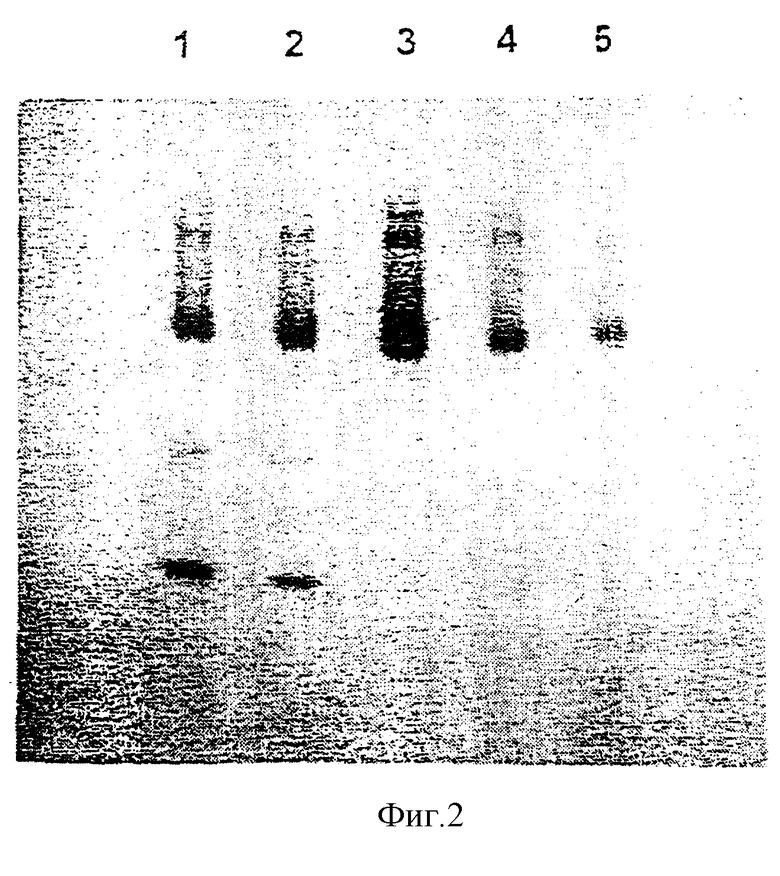


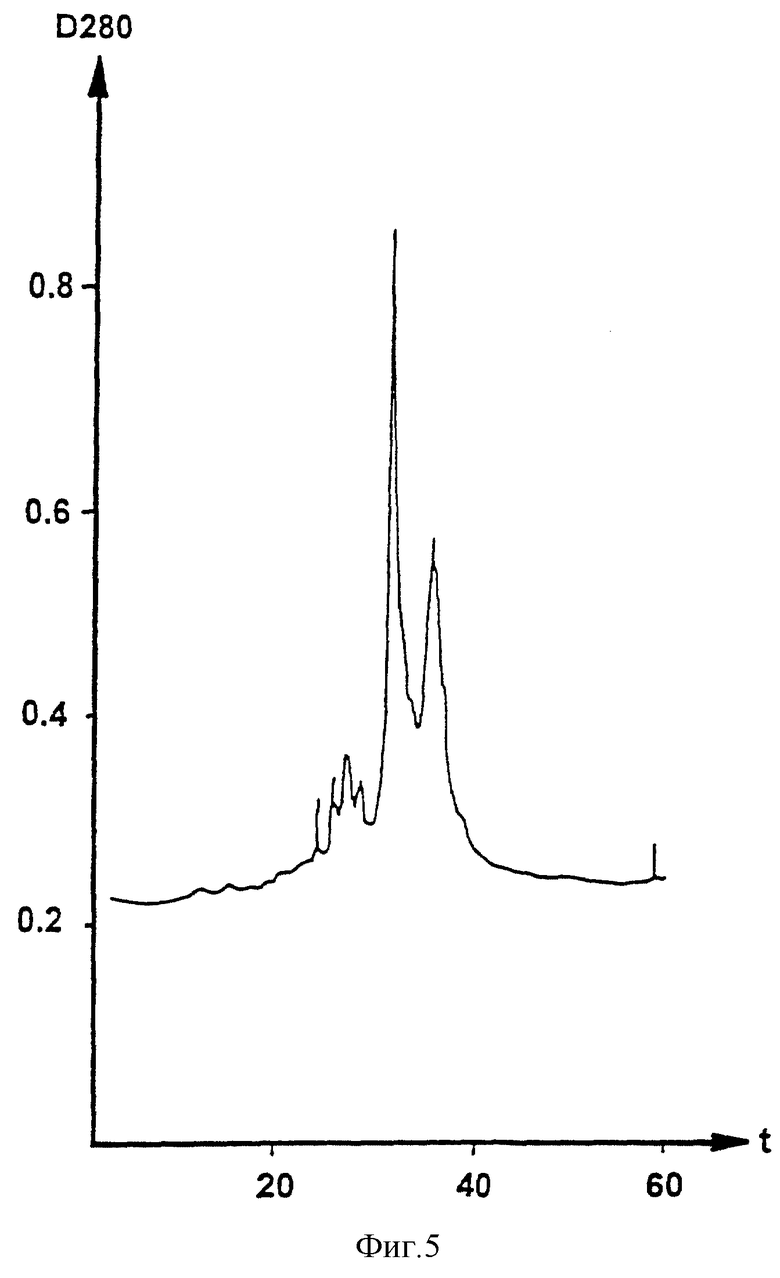




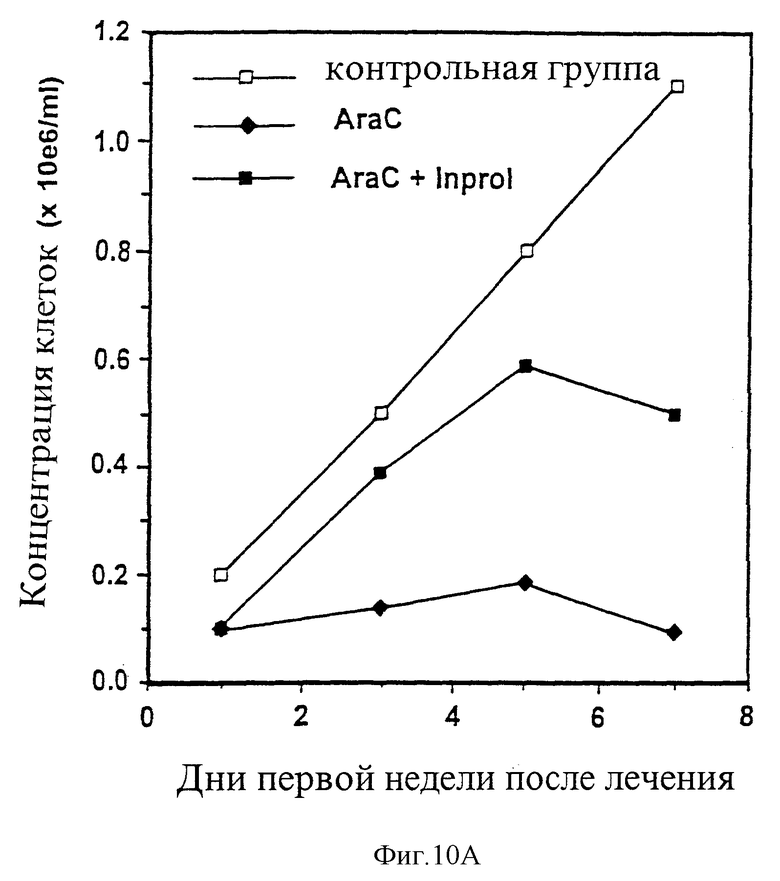


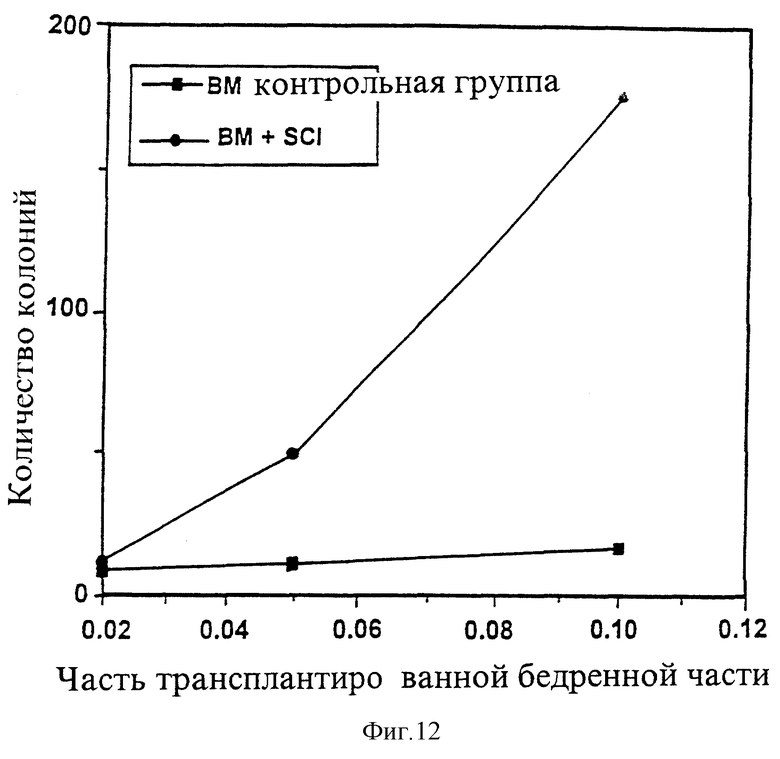

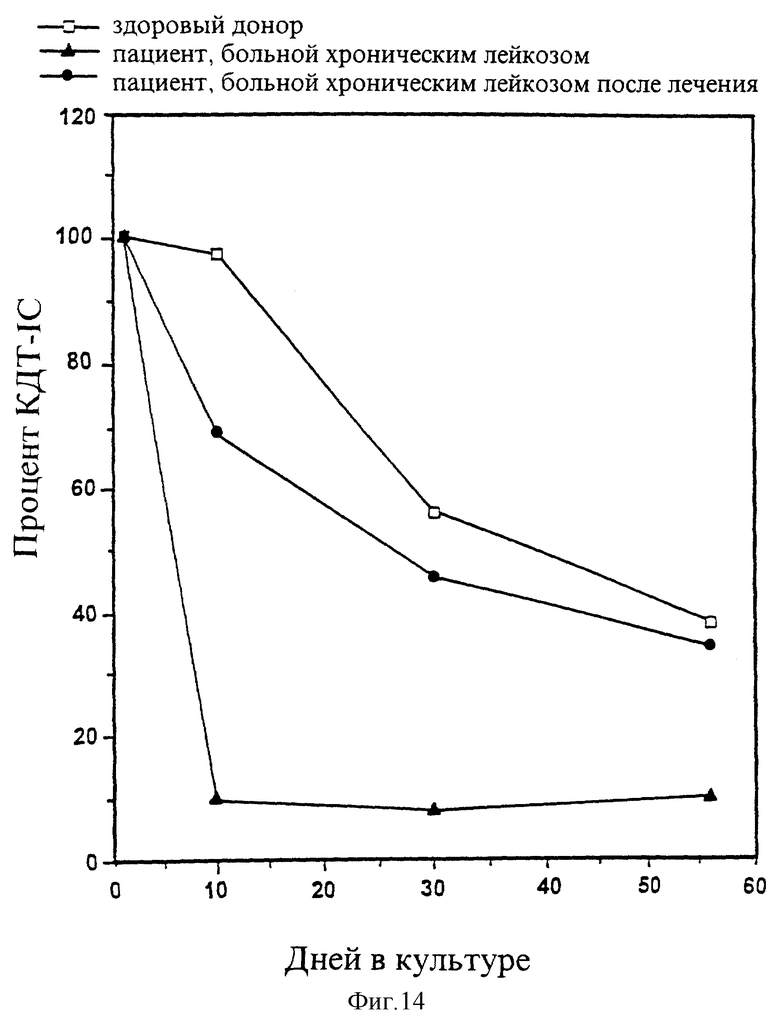
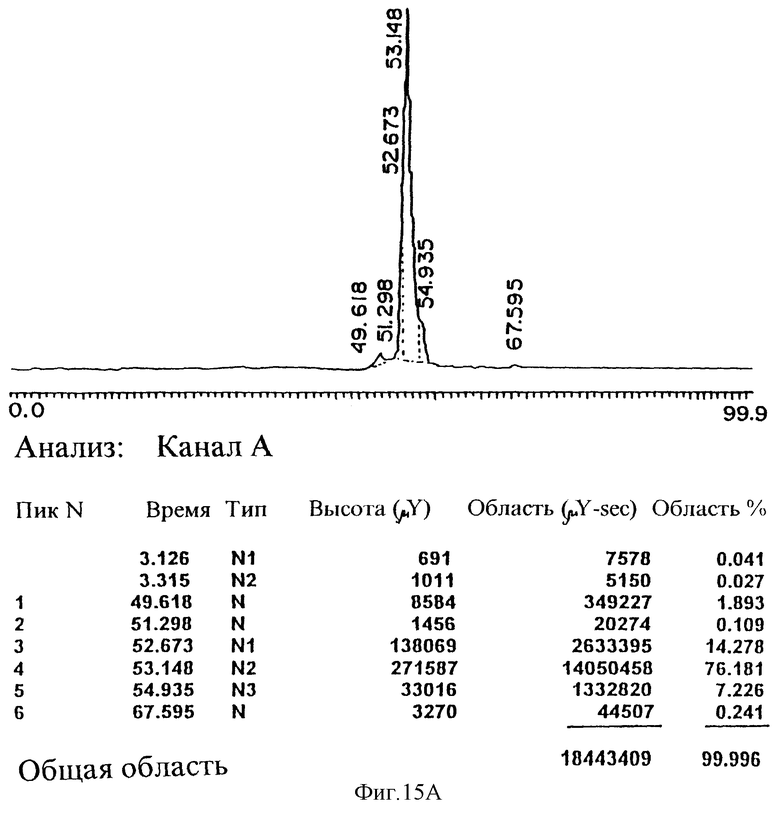





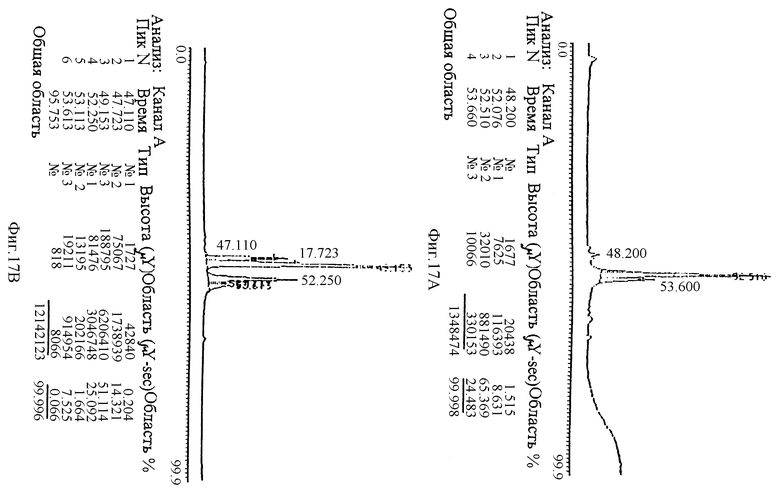
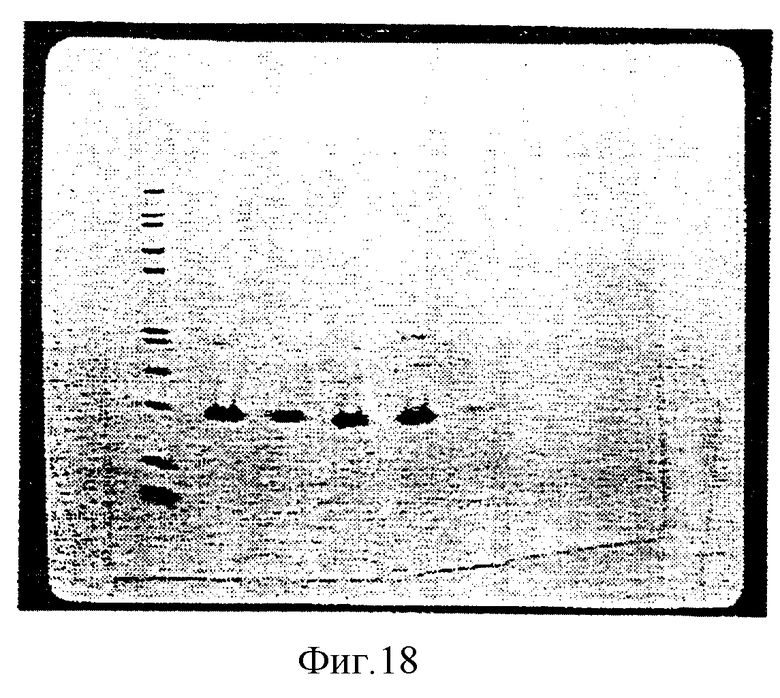
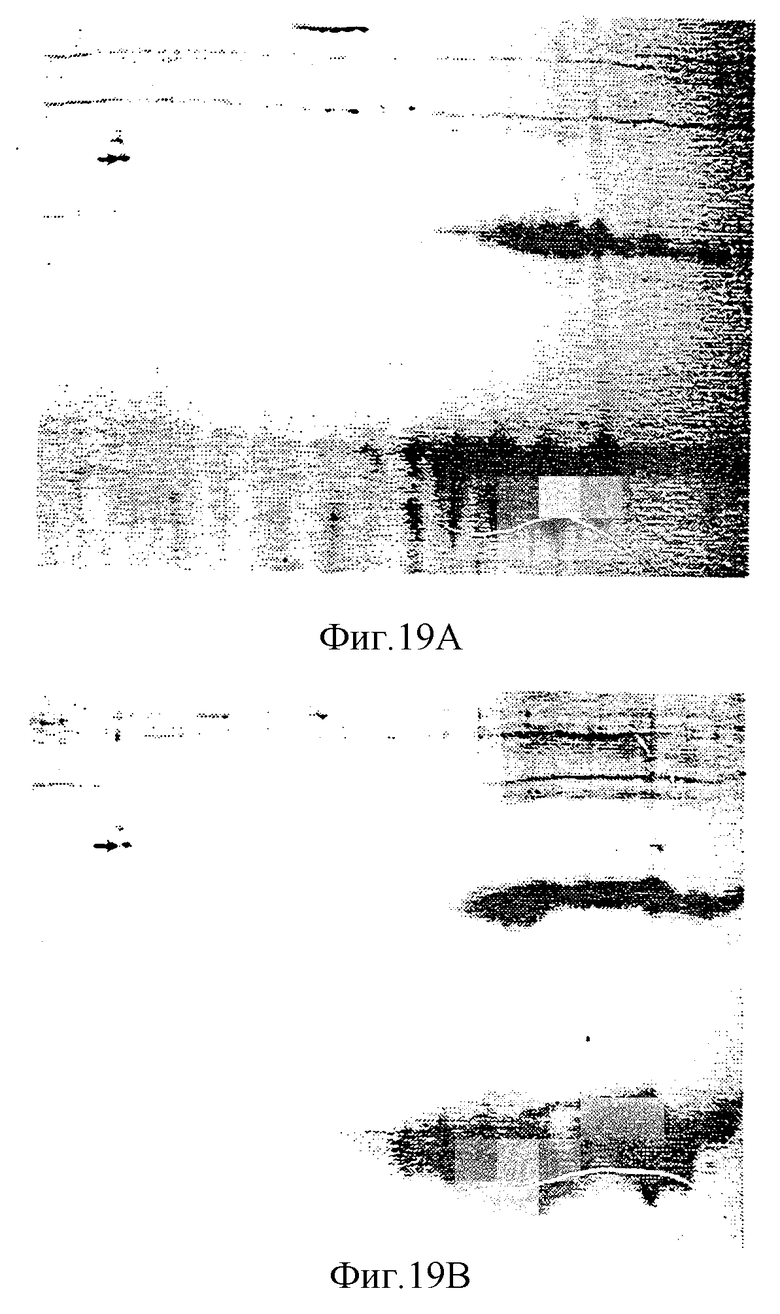


Изобретение относится к области медицины. Сущность изобретения состоит в раскрытии способов выделения и использования ингибирующих факторов стволовой клетки для регулирования цикла ненормальной стволовой клетки и акселерации выздоровления клетки периферической крови в период и после периода химиотерапии. Раскрыты также способы лечения и вакцинации. В изобретение также входят ингибиторы пролиферации стволовой клетки. Технический результат - расширение арсенала средств, ингибирующих пролиферацию стволовых клеток. 32 с. и 22 з.п.ф-лы, 19 табл., 21 ил.
Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Als-Gln-Val,
Cys-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val-Cys (в которой два остатка Cys образуют дисульфидную связь),
Asp-Ala-Leu-Thr-Asn-Ala-Val-Ala-His-Val-Asp-Asp-Met-Pro-Asn-Ala-Leu-Ser-Ala,
Leu-Val-Val-Tyr-Pro-Trp-Thr-Gln-Arg-Phe,
Leu-Val-Val-Tyr-Pro-Trp-Thr-Gln-Arg,
Leu-Val-Val-Tyr-Pro-Trp-Thr-Gln,
Leu-Val-Val-Tуr-Pro-Pro-Тhr,
Leu-Val-Val-Tyr-Pro-Trp,
Leu-Val-Val-Tyr-Pro,
Val-Val-Tyr-Pro-Trp-Thr-Gln,
Tyr-Pro-Trp-Thr-Gln-Arg-Phe,
Туr-Рrо-Тф-Thr-Gln-Arg,
Туr-Рrо-Тф-Thr-Gln, и
Туr-Рrо-Trp-Thr.
Приоритет по пунктам:
30.09.1994 по пп.2-4;
28.09.1995 по пп.1, 9-14, 16-19, 21, 26, 29, 32, 35, 38, 41, 44, 47-54.
| УСТРОЙСТВО ДЛЯ СНИЖЕНИЯ УРОВНЯ ВИБРАЦИИ ЖЕЛЕЗНОДОРОЖНОГО ПУТИ | 2002 |
|
RU2221911C2 |
| УСТРОЙСТВО для ПЕРЕВОЗКИ ДЛИННОМЕРНЫХ ТРУБ БОЛЬШОГО ДИАМЕТРА | 0 |
|
SU368092A1 |
| Торфодобывающая машина с вращающимся измельчающим орудием | 1922 |
|
SU87A1 |
| Огнетушитель | 0 |
|
SU91A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

