Область изобретения
Изобретение относится к области иммунологии, в частности к иммуногенам и антигенам от видов Haemophilus. Ссылка на родственную заявку.
Настоящая заявка является составной частью патентной заявки США серийный 08/296149 от 26 августа 1994 г., которая сама по себе является составной частью заявки серийный 08/278091 от 21 июля 1994 г.
Предпосылки изобретения
Haemophilus influenzae представляет собой организм, который ответственен за множество тяжелых заболеваний человека, такие как менингит, эпиглотит, пневмония и отит. Haemophilus influenzae тип b (Hib) является основной причиной бактериального менингита у детей в возрасте до пяти лет. Защитные антитела к этому заболеванию индуцируются капсулярным полисахаридом этого организма, и была разработана вакцина, в которой использовали в качестве антигена очищенный полирибозилрибитолфосфат (ПРФ). Эта вакцина обеспечивала 90%-ную защиту у взрослых и у детей возрастом старше 24 месяцев, но была неэффективна у детей младше 24 месяцев (Zangwill et al, 1993). (Ссылки приведены в списке литературы в конце настоящего описания, каждая из ссылок данного списка включена здесь в качестве ссылки без последующей к ней отсылки). Как и другие полисахаридные антигены, ПРФ не индуцирует пролиферацию Т-хелперных клеток, и реиммунизация не способна ни вызывать усиленный ответ, ни увеличивать клетки памяти. Конъюгация полисахарида ПРФ с белком-носителем придает зависимые от Т-клеток характеристики вакцине и существенно усиливает иммунный ответ на ПРФ-антиген. В настоящее время существует четыре доступных вакцины ПРФ-несущего конъюгата. Это вакцины, основанные на конъюгате капсулярного полисахарида Н. influenzae типа b с дифтерийным токсиоидом, тетанус токсиоидом или белком внешней мембраны Neisseria meningitidis (рассмотрены у Zangwill et al, 1993). Эти конъюгированные вакцины Н. influenzae b значительно снизили частоту случаев бактериального менингита (Schoendorf et al, 1994).
Имеется шесть серотипов Н. influenzae, обозначаемых а - f, которые определяются по их капсулярным полисахаридам. Имеющиеся в настоящие время коньюгированные вакцины Haemophilus не защищают от других инвазивных типирующихся штаммов (типы а и с) и, что важно, не защищают от нетипирующихся (NTHi) штаммов, которые являются распространенной причиной послеродового сепсиса и сепсиса новорожденных, пневмонии и отита. Отит является наиболее распространенным заболеванием раннего детства, при этом приблизительно 70% всех детей в возрасте до семи лет перенесли, по крайней мере, однажды отит. Хронический отит может привести к нарушению слуха, речи и распознавания у детей. Это вызывается бактериальной инфекцией Streptococcus pneumoniae (приблизительно 50%), нетипирующимся Н. influenzae (приблизительно 30%) и Moraxella (Branhamella) catarrhalis (приблизительно 20%). Только в одних Соединенных Штатах для лечения отитов на антибиотики и хирургические процедуры, такие как тонзиллоэктомия, аденоидоэктомия и введение тимпаностомических трубок, тратится от 1 до 2 миллиардов долларов в год. Для того, чтобы достигнуть универсальной защиты от Н. influenzae-опосредованных заболеваний, особенно в возрастной группе от двух до шести месяцев и в группах определенного высокого риска, желательно иметь запас консервированных перекрестно-реактивных некапсулярных иммуногенов Н. influenzae. Нетипированные штаммы Н. influenzae также являются важными патогенами, ответственными за пневмонию у пожилых и других индивидуумов, которые особенно предрасположены к респираторным инфекциям. Таким образом, необходимо наличие антигенов Н. influenzae, которые используются в качестве компонентов в иммуногенных препаратах, обеспечивающих защиту от множества серотипов Н. influenzae. В заявке РСТ WO 92/10936, опубликованной 9 июля 1992 г. и включенной здесь в качестве ссылки, описывается белок внешней мембраны, полученный из Н. influenzae с молекулярной массой 47000, который, как сообщается, представляет собой адгезин и обозначается как Hin47, который является иммунологически консервативным между нетипирующимися типа b и для нетипичных клинических изолятов Н. influenzae. Аминокислотная последовательность Hin47 и нуклеотидная последовательность гена, кодирующего Hin47, представлены на конференции Американского Общества Микробиологов (American Society of Microbiology - ASM), состоявшейся в Новом Орлеане 26-30 мая 1992 г. Эти последовательности также были опубликованы в заявке РСТ WO 94/00149, опубликованной 6 января 1994 г. и включенной здесь в качестве ссылки.
Поскольку Hin47 является консервативным между штаммами Haemophilus influenzae и, как сообщается, представляет собой адгезин, белок, применяющийся для диагностики и вакцинации против заболеваний, вызываемых Н. influenzae или другими бактериальными патогенами, которые продуцируют Hin47, или белок, способный увеличивать специфическую реактивность антител к Hin47.
Недостатком Hin47 при использовании в качестве антигена для диагностики, для наработки анти-Нin47 антител, применяемых для диагностики и в качестве иммуногена при вакцинации, является неожиданное обнаружение для заявителей настоящего изобретения, что Hin47 обладает протеазной активностью, которая приводит к аутоперевариванию Hin47 и протеолитической деградации других смешанных с ним антигенов.
Было бы преимуществом создать аналоги белка Hin47 (иногда обозначаемые при этом как мутанты или производные), которые проявляют существенно сниженную протеолитическую активность, для использования в качестве антигенов, иммуногенных препаратов, включая вакцины, носителей для других иммуногенов и для производства диагностических реагентов.
Описание изобретения
Настоящее изобретение касается разработки аналогов белка Haemophilus Hin47, обладающих пониженной протеазной активностью.
В соответствии с первым аспектом изобретения предложен выделенный и очищенный аналог белка Haemophilus Hin47, обладающий пониженной протеазной активностью, которая составляет менее чем 10% активности природного белка Hin47. Такой аналог Hin47 предпочтительно имеет, по существу, те же иммуногенные свойства природного белка Hin47. Аналог настоящего изобретения может быть получен путем химической, биохимической или генетической модификации природного Hin47.
При одном осуществлении настоящего изобретения, когда аналог получен путем генетической модификации, по крайней мере, одна аминокислота природного Hin47, вносящая вклад в протеазную активность, может быть удалена или заменена на другую аминокислоту для получения пониженной протеазной активности. Альтернативно пониженная протеазная активность может быть достигнута путем введения, по крайней мере, одной аминокислоты в природный белок Hin47. По крайней мере, одна удаленная или замененная аминокислота может быть выбрана среди 195-201 аминокислотного остатка Hin47, в частности это может быть serin-197, который может быть удален или заменен на аланин, цистеин или треонин. Кроме того, по крайней мере, одной удаленной или замещенной аминокислотой может быть His-91, который может быть удален или заменен на аланин, лизин или аргинин. Далее, по крайней мере, одной удаляемой или замещаемой аминокислотой может быть Asp-121, которая может быть удалена или заменена на аланин.
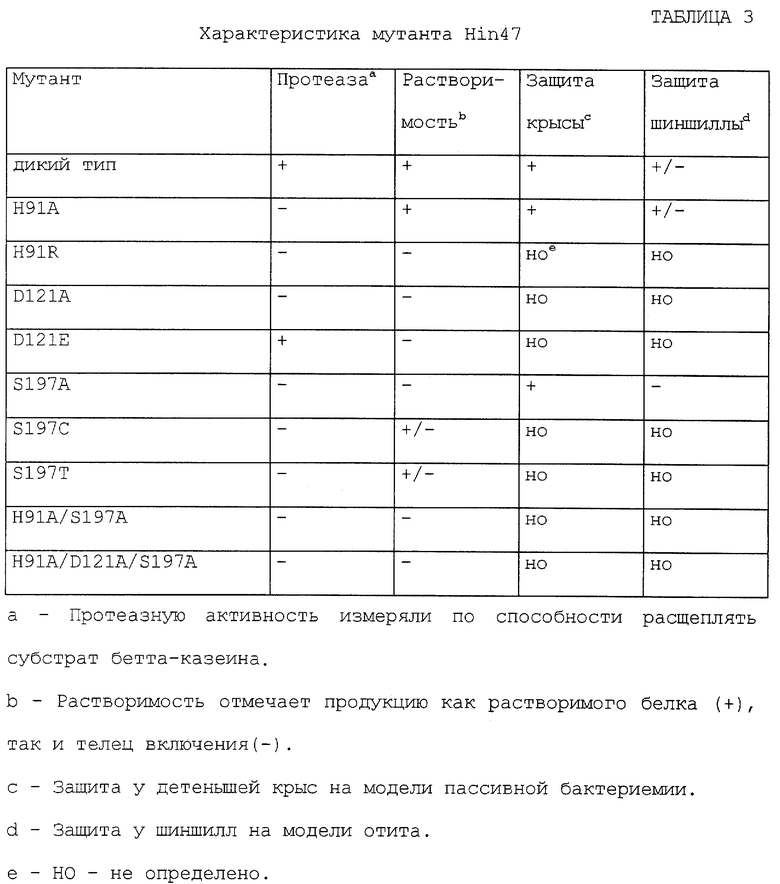
Кроме того, множество аминокислот в молекуле Hin47 могут быть удалены или заменены. Такие аминокислоты могут включать His-91 и Serin-197, и они могут быть удалены или заменены на Ала-91 и Ала-197 с получением аналога Hin47 H19A/S197A. Помимо этого, множество аминокислот могут включать His-91, Asp-121 и Ser-197 и они могут быть удалены или заменены на Ала-91, Ала-121 и Ала-197 соответственно с получением аналога Hin47 H91A/D121A/S197A. Описание некоторых свойств некоторых аналогов Hin47, предложенных здесь, представлено в таблице 3. Обнаружено, что только один мутант Hin47 D121E сохраняет существенную протеазную активность.
С другой стороны, настоящее изобретение относится к выделенной и очищенной молекуле нуклеиновой кислоты, содержащей мутантный ген Haemophilus influenzae hin47, кодирующий аналог белка Haemophilus influenzae Hin47, имеющий пониженную протеазную активность, которая составляет менее 10% от активности природного белка Hin47. Мутантный ген hin47 может кодировать любой из аналогов Hin47, обсуждаемых выше. Мутантный ген преимущественно образован путем сайт-направленного мутагенеза дикого типа гена hin47. Молекула нуклеиновой кислоты может быть включена в рекомбинантную плазмиду, адаптированную для трансформации хозяина и может быть плазмидой DS-1011-1-1 (депонированой 27 июля 1994 г. в Американскую коллекцию типов культур, American type Culture Collection, Rockwill, Maryland, USA, под входящим N 75845). Изобретение также включает трансформированную клетку, содержащую такую рекомбинантную плазмиду.
Настоящее изобретение, с другой стороны, включает способ для получения аналога белка Haemophilus influenzae Hin47, имеющего пониженную протеазную активность, которая составляет менее чем 10% от активности природного белка Hin47, который заключается в идентификации, по крайней мере, одного аминокислотного остатка белка Hin47, который вносит вклад в его протеазную активность, в осуществлении сайт-направленного мутагенеза гена hin47 для удаления или замены нуклеотидной последовательности, кодирующей, по крайней мере, одну аминокислоту, и для получения мутированного гена hin47, введении мутированного гена hin47 в клетку для получения трансформированной клетки и выращивания трансформированной клетки для получения аналога Hin47. По крайней мере, одна аминокислота, которая является выбранной, может быть любой из специфически идентифицированных аминокислот, указанных выше, в отношении к аналогу Hin47.
Введение мутированного гена hin47 предпочтительно продуцирует трансформированную клетку, в которой мутированный ген hin47 находится под контролем Т7 промотера, а выращивание трансформированной клетки и экспрессия аналога Hin47 под воздействием Т7 промотера далее предпочтительно осуществляется путем культивирования в присутствии индуцирующей концентрации лактозы. Введение мутированного hin47 предпочтительно осуществляется путем трансформации клетки рекомбинантной плазмидой DS-1011-1-1, иногда другим способом, относительно как для плазмиды pT/Hin47*.
По дальнейшему аспекту изобретения предложен способ получения выделенного и очищенного аналога Hin47, который заключается в использовании метода, описанного выше для получения аналога Hin47, с продуцированием трансформированных клеток, включающих тельца включения, содержащие аналог Hin47, разрушении выросших трансформированных клеток с получением супернатанта и телец включения, солюбилизации телец включения с получением аналога Hin47, хроматографическую очистку аналога Hin47 из раствора, не содержащего клеточных остатков, и выделении очищенного аналога Hin47.
Аналоги Hin47, предложенные здесь, с их пониженной протеолитической активностью являются применимыми в качестве антигенов в иммунологической композиции, в качестве носителей для других иммуногенов, диагностических агентов и в производстве диагностических агентов. Молекулы нуклеиновых кислот также используются в качестве проб для диагностического использования, а также в качестве иммуногенных композиций.
Следующий аспект изобретения относится к иммуногенной композиции, содержащей иммуноэффективное количество аналога Hin47 или молекулы нуклеиновой кислоты, включающей ген, кодирующий аналог Hin47. Иммуногенная композиция может быть получена в виде вакцины для введения in vivo хозяину, включая человека, для придания защиты против заболеваний, вызываемых бактериальным патогеном, который продуцирует Hin47 или белок, способный к индукции у хозяина антител, специфически реактивных к Hin47. Бактериальный патоген может быть видом Haemophilus, таким как Haemophilus influenzae. Иммуногенные композиции по изобретению могут также содержать, по крайней мере, один другой иммуногенный или иммуностимулирующий материал, такой как адъювант. При дополнительном осуществлении молекула нуклеиновой кислоты, включающая ген, кодирующий аналог Hin47, может содержать в себе живой вектор, такой как вирус оспы. Salmonella, полиовирус, аденовирус, вирус коровий оспы или BCG.
Изобретение также охватывает способ выработки иммунного ответа у хозяина, включая человека, заключающегося во введении ему иммуноэффективного количества иммуногенных композиций, предложенных здесь.
Как отмечено выше, аналог Hin47, предложенный здесь, является полезным для диагностического применения. Соответственно, в дополнительном аспекте изобретения описан способ определения наличия антител, специфически реактивных к Hin47 в образце, включающий следующие стадии:
(а) контактирование образца с аналогом Hin47, обладающим, по существу, теми же иммуногенными свойствами, что и природный белок Hin47, заявленный здесь, с получением комплексов, содержащих аналог Hin47 и любые подобные антитела, присутствующие в образце, специфически реагирующие с ним, и
(б) определение образования комплексов.
Настоящее изобретение описывает также метод определения наличия Hin47 в образце, включающий следующие этапы:
(а) иммунизация субъекта иммуногенной композицией, как описано здесь для получения антител, специфических к белку Hin47;
(б) контактирование образца с антителами для образования комплексов, включающих любой присутствующий в образце Hin47 и Hin47 специфические антитела, и
(с) определение образования комплексов.
Изобретение также охватывает диагностический набор для определения наличия антител в образце, специфически реактивных к Hin47, включающего:
(а) аналог Hin47, обладающий, по существу, теми же иммуногенными свойствами, что и природный белок Hin47, как описано здесь;
(б) устройство для контактирования аналога с образцом для получения комплекса, включающего аналог и любое подобное антитело, присутствующее в образце, и
(с) устройство для определения образования комплекса.
Краткое описание чертежей
На фиг. 1 показана рестрикционная карта плазмид JB-1031-1-14 и JB-1068-2-2 и построение плазмиды для анализа последовательности;
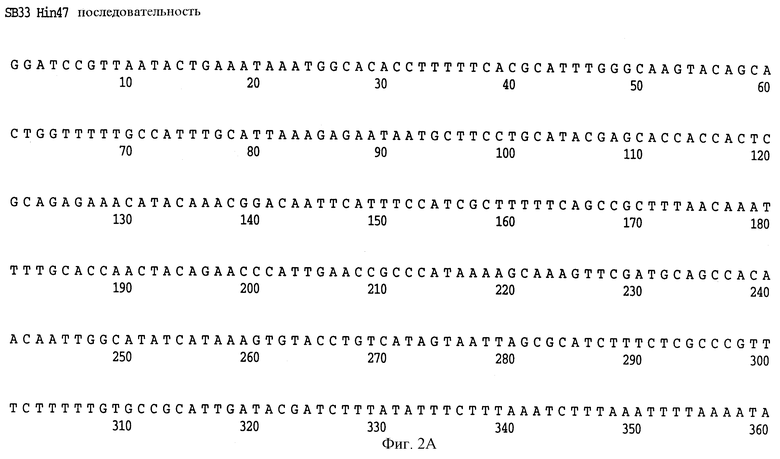
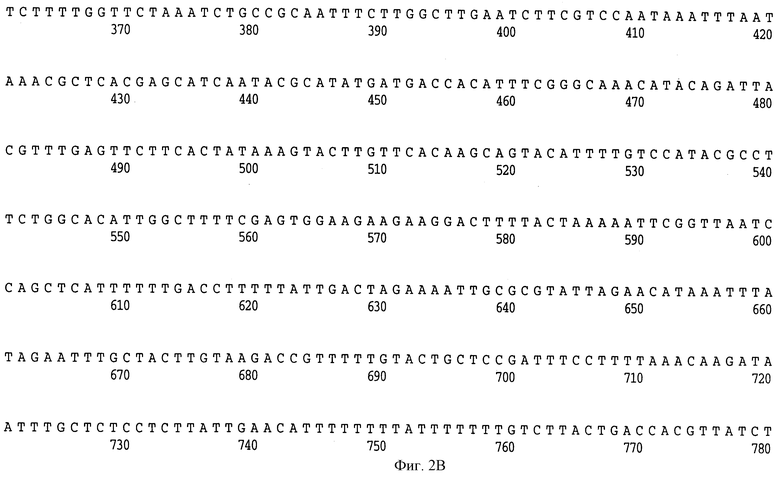
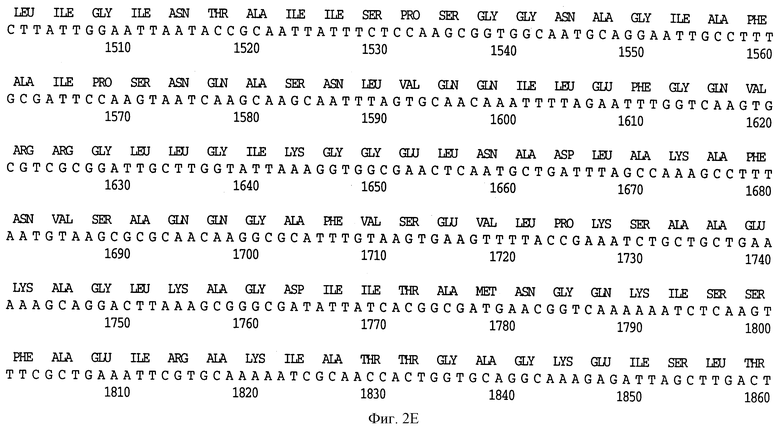
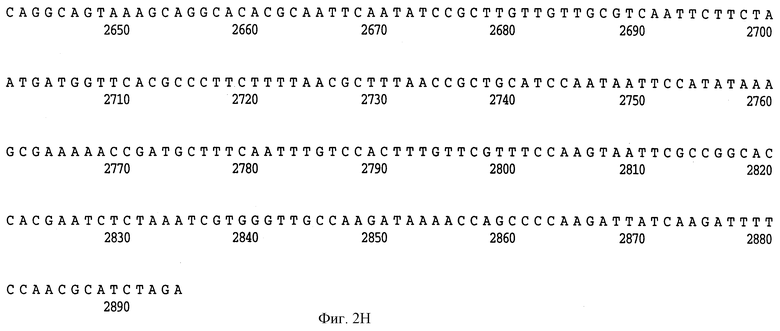
На фиг. 2 A-H представлена полная нуклеотидная (SEQ ID NO: I) и выведенная аминокислотная последовательность (SEQ ID NO: 2) Hin47 из Н. influenzae штамма SB33, как и частичная нуклеотидная последовательность (SEQ ID NO: 33) и частичная выведенная аминокислотная последовательность, скопированная здесь заявителем из материалов, представленных на конференции АОМ, как описано выше.
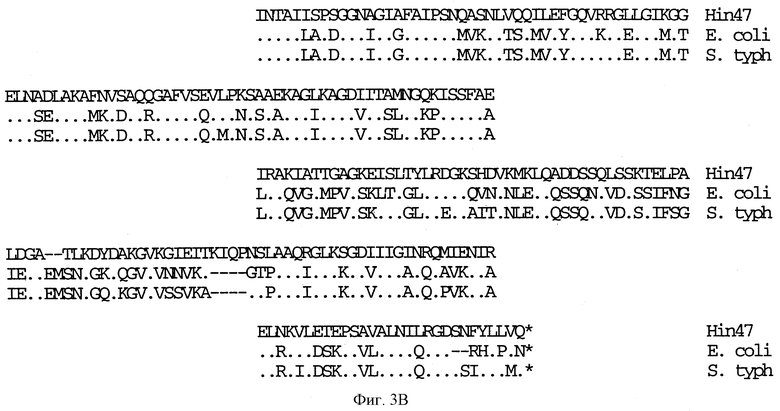
На фиг. 3 A-B представлено сравнение аминокислотных последовательностей Н. influenzae Hin47 (SEQ ID NO:2), E. coli htrA (SEQ ID NO: 5) и Salmonella typhimirium htrA (SEQ ID NO: 6).
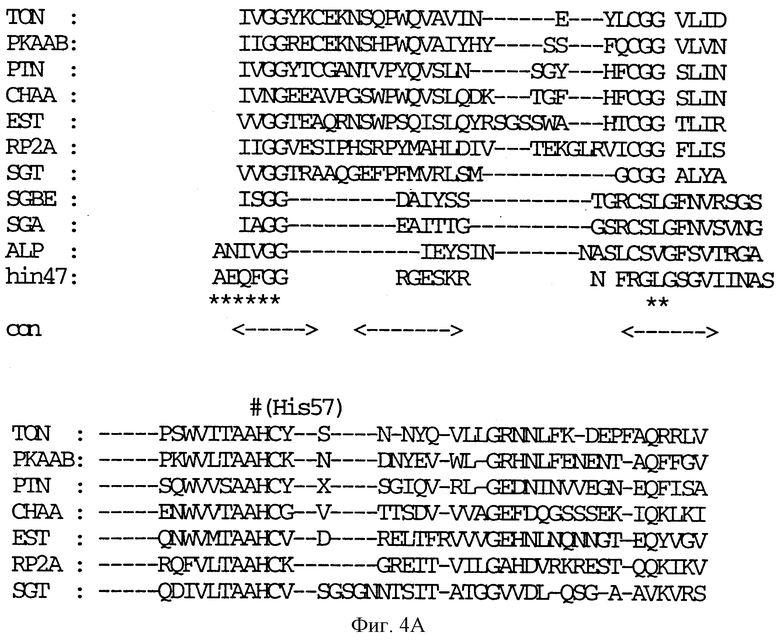
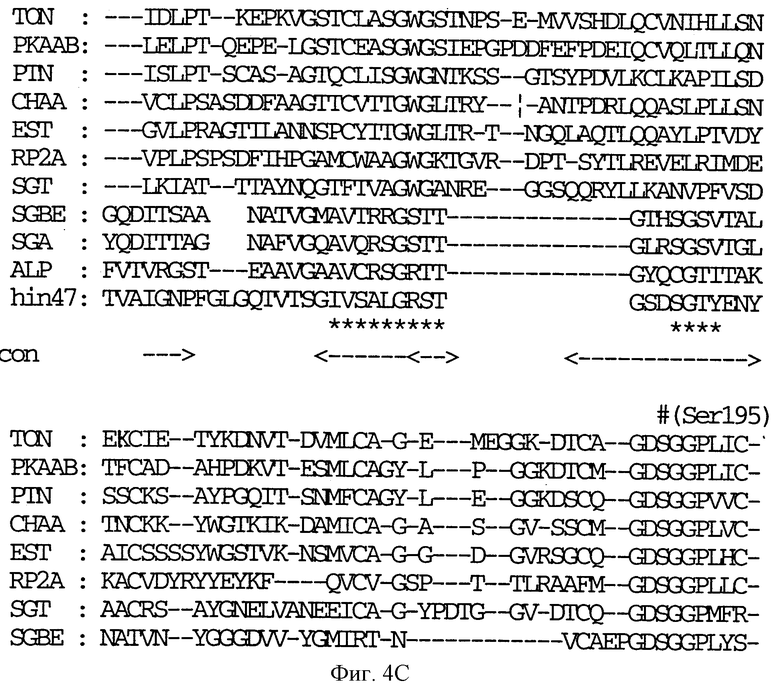
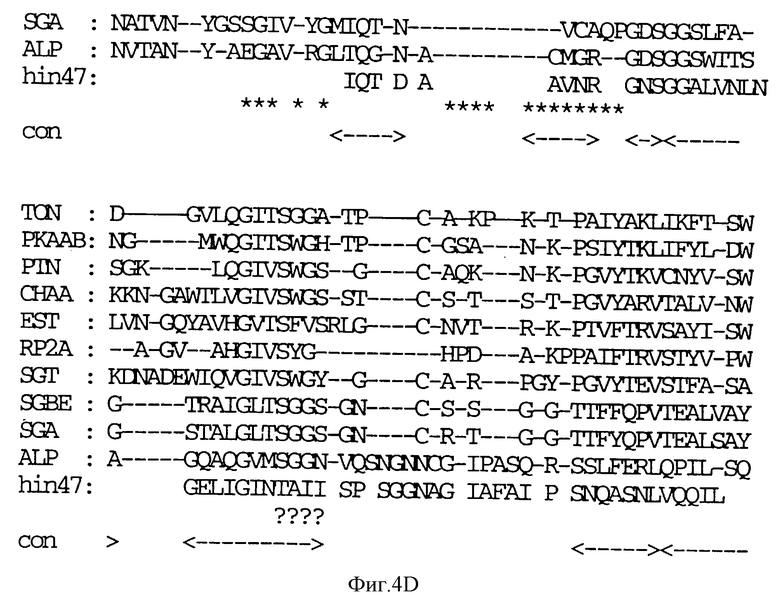
На фиг. 4 A-E показана последовательность аминокислотных остатков Hin47 от 57 до 256 в сравнении с некоторыми известными протеазами (SEQ ID NO: 7-16).
Обозначения следующие: TON, тонин крысы; РКААВ, калликреин; PTN, трипсин; СНАА, химотрипсин; EST, эластаза; RP2A, протеаза тучных клеток крысы; SGBE, протеиназа А S. griseus; SGA, протеиназа В S. griseus; ALP, альфа-литическая протеаза L. enzymogenes; hin47, остатки 57 - 256 Hin47. Звездочкой (*) обозначены структурно консервативные регионы. Триады каталитических остатков обозначены значком (#). 'con' соответствует областям структурного согласования среди протеаз млекопитающих.
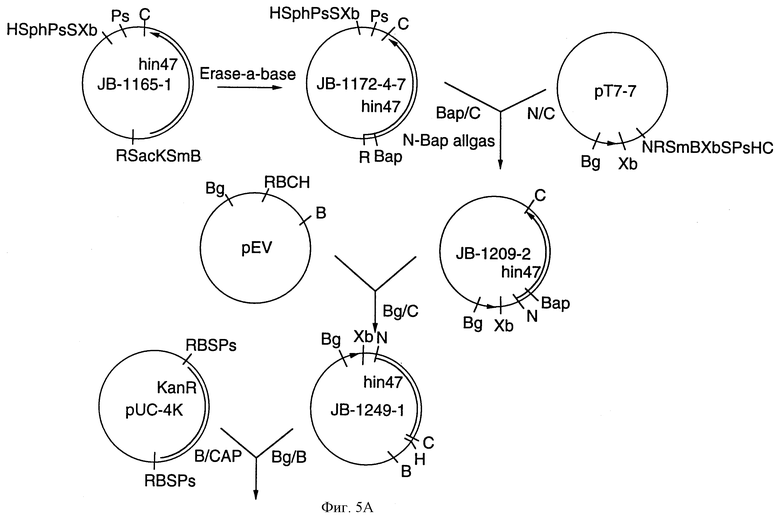
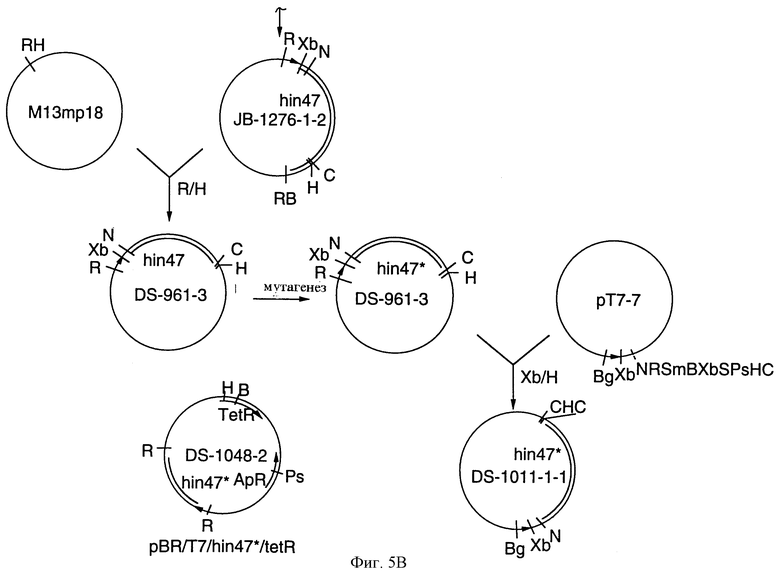
На фиг. 5 А-В представлены рестрикционные карты плазмид DS-1011-1-1 и DS-1048-2, которые экспрессируют аналог Hin47 из Е. coli и конструкционная схема для плазмиды DS-1011-1-1 (плазмида рТ7/Hin47*).
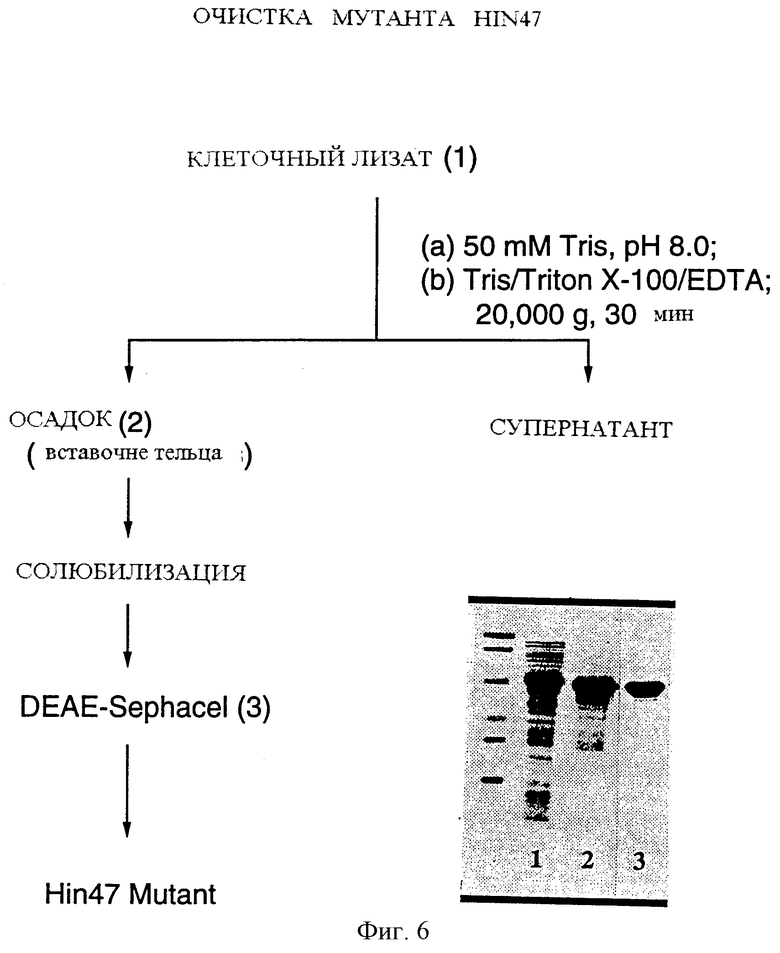
На фиг. 6 показан процесс очистки аналога Hin47 из Е. coli в соответствии с одним из описаний настоящего изобретения и анализ геля очищенного продукта.
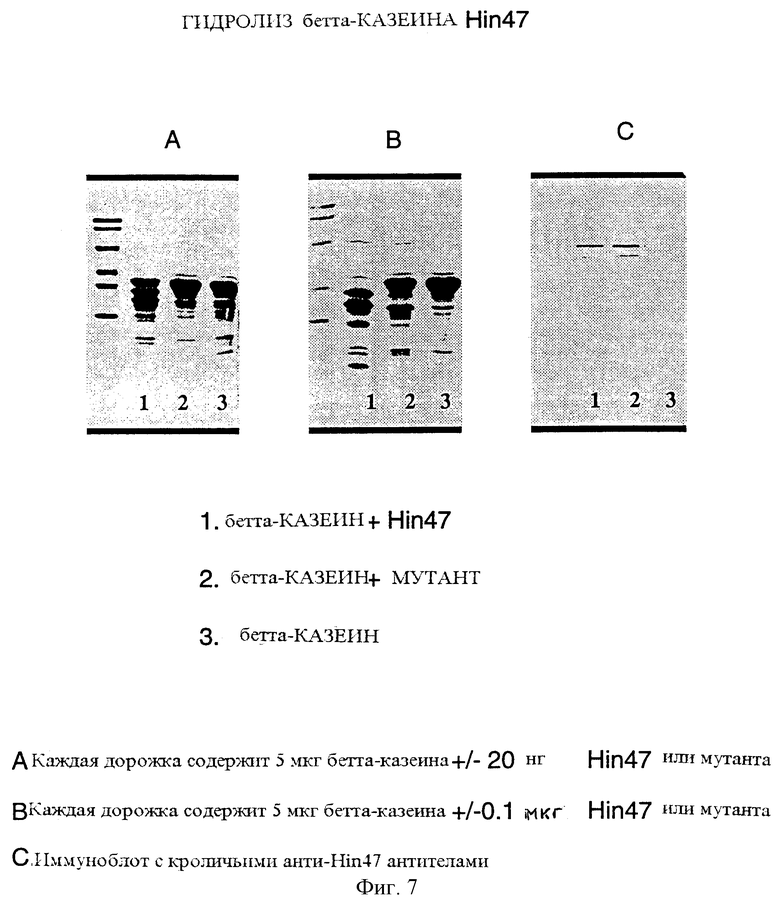
На фиг. 7 показаны протеазные активности природного Hin47 и аналога Hin47 по отношению к бетта-казеину.
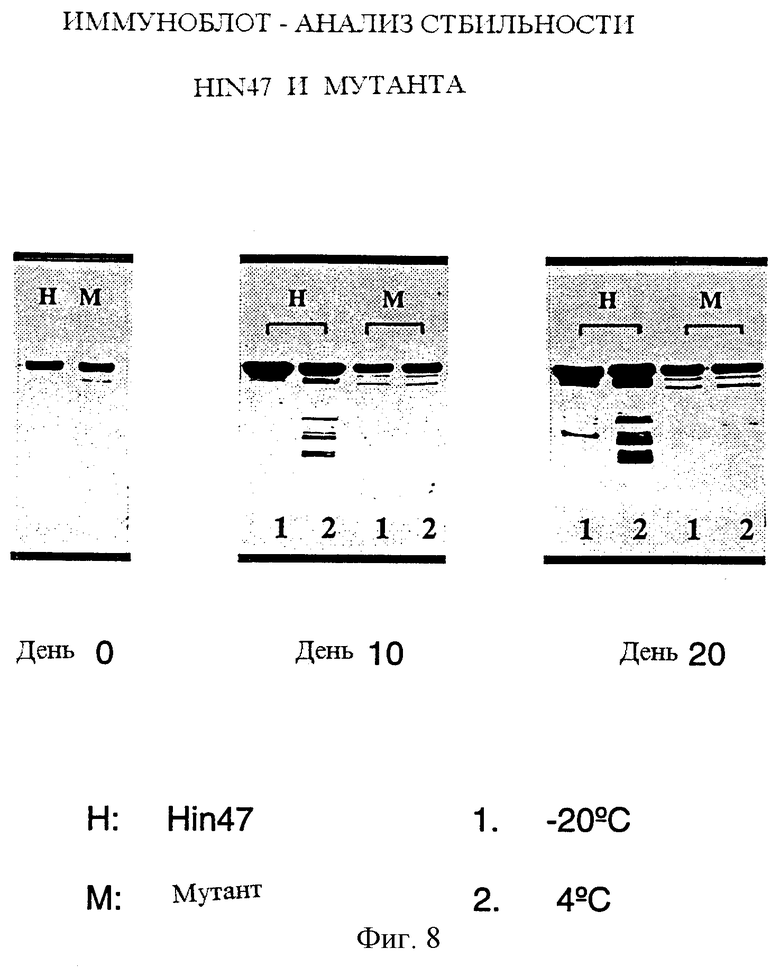
На фиг. 8 показана стабильность природного Hin47 и аналога Hin47 при различных температурах.
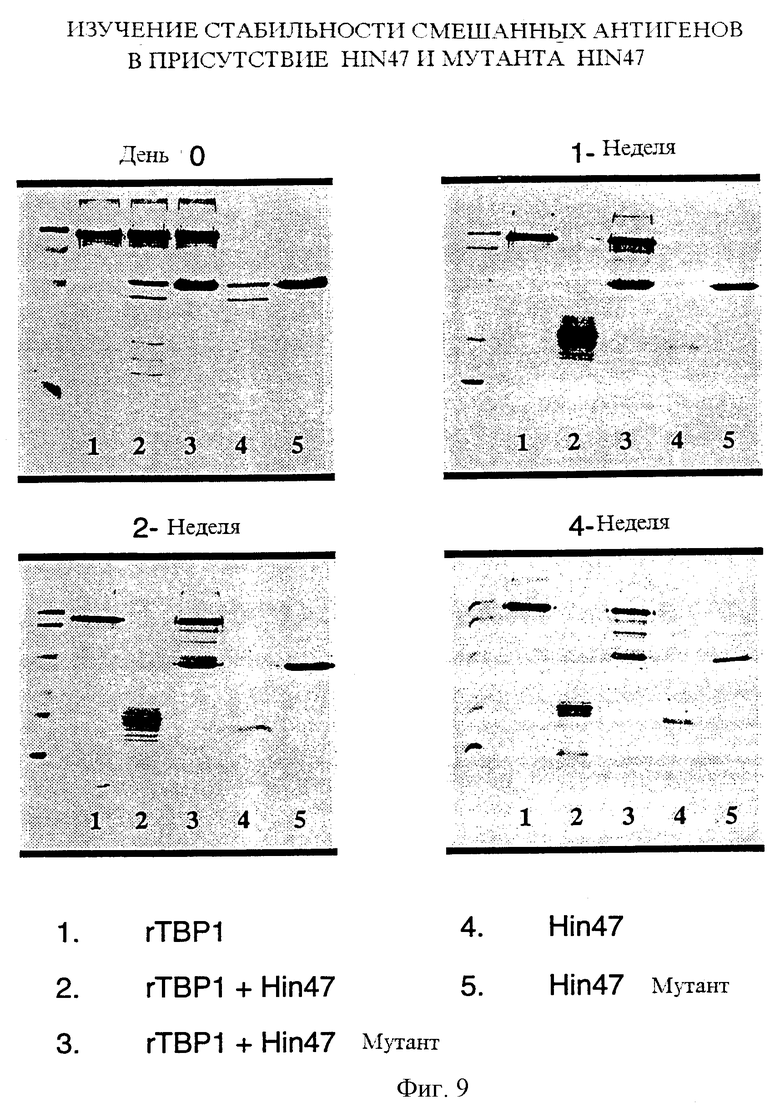
На фиг. 9 показана энзиматическая деградация рекомбинантного белка Н. influenzae под действием природного Hin47 и аналога Hin47.
На фиг. 10 представлена сравнительная иммуногенность природного Hin47 и аналога Hin47 мыши.
На фиг. 11 А-B представлено сравнение аминокислот белка Hin47, выделенного из штаммов SB33 и SB12 Н. influenzae.
На фиг. 12 показана очистка Hin47 аналога Н19А из Е. coli.
Общее описание изобретения
Любые штаммы Haemophilus, которые имеют гены Hin47, могут быть использованы общепринятым способом для очистки и выделения молекул нуклеиновых кислот (которые могут быть в форме молекул ДНК), содержащих, по крайней мере, часть, кодирующую Hin47, как типировано при осуществлениях настоящего изобретения. Такие штаммы обычно доступны из клинических источников и из коллекций бактериальных культур, таких как Американская коллекция типов культур (American Type Culture Collection). Такие штаммы включают штаммы Н. influenzae и других бактерий, которые продуцируют белок, способный образовывать антитела, которые специфически распознают фрагменты Hin47 или его аналогов. Подходящие штаммы Haemophilus могут включать:
Н. influenzae тип b штамм MinnA;
Н. influenzae тип b штамм Еаgаn;
Н. influenzae нетипирующийся штамм SB33;
Н. influenzae нетипирующийся штамм SB12 или
Н. influenzae нетипирующийся штамм РАК 12085.
Что касается фиг. 1, на нем представлены рестрикционные карты плазмид JB-1031-1-14 и JB-1068-2-2, которые содержат часть, кодирующую белок Hin47 из нетипируемого штамма Н. influenzae SB33. Была определена нуклеотидная последовательность гена Hin47, которая представлена на фиг. 2 вместе с выведенной аминокислотной последовательностью белка Hin47. Что касается фиг. 3, на нем представлена аминокислотная последовательность Hin47 Н. influenzae и сериновой протеиназы htrA из Escherichia coli и htrA из Salmonella tyрhimirium. Эта линейная последовательность в первую очередь проявляет неожиданное обнаружение заявителей настоящего изобретения о том, что Hin47 относится к бактериальным сериновым протеазам и что Hin47 обладает протеазной активностью. Как ранее сообщалось, Hin47 является адгезином. Его обнаруженная протеазная активность существенно ограничивает применимость природного Hin47 как иммуногена для вакцинации и как антигена в диагностических целях. Линейная последовательность, представленная на фиг. 3, показывает, что белки htrA и Hin47 содержат GNSGGAL (SEQ ID NO: 17) последовательность между остатками 195 и 201 зрелого белка. Согласованной последовательностью активного центра сериновых протеаз является GDSGGPK (SEQ ID NO: 18) (Brenner, 1988) и активным остатком является серин. Таким образом, Serin-197 в Hin47 мутировал с получением аналога Hin47 с пониженной протеазной активностью в соответствии с осуществлением изобретения. При частном осуществлении Serin-197 был заменен на аланин. Аминокислотные остатки от 57 до 256 Hin47 были далее сличены с известными протеазами и остатки активного центра идентифицированы из локальных гомологий, окружающих остатки каталитической триады (фиг. 4). Существует стандартная система нумерации для сериновых протеаз, у которых остатки каталитической триады пронумерованы как His-57, Asp-102 и Ser-195. Это соответствует остаткам His-91, Asp-121 и Ser-197 в последовательной системе нумерации. Таким образом, что касается фиг. 4, то там представлена линейная структурная основа десяти структурно определенных сериновых протеаз (SEQ ID NOS: 7-16), у которых гомологические остатки первично линейны в основе подобного расположения в трехмерном пространстве. Положение многих остатков в гидрофобном ядре Hin47, а также остатки вокруг активного центра могут быть выстроены вполне приемлемо для идентификации функциональных аминокислот протеазы Hin47. Следовательно, другие аминокислотные остатки в Hin47, которые вносят вклад в протеазную активность белка, включают His-91 и Asp-121. При частном осуществлении His-91 может быть заменен на аланин, лизин или аргинин. При дополнительном осуществлении Asp-121 может быть заменен на аланин или глютаминовую кислоту. При дополнительном осуществлении Serin-197 может быть заменен на аланин, серии или треонин. Хотя приготовление аналога Hin47, имеющего пониженную протеазную активность, путем частичного аминокислотного заменения в белке Hin47, было приведено здесь в примерах, обнаружение протеазной активности и методы экспрессии Hin47, очистки и анализа, предложенные здесь, позволяют получать другие аналоги, обладающие, по крайней мере, одной другой аминокислотой, удаленной или замененной, или обладающие, по крайней мере, одной дополнительной аминокислотой, введенной в белок Hin47. При конкретных применениях и осуществлениях может быть желательно одновременное изменение нескольких аминокислот в белке Hin47 для частичного уменьшения протеазной активности Hin47. Несколькими аминокислотами могут быть His-91 и Ser-197 и могут быть удалены или заменены на аланин. По альтернативному осуществлению несколькими аминокислотами могут быть His-91, Asp-121 и Ser-197 и они могут быть удалены или заменены на аланин. Соответственно, настоящее изобретение касается аналогов белка Hin47, обладающих пониженной протеазной активностью в результате единичного или множественного удаления, заменения или добавления в белке Hin47.
Как обсуждается выше, Hin47 проявляет гомологию с Е. coli htrA или S. typhimirium htrA, оба из которых являются стресс-зависимыми белками с сериновой протеазной активностью. Е. coli включается при росте при температуре 43,5oС (ссылка 13). Заявителями было показано, что белок htrA E. coli также является индуцибельным 6% этанолом. Hin47 также индуцируется 6% этанолом и, в меньшей степени, при доведении температуры до 43,5oС, как это подробно описано далее. Этот анализ экспрессии Hin47 обеспечивает дальнейшее доказательство взаимосвязи между этим белком и LtrA.
Ген hin47 также был клонирован из нетирующегося штамма Н. influenzae SB12 путем PCR амплификации. Что касается фиг. 11, то там приведено сравнение аминокислот между белками Hin47 Н. influenzae штаммов SB12 и SB33. Показана почти полная идентичность аминокислотной последовательности.
Что касается фиг. 5, то там проиллюстрированы плазмиды DS-1011-1-1 и DS-1048-2, которые экспрессируют аналог Hin47 Serin-197 --> аланин у Е. coli. Фиг. 6 показывает последовательную диаграмму метода очистки аналога Hin47 из телец включения Е. coli.
Фиг. 7 показывает пониженную протеазную активность аналога Hin47 Serin-197 --> аланин по отношению к субстрату бетта-казеина и показывает то, что аналог обладает менее чем 10% протеазной активности природного белка Hin47. Таким образом, по одному осуществлению изобретения предложен аналог Hin47, обладающей протеазной активностью, составляющей менее чем 10% от протеазной активности природного Hin47, и такой аналог специфически имеет аминокислоту Serin-197, замененную на аланин.
Что касается фиг. 8, то там представлен анализ повышенной стабильности аналога Hin47, предложенного здесь. Таким образом, одно осуществление настоящего изобретения касается аналога белка Hin47, имеющего повышенную температурную стабильность, и такой аналог может специфически иметь аминокислоту Serin-197, замененную на аланин.
Что касается фиг. 9, то там показана протеолитическая деградация не-Нin47 Haemophilus антигена под действием аналога Hin47 и Hin47, предложенного здесь. Таким образом, в соответствии со следующим осуществлением настоящего изобретения предложен аналог Hin47, совместимый со вторым белком не-Нin47, и такой аналог может специфически иметь аминокислоту Serin-197, замененную на аланин.
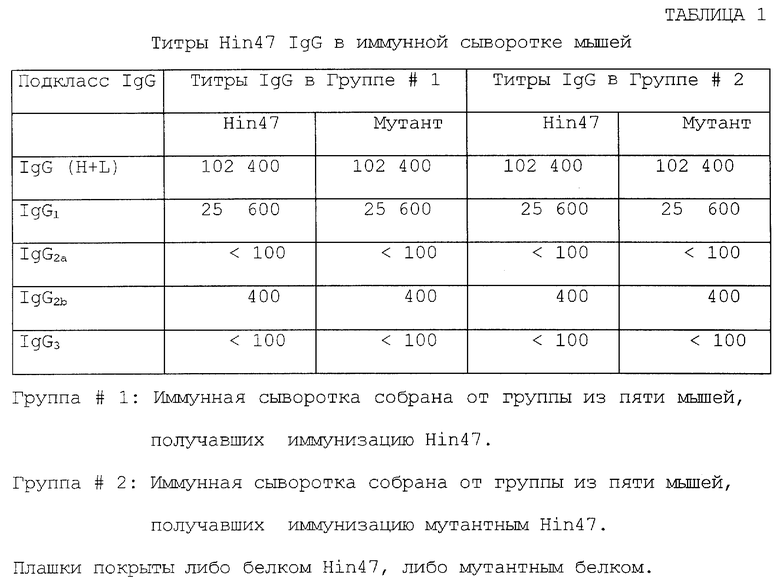
Что касается фиг. 10 и таблицы 1, то там показана сравнительная иммуногенность у мышей немодифицированного Hin47 и аналога Hin47, обладающего пониженной протеазной активностью. Белок Hin47 и аналог Hin47 S197A и Н91А имеют сравнимую иммуногенность. Таким образом, при частном осуществлении предложен аналог Hin47, обладающей пониженной протеазной активностью и имеющий существенно те же иммуногенные свойства природного белка Hin47. Подобный аналог может специфически иметь аминокислоту Serin-197, замененную на аланин.
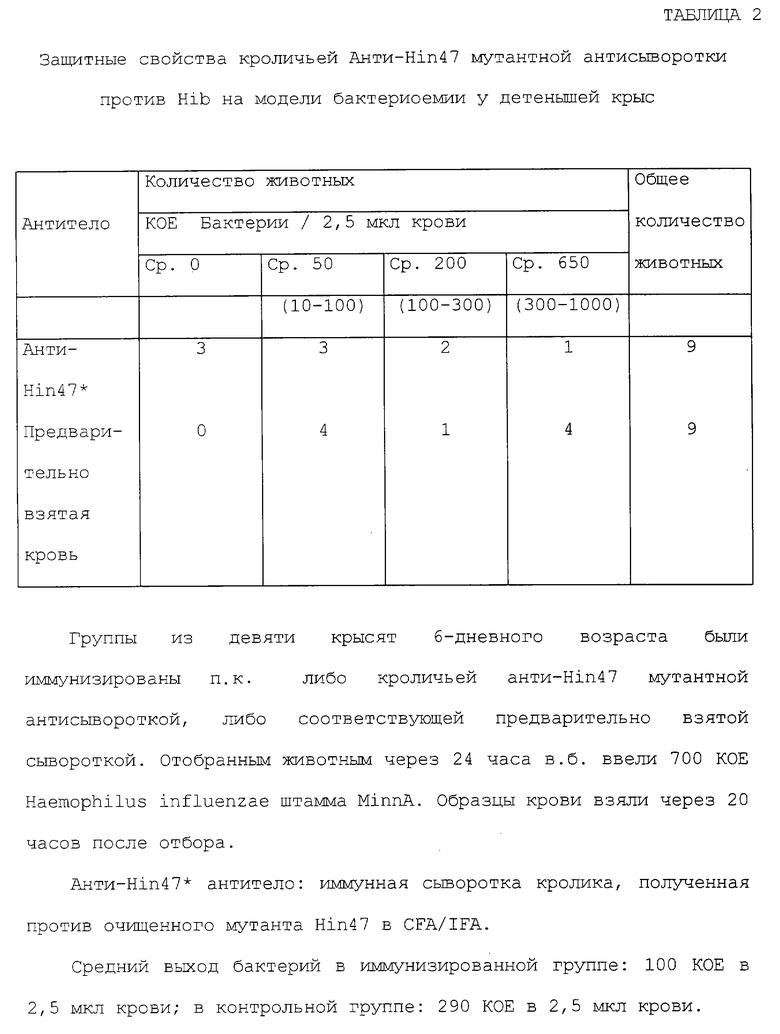
Что касается таблиц 2 и 3, то там представлены иммуногенные свойства аналогов Hin47, обладающих пониженной протеазной активностью по отношению к Hib на модели бактериемии у детенышей крыс и на модели отитов при активной иммунизации шиншилл в соответствии с конкретным осуществлением изобретения, подобный аналог может специфически иметь аминокислоту His-91, удаленную или замененную на аланин, лизин или аргинин; Asp-121, удаленный или замененный на аланин или глютаминовую кислоту; Serin-197, замененный на аланин, цистеин или треонин, или их сочетание.
В соответствии с другим аспектом настоящего изобретения предложена вакцина против Haemophilus или других бактериальных патогенов, которые производят Hin47 или белок, способный к индукции антител, которые специфически распознают Hin47, содержащая иммуногенно-эффективное количество иммунопротективного аналога Hin47, предложенного здесь, или молекулу нуклеиновой кислоты, обладающей последовательностью, кодирующей Hin47, предложенный здесь, и их физиологически приемлемый носитель. Предложенные аналоги также могут быть использованы как белки переносчики для гаптена, полисахаридов или пептидов для создания коньюгатной вакцины против антигенных детерминант, не родственных Hin47.
Как будет ясно из следующего описания, настоящее изобретение далее относится к плазмидам и новым штаммам бактерий для продукции аналогов Hin47, предложенных здесь.
Очищенные и выделенные молекулы ДНК, содержащие, по крайней мере, часть, кодирующую аналог белка Haemophilus influenzae Hin47, обладающий пониженной протеазной активностью по сравнению с природным Hin47, типированным путем осуществления описанного здесь, являются улучшенными в качестве проб нуклеиновых кислот для специфического обнаружения штаммов Haemophilus in vivo и in vitro. Аналоги Hin47, закодированные молекулами ДНК, предложенными здесь, используются в качестве диагностических реагентов, как антигены или для производства анти-Нin47 антител, антигенов для вакцинации против заболеваний, вызываемых видами Haemophilus и другими бактериальными патогенами, которые продуцируют белок, способный к продуцированию антител, которые специфически распознают Hin47 и для определения инфицирования Haemophilus и другими подобными бактериями.
При дополнительном осуществлении настоящего изобретения аналог Hin47, имеющий пониженную протеазную активность, предложенный здесь, может быть использован в качестве переносчика молекул для приготовления молекул химер и конъюгатов вакцин (включая гликоконъюгаты) против патогенных бактерий, включая инкапсулированные бактерии. Таким образом, например, гликоконъюгаты по настоящему изобретению могут быть применены для вакцинации, чтобы выработать защиту против заболевания и инфекции, вызываемой любой другой бактерией, имеющей полисахаридный антиген, включая липополисахариды (ЛПС) и PRF. Бактериальные патогены могут включать, например, Haemophilus influenzae, Strptococcus pneumonae, Niesseria meningitides. Salmonella typhi, Streptococcus mutans, Cryptococcus neophormans, Klebsiella, Staphylococcus aureus и Pseudomonas aeruginosa. Конкретные антигены, которые могут быть конъюгированы с аналогами Hin47 и методы для получения таких коньюгатов описаны в приложениях опубликованной заявки РСТ WO 94/12641, которая, кроме того, включена здесь в ссылки.
В следующем осуществлении функция переносчика аналогов Hin47 может быть использована, например, для индукции иммунитета по отношению к анормальным полисахаридам опухолевых клеток или для продукции антиопухолевых антител, которые могут быть конъюгированы к химиотерапевтическим или биоактивным агентам.
Соответственно, настоящее изобретение касается первичной последовательности и получения аналогов Hin47 H. influenzae, которые могут быть использованы для профилактики и диагностики заболеваний, вызываемых H. influenzae. В частности, заявители обнаружили, что аналоги Hin47 могут вырабатывать защитные иммунные ответы против бактериальной провокации живыми H. influenzae тира b. Таким образом, настоящее изобретение имеет полезность для вакцин. Изобретение также описывает нуклеотидные последовательности генов, кодирующих аналоги Hin47. Эти сегменты ДНК могут быть использованы для создания иммуногена, практически свободного от других антигенов H. influenzae, таких как PRP и липополисахариды (ЛПС), путем применения технологии с рекомбинантной ДНК технологии. Аналог белка Hin47 может быть получен в подходящих системах для экспрессии, таких как Е. coli, Haemophilus, Bacillus, Bordetella, грибы, дрожжи, бакуловирус, поксивирус, вирус коровий оспы, или системах экспрессии млекопитающих. Настоящее описание далее включает новые методы, которые могут быть применены для получения практически чистых аналогов Hin47.
Специалисту понятно, что различные осуществления настоящего изобретения имеют множество применений в областях вакцинации, диагностики, лечения, например, инфекций Haemophilus и инфекций других бактериальных патогенов, которые производят белки, способные для продукции антител, которые специфически распознают Hin47, и в области производства иммунологических реагентов. Дальнейшее не ограничивающее обсуждение таких применений представлено далее.
1. Препарат вакцины и применение
Иммуногенные композиции, удобные для применения в качестве вакцины, могут быть приготовлены из аналогов Hin47, как здесь описано. Вакцина усиливает иммунный ответ у субъекта, который продуцирует антитела, включая анти-Нin47 антитела и антитела, которые являются опсонизирующими или бактерицидными. Вакцинируемый объект должен быть отзывчивым на Haemophilus или другие бактерии, которые продуцируют белки, способные вырабатывать антитела, которые специфически распознают Hin47, антитела связывают и инактивируют бактерию. Более того, опсонизирующие или бактерицидные анти-Нin47 могут также обеспечивать защиту путем альтернативных механизмов.
Иммуногенная композиция, включающая вакцины, может быть получена в виде, пригодном для инъекций, в виде жидких растворов или эмульсий. Аналоги Hin47 могут быть смешаны с фармацевтически приемлемыми наполнителями, которые совместимы с аналогом Hin47. Такие наполнители могут включать воду, физраствор, декстрозу, глицерин, этанол и их сочетание. Иммуногенная композиция и вакцины могут, кроме того, содержать вспомогательные вещества, такие как увлажняющие или эмульгирующие агенты, рН буферирующие агенты или адъюванты, для улучшения их эффективности. Методы для достижения действия адъюванта включают применение агентов, таких как гидроксид или фосфат алюминия, обычно используемых в виде 0,05-0,1 процентного раствора в фосфатном буфере на физиологическом растворе. Иммуногенные композиции и вакцины могут вводиться внутрь путем инъекции подкожно или внутримышечно. Альтернативно иммуногенные композиции, образованные в соответствии с настоящим изобретением, могут быть составлены и доставлены для вызова иммунного ответа на слизистые оболочки. Таким образом, иммуногенная композиция может доставляться на слизистые оболочки, например, через носовые или ротовые (внутрижелудочные) пути. Альтернативно могут быть желательны другие методы введения, включающие суппозитории и пероральные составы. Для суппозиториев связывающие носители могут включать, например, полиалкеновые гликоли или триглицериды. Пероральные составы могут включать обычно используемые добавки, такие как, например, фармацевтически чистый сахарин, целлюлозу и карбонат магния. Эти композиции могут иметь вид растворов, суспензий, таблеток, пилюль, капсул, составов с поддерживающим высвобождением или порошков и содержать от 1 до 95 % аналогов Hin47. Иммуногенные препараты и вакцины вводятся методом, совместимым с дозировкой препарата, и в таких количествах, которые будут терапевттически эффективными, профилактическими и иммуногенными. Вводимое количество зависит от субъекта, подвергающемуся лечению, включая, например, способность индивидуальной иммунной системы для синтеза антител, и, если необходимо, для продукции клеток, опосредующих иммунный ответ. Точные количества активного ингредиента, требуемого для введения зависит от оценки лечащего врача. Однако области подходящих дозировок легко определимы специалистом и могут составлять порядка микрограмм для аналога Hin47. Подходящие режимы для начального введения и ударных доз также является вариабельными, но могут включать начальное и последующее за ним введение. Дозировки могут также зависеть от путей введения и будут варьировать в соответствии с размером хозяина.
Концентрация антигена в иммуногенной композиции, в соответствии с изобретением, составляет обычно от 1 до 95 %. Вакцина, которая содержит антигенный материал только одного патогена, являются моновалентной вакциной. Вакцины, которые содержат антигенный материал от нескольких патогенов, являются комбинированными вакцинами и также относятся к настоящему изобретению. Такие комбинированные вакцины содержат, например, материал от различных патогенов или от различных штаммов того же самого патогена, или от сочетаний различных патогенов.
Молекулы нуклеиновых кислот, кодирующие аналог Hin47 по настоящему изобретению, могут также использоваться непосредственно для иммунизации путем непосредственного введения ДНК, например путем инъекции для генетической иммунизации или путем создания живого вектора, такого как Salmonella, BCG, аденовирус, поксивирус, вирус коровий оспы или полиовирус. Обсуждение некоторых живых векторов, которые были использованы для перенесения гетерологических антигенов в иммунную систему рассматривается, например, O'Наgаn (1992). Способы прямой инъекции ДНК в исследуемые субъекты для генетической иммунизации описаны, например, Ulmer et al, 1993.
2. Иммуноанализы
Аналоги Hin47 по настоящему изобретению используются в качестве иммуногенов для наработки анти-Нin47 антител, в качестве антигенов для иммуноанализов, включая анализы на ферментах, пришитых к сорбенту (ELISA), радиоиммуноанализы (RIA) и другие анализы связывания неэнзиматически пришитых антител или методы, известные в данной области для обнаружения антибактериальных, Haemophilus и анти-Нin47 антител. При анализах ELISA аналоги Hin47 иммобилизуют на выбранной поверхности, например поверхности, способной к связыванию белков, такой как полистерольные микротитровальные планшеты. После отмывания для удаления не полностью абсорбированных аналогов Hin47 с выбранной поверхностью может быть связан неспецифический белок, такой как бычий сывороточный альбумин (BSA) в растворе, который, как известно, является антигенно нейтральным по отношению к исследуемому образцу. Это позволяет блокировать места неспецифической адсорбции на иммобилизационной поверхности и, таким образом, подавлять фон из-за неспецифического связывания антисыворотки к поверхности.
Иммобилизационная поверхность затем контактирует с образцом, таким как клинический или биологический материалы, исследуемые методом, способствующим образованию иммунного комплекса (антиген/антитело). Это может включать разбавление образца разбавителями, такими как растворы BSA, бычий гамма- глобулин (BGG) и/или фосфатный буфер на физрастворе (РВS)/Твин. Образец затем инкубируют от 2 до 4 часов при температуре, такой как положено, от 25 до 37oС. После инкубации поверхность, контактирующую с образцом, отмывали для удаления неиммунокомплексного материала. Процедура отмывки может включать отмывку раствором, таким как РВS/Твин или боратным буфером. Последующее образование специфических иммунокомплексов между исследуемым образцом и привязанными аналогами Hin47, последовательное промывание, обнаружение равного количества образования иммунокомплексов могут быть определены помещением иммунокомплекса к второму антителу, имеющему специфичность к первому антителу. Если исследуемый образец человеческой природы, второе антитело является антителом, обладающим специфичностью к человеческим иммуноглобулинам и обычно к IgG. Для обеспечения обнаружения второе антитело может иметь ассоциированную активность, такую как ферментативная активность, которая будет производить, например, развитие цветной окраски при инкубации с подходящим хромогенным субстратом. Количественное определение затем может достигаться измерением степени развития окраски, используя, например, спектрофотометр в видимом спектре.
3. Использование последовательностей как гибридизационных зондов
Молекулы нуклеиновых кислот по настоящему изобретению, имеющие последовательность аналога гена hin47, позволяют идентифицировать и клонировать гены Hin47 из любых видов Haemophilus и других бактерий, которые продуцируют белки, способные к образованию антител, которые специфически распознают Hin47.
Молекулы нуклеиновых кислот, обладающие последовательностью, кодирующей аналог Hin47 по настоящему изобретению, используются в связи с их способностью селективно образовывать дуплексные молекулы, которые комплементарно вытягивают другие гены hin47. В зависимости от применения могут использоваться разнообразные условия гибридизации для достижения различной степени селективности зондов по отношению к другим генам hin47. Для высокой степени селективности используются относительно строгие условия для образования дуплексов, такие как низкая концентрация соли и/или высокотемпературные условия, такие как создание 0,02 М-0,15 М концентрации NaCl при температуре между 50 и 70oС. Для некоторых применений требуются менее строгие условия гибридизации, такие как 0,15 М-0,9 М соль при температуре около 20-55oС. Гибридизационные условия могут также более строго поддерживаться путем добавлении увеличивающихся количеств формамида для дестабилизации гибридного дуплекса. Таким образом, конкретные гибридизационные условия могут быть легко манипулируемыми и, будут обычно методом выбора, зависящим от желаемого результата.
При клиническом диагностическом осуществлении молекулы нуклеиновых кислот, кодирующие гены hin47 по настоящему изобретению, могут быть использованы для определения гибридизации в комбинации с подходящими приспособлениями, такими как метка. В данной области известно широкое разнообразие подходящих индикаторов, включая радиоактивные, ферментативные или другие лиганды, такие как авидин/биотин, которые способны обеспечить обнаружение сигнала. При некоторых диагностических осуществлениях ферментная метка, такая как уреаза, щелочная фосфатаза или пероксидаза, может быть использована вместо радиактивной метки. В случае ферментных меток известны колориметрические индикаторные субстраты, которые могут быть использованы для создания приспособления, видимого для человеческого глаза или спектрофотометрически для идентификации специфической гибридизации с образцами, содержащими hin47 генную последовательность.
Молекулы нуклеиновых кислот, содержащие гены hin47 по настоящему изобретению, используются в качестве гибридизационных зондов в гибридизационных растворах и в осуществлениях, использующих твердофазные методы. При осуществлениях, включающих твердофазные методы, исследуемая ДНК (или РНК) из образцов, таких как клинические образцы, включая эксудаты, жидкости организма (например, сыворотка, амниотическая жидкость, выделения из среднего уха, сера, бронхоальвеолярная жидкая мокрота) или даже ткани, адсорбирована или прикреплена другим образом к выбранному матриксу или поверхности. Фиксированная однотяжевая нуклеиновая кислота подвергается затем специфической гибридизации с выбранными пробами, содержащими последовательность нуклеиновых кислот генов hin47 по настоящему изобретению в желаемых условиях. Выбранные условия будут зависеть от конкретных обстоятельств, основанных на конкретном требуемом критерии, зависящем, например, от содержания G+C, типа нуклеиновой кислоты мишени, источника нуклеиновой кислоты, размера гибридизационной пробы и т.д. После отмывания гибридизационной поверхности так, чтобы удалить не специфически связанные молекулы зонда, определяется или даже подсчитывается специфическая гибридизация посредством метки.
4. Экспрессия генов, кодирующих аналоги Hin47, обладающие пониженной протеазной активностью
Векторы, возможно, содержащие репликон и контролирующие последовательности, которые происходят от видов, совместимых с клетками хозяина, могут быть использованы для экспрессии генов аналога Hin47, как обусловлено здесь в экспрессионных системах. Вектор обычно переносит репликационный участок, а также маркированные последовательности, которые способы обеспечить фенотипическую селекцию в трансформированных клетках. Например, Е. coli может быть трансформирована с использованием pBR322, которая содержит гены резистенции для ампициллина и тетрациклина, и тем самым легко обеспечивает способы для идентификации трансформированных клеток. pRB322 плазмида или другая микробная плазмиды или фаг должны также содержать или быть модифицированными для содержания, промоторы, которые могут быть использованы клетками хозяина для экспрессии его собственных белков.
Кроме этого, фаговые векторы, содержащие репликон и контрольные последовательности, которые совместимы с хозяином, могут быть использованы как трансформирующие векторы при сношении с этими хозяевами. Например, фаги у лямбда GEMTM-11 могут использоваться в создании рекомбинантных векторов фагов, которые могут быть использованы для трансформации клеток хозяина, таких как Е. coli LE932.
Промоторы, обычно используемые в конструкциях рекомбинантной ДНК, включают бетта-лактамазу (пенициллиназу) и системы лактозного промотера (Chaang et al, 1979; Goeddel et al, 1980) и другие микробные промотерные системы (патент США 4952496). Детали, касающиеся нуклеотидных последовательностей промотеров, известны, и дают возможность специалисту функционально пришить их к плазмидным векторам. Конкретный используемый промотер обычно будет предметом выбора, зависящим от желаемых результатов. Могут быть использованы хозяева, которые подходят для экспрессии аналогов Hin47, включающие Е. coli, виды Bacillus, Haemophilus, Bordetella, грибки, дрожжи, клетки млекопитающих или систему экспрессии бакуловируса.
Таким образом, в соответствии с изобретением может быть предпочтительным создать аналог белка Hin47 рекомбинантными методами. Особенно желательные хозяева для экспрессии в соответствии с этим включают грамположительные бактерии, которые не имеют LPS и, следовательно, свободны от эндотоксина. Такие хозяева включают виды Bacillus и могут быть, в частности, использованы для продуцирования непирогенного аналога Hin47.
Биологические депозиты
Плазмида DS-1011-1-1 (pT7/Hin47*), которая содержит часть, кодирующую аналог Hin47, которая описана и приведена здесь в виде ссылки, была депонирована на хранение в American Type Culture Collection (ATCC), расположенную в Роквилле, Мэриленд, США (Rockwill), согласно Будапештскому Соглашению (Budapest Treaty)и до подачи данной заявки, являющейся продолжением части заявки от 27 июля 1994 под входящим N 75845. Образцы депонированных плазмид стали доступны общественности благодаря выдаче патента, основанного на данной патентной заявке США. Описываемое и заявляемое здесь изобретение не ограниченно депонированной плазмидой, поскольку депонированное осуществление предназначено лишь для иллюстрации изобретения. Любые эквивалентные или подобные плазмиды, которые кодируют подобные или эквивалентные антигены, как описано в этой заявке, включены в объем данного изобретения.
Примеры
Вышеприведенное описание в целом описывает настоящее изобретение. Более полное понимание может быть получено при отсылке к следующим специфическим примерам. Эти примеры описаны только для целей иллюстрации и не предназначены для ограничения объема изобретения. В зависимости от предлагаемых обстоятельств или целесообразности могут быть рассмотрены изменения в форме и замена эквивалентными. Хотя здесь использованы специфические термины, такие термины применены в описательном смысле, а не в целях ограничения.
Использованные методы молекулярной генетики, биохимии белков и иммунологии в этом описании описаны без объяснений, и данные примеры достаточно широко описаны в научной литературе и легко доступны специалисту.
Пример 1.
Данный пример иллюстрирует клонирование гена hin47 из нетипирующегося штамма SB33 Н. influenzae.
Из штамма SB33 Н. influenzae была получена хромосомальная ДНК и была получена библиотека EMBL3 и просеяна с меченным олигонуклеотидным зондом, специфическим для 5'-конца hin47. Нетипирующийся штамм SB33 Н. influenzae был выращен на Muellar-Hinton агаре или на инфузионном сердечно-мозговом бульоне, как это описано Harkness et al, 1992. Хромосомальная ДНК была получена следующим образом: клетки из 50 мл культуры были осаждены центрифугированием при 5000 об/мин в течение 15-20 мин при 4oС в Sorwall RC-3B центрифуге. Осадок клеток ресуспендировали в 10 мл ТЕ (10 мл Трис/HCl, 1 мМ ЭДТА, рН 7,5), затем добавлена проназа до 500 мкг/мл и SDS до 1%. Образец инкубировали при 37oС до получения прозрачного лизата. Лизат мягко один раз экстрагировали Трис-насыщенным фенолом (рН 7,4) и один раз Трис-насыщенным фенол/хлороформом (1:1) и один раз хлороформом. Конечную водную фазу диализовали при 4oС в течение 24 часов против 1М NaCl и затем в течение 24 часов против ТЕ.
EMBL3 библиотека была получена путем частичного расщепления SB33 хромосомной ДНК с Sau3A I и последующего фракционирования по размерам либо в градиенте сахарозы 10-30% на TNE (20 мМ Трис/HCl, 5 мM NaCl, 1 мМ ЭДТА, рН 8,0) либо препаративным гель-электрофорезом. Фракции, содержащие фрагменты ДНК более чем 5 кб длинной были собраны, осаждены и привязаны к ВаmН I плечам EMBL3 (Promega). Лигационная смесь была упакована, используя Gigapack II packaging kit и внесена к клеткам Е. coli. Библиотеку размножили и хранили при 4oС в присутствии 0,3% хлороформа.
Бляшки были помещены на нитроцеллюлозные фильтры для гибридизации с 32Р-меченными нуклеотидными зондами (3026.SL). Нуклеотидная последовательность была следующей ATGAAAAAAACACGTTTTGTATTAAATAGTATTGCACTTGG (SEQ ID NО: 3), соответствующей N-концу аминокислотной последовательности MKKTRFVLNSIALG (SEQ ID NO: 19). Фаговая ДНК была приготовлена из естественных бляшек и введенная ДНК была вырезана при расщеплении с Sal I и клонирована в pUCS-BgXb, расщепленных с Sal I. Плазмиды JB-1031-1-14 и JB-1068-2-2 (фиг. 1) были отобраны для дальнейшего анализа.
Пример 2.
Данный пример иллюстрирует характеристику и анализ последовательности гена hin47 и выведенной аминокислотной последовательности белка Hin47 из NTHi штамма SB33.
Рестрикционное картирование и Sauthern blot анализ клонов JB-1031-1-14 и JB-1068-2-2 локализовали ген hin47 на 4,7 кб ВаmН I/ВаmН I или на 2,7 кб ВаmН I/Pst I фрагменте ДНК. 4,7 кб ВаmН I/ВаmН I фрагмент из JB-1068-2-2 был субклонирован в pUC8/BgXb генерирующую плазмиду DS-755-1. Фрагмент от 3,1 кб ВаmH I до Хbа I DS-755-1 субклонировали в pUC18 генерирующую плазмиду JB-1165-1, которая имеет рестрикционные участки, подходящие для Erase-a-base (Promega) процедуры (фиг. 1). Эта техника дает успешные клоны с увеличенной усекаемостью введенной ДНК, с наблюдаемыми
делециями на том же конце. Конечный укомплектованный набор клонов может быть быстро секвенирован, используя универсальный праймер.
ДНК плазмиды JB-1165-1 была расщеплена с помощью ВаmН I и Sac I и подвергнута exoIII расщеплению, используя Erase-a-base набор. Конечный набор усеченных плазмид анализировали путем гель-электрофореза на агарозе и характерные плазмиды были отобраны для анализа последовательности.
Плазмидная ДНК для секвенирования была приготовлена согласно процедуре Holmes и Quigley, 1981, с модификациями. Коротко, осадок клеток из 50 мл культуры ресуспендировали в 10 мл STET (8% сахарозы, 5% Тритон Х-100, 50 мМ ЭДТА и 50 мМ Трис/HCl, рН 8,0), затем к смеси был добавлен лизоцим (2,5 мг) и смесь прокипятили 2 минуты. Образец открутили при 14000 об/мин в центрифуге Sorvall RC 5В в течение 20 минут и осадок преципитировали с равным количеством изопропанола, отмыли 70% этанолом и затем абсолютным этанолом, затем высушили досуха воздухом. Осадок ресуспендировали в 0,9 мл ТЕ, затем добавили 20 мкл РНКазы I в концентрации 5 мг/мл и затем смесь инкубировали при 37oС в течение 15 минут. После добавления 500 мкл 1,5 М NaCl/30% PEG смесь инкубировали на льду в течение 30 мин и осаждали ДНК центрифугированием в микрофуге Eppendorf в течение 10 минут. Осадок ресуспендировали в 400 мкл ТЕ и дважды экстрагировали Трис насыщенным фенолом (рН 7,4), дважды Трис насыщенным фенол/хлороформом (1:1) и дважды хлороформом. ДНК преципитировали путем добавления 40 мкл 3 М ацетата аммония и 1 мл этанола, промывали 70% этанолом и ресуспендировали в дистиллированной воде.
Образцы ДНК секвенировали, используя ABI модель 370А секвенатора ДНК и методы химии концевых красителей. Универсальный реверсный праймер был использован вместе с гнездованным набором клонов для определения последовательности hin47, кодирующей нити. Олигонуклеотидный праймер приблизительно в 25 баз длиной был использован для подтверждения последовательности не кодирующей нити. Нуклеотидная последовательность гена hin47 SВ33 и выведенная аминокислотная последовательность белка Hin47 представлены на фиг. 2. Нуклеотидная и аминокислотная последовательность N-конца Hin47, представленные на собрании ASM, Новый Орлеан, 26 мая 1992 года, характеризуются низкой случайностью, см. фиг. 2. Аминотерминальная последовательность SB33 Hin47 и данная представленная последовательность являются идентичными, что устанавливает идентичность клонированного гена как hin47.
Пример 3.
Данный пример описывает обнаружение сериновой протеазной активности белка Hin47.
Выведенную аминокислотную последовательность белка Hin47, определенную выше в примере 2, сравнивали с другими известными белками базы данных Genebank. Как описано выше, белок Hin47, описанный в опубликованных заявках РСТ WO 94/001149, WO 92/11367 и WO 92/110936, представляет молекулу адгезина Haemophilus. Поэтому это было удивительным и неожиданным обнаружением настоящего изобретения, что Hin47 имеет значимую гомологию (55%) с сериновыми протеазами Е. coli htrA и S. typhimurium htrA и другими протеазами. Эти гомологии аминокислотной последовательности показаны на фиг. 3 и 4. Более того, было обнаружено, что белок Hin47 самопереваривается, несмотря на то что хранится в присутствии ингибитора сериновых протеаз, такого как Pefablock.
Пример 4.
Данный пример иллюстрирует образование мутантного гена hin47 путем сайт-направленного мутагенеза.
Как объяснено выше, Н. influenzae Hin47, Е. coli htrA и S. typhimurium htrA - все являются сериновыми протеазами. Общая последовательность всех активных центров сериновых протеаз GDSGGPK (SEQ ID NO: 18) [Brenner, 1988] с серином, являющимся активным остатком. Оба белка htrA имеют последовательность GNSGGAL (SEQ ID NO: 17) и последовательность Н. influenzae Hin47 идентична последовательности между остатками 195 и 201 зрелого белка. Таким образом, сериновый остаток в положении 197 был выбран для сайт-направленного мутагенеза для получения аналога Hin47 с пониженной протеазной активностью.
Был синтезирован олигонуклеотид CGCTCCACCAGCATTACCGCGG (SEQ ID NO: 22), который должен был заменить сериновый остаток 197 на Аланин. Ген hin47 клонировали в клон М13mр18 генерирующего клона DS-981-8 и проведен мугагенез, используя набор фирмы Amersham "In Vitro Site Directed Mutagenesis Kit". Клон DS-991-8 был подтвержден анализом на содержание мутации замены Serin-197 на аланин. Мутантный hin47 ген был обозначен hin47*. Используя подходящие олигонуклеотиды, сериновый остаток 197 заменили на цистеин (мутант S197C) и на треонин (мутант S197T).
Кроме того, сравнение аминокислотной последовательности Hin47 с другими протеазами (как показано на фиг. 4), показало, что аминокислоты His-91 и Asp-121 являются местами, подходящими для мутагенеза для получения аналога Hin47, с пониженной протеазной активностью. С помощью методов мутагенеза, аналогичных описанным выше, His-91 и/или Asp-121 были удалены или заменены на различные аминокислоты. Такие замены аминокислот включали His-91 на Аланин (мутант Н91А) и Аргинин (мутант H91R) и Asp-121 на Аланин (мутант D121A) и Глютаминовую кислоту (мутант D121E). Олигонуклеотиды для получения такого мутагенеза включали: His-91 --> Ала-91 5' ATCAATAACAGCATATGGT 3' (SEQ ID NO: 21) Asp-121 --> Ала121 5' TAATGCAATTGCTGATAGTTC 3' (SEQ ID NO: 22).
Были использованы соответствующие нуклеотиды для проведения других мутаций. Были получены также множественные мутации, в которых His-91 и Ser-197 оба были заменены на Аланин (мутант H91A/S197A) и His-91, Asp-121 и Ser-197 были все заменены на Аланин (мутант H91A/D121A/S197A).
Эти дополнительные мутанты были получены, выделены, очищены и протестированы на протеазную активность, как описано для материала Hin47* в следующих примерах.
Многие сериновые протеазы выделяются в неактивной ('зимоген') форме и требуется вырезка для проявления их активных участков. Анализ N терминальной последовательности зрелого природного белка Hin47 подтверждает, что расщепление препротеина наблюдается в положении KFFFG DRFAAEQ (SEQ ID NO: 23). Модификации аминокислот, которые препятствуют расщеплению молекулы для получения активной протеазной молекулы, могут производить аналог Hin47, обладающий пониженной протеазной активностью.
Пример 5.
Данный пример иллюстрирует конструкцию плазмид, экспрессирующих аналог Ser-197 --> аланин Hin47 Е. coli.
Мутированный ген hin47* из плазмиды DS-91-8 клонировали в рТ7-7 экспрессионный вектор для получения плазммиды DS-1011-1-1 (фиг. 5). Штамм Е. coli BL21/DE3 трансформировали для получения штамма DS-1018-3-1 Е. coli, который экспрессирует аналог Ser-197 --> аланин Hin47 после индукции.
Для того чтобы использовать тетрациклиновую селекцию, ген hin47* клонировали в pBR328. Фрагмент гена Bgl II/CIa I T7/hin47* из DS-1011-1-1 бьш клонирован в pEVvrfl (Young and Davis, 1985) для того, чтобы произвести фрагмент Bgl II/BamH I, который должен быть клонирован в pUC-4K (Pharmacia), расщепленный с ВаmН I. Конечный клон DS-1034-3 был расщеплен TcoR I и фрагмент гена T7/hib47* был клонирован в pBR328 (Boehringer Mannheim Corporation) для получения плазмид DS-1048-2 и DS-1067-2. Электропорация плазмиды ДНК в штамм BL21/DE3 E.coli дает штаммы DS-1071-1-1 и DS-1071-3-1, которые экспрессируют Ser-197 --> аланин аналог Hin47.
Пример 6.
Данный пример иллюстрирует экспрессию аналога Ser-197 --> аланин Hin47 из Е. coli.
Ночные культуры штаммов DS-1018-3-1, DS-1071-1-1 или DS-1071-3-1 растили в течение ночи в среде NZCYM + 3% декстроза + антибиотики (ампициллин в концентрации 25 мкг/мл или тетрациклин в концентрации 10 мкг/мл) при 37oС при встряхивании. Разведенную 1: 40 ночную культуру привили в ту же самую среду и выращивали при 37oС при встряхивании до поглощения A578 приблизительно 0,3. Затем была добавлена 1/10 объема 10% лактозы для индукции экспрессии от Т7 промотера. Образцы клеток собрали через 4 часа после индукции путем центрифугирования культуральных образцов при 5000 об/мин в течение 10 мин при 4oС в центрифуге Sorvall RC-3B.
Пример 7.
Данный пример иллюстрирует экстракцию и очистку Hin47.
Hin47 экспрессировали как растворимый белок в Е. coli. Осадок клеток из 250 мл культуры, приготовленный, как описано в примере 6, суспендировали в 40 мл 50 мМ Трис/HCl, рН 8,0 и разрушили путем озвучивания (3•10 мин, 70% цикл нагрузки). Экстракт центрифугировали при 20 000•g и сохранили конечный супернатант, который содержал > 95% растворимого белка Hin47. Эту фракцию назвали "Hin47-экстракт".
Этот Hin47-экстракт далее очистили на колонке DEAE Sephacel. 40 мл Hin47-экстракта нанесли на колонку объемом 20 мл с DEAE Sephacel, уравновешенную 50 мМ Трис/HCl, рН 8,0. Hin47 связывается с колонкой в этих условиях. Колонку промыли 100 мл 50 мМ Трис/HCl, рН 8,0, содержащего 20 мМ NaCl. Затем Hin47 элюировали 50 мМ Трис/HCl, рН 8,0, содержащим 40 мМ NaCl. Количество Hin47 во фракциях определяли по измерению белка BSA. Чистоту Hin47 оценивали путем SDS-PAGE анализа. Фракции, содержащие Hin47, объединяли и хранили при -20oС.
Только мутант Н91А был также растворим, как и дикий тип белка Hin47, большинство других мутантов были получены как тельца включения.
Пример 8.
Данный пример иллюстрирует экстракцию и очистку аналога Hin47 Ser-197 --> аланин.
Аналог Hin47 Ser-197 --> аланин экспрессировался в тельцах включения у Е. coli. Осадок клеток, полученный из 250 мл культуры, приготовленный, как описано в примере 6, ресуспендировали в 40 мл 50 мМ Трис/HCl, рН 8,0, и разрушенный озвучиванием (3•10 мин, 70% цикл нагрузки). Экстракт центрифугировали при 20000•g и сохранили конечный осадок. Осадок реэкстрагировали 40 мл 50 мМ Трис/HCl, 0,5% Тритон Х-100, 10 мл ЭДТА, рН 8,0. Суспензию озвучивали 10 мин, 70% цикл нагрузки. Экстракт центрифугировали при 300•g в течение 5 минут. Полученный супернатант центрифугировали снова при 20000•g 30 минут и конечный осадок сохраняли. Осадок ресуспендировали в 50 мМ Трис/HCl, 0,5% Тритон Х-100, 10 мМ ЭДТА, рН 8,0. Затем суспензию перемешали в 50 мМ Трис/HCl, рН 8,0, содержащем 8 М мочевину. Конечная концентрация мочевины в смеси была доведена до 2 М 50 мМ Трис/HCl, рН 8,0. Аналог Hin47 Ser-197 --> аланин полностью растворился в этих условиях. Конечный объем раствора составил 20 мл. Эта фракция названа "Hin47 экстракт аналога". "Hin47 экстракт аналога" далее был очищен на колонке с DEAE Sephacel. 20 мл "Hin47 экстракт аналога" были нанесены на колонку объемом 10 мл с DEAE Sephacel, уравновешенную 50 мМ Трис/HCl, рН 8,0. Аналог Hin47 Ser-197 --> аланин связывается с колонкой в этих условиях. Колонка была промыта 50 мМ Трис/HCl, рН 8,0, и аналог элюировали 50 мМ Трис/HCl, рН 8,0, содержащим 30 мМ NaCl. Количество аналога Hin47 во фракциях определяли по измерению белка BSA. Чистоту аналога Hin47 оценивали путем SDS-PAGE анализа (фиг. 6). Фракции, содержащие аналог Hin47, объединяли и хранили при -20oС.
Пример 9.
Данный пример иллюстрирует протеазную активность Hin47 и Ser-197 --> аланин аналога Hin47.
Ферментативную активность Hin47 и Ser-197 --> аланин аналога Hin47 анализировали, используя бетта-казеин в качестве субстрата (фиг. 7). Реакционная смесь содержала 5 мкг бетта-казеина и Hin47 или аналог Hin47. Реакцию проводили при 37oС в течение двух часов и затем останавливали добавлением SDS-буфера и немедленным кипячением образца при 100oС в течение 5 минут. Аликвоты анализировали путем SDS-PAGE. Как показано на фиг. 7, гидролиз бетта-казеина под действием Hin47 был более выражен через два часа (панель А, прямая 1) по сравнению с фракциями, содержащими аналог Hin47 (панель А, прямая 2) или без добавления экзогенных белков (панель А, прямая 3). Наличие Hin47 и аналога Hin47 в смеси подтверждено методом иммуноблоттинга, используя моноклональное антитело к Hin47 (фиг. 7, панель С, прямые 1 и 2).
Протеазные активности Hin47 и аналога Hin47 Ser-197 --> аланин сравнивали, анализируя аутогидролиз Hin47 и аналога Hin47 при 4oС и при -20oС. Очищенные Hin47 или аналог Hin47 хранили либо при 4oС, либо при -20oС до 20 дней. На нулевой, 10 и 20 день отбирали аликвоты и анализировали стабильность Hin47 или аналога Hin47 методом иммуноблоттинга, используя моноклональные антитела к Hin47 (фиг. 8). Аналог был значительно более стабильным, чем Hin47, до 20 дней при хранении как при 4oС, так и при -20oС.
Для дальнейшего изучения протеазной активности аналога Hin47 Ser-197 --> аланин была исследовна способность Hin47 или аналога расщеплять 80-кДа рекомбинантный антиген Н. influenzae. Подобным образом было проведено изучение смешанного антигена для определения протеолитического действия Hin47 или аналога Hin47 на других антигенах. Для этого исследования был выбран рекомбинантный белок (ТВР1) массой 80 кДа Н. influenzae для того, чтобы отличить его от Hin47 или белка аналога (47 кДа). Были составлены следующие пять смесей: белок 80 кДа без добавок; белок 80 кДа + Hin47; белок 80 кДа + аналог; Hin47 без добавок; аналог без добавок. Количество каждого белка в смеси составляло 5 мкг. Смеси хранили при 4oС до четырех недель. Аликвоты отбирали на 0, 7, 14 и 28 дни для анализа методом SDS-PAGE (фиг. 9). Оба белка 80 кДа и Hin47 были заметно расщеплены через одну неделю (прямые 2 и 4). Напротив, белок 80 кДа в комбинации с аналогом Hin47 сохранился интактным через одну неделю, и только через четыре недели проявилась слабая деградация (прямая 3).
Остаточная протеазная активность других аналогов Hin47 была определена по расщеплению бетта-казеина, как описано Lipinska et аl (ссылка 13), результаты чего показаны в таблице 3. Было обнаружено, что только один мутант (D121E) сохранял сериновую протеазную активность.
Пример 10.
Данный пример иллюстрирует сравнительную иммуногенность Hin47 и аналога Hin47 у мышей.
На фиг. 10 представлены результаты изучения по определению сравнительной иммуногенности Hin47 и аналога Hin47 Ser-197 --> аланин. Так, например, группам из пяти мышей линии Balb/c ввели три раза (как отмечено стрелками) подкожно на 1, 29 и 43 дни в дозе по 1 мкг либо Hin47, либо аналога Hin47 в присутствии АlРО4 (1,5 мг на дозу). Образцы крови отбирали на 14, 28, 42 и 56 дни (как отмечено 1, 2, 3 и 4 заборами крови соответственно) для анализирования титра антител анти-Нin47 по ElAs. Определение анти-Нin47 антител в сыворотке мышей проводили, как описано Panezutti et al. (1993). Микротитровальные ячейки покрыли 1 мкг либо Hin47, либо аналогом Hin47 на 16 часов при комнатной температуре. Затем планшеты отформировали 0,1% (в/о) бычьим сывороточным альбумином на PBS. Сыворотку мышей серийно развели, добавили в ячейки, затем инкубировали один час при комнатной температуре. Аффинно очищенные F(ab')2 фрагменты антимышиного IgG (Fc специфического) антитела, конъюгированные с пероксидазой хрена, были добавлены как второе антитело. Реакцию проводили при использовании тетраметилбензидина (ТМВ/Н2O2) и измеряли поглощение при 450 нм (используя 540 нм как длину волны сравнения) на Flow Multiskan MCC microplate reader. Титр реактивности антисыворотки был определен как обратное разведение, последовательно показывающее двукратное увеличение поглощения над полученным с предварительно взятыми образцами крови. Как можно видеть из фиг. 10, и Hin47, и аналог Hin47 выявляли сравнимые титры у мышей независимо то ли Hin47, то ли мутант был использован в качестве антигена в ElAs.
Исследования иммуногенности были также проведены, используя Н91А аналог Hin47. Было обнаружено, что этот аналог дает иммунный ответ, эквивалентный ответу аналога S197A Hin47.
Для дальнейшего изучения иммунного ответа на Hin47 или Ser-197 --> аланин аналог Hin47 были определены подклассы анти-Нin47 IgG в сыворотке мышей. Микротитровальные ячейки были покрыты 1 мкг очищенного Hin47 или аналога. Конечный отбор образцов сыворотки мышей из сравнительного изучения иммуногенности (как описано выше) был собран и протестирован ElAs. Крысиные антимышиные IgG1, IgG2a и IgG2b, конъюгированные к пероксидазе хрена, и кроличьи антимышиные IgG3, коньюгированные с пероксидазой хрена, были использованы как реагенты в ElAs. Рабочие разведения каждого конъюгата определяли, используя очищенные антитела субклассов для избегания перекрестной реактивности. Титры реактивности были определены, как описано выше. Как показано ниже, в таблице 1, профиль подклассов IgG, вырабатывающийся у мышей как Hin47, так и аналогом Hin47, были идентичны, независимо от того, были ли использованы в качестве твердого антигена в EIAs Hin47 или аналог Hin47. Предоминантный ответ IgG в обоих группах сыворотки мышей представлял IgG1 изотип. Следовательно, аналог Hin47 проявляет, по существу, те же иммуногенные свойства, что и природный белок.
Пример 11.
Данный пример иллюстрирует иммунозащитные свойства Hin47 и Ser-197 --> аланин аналога Hin47.
Иммунозащитные свойства Hin47 и Ser-197 --> аланин аналога Hin47 были проанализированы по способности специфической сыворотки к Hin47 защищать детенышей крыс от Н. influenzae типа b штамма MinnA в бактериальной модели. Результаты этого исследования представлены ниже в таблице 2. Группам 6-дневных детенышей крыс Вистар подкожно ввели (п.к.) в спину рядом с шеей по 0,1 мл либо кроличьей антисыворотки к аналогу анти-Нin47, либо соответствующей предварительно взятой сыворотки. Двадцать четыре часа спустя животным внутрибрюшинно (в. б. ) ввели по 700 КОЕ свежевыращенного Hib штамма MinnA. Образцы крови собрали через 24 часа после введения и поместили в шоколадные агаровые плашки. Бактериальные колонии подсчитали через 24 часа. Как показано в таблице 2, трое из девяти животных в группе, которой вводили антисыворотку к аналогу анти-Нin47, не проявляли никакую бактериемию в крови. Только одна мышь в группе, которой вводили антисыворотку к аналогу анти-Нin47 (11%), имела высокий бактериальный выход из образцов крови в сравнении с мышами, которым вводили предварительно взятую сыворотку. Напротив, выход бактерий наблюдался у всех девяти мышей, которым вводили предварительно взятую сыворотку. Четыре из девяти животных (44%) в группе, которой вводили предварительно взятую сыворотку, проявляли высокий уровень (от 500 до 1000) выхода бактерий в образцах крови.
Модель бактериемии на детенышах крыс была использована для оценки даваемой защиты антисывороткой анти-Нin47 или мутантной анти-Нin47 антисывороткой против бактериемии, вызываемой инфекцией Н. influenzae типа b. 6 из 10 детенышей крыс были защищены антисывороткой, полученной против каждого из диких типов Hin47, Н91А Hin47 и S197A Hin47 аналогов.
Пример 12.
Данный пример иллюстрирует индукцию Hin47 в стрессовых условиях.
Штамм Еаgаn Н. influenzae выращивали при 37oС до А590=0,3 на инфузионном бульоне сердце мозг (BHI), содержащем гемин (2 мкг/мл) и NAD (2 мкг/мл). Образцы разделили на аликвоты и выращивали при 37oС, 42oС и 43,5oС или в присутствие 6% этанола, 0,2 М NaCl или 0,3 М NaCl. Штамм Е. coli JM109 выращивали при 37oС до А590 ≈ 0,3 в YT среде и разделили на аликвоты, как описано. Образцы собрали в 0 мин, 20 мин, 40 мин, 60 мин и 90 мин и анализировали по OD и SDS-PGE/Western blot. Для Western blot анализа использовали антисыворотку морских свинок, которая распознает как Н. influenzae, так и Е. coli htrA. Белок Е. coli htrA производился в больших количествах, когда организм выращивали при 43,5oС, тогда как белок Hin47 H. influenzae наблюдался в малых количествах. Оба белка Е. coli htrA и Н. influenzae Hin47 индуцировались при выращивании в среде, содержащей 6% этанол. Высокие концентрации солей были недостаточны для индукции других белков. Эти результаты показывают, что белок Hin47 является стресс-зависимым белком у Н. influenzae, индуцирующимся при сходных условиях, что и белок Е. coli htrA.
Пример 13.
Данный пример иллюстрирует очистку Н91А Hin47 белка.
Растворимый мутант Н91А очищали преимущественно, как описано для дикого типа Hin47 в примере 7, с добавлением гидроксилаппатитной (НАР) колонки. НАР колонка была уравновешена 10 мМ натрий фосфатным буфером (рН 8,0) и на нее нанесен материал, прогнанный через DEAE колонку. Н91А Hin47 связался с колонкой, а примесные белки удаляли промыванием колонки 175 мМ натрий-фосфатным буфером. Белок Н91А Hin47 элюировали 300 мM натрий-фосфатным буфером (рН 8,0) и хранили при -20oС.
Пример 14.
Данный пример иллюстрирует изучение защитного действия Hin47 и мутанта Hin47 на модели отита у шиншилл.
Шиншилл (500 г весом) иммунизировали в.м. три раза по 30 мкг/дозу Hin47 или мутантом Hin47 (H91A или S197A) с адъювантом с АlР04 и на 1, 28 и 42 дни. Животных отбирали на 56 день по образованию вздутия в организмах с 50-1000 КОЕ вирулентного NTHi штамма SB12. Животных мониторировали по тимпанометрическому и отоскопическому обследованию, а на 4 день после отбора аспирировали жидкость из среднего уха и помещали на шоколадный агар. Бактериальные колонии подсчитывали через 24 часа. Белки дикого типа Hin47 и H91A Hin47 давали защиту у 50% животных, однако S197A Hin47 был неэффективен на этой модели (таблица 3).
Заключение
В заключение настоящего описания настоящее изобретение относится к новым аналогам белка Hin47 Haemophilus influenzae, которые обладают пониженной протеазной активностью, составляющей менее чем 10% от активности природного белка Hin47, а также к выделенным и очищенным молекулам ДНК, кодирующих их. В рамках настоящего изобретения возможны модификации.
Список литературы
1. Zangwill et al, 1993. MMWR 42:1-15.
2. Schoendorf et al, 1994. Pediatrics 93:663-8.
3. Brenner et al, 1988. Nature 334:528-530.
4. O'Hagan, 1992. Clin. Pharmokinet. 22:1-10.
5. Ulmer et al, 1993. Curr. Opinion. Invest. Drugs 2:983-989.
6. Chang et al, 1978. Nature 275:617.
7. Goeddel et al, 1980. Nucl. Acid. Res. 8:4057.
8. Harkness et al, 1992. J. Bacteriol. 174:2425-2430.
9. Loeb et al, 1987. Infec. Immun. 55:2612-2618.
10. Holmes and Quigley, 1981. Analyt. Biochem. 114:193-197.
11. Young and Davis, 1985. Gene 38:31-38.
12. Panezutti et al, 1993. Infec. Immun. 61:1867-72.
13. Lipinska et al, 1985. Bacteriol. 171:1574-1584.
14. Barenkamp et al, 1986. Infect. Immun. 52:572-578.9

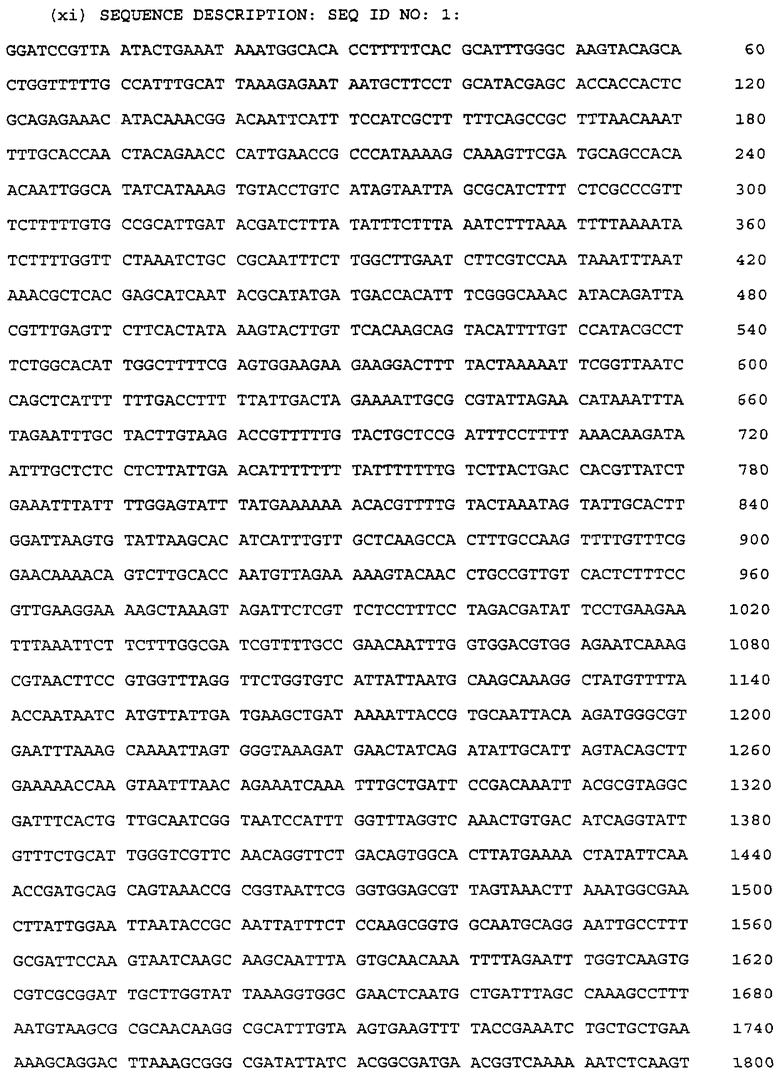



























Изобретение относится к иммунологии, а именно к иммуногенам и антигенам от видов Наеmophilus. Изолированный и очищенный аналог белка Hin47 Наеmophilus influenzae обладает пониженной протеазной активностью, которая составляет менее чем 10% от активности природного белка Hin47, и теми же иммуногенными свойствами, что и природный белок Hin47, за счет замены или включения у нативного белка Hin47, по крайней мере, одной аминокислоты, ответственной за протеазную активность. Аминокислота может быть выбрана из аминокислот 195-201, His-91 или Asp-121 нативного белка Hin47. Химерная молекула содержит аналог белка Hin47 Наеmophilus influenzae, связанный с полисахаридом. Рекомбинантная плазмида DS-1011-1-1 (рТ7/Hin47*) несет молекулу нуклеиновой кислоты, экспрессирующую аналог белка Hin47 с заменой Ser-197 на аланин. Идентифицируют, по крайней мере, один аминокислотный остаток белка Hin47, ответственный за протеазную активность. Осуществляют сайт-направленный мутагенез гена hin47 для удаления или замены нуклеотидной последовательности, кодирующей эту аминокислоту. Вводят мутанный ген в клетку с получением трансформированной клетки и культивируют последнюю с получением аналога Hin47. Иммуногенная композиция включает эффективное количество аналога Hin47. Проводят взаимодействие образца с аналогом Hin47 с получением комплексов, содержащих аналог Hin47 и специфично распознающие его антитела, если таковые присутствуют в образце. При иммунизации субъекта получают антитела, специфичные к белку Hin47. Проводят взаимодействие образца с антителами. Диагностический набор для определения наличия антител, специфично распознающих белок Hin47, в образце содержит аналог белка Hin47 и средства для взаимодействия аналога с образцом. Изобретение позволяет получать вакцинные препараты против заболеваний, вызываемых патогенными бактериями Наеmophilus influenzae. 8 с. и 10 з.п.ф-лы, 3 табл., 26 ил.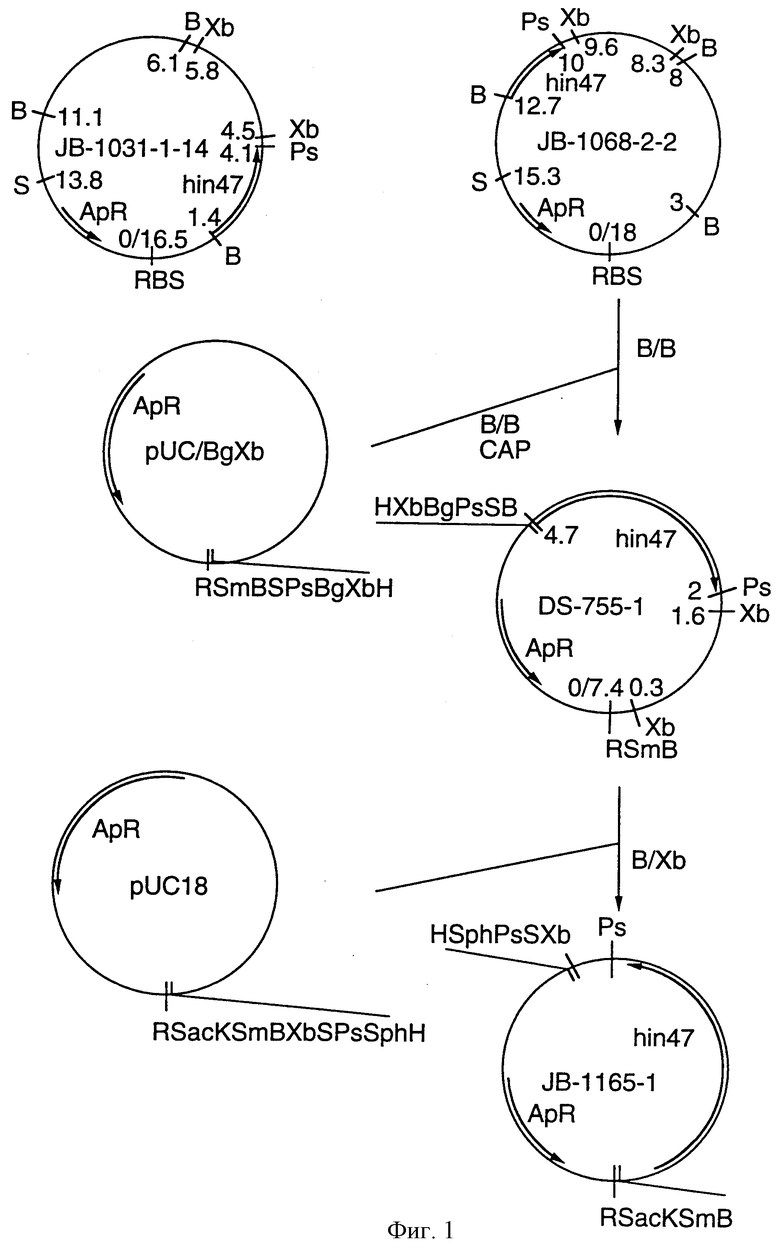
| WO 9210936 А1, 09.07.1992 | |||
| WO 9211367 А1, 09.07.1992 | |||
| WO 9412641 А1, 09.06.1994 | |||
| WO 9400149 А1, 06.01.1994 | |||
| Г.МАНИАТИС и др | |||
| Методы генетической инженерии | |||
| - М.: Мир, 1984, с.241-244. |

