Изобретение относится к медицине и может быть использовано в фармакологии для создания модели для изучения действия веществ на иммунный ответ Th2 типа.
Известен способ (прототип) получения поляризованных лимфоцитов для индукции кожной воспалительной реакции [1], заключающийся в получении поляризованных лимфоцитов Th2 типа in vitro, для чего выделяют спленоциты из селезенок трансгенных (анти-овальбумин ТКР) мышей линии BAlb/с, культивируют 3 суток в присутствии смеси антигена (овальбумина), интерлейкина-4 и антител к интерлейкину-12, после чего подготовленные таким образом клетки вводят интактным сингенным мышам и получают отек.
Недостатком данного способа является то, что он сложен в исполнении и для его осуществления требуются дорогостоящие реактивы (цитокины и антитела к ним) и животные (трансгенные мыши).
Целью предлагаемого изобретения является упрощение способа получения поляризованных лимфоцитов Th2 типа и снижение финансовых затрат.
Поставленная цель достигается тем, что поляризацию лимфоцитов осуществляют in vivo: проводят гипериммунизацию конвенциональных мышей линии BAlb/c путем трехкратного введения под кожу бедра овальбумина по 100 мкг/мышь с адъювантом гидроокисью алюминия по 5 мг/мышь с интервалом в 14 дней. Через 7 дней после последней инъекции осуществляют забор селезенок, которые содержат достаточное количество поляризованных лимфоцитов Th2 типа, что устанавливают по изменению профиля продуцируемых ими цитокинов-интерлейкина-4, интерлейкина-2 и интерферона-γ, а также по способности полученных таким образом лимфоцитов индуцировать отек при введении их в смеси с антигеном (овальбумином) в подушечку лапы интактных сингенных мышей.
Новым в предлагаемом способе является гипериммунизация овальбумином с гидроокисью алюминия путем их трехкратного введения под кожу бедра с интервалом в 14 дней между инъекциями.
Известно, что в основе многих заболеваний лежит нарушение функционирования иммунной системы. Для корректной фармакотерапии этих заболеваний необходимо учитывать их иммунопатогенез, т.е. какой тип иммунного ответа (Тh1 или Th2) развивается в том или ином случае. Поскольку меры, направленные на подавление одного типа иммунного ответа, повлекут за собой стимулирование другого типа иммунного ответа, то знание типа иммунного ответа необходимо для определения стратегии лечения.
Известно, что под действием антигена наивные Т-лимфоциты необратимо дифференцируются в два разных типа - Th1 или Th2, которые различаются по набору секретируемых цитокинов. Для Th2 характерны следующие черты: секреция интерлейкина-4, интерлейкина-5, интерлейкина-10, участие в защите от гельминтов, стимулирование генерации В-лимфоцитов, синтезирующих иммуноглобулины класса Е [2; 3]. Основным фенотипическим признаком Th1 является высокая продукция интерферона-γ, а характерным признаком Th2 - повышенная продукция интерлейкина-4 [4; 5; 6].
Баланс между дифференцировкой Т-хелперов в Th1 или Th2 является определяющим для исхода иммунного ответа [5]. Поляризация по второму типу обеспечивает реакции гиперчувствительности немедленного типа (анафилактические реакции, бронхиальная астма, гельминтозы), характерными признаками которых являются эозинофилия и повышенный синтез иммуноглобулина класса Е [7; 8], либо состояния толерантности (оральная толерантность, беременность, опухолевый рост), для которых характерна гиперпродукция Т-лимфоцитами трансформирующего фактора роста бета и интерлейкин-10 [9; 10].
Особую социально-медицинскую значимость исследованиям, посвященным регуляции функциональной активности поляризованных по второму типу лимфоцитов, придает все более увеличивающееся число пациентов, страдающих атопическими заболеваниями, такими как атонический ринит, конъюктивит, атопический дерматит. Так, например, за последние 30 лет количество пациентов с атопическим дерматитом возросло в 2-3 раза в индустриальных странах и охватывает 10-20% детей и 1-3% взрослых. Еще одним примером нозологической формы, связанной с патологическим Th2 типом ответом, является бронхиальная астма [9], идиопатический пневмофиброз [12].
В связи с изложенным выше представляется чрезвычайно актуальным получить модель иммунного ответа, связанного с поляризацией второго (Th2) типа, позволяющей проводить скрининг препаратов с иммунотропной активностью, направленной на основное патогенетическое звено иммуноглобулин-Е-зависимых аллергических заболеваний - на поляризованные по Th2 типу лимфоциты.
В настоящее время для лечения аллергических заболеваний используется симптоматическая терапия, направленная на блокирование или ингибирование дистальных механизмов, для чего широко применяются препараты с антигистаминным действием, которые либо блокируют H1-рецепторы (димедрол, фенкарол, пипольфен и т.д.), либо тормозят высвобождение и активность гистамина, а также других медиаторов аллергии (кромолин-натрий, незокромил-натрий, кетотифен, оксатомид) [13]. Однако препаратов для патогенетической терапии, направленных на регуляцию активности поляризованных лимфоцитов Th2 типа, не имеется. Для их разработки необходимы соответствующие модели, одна из которых основана на получении у интактных животных кожной воспалительной реакции, индуцированной поляризованными лимфоцитами Th2 типа, для чего требуется получение поляризованных лимфоцитов Th2 типа. Предлагаемый нами способ получения поляризованных лимфоцитов Th2 типа при помощи гипериммунизации значительно упрощает ранее описанный метод [1], позволяет обойтись без дорогостоящих реактивов и животных, а также без культивирования клеток.
Существенные признаки, характеризующие изобретение, проявили в заявляемой совокупности новые свойства, явным образом не вытекающие из уровня техники в данной области и не являющиеся очевидными для специалиста. Идентичного способа получения лимфоцитов, способных развивать локальную реакцию при введении с антигеном, не обнаружено при изучении патентной и научно-медицинской литературы.
Данное изобретение может быть использовано на практике для скрининга новых препаратов с иммунотропным действием, направленным на ключевые звенья патогенеза иммунологической дисфункции.
Исходя из выше изложенного следует считать предлагаемое изобретение соответствующим критериям патентоспособности «Новизна», «Изобретательский уровень», «Промышленная применимость».
Предлагаемый способ осуществляется следующим образом. Конвенциональным мышам линии BAlb/c (в возрасте 8-10 недель весом 18-20 г) под кожу бедра вводят смесь в объеме 0,1 мл изотонического раствора хлорида натрия овальбумина (по 100 мкг/мышь) с гидроокисью алюминия (по 5 мг/мышь) трехкратно с интервалом 14 дней. Через 7 дней после последнего введения животных забивают, в асептических условиях выделяют селезенки, гомогенизируют их в стеклянном гомогенизаторе, трижды отмывают охлажденным изотоническим раствором хлорида натрия, затем дважды фильтруют через четырехслойный капрон и оценивают жизнеспособность клеток в тесте с трипановым синим. Для дальнейших манипуляций берут спленоциты с жизнеспособностью не менее 95%. Для получения отека под апоневротическую пластинку задних лап сингенных интактных мышей вводят смесь таких спленоцитов с антигеном (овальбумин), а в контрольную лапу вводят смесь спленоцитов интактных сингенных мышей с тем же количеством овальбумина. Через 24 часа животных забивают, отрезают лапки на уровне голеностопного сустава, по разнице между массой опытной и контрольной лап судят о количестве поляризованных лимфоцитов.
Полученные в результате гипериммунизации лимфоциты соответствуют поляризации Th2 типа, поскольку 1) при введении животным разрешающей дозы антигена у них развивается анафилактический шок, 2) полученные лимфоциты вырабатывают соответствующий набор цитокинов, 3) полученные лимфоциты индуцируют отек при введении интактным животным совместно с антигеном.
Для получения поляризованных лимфоцитов Th2 типа были использованы мыши линии BAlb/c, поскольку известно, что у этих животных в ответ на антиген развивается преимущественно иммунный ответ Th2 типа [14]. В качестве стандартного антигена, вызывающего поляризацию Th2 типа [15], использовали овальбумин с гидроокисью алюминия в качестве адъюванта. Количество иммунизации, интервал между ними и сроки забора лимфоцитов подобраны экспериментально по результатам анафилактического шока и профиля продуцируемых Т-лимфоцитами цитокинов (интерферона-γ, интерлейкина-2 и интерлейкина-4).
Пример 1.
Для выбора оптимального режима иммунизации, позволяющего получить развитие поляризации Th2 типа in vivo, мышам линии BAlb/c вводили под кожу бедра овальбумин (по 100 мкг/мышь) с гидроокисью алюминия (по 5 мг/мышь) в объеме 0,1 мл один, два или три раза с интервалом между иммунизациями 14 дней. В разные сроки после иммунизации (через 7 или 14 дней) у животных вызывали анафилактический шок, вводя им внутривенно разрешающую дозу антигена (по 10 мкг/мышь овальбумина в 0,1 мл изотонического раствора хлорида натрия в ретроорбитальный синус). Наблюдалось развитие анафилактического шока разной степени тяжести.
Введение внутривенно разрешающей дозы антигена животным на 7-й и 14-й день после однократной иммунизации вызвало у всех животных (n=10 в каждой группе) анафилактическую реакцию слабой степени, что проявлялось в изменении поведения животных (они выглядели угнетенными, вялыми, неподвижными). Указанные изменения появлялись через 5-10 минут после введения антигена и наблюдались на протяжении 1-2 ч.
Введение разрешающей дозы антигена животным на 7-й и 14-й день после второй иммунизации вызывало у всех животных (n=10 в каждой группе) анафилактическую реакцию средней и тяжелой степени выраженности. Все мыши выглядели угнетенными, вялыми, теряли подвижность, их шерстяной покров был взъерошен, конечности цианозны, дыхание поверхностное и учащенное, развивались параличи конечностей, животные принимали боковое положение через 30-40 минут после инъекции. У небольшого количества животных наблюдались клонические судороги. Примерно половина мышей погибало в течение 2-4 ч, в разных сериях эксперимента количество погибших составляло 40-60%.
Введение разрешающей дозы антигена животным на 7-й и 14-й день после третьей иммунизации вызывало у всех животных (n=10 в каждой группе) анафилактическую реакцию средней и тяжелой степени выраженности. Все мыши выглядели угнетенными, вялыми, теряли подвижность, их шерстяной покров был взъерошен, конечности цианозны, дыхание поверхностное и учащенное, развивались параличи конечностей, животные принимали боковое положение через 30-40 минут после инъекции. У небольшого количества животных наблюдались клонические судороги. Большинство мышей погибало в течение 2-4 ч, в разных сериях эксперимента количество погибших составляло 80-100%.
Таким образом, наиболее тяжелый шок наблюдался у мышей, получивших три введения овальбумина.
Пример 2.
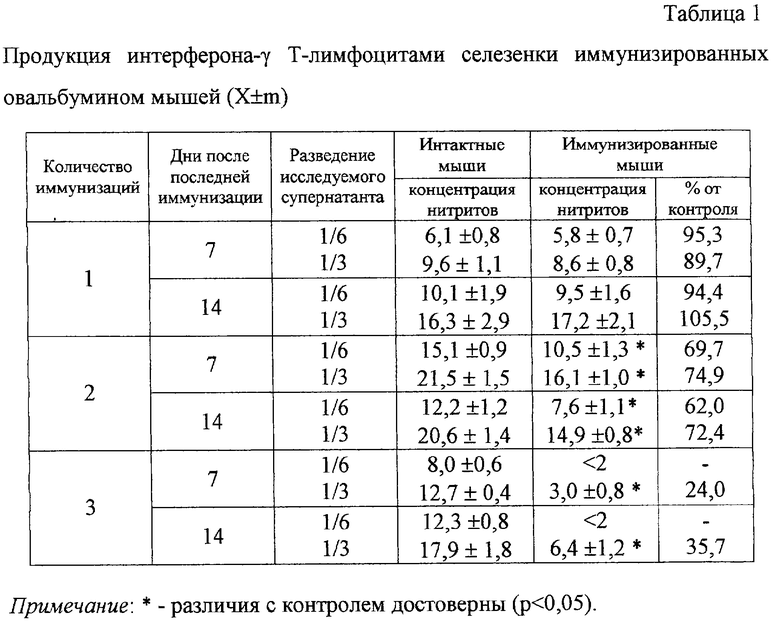
Для выбора оптимального способа получения поляризованных Th2 типа лимфоцитов оценивали функциональное состояние лимфоцитов селезенок иммунизированных мышей по продукции цитокинов (интерферон-γ, интерлейкин-2 и интерлейкин-4). Для этого мышам линии BAlb/c вводили под кожу бедра по 100 мкг овальбумина с 5 мг гидроокисью алюминия в объеме 0,1 мл один, два или три раза с интервалом между иммунизациями 14 дней. Через 7 или 14 дней после последней иммунизации животных забивали, в асептических условиях получали селезенки, гомогенизировали их для получения суспензии клеток. Продукцию цитокинов Т-лимфоцитами селезенки иммунизированных овальбумином мышей определяли в супернатантах, полученных при культивировании спленоцитов 24 ч в концентрации 5×106 клеток/мл в присутствии Т-клеточного митогена (конканавалин A, «Sigma», 4 мкг/мл). Определение продукции интерферона-γ осуществляли в функциональном тесте по способности супернатантов лимфоцитов индуцировать выработку оксида азота миелокариоцитами интактных сингенных животных [16]. Супернатант (1/3 и 1/6 от общего объема) Т-лимфоцитов добавляли в 96-луночные круглодонные планшеты («Costar»), содержащие свежевыделенные клетки костного мозга (по 2-3×105 в лунке) сингенных мышей, культивировали 48 ч, собирали супернатант и измеряли в нем количество оксида азота по содержанию нитритов в супернатантах при помощи реактива Грейса. Определение продукции интерлейкина-2Т-лимфоцитами осуществляли в функциональном тесте по способности их супернатанта стимулировать пролиферацию ИЛ-2-зависимых спленоцитов [16]. Для получения ИЛ-2-зависимых спленоцитов клетки селезенки культивировали в концентрации 4×106/мл с Кон А (4 мкг/мл) 36 ч, затем отмывали митоген центрифугированием и культивировали 48 ч в круглодонных 96-луночных планшетах (5×104 клеток/лунку) с различными разведениями исследуемого супернатанта. Затем оценивали пролиферацию клеток радиоизотопным или колориметрическим методом и выражали соответственно в имп/мин или единицах оптической плотности. При оценке радиоизотопным методом за 16 ч до окончания культивирования клеток в круглодонных 96-луночных планшетах вносили по 0,5 мкКю/лунку 3H-тимидина. Затем содержимое лунок переносили на стекловолокнистые фильтры («Titertek»), промывали изотоническим раствором хлорида натрия, высушивали и переносили в сцинтилляционную жидкость, включение изотопа оценивали на β-счетчике. Пролиферацию клеток выражали в количестве импульсов в минуту. При оценке пролиферации колориметрическим методом клетки культивировали 4 суток в круглодонных 96-луночных планшетах (2-5×104 клеток/лунку) в указанных выше условиях. За 4 ч до окончания культивирования в лунки вносили 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT, «Serva»), концентрация которого в лунках составляла 200 мкг/мл. Затем из лунок удаляли надосадок, а осадок растворяли диметилсульфоксидом (димексид, «Татхимфармпрепараты»). Абсорбцию полученных растворов замеряли при помощи многоканального спектрофотометра («Titertek») при длине волны 550 нм. Определение количества интрелейкина-4 в исследуемых супернатантах осуществляли твердофазным иммуноферментным методом при помощи тест-системы («Amersham Biosciences») согласно прилагаемым к ней протоколам.
Увеличение количества иммунизации приводило к уменьшению выработки интерферона-γ и интерлейкина-2 Т-лимфоцитами селезенки иммунизированных мышей (табл.1 и 2).
Трех иммунизаций оказалось достаточно для получения трехкратного снижения продукции этих цитокинов. Из литературы известно, что для поляризованных лимфоцитов Th2 типа характерно снижение продукции Th1 паттерна цитокинов (основные из которых интерферон-γ и интерлейкин-2) и увеличение выработки цитокинов Th2 типа (основной из которых - интерлейкин-4) [4; 5; 6]. Согласно общепринятым представлениям в наших экспериментах также на фоне понижения выработки интерферона-γ и интерлейкина-2 выявилось увеличение продукции интерлейкина-4 (табл.3).
Таким образом, как показало проведенное исследование функционального состояния лимфоцитов, наиболее оптимальным режимом иммунизации является трехкратное введение выбранного количества овальбумина с интервалом 14 дней, при этом достаточно 7 дней после последней иммунизации.


Пример 3.
Для подтверждения того, что получено достаточное количество поляризованных лимфоцитов Th2 типа через 7 дней после третьего введения овальбумина, полученные таким способом лимфоциты оценивали в адаптивной реакции Th2-индуцированного отека [1]. Для получения такой кожной воспалительной реакции интактным животным под апоневротическую пластинку задних лап вводили по 100 млн сингенных спленоцитов (из пула, состоящего из 3-4 селезенок), полученных от иммунизированных овальбумином мышей, в смеси с антигеном (по 20 мкг овальбумина) в объеме 0,03 мл изотонического раствора хлорида натрия; в противоположную (контрольную) лапу вводили в том же объеме те же количества сингенных спленоцитов (из пула, состоящего из 3-4 селезенок), полученных от интактных мышей, в смеси с теми же количествами антигена. Воспалительную реакцию оценивали через 24 ч, для чего мышей забивали, отрезали обе лапки на уровне голеностопного сустава и взвешивали на торсионных весах. Индекс реакции определяли по формуле:

где О и К - масса соответственно опытной и контрольной лап. Разрешающая доза антигена (20 мкг овальбумина) не вызывала отека при введении в лапу неиммунизированным мышам.
Как видно из таблицы 4, полученные предлагаемым способом поляризованные лимфоциты обладали тем же свойством, что и лимфоциты, полученные в наиболее близком к заявляемому способу (прототипу) - обладали способностью индуцировать Th2-зависимый отек.
Использование предлагаемого изобретения позволяет получить достаточное количество поляризованных лимфоцитов Th2 типа для моделирования иммунного ответа Th2 типа без дорогостоящих реактивов (цитокинов и антител к ним) и животных (трансгенных мышей) и упростить способ за счет исключения стадии культивирования клеток.
Литература
1. Terui T., Sano K., Okada M. et al. Production and pharmacologic modulation of the granulocyte-associated allergic responses to ovalbumin in murine skin models induced by injecting ovalbumin-specific Th1 or Th2 cells // J. Investigative Dermatology. 2001. V.117. P.236-243.
2. Le Gros G., Ben Sasson S.Z., Seder R. et al. Generation of IL-4-producing cells in vivo and in vitro: IL-2 and IL-4 are required for the in vitro generation of IL-4 producing cells // J. Exp. Med. 1990. V.172. P.921-924.
3. Sornasse T., Larenas P.V., Davis K.A. et al. Differentiation and stability of T helper 1 and 2 cells derived from naive human neonatal CD4+ T cells, analyzed at the single-cell level // J. Exp. Med. 1996. V.184. P.473-478.
4. Romagnani S. Human TH1 and TH2 subsets: doubt no more // Immunology Today. 1991. V.12. P.256-260.
5. Abbas A.K., Murphy K.M., Sher A. Functional diversity of helper T lymphocytes // Nature. 1996. V.383. P.787-793.
6. O'Garra A. Cytokines induce the development of functionally heterogeneous T helper cell subsets // Immunity. 1998. V.8. P.275-283.
7. Finkelman F.D., Katona I.M., Urban J.F. et al. IL-4 is required to generate and sustain in vivo IgE responses // J. Immunology. 1988. V.141, №7. P.2335-2341.
8. Clutterbuck E.J., Hirst E.M., Sanderson C.J. Human interleukin-5 (IL-5) regulates the production of cosinophils in human bone marrow cultures: comparison and interaction with IL-1, IL-3, IL-6, and GMCSF // Blood. 1989. V.73, №6. P.1504-1512.
9. Khoury S.J., Hancock W.W., Weiner H.L. Oral tolerance to myelin basic protein and natural recovery from experimental autoimmune encephalomyelitis are associated with downregulation of inflammatory cytokines and differential upregulation of transforming growth factor beta, interleukin 4, and prostaglandin E expression in the brain // J. Exp. Med. 1992. V.176, №5. P.1355-1364.
10. Maeda H., Shiraishi A. TGF-beta contributes to the shift toward Th2-type responses through direct and IL-10-mediated pathways in tumor-bearing mice // J. Immunology. 1996. V. 156,.№1. P.73-78.
11. Krug N., Frew A.J. The Th2 cell in asthma: initial expectation yet to be realised // Clin. Exp. Allergy. 1997. V.27. P.142-148.
12. Smith R.S., Smith T.J., Blieden T.M., Phipps R.P. Fibroblasts as sentinel cells: synthesis of chemokines and regulation of inflammation // Am. J. Pathol. 1997. V.151. P.317-320.
13. Машковский М.Д. Лекарственные средства: Пособие для врачей, пятнадцатое издание, переработанное, исправленное и дополненное. Москва: Новая волна. 2005. С.285-300.
14. Hsieh C.S., Macatonia S.E., O'Garra A., Murphy K.M. Т cell genetic background determines default Т helper phenotype development in vitro // J. Exp. Med. 1995. V.181. P.713-721.
15. Oshiba A., Hamelrnann E., Haczku A. et al. Modulation of antigen-induced В and Т cell responses by antigen-specific IgE antibodies // J. Immunology. 1997. V.159. P.4056-4063.
16. Moore S.C., Theus S.A., Barnett J.B., Soderberg L.S.F. Cytokine regulation of bone marrow natural suppressor cell activity in the suppression of lymphocyte function // Cell. Immunology. 1992. V.141. P.398-408.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРИМЕНЕНИЕ Mycobacterium vaccae ДЛЯ РЕГУЛЯЦИИ ПО ТИПУ ОТРИЦАТЕЛЬНОЙ ОБРАТНОЙ СВЯЗИ Th2-АКТИВНОСТИ ИММУННОЙ СИСТЕМЫ | 1997 |
|
RU2197989C2 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ ПРОТИВОАЛЛЕРГИЧЕСКИМ ДЕЙСТВИЕМ | 2008 |
|
RU2378004C1 |
| Средство гуминовой природы, обладающее иммуномодулирующим действием | 2022 |
|
RU2783772C1 |
| АДЪЮВАНТ | 2020 |
|
RU2736933C1 |
| Средство, обладающее иммуномодулирующей активностью | 2019 |
|
RU2734420C1 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ ПРОТИВОАЛЛЕРГИЧЕСКИМ ДЕЙСТВИЕМ | 2003 |
|
RU2240801C1 |
| СРЕДСТВО НА ОСНОВЕ ВОДОРАСТВОРИМЫХ РАСТИТЕЛЬНЫХ ПОЛИСАХАРИДОВ, ОБЛАДАЮЩЕЕ ПРОТИВОАЛЛЕРГИЧЕСКОЙ АКТИВНОСТЬЮ | 2023 |
|
RU2823873C1 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ ИММУНОМОДУЛИРУЮЩЕЙ АКТИВНОСТЬЮ | 2010 |
|
RU2421232C1 |
| МИКРОФЛЮИДИЗИРОВАННАЯ КОМПОЗИЦИЯ ДЛЯ ИНДУКЦИИ СПЕЦИФИЧНОГО ЦИТОТОКСИЧЕСКОГО Т-ЛИМФОЦИТНОГО ИММУННОГО ОТВЕТА И ЕЕ ИСПОЛЬЗОВАНИЕ ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ | 1995 |
|
RU2201253C2 |
| СПОСОБ ПОЛУЧЕНИЯ ВАКЦИНЫ ДЛЯ ЛЕЧЕНИЯ ТУБЕРКУЛЕЗА И ДРУГИХ ВНУТРИКЛЕТОЧНЫХ ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИЙ И ВАКЦИНА, ПОЛУЧЕННАЯ ПО НАЗВАННОМУ СПОСОБУ | 2001 |
|
RU2262950C2 |
Изобретение относится к медицине, а именно к получению поляризованных лимфоцитов для моделирования Th2-индуцированного отека путем сенсибилизации лимфоцитов овальбумином и может быть использовано в фармакологии. Поляризованные лимфоциты получают in vivo, для чего проводят гипериммунизацию конвенциональных мышей линии BAlb/с трехкратным подкожным введением овальбумина в количестве 100 мкг/мышь и адъюванта гидроокиси алюминия в количестве 5 мг/мышь с интервалом между инъекциями 14 дней. Забор селезенок и выделение лимфоцитов осуществляют через 7 дней после последней инъекции. Изобретение обеспечивает упрощение способа получения поляризованных лимфоцитов Th2 типа, позволяет получить достаточное количество поляризованных лимфоцитов Th2 типа для моделирования иммунного ответа Th2 типа. 4 табл.
Способ получения поляризованных лимфоцитов для моделирования Th2-индуцированного отека путем сенсибилизации лимфоцитов овальбумином, отличающийся тем, что поляризованные лимфоциты получают in vivo, для чего проводят гипериммунизацию конвенциональных мышей линии BAlb/с трехкратным подкожным введением овальбумина в количестве 100 мкг/мышь и адъюванта гидроокиси алюминия в количестве 5 мг/мышь с интервалом между инъекциями 14 дней, забор селезенок и выделение лимфоцитов осуществляют через 7 дней после последней инъекции.
| Terui Т., Sano К., Okada M | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| // J | |||
| Investigative Dermatology | |||
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| V | |||
| Аппарат для испытания прессованных хлебопекарных дрожжей | 1921 |
|
SU117A1 |
| Стеклографический печатный станок с ножной педалью | 1922 |
|
SU236A1 |
| ПРИМЕНЕНИЕ Mycobacterium vaccae ДЛЯ РЕГУЛЯЦИИ ПО ТИПУ ОТРИЦАТЕЛЬНОЙ ОБРАТНОЙ СВЯЗИ Th2-АКТИВНОСТИ ИММУННОЙ СИСТЕМЫ | 1997 |
|
RU2197989C2 |
| ПРИМЕНЕНИЕ Mycobacterium vaccae ДЛЯ РЕГУЛЯЦИИ ПО ТИПУ ОТРИЦАТЕЛЬНОЙ ОБРАТНОЙ СВЯЗИ Th2-АКТИВНОСТИ ИММУННОЙ СИСТЕМЫ | 1997 |
|
RU2197989C2 |
| Способ лечения аллергических заболеваний | 1991 |
|
SU1793923A3 |
| МХРА 04001047, 17.02.2005 | |||
| МХРА 05011399, 31.05.2006. | |||

