Настоящая заявка относится к иммортализованным фибробластам эмбриона цыпленка, к культурам клеток, включающим такие иммортализованные клетки, к вакцинам, включающим такие клетки, к способам репликации вирусов птиц в таких клетках и к способам получения таких клеток и таких вакцин.
Размножение вирусов с целью получения вакцины требует наличия восприимчивых клеток-хозяев. Обычно в зависимости от типа вируса и типа используемой клетки-хозяина эти клетки-хозяева выращивают в культуре клеток. Для размножения многих видов вирусов птиц существует дополнительная возможность размножения в оплодотворенных яйцах. Несмотря на это, на практике многие вирусы птиц выращивают на первичных клетках-фибробластах эмбриона цыпленка (CEF, ФЭЦ). (Клетки, которые культивируют напрямую от животных, известны как первичные клетки). Такие первичные Клетки ФЭЦ являются восприимчивыми ко многим различным видам вирусов, и такие вирусы часто можно выращивать до высоких титров в этих клетках. Примерами вирусов, которые часто выращивают в клетках ФЭЦ, являются вирус герпеса индеек (HVT), вирус Марека, вирус болезни Ньюкасла (NDV), вирус инфекционного бронхита (IBV), вирус инфекционного бурсита (IBDV), вирус синдрома снижения несучести (EDSV), реовирус (RV) и вирус ринотрахеита индеек (TRT).
Несмотря на это, существует несколько недостатков в использовании клеток ФЭЦ. Эти клетки имеют относительно короткий жизненный цикл in vitro, а также их получают из оплодотворенных свободных от специфической патогенной микрофлоры яиц, поэтому их производство в условиях непрерывного спроса является как чрезвычайно трудоемким, так и чрезвычайно дорогим. Таким образом, является желательным создать линии клеток птичьего происхождения для замены используемых первичных клеток ФЭЦ.
В соответствии с настоящим изобретением, иммортализованная линия клеток представляет собой популяцию клеток (в данном случае ФЭЦ), происходящих из многоклеточного организма, которые в обычных условиях не пролиферируют бесконечно, но благодаря мутации отклоняются от нормального клеточного старения и вместо него могут продолжать процесс деления клетки. Такие клетки избегают нормальных ограничений в росте только на ограниченное количество циклов деления.
Иммортализованная форма клеток ФЭЦ (линия клеток ФЭЦ) может принципиально решить проблемы, перечисленные выше, но такие клетки являются труднодоступными. Одним способом получения таких клеток является выделение и выращивание первичных клеток ФЭЦ, а также ожидание наступления спонтанной иммортализации. Тем не менее, для клеток птиц спонтанная иммортализация является очень редким явлением. Как следствие, зарегистрированы только три спонтанно иммортализованные линии клеток цыпленка; DF-1 (Патент США US 5672485), SC-1 и SC-2 (Christman, S.A. et al., Dissertation Abstracts Int. 65: 4414 (2004) (ISBN 0-496-06882-2). Помимо того, что нахождение и выделение спонтанно иммортализованных клеток ФЭЦ может быть очень длительным, также оказывается, что такие иммортализованные клетки ФЭЦ могут демонстрировать совершенно другие характеристики, такие как вариабельная скорость роста и концентрации p53 во времени, в зависимости от количества пассажей, которые они прошли (Christman, S.A. et al., FEBS letters 579: 6705-6715 (2005)). Несмотря на это, в особенности для производства вакцины, необходимы линии клеток, чьи характеристики остаются прежними с течением времени, независимо от количества пассажей. Управление по контролю качества пищевых продуктов и лекарственных средств (FDA) сообщает, что разработка минимально очищенных живых аттенуированных вирусных вакцин в неопластических клетках, которые были трансформированы при помощи неизвестных механизмов, не рекомендуется (FDA; CBER Discussion on cell substrates May 12, 2000). Таким образом, спонтанно иммортализованные клетки ФЭЦ являются менее желательными в качестве источника клеток для размножения вируса с целью получения вакцины.
Также существуют способы, известные в области техники, для преднамеренной иммортализации клеток. Преимуществом таких способов является то, что коэффициент одновременности, который является серьезной преградой в случае спонтанной иммортализации клеток, устраняется. Для клеток, не принадлежащих птицам, в частности для клеток млекопитающих, часто используемый способ для получения иммортализованных клеток основывается на инфицировании первичных клеток ретровирусами или ретровирусными векторами, или трансфекции первичных клеток молекулами ДНК, которые включают ретровирусы или по меньшей мере ретровирусные ДКП последовательности (длинные концевые повторы) и последовательности, кодирующие протеины опухолевых ДНК-вирусов, таких как вирус обезьян SV40 T и t.
SV40 T и t играют роль в инактивации белков ретинобластомы (Rb) и р53. Обзорная статья, написанная Deepika Ahuja с соавт., касающаяся SV40 Т, кодирующего большой T и t, позволяет взглянуть на механизмы действия данных белков (Oncogene 24: 7729-7745 (2005)). В целом, продукты SV40 T гена T и t ингибируют р53 и Rb-семейство онкосупрессоров.
ДКП представляют собой элементы ретровирусов, которые включают все требуемые сигналы для экспрессии ретровирусного гена: энхансер, промотор, инициация транскрипции, терминатор транскрипции и сигнал полиаденилирования.
Тем не менее, есть подозрения, что эти ДКП обладают канцерогенными эффектами. Причиной этого является тот факт, что они известны как цис-активаторы других клеточных генов, и тот факт, что они могут воссоединяться с другими ретровирусными последовательностями в клеточном геноме (Mosier, D.E., Applied Biosafety 9: 68-75 (2004)). Таким образом, сильным недостатком применения ретровирусной ДНК, включающей ДКП или по меньшей мере длинные ретровирусные последовательности, для трансфицирующих клеток является то, что в таких случаях ДКП последовательности вводят в ДНК иммортализованных клеток.
Для клеток птиц известен только один пример успешной иммортализации молекулой ДНК, включающей ДКП и экспрессирующей SV40 T антиген: такие иммортализованные клетки ФЭЦ описаны в Заявке PCT WO 97/44443.
Такие способы, как видно, являются не очень успешными. Soo-Hyun Kim описывает трансфекцию клеток ФЭЦ с ретровирусом, кодирующим SV40 T антиген, в попытке иммортализации клеток ФЭЦ путем инактивации онкосупрессоров ретинобластомы (Rb) и р53 (J. Cell Science 119: 2435-2443 (2006)). Тем не менее, это приводит только к незначительному удлинению жизненного цикла клеток ФЭЦ, но не к иммортализации.
Kim в то же время предполагает, что возможное объяснение ограниченной продолжительности жизненного цикла может заключаться в длительной эрозии теломеров. Тем не менее, ситуация осложняется тем фактом, что клетки цыпленка содержат как макро-, так и микрохромосомы с классами теломерных повторов различных размеров, некоторые из которых являются интерстициальными. Таким образом, на данном этапе динамика эрозии и восстановления теломера, а также их влияние на иммортализацию клеток цыпленка остаются неясными.
Это затрагивает основной и неразрешенный вопрос о том, играет ли важную роль в иммортализации ФЭЦ потеря р53 и Rb или активация теломеразы (Campisi, J., Exp. Gerontol. 36: 607-618 (2001) и Sherr, J.C. and DePinho, R.A., Cell 102: 407-410 (2000)).
Из последних экспериментов можно сделать вывод, что в противоположность предположению Kim роль теломеразы в иммортализации клеток ФЭЦ оказывается не такой важной. Прежде всего, поражает то, что в спонтанно иммортализованных линиях клеток ФЭЦ SC-1 (см. ранее) активность теломеразы не определяется (Christman, S.A. et al., FEBS letters 579: 6705-6715 (2005)).
Во-вторых, в нескольких несложных экспериментах, в которых клетки цыпленка трансдуцированы или трансфицированы с cTR, cTERT или и с cTR и cTERT, иммортализованные клетки цыпленка не получены (Swanberg, S.E. et al., Exp. Gerontol. 45: 647-654 (2010)).
В настоящее время вопреки всем ожиданиям обнаружено, что стабильно трансфицированные линии клеток ФЭЦ можно получить путем трансфекции клеток ФЭЦ без применения ДКП последовательностей, с молекулой ДНК, включающей инвертированные повторы транспозона для интеграции молекулы ДНК в клеточный геном, и комбинацию обоих генов, кодирующих SV40 T и t антиген или по меньшей мере Т под контролем подходящего промотора, а также гена, кодирующего теломеразу цыпленка (cTERT) под контролем подходящего промотора. Инвертированные повторы транспозона играют роль в стабильной интеграции гена, кодирующего SV40 T и t антиген или по меньшей мере T, а также гена, кодирующего (cTERT), в геном ФЭЦ, что является необходимым предварительным условиям для получения стабильно трансфицированных иммортализованных ФЭЦ в соответствии с изобретением.
Таким образом, первый вариант осуществления настоящего изобретения относится к стабильно трансфицированным иммортализованному фибробласту эмбриона цыпленка (ФЭЦ), отличающемуся тем, что стабильно трансфицированный иммортализованный ФЭЦ экспрессирует SV40 T антиген, экспрессирует теломеразу цыпленка (cTERT) и не включает экзогенную ретровирусную ДНК с Длинным концевым повтором.
Считается, что экзогенные ретровирусные ДКП ДНК представляют собой ДНК, которая попадает в фибробласты эмбриона цыпленка в процессе иммортализации, как описано ранее.
Дополнительные детали и характеристики таких иммортализованных ФЭЦ, а также способов приготовления таких ФЭЦ подробно описаны далее.
Второй вариант осуществления настоящего изобретения относится к способам приготовления таких иммортализованных линий клеток ФЭЦ.
Способы получения иммортализованной линии клеток ФЭЦ в соответствии с изобретением в целом включают следующие стадии:
а) стадию получения первичных клеток ФЭЦ. Эта стадия хорошо известна в данной области техники и описана, помимо прочего, Hernandez, R и Brown, D.T. в Current protocols in Microbiology 17: A.4i.1 - A.4i.8 (2010), а также другими авторами. Это все еще является предпочтительным способом получения ФЭЦ.
b) стадию трансфекции указанных ФЭЦ при помощи 1) молекулы ДНК, не содержащей ДКП последовательности, включающей инвертированные повторы транспозона и включающей ген, кодирующий SV40 T антиген под контролем подходящего промотора, 2) молекулы ДНК, не содержащей ДКП последовательности, включающей инвертированные повторы транспозона и включающей ген, кодирующий теломеразу цыпленка; cTERT под контролем подходящего промотора и 3) молекулы ДНК, включающей ген, кодирующий транспозазу под контролем подходящего промотора.
Молекула ДНК, включающая ген, кодирующий транспозазу под контролем подходящего промотора, не должна обязательно быть свободной от ДКП последовательностей, поскольку сама по себе данная молекула ДНК не включает последовательности транспозона и, таким образом, скорее всего не будет интегрирована в геном хозяина. Тем не менее, для избегания непреднамеренной случайной интеграции предпочтительно, чтобы молекула ДНК, включающая ген, кодирующий транспозазу под контролем подходящего промотора, не содержала ДКП последовательности.
Исходя из вопросов эффективности, на практике трансфекцию является предпочтительным проводить при помощи одной молекулы ДНК, не содержащей ДКП последовательности, включающей инвертированные повторы транспозона, включающей как ген, кодирующий SV40 T антиген под контролем подходящего промотора, так и ген, кодирующий теломеразу цыпленка под контролем подходящего промотора, а также включающей ген, кодирующий транспозазу под контролем подходящего промотора.
Активность транспозазы является необходимой только на протяжении первых стадий процесса иммортализации для интеграции ДНК в геном ФЭЦ. Сразу после интеграции ДНК (нескольких ДНК) транспозаза становится не нужна. После она может оказывать негативное влияние на стабильность клеток.
Таким образом, еще более предпочтительно, чтобы стадию трансфекции данных ФЭЦ проводили при помощи 1) одной молекулы ДНК, не содержащей ДКП последовательности, включающей инвертированные повторы транспозона и включающей как ген, кодирующий SV40 T антиген под контролем подходящего промотора, так и ген, кодирующий теломеразу цыпленка под контролем подходящего промотора, и 2) молекулы ДНК, предпочтительно не содержащей ДКП последовательности, включающей только ген транспозазы под контролем подходящего промотора без последовательностей транспозона.
Трансфекцию можно проводить многими путями, известными в области техники. Имеющиеся на рынке наборы для трансфекции в настоящее время доступны, помимо прочего, у компании Bio-Rad (Life Science (Research, Education, Process Separations, Food Science), Life Science Research, 2000 Alfred Nobel Drive, Hercules, CA 94547, USA) и Invitrogen (Life Technology, 3175 Staley Road, Grand Island, NY 14072, USA). Широко используемые методы трансфекции с помощью реагентов включают применение липидов, фосфата кальция, катионных полимеров, диэтиламиноэтилдекстрана, активированных дендримеров и магнитных микроносителей. Методы трансфекции с помощью инструментов включают электропорацию, нуклеофекцию и микроиньекции.
Молекула ДНК, не содержащая ДКП последовательности, включающая инвертированные повторы транспозона, включающая ген, кодирующий SV40 T антиген под контролем подходящего промотора, и инвертированные повторы транспозона и/или ген, кодирующий теломеразу цыпленка под контролем подходящего промотора, может представлять собой, например, плазмиду. Плазмида может иметь кольцевую или линейную форму при ее применении в стадии трансфекции.
Использование транспозонов само по себе хорошо известно в области техники. Статья, написанная Ivics, Z. и Izsvak Z., дает широкий обзор транспозонов и их применения, а также позволяет взглянуть на механизмы действия транспозонов (Mobile DNA 1: 25-39 (2010)). Транспозоны можно рассматривать как природные мобильные элементы ДНК, которые, сходным образом с интегрирующими вирусами, обладают способностью к эффективному встраиванию в геном, что регулируется транспозазой.
В основном транспозоны стабильно присутствуют в клеточном геноме после интеграции в геном. Таким образом, предпочтительно иммортализованные ФЭЦ в соответствии с изобретением включают последовательности транспозона, такие как инвертированные повторы транспозона.
Большое количество подходящих промоторов для экспрессии SV40 T антигена и сTERT известны в области техники, и отличаются своим высоким уровнем экспрессии. Известно, что промоторы, которые обладают транскрипционной активностью в клетках млекопитающих, также хорошо функционируют в клетках птиц. Такие промоторы включают классические промоторы, такие как промотор гена немедленного раннего ответа цитомегаловируса (человека) (Sun-Young Lee et al., Journal of Biomedical Science 6: 8-17 (1999), Seed, B. et al., Nature 329, 840-842, 1987; Fynan, E.F. et al., PNAS 90, 11478-11482,1993; Ulmer, J.B. et al., Science 259, 1745-1748, 1993), промотор и энхансер гена цитомегаловируса человека (Donofrio G., et al., Clinical and Vaccine Immunology 13: 1246-1254, (2006)), промотор гена немедленного раннего ответа цитомегаловируса мышей (MCMVie1), ранний промотор гена цитомегаловируса мышей (MCMVe1), промотор гена немедленного ответа SV40 (Sprague J. et al., J. Virology 45, 773,1983), промотор металлотионеина (Brinster, R.L. et al., Nature 296, 39-42, 1982), промотор гена теплового шока (Voellmy et al., Proc. Natl. Acad. Sci. USA, 82, 4949-53, 1985), главный поздний промотор вируса Ad2 и промотор β-актина (Tang et al., Nature 356, 152-154, 1992).
Предпочтительно промотор представляет собой CAG промотор (Miyazaki, J; Takaki, S; Araki, K; Tashiro, F; Tominaga, A; Takatsu, K; Yamamura, K., Gene 79 (2): 269-277 (1989), и Niwa, H; Yamamura, K; Miyazaki, J,. Gene 108 (2): 193-199 (1991)).
с) стадию отбора клеток, которые обладают способностью к непрерывной пролиферации.
Клетки ФЭЦ, которые обладают способностью к непрерывной пролиферации, представляют собой клетки, которые продолжают пролиферировать по меньшей мере до 45 удвоений популяции. Клеточный цикл или цикл деления клетки представляет собой серию событий, которые протекают в клетке, и приводят к ее делению и удвоению (репликация клетки). Отбор клеток, которые обладают способностью к непрерывной пролиферации, представляет собой очень простой процесс по следующей причине: первичные ФЭЦ даже в наиболее оптимальной ситуации являются неспособными к делению за пределами их природной окружающей среды, эмбриона птицы, более 45 раз. После начальной фазы пролиферации скорость пролиферации живых первичных клеток ФЭЦ после выделения снижается с течением времени, и, наконец, все первичные ФЭЦ входят в непролиферативную стадию. Вследствие этого они погибают после максимально количества удвоений популяции, приблизительно равных 45.
Это означает, что если имеется увеличение количества клеток, особенно после 45 удвоений популяции, это происходит благодаря тому, что одна или более клеток успешно трансфицированы, и что ген, кодирующий SV40 T антиген, и ген, кодирующий cTERT встроены в клеточный геном. Поэтому в целом процесс является само-селективным: поддержание ФЭЦ, некоторые из которых успешно трансфицированы, в подходящей среде для клеточного роста автоматически приводит к репликации данных трансфицированных клеток, в то время как неиммортализованные клетки перестают делиться и погибают. Подходящие среды для клеточного роста известны в области техники (см. Kim, 2006 и Hernandez, 2010 выше) и описаны, помимо прочего, в разделе с Примерами.
Дополнительные рекомендации по характеристикам культуры клеток можно найти в Примерах.
Обычно выбирают клетки, которые культивируют на протяжении по меньшей мере 45 клеточных циклов. Для таких клеток можно обоснованно сделать вывод, что они представляют собой успешно иммортализованные ФЭЦ, поскольку первичные ФЭЦ обычно не проходят репликацию in vitro после выделения из эмбриона цыпленка более, чем приблизительно 45 раз.
Таким образом, один вариант осуществления настоящего изобретения относится к способу приготовления иммортализованных ФЭЦ в соответствии с изобретением, где указанный способ включает стадии
а) получения первичных клеток ФЭЦ,
b) трансфекции указанных ФЭЦ при помощи 1) молекулы ДНК, не содержащей ДКП последовательности, включающей инвертированные повторы транспозона и включающей ген, кодирующий SV40 T антиген под контролем подходящего промотора, 2) молекулы ДНК, не содержащей ДКП последовательности, включающей инвертированные повторы транспозона и включающей ген, кодирующий теломеразу цыпленка; cTERT под контролем подходящего промотора и 3) молекулы ДНК, включающей ген, кодирующий транспозазу под контролем подходящего промотора.
с) отбора клеток ФЭЦ, которые культивируют в течение по меньшей мере 45 клеточных циклов.
Предпочтительная форма этого варианта осуществления относится к способу получения иммортализованных ФЭЦ в соответствии с изобретением, отличающемуся тем, что указанный способ включает стадии
а) получения первичных клеток ФЭЦ,
b) трансфекции указанных ФЭЦ при помощи одной молекулы ДНК, не содержащей ДКП последовательности, включающей инвертированные повторы транспозона и включающей ген, кодирующий SV40 T антиген под контролем подходящего промотора, а также ген, кодирующий теломеразу цыпленка под контролем подходящего промотора,
с) отбора клеток ФЭЦ, которые культивируют в течение по меньшей мере 45 клеточных циклов.
Более предпочтительная форма настоящего варианта осуществления относится к способу получения иммортализованных ФЭЦ в соответствии с изобретением, отличающемуся тем, что указанный способ включает стадии
а) получения первичных клеток ФЭЦ,
b) трансфекции указанных ФЭЦ при помощи 1) молекулы ДНК, не содержащей ДКП последовательности, включающей инвертированные повторы транспозона и включающей как ген, кодирующий SV40 T антиген под контролем подходящего промотора, так и ген, кодирующий теломеразу цыпленка под контролем подходящего промотора, и при помощи 2) молекулы ДНК, не содержащей ДКП последовательности, включающей ген, кодирующий транспозазу под контролем подходящего промотора,
с) отбора клеток ФЭЦ, которые культивируют в течение по меньшей мере 45 клеточных циклов.
В исключительных случаях клетки, которые прошли приблизительно 45 клеточных циклов, могут все еще демонстрировать нестабильность поведения, например, по причине того, что транспозон интегрирован в клеточный геном в очень важном сайте, или по причине нестабильной интеграции гена, кодирующего SV40 Т антиген или кодирующего cTERT. Таким образом, на практике предпочтительно выбирают клетки, которые культивируют по меньшей мере на протяжении 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150 или даже 160 клеточных циклов в таком порядке предпочтений. Шанс манифестации какой-либо нестабильности снижается с количеством прошедших клеточных циклов отобранных иммортализованных ФЭЦ.
Таким образом, предпочтительно выбирают клетки, которые культивируют на протяжении 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150 или даже 160 клеточных циклов в таком порядке предпочтений.
Третий вариант осуществления настоящего изобретения относится к способам репликации вируса птиц или вирусного вектора птиц, где указанные способы включают стадии
а) культивирования иммортализованных ФЭЦ в соответствии с изобретением,
b) контактирования иммортализованных ФЭЦ с вирусом птиц или вирусным вектором птиц
с) обеспечения репликации вируса птиц или вирусного вектора птиц и
d) выделения вирусного потомства.
Вирусы птиц, представляющие особый интерес, представляют собой следующие вирусы птиц: вирус герпеса индеек (HVT), вирус Марека, вирус болезни Ньюкасла (NDV), вирус инфекционного бронхита (IBV), вирус инфекционного бурсита (IBDV), вирус синдрома снижения несучести (EDSV), реовирус (RV) и вирус ринотрахеита индеек (TRT). Для всех данных вирусов в данной области техники известны вакцины, созданные либо на основании живых аттенуированных вирусов, либо инактивированных вирусов или рекомбинантных вирусов; вирусных векторов, экспрессирующих иммуногенные компоненты любых данных вирусов.
Вирусный вектор представляет собой вирус, который является носителем дополнительного гена, не представленного в форме вируса дикого типа. Вирусные векторы хорошо известны в области техники. Вирусные векторы можно применять для переноса, например, чужеродного бактериального гена или чужеродного вирусного гена. Обычно дополнительный ген размещают под контроль подходящего промотора. Примерами таких вирусных векторов являются, например, HVT вектор, включающий IBDV VP2-ген, IBV-ген белков шиловидного отростка, HA-ген птичьего гриппа, ILT-ген белка gD/gI или F-ген NDV.
Таким образом, предпочтительная форма этого варианта осуществления относится к способам репликации вируса птиц или вирусного вектора птиц в соответствии с изобретением, где вирус птиц или вирусный вектор птиц выбирают из группы вирусов птиц, состоящей из вируса болезни Марека (MDV), MDV-родственного вируса герпеса индеек (HVT), вируса болезни Ньюкасла (NDV), вируса инфекционного бронхита (IBV), вируса инфекционного бурсита (IBDV), вируса синдрома снижения несучести (EDSV), реовируса (RV), вируса ринотрахеита индеек (TRT), а также HVT вектора, включающего IBDV VP2-ген, IBV-ген белков шиловидного отростка, HA-ген птичьего гриппа, ILT-ген белка gD/gI или F-ген NDV.
Известно, что вирус болезни Марека (MDV) представляет собой вирус герпеса, существующий в трех серотипах: высоко вирулентном серотипе 1, умеренно вирулентном серотипе 2 и серотипе 3, вирусе индеек, который не является вирулентным для цыплят; родственном вирусу болезни Марека вирусе герпеса индеек (HVT).
Иммортализованные клетки ФЭЦ в соответствии с изобретением являются чрезвычайно подходящими для размножения трех MDV-серотипов.
Таким образом, более предпочтительная форма этого варианта осуществления относится к способам репликации вируса птиц в соответствии с изобретением, где вирус птиц выбирают из группы вирусов птиц, состоящей из MDV и вирусного вектора MDV.
Четвертый вариант осуществления настоящего изобретения относится к культуре клеток, включающей иммортализованные ФЭЦ в соответствии с изобретением.
Предпочтительная форма этого варианта осуществления относится к такой культуре клеток, которую инфицируют вирусом птиц или вирусным вектором птиц.
Более предпочтительная форма этого варианта осуществления связана с такой культурой клеток, которую инфицируют вирусом птиц или вирусным вектором птиц, выбираемым из группы, состоящей из вируса болезни Марека (MDV), MDV-родственного вируса герпеса индеек (HVT), вируса болезни Ньюкасла (NDV), вируса инфекционного бронхита (IBV), вируса инфекционного бурсита (IBDV), вируса синдрома снижения несучести (EDSV), вируса ринотрахеита индеек (TRT), реовируса (RV), а также HVT вектора, включающего IBDV VP2-ген, IBV-ген белков шиловидного отростка, HA-ген птичьего гриппа, ILT-ген белка gD/gI или F-ген NDV.
Еще более предпочтительная форма этого варианта осуществления относится к таким культурам клеток, где культуру клеток инфицируют MDV или вирусным вектором MDV.
Пятый вариант осуществления настоящего изобретения относится к способам получения вакцины, включающей вирус птиц или вирусный вектор птиц, где способ включает стадию смешивания культуры клеток в соответствии с изобретением, где культуру клеток инфицируют вирусом птиц или вирусным вектором птиц, с фармацевтически приемлемым носителем.
В предпочтительной форме этого варианта осуществления вирус птиц или вирусный вектор птиц выбирают из группы, состоящей из вируса болезни Марека (MDV), MDV-родственного вируса герпеса индеек (HVT), вируса болезни Ньюкасла (NDV), вируса инфекционного бронхита (IBV), вируса инфекционного бурсита (IBDV), вируса синдрома снижения несучести (EDSV), реовируса (RV), вируса ринотрахеита индеек (TRT), а также HVT вектора, включающего IBDV VP2-ген и/или F-ген NDV.
Некоторые MDV-родственные вирусы способны выжить только если они связаны с клеткой, в которой они выращены. Серотип 1 способен выжить, не будучи связанным с клеткой. Серотип 2 имеет меньшую вероятность выживания, не будучи связанным с клеткой. Серотип 3 (HVT) не выживает после отделения от клетки, в которой он выращен. Иммортализованные ФЭЦ в соответствии с изобретением являются чрезвычайно подходящими для роста этих MDV серотипов. Поэтому для MDV, особенно для серотипов 2 и 3 MDV, оказывается, что иммортализованные ФЭЦ в соответствии с изобретением, которые инфицируют этими серотипами MDV, являются чрезвычайно подходящими для роста этих серотипов MDV. Такие вакцины можно получать в соответствии со стандартными техниками, известными в области техники для приготовления вакцин, содержащих клеточно-связанный MDV, путем смешивания инфицированных MDV иммортализованных ФЭЦ и фармацевтически приемлемого носителя.
Таким образом, в более предпочтительной форме этого варианта осуществления вирус птиц или вирусный вектор птиц представляет собой MDV.
В еще более предпочтительной форме этого варианта осуществления MDV или вирусный вектор MDV находится в живой аттенуированной форме.
Шестой вариант осуществления настоящего изобретения относится к способам получения вакцины, включающей вирус птиц или вирусный вектор птиц, где способ включает стадии
a) инфицирования культуры клеток ФЭЦ в соответствии с изобретением вирусом птиц или вирусным вектором птиц
b) репликации указанного вируса птиц или вирусного вектора птиц
c) выделения вирусного потомства
d) смешивания вирусного потомства с фармацевтически приемлемым носителем.
Предпочтительная форма этого варианта осуществления относится к такой вакцине, где вирус птиц или вирусный вектор птиц выбирают из группы, состоящей из вируса болезни Марека (MDV), MDV-родственного вируса герпеса индеек (HVT), вируса болезни Ньюкасла (NDV), вируса инфекционного бронхита (IBV), вируса инфекционного бурсита (IBDV), вируса синдрома снижения несучести (EDSV), реовируса (RV), вируса ринотрахеита индеек (TRT), а также HVT вектора, включающего IBDV VP2-ген и/или F-ген NDV.
Более предпочтительная форма этого варианта осуществления относится к такой вакцине, где вирус птиц или вирусный вектор птиц представляют собой MDV или вектор MDV.
Как указано ранее, некоторые MDV-родственные вирусы способны выживать только будучи связанными с клеткой, в которой они выращены. Поэтому для MDV, особенно для серотипов 2 и 3 MDV, оказывается, что иммортализованные ФЭЦ в соответствии с изобретением, которые инфицируют этими серотипами MDV, являются чрезвычайно подходящими для применения в вакцине, включающей клеточно-связанный MDV. Такие вакцины можно получать в соответствии со стандартными техниками, известными в области техники для получения вакцин, содержащих клеточно-связанный MDV, путем смешивания инфицированных MDV иммортализованных ФЭЦ и фармацевтически приемлемого носителя.
Таким образом, седьмой вариант осуществления настоящего изобретения относится к вакцинам, включающим иммортализованную культуру клеток ФЭЦ в соответствии с изобретением, где культуру клеток инфицируют MDV, и фармацевтически приемлемый носитель.
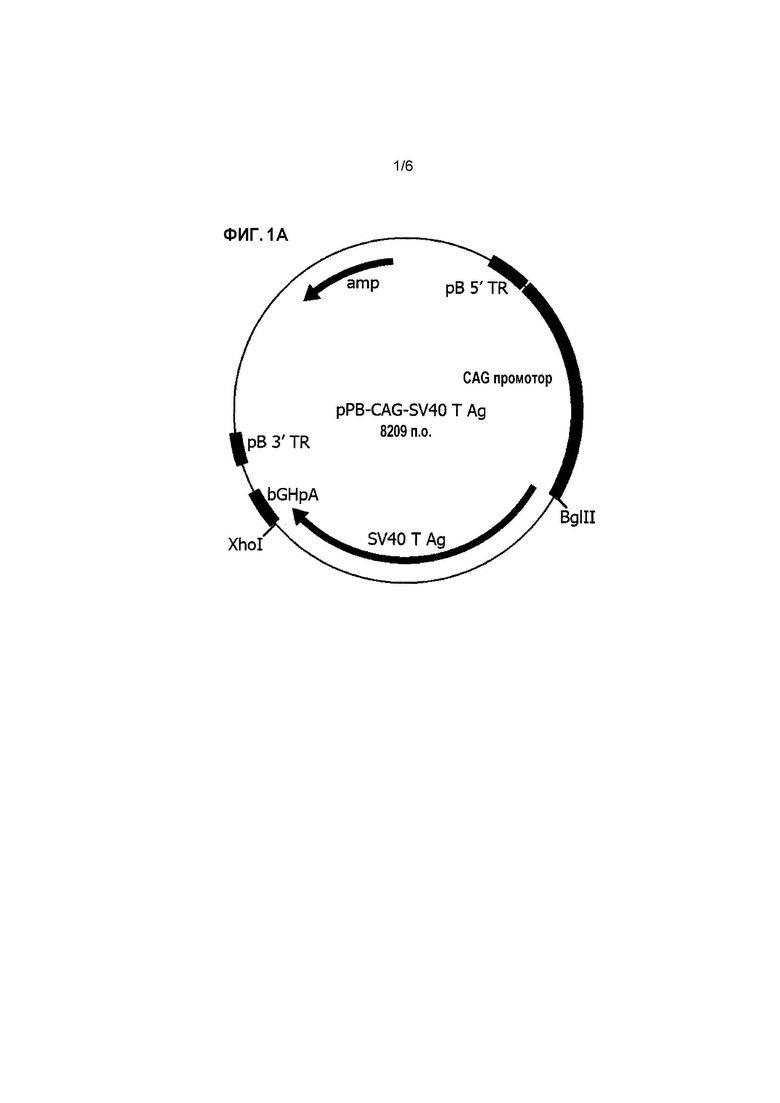
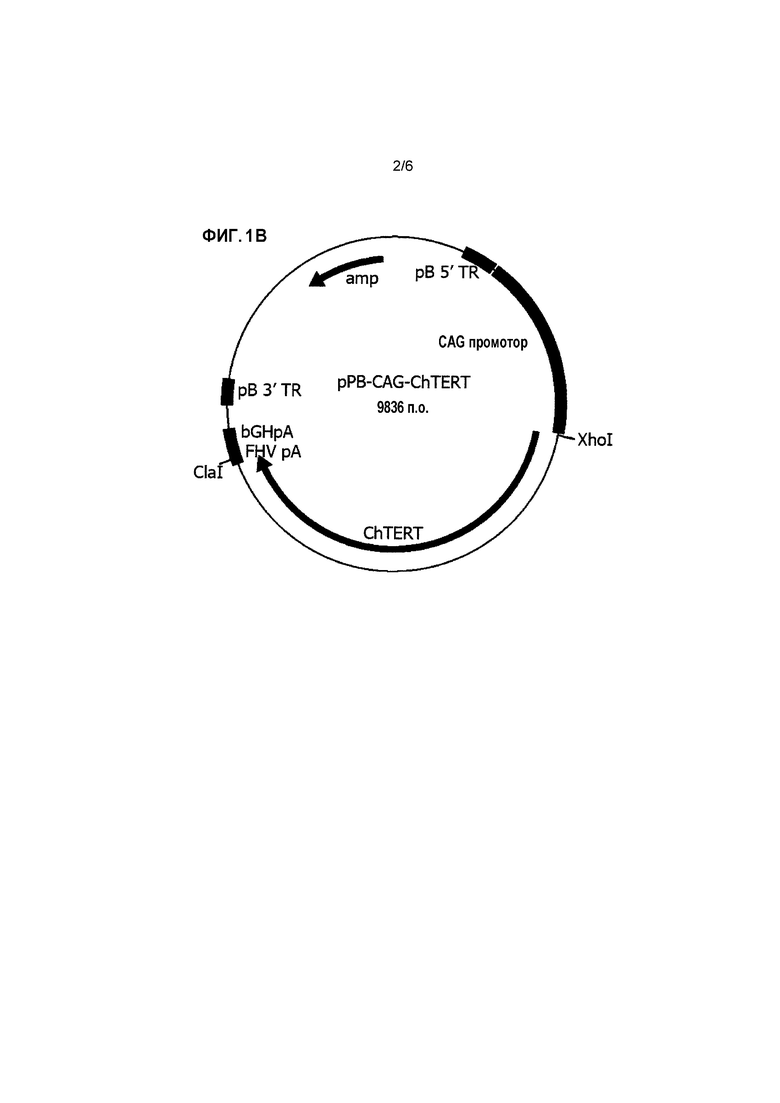
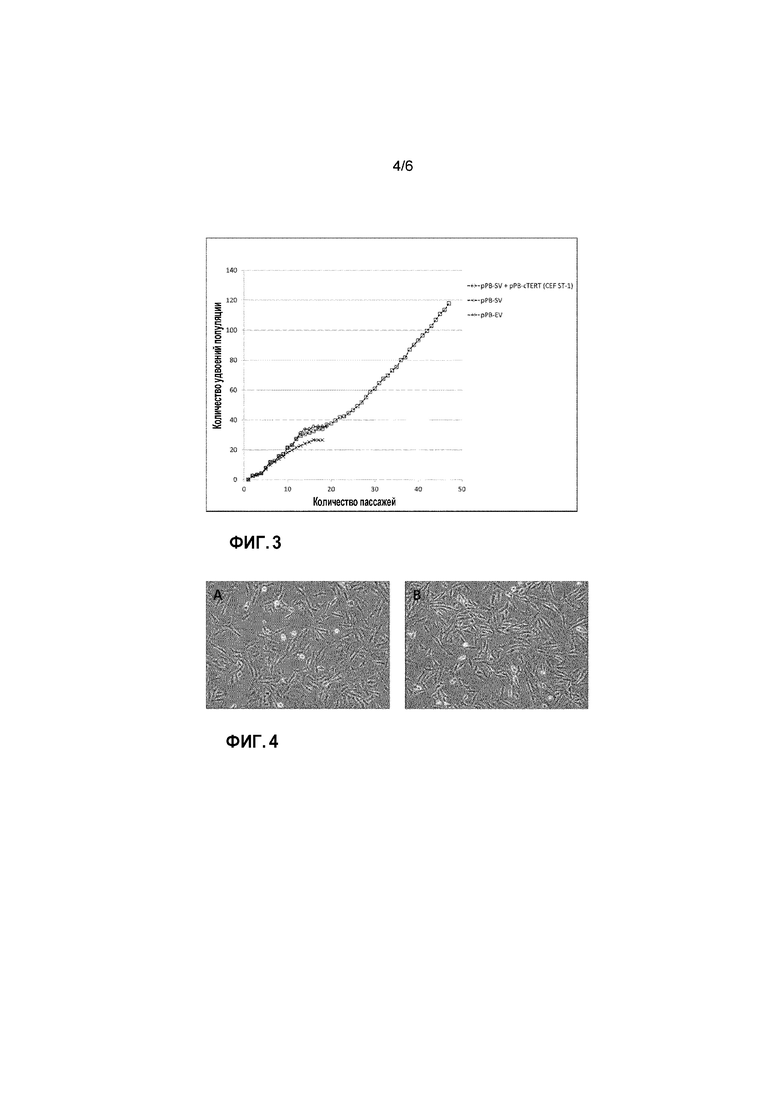
Пояснения к фигурам Фигура 1: Векторные карты для pPB-CAG-SV40 T Ag (A) и pPB-CAG-cTERT (B). Фигура 2: аминокислотная последовательность cTERT. *обозначает стоп-кодон. Фигура 3: Кривая роста линии клеток ФЭЦ ST-1. Клетки, трансфицированные с pPB-EV (крестики), pPB-SV (ромбы) или pPB-SV и pPB-cTERT (ФЭЦ ST-1) (квадраты), пассируют. Количество клеток определяют в каждом пассаже для подсчета удвоения популяции в каждом пассаже.
Фигура 4: Морфология ФЭЦ ST-1 остается стабильной на протяжении пассирования. Клетки ФЭЦ ST-1 фотографировали на 25 пассаже (100x)(A) и 44 пассаже (100x)(B).
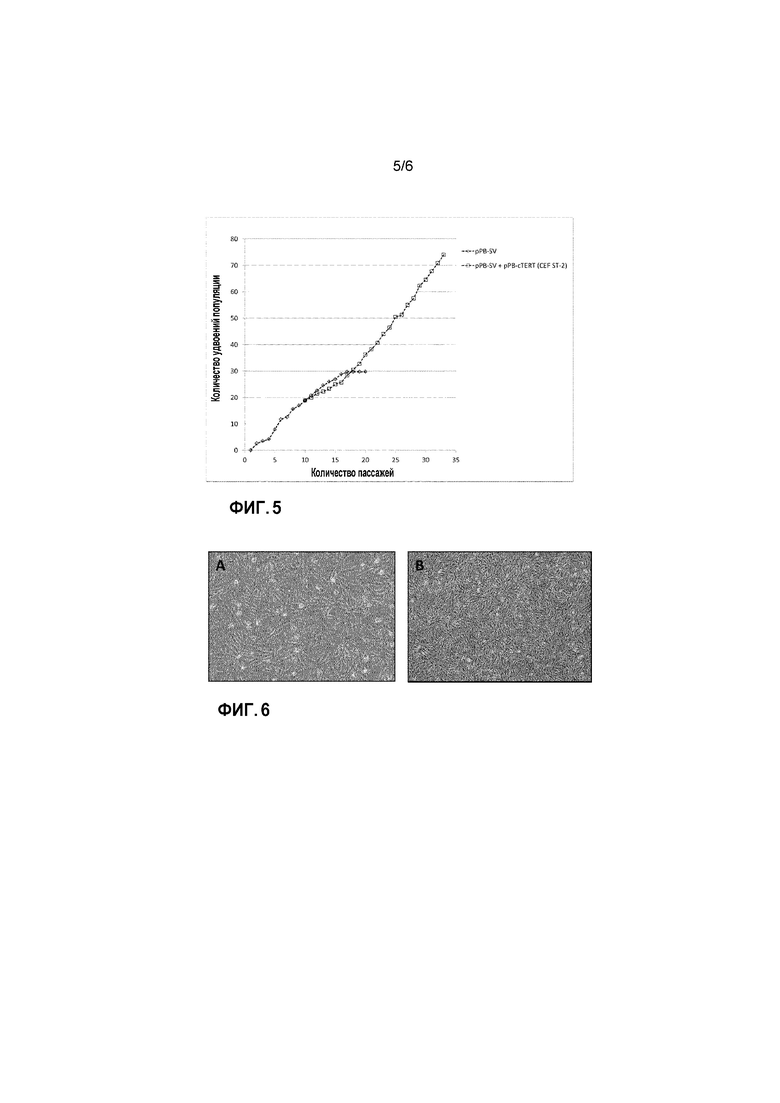
Фигура 5: Кривая роста линии клеток ФЭЦ ST-2. Клетки, трансфицированные с pPB-SV (ромбы) или pPB-SV и pPB-cTERT (ФЭЦ ST-2) (квадраты), пассируют. Количество клеток определяют в каждом пассаже для подсчета удвоения популяции в каждом пассаже.
Фигура 6: Морфология ФЭЦ ST-2 остается стабильной на протяжении пассирования. Клетки ФЭЦ ST-2 фотографируют на 19 пассаже (100x)(A) и 26 пассаже (40x)(B).
Фигура 7: Репликация HVT в клетках ФЭЦ ST-2. Клетки ФЭЦ ST-2 инфицируют HVT при T=0 и показателе множественности заражения (MOI) 0,05. Клетки собирают в указанные моменты времени и определяют титры HVT при помощи титрования.
Фигура 8: Клетки ФЭЦ ST-2 инфицируют HVT при T=0 и MOI 0,05. Клетки трипсинизируют и собирают через 96 часов и пересеивают на свежий монослой ФЭЦ ST-2. Данную процедуру повторяют 4 раза. При каждом пассаже образцы собирают для определения титра HVT.
Фигура 9: Клетки ФЭЦ ST-2 инфицируют HVT или рекомбинантными HVT вирусами при T=0 и MOI 0,05. Клетки трипсинизируют и собирают через 96 часов. Титры HVT определяют путем титрования.
Примеры.
Пример 1: Иммортализация фибробластов эмбриона цыпленка
Плазмиды.
Для создания pPB-CAG-SV40 T Ag, XhoI и BglII сайты добавляют к SV40 T антигену при помощи ПЦР с применением праймеров SV40 Tag 5'-BII (5'-GGCGAGATCTACCATGGATAAAGTTTTAAACAG-3') и SV40 Tag 3'-XI (5'-GGCGCTCGAGTTATGTTTCAGGTTCAGGGG-3'). ДНК полимеразу Phusion применяют для ПЦР в соответствии с протоколом компании-производителя (New England Biolabs). Фрагмент клонируют в pCR-Blunt (компания Life Technologies) и проверяют при помощи секвенирования. Далее SV40 T антиген вырезают из pCR-Blunt и клонируют в pPB-CAG-EBNXN (Yusaetal.,2009) при помощи BglII-XhoI сайтов для создания pPB-CAG-SV40 T Ag (Фиг. 1А). Конечную конструкцию проверяют при помощи секвенирования.
Последовательность, кодирующая cTERT, за которой следует сигнальная последовательность polyA вируса герпеса кошек (CAATAAACATAGCATACGTTATGACATGGTCTACCGCGTCTTATATGGGGACGAC) (Willemseetal.,1995), создана синтетическим путем, секвенирована и клонирована в pPB-CAG-EBNXN (Yusaetal.,2009) при помощи XhoI-ClaI сайтов для создания pPB-CAG-cTERT (Фиг. 1B и Фиг. 2). Плазмидную ДНК для трансфекции в ФЭЦ выделяют при помощи Qiagen EndoFree plasmid maxi kit (Qiagen).
Выделение и выращивание ФЭЦ.
Десять свободных от специфической патогенной микрофлоры (SPF) яиц инкубируют при 37°C в течение десяти дней и используют для выделения первичных фибробластов эмбриона. Эмбрионы собирают из яиц в стерильных условиях. После удаления головы, ног и крыльев эмбрионы трехкратно промывают в стерильном фосфатно-солевом буфере (PBS) и диссоциируют с применением раствора трипсина/этилендиаминтетрауксусной кислоты (EDTA, ЭДТК). После диссоциации для инактивации трипсина добавляют фетальную телячью сыворотку. Выделенные клетки центрифугируют в течение 15 минут при 400xg. Осажденные клетки повторно суспендируют в минимальной среде Игла, модифицированной по способу Дульбекко (DMEM), содержащей 5% фетальной телячьей сыворотки (Moregate), 1% сыворотки цыпленка (Sigma), 2 ммоль глутамина, 1 ммоль пирувата натрия и антибиотики, окрашивают для определения жизнеспособности и подсчитывают. 2,105 клеток/см2 высеивают в колбе для культур и инкубируют при 40°C и 5% CO2.
Трансфекция
После 3дней в культуре собирают ФЭЦ и подсчитывают жизнеспособные клетки перед трансфекцией. При трансфекции 1,106 жизнеспособных клеток трансфицируют в 100 мкл буфера с первичными клетками P3+добавка (Lonza Cologne AG) с применением программы CA-137 на приборе Nucleofector 4D (Lonza Cologne AG). Клетки трансфицируют с 1,6 мкг pPB-CAG-SV40 T Ag и 0,4 мкг pPB-CMV-hyPBase (Yusa et al., 2011) или, в качестве контроля, с 1,6 мкг pPB-CAG-EBNXN и 0,4 мкг 1,6 мкг pPB-CAG-hyPBase. После прохождения импульсов клетки оставляют при комнатной температуре (RT) на 5 минут. Далее к клеткам медленно добавляют 400 мкл RPMI 1640 (37°C), и клетки инкубируют при 37°C в течение 5 минут. Далее клетки аккуратно повторно суспендируют, высеивают на T25 колбы в среду для роста и инкубируют при 40°C и 5% CO2. Трансфекцию ФЭЦ+pPB-CAG-SV40 T Ag клеток с pPB-CAG-cTERT проводят с применением такого же протокола. Здесь ФЭЦ, стабильно трансфицированные с pPB-CAG-SV40 T Ag, трансфицируют с 1,6 мкг pPB-CAG-cTERT и 0,4 мкг pPB-CMV-hyPBase (Yusaetal.,2011) или, в качестве контроля, либо с 1,6 мкг pPB-CAG-EBNXN и 0,4 мкг pPB-CMV-hyPBase, либо с 2 мкг стандартного экспрессионного GFP вектора (pmaxGFP, Lonza).
Тканевая культура
После трансфекции культуры выращивают в среде для роста (DMEM, содержащей 5% фетальной телячьей сыворотки (Moregate), 1% сыворотки цыпленка (Sigma), 2 ммоль глутамина, 1 ммоль пирувата натрия) и стандартно пассируют с 80-90% конфлюентностью. После удаления среды клетки двукратно промывают при помощи PBS и трипсинизируют с применением раствора трипсина/ЭДТК. Клетки повторно суспендируют в среде для роста и осаждают. Осажденные клетки повторно суспендируют в среде для роста и подсчитывают с применением счетной камеры Бюркера (Bürker-Türk). Клетки высеивают на свежую среду в колбы для тканевой культуры с покрытием Cellbind (Corning) и инкубируют при 40°C и 5% CO2. ФЭЦ+pPB-CAG-SV40 клетки замораживают для хранения в жидком азоте на стадиях различных пассажей в стандартной среде, содержащей 42,5% фетальной телячьей сыворотки (FCS) и 10% DMSO. ФЭЦ+pPB-CAG-SV40 T Ag клетки, трансфицированные с p-PB-CAG-cTERT или с контрольными плазмидами, высеивают на поверхности, покрытые коллагеном I (Biocoat, Corning) в среде для нормального роста или в среде, не содержащей компонентов животного происхождения+1% сыворотки новорожденных телят (Hyclone, ThermoFisher). Количество удвоений популяции подсчитывают при помощи следующего уравнения:
Удвоение популяции=logNt - logN,
log 2
где Nt представляет собой количество жизнеспособных клеток в конце периода роста, и N - количество высеянных клеток (VenkatesanandPrice,1998). Клетки фотографируют при помощи камеры Olympus DP21, совмещенной с микроскопом Olympus CKX41.
Инфицирование
Клетки засеивают за 24 часа перед инфицированием с плотностью 1,105 клеток/см2 в среду, не содержащую компонентов животного происхождения, содержащую или не содержащую 0,25% сыворотки новорожденных телят, и инкубируют при 40°C и 5% CO2. Клетки инфицируют с MOI 0,05 бляшкообразующих единиц (PFU, БОЕ). Применяют вирусы HVT (FC126), рекомбинантный HVT, экспрессирующий как NDV F, так и IBDV VP2 антиген (HVT-NDV-IBDV), и рекомбинантный HVT, экспрессирующий ILT gD/gI антиген (HVT-ILT). После инфицирования клетки инкубируют при 38,5°C и 5% CO2. Среду полностью обновляют через 72 часа в случае, если клетки инкубируют более 72 часов. В зависимости от эксперимента образцы для титрования собирают через 72, 96, 120 или 144 часа после инфицирования и замораживают для хранения в жидком азоте. После удаления среды клетки двукратно промывают PBS и трипсинизируют при помощи раствора трипсина/ЭДТК. Клетки повторно суспендируют в среде для роста и осаждают. После удаления надосадочной жидкости клетки повторно суспендируют в свежей среде для роста и подсчитывают. Далее 1,107/мл клеток замораживают в среде для роста, содержащей конечную концентрацию 10% DMSO и 20% фетальной телячьей сыворотки, и хранят в жидком азоте.
Титрование
Ампулы замороженных инфицированных HVT клеток оттаивают и определяют количество БОЕ/мл путем титрования инфицированных HVT клеток на ФЭЦ. Бляшки визуализируют при помощи иммунофлуоресцентного анализа с применением моноклональных антител или поликлональной сыворотки цыпленка, распознающих HVT. Козьи антитела к иммуноглобулинам цыплят Alexa 488 или козьи антитела к иммуноглобулинам мышей Alexa 488 применяют в качестве вторичных антител, соответственно. Все титрования проводят в двух параллельных анализах.
Результаты
Экспрессия SV40 T антигена удлиняет жизненный цикл первичных ФЭЦ.
Первичные ФЭЦ трансфицируют на 1 пассаже с pPB-CAG-SV40 T Ag (pPB-SV) и экспрессионным вектором транспозазы piggyBac для получения геномной интеграции и стабильной экспрессии SV40 T Ag. В качестве контроля ФЭЦ также трансфицируют с пустым pPB-CAG-EBNXN (pPB-EV) вектором. После трансфекции клетки стандартно пассируют при 80-90% конфлюентности и подсчитывают для определения жизнеспособных клеток. Количество жизнеспособных клеток при пассировании применяют для подсчета количества удвоений популяции после высеивания (удвоения популяции (PD, УП)). Несмотря на то, что экспрессирующие SV40 T Ag клетки демонстрируют удлиненный жизненный цикл по сравнению с контролями (35 удвоений популяции по сравнению с 26 удвоениями популяции, соответственно), все культуры в конце концов прекращают рост приблизительно на 17 пассаже (Фиг. 3). После 17 пассажа пролиферирующие клетки все еще присутствуют в культурах. Тем не менее, поскольку многие клетки в культуре погибают, увеличения общего количества клеток не обнаруживают, и в конце концов все клетки погибают.
ФЭЦ+pPB-SV клетки исследуют на предмет экспрессии SV40 T Ag на этапах различных пассажей при помощи иммунофлуоресцентного анализа с применением моноклонального антитела, специфического по отношению к SV40 T Ag. На 2 пассаже, через один пассаж после трансфекции, небольшое количество клеток является SV40 T Ag позитивным. Приблизительно все клетки экспрессируют SV40 T Ag после 7 пассажа (данные не показаны).
Экспрессия cTERT в экспрессирующих SV40 T Ag клетках индуцирует иммортализацию.
Линия клеток ФЭЦ ST-1
Для получения иммортализованных ФЭЦ изобретатели трансфицируют ФЭЦ+pPB-SV клетки с pPB-CAG-cTERT (pPB-cTERT) экспрессионным вектором. Ампулы с 13 пассажем ФЭЦ+pPB-SV клеток оттаивают, клетки собирают в культуру, пассируют и трансфицируют на 18 пассаже с pPB-cTERT и экспрессионным вектором транспозазы. Общее количество ФЭЦ+pPB-SV клеток на 18 пассаже низкое. Таким образом, изобретатели применяют ФЭЦ+pPB-SV клетки, трансфицированные при помощи стандартного GFP экспрессионного вектора (EV), для определения эффективности трансфекции также в качестве пустых векторов контролей во время пассирования. После трансфекции клетки высеивают и ежедневно изучают на предмет пролиферации и роста колоний. Пролиферация не наблюдается в культурах ФЭЦ+pPB-SV+EV, и клетки в конце концов погибают (данные не показаны). В культурах ФЭЦ+pPB-SV+pPB-cTERT, тем не менее, быстро пролиферирующие колонии становятся отчетливо видны через 10 дней. Данные колонии трипсинизируют, клетки подсчитывают и высеивают в колбы для тканевых культур. Изобретатели обнаружили, что данные клетки пролиферируют лучше в среде, не содержащей компонентов животного происхождения (ACF), которая является стандартной средой для роста (данные не показаны), таким образом изобретатели продолжают выращивание данных клеток в ACF среде. Данные ФЭЦ+pPB-SV+pPB-cTERT клетки продолжают активно расти, и их пассируют до того, как они дают более 100 удвоений популяции (УП) (Фиг. 3).
На данном этапе клетки все еще здоровы и активно пролиферируют. Таким образом, изобретатели приходят к выводу, что ими получена иммортализованная линия клеток ФЭЦ, которую назвали ФЭЦ ST-1 (для ФЭЦ+SV40 T Ag+cTERT-номер1). Клетки имеют морфологию фибробластов, которая сохраняется во время пассирования (Фиг. 4).
ФЭЦ ST-1 клетки в различных пассажах замораживают в ампулах для хранения в жидком азоте. Данные клетки можно легко использовать для повторного выращивания после хранения в жидком азоте.
Линия клеток ФЭЦ ST-2
ФЭЦ ST-1 получают путем трансфицирования 18 пассажа ФЭЦ+pPB-SV клеток с cTERT экспрессионным вектором. На данном этапе ограниченное количество клеток все еще пролиферирует, и многие клетки в культуре уже погибли. Поскольку только небольшое число иммортализованных колоний выросло после трансфекции и дало рост линии клеток ФЭЦ ST-1, изобретатели приходят к выводу, что ФЭЦ ST-1 представляет собой олигоклональную линию клеток.
В целью получения поликлональной линии клеток, например, линии клеток, происходящей от многих различных иммортализованных клеток ФЭЦ, изобретатели также трансфицируют ранний пассаж ФЭЦ+pPB-SV клеток с cTERT. ФЭЦ+pPB-SV клетки, которые хранят в жидком азоте на 9 пассаже, собирают в культуру, пассируют в ACF среде и трансфицируют на 11 пассаже клеток с pPB-cTERT или pPB-EV в комбинации с экспрессионным вектором транспозазы. После трансфицирования клетки высеивают и пассируют. ФЭЦ+pPB-SV+pPB-EV клетки прекращают пролиферировать и погибают после 16 пассажа (данные не показаны). Клетки, трансфицированные в pPB-SV и pPB-cTERT, продолжают активно пролиферировать и их пассируют до того момента, как они дают более 70 УП (пассаж 33, Фиг. 5).
На данном этапе клетки все еще здоровы и хорошо пролиферируют. Таким образом, изобретатели приходят к выводу, что они создали другую иммортализованную линию клеток ФЭЦ. Данная линия клеток названа ФЭЦ ST-2 (для ФЭЦ+SV40 T Ag+cTERT-номер2). ФЭЦ ST-2 клетки имеют морфологию фибробластов, которая сохраняется на протяжении пассирования (Фиг. 6). ФЭЦ ST-2 клетки различных пассажей замораживают в ампулах для хранения в жидком азоте. Данные клетки можно легко использовать для повторного выращивания после хранения в жидком азоте.
Репликация HVT на ФЭЦ ST-1 и ФЭЦ ST-2.
Изобретатели тестируют как ФЭЦ ST-1, так и ФЭЦ ST-2 линии клеток на предмет их способности поддерживать репликацию вируса герпеса индеек (HVT). Во-первых, при помощи иммунофлуоресцентного анализа (IF, ИФ) изобретатели демонстрируют, что ФЭЦ ST-1 можно инфицировать HVT, и клетки поддерживают репликацию HVT. ФЭЦ ST-1 клетки инфицируют HVT и фиксируют для ИФ через 72, 96 и 120 часов. HVT-позитивные очаги наблюдаются в инфицированных HVT ФЭЦ ST-1 клетках во все моменты времени, что является показателем того, что ФЭЦ ST-1 клетки можно инфицировать HVT и поддерживать передачу HVT от клетки к клетке (данные не показаны).
Во втором эксперименте изобретатели сравнивают эффективность инфицирования HVT и передачу от клетки к клетке между клетками ФЭЦ ST-1 и ФЭЦ ST-2. И ФЭЦ ST-1 и ФЭЦ ST-2 инфицируют HVT и фиксируют в различные моменты времени для изучения динамики репликации HVT в данных линиях клеток. HVT-позитивные очаги присутствуют в обоих линиях клеток через 24, 48 и 72 часа, и количество и размер очагов увеличиваются во времени (данные не показаны). Из данных экспериментов очевидно, что большее количество очагов HVT, а также более крупные очаги наблюдаются в ФЭЦ ST-2 по сравнению с ФЭЦ ST-1. Таким образом, изобретатели приходят к выводу, что ФЭЦ ST-2 клетки являются лучшим субстратом для репликации HVT по сравнению с ФЭЦ ST-1.
Далее изобретатели исследуют динамику репликации в ФЭЦ ST-2 более подробно. ФЭЦ ST-2 клетки инфицируют HVT, и забор образцов осуществляют через 72, 96, 120 и 144 часа после инфицирования для определения количества бляшкообразующих единиц (БОЕ) в мл (Фиг. 7). Данные результаты показывают, что HVT реплицируется в ФЭЦ ST-2 клетках, и хотя различия в титрах в различные моменты времени сбора малы, кажется, что титры вируса достигают пиковых значений приблизительно через 96 часов после инфицирования (Фиг. 7).
HVT можно пассировать на CEF ST-2 клетках.
После того, как изобретатели установили, что ФЭЦ ST-2 является субстратом для инфицирования и репликации HVT, они исследуют, могут ли инфицированные HVT ФЭЦ ST-2 клетки инфицировать свежие монослои ФЭЦ ST-2 клеток. ФЭЦ ST-2 клетки инфицируют HVT, собирают через 96 часов и высеивают на монослой свежих ФЭЦ ST-2 клеток. Данную процедуру повторяют 4 раза, и при каждом пассаже клетки фиксируют для ИФ окрашивания, и производят забор образцов для определения количества БОЕ/мл. ИФ окрашивание демонстрирует, что HVT-позитивные очаги присутствуют после каждого пассажа, и большое количество неокрашенных очагов наблюдается после четвертого пассажа (данные не показаны). Титрование демонстрирует, что HVT титры на 2 пассаже незначительно ниже по сравнению с титрами 1 пассажа (Фиг. 8). Тем не менее, последующее пассирование на ФЭЦ ST-2 клетках приводит к повышению титров HVT с каждым пассажем, и на 5 пассаже титры HVT составляют 105,7 БОЕ/мл. Это ясно показывает, что ФЭЦ ST-2 линия клеток является подходящим субстратом для репликации HVT.
ФЭЦ ST-2 является субстратом для репликации рекомбинантных HVT конструкций.
HVT также можно применять в качестве вирусного вектора для экспрессии антигенов, например, других вирусов птиц, таких как вирус болезни Ньюкасла (NDV), вирус инфекционного бурсита (IBDV) или вирус инфекционного ларинготрахеита (ILT) (Iqbal,2012). Для того, чтобы протестировать, позволяют ли также ФЭЦ ST-2 репликацию рекомбинантных HVT конструкций, ФЭЦ ST-2 инфицируют либо HVT дикого типа, либо рекомбинантным HVT, экспрессирующим как NDV F, так и IBDV VP2 антиген (HVT-NDV-IBDV), либо рекомбинантным HVT, экспрессирующим ILT gD/gI антиген (HVT-ILT). Клетки фиксируют для ИФ окрашивания, или клетки собирают для создания образцов для титрования через 96 часов после инфицирования. ИФ окрашивание демонстрирует очаги как для HVT дикого типа, так и для рекомбинантных HVT конструкций в ФЭЦ ST-2 клетках (данные не показаны). Титрование образцов также демонстрирует, что ФЭЦ ST-2 клетки поддерживают репликацию HVT рекомбинантов (Фиг. 9), хотя репликация рекомбинантных HVT является менее эффективной по сравнению с репликацией HVT дикого типа в используемых условиях.
Список литературы
Iqbal,M. (2012). Progress toward the development of polyvalent vaccination strategies against multiple viral infections in chickens using herpesvirus of turkeys as vector. Bioengineered. 3, 222-226.
Venkatesan,R.N. and Price,C. (1998). Telomerase expression in chickens: constitutive activity in somatic tissues and down-regulation in culture. Proc. Natl. Acad. Sci. U. S. A 95, 14763-14768.
Willemse,M.J., Strijdveen,I.G., van Schooneveld,S.H., van den Berg,M.C., and Sondermeijer,P.J. (1995). Transcriptional analysis of the short segment of the feline herpesvirus type 1 genome and insertional mutagenesis of a unique reading frame. Virology 208, 704-711.
Yusa,K., Rad,R., Takeda,J., and Bradley,A. (2009). Generation of transgene-free induced pluripotent mouse stem cells by the piggyBac transposon. Nat. Methods 6, 363-369.
Yusa,K., Zhou,L., Li,M.A., Bradley,A., and Craig,N.L. (2011). A hyperactive piggyBac transposase for mammalian applications. Proc. Natl. Acad. Sci. U. S. A 108, 1531-1536.

Группа изобретений относится к области генетической инженерии, в частности к иммортализованным фибробластам эмбриона цыпленка (ФЭЦ) для размножения вирусов, способам их получения, к культурам клеток, включающим иммортализованные ФЭЦ, к способам репликации вирусов птиц в таких клетках и к способам получения вакцин. Стабильно трансфицированный иммортализованный ФЭЦ экспрессирует SV40 T антиген, экспрессирует теломеразу цыпленка (cTERT) под контролем гетерологичного промотора, не включает экзогенную ретровирусную ДНК с Длинным концевым повтором, обладает способностью к непрерывной пролиферации и дает более 45 удвоений популяции. Способы получения иммортализованных ФЭЦ включают трансфекцию первичных клеток ФЭЦ при помощи одной или нескольких молекул ДНК, не содержащих ДКП последовательности и включающих инвертированные повторы транспозона, ген, кодирующий SV40 T антиген под контролем подходящего промотора, ген, кодирующий cTERT под контролем подходящего промотора, ген, кодирующий транспозазу под контролем подходящего промотора. В результате отбирают клетки ФЭЦ, которые культивируют в течение по меньшей мере 45 клеточных циклов. Полученные клетки ФЭЦ имеют морфологию фибробластов, которая сохраняется на протяжении пассирования, и являются субстратом для репликации вирусов птиц или вирусного вектора птиц для дальнейшего получения вакцин. 9 н. и 7 з.п. ф-лы, 9 ил., 1 пр.
1. Стабильно трансфицированный иммортализованный фибробласт эмбриона цыпленка (ФЭЦ) для размножения вирусов, отличающийся тем, что указанный иммортализованный ФЭЦ экспрессирует SV40 T антиген, экспрессирует теломеразу цыпленка (cTERT) под контролем гетерологичного промотора, не включает экзогенную ретровирусную ДНК с Длинным концевым повтором, обладает способностью к непрерывной пролиферации и продолжает пролиферировать по меньшей мере до 45 удвоений популяции.
2. Культура клеток для размножения вирусов, включающая иммортализованные ФЭЦ по п.1.
3. Культура клеток по п.2, отличающаяся тем, что культуру клеток инфицируют вирусом птиц или вирусным вектором птиц.
4. Культура клеток по п.3, отличающаяся тем, что вирус птиц или вирусный вектор птиц выбирают из группы, состоящей из вируса болезни Марека (MDV), MDV-родственного вируса герпеса индеек (HVT), вируса болезни Ньюкасла (NDV), вируса инфекционного бронхита (IBV), вируса инфекционного бурсита (IBDV), вируса синдрома снижения несучести (EDSV), вируса ринотрахеита индеек (TRT), реовируса (RV), а также HVT вектора, включающего IBDV VP2-ген, IBV-ген белков шиловидного отростка, HA-ген птичьего гриппа, ILT-ген белка gD/gI или F-ген NDV.
5. Способ получения иммортализованных ФЭЦ по п.1, отличающийся тем, что указанный способ включает стадии:
а) получения первичных клеток ФЭЦ,
b) трансфекции указанных ФЭЦ при помощи 1) молекулы ДНК, не содержащей ДКП последовательности и включающей инвертированные повторы транспозона, и включающей ген, кодирующий SV40 T антиген под контролем подходящего промотора, 2) молекулы ДНК, не содержащей ДКП последовательности и включающей инвертированные повторы транспозона, и включающей ген, кодирующий теломеразу цыпленка cTERT под контролем подходящего промотора, и 3) молекулы ДНК, включающей ген, кодирующий транспозазу под контролем подходящего промотора,
с) отбора клеток ФЭЦ, которые культивируют в течение по меньшей мере 45 клеточных циклов.
6. Способ получения иммортализованных ФЭЦ по п.1, отличающийся тем, что указанный способ включает стадии:
a) получения первичных клеток ФЭЦ,
b) трансфекции указанных клеток ФЭЦ при помощи одной молекулы ДНК, не содержащей ДКП последовательности и включающей инвертированные повторы транспозона, и включающей как ген, кодирующий SV40 T антиген под контролем подходящего промотора, так и ген, кодирующий теломеразу цыпленка под контролем подходящего промотора, и включающей ген, кодирующий транспозазу под контролем подходящего промотора,
c) отбора клеток ФЭЦ, которые культивируют в течение по меньшей мере 45 клеточных циклов.
7. Способ получения иммортализованных ФЭЦ по п.1, отличающийся тем, что указанный способ включает стадии:
a) получения первичных клеток ФЭЦ,
b) трансфекции указанных клеток ФЭЦ при помощи 1) молекулы ДНК, не содержащей ДКП последовательности и включающей инвертированные повторы транспозона, и включающей ген, кодирующий SV40 T антиген под контролем подходящего промотора, и ген, кодирующий теломеразу цыпленка под контролем подходящего промотора, и при помощи 2) молекулы ДНК, включающей ген, кодирующий транспозазу под контролем подходящего промотора,
с) отбора клеток ФЭЦ, которые культивируют в течение по меньшей мере 45 клеточных циклов.
8. Способ по любому из пп. 5-7, отличающийся тем, что на стадии с) выбирают клетки, которые культивируют по меньшей мере в течение 100 клеточных циклов.
9. Способ репликации вируса птиц или вирусного вектора птиц, где указанный способ включает стадии: a) культивирования иммортализованных ФЭЦ по п.1, b) контактирования иммортализованных ФЭЦ с вирусом птиц или вирусным вектором птиц, c) обеспечения репликации вируса птиц или вирусного вектора птиц и d) выделения вирусного потомства.
10. Способ по п.9, отличающийся тем, что вирус птиц или вирусный вектор птиц выбирают из группы вирусов птиц, состоящей из вируса болезни Марека (MDV), MDV-родственного вируса герпеса индеек (HVT), вируса болезни Ньюкасла (NDV), вируса инфекционного бронхита (IBV), вируса инфекционного бурсита (IBDV), вируса синдрома снижения несучести (EDSV), вируса ринотрахеита индеек (TRT), реовируса (RV), а также HVT вектора, включающего IBDV VP2-ген, IBV-ген белков шиловидного отростка, HA-ген птичьего гриппа, ILT-ген белка gD/gI или F-ген NDV.
11. Способ по п.9, отличающийся тем, что указанный вирус птиц или вирусный вектор птиц выбирают из группы вирусов птиц, состоящей из MDV или векторного вируса MDV.
12. Способ получения вакцины, включающей вирус птиц или вирусный вектор птиц, отличающийся тем, что способ включает стадию смешивания культуры клеток по п.3 или 4 с фармацевтически приемлемым носителем.
13. Способ по п.12, отличающийся тем, что вирус птиц или вирусный вектор птиц представляет собой MDV.
14. Способ по п.12, отличающийся тем, что MDV или вирусный вектор MDV в вакцине находятся в живой аттенуированной форме.
15. Вакцина против вируса птиц, включающая эффективное количество культуры клеток по п.3 или 4 и фармацевтически приемлемый носитель.
16. Способ получения вакцины, включающей вирус птиц или вирусный вектор птиц, отличающийся тем, что данный способ включает стадии:
a) инфицирования культуры клеток по п.2 вирусом птиц или вирусным вектором птиц,
b) репликации указанного вируса птиц или вирусного вектора птиц,
с) выделения вирусного потомства,
d) смешивания вирусного потомства с фармацевтически приемлемым носителем.
| US 2014363467 A1, 11.12.2014 | |||
| WO 2007077256 Al, 12.07.2007 | |||
| WO 2009004016 A1, 08.01.2009 | |||
| WO 9744443 A1, 27.11.1997 | |||
| Холодильная камера с непосредственной холодопередачей через стенки резервуаров | 1928 |
|
SU12828A1 |

