Изобретение относится к лечению расстройств, характеризуемых сдвигом активности иммунной системы с Th1-активности на Th2-активность. Оно основано на удивительном открытии того факта, что препараты из Mycobacterium vaccae способны влиять на неспецифическое системное смещение Th1, в частности, путем регуляции по типу обратной связи отрицательной Тh2-активности без сопутствующей стимуляции Thl-активности. Расстройства, к которым может быть применено настоящее изобретение, включают синдром хронической усталости, синдром войны в (Персидском заливе) и синдром тотальной аллергии.
У пациентов с синдромом хронической усталости (Turnberg et al.) аллергические явления могут наступать чаще, вдвое увеличивается число аллергических реакций кожи, также наблюдается низкая активность NK-клеток и низкий уровень продуцирования γ-интерферона и интерлейкина 2 (Straus, 1996). Некоторые люди имеют высокий уровень антител к вирусу Эпстайна-Барра, что также характерно для пациентов с пониженной Thl-активностью. У них также наблюдаются изменения настроения и депрессии. Все эти особенности хорошо вписываются в переключение с Th1 на Th2 в профиле цитокинов. Это смещение может быть последствием современного образа жизни, в котором имеется недостаток стимулов, индуцирующих Th1, а вместо этого имеется тенденция к иммунизации и аллергенам, индуцирующим Th2.
Авторы данного изобретения полагают, что синдром войны в Персидском заливе представляет собой особый случай синдрома хронической усталости, в котором стимулы, индуцирующие Th2, могут быть идентифицированы потому, что участникам войны в Персидском заливе проводились многократные вакцинации, индуцирующие Th2. Вакцинации или инфекции могут оказывать системное влияние и неспецифически увеличивать или снижать баланс цитокинов Thl/Th2 реакции на другие, не связанные с ними антигены (Shaheen et al., 1996, Shirakawa et al. , 1996), и влиять (положительно или отрицательно) на исход не связанных с ними заболеваний (Aaby, 1996, Aaby et al., 1995). Предположение авторов изобретения, что эти вакцинации, проводимые во время указанной войны, индуцировали системное переключение Th2, подтверждается следующими четырьмя пунктами протокола вакцинации:
(i) В качестве адъюванта для Британских войск в Персидском заливе применяли Pertussis, адъювантность которого потенциально представляет собой Th2 (Мu & Sewell, 1993, Ramiya et al., 1996, Smit et al., 1996).
(ii) Британским войскам в Персидском заливе вводили иммуногены, индуцирующие Th2, против чумы, сибирской язвы, брюшного тифа, столбняка и холеры. Более того, такая большая антигенная нагрузка имеет тенденцию вести реакцию по направлению к Th2 (Aaby, 1995, Bretscher et al., 1992, Hernandez-Pando & Rook, 1994).
(iii) Вакцинации проводились после использования войск в военной зоне или непосредственно перед их отправкой туда, т.е. в тот момент, когда уровень стресса был высоким. Иммунизация в присутствии повышенного уровня глюкокортикоидов (т.е. кортизола) направляет реакцию к Th2 (Bernton et al., 1995, Brinkmann & Kristofoc, 1995, Ramirez et al., 1996).
(iv) Войска также подвергались воздействию карбаматных и фосфорорганических инсектицидов, ингибирующих явление направленного интерлейкина 2, важного для нормальной функции Th1 (Casale et al., 1993). Важность этого компонента не определена, однако предполагается, что источником инсектицидов часто являлась местность в Персидском заливе, поэтому уровень их чистоты не известен, в них даже могли присутствовать более токсичные загрязнители.
Таким образом, многократные вакцинации, проводимые в таких обстоятельствах, могли вызвать продолжительный системный дисбаланс цитокинов. Такое же действие возникает спорадически в общей массе населения в результате вакцинаций или других стимулов окружающей среды и инфекций, индуцирующих Th2, и этим может объясняться широкая распространенность синдрома хронического переутомления.
Подробное объяснение этих положений приводится ниже.
Авторы настоящего изобретения сделали неожиданное открытие, что препараты (убитых) Mycobacterium vaccae потенциально способны возмещать неспецифическим образом системное смещение Th1->Th2. Задача состоит в смещении активности иммунной системы Th2 способом, включающим иммунный ответ на антигены, не присутствующие в инъецируемом препарате М. vaccae.
Это было продемонстрировано экспериментально как на животной модели, так и на человеке. Показательные эксперименты подробно описаны ниже. В частности, удивительно показана регуляция по типу отрицательной обратной связи Тh2-активности, которая может быть не связана со стимуляцией активности Th1.
Вкратце на экспериментальных животных неспецифический системный сдвиг с Тh2-активности после введения М. vaccae может наблюдаться в виде снижения титра ответа антитела, зависимого от интерлейкина 4 (Th2), на яичный альбумин (овальбумин) (аллерген, не связанный с самим М. vaccae) у мышей, предварительно иммунизированных таким образом, чтобы обеспечить реакцию Th2. Одна инъекция М. vaccae способна вызвать такой эффект, а дальнейшие инъекции могут усилить его. Такой эффект является неспецифическим, поскольку он не требует присутствия какого-либо компонента овальбумина в инъецируемом препарате.
У людей (раковые больные) этот эффект был показан в виде появления в периферической крови лимфоцитов, спонтанно секретирующих интерлейкин 2 (характерный цитокин Th1), a также снижения количества Т-клеток, секретирующих иитерлейкин 4 (характерный цитокин Th2), после стимулирования форболмиристат-ацетатом и кальциевым ионофором. Процентное содержание лимфоцитов, демонстрирующих этот активированный Thl-фенотип, прогрессивно увеличивается после каждой последующей инъекции М. vaccae, достигая плато у многих пациентов после 3-5 инъекций 109 организмов (0, 15, 30 дни, а затем ежемесячно).
Более того, ниже приводятся ссылки на доклады о больных, страдающих синдромом хронической усталости, которым вводились препараты М. vaccae с положительным результатом.
Авторы данного изобретения ранее описывали М. vaccae для применения при лечении туберкулеза (GB-A-2156673), рака (ZA 95/2644), ВИЧ-инфекции (WO 94/06466) и хронического воспаления (GB-B-2252044). W092/08488 (этих же авторов) описывает его применение в качестве адъюванта для введения с представляющим интерес антигеном, при этом происходит переход Т-клеточного компонента реакции на антиген с Th2-профиля на Thl-профиль. В W092/08488 не содержится какого-либо упоминания о том, что М. vaccae может вызвать неспецифический сдвиг Th2/Thl в активности иммунной системы, т.е. какой-либо иной сдвиг, чем в специально вызываемой реакции на антиген, вводимый с М. vaccae. Более того, ни в W092/08488, ни где-либо еще не содержится предположения, что он способен к регуляции по типу отрицательной обратной связи Тh2-активность без сопутствующей стимуляции Thl-активности.
Обзоры по М. vaccae в качестве Тh1-адъюванта включают Abou-Zeid et al., (1997), Skinner et al, (1997a), and Skinner et al., (1997b). См. также работу Mosmann and Sad (1996), представляющую собой обзор позитивной регуляции Thl-цитокина, оказывающего ингибирующее действие на пролиферацию Тh2-клеток.
Настоящее изобретение в целом относится к регуляции по типу отрицательной обратной связи Тh2-активности иммунной системы пациента без стимуляции Thl-активности, в частности при лечении расстройств, характеризуемых сдвигом активности иммунной системы с Тh1- на Тh2-активность, в частности, когда регуляция по типу отрицательной обратной связи Тh2-активности необходима без сопутствующей стимуляции Thl-активности.
Один из аспектов настоящего изобретения предусматривает способ лечения больного, включающий введение больному антигенного и/или иммунорегулирующего материала из Mycobacterium vaccae.
Дальнейший аспект настоящего изобретения предусматривает применение антигенного и/или иммунорегулирующего материала из Mycobacterium vaccae для получения лекарственного средства для лечения больных.
Другой аспект настоящего изобретения предусматривает вещество или композицию для применения при лечении больных, при этом указанное вещество или композиция включает антигенный и/или иммунорегулирующий материал из Mycobacterium vaccae.
Препарат из М. vaccae предпочтительно предусматривается без какого-либо постороннего ("чужого") антигена, а включает иммуноген или аллерген.
Расстройства, подвергаемые лечению, могут характеризоваться общим, неспецифическим смещением иммунной системы пациента с Thl-активности на Тh2-активность. Такое смещение может быть установлено или диагностировано путем определения пониженного продуцирования интерлейкина-2(IL-2), либо повышенного продуцирования интерлейкина-4 (IL-4) или интерлейкина-5 (IL-5) у пациента, либо путем определения интерлейкина 13 (IL-13) как представителя Th2-цитокинов, и гамма-интерферона как представителя Th1-цитокина. Могут быть также рассмотрены другие Thl/Th2-цитокины. Неспецифический сдвиг с Th1- на Тh2-активность иммунной системы, возможно, не объясняется или вызывается воздействием на больного конкретного антигена или инфицированием конкретным патогеном. Больные, подвергаемые лечению, могут не страдать от туберкулеза или иной микобактериальной инфекции. Конкретные расстройства, которые могут быть подвергнуты лечению в соответствии с настоящим изобретением, включают синдром хронического переутомления, синдром войны в Персидском заливе и синдром тотальной аллергии (Straus, 1996). В соответствии с настоящим изобретением лечению могут быть также подвергнуты другие расстройства, при которых необходима регуляция по типу отрицательной обратной связи Тh2-активности без стимуляции регуляции Thl-активности.
Синдром хронической усталости может быть определен, например, с использованием так называемого критерия CDC 1994 (Fukuda et al.) или так называемого оксфордского критерия (Sharpe et al,). Современные знания о синдроме хронического переутомления суммированы и прокомментированы Turnberg et al. (опубликовано в октябре 1996 г.).
Материал из Mycobacterium vaccae может представлять собой или включать мертвые клетки М. vaccae. Такие клетки могут быть убиты, например, с применением облучения, к примеру, от 60кобальта, в дозе 2,5 мегарад, химическим или каким-либо другим способом, хотя предпочтительным является автоклавирование, например, при 69 кРа в течение 10 мин при 115-125oС. В результате автоклавирования может быть получен более эффективный препарат, чем в результате облучения.
Перед умерщвлением клетки М. vaccae могут быть выращены на подходящей твердой среде. Предпочтительной является модифицированная жидкая среда Сатона (Boyden et al.), отверждаемая агаром, предпочтительно 1,3% агаром. После аэробного инкубирования, обычно при 32oС в течение 10 дней организмы могут быть собраны, затем взвешены и суспендированы в разбавителе, готовом для введения. Если перед применением необходимо хранение, оно должно осуществляться при 4oС.
Вместо выращивания клеток на твердой среде может применяться жидкая среда, такая как модифицированная среда Сатона (Boyden et аl.), например, в ферментере.
Разбавитель может представлять собой незабуференный солевой раствор, свободный от пирогенов. Разбавитель предпочтительно забуферен боратом, предпочтительно содержащим поверхностно-активное вещество, такое как Твин 80®. Подходящий боратный буфер включает: Na2B4O7.10H20 - 3,63 г, Н3ВО3 - 5,25 г, NaCl - 6,19 г, Твин 80® - 0,0005%, дистиллированную воду - до 1 л. Эти разбавители являются фармацевтически приемлемыми.
Вышеупомянутые результаты испытаний на людях были получены путем введения М. vaccae в виде препарата GMP (гуаниловая кислота), SRL172, который может быть получен для применения на людях в нескольких исследовательских IDN. Администрации по контролю качества пищевых и лекарственных продуктов (США), а также СТХ Агентства по контролю над лекарственными препаратами (Соединенное королевство). Острая токсикология GLP проводилась Huntingdon Research. Данные по безопасности Фазы 1 и Фазы 2 были получены в США и зарегистрированы в Администрации по контролю пищевых и лекарственных продуктов. SRL172 предназначен для испытаний Фазы 3 при иммунотерапии туберкулеза. SRL172 является предпочтительным для применения в соответствии с настоящим изобретением.
SRL172 представляет собой состав М. vaccae, полученный из штамма R877R, который в соответствии с Будапештской конвенцией был депонирован в Национальной коллекции типовых культур (NCTC), Central Public Health Laboratory, Colindale Avenue, London NW9 5HT, United Kingdom, 13 февраля 1984 г. под номером NCTC 11659. R877R был первоначально выделен из образцов грязи из района Lango в Центральной Уганде (Stanford and Paul).
Вместо SRL172 могут быть использованы другие штаммы М. vaccae. Организм, принадлежащий к М. vaccae, может быть идентифицирован по биохимическим и антигенным критериям (Bonicke et al.).
Для настоящего изобретения предпочтительно, чтобы вводимый материал из М. vaccae был свободен или по существу свободен от не-М. vaccae антигенного или иммунорегулирующего материала. Другими словами, вводимое лекарственное средство или композиция может включать или по существу состоять из антигенного и/или иммунорегулирующего материала М. vaccae, такого как мертвые клетки, экстракт или его производное и фармацевтически приемлемого разбавителя.
Предпочтительно вводят "терапевтически эффективное количество", достаточное для улучшения состояния пациента. Такое улучшение может по крайней мере включать уменьшение интенсивности по меньшей мере одного симптома. Действительно вводимое количество, частота и длительность курса введения зависят от характера и тяжести подвергаемого лечению заболевания. Назначение лечения, например определение дозировки и т.д., входит в обязанности терапевтов и других врачей.
Разовая доза (при введении мертвых клеток) обычно содержит от 107 до 1010 убитых микроорганизмов М. vaccae. Пациентам может быть введена разовая доза, содержащая 108-109 убитых М. vaccae, хотя при необходимости доза может быть повторена, например, с интервалом от 2 недель до 6 месяцев.
Композиция может быть введена отдельно или в сочетании с другими видами лечения, одновременно или последовательно в зависимости от подвергаемого лечению состояния.
Фармацевтические композиции в соответствии с настоящим изобретением, а также для применения в соответствии с настоящим изобретением могут включать помимо активного ингредиента фармацевтически приемлемый наполнитель, носитель, буфер, стабилизатор или другие материалы, хорошо известные специалистам в данной области. Такие материалы должны быть нетоксичными и не влиять на эффективность активного ингредиента. Конкретная природа носителя или другого материала зависит от способа введения, который предпочтительно представляет собой инъецирование, например кожное, подкожное или внутрикожное.
Активный ингредиент для инъекций имеет вид парентерально приемлемого водного раствора, свободного от пирогенов и имеющего подходящую рН, изотоничность и стойкость. Подходящие фармацевтически приемлемые и предпочтительные разбавители были уже приведены выше.
Может также применяться пероральное введение, при котором фармацевтическая композиция может иметь вид таблетки, капсулы, порошка или жидкости. Таблетка может включать твердый носитель, такой как желатин или адъювант. Жидкие фармацевтические композиции обычно включают жидкий носитель, такой как вода, вазелин, животные или растительные масла, минеральное масло или синтетическое масло. Может быть также включен физиологический солевой раствор, декстроза или другой раствор сахаридов, либо гликоли, такие как этиленгликоль, пропиленгликоль или полиэтиленгликоль.
В дыхательные пути М. vaccae может быть введена при помощи аэрозоля с применением подходящего состава, например, включающего частицы, имеющие размер, который позволяет им достигать соответствующих частей дыхательных путей. Это скорее сухой порошок, чем водная суспензия.
Вместо мертвых клеток может быть использован материал, полученный из М. vaccae, в частности экстракт или синтетическая молекула, имеющая необходимую активность.
Как уже было вкратце отмечено выше, многократные вакцинации, стресс и другие факторы, возможно, явились причиной длительного системного цитокинового дисбаланса у ветеранов войны на Персидском заливе. Такой же эффект спорадически возникает у всего населения в результате вакцинаций или других индуцирующих Th2 стимулов окружающей среды и инфекций и может объясняться широким распространением синдрома хронического переутомления.
Сильные иммуногены могут оказывать системное длительное неспецифическое воздействие на природу иммунной реакции на не связанные с ними антигены. Например, инфекция кори снижает частоту возникновения атопии и аллергических реакций на клещи домашней пыли (Shaheen et al., 1996). Подобным образом японские дети с положительной реакцией на туберкулиновую кожную пробу менее подвержены атопии, чем дети с отрицательной реакцией на туберкулиновую пробу, при этом отношение циркулирующих в них Thl/Th2 цитокинов выше. Более того, после повторной инъекции БЦЖ дети, у которых происходит туберкулиновая конверсия, имеют повышенную вероятность потери своих атопических симптомов (Shirakawa et al., 1996).
Другим примером является вакцинация против кори. При введении стандартной дозы этой вакцины уровень смертности снижается значительно сильнее, чем уровень распространения кори у невакцинированного населения. Вакцины дифтерии, столбняка и коклюша (индуцирующие Th2) не обладают таким неспецифическим защитным действием (Ааbу et al., 1995). Однако при использовании высокотитрованной вакцины от кори смертность увеличивается, хотя защита от самой кори сохраняется (Ааbу, 1995, Aaby et al. 1995). Существуют доказательства того, что такое увеличение смертности сопровождается переключением на Th2, и зависимое от дозы увеличение индукции компонента Th2 хорошо изучено относительно нескольких других иммуногенов (Bretscher et al., 1992, Hernandez-Pando & Rook, 1994). Вакцины, использовавшиеся в войне в Персидском заливе, являлись Th2-индуцирующими вакцинами (чума, сибирская язва, брюшной тиф, столбняк, холера) и аккумулятивно составляли большую антигенную нагрузку, далее способствуя Th2. Более того, эта тенденция нарастала в результате использования pertussis в качестве адъюванта, который является сильным индуктором Th2 (Mu & Sewell, 1993, Ramiya et al., 1996, Smit et al., 1996). Это свойство pertussis недавно возбудило дискуссию о том, что, возможно, ее введение детям способствует наблюдаемому в настоящее время повышенному распространению атопии (Nilsson et al., 1996, Odent et al., 1994).
Природная тенденция этих вакцин направлять системный Th2 ответ значительно усиливалась эндокринными факторами, вторичными по отношению к стрессу, которому подвергались участники войны в Персидском заливе во время вакцинации.
Т-клеточные ответы модулируются несколькими стероидными гормонами. Дегидроэпиандростерон или его неизвестные метаболиты имеют тенденцию стимулировать профиль Th1. Таким образом, дегидроэпиандростерон способен восстанавливать иммунные функции у пожилых мышей и корректировать разрегулированное выделение цитокина, наблюдаемое у старых животных (Daynes et al., 1993, Garg & Bondada, 1993). Его исследовали на подобные свойства у старых людей (Morales et al. , 1994). Он усиливает продуцирование Thl-цитокинов, таких как интерлейкин 2 и гамма-интерферон(Daynes & Araneo, 1989, Daynes et al. , 1990, Daynes et al., 1995, Daynes et al., 1991). Дегидроэпиандростерон также усиливает секрецию иитерлейкина-2 из периферических Т-клеток крови человека (Suzuki et al., 1991).
Такие действия дегидроэпиандростеронов являются обратными действию глюкокортикоидов, таких как кортизол, которые усиливают Тh2-активность и оказывают совместное действие с Тh2-цитокинами (Fischer & Konig, 1991, Guida et al. , 1994, Padgett et al., 1995, Wu et al., 1991). Если пролиферация "наивных" Т-лимфоцитов направляется в присутствии неспецифического стимула (Brinkmann & Kristofic, 1995) или антигеном (после вакцинации), то развиваются Т-лимфоциты с профилем Тh2-цитокина. Это было довольно четко показано на клетках селезенки "чистых" лабораторных грызунов, при обычных обстоятельствах, имеющих мало клеток памяти (Ramirez et al., 1996).
В целом "нижняя линия" заключается в том, что кортизол способствует развитию профиля Тh2-цитокина из "наивных" клеток (Brinkmann & Kristofic, 1995). Это явление не следует путать с тем фактом, что цитокинсекретирующая активность установленных Тh2-клеток легко ингибируется кортизолом. Таким образом, применение аналогов кортизола в известных видах лечения Th-2-опосредованных заболеваний, таких как экзема, астма и сенная лихорадка, может срабатывать через антивоспалительное действие и путем снижения продуцирования цитокинов Тh2-клетками (Corrigan et al., 1995), и все же применение кортизола не способствует разрешению существующей проблемы путем направления вновь рекрутированных Т-клеток к Th2.
Психологический и физический стресс активирует систему гипоталамус-гипофиз-надпочечники и таким образом вызывает различные изменения, включая повышенное продуцирование кортизола. Большая физическая нагрузка и нехватка еды и сна приводит к снижению отношения дегидроэпиандростерона к кортизолу. Это непосредственно коррелирует со снижением запаздывания реактивности гиперчувствительности (Th1-маркер), при этом наблюдается одновременное повышение уровня иммуноглобулина Е сыворотки. Иммуноглобулин Е полностью зависит от продуцирования Тh2-цитокина (Bernton et al., 1995). Этого следует ожидать в свете известного действия дегидроэпиандростерона и кортизола, описанного выше.
Дальнейшим примером эффекта переключения Th1-Th2 стресса является увеличение антител к вирусу Эпстайна-Барра у студентов, реагирующих стрессом на экзамены. Этот вирус обычно контролируется ответом Thl и цитотоксичными Т-клетками, и потеря контроля приводит к репликации вируса и повышенному содержанию антител (Zwilling, 1992). Подобным образом лейкоциты периферической крови студентов-медиков во время сессии имеют пониженное содержание мРНК для гамма-интерферона, Thl-цитокина (Glaser et al., 1993).
Эти утверждения могут быть продемонстрированы более контролируемым образом на животных. Стресс, вызванный стеснением или ограничением, может увеличить рост микобактерий у мышей с туберкулезом (Brown et al., 1993, Tobach & Bloch, 1956). Эта модель является чувствительной к присутствию даже небольшого количества компонента Th2 (Rook & Hernandez-Pando, 1996). Были также описаны другие примеры (Moynihan, 1994).
Существует убедительное доказательство того, что депрессия может быть связана с излишним кортизол-опосредованным действием в мозгу (обзор Raven et al. , 1996), а стресс может привести к депрессии. Таким образом, депрессия (как видно на примере синдрома хронического переутомления и синдрома войны в Персидском заливе) имеет тенденцию быть связанной с Тh2-опосредованными расстройствами, такими как астма и экзема. Некоторые эндокринные изменения также обычны для расстройств Th2 и депрессий (Holsboer et al., 1984, Rupprecht et al., 1995).
При туберкулезе происходит системный сдвиг Th2 (Rook & Hernandez-Pando, 1996), при этом необычный тип метаболитов стероидов надпочечников выделяется в мочу (Rook et al., 1996). Таким образом появляется повышенное содержание метаболитов кортизола относительно метаболитов кортизона. Лечение заболевания восстанавливает преобладание Тh1 и исправляет тип стероидных метаболитов таким образом, что количество метаболитов кортизона увеличивается относительно метаболитов кортизола (Rook et al., 1996). Лечение депрессии лекарственным препаратом Нетирапон вызывает такие же изменения в стероидных метаболитах (т.е. увеличение метаболитов кортизона относительно метаболитов кортизола) (Raven et al., 1995). Препарат из М. vaccae SRL172 эффективен при иммунотерапии туберкулеза.
Как указано выше, вакцинации могут индуцировать системные неспецифические изменения баланса Thl/Th2, a также антигенспецифическую иммунизацию, которой обычно добиваются. Лечение синдрома хронического переутомления может потребовать индуцирования системного неспецифического смещения Th1 с целью восстановления доминирования Тh1 над Тh2, что характерно для нормальных здоровых особей. Это смещение противоположно смещению, которое, возможно, было индуцировано вакцинациями, получаемыми участниками войны в Персидском заливе.
Экспериментальное подтверждение, приводимое в данном описании, удивительным образом показывает, что препараты из Mycobacterium vaccae могут снижать объем нежелательного смещения Th1 к Th2, наблюдаемого у ветеранов войны в Персидском заливе.
Далее настоящее изобретение иллюстрируется без ограничения со ссылкой на экспериментальные примеры и следующие чертежи:
Фиг.1 показывает изменение общего количества иммуноглобулина Е сыворотки (нг/мл ± стандартное отклонение) у мышей в различные дни после первой иммунизации овальбумином, подсчитанное с учетом величины на 46-й день. Закрашенные кружочки означают реципиенты М. vaccae, чистые квадратики означают контроль солевого раствора. Для удобства указаны дни иммунизации овальбумином и введения М. vaccae или солевого раствора.
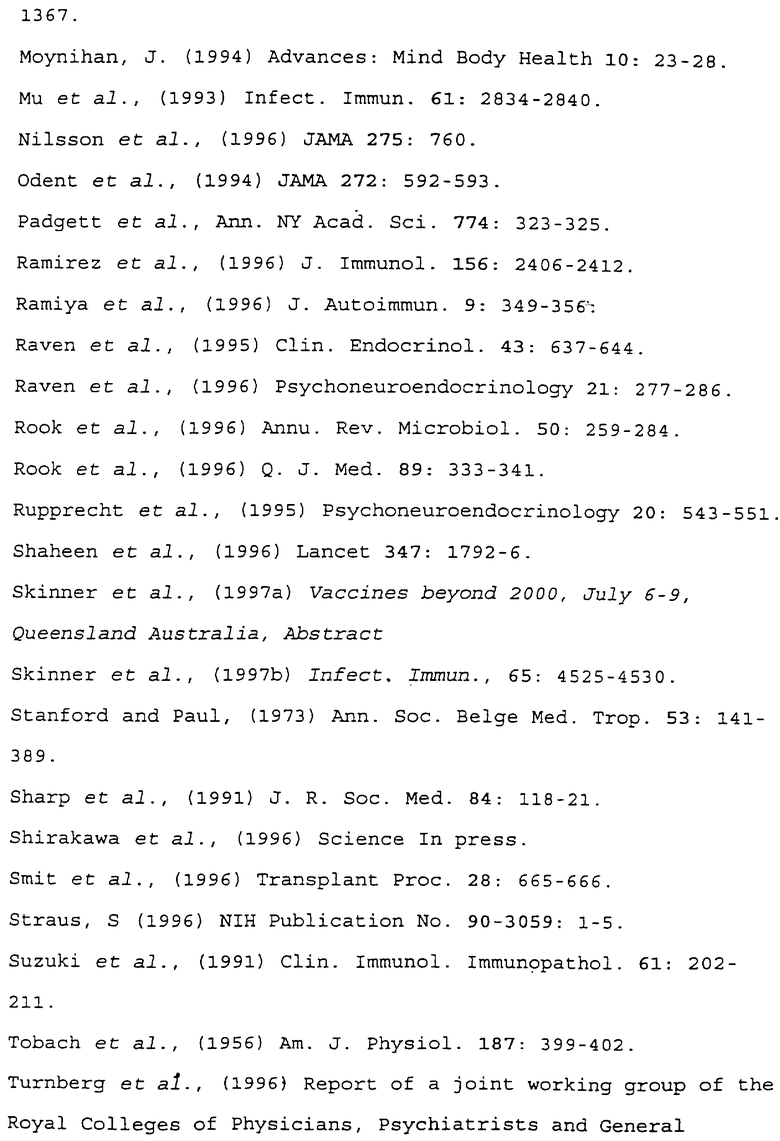
Фиг.2 показывает результаты измерений интерлейкина-5 (пг/мл ± стандартное отклонение), выделяемого in vitro из клеток селезенки, взятых у мышей, стимулированных овальбумином. Минимальная концентрация, определяемая в результате проводимого анализа, составляет 15,6 пг/мл.
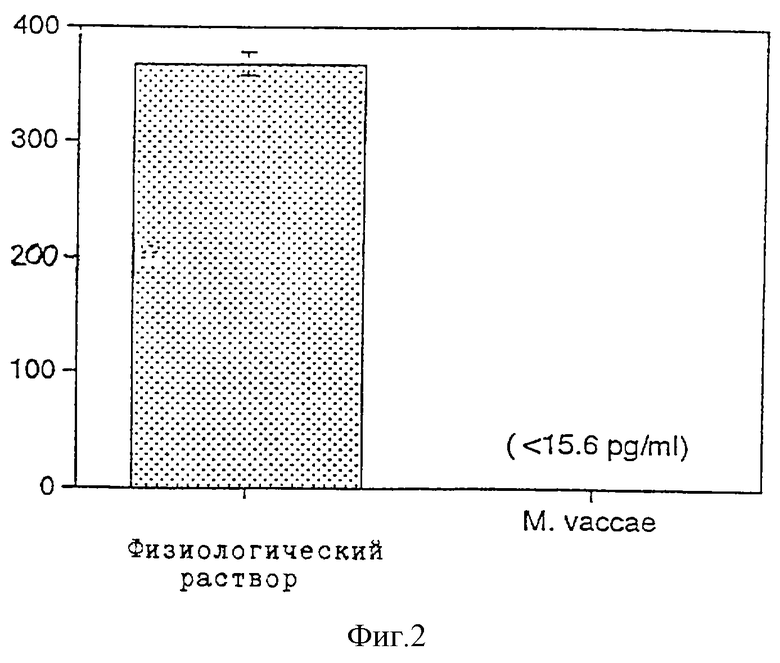
Фиг. 3 показывает эффект лечения при помощи подкожных инъекций солевого раствора (•) по сравнению с 107 препаратами из М. vaccae (о) на общее количество иммуноглобулина Е в сыворотке (нг/мл) мышей BALB/c, ранее иммунизированных OVA. Фиг.3А показывает 107 немодифицированные М. vaccae (Mv), фиг.3В - OVA-конъюгированные М. vaccae (Mv-OVA) и фиг.3С - промытые NaCl и обработанные периодатом контрольные М. vaccae (NaCl-Mv). Данные выражены в виде изменения относительно данных, полученных от образцов, взятых на 46-й день ± стандартное отклонение. Схемы иммунизации и лечения показаны на фиг. 3А - 3С (OVA: иммунизация OVA 50 мкг в IFA).
Фиг.4 показывает действие лечения при помощи
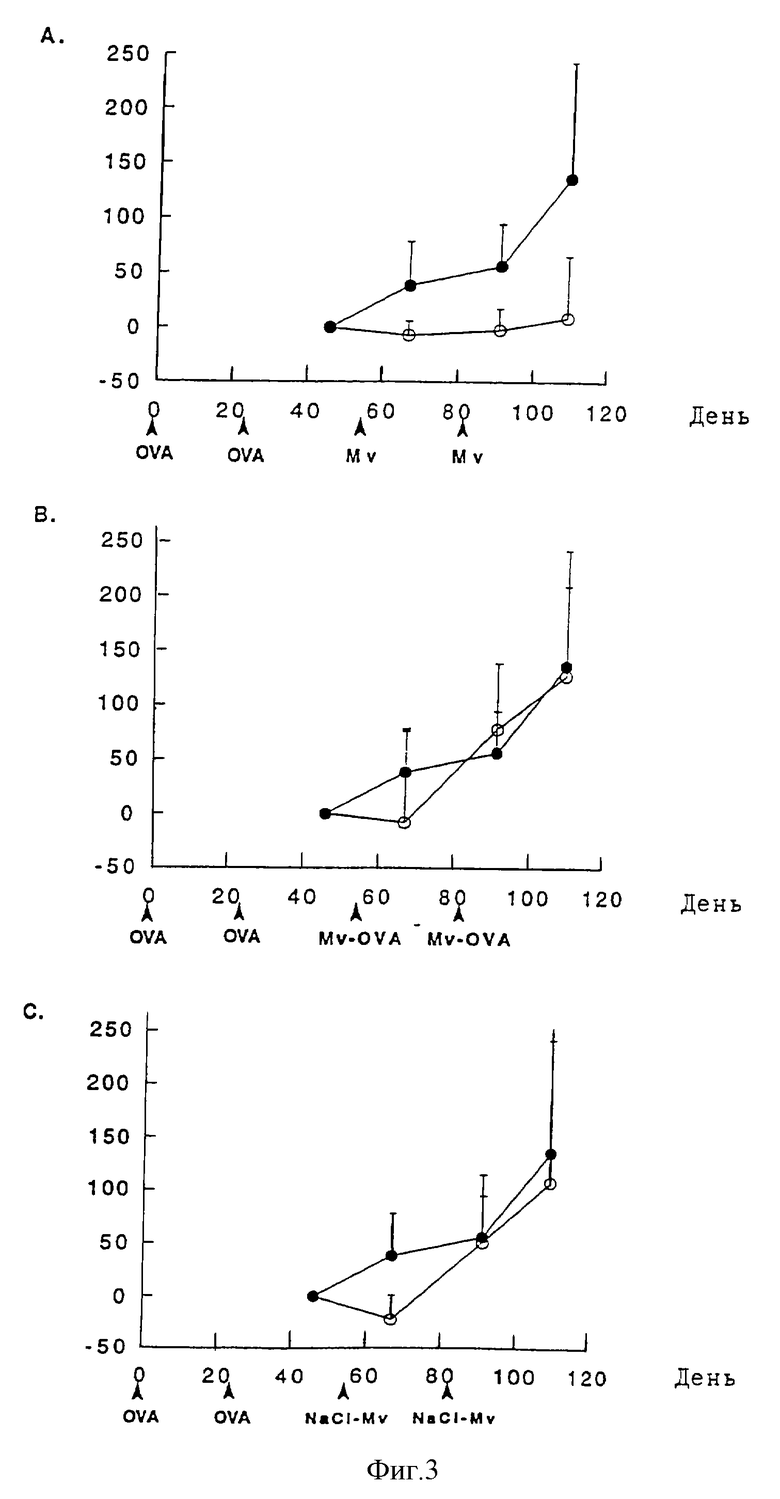
подкожных инъекций солевого раствора (•) по сравнению с 107 препаратами из М. vaccae (о) на OVA-специфический иммуноглобулин Е у мышей BALB/c, ранее иммунизированных OVA. Фиг. 4А показывает немодифицированные М. vaccae (Mv) (оптическая плотность 405 нм), Фиг. 4В - OVA-конъюгированные М. vaccae (MvOVA) (оптическая плотность 405 нм) или Фиг.4С - промытые NaCl и обработанные периодатом контрольные М. vaccae (NaCl-Mv) (нг/мл). Данные выражены в виде изменения относительно данных, полученных от образцов, взятых на 46-й день ± стандартное отклонение. Схемы иммунизации и лечения показаны на фиг. 4А-4С.
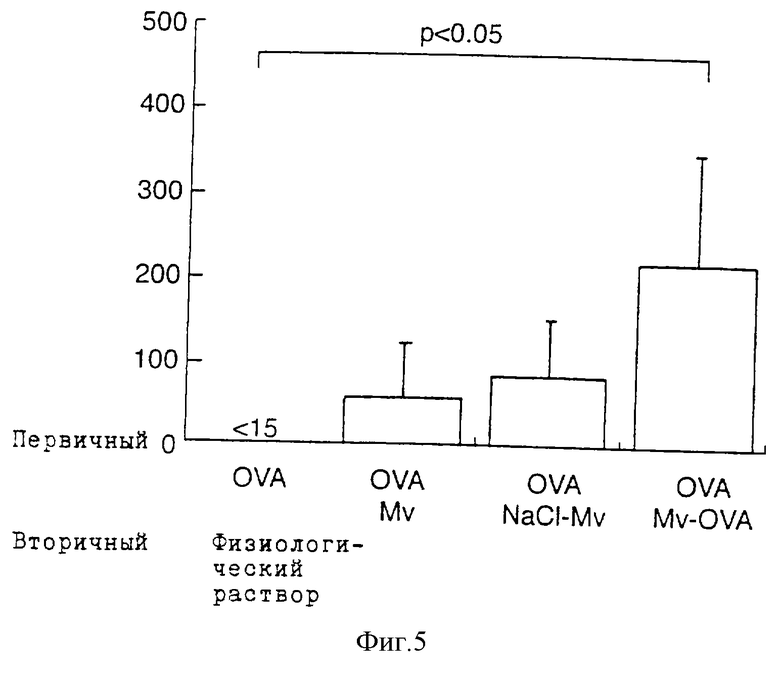
Фиг.5 показывает продуцирование интерлейкина-2 спленоцитами, культивированными с OVA. Селезенку забирают на 109-й день у мышей, иммунизированных OVA в 0-й и 24-й дни и обработанных на 53-й и 81-й дни солевым раствором, 107 немодифицированными М. vaccae (Mv), OVA, конъюгированным на М. vaccae (Mv-OVA) или соответствующими контрольными, промытыми NaCl и обработанными периодатом контрольными М. vaccae (NaCl-Mv). Через 24 ч надосадочные жидкости культур собирают для определения интерлейкина-2. Данные представлены в виде среднего уровня цитокина ± стандартное отклонение (пг/мл).
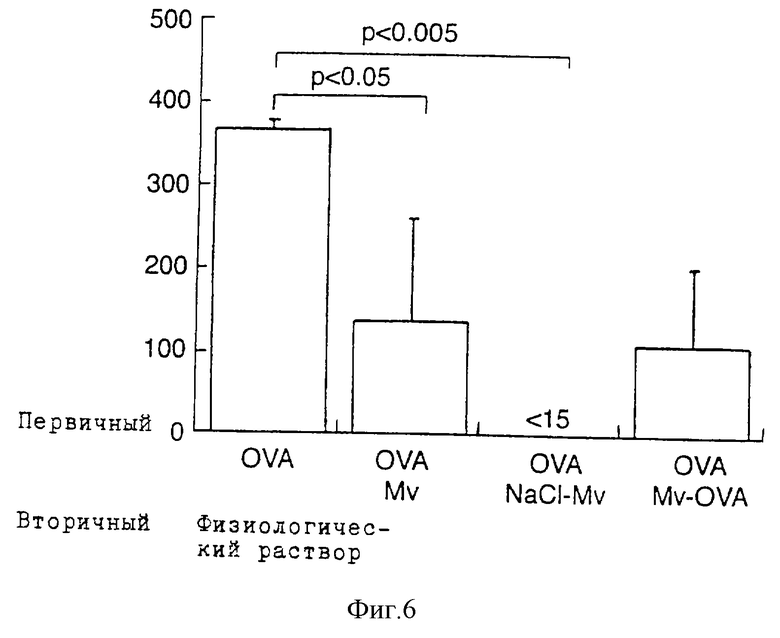
Фиг. 6 показывает продуцирование интерлейкина-5 спленоцитами, культивированными с OVA. Селезенку забирают на 109-й день у мышей, иммунизированных OVA в 0-й и 24-й дни и обработанных на 53-й и 81-й дни солевым раствором, 107 немодифицированными М. vaccae (Mv), OVA, конъюгированным М. vaccae (Mv-OVA) или соответствующими контрольными, промытыми NaCl и обработанными периодатом М. vaccae (NaCl-Mv). Через 48 ч надосадочные жидкости культур собирают для определения интерлейкина 5. Данные представлены в виде средних уровней интерлейкина 5 ± стандартное отклонение (пг/мл).
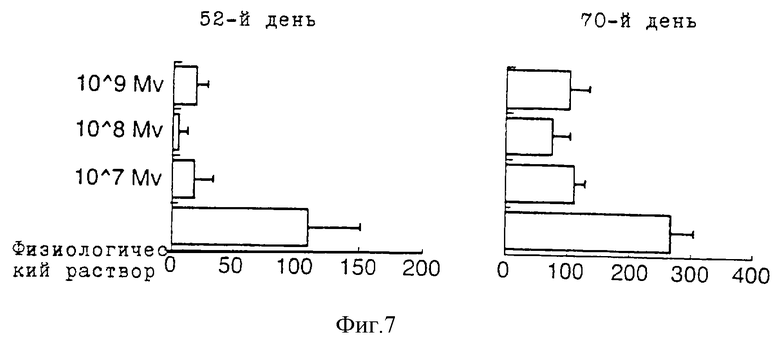
Фиг.7 показывает действие лечения разовыми подкожными инъекциями различных доз М. vaccae по сравнению с солевым раствором на уровень иммуноглобулина Е сыворотки у мышей BALB/c, ранее иммунизированных OVA. Количество иммуноглобулина Е сыворотки выражено в виде изменения (нг/мл) относительно данных, полученных от образцов, взятых на 32-й день, перед обработкой М. vaccae или физиологическим раствором (среднее ± SE). Образцы сыворотки после обработки собирают на 52-й и 70-й дни (т.е. через 10 и 28 дней после обработки физиологическим раствором или М. vaccae на 42-й день). Схемы иммунизации и лечения описаны в экспериментальных методах. Сравнение между различными группами проводилось при помощи U-теста Mann-Whitney.
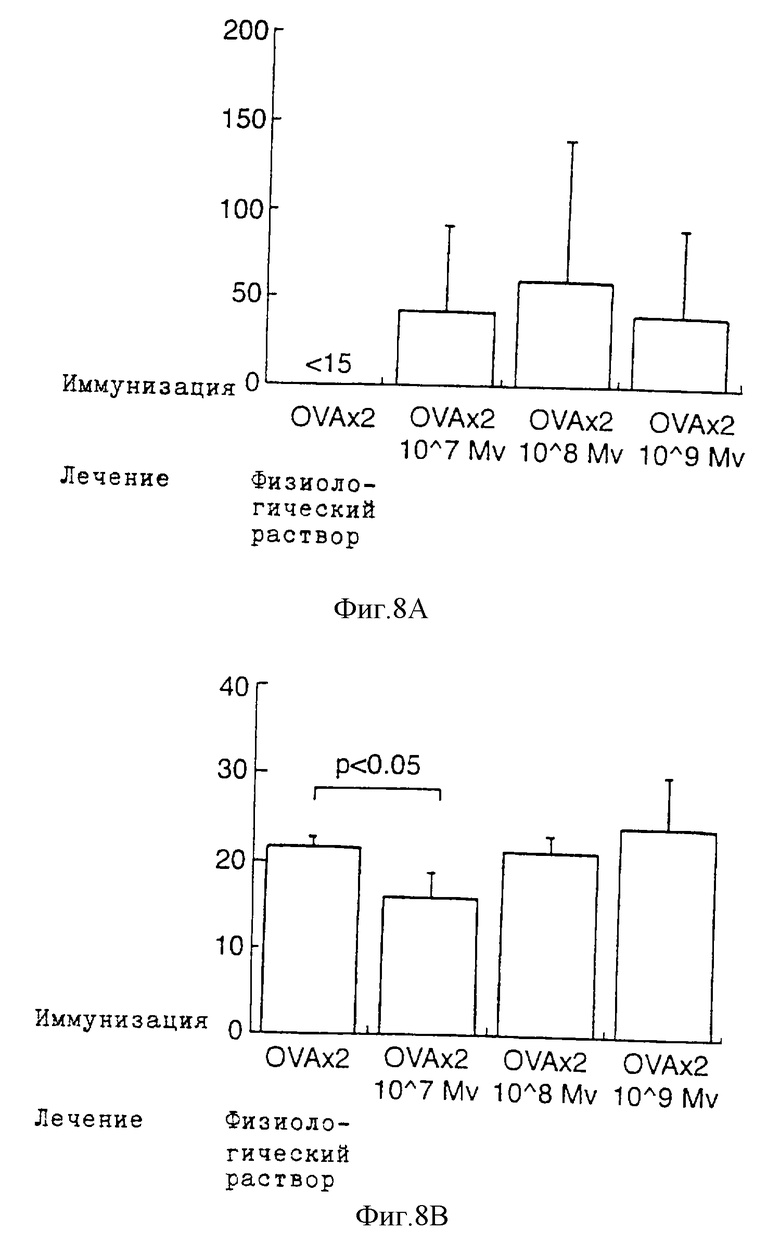
Фиг.8 показывает продуцирование интерлейкина-2 (Фиг.8А) и интерлейкина-4 (Фиг.8В) спленоцитами с OVA. Мыши BALB/c получают OVA дважды перед различными дозами М. vaccae; 107, 108 или 109 (пг/мл). Спленоциты собирают и культивируют с OVA на 82-й день. Схемы иммунизации и лечения описаны в экспериментальных методах. Через 48 ч надосадочные жидкости культур собирают для определения интерлейкина 4, а через 24 ч - интерлейкина-2. Данные представлены в виде среднего уровня цитокина ± стандартное отклонение. Сравнение между различными группами проводят при помощи t-теста Student (физиологический раствор; двухкратная иммунизация с применением OVA перед обработкой физиологическим раствором; 107 Nv: двухкратная иммунизация с применением OVA перед обработкой 107 М. vaccae.
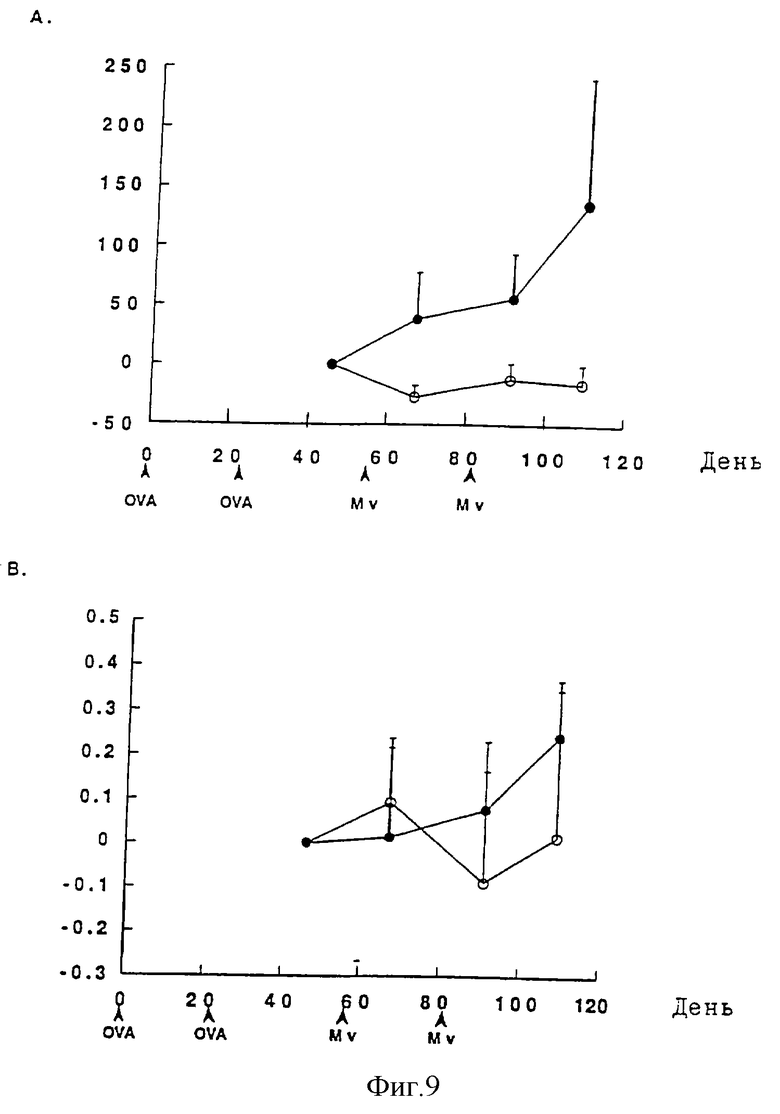
Фиг. 9 показывает действие лечения двумя подкожными инъекциями (53-й и 81-й дни) 107 М. vaccae (o) по сравнению с физиологическим раствором (•) на весь иммуноглобулин Е сыворотки (Фиг.9А) и OVA-специфический иммуноглобулин Е (Фиг.9В). Схемы иммунизации и лечения приведены на фигуре. Количество иммуноглобулина Е сыворотки выражено в виде изменения ±SD относительно данных, полученных от образцов, взятых на 46-й день (нг/мл). Сравнение между различными группами проводят при помощи U-теста Маnn-Whitney (OVA: иммунизация овальбумином, Mv: иммунизация М. vaccae).
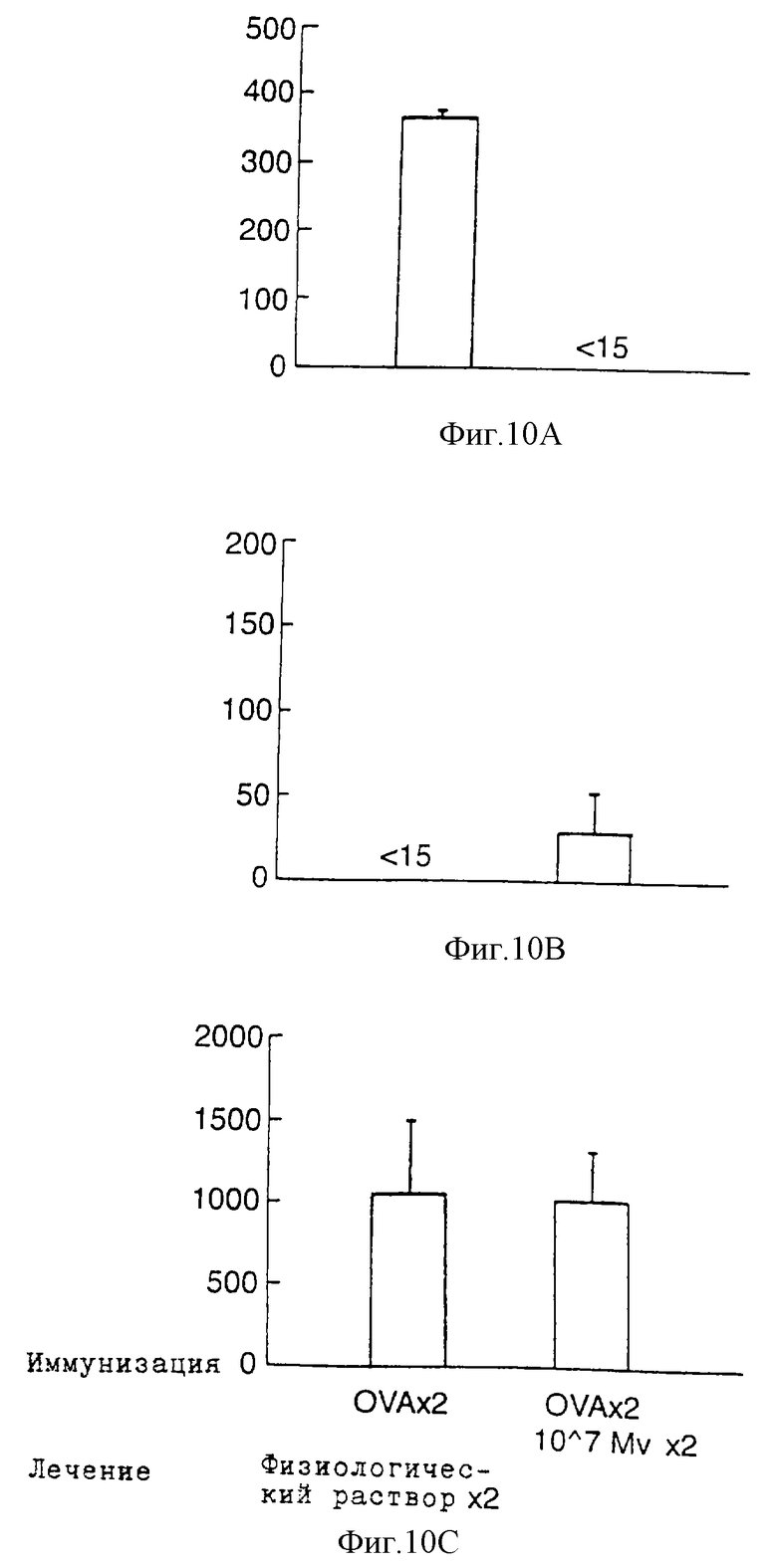
Фиг. 10 показывает продуцирование интерлейкина-5 (Фиг.10А), интерлейкина-2 (Фиг.10В) и гамма-интерферона (Фиг.10С) спленоцитами, культивированными с OVA (пг/мл). Спленоциты собирают и культивируют с OVA на 109-й день. Схемы иммунизации и лечения приведены на фиг. 9. Надосадочные жидкости культур собирают для определения интерлейкина-5 через 48 ч, а гамма-интерферона и интерлейкина-2 - через 24 ч. Данные представлены в виде среднего уровня цитокина ± стандартное отклонение. Сравнение между различными группами проводят при помощи t-теста Student (физиологический раствор: двухкратная иммунизация с применением OVA перед двухкратной обработкой физиологическим раствором, Мv: двухкратная иммунизация с применением OVA перед двухкратной обработкой 107 М. vaccae).
Все документы, упоминаемые в данном описании, приводятся в качестве ссылок.
ПРИМЕР 1
Введение М. vaccae экспериментальным животным и демонстрация достигнутого неспецифического снижения Тh2-активности иммунной системы
Мышей Balb/c в возрасте 6-8 недель в 0-й и 24-й день иммунизируют 50 мкг овальбумина, эмульгированного в масле (неполный адъювант Фрейнда). Известно, что это вызывает сильный Тh2-тип реакции, сопровождаемой продуцированием иммуноглобулина Е и примированием для выделения двух Th2-цитокинов, интерлейкина-4 и интерлейкина-5.
Затем на 53-й и 81-й дни животные получают физиологический раствор или автоклавированные 107 М. vaccae (при помощи подкожных инъекций).
Образцы сыворотки берут на 46-й день (т.е. перед обработкой физиологическим раствором или М. vaccae), а также на 67-й, 91-й и 109-й дни.
Затем клетки селезенки собирают для стимулирования овальбумином в культуральной системе in vitro. Надосадочные жидкости культур собирают через 48 ч для анализа цитокинов твердофазным иммуноферментным способом (ELISA).
Применяют анализ с захватом парами из Pharmingen.
Интерлейкин-5; захват 18051D, детектирование (биотинилированное) 18062D. Связывание детектирующего антитела обнаруживают при помощи конъюгата стрептавидин-пероксидаза из хрена (Dako Р0397) и такого хромогена, как ABTS (Sigma А-1888).
Иммуноглобулин Е; захват 02111D, детектирование (биотинилированное) 02122D. Связывание детектирующего антитела обнаруживают при помощи конъюгата авидин-щелочная фосфатаза (Sigma Е-2636) и такого хромогена, как pNPP (Sigma N-2770).
Уровень иммуноглобулина Е выражен в виде изменения в нг/мл по сравнению с концентрацией сыворотки на 46-й день (последняя величина перед обработкой). Таким образом, величину 46-го дня вычитают из величины в каждом последующем кровопускании.
Результаты
Инъекции М. vaccae снижают подъем уровня иммуноглобулина Е, вызванный иммунизацией овальбумином. Во всех исследованиях снижение, вызванное обработкой М. vaccae, является существенным. Результаты приведены на фиг.1.
Подобным образом клетки селезенки от иммунизированных животных не выделяют интерлейкин-5 in vitro в ответ на овальбумин в том случае, если животные-доноры обрабатывались М. vaccae, в то время как клетки селезенки от иммунизированных животных, обработанных физиологическим раствором, выделяют большое количество интерлейкина 5 в ответ на овальбумин. Результаты приведены на фиг.2. В контрольных лунках, культивируемых без овальбумина, интерлейкин-5 не обнаруживается (данные не приведены).
Заключение
М. vaccae снижает Тh2-тип ответа даже при введении:
(i) после иммунизации сильным аллергеном (в данном случае, овальбумином), и
(ii) без эпитопов Тh2-индуцирующей молекулы. Поэтому имеет место неспецифическая системная регуляция по типу обратной связи активности Тh2-ответа, не зависящая от прямого действия адъюванта на сам аллерген.
ПРИМЕР 2
Действие различных препаратов М. vaccae на иммуноглобулин Е сыворотки
В следующем эксперименте по определению действия конъюгированного М. vaccae-OVA на титр иммуноглобулина Е сыворотки мышей BALB/c дважды иммунизируют с применением OVA в IFA для индуцирования ответов иммуноглобулина Е, затем дважды обрабатывают физиологическим раствором, немодифицированным М. vaccae, конъюгированным М. vaccae-OVA или соответствующим псевдоконъюгированным препаратом, не содержащим OVA, промытым NaCl, обработанным периодатом М. vaccae (NaCl-Mv). На 46-й день мыши, иммунизированные OVA, имеют 112,9±10 (S. E.) нг/мл иммуноглобулина Е по сравнению с 55,4±1,7 (S.E.) нг/мл у неиммунизированных животных (р<0,01). Величину 46-го дня каждой мыши используют для нормализации данных до исходной (т.е. на 46-й день) величины, равной 0, и вычитают ее из последующей величины. Поэтому отложенные величины представляют собой изменения в нг/мл относительно 46-го дня.
Общее количество иммуноглобулина Е в сыворотке контрольных мышей (обработанных физиологическим раствором) постоянно увеличивается на протяжении эксперимента (фиг. 3А- 3С). Происходит супрессия иммуноглобулина Е у мышей, обработанных немодифицированными М. vaccae (фиг.3А). В противоположность этому промытые NaCl, обработанные периодатом препараты, не влияют на супрессию иммуноглобулина Е, независимо от того, конъюгированы ли они с OVA (фиг. 3В) или не конъюгированы (фиг.3С), несмотря на то, что одна доза контрольного NaCl-Mv влияет на временную супрессию общего количества иммуноглобулина Е в сыворотке (р<0,05) (фиг.3С). Эти результаты показывают, что способность регуляции по типу отрицательной обратной связи иммуноглобулина Е устраняется промыванием NaCl или процедурой конъюгирования.
В противоположность этому ни немодифицированные М. vaccae, ни контрольные NaCl-Mv не регулируют по типу отрицательной обратной связи OVA-специфичные иммуноглобулины Е (фиг.4А и 4С). Более того, OVA-специфический ответ иммуноглобулина Е в самом деле усиливается в результате обработки конъюгированными Mv-OVA (фиг.4В).
ПРИМЕР 3
Влияние различных препаратов М. vaccae на продуцирование интерлейкина-2 клетками селезенки
Клетки селезенки от мышей, иммунизированных OVA, собирают на 109-й день и культивируют с OVA in vitro. Клетки селезенки от OVA-сенсибилизированных мышей, получавших физиологический раствор, не продуцируют интерлейкин-2 в ответ на OVA in vitro. Клетки селезенки от иммунизированных мышей, получавших препарат из М. vaccae, не содержащий компонентов OVA, продуцирует обнаруживаемое количество интерлейкина-2 в ответ на OVA, однако увеличение является незначительным. В противоположность этому клетки селезенки от мышей, обработанных конъюгированным препаратом М. vaccae, содержащим OVA, выделяет большое количество интерлейкина-2 в ответ на OVA (p<0,05 по сравнению с реципиентами физиологического раствора) (фиг.5). Эти результаты показывают, что конъюгирование OVA на М. vaccae увеличивает продуцирование интерлейкина-2 в ответ на OVA.
Таким образом, существенного усиления ответа Тh1 на овальбумин не происходит до тех пор, пока овальбумин не включен (конъюгирован) в М. vaccae. Более того, как указано, при включении овальбумина, несмотря на то, ответ Тh1 в этом случае становится в высшей степени существенным, супрессия Th2 не улучшается и может быть снижена (см. другие эксперименты, приводимые в данном описании).
ПРИМЕР 4
Влияние различных препаратов М. vaccae на продуцирование интерлейкина-5 клетками селезенки
Клетки селезенки от мышей, сенсибилизированных OVA, вырабатывают большое количество интерлейкина-5 в ответ на OVA (Фиг.6). Однако все клетки от OVA-иммунизированных мышей, получавших либо немодифицированные, контрольные, промытые NaCl, либо OVA-конъюгированные М. vaccae, выделяют значительно меньше интерлейкина-5 в ответ на OVA. Таким образом, в противоположность результатам, получаемым при определении количества иммуноглобулина Е, ни сама процедура конъюгирования, ни присутствие OVA, конъюгированного с М. vaccae, не оказывает никакого влияния на регуляцию по типу отрицательной обратной связи выделения интерлейкина-5. В любом случае способность "выключать" интерлейкин-5 снижается путем конъюгирования аллергена на М. vaccae, поскольку соответствующим контролем для Mv-OVA является NaCl-Mv (псевдоконъюгированный).
ПРИМЕР 5
Влияние различных доз М. vaccae на иммуноглобулин Е сыворотки у животных
Для определения влияния различных доз М. vaccae на общее количество иммуноглобулина Е в сыворотке мышей BALB/c дважды иммунизируют с использованием OVA в IFA (в 0-й и 21-й дни) для индуцирования ответов иммуноглобулина Е. Им делают кровопускание на 32-й день, а затем вводят физиологический раствор или 107, 108 или 109 М. vaccae на 42-й день.
На 32-й день мыши, иммунизированные OVA, имеют 117,09± 35,81 (S.D.) нг/мл иммуноглобулина Е по сравнению с 69,27±6,09 (S.D.) нг/мл у неиммунизированных животных (р<0,001). Величину 32-го дня по каждой мыши используют для нормализации данных до исходной (т.е. на 32-й день) величины, равной 0, вычитаемой из каждой последующей величины. Поэтому отложенные величины представляют собой изменения в нг/мл относительно 32-го дня.
Ответ иммуноглобулина Е у контрольных мышей (получавших физиологический раствор) еще сильнее увеличивался на 52-й и 70-й дни, когда отбирались дальнейшие пробы (т.е. через 10 и 28 дней после введения, на 42-й день) (aиг.7). В противоположность этому увеличение уровня иммуноглобулина Е подавляется у мышей, получавших 107, 108 и 109 (aиг.7). Все величины р составляют меньше 0,01 у мышей, получавших физиологический раствор и различные дозы М. vaccae.
ПРИМЕР 6
Влияние лечения с применением М. vaccae на продуцирование цитокинов клетками селезенки
Мышей BALB/c подвергают процедурам такого же протокола, как и в примере 5. Затем на 82-й день клетки их селезенок собирают и культивируют с OVA, M. vaccae и СоnА in vitro. В ответ на OVA клетки селезенки от группы, получавшей физиологический раствор, продуцируют интерлейкин-4, но не интерлейкин-2 (Фиг. 8А/В). Клетки селезенки от мышей, иммунизированных OVA, получивших 107 автоклавированных М. vaccae в ответ на OVA, осуществляют синтез интерлейкина-2 и снижают продуцирование интерлейкина-4. При использовании клеток селезенки от мышей, получавших 108 или 109 М. vaccae, также наблюдается синтез интерлейкина-2 в ответ на OVA.
Необходимо отметить, что увеличение продуцирования интерлейкина 2 у мышей, получавших М. vaccae, является незначительным (SD превышает ответы), при этом только у нескольких мышей в группе. Большая часть обработанных мышей не продуцирует никакого интерлейкина-2.
ПРИМЕР 7
Влияние двух доз М. vaccae на иммуноглобулин Е сыворотки
Поскольку 107 М. vaccae оказалась оптимальной дозой для вызова Thl-ответов на ее собственные антигены, а также наиболее эффективной дозой в пилотных экспериментах (Фиг.7), эта доза была выбрана для дальнейших исследований. Мышей BALB/c дважды иммунизируют с применением OVA в 0-й и 24-й дни для индуцирования ответов иммуноглобулина Е и пускают им кровь на 46-й день, а затем дважды вводят им физиологический раствор или 107 М. vaccae на 53-й и 81-й дни. На 46-й день мыши, иммунизированные OVA, имеют 112,9±10 (S.E.) нг/мл иммуноглобулина Е по сравнению с 55,4±1,7 (S.E.) нг/мл у неиммунизированных животных (р<0,01). Величину 46-го дня по каждой мыши используют для нормализации данных до исходной (т. е. на 46-й день) величины, равной 0, вычитаемой из каждой последующей величины. Поэтому отложенные величины представляют собой изменения в нг/мл относительно 46-го дня.
Общий ответ иммуноглобулина Е сыворотки у контрольных мышей (получавших физиологический раствор) на протяжении эксперимента постоянно усиливается. В противоположность
этому у мышей, получавших 107 автоклавированных М. vaccae, увеличение содержания иммуноглобулина Е подавляется (фиг. 9А). Между тем подавление анти-OVA иммуноглобулина Е путем введения М. vaccae в этом исследовании является незначительным (фиг.9В). Однако дальнейшие эксперименты показывают, что OVA-сенсибилизированные мыши BALB/c, получавшие М. vaccae 4 раза, имеют значительно более низкие OVA-специфические титры иммуноглобулина Е.
Лечение с применением М. vaccae не влияет на антитела IgG1 и IgG2a к OVA.
ПРИМЕР 8
Влияние лечения с применением М. vaccae на продуцирование гамма-интерферона, интерлейкина-2 и интерлейкина-5 клетками селезенки
Мышей BALB/c подвергают процедурам такого же протокола, как и в примере 7. Затем на 109-й день клетки их селезенок собирают и культивируют с OVA in vitro. В ответ на OVA клетки селезенки от группы, получавшей физиологический раствор, продуцируют большое количество интерлейкина-5, но не интерлейкина-2. Однако клетки от OVA-иммунизированных мышей, дважды получавших 107 автоклавированные М. vaccae, не выделяют интерлейкин-5 в ответ на OVA (фиг. 10А). Продуцирование интерлейкина-4 в ответ на OVA клетками селезенки от мышей, получивших 2 дозы 107 М. vaccae, оказывается пониженным, но уровень этого цитокина слишком близок к границе детектирования иммуноанализа для того, чтобы быть надежным. Как показано выше (фиг.8А), продуцирование интерлейкина 2, индуцированное OVA, вновь становится детектируемым в надосадочной жидкости из клеток селезенки некоторых мышей, получивших М. vaccae (фиг.10В), хотя и статистически незначимым. Пониженный уровень Th2 наблюдается у животных, не имеющих повышенного уровня интерлейкина 2. Различий в OVA-индуцированном синтезе гамма-интерферона клетками селезенки в различных группах не наблюдается (Фиг.10С).
ОБСУЖДЕНИЕ
Мыши BALB/c, получившие две иммунизации с применением OVA в IFA, развивают типичный ответ типа Th2. Наблюдается повышающийся уровень иммуноглобулина Е сыворотки, а клетки селезенки от этих мышей в ответ на OVA in vitro выделяют интерлейкин-5, но не интерлейкин-2. Наиболее поразительным является тот факт, что этот непрерывный аллерген-специфический ответ у мышей BALB/c регулируется по типу отрицательной обратной связи в результате обработки низкой дозой убитых М. vaccae без какой-либо необходимости в OVA или OVA-эпитопах в препарате из М. vaccae.
Подобным образом наши эксперименты показывают, что высокий уровень индуцированного OVA иммуноглобулина Е в сыворотке подавляется широким спектром доз М. vaccae. Это открытие является неожиданным, поскольку предыдущие исследования по ответу на дозу у мышей показывают, что 107 является оптимальной величиной для индуцирования Th1-ответа на микобактериальные антигены, содержащиеся в М. vaccae без детектируемого Тh2-компонента, в то время как 109 вызывает смешанный Th1+Тh2-ответ. По всем этим причинам вероятно, что воздействие М. vaccae в этой модели не происходит через супрессивное действие Thl-цитокинов. Дальнейший уровень регуляции уровня иммуноглобулина Е может объясняться индукцией факторов, связующих иммуноглобулин Е.
Существенного усиления Th1-ответа на овальбумин не происходит до включения (конъюгирования) овальбумина в М. vaccae, и после его включения, несмотря на то, что Th1-ответ становится чрезвычайно важным, супрессия Th2 не улучшается и может быть понижена.
ПРИМЕР 9
Измерение профилей цитокинов Th1 и Th2
Существуют некоторые данные по профилю цитокинов в синдроме хронического переутомления и в синдроме войны в Персидском заливе, но они противоречивы и способы применения ненадежны, несмотря на то, что общая клиническая картина предлагает повышенное отношение Th2/Thl, как описано выше (Straus, 1996).
Оптимальной технологией для демонстрации этого является проточная цитометрия, позволяющая определить точные количества и процент Т-клеток из периферической крови, спонтанно секретирующих интерлейкин-2 (Th1) либо интерлейкин-4 или интерлейкин-5 (оба Th2), либо они могут быть индуцированы для выделения этих цитокинов после стимулирования in vitro кальциевым ионофором и форболмиристатомацетатом (РМА). (Секреция фактически блокируется ингибитором (моненсин) таким образом, что цитокин накапливается в клетке, где его и определяют (после фиксации и обеспечения проницаемости) при помощи соответствующего моноклонального антитела, меченного флуорохромом).
В результате применения этой технологии Тh2-клетки исключительно редко встречаются в периферической крови нормальных доноров, но обычно присутствуют в крови пациентов с хроническими инфекциями или раком (неопубликованные наблюдения).
Вторым способом является прямое измерение уровня цитокинов в плазме или сыворотке. Интерлейкин-13 может быть обнаружен как представитель Тh2-цитокинов, а гамма-интерферон - как представитель Thl-цитокина.
Профиль цитокинов ветеранов войны в Персидском заливе исследуют с применением одного из этих методов. Определение смещения Thl->Th2 показательно для необходимости введения М. vaccae с целью уменьшения интенсивности симптомов.
ПРИМЕР 10
Препарат М. vaccae для введения
М. vaccae выращивают на твердой среде, включая модифицированную среду Сатона (Boyden et al.), отвержденную 1,3% агара. Среду инокулируют микроорганизмами и аэробно инкубируют в течение 10 дней при 32oС для обеспечения роста микроорганизма. Затем микроорганизмы собирают, взвешивают и суспендируют в разбавителе, получая 100 мг микроорганизмов/мл разбавителя. Затем суспензию вновь разбавляют забуференным физиологическим раствором, получая суспензию, содержащую 10 мг влажного веса (около 1010 клеток) микроорганизмов/мл разбавителя, и помещают в 5-мл пузырьки, содержащие несколько доз. Затем пузырьки, содержащие живые микроорганизмы, подвергают автоклавированию (115-125oС) в течение 10 мин при 69 кРа для умерщвления микроорганизмов. Полученное таким образом терапевтическое средство хранят до использования при 4oС.
ПРИМЕР 11
Применение M. vaccae для лечения расстройств, характеризуемых смещением Тh1/Тh2-цитокинов
0,1 Мл суспензии, полученной в соответствии с примером 3 и содержащей 1 мг влажного веса (около 109 клеток) М. vaccae, подвергают энергичному встряхиванию, а затем сразу же вводят с применением интрадермальной инъекции над левой дельтоидной мышцей.
Уровень Th1- и Тh2-цитокинов у пациента может измеряться периодически для подтверждения сдвига с Тh2 на Th1, в частности регуляции по типу отрицательной обратной связи Тh2-активности, при наблюдении уменьшения интенсивности симптомов пациента.
ПРИМЕР 12
Лечение пациентов с синдромом хронического переутомления с применением М. vaccae
(1) Пациентка, подвергавшаяся лечению с применением SRL-172 от рака чешуйчатых клеток легких и имеющая сопутствующий диагноз синдром хронической усталости, с волнением сообщила об исчезновении симптомов последней после двух инъекций SRL-172. Она почувствовала себя намного лучше, и ее "болячки пропали". Будучи преподавательницей органной музыки, она взяла несколько дополнительных учеников, демонстрируя улучшение своего здоровья. Эта пациентка получала SRL172 на протяжении более 1 года и сообщает о том, что она продолжает чувствовать себя хорошо.
(2) Пациентка имела синдром хронической усталости на протяжении 7-8 лет с симптомами, включавшими общую усталость, мигрени, депрессию, пищевую аллергию и лимфаденопатию. После двух инъекций SRL172 она сообщила об улучшении. Ее заболевания горла, опухшие гланды и пищевая аллергия намного улучшились, и ей потребовалась меньшая доза Тироксина. Она разговаривала намного оживленнее и сказала, что ее депрессия уменьшилась.
(3) Симптомы пациентки выражались в головных болях, депрессии и множественных пищевых аллергиях. До лечения с применением SRL172 безуспешно пробовались многие другие виды лечения. Пытаясь контролировать свои симптомы, она регулярно принимала большие дозы Налкрона (80 капсул Налкрона по 100 мг до и после одного из приемов пищи каждый день). С тех пор, как она получает SRL172 с двухмесячными перерывами, ее симптомы синдрома хронической усталости, а также пищевая аллергия значительно улучшились, и она полагает, что будет чувствовать себя очень хорошо до тех пор, пока она будет продолжать регулярные инъекции.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИЯ, СТИМУЛИРУЮЩАЯ ИММУННЫЙ ОТВЕТ, СОДЕРЖАЩАЯ НАНОЧАСТИЦЫ НА ОСНОВЕ СОПОЛИМЕРА МЕТИЛВИНИЛОВОГО ЭФИРА МАЛЕИНОВОГО АНГИДРИДА | 2005 |
|
RU2379028C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОЛЯРИЗОВАННЫХ ЛИМФОЦИТОВ ДЛЯ МОДЕЛИРОВАНИЯ Th2-ИНДУЦИРОВАННОГО ОТЕКА | 2006 |
|
RU2318525C1 |
| Средство гуминовой природы, обладающее иммуномодулирующим действием | 2022 |
|
RU2783772C1 |
| СРЕДСТВО НА ОСНОВЕ ВОДОРАСТВОРИМЫХ РАСТИТЕЛЬНЫХ ПОЛИСАХАРИДОВ, ОБЛАДАЮЩЕЕ ПРОТИВОАЛЛЕРГИЧЕСКОЙ АКТИВНОСТЬЮ | 2023 |
|
RU2823873C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ АТТЕНУИРОВАННЫЕ ШТАММЫ Streptococcus pneumoniae, И ЕЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2772131C2 |
| КОМПЛЕКС, СОДЕРЖАЩИЙ ОЛИГОНУКЛЕОТИД, ОБЛАДАЮЩИЙ ИММУНОСТИМУЛИРУЮЩЕЙ АКТИВНОСТЬЮ, И ЕГО ПРИМЕНЕНИЕ | 2014 |
|
RU2723943C2 |
| ИНДУКЦИЯ ТОЛЕРАНТНОСТИ К АНТИГЕНАМ ЧЕРЕЗ СЛИЗИСТУЮ | 2006 |
|
RU2420569C2 |
| БИОЛОГИЧЕСКИЕ МАТЕРИАЛЫ И ИХ ПРИМЕНЕНИЕ | 2009 |
|
RU2508296C2 |
| КОМПОЗИЦИИ, СОДЕРЖАЩИЕ Actinidia, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2006 |
|
RU2423139C2 |
| УСИЛЕНИЕ ИММУННОГО ОТВЕТА И НАПРАВЛЕННОСТИ НА МИШЕНЬ С ПОМОЩЬЮ АНТИГЕНОВ И/ИЛИ ЛЕКАРСТВА | 2008 |
|
RU2548685C2 |


Изобретение относится к области медицины и касается применения Mycobacterium vaccae для регуляции по типу отрицательной обратной связи Тh2-активности иммунной системы. Сущность изобретения включает применение убитых клеток Mycobacterium vaccae или их производных, не содержащих какого-либо антигена, иммуногена или аллергена, не относящегося к M.vaccae для лечения расстройства, характеризующегося стимуляцией Th2- активности иммунной системы, путем снижения Th2- активности, без стимуляции Th1- активности иммунной системы, а именно для лечения расстройств, связанных с синдромом хронической усталости, синдромом войны в Персидском заливе или синдромом тотальной аллергии. Преимущество изобретения заключается в том, что препарат без постороннего "чужого" антигена или иммуногена смещает иммунную систему пациента с Th1- активности на Th2- активность. 3 с. и 6 з.п. ф-лы, 10 ил.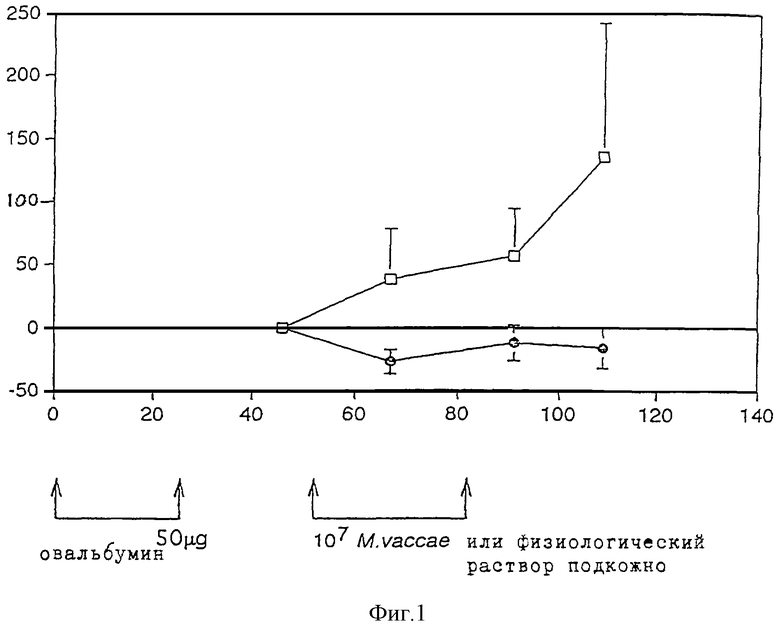
Приоритет по пунктам:
18.12.1996 по пп.1, 2, 4, 5, 7 и 8;
24.12.1996 по пп.3, 6 и 9.
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| Устройство для выпрямления опрокинувшихся на бок и затонувших у берега судов | 1922 |
|
SU85A1 |

