Настоящее изобретение относится к способу получения L-аминокислот, а именно к способу получения L-треонина, L-глутаминовой кислоты, L-гомосерина, L-метионина, L-аргинина, L-пролина или L-изолейцина с использованием бактерии, принадлежащей к роду Escherichia.
Такие L-аминокислоты, как L-треонин и L-глутаминовая кислота, обычно производятся методамии ферментации с использованием преимущественно кориноформных бактерий, принадлежащих роду Brevibacterium, Corynebacterium и Microbacterium или их мутантным разновидностям ("Amino Acid Fermentation", Gakkai Shuppan Center, p. 195-215, 1986).
С другой стороны, известен метод выведения бактерий, производящих L-аминокислоты и принадлежащих роду Escherichia, с помощью техники генетической рекомбинации.
Например, был открыт метод получения L-лизина с помощью Escherichia coli, в которой гены, кодирующие дигидродипико-линатсинтетазу и аспартокиназу, утратили чувствительность к ингибированию по типу отрицательной обратной связи L-лизином и L-треонином, а гены, кодирующие диаминопимелатдегидрогеназу (или тетрагидродипиколинатсукцинилазу и сукцинилдиаминопимелатдеацилазу), обладали повышенной активностью (WO 95/16042).
Хотя продукция L-аминокислот была значительно усовершенствована выведением микроорганизмов, указанных выше, или улучшением процессов производства, необходимо разрабатывать более эффективные процессы получения L-аминокислот, чтобы удовлетворить ожидаемое в будущем существенное повышение спроса на аминокислоты.
Пируваткарбоксилаза [пируват: диоксид углерода лигаза (ADP-образующая), ЕС 6.4.1.1] является ферментом, содержащим биотин. Она катализирует карбоксилирование пирувата с образованием оксалоацетата. Пируваткарбоксилаза из Escherichia coli неизвестна, но Brevibacterium (Diesterhaft M.D. and Freese E. , J. Biol. Chem. , 248, No. 17, p. 6062-6070 (1973)) и Corynebacterium (Peters-Wendish P.G. et al., Microbiology, 144, p. 915-927 (1988)) содержат пируваткарбоксилазу, и нуклиотидная последовательность генов, кодирующих этот фермент, была опубликована.
Кроме того, известны штаммы Corynebacterium glutamicum, в которых активность пируваткарбоксилазы увеличена, или штаммы Corynebacterium glutamicum, ген пируваткарбоксилазы которых инактивирован (Peters-Wendish, P.G. et al., Microbiology, 144, p. 915-927 (1988)). Однако эти микроорганизмы были созданы в качестве экспериментальных образцов для изучения ферментов, необходимых для роста на среде с глюкозой. Связь между активностью пируваткарбоксилазы и продукцией L-аминокислот была неизвестна. Также неизвестны попытки введения гена пируваткарбоксилазы в бактерии, принадлежащие роду Escherichia.
Настоящее изобретение выполнено на основе вышеупомянутых фактов, и его целью является разработка способа получения L-аминокислот, а именно L-треонина, L-глутаминовой кислоты, L-гомосерина, L-метионина, L-аргинина, L-пролина, L-изолейцина, с высокой степенью эффективности.
В результате кропотливого исследования с целью достижения указанной выше цели авторами настоящего изобретения было установлено, что введение гена, кодирующего пируваткарбоксилазу (здесь и далее упоминающегося как "ген рус"), в бактерию, принадлежащую роду Escherichia, может усилить продукцию бактерией L-аминокислот. Таким образом было совершено настоящее изобретение.
То есть предметом настоящего изобретения является бактерия, принадлежащая роду Escherichia, трансформированная с помощью гена, кодирующего пируваткарбоксилазу, и обладающая способностью продукции L-аминокислот.
В настоящем изобретении также представлена бактерия, описанная выше, в которой ген, кодирующий пируваткарбоксилазу, получен из бактерии, принадлежащей роду Bacillus.
Далее, в настоящем изобретении предложена бактерия, описанная выше, в которой ген, кодирующий пируваткарбоксилазу, введен в названную бактерию в малом числе копий.
Более того, в настоящем изобретении предложена бактерия, описанная выше, в которой L-аминокислота выбрана из группы, состоящей из L-треонина, L-глутаминовой кислоты, L-гомосерина, L-метионина, L-аргинина, L-пролина и L-изолейцина.
Также в настоящем изобретении предложен способ получения L-аминокислот, включающий в себя культивирование указанной выше бактерии в среде, производство и накопление в среде L-аминокислот и выделение L-аминокислот из среды.
Далее, в настоящем изобретении представлен способ получения L-аминокислот, описанный выше, в котором L-аминокислота выбрана из группы, состоящей из L-треонина, L-глутаминовой кислоты, L-гомосерина, L-метионина, L-аргинина, L-пролина и L-изолейцина.
Термин "продукция L-аминокислот" означает способность бактерии продуцировать и накапливать L-аминокислоту в среде в количестве, большем, чем у дикого штамма этого типа.
В соответствии с настоящим изобретением продукция L-аминокислоты, такой как L-треонин, L-глутаминовая кислота, L-гомосерин, L-метионин, L-аргинин, L-пролин или L-изолейцин, бактерией, принадлежащих к роду Escherichia, может быть улучшена. Более того, настоящее изобретение может быть использовано для выведения продуцентов L-аминокислот, принадлежащих роду Escherichia.
На фиг. 1 показана схема получения штамма-реципиента Bacillus subtilis pycA::KmR для клонирования гена русА.
На фиг.2 показана схема русА из Bacillus subtilis 168.
На фиг.3 показана схема конструирования плазмиды, содержащей ген русА.
Далее настоящее изобретение будет описано подробнее.
<1> Бактерия, принадлежащая роду Escherichia, трансформированная с помощью гена рус.
Бактерия, принадлежащая роду Escherichia, согласно настоящему изобретению является бактерией, трансформированной с помощью гена рус. Примером бактерии, принадлежащей роду Escherichia, является Escherichia coli. Хотя ген рус, используемый в настоящем изобретении, не ограничивается тем условием, что он может кодировать белок с активностью пируват-карбоксилаэы, примерам гена рус может служить ген рус, полученный из бактерии, принадлежащей к роду Bacillus, или ген рус из коринеподобных бактерий. Была опубликована нуклеотидная последовательность гена рус, полученного из Bacillus subtilis, (Genbank/EMBL/DDBJ Accession, Z97025, NID g2224758) и гена рус из Corynebacterium Glutamicum (Peters-Wendish P. G. et al., Microbiology, 144, p. 915-927 (1988)). Поэтому ген рус может быть получен с помощью ПЦР (полимеразной цепной реакции: White T. J. et al., Trends Genet., 5, 185 (1989)) с применением праймеров (затравок), синтезированных в соответствии с нуклеотидной последовательностью, и с использованием в качестве матрицы хромосомной ДНК бактерии, принадлежащей роду Bacillus, такой как Bacillus subtilis, или бактерии соrуnеform, такой как Corynebacterium Glutamicum.
Штамм Escherichia coli 44/pmMW119-pycA, в котором присутствует pMW119-pycA, содержащий ген рус Bacillus subtilis был депонирован в Российской национальной коллекции промышленных микроорганизмов (ВКПМ), ВНИИГенетика; Дорожный проезд, 1, 113545, Москва, Россия, под регистрационным номером ВКПМ В-7822.
Штамм Escherichia coli MG442/pmMW119-pycA, в котором присутствует pMW119-pycA, содержащий ген рус Bacillus subtilis, был депонирован в Российской национальной коллекции промышленных микроорганизмов (ВКПМ), ВНИИГенетика; Дорожный проезд, 1, 113545, Москва, Россия, под регистрационным номером ВКПМ В-7821.
В этой связи ген, кодирующий пируваткарбоксилазу из Bacillus subtilis (ген русА), был клонирован, как это описано ниже, в процессе выполнения настоящего изобретения.
Сначала был сконструирован штамм Bacillus subtilis, дефицитный по гену русА. Затем, с использованием этого штамма в качестве реципиента, ген русА был клонирован посредством выделения фрагмента ДНК, который способствовал восстановлению ауксотрофности штамма по аспартату или цитрату, из библиотеки геномной ДНК штамма Bacillus subtilis дикого типа. После того как Bacillus subtilis образует оксалоацетат с помощью пируваткарбоксилазы, Bacillus subtilis не может расти на минимальной среде, если у нее наблюдается дефицит активности этого фермента. Однако рост восстанавливается при добавлении L-аспартата, L-глутамата, цитрата или сукцината. Нуклеотидная последовательность гена русА и аминокислотная последовательность, кодированные геном, показаны в SEQ ID Nos:3 и 4.
Штамм, дефицитный по гену русА, может быть получен, например, как описано ниже. Частичный фрагмент ДНК гена ylaP, расположенный на хромосоме сразу после гена русА, был получен с помощью ПЦР. В качестве затравок для ПЦР могут быть использованы олигонуклеотиды, например SEQ ID Nos:1 и 2. Затем штамм Bacillus subtilis дикого типа был трансформирован с помощью ДНК-плазмиды, содержащей полученный ДНК и маркерный ген, с последующей интеграцией ДНК-плазмиды в ген ylaP в составе хромосомной ДНК методом гомологичной рекомбинации. Затем, с помощью фермента рестрикции, например, такого как PstI, узнающего рестрикционный сайт внутри гена русА, была сконструирована библиотека хромосомной ДНК полученного штамма. Рекомбинантная плазмида, содержащая маркерный ген и частичный фрагмент гена русА, может быть получена трансформацией штамма Bacillus subtilis дикого типа с помощью библиотеки и последующим отбором клонов, экспрессирующих маркерный ген. Затем штамм Bacillus subtilis дикого типа был трансформирован линеаризованной рекомбинантной плазмидой, и клоны, экспрессирующие маркерный ген, были отобраны для получения дефицитного по гену русА штамма, в котором ДНК-плазмида интегрирована в ген русА в хромосомной ДНК этого штамма.
Трансформация бактерии, принадлежащей роду Escherichia, геном рус может быть произведена путем введения гена рус, в подходящий вектор, который функционирует в бактерии, принадлежащей роду Escherichia, чтобы сконструировать рекомбинантный вектор, и введения этого рекомбинантного вектора в бактерию, принадлежащую роду Escherichia.
В качестве примера вектора может служить плазмида pBR322, pMW118, pMW119, pUC18, pUC19 или подобная, фаговый вектор, такой как λ, λ, М13mр9 или подобный, и транспозон, такой как Мu, Тn10, Тn 5 или подобный. Однако в настоящем изобретении плазмида с малым числом копий, такая как pMW118 или pMW119, предпочтительна в виду того, что бактерия, трансформируемая вектором, может быть нестабильной, если в качестве вектора для введения гена русА используется плазмида с высоким числом копий.
Введение ДНК в бактерию рода Escherichia может быть произведено, например, по методу D.M. Morrison (Methods in Enzymology, 68, h. 326 (1979)) или по методу, в котором бактериальные клетки, включающие ДНК, для увеличения их проницаемости для ДНК обрабатываются хлоридом кальция (Mandel M. and Higa A. , J. Mol. Biol., 53, p. 159 (1970)) или другим сходным способом.
Приготовление геномной ДНК, конструирование библиотеки геномной ДНК, гибридизация, ПЦР, приготовление ДНК-плазмиды, разрезание и лигирование ДНК, трансформация и тому подобное могут быть произведены любым способом, известным специалисту, имеющему навыки и опыт работы в этой области. Такие способы описаны Sambrook J., Frritsche E.F., Maniatis T. (Molecular Cloning, Cold Spring Harbor Laboratory Press, 1.21 (1989)).
Продукция L-аминокислот бактерией рода Escherichia может быть улучшена путем придания или усиления активности пируваткарбоксилазы в бактерии путем трансформации этой бактерии геном рус. Это происходит, вероятно, из-за того, что щавелевоуксусная кислота образуется не только из фосфоенолпировиноградной кислоты, но также из пировиноградной кислоты. А именно, транспорт молекулы глюкозы (или сахарозы) в бактериальную клетку, опосредованный системой фосфоенолпировиноградной кислоты, протекает с расходованием одной молекулы фосфоенолпировиноградной кислоты и образованием одной молекулы пировиноградной кислоты. И хотя образовавшаяся пировиноградная кислота не участвует напрямую в синтезе L-аминокислот в бактериальной клетке, продукция L-аминокислот может быть повышена вследствие образования щавелевоуксусной кислоты из пировиноградной кислоты.
В качестве бактерии, принадлежащей роду Escherichia, в которую ген рус должен быть введен, для производства L-аминокислот использован штамм, уже обладающий этим свойствам. Или же способность к продукции L-аминокислот бактерией рода Escherichia может быть придана в результате ее трансформации геном рус. Кстати ген рус из Escherichia coli неизвестен, однако, если этот ген будет найден в дальнейшем, он также может быть использован.
Примеры бактерий, принадлежащих роду Escherichia и производящих L-аминокислоты, описаны ниже.
(1) Бактерии, продуцирующие L-треонин.
Примером бактерий, продуцирукщих L-треонин, принадлежащих роду Escherichia, может служить штамм MG442 (был депонирован в Российской национальной коллекции промышленных микроорганизмов (ВКПМ), ВНИИГенетика; Дорожный проезд, 1, 113545, Москва, Россия, под регистрационным номером ВКПМ B-1628 (VKPM B-1628) (USP 4, 278, 765; Гусятинер М.М. и др., Генетика, 14, с. 947-956, (1978)). Штамм MG442 был получен как мутант, устойчивый к 2-амино-3-гидроксивалериановой кислоте - аналогу L-треонина. Этот штамм также был неполным (leaky type) ауксотрофом по изолейцину. Он продуцировал L-треонин и L-глутамат в качестве побочных продуктов.
В качестве альтернативы штамм MG442, трансформированный pVC40 (USP 5, 175, 107), предпочтительно может быть использован в качестве штамма, продуцирующего L-треонин.
(2) Бактерии, продуцирующие L-гомосерин.
Примером бактерий, продуцирующих L-гомосерин, принадлежащих роду Escherichia, может служить штамм 44 (thrB). Этот штамм был получен из известного ранее штамма С600 (thrB, leuB) (Appleyard R.K., Genetics, 39, р. 440-452) как Leu+-peвертант. Штамм 44, трансформированный с помощью плазмиды, содержащей ген thrA, кодирукший аспартаткиназу - гомосериндегидрогеназу I, может быть предпочтительно использован.
Штамм Escherichia coli 44 был депонирован в Российской национальной коллекции промышленных микроорганизмов (ВКПМ), ВНИИГенетика; Дорожный проезд, 1, 113545, Москва, Россия, под регистрационным номером ВКПМ В-2175.
(3) Бактерии, продуцирующие L-глутаминовую кислоту.
Примером бактерий, продуцирующих L-глутаминовую кислоту, принадлежащих роду Escherichia, может служить штамм, устойчивый к L-валину, например штаммы Escherichia coli B11, K-12 (АТСС10798), В (АТСС11303) и W (ATCC9637).
(4) Бактерии, продуцирующие L-изолейцин.
Примером бактерий, продуцирующих L-изолейцин, принадлежащих роду Escherichia, может служить штамм TDH6/pVIC40, pMWD5 (Hashiguchi К. et al., Biosci. Biotechnol. Biochem., 63(4), p. 672-679 (1999) или AJ12919, описанный в ЕРО 685555Al.
<2> Способ получения L-аминокислот.
Продуцирование L-аминокислот может происходить с высокой эффективностью при культивировании бактерии, принадлежащей роду Escherichia, трансформированной геном рус, как это описано выше, и обладающей способностью к продукции L-аминокислот в соответствующей питательной среде, при продуцировании и накоплении L-аминокислот в питательной среде и сборе L-аминокислот из питательной среды.
Примером L-аминокислот может служить L-треонин, L-глутаминовая кислота, L-гомосерин, L-метионин, L-аргинин, L-пролин и L-изолейцин. L-Аминокислоты могут производиться с помощью способа согласно настоящему изобретению как по отдельности, так и в смеси из двух и более аминокислот разного вида.
В способе согласно изобретению выращивание бактерии, принадлежащей роду Escherichia, накопление, выделение L-аминокислот из жидкой питательной среды и их очистка могут быть произведены способом, сходным с общепринятыми способами получения аминокислот методом ферментации с использованием бактерий. Питательная среда, используемая для культивирования, может быть как искусственной, так и натуральной, при условии, что она содержит источники углерода и азота, минеральные вещества и, если необходимо, питательные вещества, требующиеся для роста бактерий в соответствующем количестве. В качестве источников углерода могут быть использованы различные углеводы, такие как глюкоза и сахароза, а также различные органические кислоты. В зависимости от усваивающей способности используемой бактерии может быть использован спирт, такой как этанол, и глицерин. В качестве источника азота могут быть использованы аммиак, различные соли аммония, такие как сульфат аммония, другие источники азота, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов, ферментализат микробной биомассы. В качестве минеральных веществ могут быть использованы фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, карбонат кальция.
Выращивание осуществляют предпочтительно в аэробных условиях, таких как качалочная культура, а также при аэрации и перемешивании. Температура культуры обычно устанавливается в интервале от 20 до 40oС, предпочтительно в интервале от 30 до 38oС. рН среды обычно устанавливается в интервале от 5 до 9, предпочтительно в интервале от 6,5 до 7,2. рН культуральной среды может быть отрегулирован с помощью аммония, карбоната кальция, различных кислот, оснований и буферов. Обычно культивирование в течение 1-3 суток приводит к накоплению целевых L-аминокислот в питательной среде.
Сбор и очистка L-аминокислоты после культивирования могут быть произведены путем удаления из среды нерастворимых компонентов, таких как клетки, путем центрифугирования или фильтрации через мембрану с последующей очисткой на ионообменнике, концентрированием и кристаллизацией.
Наилучший способ осуществления настоящего изобретения
Настоящее изобретение будет более подробно проиллюстрировано ниже со ссылкой на примеры.
Пример 1. Клонирование гена русА штамма Bacillus subtilis 168.
<1> Конструирование штамма реципиента В. subtilis pycA::KmR для клонирования.
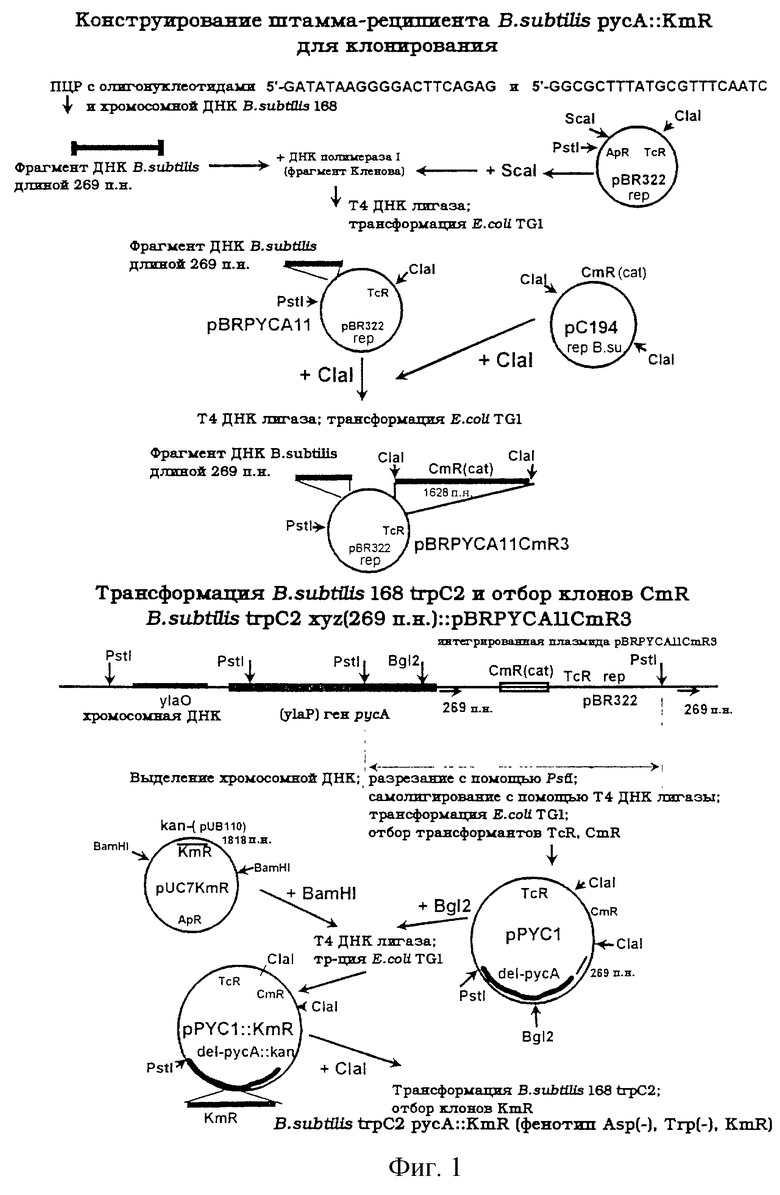
Штамм реципиент В. subtilis был сконструирован при помощи инсерционного мутагенеза гена русА. Фрагмент гена русА был клонирован в две стадии (см. фиг.1 и 2).
Одна из рамок считывания (ylaP) в базе данных проекта определения последовательности генома Bacillus subtilis была охарактеризована как ген русА, кодирующий пируваткарбоксилазу. Область, расположенная сразу после терминатора гена русА, была получена с помощью ПЦР с использованием следующей пары олигонуклеотидов: 5'-GATATAAGGGGACTTCAGAG и
5'-GGCGCXXXATGCGXXXCAATC.
Концы полученного фрагмента ДНК длиной 269 п.н. были затуплены с помощью фрагмента Кленова ДНК-полимеразы I, и фрагмент клонирован в сайт ScaI плазмиды pBR322 штамма Е. coli. Полученная плазмида pBRPYCA11 была подвержена перевариванию под действием Clal и лигирована с фрагментом ДНК длиной 1628 п. н. , содержащим ген cat, полученным после переваривания плазмиды рС194 с помощью ClaI. Затем штамм Bacillus subtilis 168 trpC2 был трансформирован полученной плазмидой pBRYCA11CmR3, и клоны CmR были отобраны. Плазмида pBRYCA11CmR3 не может самопроизвольно воспроизводиться при клеточном делении бактерии, принадлежащей роду Bacillus, но может встраиваться в хромосому рядом с геном русА ввиду гомологии с фрагментом ДНК длиной 269 п.н. Штамм был назван В. subtilis trpC2 xyz::pBRPYCACmR3. Интегрант содержал репликон pBR322, обусловливающий устойчивость к тетрациклину ген tet, полученный из pBR322, и маркер СmR.
Хромосомная ДНК штамма В. subtilis trpC2 xyz::pBRPYCACmR3 была выделена и использована для клонирования дистальной части гена русА. Обработанная с помощью PstI хромосомная ДНК была самолигирована и трансформирована в штамм Е. coli TG1. Затем были отобраны клоны с рекомбинантными плазмидами, устойчивые к тетрациклину и хлорамфениколу. Из одного клона была выделена плазмида и названа pPYCl. Плазмида pPYCl содержала единственный сайт, подверженный рестриктазе BglII, расположенный внутри гена русА. Этот сайт был использован для включения фрагмента ДНК длиной 1818 п.н., содержащего маркер устойчивости В. subtilis к канамицину. Для получения плазмиды pPYCl::KmR плазмида pPYCl была обработана рестриктазой BglII и лигирована с pUC7KmR, предварительно обработанной рестриктазой BamHI.
Инактивация гена русА в штамме дикого типа В. subtilis 168 trpC2 была произведена в одну стадию при трансформации этого штамма линеаризованной плазмидой pBYCl: : KmR с последующим отбором на бульоне Луриа (Luria) с добавлением канамицина. С помощью гомологичной рекомбинации по типу двойного кроссинговера KmR был введен в ген русА хромосомной ДНК.
Сконструированный штамм В. subtilis pycA::KmR trpC2 не был способен к росту на минимальной среде Спицайзена (Spizizen) (Anagnotopoulos С. and Spizizen J., J.Bacteriol., 81, p. 741-746 (1961)), содержащей триптофан, до тех пор пока не были добавлены цитрат или аспартат.
Состав минимальной среды Спицайзена, г/л:
К2НРO4•3Н2O - 18,3
КН2РO4 - 6,0
(NH4)2SO4 - 2,0
Цитрат натрия - 1,2
MgSO4•7H2O - 0,2
Глюкоза - 10,0
рН - 7,0
Штамм В. subtilis trpC2 русА::KmR rесЕ::TcR был сконструирован путем переноса инактивированного аллеля гена rесЕ штамма Bacillus subtilis recE:: TcR с помощью фага Е40 (фиг.2). Было необходимо инактивировать рекомбинацию в штамме-реципиенте, предназначенном для клонирования гена русА В. subtilis. Для клонирования гена русА был использован полученный штамм В. subtilis trpC2 русА::КmR rесЕ::TcR.
<2> Клонирование природного гена русА В. subtilis методом комплементации.
Ген русА был клонирован из штамма В. subtilis 168 trpC2. Хромосомная ДНК, выделенная из штамма 168 trpC2, была частично переварена EcoRI и лигирована с предварительно обработанной с помощью EcoRI pCB20 (EmR) (любезно предоставленной д-ром Ю.Йомантасом, см. "Genetic transformation and expression" L.O. Butler et al eds., Intercert Ltd., PO Box 402, Wimborne, Dorset, BH229T2, UK,1989, p. 269-281 (1989)). Затем штамм trpC2 русА::KmR recE::TcR был трансформирован лигирующей смесью (фиг.2). Клоны Asp+, ЕmR, КmR были отобраны на минимальной среде Спицайзена без добавления цитрата или аспартата в присутствии триптофана, эритромицина и канамицина. Плазмида pPYCRS, присутствующая в одном из отобранных клонов, дополненных мутацией в гене русА, имела предсказанную структуру, и подтвердила тот же фенотип Asp+, EmR после вторичной трансформации.
<3> Клонирование фрагмента ДНК, содержащего ген русА штамма B. subtilis 168 trpC2, в плазмиды pUC18 и pUC19 с высоким числом копий.
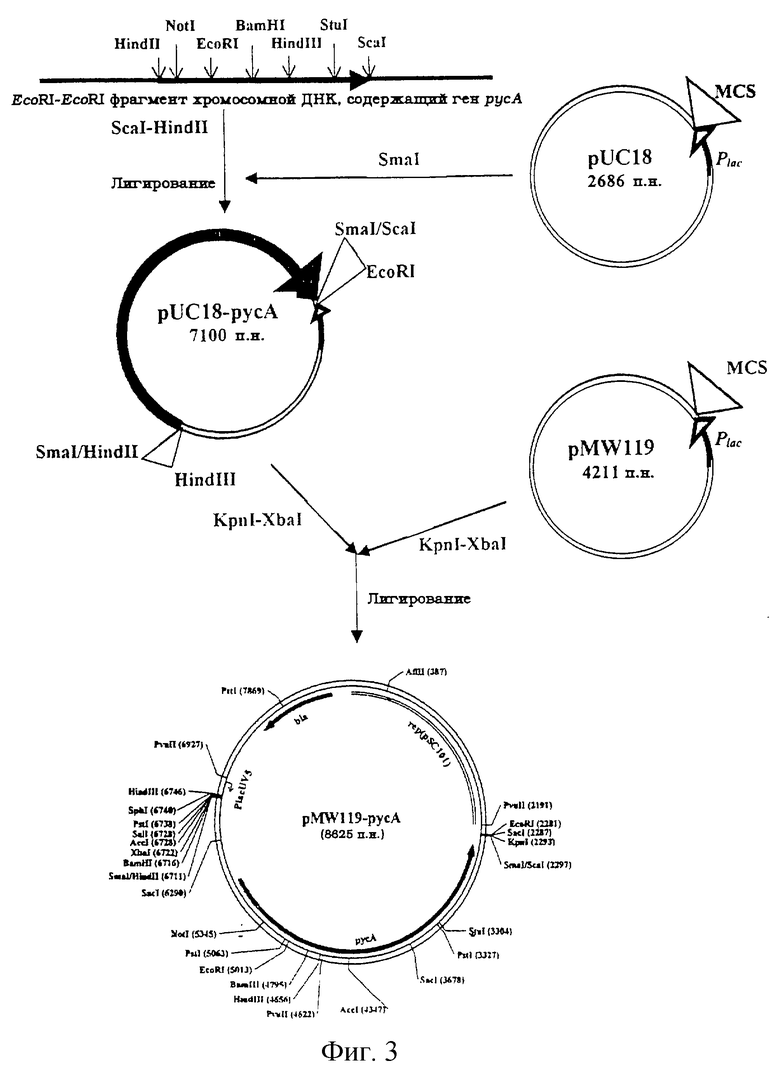
Плазмида pPYCR3 была переварена рестриктазами HindII и ScaI, и полученный HindII-ScaI фрагмент длиной 4414 п.н., содержащий ген русА с 5'-концевой нетранслируемой последовательностью, включающей в себя промотор и сайт связывания рибосомы, был клонирован в плазмиду pUC18-pycA, переваренную SmaI. Таким образом была получена плазмида pUC18-pycA (фиг.3, сверху). Реакция лигирования была проведена следующим подробно описанным способом. 90 нг плазмиды pUC18, переваренной SmaI, и 300 нг очищенного методом электрофореза в агарозном геле HindII-Scal фрагмента длиной 4414 п.н., содержащего ген русА, были лигированы с помощью Т4 ДНК-лигазы (2 единицы активности) (Pharmacia, Sweden) в 45 мкл реакционной смеси, содержащей 1-кратный буфер One-Phor-All (Pharmacia, Sweden) и 1мМ АТФ.
Штамм Е. coli TG1 был трансформирован плазмидой pUC18-русА. Полученные после трансформации клоны анализировали с помощью эндонуклеаз Kpnl, Xbal, BamHI, HindIII, EcoRI. Все клоны содержали ген русА в обратной ориентации по отношению к промотору lac плазмиды pUC18. Однако гены русА, клонированные в pUC18, могут быть экспрессированы под контролем внутреннего промотора этого гена.
Далее, для того чтобы переставить ген русА под контроль промотора lac, KpnI-XbaI фрагмент pUC18-pycA был клонирован в pUC19, расщепленную с помощью KpnI и XbaI. Однако полученные клоны не были стабильными и отторгали плазмиду после небольшого (6-8 часов) промежутка времени в процессе культивирования.
<4> Клонирование фрагмента ДНК, несущего ген русА в плазмиду pMW119 с низким числом копий.
KpnI-XbaI-фрагмент pUC18-pycA был лигирован в плазмиду pMW119 с низким числом копий, предварительно расщепленную с помощью KpnI и XbaI (фиг.3, снизу). Реакция лигирования проводилась в 50 мкл реакционной смеси, содержащей 60 нг pMW119, переваренной с помощью KpnI и XbaI, 120 нг KpnI-XbaI-фрагмента pUCl8-pycA, очищенного методом электрофореза в агарозном геле, 1-кратный буфер One-Phor-All (Pharmacia, Sweden), 1 мМ АТФ и Т4-ДНК-лигазу (1 единица активности) (Pharmacia, Sweden).
Штамм Е. coli TG1 был трансформирован с помощью лигирующей смеси. Полученные трансформанты анализировали с помощью SacI, KpnI, XbaI, BamHI, HindIII, EcoRI. Все клоны содержали ген русА, находящийся под контролем промотора lac плазмиды pMW119 и его собственного промотора. Полученный таким образом клон был назван TG1 (pMW119-pycA). Из этого клона была получена плазмида pMW119-pycA.
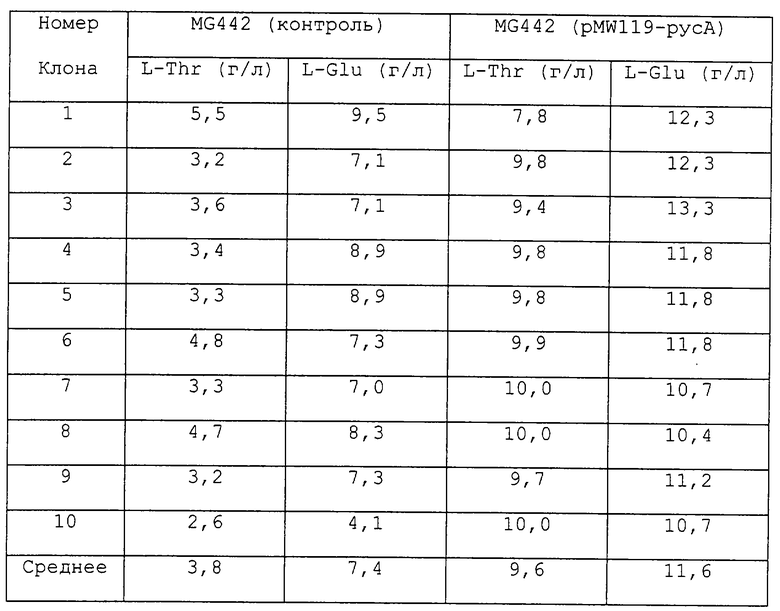
Пример 2. Продукция L-треонина и L-глутаминовой кислоты.
Штамм Е. coli MG442 был трансформирован с помощью плазмиды pMW119-pycA, и 10 клонов Аmpr были отобраны. Способность этих клонов продуцировать L-треонин и L-глутаминовую кислоту была проверена в следующей ферментационной среде.
Состав ферментационной среды, г/л:
Глюкоза - 60,0
(NH4)2SO4 - 1,5
КН2РO4 - 1,5
MgSO4•7H2O - 1,0
L-Изолейцин, мг/л - 10,0
Тиамин, мг/л - 0,1
СаСО3 (стерилизованный отдельно) - 20,0
Ферментация проводилась в пробирках (20 x 300 мм) при 32oС в течение 72 часов на роторной качалке. Одна петля каждого из 10 клонов была высеяна в пробирку, содержащую 2,0 мл среды. В качестве контроля были использованы 10 отдельных колоний штамма без плазмиды. Концентрации L-треонина и L-глутаминовой кислоты после ферментации были измерены с помощью тонкослойной хроматографии. Результаты измерений приведены в таблице.
Пример 3. Продукция L-гомосерина.
L-Гомосеринпродуцирующий штамм 44 E. coli был трансформирован плазмидой pMW119-pycA, и было получено пять резистентных к ампициллину клонов. Анализ продукции L-гомосерина проводили, как указано в примере 1, но ферментационную среду при этом дополняли 0,5 г/л L-треонина. В качестве контроля использовали пять колоний штамма 44, не содержащих плазмиду. После ферментации средние концентрации L-гомосерина составляли 4,5 г/л (штамм 44) и 5,7 г/л (штамм 44 с плазмидой pMW119-pycA).










Изобретение относится к биотехнологии. L-Аминокислоты, такие как L-треонин, L-гомосерин, L-метионин, L-аргинин, L-пролин, L-изолейцин или L-глутаминовая кислота, получают культивированием бактерии Escherichia coli, трансформированной геном, кодирующим пируваткарбоксилазу. Причем E. coli обладает способностью к продукции L-аминокислот. После накопления L-аминокислот их извлекают из питательной среды. Штамм бактерии E. coli MG 442 или 44 трансформируют плазмидой, содержащей ген, кодирующий пируваткарбоксилазу. В первом случае штамм продуцирует L-треонин и L-глутаминовую кислоту, в последнем - L-гомосерин. Изобретение позволяет получать L-аминокислоты с высокой степенью эффективности. 3 с. и 2 з.п.ф-лы, 3 ил., 1 табл.
| J | |||
| Biol | |||
| Chem | |||
| Приспособление для склейки фанер в стыках | 1924 |
|
SU1973A1 |
| Деревянная повозка с кузовом, устанавливаемым на упругих дрожинах | 1920 |
|
SU248A1 |
| УСТРОЙСТВО ДЛЯ РЕВЕРСИРОВАНИЯ НАПРАВЛЕНИЯ ВРАЩЕНИЯ ТРЕХФАЗНЫХ АСИНХРОННЫХ ДВИГАТЕЛЕЙ | 1926 |
|
SU6062A1 |
| DE 19831609 A1, 15.04.1999. | |||
