Данное изобретение относится к области молекулярной генетики и, в частности, касается клонирования белка D15 наружной мембраны HAEMOPHILUS.
Haemophilus influenzae ТИПА b(Hib) является главной причиной бактериального менингита у детей в возрасте до пяти лет. Защитные антитела против заболевания индуцируются капсульным полисахаридом микроорганизма, и была разработана вакцина, в которой используется очищенный полирибозилрибитолфосфат (ПРФ) в качестве антигена. Эта вакцина обеспечивает 90% защиту у взрослых и у детей в возрасте старше 24 месяцев, но неэффективна у детей в возрасте до 24 месяцев, Zangwill at al. 1993 (Ссылки даны в списке ссылок в конце этого описания). Подобно другим полисахаридным антигенам, PRP не индуцирует пролиферации Т-хелперных клеток, и повторная иммунизация не способна вызвать ни вторичный иммунный ответ, ни увеличения клеток памяти. Соединение PRP полисахарида с белками-носителями придает вакцине свойства Т-клеточной зависимости и значительно усиливает имиунологический ответ на PRP антиген. В настоящее время существует четыре доступные вакцины на основе конъюгатов PRP-носитель. Существуют вакцины на основе капсульного полисахарида. Н. influenzae типа b, соединенного с дифтерийным токсоидом, столбнячным токсоидом или протеином наружной мембраны Neisseria meningitidis (обзор по ним представлен у Zangwill at al. 1993).
Однако, используемые в настоящее время конъюгатные вакцины против Haemophilus защищают только от менингита, вызываемого Haemophilus influenzsae типа b. Они не защищают от других инвазивных типируемых штаммов (типов a и c) и, что более важно, от нетипируемых штаммов (HTHi), которые являются обычными возбудителями при постнатальном и неонатальном сепсисе, пневмонии и воспалении среднего уха. Только в Соединенных Штатах стоимость лечения воспаления среднего уха составляет от 1 до 2 миллиардов долларов в год на антибиотики и хирургические вмешательства, такие как тонзилэктомии, аденоидэктомии и введение тимпаностомических трубок. Для достижения общей защиты от заболеваний, связанных с H.influenzsae, в возрастной группе от 2 до 6 месяцев и в некоторых группах риска желательно обеспечение сохранных, перекрестно-реактивных некапсульных иммуногенов H.influenzsae, методы индуцирования иммунитета против заболевания постоянно совершенствуются, и в настоящее время существует стремление использовать в качестве антигенов субъединицы и более четко определенные материалы. Это предпринимается с целью сведения к минимуму или устранения возможных побочных эффектов, вызываемых некоторыми природными иммуногенами, при сохранении их иммуногенности для обеспечения защиты от болезни. Поэтому было бы очень заманчиво разработать универсальную вакцину против Haemophilus, используя перекрестно-реактивные белки наружной мембраны, фрагмент, аналоги и/или соответствующие им пептиды в качестве защитных антигенов. Такие антигены могут включаться в обычные конъюгатные вакцины против H.influenzae типа b в качестве дополнительных иммуногенов или использоваться в качестве аутологичных носителей для капсульных полисахаридов H. influenzae. Белок наружной мембраны D15 с высоким молекулярным весом, обнаруженный у нетипируемых штаммов и штаммов типа b H.influenzae, был идентифицирован как перекрестно-реактивный антиген (Thomas et. al. 1990). В естественном состоянии D15 по-видимому расположен на поверхности и имеет молекулярную массу, равную примерно 80 кра, судя по анализу с помощью электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE). Было бы желательно получить последовательность в молекуле ДНК, которая кодирует этот белок наружной мембраны D15 и пептиды, соответствующие его частям, для диагностики, иммунизации и создания диагностических и иммунологических реагентов. Заболевания, вызываемые Haemophilus, являются тяжелыми, и необходимы усовершенствованные методы предупреждения, обнаружения и лечения заболеваний, таких как воспаление среднего уха, эпиглоттит, пневмония и трахеобронхит.
Краткое изложение изобретения
Данное изобретение направлено на получение очищенных и изолированных молекул нуклеиновой кислоты, содержащих по крайней мере часть, кодирующую белок наружной мембраны D15 видов Haemophilus. Молекулы нуклеиновой кислоты, содержащие по крайней мере часть, кодирующую белок наружной мембраны D15, пригодны для специфического определения штаммов Haemophilus и для диагностики инфекции, вызванной Haemophilus. Очищенные и изолированные молекулы нуклеиновой кислоты, такой как ДНК, содержащие по крайней мере часть, кодирующую белок наружной мембраны D15, также применимы для экспрессии гена D15 посредством рекомбинантной ДНК для получения экономным путем очищенного и изолированного белка наружной мембраны D15.
Белок наружной мембраны D15 или его фрагменты или аналоги пригодны в качестве иммуногенных композиций для получения вакцин против заболеваний, вызываемых Haemophilus для диагностики инфекции, вызванной Haemophilus и в качестве средств для получения иммунологических реагентов. Моно- или поликлональные антисыворотки (антитела), индуцированные белком наружной мембраны D15, полученные в соответствии с особенностями данного изобретения, применимы для диагностики инфекции, вызнанной Haemophilus, специфического определения Haemophilus (при, например, исследованиях in vitro и in vivo) для лечения заболеваний, вызываемых инфицированием.
Пептиды, соответствующие частям белка наружной мембраны D15, или их аналоги применимы в иммуногенных композициях для получения вакцин против заболеваний, вызванных Haemophilus, диагностики инфекции, вызванной Haemophilus, и в качестве средств для получения иммунологических реагентов. Моно- и поликлональные антисыворотки, выработанные к этим пептидам, полученные в соответствии с особенностями данного изобретения, применимы для диагностики инфекции, вызванной Haemophilus, специфического определения Haemophilus (при, например, исследованиях in vitro и in vivo) и для применения при пассивной иммунизации для лечения заболевания, вызванного инфицированием Haemophilus. В соответствии с одним из аспектов этого изобретения, таким образом, создана очищенная и изолированная молекула нуклеиновой кислоты, молекула, содержащая по крайней мере часть, кодирующую белок наружной мембраны D15. Молекула нуклеиновой кислоты имеет последовательность ДНК, выбираемую из:
(а) последовательности ДНК, представленной на любой одной из фигур 1А-1Е (как описано ниже) или комплементарной ей нити, и
(b) последовательностей ДНК, которые гибридизуются в строгих условиях с последовательностями ДНК, которым дано определение в (a). Последовательности ДНК, которым дано определение в (b), предпочтительно обладают по крайней мере 90% идентичностью последовательности с последовательностями, которым дано определение в (a). Последовательность ДНК, которой дано определение в (b), в частности, могут включать консенсусную последовательность, представленную на фигуре IF (как описано ниже).
В другом аспекте этого изобретения представлен очищенный и изолированный белок наружной мембраны D15 или его часть. Белок наружной мембраны D15 может быть белком наружной мембраны D15 Haemophilus и более конкретно, белком наружной мембраны D15 H.influenzae и штамм H.influenzae может быть штаммом H. influenzae типа b, таким как штаммы Ca или Eagan или Minn A, или нетипируемым штаммом H.influenzae, таким как PAK 12085 или SB33.
В дополнительном осуществлении данное изобретение также включает рекомбинантную плазмиду, приспособленную для трансформации клетки-хозяина, рекомбинантной плазмиды, содержащей плазмидный вектор, в который встроен сегмент ДНК, содержащий очищенную и выделенную молекулу ДНК, представленную здесь. Такая рекомбинантная плазмида содержит плазмидный вектор, в который встроен сегмент ДНК, который по крайней мере включает фрагмент из 18 bp, выбираемый из молекул ДНК, перечисленных выше. Рекомбинантная плазмида может быть плазмидой DS-712-2-1, имеющей номер поступления АТСС 75604, помещенной на хранение 4 ноября 1993 г, и плазмидой JB-1042-5-1, имеющей номер поступления 65006 ATCC, помещенной на хранение 4 ноября 1993 г.
Плазмиды могут быть приспособлены для экспрессии кодируемого белка наружной мембраны D15 в клетке-хозяине, которая может быть гетерологичным или гомологичным хозяином, путем включения в рекомбинантный вектор, представляемый в соответствии с дополнительным аспектом этого изобретения. Рекомбинантный вектор может включать по крайней мере сегмент ДНК, содержащий по крайней мере фрагмент из 18 bp, выбираемый из молекул ДНК, которые перечислены выше, и средства экспрессии, действенно соединенные с сегментом ДНК для экспрессии продукта гена, кодируемого тем самым в клетке-хозяине. Плазмида для экспрессии кодируемого белка наружной мембраны 15 может быть плазмидой DS-880-1,-2, имеющей номер поступления ATCC 75605, помещенной на хранение 4 ноября 1993 г, адаптированной для экспрессии белка наружной мембраны D15 в E.coli. Выбранный сегмент ДНК может кодировать полипептид из по крайней мере 6 остатков и, в частности, может выбираться из этих сегментов, кодирующих полипептид из таблицы 2 (ниже). Сегмент ДНК может, кроме того, включать последовательность нуклеиновой кислоты, кодирующую лидерную последовательность для выделения продукта гена из клетки- хозяина. Клетка-хозяин для экспрессии может выбираться из, например, грибов, дрожжей или может использоваться бакуловирусная система экспрессии.
Дополнительные аспекты этого изобретения включают белок, кодируемый молекулой ДНК, содержащей по крайней мере часть, кодирующую белок наружной мембраны D15, фрагмент или функциональный аналог такого белка, использование этого белка или аналога для вакцинации и диагностики и создания иммуногологических реагентов. Изобретение также включает антисыворотки (антитела), вырабатываемые к белку наружной мембраны D15, кодируемому молекулой ДНК, содержащей по крайней мере часть, кодирующую белок наружной мембраны, и очищенным пептидам, соответствующим частям белка наружной мембраны D15, и которые применяются при пассивной иммунизации и лечении заболеваний, вызываемых Haemophilus.
В соответствии с другим аспектом изобретения очищенный и изолированный пептид содержит аминокислотную последовательность, соответствующую аминокислотной последовательности по крайней мере части белка наружной мембраны D15 или варианта или мутанта, который сохраняет иммуногенность. Пептид может быть получен с помощью рекомбинантных методов или с помощью пептидного синтеза, посредством чего очищенный пептид освобождается от примесей, связанных с бактериями, в норме содержащими белок наружной мембраны D15. Такие синтетические пептиды предпочтительно имеют аминокислотную последовательность, выбираемую из последовательностей, представленных в таблице 2.
В соответствии с дополнительным аспектом этого изобретения представлена имумногенная композиция, которая включает белок наружной мембраны D15, его фрагменты, его функциональные аналоги или пептиды, которые перечислены выше, и физиологически приемлемый носитель для этого. Такая иммуногенная композиция, в частности, оформляется в виде вакцины для применения in vivo с целью защиты от заболеваний, вызываемых Haemophilus.
Для подобной цели иммуногенная композиция может быть оформлена в виде препарата микрочастиц, капсульного препарата или липосомного препарата. Кроме того, такая иммуногенная композиция может быть представлена в комбинации с конъюгированной нацеливающей молекулой для доставки к специфическим клеткам иммунной системы или поверхностям слизистых оболочек.
В соответствии с дополнительным аспектом этого изобретения представлен способ индуцирования защиты от заболевания, вызываемого Haemophilus, включающего стадию введения субъекту, включая млекопитающего, такого как человек, эффективного количества иммуногенной композиции или молекул нуклеиновой кислоты, которые перечислены выше, для обеспечения защитного иммунитета против инфекции, вызываемой Haemophilus.
Данное изобретение, кроме того, включает химерную молекулу, содержащую белок D15 или пептид, ему соответствующий, который, как здесь представлено, связан с другим полипептидом или белком, или полисахаридом.
Связанный полипептид или белок может включать поверхностный белок или пептид, соответствующий этой цели из патогенных бактерий, который может быть белком наружной мембраны H.influenzae P1, P2 или P6. Связанный полисахарид предпочтительно состоит из молекулы PRP из H.influenzae.
Данное изобретение будет, к тому же, понятно из следующего описания со ссылками на чертежи, в которых:
Фигура 1A представляет нуклеотидную последовательность гена D15 из штамма Ca H.influenzae типа b (ПОСЛ ИД N:1) и его производную аминокислотную последовательность (ПОСЛ ИД N:2).
Фигура 1b представляет нуклеотидную последовательность гена D15 из штамма Eagan H.influenzae типа b (ПОСЛ ИД N:3) и его производную аминокислотную последовательность (ПОСЛ ИД N:4).
Фигура 1C представляет нуклеотидную последовательность гена D15 из штамма Minn A H. influenzae типа b (ПОСЛ ИД N:5) и его производную аминокислотную последовательность (ПОСЛ ИД N:6).
Фигура 1D представляет нуклеотидную последовательность гена D15 из нетипируемого штамма SB 33 H.influenzae (ПОСЛ ИД N:7) и его производную аминокислотную последовательность (ПОСЛ ИД N:8).
Фигура 1E представляет нуклеотидную последовательность гена D15 из нетипируемой H.influenzae.
РАК 12085 (ПОСЛ ИД N: 9) и его производную аминокислотную последовательность (ПОСЛ ИД N:10).
Фигура IF представляет сравнительный анализ первичной структуры нуклеотидных последовательностей из генов D15 (ПОСЛ ИД N:1, 3, 5, 7 и 9), полученных из различных штаммов (типируемых Ca, Eagan и Minn A, нетипируемых SB 33 и PAK 12085).
Фигура 2 представляет карты рестрикции клонов pU C19/D15 (Ca), DS-712-2-1 (Eagan, DS-691-1-5 (Minn A), JB-1042-5-1 (SB 33) и JB-1042-9-4 (PAK 12085).H = Hind III, R = EcoRI S = A San 3A 1, и Xb = Xbal.
Фигура 3 представляет сравнительный анализ первичной структуры аминокислотных последовательностей белков наружной мембраны D15 (ПОСЛ ИД NN::2, 4, 6, 8 и 10), полученных из различных штаммов H.influenzae (типируемых, Ca, Eagan и Minn A, нетипируемых SB 33 и PAK 12085). Аминокислоты представлены общепринятым однобуквенным кодом. Последовательность D15 Ca используется в качестве эталонной, и точки указывают аминокислотные остатки, которые идентичны остаткам белка наружной мембраны D15 Ca.
Фигура 4 представляет построение плазмиды (DS-880-1-2), экспрессирующей D15 SB 33 (rDI5) полной длины с использованием сильного индуцибельного промотора Т7.
Фигура 5 представляет результаты анализа с помощью SDS-PAGE природного D15, очищенного с помощью афинной хроматографии, из H.influenzae штамм 30.
Фигура 6 представляет результаты анализа с помощью SDS-PAGE последовательных фракций, полученных во время очистки D15 полной длины экспрессированного в E.coli, содержащей плазмиду DS-880-1-2.
Фигура 7 показывает Ig G - антительные ответы у морской свинки на rD15 полной длины. Стрелки показывают режим иммунизации. Отбор крови производился через 0, 1, 4, 5 и 7 недель. Ограниченные отрезки представляют стандартное отклонение.
Фигура 8 представляет антительные Ig G ответы у мышей на rD15 полной длины. Стрелки показывают схему иммунизации. Пробы крови брали через 0, 1, 4, 5 и 7 недель.
Ограниченные отрезки представляют стандартное отклонение.
Фигура 9 представляет результаты анализа с помощью SDS-PAGE концевого фрагмента rD15, очищенного из GST - (фрагмент D15) слитого белка. Дорожки: 1, предварительно окрашенные маркеры низкого молекулярного веса (14 кDa, 21 кDa, 31 кDa, 45 кDa, 68 кDa, 97 кDa), 2 стандарт GST, 3 GST- (фрагмент D15) белок слияния, 4, белок слияния, расщепленный тромбином, 5 N-концевой фрагмент rD15, 6 GST, 7, маркеры низкого молекулярного веса.
Фигура 10 (a,b) представляет антительный Ig G ответ у морских свинок на N-концевой фрагмент rD15. Стрелки показывают режим иммунизации. Отбор образцов крови производили через 2, 4, 6 и 8 недель. Ограниченные отрезки представляют стандартное отклонение, и
Фигура N представляет график гидрофильности D15, построенный с использованием окна в среднем через 7 остатков по Hope, 1986.
Основное описание изобретения
Любые штаммы Haemophilus, которые имеют гены D15, могут соответственно использоваться для получения очищенных и изолированных молекул нуклеиновой кислоты (которые могут быть в форме молекул ДНК), содержащих по крайней мере часть, кодирующую белок наружной мембраны D15, примерами чего служат осуществления данного изобретения. Такие штаммы обычно доступны из клинических источников и из коллекций бактериальных культур, таких как Американская коллекция типичных культур (American Type Culture Collection). Штаммы H. influenzae могут включать типы a, b и c, нетипируемые штаммы и другие бактерии, которые продуцируют белок D15, его фрагмент или аналог. Соответствующие штаммы Haemophilus включают:
H.influenzae типа b, штамм Ca,
H.influenzae типа b, штамм Minn A;
H.influenzae типа b, штамм Eagan;
H.influenzae нетипируемый b штамм SB 33 или,
H.influenzae нетипируемый b штамм PAK 12085.
В этой заявке термин белок наружной мембраны D15 используется для определения семейства белков D15, которое включает белки, имеющие природно встречающиеся вариации в их аминокислотных последовательностях, которые обнаружены у разных штаммов, например Haemophilus. Очищенные и изолированные молекулы ДНК, содержание по крайней мере часть, кодирующую белок наружной мембраны D15 из данного изобретения, также включают молекулы, имеющие естественно присутствующие вариации в их последовательностях нуклеиновых кислот, что обнаружено у разных штаммов, например Haemophilus, и те молекулы ДНК, которые кодируют функциональные аналоги белка наружной мембраны D15. В этой заявке первый белок является функциональным аналогом второго белка, если первый белок иммунологически родственен и/или имеет одинаковые функции со вторым белком. Функциональным аналогом может быть, например, фрагмент белка или его мутант с заменой, добавлением или делецией.
В разделах данного изобретения ген D15 выделяли из штамма Ga H.influenzae типа b, как показано на фигуре 1A, H.influenzae типа B Eagan фигура 1B, H.influenzae типа b Minn A, фигура 1C, нетипируемой H.influenzae SB 33, фигура 1D, нитипируемой H.influenzae PAK 12085, фигура 16. Сравнение последовательностей нуклеиновой кислоты генов D15 и производных аминокислотных последовательностей белков наружной мембраны D15 из этих штаммов H.influenzae показало, что гены и белки высоко консервативны (фигуры 1F и 3). Консенсусная последовательность (ПОСЛ ИД N:55) для гена 15 показана на фигуре 1F.
Очищенные и изолированные молекулы ДНК, содержащие по крайней мере часть, кодирующую белок наружной мембраны D15 видов Haemophilus, примерами чего служат осуществления, описанные здесь, применимы в качестве:
- проб нуклеиновой кислоты для специфической идентификации штаммов Haemophilus in vitro и in vivo;
- продукты, кодируемые молекулами ДНК, применимы в качестве диагностических реагентов, антигенов для продукции специфичных для Haemophilus анисывороток, для вакцинаций от заболеваний, вызываемых видами Haemophilus и обнаружения инфицирования Haemophilus и
- пептиды, соответствующие частям белка наружной мембраны D15, что подтверждено примерами осуществлений, описанных здесь, применимы в качестве диагностических реагентов, антигенов для получения специфических для Haemophilus антисывороток, для вакцинации от заболеваний, вызываемых видами Haemophilus и для определения инфекций, вызванных Haemophilus.
Теперь будет дана детальная справка по предпочтительным в настоящее время осуществлениям изобретения, которая вместе с примерами служит для объяснения этого изобретения. Для ясности раскрытия, но не путем ограничения, детальное описание изобретения делится на следующие разделы:
(I) Последовательности ДНК, кодирующие белок наружной мембраны D15 из H. influenzae типа b, штамм Ca.
Клон, продуцирующий белок наружной мембраны, обозначенный D15 H.influenzae типа b (Hib), был выделен путем просмотра геномной библиотеки с помощью поликлональных антител, специфичных для белка наружной мембраны (OMP-специфичных), как ранее описано Berns и Thomas 1965, Thomas и Rossi 1986. Фрагмент ДНК, кодирующий белок D15, был выделен, субклонирован в pUC19 с получением pUC19/D15 (фигура 2) и использовался, для трансформации E.coli HB101, как описано в примере 1. Плазмидную ДНК получали из двух отдельных колоний Е. coli HB101, содержащих pUC19/D15 плазмиду. Секвенирование выполнялось на секвенаторе ДНК ABI модель 370А с использованием химической реакции с красящим терминатором и олигонуклеотидными праймерами, которые были синтезированы на синтезаторе ДНК ABI модель 380B, и очищали с помощью хроматографии. Анализ нуклеотидной последовательности гена D15 выявил, что он содержит предполагаемый промотор и открытую рамку считывания, кодирующую 789 аминокислот (фигура 1A).
Первые 19 аминокислотных остатков транслируемой открытой рамки считывания образуют типичную лидерную последовательность, которая обнаружена для других белков наружной мембраны H.influenzae типа b, таких как PI и P2 N-концевая последовательность нативного антигена D15, очищенного с помощью иммуно-аффинитета, была определена с помощью автоматической деградации по Эдману с использованием секвенатора белков ABI 477А, и было обнаружено, что она представляет собой Ala-Pro-Phe, что идентично-N-концевой аминокислотной последовательности Ala-Pro-Phe-Val-Ala-Iys - (ПОСЛ ИД N:11), предсказанной по анализу последовательности гена D15, представленного на фигуре 1A.
(II) Последовательность генов D15 из других штаммов H.influenzae. Гены D15 были выделены из других штаммов H.influenzae путем просмотра хромосомных библиотек H.influenzae типа b, штаммов Eagan, Minn A и нетипируемых штаммов H. influenzae (HTTi) SB 33, и РАК 12085, как описано в примерах 2, 3 и 4. Клоны, положительные по гибридизации, высевали на чашки и подвергали второму кругу - отбора. Рестрикционные карты полученных клонов представлены на фигуре 2. Нуклеотидные последовательности генов D15 определены для всех этих клонов (фигуры 1B - 1E) и проверено сравнение их производных аминокислотных последовательностей (фигура 3). Аминокислотные последовательности 15 из трех штаммов H. influenzae типа b были идентичны, и наблюдалось только несколько различий аминокислот в аминокислотной последовательности белка D15 из нетипируемых штаммов (фигура 3).
(III) Экспрессия D5 и его фрагментов в E.coli
Так как D15 экспрессируется в небольших количествах штаммами H.influenzae, выгодно экспрессировать этот антиген как рекомбинантный белок в гетерологичной системе, такой как E.coli, или модифицировать микроорганизм H.influenzae для усиления природной экспрессии D15. Фрагмент Hind III/EcoRI из штамма Ca H.influenzae типа b, кодирующий белок D15 полной длины, экспрессировался в pUC19, а не в pUC18, что наводит на мысль о том, что Iac промотор помогает в экспрессии гена D15 в E.coli, даже если присутствует природный промотор гена D15. Система экспрессии Т7 - строго контролируемая, индуцибельная система, которая дает большую выгоду при экспрессии гетерологичных белков в E.coli. Система экспрессии Т7 описана в патенте США 4952496. Поэтому сконструированы клоны, которые используют систему Т7 для экспрессии зрелого белка D15, который содержит дополнительный метиониновый остаток на амино-конце. Сигнальная последовательность D15 удалялась во время этого процесса конструирования. Рекомбинант D15 (названный rD15) полной длины экспрессировался при включении частей, которые позволяют легко очищать белок D15. Гены D15 из штамма Ca H.influenzae типа b и нетипируемого штамма SB 33 H. influenzae экспрессировались на высоком уровне в E.coli с использованием системы Т7, чтобы дать возможность продукции больших количеств белка rD15. Здесь описано конструирование клона DS-880-1-2, который экспрессирует ген D15 SB 33 (смотрите фигуру 4 и пример 5).
Белок D15 был иммунологически сходным с его природным аналогом, выделенным из штаммов H.influenzae типируемых и нетипируемых (см. ниже). Таким образом, rD15 может использоваться как перекрестно-реактивный антиген в диагностическом наборе для определения многих, если не всех, штаммов H.influenzae и других бактерий, которые продуцируют белок наружной мембраны D15 или его аналог. Альтернативно D15 может использоваться в качестве антигена для специфического определения присутствия H.influenzae в образце.
Фрагмент усеченного D15 экспрессировался в Е.coli как белок слияния с глютатион-S-трансферазой (GST), как описано в примере 6. Конструкция была предназначена для экспрессии N-концевого фрагмента белка D15. Белок слияния экспрессировался на высоком уровне из конструкции pGEX-2T, и N-концевой фрагмент отщеплялся от белка-носителя GST путем обработки тромбином. Эта процедура давала молекулу, названную N-концевым фрагментом rD15, который заключает в себе аминокислоты 63-223 из белка D15. Этот N-концевой фрагмент rD15 был высокоиммуногенным и вызывал образование защитных антител от заражения живыми.
(IV) Очистка природного D15 из клеточной массы H.influenzae. Данное изобретение также представляет способ получения очищенного природного белка D15 из H.influenzae. Белок экстрагируется и очищается с использованием фаффинитета из клеточной массы или типируемых или нетипируемых штаммов с помощью процесса, включающего растворение белка в водном растворе детергента (смотрите пример 13). Природный белок D15 из штамма 30 нетипируемой H.influenzae солюбилизировали с помощью 50 мм Транс-HCl /0,5%, Тритона X-100/ 10 мМ ЭДТУ буфером pH 8,0 и далее очищали на колонке для аффинной хроматографии с D15 - специфичными моноклональными антителами (фигура 5А). Белок в 80 кDa элюировали с колонки 50 мМ диэтиламином, pH 12,0, и, как показано, он реагирует с D15 - специфическими моноклональными антителами при иммуноблоттинговом анализе (фиг. 5В). Природный D15 также высокоиммуногенен у экспериментальных животных. Кроличьи анти- D15 антисыворотки реагировали со всеми штаммами H.influenzae, что определялось с помощью иммуноблоттинга.
(V). Очистка рекомбинантного белка D 15 полной длины, экспрессированного в E.Coli.
Рекомбинантный белок D15 полной длины экспрессировался в тельцах включения в E.coli. Как показано на фигуре 6, очистка телец включения rD15 достигалась с помощью последовательной экстракции лизата клеток E.coli 50 мМ Трис-HCl, pH 6,0, затем 50 мМ Трис, содержащим 0,5% Тритон X-100 и 10 мМ ЭДТА, pH 8,0. После центрифугирования более 95% белков в подученном в результате осадке было белком в 80 кDa по анализу SDS-PAGE, который реагировал с D15 - специфическими моноклональными антителами на иммуноблоте. Как было обнаружено, N-концевая последовательность rD15 представляет собой Met-Ala-Pro-Phe-Val-Iys-Asp- (ПОСЛ ИД N:54), которая идентична предсказанной аминокислотной последовательности.
Тельца включения rD15 делали растворимыми с помощью смеси фосфатно-буферного раствора (ФБР), 0,5% Тритона X-100, 10 мМ ЭДТУ и 8М мочевины (смотрите пример 8). После диализа против ФБР для удаления мочевины более 80% белка D15 оставалось растворенным. Этот растворимый антиген rD15 использовался для исследований по иммуногенности описанных ниже. По экспериментам с выращиванием в колбах при встряхивании было установлено, что из 1 л бактериальной культуры E.coli получалось 10 мг растворимого белка rD15. Ясно, что выращивание рекомбинантных штаммов E.coli при оптимизированных условиях ферментации значительно повысит уровень продукции rD15.
(VI) Иммуногенностъ рекомбинантного белка D15.
(rD15) полной длины.
Иммуногенность белка rD15 полной длины изучали на морских свинках и мышах. При использовании схем иммунизации, описанных на фигуре 7, доза rD15 в 15 мкг вызывала высокие титры IgG у морских свинок при введении в присутствии или адъюванта Фрейнда или AlPO4. В исследовании зависимости ответа от дозы на мышах белок, по-видимому, был иммуногенным в такой низкой дозе, как 5 мкг, или в адъюванте Фрейнда (фигура 8А) или с AlPO4 (фигура 8B).
Защитная способность rD15 против инфекции, вызываемой H.influenzae типа b, изучена на модели бактеремии у крысят по существу такой, которая описана Loeb (1987). Так, крысята, пассивно иммунизированные анти- rD15 антисывороткой морских свинок, имели значительно меньшую бактеремию, чем контрольные, которым вводили предварительно взятую сыворотку, что согласуется с предыдущим сообщением Thomas et.al (1990).
(VII) Очистка и определение характеристик N-концевого фрагмента rD15.
Усеченный фрагмент rD15, соответствующий N-концу белка D15 (остатки с 22 по 223), как описано в примере 6, экспрессировался в Е.coli, как растворимый белок, слитый с GST. Слитый белок (46 кDa) легко экстрагировался при использовании фосфатно-буферного раствора (ФБР). Очистка GST-D15 фрагмента слитого белка достигалась с помощью одностадийного процесса очистки с помощью аффинной хроматографии на колонке с глютатион-сефарозой (фигура 9, дорожа 3). Расщепление 46 кDа слитого белка с помощью тромбина давало два фрагмента (фигура 9, дорожка 4), 26 кDа белок, который соответствовал очищенному стандарту GST (фигура 9, дорожка 2), и 20 кDa полипептид, который имел размер, ожидаемый для N-концевого фрагмента rD15 (аминокислотные остатки с 63 по 223), соответственно. Разделение этих двух белков достигалось с помощью второго цикла аффинной хроматографии из глютатион-сефарозе 4B. Из экспериментов с выращиванием в колбах на качалке было установлено, что из 1 л бактериальной культуры E.coli выделяется около 1 мг очищенного N-концевого фрагмента rD15. Ясно, что выращивание рекомбинантых штаммов E.coli при оптимизированных условиях ферментации значительно повысит уровень продукции N-концевого фрагмента rD15.
Идентичность 20 кDa полипептида и 26 кDa белка была подтвержена как иммуноблоттингом, так и секвенированием белка. N-концевая последовательность 20 кDa полипептида, как было обнаружено, представляет собой NH2-Ser-Leu-Phe-VaL-Ser-GLy-Arg-Phe-Asp-Asp-VaL-Lys-ALa-His- GLn-GLu-Gly-Asp-VaL-Leu-VaL-VaL-Ser (Посл. ИД N:12), которая соответствует остаткам с 63 по 85 первичной последовательности D15. Этот результат доказывает, что существует ложный сайт тромбинового расщепления в последовательности 15 и что первые 42 аминокислоты фрагмента rD15 отрезаются во время тромбинового расщепления. Таким образом, окончательный N-концевой фрагмент rD15 был длиной в 161 аминокислоту, соответствующую остаткам с 63 по 223 первичной последовательности D15. N-концевая последовательность, полученная для 26 кDa белка (NH2-Met-Ser-Pro-ILe-Leu-GLy-Tyr-Trp-Lys (ПОРСЛ ИД N:13) подтвердила, что это была GST.
(VIII) Иммуногенность N-концевого фрагмента rD15.
Иммуногенность N-концевого фрагмента rD15 была испытана на морских свинках с использованием различных адъювантов. При использовании схем иммунизации, описанных на фигуре 10, 10 мкг доза N-концевого фрагмента rD15 индуцировала хороший вторичный ответ у морских свинок почти со всеми испытанными адъювантами. Самый высокий титр против D15 наблюдался в группе морских свинок, иммунизированных N-концевым фрагментом rD15 в адъюванте Фрейнда. Вторым наилучшим адъювантом был Titermax (CytRx Inc). Другие два адъюванта, TPADA (трипалмитил-Cys-Ser-GLu4) и AlPO4 были одинаково сильнодействующими.
(IX). Защитная способность N-коцевого фрагмента rD15 от заражения H. influenzae типа b.
Моделью с заражением in vivo для оценки защитной способности антигена от заболеваний, вызываемых Haemophilus, является модель бактеремии у крысят, которая описана Loeb, 1987. Защитная способность N-концевого фрагмента rD15 от заражения H. influenzae типа b оценивали на этой модели на крысах. Как показано в таблице 1, у крысят, пассивно иммунизированных кроличьей сывороткой против N-концевого фрагмента rD15, выявлена значительно меньшая бактеремия по сравнению с бактеремией у крысят, которым вводили предварительно взятую сыворотку.
Так как пассивно вводимые антисыворотки против N-концевого фрагмента rD15, как было обнаружено, являются защитными на модели бактеремии у крысят, было интересно идентифицировать защищающий(е) эпитоп(ы) этого N-концевого фрагмента rD15. Первые девять перекрывающих пептидов из белка D15, которые перечислены в таблице 2, были химически синтезированы на основе аминокислотной последовательности, выведенной из последовательности гена D15 штамма Ca H.influenzae типа b (фигура 1). Эти синтетические пептиды оценивали по их реактивности или с кроличьей антисыворотки или с антисывороткой от морских свинок, полученных к очищенному N-концевому фрагменту rD15, с помощью ELISA. Как показано в таблице 3, как антисыворотка морских свинок, так и кроличья реагировали с группой пептидов D15, включая пептиды D15-Р4 - D15-P8, заключающие в себе остатки с 93 по 209 первичной последовательности D15.
Дополнительные исследования были выполнены, чтобы определить, может ли защита от H. influenzae типа b, наблюдаемая при применении кроличьей антисыворотки против D15 у крысят, нейтрализоваться пептидами D15. В первом эксперименте кроличью антисыворотку к N-концевому фрагменту rD15 вводили группе из семи крысят в присутствии или в отсутствие смеси девяти пептидов (D15-P2 - D15-P10). Животным в группе положительного контроля вводили кроличью антисыворотку к N-концевому фрагменту rD15, смешанную с очищенным фрагментом D15, а в группе отрицательного контроля вводили смесь только девяти пептидов. Как показано в таблице 4, у крысят, пассивно иммунизированных антисывороткой к N-концевому фрагменту rD15 (группа 1), выявлен значительно более низкий уровень бактеремии (3%, p = 1,2 • 10-7) по сравнению с уровнем в группе отрицательного контроля (группа 4,100%), что согласовывалось с ранее полученными результатами. Защита, создаваемая кроличьей антисывороткой к N-концевому фрагменту rD15, в значительной степени нейтраизовалась добавлением очищенного N-концевого фрагмента rD15 (группа 3,64%), что демонстрируется отсутствием значительного различия в уровнях бактеремии между группой 3 и группой 4 (p = 0,09). Хотя добавление смеси девяти пептидов D15 только незначительно нейтрализовало защиту, придаваемую антисывороткой (группа 2,13%) по сравнению с группой 1 (3%), различие в числе бактерий между этими двумя группами было статистически значимым (p = 0,0037).
Чтобы более четко определить защитные эпитопы N-концевого фрагмента rD15, вышеизложенный эксперимент повторяли со смесью пяти пептидов (пептиды с D15-P4 до D15-P8), которые были выбраны по их сильной реактивности с кроличьей антисывороткой к N-концевому фрагменту rD15. Результаты, полученные из этого второго эксперимента, показали, что защита, наблюдаемая при использовании кроличьей антисыворотки к N-концевому фрагменту rD15 (таблица 5, группа 1), полностью блокировалась добавлением этой смеси из пяти пептидов (таблица 5,группа 2,106%, p = 0,53 • 10-8). Эти результаты убедительно показывают, что смесь синтетических пептидов D15 может использоваться в качестве иммуногенов для индуцирования защитных антител к H.influenzae.
(X) Предсказание строения эпитопа и синтез пептида
Чтобы картировать иммунодоминантные Т-клеточные или B-клеточные эпитопы D15, перекрывающие синтетические пептиды, включающие всю последовательность белка D15 (таблица 2 - ПОСЛ ИД N:14 - 49), были синтезированы с использованием t-Boc твердофазного пептидного синтеза, как описано в примере 15. Пептиды были выбраны на основе их высокого показателя гидрофильных β-поворотов, установленного с помощью анализа по предсказанию вторичной структуры (фигура 11). Такие пептиды, вероятно, находятся на поверхности и являются антигенными. Пептиды длиной более 25 остатков были выбраны для лучшей имитации природных эпитопов.
(XI) Идентификация и определение характеристик иммунодоминантных эпитотов rD15 с использованием синтетических пептидов.
Чтобы картировать линейные B-клеточные эпитопы D15, перекрывающие синтетические пептиды, представляющие всю последовательность D15, отдельно наносили на платы для ELISA и испытывали с пробами нескольких анти rD15 антисыворотки, как описано в примере 19. Результаты суммированы в таблице 6. Мышиные антисыворотки, полученные к rD15, реагировали со всеми пептидами D15, но главные эпитопы располагались в пептидах D15 - Р8 (остатки 180-209 - ПОСЛ ИД N:21), D15 - Р10 (остатки 219-249 -ПОСЛ ИД N:23), D15- P11 (остатки 241- 270 -ПОСЛ ИД N:24) и D15 - Р26 (остатки 554-582 - ПОСЛ ИД N:39), соответственно. Кроличья анти-D15 антисыворотка распознавала только пептиды D15-Р4 (остатки 93-122- ПОСЛ ИД N:17), D15-Р14 (остатки 304-333 - ПОСЛ ИД N:27) и D15-P36 (остатки 769-798 - ПОСЛ ИД N:49). Антисыворотки морских свинок к D15 реагировали с пептидами D15-Р2 (остатки 45-72 -ПОСЛ ИД N:15), D15-Р4 (остатки 93-122 -ПОСЛ ИД N: 17), D15-Р6 (остатки 135-164 - ПОСЛ ИД N:19), D15-P8 (остатки 180-209 - ПОСЛ ИД N:21), D15-P14 (остатки 304-333 - ПОСЛ ИД N:27), D15-Р27 (остатки 577- 602 - ПОСЛ ИД N:40). Иммунодоминантные линейные В-клеточные эпитопы D15, таким образом, как было обнаружено, располагаются в пептидах D15-Р4 (остатки 93-122 - ПОСЛ ИД N:17) и D15-Р14 (остатки 304-333 - ПОСЛ ИД N:27), так как они являются единственными двумя пептидами, распознаваемыми rD15-специфическими антисыворотками от всех трех видов животных. Эти результаты показывают, что пептиды, содержащие линейные B-клеточные эпитопные последовательности, описанные выше, могут использоваться в качестве мишеневых антигенов, в, например, диагностических наборах для обнаружения присутствия антител к D15 и H.influenzae в образцах.
(XII). Идентификация и определение характеристик иммунодоминантных Т-клеточных эпитопов D15 при использовании синтетических пептидов.
Значение цитокиновых сетей в иммунном ответе и воспалительных реакциях при иммунитете и воспаление и их изменения при патологии становится более ясным, так как идентифицируются новые члены цитокинового семейства и определяются их характеристики. Mills et.al (1993) недавно сообщили, что происходит быстрое очищение легких от B.pertussis у мышей после заражения через шесть недель после респираторной инфекции или после двух иммунизаций цельноклеточной противококлюшной вакциной. Было обнаружено, что клетки селезенки от этих иммунизированных мышей секретируют на высоком уровне IL-2 и IFN-γ и на низком уровне IL-5 в присутствии коклюшного антигена (коклюшного токсоида, фимбриального гемагглютинина (ФГА) и пертактина). Этот результат наводит на мысль, что Th1-клеточная (T-клетки, продуцирующие высокие уровни IL-2 и IFN-γ) пролиферация очень важна для выздоровления от респираторной инфекции. Генерация Th1- и Th2-клеточных субпопуляций регулируется с помощью равновесия между различными группами цитокинов, преимущественно IL-2 и IL-4 (Trinchieri, 1993), IL-12 и IL-4 ответственны за дифференциацию Th1 и Th2 клеток, соответственно. Одна из ролей клеток Th2 в иммунной системе состоит в обеспечении хелперной активности для появления высоких уровней антиген-специфичных антител поело иммунизации. Антигены, содержание Th1-эпитопы, стимулируют антиген-специфичные T-клетки к продукции высоких уровней IL-2 и IFN-γ, тогда как Th2 эпитопы стимулируют экспресиию IL-4 на высоком уровне. Th0 эпитопы стимулируют синтез IFB-γ и IL-4.
Мало что известно о клеточном иммунном ответе на белки наружной мембраны H. influenzae и их роли в защите от инфицирования H.influenzae и болезней. С этой целью изобретатели выполнили исследования клеточного ответа, вызванного у мышей после иммунизации rD15. D15 - специфичные T-клеточные эпитоты были определены с использованием пептидов D15 и T-клеточных линий,
полученных от пяти мышей BAL В/с, иммунизированных rD15 (см. пример 23). Лимфоцитные пролиферативные ответы D15 - специфических Т-клеточных линий на перекрывающие пептиды D15 определялись общепринятыми исследованиями цитокинов, как описано в примере 24. Результаты, суммированные в таблице 7, показали, что стимуляция только некоторыми синтетическими пептидами вызывала пролиферативные ответы и выделение специфических цитокинов. Синтетические пептиды, соответствующие остаткам 114-143 (D15-Р5 - ПОСЛ ИД N:18), 282-312 (D15-P-13 - ПОСЛ ИД N: 26) и 577-602 (D15-Р27 - ПОСЛ ИД N:40) и 219-249 (D15-Р10 - ПОСЛ ИД N:23), 262-291 (D15-P12 - ПОСЛ ИД N:25), 390-416 (D15-P18 - ПОСЛ ИД N:31), 410-435 (D15-P19 - ПОСЛ ИД N:32), 554-582 (D15-Р26 - ПОСЛ ИД N:39), 596-625 (D15-Р28 - ПОСЛ ИД N:41), 725-750 (D15-Р34 - ПОСЛ N:47) и 745-771 (D15-Р35 - ПОСЛ ИД N:48), как было показано, обладают высоким стимуляторным действием на rD15-специфические Th0-клетки и th1-клетки мышей BALB/c, соответственно. Поэтому эти иммунодоминантные Т-клеточные эпитоты могут использоваться в качестве аутологичных носителей для PRP и/или OMP B-клеточных эпитопов для усиления их иммуногенности, Th1-клеточные эпитопы, которым дано определение выше, могут применяться в вакцинных составах против H. influenzae для индуцирования специфичных клеточных иммунных ответов к H. influenzae.
(XIII) Иммунногенность пептидов D15
Чтобы определить являются ли синтетические пептиды D15 иммуногенными, свободные пептиды оценивали индивидуально на их иммуногенность. Кроличью и морских свинок антипептидные антисыворотки испытывали на их реактивность с иммунозирующими пептидами, а также с природным D15 и rD15 с помощью ELSA и иммуноблоттинга. Как показано в таблице 8, все антисыворотки морских свинок к пептиду D15, за исключением тех, образование которых индуцировано D15-P26 (ПОСЛ ИД N: 39), D15-P29 (ПОСЛ ИД N:42), D15-P30 (ПОСЛ ИД N:43) и D15-P31 (ПОСЛ ИД N:44), являются иммуногенными, что показано с помощью ELISA. Индукция высоких титров пептид - специфических IgG антител свободными пептидами ясно показывает, что большинство пептидов содержат как функциональную Т-хелперную детерминанту, так и B-клеточные эпитопы. Кроме того, эти антипептидные антисывороки распознавали D15 в иммуноблоттинговом исследовании. Так как большинство пептидов содержат сильные функциональные Т-хелперные детерминанты и индуцируют сильный IgG антительный ответ у млекопитающих, они являются кандидатами в иммуногены для включения в вакцинный препарат против H.influenzae. Антисыворотка, специфичная к пептиду D15, перекрестно реагировала с D15 из нетипируемых штаммов H.influenzae, судя по иммуноблоттингу. Этот результат показывает, что иммуногенные пептиды D15 содержат эпитопы, которые высоко консервативны среди типируемых и нетипируемых штаммов H.influenzae. Кроме того, поликлональные антитела к этим эпитопам применимы для обнаружения H.influenzae в биологических пробах.
Поэтому эти консервативные эпитопы D15 могут использоваться или индивидуально, или в комбинации для получения перекрестно-реактивных синтетических иммуногенов для типируемых и нетипируемых штаммов H.influenzae и других бактерий, которые продуцируют белок D15, его фрагмент или аналог. Пептиды, описанные выше, можно дополнительно полимеризовать или модифицировать с помощью липидов, липопептидов или соединять с полисахаридами, включая PRP в качестве синтетических гликопептидных или липогликопептидных конъюгатов для получения альтернативных вакцин. Эти вакцины могут использоваться для иммунизации от заболеваний, вызываемых H.influenzae, при введении млекопитающим, например, внутримышечно или парентерально, или при доставке с использованием микрочастиц, капсул, липосом и молекул для направленной доставки, таких как токсины или их фрагменты и антитела, к клеткам иммунной системы или слизистых оболочек.
(XIV) Применение D15 в качестве белка-носителя для получения гликоконъюгатов.
Чтобы определить, может ли D15 служить как в качестве защитного антигена, так и носителя, выполняли эксперименты до конъюгации D15-PRP, как описано в примере 14. Конъюгаты D15-PRP, как было обнаружено, являются высокоиммуногенными у кроликов и способны вызывать как анти-D15, так и анти-РRР IgG антительные ответы, судя по D15-специфическом ELISA и иммуноисследованию с PRP-BSA (таблица 9).
Эти результаты ясно демонстрируют практическое применение D15 в качестве белка-носителя для технологии гликоконъюгации.
При предпочтительном осуществлении данного изобретения функционирование D15 в качестве носителя может в основном использоваться для получения химерных молекул и конюъгатных вакцин против патогенных бактерий, включая инкапсулированные бактерии. Таким образом, гликоконъюгаты данного изобретения могут применяться для вакцинация по созданию защиты от инфицирования любыми бактериями, имеющими полисахаридные антигены, включая, например, Haemophilus,influenzae, streptococcus, pneumoniae, Escherichia coli, Neisseria meningitidis, Salmonella typhi, Streptococcus mutans, Cryptococcus neoformans, Klebsiella, Staphylococcus aureus и Pseudomonas aeruginosa.
При другом осуществлении D15 в функции носителя может использоваться, например, для индуцирования иммунитета к ненормальным полисахаридам опухолевых клеток или для продукции противоопухолевых антител, которые можно конъюгировать с химиотерапевтическими или биоактивными препаратами.
Соответственно, данное изобретение представляет первичную последовательность и получение антигена (D15) из H.influenzae, который может использоваться для предупреждения и диагностики заболеваний, вызываемых Haemophilus. В частности, изобретатели открыли, что рекомбинантый D15 или его фрагменты могут вызывать защитные антительные ответы к H.influenzae типа b при заражении живыми бактериями. Таким образом, данные изобретения применимы в вакцинах. Изобретение также раскрывает нуклеотидные последовательности генов D15, выделенных как из штаммов H.influenzae типа b, так и из нетипируемых штаммов. Раскрыты сегменты ДНК, кодирующие D15, и показан незначительный полиморфизм как в их нуклеотидах, так и в производных аминокислотных последовательностях (фигуры 1F и 3). Эти сегменты ДНК могут использоваться для получения иммуногена, по существу свободного от других антигегов H.influenzae (таких, как PRP и липоолигосахариды (ЛОС) путем применения технологии генной инженерии. Данное раскрытие, кроме того, представляет новые методики, которые могут использоваться для получения по существу чистого D15 или его фрагментов, а также функциональных аналогов. Рекомбинантный белок D15, его фрагмент или аналог могут быть получены в подходящей системе экспрессии, такой как E.coli, Haemophilus, Bordetella, Bacillus, грибы, дрожжи, бакуловирусы, поксвирус, вирус коровьей оспы или системы экспрессии млекопитающих.
В одном из осуществлений данное изобретение касается процесса получения вакцинных композиций, которые включают очищенный рекомбинантный белок D15 (rD15) или фрагменты rD15, которые иммунологически перекрестно-реактивны с природным D15. В частности, ген, кодирующий весь белок D15, и сегмент ДНК, кодирующий N-концевой фрагмент rD15, слитый с глютатион-S-трансферазным геном, были сконструированы и экспрессированы в E.coli. Экспрессированный белок rD15 и его фрагменты, как было обнаружено, иммунологически перекрестно реагируют с природным антигеном D15, выделенным как из типируемых, так и нетипируемых штаммов H. influenzae, и таким образом представляют собой перекрестно-реактивные иммуногены для включения в вакцину против заболеваний, вызываемых H.influenzae. Кроме того, конвалесцентная сыворотка к Haemophilus распознавала очищенный D15 из H.influenzae, как описано здесь, rD15 и N-концевой фрагмент rD15.
При другом осуществлении данное изобретение представляет ген, кодирующий блок наружной мембраны D15 из H.influenzae, имеющий специфические нуклеотидные последовательности, описанные здесь, или последовательности, по существу гомологичные им (т.е. те, которые гибридизуются в строгих условиях с такими последовательностями), для получения генно-инженерных гибридов или химерных белков, содержащих фрагмент D15, слитый с другим полипептидом, или белком, или полисахаридом, такими как белки наружной мембраны H.influenzae, например, P1, P2 или P6, или PRP. В результате гибриды, химерные белок или гликоконъюгаты могут иметь более высокую защитную способность против H.influenzae, чем D15, или P1, или Р2, или Р6, или один PRP.
Таким образом, белок наружной мембраны D15 может функционировать и как защитный антиген, и как носитель в конъюгатной вакцине для обеспечения аутологичного примирования Т-клеток, причем для обеспечения аутологичного примирования Т-клеток, причем гаптеновая часть конъюгата является частью капсульного полисахарида (PRP) H.influenzae. Этот D15 - нуглеводный конъюгант может вызывать образование антител к PRP D15, и таким образом должен повышать уровень защиты против заболеваний, связанных H.influenzae, особенно у детей.
В еще одном осуществлении данное изобретение заключает в себе по существу чистую форму по крайней мере одного белка или пептида, содержащего аминокислотную последовательность, соответствующую по крайней мере одну антигенную детерминанту D15, пептида, который способен вызвать образование поликлональных антител к H.influenzae у млекопитающих. Эти D15 - специфические антитела применимы в тест-наборах для обнаружения присутствия H.influenzae в биологических образцах. Пептиды могут иметь, например, аминокислотные последовательности, соответствующие остаткам 20-49, 45-74, 68-99, 93-122, 114-143, 135-164, 157-187, 180-209, 199-288, 219-249, 241-270, 262-291, 282-312, 304-333, 325-354, 346-375, 367-369, 390-416, 410-435, 430-455, 450-477, 471-497, 491-516, 511-538, 532-559, 554-582, 577-602, 596-625, 619-646, 641-666, 662-688, 681-709, 705-731, 725-750, 745-771, 769-798 (ПОСЛ ИД NN: с 14 по 49) белка 15 штампа Ca H.influenzae типа b, соответственно, как представлено в таблице 2, ниже, или его любых части, варианта или мутанта, которые сохраняют иммуногенность.
В еще одном осуществлении данное изобретение представляет чистый природный белок D15, экстрагированнй и хроматографически очищенный из культур типируемых и нетипируемых штаммов H.influenzae. Новые процедуры включают экстракцию белка D15 из клеточной массы по методикам, известным для других белков наружной мембраны, с помощью водного раствора детергента с последующей очисткой центрифугированием и хроматографией. Очищенный природный антиген D15 может использоваться для иммунизации млекопитающих от заболеваний, вызванных H. influenzae, например, путем внутримышечного или парентерального введения или с использованием средств его доставки с применение микрочастиц, капсул, липосом и конъюгированных молекул, обеспечивающих направленную доставку, таких как токсины или их фрагменты и антитела.
Другой аспект данного изобретения состоит в том, что белок наружной мембраны, его ферменты или аналоги, или пептиды, соответствующие частям D15, могут быть компонентами поливалентной вакцины от воспаления среднего уха. Эта поливалентная вакцина включает по крайней мере одну иммуногенную детерминанту D15, которая описана здесь, наряду с по крайней мере одним защитным антигеном, выделенным из Streptococcus pneumoniae, Branhamella (Moraxella) catarrhalis, Staphilococcus aureus или респираторного синцитиального вируса, в присутствии или в отсутствие адъюванта.
Пептиды D15 (таблица 2) или его какая-то часть, вариант или мутант могут быть легко синтезированы или вручную, или с помощью коммерчески доступного пептидного синтезатора, такого как синтезатор Applied Biosystems модель 430A.
Специалисту будет ясно видно, что различные осуществления данного изобретения имеют много способов применения в области вакцинации, диагностики и лечения заболеваний, вызываемых Haemophilus инфекций, и производстве, иммунологических реагентов. Дальнейшее не ограничивающее обсуждение таких способов применения дополнительно представлено ниже.
1. Получение и применение вакцины.
Иммуногенные композиции, пригодные для использования в качестве вакцин, могут быть получены из иммуногенного белка наружной мембраны D15, его фрагменты или аналогов, и/или пептидов, соответствующих частям D15, которые раскрыты здесь. Вакцина вызывает иммунный ответ, при котором продуцируются антитела, включая антитела к белку наружной мембраны D15 и антитела к D15, которые являются опсонизирующими или бактерицидными. Если вакцинированный субъект заразится Haemophilus, антитела связываются с белком наружной мембраны D15 и тем самым инактивируют бактерии. Опсонизирующие и бактерицидные антитела являются примерами антител, полезных при защите от заболеваний.
Вакцины, содержащие пептиды, в основном, хорошо известны в этой области, примеры их представлены в патентах США 4601903, 4599231, 4599230 и 4596792, все эти работы приведены здесь в виде ссылки. Что касается любой последующей ссылки на патенты и ссылок в этом описании, они также включены здесь путем ссылки, без дальнейших указаний на это. Вакцины могут готовиться в виде инъекционных форм, в виде растворов или эмульсий. Белок наружной мембраны D15, его фрагменты или аналоги или пептиды, соответствующие частям D15, могут смешиваться c физиологически приемлемыми наполнителями, которые совместимы с белком наружной мембраны D15, его фрагментами, аналогами или пептидами. Наполнители могут включать воду, физиологический раствор, декстрозу, глицерин, этанол и их комбинации. Вакцина может, кроме того, содержать незначительные количества вспомогательных веществ, таких как смачивающие или эмульгирующие средства, забуферивающих pH средств или адъювантов для повышения эффективности вакцин. Методы достижения адъювантного эффекта для вакцины включают использование средств, таких как гидроксид алюминия или фосфат алюминия (алюм), обычно используемых в виде 0,05-0,1% раствора в фосфатно-буферном физиологическом растворе. Вакцины могут вводиться парентерально путем подкожной или внутримышечной инъекции. Альтернативно могут быть желательны другие способы введения, включая свечи и пероральные препараты. Для свечей связывающие вещества и носители могут включать, например, полиалкиленгликоли или триглицериды. Пероральные рецептуры могут включать обычно используемые составляющие, такие как, например, сахарин, целлюлоза, карбонат магния фармацевтических марок и тому подобное.
Этим композициям придают форму растворов, суспензий, таблеток, пилюль, капсул, препаратов с поддерживаемым выделением или порошков и они могут содержать 10-95% белка наружной мембраны D15, его фрагмента, аналогов и/или пептидов.
Вакцины вводятся способом, соответствующим дозированной форме, и в количестве, которое терапевтически эффективно, обладает защищающим и иммуногенным действием. Количество, которое нужно ввести, зависит от субъекта, которого нужно лечить, включая, например, способность иммунной системы индивидуума синтезировать антитела и, если необходимо, продуцировать клеточный иммунный ответ. Точные количества активного ингредиента, необходимые для введения, зависят от суждения практикующего врача. Однако подходящие интервалы дозирования легко определяются опытным специалистом и могут быть порядка микроорганизма белка наружной мембраны D15, аналога, фрагмента и/или пептидов. Подходящие режимы для первоначального введения и повторных доз также различны, но могут включать первичное введение с последующими последовательными введениями. Дозировка вакцины может такие зависеть от пути введения и меняется в соответствии с размерами индивидуума.
Молекулы нуклеиновой кислоты, кодирующие белок наружной мембраны D15 данного изобретения, могут также непосредственно применяться для иммунизации путем введения непосредственно ДНК, например, с помощью инъекций для генетической иммунизации или путем конструирования живых векторов, таких как БЦЖ, аденовирус, поксвирус или вирус коровьей оспы. Обсуждение некоторых живых векторов, которые использовались для переноса гетерологичных антигенов к иммунной системе, приводится, например, у O'Hagan (1992). Процессы по непосредственным инъекциям ДНК испытуемым субъектам для генетической иммунизации описаны, например, Ulman et al. (1993).
Применение пептидов in vivo может сначала потребовать их химической модификации, так как сами пептиды могут не обладать достаточно длительным полупериодом присутствия в сыворотке и/или ткани. Такие химические модифицированные пептиды здесь называются пептидными аналогами. Термин пептидный аналог распространяется на любой функциональный химический эквивалент пептида, характеризующийся повышенной стабильностью и/или эффективностью in vivo или in vitro, что касается практического применения изобретения. Термин пептидный аналог, используемый здесь, распространяется на любое аминокислотное производное пептидов, описанных здесь. Пептидные аналоги, рассматриваемые здесь, получают с помощью процедур, которые включают, но не ограничиваются ими, модификации в боковых цепях, включения неприродных аминокислот и/или их производных во время химического синтеза, и использование перекрестных связей и другие методы, которое придают конформационные ограничения пептидам и их аналогам.
Примеры модификаций боковой цепи, рассматриваемые в данном изобретении, включают модификацию аминогрупп, такие как с помощью восстановительного алкилирования по реакции с альдегидом с последующим восстановлением NaBH4, амидирование с помощью метилацетимидата, ацетилирование с помощью уксусного ангидрида, карбамилирование аминогрупп с помощью цианата, тринитробензилирование аминогрупп с помощью 2,4,6-тринитробензолсульфоновой кислоты (ТНБС), алкилирование аминогрупп с помощью ангидрида янтарной кислоты и тетрагидрофталевого ангидрида и пиридоксилирование лизина с помощью пиридокса -5'-фосфата с последующим восстановлением NaBH4.
Гуанидиногруппа аргипиновых остатков может быть модифицирована путем образования продуктов гетероциклической конденсации с помощью реагентов, таких как 2,3-бутадион, фенилглиоксаль и глиоксаль.
Карбоксильная группа может быть модифицирована путем карбодиимидной активации через образование о-ацилизомочевины с последующей последовательной дериватизацией, например, до соответствующего амина.
Сульфгидрильные группы могут быть модифицированы такими методами, как карбоксиметилирование с помощью иодуксусной кислоты или иодацетамида, окисление пермуравьиной кислоты до цистеиновой кислоты, образование смешанных дисульфидов с помощью других тиольных соединений, реакция с малеимидом, малеиновым ангидридом или другим замещенным малеимидом, образование производных двухвалентной ртути при использовании 4-хлормеркурибензоата, 4-хлормеркурифенилсульфоновой кислоты, фенилмеркурихлорида, 2-хлормеркури-4-нитрофенола и других производных двухвалентной ртути, карбамоилирование цианатом при щелочном pH.
Триптофановые остатки могут быть модифицированы путем, например, оксидирования N-бромсукцинимидом или алкилирования индольного кольца 2-гидрокси-5-нитробензил-бромидом или сульфонилгалогенидами. Триозиновые остатки могут быть изменены путем нитрования тетранитрометаном с образованием 3-нитротирозиновых производных.
Модификация имидазольного кольца гистидина может быть выполнена путем алкилирования производными иодуксусной кислоты или N-карбэтоксилирования диэтилпирокарбоната.
Примеры включения неприродных аминокислот и производных во время пептидного синтеза включают, но не ограничиваются ими, использование норплейцина, 4-амино-масляной кислоты, 4-амино-3-гидрокси-5-фенилпентановой кислоты, 6-аминогексановой кислоты, т-бутилглицина, норвалина, фенилглицина, орнитина, саркозина, 4-амино-3-гидрокси-6-метилгептановой кислоты, 2-тиенилаланина и/или D-изомеров аминокислот.
2. Иммуноисследования.
Белок наружной мембраны D15, аналог, фрагмент и/или пептиды данного изобретения применимы в качестве антигенов в иммуноисследованиях, включая твердофазный иммуноферментный анализ (ELISA), РИД и другие исследования или процедуры связывания антител без ферментов, известные в этой области науки, для определения антибактериальных, к Haemophilus D15 и/или пептиду антител. В исследовании ELISA белок наружной мембраны D15, его фрагмент или аналоги и/или пептиды, соответствующие частям белка наружной мембраны D15, иммобилизованы на выбранной поверхности, например, на поверхности, проявляющей аффинитет к белку, такой как ячейки полистирольной микротитровальной платы. После отмывания для удаления неполностью адсорбировавшегося белка наружной мембраны D15, аналога фрагмента и/или пептидов с избранной поверхностью может быть связан неспецифический белок, такой как бычий сывороточный альбумин (БСД) или казеин, о котором известно, что он является антигенно нейтральным в отношении испытуемого образца. Это позволяет блокировать сайты неспецифической адсорбции на иммобилизирующей поверхности и таким образом снижает фон, вызванный неспецифическим связыванием антисыворотки с поверхностью. Обычно, пептиды, используемые здесь, находятся в пределах 12 остатков и выше, и предпочтительны, от 14 до 30 остатков.
Иммобилизирующая поверхность затем приводится в контакт с образцом, таким как клинический или биологический материал, который нужно проверить способом, приводящим к образованию иммунного комплекса (антиген/антитело). Это может включать разведение образца разбавителями, такими как БСА, бычий гамма-глобулин (БГГ) и/или фосфатно-буферный физраствор (ФБР)/Твин. Образец затем подвергают инкубации в течение от 2 до 4 часов при температурах, таких как порядка от 25 до 37oC. После инкубации поверхность, проконтактировавшая с образцом, отмывается для удаления не образовавшего иммунокомплексы материала. Процедура отмывания может включать промывание раствором, таким как ФБР/Твин или боратный буфер.
После образования специфических иммунокомплексов между испытуемым образцом и связанным белком наружной мембраны, аналогом, фрагментом и/или пептидами и последующего отмывания наличие и даже количество образования иммунокомплекса может быть определено путем контакта иммунокомплекса со вторым антителом, имеющим специфичность к первому антителу. Если испытуемый образец может иметь человеческое происхождение, вторым антителом является антитело, имеющее специфичность к человеческому иммуноглобулину, и в основном к IgG. Для обеспечения средства обнаружения второе антитело может обладать связанной с ним активностью, такой как ферментативная активность, которая будет производить, например, развитие окрашивания при инкубации с соответствующим хромогенным субстратом. Количественная оценка может затем достигаться путем измерения степени развития окрашивания с использованием, например, спектрофотомера для видимого спектра.
3. Использование последовательностей в качестве проб для гибридизации.
Нуклеотидные последовательности данного изобретения, содержащие последовательность белка наружной мембраны, в настоящее время позволяет идентифицировать и клонировать гены белка наружной мембраны D15 из любых видов Haemophilus и других бактерий, которые имеют гены, кодирующие белки наружной мембраны D15.
Нуклеотидные последовательности из данного изобретения, содержащие последовательность, кодирующую белок наружной мембраны D15, полезны благодаря их способности селективно образовывать дуплексные молекулы с комплементарными отрезками других генов D15. В зависимости от применения, может использоваться ряд условий гибридизации для достижения различных степеней селективности пробы в отношении других генов D15. Для высокой степени селективности используются строгие условия для образования дуплексов, такие как низкая концентрация соли /и/или высокая температура, такие, которые обеспечиваются 0,02 М до 0,15 М NaCl при температурах в интервале между примерно 50oC и 70oC. При некоторых применениях необходимы менее строгие условия гибридизации, такие как 0,15-0,9 М раствор соли при температурах в интервале от примерно 20oC до 55oC. Условия гибридизации можно также превращать в более строгие путем добавления повышающихся количеств формамида для дестабилизации гибридного дуплекса. Таким образом, конкретными условиями гибридизации можно легко управлять, и они, в основном, будут способом выбора в зависимости от желаемых результатов.
При осуществлении клинической диагностики последовательности нуклеиновых кислот генов белка наружной мембраны D15 из данного изобретения могут использоваться в сочетании с соответствующими средствами, такими как метка, для определения гибридизации. Широкий ряд соответствующих индикаторных средств известен с этой области, включая радиоактивные, ферментные или другие лиганды, такие как авидин/биотин, которые способны обеспечить обнаруживаемый сигнал. При некоторых осуществлениях диагностики может использоваться ферментная метка, такая как уреаза, щелочная фосфатаза или пероксидаза, вместо радиоактивной метки. В случае ферментных меток, известны колориметрические индикаторные субстраты, которые могут использоваться, чтобы обеспечить средства, видимые для человеческого глаза, или регистрируемые спектрофотометрические для идентификации специфической гибридизации с образцами, содержащими последовательности гена D15.
Последовательности нуклеиновой кислоты генов D15 данного изобретения применимы в качестве проб для гибридизации при гибридизации в растворе и при осуществлении с применением твердофазных процессов. При осуществлениях, включающих твердофазные процессы, испытуемая ДНК (или РНК) из образцов, таких как клинические образцы, включая экссудаты, жидкости организма (например, сыворотка, аминиотическая жидкость, выделенная из среднего уха, мокрота, бронхоальвеолярная промывная жидкость) или даже ткани, адсорбируется или иначе прикрепляется к выбранной матрице или поверхности. Фиксированная однонитевая нуклеиновая кислота затем подвергается специфической гибридизации с выбранными пробами, содержащими последовательности нуклеиновой кислоты генов D15 или их фрагментов данного изобретения при желаемых условиях. Выбираемые условия будут зависеть от конкретных обстоятельств, обоснованных конкретными критериями, необходимыми в зависимости, например, от содержания G + C, типа мишеневой нуклеиновой кислоты, источника нуклеиновой кислоты, размера пробы для гибридизации и т.д. После отмывания поверхности гибридизации таким образом, чтобы удалить не специфически связанные молекулы пробы, специфическая гибридизация обнаруживается или даже определяется количественно с помощью метки. Выбираемая проба должна быть по крайней мере в 18 bp и может находиться в интервале от 30 bp до 90 bp длиной.
4. Экспрессия генов белка наружной мембраны D15.
Для экспрессии генов белка наружной мембраны в системах экспрессии могут использоваться плазмидные векторы, содержащие репликон и управляющие последовательности, которые происходят из видов, сходных с клеткой-хозяином. Вектор обычно несет сайт репликации, а также маркирующие последовательности, которые способны обеспечить фенотипический отбор трансформированных клеток. Например Е.coli может быть трансформирована с использованием pBR322, которая содержит гены устойчивости к ампициллину и тетрациклину, и таким образом обеспечивается средство для легкой идентификации трансформированных клеток. Плазмида рBP322 или другие микробные плазмиды или фаг должны также содержать или модифицироваться так, чтобы содержать промоторы, которые могут использоваться микроорганизмом для экспрессии его собственных белков.
Кроме того, фаговые векторы, содержащие репликон и управляющие последовательности, которые совместимы с микроорганизмом-хозяином, могут использоваться в качестве трансформирующего вектора в соединении с этими хозяевами. Например, фаг лямбда CEMTM-N может использоваться при получении рекомбинантных фаговых векторов, которые могут использоваться для трансформации клеток-хозяев, таких как E.coli LE392.
Проморы, обычно используемые при конструировании рекомбинантных ДНК, включают беталактамазные (пенициллиназные) и лактозные промоторные системы и другие микробные промоторы, такие как Т7 промоторная система. Детали, относящиеся к нуклеотидным последовательностям промоторов, известны, давая возможность опытному специалисту функционально лигировать их в плазмидные векторы. Конкретный используемый промотор является делом выбора, зависящего от желаемых результатов. Хозяева, которые подходят для экспресси генов трансферриновых рецепторов, их фрагментов, аналогов или вариантов, включают E.coli, Bacillus, Haemophilus, Bordetella, грибы, дрожжи или бакуловирусные и поксвирусные системы экспресси, которые могут использоваться.
В соответствии с особенностью этого изобретения предпочтительно получать белок наружной мембраны D15, его фрагмент или аналог с помощью методов генной инженерии, в частности, потому что природно встречающийся белок D15, который очищен от культуры видов Haemophilus, может включать нежелательные примеси, включая следовые количества токсических веществ. Эту проблему можно устранить путем использования рекомбинантно полученного белка наружной мембраны в гетерологичных системах, который можно выделять из клеток-хозяев таким образом, чтобы свести к минимуму количество токсинов в очищенном материале. Конкретно, желательные в этом отношении хозяева включают грамположительные бактерии, у которых нет липополисахаридов (ЛПС) и поэтому у них нет эндотоксинов. Такие хозяева включают виды Bacillus и могут, в частности, быть полезны для продукции непирогенного белка наружной мембраны D15, его фрагментов или аналогов.
Помещенные на хранение биологические образцы
Некоторые плазмиды, которые содержат по крайней мере часть, кодирующую белок наружной мембраны D15, из штаммов Haemophilus influenzae, которые описаны здесь и названы здесь, были помещены на хранение в Американскую коллекцию типовых культур (American Tepte Cul Luve Collection- ATCC), находящуюся в Rockyille, Navyland, США, следуя Будапештскому договору и до регистрации этой заявки. Образцы депонированных плазмид станут доступны общественности после выдачи патента на основании заявки на патент Соединенных Штатов. Изобретение, описанное и заявленное здесь, не должно ограничиваться в объеме помещенными на хранение плазмидами, так как помещенное на хранение осуществление предназначено только для иллюстрации изобретения. Любые эквивалентные или аналогичные плазмиды, которые кодируют аналогичные или эквивалентные антигены, которые описаны в этой заявке, находятся в сфере действия этого изобретения.
Депонированные клоны представлены в табл. A в конце описания.
Вышеприведенное раскрытие в основном описывает данное изобретение. Более полное понимание может быть достигнуто путем обращения к следующим конкретным примерам. Эти примеры изложены исключительно в целях иллюстрации и не предназначены для ограничения объема изобретения. Хотя здесь были использованы конкретные значения, такие значения предназначены для описательных целей, но не для целей ограничения. Иммунологические и генно-инженерные методы не могут быть подробно описаны в этом раскрытии, но достаточны в объеме для специалистов.
Примеры
Методы молекулярной генетики, белковой биохимии и иммунологии, использованные, но не описанные подробно в этом раскрытии, и эти ПРИМЕРЫ пространно изложены в научной литературе и вполне доступны для специалистов.
Пример 1
Этот пример иллюстрирует клонирование и секвенирование генов D15.
Геномную ДНК очищали из штамма Ca Haemophilus influenzae b типа путем лизиса бактерий проназой и додецилсульфатом натрия с последующей экстракцией фенолом и осаждением изопропанолом по Berns и Thomas, 1965. ДНК затем частично расщепляли с помощью E.coli и фракцию ДНК, содержащую фрагменты 6-10 kb, выделяли после электрофореза в агарозе с низкой точкой плавления. Эти фрагменты лигировали в лямбда gtII AmpI вектор (Thomas and Rossi 1986) и клонировали как лизоген в штамм BTA282 E.coli. Рекомбинантные клоны отбирали по их устойчивости к ампициллину, придаваемую вектором. Для идентификации клонов, продуцирующих антиген H.influenzae типа b, клоны с помощью репликатора наносили на нитроцеллюлозные фильтры и удвоенные колонии индуцировали к экспрессии путем температурного переключения до температуры 42oC в течение 2 часов. Колонии лизировали путем смачивания фильтров 1% раствором додецилсульфата натрия (ДСН). Фильтры затем помещали в атмосферу, насыщенную хлороформом, на 15 минут. Фильтры затем анализировались с помощью радиоиммуноисследования колоний с использованием гипериммунной кроличьей антисыворотки против H.influenzae типа b, абсорбированной с помощью лизата E.coli, для экспрессии антигена. Клоны, выявленные с помощью ауторадиографии, которые продуцируют антигены H.influenzae типа b, дополнительно очищали и их реплики снова испытывали на реактивность с гипериммунной антисывороткой против H. influenzae типа b. Антисыворотку, абсорбированную 1010 интактных бактерий H. influenzae типа b (штамм Ca), использовали в качестве отрицательного контроля.
Был идентифицирован ряд клонов, которые реагировали с неабсорбированной, но не реагировали с абсорбированной антисывороткой, и с ними проводили дальнейшие анализы. Один из клонов D15 очищали, подращивали, и, как было обнаружено, он продуцировал антиген H.influenzae типа b, который мигрировал в полиакриламидном геле с додецилсульфатом натрия с Mr, равным примерно 80 кDa. Лизаты из клона D15 соединяли с сефарозТМ-ным 4B гелем и использовали для очистки с помощью аффинной хроматографии анти-D15 антител. Эта процедура описана Thomas et al. 1990, за исключением того, что первоначально сообщалось, что Мr примерно равен 103 кDа. Антитела rD15, очищенные с помощью аффинной хроматографии, как было затем показано, реагировали с белком с Мr 80 кDa в препарате белков наружной мембраны H.influenzae типа b (фракция, нерастворимая в саркозиле-Cavtone et al. 1986). Анализы мембранных препаратов с помощью радиоиммунодот-блоттинга и Вестерн-блоттинга как из Haemophilus influenzae типа b, так и из нетипируемых штаммов показал, что анти- D15 антитела, аффинно-очищенные реагировали со всеми штаммами. Эти антитела, как было обнаружено, способны служить пассивной защитой у крысят от бактерии после внутрибрюшинной инъекции живых бактерий H.influenzae типа b. Специфичность этой защиты была подтверждена путем абсорбирования с устранением защитной активности анти- D15 антител с помощью диализата E.coli, экспрессирующей D15, соединенного с сефарозой. Изучение защитного действия было описано детально Thomas et al. 1990.
Выделяли ДНК из фага лямбда gtII Amp I D15 и фрагмент в 5,7 кb изолировали путем расщепления Е. coli. Этот фрагмент субклонировали в pUC19, и полученную в результате плазмиду трансформировали в E.coli HB101. Рекомбинантные бактерии, как было обнаружено, продуцировали ожидаемый антиген H.influenzae типа b c Mr 80 кDa по исследованию с помощью Вестерн-блоттинга. Затем определяли характеристики вставочной ДНК с помощью рестрикционного эндонуклеазного картирования. Фрагмент Hind III - E.coli 2,8 кb субклонировали в pUC19 для получения плазмиды pUCI9/D15, которую трансформировали в E.coli HB101. Рекомбинантные бактерии экспрессировали белок Mr 80 кDa, распознаваемый D15- специфическими антителами при вестерн-блоттинговом анализе лизатов E.coli.
Плазмидную ДНК получали из двух отдельных колоний рекомбинантных E.coli HB101, содержащих плазмиду pUC19/D15, с использованием стандартных методик. Олигонуклеотидные секвенируюцие праймеры длиной 17-25 оснований синтезировали на синтезаторе ДНК ABI модель 380В и очищали и помощью хроматографии с использованием картриджей ОРС, полученных от Applied Biosystem Ibn и примененных в соответствии с рекомендациями производителя. Образцы секвенировали, используя секвенатор ДНК ABI модель 370А и окрашенные терминаторные реакции по указаниям производителя. Анализ этой последовательности показал, что ген D15 содержит открытую рамку считывания, кодирующую 789 аминокислот, включая предполагаемую сигнальную последовательность (фигура 1). Производная аминокислотная последовательность, как было обнаружено, содержит последовательность внутреннего пептида, полученного путем тромбинового расщепления природного D15, который был определен химически. Аминокислотное строение D15, выведенное из последовательности гена D15, была сравнима (в пределах экспериментальной ошибки) с последовательностью природного белка, как определено с помощью анализа аминокислот.
Пример 2
Этот пример иллюстрирует получение хромосомной ДНК из штаммов Haemophilus influenzae Eagan, Minn A, SB33 и PAK 12085.
Штаммы H. influenzae подращивали на агаре Мюллера-Хинтона или в бульоне на экстракте из сердца и мозга, как описано Harkness et.al. 1992 г.
Xромосомная ДНК штамма Eagan
Бактерии из 50 мл культуры осаждали центрифугированием при 5000 об/мин течение 20 минут при 4oC. Осадок снова суспендировали в 25 мл ТЭ (10 мМ трис, 1 мМ ЭДТУ, pH 8,0) и два образца по 5 мл использовали для получения хромосомной ДНК. К каждому образцу добавляли 0,6 мл 10% каркозила и 0,15 мл 20 мг/мл протеиназы К и образцы инкубировали при 37oC в течение 1 часа. Лизат экстрагировали один раз Трис- насыщенным фенолом (pH 8,0) и три раза хлороформом : изоамиловым спиртом (24:1). Водную фазу объединяли до конечного объема, равного 7 мл. Затем, 0,7 мл 3М ацетата натрия (pH 5,2) и 4,3 мл изопропанола добавляли для осаждения ДНК, которую собирали, промывали 70% этанолом, сушили и снова суспендировали в 1 мл воды.
Хромосомная ДНК штаммов Minna, SB 33 и PAK 12085
Бактерии из 50 мл культуры осаждали центрифугированием при 5000 об/мин в течение 15-20 минут при 40oC в центрифуге Sorvall RC-3B. Клеточный осадок снова суспендировали в 10 мл ТЭ (10 мМ Трис-HCl, 1 мМ ЭДТУ, pH 7,5), добавляли проназу до 500 мкг/мл и ДСН до 1%. Образец инкубировали при 37oC в течение примерно 4 часов до получения прозрачного лизата. Лизат один раз экстрагировали насыщенным Трис фенолом, один раз насыщенным Трис фенолом/хлороформом (1: 1) и один раз хлороформом. Конечную водную фазу диализировали в течение 24 часов против 2 • 500 мл 1 М NaCl при 4oC, меняя буфер один раз и в течение 24 часов против 2 • 500 мл ТЭ при 4oC, меняя буфер один раз. Конечный диализат делили на одинаковые образцы последующего использования.
Пример 3
Этот пример иллюстрирует получение хромосомных библиотек Haemophilus influenzae
Хромосомные ДНК H.influenzae Eagan и PAK 12085 расщепляли с помощью San 3A 1 (0,5 ед/10 мкг ДНК) при 37oC в течение 15 минут и фракционировали по размеру с помощью электрофореза в агарозном геле. Слои геля, соответствующие фрагментам ДНК, равным 15-23 кв., вырезали и ДНК электроэлюировали в течение ночи в диализной трубке, содержащей 3 мл ТАЭ (40 мМ Трис- ацетата, 1 мМ ЭДТУ, pH 8,0) при 14V. ДНК осаждали дважды и повторно суспендировали в воде перед лигированием в течение ночи с плечами EMBL 3 BamHI (Promega Лигированную смесь упаковывали, используя набор Лямбда для упаковки in vitro (American), в соответствии с инструкциями производителя и высевали на чашку, засеянную E. coli NH 539 клетками. Библиотеку титровали, затем амплифицировали и хранили при 4oC под 0,3% хлороформом.
Хромосомную ДНК Minna A (10 мкг) расщепляли с помощью San 3A I (40 единиц) в течение 2,4 и 6 минут, затем фракционировали по размеру на градиенте сахарозы 10-30% в ТНЭ (20 мМ Трис-HCl), 5 мМ NaCl, 1 мМ ЭДТУ, pH 8,0). Фракции, содержащие фрагменты ДНК > 5 кb, объединяли и осаждали. Во втором эксперименте хромосомную ДНК (2,6 мкг) расщепляли с помощью San 3A I (4 единицы) в течение 1, 2 и 3 минут и фракционировали по размеру с помощью препаративного электрофореза в агарозном геле. Слои геля, содержащие ДНК фрагменты, равные 10-20 кb, вырезали и ДНК экстрагировали с помощью стандартной методики замораживания и оттаивания. Фракцонированную по размеру ДНК из двух экспериментов объединяли для лигирования с плечами Ban HI EMBL3 (Promega). Лигированную смесь упаковывали, используя набор для упаковки Gigapack II (Amersham) и высевали на чашку с клетками E.coli LE 392. Библиотеку титровали, затем амплифицировали и хранили при 4oC под 0,3% хлороформом.
Хромосомную ДНК SB 33 (20 мкг) расщепляли с помощью San 3A I (40 единиц) в течение 2,4 или 6 минут и фракционировали по размеру на 10-30% градиенте сахарозы в ТНЭ (20 мМ Трис-HCl), 5 мМ NaCl, 1 мМ ЭДТУ, pH 8,0). Фракции, содержащие фрагменты > 5 кb объединяли. Во втором эксперименте хромосомную ДНК SB 33 (2 мкг) расщепляли San 3A I (4 единицы) в течение 2,4 или 6 минут и фракционировали по размеру в препаративном агарозном геле. Слои геля, содержащие фрагменты ДНК, равные 10-20 кb, вырезали и ДНК экстрагировали с помощью стандартной методики замораживания/оттаивания. Фракционированная с плечами BamH I EMBL3 (Promega). Лигированную смесь упаковывали, используя набор для упаковки Gigapack II и высевали на чаши с клетками LE 392. Библиотеку титровали, затем амплифицировали и хранили при 4oC под 0,3% хлороформом.
Пример 4
Этот пример иллюстрирует проверку библиотек ДНК.
Библиотеки ДНК Eagan, Minn SB 33 и РАК 12085 высевали на газон клеток PE 392 на чашках NZCYM, используя 0,7% легкую агарозу в NZCYM в качестве покрытия. Снятие бляшек на нитроцеллюлозные фильтры выполняли по стандартным процедурам, фильтры обрабатывали и гибридизовали с пробой D15, меченной дигоксигенином, получаемой по описаниям производителя (Boeringer Mannheim). Проба была фрагментом EcoR I/Hind III из pUC19/D15, содержащей весь ген Ca D15 (фигура 2). Предполагаемые бляшки культивировали и подвергали второму циклу отбора, используя те же самые процедуры. ДНК фага получали из 500 мл культуры, используя стандартные методики, вставочную ДНК вырезали путем расщепления SalI и клонировали в pUC19 для получения клонов DS-712-2-1 (Eagan), DS-691-1-5 (Minn A), JB-1042-5-1 (SB 33) и JB-1042-9-4 (РАК 12085), которые показаны на фигуре 2.
Определяли нуклеотидные последовательности генов 15 из H.influenzae типа b штаммов Eagan и Minn A, нетипируемых штаммов H.influenzae 33 и РАК 12085 и сравнивали с последовательностью для штамма Ca, что видно на фигурах 1B, 1C, 1D, 1E и 1F. Желаемые аминокислотные последовательности показаны на фигурах
1B, 1C, 1D и IE и они сравниваются с аминокислотной последовательностью белка D15 H.influenzae типа b Ca (фигура 3)
Пример 5
Этот пример иллюстрирует экспрессию белка D15 в E.coli
Фрагмент Himd llll - EcoRI в 2,8 кb субклонировали в pUC19 в плазмиду pUC19/D15 трансформировали в Е.coli HB101. При индуцировании положительные клоны экспрессировали белок в 80 кDa, который распознавался D15 - специфической антисывороткой при вестерн-блотанализе. Фрагмент Hind III - Pst I также субклонировали в pUC19, и как показано, экспрессировал белок в 67 кDa. По рестрикционной карте этот белок в 67 кDa соответствовал усеченному с C-конца белку D15. При вестерн-блотанализе - этот усеченный D15 еще распознавался D15 специфической антисывороткой.
Плазмиды для экспрессии гена D15 из нетипируемого штамма SB З3 в Е.coli были сконструированы. Плазмида JB-1042-5-1, содержащая ген D15 и SB 33, и ограничивающие его области расщепляли EcoRI и Hind III и вставку D15 в 3 кb субклонировали в pUC для получения плазмиды pRY-60-1 (фигура 4). Были синтезированы соответствующие олигонуклеотиды для сохранения природной последовательности D15 между кодоном ATG плазмиды экспрессии рТ7-7 и сайтом BsrF-1 в гене D15. Эти олигонуклеотиды имели следующую последовательность:
Nde
5'-TATGGCACCTTTTGTGGCAAAAGATATTCGTGTGGATGGTGTTCAAGGTG ACCGTGGAAAACACCGTTTTCTATAAGCACACCTACCACBAGTTCCACTGAATCT ACTTAGAATCAACAAACCGAGCAAGTTTACCTGTTCGTG - SEQ ID NO:50 TGGTTGTTTAGGCTCGTTCAAATGGACAAGCACGGCC-5'- SEQ ID NO:51
BsrF I
Плазмиду рRY-60-1 расщепляли с помощью EcoRI и РsrF I и очищали фрагмент ДНК, содержащий большую часть гена D15. pUC расщепляли с помощью EcoR I и Nde I и очищали векторный фрагмент. Поликомпонентное лигирование между pUC и фрагментами D15 и олигонуклеотидами давало плазмиду DS-860-1-1, которая содержит последовательность D15 без промотора. рТ7-7 расщепляли с помощью Nde I и EcoR I и очищали векторный фрагмент. DS-860-1-1- расщепляли с помощью Nde I и EcoR I и вставку D15 очищали и лигировали с Т7-7 вектором, получая плазмиду DS-880-1-2- (фигура 4).
Плазмидные конструкции реализовали, используя E.coli JM 109 в качестве хозяина. Для экспрессии плазмиду DS-880-1-2- трансформировали в E.coli BL 21/DE 3, BL 21/DE 3/pLysS или клетки JM 109/DE3. Трансформацию клеток выполняли, используя или обработанные хлоридом кальция компетентные клетки или путем электропорации с использованием электропоратора BioRad. Трансформированные клетки выращивали в средах YT, M9 или NZCYM и индуцировали IPTG или другими индуцирующими средствами.
Пример 6
Этот пример иллюстрирует конструирование и экспрессию фрагмента GST-D15 гибридного гена в E.coli.
Использовали смысловой праймер в прямом направлении (пример 1) 5'-GGGGAATTCCAAAAGATCTTCGT (ПОСЛ ИД N:52) и обратный антисмысловой праймер CACGAATTCCCTGCAAATC-5'(праймер 7 - ПОСЛ ИД N:53) для амплификации фрагмента Hind lII-EcoR I в 2,8 кb из гена D15 с помощью полимеразной цепной реакции, который кодирует N-концевые аминокислотные остатки с 22 по 223 первичной последовательности белка D15 (фигура 1A). Нуклеотидная последовательность амплифицированного фрагмента в 609 bp подтверждалась секвенированием ДНК. Амплифицированный сегмент гена лигировали в вектор pGEX-2T в прямом направлении от гена GST и трансформировали в E.coli TG-1. Колонии, экспрессирующие антиген H. influenzae типа b, отбирали с помощью кроличьей антисыворотки к H. influenzae типа b путем радиоиммуноанализа колоний и изолировали. Слитый белок глютатион-s-трансфераза-фрагмент-D15, продуцируемый трансформированной Е.coli, выделяли с помощью аффинной очистки на глютатион-агарозе.
Пример 7
В этом примере описаны альтернативные системы экспрессии для rD15.
Ген D15 или его фрагменты также экспрессируются в E.coli под контролем других регуляторных промоторов. Ген D15 или его фрагменты экспрессируются в отсутствие лидерного пептида или в других системах, где токсичность экспрессии D15 для хозяина не является проблемой. Ген или его фрагменты синтезируются снова или путем применения полимеразной цепной реакции с использованием подходящих праймеров. Эти гены клонируют в подходящие клонирующие векторы или бактериофаговые векторы в E.coli или другие подходящие хозяева, непосредственно, если токсичности можно избежать. Системами экспрессии являются грамположительные бактерии (такие, как Bacillus species) поксвирус, аденовирус, бакуловирус, дрожжи, грибы, БЦЖ или системы экспрессии млекопитающих.
Пример 8
Этот пример иллюстрирует процедуру экстракции и очистки D15 из cистемы экспрессии E.coli.
Осадок клеток из 250 мл культуры, полученной как описано в примере 5, снова суспендировали в 40 мл 50 мМ Трис, pH 8,0 и разрушали ультразвуковым облучением (3 • 10 мин, диск 70% нагрузки). Экстракт центрифугировали при 20000 х g и полученный в результате осадок сохраняли. Первоначальный осадок снова экстрагировали 40 мл 50 мМ Трис, 0,5% Тритон-X-100, 10 мМ ЭДТУ, pH 8,0. Суспензию затем облучали ультразвуком в течение 10 минут при диске 70% нагрузки. Экстракт центрифугировали при 300 х g в течение 5 минут. Полученный в результате супернатант центрифугировали снова при 20000 х g в течение 30 минут и полученный в результате осадок сохраняли. Осадок снова суспендировали в 50 мМ Трис, 0,5% Тритон, X-100, 10 мМ ЭДТУ, pH 8,0. Суспензию затем перемешивала с ФБР/8M мочевиной до конечной концентрации мочевины, равной 6М. Раствор затем диализировали против ФБР для удаления мочевины. После диализа раствор центрифугировали при 300 х g в течение 10 мин, супернатант оставляли и хранили при 4oC.
Пример 9
Этот пример демонстрирует очистку -GST (фрагмент D15) слитого белка с использованием аффинной хроматографии на глютатион-сефарозе 4В.
Пять мг сырого экстракта слитого белка GST-(фрагмент D15), полученные, как описано в примере 6, растворяли в 5 мл фосфатно-буферного физраствора (ФБР), содержащего 1% Тритон-X-100. Раствор затем загружали на колонку глютатион-сефарозы 4В (2 мл), уравновешенную ФБР, содержащим 1% Тритон X-100. Погон, прошедший через колонку, отбрасывали. Колонку промывали 20 мл ФБР и белок слияния GST-(фрагмент D15) элюировали 50 мМ Трис-HCl буфером, pH 8,0, содержащим 5 мМ глютатиона. Элюирование контролировали по поглощению при 280 нм. Содержащие белок фракции (2 мл/фракцию) собирали и объединяли. Чистоту белка оценивали с помощью электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE)(фигура 9, дорожа 3).
Конечный объем очищенного слитого белка составил 6 мл.
Пример 10
Этот пример иллюстрирует процедуру, использованную для тромбинового расщепления белков, с целью выделения усеченной молекулы D15.
Образец слитого белка GST-(фрагмент D15) из примера 9 (0,1 - 0,5 мг белка/мл) подвергали диализу против 1 л 50 мМ Трис-HCl буфера (pH 8,5) 3 раза с по крайней мере 2-часовыми интервалами при 4oC для удаления ингибитора протеазы. После диализа раствор обрабатывали человеческим тромбином (Sigma) при соотношении 1 мл раствора на 25 единиц тромбина. Реакцию расщепления проводили при 37oC в течение 2 часов и анализировали с помощью SDS-PAGE (фигура 9, дорожка 4). Реакцию останавливали путем помещения раствора в лед.
Пример 11
Этот пример иллюстрирует процедуру, использованную для очистки N-концевого фрагмента rD15 из GST с использованием аффинной хроматографии на глютатион-сефарозе 4В.
Расщепленный тромбином образец GST-(фрагмент-D15), полученный, как описано в примере 10, загружали на колонку глютатион-сефарозы 4В (2 мл), уравновешенную ФБР, содержащим 1% Тритона X-100. Погон через колонку, содержащий N-концевой фрагмент rD15, сохраняли. После промывания колонки 20 мл ФБР, аффинную колонку регенерировали путем удаления GST с применением 50 мМ Трис-HCl буфера, pH 8,0, содержащего 5 мМ глютатион.
Чистоту фрагмента rD15 проверяли путем анализа SDS- PAGE (фигура 9, дорожка 5). Этот N-концевой фрагмент rD15 содержит аминокислоты 63-223 из белка D15 в результате расщепления в специфическом тромбиновом сайте, показанном на фигуре 1A.
Пример 12
Этот пример иллюстрирует процесс, использованный для очистки D15-специфических поликлональных антител путем аффинной хроматографии с использованием белка слияния GST-(фрагмент D15).
Рекомбинантный белок слияния - (фрагмент D15), полученный как описано в примере 9, конъюгировали с сефарозой, активированной цианогенбромидом. Аффинную колонку затем использовали для очистки антител из кроличьей гипериммунной антисыворотки против Н.influenzae типа b. Антитела, очищенные с помощью аффинной хроматографии, как было показано путем иммуноблоттинга, реагируют с компонентом в 80 кDа, присутствующим в лизатах E.coli трансформированных pUC/D15 и в лизатах нескольких типируемых и нетипируемых штаммов Н. influenzae. Эти результаты подтвердили, что сегмент ДНК, кодирующий фрагмент D15 слитого белка, был частью открытой рамки считывания гена D15.
Подобным же образом антисыворотки, полученные к рекомбинантному белку слияния (пример 9) или очищенному N-концевому фрагменту rD15 (пример 11), реагировали с белком rD15, продуцируемом штаммами H.influenzae (пример 13).
Пример 13
В этом примере описан процесс, использованный для очистки природного rD15 из H.influenzae. Клеточную массу нетипируемого штамма SB 33 H.influenzae, полученную из культуры, выращенной в среде на экстракте мозга, сердца, дополненной НАД (2 мкг/мл) и гемином (2 мкг/мл) при 37oC, как описано Panezutti et. al 1993, снова суспендировали в 50 мМ Трис-HCl, pH 8,0, содержащем 0,5% Тритон Х-100 и 10 мМ ЭДТУ (20 мл на 1 г клеточной массы). Смесь перемешивали при комнатной температуре в течение 2 часов, затем центрифугировали при 20000 х g в течение 30 минут. D15 находился в супернатанте и далее очищался.
Очистку природного D15 проводили путем аффинной хроматографии с использованием D15-специфических моноклональных антител (смотрите пример 24). Экстракт D15 (25 мл) смешивали с аффинной матрицей (1 мл) при комнатной температуре в течение 2 часов. Смесью набивали колонку и протекшие фракции отбрасывали. Колонку промывали последовательно следующими буферами: 50 мМ Трис-HCl, pH 8,0, содержащим 0,5% Тритона Х-100 и 10 мМ ЭДТУ, 1M буфером HEPES, pH 6,8, 50 мМ Трис-HCl, pH 8,0, содержащим 0,5% Тритон X-100 и 10 мМ ЭДТУ, и 10 мМ фосфатным буфером, pH 8,0, D15 затем элюировали c колонки 3 мл 50 мМ диэтиламина, pH 12,0, и раствор белка нейтрализовали 1 M HEPES, pH 6,8 (1/10 объема). Очищенный аффинной хроматографией природный D15 анализировали с помощью SDS-PAGE и хранили при - 20oC.
Пример 14
В этом примере описан процесс, использованный для получения конъюгатов D15-PRP.
Олигосахариды Haemophilus influenzae типа b (PRP), полученные путем контролируемого кислотного гидролиза, конъюгировали или с очищенным природным (пример 13) или рекомбинантным D15 (пример 8), а также с его фрагментами (пример 11), используя перииодатное окисление, как описано в патенте США 4356170 и дополнительные детали которого представлены в примере 17. Средний размер молекул PRP, использованного для конъюгации, как было определено, составлял примерно 20000 Дальтон. Конъюгацию проводили без молекулы линкера, но она может также проводиться и с молекулой линкера. Молярное соотношение PRP/15, равное примерно 7, использовали для создания избытка PRP гаптена.
Конъюгат PRP/rD15 испытывали по схеме из примера 18 на иммуногенность у кроликов и он вызывал образование как первичных, так и вторичных анти- PRP IgG и анти-D15 антительных ответов (Таблица 9). Кроличьи анти- rD15-PRP антисыворотки также сильно реагировали как с природным D15, так и с rD15, судя по анализу иммуноблота. Эти данные показывают, что rD15 могут использоваться в качестве белка-носителя в конъюгатной вакцине. Кроме того, rD15-PRP конъюгатная вакцина должна гарантировать более устойчивую защиту против заболеваний, вызываемых H.influenzae типа b, в частности, у детей, в результате дополнительной гомотипической защиты, обеcпечиваемой антителами, направленными против белка rD15.
Пример 15
В этом примере описывается получение пептидов D15.
Пептиды D15 (Таблица 2) были синтезированы с использованием синтезатора пептидов ABI 430A и оптимизированных химических процессов с т-BOC, как описано производителем, затем отделяли от смолы с помощью гидрофтористой кислоты (HF). Пептиды очищали с помощью высокоэффективной жидкостной хроматографии с обращенной фазой (RP-HPLC) на полупрепаративной колонке 1 х 30 см) Vydac C4, используя градиент ацетонитрила с 15 до 55% в 0,1% трифторуксусной кислоте (ТФУ), происходящей в течение 40 минут при скорости потока 2 мл/мин. Все синтетические пептиды (таблица 2), использованные для биохимического и иммунологического изучения, обладали чистотой > 95%, судя по аналитической ВЭЖХ. Анализы аналогичного состава этих пептидов выполняли на системе Waters Pico-Tag и они были в хорошем согласии с их теоретическим составом.
Пример 16
В этом примере описан процесс, использованный для получения специфичных антисывороток к D15 пептиду.
Морских свинок и кроликов иммунизировали отдельными пептидами (50-200 мкг), эмульгированными в полном адъюванте Фрейнда и вводимыми внутримышечно. После двух повторных введений того же самого количества пептида в неполном адъюванте Фрейнда через 14 и 28 дней антипептидую антисыворотку собирали на 42 день и испытывали с помощью ELISA и иммуноблоттинга. Как кроличьи, так и морских свинок антисыворотки, как было показано, являются моноспецифическими для соответствующих им иммунизирующих пептидов по пептид-специфическому ELIS A (Таблица 6). Кроме того, как морских свинок, так и кроличья антисыворотки, выработанные к пептидам D15, реагировали с как H. influenzae типа b, так и с D15 нетипирумых штаммов по иммуноблотанализам. Так как большинство пептидов D15 индуцировали сильный антипептидный антительный ответ по крайней мере у одного вида животных, они являются подходящими иммуногенами для включения в иммуногенные композиции, включая вакцинные препараты.
Пример 17
В этом примере описан процесс, использованный для получения конъюгатов PRP-BSA.
0,5 мл перйодат-окисленного PRP (25 мл в 1 мл 0,1 М натрий-фосфатного буфера, pH 6,0), полученного из природного PRP, обработанного водным раствором перйодной кислоты (Carlone et. al., 1986) добавляли к бычьему сывороточному альбумину (БСА) (1,32 мг, 0,02 мкмоль) в 0,5 мл 0,2 М натрий-фосфатного буфера pH 8,0 с последующим добавлением цианоборогидрида натрия (14 мкг, 0,22 мкмоль, 10 экв. к БСА). После инкубации при 37oC в течение 5 дней реакционную смесь диализировали против 4 л 0,1 М фосфатного буфера, pH 7,5. Полученный в результате раствор наносили на аналитическую колонку Superose 12 (15 х 300 мм Pharmacia), уравновешенную 0,2 М натрий-фосфатным буфером, pH 7,2 и элюировали тем же самым буфером. Фракции контролировали по поглощению при 230 нм. Первый основной белковый пик объединяли и концентрировали в Gentriprep 30 до 2,2 мл.
Количество белка определяли, используя исследование на белок bio Rad и было обнаружено, что оно равно 300 мкг/мл. Присутствие PRP во фракции белкового конъюгата подтверждалось с помощью орсинольного теста.
Пример 18
В этом примере описан процесс, использованный для продукции анти-PRP антисыворотки у животных с применением конъюгатов rD15-PRP
Кроликов иммунизировали внутримышечно конъюгатами rD15-PRP (пример 14 (5-50 мкг PRP) эквивалента), смешанных с 3 мг AlPO4 на мл, с последующими двумя повторными инъекциями (половины количества того же самого иммуногена) через двухнедельные интервалы. Антисыворотки отбирали через каждые 2 недели после первой инъекции, инактивировали нагреванием при 56oC в течение 30 минут и хранили при - 20oC.
Пример 19
Этот пример иллюстрирует реакционную способность пептидов D15 с антипептидной D15-специфическими антисыворотками при применении D15-специфических и пептид-специфических ELISA.
Микротитровальные ячейки (Nunc-Immunoplate, Nunс, Дания) покрывали 200 нг очищенного rD15 или 500 нг отдельных пептидов в 50 мкл буфера для покрытия (15 ММ Na2CO3, 35 мМ NaHCO3, pH 9,6) в течение 16 часов при комнатной температуре. Затем платы блокировали 0,1% (вес/объем) БСА в фосфатно-буферном физрастворе (ФБР) в течение 30 минут при комнатной температуре. В ячейки добавляли серийно разведенные антисыворотки и инкубировали в течение 1 часа при комнатной температуре. После удаления антисыворотки платы промывали пять раз ФБР, содержащим 0,1% (вес/объем) Твин-20 и 0,1% (вес/объем) БСА. Фрагменты F(ab')2 козьих против-кроличьих, морских свинок, мышиных или человеческих IgG антител, конъюгированных с пероксидазой хрена (Jackson ImmunoResearch Labs Inc), разводили (1/8000) отмывающим буфером и добавляли в микротитровальные платы. После 1 часа инкубации при комнатной температуре платы промывали пять раз смывающим буфером. Платы затем проявляли, используя в качестве субстрата тетраметилбензидин (ТМБ) в H2O2 (ADI, Торонто). Реакцию останавливали IN H2SO4 и оптическую плотность измеряли при 450 нм, используя Titretek Multiskan II (Flow Labs. Virginia). Два не имеющих отношения к делу пептида использовали в качестве отрицательных контролей при пептид-специфических ELISA. Исследования выполняли с троекратными повторами и реактивный титр каждой антисыворотки определяли как разведение, постоянно доказывающее 2-кратное значение поглощения свыше поглощения, получаемого для негативных контролей. Полученные результаты суммированы в таблицах 3, 6 и 8 и в подробном описании изобретения выше.
Пример 20
Этот пример иллюстрирует определение анти- PRP титров IgG в кроличьих антисыворотках к конъюгату PRP-D15 с использованием PRP- специфичного ELISA.
Ячейки для микротитрования (Дания) покрывали 200 нг очищенного PRP-БСА (смотрите пример 17) в 200 мкл покрывающего буфера (15 мМ Na2CO3, 35 мМ NaHCO3, pH 9,6) в течение 16 часов при комнатной температуре. Затем платы блокировали 0,1% (вес/объем) БСА в фосфатно-буферном физрастворе (ФБР) в течение 30 минут при комнатной температуре. Серийно разведенные кроличьи антисыворотки, полученные в конъюгатном PRP-D15, добавляли в ячейки и инкубировали в течение 1 часа при комнатной температуре. После удаления антисыворотки платы промывали пять раз ФБР, содержащим 0,1% (вес/объем) Твин-20 и 0,1% (вес/объем) БСА. Фрагмент F (ab')2 из козьих против кроличьего IgG антител, конъюгированных с пероксидазой хрена (Jackson ImmunoResearсh Labs Inc PA) разводили (1/8000) отмывающим буфером и добавляли в микротитровальные платы. После 1-часовой инкубации при комнатной температуре платы промывали пять раз отмывающим буфером. Затем платы проявляли, используя в качестве субстрата тетраметилбензидин (ТМБ) в H2O2 (ADI, Торонто). Реакцию останавливали I NH2SO4 и оптическую плотность определяли при 450 нм, используя Titretek Multiskan II (Flow Labs. Virginia). Стандартную анти-PRP антисыворотку с известным титром включали в качестве положительного контроля. Исследования проводили в троекратном повторении, и реактивный титр каждой антисыворотки определяли как величину, обратную разведению, постоянно показывающему 2-кратное повышение значения O.D. над значением, полученным с неиммунной сывороткой (таблица 9).
Пример 21
В этом примере описывается процесс, примененный для получения D15-специфических антисывороток при использовании очищенного D15, rD15 или N-концевого фрагмента rD15.
Новозеландских белых кроликов (линия MapIe) и морских свинок (Charles Rivex) иммунизировали внутримышечно (в/м) дозой в 10 мкг или очищенного аффинной хроматографией природного D15 (пример 13), рекомбинантного D15 (пример 8) или N-концевого фрагмента rD15 (пример 11), эмульгированного в полном адъюванте Фрейнда (Difco). Животным делали повторные инъекции антигена на 28 день еще одной 10 мкг дозы очищенного аффинной хроматографией D15 или rD15 или фрагмента rD15, эмульгированного в неполном адъюванте Фрейнда, и у них отбирали кровь на 42 день из краевой вены уха, D15 специфические поликлональные антитела очищали из этого материала, как описано в примере 12.
Пример 22
Этот пример иллюстрирует защитную активность введения D15-специфической антисыворотки против H.influenzae типа b при использовании модели бактеремии у крысят.
5-дневным крысятам подкожно (п/к) в тыльную сторону лапки вводили 0,15 мл двух различных кроличьих антисывороток против N-концевых фрагментов rD15. Неиммунные антисыворотки использовали в качестве отрицательных контролей. Через день после иммунизации крысятам внутрибрюшинно (в/б) вводили 200 колониеобразующих единиц(кое) Haemophilus influenzae типа b штамм Minn A (0,1 мл), свежевыращенной в среде на экстракте мозга, сердца (BHI), дополненной кофакторами и разведенной в ФБР, содержащем 0,5 мМ MgCl2 и 0,15 мМ CaCl2. Через день отбирали пробы крови путем пункции сердца под метоксифлурановой анестезией и высевали на чашки с шоколадным агаром. Число бактерий в мл крови подсчитывали через 24 часа. Статистическое значение различий, наблюдаемых в уровнях бактеремии относительно контролей, анализировали с помощью т-теста Стъюдента. Результаты суммированы в таблице I.
Пример 23
В этом примере описывается процесс, примененный для получения D15-специфических T-клеточных линий. Мышей BALB/c (H-2d), закупленных на Charles River Animal (Монреаль, Канада), индивидуально подкожно примировали 20 мкг фосфата алюминия (алюм). Животным дважды повторно вводили ту же самую дозу иммуногена через 3-недельные интервалы. Через десять дней после последнего введения антигена селезенки иммунизированных мышей удаляли. Спленоциты культивировали при нагрузке 5,75 • 105 клеток на ячейку в конечном объеме 200 мкг среды RPMI 1640 (Flow Lab), дополненной 10% инактивированной нагреванием плодной телячьей сыворотки Gibco, 2 мМ 1-глютамина (Flow Lab), 100 Ед/мл пенициллина (Flow Lab) и 5•10-5 М 2-меркаптоэтанола (Sigma) в присутствии различных концентраций (1,10 и 100 мкг на мл) отдельных пептидов D15 (таблица 2) в 96-ячеечных платах (Nunc, Дания). Культуры выдерживали во влажном инкубаторе в присутствии 5% CO2 в атмосфере. Культуры в трех повторностях выполняли для каждой концентрации каждого пептида. Через пять дней в ячейки микротитровальной платы добавляли 150 мкл 10% супернатанта крысиной с конканавалином A культуры, разведенного в культуральной среде, в качестве источника интерлейкина -2 (IL-2) для размножения пептид-специфических Т-клеток. Через шесть дней 150 мкл супернатанта удаляли из каждой микрокультуры и добавляли 150 мкл свежего супернатанта культуры, IL-2, для дальнейшего размножения и поддержания жизнеспособности пептид-специфических Т-клеток. После дополнительной 6-дневной инкубации клетки отмывали три раза, каждый раз 200 мкл культуральной среды. Каждый блок культур затем стимулировали соответствующими концентрациями (1,10 и 100 мкг на мл) пептида в присутствии 2 • 105 облученных (1500 рад) клеток селезенки BALB/c в конечном объеме 20 мкл культуральной среды. Затем из каждой ячейки удаляли шестьдесят мкл супернатанта микрокультуры. Супернатанты из каждого блока из трех повторностей культуры объединяли. Все супернатанты исследовали на 11-2, интерлейкин-4 и интерферон-гамма (FEN-γ). Определения IL-2 и IL-4 выполняли, используя наборы для ELISA мышиных IL-2 и IL-4, закупленные в Endogen. Inc. (MA, USA), соответственно. Исследование IFN- γ выполняли, используя набор для ELISA на мышиный IEN- γ, поставленный Genzyme Corporation (MA, USA). Испытуемые супернатанты культур исследовали в 1 наборе в 5 разведениях по указаниям производителей. Полученные результаты представлены в таблице 7.
Пример 25
В этом примере описана общая процедура, использованная для получения мышиных D15-специфических моноклональных антител.
Мышей BALB/с внутрибрюшинно иммунизировали 20-50 мкг N-концевого фрагмента rD15 (пример 11) эмульгированнными в полном адъюванте Фрейнда. Через две недели мышам делали еще раз инъекции того же самого количества иммуноген в неполном адъюванте Фрейнда (НАФ). За три дня до слияния мышам снова вводили то же самое количество иммуногена в НАФ. Гибридомы получали путем слияния селезеночных лимфоцитов от иммунизированных мышей с несекретирующими клетками миеломы Sp/2/0, как описано ранее Hamel et. al (1987). D15-специфические гибридомы клонировали путем последовательных ограничивающих разведений и отбирали по продукции анти D15 моноклональных антител. Были идентифицированы восемь D15-специфических гибридомных клеточных линий, размножены и заморожены и жидком азоте. Одна из гибридомных клеточных линий, 6CB-F6-C6, была частично охарактеризована. Моноклональные антитела (Маb 6CВ-Г6-C6) реагируют с пептидом D15-Р8 (ти Mab 6CB-Г6-C6) использовали для получения D15-специфичной аффинной колонки с Mab и для очистки природного D15 из клеточной массы H.influenzae (пример 13). 0,5 мл кроличьей антисыворотки к N-концевому фрагменту rD15 (Ат к фрагменту rD15) смешивали с или девятью пептидами D15 (100 мкг пептидов с D15-P2 по D15-P10, смотрите таблицу 2) или с 600 мкг N-концевого фрагмента rD15 при комнатной температуре в течение 1 часа. Антисыворотку и пептиды, смешанные с ФБР, использовали в качестве контролей. Семидневным крысятам п/к вводили 0,2 мл различных препаратов. Через 24 часа края в/б заражали 200 кое Н.influenzae типа b штамма Minn A. Образцы крови отбирали через 24 часа после заражения. Числа в скобках показывают число животных, у которых была бактеремия из общего числа зараженных животных. Уровень бактеримии выражен как среднее от значений, полученных от семи крысят, обследованных отдельно ± стандартное отклонение (CO). 0,5 мл кроличьей антисыворотки к фрагменту rD15 (Ат к rD15) смешивали с пятью пептидами D15 (пептиды с Р4 по Р8, 250 мкг каждого пептида) при комнатной температуре в течение 1 часа. Антисыворотку и пептиды, разведенные в ФБР, использовали в качестве контролей. Семидневным крысятам п/к водили 0,2 мл указанного материала. Через 24 часа крысятам в/б вводили 200 кое H. influenzae типа b штамма Minn A. Образцы крови отбирали через 24 часа после заражения. Числа в скобках показывают число животных, у которых присутствовала бактеремия из общего числа зараженных животных.
Уровень бактеремии выражен как среднее арифметическое от значений, полученных от шести крысят, обследованных отдельно ± одно CO.
БИБЛИОГРАФИЯ REFERENCES
O'Hagan (1992).
Ulman et al., 1993.
Berns C.A. and Thomas C.A. (1965) J. Mol. Biol. 11:476-490.
Thomas W.R. and Rossi A.A. (1986) Infect. Immun. 52:812-817.
Thomas W.R. et al. (1990) Infect, and Immun. 58:1090-1913.
Carlone G.M. et al. (1986) J. Clin. Microbiol. 24:330-331.
Smith, D.B. and Johnson K.S. (1988) Gene 67:31-40.
Harkness, R. et al. (1992) J. Bacteriol. 174:2425-2430.
Hamel et al. (1987) J. Med. Microbiol. 23:163-170.
Mills et al. (1993) Infect. Immun. 61:399-410.
Trinchieri, (199З) Immunol. Today 14:335-338.
Hope, T.P. (1986) J. Immunol. Methods 88:1-18.
Zangwill et al., 1993. MMWR 42:1-15.
Loeb et al. 1987. Infect. Immun. 55:2612-2618.
Panezutti, 1993. Infect. Immun. 61:1867-1872.




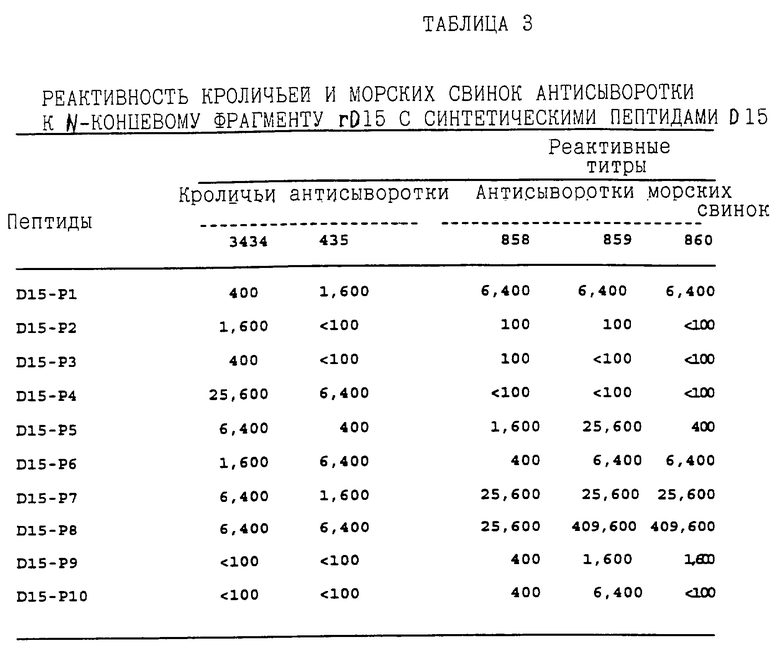


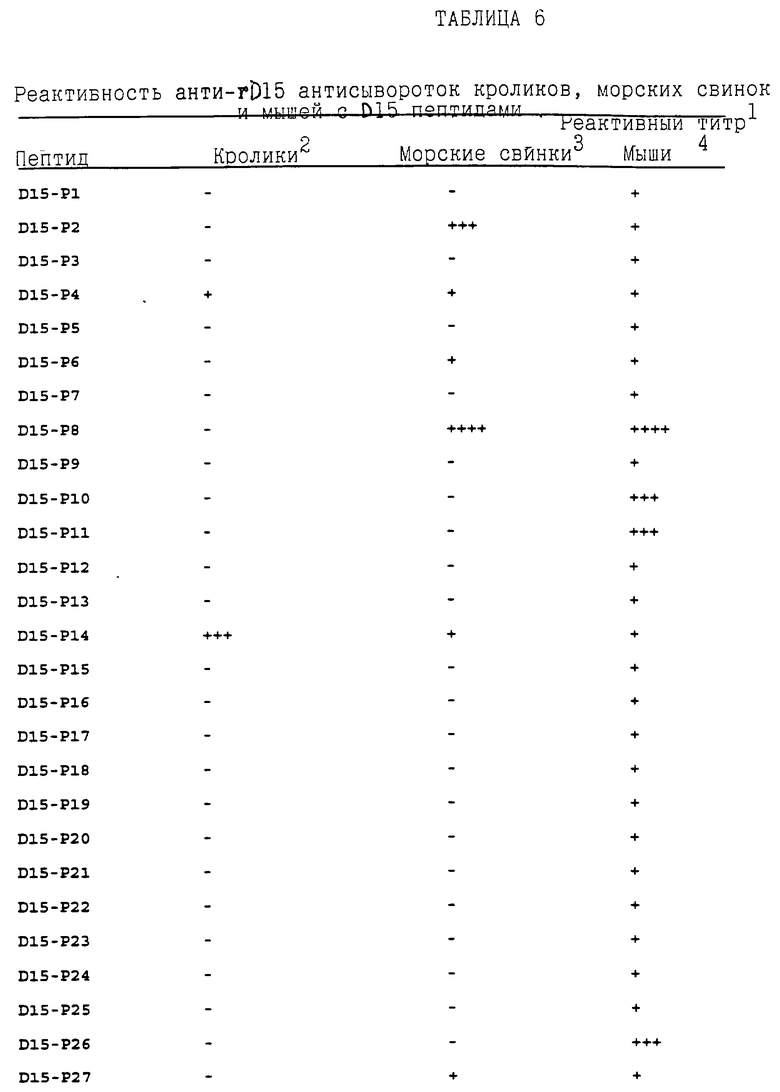

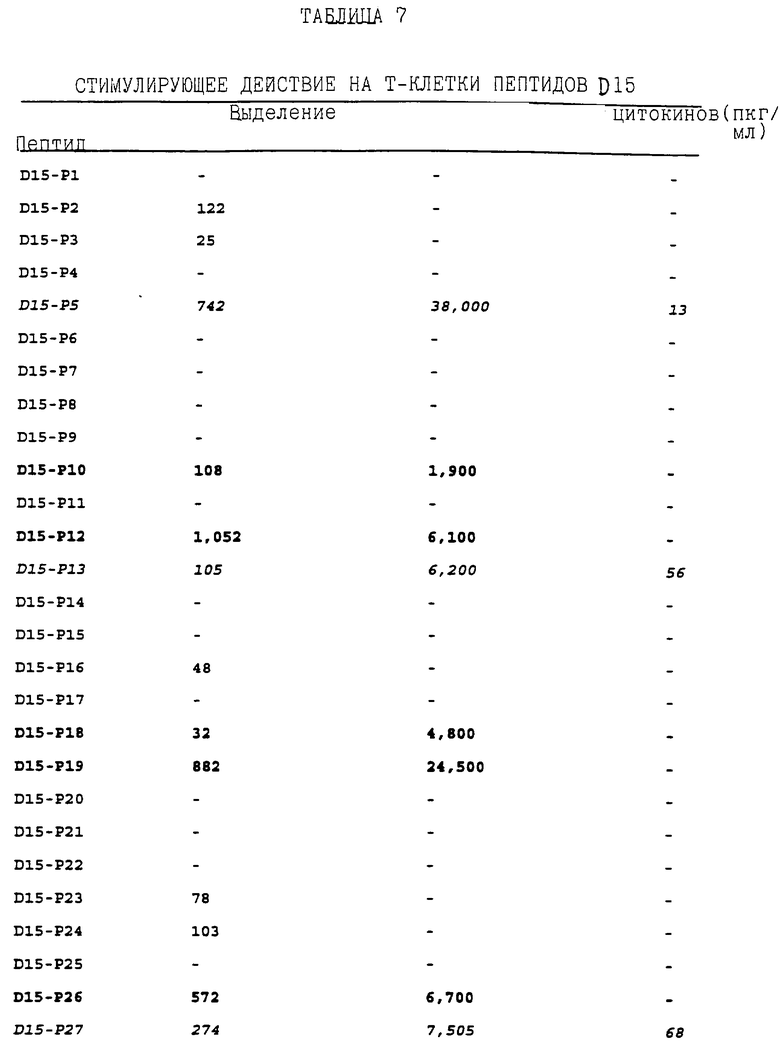


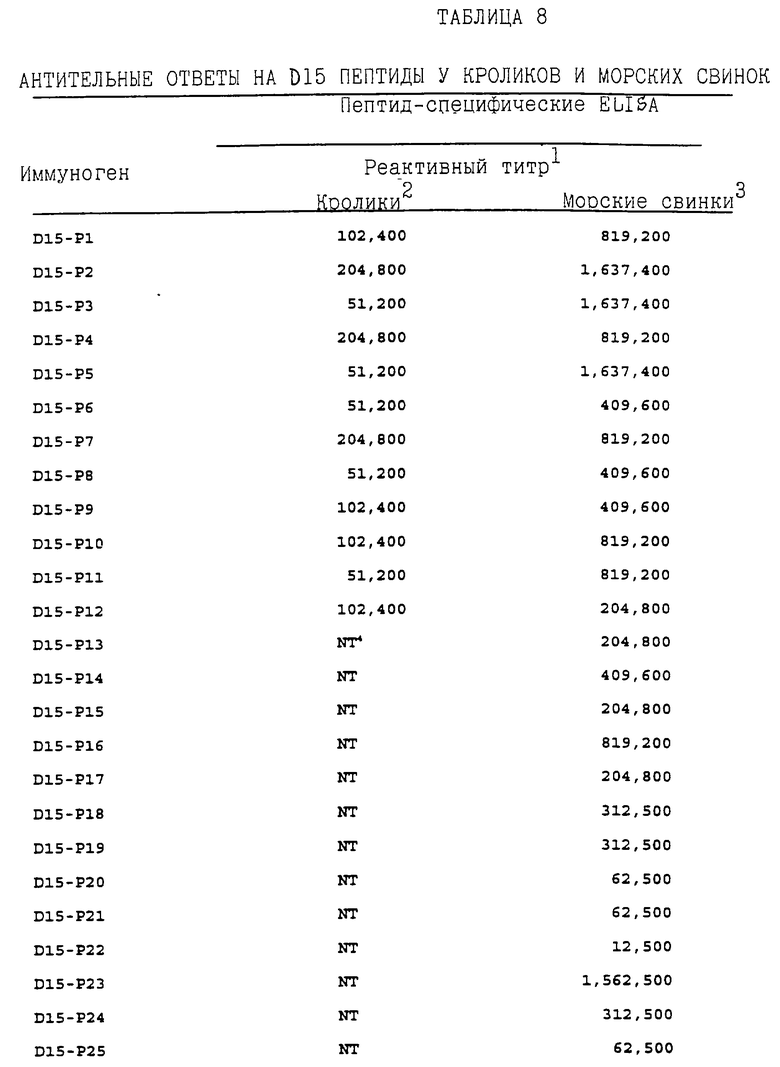

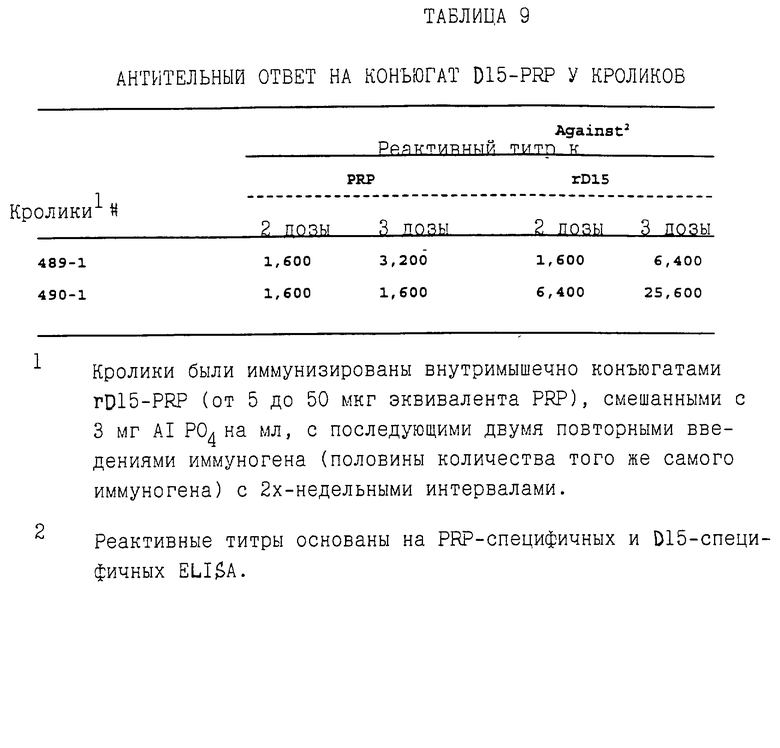






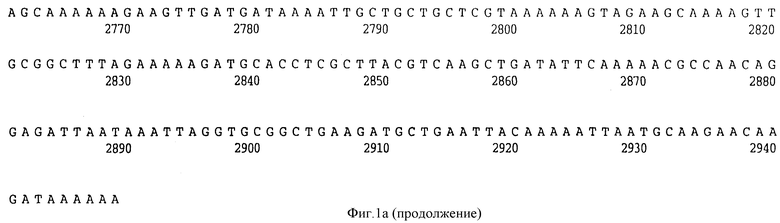







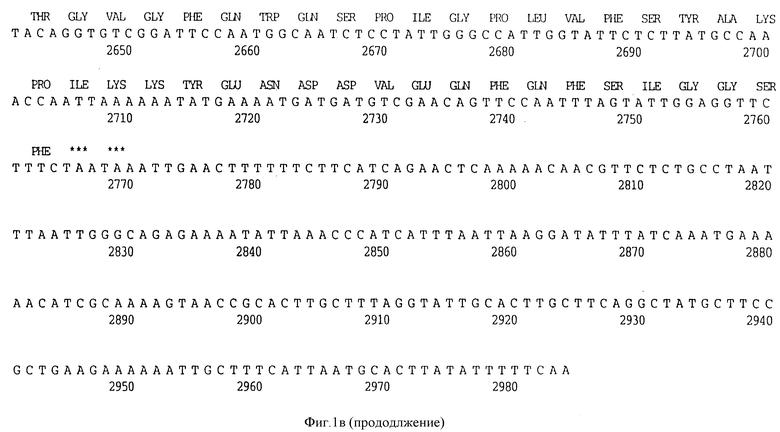

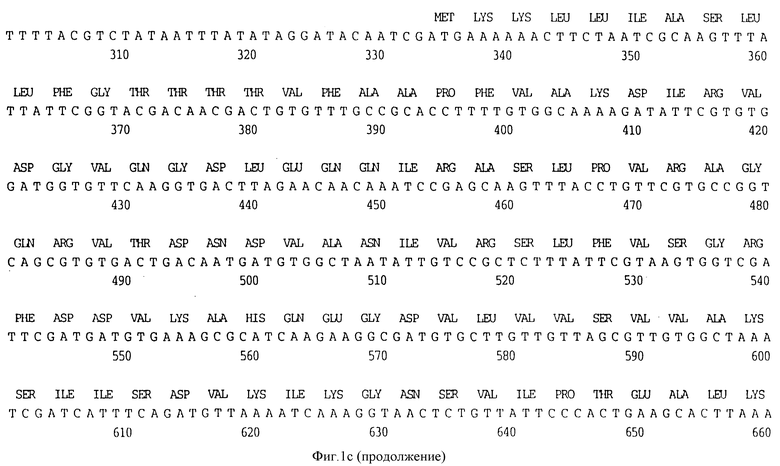





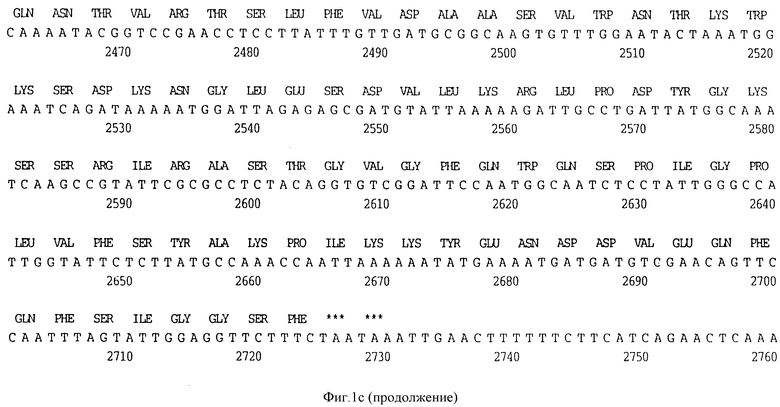
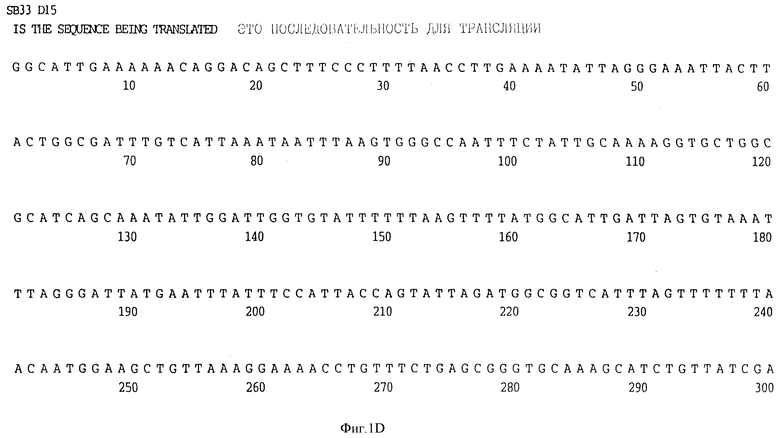
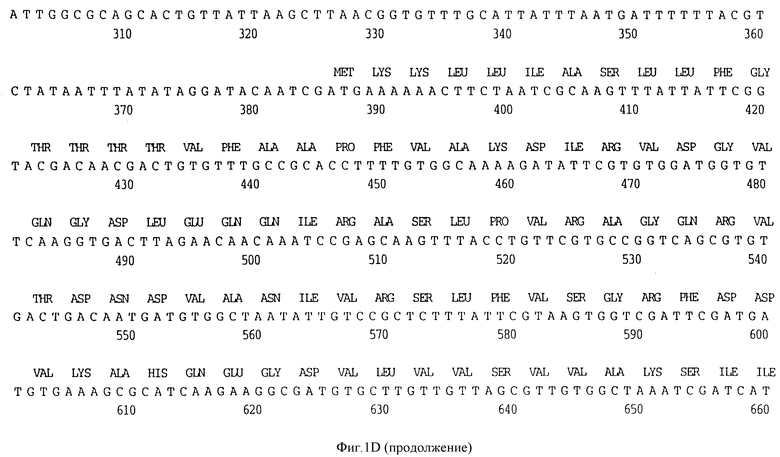


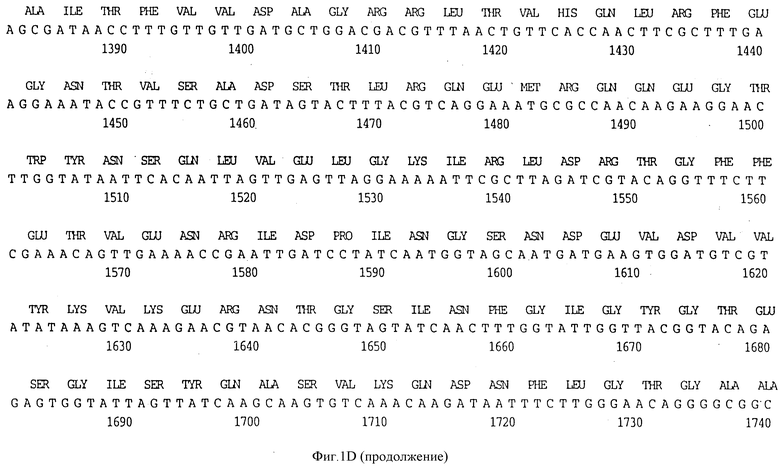



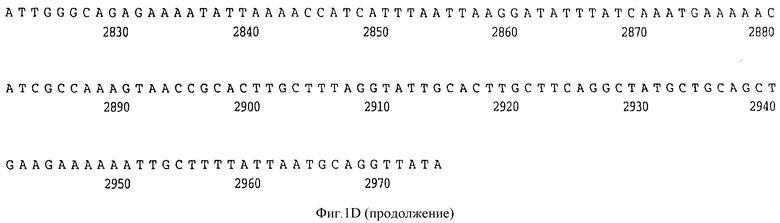

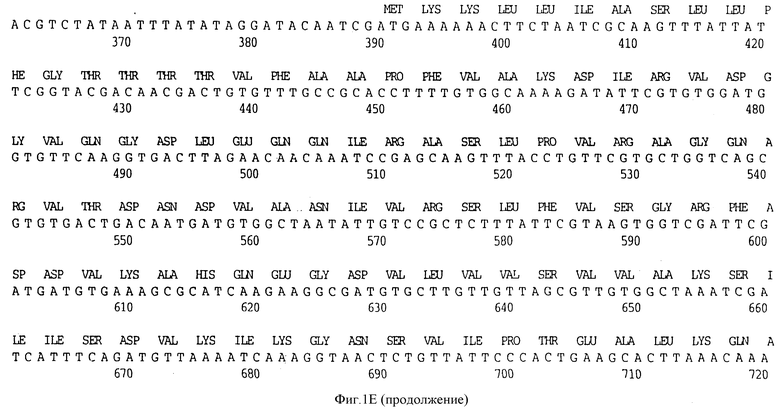
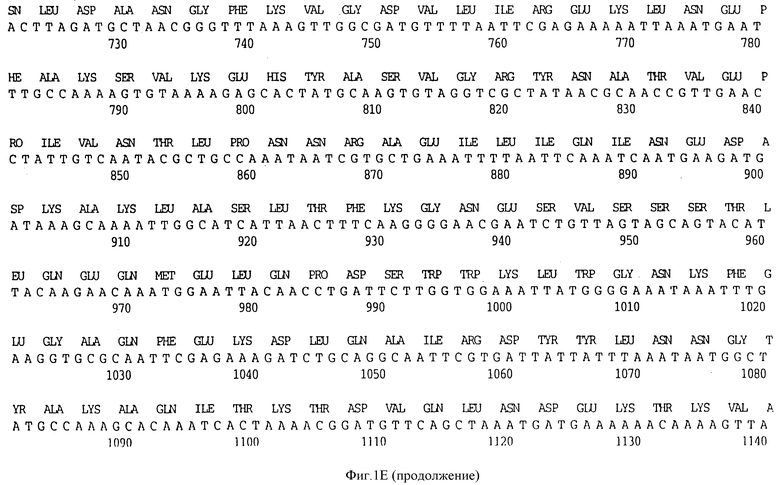




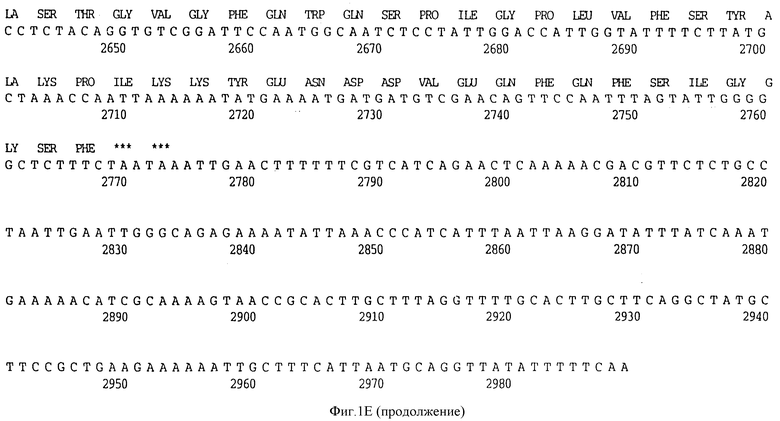


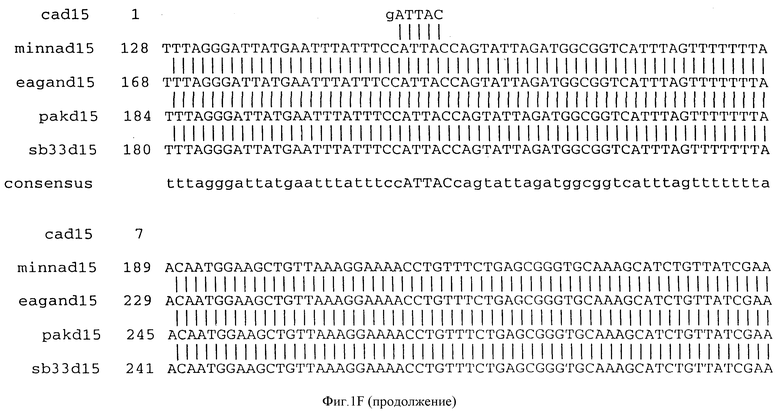
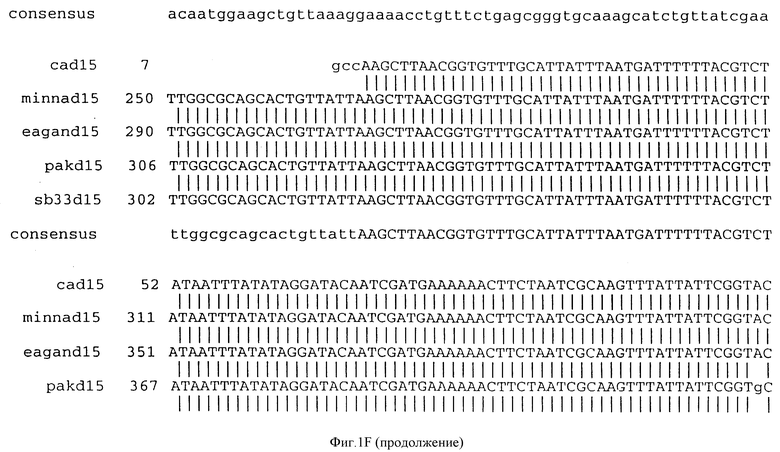
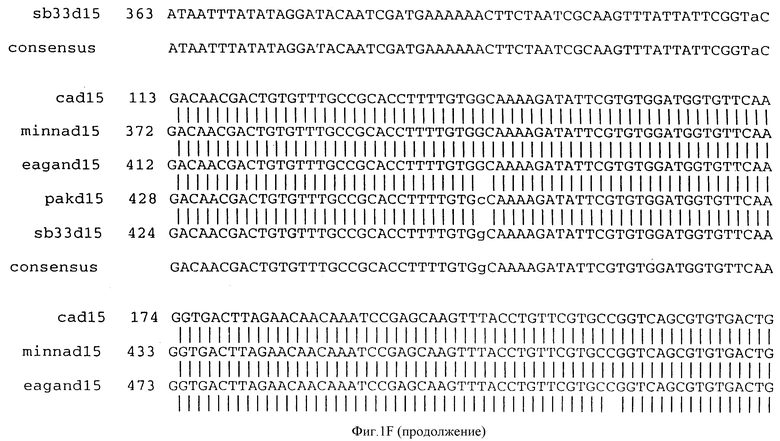
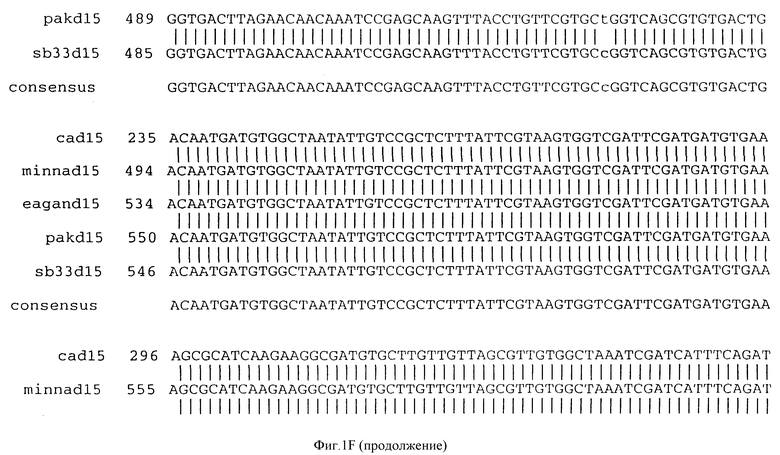
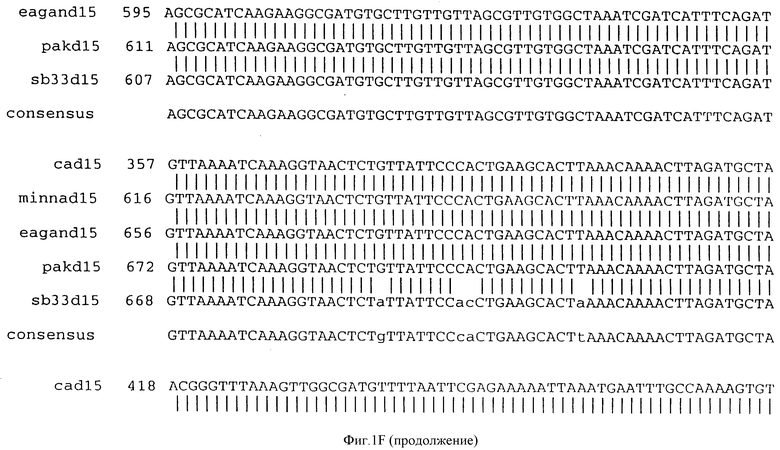
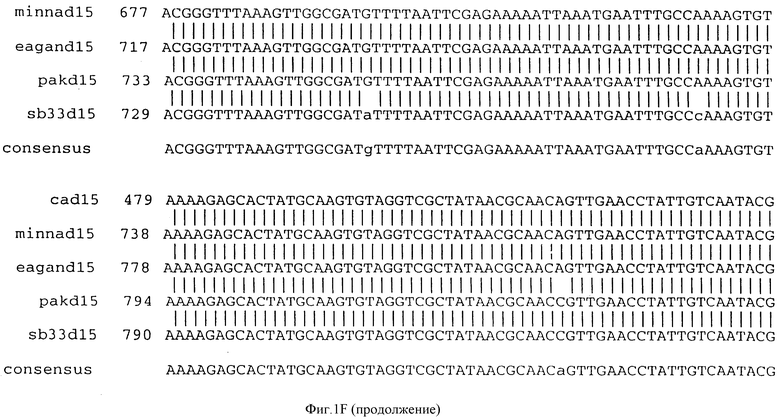
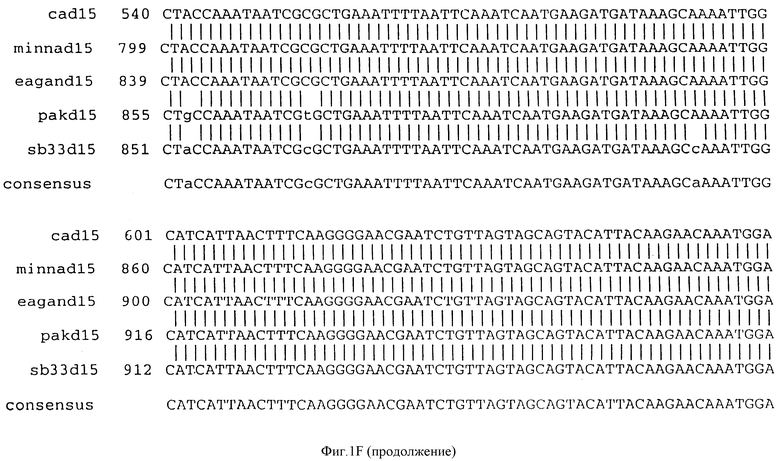
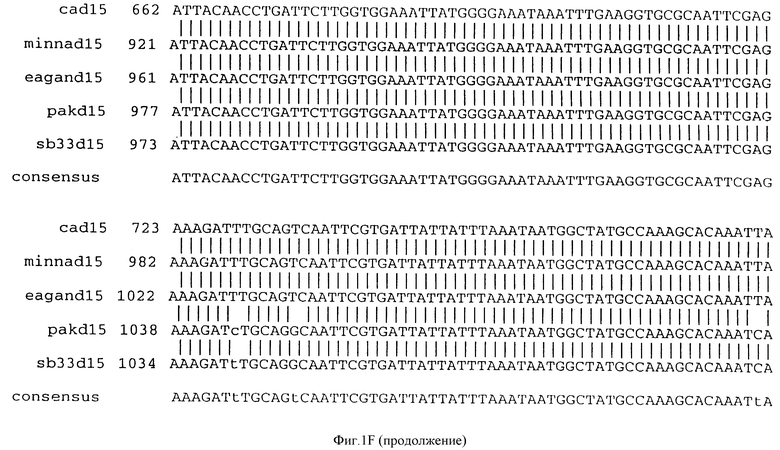
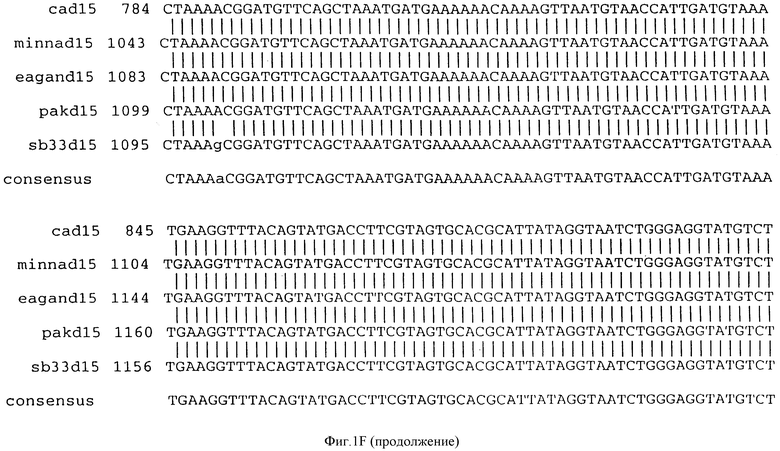
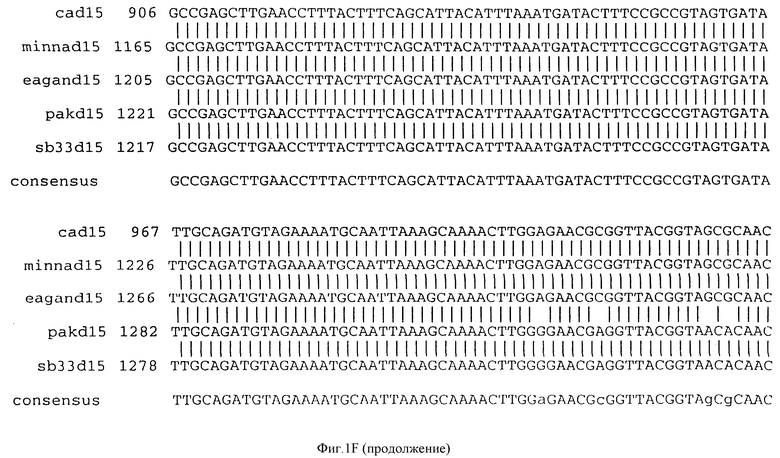
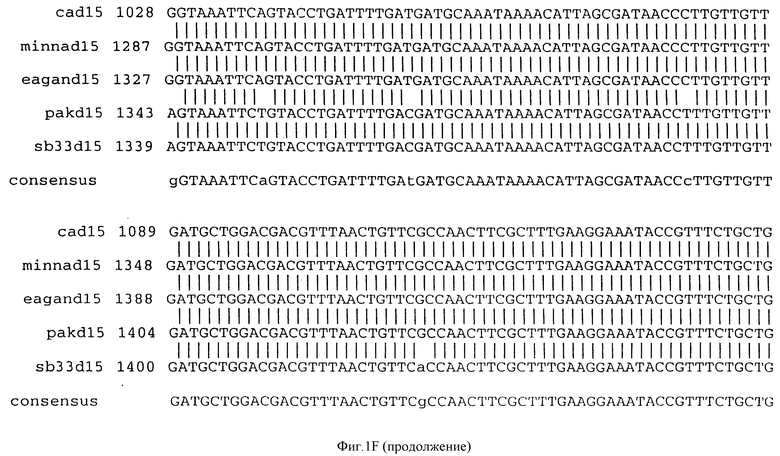
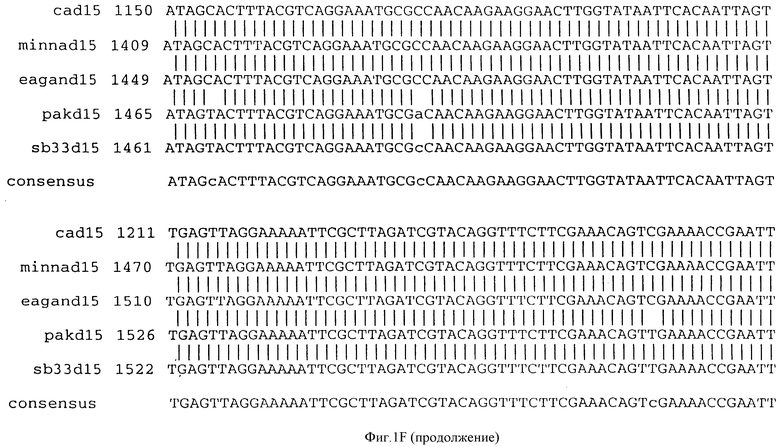
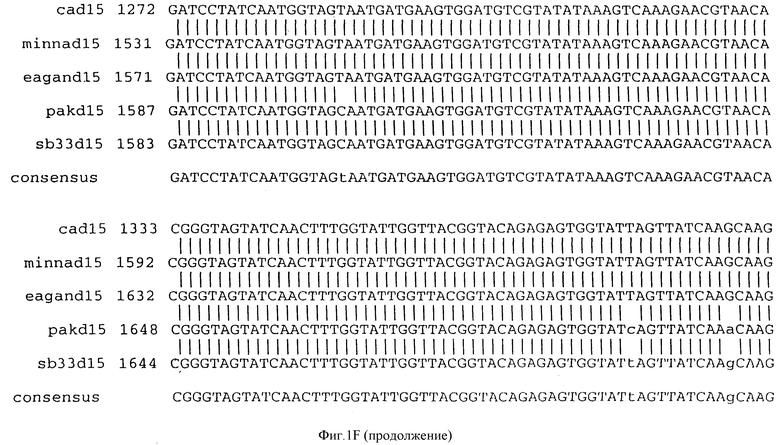
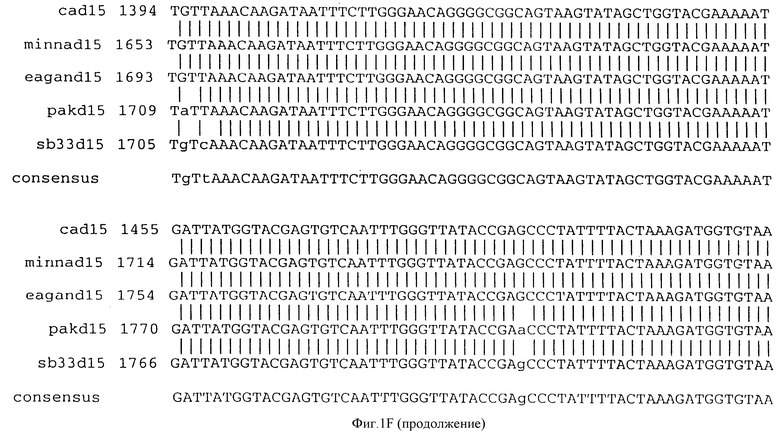
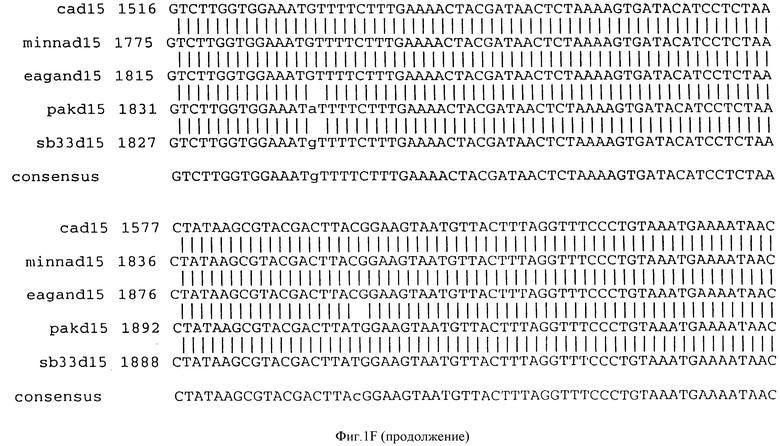
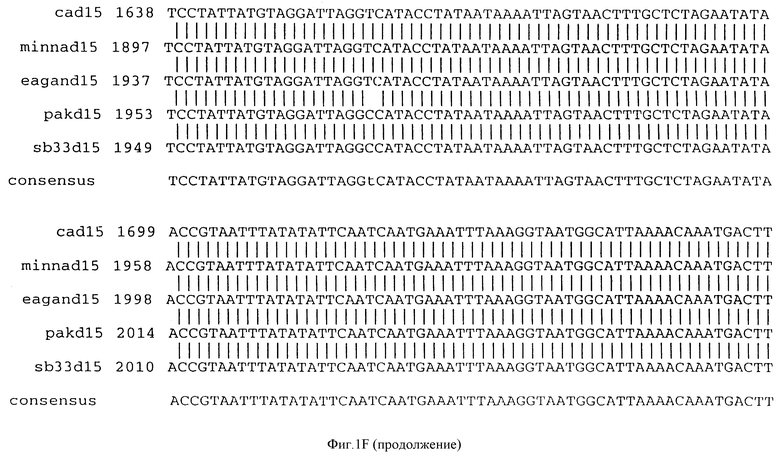
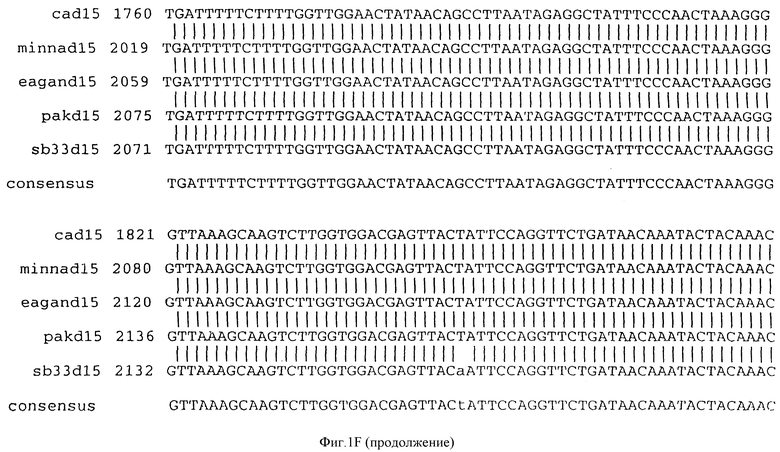
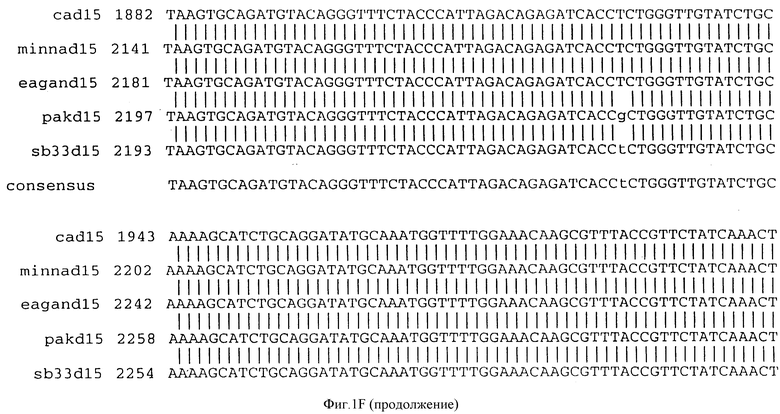
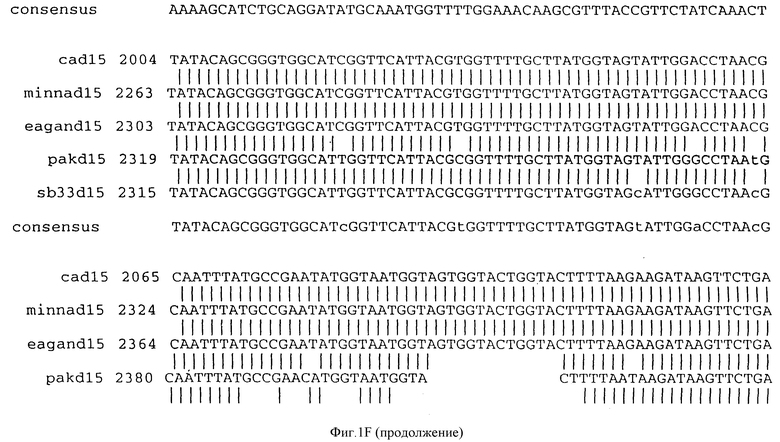

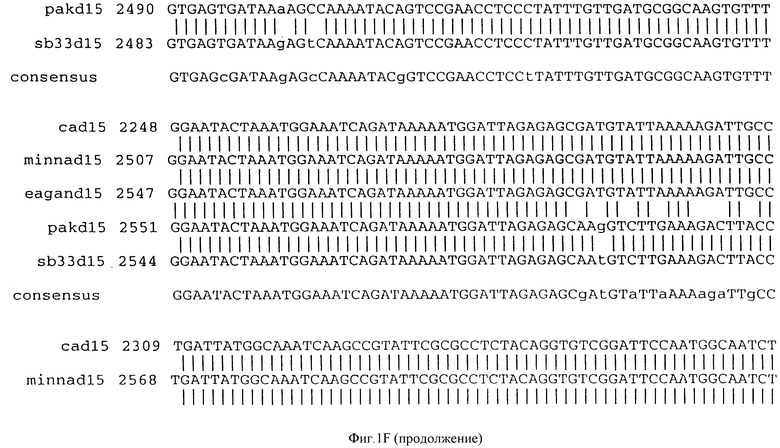
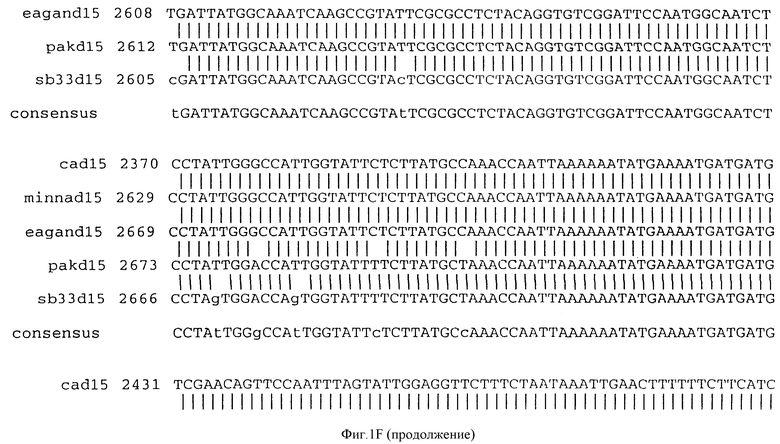
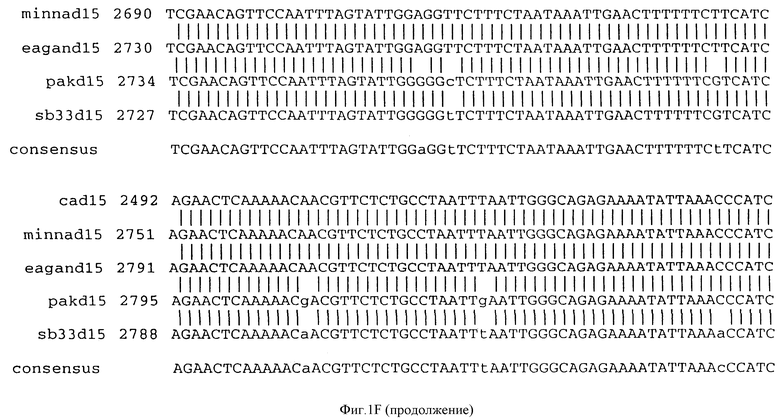
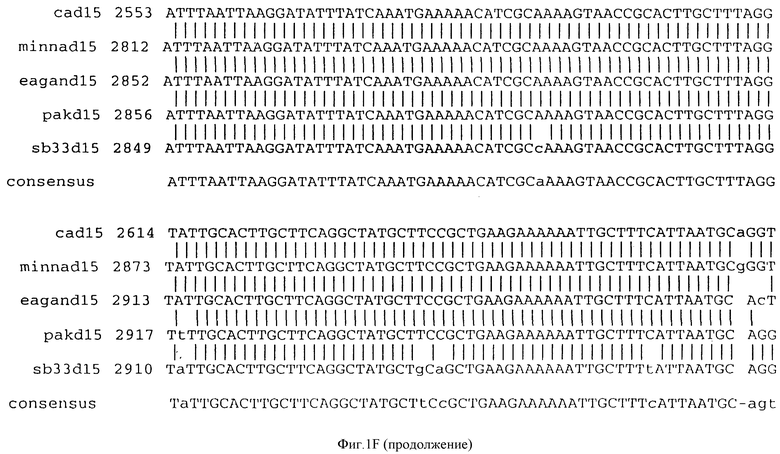


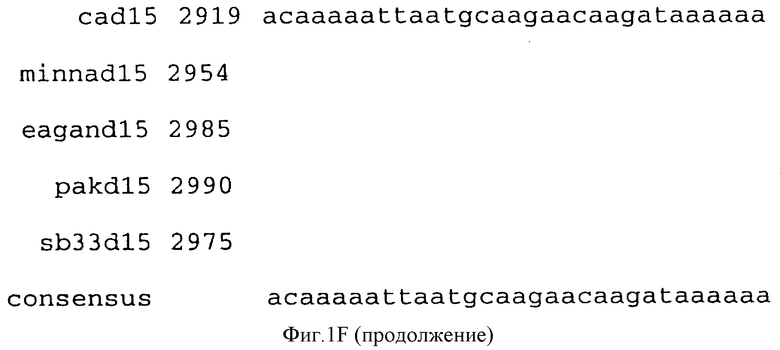
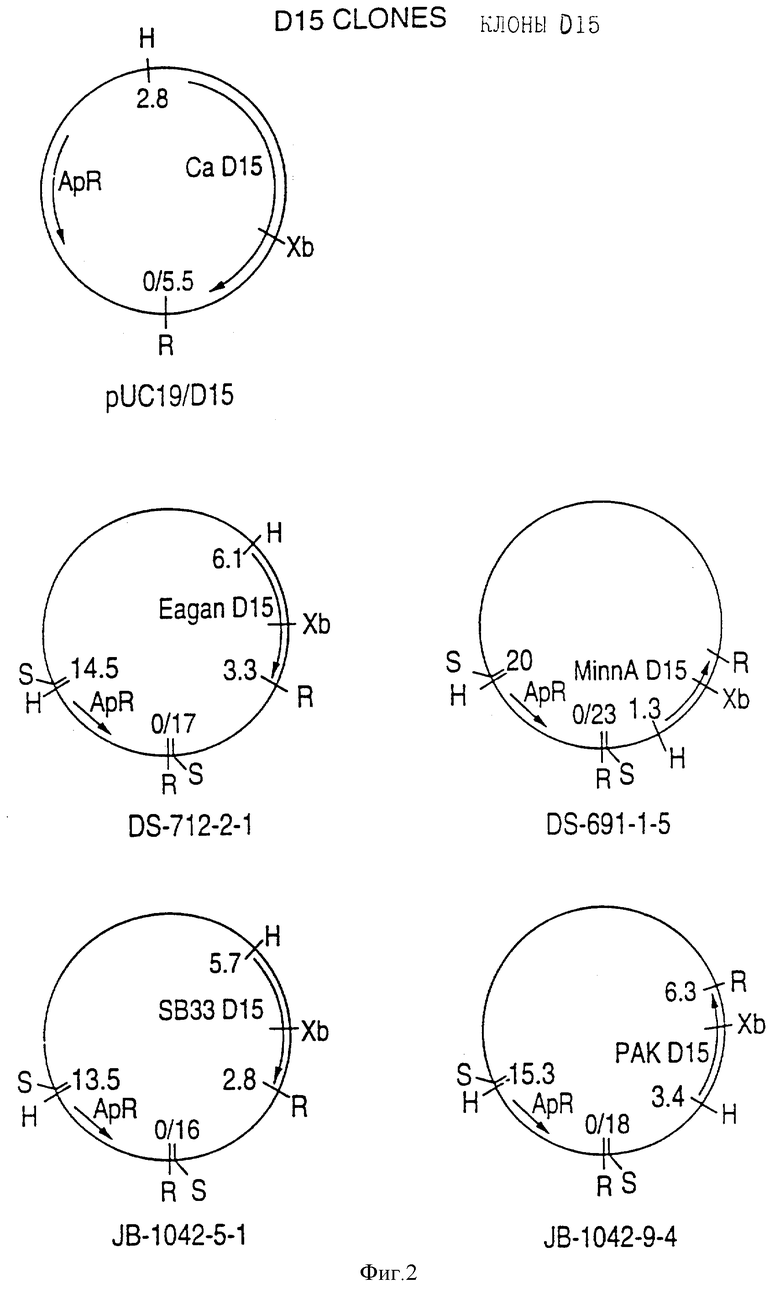


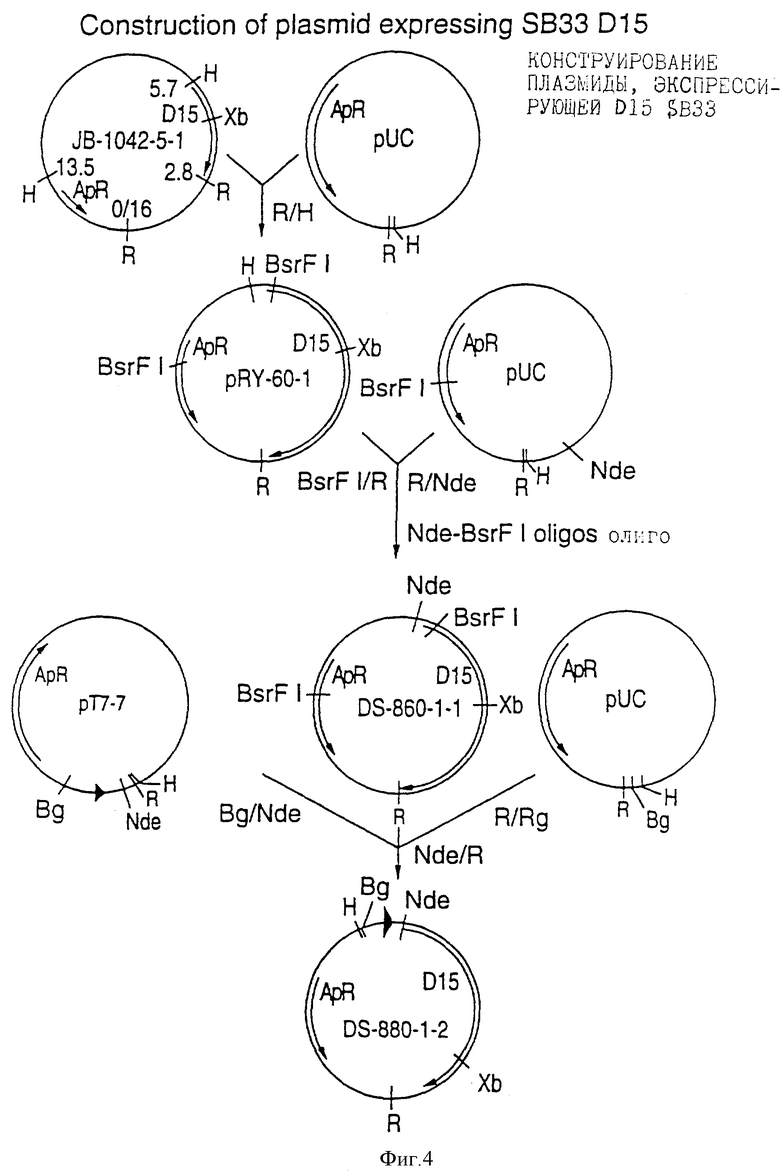
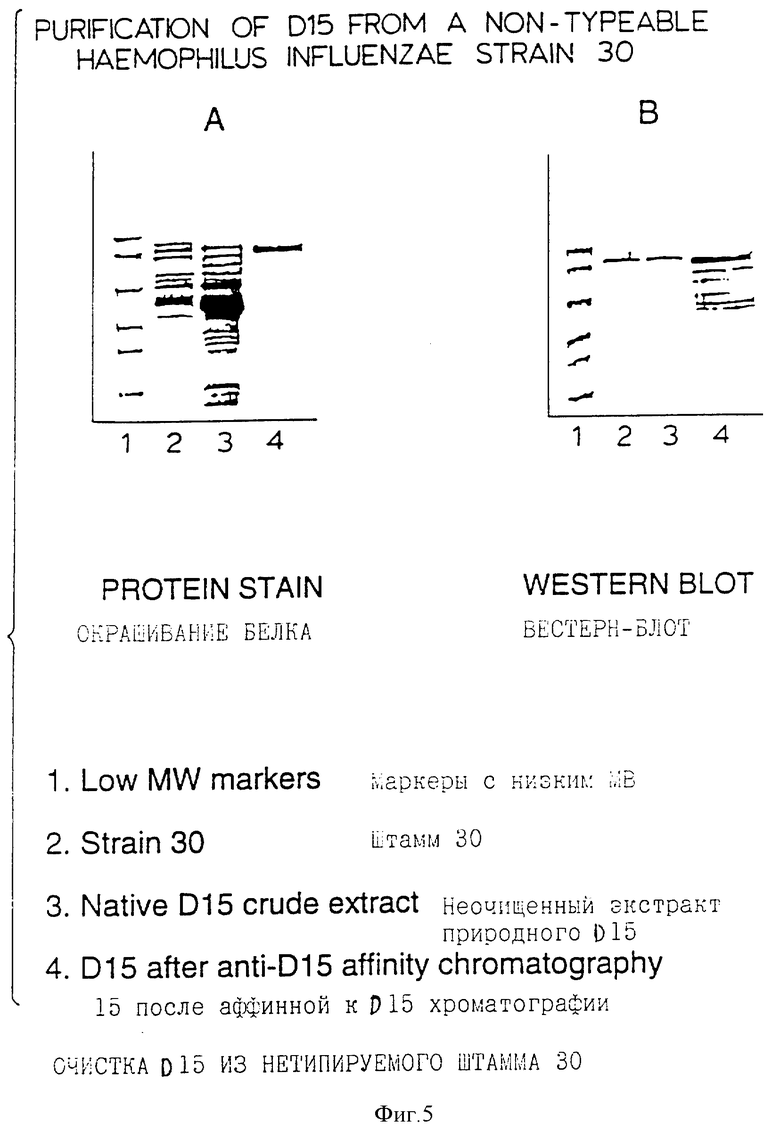

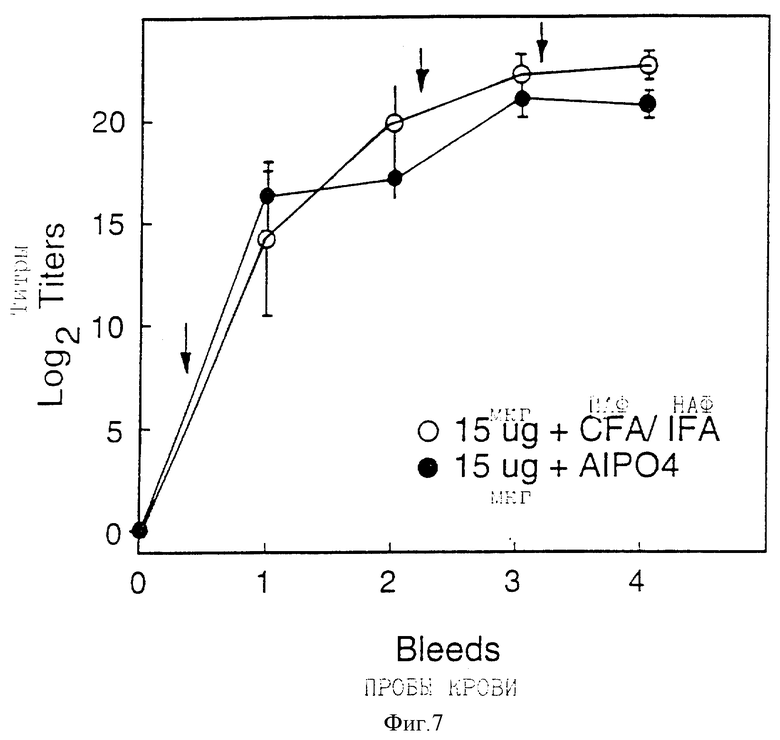
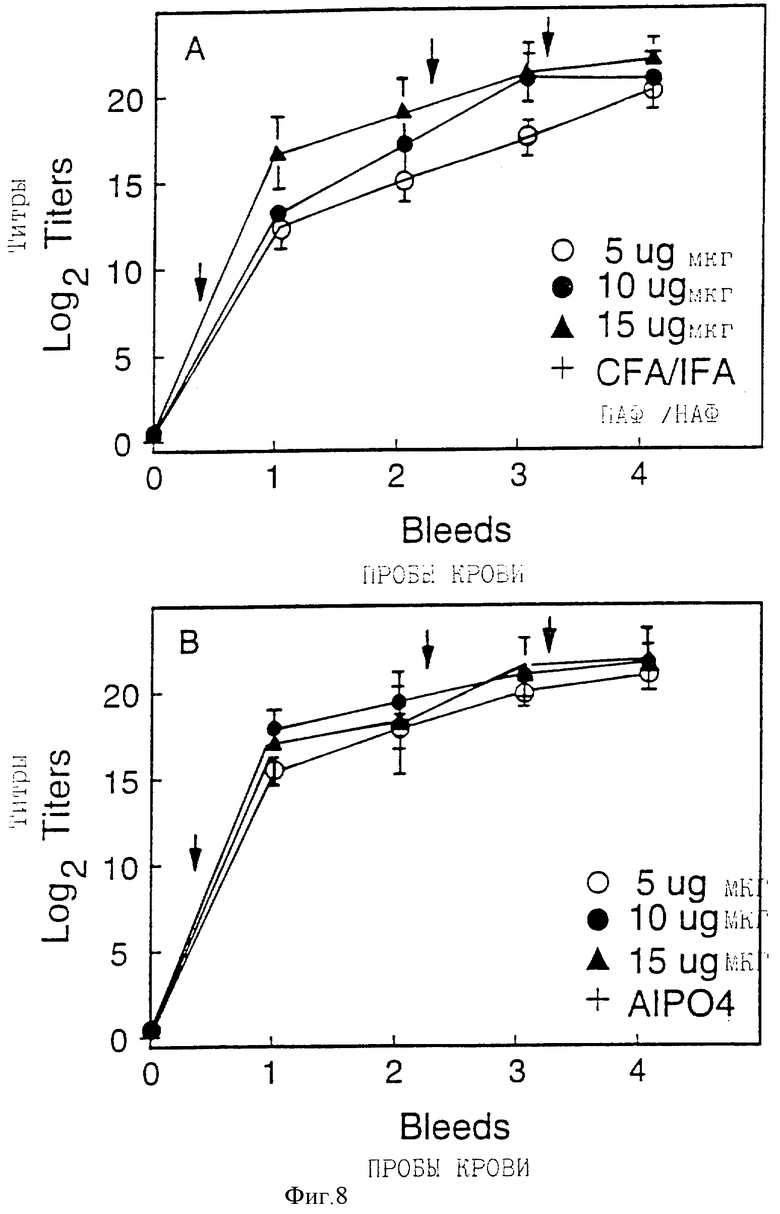

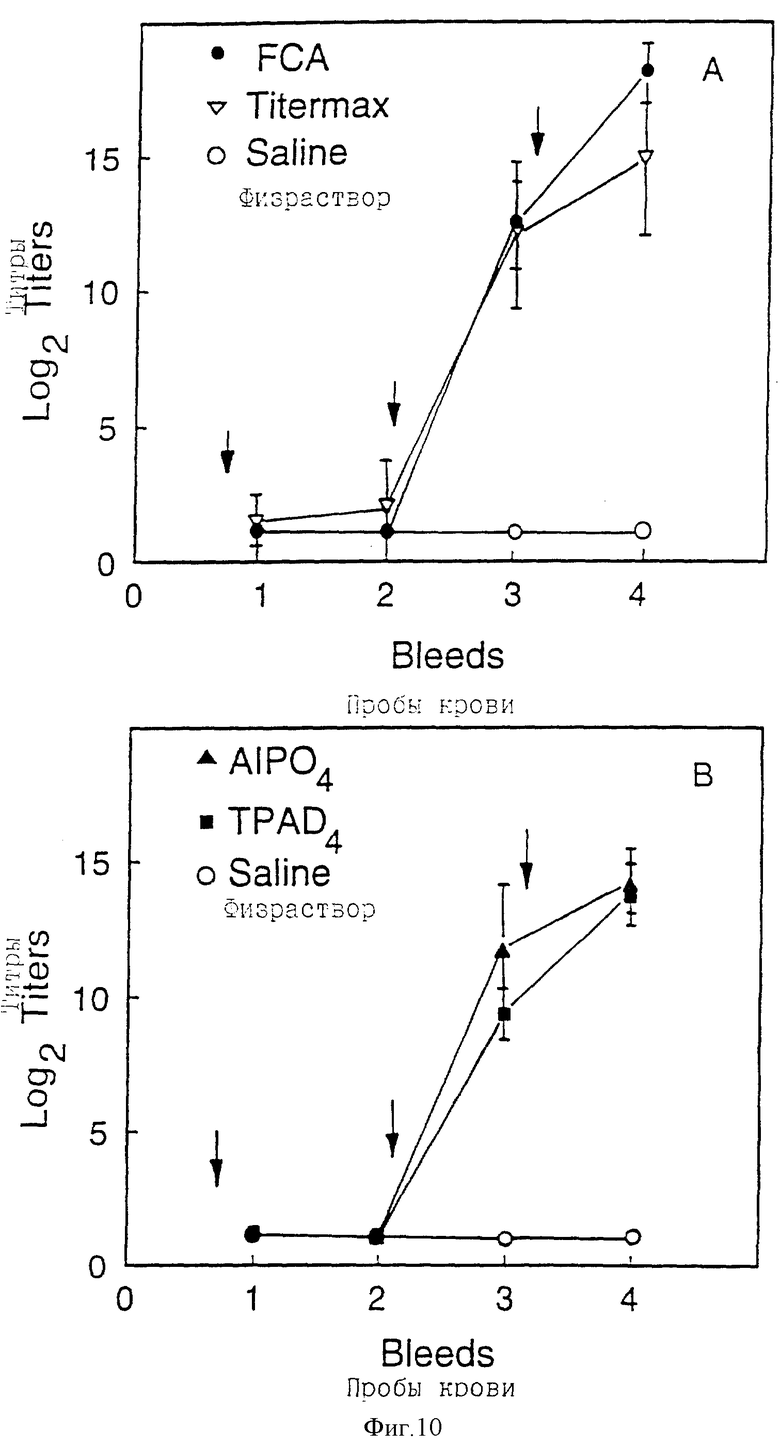
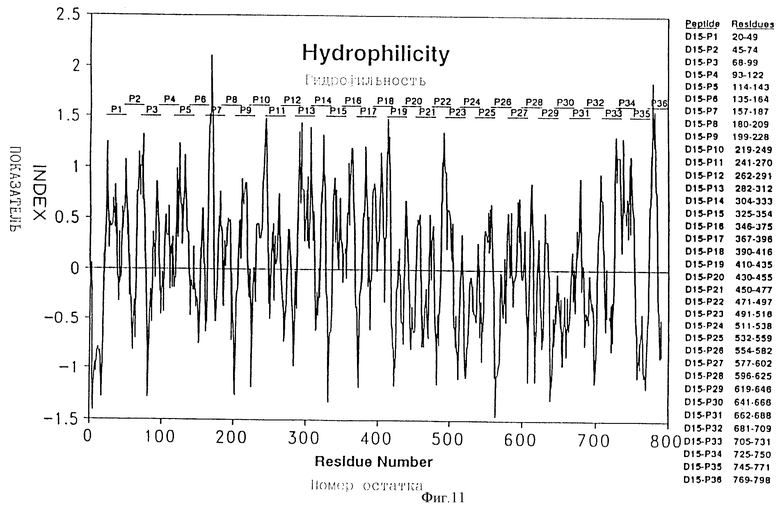
Изобретение характеризует очищенную и изолированную молекулу нуклеиновой кислоты, способную кодировать белок D 15 наружной мембраны Haemophilus. Молекула может быть встроена в рекомбинантную плазмиду или рекомбинантный вектор. Охарактеризован также белок, кодируемый нуклеиновой кислотой. Использование изобретения позволяет получить вакцины для лечения и диагностики заболеваний, вызванных Haemophilus. 9 с. и 14 з.п.ф-лы, 17 ил., 10 табл.
| УСТРОЙСТВО ДЛЯ ТЕЛЕИЗМЕРЕНИЯ УРОВНЯ^.C.V'T*'^*^' '' | 0 |
|
SU378929A1 |
| Огнетушитель | 0 |
|
SU91A1 |
| Способ получения динуклеотидов | 1983 |
|
SU1404512A1 |
