Данная заявка претендует на приоритет и эффект изобретения в соответствии с предварительной заявкой на патент США с регистрационным номером 60/081,863, поданной 15 апреля 1998 г., содержание которой включено сюда посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится, в целом, к иммуноконъюгатам, в частности к белкам, слитым из антитела и цитокина, пригодным для целенаправленной иммунотерапии и общей стимуляции иммунитета. Более конкретно, настоящее изобретение относится к использованию ингибиторов ангиогенеза для усиления иммунного ответа, опосредованного белками, слитыми из антитела и цитокина, направленного против предварительно выбранного типа клеток, например клеток плотной опухоли.
УРОВЕНЬ ТЕХНИКИ
Антитела использовали для лечения заболеваний людей в течение многих лет, главным образом для обеспечения пассивного иммунитета к вирусной или бактериальной инфекции. Однако недавно антитела и конъюгаты антител были использованы в качестве противоопухолевых агентов. Противоопухолевую активность трудно было продемонстрировать в отношении большинства типов опухолей, кроме тех случаев, когда клиническая картина представляет собой минимальное остаточное проявление заболевания (Reithmuller et al., LANCET 94:1177-1183) или когда опухоль доступна для циркулирующих антител, например в случае В-лимфомы (Maloney et al. (1994) BLOOD 84: 2457-2466). Плотные опухоли значительно более устойчивы к терапевтическому воздействию, опосредованному антителами, чем микрометастатические очаги, обнаруживаемые при картине минимальных остаточных проявлений заболевания.
Предшествующие исследования показали, что эффективность лечения опухолей антителами in vivo может быть значительно повышена за счет присоединения к молекуле антитела иммуностимулирующих цитокинов. Тем не менее белки, слитые из антитела и цитокина, были значительно менее эффективными в отношении деструкции крупных плотных опухолей, чем в отношении диссеминированных метастатических очагов (Xiang et al. (1997) CANCER RESEARCH 57: 4948-4955, и Lode et al. (1998) BLOOD 91: 1706-1715).
Поэтому в данной области техники все еще сохраняется потребность в композициях и способах, использующих такие композиции, для усиления иммунных ответов, опосредованных белками, слитыми из антитела и цитокина, и направленных против предварительно выбранных типов клеток, например типов клеток, присутствующих в плотных опухолях.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение частично основано на обнаружении того факта, что при введении млекопитающему иммуноконъюгата можно вызвать более сильный иммунный ответ против предварительно выбранного типа клеток, если иммуноконъюгат вводят вместе с ингибитором ангиогенеза. В частности, было обнаружено, что такие комбинации особенно эффективно опосредуют иммунную деструкцию предварительно выбранного типа клеток, например типов клеток, обнаруживаемых в плотных опухолях, и клеток, инфицированных вирусами.
В одном из аспектов настоящее изобретение предусматривает способ индуцирования у млекопитающего цитоцидного иммунного ответа против предварительно выбранного типа клеток. Способ включает введение млекопитающему: (1) иммуноконъюгата, содержащего сайт связывания антигенов антитела, способный соединяться с предварительно выбранным типом клеток, и цитокин, способный вызывать иммунный ответ против предварительно выбранного типа клеток, и (2) ингибитора ангиогенеза в количестве, достаточном для усиления иммунного ответа по сравнению с иммунным ответом, стимулируемым одним иммуноконъюгатом.
В предпочтительном примере осуществления изобретения предварительно выбранным типом клеток могут быть раковые клетки, присутствующие, например, в плотной опухоли, более предпочтительно в крупной плотной опухоли (а именно, больше 100 мм3). Альтернативно, предварительно выбранным типом клеток может быть инфицированная вирусом клетка, например клетка, инфицированная вирусом иммунодефицита человека (ВИЧ).
В другом предпочтительном примере осуществления изобретения ингибитор ангиогенеза может быть введен одновременно с иммуноконъюгатом. Альтернативно, ингибитор ангиогенеза может быть введен до введения иммуноконъюгата. Кроме того, предполагается, что иммуноконъюгат может быть введен совместно с несколькими различными ингибиторами ангиогенеза. Альтернативно, предполагается, что ингибитор ангиогенеза может быть введен совместно с несколькими различными иммуноконъюгатами.
В другом аспекте изобретение предусматривает композицию для индуцирования у млекопитающего цитоцидного иммунного ответа против предварительно выбранного типа клеток. Композиция включает в комбинации: (1) иммуноконъюгат, включающий сайт связывания антигенов антитела, способный присоединять предварительно выбранный тип клеток, и цитокин, способный вызывать этот иммунный ответ против предварительно выбранного типа клеток млекопитающего, и (2) ингибитор ангиогенеза в количестве, достаточном для усиления иммунного ответа, вызываемого иммуноконъюгатом комбинации, по сравнению с иммунным ответом, стимулируемым одним иммуноконъюгатом.
В предпочтительном примере осуществления изобретения сайт связывания антигенов антитела имммуноконъюгата предпочтительно включает тяжелую цепь иммуноглобулина или ее антигенсвязывающий фрагмент. Тяжелая цепь иммуноглобулина предпочтительно включает, в направлении от амино-конца к карбокси-концу, домен 1 константной области тяжелой цепи иммуноглобулина (СН1), домен 2 константной области тяжелой цепи иммуноглобулина (СН2) и факультативно может также включать домен 3 константной области тяжелой цепи иммуноглобулина (СНЗ). В более предпочтительном примере осуществления изобретения иммуноконъюгат является слитым белком, включающим тяжелую цепь иммуноглобулина или ее антигенсвязывающий фрагмент, соединенные посредством пептидной связи с цитокином. Соответственно, предпочтительный белок, слитый из антитела и цитокина, включает, в направлении от амино-конца к карбокси-концу, (1) сайт связывания антигена антитела, включающий вариабельную область иммуноглобулина и способный присоединять антиген клеточной поверхности предварительно выбранного типа клеток, домен иммуноглобулина СН1, домен иммуноглобулина СН2 (факультативно – СН3 домен), и (2) цитокин. Способы получения и использования таких слитых белков подробно описаны в работе Gillies et al. (1992) PROC. NATL. ACAD. SCI. USA 89:1428-1432; Gillies et al. (1998) J. IMMUNOL. 160:6195-6203 и Патенте США №5,650,150.
Домены константной области иммуноглобулина (а именно, СН1, СН2 и/или СН3 домены) могут быть доменами константной области, в норме связанными с тем же доменом вариабельной области в антителе, существующем в природе. Альтернативно, один или несколько доменов константной области могут быть получены из антител, отличающихся от антитела, использованного в качестве источника домена вариабельной области. Другими словами, домены вариабельной и константной областей иммуноглобулина могут быть получены из различных антител, например антител, полученных от различных видов животных. См., например, Патент США №4,816,567. Кроме того, вариабельные области иммуноглобулина могут включать последовательности каркасной области (КО), полученные от одного вида, например от человека, и последовательности области, определяющей комплементарность (ООК), вставленные между КО, полученные от второго, другого, вида, например от мыши. Способы получения и использования таких химерных вариабельных областей иммуноглобулинов описаны, например, в Патентах США №5,225,539 и №5,585,089.
Иммуноконъюгаты на основе антител, кроме того, предпочтительно включают легкую цепь иммуноглобулина, которая предпочтительно ковалентно связана с тяжелой цепью иммуноглобулина посредством, например, дисульфидной связи. Вариабельные области соединенных тяжелой и легкой цепей иммуноглобулина совместно образуют один законченный сайт связывания для присоединения предварительно выбранного антигена. В других примерах осуществления изобретения иммуноконъюгаты включают две химерные цепи, каждая из которых включает, как минимум, один участок тяжелой цепи иммуноглобулина, присоединенный к цитокину. Две химерных цепи предпочтительно ковалентно соединены друг с другом, например, при помощи одной или нескольких межцепочечных дисульфидных связей.
Таким образом, изобретение относится к слитым белкам, у которых специфичность связывания антигена и активность антитела сочетаются с высокой биологической активностью цитокина. Слитый белок согласно настоящему изобретению можно использовать для избирательной доставки цитокина к клетке-мишени in vivo, так что цитокин может оказывать локальный биологический эффект в непосредственной близости от клетки-мишени. В предпочтительном примере осуществления изобретения компонент слитого белка, представляющий собой антитело, избирательно присоединяет антиген на поверхности раковой клетки и в результате этого слитый белок проявляет локальную противораковую активность. В альтернативном предпочтительном примере осуществления изобретения компонент слитого белка, являющийся антителом, специфически присоединяет клетку, инфицированную вирусом, такую как ВИЧ-инфицированная клетка, и в результате слитый белок проявляет локальную противовирусную активность.
Предпочтительные цитокины, которые могут быть включены в иммуноконъюгаты согласно настоящему изобретению, включают, например, факторы некроза опухолей, интерлейкины, колониестимулирующие факторы и лимфокины. Предпочтительные факторы некроза опухолей включают, например, фактор некроза опухолей α (ФНОα ). Предпочтительные интерлейкины включают, например, интерлейкин-2 (ИЛ-2, IL-2), интерлейкин-4 (ИЛ-4, IL-4), интерлейкин-5 (ИЛ-5, IL-5), интерлейкин-7 (ИЛ-7, IL-7), интерлейкин-12 (ИЛ-12, IL-12), интерлейкин-15 (ИЛ-15, IL-15) и интерлейкин-18 (ИЛ-18, IL-18). Предпочтительные колониестимулирующие факторы включают, например, колониестимулирующий фактор гранулоцитов-макрофагов (ГМ-КСФ, GM-CSF) и колониестимулирующий фактор макрофагов (М-КСФ, M-CSF). Предпочтительные лимфокины включают, например, лимфотоксин (ЛТ). Другие подходящие цитокины включают интерфероны (ИФН, IFN), в том числе ИФН-α , ИФН-β и ИФН-γ , которые обладают иммунологическими эффектами, а также антиангиогенными эффектами, которые не зависят от их противовирусной активности.
Предпочтительные ингибиторы ангиогенеза, которые можно использовать при осуществлении настоящего изобретения, включают, например, эндостатин, ангиостатин, пептиды, обладающие аффинностью к αvβ3-интегрину, антитела или их фрагменты, обладающие аффиностью к αvβ3-интегрину, пептиды с аффинностью к рецептору эпидермального фактора роста (ЭФР, EGF), антитела или их фрагменты, обладающие аффинностью к рецептору ЭФР, ингибиторы циклооксигеназы (ЦОГ) СОХ-2, фумагиллин, талидомид, антиангиогенные цитокины, например ИФН-α , ИФН-β и ИФН-γ , и слитые белки, включающие цитокин, с таким антиангиогенным цитокином.
Также предусмотрены предпочтительные дозировки и схемы введения для введения иммуноконъюгатов в комбинации с ингибиторами ангиогенеза.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Вышеизложенные и прочие цели, отличительные признаки и преимущества настоящего изобретения, а также собственно изобретение можно лучше понять из нижеследующего описания предпочтительных примеров осуществления изобретения, сопровождаемого прилагаемыми графическими материалами, в которых:
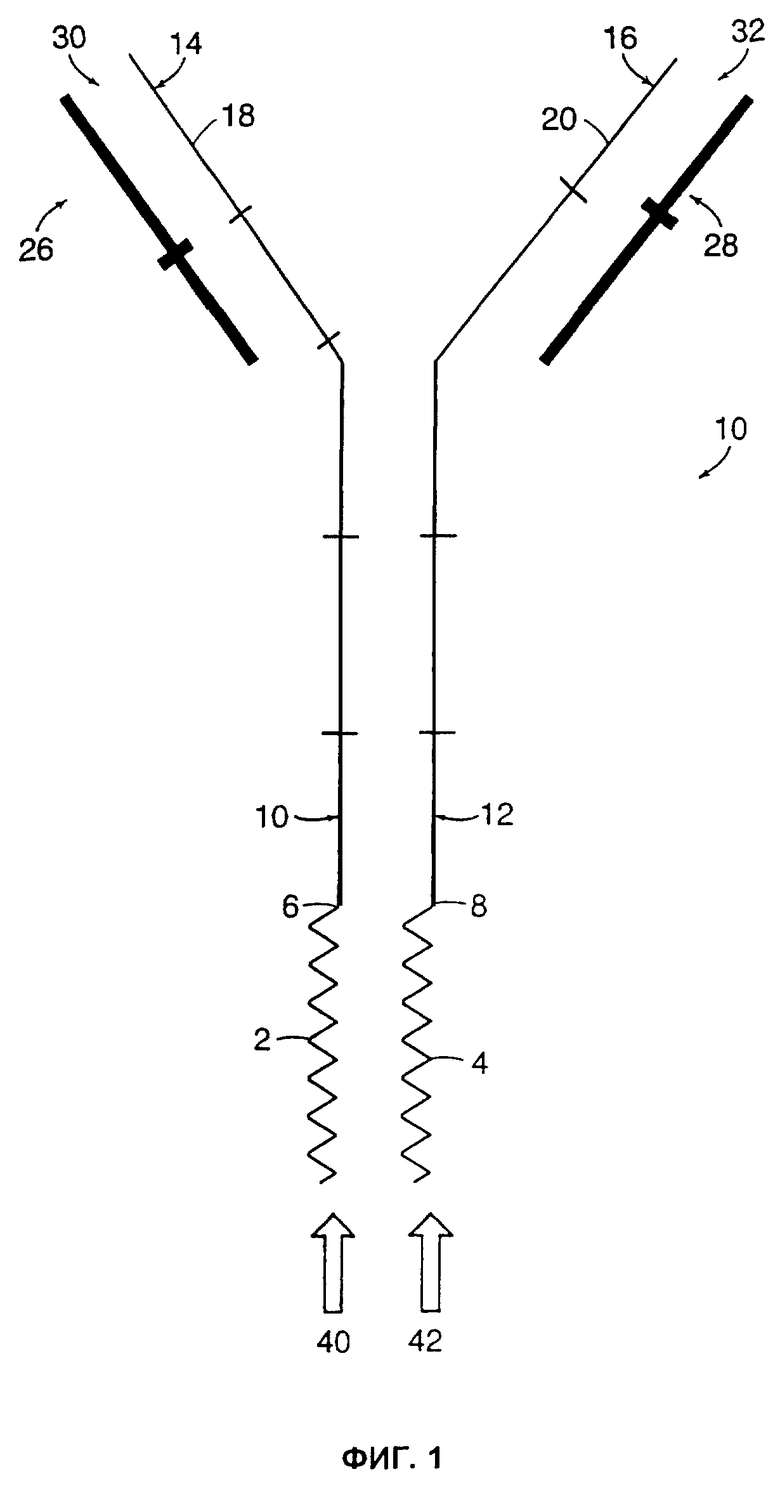
ФИГ.1 представляет собой схематическое изображение типичного иммуноконъюгата, который пригоден для использования при осуществлении изобретения.
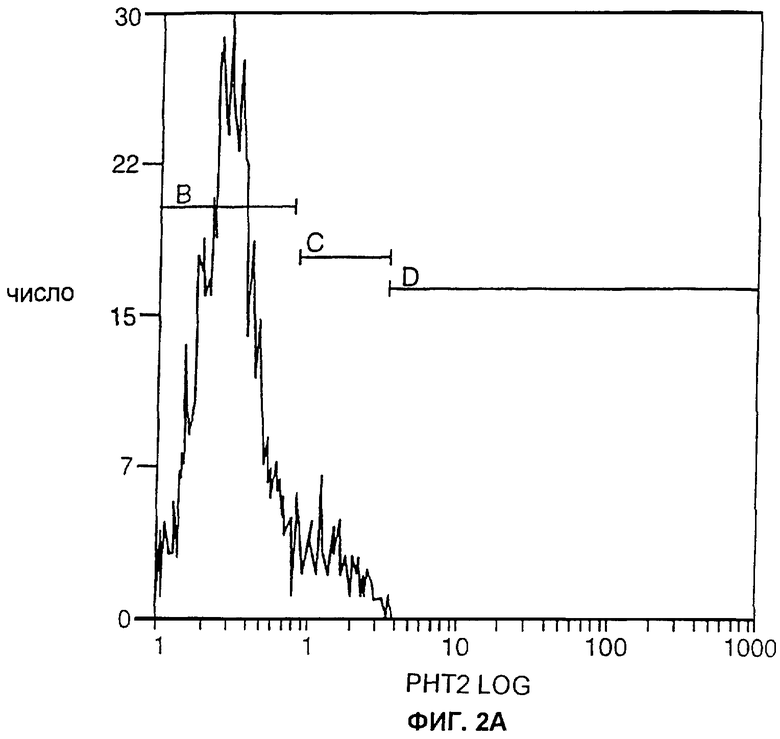
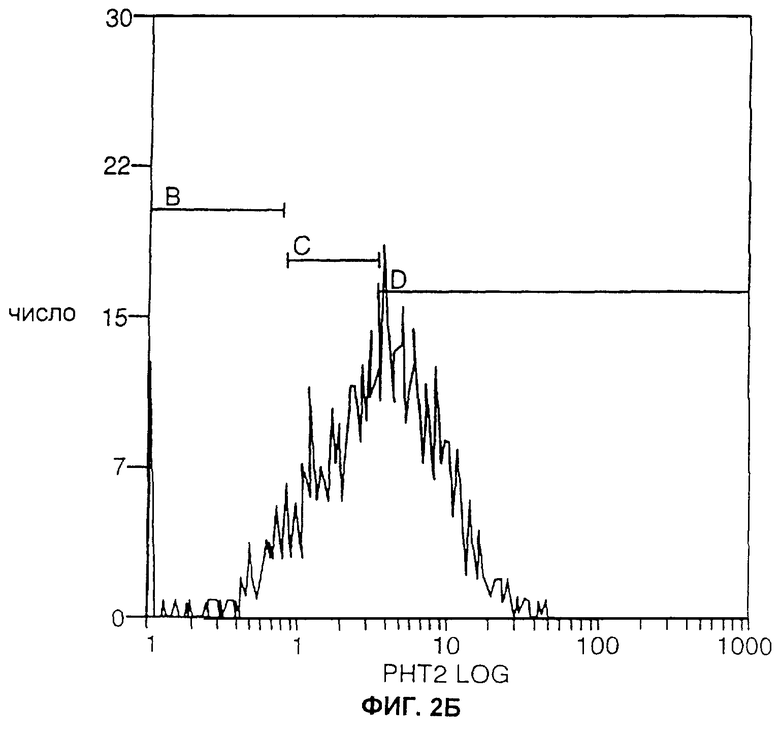
ФИГ.2А и 2Б являются графиками, отображающими экспрессию ЕрСАМ человека в трансфицированных клетках карциномы легких Левиса (КЛЛ, LLC) мышей, проанализированную с помощью сортировки флуоресцентно-активированных клеток (ФАКС, FACS). Одинаковое количество трансфицированных клеток окрашивали либо только вторичным специфическим антителом к Fc человека, меченным флуоресцеина изотиоцианатом (ФИТЦ, FITC) (ФИГ.2А), либо вначале их окрашивали белком, слитым из антитела и huKS-huIL2, а затем специфичным к Fc человека антителом, меченным ФИТЦ (ФИГ.2Б).
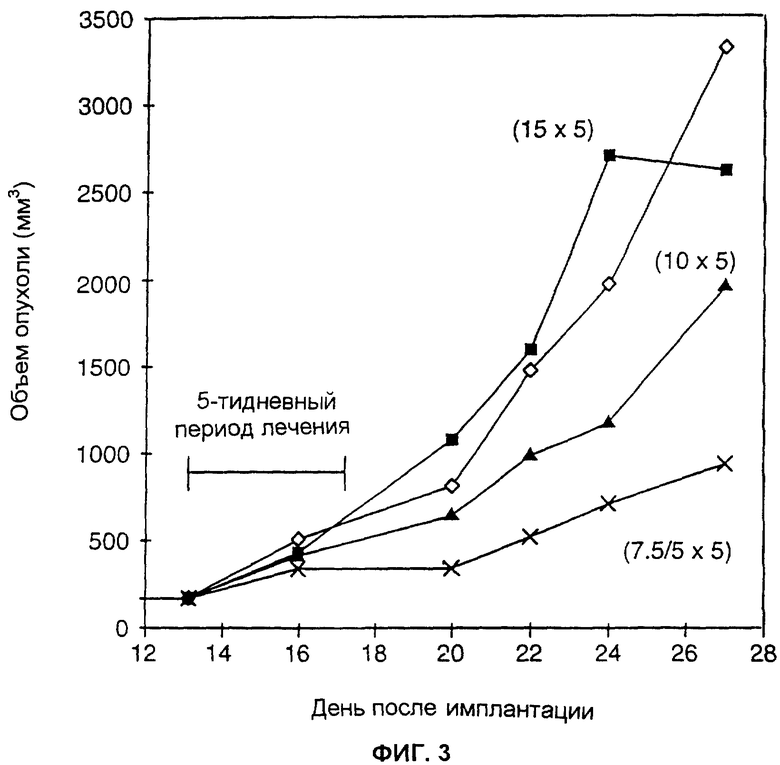
ФИГ.3 представляет собой линейный график, отображающий воздействия на подкожные опухоли белка, слитого из антитела и цитокина, вводимого или отдельно, или в комбинации со вторым белком, слитым из антитела и цитокина, в котором цитокин обладает антиангиогенной активностью. Лечение, продолжавшееся в течение 5 дней, было начато через 13 дней после имплантации КПЛ-клеток. Мышам вводили раствор фосфатного буфера (незаполненные ромбики); 15 мкг/день только слитого белка huKS-muγ 2a-muIL2 (сплошные квадратики); 10 мкг/день слитого белка huKS-muγ 2a-muIL2 (сплошные треугольники); и комбинацию 7,5 мкг/день слитого белка huKS-muγ 2a-mulL2 и 5 мкг/день huKS-muγ 2a-muIL12 (крестики).
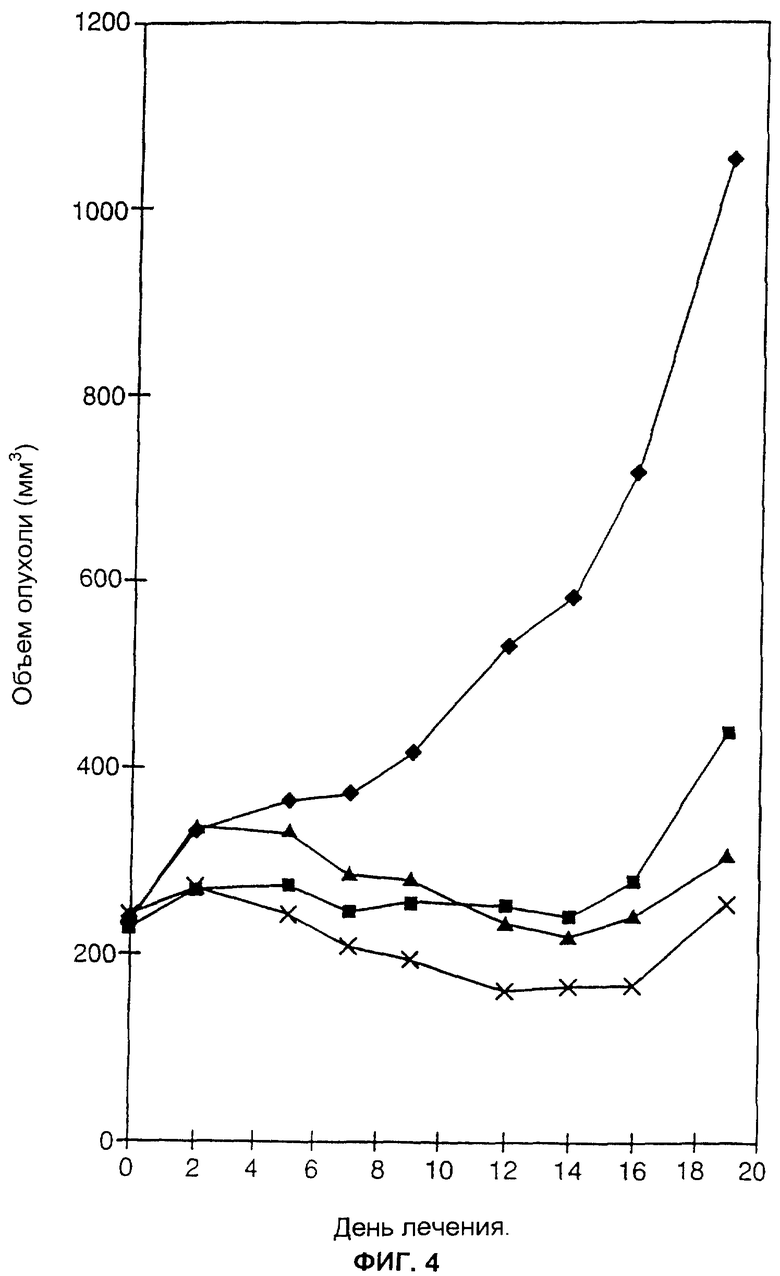
ФИГ.4 представляет собой линейный график, отображающий воздействия на подкожные опухоли белка, слитого из антитела и цитокина, вводимого либо отдельно, либо в комбинации со слитым белком, включающим эндостатин. Размер подкожных опухолей СТ26/ЕрСАМ регистрировали у мышей, которым вводили раствор фосфатного буфера (сплошные ромбики), слитый белок muFc-muEndo (сплошные квадратики), слитый белок huKS-huγ 4-huIL2 (сплошные треугольники) и комбинацию слитого белка muFc-muEndo и слитого белка huKS-huγ 4-huIL2 (крестики).
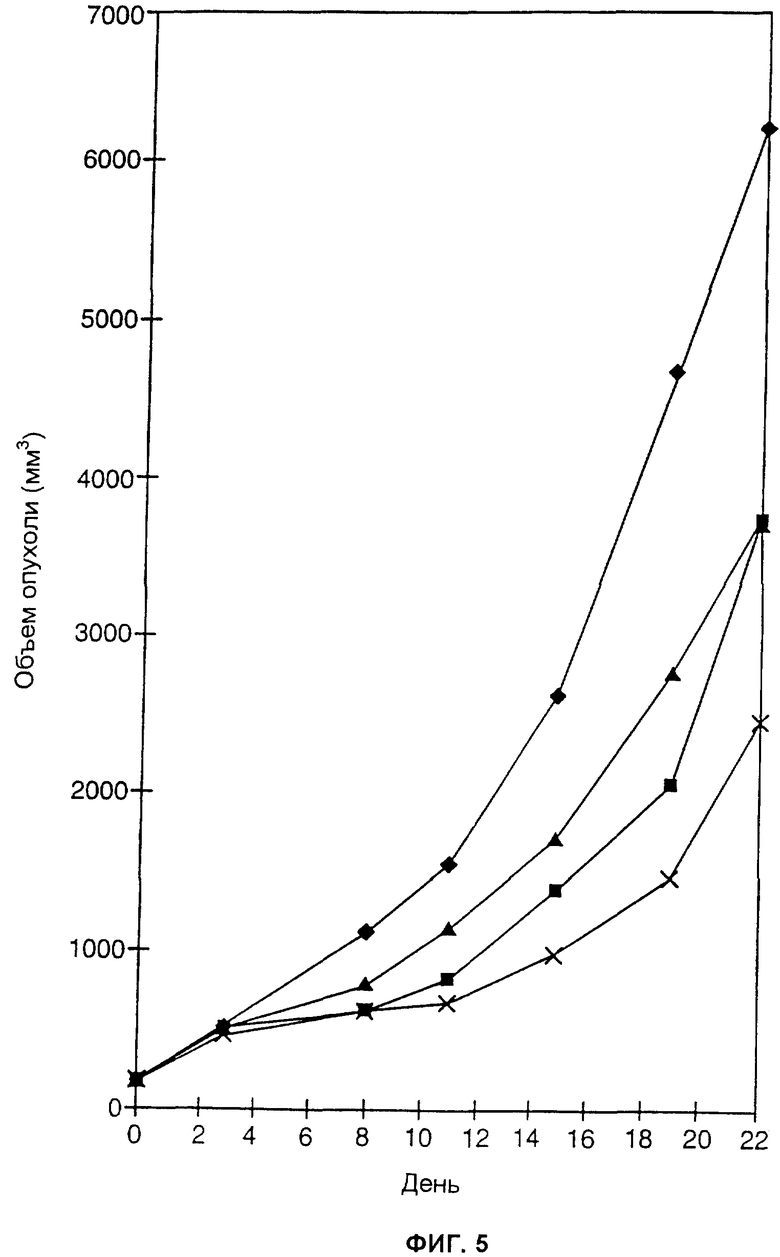
ФИГ.5 представляет собой линейный график, отображающий воздействие на подкожные опухоли белка, слитого из антитела и цитокина, вводимого либо отдельно, либо в комбинации с индометацином. Размер подкожных опухолей LLC-ЕрСАМ регистрировали у мышей, которым вводили раствор фосфатного буфера (сплошные ромбики), слитый белок huKS-huγ 1-huIL2 (сплошные квадратики), индометацин (сплошные треугольники) и комбинацию слитого белка huKS-huγ 1-huIL2 и индометацина (крестики).
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
К настоящему времени было обнаружено, что цитоцидные иммунные ответы против предварительно выбранного типа клеток, инициируемые иммуноконъюгатом, можно значительно усилить посредством введения иммуноконъюгата совместно с ингибитором ангиогенеза. Комбинированная терапия особенно эффективна при опосредовании иммунной деструкции пораженной ткани, например, определенных опухолевых или инфицированных вирусом клеток. Настоящее изобретение описывает способы получения и использования пригодных для этой цели иммуноконъюгатов, а также анализы, которые можно использовать для тестирования их фармакокинетической активности в доклинических модельных опытах на животных in vivo, если иммуноконъюгаты сочетают с подходящими ингибиторами ангиогенеза.
При использовании в настоящей работе термин “цитоцидный иммунный ответ” понимают как обозначающий любой иммунный ответ у млекопитающего, как гуморальный, так и клеточный по природе, который усиливается иммуноконъюгатом согласно настоящему изобретению и который либо убивает, либо другим способом снижает жизнеспособность предварительно выбранного типа клеток млекопитающего. В иммунный ответ может быть включен один или несколько типов клеток, в том числе Т-клетки, клетки - натуральные киллеры (НК, NK) и макрофаги.
При использовании в настоящей работе, термин “иммуноконъюгат” понимают как обозначающий конъюгат (1) сайта связывания антигенов антитела, обладающего специфичностью связывания и способного присоединять поверхностный антиген раковой клетки или инфицированной вирусом клетки, и (2) цитокина, который способен индуцировать или стимулировать цитоцидный иммунный ответ, направленный против раковых или инфицированных вирусом клеток. Соответственно, иммуноконъюгат способен избирательно доставлять цитокин к клетке-мишени in vivo, так что цитокин может опосредовать локальный иммунный ответ, направленный против клетки-мишени. Например, если компонент иммуноконъюгата, являющийся антителом, избирательно присоединяет антиген, находящийся на поверхности раковой клетки, например раковой клетки плотной опухоли, в частности крупной плотной опухоли размером более 100 мм3, то иммуноконъюгат проявляет локальную противораковую активность. Альтернативно, если компонент иммуноконъюгата, являющийся антителом, избирательно присоединяет антиген инфицированной вирусом клетки, такой как клетка, инфицированная вирусом иммунодефицита человека (ВИЧ), иммуноконъюгат проявляет локальную противовирусную активность.
При использовании в настоящей работе термин “сайт связывания антигенов антитела” понимают как обозначающий, как минимум, один участок тяжелой цепи иммуноглобулина, например вариабельную область иммуноглобулина, способный присоединять предварительно выбранный тип клеток. Сайт связывания антигенов антитела также предпочтительно включает, как минимум, один участок константной области иммуноглобулина, включающий, например, СН1 домен, СН2 домен и, факультативно, СН3 домен. Кроме того, тяжелая цепь иммуноглобулина может быть соединена либо ковалентно, либо не ковалентно с легкой цепью иммуноглобулина, включающей, например, вариабельную область легкой цепи иммуноглобулина и, факультативно, константную область легкой цепи. Соответственно, предполагается, что сайт связывания антигена антитела может включать интактное антитело или его фрагмент, способные присоединять предварительно выбранный тип клеток.
Что касается иммуноконъюгата, то предполагается, что фрагмент антитела может быть присоединен к цитокину различными способами, хорошо известными специалистам в данной области техники. Например, в искусственно полученном слитом белке сайт связывания антигена антитела предпочтительно присоединен к цитокину посредством пептидной связи. Альтернативно, сайт связывания антигена антитела можно химически присоединить к цитокину с помощью реакционноспособных групп, например, сульфгидрильных групп, в пределах боковых цепей аминокислот, присутствующих в сайте связывания антигенов антитела и цитокине.
При использовании в настоящей работе термин “цитокин” понимают как обозначающий любой белок или пептид, их аналог или функциональный фрагмент, который способен стимулировать или вызывать у млекопитающих цитоцидный иммунный ответ, направленный против предварительно выбранного типа клеток, например раковых клеток или инфицированных вирусом клеток. Соответственно, предполагается, что в иммуноконъюгаты согласно настоящему изобретению можно включать различные цитокины. Пригодные для этой цели цитокины включают, например, факторы некроза опухолей, интерлейкины, лимфокины, колониестимулирующие факторы, интерфероны, включая видовые варианты и усеченные аналоги, которые способны стимулировать или вызывать такие цитоцидные иммунные ответы. Подходящие факторы некроза опухолей включают, например, ФНОα . Подходящие лимфокины включают, например, ЛТ. Подходящие колониестимулирующие факторы включают, например, ГМ-КСФ и М-КСФ. Подходящие интерлейкины включают, например, ИЛ-2, ИЛ-4, ИЛ-5, ИЛ-7, ИЛ-12, ИЛ-15 и ИЛ-18. Подходящие интерфероны включают, например, ИФН-α , ИФН-β и ИФН-γ .
Гены, кодирующие отдельные цитокины, представляющие интерес, можно клонировать de novo, получить из стандартных источников или синтезировать посредством стандартного синтеза ДНК из известных последовательностей нуклеотидов. Например, известна ДНК-последовательность ЛТ (см., например, Nedwin et al. (1985) NUCLEIC ACID RES. 13:6361), а также последовательности ИЛ-2 (см., например, Taniguchi et al. (1983) NATURE 302:305-318), ГМ-КСФ (см., например, Gasson et al. (1984) SCIENCE 266:1339-1342) и ФНОα (см., например, Nedwin et al. (1985) NUCLEIC ACID RES. 13:6361).
В предпочтительном примере осуществления изобретения иммуноконъюгаты представляют собой рекомбинантные слитые белки, полученные с помощью обычных методик получения рекомбинантных ДНК, то есть путем формирования искусственной нуклеиновой кислоты, кодирующей химерный иммуноконъюгат. Конструирование рекомбинантных белков, слитых из антитела и цитокина, было описано в прототипах. См., например, Gillies et al. (1992) PROC. NATL. ACAD. SCI. USA 89:1428-1432; Gillies et al. (1998) J. IMMUNOL. 160:6195-6203; и Патент США №5,650,150. Предпочтительно искусственный ген, кодирующий иммуноконъюгат согласно настоящему изобретению, включает, в направлении от 5’ к 3’-концу, сегмент ДНК, кодирующий домен вариабельной области тяжелой цепи иммуноглобулина, сегмент ДНК, кодирующий константную область тяжелой цепи иммуноглобулина, и ДНК, кодирующую цитокин. Слитый ген соединяют с вектором экспрессии или внедряют в него для трансфекции соответствующей клетки-реципиента, где слитый ген экспрессируется. Гибридную полипептидную цепь предпочтительно соединяют с легкой цепью иммуноглобулина, так что вариабельная область тяжелой цепи иммуноглобулина (Vн) и вариабельная область легкой цепи иммуноглобулина (VL) объединяются с образованием одного законченного сайта связывания предварительно выбранного антигена. В предпочтительном примере осуществления изобретения тяжелая и легкая цепи иммуноглобулина соединены ковалентно, например, посредством межцепочечной дисульфидной связи. Кроме того, две тяжелых цепи иммуноглобулина, одна или обе из которых слиты с цитокином, могут быть ковалентно связаны, например, посредством одной или нескольких межцепочечных дисульфидных связей.
Фиг.1 представляет собой схематическое изображение типичного иммуноконъюгата (10). В этом примере осуществления изобретения молекулы цитокинов (2 и 4) присоединены пептидной связью к карбоксильным концам (6 и 8) СН3-областей (10 и 12) тяжелых цепей антител (14 и 16). VL-области (26 и 28) изображены спаренными с Vн-областями (18 и 20) в типичной конфигурации IgG, образуя таким образом два сайта связывания антигена (30 и 32) на аминотерминальных концах иммуноконъюгата (10) и два сайта связывания с рецепторами цитокина (40 и 42) на карбокси-концах иммуноконъюгата (10). Конечно, в широком смысле, нет необходимости спаривать иммуноконъюгаты, как это показано на чертеже, или к молекуле цитокина может потребоваться присоединить только одну из двух тяжелых цепей иммуноглобулина.
Иммуноконъюгаты согласно настоящему изобретению могут считаться химерными на основании двух особенностей их структуры. Во-первых, иммуноконъюгат является химерным, так как он включает тяжелую цепь иммуноглобулина, обладающую специфичностью связывания антигена, соединенную с определенным цитокином. Во-вторых, иммуноконъюгат согласно настоящему изобретению может быть химерным в том смысле, что он включает вариабельную область иммуноглобулина (V) и константную область иммуноглобулина (С), полученные из различных антител, так что образующийся в результате белок является химерой V/C. Например, вариабельная и константная области могут быть получены из существующих в природе молекул антител, которые можно выделить из различных видов животных. См., например, Патент США №4,816,567. Также сюда относятся конструкции, в которых либо одна, либо обе вариабельные области иммуноглобулина содержат последовательности каркасной области (КО) и последовательности области, определяющей комплементарность (ООК), полученные от различных видов животных. Такие конструкции описаны, например, в работах Jones et al. (1986) NATURE 321:522-525, Verhoyen et al. (1988) SCIENCE 239:1534-1535 и Патентах США №№5,225,539 и 5,585,089. Кроме того, предполагается, что последовательности вариабельной области могут быть получены при скрининге библиотек, например библиотек фагов, с целью поиска последовательностей вариабельной области, которые присоединяют предварительно выбранный антиген с желаемой аффинностью. Способы получения и скрининга библиотек фагов описаны, например, в работах Huse et al. (1989) SCIENCE 246:1275-1281 и Kang et al. (1991) PROC. NATL. ACAD. SCI. USA 88:11120-11123.
Домены константной области тяжелой цепи иммуноглобулина для иммуноконъюгатов могут быть выбраны из любого из пяти классов иммуноглобулинов, обозначаемых как IgA (Igα ), IgD (Igδ ), IgE (Igε ), IgG (Igγ ) и IgM (Igμ ). Однако предпочтительны константные области тяжелой цепи иммуноглобулина из класса IgG. Кроме того, предполагается, что тяжелые цепи иммуноглобулина могут быть получены из любого подкласса антител IgG, обозначаемых в данной области техники как IgG1, IgG2, IgG3 и IgG4. Как известно, каждая константная область тяжелой цепи иммуноглобулина содержит четыре или пять доменов. Домены называются последовательно как: СН1-шарнир-СН2-СН3-(-СН4). СН4 присутствует в IgM, который не имеет шарнирной области. ДНК-последовательности доменов тяжелой цепи обладают перекрестной гомологией для разных классов иммуноглобулинов, например СН2 домен IgG гомологичен СН2 домену IgA или IgD и СН3 домену IgM и IgE. Легкие цепи иммуноглобулинов могут включать либо каппа (κ ), либо лямбда (λ ) константные области. Последовательности и расположение последовательностей этих областей иммуноглобулинов хорошо известны в данной области техники (см., например, Kabat et al., "Sequences of Proteins of Immunological Interest" (“Последовательности белков, представляющих иммунологический интерес”), U.S.Department of Health and Human Services, third edition 1983, fourth edition 1987, Huck et al. (1986) NUC. ACIDS RES. 14:1779-1789).
В предпочтительных примерах осуществления изобретения вариабельную область получают из антитела, специфического для предварительно выбранного антигена клеточной поверхности (антиген, ассоциированный с больной клеткой, такой как раковая клетка или клетка, инфицированная вирусом), а константная область включает СН1 и СН2 (и, факультативно, СН3) домены из антитела, которое представляет собой то же антитело, которое является источником вариабельной области, или отличается от него. В практике осуществления настоящего изобретения участок иммуноконъюгата, представляющий собой антитело, предпочтительно является неиммуногенным или является слабо иммуногенным для намеченного реципиента. Соответственно, участок, представляющий собой антитело, предпочтительно, насколько это возможно, получают от того же вида, что и намеченный реципиент. Например, если иммуноконъюгат должен вводиться человеку, домены константной области предпочтительно должны иметь происхождение от человека. См., например, Патент США №4,816,567. Кроме того, если вариабельная область иммуноглобулина получена от другого вида, нежели намеченный реципиент, например, если последовательности вариабельной области происходят от мыши, а намеченным реципиентом является человек, тогда вариабельная область предпочтительно должна включать КО-последовательности человека, а ООК-последовательности мыши располагают между КО-последовательностями с целью создания химерной вариабельной области, которая обладает специфичностью связывания с предварительно выбранным антигеном, но минимальной иммунореактивностью у намеченного реципиента. Конструкция и синтез таких химерных вариабельных областей описаны в работах Jones et al. (1986) NATURE 321:522-525, Verhoyen et al. (1988) SCIENCE 239:1534-1535 и Патентах США №№5,225,539 и 5,585,089. Клонирование и экспрессия гуманизированного белка, слитого из антитела и цитокина, а именно слитого белка КS-1/4 антитело к ЕрСАМ - IL-12, а также его способность уничтожать подтвержденные метастазы карциномы ободочной кишки были описаны в работе Gillies et al. (1998) J. IMMUNOL 160:6195-6203.
Ген, кодирующий цитокин, присоединяют либо непосредственно, либо с помощью линкера, например посредством ДНК, кодирующей линкер (Glγ 4-Sеr)3, присоединенный к 3’-концу гена, кодирующего константную область иммуноглобулина (например, СН2 или СН3 экзон). В некоторых примерах осуществления изобретения линкер может включать последовательность нуклеотидов, кодирующую сайт протеолитического расщепления. Этот сайт, будучи расположен между константной областью иммуноглобулина и цитокином, может быть предназначен для обеспечения протеолитического высвобождения цитокина в месте назначения. Например, хорошо известно, что плазмин и трипсин производят расщепление после остатков лизина и аргинина на сайтах, доступных для протеаз. Многие другие сайт-специфические эндопротеазы и аминокислотные последовательности, которые они расщепляют, хорошо известны в данной области техники. Предпочтительные сайты протеолитического расщепления и протеолитические ферменты, которые действуют на эти сайты расщепления, описаны в Патентах США №№5,541,087 и 5,726,044.
Искусственная нуклеиновая кислота может включать эндогенный промотор и энхансер для гена, кодирующего вариабельную область, для регулирования экспрессии химерной цепи иммуноглобулина. Например, гены, кодирующие вариабельную область, могут быть получены из фрагментов ДНК, включающих лидерный пептид, VJ ген (функционально преобразованные вариабельные (V) области с соединительным (J) сегментом) для легкой цепи или VDJ-ген для тяжелой цепи, а также эндогенные промотор и энхансер для этих генов. Альтернативно, ген, кодирующий вариабельную область, может быть получен без эндогенных регуляторных элементов, а использоваться в экспрессирующем векторе, который предусматривает эти элементы.
Гены вариабельных областей можно получить при помощи стандартных процедур клонирования ДНК из клеток, которые продуцируют желаемые антитела. Скрининг геномной библиотеки на предмет поиска специфической, функционально преобразованной вариабельной области можно осуществить с использованием соответствующих пробных ДНК, таких как сегменты ДНК, содержащие ДНК последовательность J области и нижележащие последовательности. Идентификацию и подтверждение идентичности правильных клонов осуществляют путем секвенирования клонированных генов и сравнения последовательности с соответствующей последовательностью полномерной, соответствующим образом соединенной мРНК.
Целевой антиген может быть поверхностным антигеном опухолевой или раковой клетки, клетки, инфицированной вирусом или другой больной клетки. Гены, кодирующие соответствующие вариабельные области, как правило, можно получить от иммуноглобулинпродуцирующих линий лимфоидных клеток. Например, гибридомные клеточные линии, продуцирующие иммуноглобулин, специфический для антигенов, связанных с опухолями, или вирусных антигенов, могут быть получены с помощью стандартных методик гибридизации соматических клеток, хорошо известных в данной области техники (см., например, Патент США №4,196,265). Эти иммуноглобулинпродуцирующие клеточные линии являются источником генов вариабельной области в функционально преобразованной форме. Гены вариабельной области обычно будут иметь происхождение от мышей, поскольку эта система мыши приспосабливается к производству очень разнообразных иммуноглобулинов с желаемой специфичностью. Кроме того, последовательности вариабельной области могут быть получены с помощью скрининга библиотек, например библиотек фагов, с целью поиска последовательностей вариабельной области, которые связывают предварительно выбранный антиген с желаемой аффинностью. Способы получения и скрининга библиотек фагов описаны, например, в работах Huse et al. (1989) SCIENCE 246:1275-1281 и Kang et al. (1991) PROC. NATL. ACAD. SCI. USA 88:11120-11123.
Фрагмент ДНК, кодирующий ген, содержащий функционально активную вариабельную область, присоединяют к фрагменту ДНК, содержащему ген, кодирующий желаемую константную область (или ее участок). Константные области иммуноглобулина (тяжелую и легкую цепи) можно получить из клеток, продуцирующих антитело, с использованием стандартных способов клонирования генов. Гены двух классов легких цепей человека (κ и λ ) и пяти классов тяжелых цепей человека (α , δ , ε , γ и μ ) были клонированы, и поэтому константные области, имеющие происхождение от человека, легко можно получить из этих клонов.
Слитые гены, кодирующие гибридную тяжелую цепь иммуноглобулина, присоединяют или вставляют в экспрессирующий вектор для введения в клетку-реципиент. Внедрение генной конструкции в плазмидные векторы можно осуществить с помощью стандартных процедур сплайсинга генов. Химерную тяжелую цепь иммуноглобулина можно совместно экспрессировать в одной и той же клетке с соответствующей легкой цепью иммуноглобулина, так что одновременно может экспрессироваться и собираться полный иммуноглобулин. С этой целью конструкции тяжелой и легкой цепей можно поместить в один и тот же вектор или в раздельные векторы.
Линии клеток-реципиентов обычно являются лимфоидными клетками. Предпочтительной клеткой-реципиентом является миеломная (или гибридомная) клетка. Миеломы могут синтезировать, собирать и секретировать иммуноглобулины, кодируемые трансфицированными генами, и они могут гликозилировать белки. Особо предпочтительные реципиенты или клетки-хозяева включают клетки миеломы Sp2/0, которые в норме не продуцируют эндогенного иммуноглобулина, и клетки миеломы мыши NS/0. Будучи трансфицированной, клетка продуцирует только иммуноглобулин, кодируемый трансфицированными генными конструкциями.
Трансфицированные клетки миеломы могут расти в культуре или в брюшине мышей, при этом секретируемый иммуноконъюгат можно выделить из асцитной жидкости. В качестве клеток-реципиентов можно использовать другие лимфоидные клетки, такие как В-лимфоциты.
Существует несколько способов трансфекции лимфоидных клеток векторами, содержащими искусственные нуклеиновые кислоты, кодирующие химерную цепь иммуноглобулина. Например, векторы могут быть внедрены в лимфоидные клетки путем слияния со сферопластами (см., например, Gillies et al. (1989) BIOTECHNOL. 7:798-804). Другие подходящие способы включают электропорацию или преципитацию в фосфате кальция (см., например, Sambrook et al. eds. (1989) "Molecular Cloning: A Laboratory Manual" (“Клонирование молекул:
Лабораторное руководство”), Cold Spring Harbor Press).
Другие способы, пригодные для получения иммуноконъюгатов, включают получение РНК-последовательности, кодирующей конструкцию, и ее трансляцию в соответствующей экспрессирующей системе in vivo или in vitro. Подразумевается, что методологии получения рекомбинантных ДНК для синтеза генов, кодирующих белки, слитые из антитела и цитокина, внедрения генов в клетку-хозяина, экспрессии генов в клетке-хозяине и накопления образующегося в результате этого слитого белка хорошо известны и подробно описаны в данной области техники. Специфические протоколы описаны, например, в работе Sambrook et al. eds. (1989) "Molecular Cloning: A Laboratory Manual" (“Клонирование молекул: Лабораторное руководство”), Cold Spring Harbor Press.
Понятно, что химически соединенные иммуноконъюгаты можно получить с использованием разнообразных способов, хорошо известных специалистам в данной области техники. Например, антитело или фрагмент антитела можно химически присоединить к цитокину с использованием химически реакционноспособных боковых цепей аминокислот антитела или фрагмента антитела и цитокина. Боковые цепи аминокислоты можно соединить ковалентными связями, например посредством дисульфидных связей или при помощи гомо- или гетеро-бифункциональных перекрестносвязывающих реагентов, включающих, например, N-сукцинимидил-3-(2-пиридиилдитио)пропионат, сложный эфир m-малеимидобензоил-N-гидроксисукцинат, сложный эфир m-малеимидобензоил-N-гидроксисульфосукцинимид и 1,4-ди-[3’(2’-пиридилтио)пропионамидо]бутан, все эти реактивы можно приобрести в фирме Pierce, Рокфорд, Иллинойс.
Понятно, что термин “ингибитор ангиогенеза” при использовании в настоящей работе относится к любой молекуле, которая снижает или ингибирует образование новых кровеносных сосудов у млекопитающего. Что касается терапии рака, то ингибитор ангиогенеза снижает или ингибирует образование новых кровеносных сосудов в опухоли или на ней, предпочтительно в плотной опухоли или на ней. Подразумевается, что пригодные для этой цели ингибиторы ангиогенеза можно идентифицировать с использованием разнообразных анализов, хорошо известных и используемых в данной области техники. Такие анализы включают, например, анализ на основе пролиферации клеток эндотелия капилляров быка, анализ с использованием хориоаллантоисной мембраны цыпленка (ХАМ) или анализа с использованием роговицы мыши. Однако ХАМ-анализ предпочтителен (см., например, O’Reilly et al. (1997) CELL 88:277-285). В кратком изложении, эмбрионы вместе с интактными желтками удаляют из оплодотворенных три дня назад белых яиц и помещают в чашки Петри. После инкубации при 37° С и 3% СO2 в течение трех дней на хориоаллантоисную мембрану каждого отдельного эмбриона накладывают диск из метилцеллюлозы, содержащий предполагаемый ингибитор ангиогенеза. После инкубации в течение примерно 48 часов хориоаллантоисные мембраны рассматривают под микроскопом для выявления зон ингибирования.
Многочисленные ингибиторы ангиогенеза хорошо известны и подробно описаны в данной области техники. Примеры ингибиторов ангиогенеза, которые можно использовать в практике осуществления изобретения, включают, например, белково/пептидные ингибиторы ангиогенеза, такие как ангиостатин, протеолитический фрагмент плазминогена (O’Reilly et al. (1994) CELL 79:315-328 и Патенты США с №№5,733,876; 5,837,682 и 5,885,795); эндостатин, протеолитический фрагмент коллагена XVIII (O’Reilly et al. (1997) CELL 88:277-285 и Патент США №5,854,205); пептиды, содержащие трипептидную последовательность RGD и способные связывать αvβ3-интегрин (Brooks (1994) CELL 79:1157-1164); некоторые антитела, их антигенсвязывающие фрагменты и пептиды, которые взаимодействуют с αvβ3-интегрином, обнаруженным на поверхности клеток эпителия сосудов опухоли (Brooks et al., см. выше) или рецептором EGF (Ciardello et al. (1996) J. NATL. CANCER INST. 88:1770-1776). Примеры других ингибиторов ангиогенеза включают ингибиторы COX-2 (Masferrer et al. (1998) PROC. AMER. ASSOC. CANCER RES. 39:271); фумагиллин и его аналоги, такие как AGM-1470 (Ingber et al. (1990) NATURE 348:555-557); и другие мелкие молекулы, такие как талидомид (D’Amato et al. (1994) PROC. NATL. ACAD. SCI. USA 91:4082-4085). Однако в настоящее время более предпочтительны эндостатин и ангиостатин.
Сообщалось, что некоторые цитокины, включая видовые варианты и усеченные аналоги, также обладают антиангиогенной активностью, и поэтому их можно использовать при осуществлении изобретения. Примеры включают ИЛ-12, который, по имеющимся сообщениям, действует через ИФН-γ -зависимый механизм (Voest et al. (1995) J. NATL. CANC. INST. 87:581-586); сам ИФН-γ , который стимулирует хемокин (IP-10) с ангиостатической активностью (Arenberg et al. (1996) J. EXP. MED. 184:981-992). Таким образом, ИЛ-12, ИФН-γ и IP-10 представляют собой ингибиторы ангиогенеза в различных пунктах одного и того же пути ингибирования. Было показано, что другие интерфероны, особенно ИФН-α , являются антиангиогенными, по отдельности или в комбинации с другими ингибиторами (Brem et al. (1993) J. PEDIATR. SURG. 28:1253-1257). Все интерфероны ИФН-α , ИФН-β и ИФН-γ обладают иммунологическими эффектами, а также антиангиогенными свойствами, которые не зависят от их антивирусной активности. Еще один цитокин, ГМ-КСФ, по имеющимся сообщениям, ингибирует ангиогенез посредством стимуляции ангиостатина (Kumar et al. (1998) PROC. AMER. ASSOC. CANCER RES. 39:271).
В соответствии с настоящей работой должно быть понятно, что участок иммуноконъюгата, являющийся антителом, специфически связывает предварительно выбранный антиген, цитокин специфически связывает рецептор цитокина, или ингибитор ангиогенеза специфически связывает рецептор ингибитора, если сродство к антигену или рецептору более 105 М-1 и более предпочтительно более 107 М-1. При использовании в настоящей работе термины ангиостатин, эндостатин, ФНО, ИЛ, ГМ-КСФ, М-КСФ, ЛТ и ИФН относятся не только к интактным белкам, но и к их биоактивным фрагментам и/или аналогам. Биоактивные фрагменты означают участки интактного белка, которые сохраняют, как минимум, 30%, более предпочтительно, как минимум, 70% и наиболее предпочтительно, как минимум, 90% биологической активности интактного белка.
Ингибиторы ангиогенеза можно вводить совместно и одновременно с иммуноконъюгатом или вводить отдельно от него с помощью различных способов введения. Композиции согласно настоящему изобретению можно вводить любым способом, который совместим с определенными молекулами. Поэтому, если это подходит, введение может быть пероральным или парентеральным, включая внутривенный и внутрибрюшинный пути введения.
Композиции согласно настоящему изобретению могут быть введены животному любыми подходящими способами, направленно (например, местно -путем инъекции, имплантации или местного введения в тканевой локус) или системно (например, парентерально или перорально). Если композицию необходимо вводить парентерально, например путем внутривенного, подкожного, внутриглазного, внутрибрюшинного, внутримышечного, буккального, ректального, вагинального, внутриглазничного, внутримозгового, внутричерепного, внутриспинального, внутрижелудочкового, внутриоболочечного, внутрицистернального, внутрикапсулярного, назального введения или в виде аэрозоля, композиция предпочтительно должна включать часть суспензии или раствора в воде или физиологически совместимой жидкости. Поэтому носитель или растворитель должен быть физиологически приемлемым, так чтобы, помимо доставки желаемой композиции больному, он не вызывал неблагоприятных влияний на электролитный и/или объемный баланс больного. Поэтому жидкая среда для агента может включать нормальный физиологический раствор (например, 9,85% водный раствор NaCl, 0,15 М, рН 7-7,4).
Предпочтительные дозировки иммуноконъюгата на одно введение лежат в диапазоне 0,1 - 100 мг/м2, более предпочтительно 1 - 20 мг/м2 и наиболее предпочтительно 2 - 6 мг/м2. Предпочтительные дозировки ингибитора ангиогенеза в основном будут зависеть от типа используемого ингибитора ангиогенеза, тем не менее оптимальные дозировки можно определить путем рутинного экспериментирования. Введение иммуноконъюгата и/или ингибитора ангиогенеза может осуществляться путем периодических болюсных инъекций или путем продолжительного внутривенного или внутрибрюшинного введения из внешнего резервуара (например, из мешка для внутривенного введения) или внутреннего резервуара (например, из биоразрушаемого имплантата). Кроме того, подразумевается, что иммуноконъюгат согласно настоящему изобретению может быть также введен назначенному реципиенту совместно с многими различными ингибиторами ангиогенеза. Тем не менее, подразумевается, что оптимальную комбинацию иммуноконъюгатов и ингибиторов ангиогенеза, способы введения, дозировки можно легко определить путем рутинного экспериментирования в пределах современного уровня техники.
Разнообразные способы можно использовать для оценки эффективности комбинированной терапии с использованием белков, слитых из антитела и цитокина, и ингибиторов ангиогенеза в отношении иммунных ответов. Например, модельный эксперимент на животных, описанный в Примере 1, или любой другой подходящий модельный эксперимент на животных могут быть использованы специалистами в данной области техники для проверки того, какие ингибиторы ангиогенеза или комбинации ингибиторов ангиогенеза наиболее эффективны при синергичном действии с иммуноконъюгатом, например белком, слитым из антитела и цитокина (например, белком, слитым из антитела и ИЛ-2), в отношении повышения уровня иммунной деструкции диагностированных опухолей. Ингибитор ангиогенеза (или комбинация ингибиторов ангиогенеза) может вводиться до курса терапии иммуноконъюгатом или одновременно с ним, а влияние на опухоль можно регистрировать посредством измерения объема. Кроме того, по мере идентификации новых ингибиторов ангиогенеза специалисты в данной области техники смогут использовать методы, описанные здесь, для оценки потенциала этих новых ингибиторов в отношении повышения противораковой активности белков, слитых из антитела и цитокина.
Альтернативно, после терапии опухоли могут быть иссечены, из них можно приготовить срезы и окрасить стандартными гистологическими способами или специфическими иммуногистологическими реагентами с целью оценки эффекта комбинированной терапии на иммунный ответ. Например, простое окрашивание гематоксилином и эозином может выявить различия в инфильтрации лимфоцитами плотных опухолей, которая является показателем клеточного иммунного ответа. Кроме того, иммунологическое окрашивание срезов антителами к специфическим классам иммунных клеток может раскрыть природу индуцированного ответа. Например, антитела, которые связываются с CD45 (основным маркером лейкоцитов), CD4 и CD8 (для идентификации подклассов Т-клеток) и NK1.1 (маркером клеток НК), можно использовать для определения типа иммунного ответа, который опосредован иммуноконъюгатами согласно настоящему изобретению.
Альтернативно, тип иммунного ответа, опосредованного иммуноконъюгатами, можно оценить путем традиционных исследований истощения подклассов клеток, описанных, например, в работе Lode et al. (1998) BLOOD 91:1706-1715. Примеры истощающих антител включают антитела, которые реагируют с маркерами Т-клеток CD4 и CD8, а также антитела, которые присоединяют маркеры НК-клеток NK1.1 и asialoGM. В кратком изложении, эти антитела вводят млекопитающему перед началом лечения антителом-цитокином в достаточно высоких дозах (например, в дозе, примерно равной 0,5 мг/мышь) и затем вводят с недельными интервалами до окончания эксперимента. Эта методика может идентифицировать типы клеток, необходимых для того, чтобы вызвать наблюдаемый иммунный ответ у млекопитающих.
При другом подходе цитотоксическую активность спленоцитов, выделенных из животных, получавших комбинированную терапию, можно сравнить с аналогичной активностью у животных из других экспериментальных групп. Культуры спленоцитов получают путем механического измельчения выделенных стерильных селезенок по стандартным методикам, которые можно найти в большинстве лабораторных руководств по иммунологии. См., например, Coligan et al. (eds) (1988) "Current Protocols in Immunology" (“Современные схемы опытов в иммунологии”), John Wiley & Sons, Inc. Полученные клетки затем культивируют в подходящей культуральной среде (например, DMEM производства компании Gibco), содержащей сыворотку, антибиотики и низкую концентрацию ИЛ-2 (около 10 Ед/мл). Например, чтобы сравнить активность клеток НК, обычно оптимальным является 3-дневное культивирование, тогда как для того, чтобы сравнить цитотоксическую активность Т-клеток, в норме оптимальным является 5-дневное культивирование.
Цитотоксическую активность можно измерить путем мечения опухолевых клеток-мишеней (например, КЛЛ-клеток) радиоактивным изотопом 51Сr в течение 30 минут. После удаления избытка радиоактивной метки меченые клетки смешивают с различными концентрациями культивированных клеток селезенки на 4 часа. В конце инкубации 51Сr, выделившийся из клеток, измеряют с помощью гамма-счетчика, и это значение используют для количественной оценки уровня лизиса клеток, индуцированного иммунными клетками. Таким путем обычно измеряют активность цитотоксических Т-лимфоцитов (или ЦТЛ).
ОПИСАНИЕ ПРИМЕРОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Изобретение далее иллюстрируется следующими, не ограничивающими его, примерами.
Пример 1. Модельный эксперимент на животных.
Модель рака у мышей была разработана для изучения эффекта комбинации белков, слитых из антитела и цитокина, и ингибитора ангиогенеза при опосредовании эффективных иммунных ответов, направленных против опухоли. Белки, слитые из антитела и цитокина, использованные в нижеследующих примерах, связывают ЕрСАМ, антиген опухолей человека, обнаруженный в большинстве опухолей эпителиального происхождения (см., Perez and Walker (1989) J. IMMUNOL. 142:3662-3667). Чтобы проверить их эффективность в модельных опытах на иммунокомпетентных мышах, было необходимо экспрессировать антиген человека на поверхности опухолевой клетки мыши, которая сингенна мыши-хозяину. Для этой цели были выбраны клетки карциномы легких Левиса (КЛЛ), хорошо известная линия клеток раковых опухолей легких у мышей. В результате антиген опухоли человека ЕрСАМ был экспрессирован на поверхности клеток КЛЛ.
Клетки КЛЛ трансфицировали с помощью экспрессирующей плазмиды, содержащей кДНК, кодирующую антиген ЕрСАМ (распознаваемый антителом KS-1/4, как описано в работе Vurki et al. (1984) CANCER RES. 44:681), и инициировали ранним промотором цитомегаловируса (ЦМВ) (производства компании Immunogen, Карлсбад, Калифорния). Антиген KS (KSA или ЕрСАМ) клонировали с помощью полимеразной цепной реакции (ПЦР) из кДНК, полученной из клеток карциномы предстательной железы человека, LnCAP. Прямой праймер имел олигонуклеотидную последовательность 5’TCTAGAGCAGCATGGCGCCCCCGC (SEQ ID NO:1), где ATG является кодоном инициации трансляции, а обратный праймер имел олигонуклеотидную последовательность 5’CTCGAGTTATGCATTGAGTTCCCT (SEQ ID NO:2), где ТТА является антикодоном окончания трансляции. КДНК ЕрСАМ клонировали в ретровирусный вектор pLNCS (производства фирмы Clontech, Пало Альто, Калифорния) и выполняли трансфекцию в соответствии с установленными протоколами (Ausubel et al. (eds) "Current Protocols in Molecular Biology" (“Современные схемы опытов в молекулярной биологии”), John Wiley & Sons). В кратком изложении, упаковочную линию клеток РА317 (АТСС CRL 9078) трансфицировали pLNCX-EpCAM путем совместной преципитации из фосфата кальция, а кондиционированную среду, содержащую вирус, использовали для трансфекции КЛЛ-клеток. G418 (производства Sigma Chemical Co.) добавляли к трансфицированным клеткам в концентрации 1 мг/мл, чтобы отобрать устойчивые клоны. Клоны, экспрессирующие антиген человека ЕрСАМ (LLC-Ep), идентифицировали путем иммунологического окрашивания и анализа на сортировщике флуоресцентноактивированных клеток (ФАКС).
Как изображено на ФИГ.1, клоны LLC-Ep, окрашенные первоначально при помощи слитого белка hu-KS-IL2 (см. Пример 2 ниже), а затем - специфическим антителом к Fc человека, меченным флуоресцеина изотианатом (ФИТЦ) (производства Jackson ImmunoResearch Laboratories, Вест Грув, Пенсильвания), обнаруживали примерно одинаковый уровень экспрессии ЕрСАМ человека. Уровень экспрессии в этих клонах был значительно выше уровня, наблюдавшегося в клонах, окрашенных только специфическим антителом к Fc человека, меченным ФИТЦ.
Чтобы продемонстрировать, что экспрессия поверхностного белка клеток человека не увеличивает иммуногенность клеток LLC-Ep, мышам линии С 57 В 1/6 подкожно вводили различные количества клеток. У всех мышей после инъекции 5× 105 клеток было обнаружено быстрое прогрессивное развитие опухолей с примерно такой же кинетикой роста, что и у родительской линии КЛЛ-клеток. Все животные начинали агонизировать, и их безболезненно умерщвляли, чтобы избежать ненужных мучений.
Пример 2. Получение белков, слитых из антитела и цитокина или антитела и ингибитора ангиогенеза
В нижеследующих примерах обсуждаются разнообразные белки, слитые из антитела и цитокина. В частности, в Примере 3 описано использование белка, слитого из гуманизированного KS, γ 2а мыши и ИЛ-2 мыши (huKS-muγ 2a-muIL2), и белка, слитого из гуманизированного KS, γ 2а мыши и ИЛ-12 мыши (huKS-muγ 2a-muIL12). Пример 4 описывает использование слитого белка huKS-muγ 2a-muIL2 совместно с белками, слитыми из Fc мыши и ангиостатина мыши (muFc-muAngio) и из Fc мыши и эндостатина мыши (muFc-muEndo). Пример 5 описывает использование белка, слитого из гуманизированного KS, γ 4 человека и ИЛ-2 человека (huKS-huγ 4-huIL2), и слитого белка muFc-muEndo. Наконец, Пример 6 описывает использование белка, слитого из гуманизированного KS, γ 1 человека и ИЛ-2 человека (huKS-huγ 1-huIL2), совместно с индометацином. Конструирование этих слитых белков описано ниже.
huKS-huγ 1-huIL2
Ген, кодирующий слитый белок huKS-huγ 1-huIL2, был получен и экспрессирован по существу так же, как описано в работе Gillies et al. (1998) J. IMMUNOL. 160:6195-6203 и Патенте США №5,650,150. В кратком изложении, гуманизированные вариабельные области антитела мыши KS-1/4 (Varki et al. (1984) CANCER RES. 44:681-687) были сконструированы с использованием способов, описанных в работе Jones et al. (1986) NATURE 321:522-525, которые включают вставки ООК из вариабельных областей KS-1/4 в обобщающие типичные каркасные последовательности вариабельных областей человека с наивысшим уровнем гомологии. Моделирование молекул с помощью автоматизированного рабочего места Silicon Gratics Indigo, использующего программное обеспечение BioSym, подтвердило, что формы ООК были сохранены. Белковые последовательности были затем обратно транслированы, а гены сконструированы путем лигирования перекрывающихся олигонуклеотидов.
Образовавшиеся в результате вариабельные области были вставлены в экспрессирующий вектор, содержащий константные области легкой κ -цепи человека и Сγ 1 тяжелой цепи человека, по существу так же, как описано в работе Gillies et al. (1992) PROC. NATL. ACAD. SCI. USA 89:1428-1432, за исключением того, что промоторы металлотионеина и энхансеры тяжелой цепи иммуноглобулина были замещены промотором/энхансером ЦМВ для экспрессии обеих цепей. Слияния зрелых последовательностей ИЛ-2 с карбокси-концами тяжелых цепей человека были произведены, как описано в работе Gillies et al. (1992) PROC. NATL. ACAD. SCI. USA 89:1428-1432, за исключением того, что 3’-нетранслируемые области гена ИЛ-2 были получены из поли(А)-области SV40.
Слитый белок на основе ИЛ-2 был экспрессирован посредством трансфекции результирующей плазмиды в клетки миеломы линии NS/0 с использованием среды для селекции, содержащей 0,1 мкМ метотрексата. Кратко излагая, чтобы получить стабильно трансфицированные клоны, плазмидную ДНК внедряли в клетки миеломы мыши NS/0 посредством электропорации. NS/0 клетки выращивали на среде Eagle, модифицированной по Dulbecco, с добавлением 10% плодной сыворотки коров. Примерно 5× 106 клеток отмывали один раз раствором фосфатного буфера (РФБ) и повторно суспендировали в 0,5 мл РФБ. Затем десять микрограммов линеаризованной плазмидной ДНК инкубировали с клетками в Gene Pulser Cuvette (кювете генного пульсатора) (межэлектродное расстояние 0,4 см, производство BioRad) на льду в течение 10 мин. Электропорацию производили с использованием Gene Pulser (генератора импульсного напряжения) (производства BioRad, Геркулес, Калифорния) с установкой 0,25 В и 500 мкФ. Клеткам давали восстановиться в течение 10 минут на льду, после чего их повторно суспендировали в культуральной среде, а затем наносили на два 96-луночных планшета. Стабильно трансфицированные клоны отбирали по росту в присутствии 100 нМ метотрексата (МТК), который вводили в среду через два дня после трансфекции. Клетки подпитывали через каждые 3 дня еще три раза, и МТК-резистентные клоны появлялись через 2-3 недели.
Экспрессирующие клоны идентифицировали посредством твердофазного иммуноферментного анализа (ELISA) на Fc или цитокин с использованием соответствующих антител (см., например, Gillies et al. (1989) BIOTECHNOL. 7:798-804). Образующийся слитый белок очищали путем связывания и элюирования из белковой А Сефарозы (Pharmacia) в соответствии с инструкциями производителя.
huKS-huγ 4-huIL2
Ген, кодирующий слитый белок huKS-huγ 4-huIL2, конструировали и экспрессировали по существу так же, как описано в заявке на патент США №09/256,156, поданной 24 февраля 1999 г., которая претендует на приоритет в соответствии с заявкой США 60/075,887, поданной 25 февраля 1998 г.
В кратком изложении, вариант слитого белка huKS-hLγ 1-huIL2 на основе Igγ 4, описанного выше, получали путем удаления фрагмента гена Сγ 1 константной области иммуноглобулина из вектора, экспрессирующего huKS-huγ 1-huIL2, и замещения его соответствующей последовательностью из гена Сγ 4 человека. Последовательности и расположение последовательностей константных областей Сγ 1, Сγ 2, Сγ 3 и Сγ 4 тяжелой цепи иммуноглобулина человека описаны в работе Huck et al. (1986) NUC. ACID RES. 14:1779-1789.
Замену фрагмента Сγ 1 на Сγ 4 осуществляли путем ферментативного расщепления исходной Сγ 1-содержащей плазмидной ДНК при помощи Hind III и Xho I и выделения большого фрагмента, размером 7,8 тысяч пар оснований, посредством электрофореза в агарозном геле. Вторую плазмидную ДНК, содержащую ген Сγ 4, расщепляли при помощи Hind III и Nsi I и выделяли фрагмент размером 1,75 тысяч пар оснований. Третью плазмиду, содержащую кДНК ИЛ-2 человека и сайт полиА SV40, соединенный с карбоксильным концом гена Сγ 1 человека, ферментативно расщепляли с помощью Xho I и Nsi I и выделяли малый фрагмент размером 470 пар оснований. Все три фрагмента лигировали вместе примерно в эквимолярных количествах и продукт лигирования использовали для трансформации компетентных Е соli. Продукт лигирования использовали для трансформации компетентных Е. coli, а колонии отбирали по росту на чашках Петри, содержащих ампициллин. Правильно собранные рекомбинантные плазмиды были идентифицированы с помощью рестрикционных анализов препаратов плазмидных ДНК из изолированных трансформантов, а ферментативное расщепление с помощью Fsp I было использовано для различения вставок Сγ 1 (не расщепляется Fsp I) и Сγ 4 (один сайт рестрикции) генов.
Окончательный вектор, содержащий продукт замены в тяжелой цепи на Сγ 4 и ИЛ-2, внедряли в клетки миеломы мыши линии NS/0 путем электропорации (0,25 В и 500 мкФ), а трансфектанты отбирали по росту в среде, содержащей метотрексат (0,1 мкМ). Клоны клеток, экспрессирующие высокие уровни слитого белка huKS-huγ 4-huIL2, идентифицировали, расширяли и выделяли слитый белок из супернатантов культуры с использованием хроматографии на белковой А Сефарозе. Чистоту и целостность слитого белка с Сγ 4 геном определяли путем электрофореза в полиакриламидном геле с ДСН (додецилсульфатом натрия). Активность ИЛ-2 измерили в анализе с пролиферацией Т-клеток (Gillis et al. (1978) J. IMMUNOL. 120:2027-2032) и обнаружили, что она идентична активности γ 1-конструкции.
huKS-muγ 2a-muIL2
Ген, кодирующий слитый белок huKS-muγ 2a-muIL2, конструировали путем замещения константных областей антитела человека и ИЛ-2 человека в слитом белке huKS-huγ 1-huIL2, описанном выше, соответствующими последовательностями мыши. Более конкретно, ДНК, состоящую из гена Сγ 1 и гена ИЛ-2 человека, замещали фрагментом кДНК Сγ 2а мыши, соединенным с ДНК, кодирующей ИЛ-2 мыши. В кратком изложении, VH область КS человека соединяли последовательно с кДНК γ 2а мыши, выполняя ПЦР с перекрытием с использованием перекрывающихся олигонуклеотидных праймеров:
(прямое направление) 5’ СС GTC ТСС ТСА GCC ААА АСА АСА GCC ССА TCG GTC (SEQ ID NO:3);
(обратное направление) 5’ GG GGC TGT TGT TTT GGC TGA GGA GAC GGT GAC TGA CG (SEQ ID NO:4);
(прямое направление) 5’ С ТТА AGC CAG АТС CAG TTG GTG GAG (SEQ ID NO:5);
(обратное направление) 5’ СС CGG GGT CCG GGA GAA GCT СТТ AGT С (SEQ ID NO:6).
Олигонуклеотиды с последовательностями SEQ ID NO:3 и SEQ ID NO:4 были сконструированы для гибридизации к соединению Vн-домена huKS и кДНК константной области γ 2а мыши (курсив). В первом цикле ПЦР было две отдельные реакции. В одной реакции VH ДНК huKS использовали в качестве матрицы с олигонуклеотидами, имеющими последовательности SEQ ID NO:4 и SEQ ID NO:5. Праймер с последовательностью SEQ ID NO:5 внедряли в сайт рестрикции AfIII (CTTAAG) выше последовательности, кодирующей зрелый амино-конец VH huKS (полужирный шрифт). В другой реакции γ 2а-кДНК мыши использовали в качестве матрицы с олигонуклеотидами SEQ ID NO:3 и SEQ ID NO:6. Праймер с последовательностью SEQ ID NO:6 гибридизировали с кДНК, кодирующей область вокруг С-конца γ 2а и внедряли сайт рестрикции Xmal (CCCGGG) для последующего лигирования с кДНК ИЛ-2 мыши. Продукты ПЦР из двух реакций смешивали и подвергали второму циклу ПЦР с использованием олигонуклеотидов с последовательностями SEQ ID NO:5 и SEQ ID NO:6. Образующийся в результате продукт ПЦР клонировали и после подтверждения последовательности фрагмент AfIII-Xmal, кодирующий VH huKS и константную область γ 2а мыши использовали для лигирования с ДНК, кодирующей сигнальный пептид у сайта AfIII и кДНК ИЛ-2 мыши у сайта Xmal.
кДНК ИЛ-2 мыши клонировали из мРНК мононуклеарных клеток периферической крови мыши с использованием олигонуклеотидов, определенных выше как последовательности SEQ ID NO:7 и SEQ ID NO:8, а именно:
(прямое направление) 5’ GGC CCG GGT AAA GCA CCC ACT TCA AGC ТСС (SEQ ID NO:7);
(обратное направление) 5’ CCCTCGAGTTATTGAGGGCTTGTTG (SEQ ID NO:8).
Праймер с последовательностью SEQ ID NO:7 адаптировал ИЛ-2 мыши (последовательность, выделенная полужирным шрифтом) для присоединения к γ 2а мыши у сайта рестрикции Xmal (CCCGGG). Праймер с последовательностью SEQ ID NO:8 внедрял сайт рестрикции Xhol (CTCGAG) непосредственно после кодона окончания трансляции (полужирный шрифт в обратном направлении).
Сходным образом домен вариабельной области легкой цепи (VL) huKS присоединяли к последовательности кДНК κ -области мыши путем ПЦР с перекрытием. Использованные перекрывающиеся олигонуклеотиды включали
(прямое направление) 5’ G GAA АТА ААА CGG GCT GAT GCT GCA CCA ACT G (SEQ ID NO:9),
(обратное направление) 5’ GC AGC ATC AGC CCGTT TTA TT CCA GCT TGG TCC (SEQ ID NO:10);
(прямое направление) 5’ С TTA AGC GAG ATC GTG CTG ACC CAG (SEQ ID NO:11);
(обратное направление) 5’ CTC GAG СТА АСА СТС АТТ ССТ GTT GAA GC (SEQ ID NO:12).
Олигонуклеотиды были сконструированы для гибридизации к соединению VL huKS и константной области κ кДНК мыши (курсив). В первом цикле ПЦР происходило две раздельные реакции. В одной реакции VL ДНК huKS использовали в качестве матрицы с олигонуклеотидами, представленными в виде последовательностей SEQ ID NO:10 и SEQ ID NO:11, которые внедряли сайт рестрикции AfIII (CTTAAG) выше последовательности, кодирующей зрелый амино-конец VL huKS (полужирный шрифт). В другой реакции кДНК κ мыши использовали в качестве матрицы с олигонуклеотидами, представленными выше как последовательности SEQ ID NO:9 и SEQ ID NO:12, которые внедряли сайт рестрикции Xhol после кодона окончания трансляции (полужирный шрифт в обратном направлении).
Продукты ПЦР из двух реакций смешивали и подвергали второму циклу ПЦР с использованием олигонуклеотидов с последовательностями SEQ ID NO:11 и SEQ ID NO:12. Образующийся в результате продукт ПЦР клонировали и после подтверждения последовательности фрагмент AfIII-Xmal, кодирующий VH huKS, и константную область κ мыши лигировали к ДНК, кодирующей сигнальный пептид у сайта АfIII.
Как последовательности тяжелой цепи, так и последовательности легкой цепи мыши использовали для замещения человеческих последовательностей в pdHL7. Образующийся в результате вектор экспрессии антитела, содержащий маркерный ген dhfr, электропорировали (0,25 В, 500 мкФ) в клетки миеломы мыши NS/0 и клоны отбирали путем культивирования в среде, содержащей 0,1 мкМ метотрексата. Трансфицированные клоны, устойчивые к метотрексату, тестировали на предмет секреции детерминант антител с помощью стандартных способов твердофазного иммуноферментного анализа ELISA. Слитые белки очищали с помощью хроматографии на белковой Сефарозе А в соответствии с инструкциями производителя.
huKS-muγ 2a-muIL12
Ген, кодирующий слитый белок huKS-muγ 2a-muIL12, конструировали и экспрессировали в основном так же, как описано в заявке США №08/986,997, поданной 8 декабря 1997 г. и работе Gillies et al. (1998) J. IMMUNOL. 160:6195-6203. В кратком изложении это осуществляли путем присоединения субъединицы кДНК мыши р35 IL-12 к продукту соединения huKS и кодирующей области тяжелой цепи мыши muγ 2а, приготовленному предварительно. Результирующий вектор затем трансфицировали в клетки миеломы линии NS/0, предварительно трансфицированные при помощи субъединицы р40 ИЛ-12 и способные ее экспрессировать. Другими словами, клеточную линию трансфицировали только р40 и отбирали стабильные, с высоким уровнем экспрессии, клетки, которые затем использовали в качестве реципиентов для трансфекции слитым белком, содержащим р35 (то есть производили последовательную трансфекцию).
Субъединицы р35 и р40 ИЛ-12 мыши выделяли при помощи ПЦР из мРНК, полученных из клеток селезенки, активированных Конканавалином А (5 мкг/мл в культуральной среде в течение 3 дней). ПЦР-праймеры, использованные для выделения кодирующей р35 последовательности нуклеиновой кислоты, которые также адаптировали р35 кДНК как фрагмент рестрикции Xmal-Xhol, включали
5’ CCCCGGGTAGGGTCATTCCAGTCTCTGG (SEQ ID NO:13) и 5’ CTCGAGTCAGGCGGAGCTCAGATAGC (SEQ ID NO:14).
ПЦР-праймер, использованный для выделения кодирующей р40 последовательности нуклеиновой кислоты, включал
5’ TCTAGACCATGTGTCCTCAGAAGCTAAC (SEQ ID NO:15) и 5’ CTCGAGCTAGGATCGGACCCTGCAG (SEQ ID NO:16).
Плазмидный вектор (pdHL7-huKS-muγ 2a-p35) конструировали, как описано в работе Gillies et al. J. IMMUNOL. METHODS 125:191, и он включал ген избирательного маркера dhfr, единицу транскрипции, кодирующую легкую цепь гуманизированного антитела KS, и единицу транскрипции, кодирующую тяжелую цепь мыши, соединенную с субъединицей р35 ИЛ-12 мыши. Соединение осуществляли путем лигирования Xmal к фрагменту Xhol адаптированной р35 субъединицы кДНК, к уникальному Xmal сайту на конце СН3-экзона гена γ 2а мыши, полученного ранее. Единицы транскрипции Н и L-цепей включали промотор цитомегаловируса (ЦМВ) (на месте металлотионеинового промотора в исходной ссылке) на 5’-конце и сайт полиаденилирования на 3’-конце.
Сходный вектор (pNC-p40) был сконструирован для экспрессии свободной субъединицы р40, который включал маркерный ген для отбора (ген устойчивости к неомицину), но по-прежнему использовал для транскрипции промотор ЦМВ. Кодирующая область в этом случае включала природную лидерную последовательность субъединицы р40 для нормального переноса к эндоплазматическому ретикулуму и сборки со слитым белком. Плазмиду pNC-p40 электропорировали в клетки, а клетки наносили на чашки Петри и отбирали в среде, содержащей G418. В этом случае супернатанты культуры от устойчивых к лекарственному препарату клонов тестировали с помощью твердофазного иммуноферментного анализа (ELISA) на продукцию субъединицы р40.
Вектор экспрессии pdHL7-huKS-muγ 2a-p35 электропорировали в клетки линии NS/0, уже экспрессирующие р40 мыши, как описано в работе Gillies et al. (1998) J. IMMUNOL. 160:6195-6203. Трансфицированные клоны, устойчивые к метотрексату, анализировали на секрецию детерминант антител и ИЛ-12 мыши с помощью стандартных способов твердофазного иммуноферментного анализа (ELISA). Образующийся белок очищали путем присоединения и элюирования из колонки, заполненной белковой А Сефарозой, в соответствии с инструкциями производителя.
muFc-muEndo
Ген, кодирующий слитый белок muFc-muEndo, был сконструирован и экспрессирован по существу так же, как описано в Заявке США №60/097,883, поданной 25 августа 1998 г.
В кратком изложении эндостатин мыши и Fc мыши экспрессировали как слитый белок muFc-muEndostatin. ПЦР использовали для адаптации гена эндостатина для экспрессии в векторе pdCs-muFc(D4K) (Lo et al. (1998) PROTEIN ENGINEERING 11:495-500), который содержит сайт распознавания для энтерокиназы Asp4-Lys (La Vallie et al. (1993) J. BIOL. CHEM. 268:23311-23317).
Прямой праймер имел вид 5’-С ССС AAG СТТ САТ ACT CAT CAG GAC TTT С (SEQ ID NO:17), где за AAGCCT (сайтом HindIII) следовала последовательность (полужирный шрифт), кодирующая N-конец эндостатина. Обратный праймер имел вид 5’-ССС СТС GAG СТА TTT GGA GAA AGA GGT С (SEQ ID NO:18) и был сконструирован для осуществления трансляции СТОП-кодона (антикодон, СТА) сразу же после С-конца эндостатина, после чего следовал сайт Xhol (CTCGAG).
Продукт ПЦР клонировали и секвенсировали, а фрагмент HindIII-Xhol, кодирующий эндостатин, лигировали в вектор pdCs-muFc(D4K). Стабильные NS/0 клоны, экспрессирующие muFc(D4K)-muEndo отбирали и анализировали с использованием твердофазного иммуноферментного анализа ELISA с помощью антител к Fc мыши. Образующийся слитый белок экспрессировали и очищали посредством хроматографии на белковой А Сефарозе.
muFc-muAnqio
Ген, кодирующий слитый белок MuFc-muAngio, был сконструирован и экспрессирован по существу так же, как описано в Заявке США №60/097,883, поданной 25 августа 1998 г.
В кратком изложении ангиостатин мыши и Fc мыши экспрессировали как слитый белок muFc-muAngiostatin. ПЦР использовали для адаптации последовательности нуклеиновой кислоты, кодирующей ангиостатин, любезно предоставленной лабораторией Д-ра Юде Фолькмана, Детская больница, Бостон, Миннесота, для экспрессии в векторе pdCs-Fc(D4K) (Lo et al. (1998) PROTEIN ENGINEERING 11:495-500). Прямым праймером был 5’-С ССС AAG CTT GTG ТАТ CTG ТСА GAA TGT AAG ССС ТСС TGT СТС TGA GCA (SEQ ID NO:19), где за AAGCTT (сайтом HindIII) следовала последовательность (полужирный шрифт), кодирующая N-конец ангиостатина. Обратным праймером был 5’-ССС СТС GAG СТА ССС ТСС TGT СТС TGA GCA (SEQ ID NO:20), который был сконструирован для осуществления трансляции СТОП-кодона (антикодона, СТА) непосредственно после С-конца ангиостатина, после чего следовал сайт Xhol (CTCGAG).
Продукт ПЦР клонировали и секвенсировали, а фрагмент HindIII-Xhol, кодирующий ангиостатин, лигировали в вектор pdCs-muFc(D4K). Стабильные NS/0 клоны, экспрессирующие muFc(D4K)-muAngio отбирали и анализировали с использованием твердофазного иммуноферментного анализа ELISA с помощью антител к Fc мыши. Образующийся слитый белок экспрессировали и очищали посредством хроматографии на белковой А Сефарозе.
Пример 3. Комбинированная терапия с использованием слитых белков KS-IL2 и КS-11-12 для лечения опухолей LLC-Ep.
Самкам мышей С57В1/6 вводили подкожно в среднюю часть спины клетки LLC-Ep (5× 105 одной мыши), выращенные в культуре клеток. Примерно через две недели животных с пальпируемыми опухолями в диапазоне 150-400 мм3 делили на четыре группы с одинаковым распределением опухолей по размеру между группами. Животных обрабатывали следующим образом: в 1 группе животные получали только раствор фосфатного буфера (РФБ) (контрольная группа; во 2 группе животные получали только слитый белок huKS-muγ 2a-muIL2; в 3 группе животные получали только слитый белок huKS-muγ 2a-muIL12 и в 4 группе животные получали оба слитых белка - huKS-muγ 2a-muIL2 и huKS-muγ 2a-muIL12. Рост опухолей регистрировали с помощью измерений объема до тех пор, пока животные в контрольной группе не начали агонизировать и не были подвергнуты эвтаназии. Объемы опухолей были измерены с помощью штангенциркуля и рассчитаны по формуле
V=4π /3(0,5L× 0,5W× 0,5H),
где L - длина, W - ширина и Н - высота опухоли.
Результаты суммированы на ФИГ.3. Мыши, получавшие РФБ, обозначены незаполненными ромбиками; мыши, получавшие 15 мкг/день слитого белка huKS-muγ 2a-muIL2, обозначены сплошными квадратиками; мыши, получавшие 10 мкг/день слитого белка huKS-muγ 2a-muIL12, обозначены сплошными треугольниками и мыши, получавшие комбинацию 7,5 мкг/день слитого белка huKS-muγ 2a-muIL2 и 5 мкг/день слитого белка huKS-muγ 2a-rnuIL12, обозначены крестиками.
Как проиллюстрировано на ФИГ.3, лечение слитым белком huКS-muγ 2а-muIL2 (по 15 мкг в течение 5 последовательных дней) не задерживало и не уменьшало рост опухолей LLC-Ep (сплошные квадратики). Только небольшой противоопухолевый эффект был обнаружен у мышей, которых лечили только слитым белком huKS-muγ 2a-muIL12 в дозе 15 мкг/введение в течение пяти последовательных дней (сплошные треугольники). Тем не менее, если два слитых белка комбинировали, используя половину исходного количества каждого белка (7,5 мкг huKS-muγ 2a-muIL2 и 5 мкг huKS-muγ 2a-muIL12 соответственно), наблюдалась резкая задержка роста опухоли, доказывающая синергию между двумя слитыми белками.
Пример 4. Терапия с использованием белка, слитого из антитела и цитокина, и комбинации ангиостатина и эндостатина.
Самкам мышей С57В1/6 вводят подкожно в среднюю часть спины клетки LLC-Ep (5× 105 одной мыши), выращенные в культуре клеток. Примерно через две недели животных с пальпируемыми опухолями в диапазоне 150-400 мм3 делят на четыре группы с одинаковым распределением опухолей по размеру между группами. Животных обрабатывают следующим образом: (1) животные, получающие только раствор фосфатного буфера (РФБ); (2) животные, получающие смесь muFc-Angio и muFcEndo; (3) животные, получающие слитый белок huKS-muγ 2a-muIL2; и (4) животные, получающие как слитый белок huKS-muγ 2a-muIL2, так и смесь muFc-Angio и muFcEndo. Животным делают инъекции соответствующего препарата в течение пяти последовательных дней.
muFc-Angio и muFcEndo вводят мышам, имеющим опухоли, в дозе 5 мг/кг. Животным, получающим слитый белок huKS-muγ 2a-muIL2, вводят дневные дозы по 10 мкг huKS-muγ 2a-muIL2 в течение в общей сложности 5 дней. Рост опухолей регистрируют при помощи измерений объема.
Предполагается, что лечение только смесью muFc-Angio и muFcEndo будет эффективно в отношении уменьшения размеров опухоли через двухнедельный период лечения. Тем не менее предполагается, что это лечение не будет столь же эффективным через короткий промежуток времени, например через пять дней. Также предполагается, что животные, которые получают лечение только слитым белком huKS-muγ 2a-muIL2, обнаружат только минимальную противоопухолевую активность. Тем не менее предполагается, что группа, получающая и слитый белок huKS-muγ 2a-muIL2, и смесь muFc-Angio и muFcEndo, обнаружит достоверно сниженный опухолевый рост по сравнению с группой, получавшей только muFc-Angio и muFcEndo, и группой, получавшей слитый белок muKS-muγ 2a-muIL2.
Пример 5. Комбинированная терапия белком, слитым из антитела и цитокина, и эндостатином.
Клетки карциномы мышей СТ26, экспрессирующие ЕрСАМ человека, вводили подкожно в выбритые спины мышей BALB/c (2× 106 клеток за одну инъекцию). Когда опухоли достигали размеров 100-200 мм3 (примерно через 7-14 дней), мышей случайным образом делили на четыре группы, по 4 мыши в каждой группе. Группа 1 получала внутривенные инъекции 0,2 мл РФБ ежедневно. Группа 2 получала внутривенные инъекции muFc-muEndostatin (320 мкг/мышь) в РФБ ежедневно. Группа 3 получала внутривенные инъекции слитого белка huKS-huγ 4-huIL2 (10 мкг/мышь) в РФБ ежедневно только в течение 5 дней. Группа 4 получала внутривенные инъекции комбинации huKS-huγ 4-huIL2 (10 мкг/мышь) и muFc-muEndo (320 мкг/мышь) в РФБ ежедневно в течение 5 дней, а после этого ежедневные инъекции muFc-muEndo (320 мкг/мышь) в РФБ. Объемы опухолей измеряли, как описано в Примере 3.
Результаты суммированы на ФИГ.4. Мыши, получавшие РФБ, представлены сплошными ромбиками; мыши, получавшие слитый белок muFc-muEndo, представлены сплошными квадратиками; мыши, получавшие слитый белок huKS-huγ 4-huIL2, представлены сплошными треугольниками, а мыши, получавшие комбинацию слитого белка muFc-muEndo и слитого белка huKS-huγ 4-huIL2, представлены крестиками.
ФИГ.4 показывает, что комбинация белка, слитого из антитела и цитокина, и антиангиогенного белка muFc-muEndo, превосходила по эффекту любой из отдельных агентов. После лечения в течение 19 дней отношение Т/С (средний размер опухолей в группе, получавшей лечение/средний размер опухолей в контрольной группе) при комбинированной терапии huKS-huγ 4-huIL2 и muFc-muEndo было равно 0,25, что достоверно превышало Т/С, равное 0,31, в группе с huKS-huγ 4-huIL2 и 0,42 в группе с muFc-muEndo.
Пример 6. Комбинированная терапия белком, слитым из антитела и цитокина, и индометацином
Самкам мышей С57В1/6 вводили подкожно в среднюю часть спины клетки LLC-Ep (2× 106 клеток за одну инъекцию). Когда опухоли достигали размер 600-1200 мм3, мышей безболезненно умерщвляли. Кожу над опухолью промывали бетадином и бетанолом, опухоли иссекали и удаляли некротическую ткань. Суспензию опухолевых клеток в растворе фосфатного буфера готовили посредством пропускания жизнеспособной опухолевой ткани через сито, а затем - через ряд последовательно уменьшающихся гипотермических игл размера 22-30. Клетки разводили до концентрации 1× 107 клеток/мл и помещали на лед. Затем мышам C57BL/6 инъецировали 0,1 мл свежересуспензированных клеток (1× 106 клеток на одну мышь) подкожно проксимально от средней линии спины.
Когда опухоли достигали размеров 100-200 мм3 (примерно через 7-14 дней), мышей случайным образом делили на 4 группы, по 5 мышей в каждой группе. Группа 1 получала внутривенные инъекции 0,2 мл РФБ ежедневно. Группа 2 получила 5 ежедневных внутривенных инъекций huKS-huγ 1-huIL2 (25 мкг/мышь) в РФБ. Группа 3 получала индометацин перорально в питьевой воде (20 мкг/мл, или примерно 60-70 мкг индометацина в день, потребленного одной мышью) в течение всего периода лечения. Группа 4 получила 5 ежедневных внутривенных инъекций huKS-huγ 1-huIL2 (25 мкг/мышь) и индометацин перорально в питьевой воде (20 мкг/мл) в течение периода лечения. Объемы опухолей измеряли, как описано в Примере 3.
Результаты представлены на ФИГ.5. Мыши, получавшие РФБ, обозначены сплошными ромбиками; мыши, получавшие слитый белок huKS-huγ 1-huIL2, обозначены сплошными квадратиками; мыши, получавшие индометацин, обозначены сплошными треугольниками; а мыши, получавшие комбинацию слитого белка huKS-huγ 1-huIL2 и индометацина, обозначены крестиками.
ФИГ.5 показывает, что комбинация белка, слитого из антитела и цитокина, и антиангиогенного химического препарата индометацина превосходила по эффекту каждый из агентов по отдельности. После лечения в течение 22 дней отношение Т/С в случае комбинированной терапии huКS-huγ 4-huIL-2 и индометацином было равно 0,40, что было достоверно выше Т/С, равного 0,61 для huKS-huγ 4-huIL2, и 0,60 для индометацина.
ЭКВИВАЛЕНТЫ
Изобретение может быть осуществлено в других специфических формах без отклонения от его сущности или существенных характеристик. Вышеизложенные примеры осуществления изобретения следует поэтому считать во всех аспектах скорее иллюстративными, а не ограничивающими описанное здесь изобретение. Поэтому объем изобретения в большей степени определяется прилагаемой формулой изобретения, а не вышеприведенным описанием, и все изменения, которые возникают в пределах сущности и уровня эквивалентности формулы изобретения, должны быть включены в него.
ВКЛЮЧЕНИЕ ПОСРЕДСТВОМ ССЫЛОК
Все вышеупомянутые патентные документы и научные публикации включены в настоящую заявку посредством ссылок.





Изобретение относится к области иммунологии. Сущность изобретения состоит в том, что открыты композиции и способы для усиления цитоцидных иммунных ответов, направленных против предварительно выбранного типа клеток, у млекопитающих. Способы и композиции основаны на сочетании иммуноконъюгата типа антитело-цитокин и ингибитора ангиогенеза. При введении млекопитающему иммуноконъюгат вызывает иммунный ответ против предварительно выбранного типа клеток, например раковых клеток, который в результате синергии с ингибитором ангиогенеза больше иммунного ответа, вызываемого одним иммуноконъюгатом. Способы и композиции особенно подходят для уничтожения плотных опухолей или инфицированных вирусом клеток млекопитающих. Технический результат - расширение арсенала средств-иммуномодуляторов. 3 с. и 24 з.п. ф-лы, 5 ил.
(1) иммуноконъюгат, содержащий сайт связывания антитела, способный связываться с антигеном на указанном предварительно выбранном типе клеток, и цитокин, способный индуцировать иммунный ответ, направленный против предварительно выбранного типа клеток у млекопитающих, и
(2) ингибитор ангиогенеза в количестве, достаточном для усиления указанного иммунного ответа, вызываемого иммуноконъюгатом комбинации, по сравнению с иммуноконъюгатом, введенным отдельно.
| XIANG et al | |||
| Cancer Research, 1997, 57, р.4948-4955 | |||
| LODE et al | |||
| Blood, Virology, 1998, v.91, p.1706-1705 | |||
| Феррозонд | 1978 |
|
SU706799A1 |

