Изобретение относится к молекулярному клонированию генов, кодирующих рецептор трансферрина, а в частности к клонированию генов рецептора трансферрина Haemophilus influenzae.
Ссылка на родственную заявку
Настоящая заявка является частичным продолжением одновременно рассматриваемой заявки на патент США рег. 08/175116, поданной 29 декабря 1993, которая, в свою очередь, является частичным продолжением одновременно рассматриваемой заявки на патент США 08/148968, поданной 8 ноября 1993.
Предпосылки создания изобретения
Инкапсулированные штаммы Haemophilus influenzae типа b являются главными возбудителями бактериального менингита и других инвазивных инфекций у детей младшего возраста. Однако некапсулированные и нетипируемые бактерии H.influenzae (NTHi) ответственны за возникновение широкого ряда заболеваний у человека, включая воспаление среднего уха, эпиглоттит, пневмонию и трахеобронхит. Вакцины, изготовленные на основе капсульного полисахарида H. influenzae типа b, конъюгированного с дифтерийным токсоидом [Berkowitz и др., 1987. (В данной заявке, для более полного описания современного уровня науки в той области, к которой относится настоящее изобретение, приводятся ссылки (в скобках) на различные литературные источники. Полная библиографическая информация для каждой ссылки приводится в конце данного описания, непосредственно перед формулой изобретения. Раскрытие этих работ вводится в настоящее описание посредством ссылки)], столбнячным токсоидом (Classon и др. , 1989, и патент США 4496538), или с внешним мембранным белком Neisseria meningitidis (Black и др., 1991), показали свою эффективность при лечении менингита, индуцированного H. influenzae типа b, однако они оказались неэффективными при лечении NTHi-индуцированного заболевания (Bluestone, 1982).
Воспаление среднего уха (средний отит) является одним из наиболее распространенных заболеваний у детей младшего возраста; причем у 60-70% всех детей в возрасте моложе 2 лет было зарегистрировано 1-3 ушных инфекций. Хроническое воспаление среднего уха может приводить к нарушению слуха, речи, и познавательной способности у детей. Инфекции, вызываемые H. influenzae, примерно а 30% случаев являются причиной возникновения острого воспаления среднего уха и примерно в 60% случаев хронического воспаления среднего уха. Только в США на антибиотики для лечения воспаления среднего уха и на хирургические операции, связанные с этим заболеванием, такие как тонзиллэктомия, аденоидэктомия, и введение тимпаностомических трубок, затрачивается 1-2 миллиардов долларов в год. Кроме того, многие микроорганизмы, возбудители среднего отита, приобретают устойчивость к лечению антибиотиками. Поэтому разработка эффективной профилактической вакцины против среднего отита имеет исключительно важное значение. Нетипируемые штаммы H. influenzae являются также главными патогенами, ответственными за возникновение пневмонии у людей пожилого возраста, и других индивидуумов, являющихся особенно восприимчивыми к респираторным инфекциям. В связи с этим очевидно, что необходимо получить такие антигены от H. influenzae, которые можно было бы использовать в качестве компонентов в иммуногенных препаратах, предназначенных для защиты против многих серотипов H. influenzae.
Железо является главным питательным элементом, необходимым для роста многих бактерий. Некоторые патогены человека, такие как H. influenzae, Branhamella cattaraliis, n. meningitidis, n. gonorrhocae и непатогенные комменсальные штаммы neisseria могут использовать трансферрин человека в качестве источника железа (Schryvers, 1988; Schryvers и Zee 1989; Micklsen и Sparling, 1981). Рецептор бактериального трансферрина (TfR) состоит из двух цепей, Tbp1 и Tbp2. В штаммах H. influenzae, молекулярная масса Tbp1 составляет приблизительно 100000, а молекулярная масса Тbp2 варьирует в пределах от 60000 до 90000 в зависимости от штамма (Schryvers и Gray-Owen, 1992; Holland и др., 1992). Экспрессия рецептора трансферрина H. influenzae является, очевидно, железо- и/или геминрегулируемой (Norton и др., 1993), а предполагаемый сайт fur-связывания, как было идентифицировано, находится выше tbp2. Эта последовательность была обнаружена в промоторной области генов, негативно регулируемых железом, включая ген TfR N. meningitidis (Zegrain и др., 1993).
Было установлено, что в других бактериальных TfR-оперонах, за промотором следуют гены tbp2 и tbp1 (Zegrain и др., 1993; Wilton и др., 1993). Антитела, которые блокируют доступ рецептора трансферрина к его источнику железа, могут предотвращать бактериальный рост. Кроме того, антитела против TfR, которые являются опсонизирующим или бактерицидным фактором, могут также обеспечивать защиту от бактерий посредством альтернативных механизмов. Таким образом, рецептор трансферрина, его фрагменты, его составляющие цепи или белки, происходящие от этого рецептора, являются кандидатами на использование их в целях изготовления вакцины для защиты от инфекций, вызываемых H. influenzae. Мыши, иммунизированные белками TfR n. meningitidis в адьюаанте Фрейнда, обнаруживали резистентность к гомологичной антигенной стимуляции, а антисыворотки против TfR обладали бактерицидным и протективным действием в анализе на пассивный транспорт (Danve и др., 1993). Свиньи, иммунизированные рекомбинантным Tbp2 A. pleuropneumoniae, обнаруживали устойчивость к гомологичной антигенной стимуляции, но не обнаруживали устойчивость к гетерологичной стимуляции (Rossi-Campos и др., 1992). Эти данные свидетельствуют о том, что вакцина, полученная на основе TfR, обладает защитным действием. При этом, было бы желательно получить последовательности ДНК-молекулы, кодирующие рецептор трансферрина и пептиды, соответствующие определенным частям рецептора трансферрина, а также векторы, содержащие такие последовательности, в целях использования их для диагностики, иммунизации, и продуцирования диагностических иммунологических реагентов.
Полиовирус представляет собой энтеровирус, род вируса, принадлежащий к семейству Пикорнавирусов. Существует три различных серотипа полиоаирусов, и кроме того, имеется множество штаммов, принадлежащих к каждому из этих трех серотипов. Вирулентные штаммы являются возбудителями паралитического полиомиелита. Аттенюированные штаммы, т.е., штаммы, обладающие пониженной способностью к индуцированию паралитического заболевания, и инактивированные вирулентные штаммы были использованы в качестве вакцин. Инфицирование вирусом индуцирует стойкий протективный иммунитет слизистых оболочек. Вакцинация с использованием вакцин на основе инактивированного полиовируса может также индуцировать иммунный ответ, опосредованный слизистой оболочкой.
Структура полиовируса известна и является в высокой степени консервативной для различных штаммов и серотипов. Были также определены структуры некоторых других пикорнавирусов (вирусов, принадлежащих к семейству Пикорнавирусов), и было также установлено, что по своей структуре эти пикорнавирусы являются близкородственными полиовирусам. Было показано, что можно экспрессировать чужеродные эпитопы на капсиде полиовирусов (Murdin и др., 1992), и этот опыт может быть распространен на другие пикорнавирусы. Эпитопы, которые были экспрессированы, представляли собой, в основном, короткие, хорошо определенные, смежные эпитопы; причем в большинстве своем они были экспрессированы в антигенном сайте I нейтрализации полиовируса (NAg1) или а экbивалентном сайте на других пикорнавирусах. Этот сайт включает а себя петлю, связывающую бета-цепи В и С (ВС-петлю) капсидного белка VPI полиовируса. ВС-петля белка VPI представляет собой поверхностно доступную петлю, состоящую из девяти аминокислот, которые могут быть заменены и к которым могут быть присоединены, по крайней мере, двадцать пять гетерологичных аминокислот (Murdin и др. , 1991). Гибридные или химерные полиовирусы, экспрессирующие эпитопы рецептора трансферрина, и культивированные до высокого титра, а также являющиеся иммуногенными, могли бы быть использованы в качестве вакцин и как средство для продуцирования иммунологических реагентов.
Краткое описание изобретения
Настоящее изобретение относится к получению очищенных и выделенных молекул нуклеиновой кислоты, кодирующих рецептор трансферрина штамма Haemophilus, либо фрагмент или аналог белка рецептора трансферрина. Нуклеиновокислотные молекулы настоящего изобретения могут быть использованы для специфического обнаружения штаммов Haemophilus, а также для диагностики инфекций, вызываемых Haemophilus.
Выделенные и очищенные нуклеиновокислотные молекулы настоящего изобретения, такие как ДНК, могут быть также использованы для экспрессии генов TfR с применением техники рекомбинантных ДНК в целях экономически выгодного продуцирования очищенных и выделенных субъединиц рецептора трансферрина, его фрагментов, или аналогов. Рецептор трансферрина, его субъединицы, фрагменты или аналоги, а также кодирующие их молекулы нуклеиновой кислоты и векторы, содержащие эти молекулы нуклеиновой кислоты, могут быть использованы в иммуногенных композициях для защиты против заболеваний, вызываемых Haemophilus; для диагностики инфекций, вызываемых Haemophilus, а также как средство для продуцирования иммунологических реагентов. Моноклональные антитела или моноспецифические антисыворотки (антитела), вырабатываемые против белка рецептора трансферрина, продуцированного в соответствии с аспектами настоящего изобретения могут быть использованы для диагностики инфекций, вызываемых Haemophilus; для специфического обнаружения Haemophilus (например, в in vitro и in vivo-анализах); и для лечения заболеваний, вызываемых Haemophilus.
Пептиды, соответствующие определенным участкам рецептора трансферрина или его аналогам, могут быть использованы а иммуногенных композициях для лечения заболеваний, вызываемых Haemophilus, для диагностики Haemophilus-инфекций, а также как средство продуцирования иммунологических реагентов. Моноклональные антитела или антисыворотки, вырабатываемые против этих пептидов, полученных в соответствии с аспектами настоящего изобретения, могут быть использованы для диагностики Haemophilus-инфекций, для специфического обнаружения Haemophilus (например, в in vitro и in vivo-анализах) и для проведения иммунизации в целях лечения заболеваний, вызываемых Haemophilus.
В одном из своих аспектов настоящее изобретение относится к очищенной и выделенной молекуле нуклеиновой кислоты, кодирующей белок рецептора трансферрина, происходящего от штамма Haemophilus, в частности от штамма H. influenzae, а более конкретно от штамма H. influenzae типа b, такого как штамм DI63, Eagan или Minna H. influenzae типа b; или нетипируемого штамма H. influenzae, такого как штамм РАК 12085, SB33, SВ12, SВ29, SВ30 или SВ32 H. influenzae; либо фрагмент или аналог белка рецептора трансферрина.
В одном из предпочтительных вариантов осуществления изобретения молекула нуклеиновой кислоты может кодировать либо белок Tbp1, либо белок TBp2 штамма Haemophilus. В другом предпочтительном варианте осуществления настоящего изобретения нуклеиновая кислота может кодировать фрагмент белка рецептора трансферрина, происходящего от штамма Haemophilus и имеющего аминокислотную последовательность, которая является консервативной для бактерий, продуцирующих белок рецептора трансферрина. Такая консервативная аминокислотная последовательность может иметь аминокислотную последовательность, содержащуюся в аминокислотной последовательности пептидов, показанных ниже, в таблицах 2 и 3, для штамма Eagan Haemophilus influenzae типа b, а также соответствующих пептидов других штаммов H. influenzae.
В другом своем аспекте настоящее изобретение относится к очищенной и выделенной молекуле нуклеиновой кислоты, имеющей ДНК-последовательность, выбранную из группы, включающей в себя: (а) любую из ДНК-последовательностей, представленных на фиг.3, 4, 5, 6, 7, 8, 9, 10 или 11 (SEQ ID 1, 2, 3, 4, 105, 108, 110, 112, 114), или ДНК-последовательность, комплементарную любой из указанных последовательностей; (b) ДНК-последовательность, кодирующую одну из аминокислотных последовательностей, представленных на фиг.3, 4, 5, 6, 7, 8, 9, 10 или 11 (SEQ ID 5, 6, 7, 8, 9, 10, 11, 12, 106, 107, 109, 111, 113, 115), или ДНК-последовательность, комплементарную указанной последовательности; и (с) ДНК-последовательность, которая гибридизуется в жестких условиях с любой из ДНК-последовательностей, определенных в (а) и (b). Предпочтительно, чтобы ДНК-последовательность, определенная в (с), по крайней мере, примерно на 90% была идентична любой из ДНК-последовательностей, определенных в (а) и (b).
В другом своем аспекте настоящее изобретение относится к вектору, адаптированному для трансформации хозяина и содержащему молекулу нуклеиновой кислоты, определенную выше. Таким вектором может быть любой вектор, имеющий характеристики плазмиды DS-712-1-3 (ATСC, входящий номер 75603) или плазмиды JB-1042-7-6 (ATСC, входящий номер 75607).
Эти плазмиды могут быть адаптированы для экспрессии кодируемого рецептора трансферрина, его фрагментов или аналогов в липидизированной или нелипидизированной форме в гетерологичном или гомологичном хозяине. В соответствии с этим в еще одном своем аспекте настоящее изобретение относится к экспрессирующему вектору, адаптированному для трансформации хозяина, содержащему молекулу нуклеиновой кислоты настоящего изобретения; и экспрессирующий элемент, правильно присоединенный к нуклеиновокислотной молекуле для обеспечения экспрессии хозяином белка рецептора трансферрина, или его фрагмента, или аналога. В конкретных вариантах осуществления этого аспекта настоящего изобретения молекула нуклеиновой кислоты может кодировать, в основном, весь белок рецептора трансферрина, либо белок Tbp1, либо белок Tbp2 штамма Haemophilus. Экспрессирующий элемент может включать в себя часть аминокислоты, кодирующую лидерную последовательность, для секреции из хозяина белка рецептора трансферрина, или его фрагмента, или аналога. Экспрессирующий элемент может также включать в себя часть нуклеиновой кислоты, кодирующую сигнал липидизации, для экспрессии в хозяине липидизированной формы белка рецептора трансферрина, или его фрагмента, или аналога. Экспрессирующая плазмида может иметь идентифицирующие признаки плазмиды JB-1468-29, JB-1600-1 или JB-1424-2-4. Хозяин может быть выбран, например, из Escherchia coli, Bacillus, Haemophilus, грибков, дрожжей или бакуловирусов; при этом могут быть также использованы экспрессирующие системы вируса лесов Семлики.
В другом своем аспекте настоящее изобретение относится к трансформированному хозяину, содержащему вектор экспрессии, определенный выше. Этот хозяин может быть выбран из JB-1476-2-1, JB-1437-4-1 и JB-1607-1-1. Кроме того, настоящее изобретение относится к белку рецептора трансферрина, или к его фрагменту, или аналогу, продуцируемому трансформированным хозяином.
Как подробно описывается ниже, рецепторные белки Tbp1 и Тbp2 были продуцированы отдельно друг от друга. Поэтому в еще одном своем аспекте настоящее изобретение относится к выделенному и очищенному белку Tbp1 штамма Haemophilus, отделенному от белка Тbр2 штамма Haemophilus и к выделенному и очищенному белку Тbр2 штамма Haemophilus, отделенному от белка Tbp1 штамма Haemophilus. В качестве штамма Haemophilus может быть использован штамм H. influenzae типа b или нетипируемый штамм H. influenzae.
Кроме того, настоящее изобретение относится к синтетическим пептидам, соответствующим отдельным участкам рецептора трансферрина. В связи с этим в другом своем аспекте настоящее изобретение относится к синтетическому пептиду, содержащему не менее шести аминокислот и не более 150 аминокислот и имеющему аминокислотную последовательность, соответствующую лишь части белка рецептора трансферрина или аналога белка рецептора трансферрина штамма бактерий. Предпочтительным бактериальным штаммом является штамм Haemophilus, в частности штамм H. influenzae, а более конкретно штамм H. influenzae типа b, или нетипируемый штамм H. influenzae.
Пептиды, полученные в соответствии с настоящим изобретением, могут содержать аминокислотную последовательность, которая является консервативной для бактерий, продуцирующих белок рецептора трансферрина, включая штаммы Haemophilus. Этот пептид может включать в себя аминокислотную последовательность LEGGFYGP (SEQ ID 74) или последовательность LEGGFYG (SEQ ID 85). Пептиды настоящего изобретения могут иметь аминокислотную последовательность, выбранную из последовательностей, представленных ниже (в таблице 2 или 3) для штамма Eagan H. influenzae типа b, и соответствующие аминокислотные последовательности для других штаммов H. influenzae.
В другом своем аспекте настоящее изобретение относится к иммунной композиции, которая содержит, по крайней мере, один активный компонент, выбранный, по крайней мере, из одной молекулы нуклеиновой кислоты, представленной в настоящей заявке; по крайней мере, один рекомбинантный белок, представленный в настоящей заявке; по крайней мере, один из очищенных и выделенных белков Tbp1 или Тbр2, представленных в настоящей заявке; по крайней мере, один синтетический пептид, представленный в настоящей заявке; и "живой" вектор, представленный в настоящей заявке; а также фармацевтически приемлемый носитель. При введении хозяину, по крайней мере, один активный компонент продуцирует иммунный ответ.
Иммуногенные композиции настоящего изобретения могут быть изготовлены в виде вакцины для введения in vivo в целях защиты организма от болезней, вызываемых бактериальными патогенами, которые продуцируют рецепторы для трансферрина. Для этих целей композиции могут быть изготовлены в виде микрочастиц, капсул или липосомных препаратов. Альтернативно композиции могут быть изготовлены в комбинации с молекулой, обеспечивающей направленную доставку композиции к специфическим клеткам иммунной системы или к поверхности слизистых оболочек. Иммуногенная композиция может содержать множество активных компонентов для защиты от болезней, вызываемых множеством видов бактерий, продуцирующих рецептор трансферрина. Иммуногенные композиции могут также содержать адъювант.
В другом своем аспекте настоящее изобретение относится к способу защиты от инфекции или заболеваний, вызываемых Haemophilus или другими бактериями, продуцирующими рецептор трансферрина; причем указанный способ предусматривает стадию введения восприимчивому хозяину, такому как человек, эффективного количества иммуногенной композиции, определенной выше.
В еще одном своем аспекте настоящее изобретение относится к антисыворотке или антителу против рекомбинантного белка; к выделенному и очищенному белку Tbp1 или Тbp2; к синтетическому пептиду или к иммунногенной композиции.
В следующем своем аспекте настоящее изобретение относится к "живому" вектору, осуществляющему доставку рецептора трансферрина в организм хозяина и содержащему молекулу нуклеиновой кислоты, определенную выше. Этот вектор может быть выбран из Salmonella, ВСG, аденовируса, поксвируса, вакцинного вируса (вируса коровьей оспы) и полиовируса. В частности, указанным вектором может быть полиовирус, а указанная молекула нуклеиновой кислоты может кодировать фрагмент рецептора трансферрина, имеющий аминокислотную последовательность EGGFYGP (SEQ ID 74) или последовательность EGGFYG (SEQ ID 85). Кроме того, настоящее изобретение включает в себя плазмидный вектор, имеющий идентифицирующие признаки плазмиды рТ7ТВР2А, рТ7ТВР2В, рТ7ТВР2С или рТ7ТВР21 (АТСС, 75931, 75932, 75933, 75934).
В еще одном своем аспекте настоящее изобретение относится к штамму Haemophilus, который не продуцирует белок рецептора трансферрина. Этот штамм может содержать ген, кодирующий рецептор трансферрина, который является функционально блокированным, например, путем инсерционного мутагенеза. Штамм Haemophilus может быть аттенюированным штаммом, и этот аттенюированный штамм может содержать вектор для доставки рецептора трансферрина.
Как указывалось выше, в одном своем аспекте настоящее изобретение относится к выделенному и очищенному белку Tbp1 или белку Тbр2 штамма Haemophilus, а предпочтительно штамма Haemophilus influenzae; причем указанные белки являются выделенными отдельно друг от друга. В соответствии с этим в еще одном своем аспекте настоящее изобретение относится к способу получения выделенного и очищенного белка Tbp1 или Тbр2 штамма Haemophilus, предусматривающему проведение следующих стадий: (а) получение рекомбинантного хозяина, экспрессирующего в тельцах включения, либо белок Tbp1, либо белок Тbр2 (но не оба); (b) культивирование хозяйских клеток в целях получения клеточного пула; (с) дезинтеграцию клеточного пула для получения клеточного лизата; (d) фракционирования клеточного лизата для получения первого супернатанта и первого осадка, где первый супернатант, в основном, содержит большое количество растворимых хозяйских белков; (е) отделение первого супернатанта от первого осадка; (f) селективное экстрагирование первого осадка для удаления, в основном, всех растворимых белков хозяина и мембранных белков хозяина с получением второго супернатанта и экстрагированного осадка, содержащего тельца включения; (g) отделение второго супернатанта от экстрагированного осадка; (h) солюбилизацию экстрагированного осадка для получения солюбилизированного экстракта и (i) фракционирование солюбилизированного экстракта с получением фракции, содержащей белок Tbp1 или Тbр2.
Клеточный лизат может быть фракционирован с получением первого супернатанта, а первый осадок может быть получен путем, по крайней мере, одной экстракции детергентом.
Солюбилизированный экстракт может быть фракционирован путем гель-фильтрации с получением фракции, содержащей либо белок Tbp1, либо белок Тbр2, которая может быть затем диализована для удаления, по крайней мере, детергента, в результате чего получают более очищенный раствор белка Tbp1 или Тbр2.
Краткое описание чертежей
Для лучшего понимания настоящего изобретения ниже приводится подробное его описание со ссылками на следующие чертежи, где:
На фиг.1А показана рестрикционная карта двух плазмидных клонов (pBHT1 и рВНТ2) оперона рецептора трансферрина штамма DL63 Haemophilus influenzae типа b.
На фиг.1В показана рестрикционная карта клонов S-4368-3-3 и JB-901-5-3, содержащих гены ТfR от штамма Eagan H. influenzae типа b.
На фиг. 1C показана рестрикционная карта клона DS-712-1-3, содержащего ген рецептора трансферрина от штамма MinnA H. influenzae типа b.
На фиг. 1D показана рестрикционная карта клона JB-1042-7-6, содержащего ген рецептора трансферрина от нетипируемого штамма РАК 12085 H. influenzae.
На фиг.2 проиллюстрированы организация и рестриционные карты клонированных генов Тbр1 и Тbр2 идентифицированных штаммов и генетическая организация TfR-оперона с двумя последовательно расположенными генами (Tbp1 и Tbp2), образующими оперон под транскрипционным контролем единственного промотора; и кроме того, на чертеже изображен 3,0 кb-ДНК-фрагмент плазмиды рВНIТ2, используемый для зондирования библиотек генов TfR от штаммов Haemophilus.
На фиг.3 показаны нуклеотидные последовательности генов рецептора трансферрина (SEQ ID 1) и их выведенные аминокислотные последовательности (SEQ ID 5 - Tbp1 и SEQ ID 6 - Tbp2) от штамма DL63 H. influenzae типа b. Подчеркнутые аминокислотные последовательности соответствуют пептидам Тbр1, идентифицированным путем аминокислотного секbенирования. Предполагаемые сигнальные последовательности показаны двумя линиями, проведенными сверху, и соответствуют остаткам 1-17 для Tbp1 и остаткам 1-25 для Тbр2.
На фиг.4 показаны нуклеотидные последовательности генов рецептора трансферрина (SEQ ID 2) и их выведенные аминокислотные последовательности (SEQ ID 7 - Tbp1 и SEQ ID 8 - Тbр2) от штамма Eagan H. influenzae типа b. Предполагаемые -35, -10 и последовательности сайта связывания рибосомы показаны линией, проведенной сверху.
На фиг.5 показаны нуклеотидные последовательности генов рецептора трансферрина (SEQ ID 3) и их выведенные аминокислотные последовательности (SEQ ID 9 - Tbp1 и SEQ ID 10 - Тbр2) от штамма MinnA H. influenzae типа b. Предполагаемые последовательности -35, -10 и последовательность сайта связывания рибосомы показаны линиями, проведенными сверху.
На фиг.6 показаны нуклеотидные последовательности генов рецептора трансферрина (SEQ ID 4) и их выведенные аминокислотные последовательности (SEQ ID 11 - Tbp1 и SEQ ID 12 - Тbр2) от нетипируемого штамма РАК 12085 H. influenzae. Предполагаемые последовательности -35, -10 и последовательность сайта связывания с рибосомой показаны линиями, проведенными сверху.
На фиг.7 показаны нуклеотидные последовательности генов рецептора трансферрина (SEQ ID 105) и их выведенные аминокислотные последовательности (SEQ ID 106 - Tbp1 и SEQ ID 107 - Тbр2) от нетипируемого штамма SB33 H. influenzae.
На фиг.8 показана нуклеотидная последовательность гена Тbр2 (SEQ ID 108) и его выведенная аминокислотная последовательность (SEQ ID 109 - Тbр2) от нетипируемого штамма SВ12 H. influenzae.
На фиг.9 показана нуклеотидная последовательность гена Тbр2 (SEQ ID 110) и выведенная аминокислотная последовательность (SEQ ID 111 - Тbр2) от нетипируемого штамма SВ29 H. influenzae.
На фиг. 10 показана нуклеотидная последовательность гена Тbр2 (SEQ ID 112) и его выведенная аминокислотная последовательность (SEQ ID 113 - Тbр2) от нетипируемого штамма SВ30 H. influenzae.
На фиг. 11 показана нуклеотидная последовательность гена Тbр2 (SEQ ID 114) и его выведенная аминокислотная последовательность (SEQ ID 115 - Тbр2) от нетипируемого штамма SВ32 H. influenzae.
На фиг.12А показаны нуклеотидные последовательности промоторных областей и 5'-конца генов tbp2, происходящих от штаммов Eagan (SEQ ID 116), MinnA (SEQ ID 117), РАК 12085 (SEQ ID 118) и SВ33 (SEQ ID 119). Смысловой праймер, используемый для амплификации генов tbp 2 с помощью PCR, подчеркнут (SEQ ID 120).
На фиг.12В показана нуклеотидная последовательность межгенной области и 5'-конца генов tbp 1, происходящих от штаммов H. influenzae, таких как Eagan (SEQ ID 121), MinnA (SEQ ID 122), DL63 (SEQ ID 123), РАК 12085 (SEQ ID 124), SВ12 (SEQ ID 125), SB29 (SEQ ID 126), SВ30 (SEQ ID 127) и SВ32 (SEQ ID 128). Антисмысловой праймер, используемый для амплификации генов с помощью PCR, подчеркнут (SEQ ID 129).
На фиг.13 проиллюстрирован анализ методом электрофореза на агарозном геле PCR-амплифицированных генов tbp 2, происходящих от нетипируемых штаммов H. influenzae, таких как: SВ12, SВ29, SB30, SВ32 и SB33. Дорожка 1 - SB33; дорожка 2 - SВ12, дорожка 3 - SВ29, дорожка 4 - SB30, дорожка 5 - SВ32.
На фиг.14 проиллюстрировано сравнение аминокислотных последовательностей белка Tbp1, происходящего от штаммов H. influenzae, таких как Eagan DL63, РАК 12085 и SВ33 (SEQ ID 7, 5, 11 и 106); штаммов N. meningitidis, таких как В16В6 и М982 (SEQ ID 94 и 95); и штамма FA19 N. gonorrhoeae (SEQ ID 96).
На фиг.15 проиллюстрировано сравнение аминокислотной последовательности белка Тbр2, происходящего от штаммов H. influenzae, таких как Eagan, DL63, РАК 12085, SВ12, SВ29, SB30 и SВ32 (SEQ ID 8, 6, 12, 109, 110, 112, 114), штаммов N. meningitidis, таких как В16В6 и М982 (SEQ ID 97 и 98), штамма FА19 N. gonorrhoeae и штаммов Actinobacillus pleuropneumoniae, таких как АР205 и АР37 (SEQ ID 99 и 100).
На фиг.16А показана предполагаемая вторичная структура белка Tbp1 H. influenzae, а на фиг. 16В показана предполагаемая вторичная структура белка Тbр2 H. influenzae.
На фиг. 17 показана схема конструирования плазмиды JB-I468-29, которая экспрессирует в E. coli белок Tbp1, происходящий от штамма Eagan H. influenzae типа b.
На фиг. 18 показана схема конструирования плазмиды JB-I424-2-8, которая экспрессирует в E.coli белок Тbр2, происходящий от штамма H. influenzae типа b.
На фиг. 19 представлены пары олигонуклеотидов (SEQ ID 130, 131), используемых для конструирования плазмиды JB-1424-2-8.
На фиг. 20 показаны последовательности пар олигонуклеотидов: А (SEQ ID 86, 87); В (SEQ ID 88, 89); С (SEQ ID 90, 91) и D (SEQ ID 92, 93), используемых для конструирования плазмид, экспрессирующих Tbp1 и Тbр2.
На фиг.21 показана схема конструирования плазмиды JB-1600-1, которая экспрессирует в E.coli белок Тbр2, происходящий от штамма SВ12 H. influenzae.
На фиг. 22 проиллюстрирован электрофорез в ПААГ с ДСН продуктов экспрессии белка Tbp1 и белка Тbр2 штамма Eagan Haemophilus типа b, и белка Тbр2 нетипируемого штамма SВ12 H. influenzae в E.coli. Дорожка 1: JB-1476-2-1 (T7/Eagan Tbp1) в t0; дорожка 2: JB-1476-2-1, при времени индукции t=4 часа; дорожка 3: маркеры молекулярной массы: 200 кДа, 116 кДа, 97,4 кДа, 66 кДа, 45 кДа и 31 кДа; дорожка 4: JB-1437-4-1 (Т7/Eagan - Tbp2) при t0; дорожка 5: JB-1437-4-1 при времени индукции t=4 часа; дорожка 6: JB-1607-1-1 (T7/JB12-Tbp2) при t0; дорожка 7: JB-1607-1-1 при времени индукции t=4 часа.
На фиг. 23 проиллюстрирована схема очистки белков Tbp1 и Тbр2, экспрессированных в E.coli.
На фиг. 24 проиллюстрирован анализ частоты рекомбинантных белков Tbp1 и Тbр2, очищенных по схеме, проиллюстрированной на фиг.23. Дорожка 1 - маркеры молекулярной массы (106,80; 49,5; 32,5; 27,5 и 18,5 кДа); дорожка 2 - лизат целых клеток E.coli; дорожка 3 - солюбилизированные тельца включения; дорожка 4 - очищенные белки Tbp1 и Тbр2.
На фиг. 25 проиллюстрирована иммуногенность рекомбинантного белка Tbp1 (rTbp1, верхняя панель) и рекомбинантного белка Тbр2 (rТbр2, нижняя панель) у мышей.
На фиг. 26 проиллюстрирована реактивность антисывороток против rTbp1 штамма Eagan по отношению к различным штаммам H. influenzae при Вестерн-блоттинге. Дорожка 1: BL21/DЕ3; дорожка 2: SВ12-EDDA; дорожка 3: SВ12+EDDA; дорожка 4: SВ29-EDDA; дорожка 5: SВ29 + EDDA; дорожка 6: SВ33-EDDA; дорожка 7: SВ33+EDDA; дорожка 8: Eagan-EDDA; дорожка 9: Eagan+EDDA; дорожка 10: В. catarrhalis 4223-EDDA; дорожка 11: В. catarrhalis 4223+EDDA; дорожка 12: N. meningitidis 608-EDDA; дорожка 13: N. meningitidis 608+EDDA; дорожка 14: индуцированный штамм JB-1476-2-1, экспрессирующая рекомбинантный белок Tbp1 штамма Eagan; дорожка 15: маркеры молекулярных масс. Специфические ~95 кДа-полосы, прореагировавшие с антисыворотками против Tbp1, на дорожках 3, 4, 5, 7, 8 и 9 соответствуют штаммам SВ12, SВ29, SВ33 и Eagan H. influenzae; ~ 110 кДа-полосы на дорожках 10 и 11 соответствуют штамму 4223 В. catarrhalis; а ~80 кДа-полосы на дорожках 12 и 13 соответствуют 608 N. meningitidis.
На фиг.27 проиллюстрирована реактивность антисывороток против rТbр2 штамма Eagan по отношению к различным штаммам H. influenzae при Вестерн-блоттинге. Дорожка 1: маркеры молекулярных масс; дорожка 2: индуцированный штамм JB-1437-4-1, экспрессирующая рекомбинантный белок Тbр2 штамма Eagan; дорожка 3: SB12-EDDA; дорожка 4: SВ12+EDDA; дорожка 5: SВ29-EDDA; дорожка 6: SВ29+EDDA; дорожка 7: SВ30-EDDA; дорожка 8: SВ30+EDDA; дорожка 9: SВ32-EDDA; дорожка 10: SВ33-EDDA; дорожка 11: SВ33+EDDA; дорожка 12: РАК-EDDA; дорожка 13: РАК+EDDA; дорожка 14: Eagan-EDDA; дорожка 15: Eagan+EDDA. Специфические 60-70 кДа-полосы реагировали с антисыворотками против Тbр2 на дорожках 3, 6, 7, 8, 13, 14 и 15, которые соответствуют штаммам SВ12, SВ29, SВ30, РАК и Eagan.
На фиг. 28 проиллюстрировано конструирование плазмид pNH1T1KFN и pNH1T1KFP, использованных для получения штаммов H. influenzae, которые не продуцируют рецептор трансферрина.
На фиг.29 проиллюстрировано конструирование плазмид, кодирующих химерные полиовирусы, экспрессирующие эпитоп, происходящий от белка рецептора трансферрина, который является консервативным для бактерий, продуцирующих рецептор трансферрина.
На фиг. 30 показан Вестерн-блоттинг, иллюстрирующий реактивность антисывороток, продуцированных путем иммунизации кроликов химерными полиовирусами, экспрессирующими эпитоп, происходящий от белка рецептора для трансферрина, который является консервативным для бактерий, продуцирующих рецептор трансферрина. На панели А показан окрашенный кумасси бриллиантовым голубым гель, обнаруживающий очищенный рекомбинантный белок Tbp 2, происходящий от штамма SВ12 H. influenzae, и экспрессированный в E.coli (дорожка 1); очищенный белок Тbр2, происходящий от штамма 4223 Branhamella catarrhalis (дорожка 2); клеточный лизат от железолимитированного штамма 4223 В. catarrhalis (дорожка 3), клеточный лизат от штамма JM109 E.coli, выращенного в железонелимитированных условиях (дорожка 5). На панели В показаны результаты Вестерн-блоттинга реплики с геля с использованием пула сывороток, собранных на 27-ой день после иммунизации кроликов путем введения PUITBP2A (кролики 40, 41 и 42). На панели С показаны результаты, полученные для пула неиммунных сывороток, взятых от кроликов, которые обнаруживали минимальную специфическую реактивность.
На некоторых из вышеописанных фигур для сайт-специфических рестриктирующих эндонуклеаз были использованы следующие сокращения: R - Eco RI; Рs - Pst I; Н - Hind III; Вg - Bg1 II; Nde - Nde I; Ear - Ear I и Sau -Sau 3A I.
На фиг. 28 для обозначения сайт-специфических рестриктирующих эндонуклеаз были использованы следующие сокращения: А - Асc I; В - Bam HI; Е - Eco RI; O - Xho I; Н - Hind III; Ps - Pst I; V - Eco RV; Х - Хbа I; G - Вg1 II; S - Sa1 I; К - kpn I и S - SAc I.
Общее описание изобретения
Для получения очищенной и выделенной нуклеиновой кислоты, которая может представлять собой ДНК-молекулу, содержащую, по крайней мере, часть нуклеиновой кислоты, кодирующую рецептор трансферрина, типированный в соответствии с настоящим изобретением, может быть использован любой штамм Haemophilus. Такие штаммы могут быть получены из клинических источников, либо из коллекций бактериальных культур, таких как Американская коллекция типовых культур.
В соответствии с одним аспектом настоящего изобретения белок рецептора трансферрина может быть выделен из штаммов Haemophilus методами, описанными Schryvers (1989) Ogunnaviwo и Schryvers (1992), и а патенте США 5141743, раскрытие которого вводится в настоящее описание посредством ссылки. Хотя подробное описание этого способа приводится в патенте США 5141743, однако, для ясности, ниже приводится краткое изложение его сущности. Рецептор трансферрина получают путем выделения мембранной фракции из бактериального штамма, экспрессирующего активность связывания с трансферрином, и последующей очистки рецептора трансферрина аффинным методом, предусматривающим последовательные стадии предварительного связывания трансферрина с рецептором для трансферрина в мембранной фракции, солюбилизации мембраны, иммобилизации трансферрина и отделения рецептора трансферрина от иммобилизованного трансферрина. Альтернативно рецепторные белки могут быть выделены методом, представляющим собой модификацию вышеуказанного метода, в котором отсутствует стадия предварительного связывания, а буфер для солюбилизации содержит высокую концентрацию соли, что позволяет осуществить непосредственное выделение с иммобилизованным трансферрином, как описано в работе Ogunnariwo и Schryvers (1992).
В настоящей заявке термин "рецептор трансферрина" используется для определения семейства белков Tbp1 и/или Тbр2, имеющих различные отклонения в своих аминокислотных последовательностях, обусловленные их происхождением от различных природных штаммов, например Haemophilus. Другими бактериальными источниками рецептора трансферрина являются, но не ограничиваются ими такие виды, как Neisseria, Branhamella, Pasteurella и Actinobacillus.
Некоторые, если не все, из этих бактерий содержат оба белка Tbp1 и Тbр2. Под очищенными и выделенными ДНК-молекулами, содержащими, по крайней мере, часть нуклеотидной последовательности, кодирующей рецептор трансферрина настоящего изобретения, подразумеваются также ДНК-молекулы, кодирующие функциональные аналоги рецептора трансферрина. В настоящей заявке первый белок или пептид является "функциональным аналогом" второго белка или пептида в том случае, если указанный первый белок или пептид является иммунологически родственным второму белку или пептиду и/или имеет такую же функцию. Функциональным аналогом может быть, например, фрагмент белка или его мутант, образованный вследствие аминокислотного замещения, добавления или делеции.
В одном из вариантов осуществления настоящего изобретения рецептор трансферрина выделяют из штамма DL63 H. influenzae типа b и очищают методами аффинной хроматографии, как описано в работах Schryvers (1989), Ogunnariwo и Schryvers (1992) и в патенте США 5141743. Выделенный и очищенный рецептор трансферрина был использован для продуцирования у кроликов антисыворотки против TfR. Хромосомную ДНК из штамма DL63 H. influenzae типа b подвергали механической фрагментации, затем добавляли EcoRI-линкеры и конструировали библиотеку экспрессируемых последовательностей в ZAP. Эту библиотеку скринировали с использованием кроличьих антисывороток против TfR и получали два положительных клона (pBH1T1 и рВН1Т2), которые имели перекрывающиеся рестрикционные карты (фиг.1А и фиг.2). Эти клоны секbенировали, в результате чего были идентифицированы две большие открытые рамки считывания (фиг.2). Нуклеотидные последовательности генов рецепторов трансферрина Tbp1 и Тbр2 (SEQ ID 1) от штамма D63 H. influenzae и их выведенные аминокислотные последовательности (SEQ ID 5 - Tbp1 и SEQ ID 6 - Тbр2) показаны на фиг.3. Анализ этих последовательностей показал, что оперон ТfR состоит из двух последовательно расположенных генов (Tbp1 и Тbр2), транскрибированных от одного промотора (как, в частности, показано на фиг.2 и фиг.3). Молекулярная масса белка Тbр2 варьируется в зависимости от штамма, тогда как молекулярная масса белка Tbp1 остается, в основном, неизменной для различных штаммов данной бактерии, и обнаруживает некоторую вариабельность у различных бактерий, имеющих TfR-гены. Молекулярная масса Tbp1 обычно составляет 94000-106000, а молекулярная масса Тbр2, в основном, варьируется а пределах от 58000 до 98000.
Было проведено секвенирование аминокислотной последовательности N-концов и фрагментов, полученных путем расщепления бромистым цианом, рецептора трансферрина от штамма DL63 H. influenzae. N-концы Тbр2 блокировали, а аминокислотные последовательности Tbp1 идентифицировали путем секbенирования, и идентифицированные пептиды, соответствующие Tbp1, показаны на фиг. 3 (подчеркнуты). Этими пептидными последовательностями являются Glu Thr Gln Ser Ile Lys Asp Thr Lys Glu Ala Ile Ser Ser Glu Val Asp Thr (показанная на фиг.3, SEQ ID 101) и Leu Gln Leu Asn Leu Glu Lys Lya Ile Gln Gln Asn Trp Leu Thr His Gln Ile Ala Phe (показанная на фиг.3, SEQ ID 102). Сигнальная последовательность Tbp1 и предполагаемая сигнальная последовательность Тbр2 показаны на фиг.3 двойными линиями, проведенными сверху. Предполагаемая сигнальная последовательность для Tbp1 представляет собой: Met Thr Lys Lys Pro Tyr Phe Arg Leu Ser Ile Ile Ser Cys Leu Leu Ile Ser Cys Tyr Val Lys Ala (SEQ ID 103). Предполагаемая сигнальная последовательность для Тbр2 представляет собой Met Lys Ser Val Pro Leu Ile Ser Gly Gly Leu Ser Phe Leu Leu Ser Ala (SEQ ID 104). Выведенная аминокислотная последовательность N-концевой области Тbр2 показала, что это белок является липопротеином.
Из штамма Eagan H. influenzae типа b получали хромосомную ДНК и генерировали библиотеки. Первую библиотеку конструировали из ДНК, которую частично гидролизовали ферментом Sau 3А, фракционировали с получением ~5-10 кb-фрагментов и клонировали в плазмиду, полученную на основе pUC. Вторую библиотеку конструировали из Есо RI-рестриктированных хромосомных ДНК-фрагментов, клонированных в ZAP. Обе библиотеки зондировали 5'-фрагментом pBHIT-клона, показанного на фиг.2, в результате чего были получены частичные клоны TfR-генов штамма Eagan H. influenzae, обозначенные S-4368-3-3 и JB-901-5-3. Таким образом, в соответствии с другими аспектами настоящего изобретения на фиг.1В и 2 проиллюстрированы плазмидные клоны S-4368-3-3 и JB-901-5-3, кодирующие Тbр1 и Тbр2 от штамма Eagan H. influenzae типа b. ДНК-последовательности генов Tbp1 и Тbр2 (SEQ ID 2) от штамма Eagan H. influenzae типа b и их выведенные аминокислотные последовательности (SEQ ID 7 и 8) показаны на фиг.4, где последовательность Тbр2 является первым геном в опероне. На фиг.4 предполагаемые
последовательности -35, -10 и последовательность сайта связывания рибосомы показаны линиями, проведенными сверху.
Хромосомную ДНК получали из штамма MinnA H. influenzae типа b. Эту ДНК подвергали частичному гидролизу ферментом Sau 3A I, фракционировали на 10-20 кb-фрагменты и клонировали в BamHI-сайт EMBL3. Библиотеку зондировали 5'-фрагментом клона pBHIT (фиг.2) и получали полноразмерный клон, кодирующий TfR (DS-712-1-3). В соответствии с дополнительными аспектами настоящего изобретения, на фиг. 1С и 2 проиллюстрирован плазмидный клон DS 712-1-3, кодирующий Tbp1 и Тbр2 от штамма MinnA H. influenzae типа b. ДНК-последовательности Tbp1 и Тbр2 (SEQ ID 3) и их выведенные аминокислотные последовательности (SEQ ID 9 - Tbp1 и SEQ ID 10 - Тbр2) от штамма MinnA H. influenzae типа b показаны на фиг.5, где Тbр2 - последовательность является первой в опероне. На фиг.5, предполагаемые последовательности -35, -10 и последовательности сайта связывания рибосомы показаны верхними линиями.
Хромосомную ДНК получали из нетипируемого штамма РАК 12085 H. influenzae. ДНК частично гидролизовали ферментом Sau 3A, фракционировали на 10-20 кb-фрагменты и клонировали в Bam HI-сайт MBL 3. Библиотеку зондировали фрагментами клона pBHIT (фиг.2) и получали полноразмерный клон, кодирующий TfR (JB-1042-7-6). Рестрикционная карта клона JB-1042-7-6 показана на фиг.1 и 2, а нуклеотидные последовательности генов Тbр1 и Тbр2 (SEQ ID 4) от штамма РАК 12085 H. influenzae и их выведенные аминокислотные последовательности показаны на фиг.6 (SEQ ID 11, 12), при этом последовательность Тbр2 является первой. На фиг.6, предполагаемые последовательности -35, -10 и последовательность сайта связывания с рибосомой показаны линиями, проведенными сверху.
Хромосомную ДНК получали от нетипируемого штамма SВ33 H. influenzae, происходящего из отитной среды. Эту ДНК подвергали частичному гидролизу ферментом Sau 3A I, фракционировали на 10-20 кb-фрагменты и клонировали в Bam HI-сайт EMBL 3. Библиотеку зондировали фрагментами клона pBHIT (фиг.2) и получали полноразмерный клон, кодирующий TfR (В-1031-2-9). Рестрикционная карта клона JB-1031-2-9 показана на фиг.2, а нуклеотидные последовательности генов Tbp1 и Тbр2 (SEQ ID 4) от штамма SB33 H. influenzae и их выведенные аминокислотные последовательности показаны на фиг.7 (SEQ ID 11, 12), при этом последовательность Тbр2 является первой. Было обнаружено, что ген Тbр2 SВ33 имеет делецию в одно основание, которая приводит к сдвигу рамки считывания в остатке 126 и к преждевременному процессингу полученного белка в остатке 168.
Осуществляли РСR-амплификацию генов tbp 2 от штаммов SВ12, SВ28, SВ32 NTHi, происходящих из отитной среды, и эти гены секвенировали.
Нуклеотидная последовательность генов tbp 2 от нетипируемых штаммов SB12 (SEQ ID 105), SВ29 (SEQ ID 108), SB30 (SEQ ID 110) и SВ32 (SEQ ID 112). H. influenzae показаны на фиг.8, 9, 10 и 11 соответственно.
Было установлено, что все амплифицированные гены tbp 2 кодируют полноразмерные белки Тbр2, что свидетельствует о том, что дефектный ген штамма SВ33 является атипичным.
Все три штамма H. influenzae типа b имеют идентичные короткие межгенные последовательности, состоящие всего из 13 п.о. между tbp 2 и tbp 1, а штаммы РАК 12085 THi имеют более длинные межгенные последовательности, состоящие из 27 п.о. (фиг.12).
Штамм SВ12 имеет межгенную последовательность в 13 п.о., идентичную последовательности, обнаруженной в штаммах H. influenzae типа b, а штаммы SВ29, SВ30 и SВ32 содержат более длинные межгенные последовательности (27-30 п. о. ), обнаруженные в других штаммах РАК 12085 и SВ33 NTHi (фиг.2В). Все девять штаммов имеют общую консервативную последовательность из 13 п.о., расположенную между их генами tbp 2 и tbp 1.
Была идентифицирована пентапептидная последовательность возле аминоконца Tbp1 H. influenzae (фиг. 12), которая аналогична TonB-блоку. Ген tonB H. influenzae был недавно клонирован и секвенирован (Jarasic и др., 1994).
Сравнение аминокислотных последовательностей Тbр1 от штаммов Eagan/MinnA DL63, РАК 12085 и SВ33 H. influenzae проиллюстрировано на фиг.14. Белки Tbp1 штаммов Eagan и MinnA являются идентичными и имеют длину в 912 аминокислот; белок Tbp1 штамма DL63 имеет 914 остатков, белок штамма РАК 12085 имеет 914 остатков; а белок штамма SВ33 имеет 911 остатков. Белки Tbp1 H. influenzae являются в высокой степени консервативными и имеют идентичность последовательностей на 95-100%. Сравнение аминокислотных последовательностей Тbр2 от штаммов Eagan/MinnA, DL63, РАК 12085, SВ12, SВ29, SВ30 и SВ32 проиллюстрировано на фиг. 15. Белки Тbр2 штаммов Eagan и MinnA являются идентичными и содержат 660 аминокислот, белок Тbр2 штамма DL63 имеет 644 остатков, а белок штамма РАК 12085 имеет 654 остатков. В гене tbp 2 штамма SВ33 имеется делеция в одно основание, которая приводит к сдвигу рамки считывания в остатке 126 и преждевременному процессингу белка в остатке 168. Это отсутствие основания подтверждается прямым секвенированием PCR-амплифицированной хромосомной ДНК. За исключением белков Тbр2 штаммов Eagan и MinnA, которые являются идентичными, последовательности белка Тbр2 являются, в основном, менее консервативными с идентичностью лишь 66-70%, но при этом имеются несколько коротких сегментов с консервативной последовательностью, которые могут быть идентифицированы на фиг.15. Было установлено, что все РСR-амплифицированные гены tbp 2 от штаммов SВ12, SВ29, SВ30 и SВ32 кодируют полноразмерные белки Тbр2. Среди выведенных белков наблюдается гетерогенность последовательностей и размеров; так, например, белок Тbр2 штамма SВ12 имеет 648 аминокислот, белок штамма SВ29 имеет 631 остаток, белок штамма SВ30 имеет 630 остатков, а белок штамма SВ32 имеет 631 остаток.
Были определены предполагаемые структуры белков Тbр1 и Тbр2 штамма Eagan (фиг. 16А и 16В). Оба белка имеют несколько трансмембранных доменов; причем белок Tbp1 пересекает мембрану 20 раз, а белок Тbр2 пересекает мембрану 12 раз.
Три внешних консервативных эпитопа были идентифицированы в аминоконцевой области Тbр1 (DNEVTGLGK - SEQ ID 43, EQVLN/DIRDLTRID - SEQ ID 139 и 140 и GAINEIEVENVKAVEISK - SEQ ID 141) и один в С-концевой области (GI/VYNLV/LNYRYWTWE - SEQ ID 142 и 143). Только три небольших консервативных области могут быть идентифицированы в N-конце белков Tbp2 патогенов человека: CSS/LGGG(G)SFD - SEQ ID 75, 144 и 145; внутри указанных белков: LE/SGGFY/FGR - SEQ ID 74 и 146 и в С-конце этих белков: VVFGAR/K - SEQ ID 83 и 84.
Обнаружение того факта, что аминокислотная последовательность варьирует от штамма к штамму Haemophilus, позволяет подразделять Haemophilus на подгруппы, определяемые одинаковыми аминокислотными последовательностями Тbр2. Кроме того, этот факт позволяет осуществлять рациональный отбор минимального числа последовательностей Tbp1 и/или Тbр2 или синтетических пептидов, представляющих эпитопы, общие для указанных штаммов Haemophilus, которые могут быть использованы в иммуногенных композициях, например, для иммунизации против заболеваний, вызываемых Haemophilus и другими бактериями, продуцирующими рецептор трансферрина с последовательностями, аналогичными последовательностями Tbp1 и Тbр2 от штаммов Haemophilus. Таким образом, для иммунизации против многих или всех штаммов Haemophilus и других бактериальных патогенов, продуцирующих рецептор трансферрина, может быть использовано минимальное число рецептора трансферрина, его аналогов, фрагментов и/или пептидов.
Кроме того, проводили сравнение (как показано на фиг.14 и 15) аминокислотных последовательностей рецептора трансферрина, происходящего от ряда бактериальных патогенов (H. influenzae типа b, нетипируемых H. influenzae, Neisseria meningitidis, Neisseria gonorrhoe и Actinobacillus (Haemophilus) pleuropneumoniae).
В результате этих анализов были выявлены области Tbp1 и Тbр2, которые являются консервативными для всех указанных бактерий. Некоторые из таких консервативных последовательностей содержатся в пептидах, как показано в таблицах 2 и 3. В частности, в Tbp1 (таблица 1 и фиг.14) консервативными являются последовательности DNEVTGLGK (SEQ ID: 43), EQVLNIRDLTRYDPGI (SEQ ID NO 44), EQCVLNIRDLTRYDPGISVVEQGRGASSGYSIRGMD (SEQ ID NO 45), GAINEIEYENVKAVEISKG (SEQ ID NO 46) и GALAGSV (SEQ ID NO 47). Особенно консервативными последовательностями В Тbр2 являются LBGGFYGP (SEQ ID NO 74) CSGGGSFD (SEQ ID NO 75), YVYSGL (SEQ ID NO 76). СCSNLSYVKFS (SEQ ID NO 77), FLLGHRT (SEQ ID NO 78), EFNVOF (SEQ ID NO 79), NAFTGTA (SEQ ID NO 80), VNGAFYG (SEQ ID NO 81), ELGGYF (SEQ ID NO 82), VVFGAR (SEQ ID NO 83) и VVFGAK (SEQ ID NO 84) (cм. таблицу 2 и фиг.15).
Обнаружение консервативных последовательностей в рецепторе трансферрина ряда бактериальных патогенов позволяет выбрать минимальное число антигенов, имеющих конкретные аминокислотные последовательности (включая последовательности в виде синтетических пептидов), для иммунизации против заболеваний, вызываемых патогенами, имеющими рецепторы трансферринов. Такими патогенами, помимо вышеуказанных бактерий, являются и другие штаммы Neisseria, такие как Neisseria gonorrhoe и Branhamella, включая Branhamella catarrhalis. Указанные консервативные аминокислотные последовательности, содержащиеся во многих бактериальных патогенах, позволяют продуцировать TfR-специфические антитела, включая моноклональные антитела, которые распознают большинство, если не все, рецепторы трансферрина. Была продуцирована антисыворотка против пептидов, соответствующих консервативным областям, рецепторатрансферрина. Эта антисыворотка "узнавала" рецептор трансферрина в Branhamella catarrhalis. Такая антисыворотка может быть использована для обнаружения и нейтрализации большинства, если не всех бактерий, продуцирующих белок ТfR, и кроме того, она может быть также использована для пассивной иммунизации против заболеваний, вызываемых указанными патогенами. Таким образом, диагностические анализы и наборы, использующие консервативные аминокислотные последовательности, могут быть ценным инструментом для обнаружения, если не всех, то большинства бактерий, которые продуцируют рецептор трансферрина.
Эпитопы, содержащие вышеуказанные аминокислотные последовательности, могут быть доставлены в клетки иммунной системы с помощью синтетических пептидов, содержащих такие последовательности, либо с помощью живых векторов, экспрессирующих такие последовательности, либо путем прямого введения молекул нуклеиновой кислоты, кодирующих аминокислотную последовательность.
Некоторые пептиды, содержащие консервативные аминокислотные последовательности в белках Tbp1 штаммов Eagan MinnA, DL 63 и нетипируемого штамма РАК 12085 H. influenzae типа b, показаны в таблице 2. Антитела к некоторым из этих пептидов были продуцированы у морских свинок (таблица 4). Пептиды, содержащие консервативные аминокислотные последовательности в белках Тbp2 штаммов Eagan, MinnA, DL63, и нетипируемого штамма РАК 12085 H. influenzae типа b, показаны в таблице 3. Антитела к некоторым из этих пептидов были продуцированы у морских свинок (таблица 4).
Для продуцирования рекомбинантных белков, кодирующие последовательности генов Тbр1 и Тbр2, могут быть клонированы в соответствующие экспрессирующие векторы. Рекомбинантные Tbp1 и Тbр2 были экспрессированы в E.coli с использованием экспрессирующей системы Т7. Ген tbp1, кодирующий зрелый белок Tbp1 штамма Eagan, был клонирован с сохранением рамки считывания за промотором Т7, генерирующим плазмиду JB-1468-29, как показано на фиг.17. При введении гена tbp1 в клетки BL21/DE3 и индуцировании IPTG или лактозой экспрессировался белок Tbp1 штамма Eagan, показанный на фиг.22.
Ген tbp2, кодирующий зрелый белок Тbр2, был клонирован с сохранением рамки считывания за промотором Т7, генерирующим плазмиду JB-1424-2-8, как показано на фиг.18. При введении этого гена в клетки E.coli и индуцировании, как указано выше, экспрессироаался белок Тbр2, показанный на фиг.22.
Ген tbp2 от штамма NTHI SВ12 был амплифицирован с помощью полимеразной цепной реакции (PCR). Полученная амплифицированная ДНК содержала аутентичную сигнальную последовательность Тbр2 H. influenzae перед последовательностью зрелого белка. Ген tbp 2 от штамма JB12, кодирующий сигнальную последовательность и зрелый белок, был клонирован в экспрессирующую систему рТ7-7, как показано на фиг.21. При введении полученной плазмиды (JB-1600-1) в клетки BL21/LE3 E. coli и индуцировании экспрессировался белок Тbр2 SВ12, как показано на фиг.22.
Рекомбинантные белки Тbр1 и Тbр2, продуцированные в E.coli как тельца включения, были очищены в соответствии со схемой, показанной на фиг.23. Очищенные белки имели чистоту, по крайней мере, около 70%, как показано на фиг. 24. Исследования иммуногенности осуществляли на мышах с использованием очищенных рекомбинантных белков Tbp1 и Тbр2. Оба белка, введенные в дозах 3-10 мкг, продуцировали хороший иммунный ответ у мышей (фиг.25).
Антисыворотки, продуцируемые против рекомбинантных белков Tbp1 или Тbр2, происходящих от одного из штаммов H. influenzae, давали перекрестную реакцию с другими штаммами, что свидетельствует о возможности их использования в качестве ценных диагностических реагентов (фиг.26 и 27).
Плазмиды pUH1T1KFN и pUH1TKKFP, показанные на фиг.28, содержат селективный маркер устойчивости к антибиотику, клонированный в опероне рецептора трансферрина, и были сконструированы для локальной инактивации оперена рецептора трансферрина. Эти плазмиды были использованы для трансформации H. influenzae в целях получения штаммов, которые не продуцируют рецепторы трансферрина Tbp1 и/или Тbр2, как описано в примере 19. Такие штаммы могут быть использованы как негативный контроль (так как они не продуцируют TfR) в in vivo и in vitro анализах и при диагностике. Эти штаммы, по всей вероятности, окажутся также аттенюированными при их in vivo-росте и могут быть использованы в качестве живых вакцин для иммунизации против заболеваний, вызываемых Haemophilus.
Как обсуждалось выше, эпитопы белков рецепторов трансферрина могут быть доставлены в клетки иммунной системы путем использования живых векторов, экспрессирующих такие аминокислотные последовательности, причем таким живым вектором может быть провирус. На фиг.29 проиллюстрирована конструкция гибридных полиовирусов, экспрессирующих эпитоп белка рецептора трансферрина, включая консервативный эпитоп от Тbр2 LFCCFYGP (SEQ ID 74). Эти вирусы распознаются антителами, продуцированными против пептида, содержащего аминокислотную последовательность LECCFYGP (SEQ ID 74) (таблица 5), что свидетельствует о том, что указанные вирусы экспрессируют эту последовательность в антигенно распознаваемой форме. РV1ТВР2А и РV1ТВР2В были также нейтрализованы кроличьими антисыворотками против tbp2 штамма DL63 H. influenzae, что указывает на то, что, по крайней мере, два вируса экспрессироаали последовательность в форме, распознаваемой антителами, продуцированными против белка. Сыворотки против PV1 были способны нейтрализовать все вирусы, что указывало на то, что изменения в антигенной области I нейтрализации полиовируса не оказывают значительного влияния на другие антигенные области этих вирусов. Кроме того, кроличья антисыворотка, продуцированная путем иммунизации кроликов полиовирусными химерами PV1TBP2A или РV1ТВР2В, распознавала пептид, содержащий аминокислотную последовательность LEGGFYGP (SEQ ID 74). Это свидетельствует о том, что последовательности, экспрессированные химерами PV1TB2A и PV1TBP2B, являются иммуногенными и вырабатывают антитела, способные распознавать последовательность.
На фиг.30 панели А показан ДСН-ПААГ-гель, обнаруживающий очищенный рекомбинантный Tbp2 от штамма JВ12 H. influenzae и экспрессированный в E.coli (дорожка 1); tbp2 от штамма 4223 Branhamella catarrhalis (дорожка 2); клеточный лизат от железолимитированного штамма 4223 В. catarrhalis (дорожка 3), клеточный лизат от железолимитированного штамма JM109 E.coli (дорожка 4) и клеточный лизат от штамма JM109 E.coli, культивированного в железонелимитированных условиях (дорожка 5). На панели В показаны результаты Вестерн-блоттинга реплики с геля с использованием пула сывороток, собранных от кроликов, иммунизированных РV1ТВР2А. При этом наблюдалась сильная реакция с очищенными связывающимися с трансферрином белками на дорожках 1 и 2 и с полосой аналогичных размеров на дорожке 3. Однако с какими-либо белками E.coli заметной реакции не наблюдалось (дорожки 4 и 5). На панели С показаны результаты, полученные для пула сывороток, взятых от тех же самых кроликов до иммунизации, которые обнаруживали минимальную специфическую реактивность. Полученные результаты показали, что PV1TBP2A обладает способностью индуцировать антисыворотки, специфичные к белкам, связывающимся с трансферрином и происходящим от H. influenzae и В. catarrhalis, и что эти антисыворотки могут отличать В. catarrhalis от E.coli, которые не экспрессируют эквивалентный белок.
Очищенные и выделенные ДНК-молекулы, содержащие, по крайней мере, часть, кодирующую рецептор трансферрина штамма Haemophilus, типированного как описано в настоящем описании, могут быть с успехом использованы, как:
- нуклеиновокислотные зонды для специфической идентификации штаммов Haemophilus in vitro или in vivo;
- продукты, кодированные ДНК-молекулами, могут быть использованы в качестве диагностических реагентов, антигенов для продуцирования Haemophilus-специфических антисывороток, для вакцинации против заболеваний, вызываемых видами Haemophilus, и, например, для выявления инфекции, вызываемой Haemophilus;
- пептиды, соответствующие частям рецептора трансферрина, типированного как описано в настоящей заявке, могут быть с успехом использованы в качестве диагностических реагентов, антигенов для продуцирования Haemophilus-специфических антисывороток, для вакцинации против заболеваний, вызываемых видами Haemophilus, и, например, для выявления инфекции, вызываемой Haemophilus.
Рецептор трансферрина, кодированный нуклеиновокислотными молекулами настоящего изобретения, его фрагменты и аналоги, а также пептиды, содержащие последовательности, соответствующие частям рецептора трансферрина, которые являются консервативными для различных изолятов Haemophilus и других бактерий, продуцирующих рецептор трансферрина, могут быть использованы для диагностики заболеваний, вызываемых любым бактериальным штаммом, который продуцирует рецептор трансферрина, или для иммунизации против указанных заболеваний. В частности, пептиды, содержащие последовательности ЕССFYGР, являются консервативными в белках рецептора трансферрина многих бактериальных патогенов, которые продуцируют рецептор трансферрина, а поэтому могут быть с успехом использованы для диагностики и иммунизации против заболеваний, вызываемых бактериями, продуцирующими рецептор трансферрина. Такими бактериями являются, но не ограничиваются ими, штаммы Haemophilus, Neisseria (включая N. meningitidis и N. gonorrhoeae) и Branhamella (включая В. catarrhalis).
Для каждого специалиста совершенно очевидно, что различные варианты настоящего изобретения могут иметь множество применений, например, в вакцинации; диагностике; лечении инфекций, таких как Haemophilus-инфекции, и инфекций других бактериальных патогенов, продуцирующих рецептор трансферрина; а также для продуцирования иммунологических реагентов. Ниже приводится более подробное, но не исчерпывающее обсуждение применений настоящего изобретения.
1. Получение и использование вакцин
Иммуногенные композиции, подходящие для использования в качестве вакцин, могут быть получены с использованием иммуногенного рецептора трансферрина, его аналогов, фрагментов и/или пептидов, описанных в настоящей заявке. Эта вакцина продуцирует иммунный ответ посредством вырабатывания антител, включая антитела против рецептора трансферрина и опсонизирующие или бактерицидные антитела. Независимо от того, проводили ли вакцинированному индивидууму антигенную стимуляцию бактерией Haemophilus или другими бактериями, продуцирующими рецептор трансферрина, антитела связываются с рецептором трансферрина, препятствуя, тем самым, доступу бактерий к источнику железа, необходимого для их жизнеспособности. Кроме того, опсонизирующие или бактерицидные антитела против TfR могут также обеспечивать защиту организма посредством альтернативных механизмов.
Вакцины, содержащие пептиды, хорошо известны специалистам и проиллюстрированы в патентах США 4601903; 4499231; 4599230 и 4596792, которые вводятся в настоящее описание посредством ссылки. Иммуногенные композиции, содержащие вакцины, могут быть изготовлены в виде инъецируемых жидких растворов или эмульсий. Для этого рецептор трансферрина, его аналоги и фрагменты и/или пептиды могут быть смешаны с фармацевтически приемлемыми наполнителями, совместимыми с рецепторами трансферрина, фрагментами, аналогами или пептидами. Такими наполнителями являются вода, физиологический раствор, глицерин, этанол, декстроза и их комбинации. Кроме того, иммуногенные композиции и вакцины могут содержать добавки, такие как смачивающие или эмульгирующие агенты, рН-регулирующие забуферивающие агенты, или адьюванты для повышения эффективности вакцин. Иммуногенные композиции и вакцины могут быть введены парентерально путем подкожной или внутримышечной инъекции. Альтернативно иммуногенные композиции, составленные в соответствии с настоящим изобретением, могут быть изготовлены и введены таким образом, что они будут продуцировать иммунный ответ на поверхности слизистых оболочек. Так, например, иммуногенная композиция может быть нанесена на поверхности слизистых оболочек, например, через нос или перорально (внутрижелудочно). Иммуногенная композиция может быть введена а комбинации с молекулой, обеспечивающей доставку указанной композиции к специфическим клеткам иммунной системы или к поверхностям слизистых оболочек. Некоторыми из таких молекул, обеспечивающих доставку, является штамм В12 и фрагменты бактериальных токсинов, описанные в WO 92/17167 (Biotech Australia Pty. Ltd.), а также моноклональные антитела, описанные в патенте США 5194254 (Barber и др.). Альтернативно могут быть предпочтительными другие способы введения, например, в виде суппозиториев и пероральных композиций. В случае использования суппозиториев в композицию могут быть включены связующие вещества и носители, например полиалкаленгликоли или триглицериды. Пероральные композиции могут содержать инициаторы, например, такие как фармацевтические сорта сахарина, целлюлозы и карбоната магния. Эти композиции могут быть изготовлены в виде растворов, суспензий, таблеток, драже, капсул, препаратов с пролонгированным высвобождением лекарственного средства, или порошков, которые содержат 10-95% рецептора трансферрина, его фрагмента, аналога и/или пептидов.
Вакцину обычно вводят способом, соответствующим конкретно вводимой лекарственной форме, и в таких количествах, которые, как рассчитывается, являются терапевтически эффективными, протективными и иммуногенными. Вводимое количество зависит от конкретного индивидуума, подвергаемого лечению, включая, например, способность его иммунной системы синтезировать антитела, и если необходимо, продуцировать клеточный иммунный ответ. Точное количество активного ингредиента, необходимого для введения, должен определять лечащий врач. Однако подходящая доза может быть легко определена любым специалистом и составляет порядка нескольких микрограммов рецептора трансферрина, его аналога, фрагмента и/или пептидов. Подходящие режимы для введения начальных и бустер-доз также могут варьироваться, но обычно сначала вводят начальную дозу, а затем бустер-дозы. Дозы вакцины могут также варьироваться от способа введения и от веса пациента.
Молекулы нуклеиновой кислоты, кодирующие рецептор трансферрина настоящего изобретения, могут быть также использованы непосредственно для иммунизации путем прямого введения ДНК, например посредством инъекции для генетической иммунизации, либо путем конструирования живого вектора, такого как Salmonella ВСG, аденовирус, поксвирус, вирус осповакцины или полиовирус. Обсуждение некоторых живых векторов, которые были использованы для переноса гетерологичных антигенов в иммунную систему, приводится в работе O'Hagan (1992). Способы прямого введения ДНК индивидуумам для генетической иммунизации описаны, например, Ulmer и др., 1993.
Использование пептидов in vivo может, в первую очередь, потребовать их химической модификации, поскольку сами пептиды могут не иметь достаточно продолжительного периода полужизни в сыворотке и/или тканях, и/или достаточной иммуногенности. Такие химически модифицированные пептиды именуются в настоящем описании "пептидными аналогами". Термин "пептидный аналог" означает любой функциональный химический эквивалент пептида, отличающийся своей повышенной стабильностью и/или эффективностью и иммуногенностью при in vivo- или in vitro-осуществлении настоящего изобретения. Термин "пептидный аналог" относится также к любому аминокислотному производному пептидов, описанных в настоящей заявке. Пептидные аналоги, рассматриваемые в настоящем описании, получают методами, которые предусматривают (но не ограничиваются ими): модификации в боковых цепях, введение ненатуральных аминокислот и/или их производных в процессе пептидного синтеза, и использование перекрестносшивающих агентов; и другими методами, которые налагают соответствующие ограничения на пептиды или их аналоги.
Примерами модификаций боковых цепей, рассматриваемых в настоящей заявке, могут служить модификация аминогрупп, такая как восстановительное алкилирование посредством реакции с альдегидом и последующего восстановления с использованием NaBH4; амидирование с использованием метилацетимидата; ацетилирование с использованием уксусного ангидрида; карбамоилирование аминогрупп цианатом; тринитробензилирование аминогрупп 2,4,6-тринитробензолсульфоновой кислотой (TNBS); алкилирование аминогрупп ангидридом янтарной кислоты и ангидридом тетрагидрофталевой кислоты и перидоксилирование лизина с использованием пиридоксал-5-фосфата с последующим восстановлением с использованием NaBH4.
Гуанидиногруппа аргининовых остатков может быть модифицирована путем образования продуктов гетероциклоконденсации с такими реагентами, как 2,3-бутандион, фенилглиоксал и глиоксал.
Карбоксильная группа может быть модифицирована путем активации карбодиимид посредством образования о-ацилизомочевины с последующей дериватизацией, например, до получения соответствующего амида.
Сульфгидрильные группы могут быть модифицированы такими методами, как карбоксиметилирование с использованием иодоуксусной кислоты или иодоацетамида; окисление пермуравьиной кислоты до цистеиновой кислоты; образование смешанных дисульфидов с другими тиоловыми соединениями; реакция с малеимидом; ангидридом малеиновой кислоты или с другим замещенным малеидом; образование ртутных производных с использованием 4-хлоробензоата ртути, 4-хлорортутьфенилсульфоновой кислоты, хлорида фенилртути, 2-хлорортуть-4-нитрофенола и других соединений и карбамоилирование с использованием цианата при щелочном рН.
Триптофановые остатки могут быть модифицированы, например, путем окисления с использованием N-бромосукцинимида; или путем алкилирования индольного кольца с использованием 2-гидрокси-5-нитробензилбромида или сульфонилгалогенидов. Триозиновые остатки могут быть модифицированы путем нитрования тетранитрометаном с получением производного 3-нитротирозина.
Модификация имидазольного кольца гистидинового остатка может быть осуществлена путем алкилирования с использованием производных иодоуксусной кислоты либо путем N-карбэтоксилирования с использованием диэтилпирокарбоната.
Примерами введения ненатуральных аминокислот и производных в процессе пептидного синтеза могут служить, но не ограничиваются ими, использование норлейцина, 4-аминомасляной кислоты, 4-амино-3-гидрокси-5-фенилпентаноаой кислоты, 6-аминогексановой кислоты, т-бутилглицина, норвалина, фенилглицина, орнитина, сакрозина, 4-амино-3-гидрокси-6-метилгептановой кислоты, 2-тиенилаланина и/или D-изомеров аминокислот.
Иммуногенность вакцины может быть значительно увеличена путем введения антигенов вместе с адъювантами, обычно используемыми в виде 0,05-1,0%-ного раствора в забуференном фосфатом физиологическом растворе. Адъюванты усиливают иммуногенность антигена, однако, сами они необязательно обладают иммуногенностью. Адьюванты могут действовать путем удержания антигена возле места введения, создавая, тем самым, эффект "депо", способствующий медленному, пролонгированному высвобождению антигена в клетки иммунной системы. Адьюванты могут также притягивать клетки иммунной системы к "депо" антигена и стимулировать указанные клетки для продуцирования иммунного ответа.
В течение уже многих лет иммуностимулирующие агенты или адъюванты используются для повышения иммунного ответа хозяина, например, на введение вакцины. Внутренние адьюванты, такие как липополисахариды, обычно представляют собой компоненты инактивированных или аттенюированных бактерий, используемых в качестве вакцин. Внешние адъюванты представляют собой иммуномодуляторы, которые, обычно, нековалентно связываются с антигенами, и, образуя композицию, способствуют усилению иммунного ответа хозяина. Таким образом, адъюванты были идентифицированы как соединения, способствующие усилению иммунного ответа на парентерально введенные антигены. Некоторые из этих адъювантов являются токсичными и могут вызывать нежелательные побочные эффекты, что делает их неприемлемыми для использования в лечении человека и многих животных. Фактически в вакцинах для введения человеку и животным в качестве адъювантов используются лишь гидроксид алюминия и фосфат алюминия (которые имеют общее сокращенное название "алюм"). Было установлено, что алюм является эффективным для повышения гуморального ответа на токсоиды дифтерии и столбняка, а совсем недавно алюмом была стимулирована HBsAg-вакцина. Однако, хотя эффективность алюма является хорошо установленным фактом, но тем не менее, эта эффективность имеет свои ограничения. Например, алюм является неэффективным для вакцинации против гриппа и неадекватно продуцирует клеточный иммунный ответ. Антитела, продуцированные алюмстимулированными антигенами у мышей, имеют главным образом изотип IgG1 и могут быть недостаточными для выработки иммунитета в ответ на введение некоторых вакцинных агентов.
Сильный иммунный ответ на введение антигенов могут продуцировать внешние адъюванты широкого ряда. Такими адьювантами являются сапонины, конъюгированные с антигенами мембранных белков (иммуностимулирующие комплексы); полимеры-плуроники с минеральным маслом; инактивированные микобактерии в сочетании с минеральным маслом; полный адъювант Фрейнда, бактериальные продукты, такие как мурамилдипептид (MDP), и липополисахариды (ЛПС); а также липид А и липосомы.
Для эффективного индуцирования гуморального иммунного ответа (HIP) и клеточного иммунитета (СМ1) иммуногены эмульгируют в адъювантах. Многие адъюванты являются токсичными, например, при гранулемах, острых и хронических воспалительных процессах (полный адъювант Фрейнда, FСА), цитолизе (сапонины и полимеры-плуроники), пирогенных реакциях, артритах и увеитах переднего отдела сосудистой оболочки глаза (ЛПС и MDP). Хотя FСА является прекрасным адьювантом и широко используется в различных исследованиях, однако из-за его токсичности он не был лицензирован на использование в вакцинах для введения человеку или животным.
Желательными характеристиками идеальных адъювантов являются:
(1) отсутствие токсичности;
(2) способность стимулировать продолжительный иммунный ответ;
(3) простота изготовления и стабильность при длительном хранении;
(4) способность продуцировать как гуморальный, так и клеточный иммунитет в ответ на введение антигенов различными путями, если это необходимо;
(5) синергия с другими адьювантами;
(6) способность к селективному взаимодействию с популяциями антигенпредставляющих клеток (АРС);
(7) способность к специфическому индуцированию соответствующего клеточного TН1- или ТН2-специфического иммунного ответа;
(8) способность к селективному увеличению уровней соответствующего изотипа антитела (например, IgA) против антигенов.
В патенте США 4855283, выданном Lockhoff и др. 8 августа 1989 г. и вводимом в настоящее описание посредством ссылки, рассматриваются аналоги гликолипидов (включая N-гликозиламиды, N-гликозилмочевины и N-гликозилкарбаматы, каждый из которых замещен в сахарном остатке аминокислотой), которые могут быть использованы в качестве иммуномодуляторов или адъювантов. Так, например, Lockhoff и др. (1991) показали, что N-гликолипидные аналоги, обнаруживающие структурное сходство с природными гликолипидами, такими как гликосфинголипиды и гликоглицеролипиды, способны продуцировать сильный иммунный ответ на введение вакцины против вируса простого герпеса, и псевдорабической вакцины. Некоторые гликолипиды были синтезированы из длинноцепочечных алкиламинов и жирных кислот, которые непосредственно связаны с сахарами посредством аномерного атома углерода, в целях имитации функций натуральных липидных остатков.
В патенте США 4258029, выданном Moloney, переданном его правоприемнику, и вводимом в настоящее описание посредством ссылки, указывается, что гидрохлорид октадецилтирозина (ОТН) действует как адьювант при введении его в виде комплекса с противостолбнячной вакциной и инактивированной формалином вакцины против вируса полиомиелита типа I, II и III. Кроме того, в работе Nixon-George и др. (1990) указывается, что октадециловые сложные эфиры ароматических аминокислот, ассоциированные с рекомбинантным поверхностным антигеном гепатита В, усиливают иммунный ответ хозяина против вируса гепатита В.
Липидизация синтетических пептидов также была использована для повышения иммуногенности этих пептидов. Так, например, в работе Wiesmuller (1989) описываются пептиды, имеющие последовательность, гомологичную белку вируса ящура, и ассоциированные с адъювантом три-пальмитил-S-глицерил-цистеинилсерилсерином, являющимся синтетическим аналогом N-концевой части липопротеина, происходящего от грамотрицательной бактерии. Кроме того, в работе Deres и др. (1989) описывается in vivo-примирование вирусспецифических цитотоксичных лимфоцитов Т синтетической липопептидной вакциной, которая содержит синтетические пептиды, происходящие от нуклеопротеина вируса гриппа, посредством их связывания с липопептидом N-пальмитил-S-[2,3]-бис(пальмитилокси)-(2РS)-пропил-[R]-цистеином (ТРС).
2. Иммуноанализы
Рецептор трансферрина, его аналоги и фрагменты и/или пептиды настоящего изобретения могут быть использованы в качестве иммуногенов и антигенов в иммуноанализах, включая твердофазный иммуноферментный анализ (ELISA), PIA и другие неферментные анализы на связывание с антителом, или другие известные процедуры на обнаружение антител против бактерий, Haemophilus, TfR и/или пептидов. В ELISA-анализе рецептор трансферрина, его аналоги, фрагменты и/или пептиды, соответствующие частям белка TfR, иммобилизируют на выбранной поверхности, например на поверхности, способной связывать белки или пептиды, такой как стенки полистиролового планшета для микротитрования. После промывания в целях удаления не полностью адсорбированного рецептора трансферрина, его аналогов, фрагментов и/или пептидов, неспецифический белок, такой как раствор альбумина бычьей сыворотки (BSA) или казеина, который, как известно, является антигенно нейтральным в отношении испытуемого образца, может быть связан с указанной выбранной поверхностью. Это позволяет блокировать участки неспецифической адсорбции на иммобилизирующей поверхности и таким образом позволяет снизить фон, возникающий в результате неспецифического связывания антисыворотки с поверхностью. Предпочтительно, чтобы выбранные пептиды происходили из консервативных областей, показанных в таблице 2 или 3, что позволяет повысить вероятность обнаружения перекрестных видов за исключением тех случаев, когда необходимо обнаружить один конкретный бактериальный вид. В этом случае выбирают полипептид, который является уникальным для ТfR данного конкретного вида. Обычно пептиды состоят из 12 остатков и более, а предпочтительно из 14-30 остатков. Однако при этом следует отметить, что смесь пептидов может быть использована либо в качестве иммуногена в вакцине, либо в качестве диагностического агента. Это относится к тем случаям, когда смесь пептидов от консервативных областей и/или от неконсервативных областей используется для защиты от перекрестнородственных микроорганизмов, и/или для диагностики. В этом случае смесь пептидных иммуногенов обычно называют препаратом - "коктейлем" для использования в качестве вакцины или диагностического агента.
Вышеупомянутую иммобилизирующую поверхность затем подвергают контакту с образцом, таким как исследуемые клинические или биологические материалы, способом, способствующим образованию иммунного комплекса (антиген/антитело). Для этого образец разводят разбавителями, такими как BSA, коровий гамма-глобулин (BGG) и/или физиологический раствор, забуференный фосфатом (PBS)/Твин. После этого образец инкубируют в течение 2-4 часов при температуре порядка 25-37oС. После инкубирования контактируемую с образцом поверхность промывают для удаления материала, не связанного в иммунокомплексе. Процедура промывания предусматривает промывку раствором, таким как РВS/Твин или боратный буфер.
После образования специфических иммунокомплексов между испытуемым образцом и связанным рецептором трансферрина, его аналога, фрагментов и/или пептидов и последующего промывания факт образования иммунокомплекса и даже количество образованного иммунокомплекса могут быть определены путем воздействия на этот иммунокомплекс вторым антителом, специфичным к первому антителу. Если исследуемый образец происходит от человека, то вторым антителом является антитело, специфичное к иммуноглобулинам человека, в основном к IgG. Для конкретного обнаружения второе антитело может иметь ассоциированную активность, например ферментативную активность, которая будет продуцировать, например, развитие окраски после инкубирования с соответствующим хромогенным субстратом. Количественная оценка может быть затем проведена путем измерения степени генерированной окраски с использованием, например, спектрофотометра, работающего в области видимого диапазона.
3. Использование последовательностей в качестве гибридизационных зондов
Нуклеотидные последовательности настоящего изобретения, содержащие последовательность гена рецептора трансферрина, позволяют осуществлять идентификацию и клонирование гена рецептора трансферрина, происходящего от любого вида Haemophilus и других бактерий, имеющих гены рецептора трансферрина.
Благодаря своей способности к селективному образованию дуплексных молекул с комплементарными участками других генов ТfR, нуклеотидные последовательности, содержащие гены рецептора трансферрина настоящего изобретения, могут найти ценное применение. Для достижения различных уровней селективности зонда по отношению к другим TfR-генам в зависимости от его применения могут быть использованы различные условия гибридизации. Для достижения высокой степени селективности используют относительно жесткие условия для образования дуплексов, а именно условия с применением низких концентраций соли и/или высоких температур, например 0,02 М - 0,15 М NaCl при температурах примерно 50-70oC. В некоторых случаях гибридизацию осуществляют в менее жестких условиях, например, с использованием 0,15 М - 0,9 М соли при температурах примерно 20oC-55oC. Условия гибридизации могут быть сделаны более жесткими путем добавления возрастающих количеств формамида для дестабилизации гибридного дуплекса. Таким образом, условия гибридизации можно легко изменять в зависимости от результатов, которые необходимо получить. Однако, в основном, температура гибридизации в присутствии 50% формамида составляет 42oС для зондов, которые на 95-100% являются гомологичными целевому фрагменту; и 37oС для зондов, имеющих 95% гомологии; и 32oС для зондов, имеющих 85-90% гомологии.
При клиническом диагностическом применении нуклеиновокислотные последовательности генов TfR настоящего изобретения могут быть использованы в сочетании с соответствующими средствами, такими как метка, для оценки гибридизации. Специалистам известно много подходящих индикаторных средств, например радиоактивные, ферментные или другие лиганды, такие как авидин/биотин, которые способны продуцировать детектируемый сигнал. В некоторых диагностических вариантах осуществления настоящего изобретения вместо радиоактивной метки может быть использована ферментная метка, такая как уреаза, щелочная фосфатаза или пероксидаза. В случае применения ферментной метки для идентификации специфической гибридизации с образцами, содержащими последовательности гена TfR, могут быть использованы колориметрические субстраты-индикаторы, которые являются видимыми для человеческого глаза и обнаруживаемыми спектрофотометрически.
Нуклеиновокислотные последовательности генов TfR настоящего изобретения могут быть использованы в качестве гибридизационных зондов при гибридизации в растворе и при твердофазной гибридизации. В вариантах, предусматривающих осуществление твердофазных процедур, исследуемую ДНК (или РНК), взятую из образцов, таких как клинические образцы, включая физиологические жидкости и экссудаты (например, сыворотка, околоплодные воды, выпот из полости среднего уха, мокрота, бронхоальвеолярная промывная жидкость) или даже ткани, адсорбируют или связывают каким-либо другим способом с выбранной матрицей или поверхностью. Затем эту фиксированную одноцепочечную нуклеиновую кислоту подвергают специфической гибридизации с выбранными зондами, содержащими нуклеиновокислотные последовательности генов ТfR или их фрагментов, в соответствующих условиях. Выбор подходящих условий гибридизации зависит от конкретных обстоятельств, требующих соответствия определенным критериям, например от содержания G+С, типа нуклеиновой кислоты - мишени, источника нуклеиновой кислоты, размера гибридизационного зонда. После промывания гибридизационной поверхности для удаления неспецифически связанных молекул зондов проводили оценку специфической гибридизации или даже проводили ее количественную оценку с использованием метки. Что касается выбора пептидов, то предпочтительными являются такие части нуклеиновокислотной последовательности, которые консервативны для видов Haemophilus, например нуклеиновокислотные последовательности, кодирующие консервативные пептидные последовательности, представленные на фиг. 8, 9, 13 и 14 и в таблицах 2 и 3. Выбранный зонд может иметь, по крайней мере, 18 п.о., и его длина может варьироваться в пределах от 30 п.о. до 90 п.о.
4. Экспрессия генов рецептора трансферрина
Плазмидные векторы, содержащие репликон и регуляторные последовательности, которые происходят от видов, совместимых с хозяйской клеткой, могут быть использованы для экспрессии генов рецептора трансферрина в экспрессирующих системах. Указанные векторы обычно несут сайт репликации, а также последовательности-маркеры, обеспечивающие фенотипическую селекцию в трансформированных клетках. Например, E.coli могут быть трансформированы с использованием рВР322, которые содержат гены резистентности к ампициллину и тетрациклину, что обеспечивает легкую идентификацию трансформированных клеток. Плазмида рВР322, или другая микробная плазмида, или фаг должны также содержать или должны быть модифицированы таким образом, чтобы они содержали промоторы, которые могут быть использованы клеткой-хозяином для экспрессии своих собственных белков.
Кроме того, фаговые векторы, содержащие репликон и регуляторные последовательности, которые являются совместимыми с клеткой-хозяином, могут быть использованы в качестве трансформирующих векторов в отношении этих хозяев. Например, фаг в лямбда GEMTM-II может быть использован в изготовлении рекомбинантных фаговых векторов для трансформации клеток-хозяев, таких как LE392 E.coli.
Промоторами, обычно используемыми при конструировании рекомбинантных ДНК, являются промоторные системы β-лактамазы (пенициллиназы) и лактозы (Chang и др., 1978; Itakura и др., 1977; Goeddel и др., 1979; Goeddel и др., 1960), и другие микробные промоторы, такие как промоторная система Т7 (патент США 4952496). Детали, относящиеся к нуклеотидным последовательностям промоторов, являются достаточно известными, и каждый специалист в состоянии правильно осуществить функциональное лигирование этих промоторов с генами. Выбор конкретного промотора, главным образом, зависит от желаемых результатов. Хозяевами, которые могут быть использованы для экспрессии генов рецептора трансферрина, их аналогов, фрагментов или вариантов, являются E.coli, Bacillus sp. , Haemophilus, грибки, дрожжи или бакуловирусная экспрессирующая система.
В соответствии с настоящим изобретением предпочтительными методами получения белка являются рекомбинантные методы, поскольку природный белок TfR, очищенный из культуры вида Haemophilus, может содержать следовые количества токсических материалов или других примесей. Для того чтобы этого избежать, можно использовать рекомбинантно продуцированный белок в гетерологических системах, которые могут быть выделены из хозяина так, чтобы минимизировать присутствие примесей в очищенном материале. Особенно предпочтительными хозяевами для экспрессии нужного гена являются грамположительные бактерии, которые не имеют ЛПС, а поэтому не содержат эндотоксинов. Такими хозяевами являются виды Bacillus, которые могут быть, в частности, использованы для продуцирования непирогенного рецептора трансферрина, его фрагментов или аналогов. Кроме того, рекомбинантные методы продуцирования позволяют получать Tbp1 или Тbр2 или их фрагменты отдельно друг от друга, в отличие от других обычно ассоциируемых друг с другом белков, присутствующих в Haemophilus.
Биологические депозиты
Некоторые плазмиды, которые содержат, по крайней мере, часть, кодирующую рецептор трансферрина от штаммов Haemophilus influenzae и которые рассматриваются в настоящей заявке, были депонированы Американской коллекцией типовых культур (АТСС) (находящейся в Рокbилле, шт. Мэриленд, США) в соответствии с Будапештским договором до подачи настоящей заявки. Образцы депонированных плазмид станут общедоступными после выдачи патента на основании данной заявки на патент США. Настоящее изобретение не ограничивается указанными депонированными плазмидами, и эти плазмиды рассматриваются лишь в качестве иллюстрации настоящего изобретения. Любая экbивалентная или аналогичная плазмида, кодирующая аналогичные или экbивалентные антигены, описанные в настоящей заявке, также входит в объем настоящего изобретения (см. табл.I).
Штаммы Haemophilus
Гибридный штамм Eagan получали от Connaught Laboratories Limited, 1755, Stteles Ave, W, Willowdate, Ontario, Canada M2R 3T4.
Гибридный штамм MinnA был получен из коллекции Dr. Robert Munson, Department of Microbiology and Immunology, Washington University School of Medicine, Children's Hospital, St. Louis, Missouri 63110.
Гибридный штамм DL63 был получен из коллекции
Dr. Eric Hansen, Department of Microbiology, University of Texas Southern Medical Center, 5323 Harry Hines Boulevard, Dallas, Texas 75235-9048.
Штамм РАК 12085 был получен из коллекции Dr. Robert Munson (см. выше).
Штаммы SВ12, 29, 30, 32 и 33 были получены из коллекции Dr. Stephen Barenkamp, Department of Pediatrics, School of Medicine, Saint Louis University Medical Centre, St. Louis, Missouri 63104.
Примеры
В вышеприведенном описании настоящее изобретение раскрывается лишь в общих чертах. Для лучшего понимания изобретения ниже представлены конкретные примеры. Эти примеры приводятся исключительно в целях иллюстрации и не должны рассматриваться как некое ограничение объема изобретения. Изменения видов и замена экbивалентов рассматриваются как условия, которые могут иметь место в данном конкретном случае или которые, как предполагается, являются целесообразными в данном случае. Хотя в настоящем описании используются конкретные термины, они имеют лишь описательное значение и не носят ограничительного характера.
В настоящем изобретении используются методы молекулярной генетики, биохимии белков, иммунологии и техники ферментации, однако подробного их описания в данной заявке не приводится, поскольку все они широко освещаются в научной литературе и известны любому специалисту в данной области.
Пример 1
В данном примере описывается получение хромосомной ДНК из штаммов DL 63, Eagan, MinnA, РАК 12085 и SB33 H. influenzae.
Штаммы H. influenzae культивировали на агаре Мюллера-Хинтона или в бульоне с сердечно-мозговым экстрактом, как описано Harkness и др., 1992.
А. Экстрагирование хромосомной ДНК из штамма DL63 Haemophilus influenzae типа b
Хромосомную ДНК получали следующим образом. 250 мл культуры осаждали путем центрифугирования на роторе J14 Beckman при 8000 об/мин в течение 15 минут. Осадок после центрифугирования промывали 200 миллилитрами 50 мМ Трис-НСl (рН 8,0), центрифугировали как описано выше, ресуспендировали в 12,5 мл 50 мМ Трис-НСl и 50 мМ ЭДТК (рН 8,0) и замораживали при температуре -20oС. К замороженному клеточному осадку после центрифугирования добавляли 1,25 мл 10 мг/мл лизоцимного раствора в 0,25 М Трис-НСl (рН 8,0). Осадок после центрифугирования оттаивали и инкубировали на льду в течение 45 минут. После этого добавляли 2,5 мл раствора 1 мг/мл протеиназы К в 0,5% ДСН, 0,4 М ЭДГК, 50 мМ Трис-НСl (рН 7,5), и полученную смесь инкубировали при температуре 50oС в течение одного часа, периодически помешивая при этом. Лизат экстрагировали один раз 15 миллилитрами фенола, забуференного Трисом, а затем для осаждения ДНК добавляли 1,5 мл 3 М ацетата натрия и 30 мл этанола. ДНК наматывали на стеклянную палочку, а затем растворяли в 12,5 мл 50 мМ Трис-НСl и 1 мМ ЭДТК (рН 7,5) в 0,2 мг/мл РНКазы А путем качания в течение ночи. После этого образец один раз экстрагировали равным объемом хлороформа, осаждали и наматывали, как описано выше. ДНК растворяли в 2 мл 50 мМ Трис-НСl и 1 мМ ЭДГК (рН 7,5), а затем хранили при 4oС.
В. Экстрагирование хромосомной ДНК из штамма Eagan Haemophilus influenzae типа b
50 мл культуры осаждали путем центрифугирования, образовавшийся осадок ресуспендировали в 25 мл ТЕ (10 мМ Трис, 1 мМ ЭДТК (рН 7,5)), а 2•5 мл - аликbоты использовали для получения хромосомной ДНК. К каждой аликbоте добавляли 0,6 мл 10% саркозила и 0,15 мл 20 мг/мл протеиназы К, и образцы инкубировали в течение одного часа при температуре 37oС. Лизат один раз экстрагировали фенолом, насыщенным Трисом, а затем три раза экстрагировали смесью хлороформа и изоамилового спирта (24:1). Водные фазы объединяли до конечного объема 7 мл. Для осаждения ДНК добавляли 0,7 мл 3 М ацетата натрия (рН 5,2) и 4,3 мл изопропанола, после чего ДНК наматывали, промывали 70%-ным этанолом, осушали и ресуспендировали в 1 мл воды.
С. Экстрагирование хромосомной ДНК из штаммов Eagan MinnA РАК 12085 и SK33 Haemophilus influenzae
Клетки осаждали из 50 мл культуры путем центрифугирования при 5000 об/мин в течение 15-20 минут при температуре 4oС. Клеточный осадок после центрифугирования ресуспендировали в 10 мл ТЕ (10 мМ Трис-HCl, 1 мМ ЭДТК (рН 7,5), а проназу и ДСН добавляли до конечных концентраций 500 мкг/мл и 1% соответственно. Образец инкубировали при температуре 37oС в течение 4 часов до тех пор, пока не был получен прозрачный лизат. Этот лизат один раз экстрагировали фенолом, насыщенным Трис, один раз смесью насыщенного трисом фенола и хлороформа (1:1) и один раз хлороформом. Конечную водную фазу диализовали в течение 24 часов против 2•500 мл 1 M хлорида натрия при температуре 4oС с однократной заменой буфера, а затем в течение 24 часов против 2•500 мл ТЕ при температуре 4oС с однократной заменой буфера. Конечный диализат распределяли на аликbоты для дальнейшего использования.
Пример 2
В данном примере описано получение хромосомных библиотек.
A. Библиотека DL63-ZAP H. influenzae
100 мкг хромосомной ДНК DL63 H. influenzae в ТЕ подвергали механической фрагментации путем пропускания через 1-миллилитровый шприц с иглой размером 25. Фрагментированную ДНК затупляли по концам путем добавления воды до конечного объема 405 мкл, 45 мкл 10•S1 нуклеазного буфера (2 М хлорид цинка, 500 мМ NaOAc (рН 4,5), 10 мМ сульфата цинка и 5% глицерин) и 1,7 мкл нуклеазы S1 при концентрации 100 ед/мкл с последующим инкубированием в течение 15 минут при температуре 37oС. Образец один раз экстрагировали смесью фенола и хлороформа и один раз хлороформом, после чего для осаждения ДНК добавляли 1 мл этанола. Образец инкубировали на льду в течение 10 минут или в течение ночи при температуре -20oC, и ДНК собирали путем центрифугирования в микроцентрифуге в течение 30 минут. Полученную ДНК промывали 70%-ным этанолом и осушали. EcoRI-сайты в ДНК-последовательности метилировали с использованием стандартных процедур. К этой мотивированной ДНК добавляли 5 мкл мМ MgCl2, 8 мкл смеси dNTP (2,5 мМ каждого dATP, dCTP, dGTP и dTTP) и 4 мкл фрагмента Кленова при концентрации 5 ед/мкл. Полученную смесь инкубировали в течение 30 минут при температуре 12oС. После добавления 450 мкл SТЕ (0,1 М NaCl, 10 мМ Трис-HCl, 1 мМ ЭДТК) (рН 8,0) полученную смесь один раз экстрагировали смесью фенола и хлороформа и один раз хлороформом, а затем для осаждения ДНК добавляли 1 мл этанола. Образец инкубировали на льду в течение 10 минут или в течение ночи при температуре -20oС. ДНК собирали путем центрифугирования в микроцентрифуге в течение 30 минут, промывали 70%-ным этанолом и осушали.
ДНК ресуспендировали в 7 мкл ТЕ и к раствору добавляли 14 мкл фосфорилированных EcoRI-линкеров (200 нг/мл), 3 мкл 10х лигирующего буфера, 3 мкл 10 мМ АТР и 3 мкл ДНК-лигазы Т4 (4 ед/мкл). Образец инкубировали в течение ночи при температуре 4oС, а затем инкубировали в течение 10 минут при температуре 68oС для инактивации лигазы. К полученной смеси добавляли 218 мкл воды, 45 мкл 10х универсального буфера и 7 мкл EcoRI при концентрации 30 ед/мкл. После инкубирования в течение полутора часов при температуре 37oС добавляли 1,5 мкл 0,5 М ЭДТК, и эту смесь помещали на лед.
ДНК фракционировали по размеру на градиенте сахарозы, собирая фракции, содержащие ДНК размером 6-10 кb. Объединенную ДНК осаждали этанолом и ресуспендировали в 5 мкл ТЕ-буфера. 200 нг ДНК-вставки лигировали в течение 2-3 дней при температуре 4oС с 1 мкг вектора ZAP II в конечном объеме 5 мкл. Смесь для лигирования упаковывали с использованием Gigapack II Gold (Stratagene) и засевали в NZY-чашки с клетками SUPE E.coli. Затем библиотеку титровали, амплифицировали и хранили при температуре 4oС в 0,3%-ном хлороформе.
В. Библиотека Eagan - pUC H. influenzae
Хромосомную ДНК, полученную из штамма Eagan H. influenzae способом, описанным в примере 1С, гидролизовали ферментом Sau3A в течение 2, 5 и 10 минут, и образцы подвергали электрофорезу на препаративном агарозном геле. Гелевые срезы, содержащие ДНК-фрагменты длиной 3-10 кb, вырезали, а ДНК экстрагировали с помощью стандартной процедуры замораживания-оттаивания. ДНК-плазмиду из pUC 8:2 (рUC 8 с дополнительными сайтами рестриктирующих ферментов Bg1 II и Xba 1 в области множественного клонирования) гидролизовали ферментами BamHI и Bg1 II, а затем дефосфорилировали с помощью телячьей щелочной фосфатазы (CAP). Фрагменты ДНК штамма Eagan H. influenzae лигировали в pUC, и эту смесь использовали для трансформации клеток JM109 E.coli.
С. Библиотека Eagan - ZAP H. influenzae
Хромосомную ДНК штамма Eagan H. influenzae, полученную способом, описанным в примере 1В, гидролизовали ферментом EcoRI, а затем фракционировали по размеру на препаративном агарозном геле. Гелевые срезы, соответствующие ДНК-фрагментам длиной 7-23 кb, вырезали, а ДНК подвергали электроэлюированию в течение ночи в диализной трубке, содержащей 3 мл ТАЕ (40 мМ Трис-ацетат, 1 мМ ЭДТК), при 14 В. Затем ДНК дважды осаждали и ресуспендировали в воде, после чего лигировали в течение ночи с EcoRI-гидролизованной ДНК ZAP II. Смесь для лигирования упаковывали с использованием набора Gigapack II (Stratagene) и пассировали XLI-Blue E.coli. Полученную библиотеку титровали, амплифицировали и хранили при температуре 4oС в 0,3%-ном хлороформе.
D. Библиотеки ЕМВL 3
Хромосомную ДНК штамма MinnA H. influenzae (10 мкг) получали способом, описанным в примере 1С, гидролизовали ферментом Sau3A I (40 единиц) в течение 2, 4 и 6 минут, а затем фракционировали по размеру на градиенте 10-30% сахарозы в TNE-буфере (20 мМ Трис-HCl, 5 мМ NaCl, 1 мМ ЭДТК (рН 8)). Фракции, содержащие ДНК-фрагменты длиной более чем 5 кb, собирали и осаждали. Во втором эксперименте хромосомную ДНК (2,6 мкг) гидролизовали ферментом Sau3A I (4 единицы) в течение 1, 2, и 3 минут, а затем фракционировали по размеру путем электрофореза на агарозном геле. Гелевые срезы, содержащие ДНК-фрагменты длиной 10-20 кb, вырезали, а ДНК экстрагировали с использованием стандартной процедуры замораживания-оттаивания. Фракционированные по размеру ДНК от каждого эксперимента объединяли для лигирования с BamHI-ветвями EMBL 3 (Promega). Смесь для лигирования упаковывали с использованием набора для упаковки Gigapack, а затем пассировали в клетках LE392 E.coli. Полученную библиотеку титровали, амплифицировали и хранили при температуре 4oС в присутствии 0,3%-ного хлороформа.
Хромосомную ДНК штаммов РАК 12085 или SВ33 H. influenzae, полученную способом, описанным в примере 1С, гидролизовали ферментом Sau3A 1 (0,5 единиц/10 мкг ДНК) при температуре 37oС в течение 15 минут, после чего фракционировали по размеру путем электрофореза на агарозном геле. Гелевые срезы, соответствующие ДНК-фрагментам длиной 15-23 кb, вырезали, а ДНК подвергали электроэлюированию в течение ночи в диализной трубке, содержащей 3 мл ТАЕ, при 14 В. Полученную ДНК дважды осаждали и ресуспендировали в воде, а затем лигировали в течение ночи с BamHI-ветвями EMBL3 (Promega). Смесь для лигирования упаковывали с использованием набора для in vitro-упаковки Lambda (Amersham) в соответствии с инструкциями поставщика и пассировали в клетках NM539 E. coli. Библиотеку титровали, амплифицировали и хранили при температуре 4oС в присутствии 0,3%-ного хлороформа.
Пример 3
В данном примере описан скрининг библиотек.
А. Библиотека экспрессируемых последовательностей DL63-ZAP H. influenzae
Белки Tbp1 и Тbр2 подвергали аффинной очистке на твердофазном трансферрине человека (hTf). Для этого 20 мл RTf-сефарозную колонку приготовляли в соответствии с инструкциями изготовителя для связывания белковых лигандов с бромцианактивированной Сефарозой (Sigma). Полученную матрицу промывали 3 колоночными объемами 50 мМ Трис-HCl, 1 М NaCl, 6 М гуанидина/НСl (рН 8,0) для удаления нековалентно связанного твердофазного трансферрина человека. Затем колонку уравновешивали 50 мМ Трис-HCl (рН 8,0), а связанный hTf нагружали железом с использованием 1 мл 10 мг/мл FeCl3 в буфере, содержащем 100 мМ цитрата натрия и 100 мМ бикарбоната натрия, после чего колонку уравновешивали двумя колоночными объемами 50 мМ Трис-HCl (рН 8,6) и 1 М NaCl (рН 8,0). Целые бактериальные мембраны (300 мг целого белка) получали из штамма DL63 H. influenzae, выращенного на среде, не содержащей железа, как описано выше (Schryvers et al., 1989). Мембраны разводили до 2 мг/мл в 50 мМ Трис-HCl, 1 M NaCl (рН 8,0) и солюбилизировали путем добавления ЭДТК до 15 мМ и Саркозила NL 97 до 1,5%. После центрифугирования в течение одного часа при 40000•g супернатант наносили на hTf-колонку, и колонку промывали десятью колоночными объемами 50 мМ Трис-HCl, 1 M NaCl, 10 мМ ЭДТК и 0,5% Саркозила (рН 8,0). Рецепторные белки элюировали с использованием 2 М GnHCl в том же самом буфере, а проэлюированные фракции интенсивно диализовали против 25 мМ аммонийбикарбонатного буфера (5 замен буфера), лиофилизовали и хранили при температуре -20oC. Выделенные белки использовали для продуцирования антисыворотки против рецептора трансферрина у новозеландских белых кроликов с использованием стандартных процедур. Для этого кроликов иммунизировали путем 3-кратного введения подкожной инъекции с интервалами в две недели с использованием полного адъюванта Фрейнда для первой инъекции и неполного адъюванта Фрейнда для последующих инъекций.
Библиотеку DL63-ZAP высевали на клетки SUPE E.coli, и бляшки переносили на нитроцеллюлозные мембраны, предварительно смоченные в 10 мМ IPTG, для индуцирования экспрессии под контролем промотора гена LacZ pBluescript. Фильтры блокировали с использованием 0,5%-ного сепарированного молока в 50 мМ Трис-HCl, 150 мМ NaCl (рН 7,5), а затем зондировали поликлональными антисыворотками против TfR и козьими антикроличьими иммуноглобулинами (IgG), конъюгированными с пероксидазой хрена. Бляшки очищали тремя стадиями скринирования, а рекомбинантные плазмиды pBluescript (pBH1T1 и рВН1Т2; фиг.1А и 12) выделяли посредством in vivo-процедуры вырезания (Short et al., 1988).
В. Библиотеки неэкспрессированных последовательностей Eagan MinnA и РАК 12085
(i) Скрининг-библиотеки Eagan - pUC H. influenzae
Нанесение колонии на нитроцеллюлозу осуществляли с использованием стандартных процедур, а фильтры зондировали с помощью 5'-рВН1Т2-зонда гена рецептора трансферрина, показанного на фиг.2. Зонд подвергали мечению дигоксигенином (dig, Boehringer Mannheim) в соответствии с инструкциями производителя. Несколько предполагаемых клонов подвергали дотблоттированию на нитроцеллюлозе, а затем осуществляли второй цикл скринирования с использованием аналогичного 5'-рВН1Т2-зонда. Предполагаемые клоны во втором цикле анализировали посредством картирования рестриктирующим ферментом, а клон S-4368-3-3 (фиг. 1В, фиг. 2) отбирали для анализа последовательности.
(ii) Скрининг библиотеки Eagan - ZAP H. influenzae
Библиотеку фага высевали с использованием стандартных процедур в чашки с LB-средой и 0,7% агарозным слоем, в которых находились клетки Blue XLI (Stratagene). Бляшки наносили на нитроцеллюлозу с использованием стандартных процедур, а фильтры отжигали при температуре 80oC в течение 2 часов в вакууме для фиксации ДНК. 5'-рВН1Т2-зонд гена рецептора трансферрина (фиг. 2) метили дигоксигенином, а фильтры предварительно гибридизировали в течение 4 часов при температуре 42oС, а затем гибридизировали с меченым зондом в течение ночи при 42oС. Фильтры промывали при температуре 68oС и после радиоавтографии несколько бляшек отбирали для второго цикла скрининга. In vivo-процедуру вырезания фагмидной ДНК из предполагаемых клонов второго цикла осуществляли в соответствии с инструкциями с использованием ZAP системы (Promega). Примером являются полученные четыре клона JB-901-5-3 (фиг. В, фиг. 2) с идентичными EcoRI-вставками длиной примерно 2,5 кb. Предполагаемые бляшки были также амплифицированы, а фаговую ДНК очищали из 500 мл культуры. ДНК-вставку вырезали путем гидролиза ферментом Xba I и клонировали в pUC 8:2 (pUC 8, содержащий дополнительные Bg1 II- и Xba I-сайты на участке множественного клонирования), а затем гидролизовали ферментом Xba I и дефосфорилировали. Клон JB-911-3-2 (фиг. 17) содержал 3'-половину ТfR-оперона штамма Eagan H. influenzae.
(iii) Скрининг библиотек EMBL3
Библиотеку штамма MinnA H. influenzae высевали в NZCYM-чашки с клетками LE392 с использованием в качестве покрытия 0,7% агарозу в NZCYM. Нанесение бляшек на нитроцеллюлозные фильтры осуществляли с использованием стандартных процедур, а фильтры обрабатывали и зондировали с помощью 5' рВН1Т2-зонда (фиг. 2), меченного дигоксигенином. Предполагаемые бляшки высевали и подвергали второму и третьему циклам скрининга с использованием аналогичных процедур. Фаговую ДНК получали из 500 мл культуры с использованием стандартной процедуры, а затем ДНК-вставку вырезали путем гидролиза ферментом Sa1 I и клонировали в pUC, в результате чего получали клон DS-712-1-3 (рисунки 1С и 2).
Библиотеку штамма РАК 12085 H. influenzae высевали в NZCYM-чашки с клетками LE392 с использованием покрытия из 0,7% агарозы в NZCYM. Затем бляшки наносили на нитроцеллюлозу, а фильтры обрабатывали и зондировали с помощью дигоксигенинмеченного 5'-рВН1Т2-зонда (фиг. 2). Предполагаемые бляшки засевали и подвергали второму циклу скрининга с использованием аналогичных процедур. Фаговую ДНК получали из 500 мл культуры с помощью стандартной процедуры, а ДНК-вставку вырезали путем гидролиза ферментом Sa1 I, клонировали в pUC и получали клон JB-1042-7-6 (фиг.1 и 2).
Библиотеку штамма В33 H. influenzae высевали на клетки SЕ392 в NZCYM-чашки, покрытые верхним слоем 0,7% агарозы в NZCYM. Бляшки наносили на нитроцеллюлозу, и фильтры обрабатывали и зондировали диоксигенинмеченным 5'-рВН1Т2-зондом (фиг. 2). Предполагаемые бляшки планировали и подвергали второму циклу скринирования с использованием аналогичных процедур. Фаговую ДНК получали из 500 мл культуры с помощью стандартной методики, а ДНК-вставку вырезали путем гидролиза ферментом Sa1 I и клонировали в pUC, в результате чего получали клон JB-1031-2-9 (фиг. 2).
Пример 4
В данном примере описано секвенирование Тbр1- и Тbр2-генов TfR-оперона.
Плазмидную ДНК от клонов pBHIT 1, pBHIT 2, S-4368-3-3, JB-901-5-3, DS-712-1-3, JB-I042-7-6 и JB-1031-2-9 получали с использованием стандартной техники. Олигонуклеотидные праймеры для секвенирования длиной 17-25 оснований были синтезированы с помощью ДНК-синтезатора (380 В, модель АВI) и очищены посредством хроматографии с использованием ОРС-кассет, полученных от фирмы Applied Biosystems, Inc. и использованных в соответствии с инструкциями изготовителя. После этого образцы подвергали секвенированию с использованием ДНК-секвентатора (370 А, модель АВI) и терминального красителя в соответствии с инструкциями изготовителя. Последовательность TfR-оперона штамма DL63 показана на фиг.3, последовательность TfR-оперона штамма Eagan показана на фиг.4, последовательность ТfR-оперона штамма MinnA показана на фиг.5, последовательность ТfР-оперона штамма РАК 12085 показана на фиг.6, а последовательность TfR-оперона штамма SB33 показана на фиг.7.
Пример 5
В этом примере проиллюстрирована PCR-амплификация генов tbp 2 нетипируемых штаммов SВ12, SВ29, SB30 и SВ32 H. influenzae.
Хромосомную ДНК нетипируемых штаммов SВ12, SВ29, SВ30 и SВ32 H. influenzae получали, как описано выше. Гены TfR конструировали в виде оперона, содержащего ген tbp 2 и следующий за ним ген tbp 1 (см. фиг.12А и 12В). Синтезировали олигонуклеотиды к 5'-концу кодирующей последовательности tbp 2 и обратного праймера к 5'-концу кодирующей последовательности tbp 1.
Эти праймеры имели последовательности: GGАТССАТАТGАААТСТGТ АССТСТТАТСТСТGGТ (SEQ ID 120), соответствующую MKSVPLISGS (SEQ ID 147) от лидерной последовательности Tbp2; и ТСТАGААGCTTTTTTAGTСАТТТТТАGТАТТССАТ (SEQ ID 137), которая представляет собой обратный праймер лидерной последовательности МТКК (SEQ ID 138) Tbp1 и часть межгенной последовательности (фиг.12А и 12В). PCR-амплификацию осуществляли в буфере, содержащем 10 мМ Трис/HCl, рН 8,3, 50 мМ хлорида калия и 1,5 мМ хлорида магния. Каждые 100 мкл реакционной смеси содержали 5 нг хромосомной ДНК, 1 мкг каждого праймера, 5 ед amplitaq - ДНК-полимеразы (Perkin Elmer Cetus) и 0,5 мМ dNTP (Perkin Elmer Cetus). Полимеразную цепную реакцию проводили при следующих условиях: 25 циклов - при 94oС в течение 1,0 минут; при 45oС в течение 2,0 минут и при 72oС в течение 1,5 минут. Специфические 2 кb-фрагменты амплифицировали для каждого образца (фиг. 13). ДНК SВ33 использовали в качестве позитивного контроля (дорожка 1). Хромосомные ДНК, использованные для амплификации гена Тbр2, соответствуют дорожке 1 для SВ33, дорожке 2 для SВ12, дорожке 3 для SВ29, дорожке 4 для SВ30 и дорожке 5 для SВ32. Эти фрагменты были клонированы в вектор клонирования ТА (Invitrogen), и были определены их нуклеотидные последовательности. Нуклеиновокислотные последовательности Тbр2 из штаммов SВ12 (SEQ ID 108), В29 (SEQ ID 110), SВ30 (SEQ ID 112) и SВ32 (SEQ ID 114) показаны на фиг.8, 9, 10 и 11 соответственно.
Пример 6
В этом примере проиллюстрировано сравнение аминокислотных последовательностей трансферрина и идентификация потенциально доступных эпитопов белков рецептора трансферрина посредством анализа вторичной структуры.
На фиг.14 показано сравнение аминокислотных последовательностей Tbp1 из штамма Eagan и штамма DL63 H. influenzae типа b; из нетипируемых штаммов РАК 12085 и SВ33 H. influenzae; из штаммов В16В6 и М982 N. meningitidis (Legrain и др. , 1993); и из штамма FA19 N. gonorrhoe (Cornellissen и др., 1992). В результате этого анализа были выявлены участки Tbp1, которые являются консервативными для всех этих бактерий.
На фиг.15 показано сравнение аминокислотных последовательностей Тbр2 от штаммов Eagan и DL63 H. influenzae типа b; нетипируемых штаммов РАК 12085, SB12, SВ29, SВ30 и SВ32 H. influenzae, штаммов В16В6 и М982 N. meningitidis, штамма FA19 N. gonorrhoe и штаммов 205 и 37 Actinobacillus (Haemophilus) pleuropneumoniae (Gerlach и др. , 1992). В результате этого анализа были выявлены области Тbр2, которые являются консервативными для всех этих бактерий.
Анализ вторичной структуры белков осуществляли с использованием алгоритмов Chou & Fasman (1978), а диаграммы гидрофильности/гидрофобности строили с использованием алгоритма Норр (1986). Эти величины получали исходя из средних значений гептапептидных "окон", и на основании этих значений строили график для средней точки каждого фрагмента. На фиг.16А проиллюстрирована предсказанная вторичная структура Tbp1 от штамма Eagan H. influenzae типа b, а на фиг. 16В проиллюстрирована предсказанная вторичная структура Тbр2 от штамма Eagan H. influenzae типа b. Предсказанные вторичные структуры, изображенные на фиг.16А и 16В были получены с использованием процедур, описанных выше. Однако авторы настоящего изобретения пока не могли проверить точно ли соответствуют структуры, изображенные на этих чертежах, вторичной структуре этих белков.
Консервативные эпитопы белков Tbp1 и Тbр2 от нескольких различных бактерий были идентифицированы посредством сравнительного анализа первичных структур последовательностей, показанных на фиг.14 и 15 соответственно. Некоторыми из таких консервативных эпитопов являются:
TBP1 DNEVTGLGK SEQ ID NO 43
TBP1 EQVLNRLTRIDPGI SEQ ID NO 44
TBP1 GAINEIEYENVKAVEISKG SEQ ID NO 45
TBP1 GALAGSV SEQ ID NO 46
TBP2 LEGGVYGP SEQ ID NO 74
TBP2 KSGGGSFD SEQ ID NO 75
TBP2 YVYSGL SEQ ID NO 76
TBP2 CCSNLSYVKFG SEQ ID NO 77
TBP2 FLLGHRT SEQ ID NO 78
TBP2 EFNVDF SEQ ID NO 79
TBP2 NAFTGTA SEQ ID NO 80
TBP2 VNGAFYG SEQ ID NO 81
TBP2 LEGGYF SEQ ID NO 82
TBP2 VVFGAR SEQ ID NO 83
Кроме того, вместе с предсказанными вторичными структурами были идентифицированы четыре консервативных доступных эпитопа на Tbp1 и два эпитопа на Tbр2. Этими эпитопами являются:
Top1 DNEVTGLGK SEQ ID NO 43
Top1 EQVLN/DIRDLTRID SEQ ID NO 139 и 140
Top1 GAINEIEYENVKAVEISK SEQ ID NO 141
Top1 GI/VYNLF/LNIRYVTWE SEQ ID NO 142 и 143
Top2 CS/LGGG(G)SFD SEQ ID NO 75, 144 и 145
Top2 LE/SGGFY/FGP SEQ ID NO 74 и 146
Белки, полипептиды или пептиды, содержащие вышеуказанные консервативные аминокислотные последовательности, являются особенно ценными для использования их в качестве детектирующих средств при диагностике, а также в качестве иммуногенов для обнаружения заболеваний или для защиты от заболеваний, вызываемых бактериями, продуцирующими белок рецептора трансферрина. Для иммунизации вышеуказанные аминокислотные последовательности могут быть представлены иммунной системе в качестве белков или пептидов либо могут быть использованы живые векторы доставки, такие как Salmonella, ВСG, аденовирусы, поксвирусы, вирусы коровьей оспы или полиовирусы.
Пример 7
В этом примере проиллюстрировано конструирование плазмиды JB-1468-29, которая экспрессирует белок Тbр1 штамма Eagan в E.coli.
Плазмиды S-4368-3-3 (фиг. 1В и 2) и SВ-911-3-2 (фиг.17) содержат 5'- и 3'-части гена tbp 1 штамма Eagan соответственно. На фиг.17 проиллюстрирована схема конструирования для плазмиды JB-1468-29. Олигонуклеотидные последовательности, используемые для конструирования плазмиды JB-1468-29 показаны на фиг. 20 (SEQ ID 86 и 87). Плазмиду JB-1468-29 вводили в штамм BL 21/DE3 E. coli путем электропорации для продуцирования штамма JB-1476-2-1.
JB-1476-2-1 культивировали в УТ-среде и индуцировали с использованием IPTG в соответствии со стандартным протоколом. Для получения Tbp1 в целях исследования иммуногенности и других исследований, штамм JB-1476-2-1 культивировали в течение ночи в среде NZCYM, содержащей 3%-ную глюкозу. Затем к свежей среде NZXYM, не содержащей глюкозы, добавляли 1:40-инокулят, и культуру культивировали до A578=0,3 (А - оптическая плотность). После этого добавляли лактозу (до 1%), и культуру индуцировали в течение 4 часов. Анализ цельных клеточных лизатов для JB-1476-2-1, проведенный с помощью электрофореза в ПААГ с ДСН, показан на фиг. 22. Дорожка 1: JB-1476-2-1 (Tbp1 Т7/Eagan) при t0; дорожка 2: JB-1476-2-1, при времени индукции t=4 часа; дорожка 3: маркеры молекулярной массы: 200 кДа, 116 кДа, 97,4 кДа, 66 кДа, 45 кДа и 31 кДа; дорожка 4: JB-1437-4-1 (T7/Eagan - Тbр2) при t0; дорожка 5: JB-1437-4-1 при времени индукции t=4 часа; дорожка 6: JB-1607-1-1 (Т7/SB12 - Тbр2) при t0; дорожка 7: JB-1607-1-1 при времени индукции t=4 часа.
Пример 8
В этом примере проиллюстрировано конструирование плазмиды JB-1424-2-8, которая экспрессирует белок Тbр2 штамма Eagan от E.coli.
На фиг.18 показана плазмида S-4368-3-3, которая содержит целый ген tbp 2 от штамма Eagan H. influenzae типа b. На фиг.18 проиллюстрирована плазмида JB-1424-2-8, а на фиг. 19 показаны используемые олигонуклеотиды. Плазмиду JB-1424-2-8 вводили в штамм BL 21/SE3 E.coli путем электропорации для продуцирования штамма JB-1437-4-1 E.coli. После индуцирования IPTG или лактозой штамм JB-1476-2-1 E.coli экспрессировал белок Тbр2, как показано на фиг.22. Дорожка 1: JB-1476-2-1 (Т7/Eagan -Tbp1) при t0; дорожка 2: JB-1476-1-1 при времени индуцирования t=4 часа; дорожка 3: маркеры молекулярной массы: 200 кДа, 116 кДа, 97,4 кДа, 66 кДа, 45 кДа и 31 кДа; дорожка 4: JB-1437-4-1 (T7/Eagan-Тbр2) при t0; дорожка 5: JВ-1437-4-1 при времени индуцирования t=4 часа; дорожка 6: JB-1607-1-1 (T7/ АВ12-Тор2) при t0; дорожка 7: JB-1607-1-1 при времени индуцирования t=4 часа.
Пример 9
В этом примере проиллюстрировано конструирование плазмид, которые кодируют лидерную последовательность липопротеина перед последовательностью Тbр2.
Олигонуклеотиды, используемые для конструирования плазмид, имеющих лидерные последовательности липопротеина, происходящие от lpp E.coli (SEQ ID 88 и 89), rlpB E. coli (SEQ ID 90 и 91) и pa1 E.coli (SEQ ID 92 и 93) и расположенные перед Тbр2, показаны на фиг.20. Сконструированные плазмиды и продуцированные штаммы, соответствующие этим плазмидам, проиллюстрированы в таблице 1 (см. ниже).
Пример 10
В этом примере проиллюстрировано конструирование плазмиды JB-1600-1, которая экспрессирует Тbр2 от штамма SВ12 E.coli.
Плазмида DS-1047-1-2 (фиг.21) содержит PCR-амплифицированный ген tbp 2 SВ12. Ген tbp 2 вырезали как Nde1 -ЕсоR-рестрикционный фрагмент и вставляли в экспрессирующий вектор рТ7-7, в результате чего получали плазмиду JВ-1600-1. После электропорации этой плазмиды в клетки BL21/DE3 E.coli получали штамм JB-1607-1-1, который экспрессирует Тbр2 SВ12. После индуцирования IPTG или лактозой экспрессировали Тbр2 SВ12, как показано на фиг.22. Дорожка 1: JB-1476-2-1 (T7/Eagan-Tbp1) при t0; дорожка 2: SВ-1476-2-1 при времени индуцирования t= 4 часа; дорожка 3: маркеры молекулярных масс: 200 кДа, 116 кДа, 97,4 кДа, 66 кДа, 45 кДа и 31 кДа; дорожка 4: JB-1437-4-1 (Т7/ Eagan-Тbр2) при t0; дорожка 5: JB-1437/4-1 при времени индуцирования t=4 часа; дорожка 6: JВ-1607-1-1 (Т7/SВ12-Тbр2) при t0; дорожка 7: JB-1607-1-1 при времени индуцирования t=4 часа.
Пример 11
В этом примере проиллюстрированы экстракция и очистка белков Тbр1 и Тbр2.
Схема очистки для Tbp1 и Тbр2 показана на фиг.23. Оба эти рекомбинантных белка экспрессировались как тельца включения в E. coli, и процедуры их очистки являются одинаковыми. Клетки от 500 миллилитровой культуры, полученной, как описано в примере 7 для Tbp1 и в примере 8 для Тbр2, ресуспендировали в 50 мл 50 мМ Трис-HCl, рН 8,0 и разрушали с помощью ультразвука (3•10 минут, рабочий цикл 70%). Экстракт центрифугировали при 20000•g в течение 30 минут, а полученный супернатант, который содержал более 95% растворимых белков E.coli, отбрасывали.
Оставшийся осадок (фиг. 23, РРТ1) еще раз экстрагировали в 50 мл 50 мМ Трис, рН 8,0, содержащем 0,5% Тритон Х-100 и 10 мМ ЕDТА. После центрифугирования при 20 000•g в течение 30 минут супернатант, содержащий остаточное количество растворимых белков и большую часть мембранных белков, отбрасывали. После экстракции, как описано выше, полученный осадок содержал тельца включения (фиг. 23, РРТ2). Белки Tbp1 и Tbp2 солюбилизировали в 50 мМ Триса (рН 8,0), содержащего 0,1% ДСН и 5 мМ ДТТ. После центрифугирования полученный супернатант снова очищали на колонке для гель-фильтрации с Superdex 200, уравновешенной в 50 мМ Трис, рН 8,0, содержащем 0,1% ДСН и 5 мМ ДТТ. Фракции анализировали с помощью электрофореза в ПААГ с ДСН, и те фракции, которые содержали очищенный Tbp1 или Тbр2, диализовали в течение ночи при 4oС против 50 мМ Триса (рН 8,0), а затем центрифугировали при 20 000•g в течение 30 минут. Растворимый белок, оставшийся после проведения процедур в этих условиях, и очищенные Tbp1 и Тbр2 хранили при -20oС.
Анализ, проводимый с помощью электрофореза в ПААГ с ДСН в описанных условиях очистки, показан на фиг.24. Дорожка 1 - маркеры молекулярной массы (106, 80, 49,5, 32,5, 27,5, 18,5 кДа); дорожка 2: клеточные лизаты E.coli; дорожка 3: солюбилизированные тельца включения; дорожка 4: очищенные Tbp1 или Тbр2.
Пример 12
В этом примере проиллюстрированы исследования иммуногенности рекомбинантных белков Tbp1 и Тbр2, проведенные на мышах.
Группам из пяти мышей Balb/с подкожно (s,с) инъецировали (на дни: 1, 29 и 43) очищенный белок rTBp1 или rTBp2 (1 мкг - 10 мкг), полученные как описано в примере 11, в присутствии AlPO4 (1,5 мг на дозу). Пробы крови для анализа титров антител против рек. Тbр1 и против рек. Тbр2 (проводимым методом Е1А) брали в дни: 14, 28, 42 и 56. Результаты исследований иммуногенности проиллюстрированы на фиг.35.
Пример 13
В этом примере проиллюстрирован иммуноферментный анализ (ЕlА) для определения уровня антител против рек. Tbp1 и против рек. Тbр2 в мышиных сыворотках.
Титры антител к рек. Tbp1 и к рек. Тbр2 были определены способом, в основном описанным Panezzuti и др. (1993). Лунки для микротитрования покрывали 0,5 микрограммами рекомбинантного белка Tbp1 или Тbр2, полученного как описано в примере 11, и выдерживали 16 часов при комнатной температуре, после чего блокировали 0,1% (мас./об.) ВSА в РВS. Сыворотки серийно разводили, добавляли в лунки, и инкубировали в течение 1 часа при комнатной температуре. Аффинноочищенные фрагменты F(ab')2 козьих антимышиных иммуноглобулинов IgG (Fc-специфических), конъюгированных с пероксидазой хрена, использовали в качестве "второго" антитела. Реакции проявляли с использованием тетраметилбензидина (ТМВ/Н2О2), а оптическую плотность измеряли при 450 нм (используя 540 нм в качестве эталонной длины волны) с использованием спектрофотометра для прочтения микропланшетов (планшет-ридера) Flow Multiscan МСС. Реактивный титр антисыворотки определяли как величину, обратную разведению, при котором наблюдается двухкратное увеличение оптической плотности по сравнению с оптической плотностью, полученной с образцом неиммунной сыворотки.
Пример 14
В этом примере проиллюстрирована перекрестная реактивность антисывороток против Тbр1, продуцированных путем иммунизации рекомбинантным Tbp1 Eagan с различными штаммами H. influenzae.
Клеточные лизаты штаммов H. influenzae культивированы в BHI-средах, содержащих NAD и гем (Harkness и др., 1992)±EDDA, выделяли с помощью электрофореза на ПААГ с ДСН, затем переносили на нитроцеллюлозную мембрану и зондировали антисыворотками морских свинок против Tbp1, продуцированными в ответ на введение очищенного рекомбинантного белка Тbр1 Eagan (фиг.26). Дорожка 1: BL21/DE3; дорожка 2: SВ12-EDDA; дорожка 3: SВ12+EDDA; дорожка 4: SВ29-EDDA; дорожка 5: SВ29+EDDA; дорожка 6: SВ33-EDDA; дорожка 7: SВ33+EDDA; дорожка 8: Eagan-EDDA; дорожка 9: Eagan+EDDA; дорожка 10: В. catarrhalis 4223-EDDA; дорожка 11: B. meningitidis 4223+EDDA; дорожка 12: N. meningitidis 608-EDDA; дорожка 13: N. meningitidis 608+EDDA; дорожка 14: индуцированный JB-1476-2-1, экспрессирующий рекомбинантный Tbp1 Eagan; дорожка 15: маркеры молекулярной массы. Специфические 95 кДа-полосы, прореагировавшие с антисыворотками против Tbp1, на дорожках 3, 4, 5, 7, 8 и 9 соответствуют штаммам JB12, SВ29, SВ33 и Eagan; ≈100 кДа-полосы на дорожках 10 и 11 соответствуют штамму 4223 В.catarrhalis; ≈80 кДа-полосы на дорожках 12 и 13 соответствуют штамму 608 N. meningitidis.
Пример 15
В этом примере проиллюстрирована перекрестная реактивность антисывороток против Тbр2, продуцированных путем иммунизации рекомбинантным тbр2 Eagan, с различными штаммами H. influenzae.
Клеточные лизаты штаммов H. influenzae, культивированных в BHI-средах, в которые были добавлены NAD и гем (Harkness и др., 1992) ±EDDA, выделяли с помощью электрофореза на ПААГ с ДСН, затем переносили на нитроцеллюлозную мембрану и зондировали антисыворотками морских свинок против Тbр2, продуцированными в ответ на введение очищенного рекомбинантного белка Тbр2 Eagan (фиг. 27). Дорожка 1: маркеры молекулярных масс; дорожка 2: индуцированный штамм JB-1437-4-1, экспрессирующий рекомбинантный белок Тbр2 Eagan; дорожка 3: JВ12-EDDA; дорожка 4: SВ12+EDDA; дорожка 5: SВ29-EDDA; дорожка 6: SB29+EDDA; дорожка 7: SВ30-EDDA; дорожка 8: SВ30+EDDA; дорожка 9: SВ12-EDDA; дорожка
10: SВ33-EDDA; дорожка 11: SВ33+EDDA; дорожка 12: РАК-EDDA; дорожка 13: РАК+EDDA; дорожка 14: Eagan-EDDA; дорожка 15: Eagan+EDDA. Специфические ≈60-70 кДа-полосы реагировали с антисыворотками против Тbр2 на дорожках 3, 6, 7, 8, 13, 14 и 15, которые соответствуют штаммам SB12, SB29, SВ30, РАК и Eagan.
Пример 16
В этом примере проиллюстрирован синтез синтетических пептидов, соответствующих консервативным областям в Тbр2 и Тbр1.
Выведенные аминокислотные последовательности Tbp1 и Тbр2 сравнивали, как показано на фиг. 14 и 15 соответственно. В результате этого сравнения были идентифицированы области консервативных аминокислотных последовательностей в рецепторе трансферрина (см. выше) и, как показано в таблицах 2 и 3, были синтезированы пептиды, содержащие часть рецептора трансферрина. Такой синтез может быть осуществлен путем экспрессии в соответствующем хозяине рекомбинантных векторов, содержащих нуклеиновую кислоту, кодирующую указанные пептиды, или путем стандартного пептидного синтеза.
Короче говоря, пептиды были синтезированы с использованием пептидного синтезатора ABI 430A и оптимизированной t-Воc-химии в условиях, рекомендованных производителем, после чего пептиды отщепляли от полимерного субстрата с использованием фтористоводородной кислоты (HF). Эти пептиды очищали с помощью обращенно-фазовой жидкостной хроматографии высокого разрешения (ОФ-ЖХВР) на полупрепаративной колонке с Vydac С4 (1•30 см) с использованием градиента 15-55% ацетонитрила в 0,1% трифлуорилуксусной кислоты (TFA), проявляя в течение 40 минут со скоростью потока 2 мл/мин. Все синтетические пептиды, используемые в биохимических и иммунологических исследованиях, имели чистоту более 95%, что подтверждалось аналитической ЖХВР. Анализ аминокислотного состава осуществляли с использованием системы Waters Pico-Tag и этот анализ показал хорошее согласие с результатами, полученными для этого состава теоретически.
Пример 17
В этом анализе проиллюстрирована иммуногенность синтетических пептидов с использованием подопытных животных.
Морских свинок иммунизировали путем внутримышечного введения 100 мкг пептида, полученного, как описано в примере 16, и эмульгированного в полном адъюванте Фрейнда (день 0), после чего через 14 (день +14) и 28 (день +28) дней вводили бустер-дозу с использованием такого же количества пептида, эмульгированного в неполном адъюванте Фрейнда. Образцы сывороток брали на 42-й день (день +42) и с помощью твердофазного иммуноферментного анализа (ELISA) определяли титры антител. Для этого лунки для микротитрования (Nunc -Immunoplate, Nunc, Denmark) сенсибилизировали 500 нанограммами любого из конкретно исследуемых пептидов в 50 мкл буфера для сенсибилизации поверхностей (15 мМ Na2СО3, 35 мМ NaHCO3, рН 9,6) в течение 16 часов при комнатной температуре. Затем планшеты блокировали 0,1% (мас./об.) BSA в забуференном фосфатом физиологическом растворе (PBS) в течение 30 минут при комнатной температуре. Антисыворотки подвергали серийному разведению, после чего добавляли в лунки и инкубировали в течение 1 часа при комнатной температуре. После удаления антисывороток планшеты пять раз промывали PBS, содержащим 0,1% (мас. /об. ) Твин и 0,1% (мас./об.) BSA. F(ab')2-фрагменты от козьих иммуноглобулиноа IgG, направленных против морских свинок и конъюгированных с пероксидазой хрена (Jackson Immuno Research Labs Inc., PA), разводили буфером для промывания и добавляли в планшеты для микротитрования. После одночасового инкубирования при комнатной температуре планшеты пять раз промывали буфером для промывания. Эти планшеты проявляли с использованием субстрата тетраметилбензидина (ТМВ) в Н2О2 (ADL, Торонто), затем реакцию прекращали путем добавления Н2SО4 и измеряли оптическую плотность при 450 нм с использованием Titretek Multiskan II (Flow Labs., Virginia).
Два нерелевантных пептида, состоящих из 32 аминокислотных остатков, использовали в указанных ELISA-анализах в качестве негативного контроля. Эти анализы проводили в тройном дубликате, а реакционный титр каждой антисыворотки определяли как разведение, соответствующее двухкратному увеличению оптической плотностью по сравнению с оптической плотностью, полученной для негативного контроля. Антисыворотки, продуцированные у морских свинок, являются моноспецифическими для пептида, используемого для иммунизации. Титры сывороток, полученных после иммунизации, показаны в таблице 4.
Пептиды настоящего изобретения содержат единственные копии любого из пептидов, указанных в таблицах 2 и 3, либо много копий их аналогов. Кроме того, пептид может содержать множество различных пептидов, выбранных из пептидов, указанных в таблицах 2 и 3 или их аналогов, и включать в себя подходящие молекулы-носители. При этом предпочтительно, чтобы для продуцирования антител были использованы пептиды от консервативных областей, поскольку в этом случае анализ на иммуносвязывание или на связывание другого типа может быть затем использован для обнаружения нескольких видов Haemophilus. Поэтому в таблицах 2 и 3 приводятся другие консервативные области рецептора трансферрина для идентификации других пептидов, которые могли бы быть использованы для диагностики, иммунизации и медикаментозном лечении.
Пример 18
В этом примере описана способность антисыворотки, продуцированной против пептидов, соответствующих консервативным областям рецептора трансферрина, распознавать рецептор трансферрина Branhamella catarrhalis.
Морские свинки были иммунизированы пептидом, соответствующим консервативным областям рецептора трансферрина, и в результате этой иммунизации были получены антисыворотки, как описано в примере 17. Клеточный экстракт Branhamella catarrhalis подвергали иммуноблоттингу с пептидспецифической антисывороткой, которая специфически распознает рецептор трансферрина от этой бактерии. Антипептидная сыворотка, продуцированная у морских свинок, иммунизированных N-концевым пептидом Тbр2 и пептидом ТВР2-25, специфически распознавала белок Тbр2 от Branhamella catarrhalis и рекомбинантный белок, экспрессированный плазмидным клоном рВН1Т2 в E.coli. Клон рВН1Т2 экспрессировал укороченный вариант Тbр2, начинающийся с аминокислоты 80 (т.е. NKKFYSG -SEQ ID 105). Поэтому белок Тbр2 от рВН1Т2 может распознаваться только антителами, продуцированными против второго эпитопа LEGGFYGP (ТВР2-25). Этот анализ показал, что пептиды, соответствующие консервантивным последовательностям рецепторов трансферрина, могут быть с успехом использованы для обнаружения большинства, если не всех бактерий, продуцирующих рецептор трансферрина, и кроме того, они могут быть использованы в качестве компонентов в иммуногенных композициях, включая вакцины, для продуцирования иммунного ответа против рецептора трансферрина и защиты от заболеваний, вызываемых этими бактериями.
Сыворотки от кроликов тестировали с помощью анализа ELISA на специфичность к пептиду, имеющему последовательность LEGGFYGP (SEQ ID 74), или к Тbр2 штамма DL63 H. influenzae. ELISA-планшеты сенсибилизировали пептидом или белком, а затем блокировали 5%-ным сепарированным молоком. Серийные двухкратные разведения сывороток в забуференном фосфатом физиологическом растворе, 0,05% Твине-50 и 1% сухом молоке инкубировали на планшетах в течение двух часов при 37oС, после чего планшеты пять раз промывали в забуференном фосфатом физиологическом растворе, содержащем 0,05% Твин-20. Промытые планшеты зондировали ослиными антикроличьими иммуноглобулинами IgG, конъюгированными с пероксидазой хрена (HRPO) в течение 30 минут при комнатной температуре, а затем пять раз промывали в забуференном фосфатом физиологическом растворе, содержащем 0,05% Твин-20. В каждую лунку в течение 30 минут при комнатной температуре в темноте добавляли НRРО-субстрат и развитие окраски останавливали добавлением 50 мкл 1 M серной кислоты. Степень развития окраски оценивали путем определения оптической плотности при 450 нм.
Пример 19
В этом примере проиллюстрировано продуцирование штаммов H. influenzae, которые не экспрессируют рецептор трансферрина.
2,55 - EcoRI-фрагмент вставки из pBH1T1 субклонировали в EcoRI-сайт рUC4К, что приводило к удалению кластера устойчивости к канамицину (каn) Тn 903 из этого вектора (pUH1T1; фиг.28). Эта стадия субклонирования способствовала облегчению последующей инсерции либо HincII - либо Pst1-фрагмента рUС4К, содержащего каn-кластер, в Hind III или Pst1-сайт pUH1T1 (оба из которых являются уникальными сайтами в этой конструкции) в целях продуцирования pUH1T1KFH и pUH1T1KFP (фиг.28). После гидролиза ферментом EcoRI для удаления ненужных генных последовательностей эти конструкции вводили в геном H. influenzae дикого типа путем трансформации с использованием среды М-IV, как было описано ранее (Barcak и др., 1991), и трансформанты отбирали на агаре BHINH, содержащем 20 мкг/мл канамицина.
Пример 20
В этом примере проиллюстрировано конструирование полиовирусов, экспрессирующих эпитоп рецептора трансферрина.
кДНК-клон, имеющий основания 1175-2956 генома штамма Mahoney полиовируса типа I (РV1-М), разрезали рестриктирующими ферментами PVL I и Hind III. Эти ферменты вырезают фрагмент, содержащий основания 2754-2786, которые кодируют аминокислоты 1094-1102 PV1-М, как показано на фиг.29. В настоящей заявке для обозначения аминокислот полиовируса был использован четырехзначный код, например цифра 1095, обозначает аминокислоту 95 капсидного белка VP1. Новые гибридные кДНК-клоны были сконструированы путем замены вырезанного фрагмента синтетическими олигонуклеотидами, кодирующими аминокислоты белка Тbр2 H. influenzae. Эти новые гибридные кДНК-клоны разрезали рестриктирующими ферментами Nhe1 и SnaB1, которые вырезают гибридный фрагмент, содержащий ДНК-последовательности рецептора трансферрина, из оснований 2471-2956 полиовируса. кДНК-клон, например рТ7ХLD или рТ7СМСВ, целого генома РV1-М разрезали ферментами Nhe I и SnaB1 для вырезания фрагмента из оснований 2471-2956 полиовируса. Затем этот фрагмент заменяли гибридным фрагментом, содержащим ДНК-последовательности рецептора трансферрина, для продуцирования гибридного кДНК-клона генома PV1-M с основаниями 2754-2786, замененными основаниями, кодирующими гибридную ВС-петлю, включающую в себя аминокислоты рецептора трансферрина, как показано на фиг.29.
Плазмида pT7XLD и клоны, происходящие от pT7XLD, такие как рТ7СМСВ, содержат промоторную последовательность для РНК-полимеразы Т7 у 5'-конца кДНК PV1-М. РНК-транскрипты кДНК РV1-М, включая любые основания, кодирующие аминокислоты рецептора трансферрина, были получены с использованием РНК-полимеразы Т7, как описано Van der Werf и др. В результате трансфекции клеток Vero этими РНК-транскриптами продуцировали четыре жизнеспособных гибридных вируса, обозначенных РV1ТВР2А, РV1ТВР2В, PV1TBP2C и РV1TBP2D. В результате трансфекции рТ7СМСВ транскриптами получали полиовирус дикого типа, обозначенный PV1XLD (фиг.29).
Антигенные характеристики РV1ТВР2А, РV1ТВР2В, РV1ТВР2С и PV1TBP2D проиллюстрированы в таблице 5. Все указанные гибриды были нейтрализованы антисыворотками морских свинок, выработанных против пептида, содержащего последовательность LFCG-FYGP (SEQ ID 74), что свидетельствовало о том, что указанные вирусы экспрессировали эту последовательность в антигенно распознаваемой форме. Для продуцирования указанных антисывороток самок морских свинок иммунизировали 1 M 500 мкл-объемом, содержащим 200 мкг пептида, разведенного в фосфате алюминия (3 мг/мл). Животных иммунизировали в дни: 1, 14, 28 и 42, и брали кровь в дни 0, 28, 42 и 56. Сыворотки получали из крови, взятой на 56-й день, PV1TBP2A и PV1TBP2B были также нейтрализованы кроличьими антисыворотками против Тbр2 штамма DL63 H. influenzae, что свидетельствовало о том, что, по крайней мере, эти два вируса экспрессировали последовательность а форме, распознаваемой для антител, выработанных против указанного белка. Все вирусы были нейтрализованы антисыворотками против PV1, что указывает на то, что изменения в антигенном сайте нейтрализации полиовируса 1 не оказывают существенного влияния на другие антигенные сайты этих вирусов.
Пример 21
В этом примере иллюстрируется использование полиовирусных гибридов для индуцирования анти-Тbр2 антисывороток с высоким титром.
Кроликов инокулировали СsCl-очищенным PV1TBP2A (кролики 40, 41, 42). При этом следует отметить, что хотя используемые вирусы были "живыми", однако полиовирусы у кроликов не реплицировались, и любой наблюдаемый ответ был, по существу, ответом на инактивированный антиген. На день 1 кроликов инокулировали 1 микрограммами вируса в полном адъюванте Фрейнда путем подкожной инъекции а спинку, на 14-й день, кроликам вводили повторную инъекцию (подкожно, в спинку) 1 мкг вируса в неполном адъюванте Фрейнда. Пробы крови брали на день 0 (неимунная сыворотка) и на день 27. Доза вируса для инокуляции составляла 2,5•108 б.о.е и определялась исходя из величин А260, соответствующих приблизительно 3,0•1011 вирионам. Эта величина эквивалентна 0,5 пМ вируса или 30 пМ эпитопа LEGGFYG (SEQ ID 74), поскольку каждый вирион экспрессирует 60 копий эпитопа (табл.6).
Краткое изложение описания
Итак, настоящее изобретение относится к очищенным и выделенным ДНК-молекулам, содержащим гены рецептора трансферрина; к последовательностям этих генов рецептора трансферрина и к выведенным аминокислотным последовательностям рецептора трансферрина. Настоящее изобретение также относится к пептидам, соответствующим участкам рецептора трансферрина. Указанные гены, ДНК-последовательности, рекомбинантные белки и пептиды могут быть использованы для диагностики, иммунизации, и изготовления диагностических и иммунологических реагентов. Для предупреждения заболеваний, вызываемых бактериальными патогенами, продуцирующими рецептор трансферрина, могут быть получены вакцины, изготовленные на основе экспрессированных рекомбинантных белков Тbр1 и/или Тbр2, их фрагментов или пептидов, происходящих от указанных последовательностей. При этом возможны модификации, не выходящие за рамки объема настоящего изобретения.


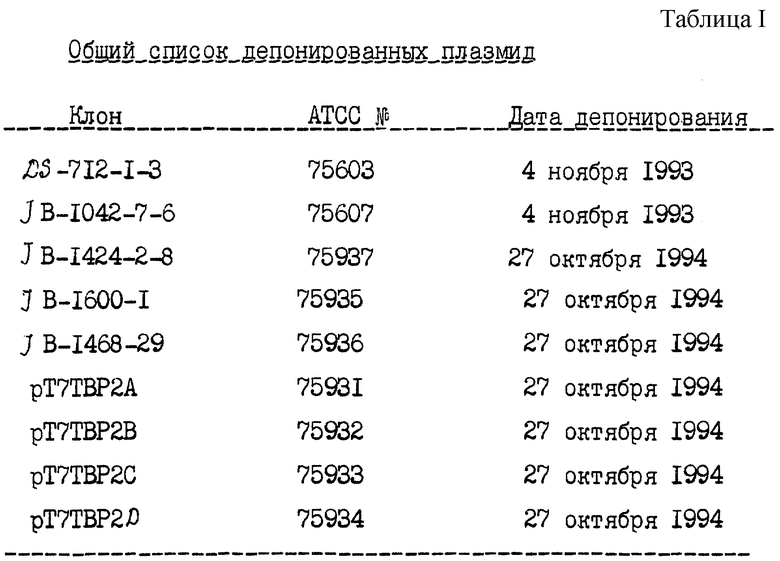
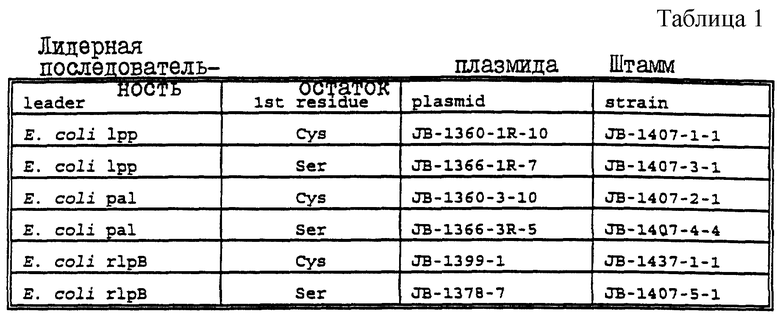





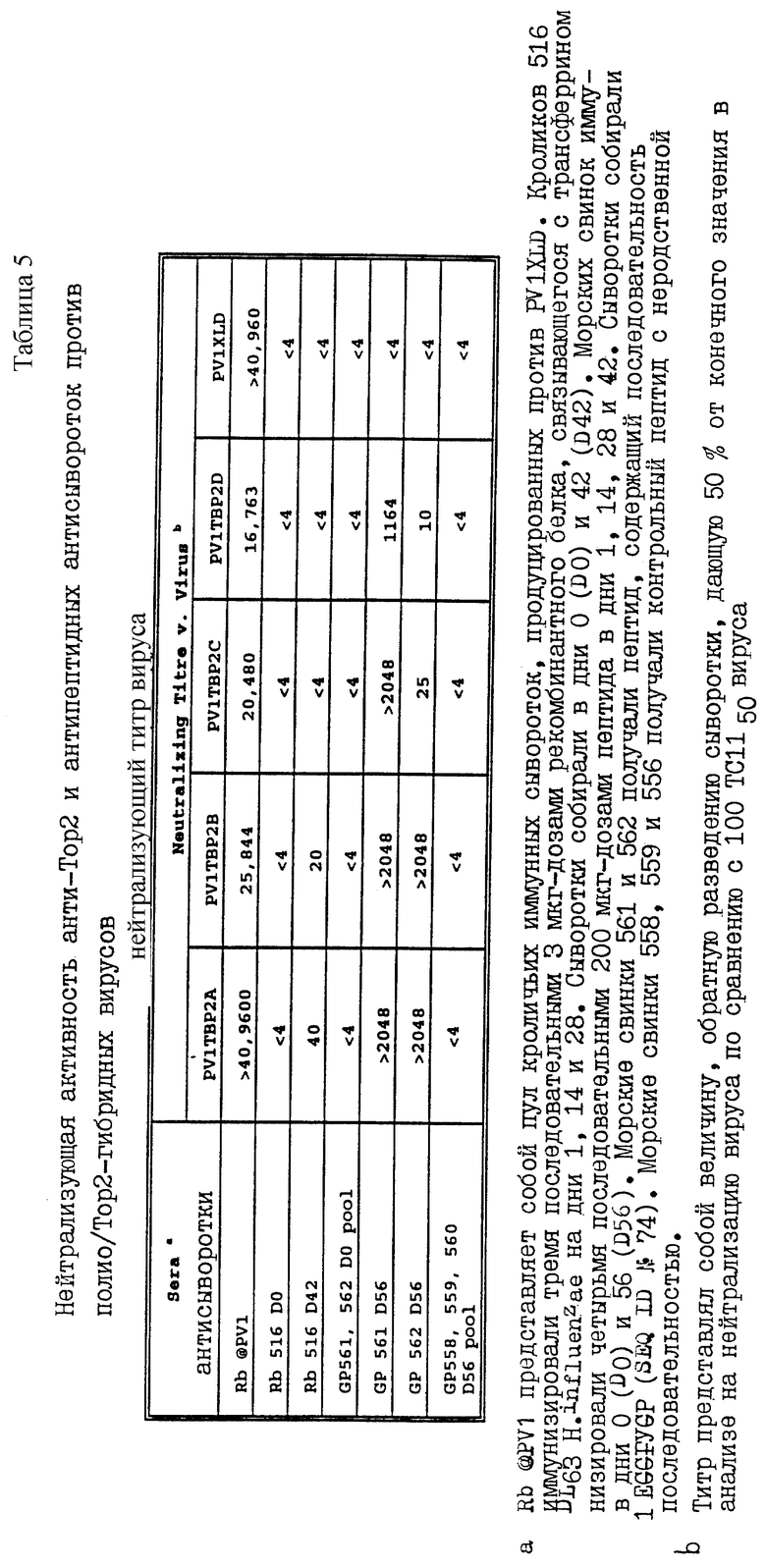
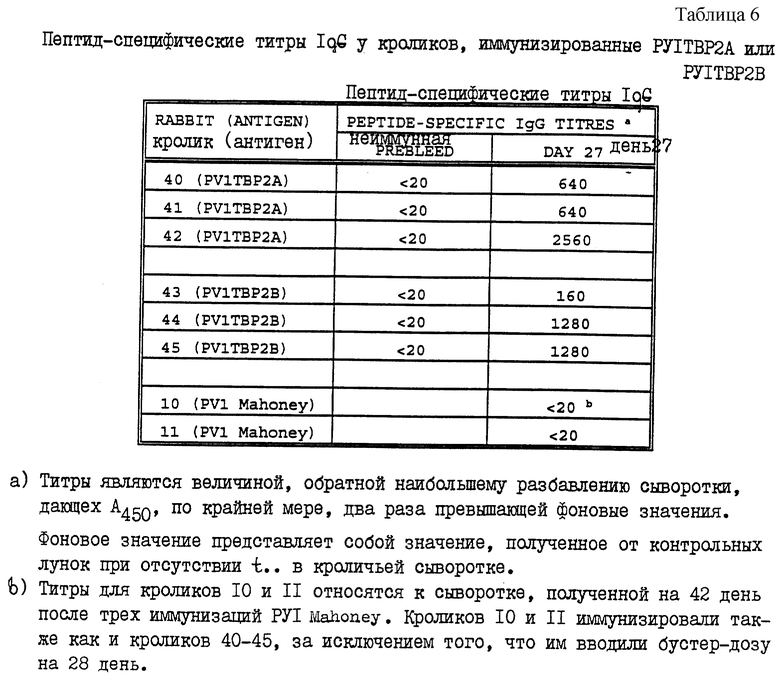
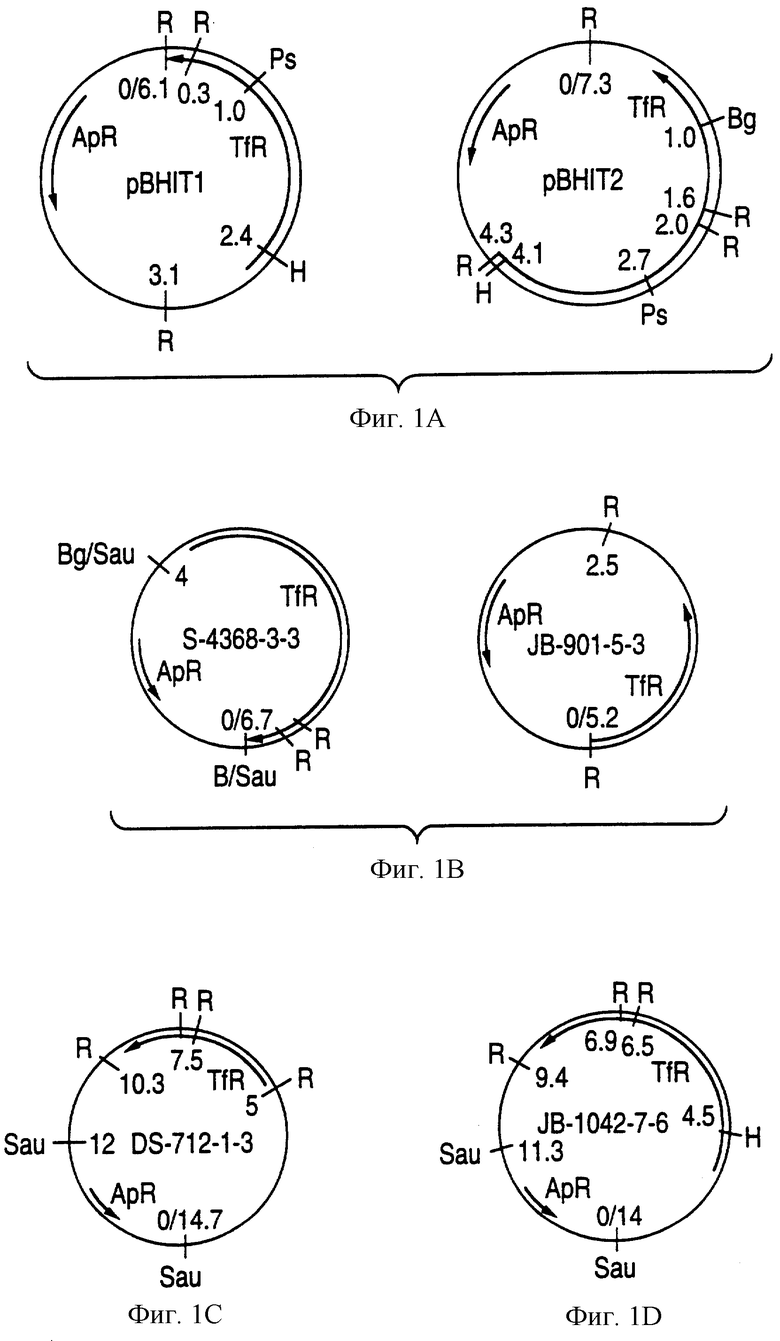
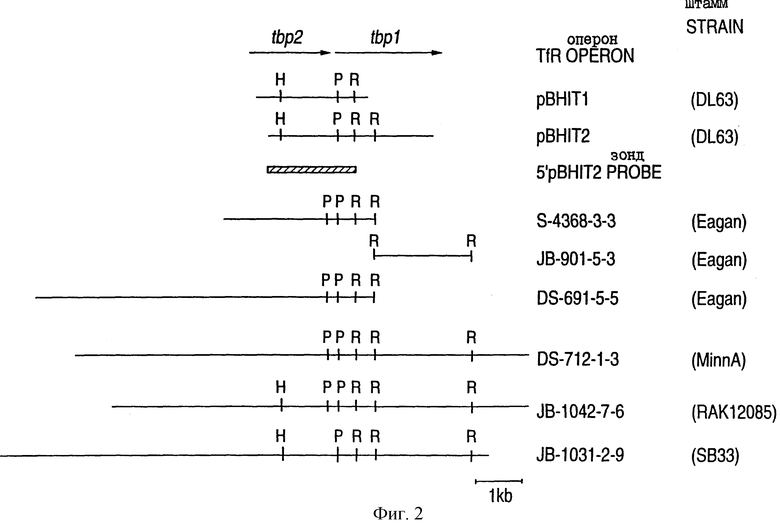


































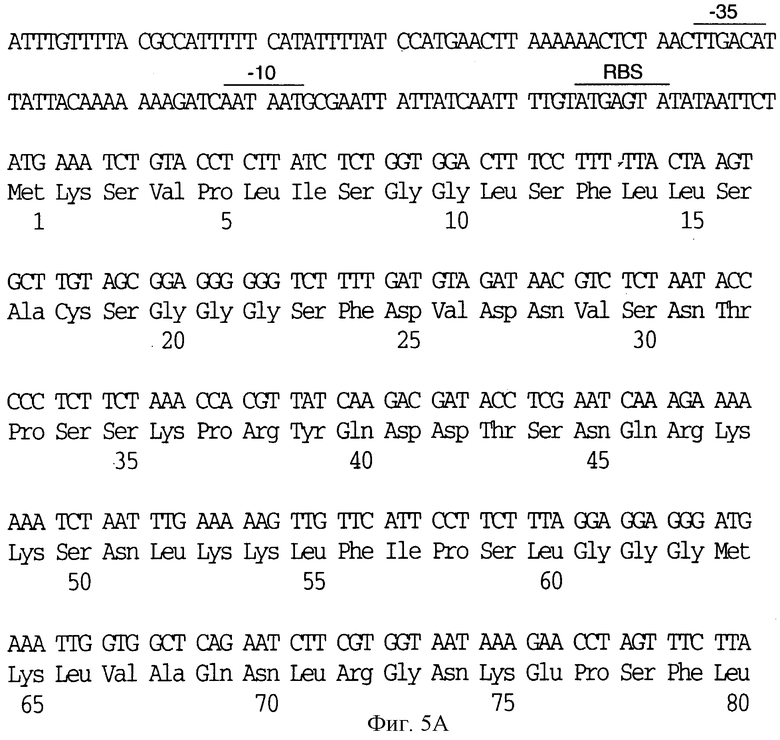





































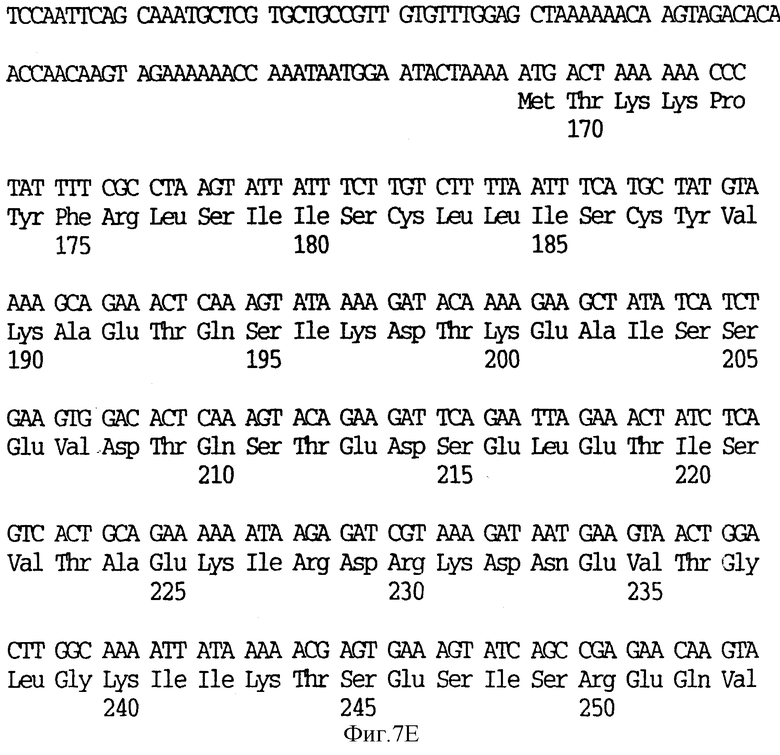







































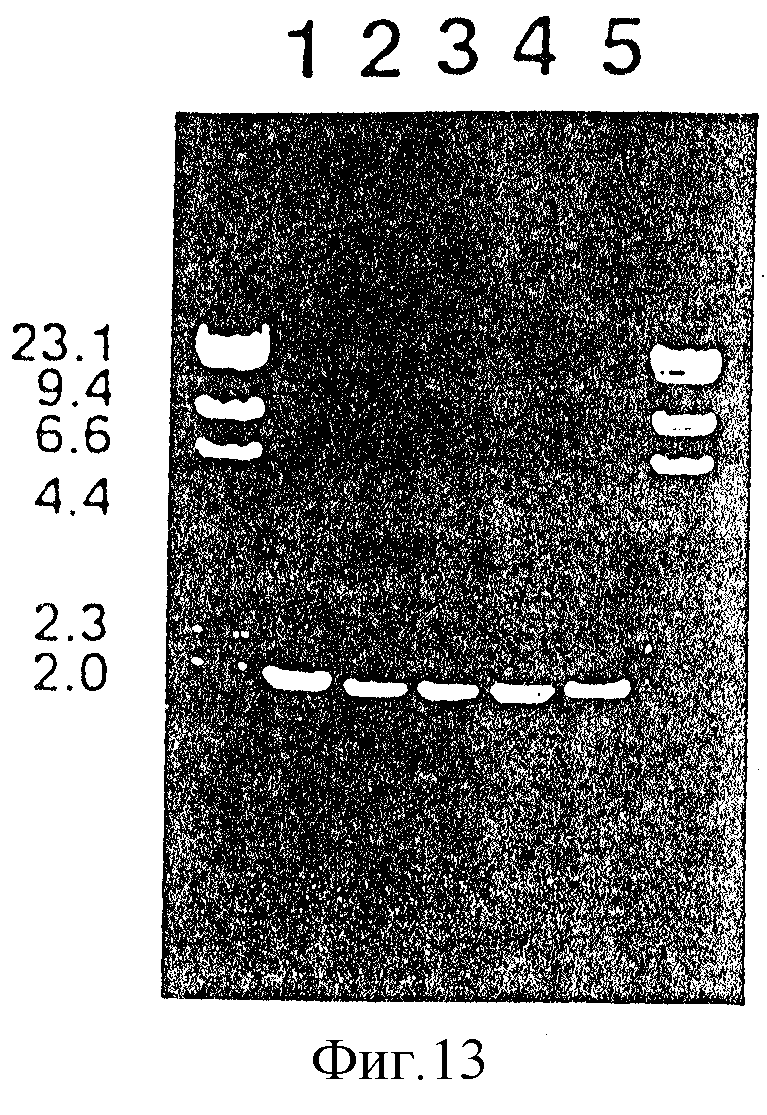
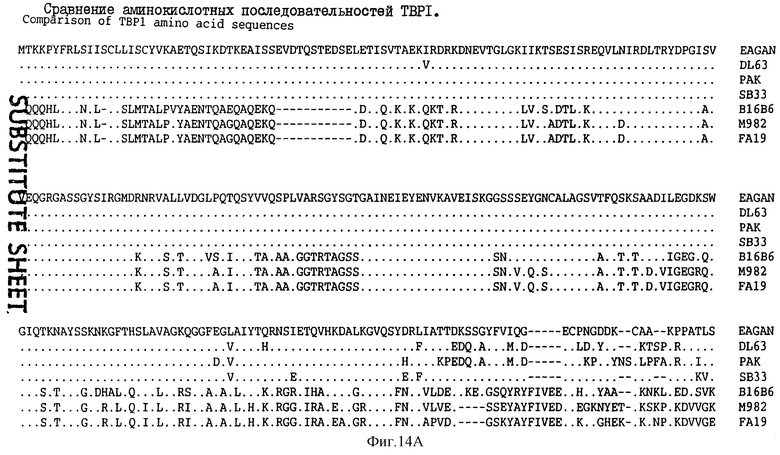

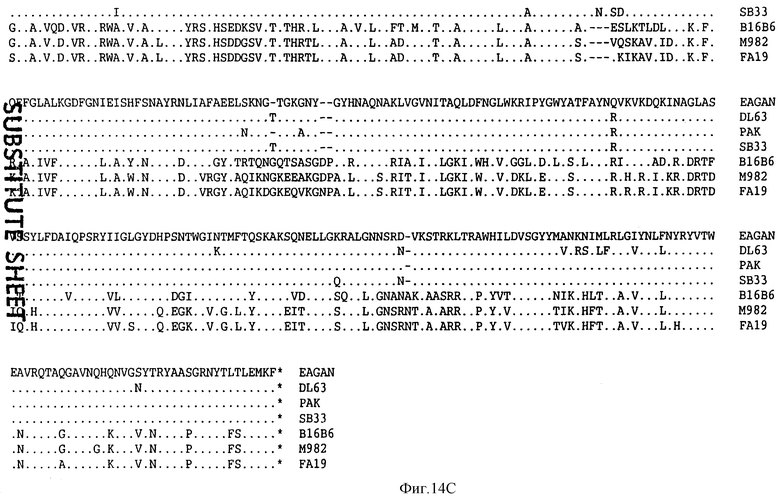



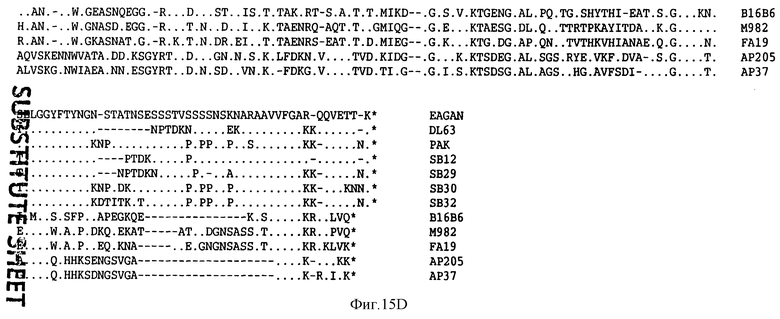
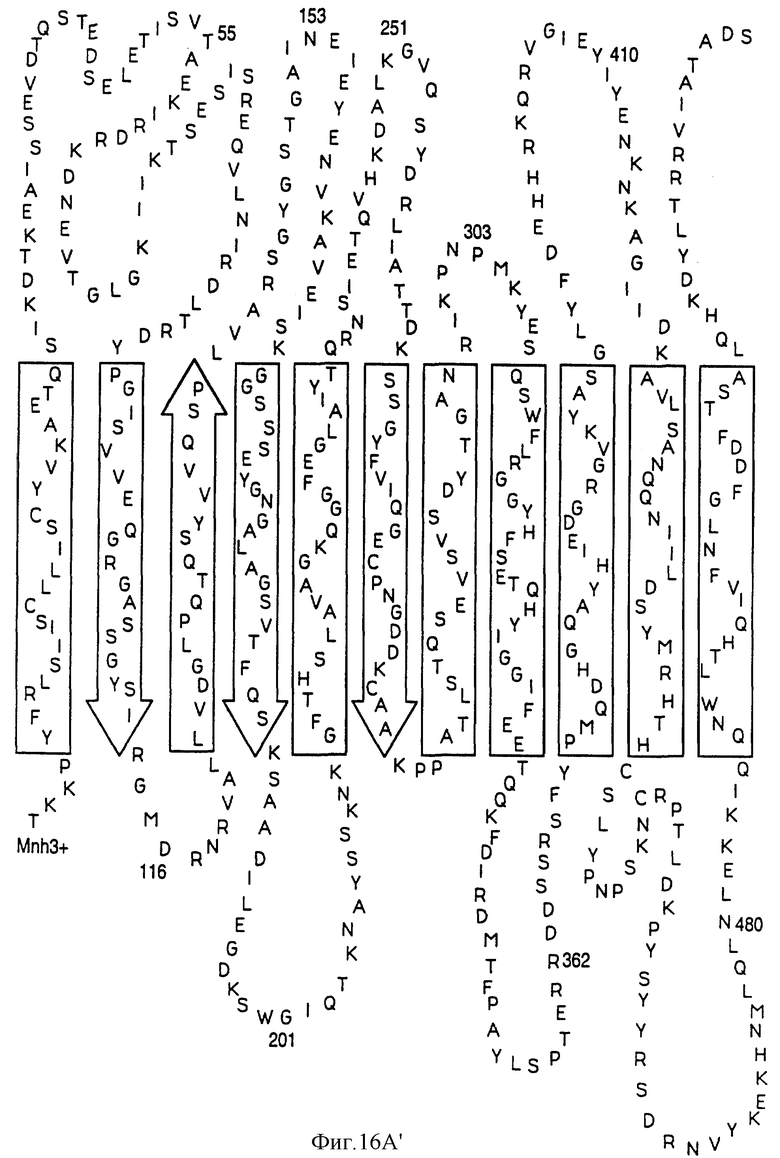
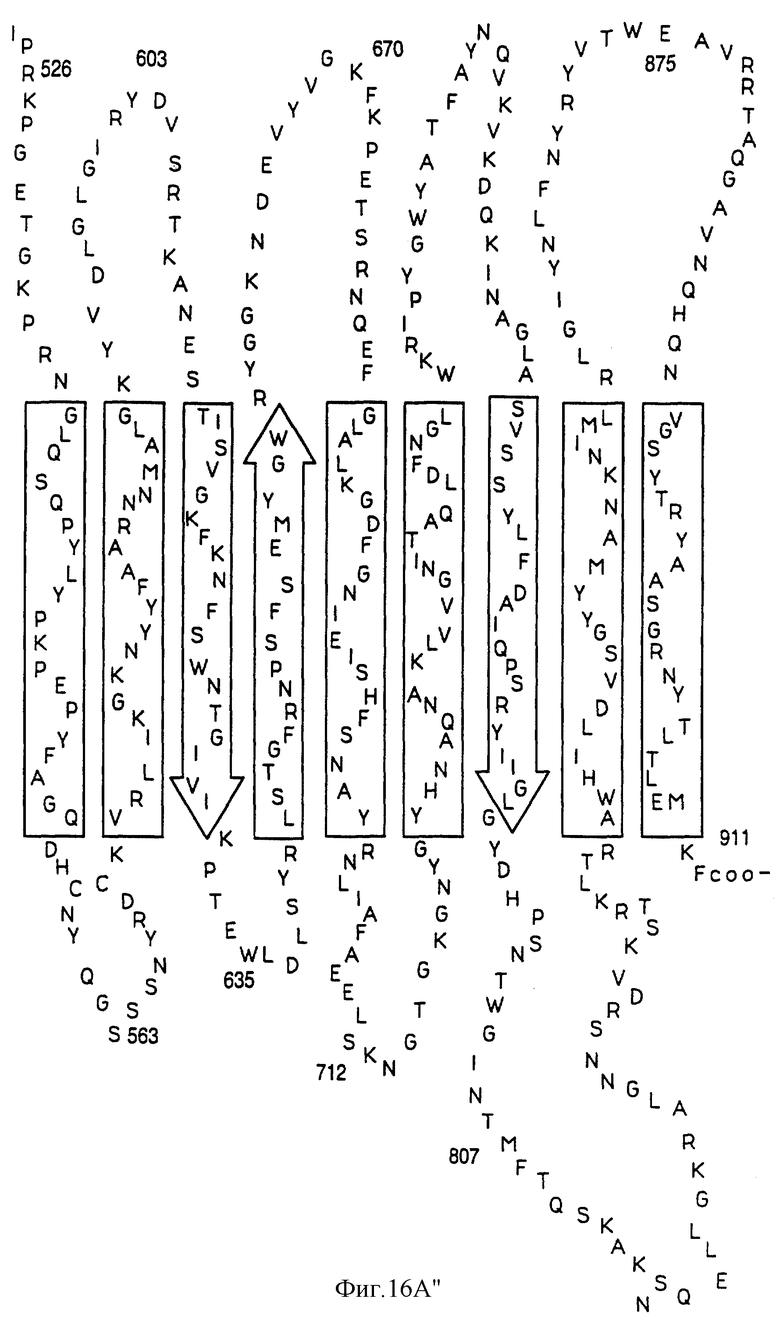
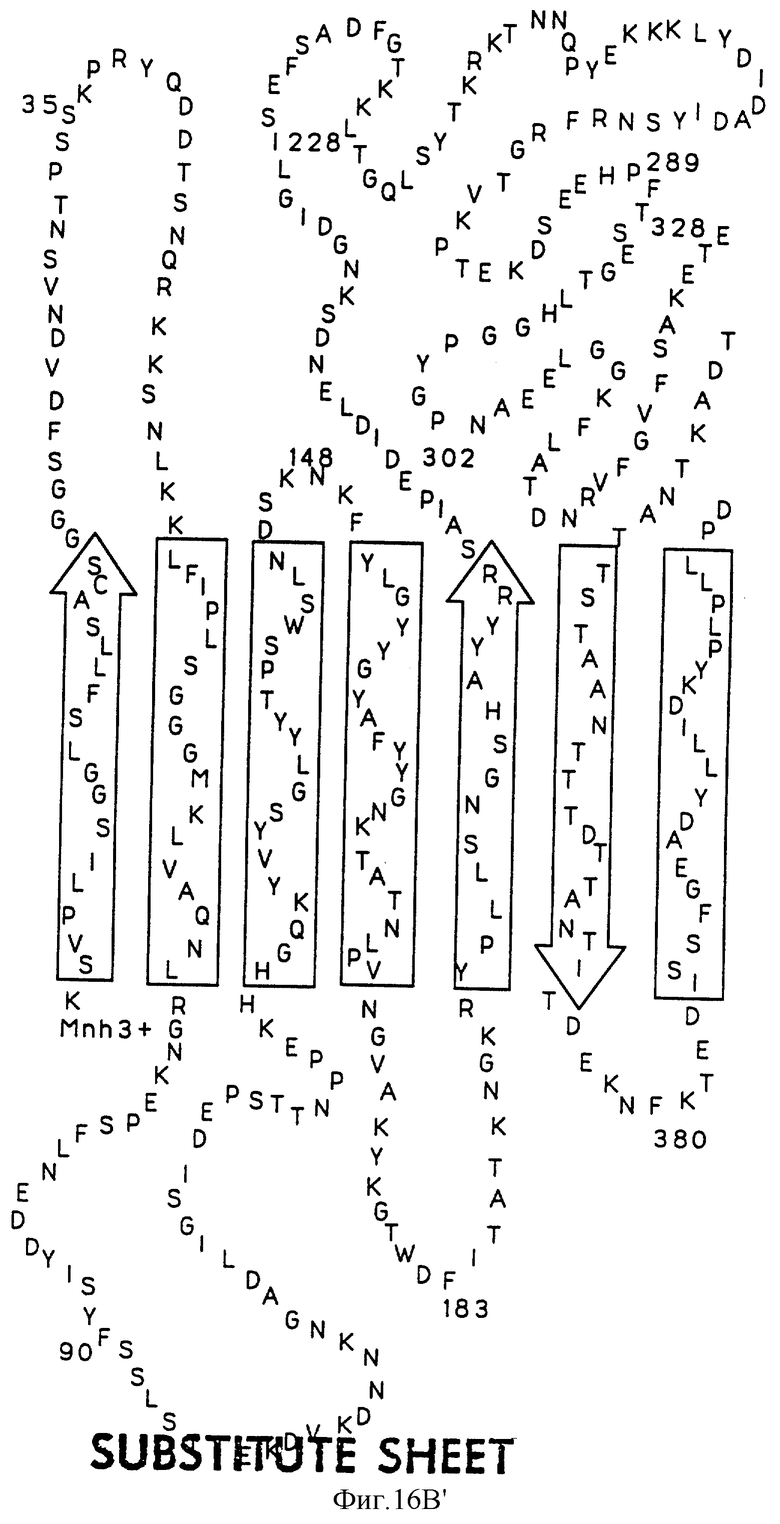
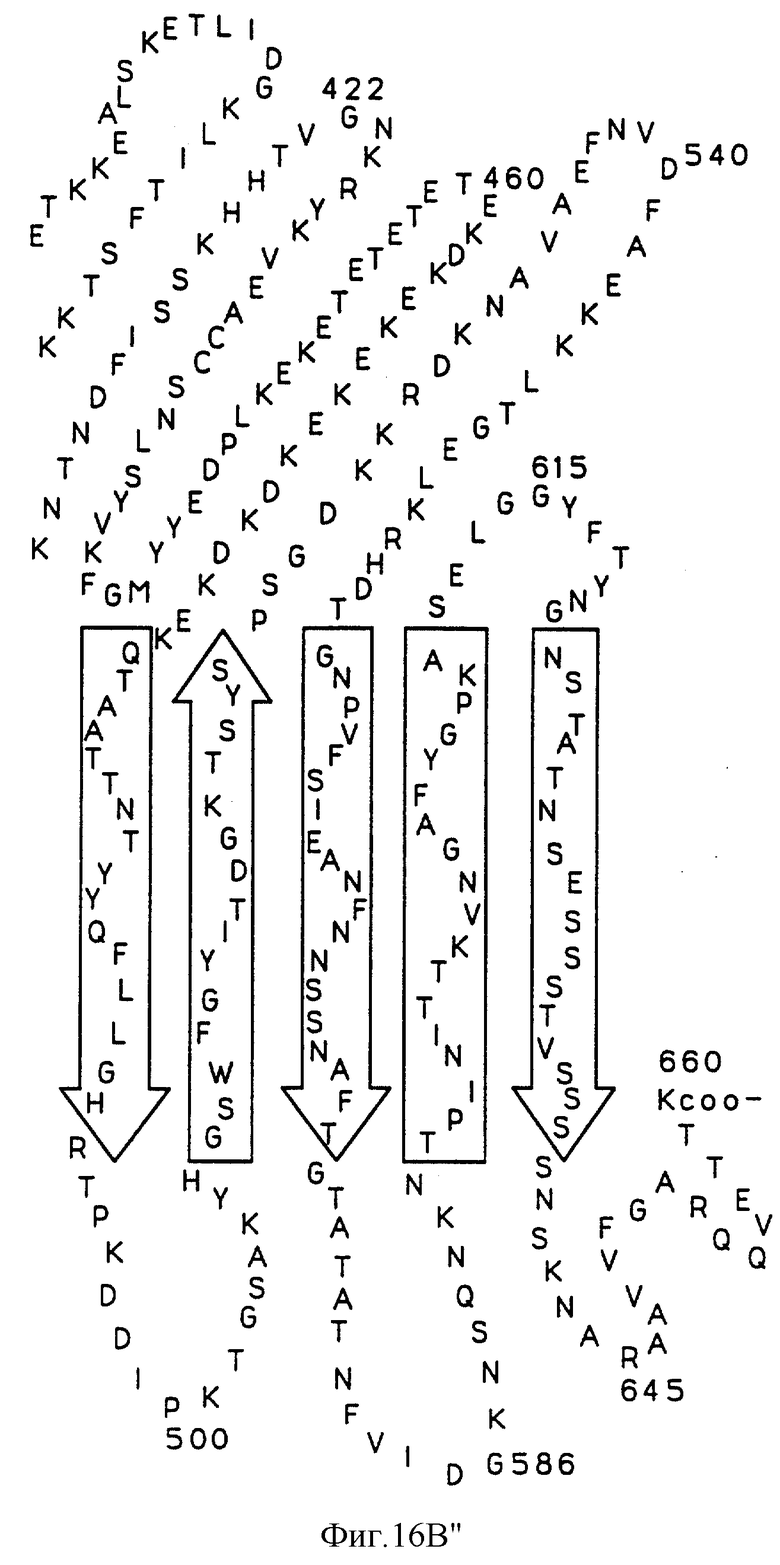
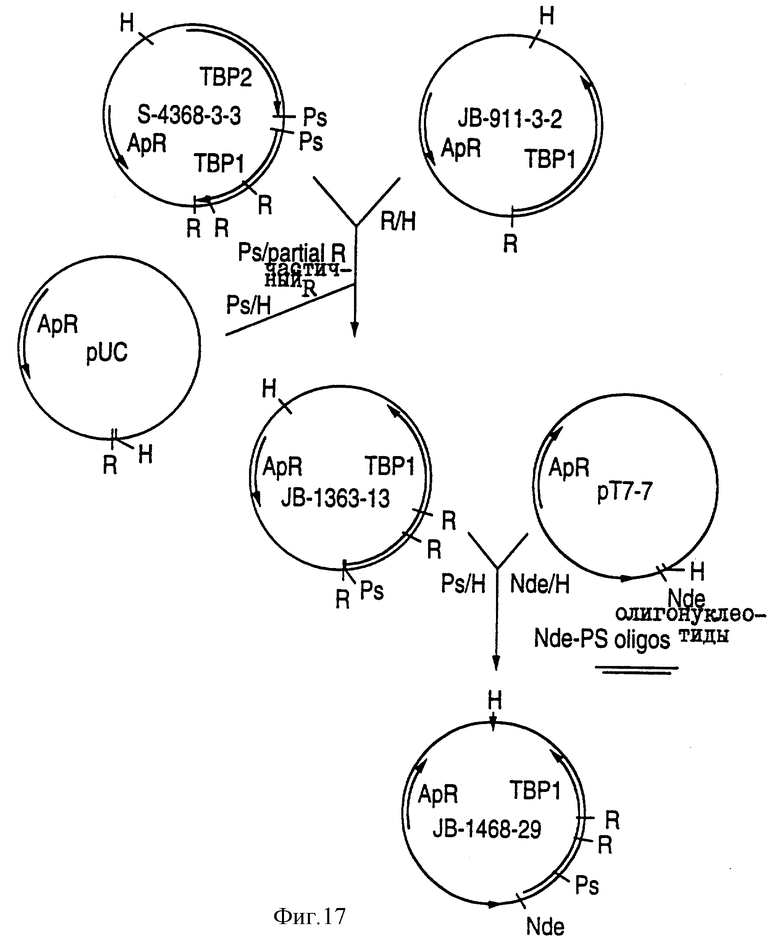
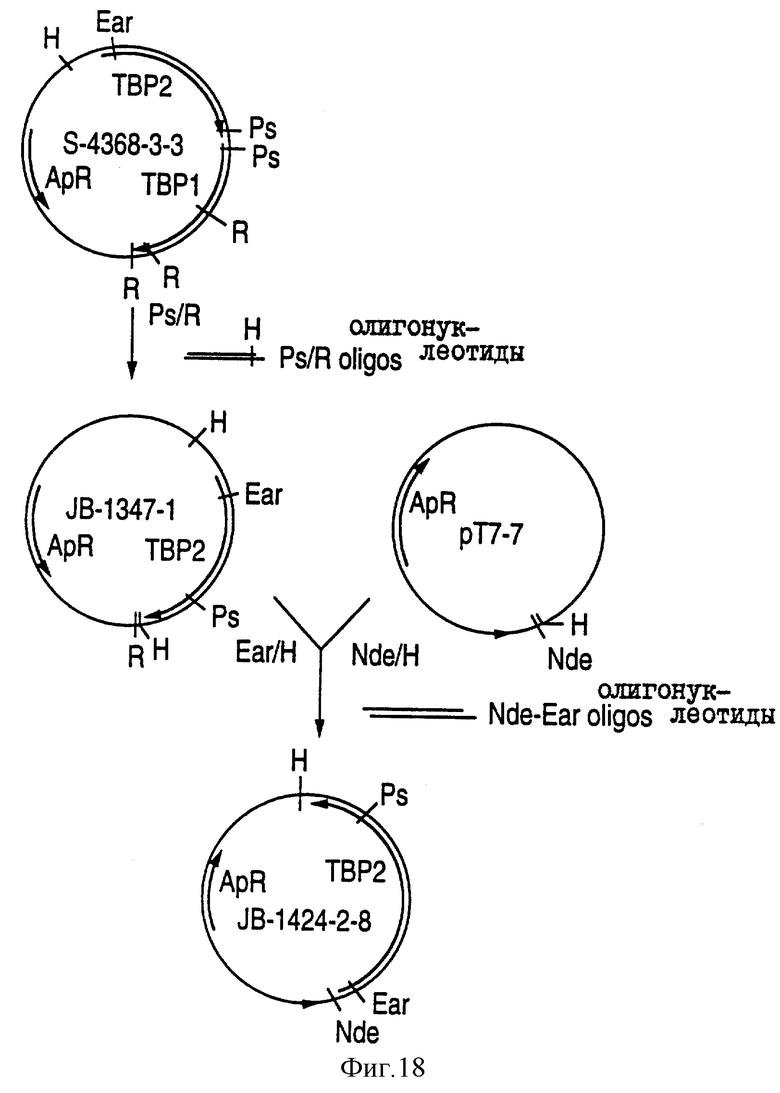



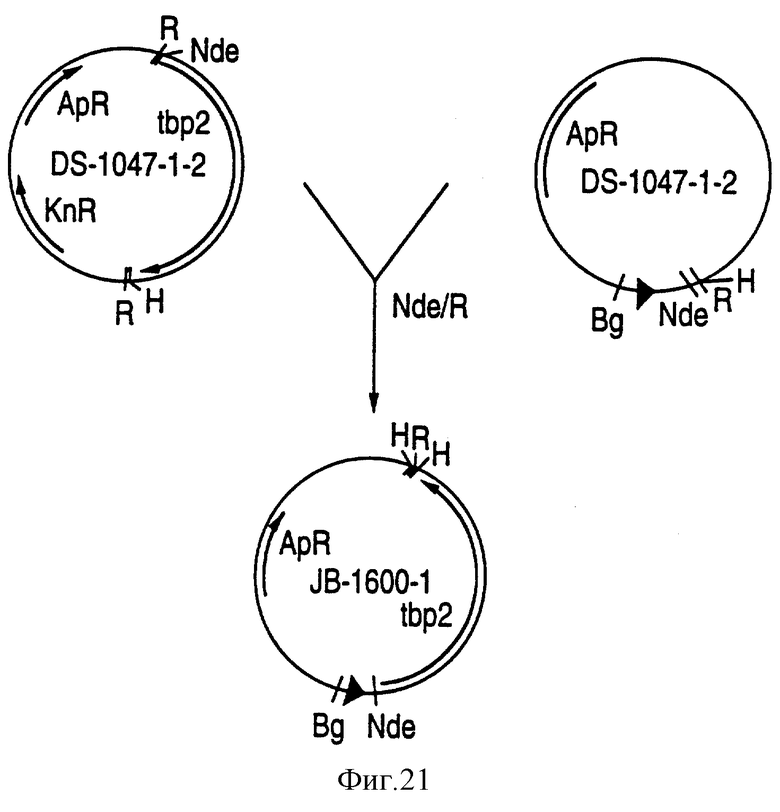

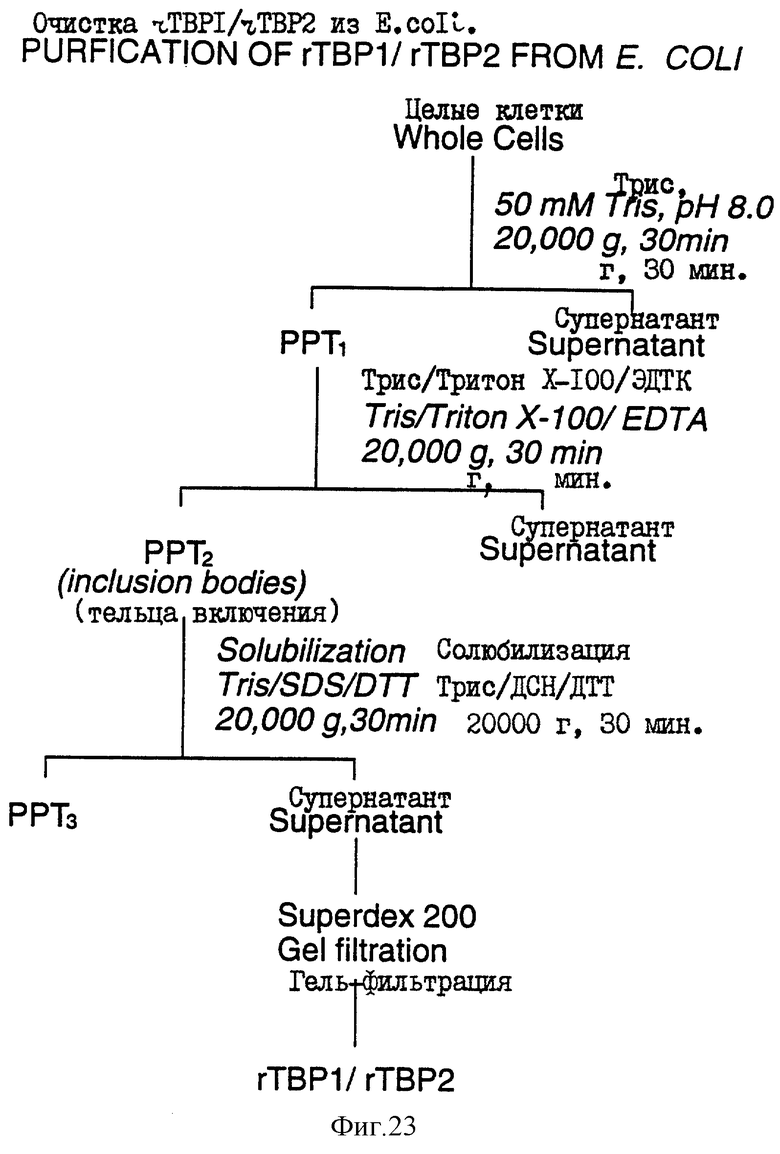
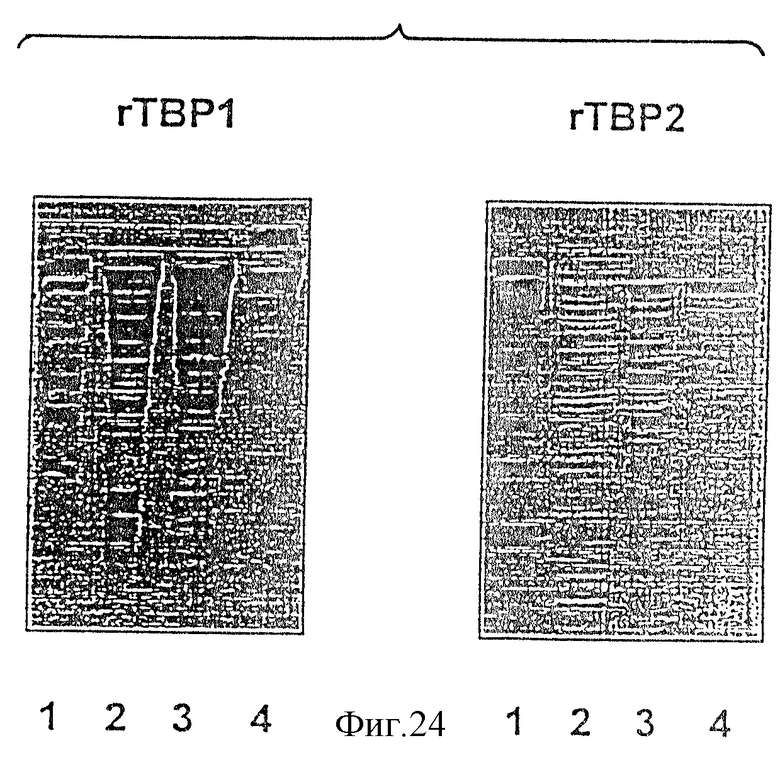
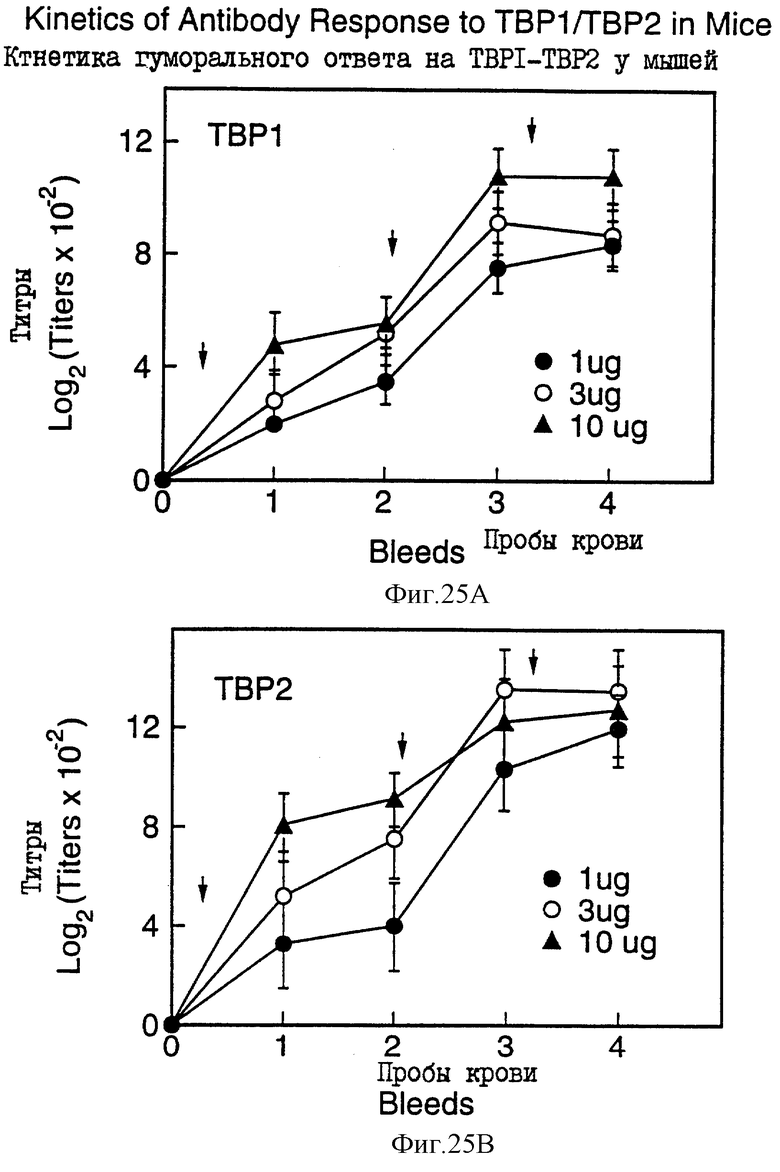

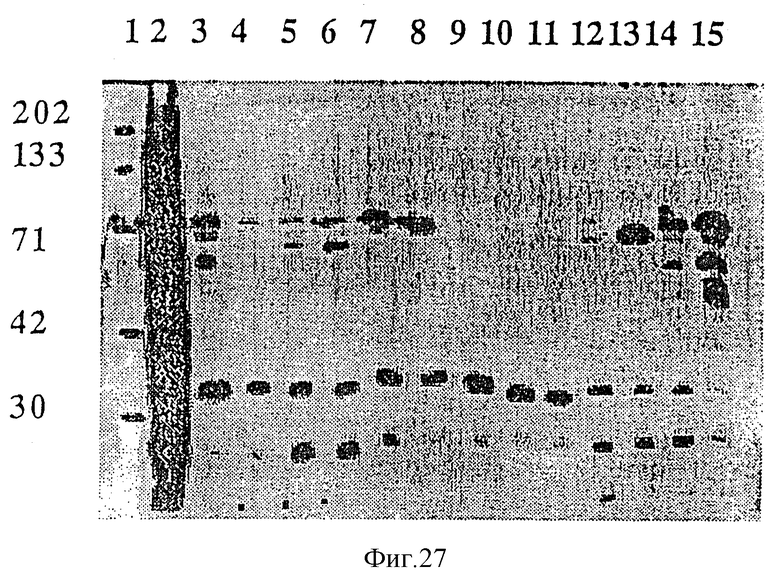
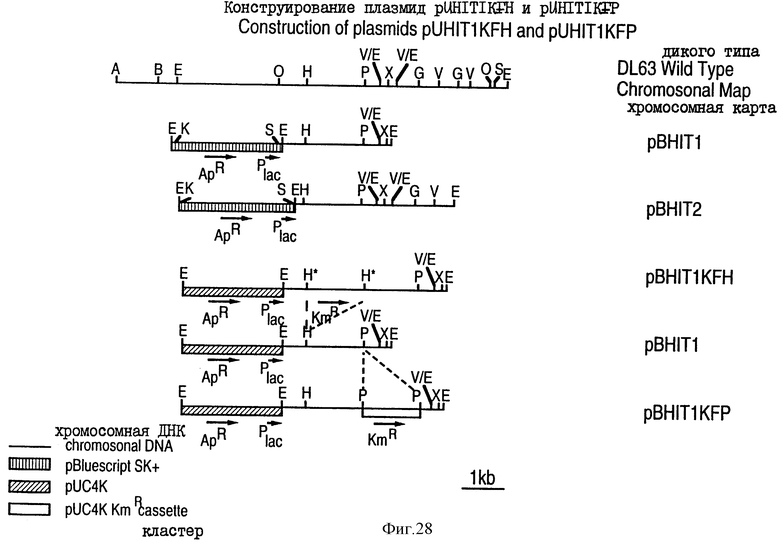
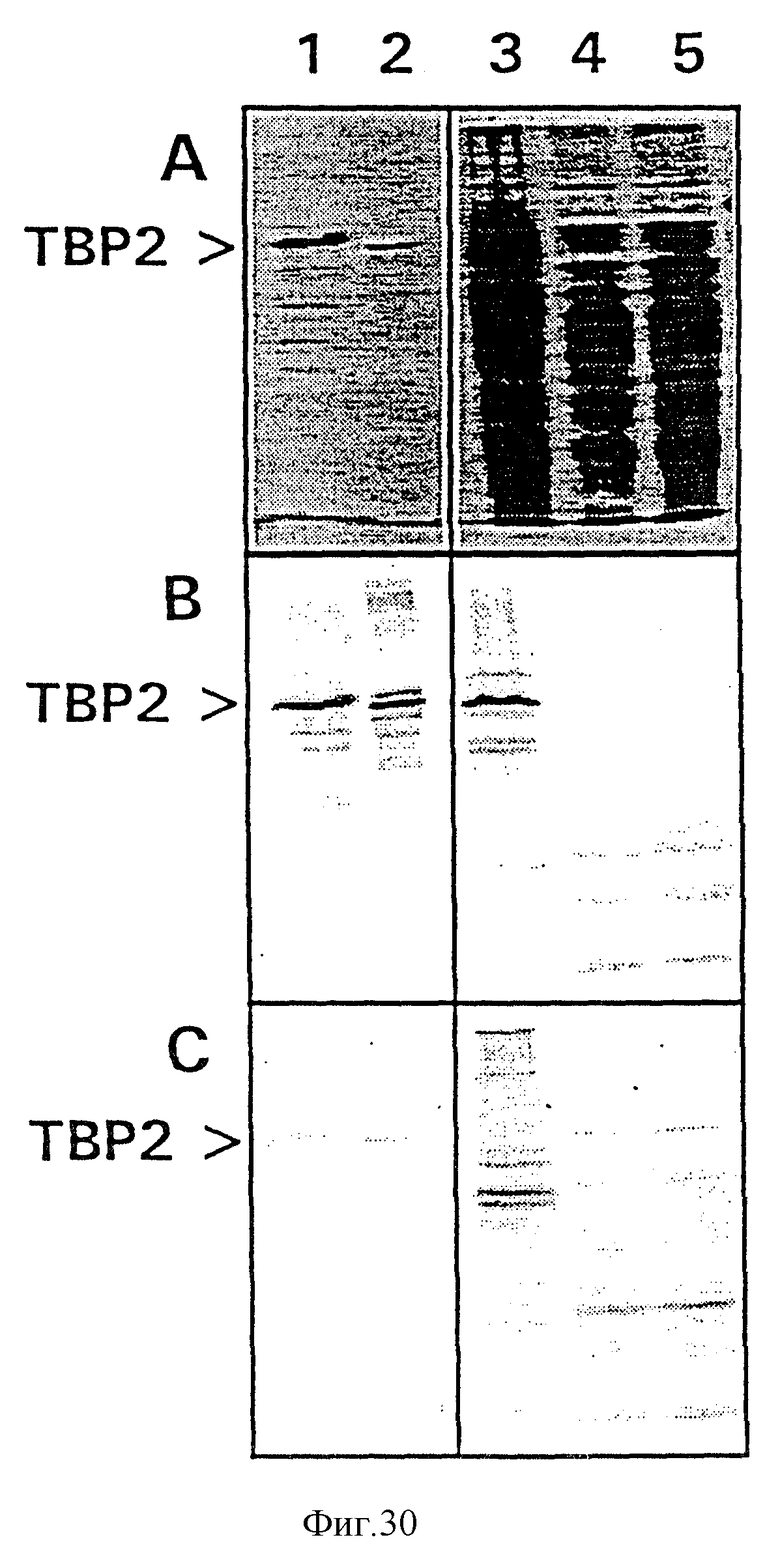
Изобретение относится к генной инженерии. Фрагмент ДНК кодирует фрагмент белка рецептора трансферрина штамма Haemophilus. Консервативная аминокислотная последовательность белка приведена в описании. Фрагменты ДНК кодируют белки рецептора трансферрина штаммов DL 63 или Eagan Haemophilus influenzae типа b или нетипируемых штаммов РАК 12085, SB33, SB12, SB29, SB30 или SB32 Haemophilus influenzae. Аминокислотные последовательности кодируемых белков приведены в описании. Плазмидный вектор адаптирован для трансформации E.coli и включает один из вышеперечисленных фрагментов ДНК. Рекомбинантный белок продуцируется Е. coli, трансформированной одним из указанных фрагментов ДНК. Выделенный и очищенный белок Тbp1 и Тbp2 с аминокислотной последовательностью приведен в описании. Иммуногенная композиция изготовлена в виде вакцины для in vivo введения и включает выделенный и очищенный белок Тp1 или Тp2 штамма Haemophilus influenzae и фармацевтически приемлемый носитель. Получают рекомбинантный хозяин, экспрессирующий в тельцах включения один из белков Тp1 или Тp2, культивируют его, разрушают клеточную массу, получают клеточный лизат и фракционируют. Проводят селективную экстракцию и вновь фракционируют с выделением фракции, содержащей белок Тbp1 или 2. Изобретение позволяет проводить диагностику инфекций, вызываемых Haemophilus. 12 с. и 13 з.п.ф-лы, 30 ил., 7 табл.
| HOLLAND J | |||
| et al | |||
| Evidence for in vivo expression of transferring-binding proteins in haemophilus influenzae type b | |||
| INFECTION AND IMMUNITY, vol.60, no.7, July 1992, p.2986-2991 | |||
| GRAY-OWEN S.D | |||
| et al | |||
| The interaction of primate transferrins with receptors on bacteria pathogenic to humans, MICROBIAL PATHOGENESIS, vol.14, may 1993, p.389-398 | |||
| SCRYVERS A.B | |||
| Characterization of the human transferring, MOLECULAR MICROBIOLOGY, vol.2, no.4, 1988 | |||
| CTEVENSON P | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| INFECTION AND IMMUNITY, vol.60, no.6, juny 1992, p.2391-2396 | |||
| МАНИАТИС Г.А | |||
| и др | |||
| Методы генетической инженерии | |||
| - М.: Мир, 1984, с.205-224. | |||

