Изобретение относится к биотехнологии, в частности касается способа получения аминокислот, а именно L-гомосерина, L-треонина, L-валина, или L-лейцина с помощью бактерий, принадлежащих к роду Escherichia.
В качестве ближайшего аналога может быть рассмотрен фрагмент ДНК, кодирующий ген rhtA, связанный с устойчивостью бактерий к L-гомосерину и L-треонину на минимальной среде, а также полученные на основе использования мутации этого гена rhtA23 (ранее обозначавшейся как thrR) штаммы Escherichia coli, продуцирующие L-треонин (Авторское свидетельство СССР N 974817; Астаурова и др. Прикладная биохимия и микробиология, т. 21, стр. 611 - 616, 1985), или L-гомосерин и L-глутаминовую кислоту (Астаурова и др. Прикладная биохимия и микробиология, т. 27, стр. 556 - 561, 1991).
Ген rhtA дикого типа обеспечивает устойчивость к L-гомосерину и L-треонину, если он клонирован в мультикопийной плазмиде, и повышение его экспрессии повышает продукцию аминокислот бактериями, принадлежащими к роду Escherihia, и способными продуцировать L-треонин, L-лизин, L-валин, или L-аргинин. Было обнаружено, что мутация rhtA23 расположена на 18 минуте карты хромосомы E.coli, а ген rhtA идентифицирован как orf1, открытая рамка считывания, локализующаяся между генами pexB и ompX. Генетическая структура, экспрессирующая белок, кодируемый orf1, была обозначена как ген rhtA (rht - resistance to homoserine and threonine - устойчивость к гомосерину и треонину). Ген rhtA включает 5'-некодирующую область, в том числе SD-последовательность, саму открытую рамку считывания, orf1, и терминатор. Мутация rhtA23 изменяет нуклеотид, непосредственно предшествующий инициаторному кодону ATG, и повышает экспрессию гена rhtA (ABSTRACTS of 17th International Congress of Biochemistry and Molecular Biology in conjugation with 1997 Annual Meeting of the American Society for Biochemistry and Molecular Biology, San Francicco, California August 24-29, 1997, N 457).
В процессе клонирования гена rhtA обнаружено, что существуют по крайней мере два участка на хромосоме Е.coli, которые в мультикопийном состоянии сообщают клеткам устойчивость к L-гомосерину и L-треонину. Один из них - это ген rhtA. Как оказалось, другой ген, rhtB, сообщает клеткам Е.coli устойчивость только к L-гомосерину (Заявка на выдачу патента в России. N 98118425).
Задачей настоящего изобретения является повышение уровня накопления аминокислот клетками бактерий рода Escherichia, продуцирующими L-гомосерин, L-треонин, L-валин или L-лейцин.
Поставленная задача решается получением фрагмента ДНК rhtC, кодирующего синтез белка RhtC, придающего повышенную устойчивость к L-треонину бактериям Е. coli, и конструированием на его основе штаммов, позволяющих разработать способ получения аминокислот с повышенным выходом целевой аминокислоты.
Предметом настоящего изобретения являются:
1. Бактерии, принадлежащие к роду Escherichia, у которых устойчивость к L-треонину повышена вследствие увеличения в клетках бактерий активности белка, характеризующегося одним из двух свойств (А) или (Б):
А - белок, который состоит из аминокислотной последовательности N 2 (фиг.2); или
Б - белок, который состоит из аминокислотной последовательности, включающей также делеции, замены, вставки или добавки из одной или нескольких аминокислот к последовательности N 2 и который имеет активность, обеспечивающую бактериям, содержащим этот белок, повышенную устойчивость к L-треонину;
2. Бактерии по п. 1, у которых устойчивость к L-гомосерину повышена путем увеличения в клетках бактерий активности белка, характеризующегося одним из двух свойств (С) или (Д):
(С) - белок, который состоит из аминокислотной последовательности N 4 (фиг. 4); или
(Д) - белок, который состоит из аминокислотной последовательности, включающей также делеции, замены, вставки или добавки из одной или нескольких аминокислот к последовательности N 4 и который имеет активность, обеспечивающую бактериям, содержащим этот белок, устойчивость к L-гомосерину;
(3). Бактерии по п. (1) или (2), где активность белка, определяемого по свойствам (А) или (Б) повышена в результате трансформации бактерий с помощью ДНК, кодирующей белок, имеющий свойства (А) или (Б);
(4). Бактерии по п. (2), где активность белка, имеющего свойства (С) или (Д), повышена путем трансформации бактерий с помощью ДНК, кодирующей белок, имеющий свойства (С) или (Д);
(5). Способ получения аминокислот, включающий этап культивирования бактерий, соответствующих любому из пунктов с 1 по 4 и обладающих способностью к продукции аминокислот, в культуральной среде, обеспечивающей продукцию и накопление соответствующей аминокислоты в этой среде, и выделения накопившейся аминокислоты из этой среды.
(6). Способ по п. 5, где аминокислота является одной из аминокислот в группе, состоящей из L-гомосерина, L-треонина, и аминокислот с разветвленной цепью;
(7). Способ по п. (6), где аминокислотами с разветвленной цепью являются L-валин или L-лейцин.
ДНК, кодирующая белок, определяемый выше по п. (А) или (Б), может рассматриваться как ген rhtC, а белок, кодируемый геном rhtC, может рассматриваться как "белок RhtC". ДНК, кодирующая белок, определяемый выше по п. (С) или (Д), может рассматриваться как ген rhtB, а белок, кодируемый геном rhtB, может рассматриваться как "белок RhtB". Активность белка RhtC, которая участвует в придании бактериям устойчивости к L-треонину (т.е. активность которая делает бактерии, имеющие белок RhtC, устойчивыми к L-треонину), может рассматриваться как "Rt активность" (от слов Resistance to threonine - устойчивость к треонину), а активность белка RhtB, которая участвует в придании бактериям устойчивости к L-гомосерину (т.е. активность, которая делает бактерии, имеющие белок RhtB, устойчивыми к L-гомосерину), может рассматриваться как "Rh активность" (от слов Resistance to homoserine - устойчивость к гомосерину).
Структурные гены, кодирующие белок RhtC и белок RhtB, обозначены соответственно как "структурный ген rhtC" или "структурный ген rhtB". Термин "повышение Rt активности или Rh активности" означает придание клеткам устойчивости к L-треонину или L-гомосерину, или повышение этой устойчивости либо путем увеличения числа молекул белка RhtC или белка RhtB, или увеличением специфической активности этих белков, или нарушением негативной регуляции экспрессии или активности этих белков и т.п. Термин "ДНК, кодирующая белок", обозначает двунитевую ДНК, одна из нитей которой кодирует белок. Устойчивость к L-треонину означает свойство бактерий расти на минимальной среде, содержащей L-треонин в концентрации, при которой штамм дикого типа, несущий природный аллель гена rhtC, не может расти, обычно это > 30 мг/мл. Устойчивость к L-гомосерину означает свойство бактерий расти на минимальной среде, содержащей L-гомосерин в концентрации, при которой штамм дикого типа, несущий природный аллель гена rhtB, не может расти, обычно это > 5 мг/мл. Способность продуцировать аминокислоту означает свойство бактерий синтезировать и накапливать аминокислоту в культуральной среде в количестве, большем чем штаммы дикого типа.
В соответствии с настоящим изобретением устойчивость к высоким концентрациям L-треонина, или L-треонина и L-гомосерина может быть придана бактериям, принадлежащим к роду Escherichia, способным продуцировать аминокислоты, в частности L-гомосерин, L-треонин, L-валин, или L-лейцин.
Настоящее изобретение представляет собой фрагмент ДНК (ген rhtC), кодирующий белок с Rt-активностью и имеющий аминокислотную последовательность, представленную на фиг. 2 ("Аминокислотная последовательность белка RhtC"). В частности, этот фрагмент может быть представлен нуклеотидной последовательностью, включающей нуклеотиды от 187 по 804 в нуклеотидной последовательности N 1 (см. формулу изобретения).
Второй фрагмент ДНК, используемый в настоящем изобретении, (ген rhtB), кодирующий белок с Rh-активностью и имеющий аминокислотную последовательность, представленную на фиг. 4 ("Аминокислотная последовательность белка RhtB"). В частности, этот фрагмент может быть представлен нуклеотидной последовательностью, включающей нуклеотиды от 557 по 1171 в нуклеотидной последовательности N 3 (фиг. 3).
Ген rhtB, имеющий нуклеотидную последовательность, указанную в Последовательности N 3, соответствует части последовательности, которая комплементарна последовательности М87049, имеющейся в базе данных GenBank, и включает известную предполагаемую открытую рамку считывания f138 (нуклеотиды с 61959 по 61543 в последовательности М-87049), функция которой неизвестна, и которая расположена на хромосоме E.coli в районе 86,8 минуты карты, а также 201 нуклеотид проксимальнее f138. Необходимо отметить, что открытая рамка считывания f138 только со 160 5'-фланкирующими нуклеотидами не обеспечивает устойчивости к L-гомосерину. Оказалось также, что указанная последовательность выше f138 не содержит стоп-кодона в рамке f138. Кроме того, одному из ATG кодонов в этой последовательности предшествует участок связывания с рибосомами (SD-последовательность, нуклеотиды с 62171 по 62166 в М87049). Эта удлиненная открытая рамка считывания (нуклеотиды 62160-61546) и представляет собой структурный ген rhtB.
Ген rhtB получают либо путем инфицирования лизогенного по Mucts штамма E. coli лизатом фазмиды миниMu d50005, как это описано Гройсманом с соавт. (Groisman et al. J. Bacteriol., 158, 357-364, 1986) с последующим выделением плазмидной ДНК из колоний, выросших на минимальной среде, содержащей 40 мкг/мл канамицина и 10 мг/мл L-гомосерина, либо из хромосомы E.coli путем гибридизации колоний, или с помощью полимеразной цепной реакции (ПЦР) (White et al. , Trends Genet., 5, 185, 1989), используя олигонуклеотид(ы), имеющие последовательность, соответствующую участку в районе 86 минуты хромосомы E. coli.
Другой подход предполагает синтез олигонуклеотида на основе последовательности N 3. Используя олигонуклеотиды, имеющие последовательность, соответствующую участку ДНК, который расположен проксимальнее нуклеотида N 557, и участку ДНК, который расположен дистальнее нуклеотида N 1171 в последовательности N 3 в качестве праймеров для ПЦР, можно амплифицировать всю кодирующую область.
Синтез олигонуклеотидов осуществляют обычным методом, например с помощью фосфоамидитного метода (см. Tetrahedron Letters, 22, 1859, 1981), с использованием коммерческого ДНК-синтезатора (например, ДНК синтезатора модели 380B, производимого Applied Biosystems). ПЦР осуществляют с использованием коммерчески доступных аппаратов для ПЦР (например, ДНК-термоциклера, модель PG 2000, производимого компанией Takara Shuzo Co., Ltd) с применением Taq ДНК полимеразы в соответствии с методикой, описанной поставщиком фермента.
Ген rhtC соответствует уточненной, как это описано ниже, последовательности о128 (нуклеотиды N 60860 - 61480 в последовательности М87049, имеющейся в базе данных GenBank), которая расположена на участке хромосомы, прилежащем к гену rhtB. Его получают одновременно с геном rhtB, как это показано в примере 1, путем инфицирования лизогенного по Mucts штамма E.coli лизатом фазмиды миниMu d50005, как это описано выше, с последующим выделением плазмидной ДНК из колоний, выросших на минимальной среде, содержащей 40 мкг/мл канамицина и 50 мг/мл треонина. Другой подход предполагает синтез олигонуклеотидов на основе последовательности N 1 описанным выше методом и использования их для гибридизации или в ПЦР. Используя олигонуклеотиды, имеющие последовательность, соответствующую участку ДНК, который расположен проксимальнее нуклеотида N 187, и участку ДНК, который расположен дистальнее нуклеотида N 804 в последовательности N 1 в качестве праймеров для ПЦР, можно амплифицировать всю кодирующую область.
ДНК, кодирующая белок RhtB по настоящему изобретению, может кодировать белок RhtB, включая делеции, замены, инсерции или добавки одной или нескольких аминокислот в одной или нескольких позициях, не нарушающих при этом Rh-активность белка RhtB. Точно так же, ДНК, кодирующая белок RhtC по настоящему изобретению, может кодировать белок RhtC, включая делеции, замены, инсерции или добавки одной или нескольких аминокислот в одной или нескольких позициях, не нарушающих при этом Rt-активность белка RhtC.
ДНК, кодирующая по существу тот же белок, что и RhtB, или тот же белок, что и RhtC, описанные выше, может быть получена, например, путем модификации нуклеотидной последовательности, в частности при помощи сайт-направленного мутагенеза, так что один или более аминокислотный остаток будет делетирован, заменен, вставлен или добавлен. ДНК, модифицированная описанным выше способом, может быть получена известными методами с помощью мутационных воздействий. Мутационная обработка включает методы обработки ДНК, кодирующей белок RhtB или белок RhtC, in vitro, например, при помощи гидроксиламина, или методы обработки микроорганизма, в частности бактерий, принадлежащих к роду Escherichia и несущих ДНК, кодирующую белок RhtB или белок RhtC, УФ-облучением или мутагенными агентами, такими как N-метил-N'-нитро-N-нитрозогуанидин (НГ) или азотистая кислота, которые обычно используются для индукции мутаций.
ДНК, кодирующую указанные варианты белка RhtB или RhtC, отбирают путем экспрессии плазмидной ДНК, несущей гены rhtC или rhtB и подвергнутой in vitro мутагенному воздействию, как описано выше, в соответствующих клетках с последующим определением их устойчивости к L-треонину или L-гомосерину и отбором ДНК, которая обеспечивает эту устойчивость. Изобретение относится также к вариантам белка RhtC, которые встречаются в разных видах, штаммах и вариантах бактерий рода Escherichia и обусловлены природным разнообразием. ДНК, кодирующие эти варианты, и гибридизуются в жестких условиях с ДНК, имеющей нуклеотидную последовательность от нуклеотида 187 по 804 нуклеотид в последовательности N 1.
Аналогичным образом, изобретение относится также к вариантам белка RhtB, которые встречаются в разных видах, штаммах и вариантах бактерий рода Escherichia и обусловлены природным разнообразием. ДНК, кодирующие эти варианты, и гибридизуются в жестких условиях с ДНК, имеющей нуклеотидную последовательность от нуклеотида 557 по 1171 нуклеотид в последовательности N 3. Термин "жесткие условия" означает здесь условия, при которых так называемая специфическая гибридизация происходит, а неспецифическая не происходит. Трудно четко выразить эти условия с помощью каких-то цифровых значений, однако, например, жесткие условия включают условия, при которых ДНК, имеющие высокую гомологию, например не менее 70% гомологии по отношению друг к другу, - гибридизуются, а ДНК, имеющие гомологию ниже указанной, - нет.
В настоящем изобретении рассматривают бактерии, принадлежащие к роду Escherichia, представленные здесь E.coli, у которых Rt активность повышена. Кроме того, настоящее изобретение включает бактерии Е.coli, у которых также повышена Rh-активность.
Повышение Rt-активности происходит, например, за счет амплификации числа копий структурного гена rhtC в клетках при трансформации их рекомбинантной ДНК, в которую включен фрагмент, содержащий структурный ген rhtC, кодирующий белок RhtC, лигированный с промоторной последовательностью, которая эффективно функционирует в бактериях рода Escherichia. Rt-активность может быть также повышена в результате замены промоторной последовательности гена rhtC на хромосоме промоторной последовательностью, которая более эффективно функционирует в бактериях рода Escherichia.
Повышение Rh-активности происходит, например, за счет амплификации числа копий структурного гена rhtB в клетках при трансформации их рекомбинантной ДНК, в которую включен фрагмент, содержащий структурный ген rhtB, кодирующий белок RhtB, лигированный с промоторной последовательностью, которая эффективно функционирует в бактериях рода Echerichia. Rh-активность может быть также повышена в результате замены промоторной последовательности гена rhtB на хромосоме промоторной последовательностью, которая более эффективно функционирует в бактериях рода Escherichia.
Амплификация числа копий структурного гена rhtC, или структурного гена rhtB в клетках может быть осуществлена также путем введения мультикопийного вектора, который несет структурный ген rhtC или структурный ген rhtB, в клетки бактерий, принадлежащих к роду Escherichia. В частности, число копий может быть увеличено путем введения плазмиды, фага или транспозона (Berg, D. E. and Berg, С.М., Bio/Technol., 1, 417, 1983), содержащих структурный ген rhtC или структурный ген rhtB, в клетки бактерий, принадлежащих к роду Escherichia. Мультикопийные вектора могут быть представлены плазмидными векторами, такими как pBR322, pMW118, pUC19 или подобными, или фаговыми векторами, такими как λ 1059, λ BF 101, m13MP9 или подобными. Транспозоны могут быть представлены фагом Mu, транспозонами Tn10, Tn5 или подобными. Введение ДНК в бактерии, принадлежащие к роду Escherichia, может быть осуществлено, например, с помощью метода Моррисона (Methods in Enzymology, 68, 326, 1979) или метода, в котором реципиентные клетки бактерий подвергают воздействию хлористого кальция для увеличения их проницаемости по отношению к ДНК (Mandel and Higa, J. Mol. Biol., 53, 159, 1970) или другими подобными методами.
Обнаруженную связь между повышением Rt-активности, а также одновременным повышением Rt-активности и Rh-активности и способностью бактерий рода Escherichia увеличивать продукцию L-аминокислот изобретатели используют для получения штаммов, обладающих повышенной продуктивностью аминокислот. При этом возможны два варианта:
1. Признак повышенной Rt-активности, или одновременно повышенной Rt-активности и Rh активности, вводят в штаммы, уже способные продуцировать желаемые аминокислоты.
2. Способность к продукции аминокислот придается штаммам, у которых повышена Rt-активность или одновременно повышены Rt-активность и Rh-активность.
Сконструированные на основе амплификации фрагмента ДНК rhtC штамм Е.coli MG442/pRhtC - продуцент гомосерина, штамм Е. coli MG442/pVIC40, pRhtC - продуцент треонина и штаммы Е.coli NZ10/pRhtBC и Е.coli NZ10/pRhtB, pRhtC - продуценты гомосерина, валина и лейцина способны к повышенной продукции указанных аминокислот по сравнению со штаммами, не содержащими амплифицированного фрагмента ДНК rhtC.
Новые штаммы депонированы во Всероссийской коллекции промышленных микроорганизмов. Штамм Е. coli MG442/pRhtC депонирован под номером ВКПМ В-7700; штамм Е.coli MG442/pVIC40, pRhtC депонирован под номером ВКПМ В-7680; штамм Е. coli NZ10/pRhtB, pRhtC депонирован под номером ВКПМ В-7681 и штамм Е.coli NZ10/pRhtBC депонирован под номером ВКПМ В-7682.
Штамм Е. coli MG442/pRhtC (ВКПМ В-7700) имеет следующие культурально-морфологические и биохимические признаки.
Морфология клеток. Грамотрицательные слабоподвижные палочки с закругленными концами, 1,5-2,0 мкм в длину.
Культуральные признаки.
Мясопептонный агар. Через 24 часа роста при 37oC образует круглые беловатые полупрозрачные колонии диаметром 1,5-3,0 мм; поверхность колоний гладкая, края ровные или слегка волнистые, центр колоний приподнят, структура однородная, консистенция пастообразная, легко эмульгируется.
Агар Лурия. Через 24 ч роста при 37oC образует беловатые полупрозрачные колонии диаметром 1,5-2,5 мм; поверхность колоний гладкая, края ровные, структура однородная, консистенция пастообразная, легко эмульгируется.
Агаризованная Среда Адамса.
Через 40-48 ч роста при 37oC образует колонии диаметром 0,5-1,5 мм; серовато-белые полупрозрачные, слегка выпуклые с блестящей поверхностью. В присутствии L-изолейцина (0,1-0,5 г/л) рост стимулируется и аналогичные колонии образуются через 18-20 ч.
Рост в мясопептонном бульоне. После 24 ч роста - сильное равномерное помутнение, характерный запах.
Физиолого-биохимические признаки.
Рост по уколу в мясопептонном агаре. Хороший рост по всему уколу. Микроорганизм является факультативным анаэробом.
Желатину не разжижает.
Рост на молоке хороший с коагуляцией молока.
Индол не образует.
Отношение к температуре. Растет на мясопептонном бульоне при температурах 20-42oC. Оптимальной температурой для роста является температура 33-37oC.
Отношение к pH среды. Растет на средах с pH от 6,0 до 8,0. Оптимальное значение pH - 7,2.
Отношение к источникам углерода. Хорошо растет на глюкозе, фруктозе, лактозе, маннозе, галактозе, ксилозе, глицерине, манните с образованием кислоты и газа.
Отношение к источникам азота. Усваивает азот в форме аммония, нитратов, а также азот некоторых органических соединений.
Устойчив к ампициллину.
Содержание плазмид. Клетки содержат многокопийную гибридную плазмиду pRhtC, несущую ген rhtC, сообщающий клеткам устойчивость к L-треонину (50 мг/мл) и детерминант устойчивости к ампициллину.
Штамм Е. coli MG442/pVIC40, pRhtC (ВКПМ В-7680) имеет те же культурально-морфологические и биохимические признаки, что и штамм ВКПМ В-7700 (MG442/pRhtC), за исключением того, что он наряду с плазмидой pRhtC содержит многокопийную гибридную плазмиду pVIC40, несущую гены треонинового оперона и детерминант устойчивости к стрептомицину. Штамм устойчив к ампициллину и стрептомицину.
Штамм Е.coli NZ10/pRhtB, pRhtC (ВКПМ В-7681) имеет те же культурально-морфологические и биохимические признаки, что и штамм ВКПМ В-7700 (MG442, pRhtC), за исключением того, что он нуждается для роста в L-треонине (0,1-5 мг/мл), рост его не стимулируется L-изолейцином, и он содержит многокопийную плазмиду pRhtB, несущую ген rhtB, сообщающий клеткам устойчивость к L-гомосерину (10 мг/мл), и детерминант устойчивости к канамицину. Штамм устойчив к канамицину и ампициллину.
Штамм Е. coli NZ10/pRhtBC (ВКПМ В-7682) имеет те же культурально-морфологические и биохимические признаки, что и штамм ВКПМ В-7681 (NZ10/pRhtB, pRhtC), за исключением того, что он содержит многокопийную плазмиду pRhtBC, несущую одновременно гены rhtB и rhtC, а также детерминант устойчивости к ампициллину. Штамм устойчив к ампициллину.
Способ получения аминокислот культивированием штаммов-продуцентов осуществляют следующим образом.
Аминокислоту получают путем культивирования бактерий, у которых Rt-активность, или одновременно Rt-активность и Rh-активность повышены, например, путем амплификации числа копий гена rhtC или rhtB, как описано выше, и которые обладают способностью к продукции аминокислоты при культивировании их в культуральной среде, где происходит накопление аминокислоты, с последующим выделением этой аминокислоты из среды (культуральной жидкости). Аминокислота представлена преимущественно L-гомосерином, L-треонином, L-аланином, L-валином или L-лейцином. В соответствии с настоящим изобретением культивирование бактерий, принадлежащих к роду Escherichia, выделение и очистку аминокислоты из культуральной жидкости осуществляют известными методами. Для культивирования используют синтетическую или натуральную среду. Такая среда включает источник углерода, азота, минеральные соли и необходимые добавки в количествах, оптимальных для роста и биосинтеза. В качестве источника углерода используют различные углеводы, такие как глюкоза, сахароза, различные органические кислоты. В зависимости от ассимилирующих способностей можно применять спирты, включая этанол или глицерол. В качестве источника азота используют аммиак, различные соли аммония, такие как сульфат аммония, или другие азотсодержащие соединения, такие как амины, а также природные источники азота, такие как пептон, гидролизат соевых бобов, или гидролизат микробных клеток. В качестве минеральных компонентов используются фосфат калия однозамещенный, сульфат магнезии, хлористый натрий, сульфат железа, сульфат марганца, карбонат кальция. Культивирование преимущественно осуществляют в аэробных условиях, таких как культивирование на мешалке, или с аэрацией и перемешиванием культуры. Температура культивирования - от 30 до 40oC, преимущественно 30-38oC. pH культуры - 5-9, преимущественно 6,5-7,2. pH культуры доводят до желаемых значений с помощью аммония, карбоната кальция, различных кислот, оснований или буферов. Культивирование осуществляют в течение 1-3 дней. После завершения культивирования выделение аминокислоты осуществляют путем удаления твердых частиц, таких как клетки, из среды с помощью центрифугирования или фильтрации через мембранные фильтры с последующим выделением и очисткой целевой аминокислоты с помощью ионообменника, фракционирования с помощью концентрации и кристаллизации.
Перечень фигур.
Фиг. 1. Клонирование и идентификация гена rhtB и гена rhtC.
Фиг. 2. Аминокислотная последовательность белка RhtC (последовательность N 2).
Фиг. 3. Нуклеотидная последовательность, содержащая ген rhtB (последовательность N 3).
Фиг. 4. Аминокислотная последовательность белка RhtB (последовательность N 4).
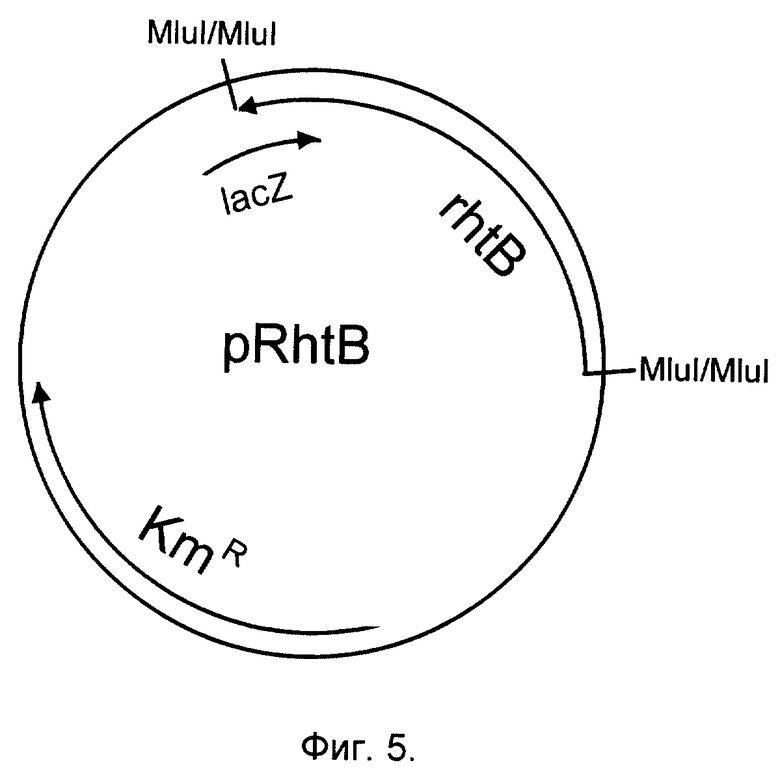
Фиг. 5. Структура плазмиды pRhtB, несущей ген rhtB.
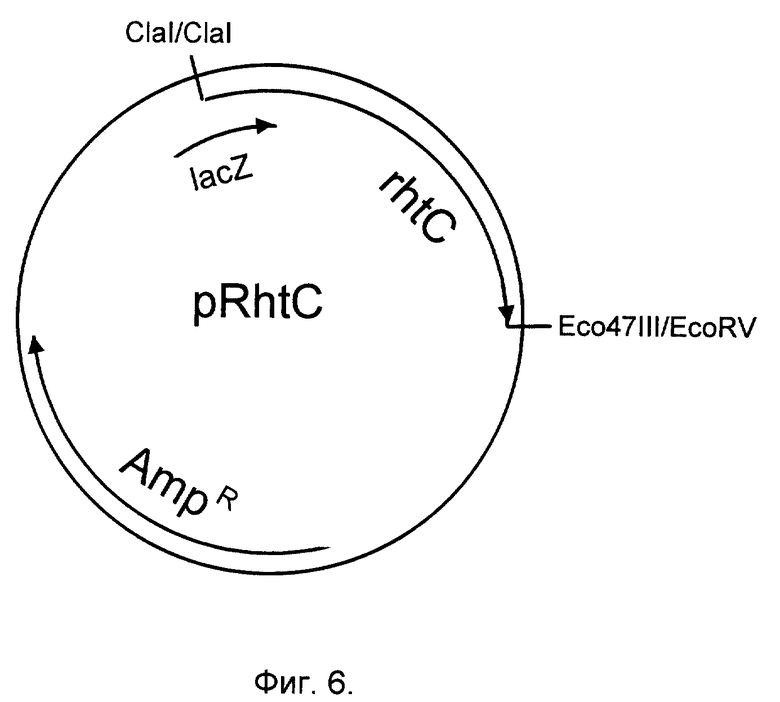
Фиг. 6. Структура плазмиды pRhtC, несущей ген rhtC.
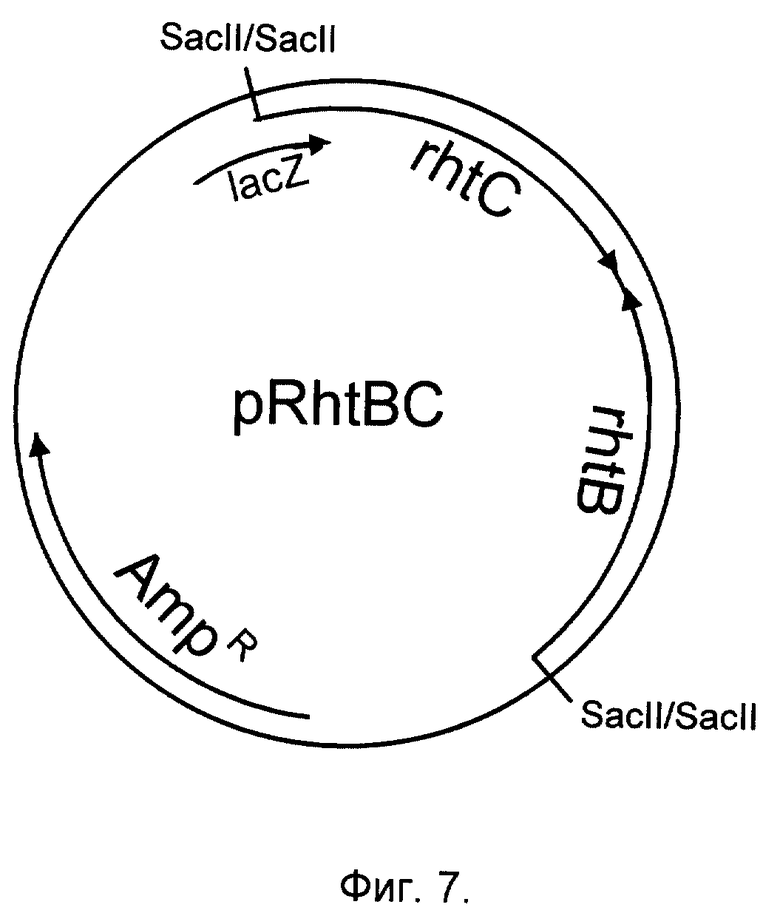
Фиг. 7. Структура плазмиды pRhtBC, несущей гены rhtB и rhtC.
Настоящее изобретение более конкретно поясняют нижеследующие примеры.
Пример 1. Получение фрагментов ДНК rhtB и rhtC
Этап 1. Клонирование генов, связанных с устойчивостью к L-треонину и L-гомосерину, на фазмиде миниMu.
Гены, связанные с устойчивостью к L-треонину и L-гомосерину, клонируют in vivo на фазмиде миниMu (Groisman et al. J. Bacteriol., 168, 357-364, 1986). В качестве донора используют штамм MG442, лизогенизированный Mu cts62. Клетки заражают фагом миниMu d5005, индуцируют профаг, полученным фаголизатом инфицируют клетки штамма ВКПМ-513 Mu cts62 (Hfr K10 metB) и высевают на минимальную среду с метионином (50 мкг/мл), L-гомосерином (10 мг/мл) и канамицином (40 мкг/мл) и культивируют при 30oC. Из выросших через 48 часов колоний выделяют плазмидную ДНК, которой трансформируют штамм Е. coli ВКПМ В-513 по стандартной методике. Трансформанты отбирают на чашках с L-агаром и канамицином (40 мкг/мл). Из трансформантов, которые устойчивы к L-гомосерину, выделяют плазмидную ДНК, которую анализируют с помощью рестриктного анализа. Из донорного штамма были отклонированы вставки (фрагменты ДНК), принадлежащие к двум разным областям хромосомы. Таким образом, на хромосоме Е. coli обнаружено по крайней мере два гена, которые при амплификации сообщают клеткам Е.coli устойчивость к L-гомосерину. Один тип вставок содержит ген rhtA, о котором уже сообщалось (ABSTRACTS of 17th International Congress of Biochemistry and Molecular Biology in conjugation with 1997 Annual Meeting of the American Society for Biochemistry and Molecular Biology, San Francicco, California August 24-29, 1997). Второй тип вставок содержит фрагмент SacII-SacII, сообщающий устойчивость к L-гомосерину и к L-треонину, или минимальный фрагмент MluI-M1uI длиной 0,8 kb, сообщающий устойчивость только к L-гомосерину (фиг. 1).
Этап 2: Идентификация генов rhtB и rhtC.
Полученный MluI-M1uI фрагмент секвенируют по двум цепям по методу Сенгера (Sanger et al. Proc. Natl. Acad. Sci. USA, 74, 5463, 1977) и выясняют, что он включает открытую рамку считывания f138 (с 61959 по 61543 нуклеотид в последовательности М87049, GenBank) и 201 нуклеотид, расположенный перед ней. Вставка, содержащая открытую рамку считывания f138 только со 160 5'-фланкирующими нуклеотидами не обеспечивает устойчивости к L-гомосерину. Указанная последовательность выше f138 не содержит стоп-кодона в рамке f138. Кроме того, одному из ATG кодонов в этой последовательности предшествует участок связывания с рибосомами (SD-последовательность, нуклеотиды с 62171 по 62166 в М87049). Эта удлиненная открытая рамка считывания (нуклеотиды 62160-61546) представляет собой структурный ген rhtB. Кодируемый им белок RhtB (фиг. 4) является сильно гидрофобным белком и содержит потенциальные трансмембранные сегменты.
Плазмида, содержащая ген rhtB, сообщает клеткам устойчивость только к высоким концентрациям гомосерина (фиг. 1). В то же время фрагмент ДНК SacII-SacII, сообщающий клеткам одновременно устойчивость к высоким концентрациям гомосерина и треонина, содержит вторую неидентифицированную открытую рамку считывания, о128. Субклонирование о128 на минимальном фрагменте ClaI-Eco47III показывает, что плазмида, несущая этот ген, сообщает клеткам устойчивость только к высоким (50 мг/мл) концентрациям L-треонина (фиг. 1). Субклонированный фрагмент был секвенирован, и оказалось, что он содержит дополнительный нуклеотид (G) в положении между нуклеотидами 61213 и 61214 последовательности М87049. Добавление этого нуклеотида к указанной последовательности элиминирует сдвиг рамки считывания и удлиняет открытую рамку считывания в направлении 5'-фланкирующей области до 60860 нуклеотида включительно. Этот новый ген (нуклеотиды N 60860-61480 в М87049) обозначен как rhtC. Оба гена, rhtB и rhtC, кодируют белки, гомологичные транспортеру, связанному с транспортом лизина из клеток Corynbacterium glutamicum.
Пример 2. Влияние амплификации гена rhtB, или гена rhtC на продукцию L-гомосерина.
<1> Конструирование L-гомосерин-продуцирующего штамма Е.coli NZ10/pAL4, pRhtB (ВКПМ В-7658) и получение L-гомосерина с его помощью.
Фрагмент ДНК rhtB клонируют в многокопийный плазмидный вектор pUK21 (Vieira, Messing, Gene, 100, 189-194, 1991). В результате получают плазмиду pRhtB (фиг. 5).
Штамм E. coli NZ10, который является Leu+ ревертантом известного штамма C600 (thrB, leuB) (Appleyard, Genetics., 39, 440-452, 1954), трансформируют плазмидой pAL4, которая представляет собой вектор pBR322, несущий ген thrA, кодирующий аспартокиназу-гомосериндегидрогеназу I. В результате получают штамм E.coli NZ10/pAL4, способный к продукции L-гомосерина. Полученный штамм трансформируют плазмидой pRhtB или вектором pUK21 и в результате получают штаммы Е.coli NZ10/pAL4, pRhtB (ВКПМ В-7658) и Е.coli NZ10/pAL4, pUK21 (ВКПМ В-7661). Штамм Е.coli NZ10/pAL4, pRhtB сообщает клеткам устойчивость к высокой концентрации гомосерина (10 мг/мл), к которой штамм Е.coli NZ10/pAL4, pUK21 остается чувствительным. Каждый из полученных штаммов культивируют при 37oC в течение 18 часов в бульоне LB (Миллер, Дж. Эксперименты в молекулярной генетике. М.: Мир, 1976, с. 395), содержащем 50 мг/л канамицина и 100 мг/мл ампициллина. По 0,3 мл полученной культуральной жидкости вносят в 3 мл ферментационной среды, имеющей состав, указанный ниже, и содержащей 50 мг/мл канамицина и 100 мг/мл ампициллина, содержащейся в пробирках 20 х 200 мм, и культивируют при 37oC 46 часов на роторной качалке.
Состав ферментационной среды, г/л:
Глюкоза - 80
(NH4)2S04 - 22
К2HPO4 - 2
NaCl - 0,8
MgSO4 • 7H2O - 0,8
FeSO4 • 7H2O - 0,02
MnSO4 • 5H2O - 0,02
Тиамин HCl - 0,0002
Дрожжевой экстракт - 1,0
CaCO3 - 30 (добавляют после стерилизации)
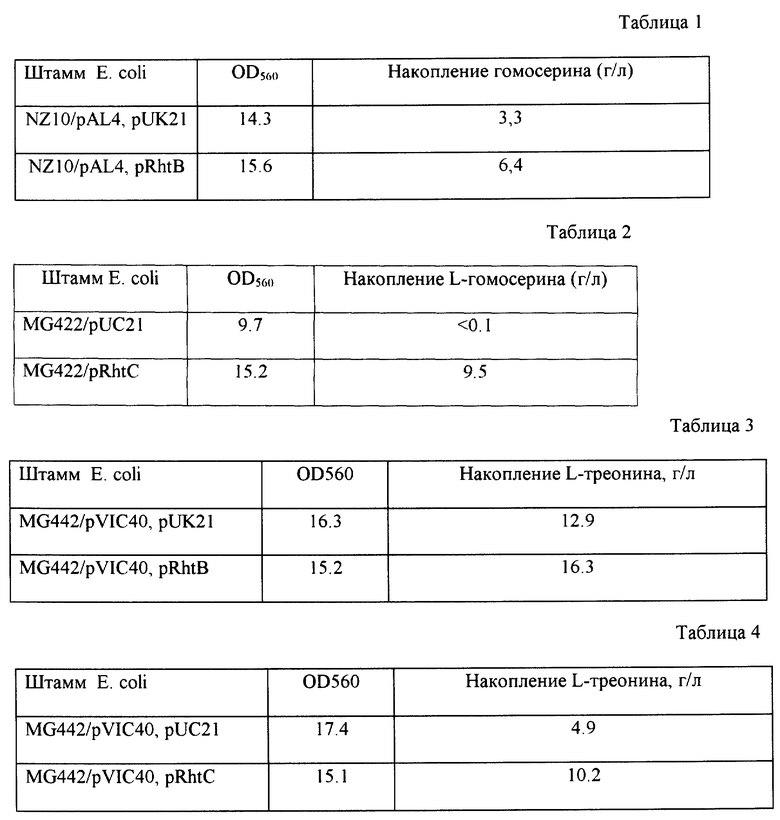
После культивирования определяют количество накопившегося в среде L-гомосерина и оптическую плотность культуральной жидкости при 560 нм известными методами. Как показано в табл. 1, штамм NZ10/pAL4, pRhtB накапливает L-гомосерин в большем количестве, чем штамм NZ10/pAL4, pUK21, в котором число копий гена rhtB не увеличено.
<2> Конструирование L-гомосерин-продуцирующего штамма Е. coli MG442, pRhtC (ВКПМ В-7700) и получение L-гомосерина с его помощью.
Фрагмент ДНК rhtC клонируют в многокопийный плазмидный вектор pUC21 (Vieira, Messing, Gene, 100, 189-194, 1991). В результате получают плазмиду pRhtC (фиг. 6).
Известный штамм Е. coli MG442 (Гусятинер и др., 1978, Генетика, 14, 947-956) трансформируют плазмидой pRhtC или вектором pUC21 и в результате получают штаммы Е.coli MG442/pRhtC (ВКПМ В-7700) и Е.coli MG442/pUC21. Штамм Е. coli MG442/pRhtC обладает устойчивостью к высокой концентрации треонина (50 мг/мл), к которой штамм Е.coli MG442/pUC21 остается чувствительным.
Каждый из полученных таким образом штаммов культивируют при 37oC 18 часов в LB бульоне с 100 мг/л ампициллина. Затем по 0,3 мл полученной культуральной жидкости вносят в пробирки 20 х 200 мм с 3 мл ферментационной среды, описанной выше, содержащей 100 мг/л ампициллина, и культивировали при 37oC 72 часа на роторной качалке. После культивирования количество накопленного в среде L-гомосерина, а также оптическую плотность культуральной жидкости при 560 нм измеряют известными методами. Результаты представлены в табл. 2.
Как показано в табл. 2, штамм MG442 после введения в него плазмиды pRhtC из продуцента треонина превращается в продуцент гомосерина и накапливает этой аминокислоты больше, чем штамм MG442/pUC21, в котором число копий гена rhtC не увеличено.
Пример 3. Влияние амплификации гена rhtB или гена rhtC на продукцию L-треонина.
<1> Конструирование штамма-продуцента L-треонина Е.coli MG442/pVIC40, pRhtB (ВКПМ В-7660) и получение L-треонина с его помощью.
Для получения нового штамма-продуцента L-треонина Е.coli MG442/pVIC40, pRhtB в качестве реципиента используют штамм Е.coli MG442 (см. пример 2). Этот штамм трансформируют известной плазмидой pVIC40 (Патент США 5175107, 1992). Трансформанты отбирают на чашках с LB агаром, содержащем 100 мг/л стрептомицина, и получают штамм MG442/pVIC40. Штамм MG442/pVIC40 трансформируют плазмидами pRhtB или pUK21 и получают штаммы MG442/pVIC40, pRhtB (ВКПМ В-7660) и MG442/pVIC40, pUK21 (ВКПМ В-7663). Штамм E.coli MG442/pVIC40, pRhtB обладает устойчивостью к высокой концентрации гомосерина (10 мг/мл), к которой штамм Е.coli MG442/pUC21 остается чувствительным.
Каждый из этих штаммов культивируют при 37oC 18 часов в LB бульоне, как в примере 2 с 50 мг/л канамицина и 100 мг/л стрептомицина. Затем 0,3 мл полученной культуральной жидкости вносят в пробирки 20 х 200 мм с 3 мл ферментационной среды, описанной в примере 2, содержащей 50 мг/л канамицина и 100 мг/л стрептомицина, и культивируют при 37oC 46 часов на роторной качалке. После культивирования количество накопившегося в среде L-треонина и оптическую плотность культур при 560 нм измеряют известными способами. Результаты представлены в табл. 3.
Как показано, в табл. 3, штамм MG442/pVIC40, pRhtB накапливает L-треонин в большем количестве, чем штамм MG442/pVIC40, pUK21, у которого число копий гена rhtB не увеличено.
<2> Конструирование штамма-продуцента L-треонина Е.coli MG442/pVIC40, pRhtC (ВКПМ В-7680) и получение L-треонина с его помощью.
Штамм MG442/pVIC40 трансформируют плазмидами pRhtC или pUC21 и получают штаммы MG442/pVIC40, pRhtC (ВКПМ В-76800) и MG442/pVIC40, pUC21.
Штамм MG442/pVIC40, pRhtC обладает устойчивостью к высокой концентрации треонина (50 мг/мл), к которой штамм Е.coli MG442/pVIC40, pUC21 остается чувствительным.
Каждый из этих штаммов культивируют при 37oC 18 часов в LB бульоне, как в примере 2 со 100 мг/л ампициллина и 100 мг/л стрептомицина. Затем 0,3 мл полученной культуральной жидкости вносят в пробирки 20 х 200 мм с 3 мл ферментационной среды, описанной в примере 2, содержащей 100 мг/л ампициллина и 100 мг/л стрептомицина, и культивируют при 37oC 46 часов на роторной качалке. После культивирования количество накопившегося в среде L-треонина и оптическую плотность культур при 560 нм измеряют известными способами. Результаты представлены в табл. 4.
Как показано в табл. 4, штамм MG442/pVIC40, pRhtC накапливает L-треонин в большем количестве, чем штамм MG442/pVIC40, pUC21, у которого число копий гена rhtC не увеличено.
Пример 4. Влияние совместной амплификации гена rhtB и гена rhtC на продукцию аминокислот.
Фрагмент ДНК SacII-SacII, содержащий одновременно гены rhtB и rhtC, клонируют в многокопийный плазмидный вектор pUC21 (Vieira, Messing, Gene, 100, 189-194, 1991). В результате получают плазмиду pRhtBC (фиг. 1, фиг. 7).
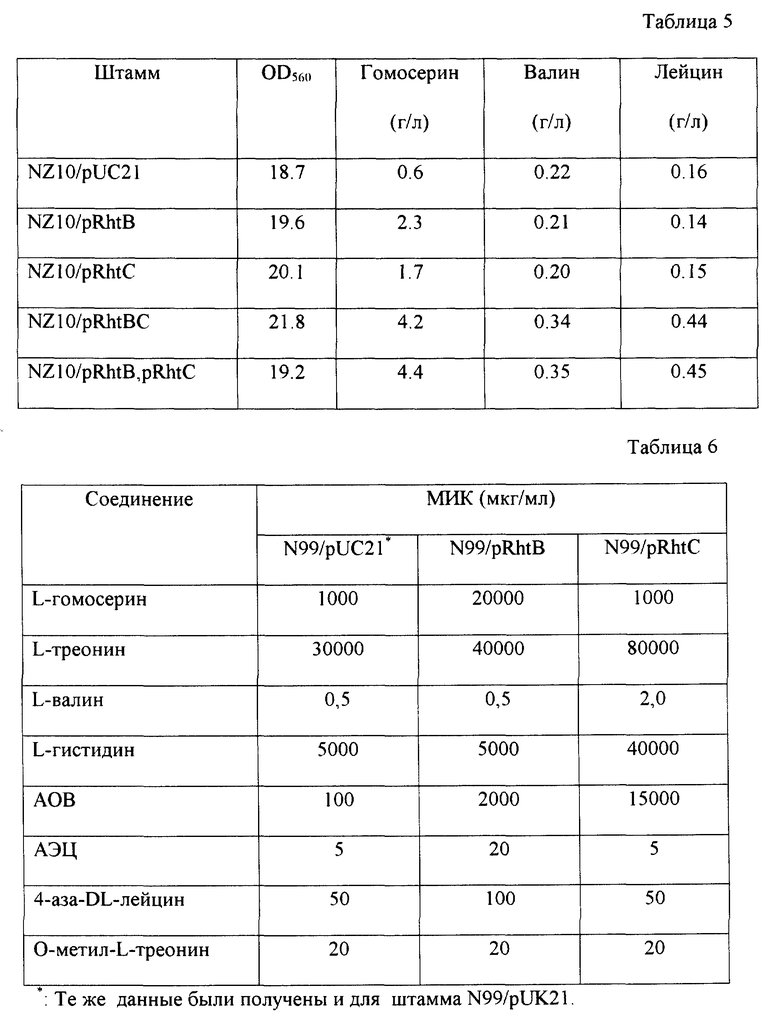
Штамм E. coli NZ10 трансформируют вектором pUC21 или плазмидами pRhtB, pRhtC, pRhtBC и в результате получают штаммы Е.coli NZ10/pUC21 (ВКПМ В-7685), Е.coli NZ10/pRhtB (ВКПМ В-7683), Е.coli NZ10/pRhtC (ВКПМ В-7684), Е. coli NZ10/pRhtB, pRhtC (ВКПМ В-7681) и Е.coli NZ10/pRhtBC (ВКПМ В-7682). Штамм Е.coli NZ10/pRhtB обладает повышенной устойчивостью к высокой концентрации гомосерина (10 мг/мл), штамм Е.coli NZ10/pRhtC обладает повышенной устойчивостью к L-треонину (50 мг/мл), а штаммы Е.coli NZ10/pRhtB, pRhtC и Е. coli NZ10/pRhtBC обладают одновременно повышенной устойчивостью к высокой концентрации гомосерина (10 мг/мл) и треонина (50 мг/мл), к которым штамм Е. coli NZ10/pUC21 остается чувствительным.
Каждый из полученных штаммов культивируют, как описано выше, внося в посевную и ферментационную среду соответствующие антибиотики. После культивирования в течение 46 часов количество накопившихся в среде аминокислот и оптическую плотность культур при 560 нм измеряют известными способами. Результаты представлены в табл. 5.
Как показано в табл. 5, одновременная амплификация генов rhtB и rhtC в клетках штамма NZ10 повышает накопление в культуральной жидкости L-гомосерина, L-валина и L-лейцина. Этот результат показывает, что в клетках продукты генов rhtB и rhtC могут между собой взаимодействовать.
Пример 5. Влияние амплификации гена rhtB и гена rhtC на устойчивость бактерий E.coli к некоторым аминокислотам и аналогам аминокислот.
Как показано выше, плазмиды, несущие гены rhtB и rhtC, оказывают положительное влияние на накопление некоторых аминокислот в культуральной жидкости различными штаммами-продуцентами. Оказалось также, что характер накапливаемых аминокислот зависит от генотипа штамма. Гомология продуктов генов rhtB и rhtC с лизиновым транспортером LysE, осуществляющим экспорт L-лизина из клеток Corynebacterium glutamicum (Vr1jic et al. , Mol. Microbiol., 22, 815-826, 1996), указывает на аналогичную функцию белков RhtB и RhtC. Известно, что повышение активности генов, контролирующих транспорт из клеток различных ингибиторов роста, увеличивает их устойчивость к соответствующим соединениям. В связи с этим определяют влияние плазмид pRhtB и pRhtC на устойчивость штамма E.coli N 99, который является StrR мутантом известного штамма E. coli W3350 (ВКПМ В-1557), к некоторым аминокислотам и аналогам аминокислот. С этой целью штамм N 99 трансформируют плазмидами pRhtB, pRhtC и векторами pUC21 и pUK21. Ночные культуры полученных штаммов N 99/pRhtB, N 99/pRhtC, N 99/pUK21 и N 99/pUC21, выращенные в минимальной среде M9 на качалке (около 109 клеток/мл) разводят 1:100 и подращивают в течение 5 часов в той же среде. Затем полученные культуры в логарифмической фазе роста разводят и приблизительно по 104 жизнеспособных клеток наносят на высушенные чашки с агаризованной (2% агара) средой M9, содержащей различные концентрации аминокислот, или аналогов аминокислот. Рост или отсутствие роста определяют через 46-48 часов. Таким образом устанавливают минимальные ингибирующие концентрации (МИК) этих соединений (табл. 6).
Как видно из таблицы, амплификация гена rhtB существенно повышает устойчивость бактерий не только к гомосерину, но и к аналогу треонина, α-амино-β-оксивалериановой кислоте (АОВ), в меньшей степени возрастает устойчивость к L-треонину и к аналогу L-лизина, (S)-2-аминоэтил-L-цистеину (АЭЦ). Кроме того, наблюдается некоторое увеличение резистентности бактерий к аналогу лейцина, 4-аза-DL-лейцину. Амплификация гена rhtC кроме L-треонина существенно повышает устойчивость бактерий к АОВ, L-гистидину и L-валину. Эти результаты свидетельствуют о том, что каждый из предполагаемых транспортеров, RhtB и RhtC, обладают специфичностью по отношению к нескольким субстратам (аминокислотам) или может обнаруживать неспецифический эффект в результате амплификации.
| название | год | авторы | номер документа |
|---|---|---|---|
| ФРАГМЕНТ ДНК rhtB, КОДИРУЮЩИЙ СИНТЕЗ БЕЛКА RhtB, ПРИДАЮЩЕГО УСТОЙЧИВОСТЬ К L-ГОМОСЕРИНУ БАКТЕРИЯМ ESCHERICHIA COLI, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 1998 |
|
RU2144564C1 |
| ФРАГМЕНТ ДНК ИЗ ESCHERICHIA COLI, ОПРЕДЕЛЯЮЩИЙ ПОВЫШЕННУЮ ПРОДУКЦИЮ L-АМИНОКИСЛОТ (ВАРИАНТЫ), И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 1999 |
|
RU2175351C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН yahN | 2007 |
|
RU2392322C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ, ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2001 |
|
RU2229513C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА, ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ ТРЕОНИНА (ВАРИАНТЫ) | 2002 |
|
RU2244007C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ ESCHERICHIA | 2003 |
|
RU2275424C2 |
| БАКТЕРИЯ РОДА ESCHERICHIA - ПРОДУЦЕНТ L-ТРЕОНИНА И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА L-ТРЕОНИНА С ЕЕ ИСПОЛЬЗОВАНИЕМ | 2012 |
|
RU2515095C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН ltaE | 2005 |
|
RU2304166C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ, ШТАММ Escherichia coli - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2001 |
|
RU2215782C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ, ШТАММ Escherichia coli - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2001 |
|
RU2215784C2 |




Изобретение относится к биотехнологии и генетической инженерии. Фрагмент ДНК rhtC кодирует синтез белка RhtC, который придает повышенную устойчивость к L-треонину бактериям Escherichia coli. На основе мультикопийной плазмиды pRhtC, содержащей этот фрагмент, сконструированы штаммы Escherichia coli, способные к повышенной продукции L-аминокислот по сравнению со штаммами, не содержащими плазмиду pRhtC. Аминокислоты получают путем культивирования соответствующих штаммов в подходящих питательных средах с последующим выделением и очисткой целевой аминокислоты. Изобретение позволяет увеличить выход L-аминокислот. 2 с. и 4 з.п. ф-лы, 7 ил., 6 табл.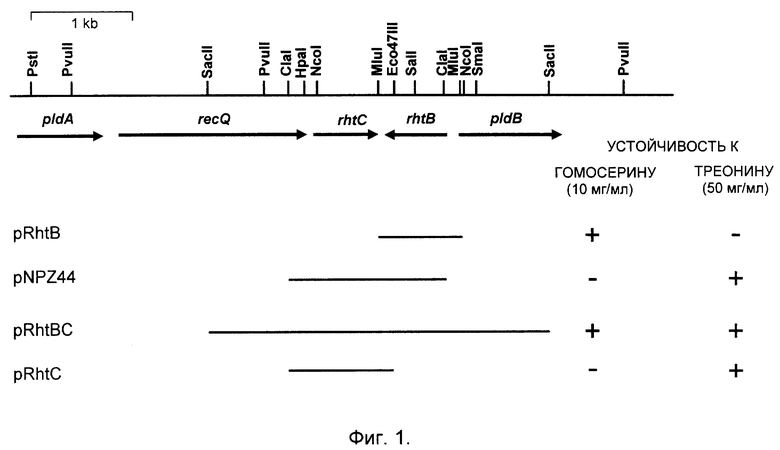
| Способ получения L-треонина | 1981 |
|
SU974817A1 |
| US 4388405 A, 14.06.83 | |||
| US 4391907 A, 05.07.83 | |||
| US 4393135 A, 12.07.83 | |||
| УСТРОЙСТВО для ИЗМЕРЕНИЯ ПАРАМЕТРОВ ВРАЩАЮЩИХСЯ ДЕТАЛЕЙ МАШИН | 0 |
|
SU301572A1 |
| JP 63273487 A, 10.11.88. | |||

