Изобретение относится к области генетической инженерии, биотехнологии, иммунологии и медицины и может быть использовано для получения противоопухолевых вакцин, профилактической вакцинации и лечения меланомы.
Изобретение включает также методы применения антигенпрезентирующих клеток, сенситизированных гибридными белками, состоящими из белков семейства HSP и специфических пептидов меланомы.
Пептиды группы MAGE представляют собой онкоспецифические маркеры, преимущественно экспрессируемые в меланомных клетках, в здоровом организме присутствующие лишь в тестикулах, которые недоступны для клеток иммунной системы благодаря отсутствию прямого контакта с иммунными клетками и отсутствию экспрессии белков главного комплекса гистосовместимости HLA класса I на поверхности клеток тестикул. Гены MAGE представлены тремя подсемействами - MAGE-A, -В и -С - из которых продукты экспрессии MAGE-A являются наиболее иммуногенными. Гены MAGE-A представлены семейством из 12 генов, локализованных на длинном плече хромосомы Х (участок Xq-28) [De Plaen E. et al., Immunogenetics 40: 360-369, 1994]. В большинстве исследованных опухолей экспрессируются только гены MAGE-A1, -А2, -A3, -А4, -А6, -A11 и -А12. Эпитопами, узнаваемыми после презентации цитотоксическими лимфоцитами, обладают лишь продукты экспрессии MAGE-A1 [van der Bruggen P. et al., Science 254: 1643-1647, 1991], MAGE-A2 [Viressen M.J. et al.. Int. J. Cancer 73: 125-130, 1997] и MAGE-А3 [Gaugler В. et al., J. Exp. Med. 179: 921-930, 1994]. Идентифицирован антигенный эпитоп MAGE-A1, презентируемый HLA-A1 (EADPTGHSY) [Yamashita N. et al., Hepatology 24: 1437-1440, 1996]. В структуре белка MAGE-A2 было идентифицировано два пептида (KWVELVHFL и YLQLVFGIEV), которые способны к связыванию с HLA-A2 и индуцируют формирование цитотоксических лимфоцитов, способных убивать клетки, экспрессирующие MAGE-A2 у трансгенных мышей.
Белок MAGE-А3 содержит четыре известных эпитопа, первый из которых (EVDPIGHLY [Gaugler В. et al, J. Exp. Med. 179: 921-930, 1994]) презентируется HLA-A1, второй (FLWGPRALV [van der Bruggen P. et al., Eur. J. Immunol. 24: 3038-3043, 1994]) - HLA-A2, третий (IMPKAGLLI [Tanaka F. et al., Cancer Res. 57: 4465-4468, 1997]) - HLA-A24 и, наконец, четвертый (MEVDPIGHLY [Herman J. et al., Immunogenetics 43: 377-383, 1996]) презентируется HLA-B44.
Особенно высокий уровень экспрессии антигенов MAGE-A отмечается в клетках меланом (65% MAGE-A3 [De Plaen E. et al., Immunogenetics 40: 360-369, 1994]), карцином молочной железы, рака головы и шеи, карцином легких и мочевого пузыря, а также сарком [Van Pel. A. et al., Immunol. Rev. 145: 229-250, 1995]. Очень высокий уровень экспрессии MAGE-A1 (80%) отмечается при раке печени [Yamashita N. et al., Hepatology 24: 1437-1440, 1996]. Отмечается также корреляция уровня экспрессии антигенов MAGE-A со степенью прогрессии опухоли [Brasseur F. et al., Int. J. Cancer 63: 375-380, 1995; Eura M. et al., Int. J.Cancer 64: 304-308, 1995; Katano M. et al., J.Surg. Oncol. 64: 195-201, 1997; Patard J.J. et al. Int. J.Cancer 64: 60-64, 1995]. Исследование способности пептидных фрагментов MAGE-A1 и MAGE-A3 к индукции антимеланомного иммунного ответа in vivo показало, что пептидный фрагмент (EVDPIGHLY) MAGE-A3 проявлял наибольшую противоопухолевую активность. Функция белков MAGE-A1 и MAGE-A3 неизвестна, хотя известно, что они локализованы в цитоплазме клеток [Kocher Т. et al. Cancer Res. 55: 2236-2239, 1995; Margolin L.F. et al., Proc. Natl. Acad. Sci. USA 91: 4509-4513, 1994] и являются сравнительно небольшими белками: MAGE-A1 - 309 аминокислот, MAGE-A2 и MAGE-A3 - 314 а.к., MAGE-A4 - 317 а.к. [Gamier J. et al., Methods Enzymol. 266: 540-553, 1996].
С иммунологической точки зрения антигены группы MAGE являются очень хорошими мишенями для проведения иммунотерапии. Они широко представлены в ряде опухолей и отсутствуют в нормальных тканях человека за исключением яичек, на которых клетки иммунной системы не действуют благодаря отсутствию прямого контакта клеток яичек и иммунных клеток, а также отсутствию HLA класса I на поверхности репродуктивных клеток [Barker C.F. and Billingham R.E., Adv. Immunol. 25: 1-54, 1977; Tomita Y. et al., J.Urol. 149: 659-663, 1993]. Белки MAGE - относительно крупные белки с рядом потенциальных эпитопов, связываемых HLA класса I. Однако при применении пептидных фрагментов этих белков in vivo вызываемый ими иммунный ответ и противоопухолевая активность остаются на весьма низком уровне.
Одним из объяснений низкой иммуногенности антигенов MAGE является стабильность их структуры - как и другие высокоорганизованные белки, MAGE имеют в составе молекулы высокий процент α-спиралей, обеспечивающих оптимальную геометрию для максимального количества и силы водородных связей. Внутри клетки для последующего представления антигена белок MAGE должен быть частично ренатурирован, убиквитирован и затем подвергнут протеолитической деградации. Очевидно, что высокоорганизованный белок со структурой, стабилизированной большим количеством связей, будет более устойчив к протеосомной деградации, а следовательно, и с большим трудом презентироваться на поверхности клеток в составе HLA типа I, что и наблюдается в случае меланомных белков MAGE [Kirkin A.F. et al., APMIS 106: 665-679, 1998]. Вероятно, что опухолевые клетки характеризуются крайне низким уровнем экспрессии презентируемого антигена из следовых количеств частично денатурированных опухолеспецифичных белков MAGE. В силу этого лимитирующим этапом может являться обучение «наивных» лимфоцитов, для которого требуется примерно в 1000 раз более высокие концентрации (уровни) антигенного пептида, презентируемого на клеточной поверхности, чем это требуется для лизиса клетки, несущей антиген, уже обученными лимфоцитами.
Значительное повышение иммуногенности пептидов MAGE может быть достигнуто при их применении в составе вакцин в связанном с белками теплового шока (стрессовыми белками, HSP) виде. Белки теплового шока обладают способностью к связыванию с широким спектром опухолеспецифических антигенных пептидов. Такой комплекс обладает способностью к интернализации в антигенпредставляющие клетки с последующей эффективной презентацией пептида на их поверхности в составе HLA класса I. Презентируемый соответствующим образом антиген после активации «наивных» лимфоцитов будет вызывать развитие клеточного иммунного ответа, направленного на клеточные белки, принимающие участие в опухолевом процессе. Преимуществом предлагаемого заявителем подхода является возможность создания комплексных противомеланомных вакцин путем смешивания набора гибридных белков, состоящих из белка теплового шока HSP70 и различных специфических антигенных пептидов меланомы. Подобные вакцины будут отличаться простотой изготовления, безопасностью и высокой эффективностью терапевтического действия, заключающегося в подавлении опухолевого роста.
Наряду с пептидами MAGE одним из наиболее перспективных для индукции клеточного иммунного ответа опухолевых антигенов является продукт экспрессии гена gp 100. Белок gp 100 представляет собой мембранный гликопротеин, состоящий из 661 аминокислоты. Ген gp 100 экспрессируется в большинстве линий меланомных клеток, а также на культивируемых меланоцитах. В линиях немеланомных раковых клеток gp 100 не экспрессируется. В нормальных тканях gp 100 обнаружен лишь в сетчатке [Adema G.J. et al., Am. J.Pathol. 143: 1579-1585, 1993], где белок преимущественно локализован в мембране и филаментном матриксе премеланосом, где, по-видимому, участвует в синтезе меланина [Schaumburg-Lever G. et al., J.Cutan. Pathol. 18: 432-435, 1991; Vennegoor С. et al., Am. J.Pathol. 130: 179-192, 1988]. Белок gp 100 распознается рестриктированными по HLA-A2 инфильтрующими опухоль лимфоцитами [Сох A.L. et al., Science 264: 716-719, 1994]. При этом идентифицирована последовательность YLEPGPVTA, распознаваемая цитотоксическими лимфоцитами. Очевидно, что использование белка gp 100 или его фрагментов в составе HSP-вакцин для профилактической вакцинации или лечения меланом имеет большие перспективы. Предпосылкой к терапевтическому использованию gp 100 является работа [Adema G.J. et al., Am. J.Pathol. 143: 1579-1585, 1993], продемонстрировавшая, что введенные больному меланомой инфильтрующие опухоль лимфоциты, использованные для клонирования гена gp 100, вызывали регрессию метастазов у больных. Применение гибридного белка, состоящего из HSP 70 и антигенного фрагмента gp 100 для активации антигенпредставляющих клеток и последующей индукции иммунного ответа может позволить сделать терапию значительно более прогнозируемой и эффективной.
В последнее время рядом авторов были проведены работы по созданию вакцин, в состав которых входят белки теплового шока. В связи с низкой иммуногенностью синтетических антигенных пептидов рядом авторов предпринимались попытки ее повышения путем создания in vitro нековалентных комплексов антигенных пептидов и человеческих или микобактериальных белков теплового шока [Barrios С.et al., Eur. J.Immunol. 22: 1365-1372, 1992; Roman E. and Moreno С., Immunol. 88: 487-492, 1996]. Эти комплексы обладают способностью вызывать сильные специфичные реакции на входящие в их состав антигенные пептиды. Комплексы HSP-белков с антигенными пептидами можно рассматривать в качестве легко доступных вакцинных препаратов, которые могут применяться без каких-либо дополнительных адьювантов. Также нековалентные комплексы HSP-белков с антигенными пептидами могут быть использованы для активации ex vivo взятых у пациентов антигенпредставляющих дендритных клеток. После введения пациенту активированных дендритных клеток может достигаться достаточно сильный клеточный иммунный ответ на антигенные пептиды, входящие в состав комплексов.
В связи с высокой потребностью в эффективных и безопасных вакцинных препаратах для профилактики и лечения онкологических заболеваний, от которых в мире ежегодно умирает около 6 млн. человек, работы в области создания таких препаратов являются чрезвычайно актуальными. Результатом работ в этой области явился ряд изобретений, которые легли в основу стратегии разработки противоопухолевых вакцин, включающих белки теплового шока.
В заявке на изобретение РФ №2003101965/14 описан способ лечения заболеваний, связанных с вирусом папилломы человека (бородавки, рак шейки матки и прямой кишки и т.д.) с помощью композиции, содержащей гибридный белок, включающий микобактериальный белок теплового шока (HSP65 или HSP70 из Mycobacterium bovis) и белок вируса папилломы. Лечение может осуществляться либо путем введения субъекту рекомбинантного гибридного белка, либо путем введения нуклеиновой кислоты, кодирующей гибридный белок, которая содержится в вирусном векторе. К недостаткам используемого авторами заявки подхода следует отнести использование вирусного вектора, последствия применения которого недостаточно изучены и могут представлять потенциальную опасность для пациента. Следует также отметить, что применение описанного изобретения ограничивается лишь новообразованиями, связанными с инфицированием вирусом папилломы человека.
В заявке США на изобретение №20030216315 описан метод ингибирования роста первичных опухолей и метастазов путем введения субъекту рекомбинантного фрагмента белка теплового шока (HSP60, HSP70, HSP90), лишенного пептидсвязывающего домена. При этом могут быть использованы гены, кодирующие указанные белки из любых организмов. Авторы изобретения рассматривают рекомбинантный фрагмент белка теплового шока в качестве средства стимуляции иммунного ответа при онкологических и иных заболеваниях. По мнению авторов, этот белок может также найти применение в качестве адьюванта для стимуляции специфического иммунного ответа на экзогенные антигены. Совместное введение белка и антигенного пептида субъекту должно вызывать специфический адаптивный иммунный ответ, направленный против антигена.
Однако нельзя не отметить, что получение в виде рекомбинантного продукта не целого нативного белка теплового шока, а его фрагмента, лишенного пептидсвязывающего домена, с технологической точки зрения является более выигрышным, поскольку процесс получения белка более прост, дешев и занимает меньше времени. Однако с точки зрения практического применения такого белка просматриваются серьезные недостатки. Хотя фрагмент и сохраняет свойства адьюванта, отсутствие в его составе пептидсвязывающего домена не позволяет использовать этот белок для создания in vitro эффетивных вакцин, в составе которых антигенные пептиды находятся в прочной связи с белком теплового шока. Поэтому более перспективным представляется создание рекомбинантных вакцин на основе химерных белков, состоящих из антигена, связанного с микобактериальным HSP70 или HSP65.
В основе настоящего изобретения лежит способность белков теплового шока стимулировать иммунные реакции. При этом ранее белки теплового шока использовались в качестве адьювантов для усиления иммуногенности вирусных и бактериальных белков [патент РФ №2229307]. При разработке новых вакцинных и иммунотерапевтических композиций заявителем решалась задача усиления иммунного ответа на пептиды - фрагменты специфических белковых антигенов меланомы - с помощью стрессовых белков (белков теплового шока). Предлагаемые заявителем гибридные белки, состоящие из белка HSP70 и специфических пептидов меланомы, перечисленных ниже, для вакцинации против меланомы и ее иммунотерапии ранее не применялись, и состав и эффективность их действия установлены заявителем опытным путем впервые. Таким образом, предлагаемое техническое решение отвечает критериям «изобретательский уровень» и «новизна».
Изобретательской задачей является создание новых вакцинных композиций для профилактики и лечения меланомы человека и методов их применения, а также методов применения антигенпрезентирующих клеток, сенситизированных компонентами этих композиций.
Изобретательская задача решается тем, что предлагаются вакцинные композиции для профилактики и лечения меланомы, содержащие инертный носитель, разбавитель и антигенный компонент в эффективном количестве, и отличающаяся тем, что в качестве последнего она содержит отдельные гибридные белки, состоящие из HSP70 Mycobacterium tuberculosis и различных пептидных фрагментов специфических белков меланомы MAGE и gp 100, или их смеси (MAGEA1-HSP70, MAGEA2-HSP70, MAGEA3-HSP70, PEPGP-HSP70).
Предлагается также способ получения указанных гибридных белков путем экспрессии их генов в составе синтетических рекомбинантных векторов в штаммах-продуцентах клеток-хозяев.
Предлагается также способ иммунотерапии и профилактической вакцинации, отличающийся тем, что стимуляцию иммунитета проводят путем введения пациенту композиции, содержащей указанные гибридные белки.
Предлагается также способ иммунотерапии и профилактической вакцинации, отличающийся тем, что стимуляцию иммунитета у пациента проводят с помощью аутологических антигенпрезентирующих клеток, сенситизированных гибридными белками, состоящими из HSP70 Mycobacterium tuberculosis и различных пептидных фрагментов специфических белков меланомы MAGE и gp 100.
Изобретение иллюстрируется следующими примерами.
Пример 1. Получение рекомбинантной плазмидной ДНК, кодирующей синтез гибридного белка MAGEA1-HSP 70
Синтез последовательности ДНК, кодирующей С-концевой фрагмент белка HSP70 (DHSP70)
В качестве источника гена HSP 70 была использована кДНК-библиотека генов М. tuberculosis.
Ген HSP70 М.tuberculosis был выделен методом амплификации с помощью следующих праймеров:

где выделенный фрагмент - сайт узнавания ферментом рестрикции XhoI, подчеркнутый - HindIII
Синтез последовательности, необходимой для создания ДНК, кодирующей гибридный белок.
Праймеры для синтеза последовательности, с сайтами рестрикции ВатН I и XhoI, необходимой для создания гибридного белка, состоящего из пептида MAGE-A1 слитого с HSP70 (MAGA1HSPD).
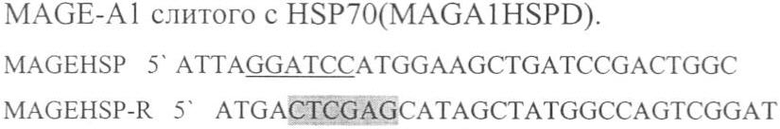
Подчеркнутый фрагмент - сайт узнавания ферментом рестрикции BamHI, выделенный - XhoI
Для синтеза нижеуказанной последовательности, соответствующие пары праймеров смешивали и амплифицировали, в результате чего был получен следующий олигонуклеотид:
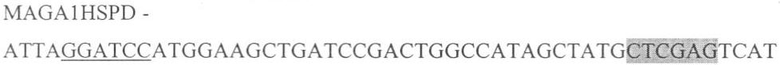 где подчеркнутый фрагмент - сайт узнавания ферментом рестрикции BamHI, выделенный - XhoI
где подчеркнутый фрагмент - сайт узнавания ферментом рестрикции BamHI, выделенный - XhoI
Синтез последовательности, с сайтами рестрикции BamHI и HindIII, кодирующей гибридный белок, состоящий из пептида MAGE-A1, слитого с HSP70 (MAGA1HSP)
Для получения MAGA1HSP фрагмент DHSP70 и олигонуклеотид MAGA1HSPD обрабатывали рестриктазой Xhol и легировали между собой с помощью лигазы фага Т4.
Последовательность, кодирующая гибридный белок, состоящий из пептида
где подчеркнутый фрагмент - сайт узнавания ферментом рестрикции BamHI, выделенный - HindIII, *** - пропущенный фрагмент последовательности DHSP70
Последовательности, кодирующие синтез гибридных белков, в состав которых входят прочие перечисленные в табл.1 специфические пептиды меланомы, синтезировали по аналогичной схеме.
Создание векторов для экспрессии гибридных белков
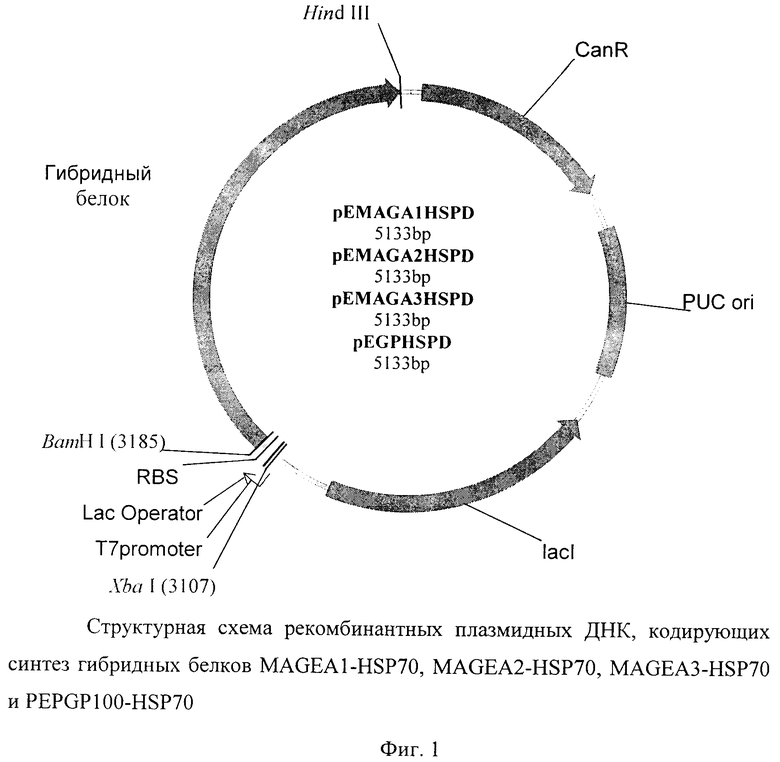
Для создания экспрессионных векторов фрагменты ДНК, кодирующие соответствующие гибридные белки, обрабатывали ферментами рестрикции BamHI и HindIII и вставили по сайтам рестрикции BamHI и HindIII в вектор рЕТ (Novagen). Полученные векторы назвали pEMAGA1HSPD, pEMAGA2HSPD, pEMAGA3HSPD, pEGPHSPD.
Общая схема полученных векторов представлена на фиг.1.
Индукция синтеза целевых гибридных белков оценивалась в 10 мл ИПТГ-индуцированных культур (выращивание до OD600 0.5, внесение ИПТГ до 0.2 мМ, затем инкубация 3 часа при 37°С на качалке). Продукция белков ожидаемого размера проверена ДСН-ПААГ электрофорезом. Для экспрессии рекомбинантных белков был использован штамм Е.coli BL21 (DE3).
Продуктами, полученными в результате выполнения данных процедур, являются гибридные белки, состоящие из HSP70 Mycobacterium tuberculosis, к С-концу которого присоединен один из перечисленных в табл.1 специфических антигенных пептидов меланомы.
Аминокислотная структура пептидов - фрагментов опухолеспецифических антигенов.
Пример 2. Индукция цитотоксических лимфоцитов и их цитотоксическая активность в отношении опухолевых клеток-мишеней.
Приготовление дендритных клеток (ДК)
ДК выделяли из лейкоконцентрата периферической крови здоровых доноров по методике [Thumer В. et al., J.Immunol. Meth. 223: 1-15, 1999]. С помощью градиентного центрифугирования (400 g, 30 мин) через раствор фиколл-пак (Pharmacia, Швеция) получали фракцию мононуклеарных лейкоцитов, которую затем промывали три раза раствором Версена (250 g, 175 g и 110 g no 15 мин), ресуспендировали в среде RPMI 1640 с добавлением 10% инактивированной нагреванием эмбриональной бычьей сыворотки, 100 ед/мл пенициллина, 100 мкг/мл стрептомицина в концентрации 50 млн клеток/мл, помещали по 10 мл в пластиковые чашки Петри и инкубировали 2 ч в увлажненной атмосфере 5%-ного CO2 при 37°С. Затем отбирали неприкрепившиеся клетки (в основном лимфоциты), центрифугировали, замораживали и хранили в жидком азоте. К монослою прикрепившихся клеток (в основном моноцитов) вносили по 10 мл свежей среды и инкубировали в стандартных условиях культивирования до следующего дня, который обозначали как "день 1". На 1 и 3 дни культивирования в инкубационную среду добавляли ГМ КСФ (800 ед/мл) и IL-4 (1000 ед/мл). В день 5 клетки собирали и переносили в 6-луночные планшеты при плотности 5·105 клеток на лунку в 3 мл свежей среды с цитокинами в той же концентрации. В день 6 добавляли индукторы созревания ДК в виде комплексного препарата цитокинов первой фазы иммунного ответа «Лейкинферон» и один из гибридных белков. На день 8 ДК собирали.
Индукция цитотоксических лимфоцитов и определение их цитотоксической активности (ЦТА)
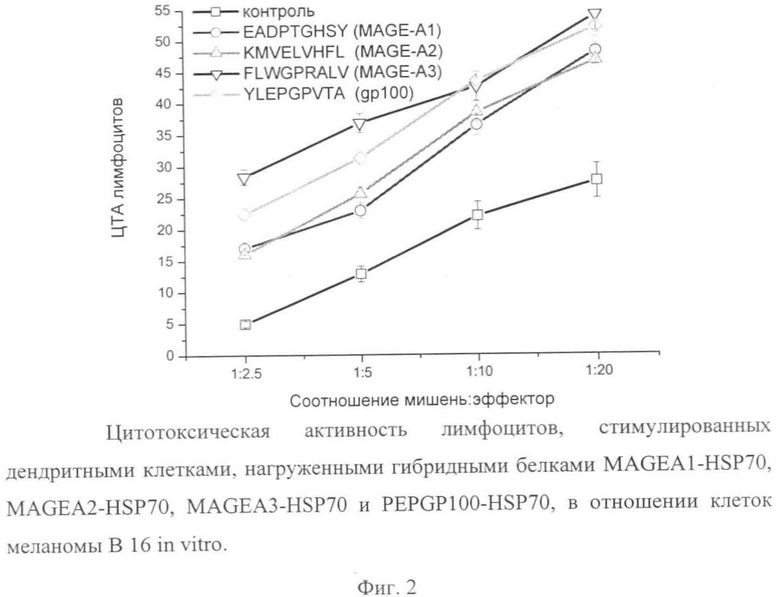
Лимфоциты, выделенные из периферической крови как было описано выше, размораживали и суспендировали в культуральной среде. Для стимуляции лимфоцитов ДК отмывали фосфатно-солевым буфером и помещали в 24-луночные планшеты вместе с лимфоцитами при соотношении ДК:лимфоциты, равном 1:20 в 1 мл среды HyQ IMDM, содержащей 10% инактивированной эмбриональной бычьей сыворотки. На следующий день после начала стимуляции и далее 1 раз в 3 дня к культуре добавляли по 30 ед/мл человеческого рекомбинантного IL-2. Через 7 дней культуры лимфоцитов рестимулировали с использованием размороженных ДК в таком же количестве, как и для первичной стимуляции. В качестве фидеров к культурам лимфоцитов после каждой стимуляции вносили сингенные лимфоциты, инактивированные ультрафиолетовым облучением (1 млн на лунку). Через 5 дней после второй стимуляции проводили определение ЦТА лимфоцитов в отношении опухолевых клеток-мишеней. Для этого в лунки 96-луночного планшета вносили суспензию лимфоцитов-эффекторов в количестве 12.5-100 тыс. клеток на лунку, а затем добавляли опухолевые клетки (5 тыс. клеток на лунку), предварительно помеченные 3H-тимидином так, чтобы соотношение клетки-мишени:клетки эффекторы составляло 1:2.5-1:20. Через 24 часа инкубации определяли выживаемость опухолевых клеток и рассчитывали ЦТА лимфоцитов-эффекторов по формуле: ЦТА=(1-Рмишень+эффектор/Рмишень)-100%, где Рмишень и Рмишень+эффектор - радиоактивность проб, в которых инкубировались только опухолевые клетки-мишени и эти же клетки вместе с клетками-эффекторами соответственно.
Результаты исследования цитотоксической активности стимулированных лимфоцитов в отношении опухолевых клеток приведены на фиг.2. В качестве опухолевых клеток-мишеней были выбраны клетки меланомы линии В16, в которых экспрессируются антигены MAGE и g 100. Как видно из фиг.2, цитотоксические лимфоциты, полученные после совместного культивирования с дендритными клетками, нагруженными соответствующими гибридными белками, обладали более высокой цитотоксической активностью по сравнению с контрольными лимфоцитами, индуцированными интактными дендритными клетками, при всех исследованных соотношениях мишень:эффектор.
Пример 3. Иммунизация мышей вакцинными композициями на основе гибридных белков.
Для иммунизации вакцинную композицию, содержащую гибридный белок MAGEA1-HSP70 (см. Пример 1), вводили мышам линии С57В1/6 внутрикожно в диапазоне доз от 0,01 до 100 мкг. Через 7 дней после иммунизации мышам подкожно вводили суспензию опухолевых клеток меланомы линии В16 в количестве 1·106 клеток/мышь. Такое количество прививаемых опухолевых клеток в 10 раз превышает дозу, обычно используемую в экспериментах по моделированию опухолей данной линии у мышей. Размеры развивающихся опухолей измеряли 3 раза в неделю. Эффективность вакцинации оценивали по замедлению роста опухолей у иммунизированных мышей по сравнению с контрольными (интактными) животными, а также по увеличению средней продолжительности жизни животных. На момент гибели контрольных животных средний объем опухолей у иммунизированных мышей составлял всего 53% от контроля, а продолжительность жизни иммунизированных животных увеличивалась на 59%. Сходные результаты были получены в результате иммунизации мышей вакцинными композициями, содержащими другие гибридные белки по Примеру. 1 (данные не приведены).
Пример 4. Сенситизация вакцинными композициями антигенпредставляющих клеток - макрофагов и дендритных клеток.
Дендритные клетки получают из лейкоконцентрата периферической крови здоровых доноров, как описано в примере 3. Макрофаги получают из мононуклеарных клеток, выделенных из периферической крови человека путем центрифугирования в градиенте Фиколла. Клетки высеиваются на культуральные чашки Петри, предварительно покрытые сывороткой человека, и инкубируются в течение 1 ч при 37°С. Затем неприкрепившиеся клетки удаляются, а к прикрепившимся добавляют охлажденный до +4°С фосфатно-солевой буфер, содержащий 1 мМ ЭДТА, и оставляют на 10-15 мин. Клетки собирают, отмывают и суспендируют в среде RPMI. Дальнейшее культивирование и сенситизацию клеток проводят, как описано в примере 3.
Сенситизированные аутологичные антигенпредставляющие клетки могут быть внутривенно введены пациенту со злокачественным новообразованием. Одному пациенту может быть одномоментно введено от 106 до 1012 сенситизированных макрофагов. Для усиления эффекта могут также применяться различные иммуномодуляторы - IFNα, IFNγ, TNFα, IL-2, -4, -6 и др.
Изобретение относится к области генетической инженерии, биотехнологии, иммунологии и медицины и может быть использовано для получения противоопухолевых вакцин и лечения меланомы. Сущность изобретения составляют вакцинные композиции на основе белков теплового шока и антигенных пептидов для лечения опухолевых заболеваний, содержащие гибридный белок, состоящий из белка теплового шока семейства HSP70 и одного из специфических пептидов меланомы MAGE (A1, А2, A3) или gp 100, а также пригодный фармацевтический носитель. Предлагается также способ получения указанных гибридных белков путем экспрессии их генов в составе синтетических рекомбинантных векторов в штаммах-продуцентах клеток Е coli BL21 (DE3). Преимущество изобретения заключается в повышении иммуногенности. 3 н.п. ф-лы, 1 табл., 2 ил.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Huang С | |||
| Yu Н | |||
| et al., Potent antitumor effect elicited by superantigen-limited tumor cells thansduced with heat shock protein 70 gene, Cancer Sci., 2004, Feb., 95 (20), pp.160-167 | |||
| US 6610659 A, 26.08.2003 | |||
| US 6468540 A, 22.10.2002 | |||
| US 6875435 A, 05.04.2005. | |||

