Разработка терапевтической вакцины на основе опухолевых клеток базируется по существу на следующих принципах: имеются качественные или количественные различия между опухолевыми клетками и нормальными клетками; иммунная система в принципе обладает способностью распознавать эти различия; иммунная система может быть стимулирована путем активной специфической иммунизации вакцинами таким образом, чтобы на основании этого узнавать эти различия и способствовать их отторжению.
Для того чтобы добиться противоопухолевого ответа, должны быть выполнены, по меньшей мере, два условия: во-первых, опухолевые клетки должны выражать антигены или неоэпитопы, которые не существуют в нормальных клетках. Во-вторых, иммунная система должна быть соответственно активирована, чтобы реагировать на эти антигены. Существенным препятствием при иммунотерапии опухолей является их низкая иммуногенность, прежде всего у людей. Это более удивительно, чем можно было ожидать, так как большое число генетических изменений злокачественных клеток должно было приводить к образованию пептидных неоэпитопов, которые узнаются в окружении с молекулами главного комплекса гистосовместимости (ГКГ-I) цитотоксическими Т-лимфоцитами.
Ранее были обнаружены ассоциированные с опухолью и специфичные к опухоли антигены, которые представляют такие неоэпитопы и поэтому должны были быть потенциальными целями для атаки иммунной системы. То, что иммунной системе все-таки не удается устранить опухоли, которые эти эпитопы выражают, может, очевидно, происходить не из-за недостатка неоэпитопов, а потому что иммунологический ответ на эти неоантигены недостаточен.
Для иммунотерапии рака на клеточной основе были разработаны две общие стратегии: с одной стороны, воспринимаемая иммунотерапия, которая служит in vitro распространению реактивных к опухоли Т-лимфоцитов и их повторному введению больным; с другой стороны, активная иммунотерапия, которая использует опухолевые клетки в ожидании, что при этом вызываются либо новые, либо усиленные иммунные ответы против опухолевых антигенов, которые приводят к системному опухолевому ответу.
Противоопухолевые вакцины, составляющие основу активной иммунотерапии, получали различными способами; примером этого являются облученные опухолевые клетки, которые перемещают с помощью иммуностимулирующих адъювантов, как, например, Corynebacterium parvum или Bacillus Calmette Guerin (BCG), чтобы вызвать иммунные реакции против опухолевых антигенов (Oettgen и Old, 1991).
В последние годы были использованы прежде всего генетически модифицированные опухолевые клетки для активной противораковой иммунотерапии, причем введенные в опухолевые клетки чужеродные гены относятся к трем категориям.
Некоторые использованные для этого опухолевые клетки модифицируют генетически, чтобы продуцировать цитокины. Местное сосуществование опухолевых клеток и цитокинового сигнала должны дать стимул, вызывающий противоопухолевый иммунитет. Обзор, относящийся к применению этой стратегии, представили Pardoll, 1993, Zatloukal и др., 1993, и Dranoff и Mulligan, 1995.
Что касается опухолевых клеток, которые были изменены генетически, чтобы секретировать цитокины, например интерлейкин-2 (IL-2), гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF) или γ-интерферон (IFN-γ), или чтобы выразить состимулирующие молекулы, на экспериментальных моделях животных было показано, что они генерируют сильный противоопухолевый иммунитет (Dranoff и др., 1993; Zatloukal и др., 1995). У человека с обнаруживаемой уже при осмотре опухолью и с развившейся толерантностью к опухоли существенно более сложно, однако, полностью охватить каскады комплексных взаимодействий так, чтобы могла происходить эффективная противоопухолевая реакция. Фактическая эффективность секретирующих цитокин противоопухолевых вакцин для применения на людях еще не доказана.
Другая категория генов, с помощью которых изменяются опухолевые клетки, имея в виду их использование в качестве противоопухолевой вакцины, кодирует так называемые добавочные белки; цель этой постановки задачи состоит в том, что для того, чтобы опухолевые клетки функционировали в представляющих антигены клетках (нео-АПК), заставить их непосредственно генерировать специфичные к опухоли Т-лимфоциты. Пример подобного подхода описывается у Ostrand-Rosenberg, 1994.
Идентификация и выделение опухолевых антигенов (ОА) или производных от них пептидов, описанные, например, Wölfel и др., 1994 а) и 1994 б); Carrel и др. , 1993, Lehmann и др., 1989, Tibbets и др., 1993, или в опубликованных международных заявках на патенты 92/20356, 94/05304, 94/23031, 95/00159, явились предпосылкой для того, чтобы использовать опухолевые антигены в качестве иммуногенов для противоопухолевых вакцин как в виде белков, так и в виде пептидов. Противоопухолевая вакцина в виде опухолевых антигенов как таковых не является, однако, удовлетворительным иммуногеном, чтобы вызвать клеточный иммунный ответ, как это было бы необходимо для элиминирования несущих опухолевый антиген опухолевых клеток; также совместное применение адъювантов предоставляло только относительную возможность для усиления иммунного ответа (Oettgen и Old, 1991).
Третья стратегия активной иммунотерапии для повышения эффективности противоопухолевых вакцин базируется на ксеногенизированных (превратившихся в чужеродные) аутологичных опухолевых клетках. В основе этой концепции лежит предположение, что иммунная система реагирует на опухолевые клетки, которые выражают чужеродный белок, и что в ходе этой реакции вызывается также иммунный ответ против тех опухолевых антигенов (ОА), которые представляются опухолевыми клетками вакцины.
Обзор этих различных вариантов, при которых опухолевые клетки в расчете на усиленную иммуногенность превращаются в чужеродные путем введения различных генов, представлен Zatloukal и др., 1993.
Центральную роль при регуляции специфичного иммунного ответа играет тримолекулярный комплекс, состоящий из компонентов, включающих рецептор антигенов Т-лимфоцитов, молекулу ГКГ и его лиганд, который является происходящим от белка пептидным фрагментом.
Молекулы ГКГ-I (или соответствующие человеческие молекулы, антигены лейкоцитов человека /HLA/) являются пептидными рецепторами, которые при строгой специфичности позволяют осуществить связывание миллионов различных лигандов. Условие для этого предоставляют специфичные к аллелям пептидные мотивы, которые обнаруживают следующие критерии специфичности: пептиды имеют в зависимости от гаплотипа ГКГ-I определенную длину, как правило, от восьми до десяти аминокислотных остатков. Обычно два из положений аминокислот представляют так называемый "якорь", они могут быть заняты отдельной аминокислотой или остатками аминокислот с похожими боковыми цепями. Точное расположение якорных аминокислот в пептиде и требование к их свойствам варьируются гаплотипами ГКГ-I. С-конец пептидных лигандов часто является алифатическим или заряженным остатком. Такие специфичные к аллелям мотивы пептидных лигандов для ГКГ-I до сих пор известны среди прочего для Н-2Кd, Кb, Кk, Кkml, Db, HLA-A*0201, А*0205 и В*2705.
В рамках обмена белка внутри клетки регулярные, дегенерированные и чужеродные генные продукты, например вирусные белки или опухолевые антигены, разлагаются в мелкие пептиды; при этом получают некоторые потенциальные лиганды для молекул ГКГ-I. Таким образом, имеется предпосылка для их представления молекулами ГКГ-I и, как следствие, появление клеточного иммунного ответа, причем до сих пор еще в отдельности не выяснено, как пептиды продуцируются в клетке в качестве лигандов ГКГ-I.
Подход, который использует этот механизм для отчуждения опухолевых клеток, в расчете на усиление иммунного ответа, состоит в том, что опухолевые клетки обрабатывают химическими мутагенами, как, например N-метил-N'-нитрозогуанидин. Это должно привести к тому, что опухолевые клетки мутированных вариантов являют собой происходящие от клеточных белков неоантигены, которые представляют чужеродные генные продукты (Van Pel и Boon, 1982). Так как, однако, мутагенные явления случайно распределяются через геном и, кроме того, чтобы ожидать, что отдельные клетки в результате различных мутагенных явлений также представляют различные неоантигены, этот способ трудно контролировать в качественном и количественном отношении.
При другом подходе опухолевые клетки превращают в чужеродные в результате того, что их подвергают трансфекции генами одного или нескольких чужеродных белков, например геном чужеродной молекулы ГКГ-I или белков ГКГ отличающегося гаплотипа, который затем содержится в определенной форме на клеточной поверхности (заявка на европейский патент 0569678; Plautz и др., 1993; Nabel и др. , 1993). Этот подход основывается на вышеупомянутом положении, что опухолевые клетки, когда они вводятся в виде вакцины из клеточной массы, распознаются как чужеродные с помощью выраженного белка или происходящих от него пептидов или что в случае экспрессии аутологичных молекул ГКГ-I с помощью повышенного количества молекул ГКГ-I на клеточной поверхности оптимизируется представление опухолевого антигена. Изменение опухолевых клеток с помощью чужеродного белка может привести к тому, что клетки представляются в окружении ГКГ происходящими от чужеродного белка пептидами и изменение от "самого себя" к "чужому" происходит в рамках узнавания комплекса ГКГ-пептид. Узнавание белка или пептида как чужеродного имеет последствием то, что в ходе иммунного узнавания вырабатывается иммунный ответ не только по отношению к чужеродному белку, но также и по отношению к собственным опухолевым антигенам опухолевых клеток. В ходе этих процессов активируются представляющие антиген клетки (АПК), которые подвергают процессингу имеющиеся в опухолевой клетке вакцины белки (включая ОА) в пептиды и используют в качестве лигандов для их собственных молекул ГКГ-I и ГКГ-II. Активированные, нагруженные пептидом АПК переходят в лимфатические узлы, где некоторые немногие из простых Т-лимфоцитов узнают в АПК происходящие от ОА пептиды и могут их использовать в качестве стимула для клоновой экспансии, другими словами, для генерации специфичных к опухоли цитотоксических Т-лимфоцитов и Т-клеток-хелперов.
В основе представленного изобретения лежит задача приготовления новой противоопухолевой вакцины на базе превращенных в чужеродные клеток опухоли, с помощью которой может быть вызван активный клеточный противоопухолевый иммунный ответ.
При решении поставленной задачи исходили из следующих соображений: в то время как иммунная система толерантна к незлокачественным нормальным клеткам организма, организм реагирует иммунной защитой на нормальную клетку, когда она, например, вследствие вирусной инфекции синтезирует чужеродные организму белки. Причина этого заключается в том, что молекулы ГКГ-I представляют чужеродные пептиды, которые происходят от чужеродных организму белков. И, как следствие, иммунная система отмечает, что нечто нежелательное, чуждое происходит с этой клеткой. Клетка ликвидируется, АПК активируются и генерируется новый специфический иммунитет против выражающих чужеродные белки клеток.
Хотя опухолевые клетки и содержат соответствующие специфичные к опухоли опухолевые антигены, они, однако, сами по себе недостаточные вакцины, так как из-за незначительной иммуногенности они игнорируются иммунной системой. Если опухолевую клетку нагружают в отличие от известных приготовлений не чужеродным белком, а чужеродным пептидом, то дополнительно к чужеродным пептидам собственные опухолевые антигены клетки также различаются этой клеткой как чужеродные. Путем отчуждения пептидом должно стать возможным, чтобы вызванный чужеродными пептидами клеточный иммунный ответ направлялся против опухолевых антигенов.
Причина недостаточной иммуногенности опухолевых клеток не может быть только качественной проблемой, но является и количественной.
Для происходящего от опухолевого антигена пептида это может означать, что, хотя он и представляется молекулами ГКГ-I, однако, в такой концентрации, которая слишком незначительна, чтобы вызвать клеточный специфичный к опухоли иммунный ответ. Повышение количества специфичных к опухоли пептидов в опухолевой клетке должно было, таким образом, также способствовать превращению опухолевой клетки в чужеродную, что приводит к возникновению клеточного иммунного ответа. В отличие от вариантов, при которых опухолевый антиген или происходящий от него пептид так представлялся на поверхности клетки, что он подвергался трансфекции с помощью ДНК, кодирующей соответствующий белок или пептид, как описано в опубликованных международных заявках на патенты 92/20356, 94/05304, 94/23031 и 95/00159, следовало бы подготовить такую вакцину, которая при однократном получении вызывает эффективный иммунный ответ.
Mandelboim и др. , 1994 и 1995, предложили инкубировать клетки RMA-S с пептидами, происходящими от опухолевых антигенов, чтобы при этом вызвать клеточный иммунный ответ против соответствующих аутогенных опухолевых антигенов. Из предложенных Mandelboim и др. для вакцинирования опухоли клеток принимаются обозначенные RMA-S ((Kärre и др., 1986), чтобы они могли выполнять функции АПК. Они характеризуются такой особенностью, что их молекулы HLA на клеточной поверхности свободны, благодаря дефекту в клеточном механизме транспорта антигенных пептидов (ТАП-механизм, ответственный за процессинг пептидов и их связывание при молекулах HLA). При этом в распоряжении находятся клетки для нагрузки пептидом, они также действуют как бы в качестве представленного носителя для введенного извне пептида. Направленное противоопухолевое действие основывается на возникновении иммунного ответа против представленного в клетках пептида, который предлагается иммунной системе без непосредственного окружения антигенным составом опухолевой клетки.
Изобретение относится к противоопухолевой вакцине для введения больному, состоящей из опухолевых клеток, которые представляются производными от опухолевых антигенов пептидами в окружении HLA и, по меньшей мере, часть которых имеет, как минимум, один гаплотип ГКГ-I больного на клеточной поверхности, и которые были нагружены одним или несколькими пептидами а) и/ или б) таким образом, что опухолевые клетки в окружении с пептидами узнаются как чужеродные иммунной системой больного и вызывают клеточный иммунный ответ, причем пептиды
а) действуют в качестве лигандов для гаплотипа ГКГ-I, одинакового для больного и опухолевых клеток вакцины, и отличаются от пептидов, являющихся производными белков, которые выражаются клетками больного, или
б) действуют в качестве лигандов для гаплотипа ГКГ-I, одинакового для больного и опухолевых клеток вакцины, и происходят от опухолевых антигенов, которые выражаются клетками больного и находятся в опухолевых клетках вакцины в концентрации, которая выше, чем концентрация пептида, происходящего от того же самого опухолевого антигена, который выражался в опухолевых клетках больного.
Молекулы ГКГ человека в соответствии с международными традициями в дальнейшем обозначаются также HLA (антиген лейкоцитов человека).
Под понятием " клеточный иммунный ответ" следует понимать опосредованный цитотоксическими Т-лимфоцитами иммунитет, который вследствие генерирования специфичных к опухоли цитотоксических позитивных к CD8 Т-лимфоцитов и позитивных к CD4 Т-хелперов вызывает разрушение опухолевых клеток.
Действие вакцины из опухолевых клеток согласно изобретению основывается прежде всего на том, что иммуногенное действие имеющегося в опухолевых клетках запаса опухолевых антигенов усиливается пептидом.
Пептиды типа а) в дальнейшем обозначают тоже как "чужеродные пептиды" или "ксенопептиды".
В одном из вариантов осуществления изобретения опухолевые клетки вакцины аутологичны. Причем речь идет о клетках, которые берутся у подвергающегося лечению больного, обрабатываются ex vivo пептидом (пептидами) а) и/или б), при необходимости инактивируются и затем опять вводятся больному. (Способы получения аутологичных противоопухолевых вакцин описываются в международной заявке на патент 94/21808, где ссылаются на их открытие).
В одном из вариантов осуществления изобретения опухолевые клетки являются аллогенными, то-есть они не принадлежат подвергающемуся лечению больному. Применению аллогенных клеток прежде всего отдают предпочтение тогда, когда играют роль экономические соображения; получение индивидуальных вакцин для каждого отдельного больного требует затрат труда и денег, кроме того, возникают трудности с отдельными больными при ex vivo культивировании опухолевых клеток, так как не получают достаточно большого количества опухолевых клеток, чтобы можно было приготовить вакцину. В случае аллогенных опухолевых клеток следует учитывать, что они должны подходить подтипу HLA больного.
При использовании чужеродных пептидов категории а) речь идет в случае аллогенных опухолевых клеток о клетках одной или нескольких клеточных линий, из которых, как минимум, одна линия клеток выражает, как минимум, один, предпочтительно несколько, опухолевых антигенов, которые идентичны опухолевым антигенам подвергающегося лечению больного, то-есть противоопухолевая вакцина соответствует признакам опухоли больного. Вследствие этого обеспечивается то, что вызванный представленным ГКГ-I чужеродным пептидом в опухолевых клетках вакцины клеточный иммунный ответ, который приводит к распространению специфичных к опухоли цитотоксических Т-лимфоцитов и Т-хелперов, направляется также против опухолевых клеток больного, так как они выражают такой же опухолевый антиген, как и клетки вакцины.
Например, если нужно подвергнуть лечению противоопухолевой вакциной согласно изобретению больную раком молочной железы с метастазами, которые обнаруживают мутацию Her2/neu (Allred и др., 1992; Peopoles и др., 1994; Yoshino и др., 1994 a); Stein и др., 1994; Yoshino и др., 1994 б); Fisk и др., 1995; Наn и др., 1995), вводят в качестве вакцины аллогенные, согласованные с гаплотипом HLA больного опухолевые клетки, которые тоже выражают мутированный Her2/neu в качестве опухолевого антигена. Ранее выделяли многочисленные опухолевые антигены и выясняли их связь с одним или несколькими раковыми заболеваниями. Известны другие примеры подобных опухолевых антигенов (Fenton и др. , 1993; Gedde Dahl и др., 1992; Jung и др., 1991; Morishita и др., 1993; Peace и др. , 1991; Skipper и др., 1993), опухолевых антигенов MAGE (Boon и др., 1994; Slingluff и др., 1994; van der Bruggen и др., 1994; международная заявка на патент 92/20356); обзор различных опухолевых антигенов в добавление к этому приводит Carrel и др., 1993.
В таблице приводится обзор известных, применяемых в рамках изобретения опухолевых антигенов и производных от них пептидов.
Опухолевые антигены больного определяют в общем стандартными способами в ходе установления диагноза и плана лечения, например с помощью анализов на основе цитотоксических Т-лимфоцитов со специфичностью к определяющему опухолевому антигену. Такие анализы были среди прочего описаны Hérin и др., 1987; Coulie и др., 1993; Сох и др., 1994; Rivoltini и др., 1995; Kawakami и др. , 1995, а также в международной заявке на патент 94/14459; эти литературные источники также включают различные опухолевые антигены или происходящие от них пептидные эпитопы. Возникающие на поверхности клеток опухолевые антигены могут быть также обнаружены с помощью иммунных анализов на основе антител. Когда опухолевые антигены являются ферментами, например тирозиназами, они могут быть обнаружены с помощью ферментных анализов.
В другом варианте осуществления изобретения может быть использована смесь аутологичных и аллогенных опухолевых клеток в качестве исходного материала для вакцины. Этот вариант осуществления изобретения находит, в частности, применение, когда выраженные больным опухолевые антигены неизвестны или только неполностью охарактеризованы и/или когда аллогенные опухолевые клетки выражают только часть опухолевых антигенов больного. Путем примешивания аутологичных, обработанных чужеродным пептидом опухолевых клеток обеспечивается то, что, по меньшей мере, часть опухолевых клеток вакцины содержит насколько возможно большое количество опухолевого антигена собственно больного. В случае аллогенных опухолевых клеток речь идет о таких, которые в одном или нескольких гаплотипах ГКГ-I совпадают с таковыми для больного.
Пептиды типа а) и б) в соответствии с требованием связываться с молекулой ГКГ-I определяют в отношении их последовательности с помощью подтипа HLA больного, которому должна вводиться вакцина. Определение подтипа HLA больного представляет, таким образом, одно из важнейших условий для выбора или построения подходящего пептида.
При применении противоопухолевой вакцины согласно изобретению в виде аутологичных опухолевых клеток автоматически проистекает подтип HLA через генетически детерминированную специфичность молекулы HLA больного. Подтип HLA больного может быть определен стандартными способами, как, например, с помощью микротеста лимфотоксичности (MLC-тест, MLC = смешанная культура лимфоцитов) (Practical Immunol., 1989). MLC-тест основывается на принципе смешивания выделенных из крови больного лимфоцитов сначала с антисывороткой или моноклональным антителом против определенной молекулы HLA в присутствии кроличьего комплемента (С). Положительные клетки подвергают лизису и окрашивают с помощью индикаторного красителя, в то время, как неповрежденные клетки остаются неокрашенными.
Для определения гаплотипа HLA больного можно также использовать полимеразную цепную реакцию с обратной транскриптазой (ОТ-ПЦР) (Curr. Prot. Mol. Biol. , глава 2 и 15). Для этого у больного берут кровь и выделяют из нее РНК. Эту РНК подвергают сначала обратной транскрипции, благодаря чему образуется кДНК больного. кДНК служит матрицей для ПЦР с праймерными парами, которые специфически способствуют амплификации фрагмента ДНК, относящегося к определенному гаплотипу HLA. Наблюдаемая полоса ДНК при электрофорезе в агарозном геле свидетельствует о том, что больной выражает соответствующую молекулу HLA. Если полосы не наблюдают, значит больной в отношении этого негативен. Для каждого больного ожидаются, по меньшей мере, две полосы.
При применении изобретения в виде аллогенной вакцины используют клетки, из которых, по меньшей мере, часть согласуется, как минимум, с одним подтипом HLA больного. Принимая во внимание по возможности широкую используемость вакцины согласно изобретению, целесообразно исходить из смеси различных линий клеток, которые выражают два или три различных наиболее часто представленных подтипа HLA, причем, в частности, имеются в виду гаплотипы HLA-А1 и HLA-A2. С помощью вакцины на основе смеси аллогенных опухолевых клеток, выражающих эти гаплотипы, можно охватить широкую популяцию больных; таким образом, можно защитить около 70% европейского населения (Mackiewicz и др., 1995).
Определение применяемых согласно изобретению пептидов через подтип HLA характеризует их относительно их якорных аминокислот и их длины; определенные якорные положения и длина гарантируют, что пептиды подходят друг к другу в линии связывания пептидов соответствующих молекул HLA так, чтобы быть представленными на клеточной поверхности образующих вакцину опухолевых клеток таким образом, что клетки узнаются как чужеродные. Вследствие этого стимулируется иммунная система и вызывается клеточная иммунная реакция также против опухолевых клеток больного.
Пептиды, которые в рамках представленного изобретения являются подходящими в качестве чужеродных пептидов согласно категории а), имеются в распоряжении в широком диапазоне. Их последовательность может происходить от встречающихся в природе иммуногенных белков или их клеточных продуктов расщепления, например вирусных или бактериальных пептидов, или от чужеродных больному опухолевых антигенов.
Подходящие чужеродные пептиды могут, например, быть выбраны на основе известных в литературе пептидных последовательностей, например, описанных Rammensee и др., 1993, Falk и др., 1991, для различных мотивов HLA, на основе производных пептидов от иммуногенных белков различного происхождения, которые подходят друг другу в линиях связывания молекул соответствующих подтипов HLA. Для пептидов, имеющих частичную последовательность белка с иммуногенным действием, можно на основе уже известной или при необходимости еще требующих определения полипептидных последовательностей твердо установить путем регулирования последовательностей, учитывая специфичные к HLA требования, какие пептиды являются подходящими кандидатами. Примеры подходящих пептидов встречаются, например, у Rammensee и др., 1993, Falk и др., 1991, и у Rammensee и др., 1995, а также в международной заявке на патент 91/09869 (ВИЧ-пептиды); производные от опухолевых антигенов пептиды были среди прочих описаны в опубликованных международных заявках на патенты 95/00159, 94/05304. Имеются ссылки на публикации этих литературных источников и цитируемые в них статьи, связанные с пептидами.
Предпочтительными кандидатами для ксенопептидов являются пептиды, иммуногенность которых уже была продемонстрирована, а также пептиды, являющиеся производными известных иммуногенов, например, вирусных или бактериальных белков. Подобные пептиды демонстрируют на основе их иммуногенности сильную реакцию в MLC-тесте.
Вместо того, чтобы применять подлинные пептиды, а также неизмененные, происходящие от природных белков, можно на основании указанных минимальных требований к последовательности подлинного пептида в отношении якорных положений и длины использовать любые варианты, в этом случае применяются также синтетические пептиды согласно изобретению, которые получают в соответствии с требованиями к лиганду ГКГ-I. Так могут, например, исходя из лиганда для типа H2-Kd Leu Phe Glu Ala Ile Glu Gly Phe Ile (LFEAIEGFI), быть изменены аминокислоты, не являющиеся якорными аминокислотами, чтобы получить пептид с последовательностью Phe Phe Ile Gly Ala Leu Glu Glu Ile (FFIGALEEI); кроме того, якорная аминокислота Ilе в положении 9 может быть замещена Leu.
Пептиды, происходящие от опухолевых антигенов, а также от белков, которые выражаются в опухолевой клетке и которые не содержатся в соответствующей нетрансформированной клетке или содержатся в значительно уменьшенной концентрации, могут применяться в рамках представленного изобретения как пептиды типа а) и/или типа б).
Длина пептида соответствует предпочтительно в отношение связывания на молекуле ГКГ-I требуемой минимальной последовательности от 8 до 10 аминокислот с необходимыми якорными аминокислотами. При необходимости пептид может быть удлинен также по С- и/или N-концу, если это удлинение не наносит вреда связывающей способности, или удлиненный на минимальную последовательность пептид может участвовать в клеточном процессинге.
В одном из вариантов осуществления изобретения пептид может быть удлинен с помощью отрицательно заряженных аминокислот или можно вставить отрицательно заряженные аминокислоты в пептид и по другим положениям, чем положения якорных аминокислот, чтобы достигнуть электростатического связывания пептида на поликатионе, например полилизине.
Под понятие "пептиды" в рамках представленного изобретения подпадают в соответствии с определением также более длинные фрагменты белков или полные белки, для которых гарантировано, что после применения АПК они превращаются в пептиды, которые подходят молекуле ГКГ.
В этом варианте осуществления изобретения антиген, таким образом, вставляется не в виде пептида, но в виде белка или фрагмента белка или в виде смеси белков или фрагментов белков. Белок представляет антиген или опухолевый антиген, от которого происходят полученные после процессинга фрагменты. Относящиеся к клеткам белки или фрагменты белков подвергаются процессингу и могут после этого в окружении ГКГ представлять иммуноэффекторные клетки и, таким образом, вызывать или усиливать иммунный ответ (Braciale и Braciale, 1991; Kovacsovics Bankowski и Rock, 1995; York и Rock, 1996).
В случае применения белков или белковых фрагментов можно подтвердить идентичность конечных продуктов после процессинга с помощью химического анализа (деградация по Эдману или масс-спектрометрия фрагментов после процессинга; сравните обзорную статью Rammensee и др., 1995, а также цитируемые в ней оригинальные источники) или биологических анализов (способность АПК к стимуляции Т-лимфоцитов, специфичных к процессированным фрагментам).
Выбор кандидатов в пептиды с точки зрения их пригодности в качестве чужеродных пептидов происходит в принципе в несколько этапов: в общем, целесообразно для серийных исследований сначала проверить кандидаты в тесте по связыванию пептида на способность связываться с молекулой ГКГ-I.
Подходящим способом исследования является, например, основывающийся на проточной цитометрии анализ, использующий установку для сортировки клеток с возбуждением флуоресценции (FACS-анализ)(Flow Cytometry, 1989; FACS VantageTM User's Guide, 1994; CELL QuestTM User's Guide, 1994). Причем пептид метят флуоресцентным красителем, например флуоресцеинизотиоцианатом (ФИТЦ), и вносят в опухолевые клетки, которые выражают соответствующую молекулу ГКГ-I. При протекании отдельные клетки подвергаются воздействию лазерного луча определенной длины волны; измеряют испускаемую флуоресценцию, она зависит от количества связанного клеткой пептида.
Другим способом определения связанного количества пептида является Скэтчард-блот. Для этого используют пептид, меченный 125J или ионами редкоземельных металлов (например, европием). Клетки нагружают при 4oС различными определенными концентрациями пептида от 30 до 240 минут. Для определения неспецифичного обменного взаимодействия пептида с клетками прибавляют к отдельным пробам избыток немеченого пептида, который препятствует специфическому взаимодействию меченого пептида. Затем клетки промывают, при этом удаляется неспецифичный ассоциированный с клетками материал. Количество связанного клеткой пептида устанавливают теперь либо в сцинтилляционном счетчике на основании измеренной радиоактивности, либо в подходящем для измерения долгоживущей флуоресценции фотометре. Обработку полученных таким образом данных производят стандартными способами.
При втором подходе кандидаты с подходящими свойствами для связывания испытываются на их иммуногенность.
Иммуногенность ксенопептидов, происходящих от белков, чье иммуногенное действие неизвестно, может, например, быть исследована с помощью MLC-теста, пептиды, используемые в этом тесте, который также целесообразно проводить в серии с различными пептидами, причем в качестве стандарта целесообразно применять пептид с известным иммуногенным действием, вызывают особенно сильную реакцию и пригодны для рассматриваемого изобретения.
Другая возможность испытания связывающих ГКГ-I пептидных кандидатов на их иммуногенность состоит в том, чтобы исследовать связывание пептидов при Т2-лимфоцитах. Подобный тест основывается на характерной особенности Т2-лимфоцитов (Alexander и др., 1989) или клеток RMA-S (Kärre и др., 1986) участвовать в недостаточном количестве в механизме транспорта антигенный пептид - пептид и сначала, кроме того, представлять стабильные молекулы ГКГ-I. когда на них наносят пептиды, которые представляются в окружении ГКГ-I. Для теста используют, например, Т2-лимфоциты или клетки RMA-S, которые стабильно подвергаются трансфекции с помощью гена HLA, например генов HLA-A1 и/или HLA-A2. Если клетки нагружают пептидами, которые являются хорошими лигандами ГКГ-I, так как они так представляются в окружении ГКГ-I, что могут узнаваться иммунной системой как чужеродные, то такие пептиды способствуют тому, что молекулы HLA обнаруживаются в значительном количестве на клеточной поверхности. Доказательство наличия антигенов лимфоцитов человека на клеточной поверхности, например, с помощью моноклональных антител позволяет идентифицировать подходящие пептиды (Malnati и др., 1995; Sykulev и др., 1994). В этом случае также целесообразно применить стандартный пептид с известной хорошей способностью к связыванию HLA или ГКГ.
В одном варианте осуществления изобретения аутологичная или аллогенная опухолевая клетка вакцины может иметь несколько ксенопептидов с различными последовательностями. Примененные пептиды могут в данном случае, с одной стороны, так отличаться, что они связываются при различных подтипах HLA. Этим может достигаться то, что охватываются некоторые или все подтипы HLA больного или большой группы больных. Вакцину вводят в форме, полученной после облучения.
Другая, при необходимости дополнительная изменчивость в отношении представленных в опухолевой клетке ксенопептидов может состоять в том, что пептиды, связанные с определенным подтипом HLA, различаются относительно их, не являющейся важной для связывания HLA, последовательности тем, что они, например, происходят от белков различного происхождения, например вирусных и/ или бактериальных белков. От одной такой изменчивости, которая предоставляет вакцинированному организму большой диапазон при отчуждении, можно ожидать усиления стимуляции иммунного ответа.
В варианте осуществления изобретения, когда противоопухолевая вакцина состоит из смеси аллогенных опухолевых клеток различных клеточных линий, а также при необходимости дополнительно из аутологичных опухолевых клеток, могут все без исключения опухолевые клетки быть обработаны одинаковым/одинаковыми пептидом/пептидами или опухолевые клетки различного происхождения могут также иметь в каждом случае различные ксенопептиды.
В рамках проведенных исследований по представленному изобретению применяли в качестве чужеродного пептида типа а) вирусный пептид с последовательностью Leu  Glu Ala Ile Glu Gly Phe
Glu Ala Ile Glu Gly Phe  , происходящий от гемагглютинина вируса гриппа и являющийся лигандом для типа H2-Кd; якорные аминокислоты подчеркнуты.
, происходящий от гемагглютинина вируса гриппа и являющийся лигандом для типа H2-Кd; якорные аминокислоты подчеркнуты.
С помощью этого встречающегося в природе вирусного пептида в качестве чужеродного пептида была получена противоопухолевая вакцина и испытана на животной модели (модель меланомы и модель рака толстого кишечника).
Другой вирусный пептид с последовательностью Ala Ser Asn Glu  Met Glu Thr
Met Glu Thr  , который происходит от нуклеопротеина вируса гриппа и является лигандом гаплотипа HLA-1 H2-Kb (Rammensee и др., 1993; якорные аминокислоты подчеркнуты), был использован для получения противоопухолевой вакцины; защитное действие вакцины было подтверждено на другой модели меланомы.
, который происходит от нуклеопротеина вируса гриппа и является лигандом гаплотипа HLA-1 H2-Kb (Rammensee и др., 1993; якорные аминокислоты подчеркнуты), был использован для получения противоопухолевой вакцины; защитное действие вакцины было подтверждено на другой модели меланомы.
Была получена другая вакцина таким образом, что опухолевые клетки превращались в чужеродные с помощью чужеродного пептида с последовательностью Phe Phe Ile Gly Ala Leu Glu Glu Ile (FFIGALEEI). При этом речь идет о синтетическом, неизвестном ранее в природе пептиде. Поэтому при выборе последовательности было важно, чтобы выполнялись требования относительно пригодности в качестве лиганда для молекулы ГКГ-I типа Н2-Кd. Пригодность пептида для того, чтобы вызвать противоопухолевый иммунитет согласно концепции активной иммунотерапии, была подтверждена на раке толстого кишечника мыши СТ-26 ( сингенный для мышиного штамма Balb/c).
В другом варианте осуществления изобретения противоопухолевая вакцина может содержать, кроме того, аутологичные и/или аллогенные опухолевые клетки и/или фибробласты, которые подвергаются трансфекции с помощью цитокиновых генов. В международной заявке на патент 94/21808, а также в работе Schmidt и др. , 1995 (на эту публикацию ссылаются), описываются эффективные противоопухолевые вакцины, которые были получены по способу транспорта ДНК, обозначенному как "трансферрин-трансфекция", с помощью экспрессирующего IL-2 вектора (этот способ основывается на опосредованном рецептором эндоцитозе и использует клеточный лиганд, конъюгированный с поликатионом, как, например, полилизин, в частности трансферрин, для образования комплекса с ДНК, а также эндосомолитически эффективный реагент, например аденовирус).
Предпочтительно смешивают обработанные пептидом опухолевые клетки и выражающие цитокин клетки в соотношении 1:1. Когда, например, смешивают секретирующую IL-2 вакцину, которая продуцирует 4.000 единиц IL-2 на 1•106 клеток, с 1•106 обработанных пептидом опухолевых клеток, можно полученную таким образом вакцину использовать для двух обработок, причем допускается оптимальная дозировка от 1.000 до 2.000 единиц IL-2 (Schmidt и др., 1995).
При комбинации цитокиновой вакцины с обработанными пептидом опухолевыми клетками может быть выгодно объединено действие обоих типов вакцины.
Обработку клеток, а также приготовление готовой лекарственной формы вакцины согласно изобретению осуществляют обычным способом, как, например, описано в работе "Биологическая терапия рака", 1991, или в международной заявке на патент 94/21808.
В другом аспекте изобретение относится к способу получения противоопухолевой вакцины, состоящей из опухолевых клеток, для введения больному.
Способ в соответствии с изобретением отличается тем, что опухолевые клетки, которые представляются производными от опухолевых антигенов пептидами в окружении HLA и, по меньшей мере, часть которых выражает, как минимум, один гаплотип ГКГ-I больного, обрабатывают одним или несколькими пептидами, которые
а) действуют в качестве лигандов для гаплотипа ГКГ-I, одинакового для больного и опухолевых клеток вакцины, и отличаются от пептидов, происходящих от белков, которые выражаются клетками больного или которые
б) действуют в качестве лигандов для гаплотипа ГКГ-I, одинакового для больного и опухолевых клеток вакцины, и происходят от опухолевых антигенов, которые выражаются клетками больного,
причем опухолевые клетки инкубируют с одним или несколькими пептидами а) и/или б) до тех пор и в таком количестве в присутствии органического поликатиона, пока пептиды свяжутся на опухолевых клетках таким образом, что они в окружении с опухолевыми клетками узнаются как чужеродные иммунной системой больного и вызывают клеточный иммунный ответ.
Количество пептида составляет преимущественно от примерно 50 мкг до примерно 160 мкг на 1•105 - 2•107 клеток. В случае использования пептида категории б) концентрация может также быть выше. Для этих пептидов важно, чтобы их концентрация в опухолевых клетках вакцины по отношению к концентрации пептида, происходящего от того же самого опухолевого антигена, в опухолевых клетках больного до такой степени увеличивалась, чтобы опухолевые клетки вакцины узнавались как чужеродные и вызывали клеточный иммунный ответ.
Подходящими поликатионами считаются гомологичные органические поликатионы, как, например, полилизин, полиаргинин, полиорнитин, или гетерологичные поликатионы с двумя или несколькими различными положительно заряженными аминокислотами, причем эти поликатионы могут иметь различную длину цепей, далее непептидные синтетические поликатионы, как полиэтиленимин, природные, связывающие ДНК белки поликатионного характера, например гистоны или протамины, или их аналоги, или их фрагменты, а также спермин или спермидины. Подходящими органическими поликатионами в рамках представленного изобретения считаются также липиды в виде поликатионов (Felgner и др., 1994; Loeffler и др. , 1993; Remy и др., 1994; Behr, 1994), которые, среди прочего, имеются в продаже, как трансфектам, липофектамин или липофектин.
В качестве поликатиона вставляют предпочтительно полилизин (пЛ), длина цепи которого составляет от примерно 30 до примерно 300 остатков лизина.
Необходимое количество поликатиона по отношению к пептиду может быть определено, в частности, эмпирически. В случае использования полилизина и ксенопептидов категории а) соотношение масс пЛ: пептид составляет предпочтительно примерно от 1:4 до примерно 1: 12.
Продолжительность инкубирования составляет в общем от 30 минут до 4 часов. Она еще регулируется в зависимости от того, к какому моменту времени достигается максимальная нагрузка пептидом; за степенью нагрузки можно следить с помощью анализа, использующего установку для сортировки клеток с возбуждением флуоресценции (FACS-анализ), и таким способом определяют необходимую продолжительность инкубирования.
В другом варианте осуществления изобретения вставляют полилизин в, как минимум, частично конъюгированной форме. Предпочтительно часть полилизина используется в конъюгированной с трансферрином (ТФ) форме (конъюгат трансферрина-полилизина, ТФпЛ, сюда также относятся данные, на которые ссылаются в международной заявке на патент 94/21808), причем массовое соотношение пЛ: ТФпЛ составляет около 1:1.
Вместо трансферрина полилизин может быть конъюгирован с другими белками, например с описанными в международной заявке на патент 94/21808 в качестве факторов интернализации клеточными лигандами.
При необходимости обработку опухолевых клеток проводят еще в присутствии ДНК. Целесообразно, чтобы ДНК существовала в виде плазмиды, предпочтительно плазмиды, независимой от последовательностей, которые кодируют функциональные эукариотные белки, а также в виде контрольного вектора. Как ДНК может быть использована в принципе каждая применяемая обычно функционально полученная плазмида.
Соотношение количества ДНК и поликатиона, при необходимости частично конъюгированного с белком, например пЛ, ТФпЛ или смеси пЛ и ТФпЛ, составляет предпочтительно примерно от 1:2 до примерно 1:5.
Продолжительность инкубирования, количество и вид поликатиона по отношению к числу опухолевых клеток и/или количеству пептида или, вернее, какая часть поликатиона или с каким белком выгодно конъюгируется, преимущество присутствия ДНК или ее количество могут быть определены эмпирически. Для этого варьируют отдельные параметры способа и пептиды в идентичных условиях доставляют в опухолевые клетки и контролируют, насколько эффективно пептиды связываются на опухолевых клетках. Для этого подходящим способом является FACS-анализ.
Способ по изобретению пригоден, кроме обработки опухолевых клеток, также и для обработки других клеток.
Вместо опухолевых клеток аутологичные, а также собственные фибробласты больного или клетки линий клеток фибробластов, которые либо согласуются с подтипом HLA больного, либо подвергаются трансфекции соответствующим геном ГКГ-I, могут быть нагружены согласно способу по изобретению одним или несколькими пептидами, происходящими от опухолевых антигенов, которые выражаются опухолевыми клетками больного. Обработанные таким образом и облученные фибробласты могут сами по себе или в смеси с обработанными пептидом опухолевыми клетками быть применены в качестве противоопухолевой вакцины.
В другом варианте осуществления изобретения вместо фибробластов способом согласно изобретению могут быть обработаны дендритные клетки. Дендритные клетки являются АПК кожи, они могут быть на выбор нагружены in vitro, т.е. выделенные у больного клетки смешивают in vitro с одним или несколькими пептидами, причем пептиды происходят от опухолевых антигенов больного и связываются на молекуле ГКГ-I или ГКГ-II больного. В другом варианте осуществления изобретения эти клетки также in vivo могут быть нагружены пептидом. Для этого инъецируют комплексы из пептида, поликатиона и при необходимости ДНК предпочтительно внутрикожно, поскольку дендритные клетки особенно часто обнаруживаются в коже.
В рамках представленного изобретения получали комплекс из пептида с ТФпЛ или пЛ для переноса в клетки СТ-26 и с ТФпЛ и нефункциональной плазмидой (контрольный вектор) для переноса в клетки М-3. В системе СТ-26 было твердо установлено, что подвергнутая отчуждению с помощью пептида облученная противоопухолевая вакцина генерировала эффективный иммунитет против опухоли: 75% вакцинированных мышей могли элиминировать симптоматику опухоли, которая у всех контролируемых животных, либо не получавших вакцины, либо получавших вакцину без ксенопептида, приводила к образованию опухоли. В системе М-3 такой же ксенопептид в условиях, обнаруживающих еще более высокую вероятность образования опухоли для организма, испытывали в эксперименте, повторявшем ситуацию для людей. Мышей с метастазами вакцинировали облученными клетками М-3 с ксенопептидом. У 87,5% вакцинированных таким образом мышей смогли исчезнуть метастазы, в то же время всем необработанным мышам и 7 из 8 мышей с опухолями вводили вакцину без ксенопептида.
Кроме того, было твердо установлено, что величина системного иммунного ответа при использовании противоопухолевой вакцины зависит от способа, с помощью которого пептид доставляется в опухолевые клетки. Когда пептид поставляли клеткам с помощью полилизина/трансферрина, эффект был более четко выражен, чем тогда, когда клетки инкубировали 24 часа с пептидом ("импульсы"). Также мало эффективной была адъювантная добавка пептида к облученным вакцинам. При трансферрин-трансфекции может быть гарантировано либо эффективное поступление пептида в клетки, либо это еще способствует такой нагрузке полилизином/трансферрином, чтобы пептид оставался прикрепленным на клеточной мембране таким образом, чтобы физически находиться близко к молекуле ГКГ-I и чтобы он затем мог с ней связаться, причем, чтобы он мог на основании его сильного сродства вытеснить более слабо связанные клеточные пептиды.
Описание чертежей
Фиг.1а-в: FACS-анализ клеток М-3, обработанных чужеродным пептидом.
Фиг.1г: Микрофотографии обработанных ФИТЦ-пептидом клеток М-3.
Фиг.2а,б: Излечение мышей DBA/2 с метастазами М-3-меланомы с помощью вакцины из нагруженных чужеродным пептидом клеток М-3.
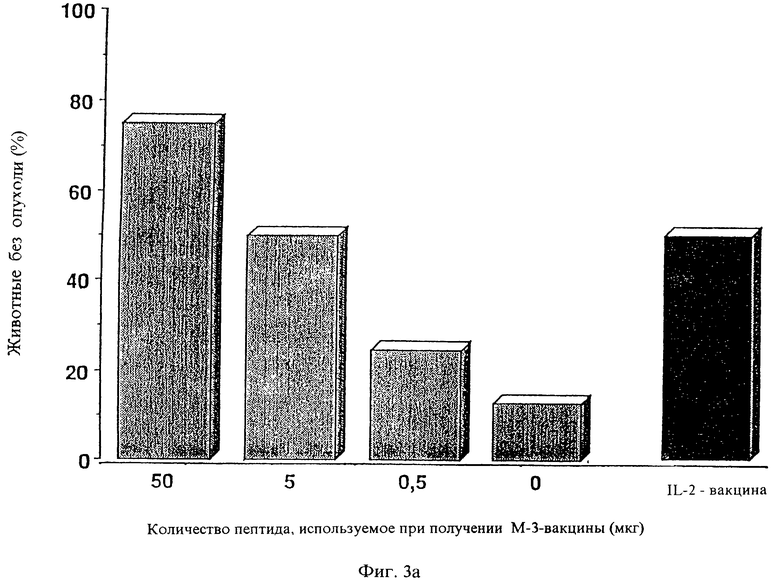
Фиг. 3а: Титрование чужеродного пептида для получения противоопухолевой вакцины.
Фиг. 3б: Сравнение противоопухолевой вакцины из нагруженных чужеродным пептидом опухолевых клеток с секретирующей IL-2 противоопухолевой вакциной.
Фиг. 4а: Защита мышей Balb/c путем предварительной иммунизации вакциной из нагруженных чужеродным пептидом клеток рака толстого кишечника.
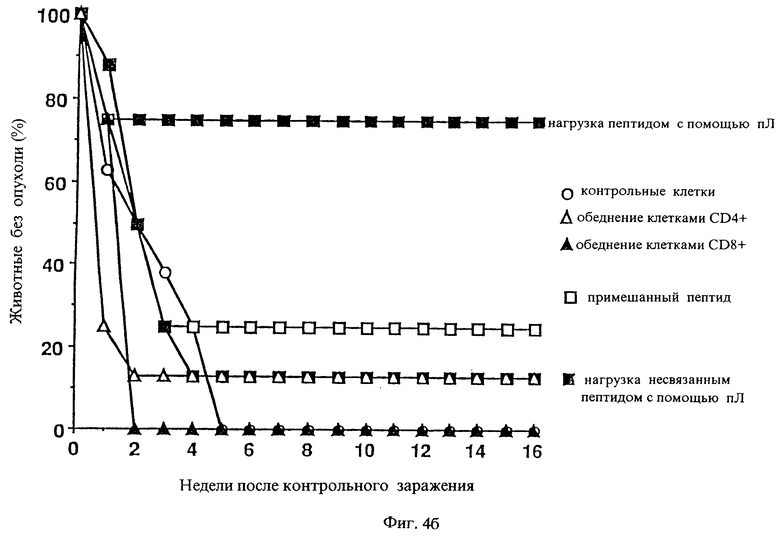
Фиг.4б: Изучение участия Т-лимфоцитов в системном иммунитете.
Фиг. 5: Защита мышей C57BL/6J путем предварительной иммунизации вакциной из нагруженных чужеродным пептидом клеток меланомы.
В следующих далее примерах применяли, если не указано иначе, следующие материалы и способы.
Линия клеток мышиной меланомы Cloudman S91 (клон М-3; Американская коллекция типовых культур /АКТК/ CCL 53.1) была приобретена в АКТК.
Линия клеток меланомы B16-F10 (Fidler и др., 1975) была приобретена в депозитарии опухолей Диагностического Центра Опухолей (ДЦО) Национального Института Здоровья США (НИЗ США).
Получение конъюгатов трансферрин-полилизин, содержащих ДНК трансфекционных комплексов, было проведено, как описано в международной заявке на патент 94/21808.
Пептиды LFEAIEGFI, FFIGALEEI, LPEAIEGFG и ASNENMETM были синтезированы в синтезаторе для пептидов (модель 433 А с монитором обратной связи, фирма Эпплайд Байесистемс, Фостер Сити, Канада) при использовании тентагеля S РНВ (Рапп, Тюбинген) в качестве твердой фазы по способу с использованием 9-флуоренилметилоксикарбонильной (Fmoc) защиты (активация HBTU, фастмокTM, масштаб 0 : 25 ммолей). Пептиды растворяли в 1М триэтиламмонийацетате (ТЭАА), рН 7,3, и очищали с помощью обращенно-фазовой хроматографии на колонке Видак С-18. Последовательности устанавливали с помощью времяпролетной масс-спектрометрии на приборе "Лазермат" фирмы MAT (Финниган, Сан-Хосе, Канада).
Испытание эффективности противораковой вакцины на ее защитное действие в отношении образования метастазов ("Терапевтическая мышиная модель"), а также испытание на профилактической мышиной модели проводили в соответствии с описанным в международной заявке на патент 94/21808 протоколом, причем в качестве мышиной модели использовали модель DBA/2 и модель Balb/c.
Пример 1. Сравнительный FACS-анализ клеток М-3, которые были обработаны различными способами с помощью чужеродного пептида.
Для этого исследования, которое представлено на фиг.1, ксенопептид LFEAIEGFI доставляли в клетки М-3 один раз с комплексами ТФпЛ/ДНК ("транснагрузка"; фиг.1а), один раз клетки инкубировали с пептидом ("импульсы"; фиг. 1б) и один раз пептид адъювантно примешивали к клеткам (фиг.1в).
Для транснагрузки смешивали 160 мкг меченного ФИТЦ ксенопептида LFEAIEGFI или немеченого контрольного пептида с 3 мкг ТФ-пЛ, 10 мкг пЛ и 6 мкг psp65 (фирма Бëрингер Маннхайм, без липополисахаридов /ЛПС/) в 500 мкл буфера, представляющего сбалансированный солевой раствор Хенкса (ССРХ-буфер). Через 30 минут при комнатной температуре вышеупомянутый раствор прибавляли в бутыль с клеточной культурой Т 75 с 1,5•106 клетками М-3 в 20 мл модифицированной по Дюльбекко среды Игла (МДСИ) (10% околоплодная сыворотка теленка /ОСТ/, 20 мМ глюкоза) и инкубировали при 37oС. Через 3 часа клетки дважды промывали забуференным фосфатом физиологическим раствором (ЗФР), растворяли в ЗФР/2мл этилендиаминтетрауксусной кислоты (ЭДТК) и повторно суспендировали для FACS-анализа в 1мл ЗФР/5% OCT.
Пульсацию клеток с пептидом проводили с 1-2•106 клетками в 20 мл МДСИ с 450 мкг пептида (меченного ФИТЦ или немеченого) в течение 3 часов при 37oС.
Для адъювантного примешивания перед FACS-анализом 106 растворенных в бутыли для культуры клеток инкубировали со 100 мкг меченного ФИТЦ пептида в 1 мл ЗФР/ 5% ОСТ 30 минут при комнатной температуре. Клетки после обмена промывали ЗФР/5% ОСТ и еще раз анализировали. FACS-анализ проводили с использованием прибора "FACS Vantage" (Бектон Дикинсон), оснащенного аргоновым лазером мощностью 5 Вт, установленным на 100 мВт при 488 нм, согласно предписанию изготовителя. Результат FACS-анализа представлен на фиг.1а-в. На фиг. 1г показаны микрофотографии цитоцентрифугированных клеток М-3: верхний рисунок представляет клетки, которые получали пептид с помощью комплекса ("транснагрузка"), на нижнем рисунке представлены клетки, которые были проинкубированы с пептидом ("импульсы"). Для контрастного окрашивания ядра применяли 4,6-диамино-2-фенилиндол (ДАФИ).
Клетки М-3, которые были нагружены с помощью содержащего пептид комплекса, показали сдвиг флуоресценции почти на 20% по сравнению с необработанными или обработанными только полилизином клетками, что указывает на эффективный перенос пептида в клетки с помощью комплекса ТФпЛ/ДНК (фиг.1а). Инкубирование с пептидом (импульсы) было менее эффективным, что выразилось в сдвиге в сторону уменьшения флуоресценции только на 10%, что практически не обнаруживали при флуоресцентной микроскопии (фиг.1г). В случае адъювантного примешивания пептид тратился после стадии промывки (фиг.1в), что поэтому означает, что связывание пептида было, в крайнем случае, незначительным.
Пример 2. Лечение мышей DBA/2 с метастазами меланомы вакциной из нагруженных чужеродным пептидом клеток меланомы ("терапевтическая мышиная модель").
а) Получение противоопухолевой вакцины из клеток М-3
160 мкг ксенопептида LFEAIEGFI смешивали с 3 мкг ТФпЛ, 10 мкг пЛ и 6 мкг psp65 (без ЛПС) в 500 мкл ССРХ-буфера. Через 30 минут выдержки при комнатной температуре вышеупомянутый раствор прибавляли в бутыль с клеточной культурой Т 75 с 1,5•106 клетками М-3 в 20 мл МДСИ (10% ОСТ, 20 мМ глюкоза) и инкубировали при 37oС. Через 3 часа клетки помещали в 15 мл свежей среды и инкубировали в течение ночи при 37oС в 5% углекислом газе. За 4 часа до применения клетки облучали дозой в 20 грей. Обработку вакцины осуществляли, как описано в международной заявке на патент 94/21808.
б) Эффективность противоопухолевой вакцины
Мышей DBA/2 в возрасте 6-12 недель с пятидневным метастазом (приобретенным в результате подкожной инъекции 104 живых клеток М-3) обрабатывали дважды на протяжении недели противоопухолевой вакциной путем подкожной инъекции (доза: 105 клеток на животное). В эксперименте участвовало 8 мышей. Результат исследования представлен на фиг.2а; показано, что 7 из 8 животных после введения вакцины, которая содержала в опухолевых клетках загруженный с помощью комплексов ТФпЛ/ДНК пептид, были излечены. В сравнительных исследованиях применялась вакцина, в которой пептид LFEAIEGFI (400 мкг или 4 мг) вводился в клетки путем инкубирования (3 ч при 37oС; "импульсы"). Из животных, которые получали вакцину с 400 мкг пептида, у трех из восьми не обнаружили опухоли, вакцина из клеток, обработанных 4 мг пептида, излечила только одно из восьми животных. Контролем служили одни облученные клетки М-3, а также клетки, которые нагружались комплексами без пептида (в каждом случае у 1/8 животных не наблюдали опухолей). В группе контрольных животных, где не проводили никакой обработки, у всех животных развились опухоли.
Чтобы, с одной стороны, исследовать актуальность способа получения вакцины, а, с другой стороны, изучить пептидную последовательность, была проведена другая серия исследований; в этих экспериментах использовали высокоонкогенный вариант клеток М-3. В испытаниях, где изучали значение способа обработки, получали вакцину, в которой пептид нагружали в клетки не посредством полилизина-трансферрина, но исключительно адъювантно подмешивали к клеткам. Для контроля пептидной последовательности якорные аминокислоты пептида в положениях 2 и 9, а именно фенилаланин и изолейцин, замещали пролином или глицином, что приводило к пептиду Leu Pro Glu Ala Ile Glu Gly Phe Gly (LPEAIEGFG); этому пептиду не хватало способности к связыванию H2-Кd. Образование метастазов контролировали, как минимум, один раз в неделю. Результат этого исследования можно наблюдать на фиг.2б. Вакцина, полученная путем нагрузки клеток LFEAIEGFI с помощью комплексов ТФпЛ/ДНК, излечивала 6 из 8 животных. Напротив, опухоли развились у 7 из 8 животных, которые получали вакцину, для которой к клеткам примешивали исключительно пептид LFEAIEGFI или которая состояла из клеток, которые нагружались с помощью комплексов ТФпЛ/ДНК измененным, не связывающимся при HLA-мотиве, пептидом LPEAIEGFG. В контрольной группе, где обработка велась только облученными клетками М-3 или вообще не проводили никакой обработки, у всех животных развились опухоли.
в) Изучение влияния количества пептида в вакцине
Были получены, как описано в а), содержащие пептид комплексы, которые содержали либо 50, 5, либо 0,5 мкг активного пептида LFEAIEGFI, и этим нагружали клетки М-3. Сравнением служила секретирующая IL-2 вакцина, которая секретировала оптимальную дозу IL-2 (см. г). Эту вакцину вводили мышам DBA/2 с пятидневным метастазом. Вакцина с 50 мкг пептида излечивала 6 из 8 мышей, с 5 мкг излечивала 4 из 8 животных, так же, как секретирующая IL-2 вакцина, в то время как вакцина, содержащая 0,5 мкг, излечивала только двух из 8 животных. Это исследование представлено на фиг.3а.
Пример 3. Сравнение содержащей чужеродный пептид вакцины с противоопухолевой вакциной из секретирующих IL-2 опухолевых клеток на профилактической мышиной модели.
В сравнительных испытаниях две группы исследуемых животных (по 8 мышей в каждой) были предварительно дважды иммунизированы в течение одной недели, с одной стороны, описанной в примере 2а вакциной, с другой стороны, вакциной из секретирующих IL-2 клеток М-3 (полученной согласно протоколу, описанному в международной заявке на патент 94/21808, доза IL-2 составляет 2.000 единиц на животное). Через неделю после последней вакцинации, судя по повышенному количеству опухолевых клеток, были установлены контралатеральные опухоли ("контрольное заражение"; доза представлена на фиг.3б). Обнаружилось, что предварительная иммунизация противоопухолевой вакциной согласно изобретению превосходила обработку секретирующей IL-2 вакциной: простые мыши, вакцинированные секретирующей IL-2 вакциной, были защищены только против одной дозы в 105 живых, высокоонкогенных клеток (M-3-W). Эффективность этой вакцины была, однако, исчерпана при контрольном заражении 3•105 клетками, в то время как опухолевые нагрузки такого масштаба были успешно преодолены животными, которые предварительно подверглись иммунизации вакциной из нагруженных чужеродным пептидом опухолевых клеток.
Пример 4. Защита мышей Balb/с путем предварительной иммунизации вакциной из нагруженных чужеродным пептидом клеток рака толстого кишечника ("профилактическая мышиная модель").
а) Получение вакцины СТ-26
160 мкг ксенопептида LFEAIEGFI или FFIGALEEI смешивали с 12 мкг пЛ или с 3 мкг ТФпЛ и 10 мкг пЛ и подвергали реакции комплексообразования в течение 30 минут при комнатной температуре в 500 мкл ССРХ-буфера, затем переносили в бутыль с клеточной культурой Т 75 с 1,5•106 клетками СТ-26 в 4 мл МДСИ (10% ОСТ, 20 мМ глюкоза), затем инкубировали при 37oС в 5% углекислом газе. Через 4 часа клетки промывали ЗФР, смешивали с 15 мл свежей среды и инкубировали в течение ночи при 37oС в 5% углекислом газе. За 4 часа до применения клетки облучали дозой в 100 грей. Обработку вакцины осуществляли, как описано в международной заявке на патент 94/21808.
б) Испытание активности противораковой вакцины на ее защитное действие против контрольного заражения СТ-26
Мышей Balb/c в возрасте 6-12 недель дважды в течение недели вакцинировали путем подкожных инъекций (доза клеток: 105/мышь). В эксперименте в каждую группу входило 8 мышей (или 7 мышей при исследовании, когда для нагрузки клеток использовали пЛ). Через неделю после последней вакцинации в контралатеральные опухоли вводили 5•104 родительских клеток СТ-26. Сравнительные исследования, в которых вакцину получали другим способом, нежели с помощью комплексов из ТФпЛ/ДНК, проводили, так же, как и контрольные, как описано в примере 2. Развитие опухоли контролировали, как минимум, раз в неделю. Результат для пептида LFEAIEGFI можно видеть на фиг.4а; шесть из восьми животных были защищены. В случае пептида FFIGALEEI (не показано на фиг.4а, были защищены 4 из 8 животных).
в) Участие Т-лимфоцитов в действии противоопухолевой вакцины
Чтобы обнаружить участие Т-лимфоцитов в вызванном СТ-26 - вакциной системном иммунитете, в другом опыте за 24 часа до вакцинирования удаляли клетки CD4+ путем внутривенной инъекции 500 мкг моноклонального антитела GK1.5 (AKTK TIB 207), клетки CD8+ удаляли путем внутривенной инъекции 500 мкг моноклонального антитела 2.43 (AKTK TIB 210). Позитивная контрольная группа получала вакцину без удаления клеток CD4+ и клеток CD8+. Результат испытания представлен на фиг. 4б: участие Т-лимфоцитов проявляется в том, что у всех животных, чьи Т-лимфоциты были удалены, развились опухоли.
Пример 5. Защита мышей C57BL/6J путем предварительной иммунизации вакциной из нагруженных чужеродным белком меланомных клеток ("профилактическая мышиная модель").
В этом примере в качестве исследуемых мышей использовали мышей штамма C57BL/6J (каждый раз по 8 животных в группе). В качестве меланомных клеток были использованы сингенные для используемого мышиного штамма клетки B16-F10 (НИЗ США, ДЦО, депозитарий опухолей; Fidler и др., 1975).
Животных всех исследуемых групп дважды за период в неделю вакцинировали путем подкожных инъекций 105 клетками B16-F10 на мышь.
В серии исследований получали вакцину таким образом, что облученные клетки B16-F10 нагружали пептидом с последовательностью ASNENMETM, как описано в примере 2 для вакцины из клеток М-3.
В параллельных исследованиях применялись секретирующие IL-2 или GM-CSF клетки B16-F10 (полученные согласно протоколу, описанному в международной заявке на патент 94/21808) в качестве вакцины для предварительной иммунизации; вакцина продуцировала 1.000 единиц IL-2 или 200 нг GM-CSF на животное.
Контрольная группа получала для предварительной иммунизации облученные и в остальном необработанные клетки B16-F10.
Через неделю после последней вакцинации подопытным животным прививали опухоль с помощью 1•104 живых, облученных клеток B16-F10 и затем следили за ростом опухоли.
Результат исследования представлен на фиг.5; нагруженные чужеродным пептидом опухолевые клетки продемонстрировали наилучшее защитное действие в отношении образования опухоли.
Литература
Alexander, J. и др., 1989, Immunogenetics 29, 380
Allred, D.C. и др., 1992, J. Clin. Oncol. 10 (4), 599-605
Behr, J.P., 1994, Bioconjug-Chem., сентябрь-октябрь, 5(5), 382-9
Biologic Therapy of Cancer, Редакторы: DeVita, V.T.Jr., Hellman, S.
Rosenberg, S.A., издательство J.B. Lippincott Company, Филадельфия, Нью-Йорк, Лондон, Хагерстаун
Boon, Т., 1993, Spektrum der Wissenschaft (май), 58-66
Boon, Т. и др., 1994, Annu. Rev. Immunol. 12, 337-65
Braciale,T.J. и Braciale, V.L., 1991, Immunol. Today 12, 124-129
Carrel, S. и Johnson, J.P., 1993, Current Opinion in Oncology 5, 383-389
Coligan, J. E. , Kruisbeek, A.M., Margulies, D.H., Shevach, Falk, К. и др., 1991, Nature 351, 290-296
Coulie, P.G. и др., 1992, Int. J. Cancer, 50, 289-297
Coulie, P. G. , Lehmann, F., Lethe, В., Herman, J., Lurquin, C., Andrawiss, M. и Boon, T. (1995). Proc. Natl. Acad. Sci. USA 92, 7976-80
Cox, A.L. и др., 1994. Science 264, 5159, 716-9
Current Protocols im Molecular Biology, 1995, ответственный редактор: Ausubel F.M. и др., John Wiley & Sons, Inc.
Dranoff, G. и др., 1993, Proc. Natl. Acad. Sci. USA 90, 3539-3543
Dranoff, G. и Mulligan, R.C., 1995, Advances in Immunology 58, 417
Falk, К. и др., 1991, Nature 351, 290-296
Felgner, J.H. и др., 1994, J. Biol. Chem. 269, 2550-2561
Fenton, R.G. и др., 1993, J. Natl. Cancer Inst. 85, 16, 1294-302
Fisk, В. и др., 1995, J. Exp. Med. 1881, 2109-2117
Flow Cytometry, Acad. Press, Methods in Cell Biology, 1989, том 33, ответственные редакторы: Darzynkiewicz, Z. и Crissman, H.A.
Gedde Dahl, T. и др., 1992, Hum. Immunol. 33, 4, 266-274
Guarini, А. и др., 1995, Cytokines and Molecular Therapy 1, 57-64
Han, X.K. и др., 1995, PNAS 92, 9747-9751
Справочник: FACS VantageTM User's Guide, апрель 1994, Бектон Дикинсон
Справочник: CELL Quest TM Software User's Guide, июнь 1994, Бектон Дикинсон
Herin. и др., 1987. Int. J. Cancer, 39, 390
Hock, Н. и др., 1993, Cancer Research 53, 714-716
Houbiers, J. G., Nijman, H. W., van der Burg, S.H., Drijfhout, J. W.,
Kenemans, P., van de Velde, C. J., Brand, A., Momburg, F., Kast, W.M. и Melief, C. J. (1993). Eur. J. Immunol. 23, 2072-7
Huang, A. Y. C. и Pardoll, D.M. (1996). Proc. Natl. Acad. Sci. USA 93, 9730-5 Jung, S. и др., 1991, J. Exp. Med. 173, 1, 273-6
Kawakami, Y. и др., 1995, The Journal of Immunol. 154, 3961-3968
Karre, К.. и др., 1986, Nature 319, 20. февр., 675
Kovacsovics Bankowski, M. и Rock, K..L., 1995, Science 267, 243-246
Lehmann, J.M. и др., 1989. Proc. Natl. Acad. Sci. USA 86, 9891-9895
Lethe, В. и др., 1992, Eur. J. Immunol. 22, 2283-2288 Lill, N.L., Tevethia, M. J., Hendrickson, W.G. и Tevethia, S. S. (1992). J. Exp. Med. 176, 449-57
Loeffler, J.-P. и др., 1993, Methods Enzymol. 217, 599-618
Mackiewicz, А. и др., 1995, Human Gene Therapy 6, 805-811
Malnati, M.S. и др., 1995, Science 267, 1016-1018
Mandelboim, О. и др., 1994, Nature 369, 5. май, 67-71
Mandelboim, O. и др., 1995, Nature Medicine 1, 11, 1179-1183
Morishita. R. и др., 1993, J. Clin. Invest. 91, 6, 2580-5
Nabel, G.J. и др., 1993, Proc. Natl. Acad. Sci. USA 90, 11307-11311
Noguchi, Y., Chen, Y. Т. и Old, L. J. (1994). Proc. Natl. Acad. Sci. USA 91, 3171-3175
Oettgen, H. F. и Old, L. J., 1991, Biologic Therapy of Cancer, под редакцией: DeVita, V. Т. Jr., Hellman, S., Rosenberg, S. A., издательство J.B. Lippincott Company, Филадельфия, Нью-Йорк, Лондон, Хагерстаун, 87-119
Ostrand-Rosenberg, S., 1994, Current Opinion in Immunology 6, 722-727
Pardoll, D. M., 1993, Immunology Today 14, 6, 310 Practical Immunology, редакторы: Leslie Hudson и Frank C. Hay, научные публикации Blackwell, Оксфорд, Лондон, Эдинбург, Бостон, Мельбурн
Peace, D. J. и др., 1991, J. Immunol. 146, 6. 2059-65
Peoples, G.E. и др., 1994, J. Immunol. 152, 10, 4993-9
Plautz, G.E. и др., 1993, Proc. Natl. Acad. Sci. USA 90, 4645-4649
Rammensee, H. G. и др., 1993, Current Opinion in Immunology 5, 35-44
Rammensee, H. G., 1995, Current Opinion in Immunology 7, 85-96
Rammensee, H. G. , Friede, Т. и Stepvanovic, S. (1995). Immunogenetics 41, 178-228
Remy, J. S. и др., 1994, Bioconjug-Chem., ноябрь-декабрь, 5(6), 647-54
Rivoltini, L. и др., 1995, The Journal of Immunology 154, 2257-2265
Robbins, P. P., el Gamil, M., Li, Y. F., Topalian, S.L., Rivoltini, L., Sakaguchi, K. . , Appella, E., Kawakami, Y. и Rosenberg, S. A. (1995). J. Immunol. 154, 5944-50
Robbins и Rosenberg. (1996). J. EXP. MED. 183, 1185-92.
Schmidt, W. и др., май 1995, Рrос. Natl. Acad. Sci. USA, 92, 4711-4714
Skipper, J. и Stauss, H. J., 1993, J. Exp. Med. 177, 5, 1493-8
Slingluff, С. L. и др., 1994, Current Opinion in Immunology 6, 733-740
Stein, D. и др., 1994, EMBO-Journal, 13, 6, 1331-40
Stuber, G. , Leder, G. H. , Storkus, W. Т., Lotze, M. Т., Modrow, S., Szekely, L. , Wolf, H. , Klein, E., Karre, К. и Klein, G. (1994). Eur. J. Immunol. 24, 765-768
Sykulev, Y. и др., 1994, Immunity 1, 15-22
Theobald, M., Levine, A. J. и Sherman, L. A. (1995) PNAS 92, 11993-7
Tibbets,L.M. и др., 1993, Cancer, январь 15., том 71, 2, 315-321
van der Bruggen, Р. и др., 1994, Eur. J. Immunol. 24, 9, 2134-40, международный стандартный серийный номер: 0014-2980
Van Pel, А. и Boon, Т., 1982, Ргос. Natl. Acad. Sci. USA 79, 4718-4722
Wolfel, T. и др., 1994 a), Int. J. Cancer 57, 413-418
Wolfel, Т. и др., 1994 б), Eur. J. Immunol. 24, 759-764
Wolfel, Т., Hauer, M., Schneider, J., Serrano, M., Wolfel, C., Klehmann Hieb, E., De Plaen, E., Hankeln, Т., Meyer zum Buschenfelde, K. H. и Beach, D. (1995). Science 269, 1281-4
York, I.A. и Rock, K. L., 1996, Ann. Rev. Immunol. 14, 369-396
Yoshino, I. и др., 1994 a), J. Immunol. 152, 5, 2393-400
Yoshino, I. и др., 1994 б), Cancer Res., 54, 13, 3387-90
Zatloukal, К. и др., 1993, Gene 135, 199-20
Zatloukal, К. и др., 1995, J. Immun. 154, 3406-3419e
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕКАРСТВЕННЫЙ СОСТАВ С ИММУНОМОДУЛЯТОРНЫМ ДЕЙСТВИЕМ, СОДЕРЖАЩИЙ ПЕПТИДЫ И ВСПОМОГАТЕЛЬНЫЕ СРЕДСТВА | 1997 |
|
RU2219942C2 |
| СРЕДСТВО И СПОСОБ ЛЕЧЕНИЯ ПЛОСКОКЛЕТОЧНОГО РАКА | 1996 |
|
RU2193779C2 |
| ИЗОЛИРОВАННАЯ НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ ПОЛИПЕПТИД TAG 7, ИЗОЛИРОВАННЫЙ ПОЛИПЕПТИД TAG 7, СПОСОБ ИНГИБИРОВАНИЯ РАЗВИТИЯ ОПУХОЛЕЙ У МЛЕКОПИТАЮЩИХ (ВАРИАНТЫ) И СПОСОБ ЛЕЧЕНИЯ РАКА У ЖИВОТНОГО(ВАРИАНТЫ) | 1998 |
|
RU2238976C2 |
| КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ РАКА ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ (РПЖ) | 2008 |
|
RU2508125C2 |
| ВАКЦИННЫЕ КОМПОЗИЦИИ, СПОСОБЫ ИХ ПРИМЕНЕНИЯ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ МЕЛАНОМЫ И ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ ДЛЯ ПОЛУЧЕНИЯ ДЕЙСТВУЮЩИХ КОМПОНЕНТОВ КОМПОЗИЦИИ | 2006 |
|
RU2333767C2 |
| КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ РАКА ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ | 2008 |
|
RU2583004C2 |
| КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ РАКА ЛЕГКИХ, ПРЕЖДЕ ВСЕГО, НЕМЕЛКОКЛЕТОЧНОГО РАКА ЛЕГКИХ (НМКРЛ) | 2008 |
|
RU2526510C2 |
| СПОСОБ ВАКЦИНОТЕРАПИИ ОНКОЗАБОЛЕВАНИЙ | 2000 |
|
RU2192883C2 |
| ПОЛУЧЕНИЕ КОМПЛЕКСОВ НУКЛЕИНОВЫХ КИСЛОТ И ПОПЕРЕЧНО СШИТЫХ ДИСУЛЬФИДНЫМИ СВЯЗЯМИ КАТИОННЫХ КОМПОНЕНТОВ, ПРЕДНАЗНАЧЕННЫХ ДЛЯ ТРАНСФЕКЦИИ И ИММУНОСТИМУЛЯЦИИ | 2011 |
|
RU2575603C2 |
| АНТИТЕЛО VFF-18 И ЕГО АНАЛОГИ | 1995 |
|
RU2204606C2 |








Изобретение относится к медицине и касается иммуностимулятора, вызывающего специфичный к опухоли клеточный иммунный ответ, и способа его получения. Иммуностимулятор содержит опухолевые клетки, по меньшей мере, часть которых имеет на клеточной поверхности, как минимум, один гаплотип ГКГ-I больного и которые были нагружены таким образом одним или несколькими пептидами, связывающими молекулу ГКГ-I, что упомянутые опухолевые клетки узнаются как чужеродные в окружении с пептидами иммунной системой больного и вызывают клеточный иммунный ответ. Нагрузка происходит в присутствии поликатиона, такого как полилизин. Преимущество изобретения заключается в повышении противоопухолевой активности. 2 с. и 24 з.п.ф-лы, 5 ил., 1 табл.
Приоритет по пунктам и признакам:
23.11.95 - по пп.1, 2, 4, 5, 7, 8 - 26;
п.3 - в части пептида, представляющего собой лиганд для Н2-Кd;
п. 6 - в части пептида с последовательностью Leu Phe Glu Аlа IIe Glu Gly Phe IIe относятся к приоритету от 23.11.1995, п. 3 - в части пептида, представляющего собой лиганд для Н2-Кb;
24.02.1996 - п.6 в части пептида с последовательностью Ala Ser Asn Glu Asn Met Glu Thr Met.
| Массонапускное устройство бумагоделательной машины | 1975 |
|
SU569678A1 |
| Способ измерения глубины залегания структур кожи у животных | 1973 |
|
SU563627A1 |
| ECSENBACH and FELDMAN | |||
| Genes and antygenes controlling tumer metastasis, Hematology and blood transfusion, Modern Trends in human Leukemia IV, ed R | |||
| Neth et al., 1985, рр | |||
| Складная пожарная (штурмовая) лестница | 1923 |
|
SU499A1 |

