Область техники, к которой относится изобретение
Данное изобретение относится к области биотехнологии, и, в частности, к слитому белку с увеличенной эритропоэтиновой (далее “ЕРО”) активностью in vivo в качестве лекарственного средства против злокачественной анемии. Более конкретно, данное изобретение относится к слитому белку с увеличенной ЕРО-активностыо, обеспеченной за счет увеличения времени полужизни в организме, причем слитый белок включает природные аминокислотные последовательности, но без увеличения уровня гликозилирования, другими словами, слитый белок включает ЕРО-молекулу, слитую с определенным пептидом, природно встречающимся in vivo.
Уровень техники
ЕРО представляет собой гликопротеин с мол.массой 30000-34000 Да и является гемопоэтическим фактором, стимулирующим продуцирование и дифференцировку эритроцитов. Этот гликопротеин связывается с рецептором клеток-предшественников эритроцитов для инициации его гемопоэтической активности и вызывает увеличение количества внутриклеточных ионов кальция, усиление биосинтеза ДНК и стимуляцию образования гемоглобина. Рекомбинантный EPO(rhEPO) был одобрен также для лечения анемии, связанной с почечной недостаточностью, недоношенностью, гипотиреозом, нарушением питания и т.д., и клиническое применение rhEPO непрерывно увеличивается. Однако интенсивное применение rhEPO ограничивается его достаточно высокой стоимостью, поскольку rhEPO необходимо вводить около трех раз в неделю вследствие его короткого периода полужизни в организме. Таким образом, частота введения rhEPO при лечении могла бы быть уменьшена путем сохранения ЕРО-активности in vivo в течение более продолжительного времени.
Биологическая активность ЕРО in vivo пропорциональна его периоду полужизни в организме in vivo, который, как известно, связан с содержанием сиаловой кислоты, расположенной на конце сахарных цепей ЕРО. Таким образом, биологическая активность ЕРО in vivo в значительной степени зависит от присутствия или отсутствия сахарных цепей. Типы сахарных цепей варьируются в зависимости от типов клеток. Так, когда один и тот же гликопротеин экспрессируется в различных клетках, типы сахарных цепей этого белка характерным образом различаются в зависимости от типов клеток. Известно, что бактериальные клетки, например Е. coli, не могут присоединять сахарные цепи к их белкам. Известно, что белки, экспрессируемые в Е. coli, не содержат сахарных цепей. ЕРО, экспрессируемый в Е. coli, также не будет содержать сахарных цепей. Было подтверждено, что в этом случае ЕРО обладает биологической активностью in vitro, но полностью не активен in vivo. Это связано с тем, что ЕРО без сахарных цепей быстрее удаляется из организма по сравнению с ЕРО с сахарными цепями, что приводит к чрезвычайно короткому периоду полужизни в организме. Таким образом, присутствие или отсутствие сахарных цепей в ЕРО играет важную роль в биологической активности ЕРО.
К настоящему времени были предприняты многочисленные исследования, направленные на увеличение биологической активности ЕРО. Большинство таких работ основано на замене некоторых аминокислот индуцированием мутаций в гене ЕРО с помощью методов направленного мутагенеза. Например, в PCT/US94/09257, озаглавленный "Аналоги эритропоэтина" (Erythropoietin Analogs), поданный Amgen Inc., раскрыт способ увеличения периода полужизни in vivo посредством увеличения содержания сахарных цепей в ЕРО с помощью мутагенеза. Были сделаны попытки увеличения периода полужизни in vivo посредством образования димеров ЕРО (A.J. Sytkowski et al. J.B.C. vol. 274, No. 35, pp 24773-24778). Другие способы увеличения биологической активности ЕРО in vivo включают слияние новых аминокислот, пептида или фрагмента белка с молекулами ЕРО методами генной инженерии и увеличение содержания сахарных цепей в ЕРО, в частности количеств сиаловых кислот. Однако разновидности аминокислот, пептидов и фрагментов белков, используемых в этих способах, являются очень ограниченными. В большинстве случаев такие генетические модификации могут приводить к снижению или потере удельной активности белка или вызывать проблемы антигенности, часто встречающиеся при применении таких веществ in vivo.
Также были проведены исследования других слитых белков или химерных белков, отличных от ЕРО, и одним из примеров которых является фолликулостимулирующий гормон, который является половым гормоном (Furuhashi et al., 1995, Mol. Endocrinol.). Однако такие белки пока еще не нашли широкого применения в данной области, поскольку с получением генетически модифицированных белков методами генной инженерии связано несколько проблем. Получение самого модифицированного белка-мишени не является легким и требует высокой профессиональной квалификации. Кроме того, в большинстве случаев активность модифицированных белков может значительно снизиться при добавлении новых или замене аминокислот.
Авторы настоящего изобретения начали интенсивные исследования по развитию нового способа увеличения активности ЕРО in vivo путем слияния новых аминокислот, пептидов или белков с молекулой ЕРО. В ходе проведения этих исследований было обнаружено, что слитый белок, полученный слиянием фрагментов С-концевого пептида (далее называемого также "СТР") (β-субъединицы хорионического гонадотропина человека (далее называемого также "HCG"), который является природным белком, встречающимся in vivo, с ЕРО существенно увеличивает период полужизни in vivo модифицированного таким образом ЕРО. Кроме того, такой ЕРО содержит аминокислоты, образующие сайты гликозилирования, при сохранении присущей ЕРО активности (см. Заявку на патент Кореи Korean Patent Application No. 10-2000-0075230).
Сущность изобретения
Авторы данного изобретения неожиданно обнаружили, что варианты СТР, из которых был удален сайт гликозилирования, также заметно увеличивают период полужизни ЕРО in vivo. В результате этого открытия были разработаны варианты СТР, которые могут увеличивать стабильность ЕРО in vivo, путем включения в ЕРО аминокислотных последовательностей, без увеличения содержания сахарных цепей в ЕРО, что позволило авторам решить задачи, поставленные в изобретении.
Данное изобретение, в котором используются варианты СТР, из которых были удалены сайты гликозилирования, отличается от предшествующего уровня техники, в котором период полужизни ЕРО in vivo увеличивается путем увеличения содержания сахарных цепей в ЕРО, и основано на открытии вариантов СТР, способных увеличивать стабильность ЕРО in vivo.
Таким образом, целью данного изобретения является получение слитого белка, обладающего увеличенной активностью ЕРО человека in vivo, включающего молекулу-ЕРО, слитую с вариантом СТР β-субъединицы HCG на ее С-конце.
Другой целью данного изобретения является получение нуклеиновой кислоты, которая кодирует указанный слитый белок, рекомбинантного вектора, содержащего указанную нуклеиновую кислоту, и клеточной линии, трансфицированной указанным рекомбинантным вектором (плазмидой).
Еще одной целью данного изобретения является разработка способа получения слитого белка с увеличенной активностью ЕРО человека путем культивирования трансформированной клеточной линии.
В одном варианте данного изобретения обеспечен слитый белок с увеличенной активностью ЕРО человека in vivo, включающий ЕРО-молекулу, слитую с СТР-вариантом β-субъединицы HCG (далее называемым также "АТР") на ее С-конце. Предпочтительно этот СТР включает все аминокислоты или часть аминокислот, соответствующих положениям 112-145, предпочтительно положениям 118-145 β-субъединицы HCG, приведенные в последовательности SEQ ID NO:l.
АТР, представляющий собой вариант СТР, выполняет функцию увеличения периода полужизни молекулы ЕРО с ее собственными аминокислотными последовательностями, т.е. без увеличения содержания сахарных цепей ЕРО.
Таким образом, до тех пор, пока не появляется негативное влияние на активность слитого белка-мишени, увеличивающего активность ЕРО in vivo, положения и типы аминокислот, меняющихся в пределах вышеуказанного диапазона, не являются специфически ограниченными. Другими словами, пока слитый белок выполняет функцию увеличения активности ЕРО in vivo, аминокислоты в положениях, относящихся к вышеуказанному диапазону, могут замещаться в любом положении любыми аминокислотами.
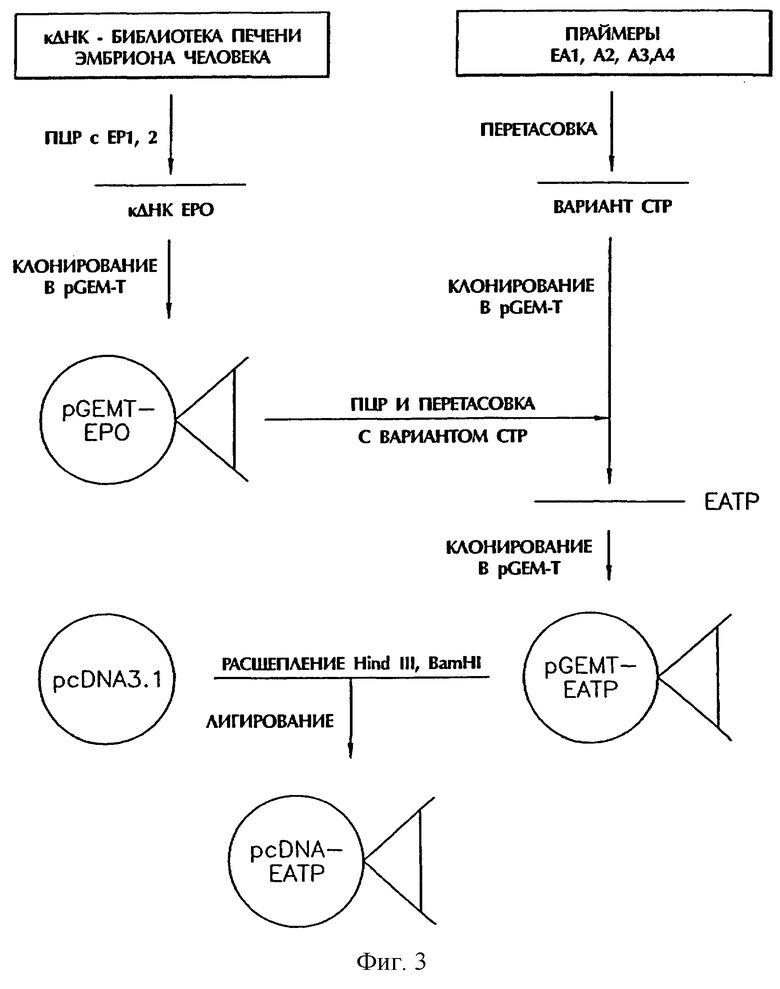
Например, предпочтительно, чтобы АТР имел одну или несколько замен в положениях 121, 127, 132 и 138 и наиболее предпочтительно имел остатки серина (Ser) в положениях 121, 127, 132 и 138, замененные остатками аланина (Аlа) (фиг.1). В этом случае слитый белок данного изобретения имеет аминокислотную последовательность, описанную SEQ ID NO:2. Тот факт, что остатки Ser в вышеуказанных положениях могут быть заменены остатками Аlа, позволяет специалисту в данной области сделать вывод, что любая другая аминокислота, имеющая размер и заряд, сходные с размером и зарядом Аlа, например глицин (Gly), может также заменять Ser.
В другом варианте данного изобретения обеспечены нуклеиновая кислота, которая кодирует слитый белок, рекомбинантный вектор, содержащий эту нуклеиновую кислоту, и линия клеток, трансфицированных рекомбинантной плазмидой, предпочтительно, клеток яичника Китайского хомячка (СНО).
Еще в одном варианте данного изобретения обеспечен способ получения слитого белка, имеющего увеличенную активность ЕРО человека путем культивирования этой линии трансформированных клеток.
Перечень фигур
Описанные выше цели и преимущества данного изобретения станут более очевидными из подробного описания предпочтительных вариантов изобретения со ссылкой на прилагаемые чертежи, в которых:
на фиг.1 представлены нуклеотидная и аминокислотная последовательности АТР как один из вариантов СТР;
на фиг.2 представлены нуклеотидная и аминокислотная последовательности слитого белка (ЕАТР), включающего последовательности ЕРО и АТР;
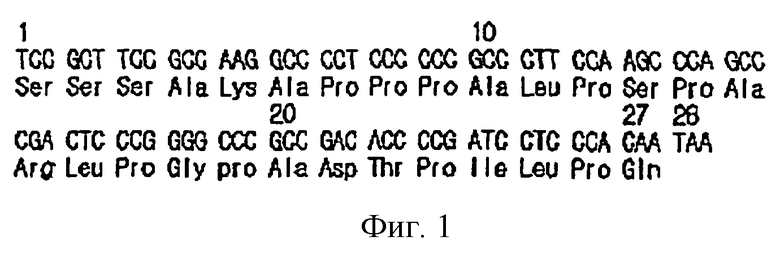
на фиг.3 показана схема получения экспрессирующего вектора pcDNA3.1-ЕАТР;
на фиг.4 приведена электрофореграмма очищенного ЕАТР;
на фиг.5 показан фармакокинетический анализ ЕАТР и ЕРО.
Подробное описание изобретения
Данное изобретение будет теперь описано более подробно. Данное изобретение описывает получение и клонирование генов слитого белка-мишени, конструирование векторов, экспрессирующих гены-мишени, трансфекция клеток животных и экспрессии ЕАТР и очистки экспрессируемого ЕАТР и измерения активности.
(1) Получение генов
Комплементарную ДНК (кДНК) ЕРО получали с помощью общепринятого способа полимеразной цепной реакции с обратной транскрипцией (OT-PCR) с использованием набора RT-PCR Premix Kit, доступного из Bioneer Corp., Korea, в котором используются праймеры ЕР1 и ЕС2, комплементарные обоим концам кДНК ЕРО, полученной ранее из кДНК-библиотеки эмбриональной печени человека (доступной из Invitrogen Corp.).
ЕР1: ATGGGGGCACGAATGTCCTGCCTGGCTGG (SEQ ID NO:3)
EC2: GTCCCCTGTCCTGCAGGCCT (SEQ ID NO:4)
кДНК ЕРО клонировали в клонирующий вектор pGEM-T (Promega Corp.), полученную плазмиду назвали pGEMT-EPO, a клонированный на ЕРО использовали в качестве матрицы в последующих экспериментах.
Варианты гена СТР β-субъединицы HCG, используемые в данном изобретении, получали с помощью химического синтеза и самопраймирующейся ПЦР. Полученные фрагменты генов были названы ЕА1, А2, A3 и А4:
ЕА1: AGGGGAGGCCTGCAGGACAGGGGACTCCTCTTCCG (SEQ ID NO:5)
А2: GGAAGGGC GGGGGGAGGGGCCTTG GC GGAAGAGGA (SEQ ID NO:6)
A3: CCGC CCTTCCAAGCCCAG CCCGACTCCCGGGGCCC (SEQ ID NO:7)
A4: TTATTGTGGGAGGATCGGGGTGTGG GC GGGCCCCG (SEQ ID NO:8)
(Жирным шрифтом отмечены аминокислоты, подходящие для замены).
1 мкл каждого из четырех генов использовали в (50 пмоль/мкл) ПЦР с использованием Taq-системы высокой точности (Boehringer Mannheim Corp.).
Фрагменты генов (модифицированные СТР-гены) размером приблизительно 100 п.н. определяли в 1% агарозном геле. Эти гены кодируют пептид, полученный заменой 4 остатков Ser в положениях 121, 127, 132 и 138 среди 28 С-концевых аминокислот в положениях 118-145 β-субъединицы HCG остатками Аlа (см. фиг.1).
ПЦР проводили с использованием pGEMT-EPO в качестве матрицы и ЕР1 и ЕС2 в качестве праймеров с получением только генов ЕРО. Затем ПЦР проводили с использованием обоих генов ЕРО и модифицированных СТР-генов в качестве матриц и с использованием праймеров ЕР11 и ЕР22 с использованием Taq-системы высокой точности, получая посредством этого желаемый слитый белок с фрагментами генов приблизительно 630 п.н. (также называемых ЕАТР-генами).
ЕР11: TAAGCTTATGGGGGTGCACGAATGT (SEQ ID NO:9)
ЕР22: TGGATCCTTATTGTGGGAGGATCGGGGT (SEQ ID NO:10)
Эти гены клонировали в клонирующие векторы pGEM-T и затем определяли нуклеотидные последовательности, полученные плазмиды были названы pGEMT-EATP (см. фиг.3).
(2) Конструирование экспрессирующих векторов
Вектор pcDNA3.1 (Invitrogen Corp.) использовали в качестве экспрессирующего вектора. Ген ЕАТР в pGEMT-EATP ограничен сайтом Hindi II и BamHI, происходящими от праймеров ЕР11 и ЕР22. pcDNA3.1 и полученную плазмиду pGEMT-EATP расщепляли HindIII и BamHI. Линеаризованную pcDNA3.1 и ген ЕАТР очищали с помощью электрофореза в агарозном геле с использованием набора для элюции Qiagen с последующим лигированием, и ими трансформировали Е. coli NM522. Плазмиды выделяли из колоний, образующихся после инкубации в течение ночи в твердой, содержащей ампициллин-LB среде, и расщепляли рестриктазами HindIII и BamHI. С помощью электрофореза в 1% агарозном геле отбирали колонии, содержащие встроенный ЕАТР. Полученные плазмиды были названы pcDNAS.1-EATP (см. фиг.3).
(3) Трансфекция клеток СНО и экспрессия ЕАТР
Клетки СНО (DG44) выращивали в чашках 60 мм до получения 40-80% конфлюентных клеток (1-4×105 клеток на чашку 60 мм). 3 мкл реагента Superfection (Boehringer Mannheim Corp.) и 97 мкл среды (α-МЕМ со средой, без сыворотки, без антибиотиков) тщательно перемешивали и к полученной смеси добавляли приблизительно 2 мкг ДНК плазмиды pcDNA3.1-EATP (более 0,1 мкг/мкл) и 0,2 мкг содержащего ген дигидрофолатредуктазы (dhfr) вектора pLTRdhfr26 (ATCC 37295) и инкубировали при комнатной температуре в течение 5-10 минут с последующим добавлением к этим клеткам. Спустя один день среду заменяли α-МЕМ без среды (содержащей 500 мкг/мл G418) с 10% ФТС. К клеткам добавляли среду с 500 мкг/мл G418 и культивировали в течение 7-10 дней. В течение этого времени G418-чувствительные клетки и клетки негативного контроля погибают. После достаточного культивирования клеток, резистентных к G418, в среде определяют экспрессируемый ЕАТР-белок с использованием набора ELISA для ЕРО (Boehringer Mannheim Corp.).
Очистка экспрессируемого ЕАТР
Смолы для аффинной очистки с использованием анти ЕРО моноклональных антител против ЕРО (R&D Inc.) получали следующим образом.
0,3 г CNBr-активированной Сефарозы 4В оставляли набухать в 1 мМ НСl в течение 20 минут и вносили в колонку с последующей промывкой 1 мМ НСl. Затем полученную смолу дополнительно промывали в 4 мл буферного раствора для связывания (0,1 М NаНСО3 и 0,5 М NaCl, pH 8,3), переносили в пробирку и сразу же смешивали с моноклональными антителами против ЕРО в буферном растворе для связывания (500 мкг на флакон) и инкубировали при комнатной температуре в течение 2 часов при энергичном перемешивании. Затем полученный продукт помещали в блокирующий буфер (0,2 М глицин, pH 8,0) и инкубировали при комнатной температуре в течение 2 часов при перемешивании. Полученную смолу промывали последовательно 6,5 мл буферного раствора для связывания, 6,5 мл ацетатного буферного раствора (0,1 М уксусная кислота, 0,5 М NaCl pH 4) и 6,5 мл буферного раствора для связывания. Приготовленную смолу помещали в колонку и затем очищали ЕРО следующим образом.
Клетки выращивали в бессывороточной среде в течение одного дня, а затем среду концентрировали приблизительно в 5 раз с помощью ультрафильтрации, например, Centriprep (имеющего номинальную предельную величину молекул (величину отсечения) 10000) (Millipore Corp.). Затем концентрированные растворы наносили на колонку, уравновешенную PBS, при скорости приблизительно 20 мл/час и промывали PBS. Белки-мишени элюировали в буферный раствор для элюции (0,1 М глицин, pH 2,8) и затем немедленно титровали 1 М Трис до pH 7,5. Чистота очищенного ЕАТР составляла 97% или больше, что подтверждали SDS-PAGG электрофорезом с последующим окрашиванием серебром (см. фиг.4).
(5) Измерение активности с помощью биоанализа и биохимического анализа
Биологические активности экспрессируемых и подходящим образом очищенных ЕРО и ЕАТР определяли с помощью биоанализа с использованием клеток селезенки мыши, обработанных фенилгидразином. Полученные результаты показали, что активность ЕАТР выше, чем активность ЕРО, что позволяет предположить, что присутствие аминокислот на С-конце в ЕАТР не ингибирует активность ЕРО.
(6) Фармакокинетический тест
Для определения, действительно ли полученные ЕАТР имеют более продолжительный период полужизни in vivo, проводили фармакокинетические тесты на мышах. При этом препараты ЕАТР вводили внутривенно четырем мышам в дозах 20 единиц для каждой мыши. Для оценки профиля концентраций ЕАТР в крови кровь забирали у каждой мыши и определяли концентрацию ЕАТР с использованием набора EIA (иммуноферментного анализа) (Boehringer Mannheim, Corp.). Результаты фармакокинетического теста, проведенного на мышах, показали, что ЕАТР имеет гораздо более продолжительный период полужизни in vivo по сравнению с ЕРО (см. фиг.5).
Далее данное изобретение иллюстрируется следующими примерами, которые не должны ошибочно рассматриваться как ограничивающие объем данного изобретения.
Сведения, подтверждающие возможность осуществления изобретения
Пример 1: Получение генов
кДНК ЕРО получали с помощью общепринятого способа полимеразной цепной реакции с обратной транскрипцией (OT-PCR) с использованием набора RT-PCR Premix Kit (Bioneer Corp., Korea), в котором использовали праймеры ЕР1 и ЕС2, комплементарные обоим концам кДНК ЕРО, полученной ранее из кДНК-библиотеки эмбриональной печени человека (Invitrogen Corp.). Выполняли 30 циклов ПЦР в условиях 35 секунд при 55°С (отжиг), 40 секунд при 72°С и 20 секунд при 94°С с получением кДНК ЕРО. Полученную кДНК ЕРО клонировали в клонирующий вектор pGEM-T ((Promega Corp.). Другими словами, продукт ПЦР элюировали из 1% агарозы, лигировали с pGEM-T с последующей трансформацией Е. coli NM522. После инкубации в течение ночи на твердой LB-ампициллин-среде с нанесенной мазком смесью X-gal/IPTG плазмидную ДНК выделяли из белых колоний и расщепляли рестриктазами SacI и SacII для отбора колоний, содержащих вставку кДНК ЕРО. Полученный вектор был назван pGEMT-EPO, а содержащийся в нем ген ЕРО был использован в качестве матрицы в последующих экспериментах.
Модифицированные СРТ-гены β-субъединицы HCG получали с помощью химического синтеза и самопраймирующейся ПЦР. Синтезированные фрагменты генов были названы EAl, A2, A3 и A4.
По 1 мкл каждого из четырех генов использовали (50 пмоль/мкл) для подвергания 15 циклам ПЦР с использованием Taq-системы высокой точности (Boehringer Mannheim Corp.) в условиях 40 секунд при 55°С (отжиг), 40 секунд при 72°С и 20 секунд при 94°С. Фрагменты генов размером приблизительно 100 п.н. определяли в 1% агарозном геле (модифицированные СТР-гены).
Эти гены кодируют пептид, полученный заменой 4 остатков Ser в положениях 121, 127, 132 и 138 среди 28 С-концевых аминокислот в положениях 118-145 β-субъединицы HCG остатками Аlа (см. фиг.1).
ПЦР проводили с использованием pGEMT-EPO в качестве матрицы и ЕР1 и ЕС2 в качестве праймеров с получением только генов ЕРО. Затем 30 циклов ПЦР дополнительно проводили с использованием как генов ЕРО, так и модифицированных СТР-генов в качестве матриц и с использованием праймеров ЕР11 и ЕР22 с применением Taq-системы высокой точности в условиях 42 секунды при 57°С (отжиг), 60 секунд при 72°С и 20 секунд при 94°С. Таким образом, получали фрагмент длиной приблизительно 630 п.н., представляющий собой слитые гены (также называемые ЕАТР-генами). Эти гены клонировали в pGEM-T при помощи вышеуказанного способа (с получением вектора pGEMT-EATP) и определяли его нуклеотидную последовательность.
Пример 2: Конструирование экспрессирующего вектора pcDNA3.1-EATP
В качестве экспрессирующего вектора использовали pcDNA3.1 (Invitrogen). Ген ЕАТР в pGEMT-EATP ограничен сайтами рестрикции HindIII и BamHI, происходящие из праймеров ЕР11 и ЕР22.
pcDNA3.1 и полученную плазмиду pGEMT-EATP расщепляли рестриктазами HindIII и BamHI. Линеаризованную pcDNA3.1 и ген EATP выделяли из агарозного геля с использованием набора для элюции Qiagen с последующим лигированием и трансформацией ими Е. coli NM522. Плазмиды выделяли из колоний, образующихся после инкубации в течение ночи в твердой LB-ампициллин-среде, и расщепляли рестриктазами HindIII и BamHI. Затем с помощью электрофореза в 1% агарозном геле отбирали колонии со встроенным EATP. Полученные плазмиды были названы pcDNA3.l-ЕАТР (см. фиг.3).
Пример 3: Трансфекция клеток СНО и экспрессия EATP
Клетки СНО (DG44) выращивали на чашке 60 мм до получения 40-80% конфлюентных клеток (1-4×105 клеток на чашку 60 мм). 3 мкл реагента Superfection (Boehringer Mannheim Corp.) и 97 мкл среды (α-МЕМ со средой, без сыворотки, без антибиотиков) тщательно перемешивали и к полученной смеси добавляли приблизительно 2 мкг ДНК плазмиды pcDNA3.1-EATP (более 0,1 мкг/мкл) и 0,2 мкг содержащего ген дигидрофолатредуктазы (dhfr) вектора pLTRdhfr26 (ATCC 37295) и проводили реакцию при комнатной температуре в течение 5-10 минут с последующим добавлением к этим клеткам. Спустя один день среду заменяли α-МЕМ без среды (содержащей 500 мкг/мл G418) с 10% ФТС. К клеткам добавляли среду, содержащую 500 мкг/мл G418, и культивировали в течение 7-10 дней. За это время все клетки, не содержащие ген устойчивости к G418, и клетки негативного контроля погибали. После достаточного культивирования клеток, отобранных из среды G418, в среде определяли экспрессируемый ЕАТР-белок с использованием набора ELISA для ЕРО (Boehringer Mannheim Corp.).
Пример 4: Очистка экспрессируемого ЕАТР
Смолы для аффинной очистки ЕРО с использованием моноклональных антител против ЕРО (R&D Inc.) получали следующим образом.
0,3 г CNBr-активированной Сефарозы 4В оставляли набухать в 1 мМ НСl в течение 20 минут и вносили в колонку с последующим промывкой 1 мМ НСl. Затем полученную смолу дополнительно промывали в 4 мл буферного раствора для связывания (0,1 М NaHCO3 и 0,5 М NaCl, pH 8,3), переносили в пробирку, сразу же смешивали с моноклональными антителами против ЕРО в буферном растворе для связывания (500 мкг на флакон) и затем инкубировали при комнатной температуре в течение 2 часов. В это время пробирку встряхивали. Затем полученный продукт помещали в блокирующий буфер (0,2 М глицин, pH 8,0) и инкубировали при комнатной температуре в течение 2 часов. Смолу промывали последовательно 6,5 мл буферного раствора для связывания, 6,5 мл ацетатного буферного раствора (0,1 М уксусная кислота, 0,5 М NaCl pH 4) и 6,5 мл буферного раствора для связывания. Приготовленную смолу помещали в колонку и затем проводили очистку следующим образом.
Клетки выращивали в бессывороточной среде в течение одного дня, а затем среду концентрировали приблизительно в 5 раз с помощью ультрафильтрации, например, Centriprep (имеющего номинальную предельную величину молекул (величину отсечения) 10000) (Millipore Corp.). Затем концентрированные растворы наносили на колонку, уравновешенную PBS, при скорости приблизительно 20 мл/час и промывали PBS. Белки-мишени элюировали раствором для элюции (0,1 М глицин, рН 2,8) и затем немедленно титровали 1 М Трис до рН 7,5. Чистота очищенного ЕАТР составляла 97% или больше, как было подтверждено SDS-PAGE электрофорезом с последующим окрашиванием серебром (см. фиг.4).
Пример 5: Определение активности с помощью биоанализа
Фенилгидразин вводили мыши один раз в день в течение 2 дней при дозе 60 мг/кг. Спустя 3 дня увеличенную селезенку извлекали и измельчали гомогенизатором с получением клеток селезенки. Клетки селезенки разбавляли до концентрации 6×106 клеток/мл и каждые 100 мкл разбавленной пробы переносили на 96-луночный планшет. Стандартные ЕРО (0-500 мЕ/мл) и экспрессируемые ЕРО и ЕАТР (в каждом случае 100 мЕ/мл) добавляли в соответствующие лунки. Затем планшет инкубировали в СO2-инкубаторе при 37°С в течение 22 часов. 50 мкл диметил-3Н-тимидина (20 мкКи/мл) добавляли в каждую лунку. Планшет инкубировали в течение 2 часов и затем растворы пробы каждой лунки абсорбировали на стеклянный фильтр (Nunc 1-73164). Этот фильтр промывали три раза солевым раствором и радиоактивность фильтра измеряли при помощи бета (β)-счетчика. Оказалось, что активность ЕАТР была по существу равной или несколько выше, чем активность ЕРО, что позволяет предположить, что присутствие дополненных аминокислот на С-конце в ЕАТР не ингибирует активность ЕРО.
Пример 6: Фармакокинетический тест
Для проверки, действительно ли полученные материалы-кандидаты имеют более продолжительный период полужизни in vivo, проводили фармакокинетические тесты на мышах. При этом слитый белок, очищенный по способу, описанному в примере 5, вводили внутривенно четырем мышам в дозах 20 единиц для каждой мыши. Для оценки профиля концентраций в крови первые 2 часа кровь собирали каждые 30 минут, а затем каждые 2 часа концентрацию в крови определяли с использованием набора EIA (иммуноферментного анализа) (Boehringer Mannheim, Corp.). Результаты фармакокинетического теста показаны на фиг.5. Как следует из фиг.5, ЕАТР имеет гораздо более продолжительный (более чем в 2,5 раза) период полужизни in vivo, чем контрольный материал ЕРО.
В соответствии с данным изобретением, активность ЕРО in vivo может быть увеличена посредством увеличения периода полужизни in vivo при сохранении присущей ЕРО активности с его собстенными аминокислотами, без увеличения содержания сахарных цепей ЕРО.






Изобретение относится к области биотехнологии и может быть использовано для получения слитого белка с увеличенной эритропоэтиновой активностью. Слитый белок представляет собой эритропоэтин человека, к С-концу которого присоединен фрагмент мутантного С-концевого пептида β-субъединицы хорионического гонадотропина человека с аминокислотной последовательностью, соответствующей положениям 7-34 последовательности SEQ ID NO.1, в которой один-четыре серина в положении 10, 16, 21 и 27 заменены аланином или глицином. Слитый белок получают путем культивирования клеточной линии, трансформированной рекомбинантным вектором, содержащим нуклеиновую кислоту, которая кодирует слитый белок. Изобретение позволяет получить слитый белок с активностью эритропоэтина, обладающий увеличенным периодом полужизни in vivo. 3 н. п. ф-лы, 5 ил.
| WO 0136489, 25.05.2001 | |||
| US 6040136, 21.03.2000 | |||
| WO 9009800, 07.09.1990 | |||
| WO 8501959, 09.05.1985 | |||
| US 5712122, 27.01.1998 | |||
| US 5959171, 28.09.1999 | |||
| СПОСОБЫ И ПРЕПАРАТЫ ДЛЯ СТИМУЛИРОВАНИЯ РОСТА И ДИФФЕРЕНЦИРОВКИ МЕГАКАРИОЦИТОВ | 1995 |
|
RU2158603C2 |
| СПОСОБ СЕРНОЙ ВУЛКАНИЗАЦИИ РЕЗИНОВЫХ СМЕСЕЙ | 0 |
|
SU248194A1 |

