Настоящее изобретение относится к области диагностики и лечения раковых заболеваний, а также к области определения трансформирующей способности потенциальных опухолеродных или способствующих возникновению опухолей агентов (эти два термина будут в дальнейшем использоваться как взаимозаменяемые).
Общей доминантой настоящего изобретения является то, что во всех отмеченных выше областях в соответствии с настоящим изобретением может быть использован апоптин или его производные и/или их фрагменты (в дальнейшем все рассматривается как апоптиновая или апоптиноподобная активность). Апоптин представляет собой белок, впервые обнаруженный в вирусе анемии цыплят (Noteborn, M. H.M., De Boer, G.P., Van Roozelaar, D., Karreman, C., Kranenburg, 0. , Vos, J., Jeurissen, S., Zantema, A., Hoeben, R., Koch, G., Van Ormondt, H., and Van der Eb, A.J. (1991). Характеристика клонированного ДНК вируса анемии цыплят, который содержит все элементы инфекции репликационного цикла. Journal of Virology 65, 3131-3139) и который впервые был назван как VP3. Апоптическая активность этого белка была раскрыта группой настоящих изобретателей (Noteborn, M.H.M., Todd, D., Verschueren, С.A.J., De Gauw, H. W. P. M. , Curran, W.L., Veldkamp, S., Douglas, A.J., McNulty, M.S., Van der Eb, A. J., and Koch, G. (1994). Один из белков вируса анемии цыплят индуцирует апоптоз. Journal of Virology 68, 346-351).
Как было отмечено выше, в настоящем изобретении используют действие апоптина, индуцирующее апоптоз.
Апоптоз - это активный и программируемый физиологический процесс уничтожения чрезмерно, сильно поврежденных или злокачественных клеток (Earnshaw, W. C., 1995. Ядерные изменения при апоптозе. Current Opinion in Cell Biology 7, 337-343; Duke, R.C., Ocjius, D.M., Young, J, D-E. (1996). Суицид клетки в здоровье и заболеваниях. Scientific American December 1996, 48-55). Апоптоз характеризуется сокращением клеток, ядерной сегментацией и фрагментацией цитоплазмы, конденсацией и расщеплением ДНК на фрагменты по размеру доменов, в большинстве случаев за этим следует межнуклеосомное разрушение. Апоптические клетки распадаются на фрагменты, представляющие собой заключенные внутри мембран апоптические тела. И наконец, соседние клетки и/или макрофаги будут быстро фагоцитировать эти умирающие клетки (Wyllie, A.H., Kerr, J.P.R., Currie, A.R. (1980). Смерть клетки: Значение апоптолиза. International Review of Cytology 68, 251-306, White, E. (1996). Жизнь, смерть и погоня за апоптозом. Genes and development 10, 1-15). Клетки, выросшие в условиях тканевой культуры, и клетки тканей могут быть проанализированы на наличие признаков апоптоза с помощью агентов, окрашивающих хромосомную ДНК, таких как, например, DAPI или пропидий иодид, который окрашивает обычную ДНК (из хроматина) сильно и равномерно, а апоптический хроматин - слабо и/или неравномерно (Noteborn, M.H.M., Todd, D., Verschueren, C.A.J., De Gauw, H.W.P.M. , Curran, W.L., Veldkamp, S., Douglas, A.J., McNulty, M.S., Van der Eb, A.J. , and Koch, G. (1994). Один из белков вируса анемии цыплят индуцирует апоптоз. Journal of Virology 68, 346-351, Telford, W.G., King, L.E., Fraker, P. J. (1992). Сравнительная оценка некоторых связывающихся ДНК красителей в определении апоптоз-ассоциированной деградации хроматина методом проточной цитометрии. Cytometry 13, 137-143).
Процесс апоптоза может быть инициирован с помощью разнообразных регулирующих стимуляторов (Wyllie, A.H. (1995). Генетическая регуляция апоптоза. Current Opinion in Genetics and Development 5, 97-104, White, E. (1996). Жизнь, смерть и погоня за апоптозом. Genes and development 10, 1-15, Levine, A. J. (1997). Р53, клеточный сторож, хранитель развития и роста. Cell 88, 323-331). Изменения в степени выживания клеток играют важную роль в человеческом патогенезе, например, при развитии раковых заболеваний, которые вызывает усиление пролиферации и/или уменьшение числа смертей клеток (Kerr, J.F. R. , Winterford, С.М., and Harmon, B.V. (1994). Апоптоз: Значение в раковых заболеваниях и раковой терапии. Cancer 73, 2013-2026, Paulovich, A.G., Toczyski, D., Hartwell, H. (1997). Когда точка инициации не действует. Cell 88, 315-321). Было продемонстрировано, что разнообразные хемотерапевтические агенты и облучение индуцируют апоптоз раковых клеток, который во многих случаях осуществляется при участии белка-супрессора опухоли р53 (Thompson, С. В. (1995). Апоптоз в патогенезе и лечении заболеваний. Science 267, 1456-1462, Bellamy, С.ОС., Malcomson, R.D.G., Harrison, D.J., and Wyllie. H. 1995. Смерть и заболевания клетки: Биология и регуляция апоптоза. Seminars on Cancer Biology 6, 3-12, Steller, H. (1995). Механизмы и гены клеточного суицида. Science 267, 1445-1449, McDonell T. J., Meyn, R.E., Robertson, L.E (1995). Использование апоптической регуляции смерти клеток в терапии рака. Seminars in Cancer Biology 6, 53-60). Однако многие опухоли подвергаются мутации в р53 в процессе своего развития, что часто связано со снижением воздействия на них противораковой терапии. Вызывающие трансформацию белки ДНК опухолевых вирусов инактивируют р53 путем косвенного или прямого связывания с ним (Teodoro, J.G. and Branton, P.E. (1997). Регуляция апоптоза посредством вирусных генных продуктов. Journal of Virology 71, 1739-1746). Примером такого агента является большой Т-антиген ДНК опухолевого вируса SV40. В определенных гемопоэтических опухолях высокий уровень экспрессии Bcl-2-онкогена сочетается с сильной устойчивостью к различным апоптоз-индуцирующим хемотерапевтическим агентам (Hockenberry, D.M. (1994). Bcl-2 в раковых клетках, развитие и апоптоз. Journal of Cell Science, Supplement 18, 51-55, Sachs, L. and Lotem, J. (1993). Контроль программированной смерти клеток для нормальных клеток и клеток лейкоза: Новые применения в терапии. Blood 82, 15-21). В отношении таких раковых заболеваний, которые устойчивы ко многим цитотоксическим агентам, развивается альтернативная противоопухолевая терапия, основанная на индукции апоптоза (Thompson, С.В. (1995). Апоптоз в патогенезе и лечении заболеваний. Science 267, 1456-1462 и Paulovich, A.G., Toczyski, D., Hartwell, H. (1997). Когда точка инициации не действует. Cell 88, 315-321).
Апоптин представляет собой небольшой белок, полученный из вируса анемии цыплят (CAV; Noteborn and De Boer, G.F. (1996). Patent USA/no. 030, 335, Noteborn, M.H.M., De Boer, G.P., Van Roozelaar, D., Karreman, C., Kranenburg, О., Vos, J., Jeurissen, S., Zantema, A., Hoeben, R., Koch, G., Van Ormondt, H., and Van der Eb, A.J. (1991). Характеристика клонированного ДНК вируса анемии цыплят, который содержит все элементы инфекции репликационного цикла. Journal of Virology 65, 3131-3139, Noteborn, M.H.M., Todd, D., Verschueren, C.A.J., De Gauw, H.W.P.M., Curran, W.L., Veldkamp, S., Douglas, A. J. , McNulty, M.S., Van der Eb, A.J., and Koch, G. (1994). Один из белков вируса анемии цыплят индуцирует апоптоз. Journal of Virology 68, 346-351), который может индуцировать апоптоз в злокачественных и трансформированных клеточных линиях человека, но не в нетрансформированных диплоидных клетках человека. In vitro апоптин не может индуцировать программируемую смерть клеток в нормальных лимфоидных, кожных фибробластных, эпидермальных, эндотелиальных клетках и клетках гладкой мускулатуры. Однако, когда нормальные клетки трансформируются, например, под воздействием вызывающих трансформацию генов SV40, они становятся восприимчивыми к апоптозу, индуцированному апоптином. (Danen-van Oorschot, A.A.A.M., Den Hollander, A., Tak-ayama, S., Reed, J. , Van der Eb, A.J., and Noteborn, M.H.M. (1997). BAG-1 ингибирует р53-индуцированный, но не апоптин-индуцированный апоптоз. Apoptosis, In Press и Noteborn, M.H.M. (1996). PCT application WO 96/41191. Апоптин индуцирует апоптоз в человеческих трансформированных и злокачественных клетках, но не в нормальных клетках, что является существенной характеристикой для развития антираковой терапии). Длительная экспрессия апоптина в нормальных человеческих фибробластах показала, что апоптин не проявляет токсической или трансформирующей активности в этих клетках. Было обнаружено, что в нормальных клетках апоптин преимущественно локализован в цитоплазме, в то время как в трансформированных или злокачественных клетках он был локализован в ядрах, что предполагает, что локализация апоптина связана с его активностью в отношении индукции клеточной смерти (Danen-van Oorschot, A.A.A.M., Den Hollander, A., Tak-ayama, S., Reed, J., Van der Eb, A.J., and Noteborn, M.H. M. (1997). BAG-1 ингибирует р53-индуцированный, но не апоптин-индуцированный апоптоз. Apoptosis, In Press).
Далее, было установлено, что апоптин может индуцировать апоптоз при отсутствии функционального р53 (Zhuang, S.-M., Landegent, J.E., Verschueren, С. A. J., Falkenburg, J.H.F., Van Ormondt, H., Van der Eb, A.J., Noteborn, M.H. M. (1995). Апоптин, белок закодированный вирусом анемии цыплят, индуцирует смерть клетки в различных гематологических злокачественных клетках in vitro. Leukemia 9 SI, 118-120) и его не могут ингибировать Bcl-2-, Bcr-abl (Zhuang, S. -M. , Shvarts, A., Van Ormondt, H., Jochemsen, A.-G., Van der Eb, A.J., Noteborn, M.H.M. (1995). Апоптин, белок выделенный из вируса анемии цыплят, индуцирует р53-независимый апоптоз в человеческих остеосаркомных клетках. Cancer Research 55, 486-489), Bcl-2-ассоциированный белок BAG-1 и белок caspase -ингибитор вакцины коровьей оспы CrmA (Danen-van Oorschot, A.A.A.M., Den Hollander, A., Tak-ayama, S., Reed, J., Van der Eb, A.J., and Noteborn, M. H. M. (1997). BAG-1 ингибирует р53-индуцированный, но не апоптин-индуцированный апоптоз. Apoptosis, In Press, Noteborn, M.H.M. (1996). PCT application WO 96/41191. Апоптин индуцирует апоптоз в человеческих трансформированных и злокачественных клетках, но не в нормальных клетках, что является существенной характеристикой для развития анти-раковой терапии). И наконец, оказывается, что клетки, которые только иммортализованы и таким образом минимально трансформированы, также могут быть убиты апоптином.
Таким образом, апоптин является потенциальным противоопухолевым агентом, также и в отношении опухолей, которые являются не восприимчивыми к действию или в меньшей степени восприимчивы к действию хемотерапевтических агентов из-за отсутствия функционального р53, (сверх) экспрессии Всl-2 или других апоптоз-ингибирующих генов. Тот факт, что апоптин не индуцирует апоптоз в нормальных клетках человека, дает возможность предположить, что токсический эффект при лечении апоптином in vivo будет очень низким. В дополнение, оказалось, что даже предраковые, минимально трансформированные клетки, могут быть восприимчивыми к воздействию апоптина, вызывающему клеточную смерть.
Зная, что апоптин совершенно безопасен в нормальных клетках, но как только клетка становится трансформированной и/или иммортализованной (эти термины могут использоваться как взаимозаменяемые), настоящие изобретатели разработали некоторые пути его использования, основываясь на этом открытии.
Таким образом, изобретение относится к способу определения трансформирующей способности агента, который предположительно может вызывать трансформацию, включающий обеспечение не трансформированной клетке индуцируемой активности на подобие активности апоптина, воздействие на клетку указанного вызывающего трансформацию агента и определение локализации указанной апоптической активности внутри этой клетки или определение индукции апоптоза в указанной клетке.
Должно быть понятно, что под указанной выше апоптической активностью подразумеваются и вещества, имеющие указанную активность.
Предпочтительно обеспечивать указанную клетку указанной апоптической активностью путем трансдукции указанной клетки с помощью молекулы рекомбинантной нуклеиновой кислоты, кодирующей указанную активность. Апоптин-подобную активность в данном случае определяют как любое (предпочтительно белковое) вещество, имеющее такую же активность, как и VP 3 или апоптин вируса анемии цыпленка. В особенности под это определение подходят аллельные варианты, производные и/или фрагменты апоптина, где производные определяют как содержащие замены аминокислот, не приводящие к полной потере апоптической активности. Должно быть понятным, что такая же активность означает, что тип активности тот же, но количество может быть другим. Способы в соответствии с изобретением особенно пригодны в случае применений, в которых указанный предположительно вызывающий трансформацию агент является белковым веществом. Это позволяет указанному белковому веществу одновременно экспессироваться в указанной не трансформированной клетке с указанной апоптической активностью. Примером белкового вещества, является большой Т-антиген SV40 или его функциональный эквивалент.
Изобретение также относится к модификации гена апоптина, приводящей к изменениям апоптинового белка, что дает возможность апоптину проникать внутрь ядра не трансформированных и трансформированных/опухолеродных клеток, что в результате приводит к индукции апоптоза. Белок апоптин увеличивают за счет присоединения сигнального пептида, определяющего ядерную локализацию, SV 40. В особенности в указанное определение апоптина включаются алелльные варианты, производные и/или фрагменты апоптина, в которых производные определены как имеющие замены аминокислот, которые не приводят к полной потере апоптиновой активности. Это позволяет апоптиновому белку экспрессироваться в нетрансформированных клетках с указанной апоптической активностью. Фрагменты апоптина с указанной апоптической активностью, которые не способны проникать внутрь ядра не трансформированных или трансформированных клеток с помощью своих собственных последовательностей, способны проникать в ядро за счет модификаций и индуцировать апоптоз.
Кроме того, изобретение относится к способу определения предрасположенности клетки к превращению в опухолевую клетку путем обеспечения указанной клетке индуцируемой апоптин-подобной апоптической активности и сообщение указанной клетке относительно мягкой опухолеродной активности и определение апоптоза в указанной клетке и/или определение локализации указанной апоптической активности в указанной клетке. В этом случае предположительно вызывающий трансформацию агент, как обсуждалось ранее, уже присутствует в указанной клетке в виде мутации, приводящей к канцерогенной или опухолеродной активности. В этом случае факт, что апоптин индуцирует апоптоз только в клетках, которые уже были трансформированы, ведет к возможности проверить, имеют ли клетки мутацию, которая приведет к имммортализации или трансформации в условиях мягкого воздействия трансформирующей активности, такой как обработки UV-излучением и рентгеновским излучением.
Таким образом может быть определена вероятность предрасположенности ряда клеток к раковым заболеваниям. Это, конечно, приводит к применению в области диагностики у людей, подверженных наследственному риску возникновения раковых заболеваний, и дает возможность проведения профилактики, что является еще одним объектом настоящего изобретения. Такого рода диагностика может быть также применена для консультации людей, дети которых имеют предрасположенность к раковым заболеваниям.
Таким образом, изобретение относится к способу определения предрасположенности пациента к наследственным формам раковых заболеваний, включающему обработку образца релевантной подгруппы клеток этого пациента с помощью способа, раскрытого здесь ранее. И, кроме того, изобретение касается способа определения генной мутации, имеющей канцерогенную и/или трансформирующую активность в клетке, включающего обработку указанной клетки способом согласно изобретению.
Как утверждалось ранее, другим объектом настоящего изобретения являются средства для профилактической обработки подгруппы клеток у пациента, когда эта подгруппа клеток имеет склонность к раковым заболеваниям. Эти средства включают нуклеиновую кислоту, кодирующую апоптин-подобную активность, предпочтительно существующую в форме инструмента доставки гена, который может иметь вирусное или другое происхождение. В уровне техники описаны многие такие инструменты, и они хорошо известны специалистам в этой области. Они включают, но не ограничиваются ими: аденовирусные векторы, предпочтительно в форме аденовирусных частиц; ретровирусные векторы, предпочтительно в форме рекомбинантных ретровирусов; этот же тип векторов, но полученных из других вирусов; липосомы или другие молекулы переносчики, и т.д.
Изобретение также относится к диагностическому тестовому набору для осуществления способа по изобретению для определения опухолеродной способности агента, включающему нетрансформированную клетку, подвергнутую трансдукции нуклеиновой кислотой, кодирующей апоптин, или его функциональное производное, или его фрагмент и, необязательно, другие материалы, необходимые для проведения теста и определения результата.
Изобретение также касается диагностического тестового набора для осуществления способа по изобретению для определения склонности клеток к раковым заболеваниям, включающего нуклеиновую кислоту, кодирующую апоптин, или его функциональное производное, или его фрагмент, способные к трансдукции эукариотической клетки и способные экспрессироваться в такой клетке, и, необязательно, все другие материалы, необходимые для проведения теста и определения результата. Предпочтительно также обеспечивается средство, с помощью которого клетку подвергают воздействию мягкой опухолеродной активности, представляющей собой UV-излучение и рентгеновское излучение.
Изобретение также относится к способу изучения индукции апоптин-индуцированного апоптоза, которая приводит к ингибированию образования опухоли, с помощью вызывающих трансформацию агентов, таких как химические соединения, вирусы, UV- и рентгеновское излучение, на модели трансгенной мыши. Апоптин-трансгенные мыши могут быть использованы для изучения противоопухолевого эффекта апоптина в трансгенных химерах, несущих наследственные типы раковых заболеваний и способных к экспрессии апоптина. Более того, воздействие экспрессии апоптина на модели in vivo может изучаться с помощью описанных апоптин-трансгенных мышей.
Подробное описание изобретения.
Ранее было показано, что вирусный белок апоптин индуцирует апоптоз в культурах трансформированных клеток как человека, так и грызунов, но не в нормальных клетках человека. Также обнаружилось, что апоптин не может индуцировать апоптоз в культуре мышиных (или крысиных) эмбриональных фибробластов (культуры клеток были получены от 16-18-дневных мышиных (крысиных) эмбрионов). Это показывает, что апоптин также может экспрессироваться в здоровом эмбрионе, не проявляя токсических свойств, по крайней мере на не слишком ранней стадии развития эмбриона.
В настоящее время имеется возможность продуцировать жизнеспособных апоптин-трансгенных мышей. Мы сделали вывод о том, что нерегулируемая экспрессия апоптина в трансгенной мыши не приводит к летальному исходу или другим угрожающим жизни, сокращающим жизнь воздействиям. Также было обнаружено, что одновременная трансфекция культивируемых нормальных фибробластов человека геном апоптина и вызывающими трансформацию генами SV40 будет активировать процесс апоптоза, который сопровождается перемещением белка апоптина из цитоплазмы, где он вначале накапливается, внутрь ядра.
Описываемое изобретение обеспечивает основу для добавления аминокислот к апоптиновому белку или его фрагментам, давая им возможность проникать в клеточное ядро и/или накапливаться там, что в результате приводит к индукции апоптоза.
Изобретение обеспечивает основу для диагностического теста для определения потенциально вызывающих трансформацию генов. В этом тесте используются нормальные диплоидные клетки млекопитающих, такие как клетки человека и/или клетки грызунов. Для этого нормальные диплоидные клетки подвергают одновременной трансфекции плазмидой, содержащей ген(ы), которые должны быть изучены, и плазмидой, кодирующей апоптин, или подвергают трансфекции геном(ами), которые должны быть изучены, и инфицируют вирусным вектором, экспрессирующим апоптин. Индукция апоптин-индуцированного апоптоза и/или присутствие апоптина в ядре показывает, что изучаемый ген обладает трансформирующей/опухолеродной способностью.
Кроме того, было обнаружено, что диплоидные клетки человека, полученные от людей, которые несут мутацию гена-супрессора опухоли в зародышевой линии, и как результат предрасположены к развитию определенного спектра раковых заболеваний (это также относится к склонности к раковым заболеваниям), устойчивы к воздействию апоптина, индуцирующему апоптоз, так же как диплоидные клетки от здоровых индивидуумов, однако они становятся чувствительными к нему, если культуры клеток облучают ультрафиолетовым светом.
Это позволило нам разработать диагностическое исследование для определения склонности к раковым заболеваниям. В семьях с наследственной предрасположенностью к раковым заболеваниям из-за мутации зародышевой линии в гене-супрессоре опухоли часто бывает невозможным без обширного анализа хромосомной ДНК, предсказать, поражен ли член семьи или несет ген заболевания. Наши результаты показали, что это может быть определено простым способом, с использованием гена апоптина. Для этого диплоидные фибробласты кожи или лимфоциты отбирают у человека для тестирования и культивируемые клетки подвергают трансфекции геном апоптина, с последующим облучением с помощью UV-света (266 нм). Если подвергшиеся трансфекции клетки становятся апоптическими после UV-облучения, но не подвергаются апоптозу без UV-экспозиции, тогда это является серьезным подтверждением того, что человек имеет склонность к раковым заболеваниям. Предрасположенность еще не ко всем типам раковых заболеваний, которые появляются из-за мутации гена-супрессора опухоли, была протестирована с помощью апоптин/UV анализа. Тем не менее нет причины предполагать, что то же самое явление не будет наблюдаться в других склонных к раковым заболеваниям клетках. Аналогичный диагностический тест для предсказания склонности к раковым заболеваниям может быть выполнен с использованием обработки рентгеновскими лучами вместо UV-облучения.
Изобретение также позволяет получить много информации о молекулярных основах склонности к раковым заболеваниям и его взаимосвязи с определенными ответными реакциями на стресс, такими как Усиленная Реактивация (Enhanced Reactivation, ER) (Abrahams, P.J., Houweling, A., Cornelissen-Steijger, P.D. M., Arwert, F., Menko, F.H., Pinedo, H.M., Terleth, C., and Van der Eb, A.J. (1996). Наследование ненормальной экспрессии SOS-подобных реакций в Xeroderma Pigmentosum и наследственные синдромы склонности к раковым заболеваниям. Cancer Research, 56, 2621-2625). Усиленная Реактивация - это одна из реакций нормальных клеток (человека) на действие определенных ДНК-повреждающих агентов, которая, по-видимому, отражает клеточную восприимчивость к онкогенной трансформации.
Изобретение будет представлено в деталях в следующей экспериментальной части. Эта часть служит только для иллюстрации и не должна интерпретироваться как ограничивающая область изобретения.
Экспериментальная часть
Клетки и условия культивирования клеток
Эмбриональные фибробласты крысы (REF) были получены из 14-дневных эмбрионов. Клетки оттаивали из жидкого азота, культивировали в DMEM, с добавлением 10% фетальной телячьей сыворотки и подвергали трансфекции ДНК плазмидой при клеточном пассаже 2.
Эмбриональные фибробласты мыши (MEF) были получены из р53+/+ мышей или из р53-/- мышей, подвергнутых нокауту (Jacks, Т. (1994). Спектральный анализ опухолей у р53-мутантных мышей. Current Biology 4, 1-7 и Jacks, T. (1996) Уроки, полученные из р53 мутированных мышей. Journal of Cancer Research and Clininal Oncology 122, 19-27; Tyler Jacks et al., 1994). Клетки выращивали на Corning чашках в среде F15 с добавлением 10% фетальной телячьей сыворотки.
Р19 клетки получены из эмбриональной мышиной карциномы/тератокарциномы (McBurney, M. W. et al. (1982) Выделение мужских мышиных эмбриональных клеток карциномы и пути репликации их хромосом. Developments in Biology 89, 503-508). Клетки выращивали на желатинированных чашках Петри в DMEM с добавлением 8% фетальной телячьей сыворотки.
BRK/xho клетки были получены из почечных клеток детенышей крысы путем трансформации с использованием 5 El участка аденовирусного типа (Schrier, P. I. , Bernards, R., Vaessen, R. Т. M. J., Houweling, A. and Van der Eb, A. J. (1983). Экспрессия класс 1 главных антиген гистосовместимости прекращается при наличии онкогенного аденовируса 12 в трансформированных клетках крысы. Nature, 305, 771-775). Клетки культивировали в среде DMEM с добавлением 10% фетальной телячьей сыворотки.
Диплоидные фибробласты крайней плоти человека VH10 и VH25 (Klein, В., Pasting, A. , Odijk, H. , Westerveld, A. , and Van der Eb, A.J. (1990). Трансформация и иммортализация диплоидных Xeroderma pigmentosum фибробластов. Experimental Cellular Research 191, 256-262) выращивали в модифицированной Dulbecco, Eagle среде (DMEM) с добавлением 10% фетальной телячьей сыворотки.
Первоначально культуры эпидермальных кератиноцитов человека (FSK-1) начинали выращивать в полной среде, как это описано (Reinwald, J.G. and Green, H. (1975). Cell 6, 331-343) с незначительными модификациями согласно Ропес, М. , Kempenaar, J. A., and De Kloet, E.R. (1981). Кортикоиды и культивированные человеческие эпидермальные кератиноциты: специфическое внутриклеточное связывание и клиническая эффективность. Journal Investmental Dermatology 76, 211-214, и после этого культивировали в кератиноцитной среде, свободной от сыворотки (KSFM). Для описанных здесь экспериментов использовали число пассажей, равное 3.
F9605 клетки представляют собой диплоидные фибробласты, р16-/-, полученные от пациентов с синдромом диспластичного невуса (DNS), для которого установлено, что он является предшественником семейного атипичного множественного синдрома меланомы невуса (FAMMM) (Gruis, N.A. et al. (1995). Гомозиготы для CDKN2(P16) мутации зародышевой линии в семейной Голландской обычной меланоме. Nature Genetics, 10: 351-353, 1995). Клетки выращивали в DMEM с 10% фетальной телячьей сыворотки.
GM1492 клетки представляют собой диплоидные фибробласты человека, которые не экспрессируют р53, и получены от пациентов с синдромом Блума (Bloom's), аутосомальным рецессивным расстройством с высокой частотой раковых заболеваний (Van Laar Т. , Steegenga wt et al. (1994). В Bloom's Syndrome клетках GM1492 нет заметного количества белка р53, но в них происходит нормальная остановка G1 клеточного цикла после UV облучения. Oncogene. 9: 981-983). Клетки выращивали в DMEM, содержащей 10% фетальной телячьей сыворотки.
LF2675T представляют собой диплоидные фибробласты кожи от пациентов, с синдромом Ли-Фраумени (Li-Fraumeni) (LFS). Это заболевание характеризуется мутацией зародышевой линии в одном аллеле гена р53 и ранним началом различных типов раковых заболеваний (Srivastava, S. , Zou. Z., et al. (1990). Трансмиссия зародышевой линии мутированного гена р53 в семьях, имеющих склонность к раковым заболеваниям с синдромом Li-Fraumeni. Nature 348, 747-749; Abrahams, P. J., Houweling, A., Cornelissen-Steijger, P.D.M., Arwert, F. , Menko, F. H. , Pinedo, H.M., Terleth, C., and Van der Eb, A.J. (1996). Наследование ненормальной экспрессии SOS-подобных реакций в Xeroderma Pigmentosum и наследственные синдромы склонности к раковым заболеваниям. Cancer Research, 56, 2621-2625). Клетки выращивали в DMEM с добавлением 10% фетальной телячьей сыворотки.
Клетки представляют собой диплоидные фибробласты кожи от людей с семейством синдромов Линча (Lynch) типа 2 с высокой частотой различных типов раковых заболеваний. Клетки были получены от людей, умерших от рака грудной железы (Abrahams, P.J., Houweling, A., Cornelissen-Steijger, P.D.M., Arwert, F., Menko, F.H., Pinedo, H.M., Terleth, C., and Van der Eb, A.J. (1996). Наследование ненормальной экспрессии SOS-подобных реакций в Xeroderma Pigmentosum и наследственные синдромы склонности к раковым заболеваниям. Cancer Research, 56, 2621-2625). Клетки выращивали в DMEM с добавлением 10% фетальной телячьей сыворотки.
Все культуральные среды были получены от GIBCO/BRL и содержали антибиотики пенициллин и стрептомицин.
Облучение культур клеток
Кондиционированную среду удаляли из культур, и клетки дважды промывали PBS. После удаления PBS культуры облучали с помощью UV-света, как описано выше (Abrahams, P.J., Huitema, В.A., and Van der Eb, A.J. (1984) Усиленная реактивация и усиленный мутагенез герпес симплекс вируса в нормальных человеческих клетках и Xeroderma Pigmentosum клетках. Molecular Cellular Biology 4, 2341-2346), или обрабатывали рентгеновскими лучами (5 gray) с использованием Andrex 225 SMART (Andrex St, Copenhagen) при 200 KV, 4 mA с 1-мм А1 фильтром. Дозу и мощность дозы регулировали с помощью PTW дозиметра. После обработки UV-излучением кондиционированную среду возвращали назад и культуры инкубировали при 37oС.
Плазмиды
Экспрессирующая плазмида pCMV-VP3 содержит CAV ДНК последовательность, кодирующую только белок апоптин (nt 427-868; Noteborn, M.H.M., De Boer, G.P. , Van Roozelaar, D., Karreman, C., Kranenburg, 0., Vos, J., Jeurissen, S., Zantema, A. , Hoeben, R., Koch, G., Van Ormondt, H., and Van der Eb, A.J. (1991). Характеристика клонированного ДНК вируса анемии цыплят, которой содержит все элементы инфекции репликационного цикла. Journal of Virology 65, 3131-3139, Noteborn and De Boer, G.F. (1996). Patent USA/no. 030, 335), а плазмида pCMV-des кодирует десмин, структурный белок мышечных клеток (Menke, A. L, Shvarts, A., Riteco, N, Van Наm, R.C.A., Van der Eb, A J., and Jochemsen, A.G. (1997). Wilms' Tumor 1 (WT1) слипшиеся варианты без КТС индуцируют апоптоз в р53-негативных и р53-позитивных HepG2 клетках. Cancer Research 57, 1353-1363). Плазмида pCMV-neo используется как "пустой" негативный контроль для плазмид, кодирующих генные продукты с потенциальным воздействием на апоптин-индуцированный апоптоз. Все экспрессированые гены регулируются (ранним) усиленным промотором цитомегаловируса.
Плазмида SV40 содержит клон первоначально дефектного (ori) раннего (early-) участка SV40, включающий участки, кодирующие и большой Т-антиген и малый Т-антиген SV40, регулируемые их собственным промотором (Dinsart, С., Cornelis, J. J. , Klein, В., van der Eb, A. J., and Rommelaere, J. (1984). Трансфекция с внеклеточной UV-поврежденной ДНК стимулируют крысиные и человеческие клетки к экспрессии мутированого фенотипа к парвовирусу Н-1. Molecular Cellular Biology, 4, 324-). Плазмида pR-s884 экспрессирует полный большой Т-антиген SV40 и усеченный малый Т-антиген под транскрипциным контролем длинного концевого повтора (LTR) вируса саркомы Роуса (Rous) (RSV; De Ronde, A. , et al. (1989). Малый Т-антиген вируса SV40 необходим для морфологической трансформации человеческих фиробластов. Virology 171, 260-263; Smits, P. H. M. et al. (1992). Модуляция трансформации и транскрипции вируса папиломы человека типа 16 при делении положения короткой ветви человеческой хромосомы 11 может быть имитирована малым антигеном t SV40. Virology 190, 40-44). Плазмида PR-SVt содержит последовательности кДНК, кодирующие малый Т-ген SV40, который был объединен с RSV LTR (Philips, В., and Rundell, К. (1988). Неудача малого t антигена вируса SV40 в попытке дезорганизовать актиновые структуры в не пермисивных клеточных линиях. Journal of Virology 62, 768-775).
Экспрессирующая плазмида 21ЕсоА, которая состоит из участков гена/промотора мышиного Н-2КЬ антигена гистосовместимости (Mellor, A.L., Golden, L. , Weiss, E. , Bullman, H., Hurst, Simpson, E., James, R. F., Townsend, A. R., Taylor, P.M., Schmldt, W., Ferluga, J., Leben, L., Santamaria, M., Atfield, G. Festenstein, H., Flavell, R.A. (1982)) и рВг327 последовательностей, подарена Prof Dr Frank Grosveld, Erasmus Университет, Ротердам. Нидерланды. Плазмида 21ЕсоА содержит Notl внутри первого экзона Н-2КЬ гена, который делает возможной интеграцию чужеродного гена, который становится регулируемым промотором Н-2Кb. Фрагмент BamHl, содержащий последовательности, кодирующие апоптин, был выделен из pCMV-VP3 и клонирован в Notl сайте плазмиды 21ЕСоА с использованием линкеров Notl-BamHl. Финальная плазмида, содержащая ген апоптина, регулируемый Н-2Кb промотором, была названа p21EcoA-VP3. Впоследствии EcoRl сайт, следующий за геном апоптина, был делегирован путем линеаризации плазмиды p21EcoA-VP3 по этому специфичному EcoRl сайту и обработки полимеразой Кленова. Плазмида названа p21EcoA-Vp3-Eco.
Фрагмент ДНК, содержащий Н-2Кb экспрессирующую единицу с геном апоптина, был отделен от последовательностей прокариотической ДНК посредством гидролиза EcoRl и электрофореза на агарозном геле.
ДНК плазмиду очищали при помощи центрифугирования в градиенте CsCl и колоночной хроматографии на Сефакриле (Sephacryl) S500 (Pharmacia, Швеция).
Транзитная Трансфекция.
Клетки были подвергнуты трансфекции в монослойных культурах агентом трансфекции DOTAP (Boehringer, Mannheim, FRG) по существу так же, как описано у Fischer, D.F., Gibbs, S., Van de Putte, P., and Backendorf, C. (1996). Внутризависимые элементы контроля транскрипции регулируют экспрессию гена SPRR2A в процессе терминальной дифференциации кератиноцитов (Molecular and Cellular Biology 16, 5365-5374) или их подвергали трансфекции с помощью ДНК плазмиды осаждением фосфата кальция, как описано у Graham, F.L. and Van der Eb, A.J. (1973). Новая методика анализа инфекционности человеческого аденовируса 5 DNA. Virology 52, 456-467.
Непрямая иммунофлуоресценция
Все клетки были выращены на предметных стеклах для микроскопа. Предметные стекла были либо без покрытия (VH10, VH25), либо с покрытием из 3-аминопропилтриэтоксисилана (TESPA; FSK-1). Клетки фиксировали 80% ацетоном в течение 10 мин при комнатной температуре и использовали для непрямой иммунофлуоресценции, как описано ранее (Van den Heuvel, S.J.L., Van Laar, Т., Kast, W.M., Melief, C.J., Zantema, A., and Van der Eb, a.]. (1990). Ассоциация между клеточным р53 и аденовирусным 5 Е1В-55 кд белками снижает онкогенность Ad-трансформированных клеток. EMBO Journal 9, 2621-2629). Для демонстрации присутствия и/или клеточной локализации апоптина в клетках, подвергнутых трансфекции, использовали мышиное моноклональное антитело (Mab) CV1-CAV-85.1 (85.1; Noteborn, M. H. M., De Boer, G.P., Van Roozelaar, D., Karreman, C., Kranenburg, О. , Vos, J., Jeurissen, S., Zantema, A., Hoeben, R., Koch, G., Van Ormondt, H., and Van der Eb, A.J. (1991). Характеристика клонированного ДНК вируса анемии цыплят, которой содержит все элементы инфекции репликационного цикла. Journal of Virology 65, 3131-3139); для человеческого десмина мышиное Mab 33 (Monosan, Uden, Нидерланды); для Т-антигенов SV40 - Pab 419, любезно предоставленное Dr A.-G Joshemsen из университета Лейдена, Нидерланды. В качестве второго антитела было использовано меченое изотиоцианатом флуоресцеина козлиное анти-мьшиное антитело (Jackson Immunoresearch Laboratories Inc., West grov PA, USA). Ядерную ДНК окрашивали 2,4-диамино-2-фенилиндолом (DAPI).
Генерация апоптин-трансгенных мышей
Для генерации апоптин-трансгенных мышей были использованы оплодотворенные ооциты из FVB линии мышей и мыши из мышиных линий в качестве fosters. Микроинъекции в мужские пронуклеусы осуществляли согласно Brinster, R.L., Chen, H. Y. , and Trumbauer, M.E. (1981). Мышиные ооциты транскрибируют инъецированный Xenopus 5S RNA ген. Science 211, 396-398. На одну микроинъекцию вводили 500 копий фрагмента ДНК EcoRl, полученного из плазмиды p21EcoA-Vp3-Есо, содержащего требуемую Н-2Кb транскрипционную единицу и полный ген апоптина.
Результаты и обсуждение.
Апоптин индуцирует апоптоз в трансформированных клетках грызунов, но не индуцирует его в нормальных эмбриональных клетках.
Чтобы исследовать, будет ли апоптин индуцировать апоптоз в нормальных эмбриональных клетках грызунов, культуры клеток зародышей мышей и клеток зародышей крыс были подвергнуты транзитной трансфекции плазмидой, кодирующей апоптин. В качестве отрицательного контроля клетки были подвергнуты трансфекции плазмидой, кодирующей десмин, который не обладает апоптической активностью. Клетки, экспрессирующие апоптин, были отобраны путем непрямой иммунофлуоресценции с использованием Маb 85.1, а клетки, экспрессирующие десмин, - с использованием мышиного Маb 33. Индукцию апоптоза в апоптин- или десмин-позитивных клетках анализировали при помощи DAPI, который вызывает равномерное окрашивание неповрежденных ядер и неравномерное и/или слабое окрашивание апоптических ядер.
Через пять дней после трансфекции примерно 10-20% десмин-позитивных клеток стали апоптическими, что является базовым уровнем, скорее всего обусловленным результатом трансфекции (данные не показаны, Menke, A.L., Shvarts, A. , Riteco, N. , Van Ham, R.C.A., Van der Eb, A. J., and Jochemsen, A.G. (1997). Wilms' Tumor 1 (WT1) слипшиеся варианты без КТС индуцируют апоптоз в р53-негативных и р53-позитивных HepG2 клетках. Cancer Research 57, 1353-1363, Danen-van Oorschot, A.A.A.M., Fischer, D., Grimbergen, J.M., Klein, В. , Zhuang, S.-M., Falkenburg, J.H.F., Backendorf, C., Quax, P.H.A., Van der Eb, A. J. , and Noteborn, M.H.M. (1997). Апоптин индуцирует апоптоз в человеческих трансформированных или злокачественных клетках, но не в нормальных клетках. Proceedings National Academy Sciences, USA: 94, 5843-5847). Через 2, 3, 4 или 5 дней после трансфекции процентное содержание апоптических апоптин-позитивных клеток не превышало в значительной степени процентное содержание апоптических клеток, наблюдавшееся в десмин-позитивных культурах, что указывает на то, что апоптин не индуцирует апоптоз в культурах нормальных эмбриональных клеток. Транзитная трансфекция трансформированных эмбриональных клеток мышей/крыс или почечных клеток детенышей крыс плазмидой, кодирующей апоптин, подтвердила, что апоптин способен индуцировать апоптоз в этих клетках. Результаты экспрессии апоптина в "нормальных" эмбриональных клетках грызунов в сравнении с трансформированными клетками грызунов показаны на фиг. 1.
Эти данные показывают, что апоптин не может вызывать апоптоз в нормальных эмбриональных клетках грызунов и клетках взрослых грызунов, но индуцирует апоптоз в клетках, полученных при помощи вирусной трансформации, по крайней мере в условиях культивирования клеток.
Одновременная экспрессия большого Т-антигена SV40 и апоптина приводит к апоптин-индуцированному апоптозу в нормальных диплоидных клетках человека.
Мы исследовали воздействие экспрессии генов, вызывающих трансформацию, на апоптин-индуцированный апоптоз в нормальных клетках человека, полученных от здоровых людей. Для этого диплоидные фибробласты VH10 и диплоидные кератиноциты FSK-1 человека были подвергнуты одновременной транзитной трансфекции плазмидой pCMV-VP3, кодирующей апоптин, и либо плазмидой pSV40, кодирующей и большой и малый Т-антигены, pR-s884, кодирующей большой Т-антиген, pR-SVt, кодирующей малый Т-антиген, либо плазмидой отрицательного контроля pCMV-neo. Клетки были проанализированы путем непрямой иммунофлуоресценции на наличие апоптин-индуцированного апоптоза. Как нормальные клетки VH10, так и клетки FSK-1 не подвергались апоптозу, когда апоптин вводили путем трансфекции контрольной плазмидой.
Как и ожидалось, результаты показали, что сама по себе экспрессия апоптина не может индуцировать апоптоз в нормальных диплоидных клетках человека, что подтверждает данные, приведенные Danen-van Oorschot, A.A.A.M., Den Hollander, A., Tak-ayama, S., Reed, J., Van der Eb, A.J., and Noteborn, M. H. M. (1997). BAG-1 ингибирует р53-индуцированный, но не апоптин-индуцированный апоптоз. Apoptosis, In Press. Однако нормальные диплоидные фибробласты и кератиноциты человека, в которых экспрессируется и апоптин и большой Т-антиген SV40, один или вместе с малым Т-антигеном, подвергаются апоптин-индуцированному апоптозу (фиг. 1). Скорость индукции апоптоза значительно увеличивалась в присутствии вирусных генов, вызывающих трансформацию. Одновременная экспрессия малого Т-антигена SV40 и апоптина не вызывала индукцию апоптоза апоптином. Переход нормальных клеток из состояния устойчивости к действию апоптина в состояние восприимчивости к действию апоптина, вероятно, может объяснить тот факт, что белок апоптин перемещается из цитоплазмы внутрь ядра. Это перемещение становится заметным уже приблизительно через 2 дня после трансфекции плазмидами SV40 (фиг. 3). Можно сделать вывод, что в данном примере имеет место какое-то событие, произошедшее благодаря экспрессии продукта из ДНК-вируса опухоли, вызывающего трансформацию, в результате которого происходит перемещение апоптина из цитоплазмы внутрь ядра, сопровождающееся индукцией апоптоза.
Одновременная экспрессия большого Т-антигена SV40 и апоптина приводит к апоптин-индуцированному апоптозу в нормальных диплоидных клетках грызунов.
Далее, мы исследовали воздействие одновременной экспрессии генов, вызывающих трансформацию, и апоптина на индукцию апоптоза в нормальных фибробластах эмбрионов мышей (MEF-клетках), полученных от р53+/+ мышей или от трансгенных р53-/- мышей. Оба вида подвергшихся транзитной трансфекции MEF-клеток, в которых одновременно экспрессировался вызывающий трансформацию ген большого Т-антигена SV40 вместе с малым Т-антигеном или без него в сочетании с апоптином, подвергались очень быстрому апоптозу, тогда как в MEF-клетках, в которых апоптин экспрессировался вместе с контрольной плазмидой или с плазмидой, кодирующей не трансформирующий малый Т-антиген, не происходил апоптин-индуцированный апоптоз. Результаты показаны на фиг. 4.
Иммунофлуоресцентный анализ также показал, что одновременная экспрессия апоптина и большого Т-антигена SV40 также приводит к внтриклеточному перемещению апоптина. В исследуемых MEF-клетках апоптин находится в цитоплазме. При экспрессии большого Т-антигена апоптин проникает внутрь ядра, что сопровождается индукцией апоптоза. Для сравнения на фиг. 5 также приведено процентное содержание апоптин-позитивных трансформированных клеток мышей.
Эти результаты показывают, что апоптин не индуцирует апоптоз в фибробластах как р53+/+, так и р53-/- мышей, но делает это при экспрессии белка, вызывающего трансформацию. Эта информация является важной, поскольку известно, что "нормальные" р53-/- клетки очень восприимчивы к самопроизвольной трансформации и легко превращаются в клетки с фенотипом, более подверженным трансформации. Однако утраты одного р53 недостаточно для создания свойства трансформируемости. Кроме того, этот вывод показывает, что апоптин может индуцировать апоптоз при экспрессии белка, вызывающего трансформацию, в других клетках млекопитающих помимо клеток человека.
Одновременная экспрессия большого Т-антигена SV40 и апоптина приводит к апоптин-индуцированному апоптозу в нормальных диплоидных клетках, полученных от людей, имеющих склонность к раковым заболеваниям.
При помощи транзитной трансфекции и иммунофлуоресценции мы также исследовали воздействие апоптина на нормальные фибробласты F9605 и GM1492, которые были получены от людей, у которых из-за генетических нарушений наблюдалась повышенная частота раковых заболеваний. Апоптин не может индуцировать апоптоз в нормальных диплоидных клетках этих людей, имеющих склонность к раковым заболеваниям. Однако при экспрессии большого Т-антигена SV40 апоптин индуцирует апоптоз (фиг. 6) после проникновения внутрь ядра (данные не показаны).
Эти данные подтверждают, что диплоидные клетки с наследственными синдромами склонности к раковым заболеваниям не восприимчивы к действию апоптина, однако они становятся таковыми, когда в них экспрессируется ген, вызывающий трансформацию. Таким образом для данного испытания диплоидные клетки с такими наследственными синдромами идентичны "нормальным" диплоидным клеткам человека.
Воздействие на индукцию апоптоза ковалентного связывания сигнального пептида, определяющего ядерную локализацию, большого Т-антигена SV40 и белка апоптина.
Далее, мы исследовали, приведет ли экспрессия химерного белка, состоящего из апоптина и сигнального пептида, определяющего ядерную локализацию, большого Т-антигена SV40 (аминокислоты N-Пролин-Пролин-Лизин-Лизин-Лизин-Аргинин-Лизин-Валин-С большого Т-антигена SV40 ковалентно связаны с N-концом апоптина), к индукции апоптоза в нетрансформированных и трансформированных клетках человека. Химерный белок называется NLS-апоптин. Для этого не трансформированные фибробласты человека VH10 и трансформированные, выделенные из остеосаркомы, клетки человека Saos-2 (Danen-van Oorschot, A.A.A.M., Den Hollander, A., Tak-ayama, S., Reed, J., Van der Eb, A.J., and Noteborn, M.H.M. (1997). BAG-1 ингибирует р53- индуцированный, но не апоптин-индуцированный апоптоз. Apoptosis, In Press), были подвергнуты трансфекции плазмидой, кодирующей химерный белок NLS-апоптин. В обоих видах клеток экспрессия NLS-апоптина привела к ядерной локализации апоптина и индукции апоптоза. Экспрессия неапоптического белка, такого как Зеленый Флуоресцентный Белок (GFP, Pines, J. (1995). GFP в клетках млекопитающих. Trends in Genetics 11, 326-327), ковалентно присоединенного к NLS, который проникал внутрь ядра, не приводила к индукции апоптоза.
Эти данные подтверждают, что модифицированный апоптин с задействованной ядерной локализацией, происходящей в независимости от трансформации клеток, способен к перемещению внутрь ядра, сопровождающемуся индукцией апоптоза.
Продукт конденсации первых 69 аминокислот N-конца апоптина и неапоптического GFP-белка не приводит к индукции апоптоза, и это согласуется с тем фактом, что этот химерный белок не проникает внутрь ядра (Noteborn, M.H.M. and Pietersen, A. M. (1998). РСТ заявка 98/00213, названная Аденовирусный вектор). Теперь мы ковалентно связали 8 аминокислот NLS с фрагментом N-конца апоптина, состоящего из 1-69 аминокислот (NLS-aпоптин/1-69). Трансфекция как нетрансформированных клеток VH10, так и опухолеродных клеток человека (таких, как выделенные из остеосаркомы клетки человека Saos-2) плазмидой, кодирующей NLS-апоптин/1-69, привела к ядерной локализации NLS-апоптина/1-69, сопровождающейся индукцией апоптоза.
Эти данные указывают на то, что кроме С-концевой части апоптина, N-концевая часть (1-69 аминокислот) также действует подобным образом, когда она перемещается внутрь ядра. В этих экспериментах, как и ожидалось, NLS-GFP-белок перемещался внутрь ядра, но это не приводило к индукции апоптоза.
Нормальные диплоидные клетки, полученные от людей, имеющих склонность к раковым заболеваниям, подвергаются апоптин-индуцированному апоптозу после UV-облучения.
Мы также исследовали воздействие UV-излучения на индукцию апоптоза апоптином в диплоидных клетках. Диплоидные фибробласты, полученные от здоровых людей (VH25) или от людей с синдромом склонности к раковым заболеваниям (клетки LF2675T от пациента с синдромом Li Fraumeni и клетки 401 от пациента с синдромом Lynch типа 2) были подвергнуты транзитной трансфекции плазмидой, кодирующей апоптин. Перед трансфекцией часть клеток подвергли UV-облучению. В качестве отрицательного контроля клетки были подвергнуты трансфекции плазмидой, кодирующей белок десмин.
Во всех трех видах клеток: VH25, LF2675T и 401, без UV-облучения не проявлялись признаки апоптин-индуцированного апоптоза. Однако в сочетании с UV-облучением клетки LF2675T и 401, но не клетки VH25, очень быстро подвергались апоптин-индуцированному апоптозу. Хотя у нас нет объяснения для этого явления, по-видимому, оно связано с другим клеточным свойством. Диплоидные клетки от пациентов, имеющих склонность к раковым заболеваниям из-за мутации гена-супрессора опухоли зародышевой линии, проявляют неожиданную реакцию на UV-излучение. Когда нормальные диплоидные фибробласты обрабатывают UV-излучением или другим реагентом, наносящим повреждение ДНК, это вызывает у них большое количество разнообразных скоротечных ответных реакций, включая активацию сигнальных путей трансдукции, индукцию экспрессии различных генов, ингибирование репликации клеточной ДНК и активацию SOS-подобных явлений, таких как усиленная реактивация (ER) и усиленный мутагенез (ЕМ). Abrahams, P. J. , Houweling, A., Cornelissen-Steijger, P.D.M., Arwert, F., Menko, F.H., Pinedo, H.M., Terleth, C., and Van der Eb, A.J. (1996). Наследование ненормальной экспрессии SOS-подобных реакций в Xeroderma Pigmentosum и наследственные синдромы склонности к раковым заболеваниям. Cancer Research, 56, 2621-2625 обнаружили, что нормальные диплоидные фибробласты от пациентов с наследственной предрасположенностью к раковым заболеваниям из-за мутации зародышевой линии гена-супрессора опухоли, показывают такие же ответные реакции на UV-излучение, как клетки от нормальных людей, кроме одной ответной реакции: усиленной реактивации. Ответная ER реакция в клетках от этих пациентов намного сильнее, чем в клетках от нормальных людей, поэтому клетки этих пациентов называются Еrсупер (+). Молекулярно-биологическое обоснование явления ER все еще не выяснено. Для обнаружения ER используется подход, требующий временных затрат, поскольку он основан на оценке (повышенного) выживания вируса, подвергшегося действию UV-излучению, в клетках, поврежденных UV- излучением (или рентгеновским излучением), по сравнению с выживанием в неповрежденных клетках. Испытание, основанное на апоптин-индуцированном апоптозе при UV-облучении, проводится значительно проще и быстрее (см. ниже). Тот факт, что апоптин становится активным в клетках, имеющих склонность к раковым заболеваниям, при UV-облучении, также дает возможность изучения процесса ER. Есть данные, указывающие на то, что ER играет важную роль в процессе индукции раковых заболеваний реагентами, повреждающими ДНК.
Нормальные диплоидные клетки, полученные от людей, имеющих склонность к раковым заболеваниям, подвергаются апоптин-индуцированному апоптозу после обработки рентгеновским излучением.
Далее, мы также исследовали воздействие обработки рентгеновским излучением на индукцию апоптоза апоптином в диплоидных клетках человека. Диплоидные фибробласты, полученные от здоровых людей (VH10) или от людей с синдромом склонности к раковым заболеваниям, такие как клетки LF2675 и 401, были подвергнуты трансфекции плазмидой, кодирующей апоптин. Перед трансфекцией часть клеток была обработана рентгеновским излучением (доза 5 Гр). В качестве отрицательного контроля клетки были подвергнуты трансфекции плазмидой, кодирующей белок десмин.
Как и ожидалось, во всех проанализированных необлученных клетках клеточных линий VH10, LF2675 и 401 не было заметно признаков апоптин-индуцированного апоптоза. Однако в сочетании с обработкой рентгеновским излучением клеточные линии, полученные от людей, имеющих склонность к раковым заболеваниям, подвергались апоптозу, а клетки, полученные от здоровых людей, - нет. Через пять дней после трансфекции большинство этих обработанных рентгеновским излучением апоптин-позитивных клеток, имеющих склонность к раковым заболеваниям, стали апоптическими. Клетки, обработанные рентгеновским излучением и экспрессируюшие неапоптический агент десмин, не подвергались апоптин-индуцированному апоптозу.
Эти результаты означают, что обработка рентгеновским излучением, вызывающая повреждение ДНК, так же, как описанная выше обработка UV-излучением, приводит к индукции апоптоза апоптином в нормальных нетрансформированных клетках человека.
Диагностическое исследование наличия генов, вызывающих раковые заболевания, основанное на апоптин-индуцированном апоптозе.
Danen-van Oorschot, A.A.A.M., Fischer, D., Grimbergen, J.M., Klein, В., Zhuang, S.-M., Falkenburg, J.H.F., Backendorf, C., Quax, P.H.A., Van der Eb, A.J., and Noteborn, M.H.M. (1997). Апоптин индуцирует апоптоз в человеческих трансформированных или злокачественных клетках, но не в нормальных клетках. Proceedings National Academy Sciences, USA:94, 5843-5847 сделали сообщение о том, что апоптин имеет различную клеточную локализацию в опухолеродных/ трансформированных клетках человека по сравнению с локализацией в нормальных нетрансформированных клетках. Более того, накопление апоптина в ядре связано с индукцией апоптоза, тогда как его локализация в цитоплазме связана с жизнеспособностью клеток и нормальной способностью к пролиферации.
На основании этого сообщения нам удалось разработать диагностическое исследование для выявления агентов и генов, вызывающих раковые заболевания и/или трансформацию. Первый вид исследования состоит в том, что "нормальные" клетки человека или грызунов подвергают трансфекции плазмидой, кодирующей апоптин, или инфицируют клетки вирусными векторами, экспрессирующими апоптин, вместе с плазмидой, кодирующей ген, предположительно вызывающий трансформацию/раковые заболевания. Впоследствии клетки анализируют на предмет (1) способности подвергаться апоптозу под действием гена апоптина и (2) изменения локализации апоптина из цитоплазмы в ядро.
Внутриклеточная локализация апоптина может быть определена с использованием иммунофлуоресцентного анализа при помощи моноклональных антител, специфичных к апоптину, таких как CVI-CAV-85.1. Если процентное содержание подвергшихся апоптозу нормальных клеток, одновременно экспрессирующих апоптин и ген, предположительно вызывающий трансформацию/раковые заболевания, значительно превышает процентное содержание подвергшихся апоптозу апоптин-позитивных контрольных клеток, экспрессирующих контрольную плазмиду, можно сделать вывод, что анализируемый ген действительно обладает вызывающим трансформацию/раковые заболевания действием.
Второй пример диагностической проверки основан на обработке культуры нормальных диплоидных клеток предполагаемым канцерогенным агентом. Агент может быть добавлен, например, в среду для культивирования на различные промежутки времени. Впоследствии клетки подвергают трансфекции плазмидой, кодирующей апоптин, или инфицируют вирусным вектором, экспресирующим апоптин. Этот подход также может быть осуществлен таким образом, что сначала проводят трансфекцию/инфицируют нормальные клетки, а затем их обрабатывают проверяемым агентом. Последующие стадии анализа такие же, как описаны для первого вида диагностического исследования.
Диагностическое исследование склонности к раковым заболеваниям.
Данные, представленные в этом сообщении, позволили нам разработать исследование для определения, имеет ли человек с неизвестной клеточной/генетической предысторией склонность к раковым заболеваниям по сравнению с нормальными здоровыми людьми. Нормальные диплоидные клетки от людей, имеющих склонность к раковым заболеваниям, не чувствительны к апоптин-индуцированному апоптозу, но становятся таковыми после обработки UV- или рентгеновским излучением или другим агентом, повреждающим ДНК. Ниже описывается пример такого диагностического исследования, основанного на воздействии UV-излучения. Это исследование можно проводить также в отношении других мутагенных/канцерогенных агентов.
Сначала нормальные диплоидные клетки выделяют из биопсии кожи человека, для того, чтобы их проанализировать, и культивируют в подходящей среде. Затем клетки облучают UV-излучением и впоследствии подвергают трансфекции плазмидой, кодирующей апоптин, или инфицируют вирусным вектором, экспрессирующим апоптин, либо клетки сначала подвергают трансфекции/инфицируют, а потом облучают. Параллельно в качестве контроля будут использованы диплоидные клетки от нормального здорового человека.
При помощи непрямого иммунофлуоресцентного анализа, основанного на применении апоптин-специфичных моноклональных антител, клетки анализируют на присутствие апоптина в ядре и/или на протекание апоптоза. Если процентное содержание клеток, подвергшихся апоптозу, среди апоптин-позитивных UV-обработанных клеток значительно превышает процентное содержание подвергшихся апоптозу UV-обработанных клеток нормального здорового человека, это будет определенным подтверждением того, что человек, чьи клетки были выделены, имеет склонность к раковым заболеваниям.
Использование апоптиновых белков в фармацевтических композициях для противораковой терапии.
На основании вышеупомянутых результатов можно также разработать способы применения апоптина для противораковой терапии не в виде гена (ДНК), а в виде белка. Апоптин является сравнительно небольшим белком, что дает возможность вводить его в клетку в виде белка (Если фрагменты белка апоптина также будут оказывать необходимое воздействие на раковые клетки, мы будем использовать фрагменты вместо целого белка). Нашей задачей является разработка эффективных фармацевтических композиций, которые обеспечивают устойчивость активного компонента (= апоптина или его фрагмента) и, по возможности, специфичность по отношению к целевым опухолевым клеткам. Новообразования, которые мы надеемся лечить при помощи подходящих апоптин-содержащих композиций, как лечебных, так и профилактических, включают наследственные формы колоректальных раковых заболеваний (врожденный семейный аденоматозный полипоз (АРС) и наследственное неполипозное колоректальное раковое заболевание (HNPCC)), раковое заболевание печени (или других органов, которые можно лечить с использованием перфузионной техники), лейкозы и лимфомы (для лечения с использованием циркуляции крови), опухоли кожи и, возможно, опухоли легких (через дыхательные пути).
Конструирование и анализ экспрессирующей плазмиды для генерации апоптин-трансгенных мышей.
Тот факт, который мы сейчас рассмотрели, свидетельствующий, что апоптин не может индуцировать апоптоз в культивируемых мышиных эмбриональных фибробластах, позволил нам сделать вывод о том, что апоптин также может экспрессироваться здоровыми эмбрионами или взрослыми мышами, не проявляя токсичных свойств, или по крайней мере зародышами на не слишком ранней стадии эмбрионального развития. Мы выбрали экспрессионную систему на основе мышиной Н-2Кb транскрипционной единицы, которая позволяет осуществлять нерегулируемую экспрессию чужеродных генов в течение эмбриогенеза и на взрослых стадиях в различных органах (Drezen, J.M., Nouvel, P., Babinet, С., and Morello, D. (1992). Различная регуляция экспрессии гена класса 1 у взрослых мышей и в течение их развития. The Journal of Immunology 149, 429-437; Morello, D., Moore, G., Salmon, A. M., Yaniv, M., and Babinet, C. (1986). Изучение экспрессии слившегося гена Н-2К/человеческого гормона роста в гигантских трансгенных мышах. The EMBO Journal 5, 1877-1883.
Таким образом мы сконструировали экспрессирующую плазмиду p21EcoA-Vp3-Eco, которая экспрессирует апоптин под влиянием регуляции со стороны мышиного Н-2Кb промотора. Кроме этого, экспрессирующий вектор содержит другие элементы Н-2КЬ, которые позволят осуществить экспрессию гена апоптина. На фиг. 8 показано схематичное представление апоптин-трансгенного экскспрессирующего вектора p21EcoA-Vp3-Eco.
При помощи транзитной трансфекции трансформированных Saos-2 клеток плазмидой p21EcoA-Vp3-Eco нам удалось подтвердить, что апоптин действительно может экспрессироваться в рамках последовательностей Н-2Кb. Кроме этого, экспрессированный апоптин приводил к индукции апоптоза в такой же степени, как апоптин, экспрессированный при помощи плазмиды pCMV-VP3(см. фиг. 9).
Эти результаты означают, что используемый экспрессирующий вектор p21EcoA-Vp3-Eco экспрессирует апоптин таким образом, что трансформированные клетки будут подвергаться апоптозу.
Генерация апоптин-трансгенных мышей.
В общей сложности в 300 оплодотворенных ооцитов при помощи микроинъекции был введен фрагмент ДНК, включающий Н-2Кb транскрипционную единицу и ген апоптина, они затем были пересажены 11 foster мышам. Мы получили потомство в общей сложности 51 новорожденных мышей.
При помощи саузерн-блот анализа (Southern, E.M. (1975). Определение специфических последовательностей среди фрагментов ДНК, разделенных посредством гель-электрофореза. Journal of Molecular Biology 98, 503-517) ДНК из хвостов мышей, гидролизованной BamHl- или Xbai-, с использованием меченого Р фрагмента ДНК, состоящего из полной копии гена VP3, было показано, что апоптин/Н-2Кb единица вошла в состав геномной ДНК в общей сложности 7 (FO) мышей-основателей. Все апоптин-трансгенные мыши выглядели здоровыми. Однако по неизвестным причинам одна апоптин-трансгенная мышь умерла в возрасте 5-6 недель. Апоптин-трансгенные мыши спаривались с FVB самцами или самками. ДНК из хвостов потомства анализировали на присутствие гена апоптина с использованием полимеразной цепной реакции (PCR), используя праймеры Р1 (5'-СТСТССААСААСАТАСТ-CCACCCGG-3') и Р2 (5'-CTTATACGCCTTTTT-GCGGTTCGGG-3'). От всех F0-мышей мы получили 1 или более апоптин-трансгенных F1-мышей (фиг. 10).
Все мыши из F1-генерации апоптин-трансгенных особей оказались жизнеспособными и в дальнейшем не проявили никаких патологических изменений, которые могли бы быть связаны с экспрессией апоптина.
При помощи нозерн-блот анализа (Noteborn, M.H.M., Kranenburg, О., Zantema, A., Koch, G., De Boer, G.F, and Van der Eb, A.J. (1992). Транскрипция генома вируса анемии цыплят (CAV) и синтез его 52 кд белка. Gene 118, 267-271) может быть определено наличие экспрессии гена апоптина в различных органах.
Описание чертежей.
На фиг.1 показано действие апоптин-индуцированного апоптоза в "нормальных" фибробластах зародышей грызунов в сравнении с клетками трансформированных клеточных линий грызунов. Клетки были подвергнуты транзитной трансфекции pCMV-VP3. Впоследствии клетки были зафиксированы через несколько промежутков времени после трансфекции и проанализированы путем непрямой иммунофлуоресценции с использованием апоптин-специфичного Mab 85.1. Процентное содержание апоптин-позитивных клеток, которые неправильно окрашиваются DAPI, приводится в качестве относительной меры индукции апоптоза.
На фиг.2 показано воздействие большого Т-антигена SV40 и/или малого Т-антигена на апоптин-индуцированный апоптоз в фибробластах и кератиноцитах от нормальных людей. Клетки VH10 и FSK-1 были подвергнуты транзитной трансфекции плазмидой pCMV-VP3 и pCMV-neo или плазмидой pSV40, экспрессирующей большой и малый Т-антигены SV40, pR-s884, экспрессирующей большой Т-антиген SV40, и pR-SVt, экспрессирующей малый Т-антиген SV40. Впоследствии клетки были зафиксированы через несколько промежутков времени после трансфекции и проанализированы путем непрямой иммунофлуоресценции с использованием апоптин-специфичного Мао 85.1. Процентное содержание апоптин-позитивных клеток, которые неправильно окрашиваются DAPI, приводится в качестве относительной меры апоптоза.
На фиг.3 показано расположение апоптина в нормальных диплоидных клетках человека, экспрессирующих только апоптин или апоптин вместе с большим и/или малым Т-антигеном SV40. Те же клетки, что были проанализированы на фиг.2 в отношении индукции апоптоза, были также исследованы на предмет расположения апоптина в ядре или в цитоплазме. Процентное содержание апоптин-позитивных клеток, содержащих апоптин в ядре и все еще не подвергшихся апоптозу, приводится в качестве относительной меры локализации апоптина в ядре.
На фиг.4 показано воздействие большого Т-антигена SV40 и/или малого Т-антигена на апоптин-индуцированный апоптоз в фибробластах мышей, которые получены от нормальной р53+/+ мыши или от трансгенной р53-/- подвергнутой нокауту мыши. Клетки были подвергнуты транзитной трансфекции плазмидой pCMV-VP3, экспрессирующей апоптин, и контрольной плазмидой pCMV-neo или pSV40, экспрессирующей большой Т-антиген SV40, pR-s884, экспрессирующей большой Т-антиген SV40, и pR-SVt, кодирующей малый Т-антиген. Впоследствии клетки были зафиксированы через несколько промежутков времени после трансфекции и проанализированы путем непрямой иммунофлуоресценции с использованием апоптин-специфичного Маb 85.1. Процентное содержание апоптин-позитивных клеток, которые неправильно окрашиваются DAPI, приводится в качестве относительной меры апоптоза.
На фиг.5 показано расположение апоптина в эмбриональных фибробластах мышей р53+/+ или р53-/-, экспрессирующих только апоптин или апоптин вместе с большим и/или малым Т-антигеном SV40. Те же клетки, что были проанализированы на фиг.4 в отношении индукции апоптоза, теперь были также исследованы на предмет расположения апоптина в ядре или в цитоплазме. Процентное содержание апоптин-позитивных клеток, содержащих апоптин в ядре и все еще являющихся апоптическими, приводится в качестве относительной меры локализации апоптина в ядре.
На фиг.6 показано воздействие большого Т-антигена SV40 на действие апоптин-индуцированного апоптоза в "нормальных " диплоидных фибробластах человека 9650 или G4905, полученных от людей, имеющих склонность к раковым заболеваниям. Клетки были подвергнуты транзитной трансфекции pCMV-VP3 и pCMV-neo или pSV40, экспрессирующей и большой и малый Т-антигены SV40, pR-s884, экспрессирующей большой Т-антиген SV40, и pR-SVt, экспрессирующей малый Т-антиген. Впоследствии клетки были зафиксированы через несколько промежутков времени после трансфекции и проанализированы путем непрямой иммунофлуоресценции с использованием апоптин-специфичного Mab 85.1. Процентное содержание апоптин-позитивных клеток, которые неправильно окрашиваются DAPI, приводится в качестве относительной меры апоптоза.
На фиг. 7 показано воздействие UV-излучения на апоптин-индуцированный апоптоз в "нормальных" диплоидных фибробластах, полученных от нормальных здоровых людей в сравнении с клетками, полученными от пациентов, имеющих склонность к раковым заболеваниям. Клетки были квази-обработаны или обработаны UV светом и впоследствии подвергнуты транзитной трансфекции pCMV-VP3 или pCMV-des. Наконец клетки были зафиксированы через несколько промежутков времени после трансфекции и проанализированы путем непрямой иммунофлуоресценции с использованием апоптин-специфичного Mab 85.1. Процентное содержание апоптин-позитивных клеток, которые неправильно окрашиваются DAPI, приводится в качестве относительной меры апоптоза.
На фиг. 8 показано схематичное представление апоптин-трансгенного экспрессионного вектора.
На фиг. 9 показано действие апоптин-индуцированного апоптоза в клетках Saos-2. Клетки были подвергнуты транзитной трансфекции p21EcoA-Vp3-Eco, pCMV-VP3 (обе плазмиды экспрессируют апоптин) или pCMV-des, экспрессирующей неапоптический белок десмин. Впоследствии клетки были зафиксированы через несколько промежутков времени после трансфекции и проанализированы путем непрямой иммунофлуоресценции с использованием апоптин-специфичного Mab 85.1. Процентное содержание апоптин-позитивных клеток, которые неправильно окрашиваются DAPI, приводится в качестве относительной меры индукции апоптоза.
Фиг.10.
Схематичное представление родословной (апоптин)-трансгенных мышей VP3. Белыми квадратиками обозначены мыши-основатели. Квадратики, раскрашенные желтым и зеленым цветом, представляют собой потомство (F1) различных апоптин-трансгенных основателей.
Литература
1. Abrahams, P.J., Huitema, B.A., and Van der Eb, A.J. (1984) Усиленная реактивация и усиленный мутагенез герпес симплекс вируса в нормальных человеческих клетках и Xeroderma Pigmentosum клетках. Molecular Cellular Biology 4, 2341-2346. Abrahams, P.J., Houweling, A., and Van der Eb, A.J. (1992). Высокие уровни усиленной реактивации герпес симплекс вируса в фибробластах кожи из различных наследственных рак-образующих синдромов склонности к раковым заболеваниям. Cancer Research 52, 53-57.
2. Abrahams, P.J., Houweling, A., Cornelissen-Steijger, P.D.M., Arwert, F., Menko, F.H., Pinedo, H.M., Terleth, C., and Van der Eb, A.J. (1996). Наследование ненормальной экспрессии SOS-лодобных реакций в Xeroderma Pigmentosum и наследственные синдромы склонности к раковым заболеваниям. Cancer Research, 56, 2621-2625.
3. Bellamy, С.ОС., Malcomson, R.D.G., Harrison/ D.J., and Wyllie-H.1995. Смерть и заболевания клетки: Биология и регуляция апоптоза. Seminars on Cancer Biology 6, 3-12.
4. Brinster, R.L., Chen, H.Y., and Trumbauer, M.E. (1981). Мышиные ооциты транскрибируют инъецированный Xenopus 5S RNA ген. Science 211, 396-398.
5. Danen-van Oorschot, A.A.A.M., Fischer, D., Grimbergen, J.M., Klein, В., Zhuang, S.-M., Falkenburg, J.H.F., Backendorf, C., Quax, P.H.A., Van der Eb, A. J. , and Noteborn, M.H.M. (1997). Апоптин индуцирует апоптоз в человеческих трансформированных или злокачественных клетках, но не в нормальных клетках. Proceedings National Academy Sciences, USA: 94, 5843-5847.
6. Danen-van Oorschot, A.A.A.M., Den Hollander, A., Tak-ayama, S., Reed, J. , Van der Eb, A.J., and Noteborn, M.H.M. (1997). BAG-1 ингибирует р53-индуцированный, но не апоптин-индуцированный апоптоз. Apoptosis, In Press.
7. Dinsart, С., Cornelis, J. J., Klein, В., van der Eb, A. J., and Rommelaere, J. (1984). Трансфекция с внеклеточной UV-поврежденной ДНК стимулируют крысиные и человеческие клетки к экспрессии мутированого фенотипа к парвовирусу Н-1. Molecular Cellular Biology, 4, 324.
8. Drezen, J.M., Nouvel, P., Babinet, С., and Morello, D. (1992). Различная регуляция экспрессии гена класса 1 у взрослых мышей и в течение их развития. The Journal of Immunology 149, 429-437.
9. De Ronde, A., et al. (1989). Малый Т-антиген вируса SV40 необходим для морфологической трансформации человеческих фиробластов. Virology 171, 260-263.
10. Duke, R. C. , Ocjius, D.M., Young, J, D-E. (1996). Суицид клетки в здоровье и заболеваниях. Scientific American December 1996, 48-55.
11. Earnshaw, W.C., 1995. Ядерные изменения при апоптозе. Current Opinion in Cell Biology 7, 337-343.
12. Fischer, D. F. , Gibbs, S., Van de Putte, P., and Backendorf, С. (1996). Внутризависимые элементы контроля транскрипции регулируют экспрессию гена SPRR2A в процессе терминальной дифференциации кератиноцитов. Molecular and Cellular Biology 16, 5365-5374.
13. Graham, F. L. and Van der Eb, A.J. (1973). Новая методика анализа инфекционности человеческого аденовируса 5 DNA. Virology 52, 456-467.
14. Gruis, N.A. et al. (1995). Гомозиготы для CDKN2(P16) мутации зародышевой линии в семейной Голландской обычной меланоме. Nature Genetics, 10: 351-353, 1995.
15. Hockenberry, D.M. (1994). Bcl-2 в раковых клетках, развитие и апоптоз. Journal of Cell Science, Supplement 18, 51-55.
16. Jacks, Т. (1996) Уроки, полученные из р53 мутированных мышей. Journal of Cancer Research and Clininal Oncology 122, 19-27.
17. Jacks, Т. (1994). Спектральный анализ опухолей у р53-мутантных мышей. Current Biology 4, 1-7.
18. Kerr, J. F.R., Winterford, C.M., and Harmon, B.V. (1994). Апоптоз: Значение в раковых заболеваниях и раковой терапии. Cancer 73, 2013-2026.
19. Klein, В., Pastink, A., Odijk, H., Westerveld, A., and Van der Eb, A.J. (1990). Трансформация и иммортализация диплоидных Xeroderma pigmentosum фибробластов. Experimental Cellular Research 191, 256-262.
20. Levine, A. J. (1997). P53, клеточный сторож, хранитель развития и роста. Cell 88, 323-331.
21. Lowe, S.W., et al. (1994). Аннулирование онкоген-зависимого апоптоза приводит к трансформации р53-дефицитных клеток. Proceedings of the National Academy of Sciences, USA 91, 2026-2030.
22. Maniatis, Т. , Fritsch, E.F., and Sambrook, J. (1982) Молекулярное клонирование. Лабораторное руководство. CSHL Press, New York, USA.
23. McBurney, M. W. et al. (1982) Выделение мужских мышиных эмбриональных клеток карциномы и пути репликации их хромосом. Developments in Biology 89, 503-508.
24. McDonell T. J., Meyn, R.E., Robertson, L.E. (1995). Использование апоптической регуляции смерти клеток в терапии рака. Seminars in Cancer Biology 6, 53-60.
25. Menke, A.L, Shvarts, A., Riteco, N., Van Ham, R.C.A., Van der Eb, A. J. , and Jochemsen, A.G. (1997). Wilms' Tumor 1 (WT1) слипшиеся варианты без КТС индуцируют апоптоз в р53-негативных и р53-позитивных HepG2 клетках. Cancer Research 57, 1353-1363.
26. Mellor, A.L., Golden, L., Weiss, E., Bullman, H., Hurst, Simpson, E. , James, R. F. , Townsend, A. R., Taylor, P.M., Schmldt, W., Ferluga, J., Leben, L., Santamaria, M., Atfield, G. Festenstein, H, Flavell, R.A. (1982). Экспрессия мышиного Н-2КЬ антигена гистосовместимости в клетках, трансформированых с клонированным Н-2 геном. Nature 298, 529-534.
27. Morello, D. , Moore, G., Salmon, A. M., Yaniv, M., and Babinet, C. (1986). Изучение экспрессии слившегося гена Н-2К/человеческого гормона роста в гигантских трансгенных мышах. The EMBO Journal 5, 1877-1883.
28. Noteborn, M.H.M. and Pietersen, A.M. (1998). PCT заявка 98/00213 названная Аденовирусный вектор.
29. Noteborn, M.H.M. (1996). PCT application WO 96/41191. Апоптин индуцирует апоптоз в человеческих трансформированных и злокачественных клетках, но не в нормальных клетках, что является существенной характеристикой для развития анти-раковой терапии.
30. Noteborn and De Boer, G.F. (1996). Patent USA/no. 030, 335.
31. Noteborn, M. H.M., De Boer, G.P., Van Roozelaar, D., Karreman, C., Kranenburg, О. , Vos, J., Jeurissen, S., Zantema, A., Hoeben, R., Koch, G., Van Ormondt, H., and Van der Eb, A.J. (1991). Характеристика клонированного ДНК вируса анемии цыплят, которой содержит все элементы инфекции репликационного цикла. Journal of Virology 65, 3131-3139.
32. Noteborn, M.H.M., Hoeben, R.C., and Pietersen, A. (1997). Инструмент доставки гена, экспрессирующий апоптоз индуцирующие белки VP2 и/или апоптин. European Patent Application no. 97201121.7.
33. Noteborn, M.H.M., Todd, D., Verschueren, C.A.J., De Gauw, H.W.P.M., Curran, W.L., Veldkamp, S., Douglas, A.J., McNulty, M.S., Van der Eb, A.J., and Koch, G. (1994). Один из белков вируса анемии цыплят индуцирует апоптоз. Journal of Virology 68, 346-351.
34. Noteborn, M.H.M., Kranenburg, О., Zantema, A., Koch, G., De Boer, G. F, and Van der Eb, A.J. (1992). Транскрипция генома вируса анемии цыплят (CAV) и синтез его 52 кд белка. Gene 118, 267-271.
35. Paulovich, A.G., Toczyski, D., Hartwell, H. (1997). Когда точка инициации не действует. Cell 88, 315-321.
36. Philips, В., and Rundell, К. (1988). Неудача малого t антигена вируса SV40 в попытке дезорганизовать актиновые структуры в не пермисивных клеточных линиях. Journal of Virology 62, 768-775.
37. Pines, J. (1995). GFP в клетках млекопитающих. Trends in Genetics 11, 326-327.
38. Ponec, М., Kempenaar, J.A., and De Kloet, E.R. (1981). Кортикоиды и культивированные человеческие эпидермальные кератиноциты: специфическое внутриклеточное связывание и клиническая эффективность. Journal Investmental Dermatology 76, 211-214.
39. Reinwald, J.G. and Green, H. (1975). Cell 6, 331-343.
40. Sachs, L. and Lotem, J. (1993). Контроль программированной смерти клеток для нормальных клеток и клеток лейкоза: Новые применения в терапии. Blood 82, 15-21.
41. Schrier, Р. I., Bernards, R., Vaessen, R. Т. М. J., Houweling, A. and Van der Eb, A. J. (1983). Экспрессия класс 1 главных антиген гистосовместимости прекращается при наличии онкогенного аденовируса 12 в трансформированных клетках крысы. Nature, 305, 771-775.
42. Smits, P.H.M. et al. (1992). Модуляция трансформации и транскрипции вируса папиломы человека типа 16 при делении положения короткой ветви человеческой хромосомы 11 может быть имитирована малым антигеном t SV40. Virology 190, 40-44.
43. Southern, E.M. (1975). Определение специфических последовательностей среди фрагментов ДНК, разделенных посредством гель-электрофореза. Journal of Molecular Biology 98, 503-517.
44. Srivastava, S., Zou. Z., et al. (1990). Трансмиссия зародышевой линии мутированного гена р53 в семьях, имеющих склонность к раковым заболеваниям с синдромом Li-Fraumeni. Nature 348, 747-749.
45. Steller, H. (1995). Механизмы и гены клеточного суицида. Science 267, 1445-1449.
46. Telford, W.G., King, L.E., Fraker, P.J. (1992). Сравнительная оценка некоторых связывающихся ДНК красителей в определении апоптоз-ассоциированной деградации хроматина методом проточной цитометрии. Cytometry 13, 137-143.
47. Teodoro, J.G. and Branton, P.E. (1997). Регуляция апоптоза посредством вирусных генных продуктов. Journal of Virology 71, 1739-1746.
48. Thompson, С.В. (1995). Апоптоз в патогенезе и лечении заболеваний. Science 267, 1456-1462.
49. Van den Heuvel, S.J.L., Van Laar, Т., Kast, W.M., Melief, C.J., Zantema, A., and Van der Eb, a.J. (1990). Ассоциация между клеточным р53 и аденовирусным 5 Е1В-55 кд белками снижает онкогенность Ad-трансформированных клеток. ЕМВО Journal 9, 2621-2629.
50. Van Laar Т., Steegenga wt et al. (1994). В Bloom's Syndrome клетках GM1492 нет заметного количества белка р53, но в них происходит нормальная остановка G1 клеточного цикла после UV облучения. Oncogene. 9: 981-983.
51. White, E. (1996). Жизнь, смерть и погоня за апоптозом. Genes and development 10, 1-15.
52. Wyllie, A.H. (1995). Генетическая регуляция апоптоза. Current Opinion in Genetics and Development 5, 97-104.
53. Wyllie, A.H., Kerr, J.P.R., Currie, A.R. (1980). Смерть клетки: Значение апоптолиза. International Review of Cytology 68, 251-306.
54. Zhuang, S.-M., Landegent, J.E., Verschueren, C.A.J., Falkenburg, J. H. F. , Van Ormondt, H., Van der Eb, A.J., Noteborn, M.H.M. (1995). Апоптин, белок, закодированный вирусом анемии цыплят, индуцирует смерть клетки в различных гематологических злокачественных клетках in vitro. Leukemia 9 SI, 118-120.
55. Zhuang, S.-M., Shvarts, A., Van Ormondt, H., Jochemsen, A.-G., Van der Eb, A.J., Noteborn, M.H.M. (1995). Апоптин, белок, выделенный из вируса анемии цыплят, индуцирует р53-независимый апоптоз в человеческих остеосаркомных клетках. Cancer Research 55, 486-489.
| название | год | авторы | номер документа |
|---|---|---|---|
| ВЕКТОР ДОСТАВКИ ГЕНОВ, СПОСОБНЫЙ ИНДУЦИРОВАТЬ АПОПТОЗ В КЛЕТКЕ | 1998 |
|
RU2252255C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pcDNA4-Apo-2NLS2, НЕСУЩАЯ СИНТЕТИЧЕСКИЙ ГЕН БЕЛКА АПОПТИНА, ИНДУЦИРУЮЩЕГО p53 НЕЗАВИСИМЫЙ АПОПТОЗ ОПУХОЛЕВЫХ КЛЕТОК | 2013 |
|
RU2541777C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pGEM-Puro-DS-Apo, СОДЕРЖАЩАЯ СИНТЕТИЧЕСКИЙ ГЕН АПОПТИНА, ФЛАНКИРОВАННЫЙ ПОСЛЕДОВАТЕЛЬНОСТЯМИ ГЕНОМА ВИРУСА ОСПОВАКЦИНЫ ИЗ РАЙОНА C10L-C12L, И РЕКОМБИНАНТНЫЙ ШТАММ VVdGF-ApoS24/2 ВИРУСА ОСПОВАКЦИНЫ, ПРОДУЦИРУЮЩИЙ АПОПТИН | 2012 |
|
RU2492238C1 |
| СПОСОБ ПОЛУЧЕНИЯ СИСТЕМЫ НАПРАВЛЕННОЙ ДОСТАВКИ БЕЛКОВЫХ МОЛЕКУЛ (ОНКОЛИТИЧЕСКИХ БЕЛКОВ) В ОПУХОЛЕВЫЕ ТКАНИ НА ОСНОВЕ АКТИВИРОВАННЫХ ЛИМФОЦИТОВ | 2013 |
|
RU2552609C1 |
| АДЕНОВИРУС-РЕКОМБИНАНТ, НЕСУЩИЙ СТРУКТУРУ АДЕНОВИРУСНОГО ВЕКТОРА (ВАРИАНТЫ), СПОСОБ ВОССТАНОВЛЕНИЯ ФУНКЦИИ ПРОТЕИНА P53 В ОПУХОЛЕВОЙ КЛЕТКЕ С ДЕФИЦИТОМ ПРИРОДНОГО P53, СПОСОБ ПРОИЗВОДСТВА АДЕНОВИРУСА-РЕКОМБИНАНТА | 1994 |
|
RU2222600C2 |
| СПОСОБ СТИМУЛИРОВАНИЯ АПОПТОЗА | 2006 |
|
RU2340349C2 |
| СИНТЕТИЧЕСКИЙ МУТАНТНЫЙ ГЕН RB (ВАРИАНТЫ), СИНТЕТИЧЕСКИЙ МУТАНТНЫЙ ГЕН P53, ПЛАЗМИДА (ВАРИАНТЫ), МУТАНТНЫЙ БЕЛОК (ВАРИАНТЫ), СПОСОБ ИНГИБИРОВАНИЯ КЛЕТОЧНОЙ ПРОЛИФЕРАЦИИ (ВАРИАНТЫ), СПОСОБ ИНДУЦИРОВАНИЯ АПОПТОЗА | 1995 |
|
RU2192431C2 |
| УНИВЕРСАЛЬНЫЕ РАКОВОСПЕЦИФИЧНЫЕ ПРОМОТОРЫ И ИХ ИСПОЛЬЗОВАНИЕ В ПРОТИВООПУХОЛЕВОЙ ТЕРАПИИ | 2013 |
|
RU2539764C2 |
| ИММОРТАЛИЗОВАННЫЕ ФИБРОБЛАСТЫ ЭМБРИОНА ЦЫПЛЕНКА | 2015 |
|
RU2694318C2 |
| Рекомбинантный штамм VV-NS1-dGF вируса осповакцины, продуцирующий белок NS1 парвовируса H-1 и обладающий онколитической активностью в отношении глиобластомы человека | 2018 |
|
RU2692628C1 |
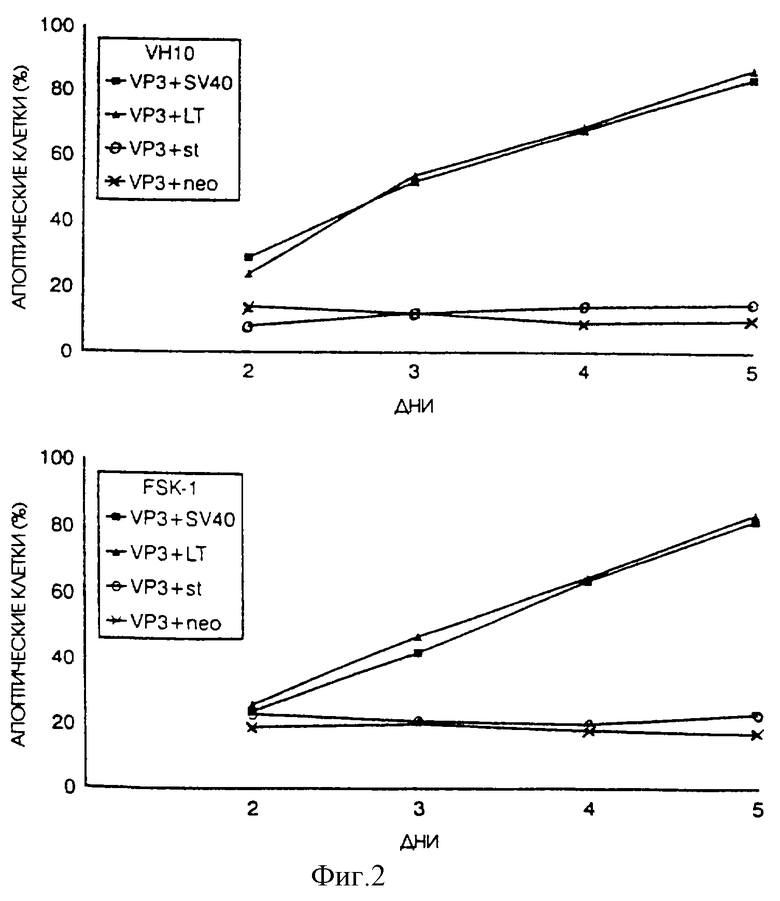
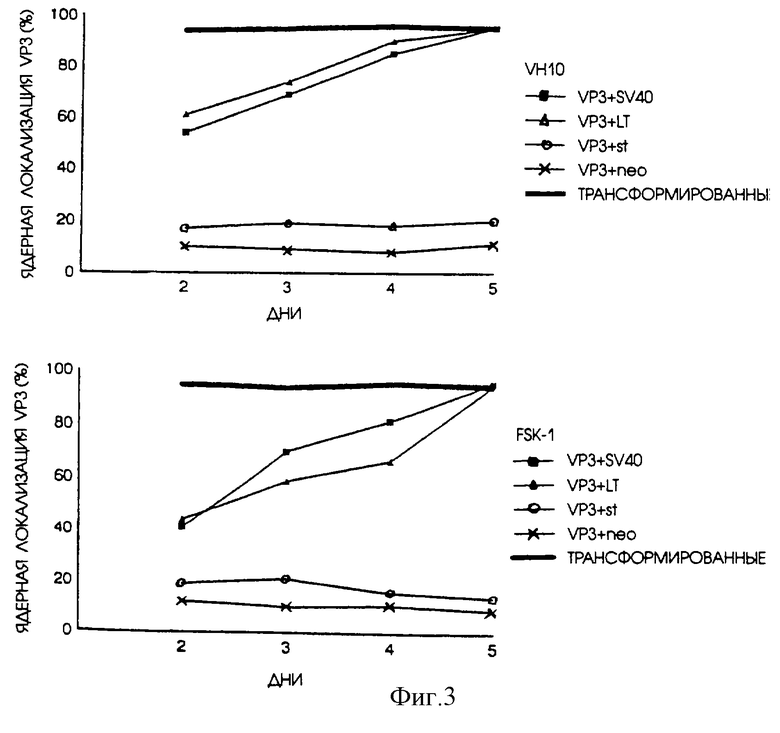
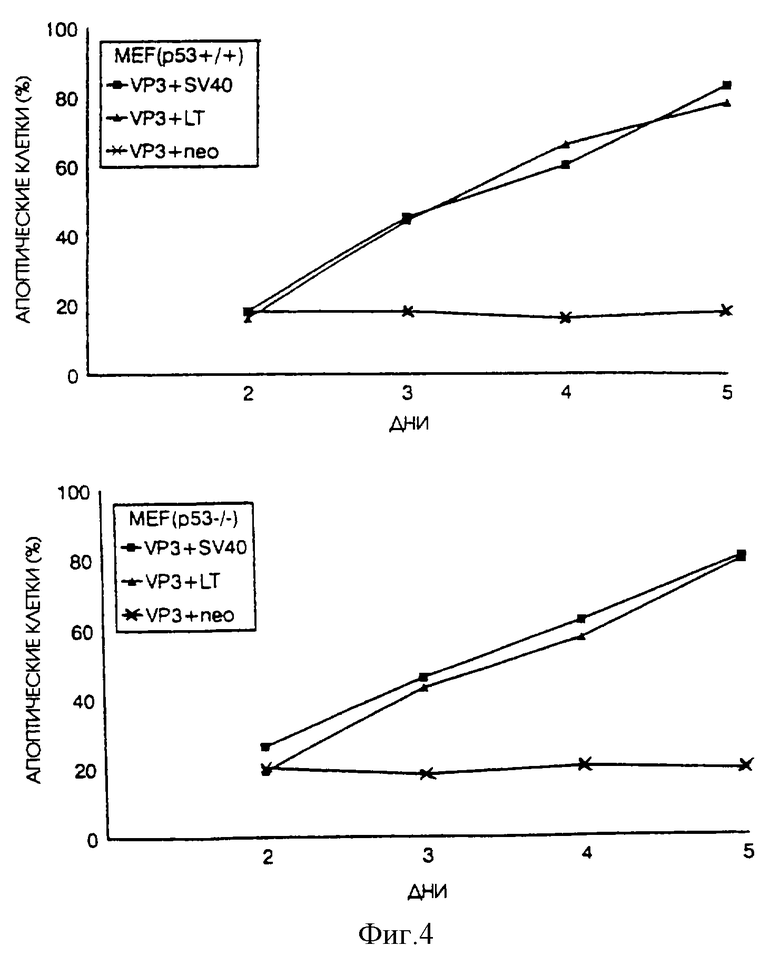
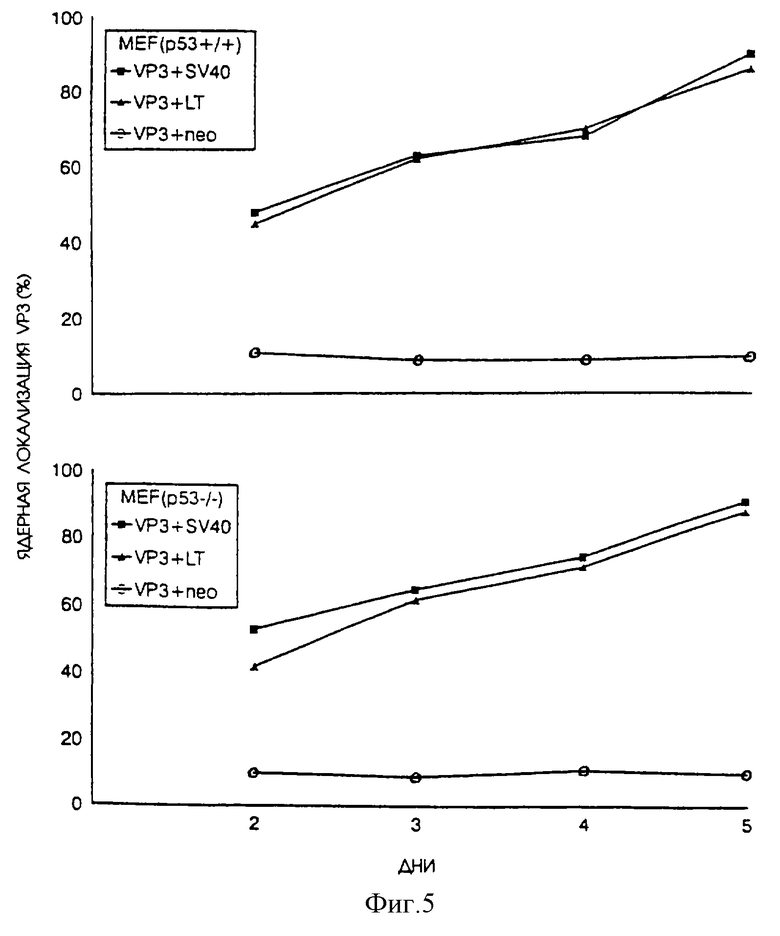
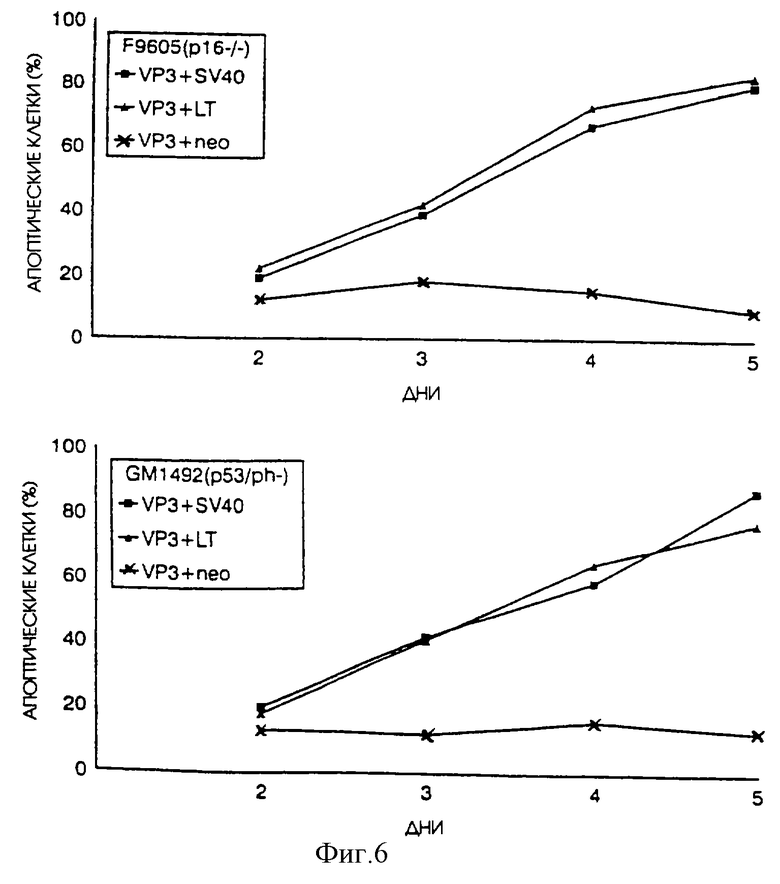
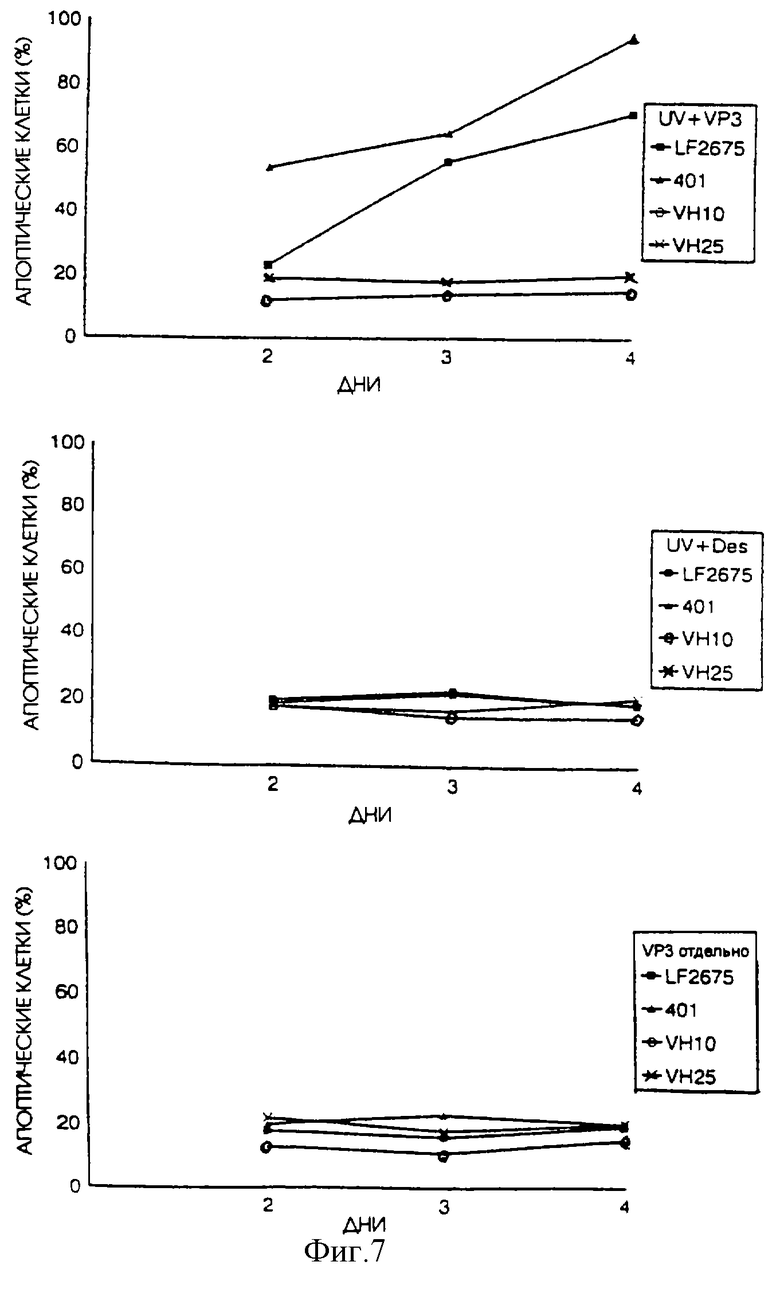
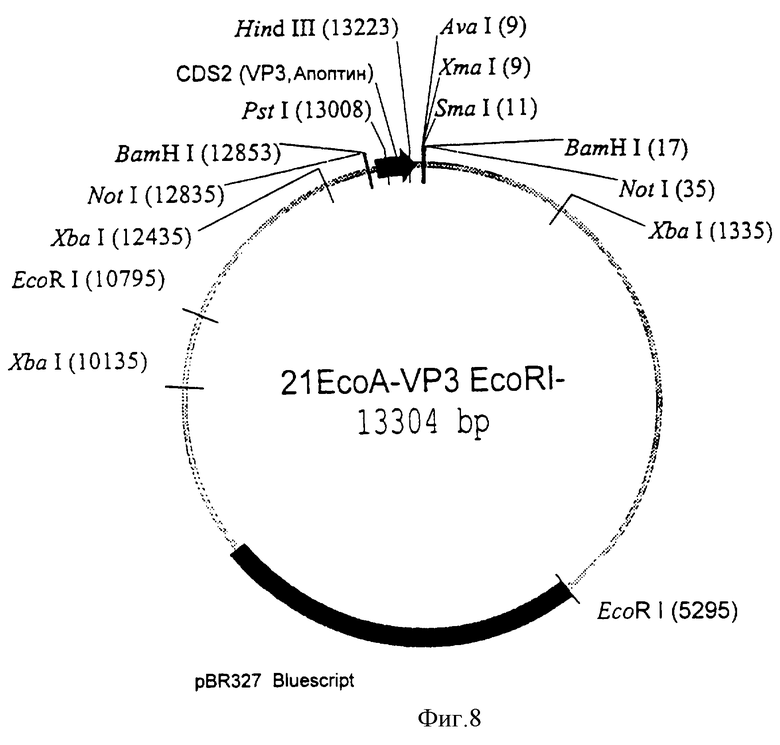
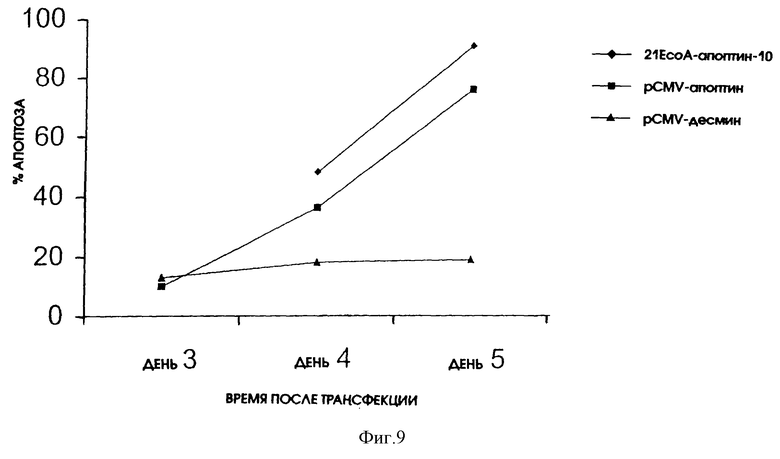
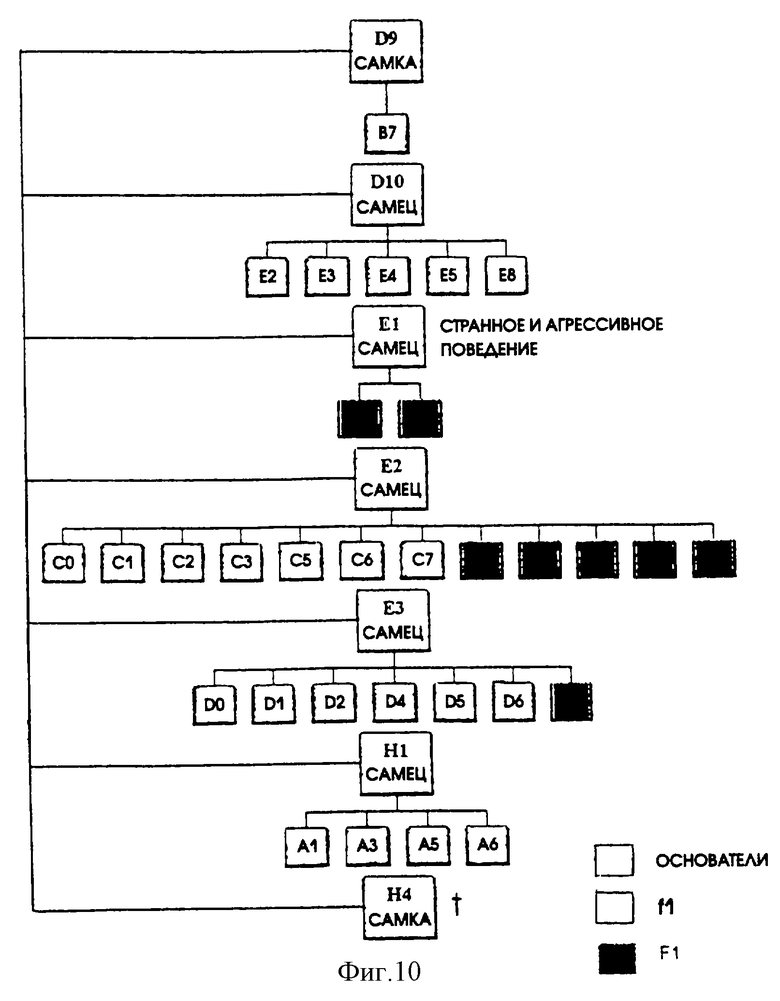
Изобретение относится к области медицинской биотехнологии, в частности активации апоптин-индуцированного апоптоза различными агентами, вызывающими трансформацию клетки в нормальных и/или склонных к раковым заболеваниям клетках. Сущность изобретения состоит в следующем. Апоптин, или так называемый VP3, является вирусным белком, выделенным из вируса анемии цыплят. Также изобретение относится к профилактической противоопухолевой терапии нормальных и/или склонных к раковым заболеваниям клеток. Обработка нормальных и/или склонных к раковым заболеваниям клеток агентами, индуцирующими развитие опухоли, будет активировать апоптин-индуцированный апоптоз, приводя к уничтожению потенциальных опухолевых клеток. Также изобретение относится к диагностике агентов, вызывающих раковые заболевания. Агенты, обладающие опухолеродной активностью, могут быть исследованы путем их экспрессии в нормальных клетках и анализа их способности вызывать апоптин-индуцированный апоптоз. Технический результат - расширение арсенала средств для диагностики и терапии опухолей. 6 с. и 6 з.п. ф-лы, 10 ил.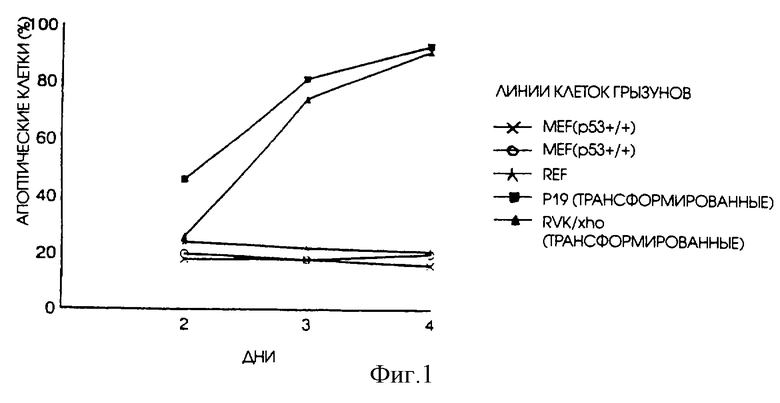
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |
| DANEN-VAN OORSCHOT A.A.M | |||
| et al | |||
| Apoptin induces apoptosis..., Proc | |||
| Nat | |||
| Acad | |||
| Sci | |||
| USA, may l997, p | |||
| Ударниковое приспособление к автоматическому оружию с коротким откатом ствола | 1926 |
|
SU5843A1 |

