Изобретение относится к синтетическим вакцинам против инфекции Haemophilus influenzae (Hi). В частности, изобретение относится к использованию потенциальных T-хелперных детерминант (THDs), и B-клеточных эпитопов (BE) белков внешней мембраны Hi (OMP) P1, P2 и P6, ковалентно связанных с синтетическим олигосахаридом, содержащим повторяющиеся фрагменты полирибозилрибитол-фосфата (sPRP), образуя таким образом иммуногенные синтетические PRP-пептидные конъюгатные вакцины, которые вызывают в млекопитающих образование анти-PRP и анти-OMP антител с высоким титром.
Haemoph ilus influenzae типа в (Hib)-основная причина бактериального менингита у детей в возрасте до пяти лет [1,2]. Ссылки указаны в конце описания. От фагоцитоза бактерии защищены полисахаридной капсулой, которая представляет собой полимер полирибозилрибитол-фосфата (PRP). Антитела, продуцируемые на капсульный полисахарид этого организма, обладают защитным свойством [3] . Были получены эффективные конъюгатные вакцины, в которых PRP связан с различными белками-носителями, такими как дифтерийный токсин (PRP-D), тетанусный токсин (PRP-T), CRM197 (HbOC) Neisseria meningidis и белок внешней мембраны Neisseria meningidis [4,5]. Однако эти конъюгатные вакцины не являются защитой от других распространенных инкапсулированных H.influenzae - типа a- и c-штаммов и, что более важно, от не имеющих капсулы неклассифицированных разновидностей H. influenzae, которые являются причиной воспаления среднего уха, против которого нет вакцины. Следовательно, необходимо включение отобранных неинкапсулированных H.influenzae-иммуногенов в существующие Hib - вакцины, чтобы разработать универсальную Hi вакцину.
Granoff и Munson [6] показали, что антитела против внешних мембранных белков (OMP) Hib PI, P2 и P6 обладают защитным свойством при модельном заражении новорожденных крыс бактериемией. Значит многообещающей стратегией для получения универсальной вакцины с усиленной защитной способностью против H. influenzae было бы использование в качестве дополнительных иммуногенов и носителей для PRP либо очищенных OMP, либо их защитных эпитопов. Кодирующий ген Р1 был клонирован из различных подтипов Hib [7,8]. Сравнительный анализ последовательностей Pi-белков из этих Hib показал существование трех гипервариабельных участков. В действительности Pi-специфичные MA, о которых сообщал Hansen, узнают только 50% тестированных Hib [7,9] изолятов. Что касается Р2-белка, хотя нуклеотидные последовательности P2 гена, выделенного из двух различных подтипов Hib (1Н и 3L), оказались идентичны [10,11], найдена некоторая вариабельность аминокислот в Р2-последовательностях двух других подтипов (2L и 6U) [II]. Напротив, анализ антигенных детерминант, генных последовательностей и длин рестрикционных фрагментов показал, что P6 белок высоко консервативен из штаммов Hib [12].
Недавние исследования показали, что мышиные Pl-специфичные моноклональные антитела (MAB 7C8) и кроличьи антитела на очищенный PI белок из определенной и из неопределенной разновидностей H.influenzae имели защитный эффект в животных моделях [9, 13, 14]. Murphy и Bartos [15] также показали, что моноклональные антитела, узнающие экспонированную на клеточной поверхности неопределенного H.influenzae, имеют бактерицидную активность in Vitro. Для анти-Р1 и анти P2 моноклональных антител было найдено, что они перекрестно реагируют с определяемыми и неопределяемыми H.influenzae [16-18]. В действительности все еще существуют серьезные сомнения относительно использования всех нативных ОМР Hib как эффективной универсальной вакцины против обеих ветвей H. influenzae. Во-первых, у детей, выздоравливающих от отита, вызванного неопределяемыми Hi, вырабатываются бактерицидные антитела к различным антигенам, таким как Р2 и липополисахариды. Во-вторых, Р1- и Р2-кросс-реагирующие эпитопы, описанные выше, еще не идентифицированы. В-третьих, сообщалось, что [12] эпитоп(ы), которые узнаются анти-Р6 бактерицидными антителами, экспрессируются в малых количествах на клеточной поверхности и возможно таким образом возобновление инфекции. В-четвертых, недостаточно известно пока о роли клеточного иммунного ответа на ОМР. Иммунодоминантные Т-хелперные клеточные эпитопы ОМР Hi пока не охарактеризованы. Значит необходима идентификация функциональных T-хелперных клеточных эпитопов и сохраняющихся экспонированных на поверхности и/или защитных B-клеточных эпитопов Р1, Р2 и Р6-белков, чтобы определить, могут ли они вызывать иммунный ответ против Hi инфекции.
Методы индукции иммунности к болезням постоянно улучшаются и в настоящее время основное направление - использовать в качестве антигена меньший и хорошо очищенный материал. Цель - устранить возможные побочные эффекты определенных природных иммуногенов, сохраняя их иммунногенность и способность к защите от болезней. Последние исследования показали, что иммунизация экспериментальных животных синтетическими пептидами, представляющими собой определенные участки вирусных и бактериальных белков, могут вызывать иммунный ответ на родительские белки и нейтрализовать их функции [19-22]. Так что синтетические пептиды являются потенциальными антигенами для производства недорогих и надежных вакцин против инфекционных возбудителей. Недавний прогресс в фундаментальной иммунологии свидетельствует о том, что хорошие и эффективные иммуногены должны содержать две различные функциональные антигенные детерминанты (эпитопы). Один эпитоп (Т-клеточный) служит для того, чтобы представлять комплекс соответствующего МНС II класса антигена иммунной системе и вызывать активизацию Т-хелперных клеток. Другой эпитоп (В-клеточный) должен узнаваться В-клеточным антигенным рецептором, чтобы вызывать продукцию антител [23-26]. Таким образом, чтобы получить сильнодействующую и эффективную синтетическую вакцину, в синтетическую конструкцию должны включаться оба функциональных эпитопа: Т-хелперный и В-клеточный.
Синтетические PRP-димер, тример и тетрамер были синтезированы, очищены и конъюгированы с белками-носителями для изучения иммуногенности в животных. Эти исследования показали, что конъюгаты из -тримеров PRP-белка в присутствии сильных добавок таких, как полный адъювант Фрейнда (CFA), могут вызвать в экспериментальных животных отклик анти-PRP антител.
Вместо использования условных гетерологичных белковых носителей наша стратегия использует синтетические пептиды, которые содержат иммунодоминантные эпитопы из ОМР Hi как дополнительные антигены и в качестве носителей PRP, чтобы разработать первое поколение полностью синтетических PRP-пептидных конъюгатных вакцин с усиленной защитной способностью и аутологичным Т- клеточным примированием. Подобные вакцины имеют и другие возможные преимущества перед существующими вакцинами, в которых PRP связан с чужеродным белком (дифтерийным токсином (PRP-D), тетанусным токсином (PRP-T) или CRM197 (Hb OC или OMP Neisseria meningidis). Во-первых, использование синтетических Hi должно сократить количество D или Т в любой будущей мультивалентной комбинированной вакцине, уменьшая таким образом риск гипериммунизации против этих белков-носителей. Во-вторых, PRP может быть привязан к сохранившемуся защитному эпитопу для получения вакцины против Hi-заболевания и отита.
Сокращения и определения:
CRM197 - нетоксичный белок антигенно кросс-реагирующий с дифтерийным токсином
Hi - Н.influenzae
Hib - Н.influenzae типа В
MAP - множественный антигенный пептид
MBS - малеимидобензоил-N-гидроксилсукцинимид
OMP - белок внешней мембраны
PEG - полиэтиленгликоль монометиловый эфир
PRP - полирибозирибитол фосфат
Один из аспектов настоящего изобретения направлен на обеспечение иммунногенных синтетических конъюгантных вакцин, включающих синтетические PRP-олигомеры и антигенные детерминанты белков внешней мембраны Hi.
Настоящее изобретение, с другой стороны, направлено на обеспечение вакцин на основе конъюгатов синтетических PRP с пептидами, включающих олигомеры синтетических PRP определенной длины.
Следующий аспект настоящего изобретения состоит в обеспечении химического процесса, в котором эффективно ведется производство синтетических PRP с химически реакционными функциональными группами, позволяющими сайт-направленно конъюгировать с антигенными детерминантами наружной мембраны Hi с использованием полиэтиленгликольмонометилового эфира (PEG) в качестве твердого носителя.
Следующий аспект настоящего изобретения состоит и в обеспечении метода, который может быть использован для улучшения иммуногенности конъюгатов синтетических PRP с пептидами путем выбора правильной ориентации сахарной части по отношению к Т-клеточному эпитопу.
Дополнительным аспектом настоящего изобретения является обеспечение такого химического процесса, который может усилить иммуногенность углеводов, используя множественные антигенные пептидные системы (MAP), содержащие антигенные детерминанты Hib как носители, чтобы увеличить плотность углеводной части в конъюгатах с синтетическими PRP.
Настоящее изобретение в дополнительном аспекте состоит в обеспечении универсальной вакцины против Hi, которая включает иммуногенные конъюгаты PRP с пептидами и обладающие кроссзащитными свойствами Hi антигены.
Следующий аспект настоящего изобретения состоит в получении нового поколения поливалентных вакцин, в составе которых иммуногенные конъюгаты PRP с пептидами и Hi антигенами, комбинированными с другими вакцинами, такими, как DTP -polio, Neisseria meningidis типа A, B, C, abd W и S. pneumoniae серотипа 6B, 14, I9F и 23F.
Настоящее изобретение в следующем аспекте направлено на получение конъюгатов синтетического PRP с пептидами, которые могут быть использованы в диагностических иммунотестах для определения присутствия анти- Hib антител, например, анти-PRP и анти-ОМР антител.
И следующий аспект этого изобретения состоит в получении смеси PRP-специфичных и ОМР-специфичных антител как компонента в наборе для диагностического иммуноанализа для определения присутствия в биологических образцах определяемых и неопределяемых штаммов Hi.
Краткая сущность изобретения.
Настоящее изобретение относится к обеспечению иммуногенов и возможных вакцин на основе пептидов, содержащих аминокислотные последовательности разнообразных антигенных детерминант (Т-хелперных и В-клеточных эпитопов) белков внешней мембраны (Р1, Р2 и Р6)Hib. Открыты синтетические вакцины, включающие один или больше таких пептидов, которые могут быть введены как свободные пептиды или ковалентно связанные с синтетическими PRP-олигомерами в качестве синтетических гликоконъюгатных вакцин и/или привязанные к липидной части с целью увеличения их иммуногенности.
В одном из аспектов настоящего внедрения обеспечивается синтетический пептид с аминокислотной последовательностью, соответствующей по крайней мере одной антигенной детерминанте по крайней мере одного белка H.influenzae, предпочтительно белка внешней мембраны H.unfluenzae b.
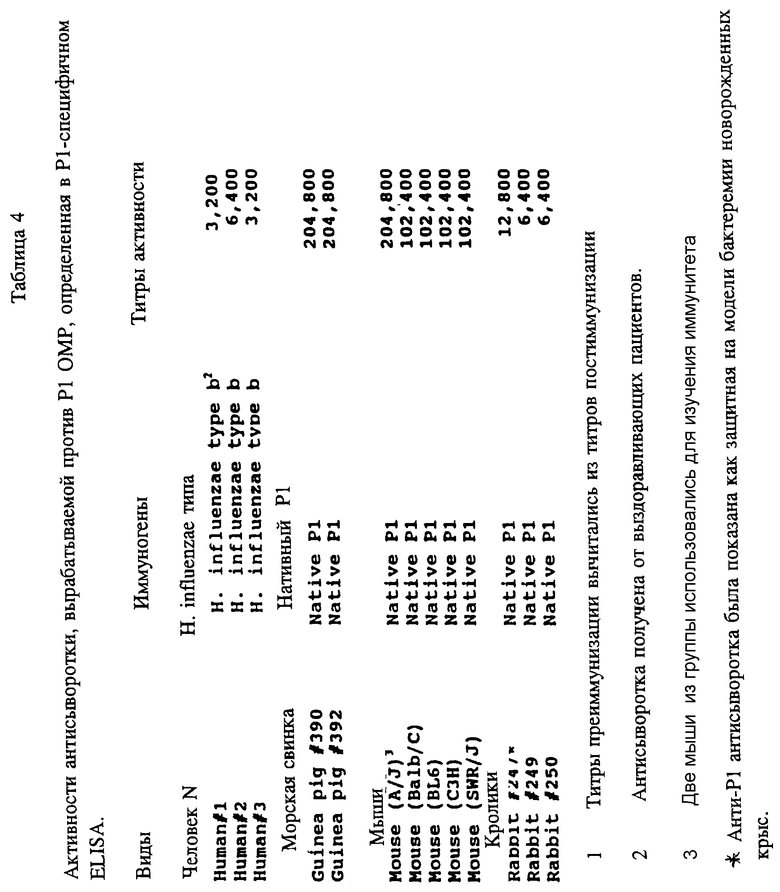
С одной стороны, настоящее изобретение включает значительно очищенную форму пептида, содержащего по крайней мере одну аминокислотную последовательность, относящуюся к сохранившейся антигенной детерминанте PI белка Hi, пептид которого может вызывать образование поликлональных антител, которые узнают Hi in vitro в млекопитающих. Эти PI-специфичные поликлональные антитела могут быть использованы в качестве компонентов тест-наборов для определения Hi в биологических образцах. Пептиды могут содержать, например, аминокислотные последовательности из следующих аминокислот: с 1 по 29, 39-64, 103-137, 165-193, 189- 218, 226-253, 248-283, 307-331, 400-437 и 179-218 зрелого PI- белка Hib MinnA-штамма в соответствии, как показано в табл. 1 (SEQ ID NOS : 1, 12, 3, 4, 5, 6, 7, 9, 13 или 14 и 15 соответственно) или же любая часть или комбинация из них сохраняющая иммуногенность.
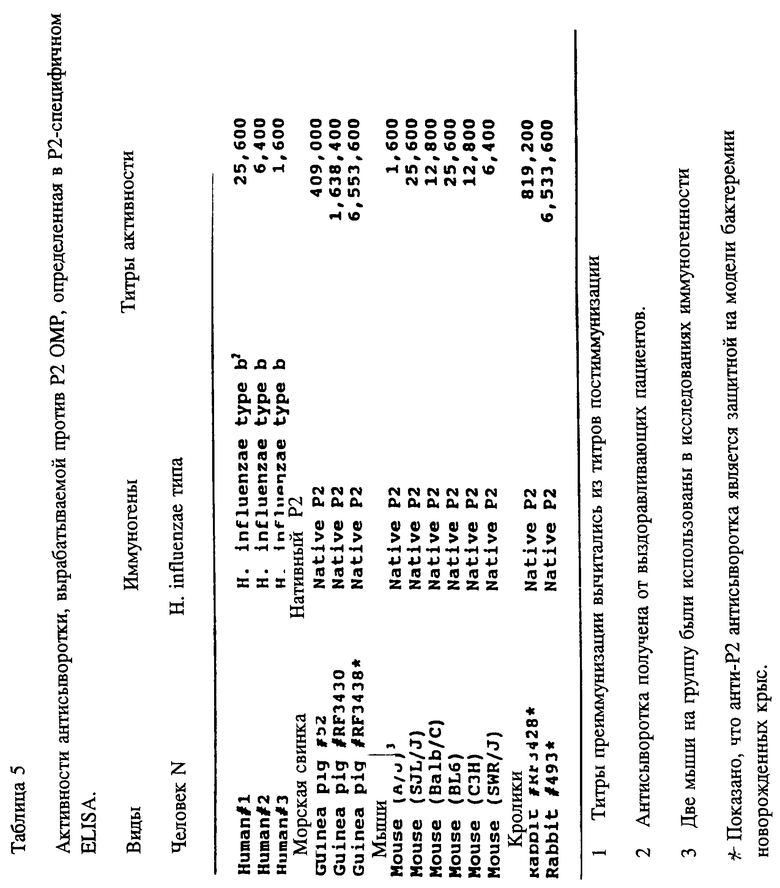
С другой стороны, настоящее изобретение включает существенно очищенную форму пептида, содержащего по крайней мере одну аминокислотную последовательность, относящуюся к консервативной последовательности P2-белка, пептид которой способен вызывать у млекопитающих образование поликлональных антител, которые узнают Hi in vitro. Эти P2-специфичные поликлональные антитела могут быть использованы в качестве компонентов тест-наборов для определения Hi в биологических образцах. Пептиды могут содержать, например, аминокислотные последовательности из следующих аминокислот: с 1 по 14, 125-150, 241-265, 263-289, 285-306, 302-319, 314-341 зрелого P2-белка Hib MinnA-штамма, как показано соответственно в табл. 2 (SEQ ID NOS : 16, 23, 28, 29, 30, 31 и 32 соответственно) или же любая часть, или их вариация их, сохраняющая иммуногенность.
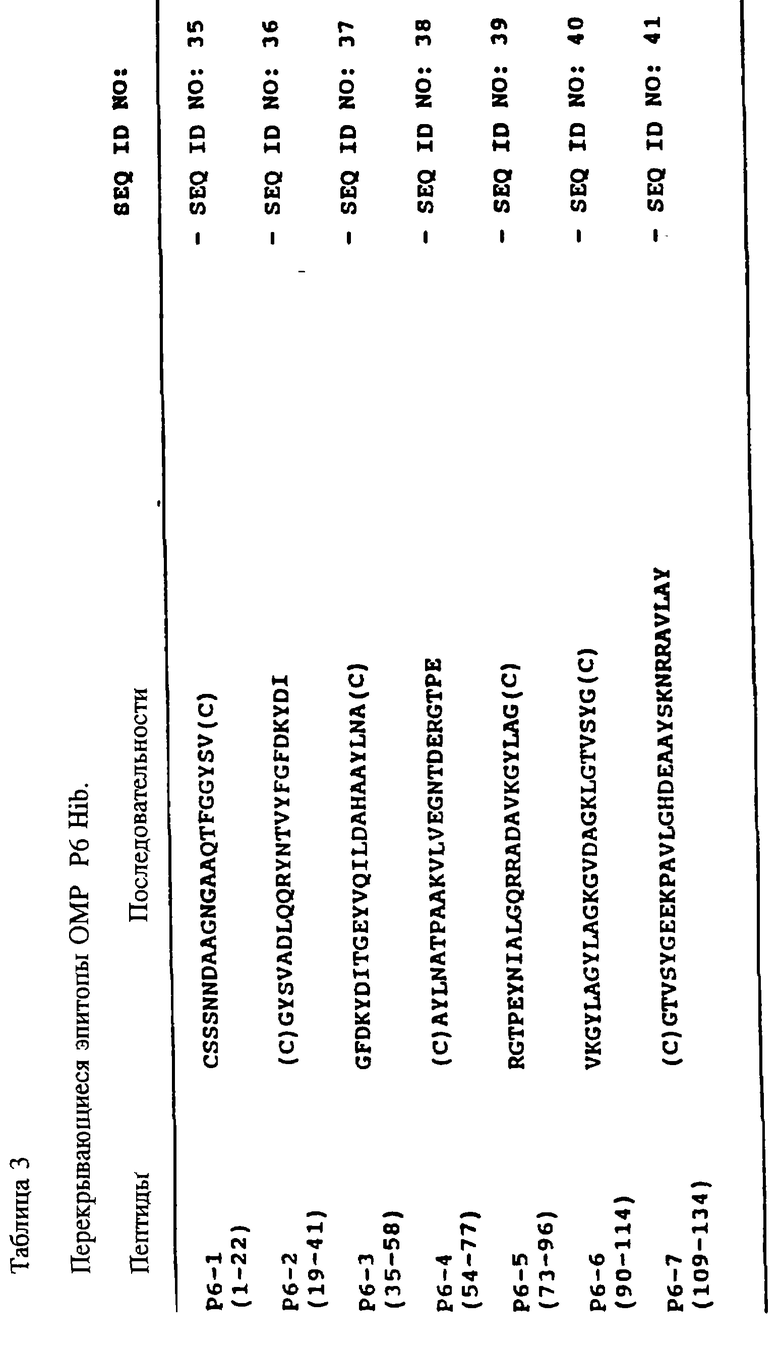
Другим воплощением настоящего изобретения является наличие существенно очищенной формы пептида, содержащего по крайней мере одну аминокислотную последовательность, относящуюся к консервативной последовательности P6-белка, пептид которой способен вызывать у млекопитающих образование поликлональных антител, которые узнают Hi vitro. Эти P6-специфичные поликлональные антитела могли бы быть использованы в качестве компонентов тест-наборов для определения H в любых биологических образцах. Пептиды могут содержать, например, аминокислотные последовательности из следующих аминокислот: с 1 по 22, 19-41, 35- 58, 54-77, 73-96, 90-114, и 109-134 зрелого P6-белка Hib MinnA штамма, как показано соответственно в табл. 3 (SEQ ID NOS : 35-41 соответственно) или же любая часть, или комбинация из этого, обладающая иммуногенностью.
Другим воплощением настоящего изобретения является наличие по крайней мере одного PI-пептида, который содержит по крайней мере одну аминокислотную последовательность, относящуюся к иммунодоминантному линейному В-клеточному эпитопу PI-белка. Эти эпитопы можно использовать в качестве антигенов-мишеней в диагностических наборах для определения анти-Hi антител, к примеру, защитных антител. Пептиды могут содержать, например, аминокислотные последовательности из следующих аминокислот: с 39 по 64, 103-137, 165-193, 248-283, 307-331, 400- 437 и 179-218 зрелого PI-белка Hib MinnA- штамма, как показано в табл. 1 (SEQ ID NOS : 12, 3, 4, 7, 9, 13 или 14 и 15, соответственно) или же любая часть, или комбинация их, обладающая иммуногенностью.
Другое воплощение настоящего изобретения включает наличие по крайней мере одного P2-пептида, который содержит по крайней мере одну аминокислотную последовательность, относящуюся к иммунодоминантному линейному B-клеточному эпитопу Р2-белка. Эти эпитопы можно использовать в качестве антигенов-мишеней в диагностических наборах для определения анти-Hi антител, к примеру, защитных антител. Пептиды могут содержать, например, аминокислотные последовательности из следующих аминокислот: с 53 по 81, 148-174, 241-265 и 314-342 зрелого P2-белка Hib MinnA-штамма, как показано в табл. 2 (SEQ ID NOS) : 20, 24, 28 и 32 соответственно) или же любая часть, или комбинация их, обладающая иммуногенностью.
Другим аспектом настоящего изобретения является наличие по крайней мере одного P6-пептида, который содержит по крайней мере одну аминокислотную последовательность, относящуюся к иммунодоминантному линейному B-клеточному эпитопу P6-белка. Эти эпитопы можно использовать в качестве исследуемых антигенов в диагностических наборах для определения анти-Hi антител, к примеру, защитных антител. Пептиды могут содержать, например, аминокислотные последовательности из следующих аминокислот: с 73 по 96, 90-114 и 109-134 зрелого P6-белка Hib MinnA-штамма, как показано в табл. 3 (SEQ ID NOS: 39, 40 и 41 соответственно) или же любая их часть, или комбинация, обладающая иммуногенностью.
Другим аспектом настоящего изобретения является наличие пептидов, которые можно определить как иммунодоминантные Т- клеточные эпитопы PI. Эти пептиды могут быть использованы в качестве аутологичных носителей для PRP, или как носители для аутологичных и гетерологичных B-клеточных эпитопов. Пептиды могут содержать, например, аминокислотные последовательности из следующих аминокислот: с 39 по 64, 226-253, 339-370 и 400-437 и 179-218 зрелого PI-белка Hib MinnA-штамма в соответствии, как показано в табл. 1 (SEQ ID NOS : 12, 6, 10 и 13 или 14 соответственно) или же любая часть, или комбинация их, обладающая иммуногенностью.
Другим свойством настоящего изобретения является наличие пептидов, которые можно определить как иммунодоминантные Т-клеточные эпитопы Р2. Эти пептиды могут быть использованы в качестве аутологичных носителей для PRP, или как носители для аутологичных и гетерологичных В-клеточных эпитопов. Пептиды могут содержать, например, аминокислотные последовательности из следующих аминокислот: с 125 по 150, 193-219, 219-244 и 241-265, зрелого Р2-белка Hib MinnA-штамма, как показано в табл. 2 (SEQ ID NOS : 26, 27 и 28 соответственно) или же любая часть, или их комбинация, обладающая иммуногенностью.
Другим свойством настоящего изобретения является наличие пептидов, которые можно определить как иммунодоминантные Т-клеточные эпитопы Р6. Эти пептиды могут быть использованы в качестве аутологичных носителей для PRP, или как носители для аутологичных и гетерологичных B-клеточных эпитопов. Пептиды могут содержать, например, аминокислотные последовательности из следующих аминокислот: с 19 по 41, 35-58, 73-96 и 109-134, зрелого P6-белка Hib MinnA-штамма в соответствии, как показано в табл. 2 (SEQ ID NOS: 36, 37, 39 и 41, соответственно) или же любая часть, или их комбинация, сохраняющая иммуногенность.
Следующий аспект, следовательно, это то, что настоящее изобретение предлагает иммуногенный конъюгат, включающий синтетический пептид, который имеет аминокислотную последовательность, относящуюся по крайней мере к одному T-клеточному эпитопу по крайней мере одного белка Haemophilus influenzae, привязанного по крайней мере к одному B-клеточному эпитопу.
Другой чертой изобретения является то, что здесь обеспечивается высокоэффективный химический синтетический способ получения синтетических PRP-олигомеров. Этот процесс является комбинацией твердофазного синтеза с использованием полиэтиленгликольмонометилового эфира (PEG) в качестве твердой подложки. Твердофазная подложка содержит большое количество химически активных функциональных групп, в диапазоне от приблизительно 200 до 500 мкмоль/г носителя в сравнении с от 30 до 35 мкмоль реакционных групп на 1 г традиционных подложек, таких как стекло с контролируемым размером пор. Только стехиометрические количества синтетического PRP повторяющегося звена в каждом цикле связывания в сравнении с 5-10-кратным молярным избытком в обычном твердофазном синтезе. Кроме того, настоящий новый способ является быстрым и эффективным в коммерческих целях, в противоположность твердофазному синтезу, который представляет собой трудоемкий, дорогостоящий и занимающий время процесс.
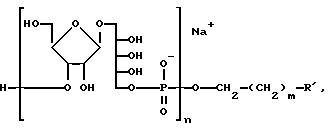
Продукт этого аспекта изобретения включает химически реактивный синтетический PRP олигосахарид, представленный следующей формулой: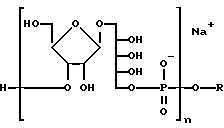
где n - целое число, предпочтительно от 3 до 20, и R - связующий фрагмент, определяемый -CH2-((CH2)-m -X, где m - целое число, предпочтительно от 3 до 5, а X - химически реактивная функциональная группа, такая как -CH2NH2, -CH2 SH2 или аминоактивная группа, такая как галоген, метаносульфонил, трифторметаносульфонил, или толуол-сульфонил и т.п. или фотоактивируемая группировка, такая как фенилазид, нитрофенил, бензилфенил и т.п. Реактивная функциональная группа позволяет синтетическому PRP связываться с другими молекулами.
Следующей чертой изобретения является то, что предложен иммуногенный конъюгат, куда входит синтетический углеводный антиген, связанный с по крайней мере одним синтетическим Т-клеточным эпитопом. Углеводный антиген может быть получен из бактериального материала, в частности синтетический рибозорибитолфосфат (PRP) олигомер.
Следующим воплощением настоящее изобретение обеспечивает иммуногенный конъюгат синтетического PRP и пептида, который способен вызывать образование анти-PRP антител с высоким титром у млекопитающих. Вакцина на основе конъюгата синтетического PRP с носителем имеет в составе вещество следующей формулы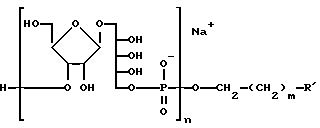
где n и m определены выше и R' - синтетический пептид, который содержит по крайней мере один Т-хелперный эпитоп, включающий последовательность GPKEPFRDYVDRFYK (SEQ ID NO 50) из HIV-I gag-белка P2 или Т-клеточный эпитоп из Hi OMP. Носитель может быть пептидом, который содержит Т-хелперный и В-клеточный эпитопы.
Следующим аспектом является то, что настоящее изобретение включает иммуногеный синтетический гликоконъюгат синтетического PRP-олигомера определенной длины и Hib PI-пептида, содержащего Т- и/или Т-В эпитопы. Размер синтетического PRP-олигомера составляет по крайней мере три повторяющихся звена PRP, но предпочтительным является повтор из шести звеньев. Пептиды могут содержать, например, аминокислотные последовательности из следующих аминокислот: с 39 по 64, 165-193, 189-218, 226-253, 339-370, 4OO-437, PI-белка Hib MinnA-штамма, как показано в табл. 1 (SEQ ID NOS : 12, 4, 5, 6, 10, 13 или 14 соответственно) или же любая часть, или их вариант, сохраняющий иммуногенность.
Один из аспектов настоящего изобретения состоит в том, что оно включает в себя иммуногенный синтетический гликоконъюгат синтетического PRP-олигомера определенной длины и P2-пептида, содержащего T- и/или T-B эпитопы. Размер синтетического PRP- олигомера составляет по крайней мере три повторяющихся звена PRP, но предпочтительным является повтор из шести звеньев. Пептиды могут содержать, например, аминокислотные последовательности из следующих аминокислот: с 125 по 150, 193-219, 219-244 и 241-265 зрелого P2-белка Hib MinnA-штамма, как показано в табл. 2 (SEQ ID NOS : 23, 26, 27 и 28 соответственно) или же любая часть, или вариант из них, проявляющий иммуногенность.
Один из аспектов настоящего изобретения состоит в том, что оно включает в себя иммуногенный синтетический гликоконъюгат синтетического PRP-олигомера определенной длины и P6-пептида, несущего T- и/или T-B-эпитопы. Размер синтетического PRP-олигомера составляет по крайней мере три повторяющихся звена PRP, но предпочтительным является повтор из шести звеньев. Пептиды могут содержать, например, аминокислотные последовательности из следующих аминокислот: с 19 по 41, 35-58, 73-96 и 109-134 зрелого P6-белка Hib MinnA-штамма в соответствии, как показано в табл. 3 (SEQ ID NOS : 36, 37, 39 и 41 соответственно) или же любая часть, или вариант из них, проявляющий иммуногенность.
Другое воплощение настоящего изобретения состоит в том, что предлагает концепцию того, что иммуногенность углеводного антигена, например, синтетического PRP может усиливаться при использовании множественной антигенной пептидной системы (MAP), в составе которой содержатся функциональные T-хелперные эпитопы как носитель для увеличения углеводной плотности внутри синтетического гликопептидного конъюгата. MAP могут содержать, например, (фиг. 1) последовательность
DIVAKIAYGRTNYKYNESDEHKQQLNG
(SEQ ID NO : 26),
которая относится к аминокислотам 193-219 P2-белка Hib MinnA-штамма или их любой фрагмент.
В другом аспекте настоящее внедрение включает конъюгат синтетического PRP-и липопептида (или смесь синтетических PRP и липопептидов), который способен вызывать клеточный и гуморальный иммуногенный отклик против Hi у млекопитающих. Липопептиды могут иметь, например, последовательность Трипальметил
CSSYAKAQVERNAGLIADSVKDNQITSALSTQC
(SEQ ID NO: 43),
которая относится к аминокислотам 165-193 PI-белка Hib MinnA-штамма или его любого фрагмента.
Следующая черта изобретения состоит в том, что оно включает иммуногенные химерные пептидные вакцины, которые состоят из идентифицированных T-B-эпитопов Р1, Р2 или Р6 Hib, и могут быть использованы для иммунизации против Hi-инфекции. Пептиды содержат, например, последовательности I (см в конце текста) или любую часть, или вариант, являющийся иммуногенным. Пептиды изобретения могут также иметь последовательность, соответствующую фрагментам, аналогичным изомерам штаммов Hi, отличных от MinnA.
Предложенные здесь новые синтетические пептиды и конъюгаты могут сформировать вакцину против заболевания, индуцированного патогеном, в частности, Н. influenzae, включающую по крайней мере один синтетический пептид и/или по меньшей мере один синтетический конъюгат, как описывается здесь, и физиологический носитель. Вакцину можно использовать для иммунизации хозяйского организма против патогенного заболевания путем введения ему эффективного количества вакцины. Далее вакцина может включать по крайней мере еще одну иммуногенную и/или иммуностимулирующую молекулу. B изобретении также предлагается метод иммунизации хозяйского организма против Hi-инфекции путем введения эффективного количества вакцины.
Пептиды, описанные в изобретении, могут быть далее модифицированы липидами подобно липопептидам или пришиты к синтетическим PRP (и/или полимеризованы) как синтетические липогликопептидные конъюгаты для производства модифицированных вакцин. Вакцины могут использоваться для иммунизации против инфекции путем введения млекопитающим, например, внутримышечно или перорально, или когда доставляются на слизистую с использованием микрочастиц, капсул, липосом и молекул, имеющих мишень, таких, как токсины и антитела.
Настоящее изобретение включает далее жизнеспособный вектор для доставки антигена, содержащий ген, кодирующий аминокислотную последовательность любого из синтетических пептидов, представленных здесь. Природный вектор может быть вирусного происхождения, например, поксовирусным, аденовирусным, полиовирусным или ретровирусным вектором. Также может быть бактериальным вектором, допустим, из сальмонеллы и микобактерий. Природный вектор может быть включен в вакцину, куда входит также и физиологически приемлемый носитель.
Как отмечено ранее, синтетические пептиды, предлагаемые здесь, могут быть использованы как диагностические реагенты в методах, определения заражения Haemophilus influenzae.
Антитела, которые вырабатываются против любого синтетического пептида и конъюгатов, описываемых здесь, включены в изобретение.
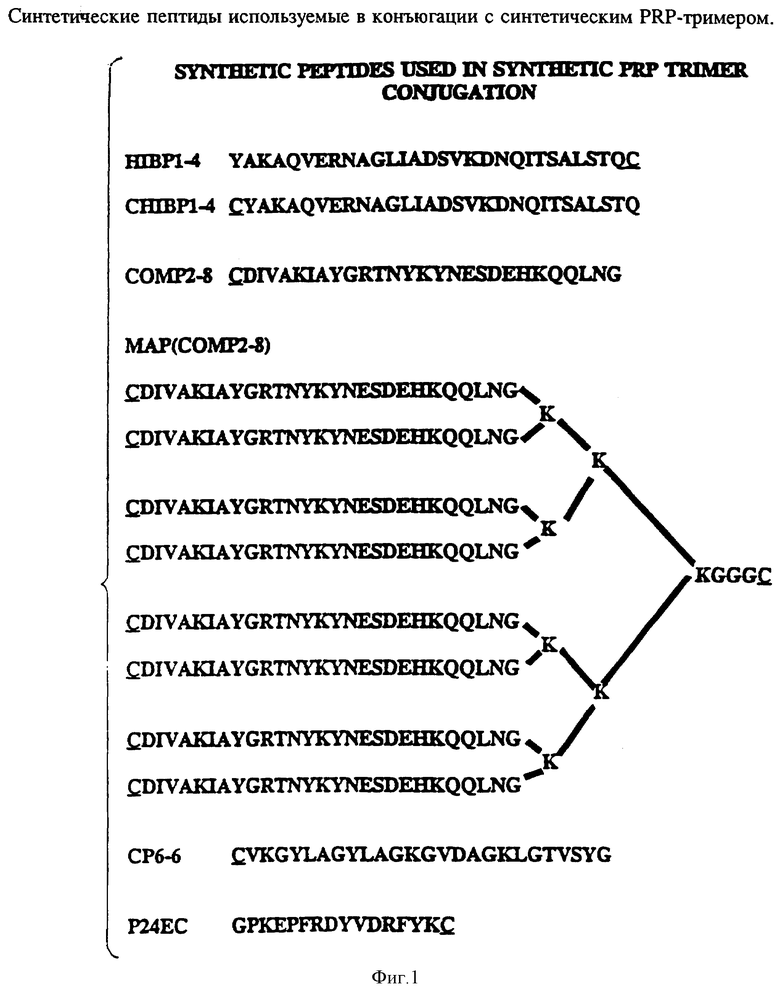
На фиг. 1 показаны аминокислотные последовательности пептидных носителей, использованных в исследовании конъюгатов синтетических PRP и пептидов, о которых здесь говорится. Как следует из фиг. 1, синтетические пептиды, обозначенные там, имеют следующие SED ID NOS :
Пептид - SEQ ID NOS:
HIBP-4 - 51
CHIBPI-4 - 52
COMP2-8 - 53
MAP (COMP2-8) - 54
CPC-6 - 55
PZ4EC - 56
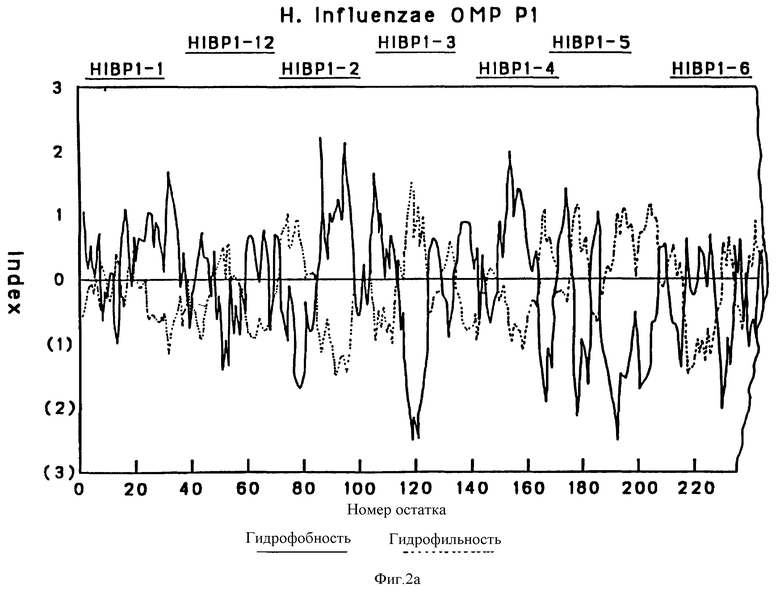
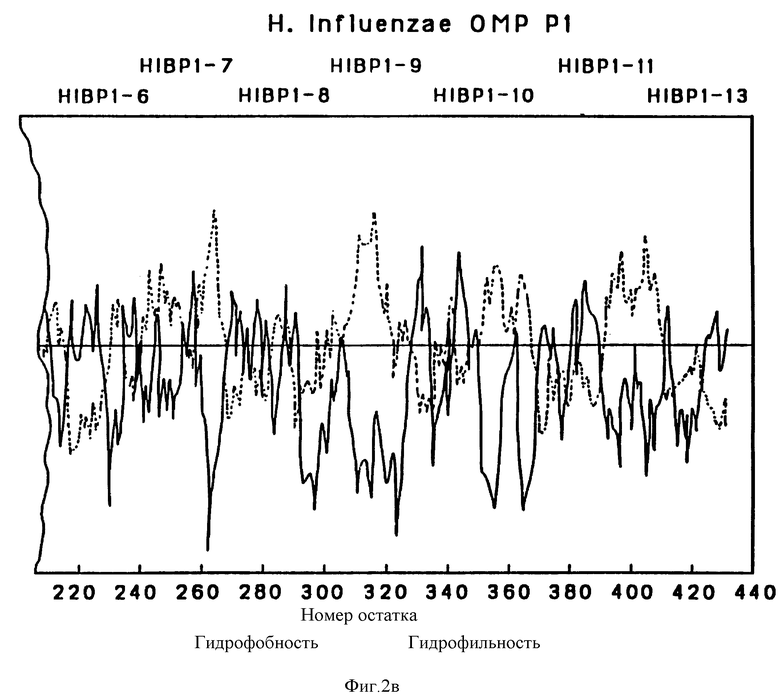
На фиг. 2 представлена предсказанная по общепринятому алгоритму структурного анализа структура OMP P1.
Профиль гидрофобности предсказан Hopp [30]. Значения получены из средних для гептапептидных фрагментов и обозначены в центре каждого сегмента.
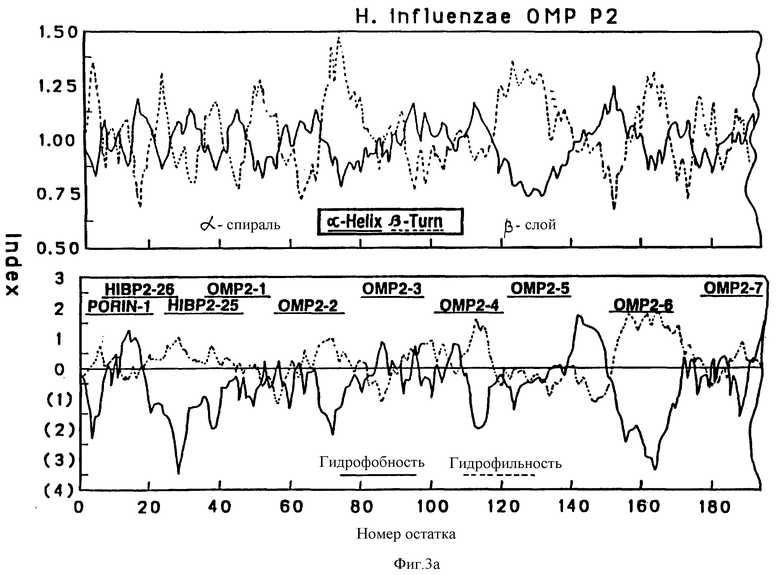
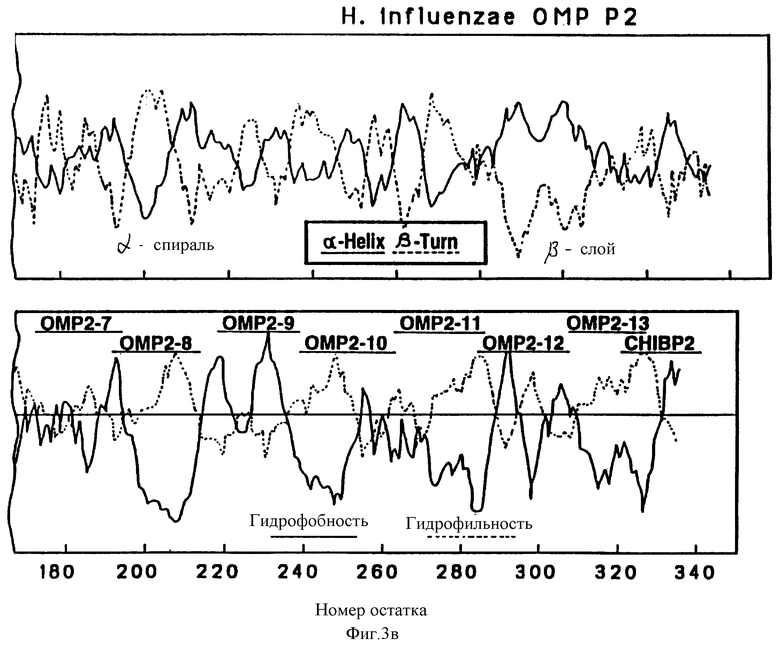
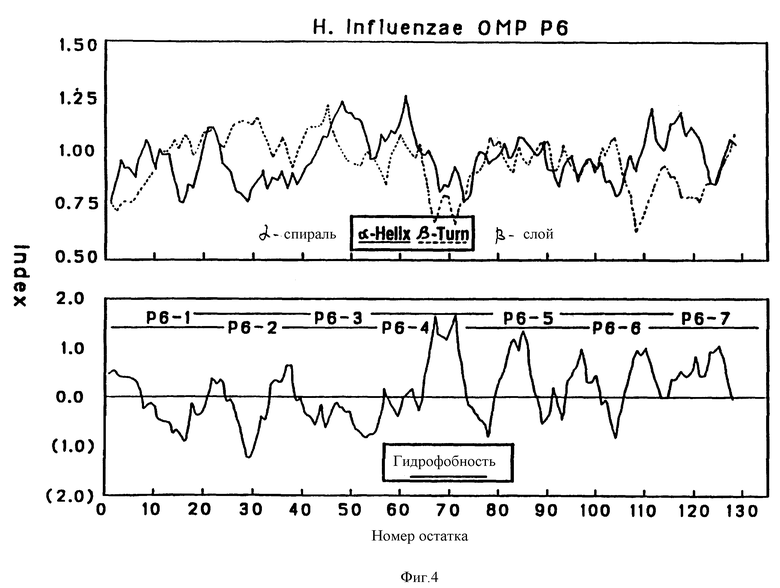
На фиг. 3 и 4 показаны соответственно предсказанные структуры Р2 и Р6 OMP, полученные по традиционному алгоритму структурного анализа. Верхняя часть, структурный анализ вторичной структуры по локальным средним потенциалам альфа-спирали и бета-слоев по Chou и Fasman [29]. Нижняя часть - профили гидрофобности, предсказанные Hopp и Woods (30). Значения получены из средних для гептапептидных фрагментов и обозначены в центре каждого сегмента.
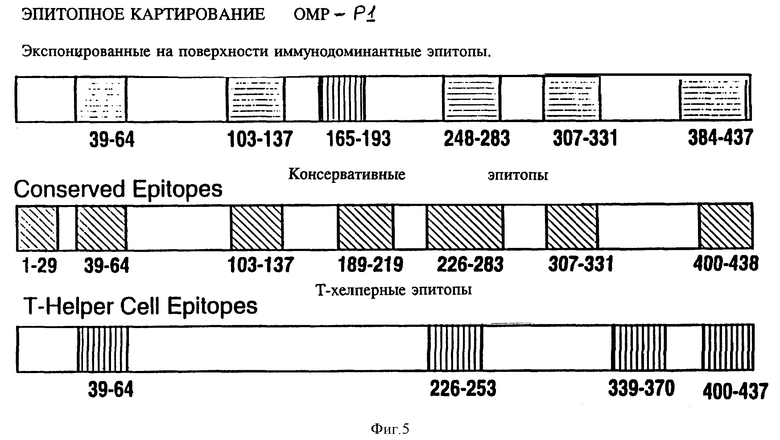
На фиг. 5 представлено диаграммное воспроизведение иммунодоминантных B- и T-клеточных эпитопов Hib OMP P1;
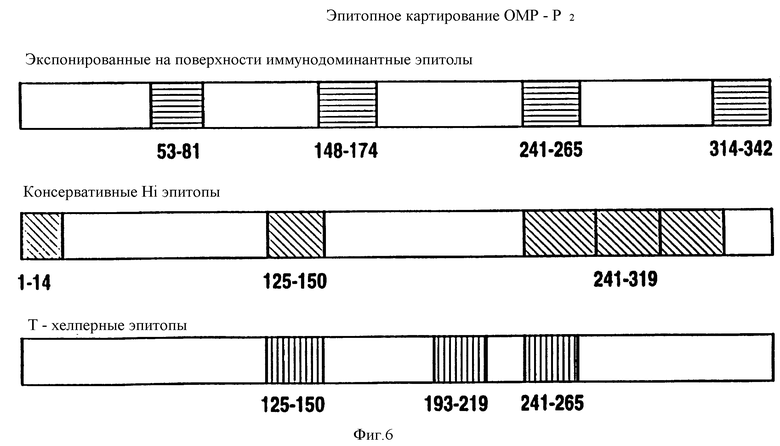
на фиг. 6 - диаграммное воспроизведение иммунодоминантных B- и T-клеточных эпитопов Hib OMP P2;
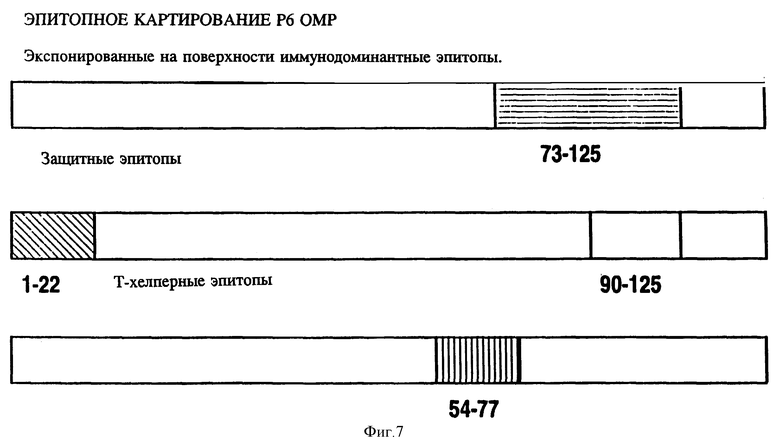
на фиг. 7 - диаграммное воспроизведение иммунодоминантных B- и T-клеточных эпитопов Hi OMP Р6;
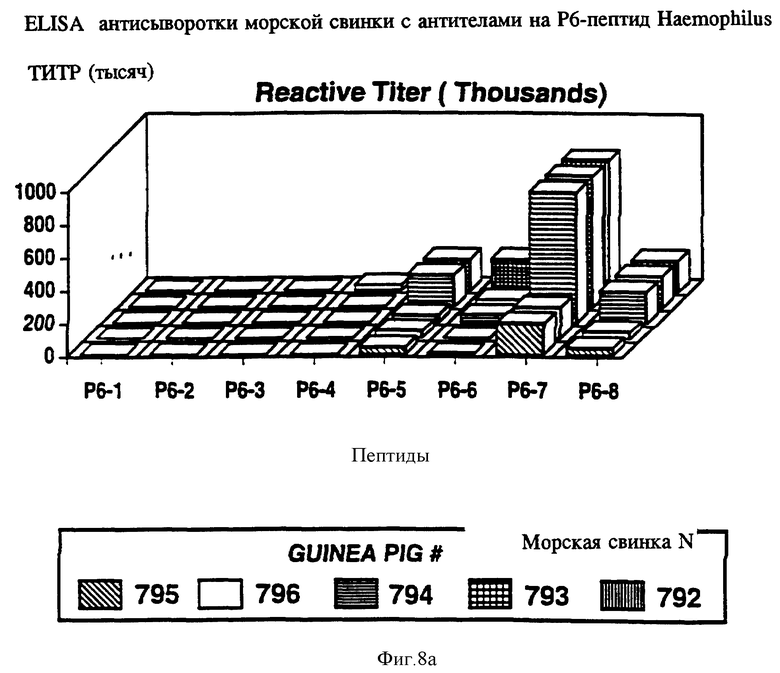
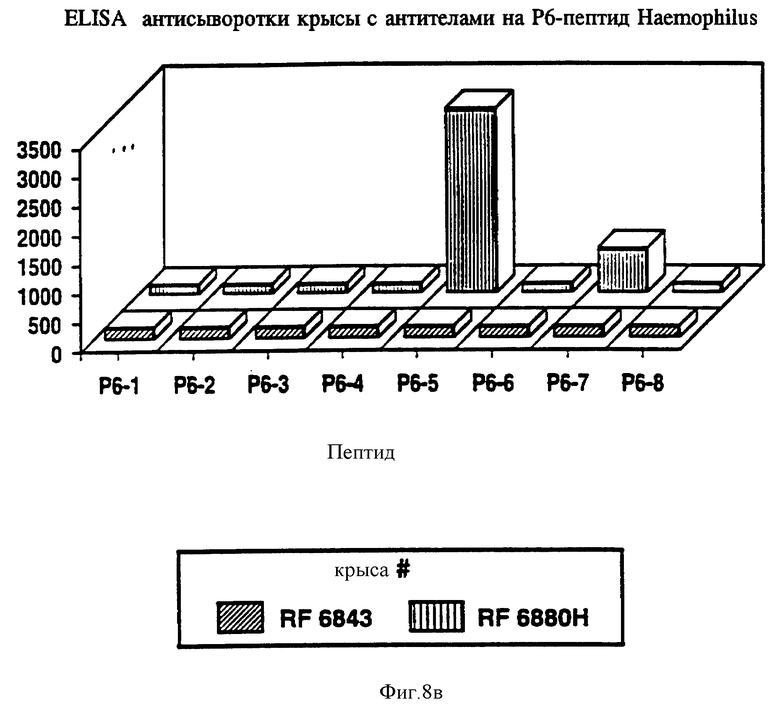
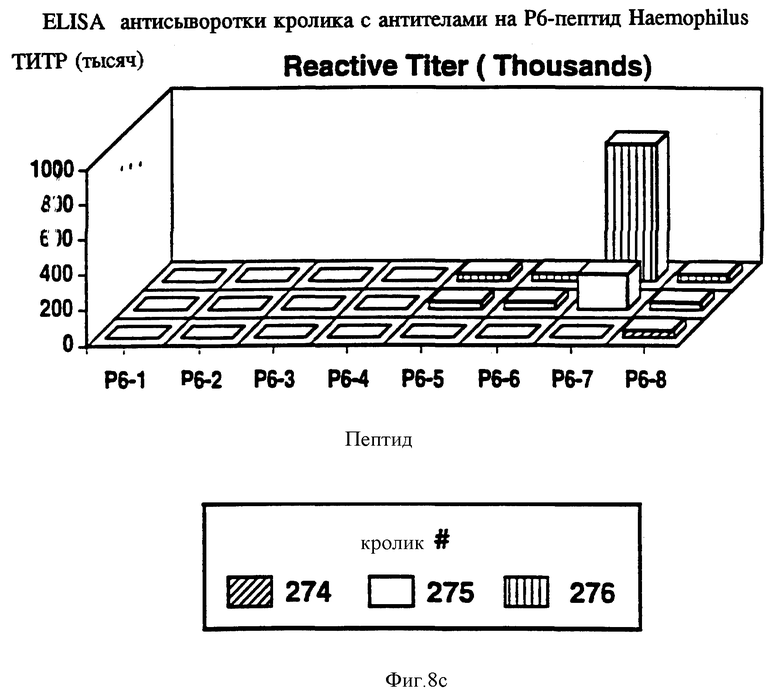
на фиг. 8 показана активность P6-пептидов в отношении анти-P6 антител морской свинки, крысы и кролика в ELISA;
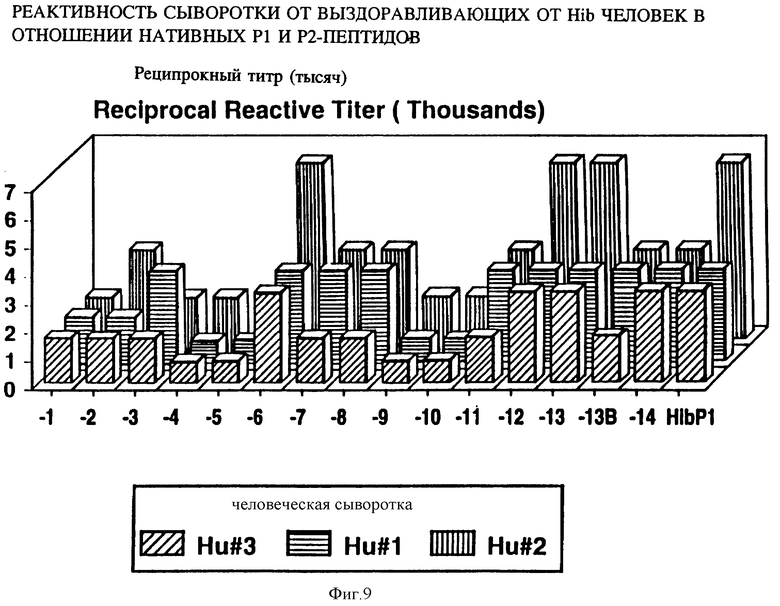
на фиг. 9 показана активность P1-пептидов в отношении сывороток от трех выздоравливающих человек в ELISA;
на фиг. 10 - активность Р2-пептидов в отношении сывороток от трех выздоравливающих человек в ELISA;
на фиг. 11 представлен пролиферативный ответ у Р1-специфичной мышиной T-клеточной линии на P1-пептиды с иммунодоминантными T-клеточными эпитопами, выделенными звездочкой;
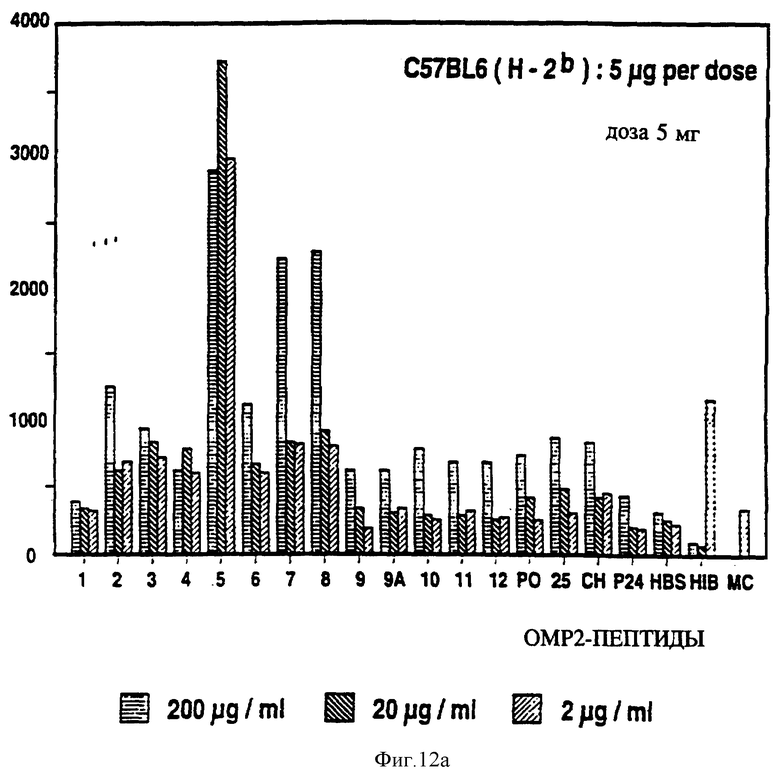
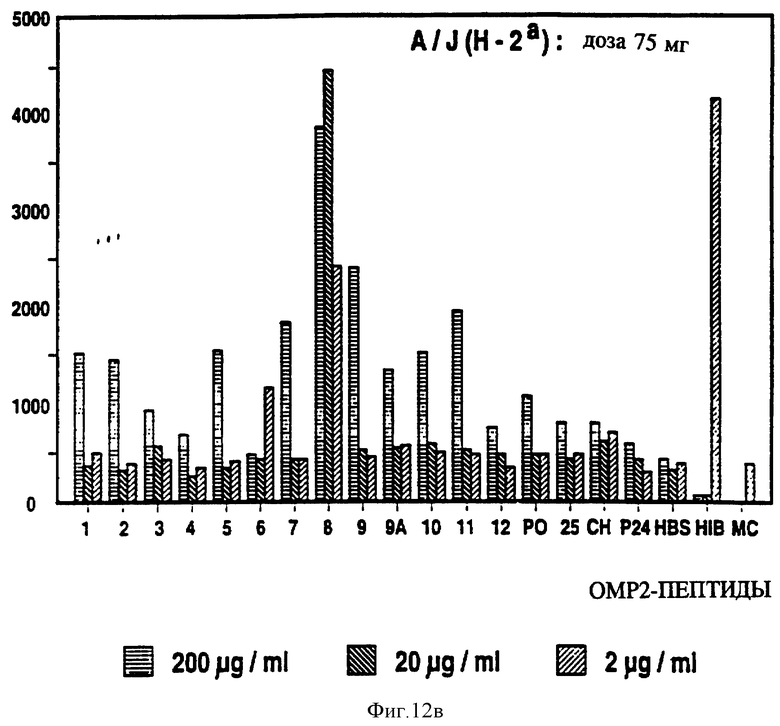
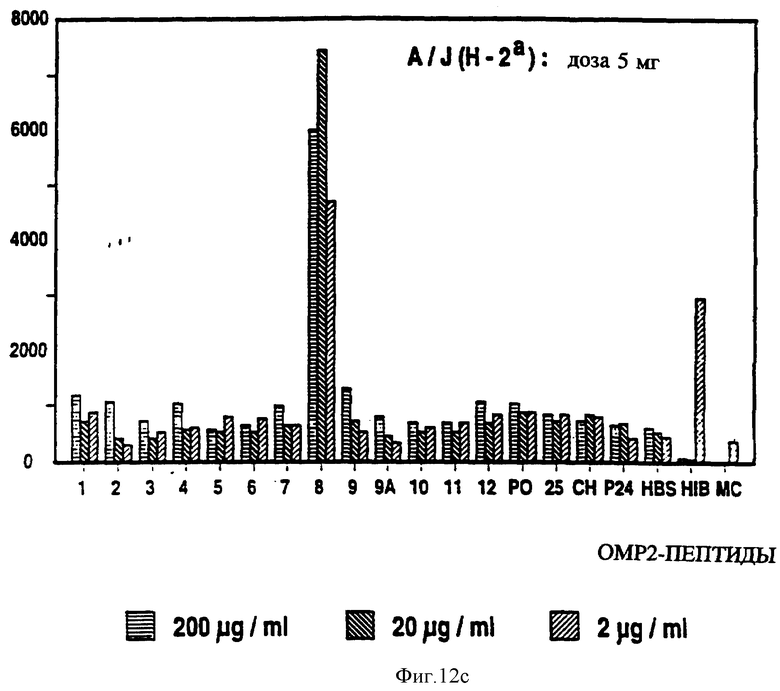
на фиг. 12 представлен пролиферативный ответ в Р2-спе- нифичной мышиной T-клеточной линии на Р2-пептиды с иммунодоминантными T-клеточными эпитопами, выделенными звездочкой;
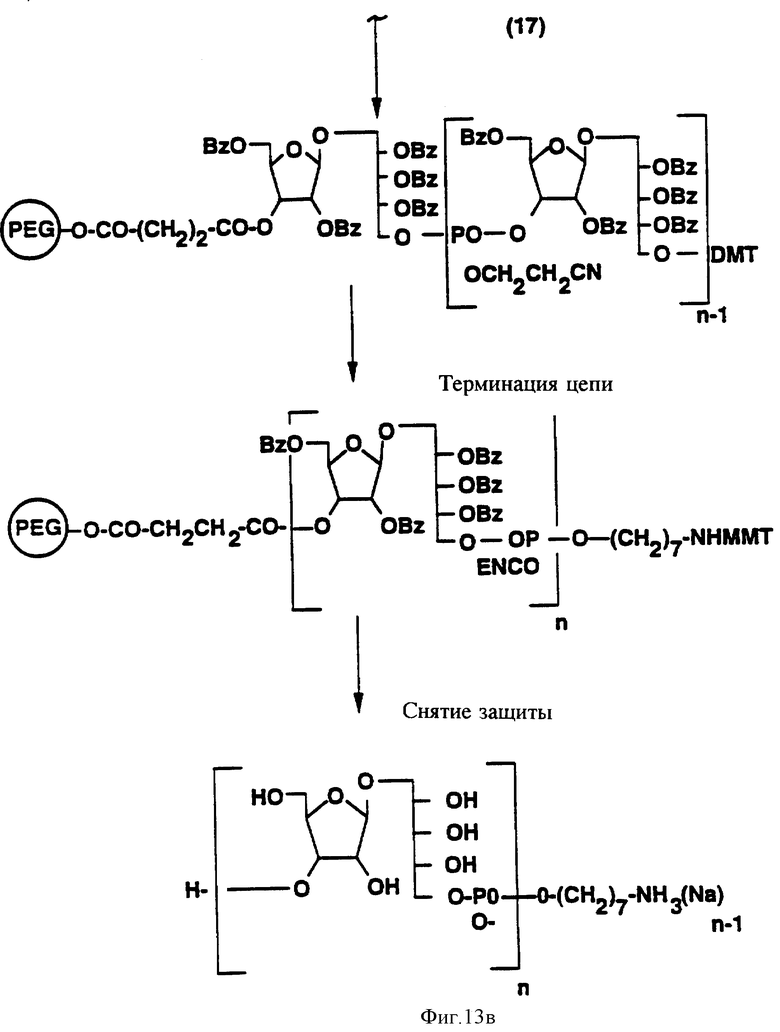
на фиг. 13 представлена последовательная схема синтеза PRP с использованием PEG как твердой фазы;
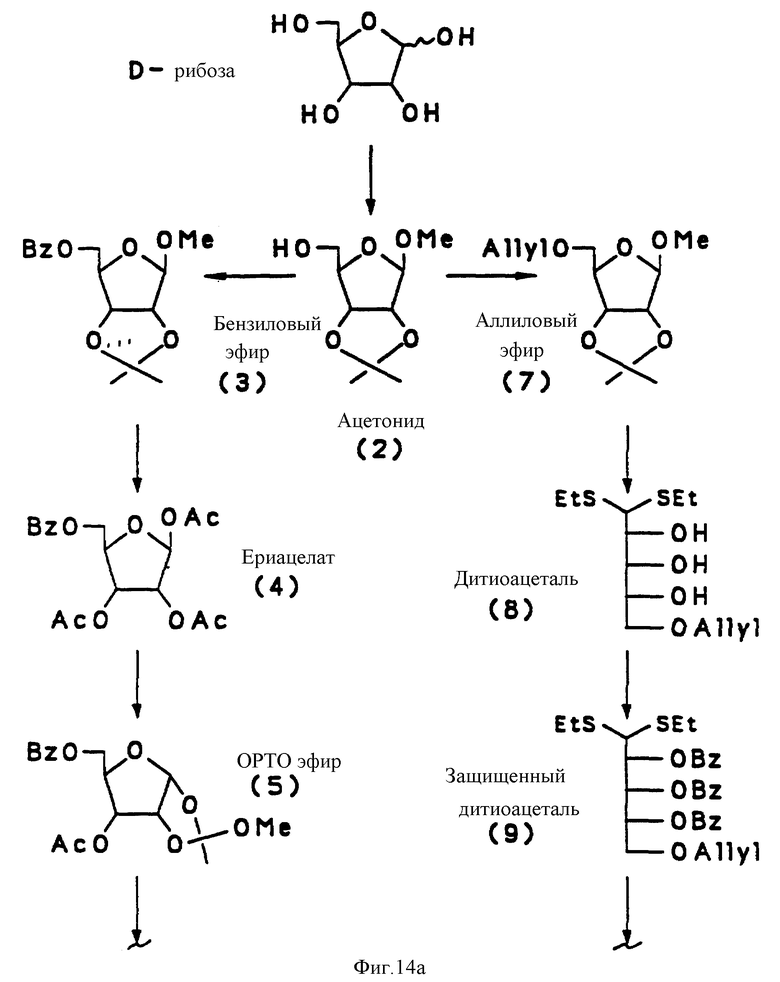
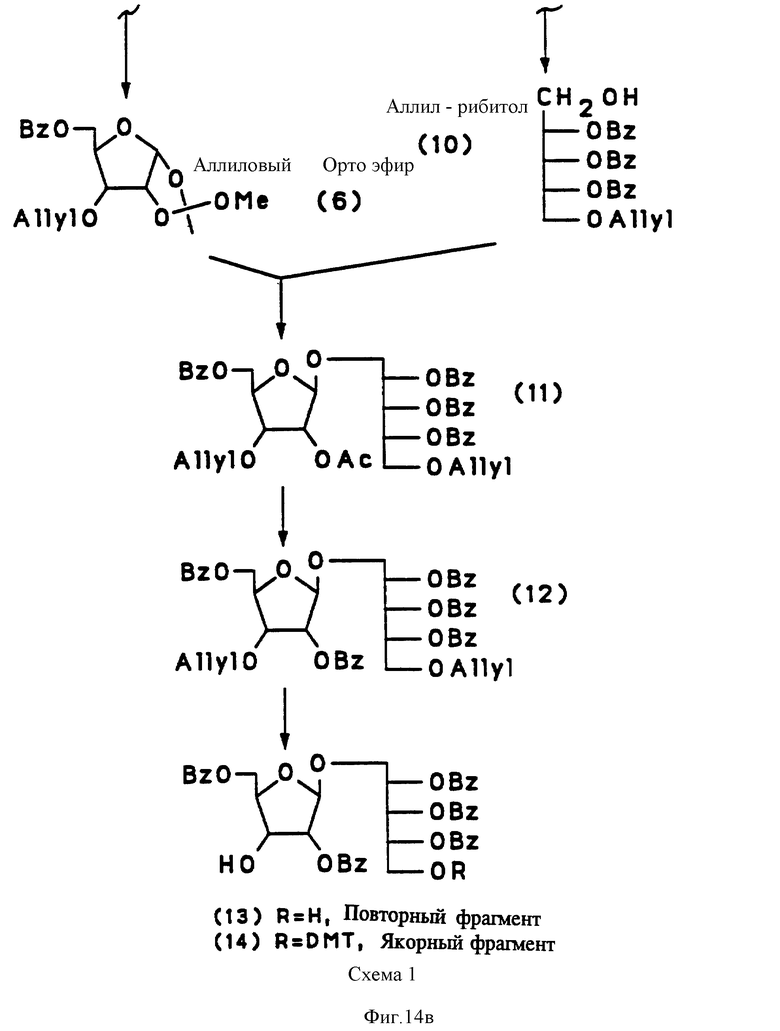
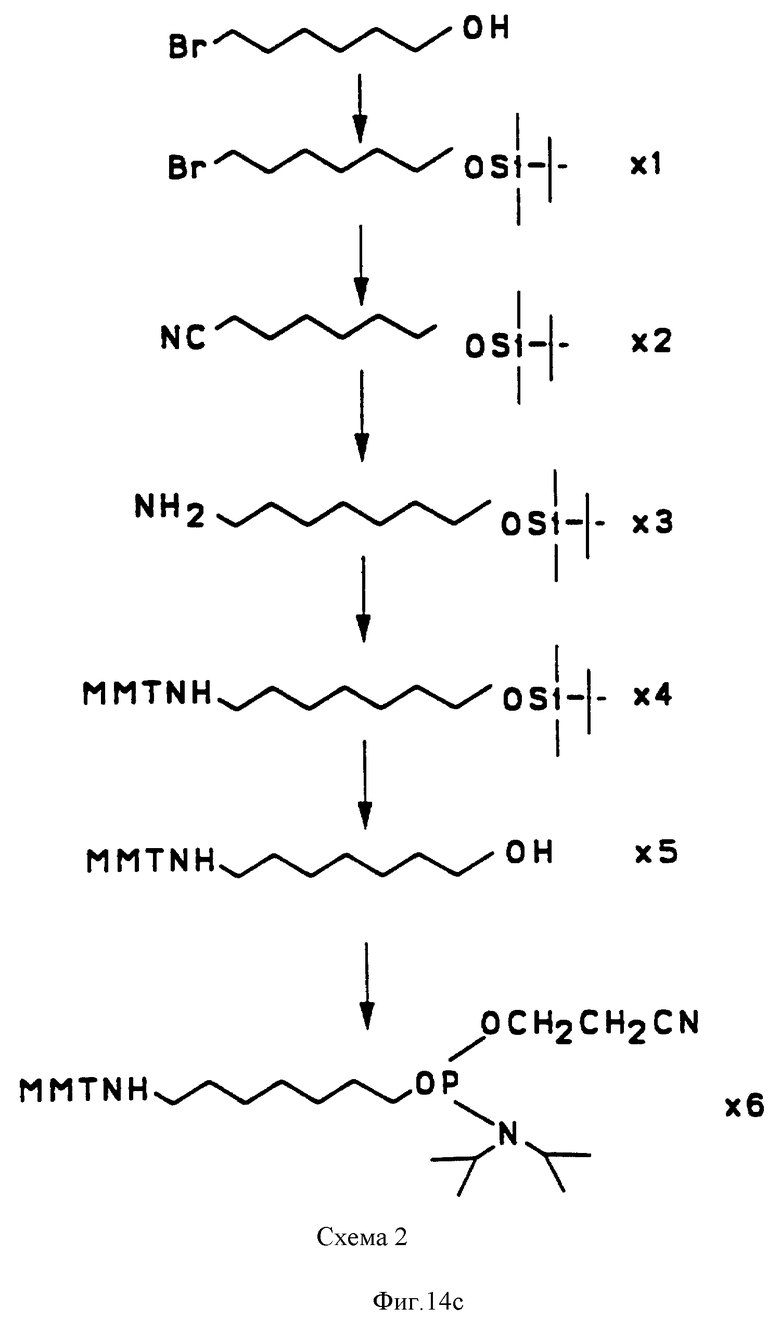
на фиг. 14 представлена последовательная схема синтеза PRP-интермедиатов (Br, бензил; Ac, ацетил; ETS, этилтио; AIIye, аллил; Me, метил; DMT, 4,4'-деметоксилтритил; NCE, цианоэтил; MMT, 4-метокситритил;
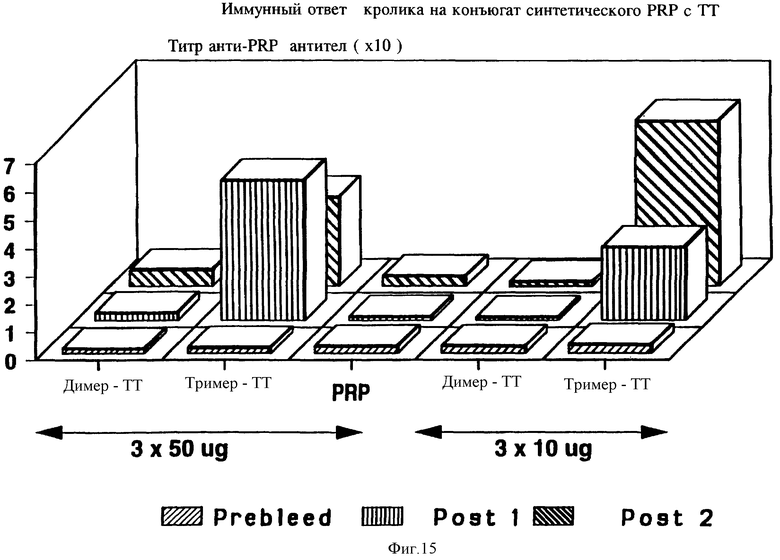
на фиг. 15 показан иммунный ответ кролика на конъюгат синтетических PRP-димера и тримера с тетанусным токсином;
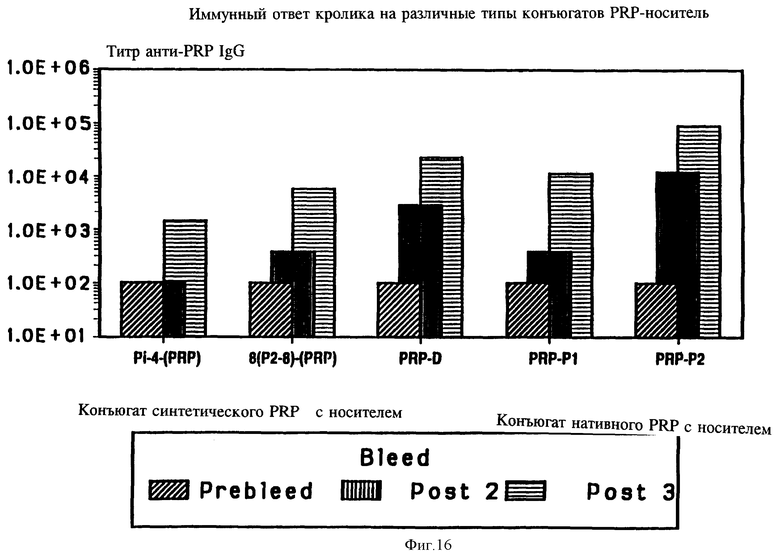
на фиг. 16 - иммунный ответ кролика на различные типы конъюгатов PRP с носителями;
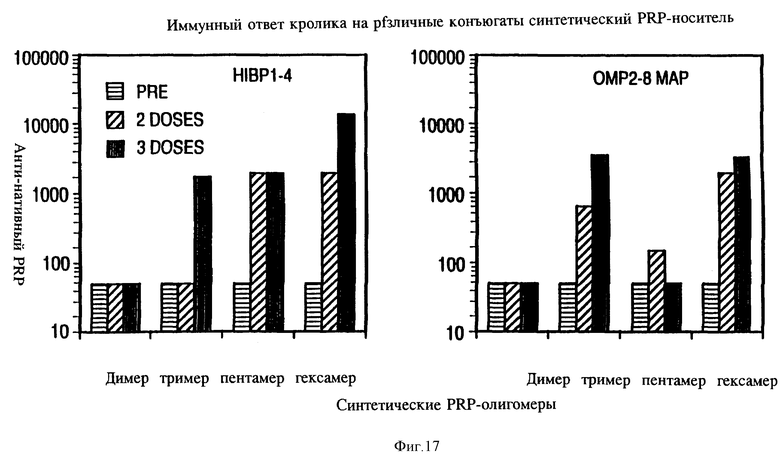
на фиг. 17 - иммунный ответ кролика на различные типы конъюгатов синтетических пентамеров и гексамеров с HibP1-4 и MAP OMP.
Детальное описание изобретения.
Настоящее изобретение относится к идентификации иммуногенных эпитопов OMP Hib, новых конъюгатов синтетических PRP с пептидами и приготовленных таким образом вакцин. Эти новые иммуногенные агенты подготавливаются путем химического синтеза пептидов, имеющих общие антигенные детерминанты с OMP Hib P1, P2 и P6. Пептиды или липопептиды используются либо независимо, либо привязанными к синтетическим PRP-олигомерам в качестве вакцин. Также они могут быть полимеризованы для производства других вакцин. Такие вакцины могут использоваться для иммунизации против в Hi -инфекции, если их вводить млекопитающим, например, внутримышечно или парентерально или доставлять к слизистой оболочке, используя микрочастицы, капсулы, липосомы и молекулы имеющие мишень, такие как токсины и антитела.
Первоочередные черты изобретения, которые наряду с последующими примерами служат для объяснения принципов внедрения. Для ясности изложения, но не для ограничения изобретение разделено на следующие пункты:
(i) предсказание эпитопов и пептидный синтез;
(ii) идентификация и характеристика иммунодоминантных B-клеточных эпитопов OMP Hib P1, P2 и P6 с использованием синтетических пептидов;
(iii) идентификация и характеристика иммунодоминантных T-клеточных эпитопов OMP Hib Р1, Р2 и Р6 с использованием синтетических пептидов;
(iv) иммуногенность OMP пептидов Hib;
(v) твердофазный углеводный синтез PRP-олигомеров с использованием PEG как подложки;
(vi) конъюгация синтетических PRP-олигомеров с пептидами и иммунохимическая характеристика гликоконъюгатов;
(vii) использование Hi конъюгатных PRP-пептидных вакцин.
Предсказание эпитопов и пептидный синтез.
Чтобы картировать иммунодоминантные T-клеточные или B-клеточные эпитопы Hi OMP, 13, 17 и 7, перекрывающиеся пептиды, охватывающие большую часть последовательностей Р1, Р2 и Р6-белков (табл. 1, 263), соответственно были синтезированы с использованием t-Вос твердофазного пептидного синтеза, как подробно описано в примере 12. Длина пептидов была выбрана на основании высокого показателя гидрофильных бета-слоев, оцененного из предсказательного анализа вторичной структуры согласно принятым алгоритмам (29-31) (фиг. 2, 3 и 4). Вероятно эти пептиды Van экспонированы на поверхности и являются антигенными. Пептиды длиной более 25 остатков были выбраны, чтобы маскировать нативные эпитопы как предлагалось в работе Regenmortel [32]. Иногда в целях сайт-специфической конъюгации к цепи добавлялся дополнительный цистеиновый остаток либо к N-концу, либо к C-концу пептида.
Идентификация и характеризация иммунодоминантных эпитопов OMP Hi Р1, Р2 и Р6 с использованием синтетических пептидов.
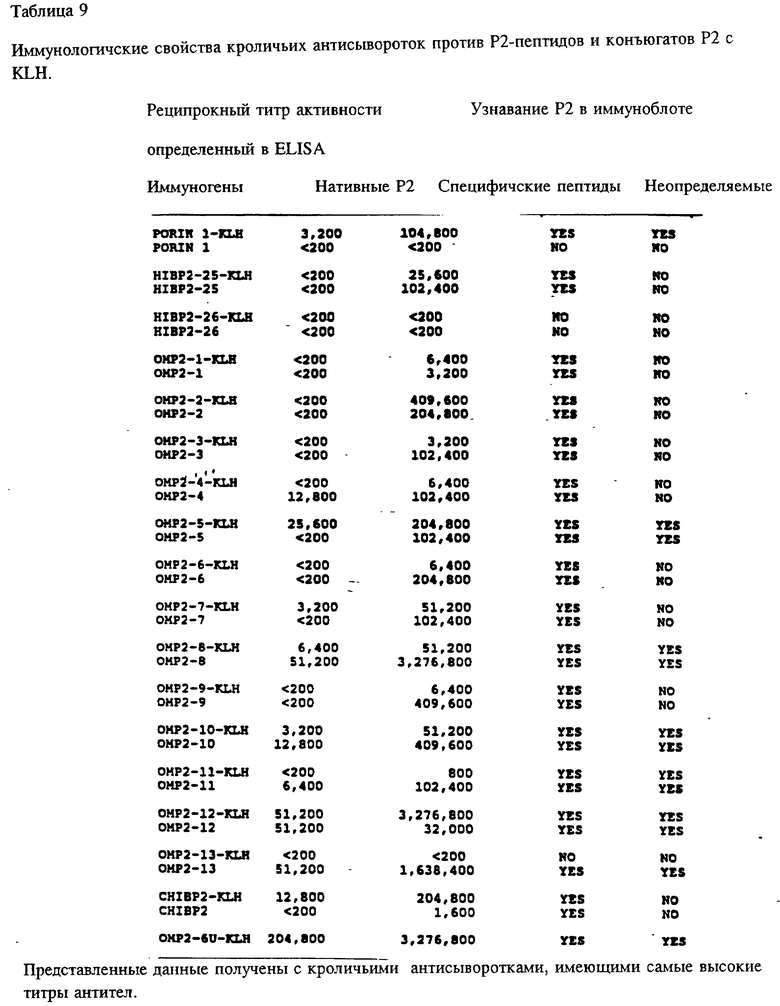
Чтобы идентифицировать иммунодоминантные B-клеточные эпитопы OMP Hib кроликов, морских свинок и мышей различных гаплотипов (H-2, H-2, H-2, H-2, H-2, H-2), иммунизировали очищенным Р1, Р2 или P6-белком в присутствии адъюванта Фрейнда. После первичной и вторичной иммунизации у всех животных развивался сильный и специфичный ответ анти-OMP антителами как было проверено в Р1-, Р2- и Р6-спепифичном ELISA (табл. 4, 5, 6) и иммуноблотном анализе. Как ранее показано Granoff Munson [6] , кроличьи Анти- PI, анти-Р2 и анти-P6 антитела постоянно защищают новорожденных крыс от заражения неубитыми Hib. Анти-Р2 антитела морской свинки также обладали защитными свойствами в этой модели.
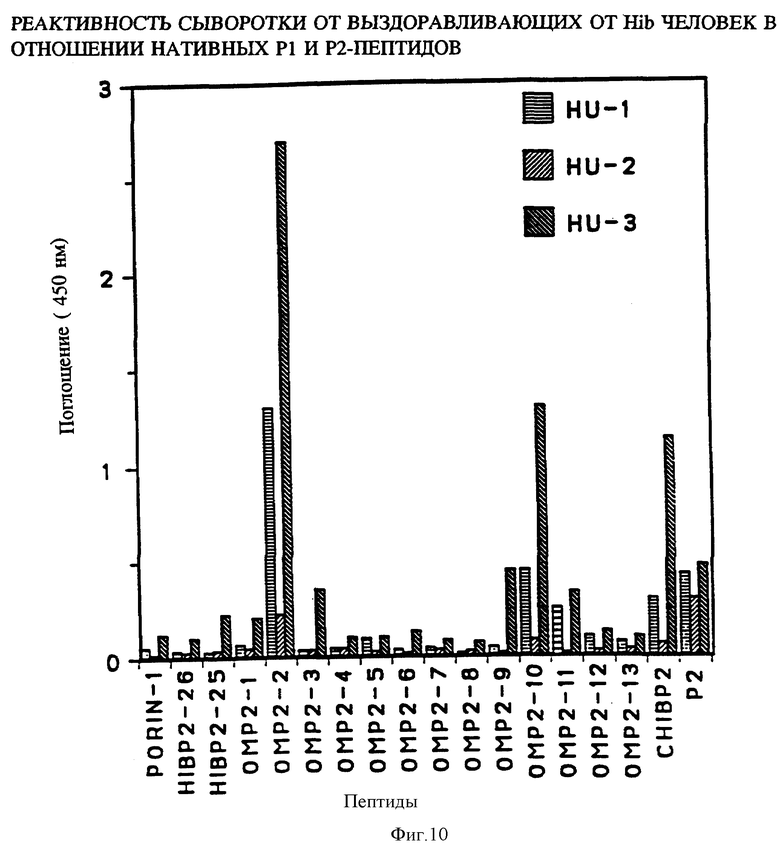
Чтобы картировать линейные B-клеточные эпитопы OMP Hib, перекрывающиеся пептиды, охватывающие большую часть последовательностей Р1, Р2 и Р6, были нанесены на планку ELISA и протестированы с различными анти-P1, анти-Р2 и анти-Р6 антисыворотками, как описано в примере 17. На фиг. 5, 6, 7 суммированы результаты. Было найдено, что иммунодоминантные линейные B-клеточные эпитопы Р1 локализованы среди пептидных последовательностей, относящихся к аминокислотным остаткам 39-64, 103-137, 165-193, 248-283, 307-331, 400-437 и 179-218 зрелого P1-белка Hib MA-штамма (см. табл. 1). Иммунодоминантные линейные B-клеточные эпитопы Р2 локализованы среди пептидных последовательностей, относящихся к аминокислотным остаткам 53-81, 148-174, 241-265 и 314-342 зрелого Р2-белка Hib MA штамма (см. табл. 2). Аналогично, P6-пептиды, содержащие иммунодоминантные линейные B-клеточные эпитопы есть аминокислотные остатки 73-96, 90-114 и 109-134 зрелого Р6-белка Hib MA-штамма (см. табл. 3). Интересно, что сыворотки трех выздоравливающих человек стабильно реагировали с Р1 и Р2 иммунодоминантными эпитопами, описанными выше (фиг. 9 и 10). Кроме того, штамм-специфичные протективные в отношении Р1 B-клеточные эпитопы были картированы в районе, относящемся к 165-193 остаткам P1-белка. Эти результаты свидетельствуют, что B-клеточные эпитопы, описываемые выше, могут использоваться как антигены-мишени в диагностических наборах для определения анти-Hi антител в биологических жидкостях.
Идентификация и характеристика иммунодоминантных T-клеточных эпитопов OMP Hib Р1, Р2 и Р6 с использованием синтетических пептидов.
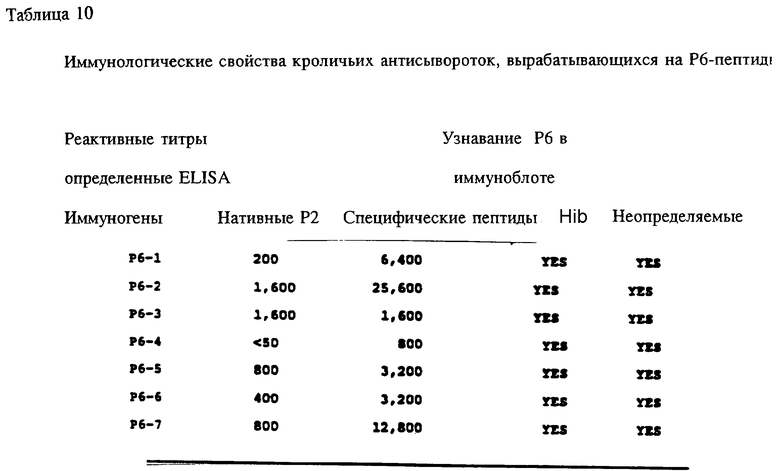
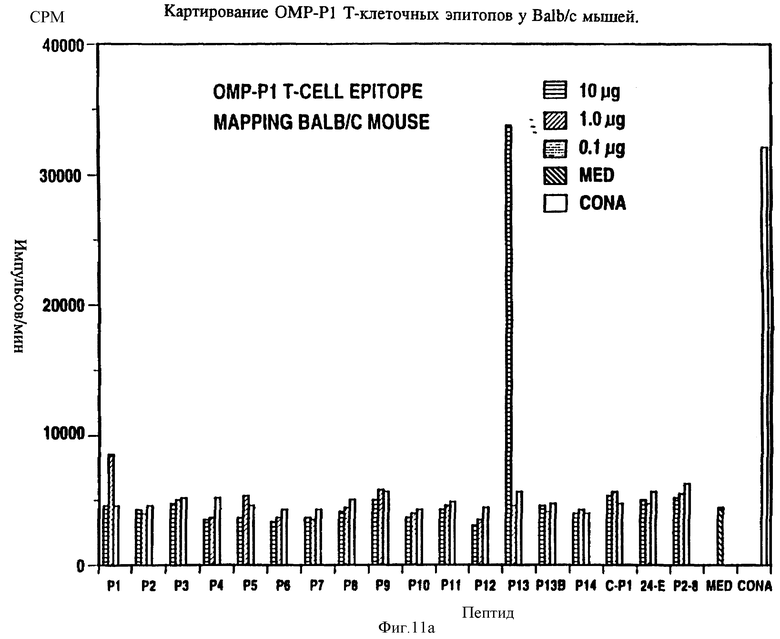
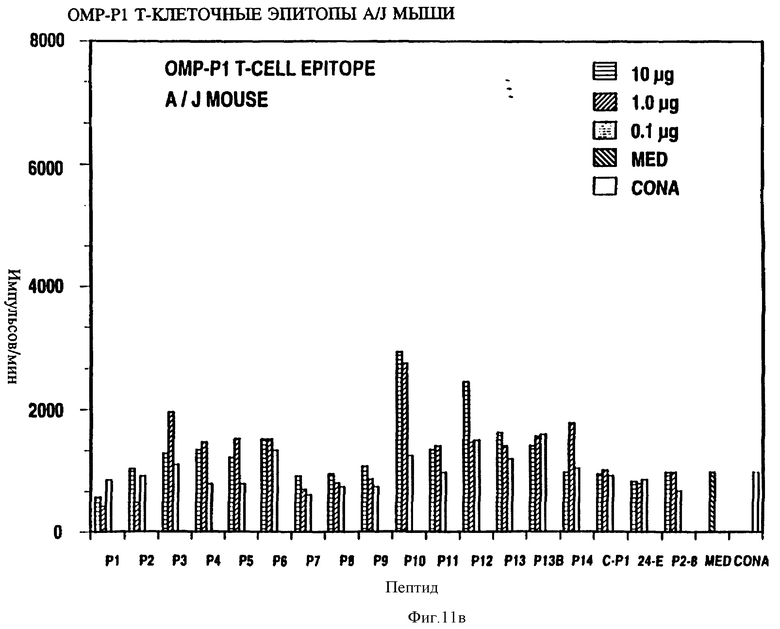
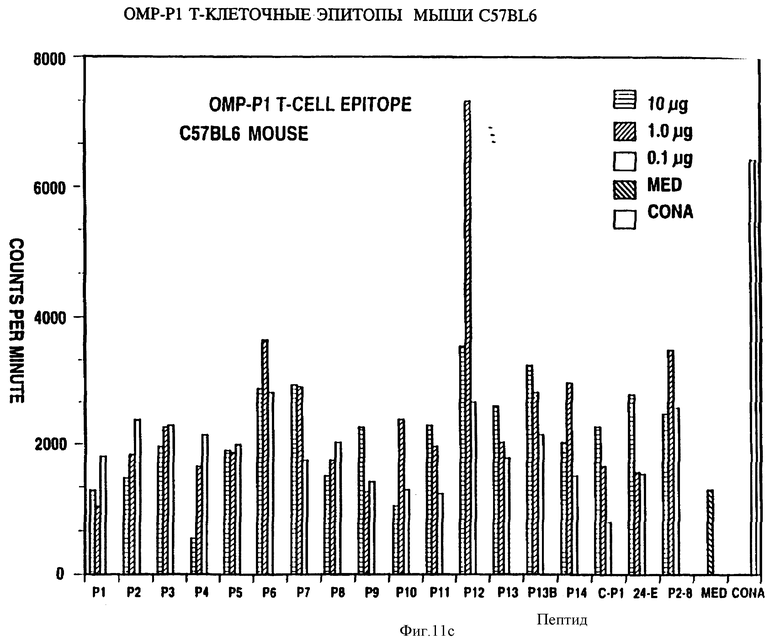
Специфичные к OMP Hib T-клеточные эпитопы были определены с использованием Р1, Р2 и Р6 пептидов и T-клеточных линий из панели с различных мышей, иммунизированных нативными OMP. Лимфоцитные пролиферативные ответы OMP-специфичных Т-клеточных линий на перекрывающиеся Р1-пептиды (13 пептидов), Р2-пептиды (17 пептидов), и Р6- пептиды (7 пептидов) были определены в обычном анализе пролиферации, как описывается в примере 19. Результаты (фиг. 11 и 12 и табл. 7) показали, что определенные синтетические пептиды только вызывают пролиферативный ответ и узнавание E-клеточных эпитопов, рестриктированных MHC. Синтетические пептиды, которые относятся к остаткам 33-64, 226-253, 339-370 и 400-437 Р1; остаткам 125-150, 193-219, 219-244 и 241-264 Р2; остаткам 19-41, 35-58, 73-96 и 109-134 Р6, при представлении в составе соответствующего мышиного MHC, являются, как показано, высокостимуляторными для всех соответствующих OMP-специфичных мышиных T-клеточных линий. Следовательно, эти иммунодоминантные T-клеточные эпитопы могут быть использованы как аутологичные носители для PRP и/или OMP B- клеточных и T-клеточных эпитопов для увеличения их иммуногенности.
Иммуногенность OMP пептидов Hib.
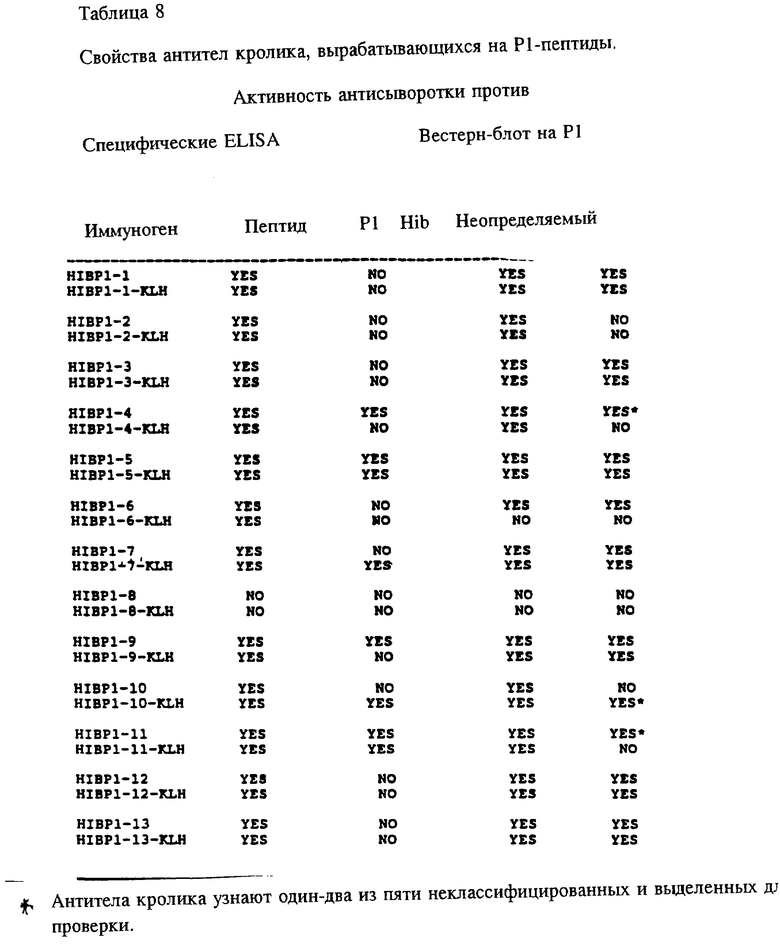
Чтобы определить, могут ли синтетические OMP-пептиды служить возможными вакцинами, на иммуногенность оценивались раздельно свободные пептиды и KLH-конъюгаты. Кроличьи антипептидные сыворотки были тестированы на реактивность в отношении иммунизирующих пептидов и их родительских белков в ELISA и иммуноблотном анализе. Как показано в табл. 8, все анти-PI пептид-сыворотки, кроме тех, которые получены в ответ на HIBPI-8 или конъюгат HIBPI-8-KIH, специфичны по отношению к соответствующим иммунизирующим пептидам в ELISA. Индукция высокого титра пептидспецифичных lgG-антител свободными пентидами свидетельствует в пользу того, что пептид включает и функциональную T-хелперную детерминанту и B-клеточный эпитоп(в). В дополнение, анти-HIBPI-4, анти-HIBPI-5, анти-HIBPI-7, анти-HIBPI-9, анти-HIBPI-10, анти-NIBPI-11, анти-HIBPI-14-антитела узнавали Р1, во всех использованных методах, которые указывают на антигенность этих районов, и свободны при взаимодействии с антителами. Поскольку для этих пептидов показано, что они содержат потенциальные T-хелперные детерминанты и пептид-KLH-конъюгаты индуцируют сильный lgG ответ у кроликов, очевидно, что они могут действовать как антигены в приготовлении вакцин.
Представляло интерес определить, будут ли P1 Hib -специфичные антитела кроссреагировать с нативным Р1 из неопределяемого штамма Н.influenzae. Кроличьи антитела, образующиеся на синтетические пептиды HIBPI-1, HIBPI-3, HIBPI-5, HIBPI-6, HIBPI-7, HIBPI-9, HIBPI-12, HIBPI-13, узнают Р1-белок из дефинированных и недефинированных культур Н. influenzae. Эти результаты предполагают, что пептиды, относящиеся к аминокислотным остаткам 39-64, 103-137, 165-193, 248-283, 307-331, 400-437 и 179-218 зрелого Pi-белка, содержат высококонсервативные среди дефенированных и недефинированных культур Н. influenzae эпитопы.
Антитела кролика, вырабатывающиеся на конъюгат Р2-пептида и KIH, были исследованы на активность против нативных Р2 в Р2-специфичных ELISA и в иммуноблоттинге. Хотя все пептидспецифичные сыворотки, кроме сывороток, вырабатывающихся на HIBP2- 26-КIH- и ОМР2-13 KIH конъюгаты узнавали Р2 в иммуноблотах, только для Porin-1, ОМР2-5, -7, -8, -10-12 и CHIBP2 пептид конъюгатов с KLH было найдено, что они вызывают образование антител, которые кросс-реагируют с нативным Р2 в Р2-спепифичном ELISA (табл. 9). Все неконъюгированные пептиды, кроме Porin -1 и HIBP2-26, эмульсифицированные полным адъювантом Фрейнда, вызывали сильный пептид-специфичный ответ антителами против Р2 в иммуноблотах (табл. 3). Также, антисыворотка на неконъюгированные пептиды ОМР2-5, -8, -10, 11, -12 и -13 строго реагировала с очищенным Р2 в Р2-спепифичном ELISA. Эти данные указывают на то, что эти пептиды несут потенциальные функциональные T-хелперные эпитопы и B-клеточные эпитопы. Далее, Р2, выделенный из трех различных неидентифицируемых культур SB30, SB32 SB33, был использован как мишень-антиген в иммуноблотах. Кроличьи анти- Porin-1, OMP2-5, -8, -8, -10, -11, -12 и -13 антисыворотки четко реагировали с Р2 из всех неопределяемых штаммов. Эти результаты предполагают, что пептиды, относящиеся к аминокислотным остаткам 1-19, 125-150, 183-219, 241-265, 263-289, 285-306 и 302-319 зрелого Р2-белка, содержат высококонсервативные среди дефинированных и недефинированных культур Н. influenzae эпитопы.
Антисыворотка кролика, полученная на Р6-пептиды, была проверена на активность против Р6 в Р6-специфичном ELISA и в иммуноблотном анализе. Все пептидспецифичные антисыворотки, кроме тех, которые образовались в ответ на Р6-4, узнавали нативный Р6 в Р6-специфичном ELISA и обнаруживали кроссреактивность и с определяемым и неопределяемым Р6 в иммуноблотах (табл. 10). Эти данные свидетельствуют о том, что Р6-пептиды несут потенциальные T-хелперные эпитопы и иммуногенные B-клеточные эпитопы. Далее эти результаты подтверждают, что Р6-белок высококонсервативен среди дефинированных и недефинированных штаммов H.influenzae. Следовательно, эти консервативные эпитопы Р1, Р2 и Р6 можно использовать либо по отдельности, либо в комбинации для приготовления кроссреагирующих синтетических вакцин.
Пептиды, описанные выше, могут в дальнейшем быть полимеризованы или модифицированы липидами подобно липопептидам или привязаны к синтетическому PRP как синтетический гликопептид, либо липогликопептидный конъюгат для производства дополнительных вакцин. Эти вакцины могут использоваться для иммунизации против Hi-инфекции при введении их млекопитающим, например, внутримышечно или парентерально, или когда доставляются на слизистую, с использованием микрочастиц, капсул, липосом и молекул, имеющих мишень, таких, как токсины и антитела.
Дальнейшие эксперименты были произведены, чтобы определить, могут ли синтетические пептиды, несущие идентифицированные иммунодоминантные Т- и B-клеточные эпитопы из либо P1, либо P2 или P6, сшитые в тандеме, вызывать сильный ответ в виде пролиферации защитных от Hi- инфекции антител.
Пептиды, имеющие аминокислотную последовательность II (см. в конце описания).
(SEQ ID NOS : 42-49 соответственно) были синтезированы, очищены и использованы для иммунизации кроликов в присутствии CFA или алюминия. Итоги подведены в табл. 11. Все антипептидные сыворотки достоверно реагировали с соответствующими иммунизирующими пептидами, но не все химерные пептиды вызывали образование антител на нативные OMP. Наилучшими иммуногенами являлись 1P13-2Р8 и 2Р6-IРI3, которые вызывали образование антител, узнающих нативные Р1 и Р2-белки, при введении с алюминием. Поскольку эти пептиды содержали консервативные среди штаммов Hi-эпитопы, они могут быть использованы как дополнительные антигены или модифицированные как липопептиды, или сшитые с синтетическими PRP-олигомерами в качестве вакцин. Эти вакцины можно использовать для иммунизации против Hi-инфекции при введении их млекопитающим, например, внутримышечно или парентерально, или, когда доставляются на слизистую, с использованием микрочастиц, капсул, липосом и молекул, имеющих мишень, таких, как токсины и антитела.
Синтез PRP-олигосахаридных фрагментов с использованием PEG.
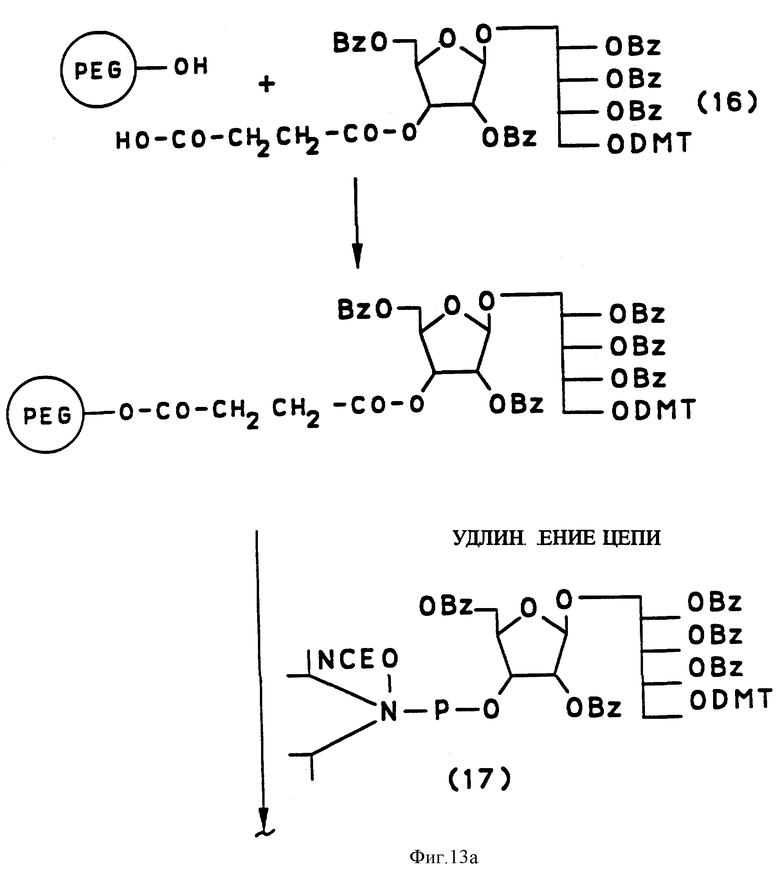
Синтетический PRP приготовлен путем комбинации жидкотвердофазного синтеза и высокоэффективного фосфорамидитного метода, как схематически показано на фиг. 13 и 14. Это новый процесс, в котором используется полиэтиленгликоль монометиловый эфир (PEG) в качестве твердого носителя. Твердофазный носитель содержит большое количество химически активных функциональных групп в диапазоне от приблизительно 200 до 500 мкмоль/г носителя в сравнении с от 30 до 35 мкмоль реакционных групп на грамм традиционных носителей, таких, как стекло с контролируемым размером пор только стехиометрические количества синтетического PRP повторяющегося звена в каждом цикле связывания, сравнивая с 5-10-кратным молярным избытком в обычном твердофазном синтезе. Далее PEG растворим в реакционных растворах, так что эффективность связывания в каждом цикле 35-98%. В конце цикла PEG-связанный, синтезированный PRP, осаждается эфиром, чтобы убрать все побочные продукты. Для синтетического PRP-гексамера конечный выход составил около 70%. Таким образом, настоящий синтез является очень быстрым, недорогим и простым для масштабирования в коммерческих целях в противоположность твердофазному синтезу, который представляет собой трудоемкий, дорогостоящий и занимающий время процесс.
Далее процесс синтеза описывается более подробно. PRP-мономер для инициации полимеризации представляет собой соединение, формула которого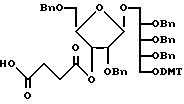
где Вn и DMT это бензил и диметокситритиловые группы соответственно. Этот мономер привязан к PEG, как описано в примере 10, детритилирование производится трихлоруксусной кислотой (TCA) и затем связывается с еще одним PRP-мономером для элонгации цепи, как представлено формулой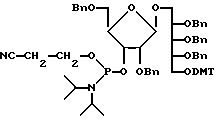
Конечное соединение затем детритилируется TCA. В каждом цикле элонгация цени сопровождается связыванием детритилированной цепи в присутствии катализатора, главным образом, тетразола. После каждого шага связывания происходит окисление фосфора окислительным агентом, обычно t-бутил гидроперекисью. Цикл синтеза (стадии детритилирования, связывания, окисления) повторяется до тех пор, пока не получен олигомер нужной длины. Терминация PRP-олигомера происходит в результате реакции с терминатором цепи, имеющего следующую формулу: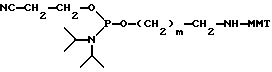
где m-целое число, обычно от 4 до 6 и MMT-монометокситритил. После терминации цепи конечный олигомер на PEG-носителе, который является одной из характеристик данного изобретения, снимается с твердого носителя, главным образом, путем аммонолиза. Получаемый материал представлен следующей формулой: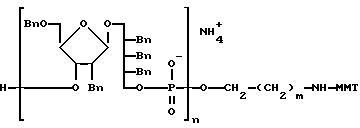
где n- это целое число, обычно от 3 до 20 и m-целое число, обычно от 4 до 6. Вn-бензил, MMT-монометокситритил. Вещество ассоциировано с заряженным ионом. Главным образом, это ион аммония, как на иллюстрации, или замещенный аммоний.
Боковые защитные группировки удаляются гидрогенированием палладием на древесном угле в присутствии воды/ацетиловой кислоты/ t-бутилового спирта как описывается в примере 10. Конечный олигомер может быть очищен стандартными методами, обычно, комбинированием гель-ион-обменной хроматографией.
Как описано выше, при связывании веществ в X6 (фиг. 14) на последнем шаге перед терминацией цепи очень легко превратить синтезированный PRP-олигомер в соединение, несущее химически активные группы, воспроизведеном следующей формулой: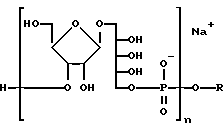
где n- это целое число обычно от 3 до 20 и R-линкерный фрагмент, определяемый как -CH2- (CH2)m-X), где m - целое число обычно от 3 до 5 включительно и X-химически активная функциональная группа подобно- CHY2NH2, - CH2 SH2, или аминореактивные группы, как галоген, метансульфонил, трифторметансуфонил или толуенсульфонил или фотоактивируемые группы, как фенилазид, нитрофенил, бензилфенил и им подобные.
Соединение, в котором присутствует функциональная группа, может быть сформировано как конъюгат, что является одной из главных черт изобретения, конъюгат представлен формулой: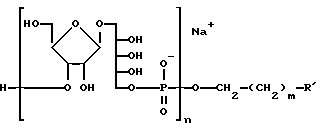
где n - целое число, обычно от 3 до 20, и m - целое число, обычно от 3 до 5, и R' - это (CH2-носитель), где Y-линкерная молекула, которая может быть m -малеимидибензоил-N-гидроксисукциимид и носитель- Hi -пептид или MAP-система из того. Конъюгат будет ассоциирован с заряженным ионом. Главным образом, ионом Na, как на иллюстрации.
Очевидно, что существует множество путей получения синтетического PRP. Технология, которая включает те, что известны, например Публикации Европейского Патентного Бюро 0 320 942 [28] и 0 276 516 [27] так же, как и те, которые могли использоваться в связи с настоящим изобретением, лежат в рамках изобретения.
Иммунохимическая характеристика синтезированного PRP-олигосахарида, конъюгированного с пептидом, содержащим T-хелперный эпитоп(ы).
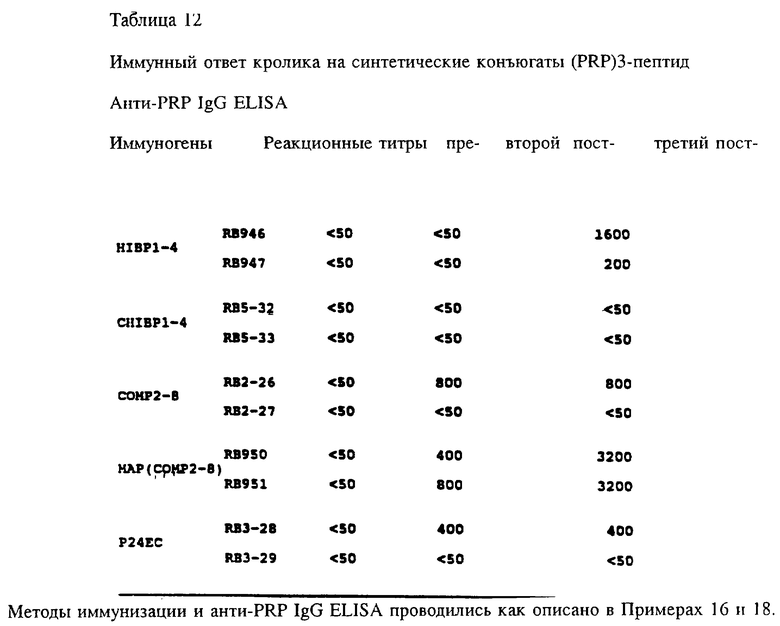
Пептиды, которые можно использовать согласно изобретению, включают любой пептид, который сохраняется при введении молодым млекопитающим, и может служить эффективным T-клеточным эпитопом, например, Р24Е, человеческий T-клеточный эпитоп из gag-белка р24 HIV-1 (фиг. 1). В данном случае были использованы белки внешней мембраны Hib, технология конъюгации подробно описана в примерах 11 и 13. Чтобы определить минимальное число мономеров, требующихся для генерации анти-PRP lgG ответа, синтезированные олигомеры (димер и тример) привязывались к тетанусному токсину и делали инъекцию гликоконъюгата в присутствии алюминия кроликам. Результаты, представленные на фиг. 15, показывают, что для иммуногенности систетический олигомер должен состоять по крайней мере из трех мономеров.
Согласно изобретению кандидаты в конъюгатные вакцины: на основе полностью синтетических PRP-олигомеров были получены путем связывания синтезированных PRP-олигомеров с хорошо охарактеризованными синтетическими T-клеточными эпитопами Hib OMP через цистеиновый остаток, добавляемый либо к N-концу, либо C- концу пептидов, например пептида HIBPI-4 (остатки 165-193 Р1-белка), который был идентифицирован как Hib-штамм-специфичный B-клеточный защитный эпитоп и по крайней мере один функциональный T-хелперный эпитоп.
Чтобы получить эффективную PRP-пептид конъюгатную вакцину, поскольку несколько факторов могут действовать на иммуногенность углевода, антиген должен быть тщательно проверен. К этим факторам относятся: (i) длина цепи олигосахарида; (ii) сайт конъюгации сахаридной части по отношению к Т-клеточным эпитопам; (iii) плотность углеводного антигена на пептиде; (iv) методология конъюгации, которая влияет на стабильность гликоконъюгата; (v) необходимость линкеров или спейсеров между углеводной частью и пептидным носителем для оптимальной антигенной презентации и процессинга. С этими целями два пептида, HIBPI-4 CHIBPI-4 (фиг. 1), которые различались дополнительными цистеиновыми остатками, добавленными к N-концу (HIBPI-4 и SEQ ID NO: 51) и к C-концу (CHIBPI-4 - SEQ ID N0:52), соответственно были синтезированы, очищены и использованы как носители T-клеточных эпитопов для проверки эффекта ориентации сахаристой части относительно T-клеточного эпитопа на конструированную иммуногенность. Синтетический PRP-тример использовался как углеводный антиген. Были приготовлены два PRP-пептидных конъюгата (PRP-CHIBPI-4 и НIВРI-4-PRP) и инъектированы кроликам в присутствии алюминия. После трех иммунизаций исследовали титры анти-PRP и антипептидных IgG антитела кроличьей антисыворотки. Оба конъюгата вызывали сильное образование антипептидных и анти-PPP антител в ответ, но только синтетический HIBPI-4-PRP вызывал анти-PRP lqG ответ. Эти результаты предполагают, что ориентация сахаристой части относительно T-клеточного эпитопа может значительно влиять на хозяйский иммунный ответ на углеводный антиген. Чтобы определить все ли пептиды, несущие функциональные T-клеточные эпитоп(ы) могут эффективно представлять синтезированные PRP-олигомеры иммунной системе, еще два пептида (СОМР2-8- SEQ ID NO: 53 и Р24ЕС ID N0:56), о которых известно, что они несут функциональные T-клеточные эпитоп(ы), были конъюгированы с синтетическим PRP-тримером. Гликопептидные конъюгаты были адсорбированы на алюминии и ими иммунизировали кроликов. Результаты представлены в табл. 12. Оба гликопептидных конъюгата (СОМР2-8-PRP и P24EC-PRP) вызывали образование анти-PRP lgG антител.
Чтобы определить эффект углеводной плотности на иммунногенность гликопептидных конъюгатных вакцин, синтезированный PRP-тример был конъюгирован с множественной антигенной пептидной системой (MAP), содержащей восемь разветвленных ОМР2-8 пептидов (остатки 193-219 Р2-белка) (фиг. 1 - SEQ ID N0:54). Хотя для конъюгации были доступны девять цистеиновых остатков, было обнаружено только пять PRP-тримеров с одной MAP-молекулой. Тем не менее после трех инъекций по 50 мг синтетического гликопептидного конъюгата в присутствии алюминия, у обоих кроликов наблюдался сильный анти-PRP lgG-ответ. Титр анти-PRP IgG был в четыре раза выше, чем полученный с конъюгатом линейного пептида с PRP (табл. 12). Более того ответ антипептидными и анти-Р2 антителами был на 1-2 порядка выше, чем таковой, полученный на конъюгат линейного пептида с PRP. Дальнейший анализ результатов, представленных на фиг. 16, показал, что MAP Hib, конъюгированные с синтетическими PRP-олигомерами, хороши в качестве вакцин, на которые образуются высокие титры анти-PRP lqG антител, сравнимые с полученными на нативный PRP, пришитый к дифтерийному токсину или Р1 или Р2-белкам.
Чтобы определить влияет ли длина углеводного мономера на иммуногенность углеводного антигена в гликоконъюгате, синтезированные PRP-димер, тример, пентамер, гексамер и нативный PRP (молекулярная масса 30 кДа) связывались или к линейному пептиду HIBPI-4 или ОМР2-8 MAP соответственно. Удивительно, что оба пептида, конъюгированные с нативным PRP, не вызывали образования анти-PRP IgG антител. Напротив, PRP-пентамер и гексамер, конъюгированные с линейным пептидом НIВРI-4 вызывали стойкое и продолжительное образование анти-PRP lqG антител в ответ (фиг. 17). ОМР2-8 MAP, конъюгированные с синтетическим PRP-гексамером, также оказались высокоиммуногенными. Синтетический PRP-димер был неиммуногенным, что согласовалось с результатами, полученными ранее.
Использование технологии конъюгации синтетических гликопептидов.
В основных чертах настоящего изобретения технология гликоконъюгации в общем может использоваться для получения конъюгатных вакцин против патогенных инкапсулированных бактерий. Таким образом, технология гликоконъюгации в настоящем изобретении может применяться в вакцинации, чтобы обеспечивать защиту от инфекции от любого микроорганизма, у которого выражены возможные протективные полисахаридные антигены, включая Haemophilus influenzae, Streptococcus pneumoniae, Escherichia coli, Neisseria meningidis, Salmonellantyphi, Streptococcus mutans, Cryptococcus neoformans, Klebsiella, Staphilococcus aureus и Rseudomonas aerogenosa.
Как частное применение, технология конъюгации синтетических гликопептидов может использоваться для получения вакцин, вызывающих образование антител на белки или олигосахариды. Такие вакцины можно использовать, например, для обеспечения иммунности в отношении опухолевых клеток или для получения антиопухолевых антител, которые могут быть конъюгированы с химотерапевтическими или биоактивными агентами.
Понятно, что применение методик настоящего изобретения не выходит за пределы возможностей тех, кто владеет обычной технологией в данной области. Продукты и методы их получения и использования в настоящем изобретении приводятся в последующем разделе "примеры".
Также ясно, что в рамках этой разработки существуют разнообразные варианты или функционально эквивалентные варианты указанных выше пептидов. Термины "варианты" или "функционально эквивалентные варианты", использованные выше, означают, что если пептид модифицирован путем добавления, удаления или изменения одного или больше аминокислотных остатков в любом отношении и тем не менее действует сходным образом с таковыми Р1, Р2 и Р6-пептидами для любого штамма Haemophilus influenzae, то он попадает в объем изобретения.
Приведенные аминокислотные последовательности этих пептидов (табл. 1 - 3 и 11), а также любого подобного пептида, могут быть легко синтезированы на коммерчески доступных пептидных синтезаторах, таких как Applied Biosystems Model 430А, или могут быть получены с использованием техники рекомбинантной ДНК.
Описание, в общем, раскрывает настоящее изобретение. Более полное понимание можно получить, обратившись к последующим примерам. Эти примеры описываются только в иллюстративных целях и не ограничивают рамок рассмотрения изобретения. Изменения по форме и эквивалентные замены не исключаются, если это диктуется обстоятельствами и является подходящим. Несмотря на применение здесь специфических терминов, эти термины применяются в целях описательных, а не ограничительных. Иммунологические методы могут быть не абсолютно четко описаны здесь в описании, но они полностью соответствуют обычным техническим умениям в данной сфере.
Пример 1.
Получение 2,3,4-три-0-бензил-1-0-[2,5,-ди-0-бензил- -β-D- рибофуранозил] -5-0-(4,4'-диметокситритил)-D-рибитол (соединение 14, фиг. 14).
При комнатной температуре 4, 4'-диметокситритилхлорид (6.2 г) добавили к 200 мл раствора дихлорметана, содержавшего 10.2 г 2, 5,-ди-0-бензил- -β-D- рибофуранозил-2, 3, 4-три 0-бензил-D-рибитола (соединение 13, фиг.14), полученного из D-рибозы: через 12 промежуточных продуктов, как описано ранее [33-36] , пиридин (3.4 мл) и 4-диметиламинопиридин (860 мг). После перемешивания 18 - 24 часов реакционную смесь вылили в насыщенный раствор бикарбоната натрия. Водную фазу экстрагировали дихлорметаном, высушивали и растворители выпаривали. Продукт очищали хроматографией на силикагеле, его структура подтверждена ЯМР.
Пример 2.
Получение-2,3,4-три-0-бензил-D-рибитол[2,5-ди-0-бензил-3- 0-)сукцинил- -β-D -рибофуранозил]-5-0-(4,4'-диметокситритил)-D- рибитола (соединение 16, фиг. 13).
К раствору 1.34 г продукта из примера 1 в безводном пиридине (4.5 мл) был добавлен янтарный ангидрид (390 мг) и 4-диметиламинопиридин (240 мг). Реакционную смесь перемешивали на водяной бане от 50 до 80oC от 3 до 10 ч. После добавления воды реакционную смесь концентрировали на роторном испарителе. Хроматографией на колонке силикагеля с использованием дихлорметана: метанола: триэтиламина в отношении 95:5:2.5 (об/об/об) получен продукт в виде триэтиламмониевой соли, структура подтверждена ЯМР.
Пример 3.
Получение рибозилрибитолфосфорамидита.
К раствору соединения 16 (1.2 г в 5 мл безводного диоксана) были добавлены N,N-диизопропилэтиламин (1.4 мл) и 2-циано-этил- N,N-диизопропил хлорофосфорамидит (640 мл). После перемешивания 1- 3 ч, дополнительное количество N, N-диизопропилэтиламина (430 мл) и 2-цианоэтил-N,N-диизопропил хлорофосфорамидита (250 мл) было добавлено. Реакционная смесь была разбавлена в три раза дихлорметаном и экстрагирована равным объемом IM триэтиламмония бикарбоната, рассолом и высушена безводным сульфатом натрия. Продукт был очищен на силикагеле и его структура была подтверждена ЯМР.
Пример 4.
Получение I-t-бутилдиметилсилилокси-6-циано-гексана (соединение Х2, фиг. 14)
Цианид натрия (1.2 г) растворили в диметилсулфоксиде и нагревали 30 мин при 90oC. Твердый I-t-бутилдиметилсилилокси- 6-бромгексан (5.8 г, соединение XI) полученный методом, описанным ранее [37], добавляли к раствору цианида натрия. После нагревания при 120-130oC в течение 20-180 мин, реакционную смесь выливали в ледяную воду и водную фазу экстрагировали эфиром, промывали рассолом, высушивали и концентрировали. Продукт перегоняли при 0.5 Topp (мм рт.ст.) и 107oC для получения бесцветного масла. Масс-спектрометрией высокого разрешения для C12H24ONSi получено 226.1624 при вычисленном 226.1627.
Пример 5.
Получение 7-амино-1- t-бутилдиметилсилилокси-6-циано-гептана (соединение 13, фиг. 14).
К раствору алюмогидрида лития (600 г, AIdrich) в эфире (50 мл) по каплям добавляли продукт (3.7 г) из примера 4 в эфире (50 мл). После 1-3 ч смесь перелили в воду и перемешивали 30 мин. Нерастворимый гидроксид алюминия отфильтровали через цеолитную прокладку, водную фазу экстрагировали трижды эфиром. Эфирный экстракт промывали раствором соли, обезвоживали и концентрировали. Неочищенный продукт перегоняли при 0.25 Topp и 82oC. Масс-спектрометрией высокого разрешения для C13H31ONSi получено 245.2175 при вычисленном 245.2159.
Пример 6.
Получение (N-Монометокситритил)-7-амино-I-t-бутилдиметилсилилокси- 6-гептана (соединение Х4 фиг. 14).
Монометоксиметил хлорид (3.7 г, AIdrich) добавляли к раствору продукта (2.3 г) из примера 5 в дихлорэтане (40 мл). После перемешивания при комнатной температуре в течение 10-24 ч, раствор перелили в насыщенный раствор бикарбоната натрия. Водную фазу экстрагировали дихлорметаном. Дихлорметановый экстракт промывали раствором соли и высушивали. Растворитель испаряли и продукт был очищен хроматографией на силикагеле. Очищенное соединение X4 анализировали на масс-спектрометре высокого разрешения. C33H47O2NSi получено 517.3355 при вычисленном 517.3376.
Пример 7.
Синтез (N-Монометокситритил)-7-аминогептанола (соединение 15, фиг. 14).
IМ раствор тетрабутиламмоний фторида (25.8 мл) медленно добавляли к раствору соединения 14 (4.3 г) в тетрагидрофуране (46 мл). После перемешивания при комнатной температуре в течение 4-18 ч раствор перелили в 100 мл воды и перемешивали еще 30 мин. Органическую фазу экстрагировали раствором соли и высушили. Неочищенный продукт затем был очищен на силикагеле. Очищенный продукт был анализирован на масс-спектрометре высокого разрешения. C27H33O2N, получено 403.2514 при вычисленном 403.2511.
Пример 8.
Получение (N-Монометокситритил)-7-аминогептил (2-цианоэтил)- N-диэтилфосфорамидита (соединение X6, фиг. 14).
К раствору соединение X5 (240 мг) в диоксане (10 мл) был добавлен диизопропилэтиламин (840 мкл) и цианоэтил-N, N-диэтилфосфорамидит (270 мкл). После перемешивания в течение 1 ч, реакционная смесь была разбавлена дихлорметаном и ее промывали IM раствором триэтиламмония бикарбоната и, наконец, раствором соли. После высушивания и концентрирования осадок был очищен хроматографией на силикагеле. Продукт был анализирован на масс-спектрометре высокого разрешения. C36H50N3O3 получено 603.3620 при вычисленном 603.3620. Структура продукта также подтверждена ЯМР-анализом.
Пример 9.
Получение сукцинилрибозилрибитол-PEG- (фиг. 14).
К раствору соединения 16 (1.8 г) в дихлорметане (18 мл) были добавлены N-гидроксибензотриазол (295 мл) и дициклогексилкарбодиимид (450 мл). Реакционную смесь перемешивали при комнатной температуре. После 2-8 ч дициклогексимочевину удаляли фильтрацией. Фильтрат, N-метилимидазол (522 мкл) и диизопропилэтиламин (600 мкл) были добавлены к полиэтиленгликольмонометиловому эфиру, PEG (средняя Мг 5000; 2.1 г, Fluka). Смесь перемешивали ночь под аргоном при комнатной температуре. Функциональный PEG был осажден холодным эфиром и отфильтрован. Емкость носителя была определена спектрофотометрически по методу Gait и др [36] и составила около 200 ммоль/г. Свободные остаточные гидроксильные группы были закрыты смесью 20%-ного уксусного ангидрида/пиридина в дихлорметане в течение 1-3 ч при комнатной температуре. Носитель осаждали холодным эфиром, фильтровали и промывали холодным эфиром.
Пример 10.
Получение синтетического PRP с использованием растворимого полимерного носителя (фиг. 13).
1 г PEG-PRP-DMT (продукт примера 9) выпаривали дважды с пиридином и растворяли в ацетонитриле под аргоном. Удлинение PRP-олигосахарида проходило в цикле из четырех стадий, каждому шагу (стадии) предшествовала преципитация функционального PEG-холодным эфиром для освобождения от побочных продуктов с последующей кристаллизацией из дихлорметана эфира.
Первый шаг синтеза включал снятие диметокситретиловой группировки с использованием 3%-ного толуолсульфоновой кислоты в хлороформе/муравьиной кислоте, сопровождавшееся рибозилрибитол фосфорамидитным продуктом из примера 3 в присутствии тетразола (180 мин). Эффективность связывания определена как 95%. Окисление (3 стадия) проводилось с использованием 70% раствора t-бутил гидропероксида (120 мин) и конечное кэппирование (стадия 4) в 20%-ном уксусном ангидриде/пиридин в дихлорметане (60 мин). Было проведено два цикла синтеза, которые сопровождались связыванием спейсерного фосфорамидитного продукта из примера 8. Смолу нагревали с водным концентрированным аммиаком и тетрагидрофураном в течение 17 - 24 ч при 500-100oC. Смесь фильтровали чтобы отделить PEG, промывали и выпаривали растворители. Гидрогенолиз продукта в присутствии 10%-ного Pd/древесный уголь в t-бутиловый спирт/вода/уксусная кислота (4:4:1) с использованием аппарата гидрогенации среднего давления при 40 psi обеспечивает чистый продукт после фильтрации. Продукт был лиофилизирован, затем очищен комбинацией гель-фильтрации на колонке Sephadex G-25 в 0.01 М триэтиламмоний бикарбонате, pH 7.0 и последующей ионобменной хроматографией на Sephadex G-25 с использованием воды. После лиофилизации соответствующих фракций получен твердый продукт, структура которого анализирована ЯМР. Спектр рибозилрибитолфосфатного тримера был получен и оказался сходным с таковым, полученным Hoogerhout (J. Carbohydr. Chem. 7,399, 1988).
Пример 11.
Модификация синтетического (PRP)3.
m -Малеимидобензоил-N-гидроксисукцинимид (20 мг, 63.3 мкмоль) в тетрагидрофуране (1 мл) был добавлен к раствору синтезированного (PRP) (5.2 мг, 4.3 мкмоль) в 0.1 М фосфатном буфере (1 мл) pH 7.5. После перемешивания раствора 30 мин при комнатной температуре под аргоном реакционную смесь экстрагировали эфиром (4х5 мл) и образовавшуюся водную фазу нанесли на колонку Sephadex G-25 (Pharmaci а) (2х30 см), уравновешенную 0.1 M триэтиламмонийацетатным буфером, pH 7.2, и элюировали тем же буфером. Элюирование контролировали спектрофотометрически при 254 нм. Первый пик элюиции был собран и лиофилизирован. Количество малеимидных групп, включившихся в (PRP)3, определялось путем модифицированного метода Эдмана [39] и составило 90% включения.
Пример 12.
Пептидный синтез.
Пептиды из OMP Р1, Р2 и Р6 (табл. 1 - 3) были синтезированы с использованием пептидного синтезатора АВ1 430А и оптимимизированной t-ВОС химии, как описано производителем, затем очищены от смолы фтористоводородной кислотой (HF). Пептиды были очищены обратно-фазной жидкостной хроматографией высокого давления (RP-HPLC) на полупрепаративной колонке Yydac С4 (1х30 см) в 15-55%-ном градиенте ацетонитрила в 0.1%-ной трифторуксусной кислоте (TFA) за 40 мин при скорости 2 мл/мин. Все синтетические пептиды (табл. 1 - 3), использовавшиеся в биохимических и иммунологических исследованиях, были > 95% очищены, как показала аналитическая хроматография. Аминокислотный анализ производился на Waters Pico-Tag - системе и хорошо согласовался с теоретическими составами. Синтетические MAP (ОМР2-8) были получены вручную с использованием t-ВОС химии твердофазного пептидного синтеза по методу, описанному ранее Tam и др. [40]. Цистеиновые остатки были добавлены к C- и N-концам пептидов в целях конъюгации с PRP. MAP - пептиды были очищены как ранее описано RP-HPLC.
Пример 13.
Получение полностью синтетических пептид-PRP-конъюгатов.
От одного до двух миллиграммов индивидуальных синтетических пептидов (ОМР2-8) и HIBPI-4 растворяли в 0.5 мл хорошо дегазированной воды, затем добавляли 0.8 мл MBS -(PRP)3 (1.6 мг) хорошо дегазированной воды. Полученную смесь перемешивали в течение ночи при комнатной температуре под аргоном. Нерастворимый осадок удаляли центрифугированием, и супернатант наносили на колонку Saphadex G-50 (2х30 см), уравновешенную 0.1М триэтиламмоний ацетатным буфером, pH 7.2, чтобы удалить избыточный MBS - (PRP)3 гель-фильтрацией. Синтетические пептид- (PRP)3 конъюгаты были собраны и анализированы обратно-фазной HPLC в тесте OrcinoI и аминокислотном анализе. Молярное отношение пептида к PRP было 1:1 и 1:5 для HIBPI-4 и MAP пептидных конъюгатов соответственно. Синтетические пептид-PRP конъюгаты затем адсорбировались на алюминии для исследования иммуногенности.
Пример 14.
0.5 мл окисленного перийодатом PRP (25 мг в 1 мл 0.1М натрий фосфатном буфере, pH 6.0), полученного из нативного PRP, обработанного водной перийодной кислотой (4), добавили к бычьему сывороточному альбумину (BSA) (1.32 мг, 0.02 мкмоль в 0.5 мл 0.2 М натрий фосфатного буфера, pH 8.0) с последующим добавлением цианоборгидрида натрия (14 мг, 0.22 мкмоль; 10 экв. к В А). После инкубации при 37oC 5 дней реакционную смесь диализовали против 0.1 М фосфатного буфера (4х1 л), pH 7.5, и получившийся раствор нанесли на аналитическую колонку Syperose 12 (15х300 мм, Pharmaci a), уравновешенную 0.2М натрийфосфатным буфером, pH 7.2, и элюировали тем же буфером. Выход фракций определялся по поглощению в 230 нм. Главный пик белка собран и сконцентрирован в Centriprep 30 (Pierce) до 2.2 мл. Количество белка было определено в Bio Rad и составило 300 мкг/м. Подтверждение производного PRP получено в тесте OrcinoI.
Пример 15.
Получение антипептид и анти-OMP антисывороток.
Кролик, мыши (Balb/C и морские свинки были иммунизированы внутримышечно нативными Р1 или Р2 или Р6 или индивидуальными пептидами, эмульгированными в полном адъюванте Фрейнда и затем двумя. booster дозами (половинное количество того же иммуногена в неполном адъюванте Фрейнда) с интервалом в две недели. Антисыворотки были собраны и сохранялись, как описано выше.
Пример 16.
Получение анти-PRP антител.
Кролики были иммунизированы внутримышечно индивидуальными конъюгатами PRP-носитель (5-50 мкг PRP эквивалента), смешанными с 3 мг AIPO4 (в мл) с последующими boozter дозами (половинное количество того же иммуногена) с интервалом в две недели. Антисыворотку отбирали каждые две недели первой инъекции, инактивировали нагреванием при 56oC 30 мин и хранили при -20oC.
Пример 17.
P1-, Р2-, Р6- и пептид-специфичный ELISA.
На микротитровые плашки (чашки с лунками) (Nunc-immunoplate, Nunc, Denmark) были нанесены 200 нг очищенных OMP или 500 нг индивидуальных пептидов в 50 мкл в буфере нанесения (15 мМ Na2CO3, 35 мM NaHCO3, pH 9.6) и оставлены при комнатной температуре на 16 ч. Затем панели блокировали 0.1% (w/v) BSA в натрийфосфатном буфере (PBS) в течение 30 мин при комнатной температуре. Последовательно разведенные антитела были добавлены к лункам и инкубированы 1 ч при комнатной температуре. После удаления антисыворотки плашки промывали пять раз PBS, содержащим 0.1% (w/v) Twееп-20 и 0.1% (w/v) BSA. Козлиные, антикроличьи, мышиные и морской свинки F(ab')2, человеческие IgG были конъюгированы с пероксидазой хрена (Jackson Immunoresearch Labs Inc., PA), разведены буфером промывки (1/8000), нанесены на микротитровые плашки. После 1 ч инкубации при комнатной температуре плашки промывали пять раз буфером промывки. Затем их проявляли с использованием тетраметилбензидина (TMB) в H2O2 (ADI, Toronto) в качестве субстрата. Реакцию останавливали IN H2SO4, оптическую плотность измеряли при 450 нм на Ti retek MuItis can II (Flow L abs., Virginia). Два не имеющих отношения пептида токсина пертусис NAD-SI (19 аминокислот) и S3 (123-154) (32 остатка) были использованы как негативные контроли в пептидспецифических ELISA. Эксперименты были проведены трижды, и реактивный титр антител определялся по мере разбавления в соответствии с двукратным увеличением оптической плотности по сравнению с величиной, полученной в преиммунной сыворотке.
Пример 18.
Измерение анти-PRP антител.
На микротитровые чашки с лунками (Nunc-Immunoplate, Nunc, Denmark) было нанесено 200 нг очищенного PRP-BSA в 200 мкл буфера нанесения (15 мм Na2CO3, 35 мM NaHCO3, pH 9.6) и оставлены при комнатной температуре на 16 ч. Затем панели блокировали 0.1% (w/v) BSA в натрийфосфатном буфере (PBS) в течение 30 мин при комнатной температуре. Последовательно разведенные антитела на PRP-носитель были добавлены к лункам и инкубированы 1 ч при комнатной температуре. После удаления антисыворотки плашки промывали пять раз PBS, содержащим 0.1% (w/v) Tween-20 и 0.1% (w/v) BSA. Козлиные, антикроличьи, мышиные и морской свинки F (ab')2, человеческие lqG были конъюгированы с пероксидазой хрена (Jackson ImmunoResearch Labs Inc., PA), разведены буфером промывки (1/8000), нанесены на микротитровые плашки. После 1 ч инкубации при комнатной температуре плашки промывали пять раз буфером промывки. Затем их проявляли с использованием тетраметилбензидина (TMB) в Н202 (ADI, Toronto) в качестве субстрата. Реакцию останавливали IN H2SO4, оптическую плотность измеряли при 450 нм на Titretek MuItiscan II (Flow Labs., Virginia). Стандартная анти-PRP антисыворотка использовалась как позитивный контроль. Эксперименты были проведены трижды, и реактивный титр антител определялся по мере разбавления в соответствии с двукратным увеличением оптической плотности по сравнению с величиной, полученной в преиммунной сыворотке.
Пример 19.
Пролиферативный тест для синтетических T-клеточных эпитопов.
Картирование T-клеточных эпитопов было проведено путем праймирования Balb/c, С57B1/6 и A/J мышей 5 мг индивидуальных OMP (Р1, Р2 или Р6). Через три недели селезенку удаляли и культивировали спленоциты в RPMI 1640 (Flow Lab) в 10%-ной телячьей фетальной сыворотке, инактивированной нагреванием (Gibco) 2мМ L-глутамине (Flow Lab), 100 U/мл пенициллина (FIow Lab), 100 мг/мл стрептомицина (Flow Lab), 10 ед/мл rlL-2 и 50 мкМ 2-меркаптоэтанола (Sigma) в течение 5-7 дней. Пролиферативный ответ в праймированных спленоцитах на ОМР-пептиды определялся в стандартном анализе in vitro [41]. Кратко, 104 спленоцитов были помещены на 96-луночную микротитровую плашку вместе с 5х10 (1700 Rad) свежими сингенными клетками селезенки, которые использовались как антигенпрезентирующие клетки (АРС) в присутствии повышающихся молярных концентраций (0.03-3 мкМ пептида, растворенного в культуральной среде без IL-2). Культуры выдерживались 40 ч в увлажняемом инкубаторе в 5% СО2/воздух при 37oC. Во время последних 16 ч инкубирования в каждую лунку добавляли 0.5 мкКи [H3] -Tdr (5 Ки/ммоль, NEN). Содержимое лунок перенесли на фильтры из волоконного стекла, включение H3-тимидина в клеточную ДНК измерялось сцинцилляционным счетчиком β- излучения (Beckman). Результат как среднее из трех показаний для каждой пептидной концентрации. Стандартное отклонение было всегда < 15%. Пролиферативный ответ считался позитивным, когда включение было в три раза выше, чем полученное с неотносящимися пептидами или культуральной средой.
Пример 20.
Иммуноблотный анализ.
Иммуноспецифичность антисыворотки, вырабатывающейся на пептиды или конъюгаты PRP с носителем, определялись иммуноблотным анализом, как описано ранее [42].
Настоящим изобретением предлагаются иммуногенные синтетические пептиды, которые могут быть использованы индивидуально или в PRP-конъюгатах, в вакцинах против инфекции Hi. В рамках изобретения возможны модификации.












Синтетический пептид имеет аминокислотную последовательность, относящуюся по крайней мере к одной антигенной детерминанте по крайней мере одного белка, обычно структурного, в особенности, Р1, Р2 и Р6-белкам Halmophilus influenzae (Нi), в частности типа В. Его используют в виде химерной липофильной Т-В формы, привязанной к молекуле носителя, например синтетической РRР-молекуле и/или полимеризованного и образующего молекулярные агрегаты, как вакцина против Нi. 6 с. и 29 з.п.ф-лы, 12 табл., 17 ил.
где n - целое число;
m - целое число;
R1 - синтетический пептид, содержащий по крайней мере один Т-клеточный эпитоп.
2. 24. Коньюгат по п.22, отличающийся тем, что указанный синтетический пептид содержит аминокислотную последовательность GPKEPFRDYVDRFYK из gag p24 белка HIV-I.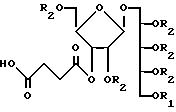
где R1 - первая защитная группа;
R2 - вторая защитная группа,
к твердой подложке монометилового эфира полиэтиленгликоля (REG) с образованием соединения, связанного с REG, растворения указанного связанного с REG соединения в растворителе, удаления указанной первой защитной группы из связанного с REG соединения с образованием незащищенного связанного с REG соединения, связывания незащищенного, связанного с PEG соединения с повторяющимся фрагментом, в целях элонгации цепи, имеющего формулу: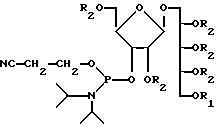
удаления защитных групп с атома фосфора с образованием синтетического, связанного с REG PR P, удаления указанного синтетического связанного с REG PR P в твердой форме из указанного растворителя для разделения REG - связанного синтетического PR P от побочных продуктов, повторное растворение REG - связанного синтетического PR P в твердой форме в растворителе, повторения указанных стадий снятия указанной первой защитной группы, связывания с повторяющимся фрагментом, удаления защитной группы от атома фосфора, удаления REG - связанного синтетического PR P в твердой форме из растворителя и повторного растворителя REG - связанного синтетического PR P в твердой форме до тех пор, пока в олигомере P R P не соединится желаемое число повторяющихся фрагментов, терминацию олигомеризации цепи терминирующей молекулой с формулой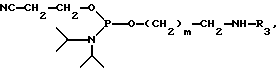
где m - целое число от 4 до 6;
R3 - третья защитная группа,
для получения PEG - связанного защищенного олигомера PR P, удаления защитной группы от атома фосфора и удаление указанного PEG - связанного защищенного олигомера PR P в твердой форме из указанного растворителя для разделения PEG - связанного защищенного олигомера PR P, имеющего формулу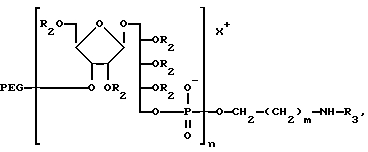
где n является целым числом от 3 до 20;
X+ является контрионом.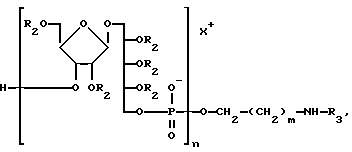
где m и n целые числа;
X+ противоион,
и удаляют вторую и третью защитные группы, где R2 - вторая защитная группа, а R3 - третья защитная группа.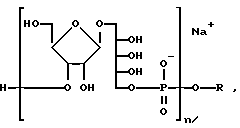
где R - связывающий фрагмент.
-CH2(CH2)m -X,
где m - целое число;
X - химически активная функциональная группа, аминореактивная или фотоактивируемая группа.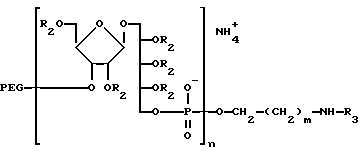 ,
,
где R2 и R3 защитные группы; m и n - целые числа.
| Огнетушитель | 0 |
|
SU91A1 |
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| ЛОГИЧЕСКИЙ АВТОМАТ | 0 |
|
SU276516A1 |

