Изобретение относится к биотехнологии и, в частности, к способу получения аминокислот методом ферментации с использованием бактерий - продуцентов аминокислот, принадлежащих к роду Escherichia coli и способных к утилизации сахарозы в качестве единственного источника углерода.
Сахароза и субстраты, содержащие сахарозу (такие как меласса), часто используются в качестве исходного соединения для получения с помощью микроорганизмов коммерческих продуктов, таких как аминокислоты, витамины и органические кислоты. В процессе получения аминокислот из углеводов стараются добиться максимальной эффективности, с которой углеродный скелет углевода превращается в желаемый продукт.
Большинство сахароза-положительных бактерий включает и фосфорилирует сахарозу с помощью фосфоенолпируват-зависимой, сахароза-6-фосфотрансферазной системы (сахарозная PTS) с получением внутриклеточного сахароза-6-фосфата. Этот фосфат гидролизуется сахароза-6-фосфат гидролазой (инвертазой или сахаразой) с получением D-глюкоза-6-фосфата и D-фруктозы, которая сама фосфорилируется АТФ-D-фруктоза-6-фосфат фосфотрансферазой (фруктокиназой). Такие системы и метаболитические пути были описаны на молекулярном уровне для грамположительных бактерий Bacillus subtilis и Streptococcus mutans (Debarboulli et al., 1991, Res. Microbiol., 142: 757-764; Sato et al., 1989, J. Bacteriol 171:263-271) и для грамотрицательных бактерий. Кроме того, описана система, кодируемая плазмидой pUR400 из кишечных бактерий (Aulkemeyer et al. , 1991, Mol. Microbiol., 5:2913-2922; Schmid et al. l988. Mol. Microbiol., 2: l-8: Schmid et al. 1991. Mol. Microbiol., 5: 941-950).
Хотя около 50% природных изолятов Escherichia coli являются сахароза-положительными, лабораторные штаммы Escherichia coli, такие как E.coli К-12, E. coli В, E. coli С, которые в настоящее время используются для выведения промышленно значимых штаммов-продуцентов, не могут утилизировать сахарозу. Однако эта способность может быть легко придана указанным штаммам путем введения генов, утилизирующих сахарозу, из сахароза-положительных штаммов E. coli или Salmonella с использованием методов конъюгации, трансдукции или клонирования (Wohlhieter et al., 1975, J. Bacteriol., 122: 401-406; Parsell and Smith), 1975, J. Gen. Microbiol., 87: 129-137;
Alaeddinoglu and Charles, 1979, J. Gen. Microbiol., 110:47-59; Livshits et al. , 1982, in: Metabolic plasmids, p. 132-134; Garsia, 1985, Mol. Gen. Genet., 201:575-577; Патент США 5, 175, 107).
Фосфоенолпируват (PEP) является одним из главных структурных блоков в нескольких путях биосинтеза. PEP связывается с диоксидом углерода с образованием оксалоацетата. Оксалоацетат служит углеродным скелетом для аспарагиновой кислоты, аспарагина, треонина, изолейцина, метионина и лизина. Кроме того, эквимолярное количество PEP конденсируется с эритроза-4-фосфатом с образованием 3-дезокси-D-арабиногептулозонат-7-фосфата (DAHP) первого промежуточного соединения в общей части пути синтеза ароматических соединений. С использованием этого метаболического пути могут быть получены такие коммерчески важные аминокислоты, как триптофан, фенилаланин и тирозин. Выход этих метаболитов может быть ограничен доступностью PEP.
В процессе гликолиза из двух молей глюкозы получаются четыре моля PEP и половина PEP обязательно расходуется для энергетического обеспечения включения глюкозы. В случае усвоения сахарозы из двух молей гексозы (глюкозы и фруктозы), образующихся из одного моля сахарозы, также получаются четыре моля PEP, но только один моль расходуется для транспорта сахарозы и, таким образом, в 1,5 раза большее количество PEP становится доступным в качестве источника углеродного скелета для биосинтеза. Вследствие этого можно увеличить выход аминокислот путем придания штаммам E.coli - продуцентам аминокислот способности к утилизации сахарозы и использования сахарозы или содержащих сахарозу субстратов в качестве источника углерода.
К настоящему моменту известен штамм ВКПМ В-3996 - продуцент треонина, созданный на основе E.coli К-12, способный утилизировать сахарозу (Патент США 5,705,371). Рестриктазный анализ и анализ нуклеотидной последовательности клонированных сахарозных генов из штамма ВКПМ В-3996 показал, что они практически идентичны генам pUR400 (инвентарные номера последовательностей в базах данных: EMBL X61005; EMBL X67750, GB M38416), кодирующим PTS-зависимый транспорт сахарозы и ее метаболизм (Lengeler et al., 1982, J. Bacteriol, 151: 468-471; Schmid et al., 1988, Mol. Microbiol., 2: 1-8; Schmid et al., 1991. Mol. Microbiol 5: 941-950).
У Escherichia coli также был найден кодируемый генами, расположенными на хромосоме, не зависимый от PTS метаболический путь утилизации сахарозы - сахарозная non-PTS система (сахарозная non-PTS) (Bockmann et al., 1992, Mol. Gen. Genet., 235:22-32). Указанный путь включает в себя систему транспорта в симпорте с протоном (пермиаза типа LacY), инвертазу, фруктокиназу и сахароза-специфический репрессор. С использованием этого non-PTS метаболического пути выход аминокислот, полученных из предшественника PEP, может быть повышен в еще большей степени потому, что транспорт сахарозы в клетку не будет связан с PEP. Однако этот подход не был использован ранее для улучшения штаммов-продуцентов аминокислот.
Целью настоящего изобретения является разработка метода получения аминокислот с использованием штаммов Escherichia coli, содержащих гены, кодирующие метаболические пути утилизации сахарозы, в особенности non-PTS метаболический путь утилизации сахарозы.
Авторами изобретения было установлено, что бактерия, принадлежащая к роду Escherichia и обладающая способностью к продукции аминокислот, производит аминокислоты более эффективно при введении сахарозных генов в бактерию. Таким образом было выполнено настоящее изобретение.
Вот то, что представляет собой настоящее изобретение:
(1) Бактерия, принадлежащая к роду Escherichia, сконструированная из не усваивающего сахарозу штамма, принадлежащего к роду Escherichia, несущая гены сахарозной PTS и обладающая способностью к продукции аминокислоты, отличной от треонина.
(2) Бактерия по п.1, где бактерией, принадлежащей к роду Escherichia, является Escherichia coli.
(3) Бактерия по п.1 или п.2, где аминокислота выбрана из группы, состоящей из гомосерина, изолейцина, лизина, валина и триптофана.
(4) Бактерия, принадлежащая к роду Escherichia. сконструированная из не усваивающего сахарозу штамма, принадлежащего к роду Escherichia, несущая гены сахарозной non-PTS и обладающая способностью к продукции аминокислот.
(5) Бактерия по п.4, где гены сахарозной non-PTS представлены по крайней мере генами, кодирующими систему транспорта в симпорте с протоном, инвертазу и фруктокиназу.
(6) Бактерия по п.4 или п.5, где бактерией, принадлежащей к роду Escherichia, является Escherichia coli.
(7) Бактерия но любому из п.4-6, где аминокислота выбрана из группы, состоящей из треонина, гомосерина, изолейцина, лизина, валина и триптофана.
(8) Способ получения аминокислот, включающий стадии выращивания бактерии по любому из п.1-7 в питательной среде с целью продукции и накопления аминокислоты в этой среде и выделения аминокислоты из культуральной жидкости.
В настоящем изобретении аминокислотами являются аминокислоты в L-конфигурации, если не сказано иное.
Настоящее изобретение будет более подробно описано ниже.
Бактерия, принадлежащая к роду Escherichia, согласно настоящему изобретению представляет собой штамм, сконструированный из не усваивающего сахарозу исходного штамма Escherichia coli, который содержит сахарозные гены, главным образом гены сахарозной non-PTS, и обладает способностью к продукции аминокислоты.
Круг бактерий Escherichia coli, не усваивающих сахарозу, не ограничен каким-либо образом при условии, что бактерия обладает способностью к продукции аминокислот или такая способность может быть ей придана. Примерами таких штаммов являются E.coli К-12, E.coli В и E.coli С, а также производные от них штаммы, более конкретно штаммы-продуценты аминокислот, упоминающиеся позже.
Бактерия согласно настоящему изобретению может быть получена путем введения генов сахарозной PTS или генов сахарозной non-PTS в штаммы-продуценты аминокислот, например, такие, как указано выше. С другой стороны, бактерия согласно настоящему изобретению может быть получена путем придания способности к продукции аминокислот бактерии, принадлежащей к роду Escherichia, в которую гены сахарозной PTS или гены сахарозной non-PTS уже введены.
Гены сахарозной non-PTS не ограничены каким-либо образом при условии, что они могут функционировать в бактерии, принадлежащей к роду Escherichia. Примером таких генов являются гены сахарозной non-PTS (csc), присутствующие в E. coli ЕС3132 (Bockmann et al., 1992, Mol. Gen. Genet., 235:22-32). Гены csc могут быть получены из E.coli К-12 W3350csc. Штамм W3350csc был депонирован во Всероссийской коллекции промышленных микроорганизмов (Россия, 113545 Москва, 1-й Дорожный проезд, 1) в соответствии с Будапештским договором под инвентарным номером BKПM В-7914.
Гены csc включают в себя гены, кодирующие систему транспорта в симпорте с протоном (пермеаза типа LacY), инвертазу, фруктокиназу и сахароза-специфический репрессор. Среди них для осуществления настоящего изобретения требуются, по крайней мере, гены, кодирующие пермеазу, инвертазу и фруктокиназу.
Аминокислота также может быть эффективно произведена путем введения сахарозной PTS в бактерию, принадлежащую к роду E.coli. В качестве примеров генов сахарозной PTS могут быть приведены гены scr, включенные в систему pUR400, кодируемую плазмидой, полученной из кишечных бактерий (Aulkemeyer et al. , 1991, Mol. Microbiol.. 5: 2913-2922: Schmid et al. 1988. Mol. Microbiol, 2: 1-8; Schmid et al, 1991, Mol. Microbiol., 5: 941-950). С другой стороны, гены сахарозной PTS могут быть получены из транспозона Tn2555 (Дорошенко и др., 1988, Молекулярная биология, 22: 645-658).
Гены сахарозной PTS и гены сахарозной non-PTS могут быть встроены в бактерию, принадлежащую роду Escherichia, например, с помощью введения рекомбинантной плазмиды, содержащей желаемые гены. В частности, желаемые гены могут быть встроены в бактерию, принадлежащую к роду Escherichia, путем введения в бактериальную клетку плазмиды, фага или транспозона, несущего желаемые гены (Berg, D.E. and Berg, C.M., 1983, Bio/Technol., 1: 417).
В качестве примера вектора могут быть приведены плазмидные векторы, такие как pBR322, pMW118, pUC19 или подобные им, и фаговые векторы, включающие фаг PIvir, mini-Mud, такой как pMu4041 или подобный ему. Примерами транспозопов являются Мu, Тn10, Тn5 или подобные им.
Введение ДНК в бактерию, принадлежащую к роду Escherichia, может быть осуществлено, например, методом D.A.Morrison (Methods of Enzymology, 68, 326 (1979)) или методом, в котором бактериальная клетка-реципиент обрабатывается хлоридом кальция для повышения проницаемости для ДНК (Mandel M. and Higa A., 1970, J. Mol. Biol, 53: 159) или подобным методом. В качестве альтернативы введение ДНК также может быть осуществлено путем трансдукции с использованием фагового вектора.
Гены сахарозной non-PTS и гены сахарозной PTS вводятся в принадлежащую к роду Escherichia бактерию-продуцент аминокислоты, что приводит к такому результату, что аминокислота получается из сахарозы. В качестве бактерии, принадлежащей к роду Escherichia, в которую вводятся гены сахарозной non-PTS и гены сахарозной PTS, могут быть использованы штаммы, уже обладающие способностью к продукции желаемой аминокислоты. Кроме того, способность к продукции аминокислоты может быть придана бактерии, в которую уже введены гены сахарозной non-PTS и гены сахарозной PTS. Примеры принадлежащих к роду Escherichia бактерий-продуцентов аминокислот приведены ниже.
(1) Бактерия - продуцент треонина
В качестве примеров принадлежащих к роду Escherichia бактерий - продуцентов треонина могут быть приведены штаммы MG422 (Гусятинер и др., 1978. Генетика, 14, 947-956), VL643 и VL2055 (см. примеры 2 и 3).
(2) Бактерия - продуцент гомосерина
В качестве примеров принадлежащих к роду Escherichia бактерий - продуцентов гомосерина могут быть приведены E.coli NZ10 и NZ10rhtA23/pAL4. Штамм NZ10 был получен как Leu+-ревертант из известного штамма С600 (Appleyard R. K., 1954, Genetics, 39:440-452). Штамм NZ10rhtA23/pAL4 был получен из штамма NZ10 (см. пример 4).
(3) Бактерия - продуцент изолейцина
В качестве примеров бактерий-продуцентов изолейцина могут быть приведены штамм E. coli 44-3-15, штамм КХ141 (ВКПМ В-4781) (ЕР-А-519113) и TDH-6/pVIC40,pMWD5 (WО97/08333).
(4) Бактерия - продуцент лизина
В качестве продуцента лизина предпочтителен штамм E.coli VL612 (пример 5). В дополнение, в качестве конкретных примеров принадлежащих к роду Escherichia бактерий - продуцентов лизина могут быть приведены мутантные штаммы, обладающие устойчивостью к аналогам лизина. Аналоги лизина - это соединения, подавляющие размножение бактерий, принадлежащих к роду Escherichia, но это подавление частично или полностью снимается, если лизин присутствует в питательной среде. Это, например, оксализин, гидроксамат лизина, S-2-аминоэтил-L-цистеин (АЕС), γ-метиллизин, хлорокапролактам и подобные им. Мутантные штаммы, обладающие устойчивостью к таким аналогам лизина, получаются с применением обычных методов получения искусственных мутаций в бактериях, принадлежащих к роду Escherichia. Примерами бактериальных штаммов, которые могут быть использованы для продукции лизина, являются Escherichia coli АJ11442 (депонирован как FERM P-5084 и NRRL В 12185, см. выложенную патентную заявку Японии 56-18596 или патент США 4,346,170) и Escherichia coli VL611. Штамм Escherichia coli AJ11442 был депонирован в Национальном институте биологической науки и технологии человека при Агентстве промышленной науки и технологии (305, 13, Higashi Ichome, Tsukubashi, Ibarakiken, Japan) 5 мая 1981 (инвентарный номер FERM P-5084) и переведен из первоначального депозитария в Международный депозитарий 29 октября 1987 в соответствии с Будапештским договором с присвоением инвентарного номера FERM BP-1543. В микроорганизме, описанном выше, у аспартокиназы утрачена чувствительность к ингибированию лизином по типу обратной связи.
Кроме того, в качестве примера могут быть приведены бактерии - продуценты треонина, потому что у аспартокиназы этих бактерий - продуцентов треонина также, как правило, утрачена чувствительность к ингибированию лизином. Примером бактерии - продуцента треонина, принадлежащей к роду E.coli, является MG442 (Гусятинер и др., 1978. Генетика, 14:947-956).
Ген(ы), кодирующие фермент(ы) биосинтеза лизина, могут быть улучшены в бактериях, упомянутых выше. Например, такой ген может быть представлен геном, кодирующим фосфоенолпируваткарбоксилазу, в котором была получена мутация, чтобы чувствительность к ингибированию аспарагиновой кислотой по типу обратной связи была утрачена (см. выложенную патентную заявку Японии 7-83714).
(5) Бактерия - продуцент валина
Конкретным примером бактерий - продуцентов валина являются E.coli VL1970 (ВКПМ В-4411) (см. ЕР-А-519113) и VL1971 (см. пример 6). Кроме того, в качестве примера могут быть приведены бактерии, принадлежащие к роду Escherichia, содержащие гены биосинтеза валина, у которых механизм регуляции в значительной степени нарушен. Такие бактерии могут быть получены при введении в бактерии, принадлежащие к роду Escherichia, оперона ilvGMEDA, который, предпочтительно, не эскпрессирует треонин дезаминазу и в котором нарушена аттенуация (выложенная патентная заявка Японии 8-47397).
Оперон ilvGMEDA может быть получен из хромосомной ДНК E.coli путем гибридизации колоний или с помощью ПЦР с использованием олигонуклеотидов, приготовленных в соответствии с нуклеотидной последовательностью оперона, полная последовательность которого опубликована (Nucleic Acid Res., 1987, 15: 2137). Введение фрагмента ДНК, включая оперон ilvGMEDA, может быть осуществлено способами с использованием плазмиды, фага или транспозона, как это описано выше.
(6) Бактерия - продуцент триптофана
Конкретным примером бактерий - продуцентов триптофана являются E.coli SV164/pGH5 (см. пример 7), AGX17/pGX44 (NRRL В-12263) и AGX6/pGX50 aroP (NRRL В-12264) (патент США 4,371,614), AGX17/pGX50,pACKG4-pps (WО97/08333).
(7) Бактерия - продуцент фенилаланина
Примером бактерий-продуцентов фенилаланина являются E.coli АJ12604 (FERM ВР-3579) (ЕР-А-488424).
Аминокислоты могут быть получены из сахарозы с высокой эффективностью при выращивании описанных выше бактерий, в которых введены гены сахарозной non-PTS и гены сахарозной PTS и которые обладают способностью к продукции аминокислот в питательной среде, содержащей сахарозу, для получения и накопления аминокислоты в питательной среде и сбора аминокислоты из питательной среды. Примерами таких аминокислот являются, предпочтительно, треонин, гомосерин, изолейцин, лизин, валин, триптофан, тирозин, фенилаланин и метионин, а более предпочтительно, треонин, гомосерин, изолейцин, лизин, валин, триптофан.
В способе получения аминокислот согласно настоящему изобретению выращивание бактерии, принадлежащей к роду Escherichia, накопление и очистка аминокислоты из жидкой питательной среды могут быть осуществлены способом, подобным обычным методам ферментации, в которых аминокислота получается с использованием бактерии. Питательная среда, используемая для выращивания, может быть как искусственной, так и натуральной, при условии, что в нее включены источники углерода и азота, минеральные добавки и, если необходимо, умеренные количества питательных добавок, необходимые для роста используемых бактерий. Сахароза используется в качестве главного источника углерода. Небольшое количество отличных от сахарозы источников углерода может содержаться в питательной среде в качестве вспомогательного источника углерода. В качестве источника азота используются различные соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов и микробный ферментализат. В качестве минеральных добавок используются фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, карбонат кальция.
Выращивание происходит предпочтительно при аэробных условиях, таких как перемешивание и аэрация с взбалтыванием при температуре от 20 до 40oС, предпочтительно от 30 до 38oС. рН среды обычно находится в интервале от 5 до 9, предпочтительно между 6,5 и 7,2. рН среды может быть подкорректнрован с помощью аммиака, карбоната кальция, различных кислот, оснований и буферных растворов. Обычно, выращивание в течение от 1 до 3 дней приводит к накоплению целевой аминокислоты в жидкой питательной среде.
После выращивания твердые остатки, такие как клетки, удаляются из жидкой питательной среды методом центрифугирования и фильтрации на мембранах и затем целевая аминокислота может быть собрана и очищена методами ионообменной хроматографии, концентрации и фракционной кристаллизации.
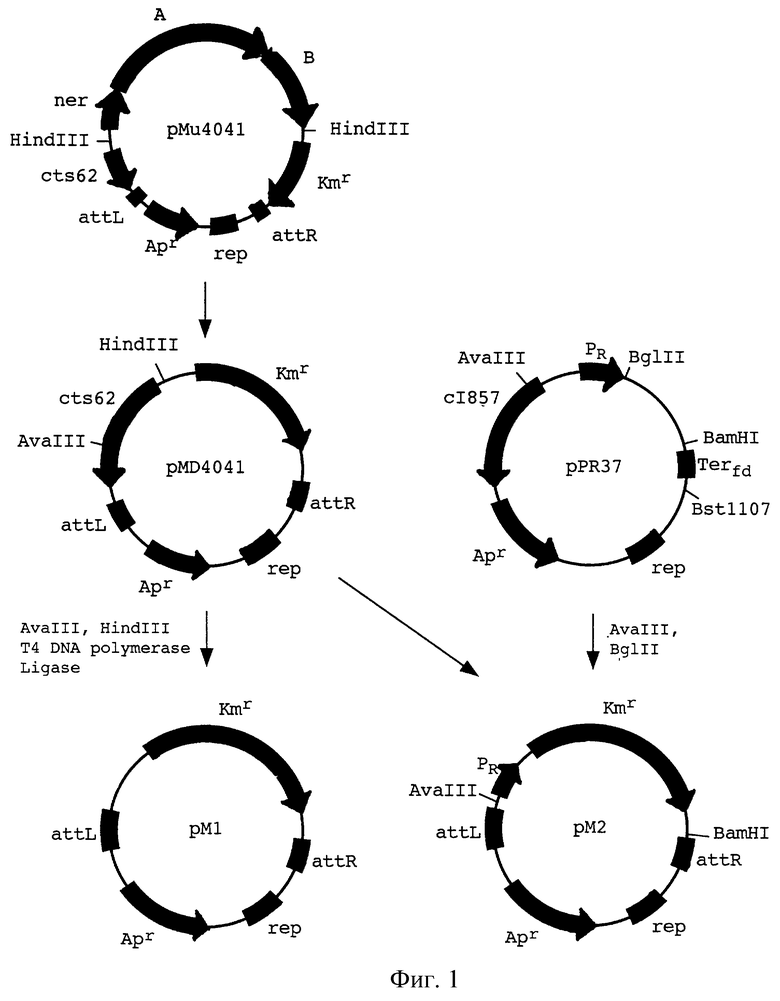
На фиг.1 показана конструкция плазмид рМ1 и рМ2 - производных от mini-Mud 4041.
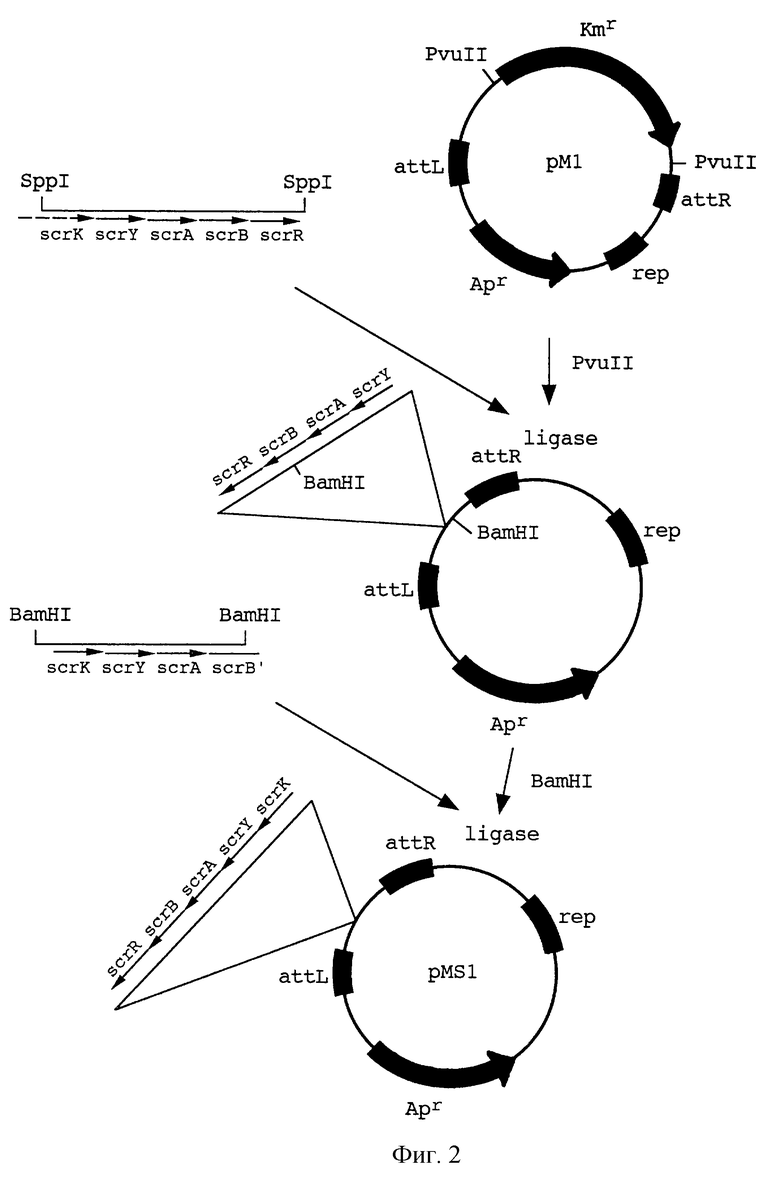
На фиг.2 показана схема клонирования гена scr в плазмиду рМ1.
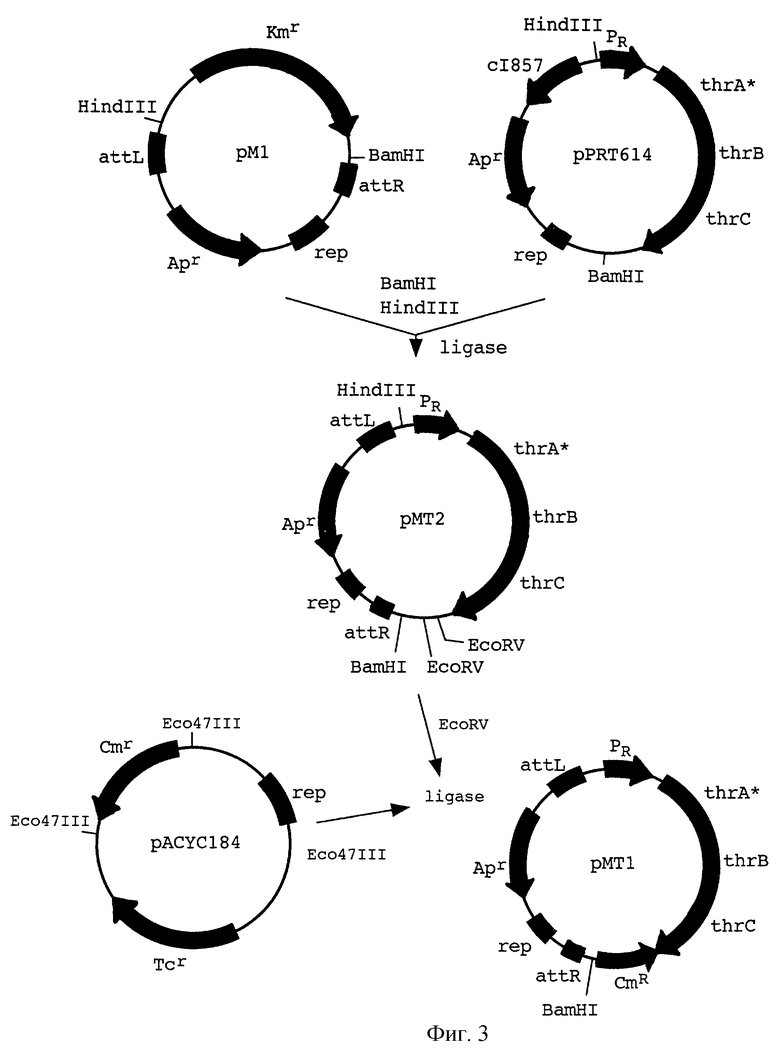
На фиг.3 показана конструкция плазмид рМТ1 и рМТ2.
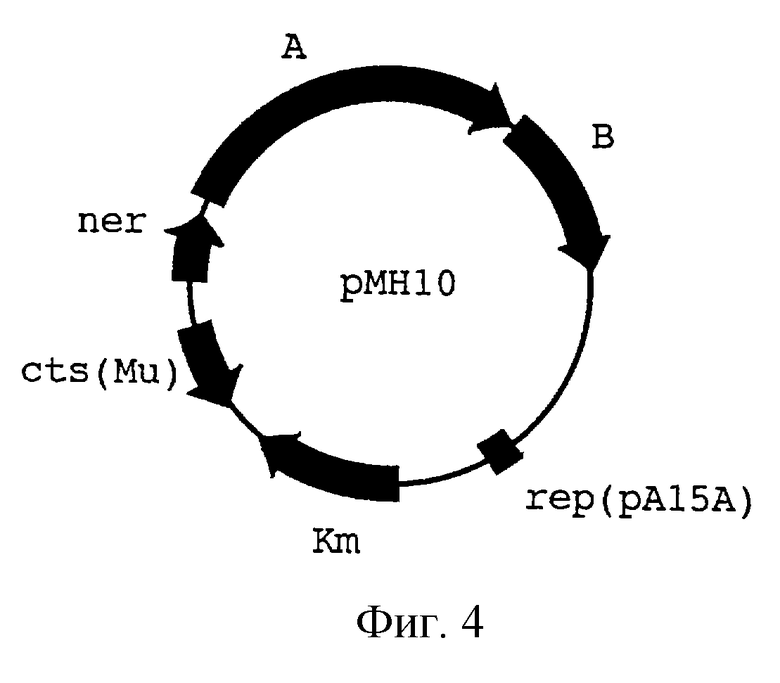
На фиг.4 показано устройство плазмиды рМН10, несущей ген KmR, гены А и В фага Мu, кодирующие транспозазу фага Мu, ген ner, кодирующий негативный регулятор, и ген cts62, кодирующий репрессор фага Мu.
Здесь и далее, настоящее изобретение будет более конкретно разъяснено со ссылками на следующие примеры.
Пример 1. Приготовление донорных генов сахарозной non-PTS или генов сахарозной PTS
(1) Гены сахарозной PTS
В качестве донора генов scr утилизации сахарозы с помощью сахарозной PTS был использован штамм VD1. Этот штамм был получен следующим образом. Транспозон Тn2555 содержит гены scr (Дорошенко и др., 1988. Молекулярная биология, 22: 645-658). Рестриктазный анализ и частичное определение нуклеотидной последовательности выявили, что гены scr транспозона Тn2555 идентичны генам в pUR400 (инвентарные номера: EMBL X61005; EMBL X67750, GB М38416), контролирующим транспорт и метаболизм сахарозы через систему PTS.
Гены scr из транспозона Тn2555 были клонированы в плазмиду рМ1, которая является производной mini-Mud вектора рМu4041, полученной в результате делеции генов фага Мu, кодирующих транспозазу и репрессор (М. Faelen. Useful Mu and mini-Mu derivatives. In: Phage Mu. Symonds et al., eds. Cold Spring Harbor Laboratory, New York, 1987, pp.309-316). Это было сделано в две стадии. На первой стадии SspI фрагмент pBRS5.2 (pBR325::Tn2555) (Дорошенко и др. , 1988. Молекулярная биология, 22:645-658), содержащий гены scrYABR и только часть гена scrK, был введен в рМ1, предварительно обработанную PvuII, с заменой гена kan.
Указанная выше плазмида рМ1 была получена следующим образом (фиг.1). Плазмида рМи4041 расщеплялась рестриктазой HindIII и закольцовывалась обратно, чтобы удалить гены A и В, кодирующие транспозазу фага Mu, и ген ner, кодирующий негативной регулятор, с получением плазмиды pMD4041. Затем плазмида pMD4041 расщеплялась рестриктазами AvaIII и HindIII, концы затуплялись ДНК полимеразой фага Т4 с последующем закольцовыванием с целью удаления гена сts62, кодирующего репрессор фага Мu.
На второй стадии, BamHI фрагмент полученной плазмиды был замещен BamHI фрагментом плаэмиды pBRS5.2, восстанавливающим ген scrK. Таким образом, весь сахарозный кластер из Тп2555 был клонирован в плазмиду, содержащую также маркер ampR и концы фага Мu. Эта плазмида, обозначенная как pMSl, содержит транспозируемый фрагмент ДНК mini-Mu-scrKYABR (фиг.2).
Для введения фрагмента mini-Mu-scrKYABR в бактериальную хромосому была использована стандартная процедура. Плазмида pMSl была введена в клетки MG1655/pMH10. Mu-транспозаза, кодируемая плазмидой рМН10 (производная pACYC177, несущая ген КmR гены А и В фага Мu, кодирующие Мu-транспозазу, ген сts62, кодирующий репрессор фага Мu, и ген сI857, кодирующий репрессор фага лямбда), была индуцирована путем инкубации в течение 15 мин при 42oС немедленно после трансформации. Сахароза-положительные (Scr+) клоны отбирались на чашках с агаром М9, содержащим 0,2% сахарозы в качестве единственного источника углерода, при 30oС, отмывались и выращивались в LB бульоне (J. Miller. Experiments in molecular genetics. Cold Spring Harbor laboratory. New York, 1972), не содержащем антибиотиков, в течение 48-72 часов. Затем соответствующие разведения культуры высевались на чашки с агаром М9, содержащим 0,2% сахарозы. Было отобрано и проверено несколько десятков клонов AmpS, KmS. Было показано, что они не содержат плазмид. Среди них был выбран штамм VD1 (MG1655: :mini-Mu-scrKYABR), являвшийся прототрофным, быстрорастущим сахароза-положительным штаммом.
Кроме того, штамм VL478, несущий плазмиду pVG478, содержащую сахарозные гены в транспозоне Тn2555 (Молекулярная генетикаб микробиология и вирусология, 6:23-28, 1987), также был использован в качестве донора генов scr. Штамм VL478 был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) под инвентарным номером ВКПМ В-7915.
Указанные выше штаммы были использованы в качестве доноров генов scr в последующих примерах.
(2) Гены сахарозной non-PTS
В качестве источника генов утилизации сахарозы non-PTS (csc) был использован штамм E.coli К 12 W3350csc. Этот штамм содержит гены csc из E.coli EC3132 (Bockmann et al., 1992, Mol. Gen. Genet., 235:22-32). Штамм W3350csc был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) под инвентарным номером ВКПМ В-7914. Гены csc содержат гены, кодирующие пермеазу, фруктокиназу, инвертазу и репрессор.
Пример 2: Получение штамма E.coli - продуцента треонина, способного к утилизации сахарозы, и продукция треонина с использованием штамма (1)
Штаммом-реципиентом, в который были введены гены PTS, был следующим образом заново сконструированный штамм E.coli VL643. В известный штамм E.coli MG442 (Гусятинер и др., Генетика 14:947-956, 1978) ВКПМ В-1628) трансдуцировали мутацию rhtA23 из штамма 472T23/pYN7 (ВКПМ В-2307) с целью получения штамма VL643. Мутация rhtA23 придает устойчивость к высокой концентрации треонина (>40 мг/мл) или гомосерина (>5 мг/мл) и усиливает продукцию треонина (ТЕЗИСЫ 17 международного конгресса по биохимии и молекулярной биологии совместно с ежегодной конференцией американского общества по биохимии и молекулярной биологии, Сан-Франциско, Калифорния, 24-29 августа 1997, тезисы 457).
Затем полученный штамм VL643 - продуцент треонина был инфицирован фагом P1vir, выращенным на донорном штамме VL478. Трансдуктанты отбирались на минимальной среде М9, содержащей 0.2% сахарозы, в качестве единственного источника углерода. Таким образом был получен штамм VL644. Этот штамм и исходный штамм выращивались при 37oС в течение 18 часов в питательном бульоне и по 0,3 мл каждой из полученных культур засевались в 3 мл питательной среды для ферментации следующего состава (см. ниже), содержащейся в пробирках 20•200 мм, и выращивались при 37oС в течение 72 часов в роторной мешалке.
Состав питательной среды для ферментации, г/л:
Сахароза (или глюкоза) - 50,0
(NH4)2SO4 - 10,0
К2НРO4 - 1,0
NaCl - 1,0
MgSO4*7H2O - 0,8
FeSO4*7H2O - 0,02
MnSO4*5H2O - 0,02
Гидрохлорид тиамина - 0,002
СаСО3 - 20,0
(Каждое из соединений, MgSO4*7H2O и СаСО3, стерилизовалось по отдельности).
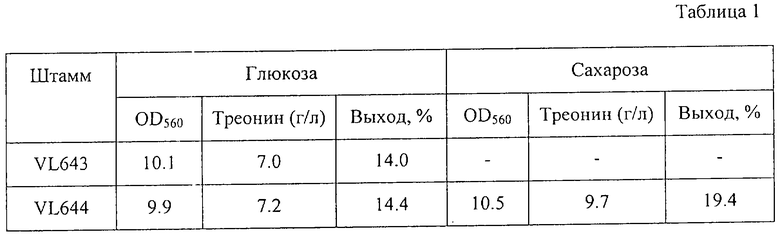
С помощью известных методов определялись накопленное в питательной среде количество треонина после выращивания и поглощение питательной среды (OD) при 560 нм. Результаты представлены в Таблице 1.
Как видно из Таблицы 1, оба штамма, VL643 и VL644, росли одинаково в питательной среде с глюкозой и накапливали примерно одинаковое количество треонина. Кроме того, штамм VL644 хорошо рос в питательной среде с сахарозой и накапливал в этих условиях больше треонина с более высоким выходом.
Пример 3: Получение штамма E.coli - продуцента треонина, способного к утилизации сахарозы, и продукция треонина с использованием штамма (2)
В качестве реципиентного штамма, в который вводились гены PTS, был сконструирован штамм E.coli VL2055.
Штамм VL2055 был получен из известного штамма E.coli ВКПМ В-3996 (патент США 5,705,371). Штамм ВКПМ В-3996, исходным для которого был штамм E.coli TDH-6, является дефектным по гену thrC и ассимилирующим сахарозу, в котором ген ilvA имеет мутацию типа "leaky". Штамм ВКПМ В-3996 содержит плазмиду pVIC40, полученную интеграцией оперона thrA*BC, включающего ген thrA*, кодирующий фермент аспартокиназу I - гомосериндегидрогеназу I (AKI-HDI), у которого существенно понижена чувствительность к ингибированию треонином, в вектор, полученный на основе RSF1010.
Штамм VL2055 был сконструирован из В-3996 в две следующие стадии.
Сначала был отобран безплазмидный производный штамма ВКПМ В-3996, TDH-6, после спонтанной утраты плазмиды pVIC40. Затем известным методом (NTG мутагенез) была получена мутация, инактивирующая ген kan в транспозоне Тn5, который интегрирован в ген tdh штамма TDH-6. Затем, не утилизирующий сахарозу штамм, производный полученного штамма, был отобран после удаления генетических детерминант ассимиляции сахарозы. Так был получен штамм VL2053.
С другой стороны, плазмида pPRT614 (EP 0593792), содержащаяся в E.coli ВКПМ В-5318, расщеплялась рестриктазами HindIII и ВатHI для того, чтобы вырезать фрагмент, содержащий треониновый оперон, находящийся под контролем промотора РR фага лямбда. Треониновый оперон содержит мутацию в гене thrA (thrA*), которая сообщает ферменту аспартокиназа I - гомосериндегидрогеназа I нечувствительность к ингибированию треонином но типу обратной связи. Полученный фрагмент был клонирован в плазмиду рМ1, производную mini-Mud вектора pMu4041 (M.Faelen. Useful Mu and mini-Mu derivatives. In: Phage Mu. Symonds et al. , eds. Cold Spring Harbor Laboratory, New York, 1987, pp.309-316) с целью получения плазмиды рМТ2 (фиг.3).
В дополнение, ген cat из Тп9, придающий устойчивость к хлорамфениколу, из pACYC184 был клонирован в рМТ2. Таким образом была получена плазмида рМТ1, содержащая транспозируемую конструкцию: гены РR-thrА*ВС и cat, фланкированные концами фага Mu (mini-Mu-thrA*BC-cat).
Эта плазмида была введена в клетки E.coli С600 (рМН10). Mu транспозаза, кодируемая плазмидой рМН10 (производная pACYC177, содержащая ген КmR, гены фага Mu А и В, кодирующие транспозазу, ген ner, кодирующий негативный регулятор, и ген cts62, кодирующий репрессор фага Мu, см. фиг.4), была индуцирована инкубацией в течение 15 мин при 42oС немедленно после трансформации.
Клоны (СmR), устойчивые к хлорамфениколу, были отобраны на чашках с LB агаром, содержащим 15 мг/л хлорамфеникола, при 30oС. Несколько десятков клонов KmS были отобраны и проверены. Было показано, что большинство из них не содержало плазмиды. Затем гены PR-thrA*BC-cat из хромосомы одного из отобранных клонов С600 Thr+, CmR были трандуцированы с помощью P1vir в штамм VL2053, полученный на первой стадии, что дало новый безплазмидный штамм VL2055 - продуцент треонина.
Штамм VL2055 продуцент треонина был инфицирован фагом P1vir, выращенным на донорных штаммах VD1 или W3350csc. Трансдуктанты отбирались на минимальной среде М9, содержащей 50 мг/л изолейцина и 0,2% сахарозы в качестве единственного источника углерода. Таким образом были получены штаммы VL2055 Scr и VL2055 Csc соответственно. Каждый из этих штаммов и исходный штамм выращивались при 37oС в течение 18 часов в питательном бульоне и по 0,3 мл полученной культуры вносились в 3 мл питательной среды для ферментации, имеющей следующий состав (см. ниже), содержащейся в пробирках 20•200 мм, и выращивались при 37oС в течение 72 часов на роторной мешалке.
Состав питательной среды для ферментации (г/л):
Сахароза (или глюкоза) - 80,0
Изолейцин - 0,1
(NH4)2SO4 - 22,0
К2НРO4 - 2,0
NaCl - 0,8
MgSO4*7H2O - 0,8
FeSO4*7H2O - 0,02
MnSO4*5H2O - 0,02
Гидрохлорид тиамина - 0,2
Дрожжевой экстракт - 1,0
СаСО3 - 30,0
(Каждое из соединений, MgSO4*7H2O и СаСО3, стерилизовалось по отдельности)
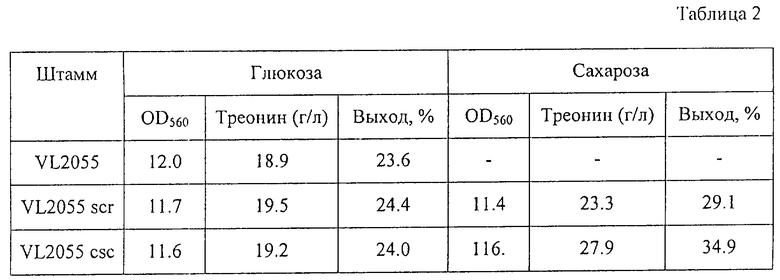
С помощью известных методов были определены количество накопленного в питательной среде треонина после выращивания и поглощение питательной среды (OD) при 560 нм. Результаты представлены в Таблице 2.
Как видно из таблицы, оба штамма, утилизирующих сахарозу, VL2055 Scr и VL2055 Csc, имели одинаковые ростовые характеристики и накапливали почти такое же количество треонина, как их исходный штамм VL2055, в случае, когда они выращивались в питательной среде, содержащей глюкозу. Однако эти штаммы накапливали больше треонина с более высоким выходом при выращивании их в питательной среде, содержащей сахарозу. Кроме того, штамм VL2055 Csc (содержащий гены сахарозной non-РTS) был более продуктивен в этих условиях, чем штамм VL2055 Scr (содержащий гены сахарозной PTS).
Пример 4. Получение штамма E.coli - продуцента гомосерина, способного к утилизации сахарозы, и получение гомосерина с использованием этого штамма.
В качестве реципиентного штамма, продуцирующего гомосерин, в который были введены гены сахарозной PTS, был сконструирован штамм E.coli NZ10 rhtA23/pAL4 на основе штамма NZ10. Штамм NZ10 (thrB) является leuB+-ревернатном, полученным из штамма E.coli С600 (thrB, leuB) (Appleyard R. K. , Genetics, 1954, 39:440-452). Затем в этот штамм была взедена мутация rhtA23, как описано в Примере 2, с получением штамма NZ10 rhtA23. Этот штамм был трансформирован плазмидой pAL4, представляющей собой вектор pBR322, в который вставлен ген thrA, кодирующий аспартокиназа-гомосерин дегидрогеназу I.
Штамм Е.coli NZ10 rhtA23/pAL4 - продуцент гомосерина был инфицирован фагом P1vir, выращенным на донорном штамме VD1. Трансдуктанты отбирались на минимальной среде М9, содержащей 0,2% сахарозы и 50 мг/л треонина. Таким образом был получен штамм NZ10 rhtA23 scr/pAL4. Этот штамм и исходный штамм выращивались при 37oС в течение 18 часов в питательном бульоне и по 0,3 мл полученной культуры вносились в 3 мл питательной среды для ферментации, содержащейся в пробирках 20•200 мм, и выращивались при 37oС в течение 48 часов на роторной мешалке. Питательная среда для ферментации имела такой же состав, как описано в Примере 3, за исключением того, что 0,2 г/л треонина было добавлено вместо изолейцина.
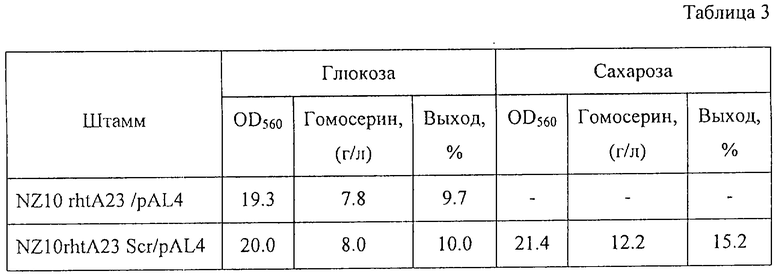
С помощью известных методов были определены количество накопленного в питательной среде гомосерина после выращивания и поглощение питательной среды при 560 нм. Результаты представлены в Таблице 3.
Как показано в Таблице 3, штамм NZ10 rhtA23 Scr/pAL4 и штамм исходный NZ10 rhtA23 /pAL4 росли примерно одинаково и накапливали примерно одинаковое количество гомосерина при выращивании в ферментационной среде, содержащей глюкозу. Однако штамм NZ10 rhtA23 Scr/pAL4 накапливал больше гомосерина с большим выходом при выращивании в ферментационной среде, содержащей сахарозу.
Пример 5. Получение штамма E.coli - продуцента изолейцина, способного к утилизации сахарозы, и получение изолейцина с использованием этого штамма
В качестве бактерии - продуцента изолейцина, принадлежащей к роду Escherichia, был использован штамм E.coli K-12 44-3-15. Этот штамм был сконструирован следующим образом. В качестве исходного был использован штамм дикого типа E.coli K-12 ВКПМ В-7. После последовательных процедур мутагенеза с использованием NTG и отбора на устойчивость к валину, 4-аза-DL-лейцину и 3-гидрокси-DL-лейцину был получен штамм 44, содержащий по крайней мере две мутации в опероне ilvGMEDA: мутацию в гене ilvG (ilvG*), восстанавливающую активность синтетазы II ацетогидроксикислот, и мутацию в гене ilvA (ilvА*), придающую треониндезаминазе нечувствительность к ингибированию изолейцином по типу обратной связи. Этот штамм может продуцировать некоторое количество изолейцина.
С другой стороны, на основе плазмиды pVR4 (Гаврилова и др., 1988, Биотехнология, 4:600-608), содержащей гены ilvG5MEDA7434YC, путем введения BamHI - линкеров по местам узнавания DraIII и XmaIII была сконструирована плазмида pVR72. Затем BamHI фрагмент плазмиды pVR72, содержащий гены ilvG5MEDA7434YC (ilvG*MEDA*YC) с удаленным промотором и аттенуатором, был клонирован в плазмиду рМ2 - производную mini-Mud вектора рМu4041, содержащую промотор PR фага лямбда. Полученная плазмида была использована для введения конструкции mini-Mu-PR-ilvG*MEDA*YC в хромосому штамма 44(рМН10), как это описано выше. После индукции транспозазы Мu была проверена способность клонов к продукциии изолейцина. Среди них был выбран наиболее продуктивный штамм 44-3. В заключение, конструкция mini-Mu-PR-thrA*BC-cat из С600 Thr+, CmR была введена в штамм 44-3, как это описано выше. Таким образом был получен штамм 44-3-15.
Штамм 44-3-15 - продуцент изолейцина был инфицирован фагом P1vir, выращенным на донорных штаммах VD1 или W3350csc. Трансдуктанты были отобраны на минимальной среде М9, содержащей 0,2% сахарозы в качестве единственного источника углерода. Таким образом были получены штаммы 44-3-15 Scr и 44-3-15 Csc.
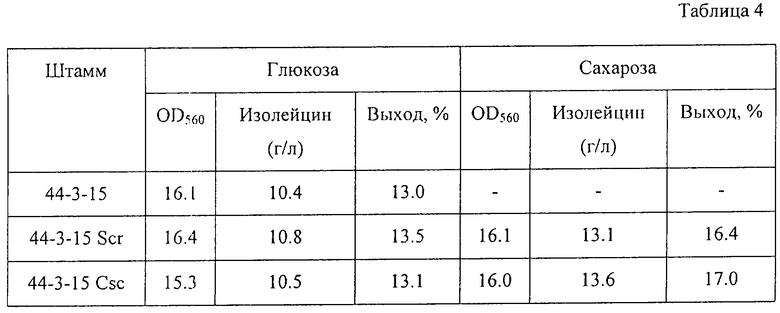
Эти штаммы и исходный штамм выращивались при 37oС в течение 18 часов в питательном бульоне и по 0,3 мл полученной культуры вносились в 3 мл питательной среды для ферментации, содержащейся в пробирках 20•200 мм, и выращивались при 37oС в течение 72 часов на роторной мешалке. Ферментационная среда имела такой же состав, как описано в Примере 3, за исключением того, что изолейцин не добавлялся. С помощью известных методов были определены количество накопленного в среде изолейцина после выращивания и поглощение среды при 560 им. Результаты представлены в Таблице 4.
Как видно из таблицы, оба штамма, утилизирующих сахарозу, 44-3-15 Scr и 44-3-15 Csc, имели одинаковые ростовые характеристики и накапливали почти одинаковое количество изолейцина в сравнении с их исходным штаммом 44-3-15 в случае, когда они выращивались в среде, содержащей глюкозу. Однако эти штаммы накапливали больше треонина с более высоким выходом при выращивании их в среде, содержащей сахарозу. Кроме того, штамм 44-3-15 Csc (содержащий гены сахарозной non-PTS) был немного более продуктивен в этих условиях, чем штамм 44-3-15 Scr (содержащий гены сахарозной PTS).
Пример 6. Получение штамма E. coli - продуцента лизина, способного к утилизации сахарозы, и получение лизина с использованием этого штамма
В качестве реципиента был использован штамм E.coli VL612 - продуцент лизина. Этот штамм был получен из известного штамма E.coli Gif102 (Theze, J. and I Siant-Girons, I. 1974. J. Bacteriol., 118:990-998) в две стадии. На первой стадии были отобраны мутантные штаммы, устойчивые к S-(2-аминоэтил)-L-цистеину в концентрии 2 мг/мл, и среди них найден штамм VL611, способный к продукции лизина. На второй стадии в штамм VL611 была введена мутация rhtA23, как описано выше, с получением штамма VL612.
Штамм VL612 был инфицирован фагом P1vir, выращенным на донорном штамме VL478. Трансдуктанты были отобраны на минимальной среде М9, содержащей 50 мг/л гомосерина и 0,2% сахарозы в качестве единственного источника углерода. Таким образом был получен штамм VL613 (ВКПМ В-3423). Этот штамм и исходный штамм выращивались при 37oС в течение 18 часов в питательном бульоне и по 0,3 мл полученной культуры вносились в 3 мл питательной среды для ферментации, содержащейся в пробирках 20•200 мм, и выращивались при 37oС в течение 72 часов на роторной мешалке. Ферментационная среда имела такой же состав, как описано в Примере 2, за исключением того, что был добавлен гомосерин в концентрации 0.2 г/л. С помощью известных методов были определены количество накопленного в питательной среде изолейцина после выращивания и поглощение среды при 560 нм.
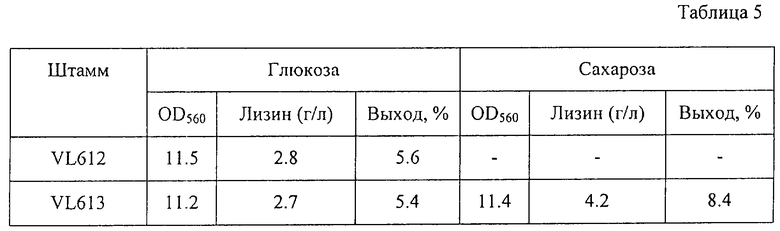
Результаты представлены в Таблице 5.
Как видно из Таблицы 5, штаммы VL612 и VL613 росли примерно одинаково в среде, содержащей глюкозу, и накапливали примерно одинаковое количество лизина. Однако штамм VL613 накапливал больше лизина с более высоким выходом при выращивании в среде, содержащей сахарозу.
Пример 7. Получение штамма E. coli - продуцента валина, способного к утилизации сахарозы, и получение валина с использованием этого штамма.
В качестве бактерии - продуцента валина, принадлежащей к роду Escherichia, был использован штамм VL1971. Этот штамм является производным от известного штамма VL1970 (ВКПМ В-4411, патент США 5,658,766), в который способом, описанным в Примере 1, была введена мутация rhtA23.
Штамм E. coli VL1971 был инфицирован фагом P1vir выращенным на донорном штамме VL478, и высеян на минимальную среду М9, содержащую 0,2% сахарозы в качестве единственного источника углерода. Трансдуктанты, выросшие после 40 часов, были отобраны, очищены и среди них был выбран штамм VL1972 (ВКПМ В-4413) - продуцент валина, способный утилизировать сахарозу.
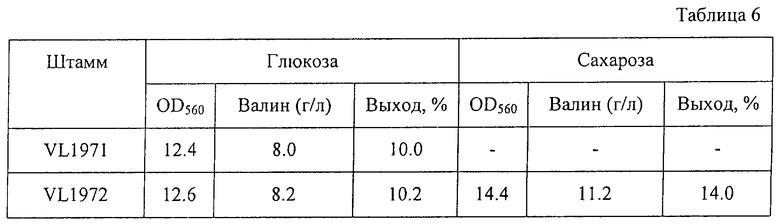
Штаммы VL1971 и VL1972 выращивались при 37oС в течение 18 часов в питательном бульоне и по 0,3 мл полученной культуры вносились в 3 мл питательной среды для ферментации, содержащейся в пробирках 20•200 мм, и выращивались при 37oС в течение 72 часов на роторной мешалке. Ферментационная среда имела такой же состав, как описано в Примере 3. С помощью известных методов были определены количество накопленного в среде валина после выращивания и поглощение среды при 560 нм. Результаты представлены в Таблице 6.
Как видно из Таблицы 6, штаммы VL1971 и VL1972 росли примерно одинаково и накапливали примерно одинаковое количество валина. Однако штамм VL1972 накапливал больше валина с более высоким выходом при выращивании в среде, содержащей сахарозу.
Следует отметить, что гены сахарозной PTS придают большую продуктивность продуцентам валина, при том что фосфоенолпируват не требуется для синтеза валина.
Пример 8. Получение штамма E.coli - продуцента триптофана, способного к утилизации сахарозы, и получение триптофана с использованием этого штамма.
В качестве бактерии-реципиента, принадлежащей к роду Escherichia, был использован штамм SV164/pGH5 (WО94/08031).
Штамм SV164/pGH5 - суперпродуцент триптофана был инфицирован фагом P1vir, выращенным на донорных штаммах VD1 или W3350csc. Трансдуктанты были отобраны на минимальной среде М9, содержащей 50 мг/л тирозина, 50 мг/л фенилаланина, 0.2% сахарозы в качестве единственного источника углерода и 15 мг/л тетрациклина. Таким образом были получены штаммы SV164scr/pGH5 и SV164csc/ pGH5 соответственно.
Эти штаммы и исходный штамм выращивались при 29oС в течение 18 часов в питательном бульоне и по 0,3 мл полученной культуры вносились в 3 мл питательной среды для ферментации, содержащейся в пробирках 20 х 200 мм, и выращивались при 29oС в течение 40 часов на роторной мешалке.
Состав питательной среды для ферментации, г/л:
Сахароза (или глюкоза) - 40,0
Фенилаланин - 0,1
Тирозин - 0,1
(NH4)2SO4 - 15,0
К2НРO4 - 1,5
NaCl - 0,5
MgSO4*7H2O - 0,3
CaCl2*2H2O - 0,015
FeSO4*7H2O - 0,075
Цитрат натрия - 1,0
Na2MoO4*2H2O - 0,00015
H3ВО3 - 0,0025
CoCl2*6H2O - 0,0007
CuSO4*5H2O - 0,00025
MnCl2*4H2O - 0,0016
ZnSO4*7H2O - 0,0003
Гидрохлорид тиамина - 0,005
Пиридоксин - 0,03
Кукурузный экстракт сухой - 2,0
(AJINOMOTO) СаСО3 - 30,0
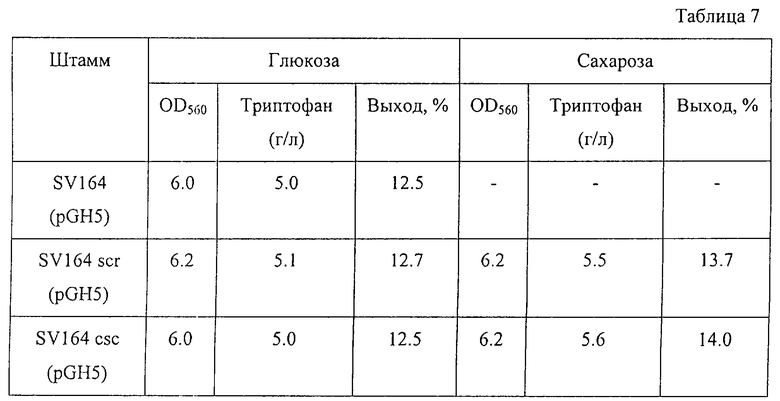
Тетрациклин - 0,015
С помощью известных методов были определены количество накопленного в питательной среде триптофана после выращивания и поглощение среды при 560 нм. Результаты представлены в Таблице 7. Как видно из Таблицы 7, оба штамма SV164scr (pGH5) и SV164csc (pGH5), утилизирующих сахарозу, имели примерно одинаковые характеристики роста и накапливали примерно такое же количество триптофана, как и исходный штамм SV164/pGH5 при выращивании в питательной среде, содержащей глюкозу. Однако эти штаммы накапливали больше триптофана с более высоким выходом при выращивании в питательной среде, содержащей сахарозу. Более того, штамм SV164csc/pGH5 (содержащий гены сахарозной non-PTS) был более продуктивен в этих условиях, чем штамм SV164scr/pGH5 (содержащий гены сахарозной PTS).
Изобретение относится к биотехнологии, а именно к способу получения аминокислот методом ферментации с использованием бактерий-продуцентов аминокислот, принадлежащих к роду E.coli. Штамм бактерии E.coli, сконструированный на основе не усваивающего сахарозу штамма E.coli путем введения генов фосфоенолпируват-зависимой, сахароза-6-фосфотрансферазной системы (сахарозная РТС) или генов системы транспорта в симпорте с протоном (пермеаза типа Lac Y), инвертозы, фруктокиназы и сахароза-специфического репрессора (сахарозная non-PTS), выращивают в питательной среде и выделяют аминокислоту из культуральной жидкости. Штамм E.coli VL 2055, содержащий гены сахарозной non-PTS, - продуцент треонина. Штамм E.coli NZ10 rht A23/pAL4, содержащий гены сахарозной PTS, - продуцент гомосерина. Штамм E.coli 44-3-15, содержащий гены сахарозной non-PTS, - продуцент изолейцина. Штамм E.coli VL612, содержащий гены сахарозной PTS, - продуцент лизина. Штамм E.coli VL613 (ВКПМ В-3423) - продуцент лизина. Штамм E.coli VL 1971, содержащий гены сахарозной PTS, - продуцент валина. Штамм E.coli VL 1972 (ВКПМ В-4413 - продуцент валина. Штамм E. coli SV 164/pGH5, содержащий гены сахарозной non-PTS, - продуцент триптофана. Изобретение позволяет повысить экспрессию аминокислот. 10 с. и 3 з.п. ф-лы, 4 ил., 7 табл.
| TSUNEKAWA H | |||
| et al | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Appl Environ Microbiol, 1992 Jun; 58 (6); р | |||
| Регулятор перегретого пара | 1925 |
|
SU2081A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| J | |||
| Устройство для видения на расстоянии | 1915 |
|
SU1982A1 |
| US 5705371 А, 06.01.1998. | |||
