ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Изобретение относится к поливалентным вакцинам, в частности, для применения в педиатрии.
ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Эта заявка является частичным продолжением патентной заявки с серийным номером United States Patent Application No. 08/501743, поданной 12 июля 1995 в патентное ведомство США, которая сама является частичным продолжением патентной заявки США с серийным No. 08/433646, поданной 4 мая 1995.
УРОВЕНЬ ТЕХНИКИ
Коклюш представляет собой тяжелую очень заразную инфекцию верхних дыхательных путей, вызываемую Bordetella pertussis. Согласно оценке Всемирной организации здравоохранения имеется 60 миллионов случаев коклюша в год и 0,5-1 миллион связанных с коклюшем смертей (ссылка 1).( Во всей этой публикации даются многочисленные ссылки в скобках для более полного описания состояния данной области, к которой относится данное изобретение. Полная библиографическая информация для каждого цитирования находится в конце заявки, сразу же после формулы изобретения. Описания этих ссылок включены тем самым в качестве ссылки в данное описание). В невакцинированных популяциях наблюдали такой высокий коэффициент заболеваемости, как 80%, у детей в возрасте менее 5 лет (ссылка 2). Хотя коклюш считается детским заболеванием, имеются все увеличивающиеся доказательства клинического и бессимптомного заболевания у подростков и взрослых (ссылки 3, 4 и 5).
Введение цельноклеточных вакцин, состоящих их химически инактивированных и инактивированных нагреванием организмов В. pertussis, в 1940-х годах вызывало драматичное снижение частоты заболеваний коклюшем, вызываемых В. pertussis. Коэффициенты эффективности для цельноклеточных вакцин были оценены как 95% в зависимости от конкретного случая распознавания заболевания (ссылка 6). Хотя инфекция В. pertussis вызывает пожизненный иммунитет, появляется все больше доказательств уменьшения иммунитета после иммунизации цельноклеточными вакцинами (ссылка 3). Несколько сообщений, указывающих на взаимосвязь между цельноклеточной вакцинацией против коклюша, реактогенностью и серьезными побочными эффектами, привели к отказу от приема вакцин и, как следствие, возобновлению эпидемий. Относительно недавно были разработаны вакцины, состоящие из коклюшных компонентов.
Антигены для описанных коклюшных вакцин
Были разработаны многочисленные бесклеточные коклюшные вакцины, включающие в себя антигены Bordetella pertussis, коклюшный токсин (РТ), нитевидный гемагглютинин (FHA), белок наружной мембраны 69 кДа (пертактин) и фимбриальные (жгутиковые) агглютиногены (см. таблицу 1. Таблицы помещены в конце описания).
Коклюшный токсин
Коклюшный токсин является экзотоксином, который является членом семейства А/В бактериальных токсинов с АДФ-рибозилтрансферазной активностью (ссылка 8). А-часть этих токсинов проявляет АДФ-рибозилтрансферазную активность, а В-часть медиирует связывание токсина с рецепторами клеток-хозяина и транслокацию А к месту действия. РТ облегчает также прикрепление В. pertussis к реснитчатым эпителиальным клеткам (ссылка 9) и также играет роль в инвазии макрофагов В. pertussis (ссылка 10).
Все бесклеточные коклюшные вакцины включали в себя РТ, который считали основным фактором вирулентности и иммунным антигеном (ссылки 11, 12). Природная инфекция В. pertussis, генерирует как гуморальный, так и клеточно-опосредованный ответы на РТ (ссылки 13-17). Младенцы имеют антитела против РТ трансплацентарного происхождения (ссылки 16, 18), и молозиво человека, содержащее антитела против РТ, было эффективно в пассивном иммунитете мышей против аэрозольной инфекции (ссылка 19). Клеточно-опосредованный иммунный ответ (СМI) на субъединицы РТ, был продемонстрирован после иммунизации бесклеточной вакциной (ссылка 20), а ответ CMI на РТ был вызван после вакцинации цельноклеточными вакцинами (ссылка 13). Химически инактивированный РТ в цельноклеточных или компонентных вакцинах является иммунным в животных моделях и в людях (ссылка 21). Кроме того, моноклональные антитела, специфические для субъединицы 31, защищают против инфекции В. pertussis (ссылки 22 и 23).
Основные патофизиологические эффекты РТ обусловлены его АДФ-рибозилтрансферазной активностью. РТ катализирует перенос АДФ-рибозы от НАД к G1-гуаниннуклеотидсвязывающему белку, разрушая тем самым клеточную аденилатциклазную регуляторную систему (ссылка 24). РТ также предотвращает миграцию макрофагов и лимфоцитов к местам воспаления и препятствует медиируемому нейтрофилами фагоцитозу и уничтожению бактерий (ссылка 25). Ряд тестов in vitro и in vivo использовали для оценки ферментативной активности 31 и/или РТ, в том числе ДДФ-рибозилирование бычьего трансдуцина (ссылка 26), тест образования кластеров клеток яичника Китайского хомячка (СНО) (ссылка 27), сенсибилизацию гистамином (ссылка 28), лейкоцитоз и НДД-гликогидролазу. При экспонировании с РТ клетки СНО развивают характерную кластерную морфологию. Этот феномен зависит от связывания РТ, последующей его транслокации и АДФ-рибозилтрансферазной активности S1 и поэтому тест образования кластеров СНО широко используется для тестирования целостности и токсичности голотоксинов РТ.
Нитевидный гемагглютинин
Нитевидный гемагглютинин представляет собой большой (220 кДа) нетоксичный полипептид, который медиирует присоединение В. pertussis к реснитчатым клеткам верхних дыхательных путей во время бактериального заселения (ссылки 9, 29). Природная инфекция индуцирует антитела против FHA и клеточно-опосредованный иммунитет (ссылки 13, 15, 17, 30 и 31). Антитела против FHA находят в молозиве человека, и они также передаются трансплацентарно (ссылки 17, 18 и 19). Вакцинация цельноклеточными или бесклеточными коклюшными вакцинами генерирует антитела против FHA, а бесклеточные вакцины, содержащие FHA, также индуцируют CMI-ответ на FHA (ссылки 20, 32). FHA представляет собой иммунный антиген в мышиной модели респираторного заражения после активной или пассивной иммунизации (ссылки 33, 34). Однако один FHA не создает иммунитета в мышином тесте эффективности интрацеребрального заражения (ссылка 28).
Белок наружной мембраны 69 кДа (пертактин)
Белок 69 кДа является белком наружной мембраны, который первоначально был идентифицирован из В. bronchiseptica (ссылка 35). Этот белок известен также как пертактин и Р.69. Было показано, что он является иммунным антигеном против В. bronchiseptica и он был впоследствии идентифицирован как в В. реrtussis, так и в В. parapertussis. Белок 69 кДа связывается непосредственно с эукариотическими клетками (ссылка 36), и природная инфекция B. pertussis индуцирует гуморальный ответ в виде антител против Р.69 (ссылка 14) и Р.69 индуцирует также клеточно-опосредованный иммунный ответ (ссылка 17, 37, 38). Вакцинация цельноклеточными или бесклеточными вакцинами индуцирует антитела против Р.69 (ссылки 32, 39), а бесклеточные вакцины индуцируют CMI на Р. 69 (ссылка 39). Пертактин защищает мышей против аэрозольного заражения В. pertussis (ссылка 40) и в сочетании с FHA защищает в тесте интрацеребрального заражения против В. pertussis (ссылка 41). Пассивный перенос поликлональных или моноклональных антител против Р.69 также защищает мышей против аэрозольного заражения (ссылка 42).
Агглютиногены
Серотипы В. pertussis определяются их агглютинирующими фимбриями (жгутиками). Всемирная организация здравоохранения (WHO) рекомендует, чтобы цельноклеточные вакцины включали в себя типы 1, 2 и 3 агглютиногенов (Aggs), так как они не являются перекрестно-иммунными (ссылка 43). Agg 1 является нефимбриальным и обнаружен на всех штаммах В. pertussis, тогда как Agg серотипов 2 и 3 являются фимбриальными. Природная инфекция или иммунизация цельноклеточными или бесклеточными вакцинами индуцирует антитела против Agg (ссылки 15, 32). Специфический клеточно-опосредованный иммунный ответ может быть генерирован в мышах Agg 2 или Agg 3 после аэрозольной инфекции (ссылка 17). Аgg 2 и 3 являются иммунными в мышах против респираторного заражения, и молозиво человека, содержащее антиагглютиногены, будут также защищать в этом тесте (ссылки 19, 44, 45).
Бесклеточные коклюшные вакцины
Первой разработанной бесклеточной коклюшной вакциной была двухкомпонентная РТ+FHA вакцина (JNIH 6) Sato et al. (ссылка 46). Эта вакцина была приготовлена совместной очисткой антигенов РТ и FHA из культурального супернатанта штамма В. pertussis Tohama с последующим превращением в токсоид (обезвреженный токсин) формалином. Бесклеточные вакцины от различных изготовителей и различные композиции использовали с успехом для иммунизации японских детей против коклюша с 1981 года, что привело к серьезному снижению частоты заболевания (ссылка 47). Вакцина JNIH 6 и монокомпонентная РТ-токсоидная вакцина (JNIH 7) были испытаны в большом клиническом испытании в Швеции в 1986 году. Первоначальные результаты показали более низкую эффективность, чем эффективность, сообщенная для цельноклеточной вакцины, но последующие исследования показали, что они были более эффективными против заболевания легкой тяжести, диагностируемого серологическими способами (ссылки 48, 49, 50, 51). Однако было обнаружено возвращение к токсичности инактивированного формалином РТ в этих вакцинах. Было также обнаружено, что эти вакцины защищают скорее от заболевания (защищают уже заболевшего), чем от инфекции.
В настоящее время оценивается ряд новых бесклеточных и компонентных коклюшных вакцин, которые включают в себя комбинации РТ, FHA, Р. 69 и/или агглютиногены, и которые представлены в таблице 1. Несколько способов химической детоксикации были использованы для РТ, в том числе инактивация формалином (ссылка 46), глутаровым альдегидом (ссылка 52), перекисью водорода (ссылка 53) и тетранитрометаном (ссылка 54).
Столбняк
Столбняк представляет собой острую инфекцию, вызываемую Clostridium tetani. Это заболевание характеризуется тяжелыми болезненными мышечными сокращениями, сопровождающимися гиперчувствительностью (аллергией), гиперрефлексией и повышенной автономной стимуляцией пораженных частей тела. Легкие стимулы могут вызвать тяжелые непроизвольные мышечные спазмы. Может присутствовать лихорадочное состояние, вызываемое мышечным спазмом. Столбняк может быть генерализованным, включающим в себя лицо, шею, живот и туловище, или локализованным в конкретной части тела (месте поражения). Вовлечение в патологический процесс жевательной мышцы лица приводит к тризму или спазму жевательных мышц, приводящему к возникновению классического выражения лица, известного как "risus sardonicus" (сардоническая улыбка) (ссылка 78).
Clostridium tetani существует в виде непатогенного организма в кишечнике людей и животных. Этот организм обнаружен также в почве, загрязненной фекалиями, и может выживать в почве в течение нескольких лет в виде инфекционных спор (ссылка 79).
Столбняк возникает от анаэробного роста Clostridium tetani и образования нейротоксина в загрязненных тканях. Инфекция вызывается при введении веществ, загрязненных организмами или спорами, в ткань. Наиболее обычным сценарием является инфекция через проникающее повреждение. Однако во многих случаях неизвестна история повреждения. Присутствие некротической или ишемической ткани облегчает рост этой бациллы (ссылка 78).
Предотвращением инфекции является вакцинация и хороший уход за раной, включающий в себя тщательную очистку и хирургическую обработку мертвых тканей. Индивидуумы с загрязненными ранами и те, у кого не были завершены вакцинации, должны получить столбнячную вакцину и столбнячный иммуноглобулин.
Лечение этого синдрома является в основном поддерживающим и может включать в себя поддержку дыхания, введение столбнячного антитоксина и тщательную очистку инфицированных ран. Несмотря на современный медицинский уход, коэффициенты смертности все еще бывают такими высокими, как 30-90% (ссылка 79). Это, в частности, верно для людей пожилого возраста. Природная инфекция не всегда создает иммунитет против последующих инфекций.
Предупреждение инфекции вакцинацией является наиболее эффективным способом борьбы с этим заболеванием. Со времени введения всеобщей вакцинации столбняк стал чрезвычайно редким заболеванием в развитых странах. Случаи столбняка встречаются почти исключительно у индивидуумов, которые не завершили ряда вакцинаций или которые не получили соответствующих бустердоз (повторных доз). Индивидуумы должны получать бустер-дозу один раз каждые 10 лет.
Дифтерия
Дифтерия является острой инфекцией, вызываемой бактерией Corynebacterium diphtheriae. Основное место инфекции находится в верхних дыхательных путях (нос, глотка, гортань и трахея) (ссылка 80). Характерным повреждением, результатом бактериального цитотоксина, являются бляшки сероватой псевдомембраны, окруженные воспалением. Это сопровождается цервикальной лимфоаденопатией, набуханием и отеком горла. В тяжелых случаях (набухание) может прогрессировать до обструкции (закупорки) (дифтерийный круп). Другие осложнения включают в себя миокардит, эффекты центральной нервной системы (краниальную невропатию, двигательную и сенсорную невропатии, такие как восходящий паралич) и тромбоцитопению. Другие слизистые оболочки могут быть менее часто поражены. Клиническое проявление может варьировать от бессимптомной инфекции до молниеносной, внезапно развивающейся мультисистемы и смерти (ссылка 79). Кожные и раневые инфекции с дифтерией являются обычными в тропиках. Единственным резервуаром инфекции для С. diphtheriae является человек (ссылка 79).
Предполагаемый диагноз может быть сделан на основании клинического наблюдения характерных повреждений, но он должен быть подтвержден бактериальным исследованием повреждений. Если имеется сильное клиническое подозрение на дифтерию, лечение должно быть начато немедленно антибиотиками (пенициллин или эритромицин) и дифтерийным антитоксином (нейтрализующим токсин антителом), даже если этот диагноз еще не подтвержден. Смертность увеличивается, чем дольше выжидают после появления клинических симптомов (ссылка 80). Коэффициент смертности находится в диапазоне от 5 до 10%, несмотря на современный медицинский уход (ссылка 79) и смертность встречается в основном в случае очень молодых пациентов и пожилых пациентов. Природная инфекция не всегда вызывает иммунитет от последующей инфекции (ссылка 80).
Передача инфекции происходит путем прямого контакта с секрециями или выделениями из инфицированных индивидуумов. Индивидуумы являются заразными до тех пор, пока в секрециях наблюдаются бактерии. Это может продолжаться до 4 недель после инфекции. Передача инфекции имеет также место с инфицированными предметами (ссылка 79). Рекомендуется строгая изоляция пациентов.
Изредка индивидуумы могут становиться носителями и распространяют организмы вплоть до 6 месяцев после инфекции. Неиммунизированные носители должны быть немедленно вакцинированы по полной схеме вакцинации. Лечение антибиотиками устраняет ношение возбудителя и инфекционность пациентов в течение 4 дней (ссылка 80).
Полиомиелит
Как инактивированные (IPV), так и живые аттенуированные (OPV) полиовирусные вакцины были эффективными в борьбе с полиомиелитом во всем мире. Комбинированная DPT-IPV вакцина в настоящее время лицензирована в Европе и в Канаде, и было показано, что она была безопасной и эффективной для миллионов детей по всему миру.
Haemophilus influenzae типа b
До того как стали доступны эффективные вакцины, Haemophilus influenzae типа b (Hib) был основным возбудителем инвазивных переносимых кровью инфекций менингита в первые 2 года жизни (ссылка 81). Приблизительно 10% жертв менингита, вызываемого Haemophilus influenzae, умирают несмотря на медицинский уход. Перманентные осложнения являются обычными у выживших. Иммунизация против Haemophilus influenzae началась в Канаде в 1987 году полисахаридной вакциной (полирибозорибитолфосфат [PRP] из Haemophilus influenzae типа b). Улучшенная иммуногенность была достигнута у детей в возрасте 18 месяцев и старше с введением в 1988 году вакцины, состоящей из PRP, конъюгированным с дифтерийным токсоидом (PRP-D). С 1992 года была возможна иммунизация младенцев лицензированными PRP-конъюгатными вакцинами, иммуногенными для младенцев в возрасте менее 1 года (PRP, конъюгированный со столбнячным токсоидом, или PRP-Т). Использование этих Haemophilus influenzae-конъюгатных вакцин было ассоциировано с серьезным уменьшением частоты заболевания инвазивной инфекцией Haemophilus influenzae в Канаде и других местах (ссылка 82). Два Канадских клинических исследования с участием почти 900 детей в Британской Колумбии и Альберте показали, что лиофилизированный PRP-T может быть реконституирован (воссоздан из порошка) при помощи DPT (COMBIPACK) (ссылка 83) или при помощи DPT-Polio-адсорбированной вакцины (PENTAТМ) (ссылка 84), кроме обычного солевого разбавителя. Клинические исследования с участием более 100000 детей по всему миру продемонстрировали эффективность лиофилизированного PRP-T (ActHibТМ). Свыше 90% достигаемых уровней антител против PRP считаются иммунными (больше или равно 0,15 мкг/мл) после 3 доз PRP-T (ActHibТМ), начинающихся при 2 месяцах, или после однократной дозы PRP-T, вводимой после 12-месячного возраста. Соотношение, достигающее уровней, указывающих на долгосрочный иммунитет (более 1,0 мкг/мл), варьирует от 70 до 100% в зависимости от исследования. Миллионы доз PRP-T были проданы в Канаде с 1992 года. Случаи инвазивной инфекции Haemophilus influenzae после вакцинации PRP-T являются редкими и могут быть связаны с такими заболеваниями, как иммунодефицит (ссылка 85).
Комбинированные вакцины
Хотя имеется много фактических и потенциальных выгод вакцин, которые объединяют антигены для придания иммунитета против множественных патогенов, эти комбинации могут оказывать неблагоприятное действие на иммуногенность индивидуальных компонентов. Комбинации дифтерийного и столбнячного токсоидов с цельноклеточной противококлюшной вакциной (DTP) были доступны на протяжении 50 лет, и образование антител на эту комбинацию превосходило образование антител на индивидуальные компоненты, возможно, в результате адъювантного эффекта цельноклеточной противококлюшной вакцины. Комбинации DTP, которые также включают в себя инактивированную полиовирусную вакцину, лицензированы для многих случаев применения, хотя образование антител на антигены коклюша может быть уменьшено этой комбинацией (ссылки 69-71). Действие объединенных вакцин DTP с конъюгатными вакцинами Hib было вариабельным. Исследования с французским DTP и PRP-T показало такую же безопасность, но уменьшенное образование антител против PRP (ссылки 72-73), тогда как исследования с вакциной Канадского DTP и PRP-T не обнаружили влияния на ответ на PRP, но показали снижение коклюшных агглютиногенов и увеличенную болезненность места инъекции в группе комбинированной иммунизации (ссылки 74, 75).
В настоящее время становятся доступными данные о влиянии комбинирования APDT-вакцин с Hib-конъюгатной вакциной. У двух месячных младенцев, получивших три дозы бесклеточной коклюшно-дифтерийно-столбнячной вакцины (APDT), комбинированной с Hib-конъюгатной вакциной (PRP-T), образование антител к PRP было значимо более низким, чем в группе, получавшей отдельные инъекции в тот же самый день (ссылка 76). Сходные результаты сообщались и другой бесклеточной коклюшно-дифтерийно-столбнячной вакцине, комбинированной с PRP-T, предоставляемой в виде первых трех доз (ссылка 77).
В противоположность другим сообщенным исследованиям, дети, иммунизированные комбинированной вакциной, имели превосходящий титр антител к FHA, антигену дифтерии, и нескольким антигенам коклюша по сравнению с детьми, получавшими PRP в виде одновалентной вакцины. Существует несколько причин для эквивалентной или лучшей иммуногенности для этих вакцин, предоставляемых в виде комбинированной инъекции, иных, чем уменьшенная иммуногенность, сообщенная с другими продуктами. Все бесклеточные и компонентные коклюшные вакцины неодинаковы по их антигенному содержанию, способу токсоидирования, адъювантам или консервантам. Однако уменьшенная иммуногенность сообщалась бесклеточными коклюшными вакцинами, содержащими РТ, FHA и 69К и (ссылка 77) и содержащими РТ, FHA, 69K и фимбриальные агглютиногены (ссылка 76).
Было обнаружено, что пятикомпонентная вакцина APDT, исследованная в данном исследовании, имеет защитную эффективность 85% (пример 5), (95% CI 81/89) в фазе III клинического испытания, недавно завершенного в Швеции при содействии Национального Института Здоровья (ссылка 78).
Современные коммерчески доступные комбинированные вакцины могут не содержать подходящих форм подходящих антигенов в подходящих иммуногенных формах для достижения желательного уровня эффективности в восприимчивой к коклюшу популяции человека.
Было бы желательно обеспечение эффективных комбинированных вакцин, включающих в себя бесклеточные коклюшные компоненты, содержащие выбранные относительные количества выбранных антигенов.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к комбинированным или поливалентным вакцинам, содержащим бесклеточные компоненты коклюшной вакцины, и способам их применения.
В соответствии с одним аспектом данного изобретения обеспечена поливалентная иммуногенная композиция для создания иммунитета у хозяина против заболевания, вызываемого инфекцией Bordetella pertussis, Clostridium tetani, Corynebacterium diphtheriae, полиовирусом и/или Haemophilus influenzae, содержащая:
(a) коклюшный токсоид, нитевидный гемагглютинин, пертактин и агглютиногены в очищенном виде,
(b) столбнячный токсоид,
(c) дифтерийный токсоид,
(d) инактивированный полиовирус и необязательно
(e) конъюгат молекулы-носителя, выбранной из столбнячного токсоида и дифтерийного токсоида, и капсульного полисахарида Haemophilus influenzae типа b.
Эта иммуногенная композиция может быть приготовлена в виде вакцины для введения in vivo хозяину, причем индивидуальные компоненты этой композиции приготовлены таким образом, что иммуногенность индивидуальных компонентов не ухудшается другими индивидуальными компонентами композиции.
Эта иммуногенная композиция может дополнительно содержать адъювант, в частности гидроксид алюминия или фосфат алюминия.
Такая иммуногенная композиция может содержать приблизительно от 5 до приблизительно 30 мкг азота коклюшного токсоида, от примерно 5 до примерно 30 мкг азота нитевидного гемагглютинина, от примерно 3 до примерно 15 мкг азота пертактина и от примерно 1 до примерно 10 мкг азота агглютиногенов.
В одном конкретном варианте, иммуногенная композиция может содержать коклюшный токсоид, нитевидный гемагглютинин, белок 69 кДа и фимбриальные агглютиногены Bordetella pertussis в весовом соотношении примерно 20:20:5:3, обеспечиваемом примерно 20 мкг коклюшного токсоида, примерно 20 мкг нитевидного гемагглютинина, примерно 5 мкг фимбриальных агглютиногенов и примерно 3 мкг белка 69 кДа, в дозе для одного приема для человека. В другом специфическом варианте, вакцина может содержать коклюшный токсоид, нитевидный гемагглютинин, белок 69 кДа и фимбриальный агглютиноген Bordetella pertussis в весовом соотношении 10:5:5:3, обеспечиваемом примерно 10 мкг коклюшного токсоида, примерно 5 мкг нитевидного гемагглютинина, примерно 5 мкг фимбриального агглютиногена и примерно 3 мкг белка 69 кДа, в дозе для одного приема для человека. В одном варианте иммуногенной композиции, обеспеченной здесь, вакцина содержит примерно 15 Lf дифтерийного токсоида и примерно 5 Lf столбнячного токсоида.
Инактивированный полиовирус, применяемый в иммуногенной композиции данного изобретения, обычно включает в себя смесь инактивированных типов полиовируса 1, 2 и 3. Такая смесь инактивированных типов полиовируса 1, 2 и 3 может применяться в композициях:
от примерно 20 до примерно 50 единиц антигена D полиовируса типа 1;
от примерно 5 до примерно 10 единиц антигена D полиовируса типа 2;
от примерно 20 до примерно 50 единиц антигена D полиовируса типа 3
в однократной дозе для человека. В одной композиции такие смеси инактивированных типов полиовируса могут содержать:
примерно 40 единиц антигена D полиовируса типа 1;
примерно 8 единиц антигена D полиовируса типа 2;
примерно 32 единицы антигена D полиовируса типа 3
в разовой дозе для человека.
Компонент конъюгированной молекулы иммуногенной композиции может содержать конъюгат столбнячного токсоида или дифтерийного токсоида и полирибозорибитолфосфата (PRP) Haemophilus Influenzae типа b. Такая молекула-конъюгат может быть обеспечена в лиофилизированной форме, которая воссоздается (реконституируется) для введения путем комбинирования с другими компонентами. Эта иммуногенная композиция может содержать конъюгат в количестве от примерно 5 до примерно 15 мкг PRP, конъюгированного с от примерно 15 до примерно 35 мкг столбнячного токсоида, в разовой дозе для человека. В одной композиции этот конъюгат применяют в количестве примерно 10 мкг PRP, конъюгированного с примерно 20 мкг столбнячного токсоида.
В таких конкретных вариантах иммуногенные композиции обеспечивают профиль иммунного ответа на каждый из антигенов коклюша, содержащихся в них, и этот профиль ответа, обеспечиваемый бесклеточными компонентами, по существу, эквивалентен профилю, создаваемому цельноклеточной коклюшной вакциной.
В предпочтительном варианте данного изобретения обеспечена поливалентная вакцинная композиция, содержащая на дозу 0,5 мл 20 мкг коклюшного токсоида; 20 мкг нитевидного гемагглютинина; 5 мкг фимбриальных агглютиногенов 2 и 3; 3 мкг мембранного белка пертактина; 15 Lf дифтерийного токсоида; 5 Lf столбнячного токсоида; 40 единиц антигена D полиовируса 1; 8 единиц антигена D полиовируса типа 2; 1,5 мкг фосфата алюминия.
Такие композиции могут дополнительно содержать на дозу 0,5 мл 10 мкг очищенного капсульного полисахарида полирибозорибитолфосфата (PRP) Haemophilus influenzae типа b, ковалентно связанного с 20 мкг столбнячного токсоида. Кроме того, такие композиции могут содержать на дозу в 0,5 мл 0,6% 2-феноксиэтанол.
В дополнительном аспекте данного изобретения обеспечен способ иммунизации хозяина против многочисленных заболеваний, предусматривающий введение хозяину, которым может быть человек, иммуноэффективного количества иммуногенной композиции или вакцины, обеспеченной здесь.
Преимущества данного изобретения включают в себя поливалентную вакцину, которая может вызывать безопасным и эффективным образом иммунитет против ряда обычных педиатрических заболеваний. Способность обеспечивать одну вакцинацию против множества заболеваний без мешающих взаимодействий между иммуногенными ответами на различные иммуногены является важным преимуществом данного изобретения.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
Данное изобретение будет более хорошо понято из последующего подробного описания и примеров со ссылкой на сопутствующий чертеж, на котором дано схематическое изображение процедуры выделения препарата агглютиногена из штамма Bordetella.
СВЕДЕНИЯ,ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Приготовление агглютиногена
На чертеже показана схема способа получения препарата агглютиногена из штамма Bordetella. Как видно из чертежа, клеточную пасту Bordetella, содержащую агглютиногены, такую как клеточная паста В. pertussis, экстрагируют, например, содержащим мочевину буфером, таким как 10 мМ фосфат калия, 150 мМ NaCI и 4 М мочевина, для селективной экстракции агглютиногенов из клеточной пасты с получением первого супернатанта (sp1), содержащего агглютиногены, и первого оставшегося осадка (ppt1). Первый супернатант (sp1) отделяют от первого оставшегося осадка (ppt1), например, центрифугированием. Оставшийся осадок (ppt1) отбрасывают. Затем осветленный супернатант (sp1) может быть сконцентрирован и диафильтрован против, например, смеси 10 мМ фосфат калия/150 мМ NaCl/0,1% Тритон Х-100, с применением, например, мембранного фильтра NMWL 100-300 кДа.
Затем первый супернатант инкубируют при температуре и в течение времени, достаточных для получения осветленного супернатанта (sp2), содержащего агглютиногены, и второго отбрасываемого осадка (ppt2), содержащего примеси, не являющиеся агглютиногенами. Подходящие температуры находятся в пределах от приблизительно 50 до приблизительно 100oС, в том числе от приблизительно 75 до приблизительно 85oС, а подходящие периоды инкубации включают периоды времени от приблизительно 1 до приблизительно 60 минут. Затем осветленный супернатант концентрируют, например, добавлением полиэтиленгликоля с молекулярной массой приблизительно 8000 (ПЭГ 8000) до конечной концентрации приблизительно 4,5±0,2% и перемешивают осторожно в течение минимально приблизительно 30 минут с получением третьего осадка (ppt3), который может быть собран центрифугированием. Оставшийся супернатант sp3 отбрасывают.
Этот третий осадок (ppt3) экстрагируют, например, буфером, содержащим 10 мМ фосфат калия/150 мМ NaCl, с получением неочищенного фимбриального агглютиногенсодержащего раствора. К неочищенному фимбриальному раствору может быть добавлен 1 М фосфат калия, чтобы сделать его приблизительно 100 мМ относительно фосфата калия. Альтернативно осветленный супернатант обработанных нагреванием агглютиногенов может быть очищен без осаждения гель-фильтрационной хроматографией с применением геля, такого как Sepharose CL6B. Фимбриальные агглютиногены в неочищенном растворе очищают затем колоночной хроматографией, например пропусканием через колонку диоксида кремния PEI, с получением препарата фимбриального агглютиногена в прошедшей через колонку фракции.
Эта прошедшая через колонку фракция, содержащая фимбриальный агглютиноген, может быть дополнительно сконцентрирована и диафильтрована против, например, буфера, содержащего 10 мМ фосфат калия/150 мМ NaCl, с применением мембраны NMWL 100-300 кДа. Препарат агглютиногена может быть простерилизован фильтрованием через мембранный фильтр менее или равный 0,22 мкМ с получением конечного очищенного препарата фимбриального агглютиногена 2 и 3, по существу, не содержащего агглютиногена 1. Весовое отношение Аgg 2 к Аgg 3 может быть от примерно 1,5:1 до примерно 2:1. Вакцины могут содержать другие очищенные иммуногены Bordetella, в том числе нитевидный гемагглютинин, белок наружной мембраны 69 кДа и коклюшный токсин или его токсоид, в том числе генетически детоксифицированные аналоги РТ, описанные, например, в ссылке 68.
Другие иммуногены Bordetella, коклюшный токсин (в том числе его генетически обезвреженные аналоги, описанные, например, в Klein et al. Патент США No. 5085862, принадлежащем заявителю данной заявки и включенном здесь в качестве ссылки), FHA и белок 69 кДа могут быть получены в очищенном виде различными способами, такими как описанные ниже.
Очистка РТ
РТ может быть выделен из культурального супернатанта штамма В. pertussis при помощи общепринятых способов. Например, может быть использован способ Sekura et al. (ссылка 55). РТ выделяют сначала абсорбцией культурального супернатанта на колонке, содержащей гелевый матрикс с красителем-лигандом, Affi-Gel Blue (Bio-Rad Laboratories, Richmond, CA). РТ элюируют из этой колонки солевым раствором высокой концентрации, таким как 0,75 М хлорид магния, и после удаления соли пропускают через колонку аффинного матрикса fetuin-Sepharose, состоящего из фетуина, связанного с CNBr-активированной Сефарозой. РТ элюируют из фетуиновой колонки при помощи 4 М соли магния.
Альтернативно может быть использован способ Irons et al. (ссылка 56). Культуральный супернатант абсорбируют на колонке CNBr-активированной Сефарозы 4В, с которой сначала связывают гаптоглобин. РТ связывается с абсорбентом при рН 6,5 и элюируется из колонки буфером 0,1 М Трис/0,5 М NaCI ступенчатым изменением до рН 10.
Альтернативно может быть использован способ, описанный в Патенте США No. 4705686, выданном Scott et al. 10 ноября 1987 и включенном здесь в качестве ссылки. В этом способе культуральные супернатанты или клеточные экстракты В. pertussis пропускают через колонку анионообменной смолы достаточной емкости для адсорбции эндотоксина, но позволяющей антигенам Bordetella протекать через нее или отделяться от эндотоксина другим способом.
Альтернативно РТ может быть очищен при помощи хроматографии на перлите, как описано в Патенте ЕР No. 336736, принадлежащем заявителю данной заявки и включенном здесь в качестве ссылки.
Детоксикация РТ
РТ обезвреживают для удаления нежелательных активностей, которые могли бы вызывать побочные реакции конечной вакцины. Могут быть использованы любые из разнообразных общепринятых способов химической детоксикации, такие как обработка фомальдегидом, перекисью водорода, тетранитрометаном или глутаровым альдегидом.
Например, РТ может быть обезврежен глутаровым альдегидом согласно модификации процедуры, описанной в Munoz et al. (ссылка 57). В этом способе детоксикации очищенный РТ инкубируют в растворе, содержащем 0,01 М забуференный фосфатом солевой раствор (ЗФР). Раствор готовят 0,05% в отношении глутарового альдегида и эту смесь инкубируют при комнатной температуре в течение 2 часов и затем добавляют L-лизин до концентрации 0,02 М. Эту смесь инкубируют дополнительно в течение 2 часов при комнатной температуре и затем диализуют в течение 2 дней против 0,01 М ЗФР (PBS). В конкретном варианте можно использовать способ детоксикации ЕР Патента No. 336736. Вкратце, РТ может быть детоксифицирован глутаровым альдегидом следующим образом.
Очищенный РТ в 75 мМ фосфате калия при рН 8,0, содержащем 0,22 М хлорида натрия, разбавляют равным объемом глицерина до концентрации белка приблизительно 50-400 мкг/мл. Этот раствор нагревают до 37oС и детоксифицируют добавлением глутарового альдегида до конечной концентрации 0,5% (в/об). Смесь выдерживают при 37oС в течение 4 часов и затем добавляют аспарагиновую кислоту (1,5 М) до конечной концентрации 0,25 М. Эту смесь инкубируют при комнатной температуре в течение 1 часа и затем диафильтруют с 10 объемами 10 мМ фосфата калия при рН 8,0, содержащего 0,15 М хлорид натрия и 5% глицерин, для снижения глицерина и удаления глутарового альдегида. Токсоид РТ стерильно фильтруют через мембрану 0,2 мкМ.
Если используют рекомбинантные способы для получения мутантной молекулы РТ, не обнаруживающей токсичности или имеющей низкую токсичность, для применения в качестве превращенной в токсоид молекулы, то химическая детоксикация не является необходимой.
Очистка FHA
FHA может быть очищен из культурального супернатанта, как описано Cowell et al. (ссылка 58). Для увеличения выхода FHA в культуральных супернатантах могут быть использованы промоторы роста, такие как метилированные бета-циклодекстрины. Культуральный супернатант наносят на колонку гидроксилапатита. FHA адсорбируется на колонке, а РТ не адсорбируется. Колонку интенсивно промывают Тритоном Х-100 для удаления эндотоксина. Затем FHA элюируют при помощи 0,5 М NaCl в 0,1 М фосфате натрия и, если требуется, пропускают через колонку fetuin-Sepharose для удаления оставшегося РТ. Дополнительная очистка может включать в себя пропускание через колонку Sepharose CL-6B.
Альтернативно FHA может быть очищен при помощи моноклональных антител к этому антигену, причем антитела фиксируют на CNBr-активированной аффинной колонке (ссылка 59).
Альтернативно FHA может быть очищен при помощи хроматографии на перлите, как описано в вышеупомянутом ЕР 336736.
Очистка белка наружной мембраны 69 кДа (пертактина)
Белок наружной мембраны 69 кДа (69К или пертактин) может быть извлечен из бактериальных клеток инактивацией сначала этих клеток бактериостатическим агентом, таким как тимеросал, как описано в опубликованном ЕР 484621, включенном здесь в качестве ссылки. Инактивированные клетки суспендируют в водной среде, такой как ЗФР (рН 7-8), и подвергают повторяемой экстракции при повышенной температуре (45-60oС) с последующим охлаждением до комнатной температуры или до 4oС. Экстракции высвобождают белок 69К из клеток. Материал, содержащий белок 69К, собирают осаждением и пропускают через колонку Affi-Gel Blue. Белок 69К элюируют солью с высокой концентрацией, такой как 0,5 М хлорид магния. После диализа его пропускают через хроматографический фокусирующий носитель.
Альтернативно белок 69 кДа может быть очищен из культурального супернатанта культуры В. pertussis, как описано в опубликованной Международной заявке РСТ WO 91/15505, переуступленной автору и включенной здесь в качестве ссылки. Такой способ является предпочтительным, так как пертактин получают свободным от хроматографических материалов аденилатциклазы и красителя.
Другие подходящие способы очистки белка наружной мембраны 69 кДа из В. pertussis описаны в Патенте США No. 5276142, выданном Gotto et al. 4 января 1984 и в Патенте США No. 5101014, выданном Burns 31 марта 1992.
Другие компоненты изобретения
Вакцины данного изобретения содержат также иммуногены не Bordetella, в том числе столбнячный токсоид, дифтерийный токсоид, инактивированный полиовирус (IPV) и необязательно конъюгат дифтерийного токсоида или столбнячного токсоида и капсульного полисахарида Haemophilus influenzae типа b. Другие потенциальные компоненты поливалентных вакцин включают в себя белки наружной мембраны Haemophlivs influenzae, поверхностный антиген вируса гепатита В, эпидемического паротита, кори и коревой краснухи.
Конъюгаты столбнячного токсоида или дифтерийного токсоида и капсульного полисахарида Hib могут быть образованы выделением полирибозорибитолфосфата (PRP) из Haemophilus influenzae типа b дериватизацией PRP с получением дигидразида адипиновой кислоты и ковалентным конъюгированием со столбнячным токсоидом или дифтерийным токсоидом с получением соответственно конъюгатов PRP-T или PRP-D.
Каждый из этих антигенов индивидуально абсорбируют на адъюванте, таком как фосфат алюминия или гидроксид алюминия, вместе называемые квасцами, для обеспечения удобного и быстрого получения вакцин, содержащих выбранные относительные количества этих антигенов в вакцинах, описанных здесь.
Выбранные поливалентные вакцинные композиции
В выбранных вариантах данное изобретение обеспечивает вакцины со следующими характеристиками (мкг белка, используемые здесь, основаны на результатах теста Къельдаля, выполненного на очищенных концентратах, и выражены в мкг белкового азота), все из которых могут вводиться при помощи внутримышечной инъекции.
(a) CP20/20/5/3DT-mIPV (ГИБРИД):
Одна композиция содержит комбинацию компонента коклюшной вакцины (СР), комбинированного с дифтерийным (DI) и столбнячным (Т) токсоидами и инактивированным полиовирусом (mIPV), названную СР20/20/5/3DТ-mIРV (ГИБРИД). Полиовирус, выращенный на клетках MRC-5, обозначен mIPV, тогда как полиовирус, выращенный на клетках Vero, обозначен IPV или vIPV. Любой инактивированный полиовирусный материал может быть использован взаимозаменяемо в этих композициях.
Каждую дозу 0,5 мл для человека CP20/20/5/3DT-mIPV (ГИБРИД) готовили таким образом, чтобы она содержала приблизительно:
20 мкг коклюшного токсоида (РТ),
20 мкг нитевидного гемагглютинина (FHA),
5 мкг фимбриальных (жгутиковых) агглютиногенов 2 и 3 (FIM),
3 мкг белка пертактина наружной мембраны (69 кДа),
15 Lf дифтерийного токсоида,
5 Lf столбнячного токсоида,
40 единиц антигена D полиовируса типа 1,
8 единиц антигена D полиовируса типа 2,
32 единицы антигена D полиовируса типа 3,
1,5 мг фосфата алюминия,
0,6% 2-феноксиэтанол, в качестве консерванта.
(b) СР20/20/5/3РТ-mIРУ (ГИБРИД) + PRP-T:
Другая композиция содержит комбинацию компонента коклюшной вакцины (СР), комбинированного с дифтерийным (D) и столбнячным (Т) токсоидами и инактивированным полиовирусом (mIPV), названную CP20/20/5/3DT-mIPV (ГИБРИД), и используется для воссоздания (реконституирования) лиофилизированного PRP-T. Полученная композиция содержит на дозу 0,5 мл приблизительно:
20 мкг коклюшного токсоида (РТ),
20 мкг нитевидного гемагглютинина (FHA),
5 мкг фимбриальных (жгутиковых) агглютиногенов 2 и 3 (FIM),
3 мкг белка пертактина наружной мембраны (69 кДа),
15 Lf дифтерийного токсоида,
5 Lf столбнячного токсоида,
10 мкг очищенного капсульного полисахарида полирибозорибитолфосфата (PRP) Haemophilus influenzae типа b, ковалентно связанного с 20 мкг столбнячного токсоида,
40 единиц антигена D полиовируса типа 1,
8 единиц антигена D полиовируса типа 2,
32 единицы антигена D полиовируса типа 3,
1,5 мг фосфата алюминия,
0,6% 2-феноксиэтанол.
(с) CP20/20/5/3DT-PRP-T-IPV (ГИБРИД):
Дополнительная композиция содержит комбинацию компонента коклюшной вакцины (СР), комбинированного с дифтерийным (D) и столбнячным (Т) токсоидами и инактивированным полиовирусом (mIPV) и PRP-T, и названа CP20/20/5/3DT-PRP-T-IPV (ГИБРИД). Каждая доза 0,5 мл для человека этой композиции содержит приблизительно:
20 мкг коклюшного токсоида (РТ),
20 мкг нитевидного гемагглютинина (FHA),
5 мкг фимбриальных (жгутиковых) агглютиногенов 2 и 3 (FIM),
3 мкг белка пертактина наружной мембраны (69 кДа),
15 Lf дифтерийного токсоида,
5 Lf столбнячного токсоида,
10 мкг очищенного капсульного полисахарида полирибозорибитолфосфата (PRP) Haemophilus Influenzae типа b, ковалентно связанного с 20 мкг столбнячного токсоида,
40 единиц антигена D полиовируса типа 1,
8 единиц антигена D полиовируса типа 2,
32 единицы антигена D полиовируса типа 3,
1,5 мг фосфата алюминия,
0,6% 2-феноксиэтанол.
Клинические испытания
(а) DРТ-компонентная коклюшная вакцина
Ряд клинических испытаний проводили на людях, как описано здесь, для установления безопасности, нереактогенности и применимости компонентных коклюшных вакцин, содержащих фимбриальные агглютиногены, полученные, как описано здесь, для иммунитета против коклюша. В частности, были получены иммунные ответы на каждый из антигенов, содержащихся в этих вакцинах (как показано, например, в таблице 3). Одну конкретную поливалентную коклюшную вакцину CP10/5/5/3DT анализировали при двукратном повторе клинического испытания на людях группы риска при контроле с плацебо для оценки эффективности этой вакцины против типичного коклюша.
Определение случая типичного коклюшного заболевания было следующим.
Двадцать один день или более спазматического кашля и либо подтвержденный в культуре В. pertussis, либо серологическое доказательство специфической для Bordetella инфекции, о которой свидетельствовало увеличение на 100% антител IgG или IgA в ELISA против FHA или РТ в параллельных сыворотках, либо, если серологические данные отсутствуют, исследуемый ребенок был в контакте с ребенком с подтвержденным в культуре В. pertussis с приступами кашля в пределах 28 дней до или после приступов кашля у обследуемого ребенка.
Результаты этого исследования показали, что вакцина CP10/5/5/3DT является на примерно 85% эффективной в предотвращении коклюша, определенного в определении типичного заболевания коклюша, приведенного выше. В том же самом исследовании двухкомпонентная коклюшная бесклеточная вакцина, содержащая только РТ и FHA, была эффективна на 58% (PT25.FHA25.DT) и цельноклеточная коклюшная вакцина (DTP) была эффективна на примерно 48% (см. таблицу 4 ). Кроме того, вакцина CP10/5/5/3DT предотвращала слабый коклюш, определяемый как кашель с продолжительностью, по меньшей мере, 1 день, с эффективностью примерно 77%. В частности, показатели полученного иммунного ответа были, по существу, такими же, что и показатели, полученные после иммунизации цельноклеточными коклюшными вакцинами, которые, как сообщалось, являются высокоэффективными против коклюша.
(b) Поливалентная DPT-PRP-T-IPV-вакцина
(I) Безопасность и иммуногенность компонентных коклюшных вакцин в комбинации с адсорбированными дифтерийным токсоидом и столбнячным токсоидом, содержащей конъюгат Haemophilus influenzae типа b - столбнячный токсоид вакциной и инактивированной вакциной против полиомиелита, выращенной на клетках Vero (CP20/20/5/3DT-PRP-T-IPV), сравнивали с цельноклеточной коклюшной вакциной (а) в комбинации с адсорбированными дифтерийным и столбнячным токсоидами и инактивированной вакциной против полиомиелита, (b) в комбинации с адсорбированными дифтерийным и столбнячным токсоидами и инактивированной вакциной против полиомиелита, выращенной на клетках MRC-5 (DPT-Polio-адсорбированной), используемой для воссоздания лиофилизированной, содержащей конъюгат Haemophilus influenzae типа b - столбнячный токсоид вакцины (PENTAТМ), или (с) компонентной коклюшной вакциной в комбинации с адсорбированными дифтерийным и столбнячным токсоидами, содержащей конъюгат Haemophilus influenzae типа b - столбнячный токсоид вакциной и с инактивированной полиомиелитной вакциной, выращенной на клетках MRC-5 (CP20/20/5/3DT-mIPV), предоставляемой отдельно от содержащей конъюгат Haemophilus Influenzae типа b - столбнячный токсоид вакцины или используемой для воссоздания такой лиофилизированной вакцины (PRP-T), на младенцах в возрасте 2, 4, 6 и 18 месяцев.
Это рандомизированное, имеющее контроли испытание охватывало 897 двухмесячных младенцев, получавших один из восьми различных вакцинных вариантов (групп): СР20/20/5/3DТ-РRР-Т-IPV (жидкую); СР20/20/5/3DТ, даваемую параллельно, но в отличающемся месте от PRP-T, или контрольную вакцину, цельноклеточную DPT-Polio, используемую для воссоздания PRP-T (PENTAТМ).
Все исследуемые вакцины хорошо переносились. Не были заметны значимые различия в скоростях реакции между двумя типами рекомбинантных коклюшных комбинаций. Дети, которые получали комбинированную вакцину CP20/20/5/3DT-mIPV, используемую для воссоздания PRP-T, имели слегка более высокие скорости локальных реакций в сравнении с теми же продуктами, вводимыми в различных местах. Все компонентные коклюшные комбинации имели устойчиво более низкие скорости локальных и системных реакций, чем цельноклеточная комбинация. Различия в скоростях реакции между компонентной коклюшной вакциной и цельноклеточной вакциной были наиболее очевидными через 24 часа непосредственно после вакцинации.
Обе компонентные коклюшные комбинации вызывали превосходные ответные реакции на все антигены. Во всех ситуациях ответы; на коклюшный РТ, FHA и пертактин превосходили ответы, наблюдаемые на цельноклеточных комбинациях. Не наблюдали значимых различий между компонентными и цельноклеточными комбинациями. Не наблюдали значимых различий между компонентными и цельноклеточными композициями для антител против PRP, дифтерии и полиомиелита 1 и 2. Обе компонентные коклюшные композиции производили более высокие ответы на антиген столбняка, чем PENTAТМ. Обе компонентные композиции вызывали сходные серологические ответы на все антигены, за исключением полио 3, для которого CP20/20/5/3DT-mIPV, использованная для воссоздания PRP-T, производила более высокие ответы, чем СР20/20/5/3DТ-РРР-T-IPV. Способ введения не влиял на серологические ответы на любой антиген, за исключением столбняка. Как в комбинированных, так и в отдельных вариантах 100% детей были защищены (более 0,01 ЕЕ/мл) против столбняка после 3 доз вакцины.
Наиболее важно, что все вакцинные группы давали хорошие ответы на PRP-T, причем 98,3% детей достигали уровней более 0,15 мкг/мл и более 86,1% детей достигали уровней более 1,0 мкг/мл. Эти цифры сравнимы с цифрами, наблюдаемыми в прежних исследованиях, в которых цельноклеточную вакцину использовали с PRP-T.
Полученные и описанные выше серологические ответы представлены в таблицах 5-7 (Н = гибрид).
(II) Безопасность и иммуногенность компонентной коклюшной вакцины в комбинации с адсорбированными дифтерийным и столбнячным токсоидами и инактивированной полиомиелитной вакциной, выращенной на клетках MRC-5 (СР20/20/5/3DТ-mIРV), предоставляемой отдельно или используемой для воссоздания лиофилизированной, содержащей конъюгат Haemophilus influenzae типа b - столбнячный токсоид вакцины (PRP-T), сравнивали с цельноклеточной коклюшной вакциной в комбинации с адсорбированными дифтерийным и столбнячным токсоидами и инактивированной полиомиелитной вакциной, выращенной на клетках MRC-5 (DPT-Polio-адсорбированной), используемой для воссоздания лиофилизированной, содержащей конъюгат Haemophilus influenzae типа b - столбнячный токсоид вакцины (PENTAТМ), на детях в возрасте 18-19 месяцев.
Это исследование в пяти вариантах (группах) включало в себя группу цельноклеточной вакцины PENTAТМ в качестве контроля и вакцину СР20/20/5/3DТ-mIРV, используемую для реконституирования PRP-T. Пятой группе вводили CP20/20/5/3DT-mIPV вместе с PRP-T, но в другом месте. 489 субъектов получали вакцину в возрасте 18-19 месяцев, из них 466 (95%) завершили это исследование согласно протоколу. Вакцина СР20/20/5/3DТ-mIРV, использованная для воссоздания PRP-T, была значимо менее реактогенной, чем PENTAТМ, в первые 24 часа после вакцинации. Воссозданный продукт давал слегка более высокие скорости местных реакций, чем отдельно вводимый продукт.
PENTAТМ вызывала более высокие ответы на полио 1, чем вакцина СР20/20/5/3DТ-mIРV, используемая для воссоздания PRP-T. Не наблюдали значимых различий для антител против PRP, дифтерии, коклюшного агглютинина, фимбриальных агглютиногенов, полио 2 или полио 3. СР20/20/5/3DТ-mIРV, используемая для воссоздания PRP-T, давала значимо более высокие серологические ответы на РТ коклюша, FHA и пертактин. CP20/20/5/3DT-mIPV, используемая для воссоздания PRP-T, давала стойкие серологические ответы на все испытанные антигены. Не наблюдали значимых различий между вакциной CP20/20/5/3DT-mIPV, предоставляемой отдельно и используемой для воссоздания PRP-T, за исключением столбнячного антитоксина (6,78 против 4,91 ЕЕ/мл).
Это исследование продемонстрировало, что вакцина СР20/20/5/3DТ-mIРV, используемая для реконституирования PRP-T, вызывала устойчивые серологические ответы в трех сериях опытов и была более иммуногенной, чем вакцина PENTAТМ, в отношении ответов на антиген коклюша. Вакцина CP20/20/5/3DT-mIPV вызывала также значимо более низкие скорости местных и системных реакций, чем PENTAТМ.
(III) Безопасность и иммуногенность компонентной коклюшной вакцины, комбинированной с адсорбированными дифтерийным и столбнячным токсоидами и инактивированной полиомиелитной вакциной, выращенной на клетках MRC-5 (CP20/20/5/3DT-mIPV), сравнивали с цельноклеточной коклюшной вакциной в комбинации с адсорбированными дифтерийным и столбнячным токсоидами и инактивированной полиомиелитной вакциной, выращенной на клетках MRC-5 (DPT-polio-адсорбированной), на детях в возрасте 4-6 лет.
164 субъекта были рандомизированно распределены в отношении 4:1 для получения либо CP20/20/5/3DT-mIPV (n= 131), либо DPT-Polio (n=33). В этом исследовании не наблюдали значимых или тяжелых побочных событий. CP20/20/5/3DT-mIPV давала устойчиво более низкие скорости как местных, так и системных реакций, в частности, в период времени 0-24 часа. Местные реакции были обычными для обеих групп с 97% реципиентами DPT-Polio и 76,9% реципиентами СР20/20/5/3DТ-mIРУ, имеющими некоторую местную реакцию в периоде времени 0-24 часа. Местные реакции CP20/20/5/3DT-mIPV были обычно слабыми или умеренными. В противоположность этому более половины реципиентов DPT-Polio имели местные реакции, классифицируемые как тяжелые. Болезненность мест инъекции обычно исчезала за 72 часа, но краснота или набухание держались стойко в период 24-72 часа.
Системные реакции в интервале 0-24 часа были менее обычными у реципиентов CP20/20/5/3DT-mIPV (38,5%), чем у реципиентов DPT-Polio (90,9%). Системные реакции в интервале времени 24-72 часа не были обычными для обеих групп.
Ответы на антигены дифтерии, столбняка, полио 2 и 3 были сравнимы между двумя вакцинами. Реципиенты DPT-Polio имели значимо более высокую реакцию на полио 1 (15462), чем реципиенты CP20/20/5/3DT-mIPV (10903). Все субъекты имели превосходные ответные реакции и могли рассматриваться как имеющие иммунитет против соответствующих заболеваний. Серологические ответы на все коклюшные антигены были значимо более высокими у реципиентов CP20/20/5/3DT-mIPV.
(IV) Безопасность и иммуногенность компонентных коклюшных вакцин в комбинации с адсорбированными дифтерийным токсоидом и столбнячным токсоидом, содержащей конъюгат HaemophiIus influenzae типа b - столбнячный токсоид вакциной и инактивированной вакциной против полиомиелита (с полиовирусом, выращенным на клетках MRC-5 (CP20/20/5/3DT-PRP-T-mIPV), сравнивали с цельноклеточной коклюшной вакциной в комбинации с адсорбированными дифтерийным и столбнячным токсоидами и инактивированной вакциной против полиомиелита (с полиовирусом, выращенным на клетках MRC-5) (DPT-Polio-адсорбированной), используемой для воссоздания лиофилизированной, содержащей конъюгат Haemophilus Influenzae типа b - столбнячный токсоид вакцины (PENTAТМ), или компонентной коклюшной вакциной в комбинации с адсорбированными дифтерийным и столбнячным токсоидами и с инактивированной полиомиелитной вакциной, выращенной на клетках MRC-5 (CP20/20/5/3DT-mIPV), используемой для воссоздания лиофилизированной, содержащей конъюгат Haemophilus influenzae типа b - столбнячный токсоид вакцины (PRP-T), на младенцах в возрасте 18-19 месяцев.
Целью этого рандомизированного, с тремя группами, контролируемого, одинарно слепого исследования была оценка безопасности и иммуногенности двух новых бесклеточных коклюшных комбинаций, CP20/20/5/3DT-PRP-IPV и CP20/20/5/3DT-mIPV, используемой для реконституирования PRP-T, с PENTAТМ (цельноклеточной коклюшной DPT-Polio-вакциной, используемой для реконституирования PRP-T), на детях в возрасте 18-19 месяцев. В трех вакцинируемых группах участвовали всего 99 детей, по 33 в каждой из групп, из которых 97 (98%) завершили исследование согласно протоколу.
В этом исследовании не наблюдали серьезных реакций. Реципиенты PENTAТМ, более заметно, испытывали значимо умеренные или тяжелые местные и системные реакции, чем реципиенты двух других вакцин. Различия были наиболее выраженными к 24 часам и достигали статистической значимости в отношении лихорадочного состояния, покраснения, набухания, болезненности, суетливости, пониженной активности и понижения аппетита. Реакции имели тенденцию в сторону слабых реакций у детей, получавших компонентные коклюшные комбинации. Значимых различий в скоростях реакции не наблюдали между двумя компонентными коклюшными композициями, хотя суетливость наблюдали более часто к 24 часам у реципиентов CP20/20/5/3DT-mIPV, используемой для реконституирования PRP-T, по сравнению с CP20/20/5/3DT-PRP-T-mIPV (18% против 3%).
Серологические ответы были удовлетворительными у 100% участников, достигших уровней, рассматриваемых как защитные для антител против дифтерийного токсина (более или равно 0,01 МЕ/мл), против столбнячного токсина (более или равно 0,01 МЕ/мл) и против PRP (более или равно 1,0 мкг/мл). Детектируемые нейтрализующие антитела к полио 1, 2 и 3 были отмечены у всех участников после иммунизации.
Дифтерийные ответные реакции были более высокими у реципиентов PENTAТМ, что отражало более высокое содержание антигенов этой вакцины (25 Lf против 15 Lf).
Количество коклюшных антител было устойчиво выше в случае двухкомпонентных коклюшных комбинаций в сравнении с PENTAТМ, достигая статистической значимости для ответов антител против РТ, против FHA и против пертактина GMT. Антитела против фимбриальных агглютиногенов и коклюшного гемагглютинина были также в большем количестве у реципиентов компонентной коклюшной вакцины, хотя различия не достигали статистической значимости.
В целом, это исследование показало, что эти две бесклеточные коклюшные комбинации: CP20/20/5/3DT-mIPV, используемая для реконституирования PRP-T, и CP20/20/5/3DT-PRP-T-IPV, были сравнимы и вызывали удовлетворительно низкие скорости реакции и высокие серологические ответы при предоставлении в виде бустер-инъекции детям в возрасте 18-19 месяцев.
(V) Безопасность и иммуногенность компонентной коклюшной вакцины в комбинации только с адсорбированными дифтерийным и столбнячным токсоидами (СР20/20/5/3DТ) или в комбинации с одним или двумя инактивированными полиовирусами, одним mIPV, выращенным на клетках MRC-5, и другим vIPV, выращенным на клетках Vero, или пероральной полиомиелитной вакциной (OPV), исследовали на детях в возрасте 17-19 месяцев.
Это исследование с пятью группами было предназначено для исследования взаимодействия между СР20/20/5/3DТ и двумя IPV (выращенными на клетках Vero или на клетках MRC-5). Оба IPV объединяли в виде единого жидкого продукта с CP20/20/5/3DT-mIPV или CP20/20/5/3DT-vIPV или вводили совместно, но в различные места инъекции (СР20/20/5/3DТ+mIРV и CP20/20/5/3DT+vIPV). Пятая группа исследования получала СР20/20/5/3DТ и OPV одновременно. Все субъекты получали PRP-T при постиммунизационном взятии крови. В этом исследовании не оценивали образования антител против PRP (см. таблицу I).
В общем, не было различий в скоростях побочных реакций, сообщаемых инактивированными полиомиелитными вакцинами, полученными из клеток MRC-5 или из клеток Vero, вводили ли инъекцию в виде отдельной инъекции или в сочетании с вакциной CP20/20/5/3DT (ГИБРИД).
Не наблюдали значимых различий между группами для РТ, FHA и пертактина. Ответы у детей, получавших СР20/20/5/3DТ (ГИБРИД) и OPV были слегка, но не значимо выше, чем у детей, получавших СР20/20/5/3DТ (ГИБРИД) и IPV из клеток Vero, для FIM, агглютинина коклюша, дифтерии и столбняка. Полиоответы были обычно сравнимыми или более высокими у детей, получавших вакцину IPV, против вакцины OPV. Все, кроме одного индивидуума, имели коклюшный агглютинин более 1: 64. Все, кроме одного субъекта, достигали уровней дифтерийного антитоксина, более или равных 0,1 Е/мл, и все достигали уровней столбнячного антитоксина более или равных 0,1 ЕЕ/мл.
Результаты этого исследования показали, что СР20/20/5/3DТ (ГИБРИД) в комбинации с IPV (из клеток MRC-5 или из клеток Vero) является безопасной и иммуногенной для детей в возрасте 17-19 месяцев. Комбинированные вакцины были, по меньшей мере, такими же иммуногенными, как вакцины, вводимые в виде отдельных инъекций, и в некоторых случаях более иммуногенными. Комбинирование такой вакцины в виде единой инъекции не было связано со значимым увеличением локальных побочных реакций. Не были обнаружены существенные различия в неблагоприятных побочных реакциях или в иммунном ответе на эти два препарата IPV ни при отдельных инъекциях, ни при комбинированных продуктах. Включение IPV в вакцину не увеличивало интенсивности побочных вредных реакций в сравнении с вакциной СР20/20/5/3DТ (ГИБРИД), вводимой отдельно (т.е. с OPV).
Серологические результаты, полученные в этом исследовании, сведены в таблицу 8 (Н = гибрид).
(VI) Безопасность и иммуногенность компонентных коклюшных вакцин в комбинации только с адсорбированными дифтерийным и столбнячным токсоидами (СР20/20/5/3DТ и CP10/5/5/3DT) или в комбинации с содержащей конъюгат Haemophilus influenzae типа b - столбнячный токсоид вакциной определяли на детях в возрасте 17-19 месяцев.
Исследования на шести группах были предназначены для исследования взаимодействия между классическими (СР10/5/5/3DТ) и гибридными (СР20/20/5/3DТ) компонентными коклюшными композициями и содержащей конъюгат Haemophilus influenzae типа b - столбнячный токсоид вакциной (PRP-T) на детях в возрасте 18-19 месяцев.
Использовали три схемы, в которых (а) каждую из этих компонентных вакцин использовали для реконституирования PRP-T, (b) вводили одновременно, но в разные места с PRP-T или (с) PRP-T вводили через месяц после введения компонентной коклюшной вакцины. Все дети получали OPV при первом визите и были примированы той же самой компонентной коклюшной вакциной в возрасте 2, 4 и 6 месяцев. Все дети предварительно участвовали в большом исследовании безопасности этих двух вакцинных композиций.
В этом исследовании были зарегистрированы всего 545 субъектов, из которых 542 (99%) завершили исследование (см. таблицу II).
Серологические ответы были, как правило, более высокими на большинстве антигенов, когда компонентную коклюшную комбинированную вакцину и PRP-T вводили в один и тот же день по сравнению с введением в разные дни (см. таблицу 9).
Важно, что образование антител против PRP не уменьшалось, когда PRP-T вводили отдельно, в сравнении с введением его с компонентой коклюшной комбинированной вакциной, в тот же самый день. GMT после иммунизации у детей, получавших эти вакцины, были значимо более низкими. Различия в антителах к PRP между комбинированными инъекциями и отдельными инъекциями наблюдали, когда субъекты были разделены компонентной коклюшной вакцинной композицией. Реципиенты вакцины СР20/20/5/3DТ (ГИБРИД) показали более низкие уровни антител против PRP, когда эту вакцину вводили в комбинированном виде, а не отдельно. Эти различия не наблюдались с реципиентами СР10/5/5/3DТ, и различия исчезали, когда эти группы объединяли. Все участники достигали уровней антител против PRP более или равно 0,15 мкг/мл и более 98% каждой группы имели уровень более или равно 1,0 мкг/мл. Только четыре участника (0,7 %) в этом исследовании не достигали этого уровня; трое в группе отдельных инъекций в разные дни и один в группе комбинированных инъекций. Более 82% каждой группы превышали титры 10 мкг/мл антител против PRP.
Из индуцированных локальных реакций только болезненность сообщалась более часто в объединенной группе (27,8%) в сравнении с группой с вакцинациями в разные дни (16,7%). Эта частота не отличалась от частоты, наблюдаемой в группе, в которой вакцины вводили в тот же самый день, когда и отдельные инъекции (24,2%).
В целом, системная реакция сообщалась с одинаковой частотой (60-62,1%) участникам каждой из этих вакцинных групп. Лихорадочное состояние сообщалось приблизительно одной трети участников. Только суетливость сообщалось чаще в группе комбинированной инъекции (33,3%) в сравнении с группой отдельных инъекций (22,0%) или группе инъекций в разные дни (22,8%).
Обобщая результаты этого испытания, можно отметить, что совместное введение СР10/5/5/3DТ или CP20/20/5/3DT и PRP-T в один и тот же день не мешает образованию антител против PRP, но может фактически усиливать его. Серологические ответы на другие антигены также были превосходными. Столбняк был единственным антигеном, на который влияло смешивание двух вакцин вместе, но все дети имели высокие уровни иммунитета.
Для этих последних клинических испытаний готовили CP10/5/5/3DT-IPV, росший на клетках MRC-5, для реконституирования с ActHib (PRP-T) и A5I (CP10/5/5/3DT-PRP-T-IPV), росший на клетках Vero (3 мкг/мл). Компонентные коклюшные антигены индивидуально адсорбировали на фосфате алюминия 3 мг/мл в отсутствие консерванта. В связи с этим РТ был в 10 мМ фосфате калия, 0,15 М NaCl, 5% глицерине, FHA был в 10 мМ фосфате калия, 0,5 М NaCI, 69K был в 10 мМ фосфате калия, 0,15 М NaCl, фимбриальные агглютиногены были в 10 мМ фосфате калия, 0,15 М NaCl. D был адсорбирован на фосфате алюминия (6,25 мг/мл) в концентрации 300 Lf/мл. 2-Феноксиэтанол добавляют в качестве консерванта до 0,6%. Т был адсорбирован на фосфате алюминия (6,25 мг/мл) при концентрации 300 Lf/мл. 2-Феноксиэтанол добавляют до 0,6%.
Адсорбированные компонентные коклюшные антигены комбинировали с адсорбированным D и адсорбированным Т при концентрации 3,65 доз/мл или 55% конечного объема. Содержание 2-феноксиэтанола было 0,6%. Перед объединением с mIPV или vIPV/PRP-T проверяли стерильность, содержание алюминия и содержание 2-феноксиэтанола. Для 5 мл добавляли mIPV и 2-феноксиэтанол и разбавляли до конечной концентрации. Для A5I добавляли vIPV, PRP-T и 2-феноксиэтанол и разбавляли до конечной концентрации.
Обобщая результаты клинических испытаний, можно отметить, что CP20/20/5/3DT-mIPV, использованная для воссоздания PRP-T, производит сравнимые серологические ответы на антигены дифтерии, столбняка и полио 1, 2 и 3, в сравнении с PENTAТМ (содержащей цельноклеточную коклюшную вакцину). Ответы в виде образования антител против PRP сравнимы или более высокие, чем ответы, наблюдаемые с PENTAТМ как при младенческих дозах, так и при бустер-дозах. Ответы на столбняк являются более низкими, чем для CP20/20/5/3DT-mIPV, используемой для воссоздания PRP-T, при сравнении с CP20/20/5/3DT-mIPV, вводимой отдельно от PRP-T, но это снижение клинически не имело значения. В соответствии с другими исследованиями цельноклеточная вакцина производит сравнимые или более высокие фимбриальные и агглютининовые ответные реакции, чем компонентная коклюшная вакцина, однако известно, что используемая цельноклеточная вакцина содержит высокоиммуногенный фимбриальный компонент. Все остальные коклюшные ответные реакции были устойчиво более высокими при использовании CP20/20/5/3DT-mIPV для воссоздания PRP-T, чем PENTAТМ. Таким образом, данное изобретение обеспечивает поливалентные иммуногенные композиции, в которых иммунные ответные реакции на антигены не являются сниженными или ухудшенными другими компонентами или их включением в поливалентную вакцину. Уменьшенные иммунные ответы иногда относят к примесям.
Получение и применение вакцин
Таким образом, иммуногенные композиции, пригодные для использования в качестве вакцин, могут быть получены из иммуногенов, как описано здесь. Такая вакцина вызывает иммунный ответ в субъекте, который продуцирует антитела.
Иммуногенные композиции, в том числе вакцины, могут быть приготовлены в виде инъекционных растворов, в виде жидких растворов или эмульсий. Эти иммуногены могут быть смешаны с фармацевтически приемлемыми наполнителями, которые совместимы с иммуногенами. Такие наполнители могут включать в себя воду, солевой раствор, декстрозу, глицерин, этанол и их сочетания. Иммуногенные композиции и вакцины могут дополнительно содержать вспомогательные вещества, такие как увлажняющие или эмульгирующие агенты, буферящие рН агенты или адъюванты для усиления их эффективности. Иммуногенные композиции и вакцины могут вводиться парентерально, подкожной или внутримышечной инъекцией. Иммуногенные препараты и вакцины вводят способом, совместимым с дозированной формой, и в таком количестве, которое будет терапевтически эффективным, иммуногенным и защитным. Количество, необходимое для введения, зависит от субъекта, подлежащего лечению, в том числе, например, от способности иммунной системы индивидуума синтезировать антитела и, если требуется, производить клеточно-опосредованный ответ. Точные количества активного ингредиента, подлежащего введению, зависят от суждения практикующего врача. Однако подходящие диапазоны доз легко могут быть определены специалистом в данной области и могут быть порядка микрограммов иммуногенов. Подходящие схемы для первоначального введения и бустер-доз также варьируют, но могут включать в себя первоначальное введение, за которым следуют последующие введения. Доза может также зависеть от способа введения и будет также меняться в соответствии с размером хозяина.
Концентрация иммуногенов в иммуногенной композиции согласно данному изобретению составляет обычно от примерно 1 до примерно 95%. Иммуногенность может быть значимо улучшена, если антигены вводят совместно с адъювантами, обычно применяемыми в виде 0,005-0,5% раствора в забуференном фосфатом солевом растворе. Адъюванты усиливают иммуногенность антигена, но не являются обязательно сами иммуногенными. Адъюванты могут действовать путем удерживания антигена локально вблизи места введения с образованием эффекта депо, облегчающего медленное, поддерживаемое высвобождение антигена в клетки иммунной системы. Адъюванты могут также аттрагировать клетки иммунной системы к депо антигенов и стимулировать такие клетки к индуцированию иммунных ответов.
Иммуностимуляторные агенты или адъюванты использовали в течение многих лет для улучшения иммунных реакций хозяина, например, на вакцины. Внутренние адъюванты, такие как липополисахариды, обычно являются компонентами убитых или аттенуированных бактерий, используемых в качестве вакцин. Внешние адъюванты являются иммуномодуляторами, которые обычно нековалентно связывают с антигенами и готовят для усиления иммунных ответов хозяина. Таким образом, были идентифицированы адъюванты, которые усиливают иммунный ответ на парентерально доставляемые антигены. Однако некоторые из этих адъювантов являются токсичными и могут вызывать нежелательные побочные эффекты, что делает их непригодным для применения на людях и многих животных. Действительно, только гидроксид алюминия и фосфат алюминия (вместе обычно называемые квасцами) рутинно используют в качестве адъювантов в вакцинах для человека и в ветеринарных вакцинах. Эффективность квасцов в увеличении образования антител на дифтерийный и столбнячный токсоиды является хорошо установленной.
Широкий диапазон внешних адъювантов может вызывать сильные иммунные ответы на антигены. Они включают в себя сапонины в комплексе с мембранными белковыми антигенами (иммуностимулирующие комплексы), блоксополимер полиоксиэтилена и полиоксипропилена (pluronic) с минеральным маслом, убитые микобактерии в минеральном масле, полный адъювант Фрейнда, бактериальные продукты, такие как мурамилдипептид (MDP) и липополисахарид (LPS), а также липид А и липосомы.
Для эффективной индукции гуморальных иммунных ответов (HIR) и клеточно-опосредованного иммунитета (CMI) иммуногены часто эмульгируют в адъювантах. Многие адъюванты являются токсичными, индуцирующими гранулемы, острые и хронические воспаления (полный адъювант Фрейнда, FCA), цитолиз (сапонины и полимеры Pluronic), и обладают пирогенностыо, индуцируют артрит и передний увеит (LPS и MDP). Хотя FCA является превосходным адъювантом и широко используется в исследованиях, он не лицензирован для применения в вакцинах для человека и в ветеринарных вакцинах вследствие его токсичности.
Желательные характеристики идеальных адъювантов включают в себя:
(1) отсутствие токсичности;
(2) способность стимулировать долгосрочный иммунный ответ;
(3) простоту изготовления и стабильность при долгосрочном хранении;
(4) способность вызывать как CMI, так и HIR на антигены, вводимые различными путями;
(5) синергизм с другими адъювантами;
(6) способность селективного взаимодействия с популяциями антигенпредставляющих клеток (АРС);
(7) способность специфической индукции подходящих специфических для Тн1- или Тн2-клеток иммунных ответов и
(8) способность селективного увеличения уровней подходящих изотипов антител (например, IgA) против антигенов.
Патент США No. 4855283, выданный Lockhoff et al. 8 августа 1989, который включен здесь в качестве ссылки, описывает аналоги гликолипидов, в том числе N-гликозиламиды, N-гликозилмочевины и N-гликозилкарбаматы, каждый из которых замещен в сахарном остатке аминокислотой, в качестве иммуномодуляторов или адъювантов. Так, Lockhoff et al. (Патент США No. 4855283 и ссылка 60) сообщили, что аналоги N-гликолипида, проявляющие структурное сходство с природно-встречающимися гликолипидами, способны вызывать сильные иммунные ответы как в вакцине вируса простого герпеса, так и в вакцине вируса псевдобешенства. Некоторые гликолипиды были синтезированы из алкиламинов с длинной цепью и жирных кислот, которые непосредственно связаны с сахарами через аномерный атом углерода, для имитации функций природно-встречаюшихся липидных остатков.
В Патенте США No. 4258029, выданном Moloney, переуступленном заявителю настоящего изобретения и включенном здесь в качестве ссылки, сообщается, что гидрохлорид октадецилтирозина (ОТН) функционирует как адъювант при образовании комплекса со столбнячным токсоидом и инактивированной формалином вакциной вируса полиомиелита типа I, II и III. Nixon-George et al. (ссылка 61) также сообщили, что октадециловые эфиры ароматических аминокислот, комплексированные с рекомбинантным поверхностным антигеном вируса гепатита В, усиливали иммунные ответы хозяина против вируса гепатита В.
ПРИМЕРЫ
Приведенное выше описание в общих чертах описывает данное изобретение. Более полное понимание может быть получено со ссылкой на следующие далее характерные примеры. Эти примеры описаны лишь для целей иллюстрации и не предназначены для ограничения объема данного изобретения. Изменения по форме и замена эквивалентов предполагаются, так как обстоятельства могут делать их целесообразными. Хотя здесь использовались специфические термины, такие термины предназначены для описания, а не для целей ограничения.
Способы белковой биохимии, ферментации и иммунологии, использовались, но не были подробно описаны в данном описании; эти примеры достаточно описаны в научной литературе и находятся вполне в пределах понимания специалистами в данной области.
Пример 1.
Этот пример описывает рост Bordetella pertussis.
Основной посевной материал
Культуры основного посевного материала Bordetella pertussis хранили в виде лиофилизированных партий материала при 2-8oС.
Рабочий посевной материал
Лиофилизированную культуру переводили в среду Hornibrook и использовали для засева чашек с агаром Bordet-Gengou (BGA). Среда Hornibrook имеет следующий состав, г/л:
Гидролизат казеина (обработанный углем) - 10,0
Никотиновая кислота - 0,001
Хлорид кальция - 0,002
Хлорид натрия - 5,0
Гексагидрат хлорида магния - 0,025
Хлорид калия - 0,200
Двухосновный фосфат калия - 0,250
Крахмал - 1,0
Дистиллированная вода, л - До 1
Доводили рН до 6,9±0,1 1% раствором карбоната натрия. Среду разливали в пробирки и стерилизовали паром в автоклаве в течение 20 минут и автоклавировали в течение 20 минут при 121 -124oС. Посевной материал субкультивировали дважды, сначала на чашках BGA, затем на компонентном коклюшном агаре (СРА). Компонентный коклюшный агар имеет следующий состав, г/л:
NaCl - 2,5
KH2PO4 - 0,5
КСl - 0,2
MgCl2(H2O)6 - 0,1
Трис-основание - 1,5
Казаминокислоты - 10,0
NaH-глутамат - 10,0
Концентрированная НСl - До рН 7,2
Агар - 15,0
Факторы роста (CPGF), мл/л - 10,0
Факторы роста для компонентной коклюшной вакцины (CPGF)-100X имеют следующий состав, г/л:
L-Цистеин НСl - 4,0
Ниацин - 0,4
Аскорбиновая кислота - 40,0
Глутатион восстановленный - 15,0
Fe2SO4(Н2O)7 - 1,0
Диметил-β-циклодекстрин - 100
CaCl2(H2O)2 - 2,0
Конечную культуру суспендировали в буфере для суспензии коклюшного посевного материала (CPSB), разливали в виде аликвот 2 - 4 мл и хранили замороженными при -60 -85oС. Буфер для суспензии коклюшного посевного материала (PSSB) имеет следующий состав:
Казаминокислоты, г/л - 10,0
Трис-основание, г/л - 1,5
Безводный глицерин, мл/л - 100
Концентрированная НСl - До рН 7,2
Эти суспензии в глицерине обеспечивали исходный материал для получения рабочего посевного материала.
Процесс культивирования
Размножение рабочего посевного материала проводили во флаконах Roux для компонентной коклюшной вакцины в течение 4-7 дней при 34-38oС. После этого культивирования клетки смывали с агара компонентным коклюшным бульоном (СРВ). Пробы наблюдали на окрашивание по Граму, на чистоту культуры и непрозрачность.
Клетки переносили в 4-литровые конические колбы, содержащие СРВ, и инкубировали при 34-38oС в течение 20-26 часов при встряхивании. Пробы наблюдали на окрашивание по Граму и проверяли чистоту культуры. Колбы сливали вместе и эту суспензию использовали для засева двух ферментеров, содержащих СРВ (объем 10 литров, исходная OD600 0,1-0,4). Этот посевной материал выращивали до конечной OD600 5,0-10,0. Пробы тестировали окрашиванием по Граму на чистоту культуры, при помощи ELISA на специфический антиген и на стерильность.
Пример 2.
Этот пример описывает очистку антигенов из культуры клеток Bordetella pertussis.
Получение бульона и концентратов клеток
Бактериальную суспензию выращивали в двух производственных ферментерах при 34-37oС в течение 35-50 часов. Из ферментеров брали пробы для тестирования стерильности среды. Суспензию подавали в центрифугу непрерывного действия с перегородками в роторе (12000•g) для отделения клеток от бульона. Клетки собирали для последующей экстракции фимбриального (жгутикового) компонента. Осветленную жидкость пропускали через мембранный фильтр, менее или равный 0,22 мкм. Отфильтрованную жидкость концентрировали ультрафильтрацией при помощи мембраны с номинальным ограничением молекулярной массы 10-30 кДа (NMWL). Концентрат хранили для последующего разделения и очистки компонентов коклюшного токсина (РТ), нитевидного гемагглютинина (FHA) и пертактина 69 кДа.
Разделение компонентов бульона
Компоненты бульона (69 кДа, РТ и FHA) разделяли и очищали при помощи хроматографии на перлите и селективных стадий элюции, по существу, как описано в Патенте ЕР No. 336736 и опубликованной заявителями заявке РСТ No. WO 91/15505, описанной выше. Конкретные выполняемые операции очистки описаны ниже.
Коклюшный токсин (РТ)
Колонку перлита промывали 50 мМ Трис-, 50 мМ Трис/0,5% Тритон Х-100- и 50 мМ Трис-буферами. Фракцию РТ элюировали из перлитной колонки 50 мМ Трис/0,12 М NaCl-буфером.
Фракцию РТ из хроматографии на перлите наносили на гидроксилапатитную колонку и затем промывали содержащим 30 мМ фосфат калия буфером. РТ элюировали буфером 75 мМ фосфат калия/225 мМ NaCl. Колонку промывали смесью 200 мМ фосфат калия/0,6 М NaCl с получением фракции FHA, которую отбрасывали. К очищенному РТ добавляли глицерин до 50% и смесь хранили при 2-8oС до детоксикации в пределах одной недели.
Нитевидный гемагглютинин (FHA)
Фракцию FHA элюировали из перлитной колонки раствором 50 мМ Трис/0,6 М NaCl. Нитевидный гемагглютинин очищали хроматографией на гидроксилапатите. Фракцию FHA из перлитной колонки наносили на гидроксилапатитную колонку, затем промывали 30 мМ фосфатом калия, содержащим 0,5% Тритон Х-100, и затем буфером с 30 мМ фосфатом калия. Фракцию РТ элюировали буфером с 85 мМ фосфатом калия и отбрасывали. Затем фракцию FHA элюировали раствором 200 мМ фосфат калия/0,6 М NaCl и хранили при 2-8oС до детоксикации в пределах одной недели.
69 кДа (пертактин)
Концентрат бульона разбавляли водой для инъекции (WFI) с получением электропроводности 3-4 мС/см и нагружали на перлитную колонку при нагрузке 0,5-3,5 мг белка на 1 мл перлита. Проходящую через колонку фракцию (фракцию компонента 69 кДа) концентрировали ультрафильтрацией при помощи мембраны NMWL 10-30 кДа. К проходящему через колонку концентрату добавляли сульфат аммония до 35±3% (м/об) и полученную смесь выдерживали при 2-8oС в течение 4±2 дней или центрифугировали (7000•g) немедленно. Избыточный супернатант декантировали и осадок собирали центрифугированием (7000•g). Осадок 69 кДа либо хранили замороженным при -20 - -30oС, либо растворяли в Трис-буфере или фосфатном буфере и использовали сразу.
Белок наружной мембраны 69 кДа, полученный осаждением 35% (м/об) сульфатом аммония концентрированного проходящего через перлитную колонку раствора, использовали для очистки. Сульфат аммония (100±5 г на литр) добавляли к фракции 69 кДа и смесь перемешивали в течение, по меньшей мере, 2 часов при 2-8oС. Смесь центрифугировали (7000•g) для извлечения супернатанта. Сульфат аммония (100-150 г на литр) добавляли к этому супернатанту и смесь перемешивали в течение, по меньшей мере, 2 часов при 2-8oС. Смесь центрифугировали (7000•g) для извлечения осадка, который растворяли в 10 мМ Трис-HCl, рН 8. Ионную силу раствора доводили до эквивалента 10 мМ Трис-НСl (рН 8), содержащего 15 мМ сульфат аммония.
Белок 69 кДа наносили на гидроксилапатитную колонку, соединенную тандемно с колонкой Q-Sepharose. Белок 69 кДа собирали в проходящем через колонку растворе, вымывали из этих колонок 10 мМ Трис-HCl (рН 8), содержащим 15 мМ сульфат аммония, и объединяли с белком 69 кДа в проходящем через колонки растворе. Пул белка 69 кДа диафильтровали с 6-10 объемами 10 мМ фосфата калия (рН 8), содержащего 0,15 М NaCl на мембране NMWL 100-300 кДа. Ультрафильтрат собирали и белок 69 кДа в ультрафильтрате концентрировали.
Белок 69 кДа подвергали смене растворителя на 10 мМ Трис-HCl (рН 8) и адсорбировали на Q-Sepharose, промытой раствором 10 мМ Трис-HCl (рН 8)/5 мМ сульфат аммония. Белок 69 кДа элюировали 50 мМ фосфатом калия (рН 8). Белок 69 кДа диафильтровали 6-10 объемами 10 мМ фосфата калия (рН 8), содержащего 0,15 М NaCl, на мембране NMWL 10-30 кДа. Белок 69 кДа стерильно фильтровали через фильтр менее или равный 0,22 мкм. Этот стерильный материал хранили при 2-8oС и адсорбцию проводили в пределах трех месяцев.
Фимбриальные агглютиногены
Агглютиногены очищали из клеточной пасты после отделения от бульона. Клеточную пасту разбавляли до фракции 0,05 объема клеток в буфере, содержащем 10 мМ фосфат калия, 150 мМ NaCl и 4 М мочевину, и смешивали в течение 30 минут. Клеточный лизат осветляли центрифугированием (12000•g), затем концентрировали и диафильтровали против раствора 10 мМ фосфат калия/150 мМ NaCl/0,1% Тритон Х-100 с применением мембранного фильтра NMWL 100-300 кДа.
Концентрат обрабатывали нагреванием при 80oС в течение 30 минут, затем осветляли повторно центрифугированием
(9000•g). ПЭГ 8000 добавляли к осветленному супернатанту до конечной концентрации 4,5±0,2% и перемешивали осторожно минимально в течение 30 минут. Полученный осадок собирали центрифугированием (17000•g) и осадок экстрагировали буфером 10 мМ фосфат калия/150 мМ NaCl с получением раствора неочищенных фимбриальных агглютиногенов. Фимбриальные агглютиногены очищали пропусканием через диоксид кремния PEI. Неочищенный раствор делали 100 мМ в отношении фосфата калия при помощи 1 М фосфата калия (буфера) и пропускали через колонку диоксида кремния PEI.
Проходящую фракцию из колонок концентрировали и диафильтровали против буфера 10 мМ фосфат калия/150 мМ NaCl с применением мембранного фильтра NMWL 100-300 кДа. Этот стерильный материал хранили при 2-8oС и адсорбцию проводили в пределах трех месяцев. Препарат фимбриальных агглютиногенов содержал фимбриальный Аgg 2 и фимбриальный Аgg 3 в весовом соотношении от примерно 1,5 до примерно 2:1 и было обнаружено, что он, по существу, не содержит Аgg 1.
Пример 3.
Этот пример описывает токсоидирование (превращение в токсоид) очищенных антигенов Bordetella pertussis, PT и FHA.
РТ, полученный в чистом виде, как описано в примере 2, токсоидировали доведением концентрации глутарового альдегида в растворе РТ до 0,5±0,1% и инкубированием при 37±3oС в течение 4 часов. Реакцию останавливали добавлением L-аспартата до 0,21±0,02 М. Затем эту смесь выдерживали при комнатной температуре в течение 1±0,1 часа и затем при 2-8oС в течение 1-7 дней.
Полученную смесь диафильтровали против буфера 10 мМ фосфат калия/0,15 М NaCl/5% глицерин на мембранном фильтре NMWL 30 кДа и затем стерилизовали пропусканием через мембранный фильтр, менее или равный 0,22 мкм. Этот стерильный материал хранили при 2-8oС и адсорбцию проводили в пределах трех месяцев.
Фракцию FHA, полученную в чистом виде, как описано в примере 2, токсоидировали доведением концентраций L-лизина и формальдегида до 47±5 мМ и 0,24±0,05% соответственно и инкубированием при 35-38oС в течение 6 недель. Затем эту смесь диафильтровали против буфера 10 мМ фосфат калия/0,5 М NaCl с применением мембранного фильтра NMWL 30 кДа и стерилизовали пропусканием через мембранный фильтр. Этот стерильный материал хранили при 2-8oС и адсорбцию проводили в пределах трех месяцев.
Пример 4.
Этот пример описывает адсорбцию очищенных антигенов Воrdetella pertussis.
Для индивидуальной адсорбции РТ, FHA, Аgg и 69 кДа на фосфате алюминия (квасцах) готовили исходный раствор фосфата алюминия в концентрации 18,75 ± 1 мг/мл. Готовили подходящий сосуд и любой один из антигенов асептически помещали в этот сосуд. Асептически добавляли 2-феноксиэтанол с получением конечной концентрации 0,6±0,1% (об/об) и перемешивали до гомогенности. В сосуд асептически добавляли подходящий объем фосфата алюминия. Добавляли подходящий объем стерильной дистиллированной воды до конечной концентрации фосфата алюминия 3 мг на 1 мл. Контейнеры закрывали герметично, метили и давали им перемешиваться при комнатной температуре в течение 4 дней. Затем сосуд хранили до конечной стадии приготовления композиции.
Пример 5.
Этот пример описывает приготовление компонентной коклюшной вакцины, объединенной с дифтерийным и столбнячным токсоидами.
Антигены В. pertussis, полученные, как описано в предыдущих примерах, готовили с дифтерийным и столбнячным токсоидами для обеспечения нескольких компонентных коклюшных (СР) вакцин, как описано ниже.
Компоненты коклюша получали из Bordetella pertussis, выращенной в погруженной культуре, как описано подробно в примерах 1-4 выше. После завершения роста культуральный бульон и бактериальные клетки разделяли центрифугированием. Каждый антиген очищали индивидуально. Коклюшный токсин (РТ) и нитевидный гемагглютинин (FHA) очищали из бульона последовательной хроматографией через перлит и гидроксилапатит. РТ детоксифицировали глутаровым альдегидом, а оставшийся РТ (примерно 1%), присутствующий во фракции FHA, детоксифицировали формальдегидом. Фимбриальные агглютиногены (2+3) (Аgg) получали из бактериальных клеток. Клетки разрушали мочевиной и обрабатывали нагреванием, фимбриальные агглютиногены очищали осаждением полиэтиленгликолем и хроматографией через полиэтиленимин-диоксид кремния. Компонент белок 69 кДа (пертактин) выделяли из проходящей через колонку фракции из стадии перлитной хроматографии (пример 2) осаждением сульфатом аммония и очищали последовательной хроматографией через гидроксилапатит и Q-Sepharose. Все компоненты стерилизовали фильтрованием через мембранный фильтр 0,22 мкм.
Дифтерийный токсоид получали из Corynebacterium diphtheriae, выращеной в погруженной культуре, стандартными способами. Получение дифтерийного токсоида разделено на пять стадий, а именно поддержание рабочего посевного материала, выращивание Corynebacterium diphtheriae, сбор дифтерийного токсина, детоксикация дифтерийного токсина и концентрирование дифтерийного токсоида.
Получение дифтерийного токсоида
(I) Рабочий посевной материал
Штамм Corynebacterium diphtheriae поддерживали в виде лиофилизированной партии посевного материала. Реконституированный (воссозданный) посевной материал выращивали на косяках Loeffler в течение 18-24 часов при 35±2oС и затем переносили в колбы с "дифтерийной" средой. Затем культуру тестировали на чистоту и содержание Lf. Оставшийся посевной материал использовали для инокулирования ферментера.
(II) Рост Corynebacterium diphtheriae
Культуру инкубировали при 35±2oС и перемешивали в ферментере. К культуре добавляли заранее определенные количества растворов сульфата железа (II), хлорида кальция и фосфата. Фактические количества каждого раствора (фосфата, сульфата железа (II), хлорида кальция) определяли экспериментально для каждой партии среды. Уровни выбирали такие, которые давали наивысшее Lf-содержание. В конце цикла роста (30-50 часов) из культур брали пробы на чистоту и Lf-содержание.
Доводили рН бикарбонатом натрия и культуру инактивировали 0,4% толуолом в течение 1 часа при поддержании температуры 35±2oС. Затем выполняли тест на стерильность для подтверждения отсутствия живой Corynebacteriun diphtheriae.
(III) Сбор дифтерийного токсина
Обработанные толуолом культуры из одного или нескольких ферментеров объединяли в большом чане. Добавляли приблизительно 0,12% бикарбонат натрия, 0,25% уголь и 23% сульфат аммония и определяли рН.
Эту смесь перемешивали в течение примерно 30 минут. Добавляли диатомовую землю и смесь накачивали в глубинный фильтр. Фильтрат рециркулировали до прозрачности, затем собирали и брали пробы для тестирования содержания Lf. К фильтрату добавляли дополнительно сульфат аммония до концентрации 40%. Добавляли также диатомовую землю. Эту смесь выдерживали в течение 3-4 дней при 2-8oС для оседания осадка. Осажденный токсин собирали и растворяли в 0,9% солевом растворе. Диатомовую землю удаляли фильтрованием и токсин диализовали против 0,9% солевого раствора для удаления сульфата аммония. Диализованный токсин объединяли и брали пробы для тестирования содержания Lf и чистоты.
(IV) Детоксикация дифтерийного токсина
Детоксикация имеет место сразу же после диализа. Для детоксикации токсин разбавляли таким образом, что конечный раствор содержал:
a) дифтерийный токсин при 1000±10% Lf/мл,
b) 0,5% бикарбонат натрия,
c) 0,5% формалин,
d) 0,9% (м/об) моногидрохлорид L-лизина.
Раствор доводили до объема солевым раствором и рН доводили до 7,6 ± 0,1.
Токсоид фильтровали через фильтровальные подушки из целлюлозы-диатомовой земли и/или мембранный предварительный фильтр и мембранный фильтр 0,2 мкм в сосуд для сбора и инкубировали в течение 5-7 недель при 34oС. Брали пробу для тестирования на токсичность.
(V) Концентрирование очищенного токсоида
Токсоиды объединяли, затем концентрировали ультрафильтрованием и собирали в подходящий контейнер. Брали пробы для тестирования содержания Lf и чистоты. Добавляли консервант (2-феноксиэтанол) с получением конечной концентрации 0,375% и рН доводили до 6,6-7,6.
Токсоид стерилизовали фильтрованием через предварительный фильтр и мембранный фильтр 0,2 мкм (или его эквивалент) и собирали. Затем брали пробу стерильного токсоида для тестирования необратимости содержания Lf токсоида, содержания консерванта, чистоты (содержания азота), стерильности и токсичности. Стерильный концентрированный токсоид хранили при 2-8oС до конечной стадии приготовления вакцины.
Получение столбнячного токсоида
Столбнячный токсоид (Т) получали из Clostridium tetani, росшего в погруженной культуре.
Получение столбнячного токсоида может быть разделено на пять стадий, а именно поддержание рабочего посевного матриала, рост Clostridium tetani, сбор столбнячного токсина, детоксикация столбнячного токсина и очистку столбнячного токсоида.
(I) Рабочий посевной материал
Штамм Clostridium tetani, используемый в получении столбнячного токсина для превращения в столбнячный токсоид, поддерживали в лиофилизированной форме в партии посевного материала. Этот посевной материал инокулировали в тиогликолатную среду и давали расти в течение приблизительно 24 часов при 35±2oС. Брали пробу для тестирования чистоты культуры.
(II) Рост Clostridium tetani
"Столбнячную" среду заливали в ферментер, обрабатывали нагреванием и охлаждали. Затем ферментер засевали и культуре давали расти в течение 4-9 дней при 34±2oС. Брали пробу для тестирования чистоты культуры и содержания Lf.
(Ill) Сбор столбнячного токсина
Токсин отделяли фильтрованием через фильтровальные подушки из целлюлозы-диатомовой земли и затем осветленный токсин стерилизовали с применением мембранных фильтров. Брали пробы для тестирования содержания Lf и стерильности. Токсин концентрировали ультрафильтрованием с применением размера пор 30000 дальтон.
(IV) Детоксикация столбнячного токсина
Брали пробы токсина для тестирования содержания Lf перед детоксикацией. Концентрированный токсин (475-525 Lf/мл) детоксифицировали добавлением 0,5% (м/об) бикарбоната натрия, 0,3% (об/об) формалина и 0,9% (м/об) моногидрохлорида L-лизина и доводили до требуемого объема солевым раствором. рН доводили до 7,5±0,1 и смесь инкубировали при 37oС в течение 20-30 дней. Брали пробы для тестирования стерильности и токсичности.
(V) Очистка токсоида
Концентрированный токсоид стерилизовали через предварительные фильтры, затем при помощи мембранных фильтров. Брали пробы для тестирования стерильности и содержания Lf.
Оптимальную концентрацию сульфата аммония определяли на основании S-кривой фракционирования, определенной из проб токсоида. Первую концентрацию добавляли к токсоиду (разбавленному до 1900-2100 Lf/мл). Эту смесь выдерживали в течение, по меньшей мере, 1 часа при 20-25oС и супернатант собирали, а осадок, содержащий высокомолекулярную фракцию, отбрасывали.
Вторую концентрацию сульфата аммония добавляли к супернатанту при втором фракционировании для удаления низкомолекулярных примесей. Эту смесь выдерживали в течение, по меньшей мере, 2 часов при 20-25oС и затем могли выдерживать при 2-8oС в течение максимально трех дней. Осадок, который представляет собой очищенный токсоид, собирали центрифугированием и фильтрованием.
Сульфат аммония удаляли из очищенного токсоида диафильтрованием с применением ультрафильтрационных мембран Amicon (или равноценных) с PBS до тех пор, пока не могли больше детектировать сульфат аммония в растворе токсоида. рН доводили до 6,6-7,6 и добавляли 2-феноксиэтанол до конечной концентрации 0,375%. Токсоид стерилизовали мембранным фильтрованием и брали пробы для тестирования (необратимости токсоида, содержания Lf, рН, содержания консерванта, чистоты, стерильности и токсичности).
Приготовление поливалентной вакцины
Одна композиция компонентной коклюшной вакцины, объединенной с дифтерийным и столбнячным токсоидами, была названа CP10/5/5/3DT (иногда называемая CLASSIC). Каждая доза 0,5 мл СР10/5/5/3DТ была приготовлена таким образом, что она содержит:
10 мкг коклюшного токсоида (РТ),
5 мкг нитевидного гемагглютинина (FHA),
5 мкг фимбриальных агглютиногенов 2 и 3 (FIMB),
3 мкг белка 69 кДа наружной мембраны,
15 Lf дифтерийного токсоида,
5 Lf столбнячного токсоида,
1,5 мг фосфата алюминия,
0,6% 2-феноксиэтанол в качестве консерванта.
Другая композиция компонентной коклюшной вакцины, объединенной с дифтерийным и столбнячным токсоидами, была названа CP10/5/5DT (иногда называемая HYBRID). Каждая доза 0,5 мл СР10/5/5DТ была приготовлена таким образом, что она содержит:
10 мкг коклюшного токсоида (РТ),
5 мкг нитевидного гемагглютинина (FHA),
5 мкг фимбриальных агглютиногенов 2 и 3 (FIMB),
15 Lf дифтерийного токсоида,
5 Lf столбнячного токсоида,
1,5 мг фосфата алюминия,
0,6% 2-феноксиэтанол в качестве консерванта.
Другая композиция компонентной коклюшной вакцины, объединенной с дифтерийным и столбнячным токсоидами, была названа СР20/ 20/5 /3DТ. Каждая доза 0,5 мл СР20/20/5/3DТ была приготовлена таким образом, что она содержит:
20 мкг коклюшного токсоида (РТ),
20 мкг нитевидного гемагглютинина (FHA),
5 мкг фимбриальных агглютиногенов 2 и 3 (FIMB),
3 мкг белка 69 кДа наружной мембраны,
15 Lf дифтерийного токсоида,
5 Lf столбнячного токсоида,
1,5 мг фосфата алюминия,
0,6% 2-феноксиэтанол в качестве консерванта.
Другая композиция компонентной коклюшной вакцины, объединенной с дифтерийным и столбнячным токсоидами, была названа СР20/ 10/10 /6DТ. Каждая доза 0,5 мл СР20/ 10/ 10/ 6DТ была приготовлена таким образом, что она содержит:
20 мкг коклюшного токсоида (РТ),
10 мкг нитевидного гемагглютинина (FHA),
10 мкг фимбриальных агглютиногенов 2 и 3 (FIMB),
6 мкг белка 69 кДа наружной мембраны,
15 Lf дифтерийного токсоида,
5 Lf столбнячного токсоида,
1,5 мг фосфата алюминия,
0,6% 2-феноксиэтанол в качестве консерванта.
Пример 6.
Этот пример описывает клиническую оценку компонентных бесклеточных коклюшных вакцин.
(а) Исследования на взрослых
Исследования на взрослых и детях в возрасте 16-20 месяцев показали, что поливалентные вакцины, содержащие фимбриальные агглютиногены, являются безопасными и иммуногенными (таблица 2).
Клиническое исследование Фазы I проводили на детях в возрасте 17-18 месяцев в Калгари, Альберте с пятикомпонентной коклюшной вакциной (СР10/5/5/3DТ) и оно показало неблагоприятную реакцию. Тридцать три ребенка получали эту вакцину и еще 35 получали ту же самую вакцину без белкового компонента 69 кДа.
Местные реакции были редкими. Системные побочные (вредные) реакции, первично состоящие из раздражимости, присутствовали приблизительно у половины участников исследования, независимо от того, какую вакцину вводили. Значимые увеличения антител измеряли для антител IgG против РТ, против FHA, против фимбриальных агглютиногенов и против 69 кДа посредством иммуноферментного анализа и антител против РТ в тесте нейтрализации на клетках СНО. Не обнаруживали различий в ответе антител у детей, получавших четырехкомпонентную вакцину СР10/5/5DT или пятикомпонентную вакцину CP10/5/5/3DT, за исключением антител против 69 кДа. Дети, которые получали пятикомпонентную вакцину, содержащую белок 69 кДа, имели значимо более высокий уровень антител против 69 кДа после иммунизации.
Исследование зависимости ответа от дозы было предпринято с 4-компонентной вакциной в Виннипеге и Манитоба (Канада). Использовали две композиции компонентных вакцин: CP10/5/5/3DT и CP20/ 10/ 10/6DT. В качестве контроля включали также цельноклеточную вакцину.
Это исследование было дважды слепым исследованием на 91 младенце в возрасте 17-18 месяцев в момент их бустер-дозы коклюшной вакцины. Как CP10/5/5/3DT, так и СР20 /10/ 10/6DТ хорошо переносились этими детьми. Не обнаружили различий у ряда детей, которые имели какую-либо местную реакцию или системные реакции после любой из этих компонентных вакцин. В противоположность этому, значимо больше детей, которые получали цельноклеточную вакцину, имели местные и системные реакции, чем среди детей, которые получали вакцины CP20 /10/ 10/6DT.
(b) Исследования на младенцах
Фаза II:
Исследование проводили с использованием вакцины CP10/5/5/3DT в Калгари, Альберте и Британской Колумбии в Канаде. В этом исследовании 432 младенца в возрасте 2, 4 и 6 месяцев получали компонентную коклюшную вакцину или цельноклеточную/контрольную вакцину DPT. Вакцина СР10/5/5/3DТ хорошо переносилась этими младенцами. Местные реакции были менее распространенными в случае компонентной вакцины, чем в случае цельноклеточной вакцины, после каждой дозы.
Значимый ответ в виде образования антител на все антигены был показан после вакцинации компонентной коклюшной вакциной. Реципиенты цельноклеточной вакцины имели сильный ответ антител на фимбриальные агглютиногены D и Т. При семи месяцах 82-89% реципиентов компонентной вакцины и 92% реципиентов цельноклеточной вакцины имели 4-кратное увеличение или еще большее повышение титра антител к фимбриальным агглютиногенам. В противоположность этому, ответ антител на FHA был 75-78% в случае компонентных вакцин и 31% в случае реципиентов цельноклеточной вакцины, 4-кратное увеличение антител против 69 кДа было отмечено в 90-93% реципиентов компонентной вакцины и 75% реципиентов цельноклеточной вакцины. 4-кратное повышение антител против РТ наблюдали иммуноферментным анализом в 40-49% реципиентов компонентной вакцины и 32% реципиентов цельноклеточной вакцины; 4-кратное повышение антител к РТ согласно СНО-нейтрализации было обнаружено в 55-69% реципиентов компонентной вакцины и 6% реципиентов цельноклеточной вакцины (таблица 2).
Фаза IIB:
Вакцины СР20/20/5/3DТ и CP10/10/5/3DT оценивали на 100 младенцах в возрасте 2, 4 и 6 месяцев в рандомизированном слепом исследовании против контроля D15PT с более низким содержанием дифтерийного антигена 15 Lf по сравнению с композицией 25 Lf. Не обнаружили различий между этими двумя компонентными композициями; обе были значимо менее реактогенными, чем цельноклеточный контроль. Более высокие титры антител против РТ как в иммуноферментном анализе, так и в СНО-нейтрализации и против FHA были достигнуты у реципиентов вакцины СР20/20/5/3DТ с увеличенным содержанием антигена. При 7 месяцах среднее геометрическое значение титра антител против FHA было 95,0 у реципиентов СР20/20/5/3DТ, 45,2 у реципиентов СР10/5/5/3DТ и было только 8,9 у реципиентов D15PT. Титры антител против РТ были 133,3, 58,4 и 10,4 в иммуноанализе и 82,4, 32,7 и 4,0 согласно СНО-нейтрализации соответственно (таблица 2).
Это исследование показало, что компонентная коклюшная вакцина, объединенная с дифтерийным и столбнячным токсоидами, с увеличенным содержанием антигенов, была безопасной и иммуногенной для младенцев и что увеличенное содержание антигенов увеличило иммунный ответ на полученные антигены (РТ и FHA) без увеличения реактогенности.
NIAID, ФАЗА II, США, Сравнительное испытание:
Исследование Фазы II выполняли в Соединенных Штатах при содействии Национального Института Аллергии и Инфекционных Заболеваний (NIAID) в качестве вступления к широкомасштабному испытанию эффективности бесклеточных коклюшных вакцин. Одна компонентная коклюшная вакцина данного изобретения в сочетании с адсорбированными дифтерийным и столбнячным токсоидами (CP10/5/5/3DT) была включена в это испытание вместе с 12 другими бесклеточными вакцинами и двумя цельноклеточными вакцинами. Результаты по безопасности сообщались для 137 детей, иммунизированных в возрасте 2, 4 и 6 месяцев компонентной вакциной CP10/5/5/3DT.
Как видно в прежних исследованиях, было обнаружено, что компонентная вакцина является безопасной, имеет низкую реактогенность и хорошо переносится реципиентами.
В возрасте 7 месяцев уровни всех антител, антител против РТ, антител против FHA, антител против 69 кДа и антител против фимбриальных агглютиногенов, были более высокими, чем уровни, достигаемые после цельноклеточных вакцин (ссылка 71 и таблица 2), или равноценные им. Проводили дважды слепое исследование, в котором детей рандомизированно распределяли для получения вакцинной композиции либо CP20/20/5/3DT, либо CP10/5/5/3DT. Всего 2050 младенцев были зарегистрированы в Соединенных Штатах и Канаде; 1961 младенец завершил это исследованте. Иммуногенность оценивали в подгруппе из 292 субъектов. Повышение антител индуцировалось всеми антигенами, содержащимися в вакцине, в случае обеих вакцинных композиций. Композиция СР20/20/5/3DТ индуцировала более высокие титры антител против FHA, но не против РТ. Композиция СР10/5/5/3DТ индуцировала более высокие титры против фимбрий и более высокие титры против агглютиногенов.
Дальнейшее исследование безопасности и иммуногенности проводили во Франции. Схема исследования сходна с Североамериканским исследованием, описанным выше, за исключением того, что вакцины вводили в возрасте 2, 3 и 4 месяца. Местные и системные реакции были обычно небольшими. В целом, эта вакцина хорошо воспринималась участниками французского исследования с применением этого режима введения.
Испытание эффективности с плацебо в качестве контроля двух бесклеточных коклюшных вакцин и цельноклеточной вакцины на 10000 младенцев
После результатов сравнительного испытания NIAID Фазы II, США, были выбраны двухкомпонентная и пятикомпонентная бесклеточные вакцины для многоцентрового, контролируемого, дважды рандомизированного, контролируемого плацебо испытания эффективности. Это клиническое испытание проводили в Швеции, где существует высокая частота заболеваний коклюшем. Двухкомпонентная вакцина содержала инактивированный глицеральдегидом и формалином РТ (25 мкг), обработанный формалином FHA (25 мкг) и дифтерийный токсоид (17 Lf), и столбнячный токсоид (10 Lf). Пятикомпонентная коклюшная вакцина была CP10/5/5/3DT. Для этого испытания были зарегистрированы 10000 младенцев, представляющих 1/2 младенцев этой возрастной группы в Швеции, в 14 географически определенных местах исследования согласно регистрации рождения.
Дети, родившиеся в январе и феврале 1992 года, были рандомизированы в 3-групповом испытании. После получения согласия родителей 2/3 младенцев получали один из двух дифтерийно-столбнячных бесклеточных коклюшных препаратов в возрасте 2, 4 и 6 месяцев. Контрольная группа получала только DT. В мае 1992 года лицензированную в США коммерчески доступную цельноклеточную DPT (КДС) вакцину вводили детям, родившимся в марте-декабре 1992 года, рандомизированным в 4-групповое испытание. После согласия родителей 3/4 этих младенцев получали один из трех препаратов DTP в возрасте 2, 4 и 6 месяцев. Контрольная группа получала только DT (ДС).
Каждую вакцину вводили приблизительно 2500 детям. Вакцины вводили в виде трех доз. Первую дозу вводили в возрасте 2 месяцев и не позднее возраста 3 месяцев. Последующие дозы вводили с интервалами 8 недель. Вакцины вводили внутримышечной инъекцией.
Эти дети и их домашние наблюдались в течение 30 месяцев. Если было подозрение на коклюш, собирали клинические данные и старались получить лабораторную проверку при помощи назальных аспиратов бактериологической культуры и диагностики с применением полимеразной цепной реакции (ПЦР). Пробы крови острых случаев и выздоравливающих детей брали для серологической диагностики.
До этого исследования уровень пертактина, допустимого в компонентных коклюшных вакцинах данного изобретения в группе риска людей (в частности, новорожденных), был неизвестен. В частности, вклад различных компонентов Bordetella и их присутствия в коклюшных вакцинах в выбранных относительных количествах на эффективность вакцин был неизвестен.
Основная цель этого испытания заключалась в оценке способности бесклеточных коклюшных вакцин и цельноклеточной вакцины защищать против типичного коклюша в сравнении с плацебо.
Второй конечной целью было исследование эффективности вакцины против установленной коклюшной инфекции различной степени тяжести.
Эффективность вакцины определяется как снижение (в %) вероятности заражения коклюшем среди реципиентов вакцины относительно невакцинированных детей.
Относительный риск коклюша в двух вакцинных группах выражается в виде отношения вероятности заболевания в этих двух группах.
Вероятность заражения коклюшем, называемая также скоростью атаки, может быть определена различными путями. В расчете объема выборки, вероятность заражения коклюшем в конкретной группе исследования оценивали отношением между числом детей с коклюшем и детей, остающихся в этой группе исследования при завершении катамнестического исследования.
Эффективность компонентной вакцины CP10/5/5/3DT в этом испытании показана в таблице 4 и она была примерно 85%. В том же самом испытании двухкомпонентная коклюшная бесклеточная вакцина, содержащая только РТ и FHA, имела эффективность примерно 58%, а цельноклеточная вакцина имела эффективность примерно 48%. СР10/5/5/3DТ была также эффективна в предотвращении легкого коклюша с оцененной эффективностью примерно 77%.
Пример 7.
Этот пример описывает приготовление и иммуногенность поливалентной комбинированной вакцины, содержащей капсульный полисахарид Haemophllus influenzae.
(а) Получение компонента PRP-T
Капсульный полисахарид (PRP) из Н. influenzae очищали и конъюгировали со столбнячным токсоидом следующим образом. Из лиофилизированных ампул партии рабочего посевного материала Н. influenzae получали три последовательные предварительные культуры. Первая предварительная культура была на твердой среде. Ампулы инокулировали на чашки с агаром с угольной пылью + прокипяченной кровью (10% лошадиной крови, нагреваемой в течение 15 минут при 80oС) и инкубировали в течение 20±4 часов при 36-37oС в атмосфере СO2. Вторая предварительная культура была в жидкой среде в течение 8 часов при 37oС. Жидкая среда имела следующий состав на литр:
1. Казаминокислоты Difco, г - 10
Мононатрийфосфат•2H2O, г - 2,03
Динатрийфосфат•12Н2O, г - 31,14
Лактат натрия (60% раствор), мл - 1,5
L-Цистин, г - 0,07
L-Триптофан, г - 0,02
СаСl2•2Н2O, г - 0,02
(NH4)2SO4, г - 1
MgSO4•7H2O, г - 0,4
Антивспениватель Antifoam Dow Corning M.S.A. при 25% в парафиновом масле - 0,15 мл
2. Ультрафильтрат гемина + декстроза в соотношении 20 г декстрозы и 1 мг гемина. К этому раствору добавляют 5 мг никотинамидадениндинуклеотида. Стерилизуют при помощи фильтра.
3. Дрожжевой экстракт Difco - 5 г
Стерилизуют при помощи фильтра.
Третья предварительная культура была в жидкой среде, перемешиваемой в течение 4 часов при 37oС. Третью предварительную культуру использовали для инокуляции ферментера и эту культуру поддерживали при перемешивании при 37oС в течение 12-14 часов. Культуру собирали в охлаждаемом чане. Добавляли формалин в концентрации 10 мл/литр. Культуру поддерживали при осторожном перемешивании при 4oС в течение 2-24 часов и затем центрифугировали. Добавленный формалин был предназначен не для полной инактивации бактерий, а для остановки роста и ингибирования метаболизма. Это добавление уменьшало лизис клеток и сопутствующее ему загрязнение внутриклеточными компонентами. Продолжительность этой фиксации была между 2 и 24 часами и, как правило, эту культуру оставляли на ночь перед центрифугированием. Супернатант, содержащий полисахарид, собирали, а осадок бактерий отбрасывали. Процесс очистки обычно проводили в холодной комнате или в таких условиях, при которых температура продуктов и реагентов ниже или равна 10oС, за исключением стадии фенольной очистки, которую проводили при комнатной температуре. После центрифугирования культуры супернатант этой культуры концентрировали. Капсульный полисахарид осаждали из полученного концентрата добавлением центримида (centrimide) с получением конечной концентрации 5% (м/об). Центримид осаждал PS из концентрированной жидкости (SNF). Также соосаждались некоторые количества белка, нуклеиновой кислоты и липополисахарида (LPS). Осадки собирали центрифугированием, причем некоторые другие примеси и некоторое количество белка оставались в SNF. Полученный осадок собирали центрифугированием и хранили при температуре, менее или равной -20oС.
Осадки ресуспендировали в 0,3 М растворе NaCl и суспензию опять центрифугировали. NaCl избирательно диссоциировал комплексы полисахарид-центримид. Некоторые примеси (нуклеиновая кислота, LPS, белок) также диссоциировали в этом процессе. К супернатанту добавляли предварительно охлажденный абсолютный этанол до конечной концентрации 60%. Полученный осадок собирали центрифугированием и промывали холодным абсолютным этанолом. Осадок сушили в вакууме при 0-4oС, и он представлял собой промежуточный продукт. Промежуточный продукт растворяли в натрий-ацетатном буфере и экстрагировали фенолом при комнатной температуре. Водную фазу собирали продолжительным центрифугированием. Фенольная экстракция и центрифугирование могут повторяться несколько раз и водную фазу диализуют и диафильтруют. Капсульный полисахарид из диафильтрованного раствора осаждали добавлением предварительно охлажденного этано-ла до конечной концентрации 60% в присутствии 0,3 М NaCl. Осадок собирали центрифугированием, промывали предварительно охлажденным абсолютным этанолом, ацетоном и эфиром и сушили под вакуумом при 4oС. Затем высушенный осадок растирали до тонкоизмельченного порошка при низкой влажности и этот продукт представлял собой очищенный полисахарид Haemophilus influenzae типа b.
Очищенный полисахарид растворяли в воде для получения 5 мг полисахарида на 1 мл раствора и рН доводили до 10,8±0,2 при помощи NaOH. Добавляли цианогенбромид в виде раствора в воде в соотношении 0,5 мг CNBr/мг полисахарида. рН реакционной смеси поддерживали NaOH при 10,8±0,2 в течение 35-40 минут при 23±3oС. рН снижали до рН 9 добавлением НСl. Дигидразид адипиновой кислоты добавляли до конечной концентрации 3,5 мг ADH/мг полисахарида и рН доводили до 8,5. Реакционную смесь инкубировали при 23±3oС в течение 15 минут (рН поддерживали при 8,5) и затем этот раствор инкубировали в течение ночи при 4oС с осторожным перемешиванием. Реакционную смесь диализовали против раствора NaCl и затем концентрировали. Затем раствор фильтровали через фильтр 0,45 μ и замораживали при температуре, менее или равной -40oС. Это был АН-полисахарид, и его хранили при температуре, менее или равной -40oС.
Для получения компонента столбнячного токсина штамм Clostridium tetani инокулировали в ряд пробирок, содержащих 10 мл среды Rosenow или тиогликолатной среды. Среда Rosenow имеет следующий состав:
Состав (формула) (в граммах на литр дистиллированной воды)
Пептон - 10
Мясной экстракт - 3
Глюкоза - 2
Хлорид натрия - 5
Индикатор "Andrade" (5% Фуксин-кислота), мл - 10
Белый мрамор - 1 кусочек
Мозг - 1 кусочек
Этой средой, приготовленной непосредственно перед использованием из готовых к употреблению продуктов, заполняли пробирки и стерилизовали при 120oС в течение 20 минут.
Бутыль на 5 литров, содержащую 3 литра среды "Massachusetts", инокулировали С. tetani и инкубировали в течение 16-18 часов при 35±1oС. Затем содержимое переносили в бутыль на 20 литров, содержащую 15 литров стерильной среды "Massachusetts", и инкубировали в течение 8 часов при 35±1oС. Каждую бутыль использовали для инокуляции ферментера, содержащего 582 литра среды "Massachusetts", и инкубировали при 35oС в течение 5-6 дней при аэрации. Ферментеры охлаждали и к культуре добавляли 12 кг хлорида натрия, 8 кг тринатрийцитрата. Встряхивание поддерживали в течение 1 дня, затем останавливали и этот процесс позволил экстрагировать оставшийся токсин из бактерий в конце культивирования. Токсин осветляли фильтрованием или пропусканием через центрифугу непрерывного действия.
Супернатант из 1200 л культуры концентрировали ультрафильтрацией и концентрированный токсин диафильтровали против 0,07 М раствора динатрийфосфата, рН 8,2. Конечный объем доводили до 500 Lf/мл.
Двойное осаждение сульфатом аммония проводили для получения очищенного столбнячного токсина. Так, сульфат аммония и 10 г угля медленно добавляли на литр полученного предварительно диафильтрованного токсина. После 16-24 часов инкубации при 4oС токсин фильтровали на картриджах (наборах фильтрующих дисков) для удаления осадка. Затем количество сульфата аммония, достаточное для получения 320 г/л, медленно добавляли на литр предварительно полученного супернатанта. После примерно 48 часов при 4oС осадок собирали центрифугированием и растворяли в 0,05 М растворе динатрийфосфата, рН 8,2. Раствор диафильтровали против 0,05 М раствора динатрийфосфата, рН 8,2, и доводили до 300 Lf/мл. Затем раствор стерилизовали фильтрованием. 7,5 мкмоль (0,225%) формальдегида добавляли на 1 мл раствора токсина. Детоксикация достигалась после инкубации в течение 24 дней при 37oС, включающей в себя промежуточные периоды при 4 и 22oС. Для получения столбнячного токсоида проводили стерилизацию фильтрованием (0,22μ). Столбнячный токсоид диализовали и концентрировали против NaCl с применением мембраны, имеющей отсечение молекулярной массы, менее или равной 50000. Затем концентрированный белок асептически фильтровали и хранили при 4oС.
Равные количества АН-полисахарида и столбнячного токсоида (± 20%) смешивали с 0,05 М NaCl с получением концентрации 7,5 мг полисахарида на 1 мл. рН раствора доводили до 5,7±0,2 при помощи НС1 и добавляли 1-этил-3-(3-диметиламинопропил)карбодиимид (EDAC) с получением конечной концентрации 19,17 мг EDAC/мл реакционной смеси. Карбоксильные группы столбнячного белка активировали связыванием с EDAC. При слабокислых условиях реакции происходит реакция конденсации, в которой AH-PS и EDAC-активированный столбнячный белок становятся ковалентно связанными. Смесь инкубировали при постоянном рН (5,7) в течение 60 минут при 4oС и затем рН доводили до 6,9±0,2 NaOH и реакционную смесь диализовали против NaCl при 4oС. Конъюгат очищали зональным центрифугированием на сахарозном градиенте (4%-60%) для удаления EDAC, свободного АН-полисахарида, свободного столбнячного белка и низкомолекулярного конъюгата. Затем к фракции, содержащей конъюгат полисахарида, добавляли апирогенную воду, сахарозу, Трис-НСl-буфер с получением раствора конъюгата следующего состава: сахароза 8,5 (м/об) ± 0,5%, пoлисахарид в концентрации примерно 200 мкг/мл, Трис-НС1-буфер 10 мМ, рН 7,0±0,5.
Затем этот раствор асептически фильтровали с применением фильтра 0,1μ и хранили при -40oС.
Материал конъюгата полисахарида Haemophilus influenzae типа b разбавляли в стерильных условиях разбавителем для получения конечного состава:
Концентрированный материал конъюгата полисахарида Haemophilus influenzae полисахарида типа b, мг - До 200
Трис-НСl-буфер 200 мМ до рН 7,2, мМ - До 10
Сахароза, г - До 850
Вода для инъекций, л - До 10
Конечный материал помещали во флаконы и лиофилизировали. (Лиофилизированную вакцину реконституировали 0,5 мл или 0,4% NaCl для использования).
(b) Композиция
Испытывали две композиции поливалентной вакцины (APDT) (АКДС). Первая композиция (APDT-низкая) содержала 10 мкг коклюшного токсоида (РТ), 5 мкг нитевидного агглютинина (FHA), 5 мкг фимбриального агглютиногена 2 и 3 мкг белка 69 кДа (69К) на дозу 0,5 мл (CLASSIC). Вторая композиция (APDT-высокая) содержала двойное количество РТ (20 мкг) и идентичные количества FIM и 69К (HYBRID). Обе композиции содержали 15 порогов флокуляции (Limit of flocculation) (Lf) дифтерийного токсоида, 5 Lf столбнячного токсоида, 1,5 мг фосфата алюминия в качестве адъюванта и 0,6% 2-феноксиэтанол в качестве консерванта. Содержащую конъюгат Hib-столбнячный токсоид вакцину (PRP-T) получали описанным выше способом Connaught Laboratories Inc. (Swiftwater, USA).
Популяция
Здоровые 17-21-месячные дети, которые были иммунизированы тремя дозами вакцин APDT-низкая или PRP-T в виде отдельных инъекций в возрасте 2, 4 и 6 месяцев в первичном клиническом испытании, были привлечены для этого исследования. После письменного сообщения родителей о согласии детей распределяли при помощи созданного компьютером сбалансированного блок-списка случайных номеров для получения PRP-T либо в виде отдельной инъекции в тот же самый день, отдельной инъекции PRP-T, вводимой через месяц после вакцины APDT, либо в виде одной инъекции (лиофилизированного PRP-T, реконституированного в APDT). Композиция APDT (высокая или низкая) для каждого ребенка оставалась той же самой, которая давалась ему для первых трех доз (отношение распределения в выборке 6:1 APDT-высокая; APDT-низкая). Вакцины вводили внутримышечно (i.m.) иглой 25 мм в дельтовидную мышцу плеча или в широкую латеральную мышцу бедра, если дельтовидная мышца имела недостаточную массу. Если требовалась вторая инъекция PRP-T, инъекцию выполняли в противоположную конечность.
Клинический и лабораторный мониторинг
Участников подвергали мониторингу на местные и системные неблагоприятные побочные реакции сразу же после иммунизации и (родителями) в течение 72 часов после иммунизации. Данные собирали при помощи спланированных телефонных интервью при 24 и 72 часах. Температуру тела измеряли, по меньшей мере, один раз в день или, когда родителям казалось, что у ребенка повышена температура. Болезненность и системные реакции (раздражимость, пониженная активность, пониженный аппетит) распределялись по следующим градациям: слабая, умеренная или тяжелая в соответствии с предварительно установленными критериями, при помощи которых родитель выбирал тяжесть на основе структурированных примеров. Измеренные местные реакции классифицировались по их размеру и продолжительности плача.
Пробы крови брали посредством венепункции или проколом из пальца до и после 28 дней после иммунизации; у детей, получивших инъекцию PRP-T (следовательно, через 2 месяца после APDT). Антитела к капсульному полисахариду Hib (PRP) измеряли при помощи РИА. Антитела IgG к РТ измеряли при помощи EIA и РТ-нейтрализующие антитела нейтрализацией цитотоксичности клеток яичника китайского хомячка (СНО). IgG-антитела против FHA, против FIM и антитела против 69К измеряли при помощи EIA; количество единиц рассчитывали с применением сравнительной антисыворотки US FDA (#3). Измеряли также коклюшные агглютинины. Дифтерийный антитоксин измеряли в тесте микронейтрализации и столбнячный антитоксин при помощи EIA. Титры антител выражали в виде средних геометрических значений титров; пробы сывороток с титрами менее порога детектирования теста относили к величине 1/2 нижнего порога детектирования для целей статистических расчетов.
Статистический анализ
Побочные реакции анализировали после группирования по клинической значимости. Скорости побочных реакций сравнивали посредством оценок Mantel-Haenszel относительного риска применением центра выборки и вакцинной композиции в качестве расслоенных переменных. В каждом случае оценивали точечные оценки и 95% CI RR. CI, которые не включают 1,00, являются статистически значимыми.
Геометрические средние титров антител и 95% CI (доверительные интервалы) вычисляли для титра антител для каждого вакцинного антигена до и после иммунизации. Средние логарифмические титры сравнивали трехфакторным дисперсионным анализом. Соотношение субъектов, достигающих предварительно заданных уровней в каждой группе, сравнивали при помощи логистической регрессии. Для множественных сравнений не делались никакие корректировки.
В целом, 545 детей (44% женского пола) были зарегистрированы в этом исследовании, 74% из которых завершили серийное исследование. Соотношение участников в этом исследовании, которые получали две композиции, оставалось 6:1 (468 APDT-высокая, 77 APDT-низкая). Средний возраст был 18,9 месяцев (диапазон 17-21 месяцев); все, кроме 3 детей (99,4%), завершили это исследование.
Побочные реакции
Показатели побочных реакций не отличались в группах, иммунизированных APDT-высокой или APDT-низкой вакциной; скорости были также одинаковыми, проводились ли иммунизации при одном визите, при различных визитах или в виде единой комбинированной инъекции.
Гуморальный иммунный ответ (образование антител)
Перед иммунизацией уровни антител против всех антигенов, за исключением FHA, были одинаковыми у детей, которые получали вакцины APDT-высокую или APDT-низкую для их первых трех доз. Дети, иммунизированные APDT-высокой с 4-кратным содержанием в ней FHA, имели значимо более высокие титры FHA, чем дети, иммунизированные APDT-низкой вакциной (р=0,0001). После иммунизации APDT-высокая индуцировала более высокие титры антител, чем APDT-низкая (р= 0,0001). В противоположность этому, предиммунизационные титры антител против РТ, измеренные по нейтрализации СНО или EIA, были одинаковыми в обеих этих группах. Парадоксально, несмотря на вдвое большее содержание антигена, титры антител против РТ были более низкими после иммунизации APDT-высокой, чем APDT-низкой вакциной (р=0,038). Подобным образом титры антител против FIM и агглютинина после иммунизации были более высокими в группе с APDT-низкой вакциной (р=0,01 и р=0,04 соответственно) вопреки идентичным
количествам фимбриального антигена в обеих вакцинных композициях.
Перед иммунизацией были небольшие различия в антителах против антигена коклюша среди группы, рандомизированной для получения PRP-T, комбинированного с APDT в виде одной инъекции, или предоставляемого в виде отдельных инъекций в те же самые или в другие дни. Данные представлены отдельно для реципиентов APDT-высокой или APDT-низкой вакцины; однако, вследствие малого числа детей в группе APDT-низкой вакцины, эти результаты не будут обсуждаться далее. Группа, рандомизированная для получения отдельных инъекций в тот же самый день, имела более высокие титры антител против РТ в тесте СНО-нейтрализации, чем группа, которая получала низкие инъекции в отдельные дни (6,14 против 4,80 единиц; р менее 0,05). Постиммунизационные уровни антител были также более высокими в этой группе (176 единиц), чем в группе с отдельными инъекциями в другие дни (122 единицы; р менее 0,01), хотя аналогично более высокие уровни были обнаружены в группе, получавшей единую комбинированную иммунизацию (171 единица; р менее 0,01). Образование антител против 69К детектировали в этой группе после иммунизации, хотя дети, иммунизированные двумя инъекциями в один и тот же день, имели более высокое образование антител, чем дети, иммунизированные комбинированной единой вакциной, и дети, иммунизированные двумя инъекциями в один и тот же день, имели более высокое образование антител, чем дети, иммунизированные в разные дни (243 против 190 единиц; р менее 0,001), чем дети иммунизированные двумя инъекциями в разные дни.
Уровни антител против PRP были одинаковыми среди трех групп перед иммунизацией. После иммунизации титры были более высокими у детей, иммунизированных отдельными инъекциями в один и тот же день (66,0 мкг/мл), чем у детей, иммунизированных в разные дни (28,4 мкг/мл; р менее 0,001), или у детей, получивших единую комбинированную иммунизацию (47,1; р менее 0,05). Комбинированная иммунизация индуцировала также значимо более высокие уровни антител, чем вакцины, предоставленные в раздельные дни (р менее 0,05). Различий не обнаружили среди групп в процентах, которые достигали "защитных" уровней; все участники имели постиммунизационные титры, превышающие 0,15 мкг/мл и только 4 участника (0,7%) не смогли достичь титра, превышающего 1 мкг/мл (3 в группе, получавшей инъекции в различные дни, и один в группе, получившей единую комбинированную инъекцию). Более 82% детей в каждой группе превышали уровень антител против РТ 10 мкг/мл.
Сильное образование антител индуцировалось также против дифтерийного и столбнячного токсоидов. По сравнению с группой, получавшей иммунизацию в различные дни (2,1 МЕ/мл), значимо более высокие уровни антител против дифтерийного токсоида индуцировались в детях, иммунизированных двумя инъекциями в один и тот же день (3,1 МЕ/мл; р менее 0,01) или комбинированными едиными инъекциями (3,3 МЕ/мл; р менее 0,001). Антитела против столбняка были более высокими в реципиентах с двумя инъекциями в один и тот же день (6,7 МЕ/мл), чем в реципиентах, иммунизированных в различные дни (5,2 МЕ/мл; р менее 0,01), или в детях, получавших комбинированную единую инъекцию (4,8 МЕ/мл; р менее 0,001). Все дети имели после иммунизации титры антител против дифтерии и против столбняка более 0,1 МЕ/мл, уровень, в 10 раз превышающий предполагаемый защитный уровень. Более 96% титров антител против столбняка и более 74% титров антител против дифтерии превышали уровень 1,0 МЕ/мл; среди групп иммунизации не были обнаружены различия.
Пример 8.
Этот пример описывает состав и иммуногенность поливалентной комбинированной вакцины, содержащей инактивированную полиовакцину.
(а) Получение инактивированного полиовируса,
(i) выращенного на клетках MRC-5
Инактивированный полиовирус, росший в клетках MRC-5, получали следующим образом. Клетки были клетками почек зеленой мартышки (Ceropithacus aethiops).
Трехвалентная полиовирусная вакцина, инактивированная, содержала компоненты Типа I (Mahoney), Типа II (MEF) и Типа III (Saukett), которые выращивали на клетках MRC-5 на гранулах-микроносителях, обрабатывали и инактивировали перед комбинированием в трехвалентную полиовирусную вакцину.
Суспензию клеток MRC-5 добавляли к среде для выращивания клеток в ферментер при рН 7,2 (6,9-7,6) и температуре 37±0,5oС. Среда для выращивания клеток имела следующий состав:
Среда CMRL 1969
Бикарбонат натрия,% - 0,15
Сыворотка взрослых коров,% - 5,00-7,00
Сульфат неомицина (мкг активности), МЕ/мл - 10
Полимиксин В, МЕ/мл - 200
Среда CMRL имела следующий состав:
СУХОЙ ПОРОШОК
Ингредиенты:
Аминокислоты - мг/л
L-Аланин - 25,0
L-Аргинин (свободное основание) - 58,0
L-Аспарагиновая кислота - 30,0
L-Цистеин•НСl - 0,1
Динатриевая соль L-цистеина - 24,0
L-Глутаминовая кислота•H2O - 67,0
L-Глутамин - 200,0
L-Глицин - 50,0
L-Гистидин (свободное основание) - 16,2
L-Гидроксипролин - 10,0
L-Изолейцин - 20,0
L-Лейцин - 60,0
L-Лизин•НСl - 70,0
L-Метионин - 15,0
L-Фенилаланин - 25,0
L-Пролин - 40,0
L-Серин - 25,0
L-Треонин - 30,0
L-Триптофан - 10,0
L-Тирозин - 40,0
L-Валин - 25,0
Витамины
п-Аминобензойная кислота - 0,05
Аскорбиновая кислота - 0,05 -Биотин - 1,00
-Биотин - 1,00
Пантотенат кальция - 1,00
Холиндигидрогенцитрат - 2,12
Фолиевая кислота - 1,00
Глутатион - 0,05 -Инозит - 2,00
-Инозит - 2,00
Никотинамид - 1,00
Пиридоксаль•НСl - 1,00
Рибофлавин-5-фосфат - 0,10
Тиамин•НСl - 1,00
Ингредиенты - мг/л
Хлорид натрия - 8000,0
Хлорид калия - 400,0
Хлорид кальция (безводный) - 140,0
Сульфат магния•7H2O - 200,0
Фосфат натрия, двухосновный, безводный - 180,0
Фосфат натрия, неосновный - 70,0
D-Глюкоза (безводная) - 1000,0
Феноловый красный - 20,0
10,852 г дают 1 литр Среды CMRL 1969.
Эту среду готовят следующим образом.
450 литров дистиллированной апирогенной воды добавляют к 905 мл 1 н. хлористоводородной кислоты. К этой смеси добавляют 5426,5 г сухого порошка CMRL 1969 при непрерывном перемешивании до растворения до прозрачного раствора.
Следующие химикалии добавляли в указанном порядке с непрерывным перемешиванием, давая каждому химикалию раствориться перед добавлением следующего:
Неомицин, мкг/мл - 10
Полимиксин В, единиц/мл - 200
Буферный раствор TES ,мл - 5000,0
Бикарбонат натрия, г - 750,0
Бычья сыворотка, л - 30,0
Объем доводят до 500 л свежедистиллированной водой и перемешивают до однородного смешивания.
Рост клеток наблюдают и, когда клетки находятся в фазе логарифмического роста, отработанную среду для выращивания отбрасывают и заменяют средой для роста вируса. Среда для роста вируса имеет следующий состав.
Химикалии Среды 199 с солями Ирла:
Бикарбонат натрия,% - 0,26
Твин 80, м.д. - 20
Сульфат неомицина (мкг активности), МЕ/мл - 10
Полимиксин В, МЕ/мл - 200
L-Глутамин, мг/л - 100
L-Аргинин, мг/л - 29
L-Лейцин, мг/л - 30
L-Изолейцин, мл/л - 10
L-Метионин, мг/л - 7,5
L-Серин, мг/л - 12,5
L-Треонин, мг/л - 15
L-Цистин, мг/л - 10
Холин-диН-цитрат, мг/л - 107
Среда CMRL 199 имела следующий состав.
СУХОЙ ПОРОШОК
Ингредиенты - мг/л
L-Аланин - 25,0
L-Аргинин (свободное основание) - 58,0
L-Аспарагиновая кислота - 30,0
L-Цистеин•НСl•Н2O - 0,1
Динатриевая соль L-цистеина - 24,0
L-Глутаминовая кислота•Н2O - 67,0
L-Глутамин - 100,0
Глицин - 50,0
L-Гистидин (свободное основание) - 16,2
L-Гидроксипролин - 10,0
L-Изолейцин - 20,0
L-Лейцин - 60,0
L-Лизин - 70,0
L-Метионин - 15,0
L-Фенилаланин - 25,0
L-Пролин - 40,0
L-Серин - 25,0
L-Треонин - 30,0
L-Триптофан - 10,0
L-Тирозин - 40,0
L-Валин - 25,0
п-Аминобензойная кислота - 0,050
Аскорбиновая кислота - 0,050 -Биотин - 0,010
-Биотин - 0,010
Пантотенат кальция - 0,010
Холиндигидроцитрат - 1,060
Фолиевая кислота - 0,010
Глутатион - 0,050 -Инозит - 0,050
-Инозит - 0,050
Менадион - 0,010
Никотинамид (ниацинамид) - 0,025
Никотиновая кислота (ниацин) - 0,025
Пиридоксаль•НСl - 0,025
Пиридоксин•НСl - 0,025
Рибофлавин-5-фосфат - 0,010
Тиамин•НСl - 0,010
Ацетат витамина А - 0,100
Витамин D (кальциферол) - 0,100
Витамин Е (α-токоферолфосфат) - 0,010
Аденинсульфат - 10,000
Аденозинтрифосфат - 1,000
Аденозин-5-фосфорная кислота - 0,200
Дезокси-2-рибоза - 0,500 -Рибоза - 0,500
-Рибоза - 0,500
Холестерин - 0,200
Гуанин - 0,300
Гипоксантин - 0,300
Культуры инфицировали соответствующим посевным вирусом при множественности инфицирований. Инфицирование проводили при 36±1oС. Когда С.Р.Е. вируса было полным, культуру охлаждали до 2-15oС.
Собранный вирус осветляли фильтрованием. Объем собранного вируса уменьшали мембранной ультрафильтрацией с номинальным отсечением молекулярной массы 100000 до объема, пригодного для диафильтрации против 0,04 М фосфатного буфера. После диафильтрации объем дополнительно концентрировали до объема, пригодного для гель-фильтрации. Производили выборку концентрата живого вируса и хранили его при 2-8oС.
Концентрат живого вируса наносили на гель-фильтрационную колонку и элюировали из колонки 0,04 М фосфатным буфером. Фракцию вируса собирали путем мониторинга оптической плотности элюата колонки при 254 и 280 нм.
Вторую стадию очистки проводили с применением ионообменной среды ДЭАЭ с 0,04 М фосфатом в качестве буфера для элюции. Эту стадию повторяли дважды, если количество ионообменной среды было недостаточным, как было определено мониторингом при 254 и 280 нм.
Собранную фракцию вируса концентрировали и диализовали против Специальной Среды Хенка для снижения содержания фосфата.
Специальная Среда Хенка имеет следующий состав:
АМИНОКИСЛОТЫ - мг/л
D,L-Аланин - 25,0
L-Аргинин•НСl - 58,0
D,L-Аспарагиновая кислота - 30,0
L-Цистеин•НСl•Н2O - 0,10
L-Цистин•2НСl - 26,00
D,L-Глутаминовая кислота - 67,0
L-Глутамин - 100,0
Глицин - 50,0
L-Гистидин•HCl•H2O - 16,2
L-Гидроксипролин - 10,00
D,L-Изолейцин - 20,00
D,L-Лейцин - 60,0
L-Лизин•НСl - 70,00
D,L-Метионин - 15,0
D,L-Фенилаланин - 25,00
L-Пролин - 40,00
D,L-Сepин - 25,00
D,L-Треонин - 30,00
D,L-Триптофан - 10,00
L-Тирозин (динатриевая соль) - 40,00
D,L-Валин - 25,00
ВИТАМИНЫ
Аскорбиновая кислота - 0,050 -Биотин - 0,010
-Биотин - 0,010
Витамин D (кальциферол) - 0,100
D-Пантотенат кальция - 0,010
Холинхлорид - 1,060
Фолиевая кислота - 0,010 -Инозит - 0,050
-Инозит - 0,050
Минеральные соли - мг/л
Хлорид кальция (безводный) - 40,00
Нитрат железа (III)•9Н2O - 0,10
Хлорид калия - 400,00
Хлорид натрия - 8000,00
Сульфат магния•7Н2О - 200,00
Другие ингредиенты
Аденинсульфат - 10,000
Аденозинтрифосфат (динатриевая соль) - 1,000
Адениловая кислота - 0,200
d-α-Токоферолфосфорная кислота (натриевая соль) - 0,010
Холестерин - 0,200
Дезоксирибоза - 0,500
Глюкоза - 1000, 000
Глутатион - 0,050
Гуанин•НСl - 0,300
Гипоксантин (натриевая соль) - 0,300
Рибоза - 0,500
Ацетат натрия•3Н2O - 81,500
Тимин - 0,300
Твин 80 - 20,000
Урацил - 0,300
Ксантин (натриевая соль) - 0,300
Менадион - 0,010
Никотиновая кислота - 0,025
Никотинамид - 0,025
п-Аминобензойная кислота - 0,050
Пиридоксаль•НСl - 0,025
Пиридоксин•НСl - 0,025
Рибофлавин-5-фосфат - 0,010
Тиамин•НСl - 0,010
Витамин А (ацетат) - 0,140
Очищенную вирусную фракцию фильтровали через фильтр с порами 0,2 μ.
Одну или более фракций очищенного концентрата вируса объединяли для инактивации. На основании результатов теста ELISA пул моновалентного вируса разбавляли до:
Тип I: 1750±250 ДЕ/мл,
Тип II: 1500±250 ДЕ/мл и
Тип III: 1250±250 ДЕ/мл
Специальной Средой Хенка.
Моновалентный пул нагревали до 37±1oС, затем фильтровали через фильтр с порами 0,2μ.
Добавляли необходимое количество формалина для получения концентрации 1: 4000. Пул вируса и формалин смешивали и перемешивали непрерывно при 37±1oС. Из моновалентного пула вируса брали пробы на жизнеспособность. На шестой день пул инактивируемого вируса фильтровали через фильтр 0,2μ и поддерживали при 37±1oС. На тринадцатый день инактивации пул вируса фильтровали через фильтр 0,2 μ.
Отбирали один или несколько инактивированных моновалентных компонентов и асептически соединяли с чаном для объединения. Моновалентный пул дополнительно концентрировали мембранной ультрафильтрацией с номинальным отсечением молекулярной массы 100000. Затем проводили диализ против разбавителя RIV-PBS:
Na2HPO4 (динатрийгидрогенфосфат), 0,346 г/CCml,
KH2PO4 (калийдигидрогенфосфат), 0,187 г/CCml
с Твином для получения однородного конечного продукта.
Альбумин (человека) добавляли до конечной концентрации 0,5%. Затем объединенный моновалентный концентрат фильтровали через фильтр 0,2μ. Разбавитель RIV-PBS с Твином добавляли для достижения оцененной (посредством расчета) концентрации 10-15 доз на 0,5 мл. Объединенный концентрат хранили при 2-8oС до последующего использования.
Рассчитывали подходящие объемы моновалентных компонентов Типов I, II и III и объединяли. Трехвалентная вакцина должна была содержать:
Тип I: 40 ДЕ/дозу 0,5 мл,
Тип II: 8 ДЕ/дозу 0,5 мл,
Тип III: 32 ДЕ/дозу 0,5 мл.
Трехвалентный концентрат хранили при 2-8oС до использования. Добавляли формальдегид и 2-феноксиэтанол и смешивали. Добавляли альбумин (человека) согласно расчету с получением конечной концентрации 0,5%.
(ii) выращенного на клетках Vеrо
Ампулы банка рабочих клеток Vero субкультивировали до уровня выбранного клеточного пассажа. Ампулы с клетками хранили в жидком азоте. Клетки выращивали с применением гранул-микроносителей, которые представляли собой сферические гранулы со средним диаметром приблизительно 100 микрометров, состоящие из декстрановых полимеров, имеющих радикалы ДЭАЭ, привитые на их поверхности (диэтиламиноэтил), что делало их положительно заряженными.
Основная среда для выращивания клеток была Минимальной Эссенциальной Средой (MEM) Игла в солевом растворе Ирла, обогащенная 0,2% гидролизатом лактальбумина, 0,1% декстрозой, 5% телячьей сывороткой. Каждый миллилитр среды содержит следующие антибиотики:
Стрептомицин: 75 единиц на 1 мл,
Неомицин: 14 единиц на 1 мл,
Полимиксин В (сульфат): 35 единиц на 1 мл.
Клетки Vero прогрессивно культивировали в Биогенераторах увеличивающегося размера. Затем культуральную среду и достаточный объем гранул-микроносителей вводили в промышленный биогенератор. Температуру стабилизировали при 37oС. Клетки, собранные трипсинизацией, добавляли и подвергали перемешиванию. Культивирование продолжали в течение 4-7 дней при 37oС, причем перемешивание прогрессивно увеличивали. Обычно в конце культивирования наблюдали увеличение клеточного роста в 6-20 раз. Средой, используемой для роста вируса, была среда 199 (Parker) в солевом растворе Ирла, обогащенная 0,1% декстрозой. Эта среда содержит те же самые антибиотики, в той же самой концентрации, что и среда для роста клеток, но она не содержит телячьей сыворотки. На 4-7-й день клеточного роста перемешивание в биогенераторе на промышленной стадии останавливали; гранулы оседали на дне чана, старую среду удаляли. Затем некоторое количество бессывороточной среды 199 вводили в каждый биогенератор и перемешивали. Затем эту среду удаляли, выполняя тем самым промывание гранул и клеток. Некоторое количество бессывороточной среды 199 переносили в биогенератор вместе с необходимым объемом партии посевного материала. Вирус адсорбировали в клетках медленным перемешиванием. В конце культивирования вируса перемешивание останавливали. Суспензию вируса оттягивали и собирали, а гранулы удерживались фильтрованием. Суспензию вируса гомогенизировали. Сбор, отфильтрованный на органической мембране со средним размером пор 0,20 мм, хранили при 4oС. Вирус концентрировали ультрафильтрацией.
Вирус дополнительно очищали ионообменной хроматографией с применением носителя ДЭАЭ-декстран-Сферосил, уравновешенного 0,04 М фосфатным буфером, рН 7,00. Затем вирус очищали гель-фильтрационной хроматографией с применением колонки, содержащей агарозный гель Sepharosis CL-6B, забуференный 0,04 М фосфатом, рН 7,00. Вирус дополнительно очищали хроматографией с применением носителя ДЭАЭ-декстран-Сферосил, забуференного 0,04 М фосфатным буфером, рН 7,00. Как только последняя очистка завершалась, суспензию вируса доводили до требуемого объема некоторым количеством Среды М-199, рН 7,0, без фосфата, концентрировали в 10 раз в ЭДТА 5 мМ, глицине при 0,5% и Твине 80 в конечной концентрации 50 мг/литр (Среда для инактивации). Эта смесь является "Концентрированной Вирусной Смесью" и ее фильтруют на мембране 0,2 мкм. Концентрированную вирусную суспензию хранят при 4oС в ожидании инактивации.
Одну или несколько партий "Концентрированной Вирусной Смеси" одного и того же типа смешивали и необязательно разбавляли или доводили некоторым количеством "Среды для инактивации" в подходящем чане. Разбавление доводили до правильного объема в соответствии с типами для получения титра D антигена:
1500 и 2000 D единиц в типе 1,
800 и 1000 D единиц в типе 2,
1000 и 1500 D единиц в типе 3,
и уровень белка:
менее или равен 40 мкг/мл в типе 1,
менее или равен 70 мкг/мл в типе 2,
менее или равен 30 мкг/мл в типе 3.
Доведенную суспензию очищенного концентрированного вируса фильтровали на мембране 0,22 мкм в течение максимально 72 часов перед началом инактивации. Затем эту вирусную суспензию нагревали снова при 37oС. Для инактивации добавляли раствор формальдегида с получением концентрации 1/4000. Для прослеживания кинетики инактивации после 24, 48, 72 и 96 часов во время первых четырех дней брали пробы. Проводили взятие проб по 10 мл с немедленной нейтрализацией формальдегида бисульфитом натрия и немедленным хранением при -20oС до титрования.
На 6-й день вирусную суспензию во время инактивации фильтровали при помощи фильтра 0,22 мкм. После фильтрования проводили инкубирование жидкости при 37oС еще в течение 6 дней с постоянным перемешиванием. На 9-й день инактивации брали 3-кратный объем относительно объема, соответствующего 3000 доз для человека, и 500 мл минимально неочищенного индивидуального сбора. Этот объем был рассчитан в соответствии с титром D антигена "Концентрированной Вирусной Смеси". Пробу сразу же нейтрализовали некоторым количеством бисульфита натрия для остановки действия оставшегося формальдегида. Затем гомогенизированную и инактивированную вирусную суспензию вынимали из термостата при 37oС после 12-го дня инактивации. Объем сразу же нейтрализовали некоторым количеством бисульфита натрия и хранили при 4oС.
Для приготовления концентрированной трехвалентной партии IPV моновалентные препараты комбинировали таким образом, чтобы обеспечить:
Тип 1 (Mahoney) 400 единиц антигена D,
Тип 2 (MEF-1) 80 единиц антигена D,
Тип 3 (Saukett) 320 единиц антигена D,
Среда 199, рН 7,2, q.s. до 1 мл.
Эту смесь перемешивали до гомогенного состояния, фильтровали на мембране с порами 0,22 мкм.
Конечную массу продукта получали из концентрированных трехвалентных партий, описанных выше, разбавлением средой 199, рН 7,2, без фенолового красного таким образом, чтобы стандартная доза содержала на 0,5 мл:
40 единиц D антигена для типа 1,
8 единиц D антигена для типа 2,
32 единицы D антигена для типа 3.
(b) Композиции
Поливалентные вакцинные композиции (APDT) содержали пять коклюшных антигенов (10 мкг РТ, 5 мкг FHA, 5 мкг FIM 2 и 3,3 мкг 69К), 15 Lf дифтерийного токсоида, 5 Lf столбнячного токсоида, 1,5 мг фосфата алюминия в качестве адъюванта и 0,6% 2-феноксиэтанол в качестве консерванта на 0,5 мл (CLASSIC). Эту вакцину использовали отдельно или в комбинации либо с IPV, продуцируемым на клетках MRC-5 (mIPV), полученным, как описано выше, IPV, продуцируемым на клетках Vero (vIPV), полученным, как описано выше, либо с OPV (Connaught Laboratories Limited). Для того чтобы не нарушать рутинной схемы иммунизации, содержащую конъюгат Haemophilus influenzae b-столбнячный токсоид вакцину вводили во время следующего визита.
Популяция
Здоровые 17-19-месячные дети, которые были иммунизированы тремя дозами DTP и 2 дозами OPV или 3 дозами DTP-IPV в возрасте 8 месяцев были вовлечены в это исследование. После письменного сообщения родителей или опекунов о согласии детей распределяли при помощи созданного компьютером сбалансированного блок-списка случайных номеров для получения одной из пяти вакцинных схем. Комбинированные вакцины, содержащие A3DT, вводили внутримышечно иглой 25 мм в дельтовидную мышцу плеча или в широкую латеральную мышцу бедра, если дельтовидная мышца имела недостаточную массу. IPV (0,5 мл; mIPV или vIPV) при отдельном введении вводили подкожно иглой длиной 1/2-5/8 дюйма (12,5-16 мм). Содержащие APDT вакцины вводили в левую конечность; правую конечность использовали для вакцин инактивированного полиовируса при раздельном введении и для конъюгатных вакцин Haemophilus Influenzae типа b при втором визите.
Клинический и лабораторный мониторинг
Пробы крови брали венепункцией или проколом пальца до и через 28 дней после иммунизации. Антитела IgG на РТ измеряли иммуноферментным анализом, а РТ-нейтрализующие антитела при помощи СНО-нейтрализации. Антитела IgG против FHA, против FIM и против 69К измеряли иммуноферментным анализом; единицы рассчитывали с применением сравнительной антисыворотки US FDA (#3). Измеряли также агглютинины коклюша. Дифтерийный антитоксин измеряли в тесте микронейтрализации и столбнячный антитоксин при помощи иммуноферментного анализа. Антитела против полиовируса типа 1, 2 и 3 измеряли вирусной нейтрализацией. Титры антител выражали в виде средних геометрических значений титров; пробы сывороток с титрами менее порога детектирования теста относили к величине 1/2 нижнего порога детектирования для целей статистических расчетов.
Геометрические средние титров антител и 95% доверительные интервалы вычисляли для титра антител для каждого антигена вакцины до и после иммунизации. Средние логарифмические титров и средние log-кратные повышения титров антител сравнивали анализом профиля и дисперсионным анализом. Соотношение субъектов, достигающих заранее заданных уровней в каждой группе, сравнивали при помощи логистической регрессии. Проводили сравнения между каждой полиовирусной вакциной, даваемой отдельно или в виде комбинированной инъекции, между mIPV и vIPV (в обоих случаях при отдельном и комбинированном введении) и между комбинированными вакцинами IPV и OPV. Для множественных сравнений не делали корректировок.
Всего 425 детей (52% женского пола) были зарегистрированы в этом исследовании и получали бустер-иммунизацию. Средний возраст при регистрации был 17,8 месяцев (диапазон 17,0-20,0). Образцы сывороток после иммунизации получали из 422 (99,3%) участников в среднем через 29,2 дня после иммунизации (диапазон 28-41 день). Побочные события, классифицированные как тяжелые, не были обычными в этом исследовании.
Перед иммунизацией уровни антител были эквивалентными среди групп для большинства антигенов. Исключением было то, что участники, распределенные для получения APDT и mIPV в виде отдельных инъекций, имели значимо более низкие уровни антител против FIM, агглютинина, против дифтерийного и столбнячного антигенов, чем группа, распределенная для получения комбинированной вакцины APDT-mIPV. Подобным образом, группа, рандомизированная для получения отдельных инъекций APDT и vIPV, имела более низкие уровни противостолбнячных антител, чем группа, получавшая комбинированную вакцину APDT-vIPV.
После иммунизации было значимое повышение антител во всех вакцинных группах против всех антигенов, включенных в вакцины. Были небольшие различия в образовании антител на коклюшные антигены в зависимости от группы полиовакцины. Не было различий в образовании антител против РТ в иммуноферментном анализе или СНО-нейтрализации или в антителах против FHA. Антитела против 69К были значимо выше в группе, получавшей вакцину mIPV, комбинированную с APDT (77,7 единиц), чем в группе, получавшей mIPV в виде отдельной инъекции (37,9 единиц; р менее 0,001) или в группе, получавшей OPV (47,7 единиц; р менее 0,05). Антитела против FIM и агглютининов были также выше в группе, получавшей комбинированную вакцину APDT-mIPV, чем в группе, получавшей отдельные инъекции, однако, те же самые различия были также обнаружены в сыворотках перед иммунизацией.
Были обнаружены различия в образовании антител против полиовируса. Обе вакцины APDT-mIPV и APDT-vIPV индуцировали более высокие уровни антител против полиовируса типа 1 и типа 3 (р менее 0,001 для всех сравнений). Уровни антител против полиовируса типа 2 были также более высокими после APDT-mIPV (10633 реципрокное разведение) и APDT-vIPV (10256), чем OPV (7185); однако, эти различия достигали уровня значимости только для APDT-mIPV (р менее 0,05). Титры антител против полиовируса, достигнутые с комбинированной вакциной APDT-mIPV, были также более высокими, чем в том случае, когда mIPV давалась в виде отдельной инъекции (6620-1; р менее 0,05).
Титры противостолбнячных антител были более высокими у реципиентов OPV, чем у реципиентов любой комбинации IPV (р менее 0,05). Антидифтерийные титры были также более высокими у реципиентов OPV, но этот результат достигал статистической значимости только в сравнении с группой APDT-vIPV (р менее 0,05). После иммунизации все дети имели уровни антител против дифтерии и столбняка более 0,01 МЕ/мл и все, кроме одного ребенка, имели уровни более 0,1 МЕ/мл.
Результаты этого исследования продемонстрировали, что компонентная коклюшная вакцина, содержащая РТ, FHA, 69K и FIM, комбинированная с дифтерийным и столбнячным токсоидами, может быть также комбинирована с инактивированной полиовирусной вакциной без значимого увеличения реактогенности или потери иммуногенности. В противоположность результатам с цельноклеточной вакциной DTP, не было снижения образования антител на антигены Bordetella pertussis. He было существенных различий между вакцинами IPV, полученными на клеточных линиях MRC-5 или Vero; обе вакцины индуцировали более высокие уровни сывороточных антител против полиовируса, чем OPV. Демонстрация равноценности mIPV и vIPV облегчает внедрение бесклеточной коклюшной вакцины в юрисдикции с предпочтительностью для IPV, происходящей из конкретной клеточной линии.
Выводом из экспериментальных данных, сообщенных в этом примере, является то, что они демонстрируют, что пятикомпонентная бесклеточная вакцина может безопасно комбинироваться с любой их двух вакцин IPV для четвертой дозы вакцины между возрастом 17 и 19 месяцев.
РЕЗЮМЕ ИЗОБРЕТЕНИЯ
Данное изобретение обеспечивает новые препараты антигенов Bordetella и не-Bordetella для получения поликомпонентных коклюшных вакцин. Такие вакцины являются безопасными, нереактогенными, иммуногенными и создающими иммунитет у людей. В объеме данного изобретения возможны модификации.











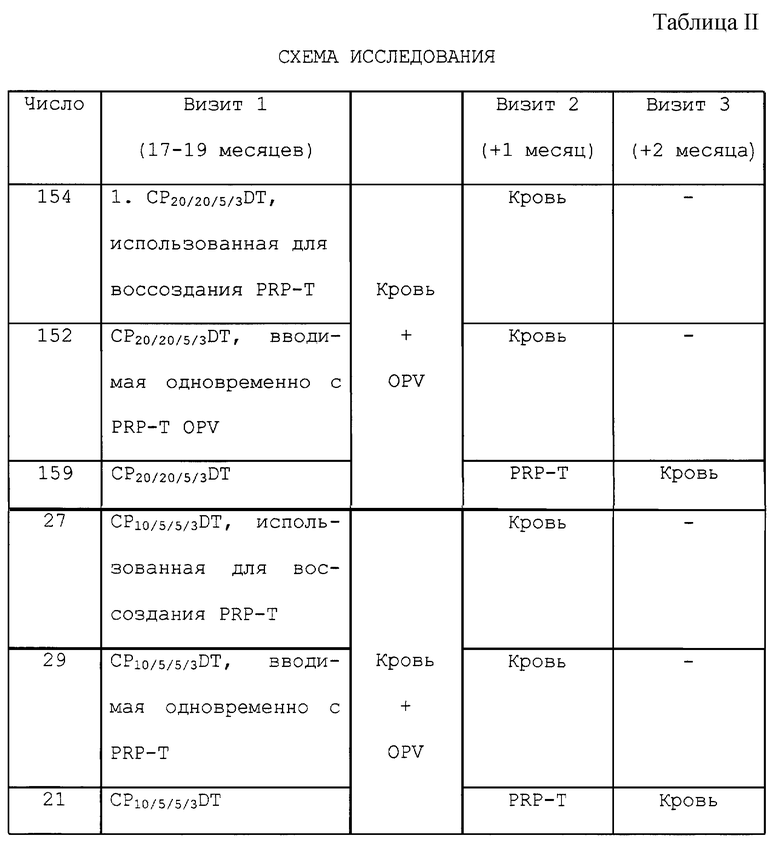
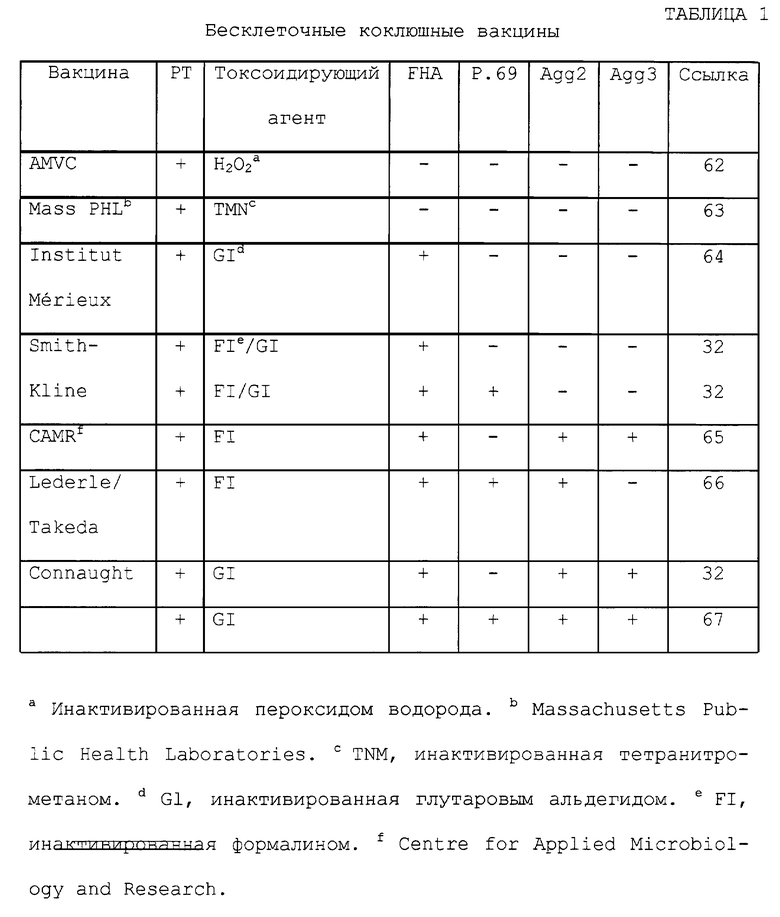
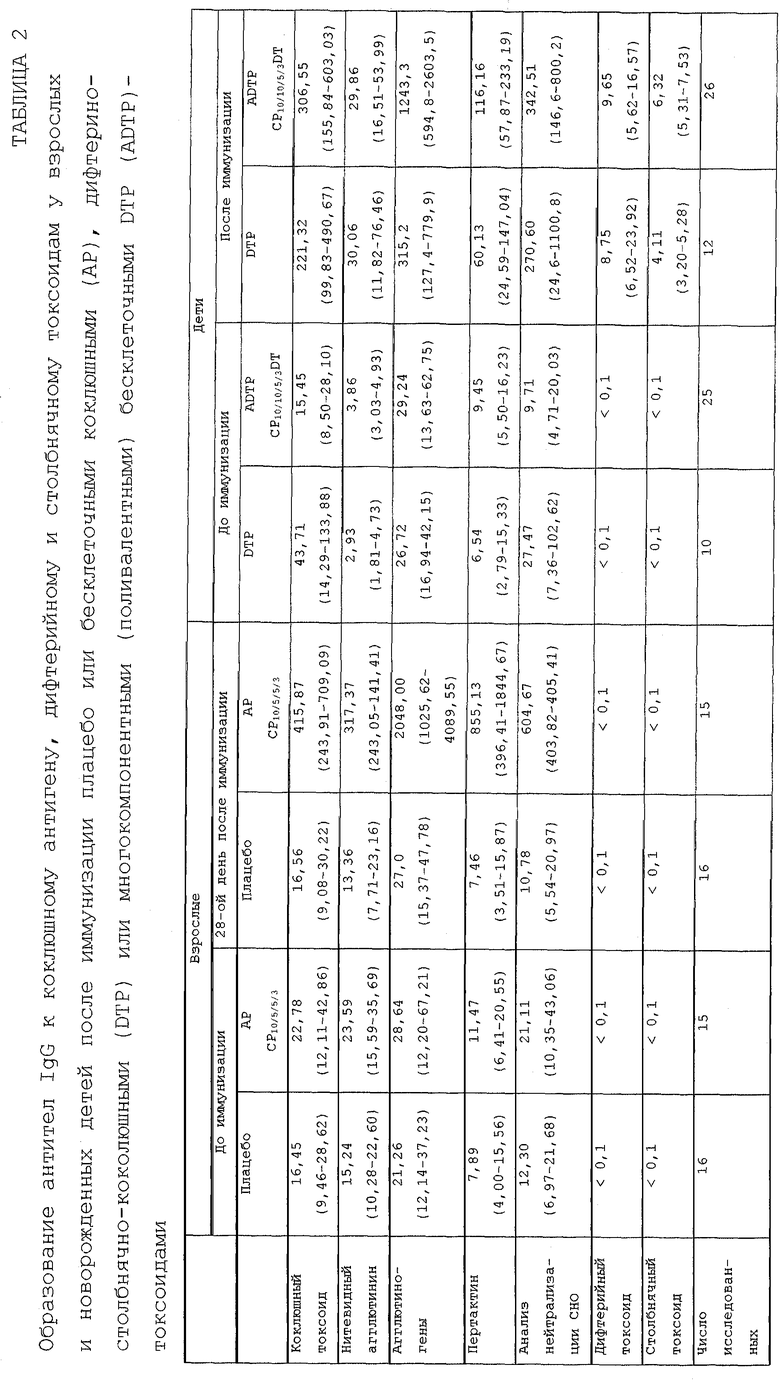
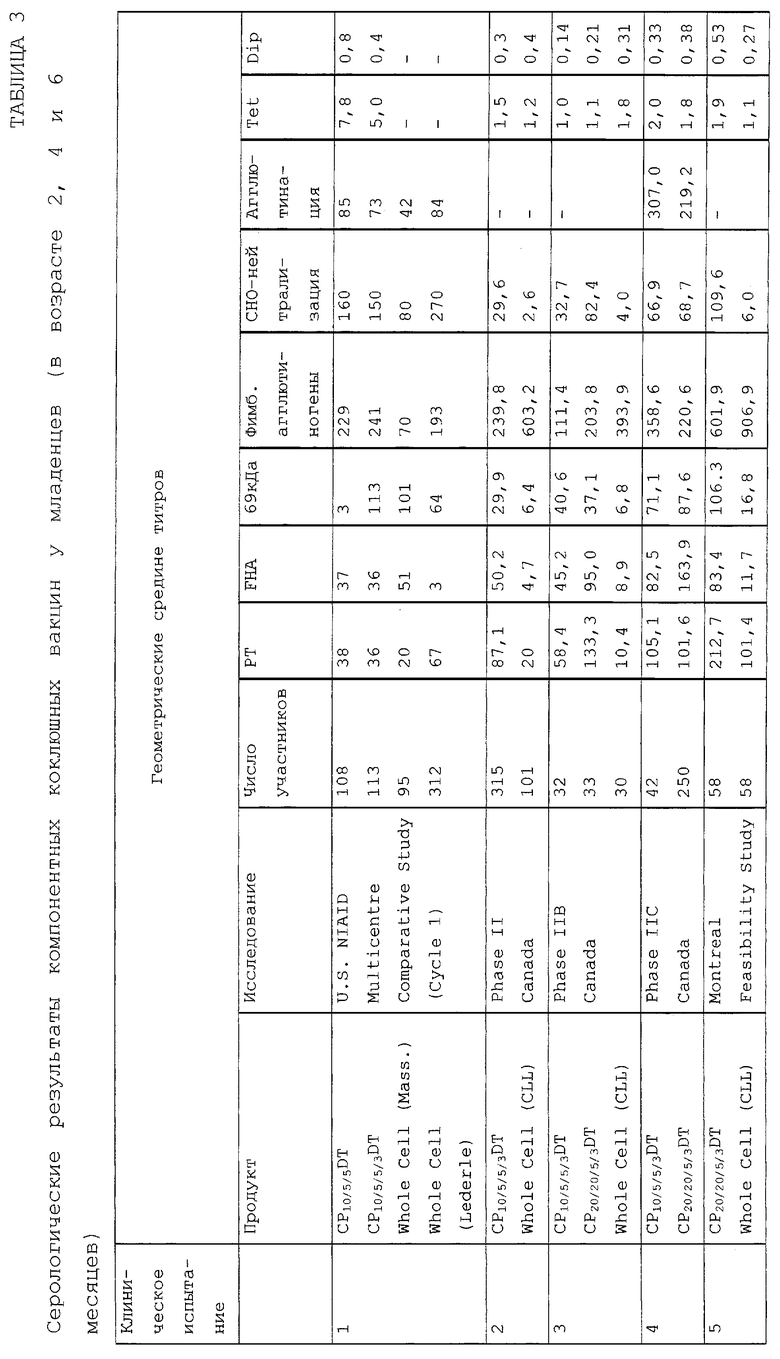





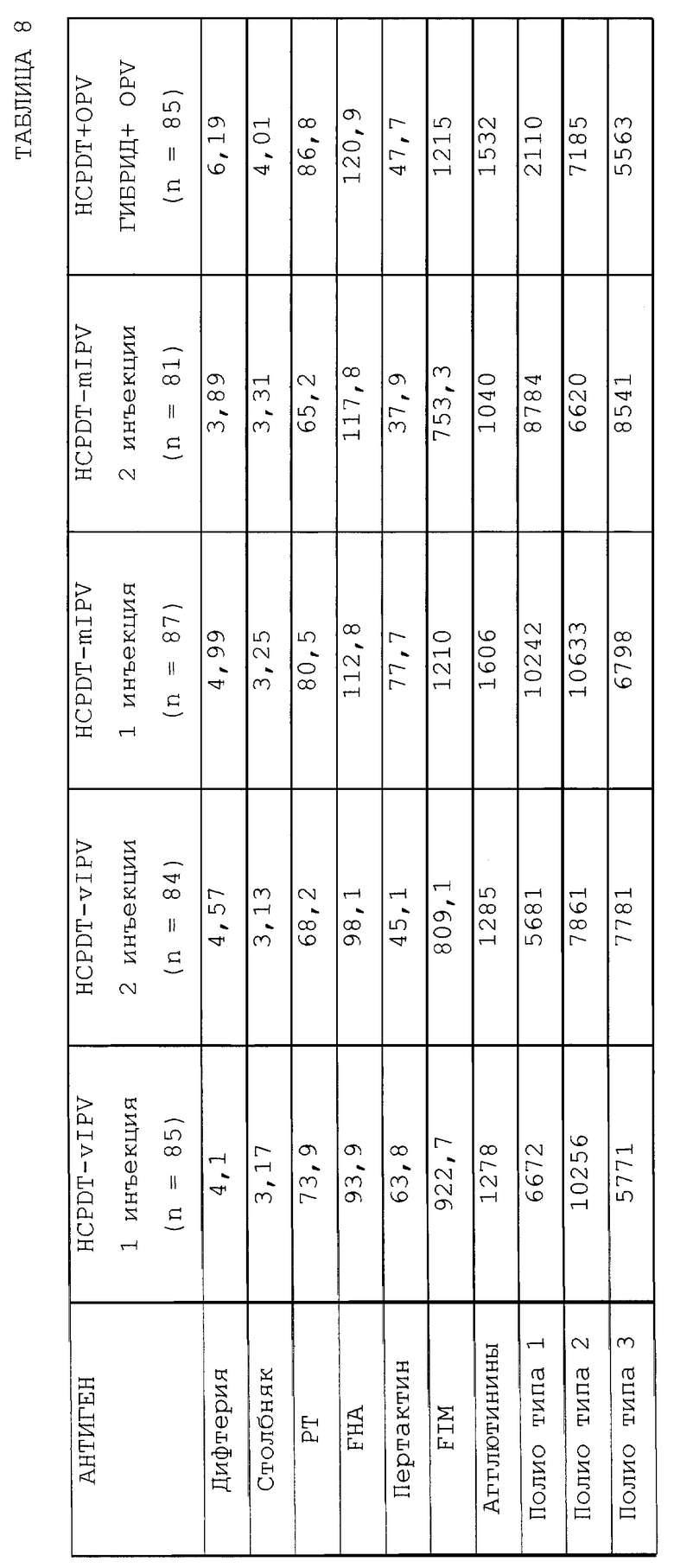

Изобретение относится к области медицины и касается поливалентной вакцинной композиции, содержащей бесклеточные компоненты коклюшной вакцины, дифтерийный токсоид, столбнячный токсоид и инактивированный полиовирус. Композиция может также содержать конъюгат капсульного полисахарида Haemophilus influenzae типа b и столбнячного или дифтерийного токсоида, который может быть реконструирован (воссоздан) из лиофилизированного состояния другими компонентами этой вакцины. Введение состоящей из многих компонентов вакцины не приводит к уменьшению иммуногенности любого компонента в результате взаимодействия с другими компонентами вакцины. Преимущество изобретения заключается в создании высокоимммуногенного препарата вакцины, включающего иммуногенные индивидуальные компоненты. 3 с. и 22 з.п.ф-лы, 11 табл., 1 ил.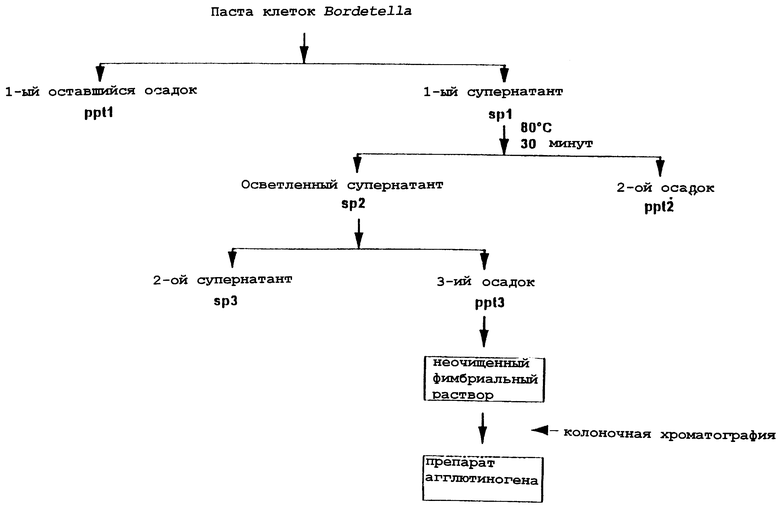
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| Частотно-фазовый компаратор | 1974 |
|
SU484621A1 |
| Руководство по вакцинному и сывороточному делу | |||
| /Под ред | |||
| П.Н.Бургасова | |||
| - М.: Медицина, 1978, с.157-175. | |||
