Область техники, к которой относится изобретение
Настоящее изобретение относится к биотехнологии, в частности генной инженерии, к искусственным ДНК-конструкциям, демонстрирующим высокую активность транскрипционного промотора, с которого осуществляется экспрессия расположенной ниже произвольной нуклеотидной последовательности, транскрибируемой РНК-полимеразой II. Активность промоторов ограничена только раковыми клетками.
Промоторы могут быть использованы для генной терапии новообразований различной природы.
Уровень техники
Рак представляет собой в настоящее время наиболее серьезную проблему медицины. Становится все более очевидным, что господствовавшая до последнего времени концепция молекулярного нацеливания (таргетинга) противораковых препаратов на предположительные ключевые молекулы раковой клетки, которые вовлечены в развитие опухоли, находится в кризисном состоянии (Salk JJ and Fox EJ et al. (2010) Annu Rev Pathol Mech Dis 5:51-75). Подавляющее большинство агентов этой категории, которые проходили доклинические и клинические испытания, оказались малоэффективными (Hambley TW and Hait WN (2009) Cancer Res 69:1259-62). Это вполне ожидаемый кризис: рак как заболевание сочетает в себе сложность клеточной организации, присущую и другим болезням, со сложностью растущей системы, способной адаптироваться к воздействиям лекарствами путем существования микрогетерогенности внутри опухоли (Merlo LM and Pepper JW et al. (2006) Nat Rev Cancer 6:924-35). Это приводит к тому, что большинство раковых генов принимают только незначительное участие в развитии рака. Вследствие этого их трудно идентифицировать для использования в качестве мишеней действия противораковых агентов (Salk JJ and Fox EJ et al. (2010) Annu Rev Pathol Mech Dis 5:51-75). В результате в терапии рака сложилась критическая ситуация: с одной стороны, медицина располагает достаточно эффективными, но высокотоксичными средствами химиотерапии, с другой - малотоксичными, но малоэффективными средствами молекулярной таргетной терапии (МТТ).
В последнее десятилетие активно начала развиваться генная терапия рака (ГТ). Общий принцип ГТ заключается в доставке регулируемого генетического материала в раковые клетки, где в результате продуцируются продукты, способные к уничтожению раковых клеток. Подходы ГТ могут быть разделены на две широкие категории (Sverdlov ED (2009) Mol Gen Mikrobiol Virusol 24:93-113) / (http://link.springer.com/article/10.3103%2FS089141680903001X). В первой используют стратегию таргетной терапии. При этом в роли таргетного агента выступает продукт гена, вводимого каким-либо способом в клетку опухоли, который является ингибитором того или иного продукта, концентрация которого в раковой клетке повышена, и это является одной из причин ракового процесса. К этой же категории можно отнести обратную технологию -доставка в опухоль гена, продукт которого компенсирует недостаток определенного белка в раковых клетках. В обоих вариантах мишенью воздействия является определенное звено в сигнальных системах клетки, которое меняется при раковом перерождении и способствует ему. Варианты ГТ, основанные на таргетном принципе, страдают от тех же недостатков, что иМТТ.
Первый генно-терапевтический вирус для терапии рака головы и шеи был одобрен для клинического применения в Китае в 2003 г. под названием Гендицин (Gendicine). Вирус содержит в качестве терапевтического гена р53 (Peng Z (2005) Hum Gene Ther 16:1016-27, Wilson JM (2005) Hum Gene Ther 16:1014-5). В сочетании с радиотерапией Гендицин вызывал полную регрессию опухоли у 64% пациентов и частичную - у 29%, тогда как одна радиотерапия давала полную регрессию у 19% и частичную у 60%, что, по-видимому, показывает заметное улучшение результата при комбинированной терапии (Peng Z (2005) Hum Gene Ther 16:1016-27). Но эти результаты показывают также, что только 64% откликаются на обработку, и дальнейшее поведение их болезни неизвестно. Есть основания ожидать появления вторичного роста опухоли, поскольку было показано, что при введении р53 в опухоли, в которых он исходно поврежден, вызывает появление устойчивых к р53 вариантов (Martins CP and Brown-Swigart L et al. (2006) Cell 127:1323-34). Полученные данные с использованием ГТ в качестве молекулярной таргетной терапии предполагают, что также как и классическая МТТ и вследствие тех же причин, она едва ли будет высокоэффективной или универсальной.
Вторая стратегия генной терапии, направленная на уничтожение опухолевых клеток, как таковых, путем использования их свойств, которые характерны для всех раковых клеток, например, повышенная скорость митотических делений, в этом отношении подобна химиотерапии. Однако, в отличие от последней, токсин, убивающий раковые клетки путем ингибирования систем репликации, образуется внутри них, так что свойственная химиотерапии токсичность в данном случае резко снижается. Этот подход известен как ген-направленная энзиматическая пролекарственная терапия, ГНЭПТ (Gene-directed enzyme prodrug therapy, GDEPT) или генная терапия с использованием генов самоубийства опухоли (suicide gene therapy) (Altaner С (2008) Cancer Lett 270:191-201, Fillat С and Carrio M et al. (2003) Curr Gene Ther 3:13-26, Portsmouth D and Hlavaty J et al. (2007) Mol Aspects Med 28:4-41, Seth P (2005) Cancer Biol Ther 4:512-7). Подход не является молекулярно таргетированным и потому лишен всех недостатков МТТ.
Одним из важных элементов успеха генной терапии рака является адекватная система экспрессии терапевтических генов, поскольку данные гены должны работать в раковых клетках и не работать в нормальных клетках организма. В идеальном случае система контроля терапевтических генов в организме должна обеспечивать (i) строго тканеспецифическую и (ii) достаточно сильную экспрессию трансгена, чтобы с одной стороны обеспечить безопасность, а с другой - эффективность системы. Для этого при конструировании векторов используют промоторы и энхансеры, специфично работающие в опухолях данной ткани (Robson Т and Hirst DG (2003) J Biomed Biotechnol 2003:110-137, Saukkonen К and Hemminki A (2004) Expert Opin Biol Ther 4:683-96). Однако все природные опухолеспецифичные промоторы обладают двумя существенными недостатками: с одной стороны, являются слабыми, а с другой - имеют высокий уровень активности только в определенных раковых клеточных линиях, то есть не являются универсальными. Так, AFP промотор наиболее активен в клетках карциномы печени, PSA промотор - в опухолевых клетках простаты, промотор Сох-2 - в раковых клетках желудка и двенадцатиперстной кишки, МК промотор - в клетках нейробластомы (Adachi Y and Reynolds PN et al. (2001) Cancer Res 61:7882-8).
Большинство опухолеспецифичных промоторов обладает низкой активностью по сравнению с конститутивными сильными промоторами, такими как промоторы вирусов SV40 и CMV (Van Houdt WJ and Haviv YS et al. (2006) J Neurosurg 104:583-92, Lu В and Makhija SK et al. (2005) Gene Ther 12:330-8, Rein DT and Breidenbach M et al. (2004) J Gene Med 6:1281-9).
Есть и другая проблема. Даже сравнительно сильные опухолеспецифические промоторы, такие как промотор гена BIRC5 (hSurv), кодирующий ингибитор апоптоза сурвивин, и промотор гена обратной транскриптазы теломеразы человека (hTERT) с достаточно широким спектром активности, проявляют ее не во всех раковых клетках. Так, например, промотор hSurv активен в опухолях только примерно 60- 80% пациентов с немелкоклеточным раком легких (NSLC), а промотор TERT - в 60% пациентов с NSLC (Hsu СР and Miaw J et al. (2003) Eur J Surg Oncol 29:594-9). При этом наблюдается значительная вариабельность относительной активности данных промоторов в различных опухолевых клеточных линиях. Так, активность промотора сурвивина варьирует в пределах от 0,3 до 16% от активности промотора CMV (Chen JS and Liu JC et al. (2004) Cancer Gene Ther 11:740-7, Konopka К and Spain С et al. (2009) Cell Mol Biol Lett 14:70-89, Zhu ZB and Makhija SK et al. (2004) Cancer Gene Ther 11:256-62), а эффективность работы промотора hTERT может различаться до 20 раз в зависимости от раковой клеточной линии (Gu J and Fang В (2003) Cancer Biol Ther 2:S64-70).
Вариабельность природных опухолеспецифичных промоторов в разных опухолях затрудняет подбор доз пролекарства для получения терапевтического эффекта и меняет терапевтический индекс препарата от опухоли к опухоли.
Для увеличения эффективности опухолеспецифической экспрессии терапевтических генов используют комбинированные и двойные (химерные) промоторы. Химерные промоторы могут включать в себя комбинации известных промоторов друг с другом или с отдельными гетерологичными регуляторными элементами с целью увеличить силу и специфичность экспрессии в раковых клетках (Wu С and Lin J et al. (2009) Mol Ther 17:2058-66). Примером химерного промотора может служить комбинация PhTERT с минимальным промотором цитомегаловируса (PhTERT-CMV) (Davis JJ and Wang L et al. (2006) Cancer Gene Ther 13:720-3); или с ТАТА боксом, который отсутствует в нативном PhTERT. Как правило, авторы различных исследований, направленных на эффективное использование гибридных промоторов, идут по пути максимального увеличения эффективности и специфичности экспрессии в определенном виде рака. Примером такого подхода служит работа Poulsen et al. (Poulsen TT and Pedersen N et al. (2008) Cancer Gene Ther 15:563-75). В этой работе было идентифицировано два гена, высокоэкспрессируемых в мелкоклеточном раке легких (SCLC). Один из них кодирует фактор транскрипции hASH1, а другой, EZH2 относится к члену семейства Поликомб. Когда промоторы этих генов были соединены в одну конструкцию, то полученный химерный промотор был способен вызывать сильную экспрессию трансгена специфично в клетках SCLC. Такой промотор способен инициировать экспрессию генов-убийц в SCLC, но не в других видах рака.
Такая идеология максимальной специфичности активности промотора в определенном типе рака является распространенной (см., например, Farokhimanesh S and Rahbarizadeh F et al. (2010) Biotechnol Prog 26:505-11). Использование строго специфичных к данной опухоли промоторов и других регуляторных элементов имеет в качестве преимущества максимальное снижение побочных эффектов за счет снижения экспрессии трансгенов в нормальных тканях. Однако недостатком таких подходов является их неуниверсальный характер и связанное с этим неизбежное увеличение стоимости препаратов, основанных на таких промоторах. Компромиссным вариантом является использование более универсальных раковоспецифических промоторов, способных работать в широком спектре опухолей, но не в нормальных клетках. Несколько увеличивая риск поражения нормальных тканей, такой подход является более экономически оправданным: одни и те же конструкции могут использоваться в лечении широкого спектра опухолей. Существует и еще одно важное соображение в пользу использования промоторов более широкого спектра действия. Оно связано с плохо изученной специфичностью экспрессии генов в метастазах данной опухоли. Нет строгой гарантии, что узкоспецифический промотор, хорошо работающий в первичной опухоли, сохранит эту способность во всех ее метастазах. Использование универсальных промоторов снижает вероятность инактивации промотора в метастазах.
Раскрытие изобретения
Настоящее изобретение направлено на устранение недостатков, свойственных промоторам из уровня техники, строго специфичным в отношении того или иного типа рака, и на создание универсальных промоторов, обладающих широким спектром действия в отношении различных типов рака и их метастазов, и при этом все еще сохраняющих способность функционировать по существу только в опухолевых, но не в нормальных клетках.
Настоящее изобретение обеспечивает искусственные универсальные опухолеспецифичные двойные промоторы, обладающие высокой активностью в широком спектре раковых клеток вне зависимости от их типа.
Опухолеспецифичные двойные промоторы созданы на основе модифицированных промоторов генов обратной транскриптазы теломеразы человека (hTERT) и сурвивина человека и мыши (Surv). Ген сурвивина человека (BIRC5) кодирует белок сурвивин, который относится к семейству белков-ингибиторов апоптоза и является одним из ключевых участников опухолеобразования. Промотор сурвивина обладает высокой опухолеспецифичностью и активен в подавляющем большинстве (85-90%) опухолей (Takakura M and Kyo S et al. (1999) Cancer Res 59:551-7, Ambrosini G and Adida С et al. (1997) Nat Med 3:917-21, Fukuda S and Pelus M (2006) Mol Cancer Ther 5:1087-98). Ген hTERT кодирует каталитическую субъединицу теломеразы человека. Транскрипционная активность данного гена наблюдается во время эмбрионального развития и часто примерно в 85% случаев присутствует в опухолевых клетках, тогда как в подавляющем большинстве нормальных клеток организма экспрессия hTERT подавлена (Cong YS and Wright WE et al. (2002) Microbiol Mol Biol Rev 66:407-25).
Показано, что созданные промоторы по количественным и качественным критериям отличаются от составляющих их промоторов hTERT и Surv.
В экспериментах по транзиентной трансфекции раковых клеточных линий плазмидными векторами, содержащими репортерный ген люциферазы светлячка под контролем созданных промоторов, продемонстрирована высокая активность созданных промоторов, превышающая активность как сильного конститутивного промотора вируса SV40, так и одиночных промоторов генов hTERT и Surv. При этом в экспериментах с использованием нормальных клеток (фибробластов легкого) доказана опухолеспецифичность созданных двойных промоторов.
Также на основе созданных двойных промоторов получены терапевтические конструкции, содержащие ген-убийцу или ген-убийцу в сочетании с геном цитокина.
На панели раковых и нормальных клеточных линий продемонстрирован терапевтический опухолеспецифический эффект полученных конструкций.
На основе созданных двойных промоторов могут быть получены плазмидные, вирусные или иные векторные конструкции для опухолеспецифичной экспрессии любого гена или различных генов в раковых клетках. При этом размер промотора может быть увеличен, а к нуклеотидной последовательности промотора могут быть добавлены другие регуляторные и генетические элементы, обеспечивающие повышение активности промотора.
Таким образом, настоящее изобретение в своем первом аспекте относится к опухолеспецифичному промотору, состоящему из:
А) нуклеотидной последовательности SEQ ID NO:2 или ее функционального производного, и
Б) нуклеотидной последовательности, выбранной из группы, включающей:
(i) нуклеотидную последовательность SEQ ID NO:1 или ее функциональное производное;
(ii) нуклеотидную последовательность SEQ ID NO:3 или ее функциональное производное.
В промоторе настоящего изобретения последовательность SEQ ID NO:2 может быть расположена выше (в 5′-направлении) относительно последовательности SEQ ID NO:1 или SEQ ID NO:3, или последовательность SEQ ID NO:2 может быть расположена ниже (в 3′-направлении) относительно последовательности SEQ IDNO:1 или SEQ ID NO:3. Промотор настоящего изобретения может содержать другие нуклеотидные последовательности, не влияющие на его активность, например линкерную последовательность.
Наиболее предпочтительные последовательности промоторов настоящего изобретения выбраны из группы, включающей SEQ ID NO:4- 7.
В своем втором аспекте настоящее изобретение относится к экспрессионной кассете, содержащей в направлении от 5′ к 3′ промотор настоящего изобретения, кодирующую последовательность, функционально связанную с промотором, и сигнал полиаденилирования, причем экспрессия кодирующей последовательности находится под контролем указанного промотора.
В экспрессионной кассете настоящего изобретения кодирующая последовательность может кодировать различные терапевтические белки или пептиды, антигены, антисмысловую РНК или рибозимы. В частности, экспрессионная кассета может содержать кодирующую последовательность, которая включает гены терапевтического вмешательства, выбранные из группы, включающей: негативные доминантные мутанты (НДМ) генов, участвующих в онкогенезе; гены-убийцы, гены - ингибиторы ангиогенеза, гены-онкосупрессоры, гены-иммуностимуляторы, гены siRNA.
Негативные доминантные мутанты (НДМ) генов могут быть выбраны из группы, включающей НДМ гена сурвивина, НДМ гена c-Jun, НДМ онкогена RAS. Гены-убийцы могут быть выбраны из группы, включающей гены токсинов и гены ферментов, способных превращать нетоксичный агент (пролекарство) в токсин. Последние могут быть выбраны из группы, включающей ген тимидинкиназы вируса простого герпеса и ген цитозиндезаминазы дрожжей или Е. coli. Гены-ингибиторы ангиогенеза могут быть выбраны из группы, включающей гены белков ангиостатина и эндостатина. Геном-онкосупрессором может быть, например, ген белка р53. Гены-иммуностимуляторы могут быть выбраны из группы, включающей гены IL-1, IL-2, IL-4, IL-6, TNF, GM-CSF, или гамма-интерферона. Гены siRNA могут быть выбраны из группы, включающей siRNA к гену сурвивина.
В третьем аспекте настоящее изобретение относится к экспрессионному вектору, содержащему экспрессионную кассету настоящего изобретения. При этом экспрессионный вектор настоящего изобретения может быть вирусным или невирусным. В том случае, когда вектор настоящего изобретения является вирусным вектором, он может быть выбран из группы, включающей ретровирусный, аденовирусный, аденоассоциированный или герпесвирусный вектор. Невирусный вектор может быть представлен, например, плазмидой.
В четвертом аспекте настоящее изобретением относится к фармацевтической композиции, содержащей терапевтически эффективное количество экспрессионной кассеты или вектора настоящего изобретения и фармацевтически приемлемое вспомогательное вещество, которое без ограничения может быть представлено носителем, растворителем, эксципиентом, наполнителем, буферным агентом, стабилизатором, консервантом и т.д.
Фармацевтическая композиция настоящего изобретения может быть использована для лечения широкого спектра онкологических заболеваний, причем заболевание без ограничения может быть выбрано из группы, включающей рак легкого, рак поджелудочной железы, меланому, фибросаркому, саркому, рак шеи и головы.
В пятом аспекте настоящее изобретение относится к способу лечения онкологического заболевания, включающему введение пациенту фармацевтической композиции настоящего изобретения.
Способ лечения настоящего изобретения может предусматривать без ограничения внутривенное, внутриопухолевое внутримышечное, внутрибрюшинное, подкожное, пероральное введение фармацевтической композиции настоящего изобретения. Способ лечения может быть использован для лечения онкологического заболевания, без ограничения выбранного из группы, включающей рак легкого, рак поджелудочной железы, меланому, фибросаркому, саркому, рак головы и шеи.
Наконец, в шестом аспекте настоящее изобретение относится к применению экспрессионной кассеты или вектора настоящего изобретения для изготовления лекарственного средства для лечения онкологического заболевания. Онкологическое заболевание без ограничения может быть выбрано из группы, включающей рак легкого, рак поджелудочной железы, меланому, фибросаркому, саркому, рак головы и шеи.
Краткое описание чертежей
Фиг.1. Схематическое изображение экспрессионной конструкции PhSurv269-pGL3. PhSurv 269 - фрагмент промоторной области гена сурвивина человека; LUC - ген люциферазы светлячка, polyA - сайт полиаденилирования вируса SV40.
Фиг.2. Схематическое изображение экспрессионных конструкций. Слева приведено название конструкций. PhSurv269 - фрагмент промотора гена сурвивина человека; PhTERT -фрагмент промотора гена обратной транскриптазы теломеразы человека; LUC - ген люциферазы светлячка.
Фиг.3. Схематическое изображение экспрессионной конструкции PhTSurv269-HSVtk-pGL3. PhTERT - фрагмент промоторного региона гена обратной транскриптазы теломеразы человека (hTERT); PhSurv269 - фрагмент промоторного региона гена сурвивина человека; HSVtk - ген тимидинкиназы вируса простого герпеса типа 1; SV40 late poly(A) -сигнал полиаденилирования вируса SV40; Amp(r) - ген р-лактамазы.
Фиг.4. Схематическое изображение экспрессионной конструкции PhTSurv269-HSVtk-mGM-CSF-pGL3. PhTERT - фрагмент промоторного региона гена обратной транскриптазы теломеразы человека (hTERT); PhSurv269 - фрагмент промоторного региона гена сурвивина человека; HSVtk - ген тимидинкиназы вируса простого герпеса типа 1; IRES - участок посадки рибосомы вируса энцефаломиокардита человека; mGMCSF - ген гранулоцитарно-макрофагального колониестимулирующего фактора мыши; SV40 late poly(А) - сигнал полиаденилирования вируса SV40; Amp(r) - ген β-лактамазы.
Фиг.5. Схематическое изображение экспрессионной конструкции PhTSurv269-FCU1-mGM-CSF-pGL3. PhTERT - фрагмент промоторного региона гена обратной транскриптазы теломеразы человека (hTERT); PhSurv269 - фрагмент промоторного региона гена сурвивина человека; FCU1 - гибридный ген цитозиндезаминазы/урацилфосфорибозилтрансферазы; IRES - участок посадки рибосомы вируса энцефаломиокардита человека; mGMCSF - ген гранулоцитарно-макрофагального колониестимулирующего фактора мыши; SV40 late poly(А) - сигнал полиаденилирования вируса SV40; Amp(r) - ген р-лактамазы.
Фиг.6. Схема модифицированной системы PhTSurv269-Cre-LoxP//pCMV-Stop-FCU1. При одновременном попадании в опухолевую клетку киллерного и активационного векторов будет инициироваться следующий каскад событий:1) активация промотора гена PhTSurv269, которая будет приводить к экспрессии гена Cre рекомбиназы; 2) наработанная Cre рекомбиназа будет узнавать loxP-сайты в составе киллерного вектора, а затем вырезать фрагмент, фланкированный этими сайтами; 3) в результате удаления ″Stop″-сигнала, промотор CMV будет ″запускать″ наработку киллерного белка CD-UPRT (продукт экспрессии FCU1). Stop - последовательность длиной 705 п.о., состоящая из тройного повтора (235 п.о. ×3) сигнала полиаденилирования вируса SV40. NLS (Nuclear Localization Signal) - сигнал ядерной локализации. Черными стрелками обозначены LoxP-сайты, пустыми стрелками - промоторы, пустыми прямоугольниками - структурные части генов, пустой окружностью обозначена Stop-последовательность.
Фиг.7. Диаграмма относительной активности исследованных промоторов в различных клеточных линиях. Названия промоторов приведены внизу рисунка. По оси ординат приведена относительная люциферазная активность, нормализованная на люциферазную активность в клетках, трансфицированных конструкцией PV-pGL3. Названия клеточных линий приведены по оси абсцисс. Calu-1 - эпидермоидная карцинома легкого человека, A375 - меланома человека, А549 - карцинома легкого человека, PANC1 - карцинома экзокринной части поджелудочной железы, НТ1080 - фибросаркома.
Фиг.8. Активность универсальных опухолеспецифичных промоторов в различных клеточных линиях. По оси ординат приведена активность люциферазы относительно люциферазной активности в клетках, трансфицированных вектором PV-pGL3. По оси абсцисс обозначены типы клеточных линий, в которых проводилось измерение промоторной активности. В правом верхнем углу приведены названия промоторов, используемых в работе. Высота столбцов отражает среднее значение люциферазной активности в трех независимых экспериментах, приведены стандартные ошибки среднего значения (SEM). PV - промотор вируса SV40, mSurv - фрагмент промоторной области гена сурвивина мыши, hTERT - фрагмент промоторной области гена обратной транскриптазы теломеразы человека.
Фиг.9. Активность созданных опухолеспецифичных промоторов в различных клеточных линиях. По оси ординат приведена активность люциферазы относительно люциферазной активности в клетках, трансфицированных вектором PV-pGL3. По оси абсцисс обозначены типы клеточных линий, в которых проводилось измерение промоторной активности. В правом верхнем углу приведены названия промоторов, используемых в работе. Высота столбцов отражает среднее значение люциферазной активности в трех независимых экспериментах, приведены стандартные ошибки среднего значения (SEM).
Фиг.10. Вестерн-блот анализ содержания белка FCU1 в клетках линии S37, трансфицированных конструкциями, экспрессирующих ген FCU1. 1 - pCMV-FCU1; 2-3 - различные соотношения векторов PhTSurv269-Cre и pCMV-STOP-FCU1 (котрансфекция): 2:2, 1:3. В качестве первичных антител использовали овечьи IgG (″Santa-Cruz″) (1:5000). Вторичные антитела к овечьим IgG были конъюгированы с пероксидазой хрена (″Santa-Cruz″). Клеточные лизаты предварительно нормализовали по суммарному белку.
Фиг.11. Выживаемость раковых клеток линии S37 (саркома мыши) в присутствии 5-фторцитозина, трансфицированных следующими векторами: вектором pCMV-FCU1 (положительный контроль), системой векторов PhTSurv269-Cre (активатор)/pCMV-STOP-FCU1 (эффектор) при соотношении активатор/эффектор 2:2 и 1:3 по количеству мкг.
Фиг.12. Выживаемость раковых клеток линии S37 (саркома мыши) в присутствии 5-фторцитозина, трансфицированных следующими векторами: вектором pCMV-FCU1 (положительный контроль), системой векторов PhTSurv269-Cre/pCMV-STOP-FCU1 и PhSurv-Cre/pCMV-STOP-FCU1.
Фиг.13. Схемы двойных промоторов и соответствующих вариантов транскриптов. Прямоугольниками обозначены промоторы PhTERT, PhSurv, PhSurv269, PmSurv и ген люциферазы светлячка - Luc. Слева указаны названия двойных промоторов. Под схемой каждого двойного промотора линиями обозначены транскрипты, инициируемые с данных промоторов. Утолщенная часть линии обозначает последовательность транскрипта, комплементарную последовательностям промоторов или гена люциферазы, тонкая - линкерным участкам. Изломанной стрелкой обозначены точки инициации транскрипции. Стрелками обозначены праймеры. Над стрелками указаны названия праймеров.
При использовании пар праймеров TSL-F/hSurv_150R, TSL-F/hS269_122R and TSL-F/mS_122R амплифицируются только транскрипты, инициированные с дистальных промоторов. Пара праймеров UPF/Luc_202R позволяет амплифицировать суммарные транскрипты.
Фиг.14. Электрофореграммы продуктов, полученных в результате ОТ-ПЦР и инициированных с дистального и проксимального промоторов. Названия двойных промоторов указаны сверху. Положение по отношению к контролируемому гену отдельных промоторных элементов, составляющих двойной промотор, и количество циклов ПЦР указаны слева и внизу, соответственно. Дистальный - продукты амплификации транскриптов, инициированных с промотора PhTERT с использованием праймеров TSL-F/hSurv_150R, TSL-F/hS269_122R для промотора PhTS и праймеров TSL-F/mS_122R для промоторов, PhTSurv269 и PhTmS. Проксимальный - продукты амплификации с использованием праймеров UPF и Luc_202R суммарных транскриптов и/или транскриптов, инициированных с дистального промотора.
Фиг.15. Выживаемость в присутствии ганцикловира раковых клеток линии НТ1080 (фибросаркома человека), трансфицированных векторами: PhSurv-HSVtk, PhTSurv269-HSVtk, контроль.
Фиг.16. Выживаемость в присутствии 5-фторцитозина раковых клеток линии Calu-1, трансфицированных векторами: PhSurv-FCU1-mGM-CSF, PhTSurv269-FCU1-mGM-CSF, CMV-FCU1-mGM-CSF, контроль.
Фиг.17. Вестерн-блот анализ содержания белка HSVtk в клетках линии НТ1080, трансфицированных конструкциями, экспрессирующими ген HSVtk. 1 - pCMV-HSVtk; PhSurv-HSVtk, PhTSurv269-HSVtk. В качестве первичных антител использовали овечьи IgG (″Santa-Cruz″) (1:5000). Вторичные антитела к овечьим IgG были конъюгированы с пероксидазой хрена (″Santa-Cruz″), Клеточные лизаты предварительно нормализовали по суммарному белку.
Фиг.18. Влияние трансформации клеток карциномы легкого Льюис мыши (LLC) генно-инженерньми конструкциями на скорость роста опухолей, вызванных трансплантацией данных клеток мышам С57В1/6. Нетрансформированные (Control), трансформированные конструкцией CMV-HSVtk-mGM-CSF-pGL3 (CMV), трансформированные конструкцией PhSurv-HSVtk-mGM-CSF-pGL3 (PhSurv) и трансформированные конструкцией PhTSurv269-HSVtk-mGM-CSF-pGL3 (PhTSurv269) клетки трансплантировали мышам. Внутрибрюшинные инъекции ганцикловира (GCV) в количестве 75 мг/кг дважды в день проводили 10 дней начиная с третьего дня после трансплантации клеток. Размеры опухоли измеряли с 6 дня после трансплантации клеток. Данные представляют собой средние значения для группы из 10 животных. По оси абсцисс указано время, прошедшее с момента трансплантации клеток.
Фиг.19. Влияние трансформации клеток карциномы легкого Льюис мыши (LLC) генно-инженерными конструкциями на скорость роста опухолей, вызванных трансплантацией данных клеток мышам С57В1/6. Трансформированные конструкцией CMV-HSVtk-mGM-CSF-pGL3 (CMV), трансформированные конструкцией PhSurv-HSVtk-mGM-CSF-pGL3 (PhSurv) и трансформированные конструкцией PhTSurv269-HSVtk-mGM-CSF-pGL3 (PhTSurv269) клетки трансплантировали мышам. Внутрибрюшинные инъекции ганцикловира (GCV) в количестве 75 мг/кг дважды в день проводили 10 дней начиная с третьего дня после трансплантации клеток. Размеры опухоли измеряли с 6 дня после трансплантации клеток. Данные представляют собой средние значения для группы из 10 животных. По оси абсцисс указано время, прошедшее с момента трансплантации клеток.
Осуществление изобретения
Созданные двойные промоторы являются химерными двойными промоторами, в которых промоторы генов hSurv и hTERT находятся в двух положениях относительно друг друга. В первом случае промотор гена hSurv расположен выше промотора hTERT относительно старт-кодона подконтрольного гена, во втором случае промотор гена hTERT расположен выше промотора hSurv относительно старт-кодона подконтрольного гена. Созданные промоторы обладают высокой активностью в раковых клетках человека, причем активность промоторов превышает активность промотора вируса SV40. Промоторы могут иметь функциональные производные, то есть модификации промоторов как результат делеции, замены, инсерции или иной мутации в исходном промоторе, не изменяющие или сохраняющие достаточно высокий уровень активности по сравнению с исходным промотором (примеры 2 и 3, таблица 2).
Полученные промоторы могут быть использованы для преимущественной транскрипции различных генов в раковых клетках в составе экспрессионной кассеты. Экспрессионная кассета (ЭК), предназначенная для экспрессии терапевтических генов в клетках млекопитающих, представляет собой фрагмент ДНК, содержащий все необходимые генетические элементы для экспрессии внедренной в него генетической информации. Экспрессионная кассета, в направлении от 5′ к 3′-концу, состоит из 1) промотора (промоторов); 2) одного или нескольких генов, экспрессию которых предполагается обеспечить; 3) сигнала полиаденилирования, необходимого для окончания транскрипции и посттранскрипционного процессинга РНК. В состав ЭК могут входить также дополнительные элементы, обеспечивающие регулировку условий экспрессии интересующего гена. Так, для усиления экспрессии терапевтического гена иногда используется химерный интрон, расположенный между последовательностями промотора и терапевтического гена (Gross MK and Kainz MS et al. (1987) Mol Cell Biol 7:4576-81, Buchman AR and Berg P (1988) Mol Cell Biol 8:4395-405). Используемый сигнал полиаденилирования также может оказывать эффект на уровень экспрессии гена путем влияния на устойчивость синтезируемого транскрипта (Azzoni AR and Ribeiro SC (2007) J Gene Med 9:392-402).
В качестве генов, подконтрольных опухолеспецифичным промоторам в составе экспрессионных кассет, обычно используют гены терапевтического вмешательства следующих групп:
1. Негативные доминантные мутанты (НДМ) генов, участвующих в онкогенезе
2. Гены-убийцы
3. Гены - ингибиторы ангиогенеза
4. Гены-онкосупрессоры
5. Гены-иммуностимуляторы
6. Гены siRNA
1. Негативные доминантные мутанты (НДМ) генов используются для подавления действия белков генов, участвующих в онкогенезе, таких как онкогены или ингибиторы апоптоза. Так, например, введение НДМ гена сурвивина мыши в раковые клетки простаты ингибирует их рост (Pan L and Peng XC et al. (2011) J Cancer Res Clin Oncol 137:19-28). Введение НДМ гена с-Jun в опухоль толстой кишки приводит к ее частичной регрессии (Suto R and Tominaga К et al. (2004) Gene Ther 11:187-93). НДМ онкогена RAS подавляет развитие ряда линий раковых клеток человека, такие как рак поджелудочной железы, толстой кишки, языка.
2. Генами-убийцами являются гены токсинов (I тип), а также гены ферментов, способных превращать нетоксичный агент (пролекарство) в токсин, убивающий раковые клетки (II тип). Наиболее эффективными генами II типа являются гены тимидинкиназы вируса простого герпеса (HSVtk) и цитозиндезаминазы дрожжей или Е.coli. Цитозиндезаминаза - фермент, катализирующий гидролитическое дезаминирование 5-фторцитозина с образованием 5-фторурацила (5-ФУ), убивающего раковые клетки. Противоопухолевая активность комбинации цитозиндезаминаза/5-фторурацил была показана на животных с таким опухолями, как: фибросаркома (Mullen CA and Coale MM et al. (1994) Cancer Res 54:1503-6), карцинома (Huber BE and Austin EA et al. (1994) Proc Natl Acad Sci US A 91:8302-6, Bentires-Alj M and Hellin AC et al. (2000) Cancer Gene Ther 7:20-6, Huber BE and Austin EA et al. (1993) Cancer Res 53:4619-26, Ohwada A and Hirschowitz EA et al. (1996) Hum Gene Ther 7:1567-76, Kanai F and Lan KH et al. (1997) Cancer Res 57:461-5), глиома (Ichikawa Т and Tamiya Т et al. (2000) Cancer Gene Ther 7:74-82, Ge К and Xu L et al. (1997) Int J Cancer 71:675-9) и другие.
Тимидинкиназа вируса простого герпеса способна превращать нетоксичный противогрибковый агент - Ганцикловир (GCV) в токсичные метаболиты, которые встраиваясь в растущую цепь ДНК раковых клеток во время деления, ингибируют ее синтез, тем самым обеспечивая гибель раковой клетки. HSVtk/GCV - единственная генно-терапевтическая комбинация, достигшая третьей фазы клинических испытаний (Immonen A and Vapalahti M et al. (2004) Mol Ther 10:967-72, Evrard A and Cuq P et al. (1999) Int J Cancer 80:465-70). В фазе I клинических испытаний восьми пациентам с местными рецидивами рака простаты, предварительно прошедшим гормональную терапию, вводили аденовирус, несущий экспрессионную кассету с геном HSVtk. Последующее введение GCV приводило к значительной частичной регрессии опухоли (Nasu Y and Saika Т et al. (2007) Mol Ther 15:834-40).
Исследования фазы II клинических испытаний, в которой в качестве вектора для HSVtk был использован репликативно дефектный аденовирус, показали, что совместное применение стандартной и генотерапии приводит к статистически значимому увеличению средней выживаемости: с 38 до 62 недель (Immonen A and Vapalahti M et al. (2004) Mol Ther 10:967-72).
3. Экспрессия генов - ингибиторов ангиогенеза в клетках опухоли приводит к подавлению образования в ней новых кровеносных сосудов в опухоли ведет к замедлению ее роста и деградации из-за недостаточного поступления питательных веществ и кислорода. Ингибиторами ангиогенеза являются гены белков ангиостатина (подавляет развитие лимфомы EL-4) и эндостатина (рост карциномы почки, меланомы, колоректальной карциномы) (Sun X and Kanwar JR et al. (2001) Gene Ther 8:638-45, Shi W and Teschendorf C et al. (2002) Cancer Gene Ther 9:513-21, Cichon Т and Jamrozy L et al. (2002) Cancer Gene Ther 9:771-7).
4. Гены-онкосупрессоры играют ключевую роль в поддержании стабильности клеточного генома и в регуляции клеточного деления и апоптоза (запрограммированная клеточная гибель). Клетки опухоли подавляют работу генов-онкосупрессоров и т.о. избегают апоптоза. Введение генов-онкосупрессоров в раковые клетки приводит к подавлению их роста и гибели. Белок р53 является самым известным опухолевым супрессором. Первый в мире генно-терапевтический препарат ″Gendicine ™″ (Chinese Shenzhen SiBiono Genetechnologies), созданный на основе нереплицирующегося аденовируса, который несет ген р53, был введен в клиническую практику в 2003 году в Китае и используется для лечения плоскоклеточного рака головы и шеи.
5. Гены-иммуностимуляторы способны активировать клетки иммунной системы. Предполагается, что введение данных генов в раковые клетки обеспечивает стимулирование иммунной системы к узнаванию опухолевых клеток. Примерами генов-иммуностимуляторов являются IL-1, IL-2, IL-4, IL-6, TNF, GM-CSF, гамма-интерферон.
Так, например, использование GM-CSF в комплексе с суицидальными генами приводит к увеличению эффективности терапии онкологических заболеваний. В ряде работ показано, что регрессия опухоли значительно возрастает при совместном использовании гена-убийцы и GM-CSF (Hamilton JA (2002) Trends Immunol 23:403-8, Hamilton JA and Anderson GP (2004) Growth Factors 22:225-31). Более того, в некоторых исследованиях продемонстрировано развитие специфического противоопухолевого иммунитета после совместной экспрессии HSVtk и GM-CSF в клетках опухоли (Guo SY and Gu QL et al. (2003) World J Gastroenterol 9:233-7).
6. Введение в раковые клетки генов siRNA позволяет подавлять экспрессию онкогенов и как следствие индуцирует клеточную смерть. Например, введение в раковые клетки siRNA к гену сурвивина человека в составе рекомбинантной плазмиды подавляет экспрессию гена сурвивина, стимулирует апоптоз раковых клеток и подавляет их пролиферацию.
Для транспортировки терапевтического гена в составе экспрессионной кассеты внутрь раковой клетки используют экспрессионные векторы.
Экспрессионный вектор представляет собой молекулу нуклеиновой кислоты, способную транспортировать экспрессионную кассету внутрь клетки. Обычно, экспрессионный вектор - это плазмида или другая форма ДНК или РНК, обладающая способностью к автономной репликации в определенных клетках. Векторная молекула должна обладать некоторыми свойствами: 1) вектор должен длительное время существовать в популяции клеток-хозяев, т.е. реплицироваться автономно или вместе с хромосомами клеток; 2) в любом векторе должны быть биохимические или генетические маркеры, которые позволяли бы обнаруживать его присутствие в клетках; 3) структура векторной молекулы должна допускать встраивание в нее чужеродной последовательности нуклеотидов без нарушения ее функциональной целостности.
В целях генной терапии используется несколько типов векторов, отличаемых по способу доставки генетического материала. Так, векторы бывают вирусными и невирусными. В настоящее время способ доставки ДНК в раковые клетки с помощью вирусов является наиболее распространенным. Так, 76% генно-терапевтических испытаний проводят с разного рода вирусными векторами и 24% - с невирусными. Вирусный вектор - это такой тип вектора, в котором для доставки и реализации заложенной в него генетической информации используется генетический аппарат какого-либо вируса. По типу используемых вирусных векторов наиболее часто используются ретровирусные, аденовирусные, аденоассоциированные и герпесвирусные векторы (Balicki D and Beutler E (2002) Medicine (Baltimore) 81:69-86, Kovesdi I and Brough DE et al. (1997) Curr Opin Biotechnol 8:583-9).
Для репликации и упаковки аденовирусной ДНК необходимы только небольшие последовательности на обоих концах, содержащие области начала репликации, и последовательность, специфичная для упаковки ДНК. Поэтому почти весь вирусный геном можно заместить чужеродной ДНК, выращивая гибридный вирус в присутствии вируса-помощника. Одним из недостатков подобной системы является определенная трудность при избавлении от вируса-помощника. При использовании ретровирусов обычно используются вирусы, в которых удалены гены структурных белков и ревертазы. Наиболее прогрессивными в качестве систем доставки являются векторы на основе аденоассоциированных вирусов, эти вирусы не вызывают заболеваний у людей и потому являются весьма перспективными с точки зрения безопасности использования. К минусам этой системы нужно отнести малую емкость для введения генетической информации которая ограничивается 4800 п.о. Достоинством вирусных систем является их высокая эффективность. Недостатком - иммуногенность, а также сложность и высокая себестоимость производства (Prestwich RJ and Errington F (2008) Clin Med Oncol 2:83-96).
В качестве вектора для невирусной доставки обычно используются плазмиды, которые можно нарабатывать в больших количествах в бактериальных культурах.
Плазмида должна иметь специальные последовательности, необходимые для ее наработки вне организма, - точку начала репликации, ген устойчивости к антибиотику, сайт множественного клонирования (MCS). В отличие от рекомбинантных вирусов, плазмиды довольно просто конструировать и нарабатывать в больших количествах. Кроме того, плазмиды обеспечивают довольно высокий уровень безопасности при использовании по сравнению с вирусными векторами (Williams PD and Kingston PA (2011) Cardiovasc Res 91:565-76).
Кроме плазмидных и вирусных систем в последнее время появляются другие системы доставки. Так, например, предложены системы бактериальной доставки, которые используют натуральное свойство бактерий накапливаться в раковых клетках.
Для доставки плазмидных векторов в клетки опухоли используется два подхода.
Первый - это заключение плазмиды в искусственные липидные везикулы - липосомы, способные проникать через плазматическую мембрану. Второй путь заключается в создании комплекса ДНК с положительно заряженным носителем, например полиэтиленимином или поликатионными липосомами, однако при этом образуются большие по размеру комплексы, которые достаточно сложно доставить внутрь клетки (Wagstaff KM and Jans DA (2007) Biochem J 406:185-202).
Также для доставки используют пептиды/белки, проникающие в клетки (СРР). Данные пептиды/белки при образовании комплекса с ДНК обеспечивают ее прямую доставку в ядра клеток (Wagstaff KM and Jans DA (2006) Curr Med Chem 13:1371-87, Gupta В and Levchenko TS et al. (2005) Adv Drug Deliv Rev 57:637-51, Morris MC and Chaloin L et al. (2000) Curr Opin Biotechnol 11:461-6, Wagstaff KM and Jans DA (2007) Biochem J 406:185-202, WagstaffKM and Fan JY et al. (2008) FASEB J 22:2232-42). CPP не обладают выраженными антигенными свойствами и способны переносить в клетку большие по размеру молекулы ДНК (Balicki D and Putnam CD et al. (2002) Proc Nati Acad Sci USA 99:7467-71). Для доставки ДНК в ядра клеток могут быть использованы белки-гистоны. Наилучшей активностью в отношении экспрессии доставленных генов, по имеющимся данным, обладают гистоны Н2А и Н2В (Kaouass M and Beaulieu R et al. (2006) J Control Release 113:245-54). Большим преимуществом гистонов перед другими системами доставки генов является их низкая токсичность.
В целом, для эффективной невирусной доставки вектор должен обладать комбинацией таких свойств, как способность образовывать комплекс с ДНК, конденсировать ДНК в более компактное состояние, защищать ее от действия нуклеаз, обеспечивать эффективное прохождение через клеточные и ядерные мембраны и при этом не ингибировать транскрипцию ДНК. Генные противораковые препараты могут вводиться пациенту внутривенно, внутрибрюшинно, внутриопухолево, подкожно или внутримышечно в зависимости от локализации опухоли и распространенности заболевания. Примером эффективного использования внутриопухолевого введения генетических конструкций может служить испытание аденовирусного препарата, содержащего гены бактериальной цитозиндезаминазы и тимидинкиназы вируса простого герпеса - Ad5-CD/TKrep. Препарат вводили в простату и затем проводили терапию 5-фторцитозином и ганцикловиром (Freytag SO and Khil M et al. (2002) Cancer Res 62:4968-76). Для лечения опухоли простаты также использовали подкожное и внутривенное введение генетических конструкций (Small EJ and Carducci MA et al. (2006) Mol Ther 14:107-17) (http://www.asco.org/ASCOv2/Meetings/Abstracts?&vmview=abst_detail_view&confID=26&ab stractID=890).
При лечении злокачественных плевральных мезотелиом и рака легких используют внутриплевральное введение конструкций (Sterman DH and Recio A et al. (2005) Clin Cancer Res 11:7444-53, Tan Y and Xu Metal. (1996) Anticancer Res 16:1993-8).
Часто, генно-терапевтический препарат представляет собой лиофилизованную ДНК, поскольку ДНК в виде лиофилизованного порошка является наиболее удобной для хранения и использования в качестве инъекций. Для предотвращения потери трансфекционной активности ДНК в процессе ее лиофилизации используют моно и дисахариды, такие, как лактоза, глюкоза и сахароза в качестве добавок к раствору плазмидной ДНК перед процессом лиофилизации. Сахара также используют при лиофилизации комплексов ДНК и поликатионов.
Таким образом, генно-терапевтическая фармацевтическая композиция должна содержать: 1) экспрессионную кассету в составе экспрессионного вектора, несущую терапевтический ген под контролем промотора; 2) носитель для обеспечения проникновения экспрессионного вектора, содержащего экспрессионную кассету, через мембрану клетки. Носителем может выступать вирус. В случае невирусного носителя он должен быть положительно заряженным (липосомы, полиэтиленимин и другие поликатионы). Вспомогательными компонентами в генно-терапевтической фармацевтической композиции могут быть сахара (моно и дисахариды), буферные растворы, антибактериальные агенты и другие.
Примеры
Приведенные ниже примеры предназначены исключительно для иллюстрации отдельных предпочтительных воплощений, но не должны рассматриваться в качестве ограничивающих объем настоящего изобретения.
Пример 1. Создание конструкции PhSurv269-pGL3
Фрагмент 269 п.о. проксимальной промоторной области гена BIRC5 (сурвивин - Surv) амплифицировали с помощью ПЦР, используя в качестве матрицы геномную ДНК мозга человека и синтезированные праймеры (pSurv269For -BglII CGGAGATCTCGCGTTCTTTGAAAGCAGTCGA, pSurv269Rev-HmdIII CCCAAGCTTGCCGCCGCCGCCACCTCTG), фланкирующие фрагмент 269 п.о. и содержащие сайты узнавания рестриктаз BglII и HindIII (Fermentas, Канада). Полученный ПЦР-продукт промежуточно клонировали в вектор pAL-TA (Евроген, Россия) и полученную промежуточную конструкцию PhSurv269-pAL-TA секвенировали для исключения ошибок работы Taq-полимеразы (Евроген, Россия). Далее промежуточную конструкцию PhSurv269-pAL-TA обрабатывали рекстриктазами BglII/HindIII, требуемую вставку 269 п.о. выделяли из агарозного геля и клонировали в вектор pGL3, несущий репортерный ген люциферазы светлячка, также обработанный рекстриктазами BglII/HindIII. Далее определяли нуклеотидную последовательность конечной конструкции PhSurv269-pGL3 для исключения нуклеотидных замен (Фиг.1).
Пример 2. Создание конструкции PhSurv269+130-pGL3, функционального производного промотора PhSurv269, модифицированного добавлением инсерции
Был создан промотор PhSurv269+130, являющийся функциональным производным промотора PhSurv269. Модифицированный промотор отличается от исходного промотора PhSurv269 наличием инсерции размером в 130 п.о. Инсерция расположена в проксимальном положении относительно 5′-конца промотора PhSurv269 и представляет собой фрагмент+269/+399 последовательности нативного промотора сурвивина человека. Инсерция располагается в ″прямом″ положении, т.е. в той же ориентации, что и в нативном промоторе.
Фрагмент проксимальной промоторной области гена BIRC5 (сурвивина, Surv) амплифицировали с помощью ПЦР, используя в качестве матрицы геномную ДНК мозга человека и синтезированные праймеры (pSurv269+130For-BglII CATAGATCTATTTTTAGTAGAGACAAGGTTTCACCGTG и pSurv269+130Rev-HindIII CCCAAGCTTGCCGCCGCCGCCACCTCTG), содержащие сайты узнавания рестриктаз BglII и HindIII (Fermentas, Канада). Полученный ПЦР-продукт промежуточно клонировали в вектор pAL-TA (Евроген, Россия) и полученную промежуточную конструкцию PhSurv269+130-pAL-TA секвенировали для исключения ошибок работы Taq-полимеразы (Евроген, Россия). Далее промежуточную конструкцию PhSurv269+130-pAL-TA обрабатывали рестриктазами BglII/HindIII, требуемую вставку выделяли из агарозного геля и клонировали в векторе pGL3, несущем репортерный ген люциферазы светлячка, также обработанный рекстриктазами BglII/HindIII. Далее определяли нуклеотидную последовательность конечной конструкции PhSurv269+130-pGL3 для исключения нуклеотидных замен.
Пример 3. Сравнение активности PhSurv269+130, модифицированного добавлением инсерции, с немодифицированным промотором PhSurv269
Для сравнения активности функционального производного промотора PhT269 (PhT269+130) и его немодифицированного варианта PhT269 были использованы следующие раковые клеточные линии различного генеза: Calu-1 (карцинома легкого человека), А375 (меланома кожи человека), А549 (аденокарцинома легкого человека), PANC-1 (карцинома поджелудочной железы человека), HepG2 (гепатоцеллюлярная карцинома человека), LLC (эпидермоидная карцинома легкого Льюис).
Эксперимент проводили, как описано в Примере 9.
Относительная активность исследованных промоторов в разных клеточных линиях приведена в таблице 1. Как видно из таблицы 1, активности промоторов PhT269 и PhT269+130 практически совпадают во всех исследованных клеточных линиях, при этом активности промоторов PhT269 и PhT269+130 различаются в разных клеточных линиях, но в каждой клеточной линии активности этих промоторов практически одинаковы. Таким образом, функциональное производное промотора PhT269 обладает такой же активностью, как и исходный промотор PhT269.
Полученные данные позволяют заключить, что возможно использование функциональных производных промоторов, характеризующихся практически одинаковой или достаточно высокой активностью по сравнению с исходным промотором.
Пример 4. Конструирование экспрессионных векторов на основе двойных промоторов PhTSurv269 и PhSurv269T
Фрагмент промотора гена сурвивина человека, PhSurv269 (SEQ ID NO: 1) длиной 269 п.о. клонировали в конструкцию PhTERT-pGL3, полученную на основе плазмиды pGL3-basic (Promega, США), несущую фрагмент промотора обратной транскриптазы теломеразы человека (PhTERT) и репортерный ген люциферазы светлячка. Промотор PhSurv269 вставляли в конструкцию PhTERT-pGL3 в прямой ориентации в двух положениях относительно промотора PhTERT (SEQ ID NO:2). Для этого получали кДНК промоторной области гена сурвивина человека длиной 269 п.о. в результате гидролиза плазмиды PhSurv269-pGL3 ферментами рестрикции BglII и HindIII с последующей обработкой фрагментом Кленова. Затем полученный фрагмент PhSurv269 лигировали с вектором PhTERT-pGL3, предварительно линеаризованным по сайту узнавания рестриктазой HindIII, либо по сайту узнавания рестриктазой XhoI и обработанным большой субъединицей ДНК полимеразы I E.coli (Fermentas, Канада). Таким образом, в первом случае был получен вектор PhTSurv269-pGL3, несущий ген люциферазы под контролем двойного промотора PhTSurv269, где промотор PhTERT находится выше промотора PhSurv269 относительно старт-кодона гена люциферазы светлячка. Во втором случае получили вектор PhSurv269T-pGL3, в котором промотор PhSurv269 находится выше промотора PhTERT относительно старт-кодона гена люциферазы. Структура полученных экспрессионных кассет проиллюстрирована на Фиг.2.
Пример 5. Конструирование экспрессионных векторов на основе двойных промоторов PhTmSurv и PmSurvhT
Фрагмент промотора гена сурвивина мыши (PmSurv) длиной 197 п.о. (SEQ ID NO:3), клонировали в конструкцию PhTERT-pGL3, созданную на основе плазмиды pGL3-basic (Promega, США) ранее, несущую промотор обратной транскриптазы теломеразы человека (PhTERT) и репортерный ген люциферазы светлячка. Промотор PmSurv клонировали в конструкцию PhTERT-pGL3 в прямой ориентации в двух положениях относительно промотора PhTERT. Для этого получали кДНК сурвивина мыши (pmSurv) в результате гидролиза плазмиды PmSurv-pGL3 ферментами рестрикции HindIII с последующей обработкой фрагментом Кленова и NoiI (Fermentas, Канада). Затем полученный фрагмент, содержащий кДНК гена сурвивина мыши (420 п.о.), лигировали с вектором PhTERT-pGL3, предварительно линеаризованным по сайту узнавания рестриктазой XhoI и обработанным большой субъединицей ДНК полимеразы I Е.coli, а затем обработанным рестриктазой NotI. Таким образом, был получен вектор PmSurvhT-pGL3, несущий ген люциферазы под контролем двойного промотора PmSurvhT, где промотор PhTERT находится ниже промотора PmSurv относительно старт-кодона гена люциферазы светлячка.
Для получения плазмиды, несущей ген люциферазы под контролем двойного промотора PhTmSurv, где промотор PhTERT находится выше промотора pmSurv относительно старт-кодона гена люциферазы светлячка, получали кДНК обратной транскриптазы теломеразы человека (PhTERT) длиной 243 п.о. в результате гидролиза плазмиды PhTERT-pGL3 ферментами рестрикции HindIII с последующей обработкой фрагментом Кленова и NotI. Полученный фрагмент, содержащий PhTERT (437 п.о.), лигировали с вектором pmSurv-pGL3, предварительно обработанным ферментами рестрикции BglII с последующей обработкой фрагментом Кленова и NotI. Таким образом, был получен вектор PhTmSurv-pGL3, несущий ген люциферазы под контролем двойного промотора PhTmSurv, где промотор PhTERT находится выше промотора PmSurv относительно старт-кодона гена люциферазы светлячка.
Пример 6. Конструирование экспрессионных суицидальных векторов на основе двойного раковоспецифичного промотора PhTSurv269: PhTSurv269-HSVtk и PhTSurv269-HSVtk-mGM-CSF
кДНК гена тимидинкиназы вируса простого герпеса типа 1 (HSVtk) клонировали в конструкцию PhTSurv269-pGL3 на основе плазмиды pGL3-basic (Promega, США), несущую двойной промотор, состоящий из промоторного региона гена обратной транскриптазы теломеразы человека (hTERT) и укороченного промоторного региона гена сурвивина человека (PhSurv269, координаты -268+1 [где +1 аденин триплета, кодирующего инициирущий метионин]). кДНК гена HSVtk вставляли в конструкцию PhTSurv269-pGL3 в прямой ориентации. Для этого вектор PhTSurv269-pGL3 обрабатывали рестриктазами NcoI/BamHI (Fermentas, Канада), в результате из вектора была полностью удалена последовательность кДНК гена люциферазы светлячка. Плазмиду PhSurv-HSVtk-pGL3, полученную ранее, обрабатывали рестриктазами NcoI/BamHI. Затем полученный фрагмент, содержащий последовательность HSVtk, лигировали с линеаризованным вектором PhTSurv269-pGL3. В результате была получена конструкция PhTSurv269-HSVtk-pGL3. Последовательность полученной конструкции подтверждалась методом секвенирования по Сэнгеру. Структура полученной экспрессионной кассеты проиллюстрирована на Фиг.3.
Вставку HSVtk-IRES-mGM-CSF, содержащую ген тимидинкиназы вируса простого герпеса типа 1 (HSVtk), IRES (участок посадки рибосомы вируса энцефаломиокардита человека) и ген mGM-CSF (гранулоцитарно-макрофагальный колониестимулирующий фактор мыши), клонировали в конструкцию PhTSurv269-pGL3. Указанный фрагмент клонировали в конструкцию PhTSurv269-pGL3 в прямой ориентации. Для этого вектор PhTSurv269-pGL3 линеаризовали с помощью обработки рестриктазой BamHI в сочетании с фрагментом Кленова и далее рестриктазой NcoI. Фрагмент HSVtk-IRES-mGM-CSF был получен при гидролизе плазмиды CMV-HSVtk-IRES-mGM-CSF-pGL3 эндонуклеазами рестрикции NcoI/XhoI с ″затуплением″ остатка сайта XhoI большой субъединицей Кленова. Полученный фрагмент ДНК был лигирован в PhTSurv269-pGL3. В результате была получена конструкция PhTSurv269-HSVtk-IRES-mGM-CSF-pGL3. Последовательность полученной конструкций подтверждалась методом секвенирования по Сэнгеру. Структура полученной экспрессионной кассеты проиллюстрирована на Фиг.4.
Пример 7. Создание экспрессионных векторов, несущих генную кассету FCU1-mGM-CSF под контролем двойного промотора PhTSurv269
Конструкция PhTSurv269-FCU1-mGM-CSF состоит из: 1) промотора PhTSurv269; 2) химерного интрона длиной 133 п.о., расположенного между промотором и геном FCU1; 3) кДНК гена FCU1 (фрагмент гена цитозиндезаминазы (FCY1) и фрагмента гена урацил-фосфорибозилтрансферазы (UPRT, FUR1) из организма Saccharomyces cerevisiae, координаты 1002-2120 п.о.) ; 4) IRES (участок посадки рибосомы вируса энцефаломиокардита человека) и 5) кДНК гена mGM-CSF
Для получения конструкции PhTSwv269-FCU1-mGM-CSF-pGL3 плазмиду pCMV-FCU1-pGL3, полученную ранее, линеаризовали посредством обработки рестриктазами HindIII (с последующей обработкой фрагментом Кленова) и NotI (Fermentas, Канада). Вставка PhTSurv269 была получена при обработке вектора PhTSurv269-pGL3 рестриктазами NcoI (с последующей обработкой фрагментом Кленова) и NotI (Fermentas, Канада). Затем полученный фрагмент, содержащий промотор PhTSurv269, дотировали с линеаризованным вектором CMV-FCU1-pGL3. В результате была получена конструкция PhTSurv269-FCU1-pGL3. Последовательность полученной конструкции подтверждалась методом секвенирования по Сэнгеру.
Целевую конструкцию PhTSurv269-FCU1-mGM-CSF (Фиг.5) получали следующим образом: плазмиду PhTSurv269-FCU1-pGL3, обрабатывали рестриктазой BamHI (Fermentas, Канада), в результате из вектора была удалена часть гена FCU1 и сигнал полиаденилирования. Вставка FCU1*(ahfuvtyn)-mGM-CSF была получена при обработке вектора pCMV-FCU1-IRES-mGM-CSF-pGL3 рестриктазой BamHI. Затем полученный фрагмент, содержащий фрагмент гена FCU1, IRES, mGM-CSF, и сигнал полиаденилирования дотировали с линеаризованным вектором PhTSurv269-FCU1-pGL3 (из которого вырезана часть гена FCU1 и сигнал полиаденилирования). При лигировании происходило восстановление целостного гена FCU1 (ген содержит в себе сайт рестрикции BamHI). В результате была получена конструкция PhTSurv269-FCU1-mGM-CSF-pGL3. Последовательность полученной конструкции подтверждалась методом секвенирования по Сэнгеру.
Пример 8. Создание конструкции PhTSurv269-Cre
Для усиления опухолеспецифической экспрессии гибридного гена цитозиндезаминазы/урацилфосфорибозилтрансферазы (FCU1) в клетках опухоли мы создали модифицированную систему Cre-LoxP. Были сконструированы два вектора: PhTSurv269-Cre-pQC (активатор) и CMV-LoxP-Stop-LoxP-FCU1-pGL3 (эффектор).
Первый, активационный вектор бинарной системы несет ген Сге-рекомбиназы с сигналом ядерной локализации, под контролем созданного нами опухолеспецифичного промотора PhTSurv269. Второй, эффекторный, или киллерный, вектор, содержит промотор цитомегаловируса (CMV), отделенный от гена FCU1 ″Stopw-CHraanoM, который состоит из тандема трех сигналов полиаденилирования вируса SV40. Кроме того, ″Stop″-сигнал с обеих сторон фланкирован находящимися в одной ориентации последовательностями, узнаваемыми белком Cre - LoxP-сайты. Принципиальная схема усиления экспрессии суицидального гена FCU1 с помощью Cre/LoxP системы приведена на Фиг.6.
Конструкция PhTSurv269-Cre была получена на основе плазмид PhSurv-Cre-pQXIX и PhTSurv269-pGL3, полученных ранее.
ДНК плазмиды PhSurv-Cre-pQXIX гидролизовали по сайтам узнавания рестриктазами BglII/NotI, затем обрабатывали большой субъединицей Кленова. С оставшейся векторной частью дотировали вставку PhTSurv269, полученную при гидролизе ДНК плазмиды PhTSurv269-pGL3 рестриктазами XhoI/NcoI с последующей обработкой фрагментом Кленова. Отбор клонов в правильной ориентации проводили методом ПЦР с использованием специфических праймеров, комплементарных гену Cre-рекомбиназы (CreSeq-Rev!) и промотору PhSurv269 (Surv269For). Правильность полученной конструкции подтверждали с помощью определения нуклеотидной последовательности с использованием специфических праймеров UP-L1, Surv269For и CreSeq-F2.
Пример 9. Определение активности универсальных опухолеспецифичных промоторов PhTSurv269 и PhSurv269T в раковых клетках
Для тестирования активности полученных промоторов были использованы следующие раковые клеточные линии различного генеза: Calu-1 (карцинома легкого человека), А375 (меланома кожи человека), А549 (аденокарцинома легкого человека) НТ1080 (фибросаркома человека), PANC-1 (карцинома поджелудочной железы человека).
Эукариотические клетки культивировали при 37°С в СО2 инкубаторе для достижения 90% конфлюэнтности в 25 см2 культуральном флаконе. Далее готовили растворы, содержащие 9 мкг плазмиды из Примеров 1, 4 или 5 или контрольных плазмид BV-pGL3, PV-pGL3 и 1 мкг нормировочной плазмиды pRL-TK, несущей ген люциферазы Remlla reniformis, в 200 мкл бессывороточной среды Opti-MEM (Invitrogen, США). Далее к раствору плазмид добавляли 200 мкл раствора LFA2000 в Opti-MEM (25 мкл LFA+175 мкл Opti-MEM), инкубировали 5 мин при комнатной температуре. Эукариотические клетки инкубировали с полученными липоплексами в течение 3-4 часов в зависимости от клеточной линии.
Через 48 часов после введения в клетки одной из конструкций из Примеров 1,4 или 5, контрольных плазмид BV-pGL3, PV-pGL3 и нормировочной плазмиды pRL-TK измеряли активности люциферазы светлячка и R. reniformis в клеточных экстрактах с помощью набора реактивов Dual-Luciferase Reporter Assay System (Promega, США). Значения активности люциферазы светлячка нормировали относительно активности люциферазы R. reniformis плазмиды pRL-TK с целью снижения погрешности, связанной с различной эффективностью доставки экспрессионных конструкций в клетки в серии независимых экспериментов. С целью увеличения наглядности и дополнительного уменьшения погрешности эксперимента данные нормировались по активности контрольных конструкций BV-pGL3 и PV-pGL3.
Среднюю люциферазную активность и стандартную ошибку среднего рассчитывали по формуле Sx-bar=(s2/n)1/2, где s - совокупная дисперсия, n - число наблюдений.
В результате проведенного анализа было установлено, что активность двойных тандемных промоторов PhTSurv269 и PhSurv269T превосходит активности одиночных PhTERT и PhSurv269 промоторов во всех клеточных линиях кроме линии Calu-1. Относительная активность исследованных промоторов в разных клеточных линиях приведена на Фиг.7, 8 и в таблице 2. Как видно из таблицы 2, активности двойных промоторов PhTSurv269 и PhSurv269T примерно соответствуют сумме активностей отдельных промоторов. Особенно это соблюдается для PhTSurv269 тандема. Активность PhSurv269T тандема обычно немного ниже активности PhTSurv269 тандема.
Также было показано, что активность двойного промотора PmSurvhT в раковых клетках в 2-3 раза выше, чем активность одиночного промотора PmSurv в зависимости от типа клеточной линии (Фиг.7).
Полученные данные позволяют заключить, что использование тандемных промоторов для экспрессии генов предпочтительнее, чем использование каждого из одинарных промоторов, и PhTSurv269 тандем предпочтительнее PhSurv269T тандема по активности экспрессии в разных типах раковых клеток (Фиг.8).
Пример 10. Сравнение эффективности двойных промоторов PhSurv269T и PhTSurv269 с эффективностью двойных промоторов PhTS и PhST, сконструированных нами ранее
Для сравнения активности промоторов были использованы следующие раковые клеточные линии различного генеза: Calu-1 (карцинома легкого человека), A549 (аденокарцинома легкого человека) HT1080 (фибросаркома человека), PANC-1 (карцинома поджелудочной железы человека).
Эксперимент проводили как описано в Примере 9. На Фиг.9 видно, что во всех исследованных клеточных линиях активность промотора PhTSurv269 выше активности одиночных промоторов PhSurv, PhTERT, а также активности искусственных двойных промоторов PhTS и PhST.
Пример 11. Детекция гибридного белка цитозиндезаминазы/ урацилфосфорибозилтрансферазы (FCU1) с помощью вестерн-блот анализа в трансфицированных раковых клетках
На первом этапе проводили трансфекцию клеток линии S37 генно-инженерными конструкциями PhTSurv269-Cre//pCMV-STOP-FCU1 (котрансфекция векторами pCMV-STOP-FCU1 и PhTSurv269-Cre). Трансфекцию клеток проводили в культуральных флаконах Т-25 Липофектамином 2000 (″Invitrogen″, США) согласно рекомендациям производителя. Для трансфекции обычно использовали 10 мкг плазмидной ДНК. После проведения липофекции клетки культивировали в среде DMEM/F12 (1:1) без антибиотиков, содержащей 10% эмбриональной телячьей сыворотки в течение 48 часов, далее получали клеточные осадки для проведения вестерн-блот анализа.
В качестве вектора для сравнения мы использовали вектор pCMV-FCU1, в котором экспрессия гена FCU1 направлялась сильным конститутивным промотором pCMV. Через 48 часов после введения конструкций клетки открепляли от поверхности культуральных флаконов посредством раствора трипсина, затем суспендировали их в среде DMEM:F12 с антибиотиками и определяли количество клеток в полученной суспензии с помощью счетной камеры Neubauer, добавляя к аликвоте клеточной суспензии трипановый синий в соотношении 1:1. Далее центрифугировали 5 минут при 7°С и 160g, клеточные осадки двукратно промывали 1X фосфатно-солевым буфером (1,7 мМ КН2РО4, 5,2 мМ Na2HPO4,150 мМ NaCl) (см. ниже).
Далее клетки суспендировали в буфере для нанесения (концентрация клеток в буфере должна быть 107 кл/мл) и инкубировали 5 минут при 95°С.
Полученные лизаты клеток подвергали денатурирующему электрофорезу белков в полиакриламидном геле по методу Лэммли (Laemmli U.K. (1970) Nature, Aug 15;227(5259):680-5) с последующим переносом фракционированных белков из геля на мембрану. Количество белка FCU1 в лизатах (нормализованных по суммарному белку) трансфицированной клеточной линии анализировали при помощи вестерн-блот анализа. В качестве первичных антител использовали овечьи IgG (″Abeam″), связывающиеся с N-концевой областью белка FCU1. Вторичные антитела к овечьим IgG были конъюгированы с пероксидазой хрена (″Santa-Cruz″). Для контроля количества белка в клеточных экстрактах использовали окраску с антителами к актину. Результат вестерн-блот анализа приведен на Фиг.10.
Для того чтобы определить наиболее эффективное соотношение вектора-эффектора и вектора-активатора, мы провели транзиентные трансфекции векторами PhTSurv269-Cre и pCMV-STOP-FCU1 в соотношениях по количеству мкг 2:2 и 1:3. Максимальное количество ДНК, которое могло быть взято для трансфекции, составляет 4 мкг (согласно рекомендациям производителя). Выбранные соотношения эффекторного (pCMV-STOP-FCU1) и активаторного (PhTSurv269-Cre) векторов описывают две возможные ситуации: 2:2 - для эффективной работы системы необходимо равное количество эффекторного и активаторного векторов; 1:3 - для наибольшего усиления экспрессии гена FCU1 необходим избыток эффекторного гена.
В лизатах клеток S37, трансфицированных вектором pCMV-FCU1, количество белка FCU1 больше, чем в лизатах клеток тех же линий, котрансфицированных бинарной системой векторов PhTSurv269-Cre//pCMV-STOP-FCU1 в обоих использованных соотношениях, тем не менее соотношение активатор/эффектор 1:3 представляется несколько более предпочтительным с точки зрения продукции белка FCU1.
Пример 12. Определение цитотоксичности созданной системы векторов PhTSurv269-Cre//DCMV-STOP-FCU1
Для определения цитотоксичности системы PhTSurv269-Cre//pCMV-STOP-FCU1 измеряли чувствительность трансфицированных клеток S37 (саркома мыши) к 5-фторцитозину (5-ФЦ). Для количественной оценки чувствительности клеток определяли процент выживших клеток при различных концентрациях 5-ФЦ. Сравнив выживаемость клеток, трансфицированных вектором pCMV-FCU1 и трансфицированных бинарной системой векторов PhTSurv269-Cre//pCMV-STOP- FCU1, можно сделать выводы об эффективности конструкций с терапевтической точки зрения.
Клетки транзиентно трансфицировали следующими экспрессионными конструкциями: системой векторов PhTSurv269-Cre//pCMV-STOP-FCU1 (котрансфекция, соотношение векторов 2:2 и 1:3 по количеству мкг) и вектором pCMV-FCU1 (положительный контроль). Далее трансфицированные клетки в течение 120 ч инкубировали в среде с различными концентрациями 5-ФЦ (0, 50, 200, 500 и 1000 мкМ). В качестве контроля использовали нетрансфицированные клетки S37.
Выживаемость клеток оценивали при помощи MTS-теста согласно рекомендациям производителя (Promega, США). Эффективность трансфекции определяли, проведя FACS-анализ (Fluorescence-Activated Cell Sorting, FACS) клеток, трансфицированных репортерным вектором pEGFP.
В результате проведенных экспериментов было показано, что эффект системы векторов PhTSurv269-Cre//pCMV-STOP-FCU1 сравним с эффектом, вызываемым конструкцией pCMV-FCU1. При концентрации в среде для культивирования раковых клеток 5-фторцитозина 200 мкМ 90% раковых клеток погибает после обработки системой PhTSurv269-Cre//pCMV-STOP-FCU1 (Фиг, 11).
Пример 13. Сравнение цитотоксичности систем векторов Promoter-Cre//pCMV-STOP-FCU1, где Promoter - PhSurv или PhTSurv269
Для сравнения цитотоксичности систем PhTSurv269-Cre//pCMV-STOP-FCU1, PhSurv-Cre//pCMV-STOP-FCU1 клетки линии S37 трансфицировали полученными системами и контрольной конструкцией CMV-FCU1-pGL3, далее трансфицированные клетки обрабатывали раствором 5-фторцитозина различной концентрации и через 196 часов после обработки измеряли и определяли процент выживших клеток в каждой из групп.
Выживаемость клеток оценивали при помощи MTS-теста согласно рекомендациям производителя (Promega, США). Эффективность трансфекции определяли, проведя FACS-анализ (Fluorescence-Activated Cell Sorting, FACS) клеток, трансфицированных репортерным вектором pEGFP.
Показано, что суицидальный эффект системы векторов PhTSurv269-Cre//pCMV-STOP-FCU1 превосходит эффект системы PhSurv-Cre//pCMV-STOP-FCU1 и сравним с эффектом, вызываемым конструкцией pCMV-FCU1 (Фиг.12).
Пример 14. Определение транскрипционной активности двойных промоторов PhTSurv269, PhTmSurv
Клетки линии А375 (меланома кожи человека) трансфицировали конструкциями PhTSurv269-pGL3, PhTmSurv-pGL3 и контрольной конструкцией PhTSurv-pGL3, через 48 часов после введения конструкций клетки собирали и выделяли суммарную РНК с использованием набора RNeasy Mini Kit (Qiagen, США), затем обрабатывали ДНКазой I (Quiagen, США) согласно протоколам производителя. Полученную суммарную РНК использовали для синтеза кДНК с затравкой, состоящей из статистических гексамеров (Perkin Elmer, США). Данная кДНК использовалась для определения наличия транскриптов с промоторов PhTERT, PmSurv и PhSurv269 в составе тандемов. Для определения наличия транскриптов с промоторов была использована методика полуколичественного ОТ-ПЦР, описанная ранее (Pleshkan VV and Vinogradova TV et al. (2008) Biochim Biophys Acta 1779:599-605).
Для определения наличия транскриптов с дистального промотора PhTERT тандема подобрали общий праймер TSL-F, который находится в линкере сразу за исследуемым промотором PhTERT и парные к нему обратные праймеры для каждого из промоторов гена сурвивина: праймер hS269_128R для конструкции с промотором PhTSurv269, праймер mS_122R для конструкции с промотором PhTmSurv и праймер hSurv_150R для конструкции с промотором PhTS. Обратные праймеры находились в проксимальном промоторе на расстоянии не более 150 п.о. от его 5′-конца, заведомо до сайтов инициации транскрипции проксимального промотора (Фиг.13). Полученные в результате проведения полимеразной цепной реакции продукты анализировали с помощью электрофоретического разделения в агарозном геле. Для каждой конструкции было проанализировано не менее трех независимых результатов ОТ-ПЦР.
В результате проведенного анализа было показано, что в конструкциях PhTSurv269-pGL3 и PhTmSurv-pGLS работает дистальный PhTERT промотор, в то время как транскрипты дистального PhTERT промотора в конструкции PhTS-pGL3 не детектируются (Фиг.14, панель ″дистальный″).
Для определения наличия транскриптов, инициированных с проксимального и/или обоих промоторов тандема, использована другая пара праймеров. Использовали прямой UPF праймер, расположенный сразу за проксимальным промотором и обратный к нему праймер Luc_202R. Транскрипты были детектированы как в случае конструкций PhTSurv269-pGL3 и PhTmSurv-pGL3, так и в случае контрольной конструкции PhTS-pGL3 (Фиг.14, панель ″проксимальный″).
Таким образом, мы показали, что в исследованных конструкциях PhTSurv269-pGL3 и PhTmSurv-pGL3 работают оба промотора.
Пример 15. Функциональный тест на цитотоксичность HSVtk, контролируемого двойным тандемным промотором PhTSurv269, при последующей обработке ганцикловиром
Эукариотические клетки трансфицировали в шестилуночных планшетах конструкцией PhTSurv269-HSVtk-mGM-CSF-pGL3 и контрольной конструкцией PhSurv-HSVtk-mGM-CSF-pGL3 с помощью Липофектамина-2000 (Invitrogen, США). Спустя 48 часов после трансфекции к клеткам добавляли раствор ганцикловира (″Cymeven, Roche″, Швейцария) в среде DMEM/F12 (1:1) с антибиотиками в концентрации 0, 12,5, 50 мкМ. Через 192 часа после добавления ганцикловира проводили MTS-тест с целью определения количества живых клеток (Promega, США). Результаты проведенного эксперимента приведены на Фиг.15. На клеточной линии НТ1080 было показано, что выживаемость контрольной группы клеток (нетрансфицированные клетки) при концентрации ганцикловира от 0 до 50 мкМ была около 100%, выживаемость клеток, трансфицированных конструкцией PhTSurv269-HSVtk-mGM-CSF-pGL3, была в каждой из групп приблизительно в 2 раза ниже, чем выживаемость клеток, трансфицированных конструкцией PhSurv-HSVtk-mGM-CSF-pGL3.
Таким образом, в данном эксперименте показано, что использование двойного тандемного промотора PhTSurv269 для контроля терапевтических генов более перспективно, чем использование полноразмерного промотора сурвивина.
Пример 16. Функциональный тест на цитотоксичность FCU1, контролируемого двойным тандемным промотором PhTSurv269, при последующей обработке 5-фторцитозином
Эукариотические клетки линии Calu-1 трансфицировали в 25 см2 флаконах конструкцией PhTSwv269-FCU1-mGM-CSF-pGL3 и контрольными конструкциями pCMV-FCU1-mGM-CSF-pGL3 и PhSurv-FCU1-mGM-CSF-pGL3 с помощью Липофектамина-2000 (Invitrogen, США). Спустя 48 часов после трансфекции к клеткам добавляли раствор 5-фторцитозина (″Sigma-Aldrich″, США) в среде DMEM/F12 (1:1) с антибиотиками в концентрации 0, 10, 50, 200, 500 мкМ. Через 144 часа после добавления 5-фторцитозина проводили MTS-тест с целью определения количества живых клеток (Promega, США). Результаты проведенного эксперимента приведены на Фиг.16. Было показано, что выживаемость контрольной группы клеток (нетрансфицированные клетки) при концентрации 5-фторцитозина от 0 до 500 мкМ была около 100%, выживаемость клеток, трансфицированных конструкцией PhTSurv269-FCU1-mGM-CSF-pGL3, была ниже, чем выживаемость клеток, трансфицированных конструкцией PhSurv- FCU1-mGM-CSF-pGL3.
Таким образом, использование двойного тандемного промотора PhTSurv269 для контроля терапевтических генов более предпочтительно, чем использование полноразмерного промотора сурвивина (PhSurv). В сочетании с таким свойством двойного тандемного промотора PhTSurv269, как более широкая опухолевая специфичность, это особенно актуально.
Пример 17. Определение количества синтезируемого цитокина GM-CSF под контролем двойного тандемного промотора PhTSurv269
Эукариотические клетки трансфицировали в шестилуночных планшетах конструкцией PhTSurv269-HSVtk-mGM-CSF-pGL3 и контрольной конструкцией PhSurv-HSVtk-mGM-CSF-pGL3 с помощью Липофектамина-2000 (Invitrogen, США). Спустя 48 часов после трансфекции собирали кондиционированную среду для определения количества белка GM-CSF.
Определение продукции GM-CSF в трансфицированных клетках проводили методом иммуноферментного анализа культуральной жидкости при помощи коммерческого набора ELISA производства R&D Systems (США). Результаты эксперимента приведены в таблице 3. Было показано, что под контролем промотора PhTSurv269 при использовании конструкции PhTSurv269-HSVtk-mGM-CSF-pGL3 в зависимости от клеточной линии синтезируется в 2,5-4 раза больше белка GM-CSF, чем под контролем промотора PhSurv при использовании конструкции PhSurv-HSVtk-mGM-CSF-pGL3.
Пример 18. Определение количества синтезируемого цитокина GM-CSF конструкции PhTSurv269-FCU1-mGM-CSF-pGL3
Эукариотические клетки линии Calu-1 трансфицировали конструкциями PhTSurv269-FCU1-mGM-CSF-pGL3, PhSurv-FCU1-mGM-CSF-pGL3 и контрольной конструкцией CMV-FCU1-mGM-CSF-pGL3 с помощью Липофектамина-2000 (Invitrogen, США). Спустя 48 часов после трансфекции собирали кондиционированную среду для определения количества белка mGM-CSF.
Определение продукции mGM-CSF в трансфицированных клетках проводили методом иммуноферментного анализа культуральной жидкости при помощи коммерческого набора ELISA производства R&D Systems (США). Результаты эксперимента приведены в Таблице 4. Было показано, что под контролем промотора PhTSurv269 при использовании конструкции PhTSurv269-FCU1-mGM-CSF-pGL3 в зависимости от клеточной линии синтезируется в 2,6 раза больше белка GM-CSF, чем под контролем промотора PhSurv при использовании конструкции PhSurv-FCU1-mGM-CSF-pGL3.
Пример 19. Определение количества HSVtk, синтезируемого конструкциями PhTSurv269-HSVtk-mGM-CSF-pGL3 и PhSurv-HSVtk-mGM-CSF-pGL3
На первом этапе работы клетки трансфицировали в культуральных флаконах Т-25 Липофектамином 2000 (″Invitrogen″, США) согласно рекомендациям производителя. Для трансфекции обычно использовали 10 мкг плазмидной ДНК. После проведения липофекции клетки культивировали в среде DMEM/F12 (1:1) без антибиотиков, содержащей 10% эмбриональной телячьей сыворотки в течение 48 часов, далее клетки открепляли от поверхности культуральных флаконов посредством раствора трипсина, затем суспендировали их в среде DMEM:F12 с антибиотиками и определяли количество клеток в полученной суспензии с помощью счетной камеры Neubauer, добавляя к аликвоте клеточной суспензии трипановый синий в соотношении 1:1. Затем центрифугировали 5 минут при 7°С и 160g, клеточные осадки двукратно промывали IX фосфатно-солевым буфером (1,7 мМ KH2PO4, 5,2 мМ Na2HPO4, 150 мМ NaCl). Далее клетки суспендировали в буфере для нанесения и инкубировали 5 минут при 95°С. Нормализованные по концентрации белковые образцы фракционировали с помощью электрофореза в 12-13% ПААГ в денатурирующих условиях. В качестве маркера использовали набор белков с молекулярными массами 6,5-200 кДа. После фракционирования в ПААГ белки из геля переносили на мембрану PVDF Immobilon-P (″Millipore″, Англия), используя прибор Bio-Rad Trans-Blot SD cell. Мембрану предварительно обрабатывали метанолом в течение 1 мин, отмывали бидистиллированной водой в течение 3 минут и помещали в буфер для переноса. Далее мембрану инкубировали с антителами к тимидинкиназе вируса простого герпеса HSV-tk (SC-28038) и GAPDH (SC-47724) (″Santa Cruz Biotechnology″, США) в разведении 1:1000 в течение 16 часов. Затем отмывали мембрану IX фосфатно-солевым буфером (PBS), содержащим 0,5% Tween-20, 4 раза по 10 минут. Далее мембрану инкубировали с вторичными антителами осла, коньюгированными с пероксидазой хрена, против иммуноглобулинов козы в случае белка HSV-tk и антителами козы против иммуноглобулинов мыши в случае белка GAPDH (SC-2302) (″Santa Cruz Biotechnology″, США) в разведении 1:1000 и отмывали, как описано выше. Белки на мембране детектировали хемилюминесцентным методом, используя реактивы фирмы ″Bio-Rad″ (США). Обсчет количества белка проводили с помощью программы Quantity One 4.6.1 (″Bio-Rad″, США). Результаты эксперимента приведены на Фиг.17. Видно, что количество HSVtk, нарабатываемое под контролем двойного промотора PhTSurv269 (дорожка 3) выше, чем количество HSVtk, нарабатываемое под контролем промотора PhSurv (дорожка 2).
Пример 20. Сравнительный анализ эффективности следующих промоторов: CMV (промотор цитомегаловируса), промотор гена сурвивина человека (PhSurv), искусственный двойной промотор (PhTSurv269) ex vivo на мышах
Для сравнения эффективности промоторов CMV, PhSurv и PhTSurv269 на основе вектора pGL3 (Promega, США) были созданы конструкции, содержащие кассету терапевтических генов HSVtk-mGM-CSF под контролем данных промоторов. Были созданы следующие конструкции CMV-HSVtk-mGM-CSF-pGL3, PhSurv-HSVtk-mGM-CSF-pGL3, PhTSurv269-HSVtk-mGM-CSF-pGL3 (конструирование вектора описано в Примере 4). Кассета терапевтических генов HSVtk-mGM-CSF содержала ген тимидинкиназы вируса простого герпеса (HSVtk). Продукт данного гена способен фосфорилировать ганцикловир, низкотоксичный аналог гуанина. Клетки, трансформированные HSVtk, погибают в присутствии ганцикловира, поскольку клеточные киназы превращают фосфорилированный ганцикловир в трифосфаты, которые при клеточном делении включаются во вновь синтезированную ДНК и обрывают ее дальнейший синтез. Продукт гена гранулоцитарно-макрофагального колониестимулирующего фактора (GM-CSF) способен активировать рост, развитие и дифференцировку гранулоцитов и макрофагов, стимулируя тем самым иммунный ответ.
В эксперименте были использованы:
1) Мыши (самки) линии С57В1/6, средний вес на начало эксперимента - 17,9±1,4 (здесь и далее среднее ± стандартная ошибка среднего), содержались на стандартном гранулированном сухом корме.
2) Клетки линии LLC (карцинома легких Льюис мыши).
3) Конструкции CMV-HSVtk-mGM-CSF-pGL3, PhSurv-HSVtk-mGM-CSF-pGL3, PhTSurv269-HSVtk-mGM-CSF-pGL3.
Клетки линии LLC трансфицировали в культуральных флаконах (75 см2) плазмидами CMV-HSVtk-mGM-CSF-pGL3, PhSurv-HSVtk-mGM-CSF-pGL3, PhTSurv269-HSVtk-mGM-CSF-pGL3, нетрансфицированные клетки использовали в качестве контроля. Для трансфекции использовали 30 мкг плазмидной ДНК и 75 мкл Липофектамина-2000 в расчете на культуральный планшет. Трансфекцию проводили в течение 3 ч, через 24 часа после трансфекции клетки открепляли от поверхности культурального флакона с помощью раствора трипсина и суспендировали в среде DMEM/F12 с антибиотиком, клетки переносили во флаконы и центрифугировали (1000 об/мин, 160g,) 5 мин. Далее клетки дважды промывали PBS и готовили суспензию клеток LLC в PBS с концентрацией 2·105 клеток/ мл для введения животным.
Трансплантация клеток самкам мышей линии С57В1/6 проводилась в день 1 подкожно в заднюю часть спины в количестве 2·105 клеток в 100 мкл PBS. Через 48 часов после инокуляции опухоли начинали введение раствора ганцикловира (GCV) внутрибрюшинно в течение 10 дней из расчета 75 мкг на грамм веса (2 введения по 100 мкл на 1 мышь в день, концентрация ганцикловира 5 мг/мл) 2 раза в день.
Начиная с 5-го дня после трансплантации клеток проводили измерение размера опухоли в 2-х измерениях (меньший (А) и больший (В) диаметр опухоли) электронным штангенциркулем и массы животных. Размер опухоли вычисляли по формуле A·B2/2. Замер диаметров опухоли проводили через день. Критерием эвтаназии служило достижение опухоли размером более 10% от массы животного.
Всего было сформировано 4 группы животных: 1) мыши с трансплантированными клетками LLC, транзиентно трансфицированными конструкцией CMV-HSVtk-mGM-CSF-pGL3, 2) мыши с трансплантированными клетками LLC, транзиентно трансфицированными конструкцией PhSurv-HSVtk-mGM-CSF-pGLS, 3) мыши с трансплантированными клетками LLC, транзиентно трансфицированными конструкцией PhTSurv269-HSVtk-mGM-CSF-pGL3, 4) контрольная группа (нетрансфицированные клетки LLC). Каждая группа состояла из 10 животных. Животные были равномерно распределены по группам согласно их весам.
Результаты эксперимента приведены на Фиг.18 и 19. Как видно на Фиг.18, 19, рост опухоли в контрольной группе начинается на 10 день эксперимента, тогда как в опытных группах опухоль появляется у животных только на 20 сутки с начала эксперимента. Средний размер опухоли на 20 день эксперимента в контрольной группе составлял 1343 мм3, что являлось основанием выведения животных группы из эксперимента (масса опухоли у животных превысила 10% от массы тела). На 36 день эксперимента в группе CMV опухоль пальпировалась у 1 из 10 животных, в группе PhSurv - у 2 животных, в группе PhTSurv269 не наблюдалось животных с пальпируемыми опухолями. Таким образом, созданный искусственный тандемный промотор PhTSurv269 в системе с терапевтическими генами HSVtk-mGM-CSF обладает высокой эффективностью. Он является опухолеспецифичным в отличие от промотора CMV и активен в более широком спектре раковых клеточных линий, чем промотор PhSurv.
Свободный текст перечня последовательностей:
SEQ ID NO:1 -
Human survivin gene promoter fragment sequence (PhS269, positions -268 to +1, wherein +1 corresponds to adenin of an initiating methionine encoding triplet)
SEQ ID NO:2 -
Human reverse transcriptase telomerase gene promoter fragment sequence (hTERT)
SEQ ID NO:3 -
Murine survivin gene promoter fragment sequence
SEQ ID NO:4 -
PhTSurv269 promoter sequence
SEQ ID NO:5 -
PhSurv269T promoter sequence
SEQ ID NO:6 -
PhTmSurv promoter sequence
SEQ ID NO:7 -
PmSurvhT promoter sequence
| название | год | авторы | номер документа |
|---|---|---|---|
| МНОГОПРОФИЛЬНЫЙ ПРОМОТОР, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР И СПОСОБ ИЗБИРАТЕЛЬНОГО УБИЙСТВА РАКОВЫХ КЛЕТОК С ИХ ИСПОЛЬЗОВАНИЕМ | 2012 |
|
RU2476596C1 |
| УНИВЕРСАЛЬНЫЙ ПРОМОТОР ДЛЯ ЭКСПРЕССИИ ТЕРАПЕВТИЧЕСКИХ ГЕНОВ В КЛЕТКАХ МЛЕКОПИТАЮЩИХ | 2013 |
|
RU2551784C1 |
| Генотерапевтический препарат FCU1-BsFm/PP для лечения солидных опухолей | 2023 |
|
RU2824977C1 |
| ФАРМАКОЛОГИЧЕСКАЯ КОМБИНАЦИЯ ПОЛИКАТИОННОГО НОСИТЕЛЯ ПЭГ-ПЭИ-ТАТ С ЗАКЛЮЧЕННОЙ В НЕМ ПЛАЗМИДОЙ, НЕСУЩЕЙ ТЕРАПЕВТИЧЕСКИЕ ГЕНЫ HSVtk И GM-CSF ДЛЯ ЦЕЛЕЙ ГЕНОТЕРАПИИ ОПУХОЛЕВЫХ ЗАБОЛЕВАНИЙ | 2013 |
|
RU2575077C2 |
| Гистоны и биодеградируемые липиды как средство для доставки нуклеиновых кислот в клетки эукариот | 2015 |
|
RU2637371C2 |
| Лекарственная комбинация для ген-иммунной терапии | 2022 |
|
RU2792683C1 |
| ИММОРТАЛИЗОВАННАЯ КЛЕТОЧНАЯ ЛИНИЯ ПТИЦ | 2008 |
|
RU2475536C2 |
| СПОСОБ ВЫЯВЛЕНИЯ ИНГИБИТОРОВ СУПРЕССИИ СУПРЕССОРА ОПУХОЛЕВОГО РОСТА Pdcd4 В ОПУХОЛЕВЫХ КЛЕТКАХ И ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ pLucPdcd4 ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2016 |
|
RU2640909C1 |
| ПРОМОТОР ДЛЯ ТКАНЕСПЕЦИФИЧЕСКОЙ ЭКСПРЕССИИ ГЕНОВ В ГЕРМИНАЛЬНЫХ ТКАНЯХ МЛЕКОПИТАЮЩИХ | 2010 |
|
RU2459870C2 |
| АЛЛОГЕННОЕ ПРОТИВООПУХОЛЕВОЕ ТЕРАПЕВТИЧЕСКОЕ СРЕДСТВО | 2004 |
|
RU2354694C2 |




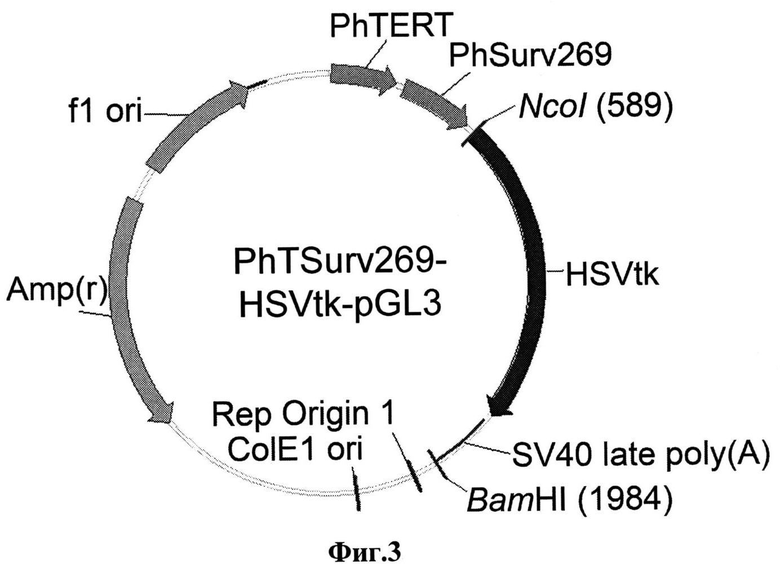









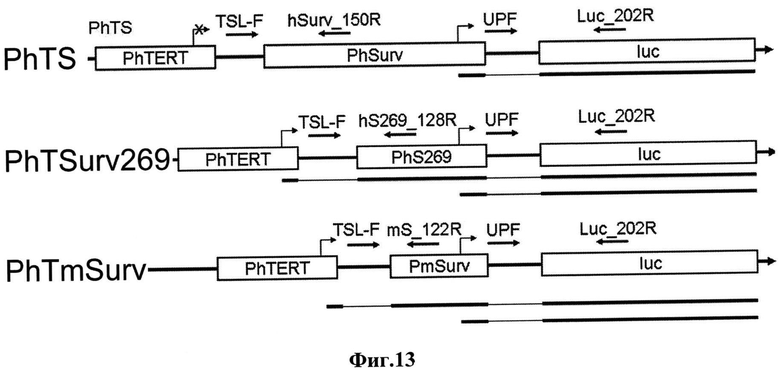




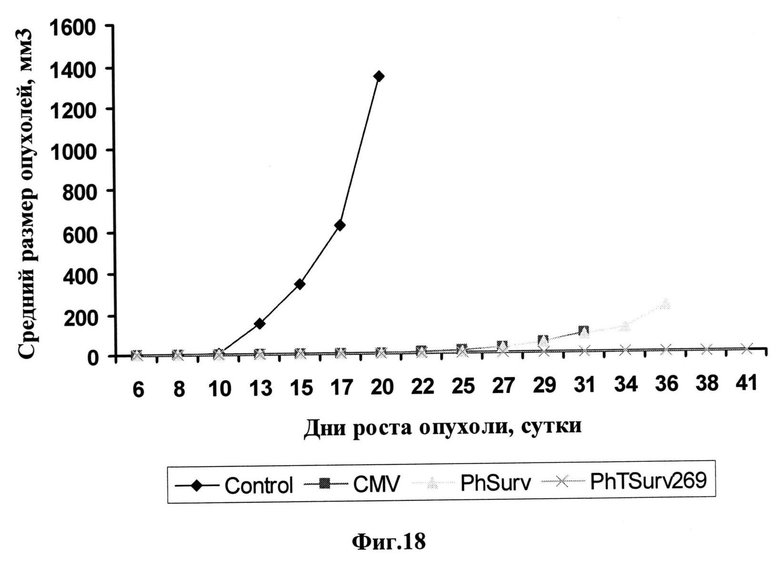

Изобретение относится к биотехнологии, в частности к опухолеспецифичным промоторам, и может быть использовано в противоопухолевой терапии. Сконструированы опухолеспецифичные промоторы широкого спектра действия, обеспечивающие экспрессию терапевтического гена внутри раковой клетки. Изобретение включает также экспрессионные кассеты, экспрессионные векторы, фармацевтические композиции, способы лечения раковых заболеваний и применения экспрессионных кассет и векторов. Промоторы настоящего изобретения обеспечивают высокий уровень экспрессии функционально связанного с ними терапевтического гена в раковых клетках различного генеза, при этом экспрессия в нормальных клетках отсутствует или находится на низком уровне. 5 н. и 24 з.п.ф-лы, 19 ил., 4 табл., 20 пр.
1. Опухолеспецифичный промотор, содержащий:
А) нуклеотидную последовательность SEQ ID NO:2 или ее функциональное производное; и
Б) нуклеотидную последовательность, выбранную из группы, состоящей из:
(i) нуклеотидной последовательности SEQ ID NO:1 или ее функционального производного; и
(ii) нуклеотидной последовательности SEQ ID NO:3 или ее функционального производного,
соединенные непосредственно или посредством линкерной последовательности.
2. Промотор по п.1, в котором последовательность SEQ ID NO:2 расположена выше (в 5′-направлении) относительно последовательности SEQ ID NO:1 или SEQ ID NO:3.
3. Промотор по п.1, в котором последовательность SEQ ID NO:2 расположена ниже (в 3′-направлении) относительно последовательности SEQ ID NO:1 или SEQ ID NO:3.
4. Промотор по любому из предшествующих пунктов, который дополнительно содержит линкерную последовательность между последовательностью согласно подпункту А) и последовательностью согласно подпункту Б).
5. Промотор по п.2, последовательность которого представлена в SEQ ID NO:4 или SEQ ID NO:5.
6. Промотор по п.3, последовательность которого представлена в SEQ ID NO:6 или SEQ ID NO:7.
7. Экспрессионная кассета для направленной экспрессии в клетках опухоли противоопухолевых агентов, содержащая в направлении от 5′ к 3′ промотор по любому из пп.1-6, соединенный с кодирующей последовательностью ДНК, которая при транскрипции продуцирует информационную РНК, кодирующую противоопухолевый агент.
8. Экспрессионная кассета по п.7, в которой кодирующая последовательность кодирует терапевтические белки или пептиды, антигены, антисмысловую РНК или рибозимы, обладающие противоопухолевой активностью.
9. Экспрессионная кассета по п.8, в которой кодирующая последовательность включает гены терапевтического вмешательства, выбранные из группы, включающей: негативные доминантные мутанты (НДМ) генов, участвующих в онкогенезе; гены-убийцы; гены-ингибиторы ангиогенеза; гены-онкосупрессоры; гены-иммуностимуляторы; гены siRNA.
10. Экспрессионная кассета по п.8, в которой негативные доминантные мутанты (НДМ) генов выбраны из группы, включающей НДМ гена сурвивина, НДМ гена c-Jun, НДМ онкогена RAS.
11. Экспрессионная кассета по п.8, в которой гены-убийцы выбраны из группы, включающей гены токсинов и гены ферментов, способных превращать нетоксичный агент (пролекарство) в токсин.
12. Экспрессионная кассета по п.11, в которой гены ферментов, способных превращать нетоксичный агент (пролекарство) в токсин, выбраны из группы, включающей ген тимидинкиназы вируса простого герпеса и цитозиндезаминазы дрожжей или Е. coli.
13. Экспрессионная кассета по п.8, в которой гены-ингибиторы ангиогенеза выбраны из группы, включающей гены белков ангиостатина и эндостатина.
14. Экспрессионная кассета по п.8, в которой гены-онкосупрессоры выбраны из группы, включающей ген белка p53.
15. Экспрессионная кассета по п.8, в которой гены-иммуностимуляторы выбраны из группы, включающей гены IL-1, IL-2, IL-4, IL-6, TNF, GM-CSF, гамма-интерферона.
16. Экспрессионная кассета по п.8, в которой гены siRNA выбраны из группы, включающей siRNA к гену сурвивина.
17. Экспрессионный вектор, содержащий экспрессионную кассету по любому из пп.7-16.
18. Экспрессионный вектор по п.17, который представляет собой вирусный вектор.
19. Экспрессионный вектор по п.18, где вектор выбран из группы, включающей ретровирусный, аденовирусный, аденоассоциированный, герпесвирусные вектор.
20. Экспрессионный вектор по п.17, который представляет собой невирусный вектор.
21. Экспрессионный вектор по п.20, где вектор выбран из группы, включающей плазмиду.
22. Фармацевтическая композиция, предназначенная для лечения онкологических заболеваний, содержащая терапевтически эффективное количество экспрессионной кассеты по любому из пп.7-16 или вектора по любому из пп.17-21 и фармацевтически приемлемое вспомогательное вещество.
23. Фармацевтическая композиция по п.22, предназначенная для лечения онкологических заболеваний.
24. Фармацевтическая композиция по п.23, где заболевание выбрано из группы, включающей: рак легкого, рак поджелудочной железы, меланому, фибросаркому, саркому.
25. Способ лечения онкологического заболевания, включающий введение пациенту фармацевтической композиции по любому из пп.22-24.
26. Способ лечения по п.25, где фармацевтическую композицию вводят внутривенно, внутриопухолево, внутримышечно, внутрибрюшинно, подкожно, перорально.
27. Способ лечения по любому из пп.25-26, где онкологическое заболевание выбрано из группы, включающей: рак легкого, рак поджелудочной железы, меланому, фибросаркому, саркому.
28. Применение экспрессионной кассеты по любому из пп.7-16 или вектора по любому из пп.17-21 для изготовления лекарственного средства для лечения онкологического заболевания.
29. Применение по п.28, где онкологическое заболевание выбрано из группы, включающей: рак легкого, рак поджелудочной железы, меланому, фибросаркому, саркому.
| RU 2007137122 A1, 20.04.2009 | |||
| US 2010256225 A1, 07.10.2010 | |||
| RU 2011130511 A1, 27.01.2013 | |||
| DAVIS J | |||
| et al., Oncolysis and suppression of tumor growth by a GFP-expressingoncolytic adenovirus controlled by an hTERT and CMV hybridpromoter, Cancer Gene Therapy, 2006, v | |||
| Насос | 1917 |
|
SU13A1 |
| ПАРОВОЗНАЯ ДРОВЯНАЯ ТОПКА | 1923 |
|
SU720A1 |
| CHUNXIAO WU et al., Combinatorial Control of Suicide Gene | |||

