Область техники, к которой относится изобретение
Настоящее изобретение относится в целом к композициям и способам, которые являются пригодными для модулирования развития сосудов. Конкретно, настоящее изобретение относится к антителам, которые связываются с полипептидом домена 7, подобного эпидермальному фактору роста (EGFL7). Настоящее изобретение, кроме того, относится к диагностике и лечению состояний и заболеваний, связанных с ангиогенезом.
Уровень техники
Развитие переноса с помощью сосудов является фундаментальным требованием для множества физиологических и патологических процессов. Активно растущие ткани, такие как эмбрионы и опухоли, требуют адекватной подачи крови. Они удовлетворяют эту потребность посредством продуцирования проангиогенных факторов, которые способствуют формированию новых кровеносных сосудов с помощью процесса, называемого ангиогенезом. Формирование трубочек сосудов представляет собой сложное, но упорядоченное биологическое событие, включающее в себя все следующие далее стадии или многие из них: a) эндотелиальные клетки (EC) пролиферируют из существующих EC или дифференцируются от прогениторных клеток; b) EC мигрируют и коалесцируют с формированием тяжеобразных структур; c) затем сосудистые тяжи подвергаются тубулогенезу с формированием сосудов с центральным просветом; d) существующие тяжи или сосуды высылают отростки для формирования вторичных сосудов; e) примитивное сосудистое сплетение подвергается дальнейшей перестройке и переформированию; и f) периэндотелиальные клетки рекрутируются для окружения эндотелиальных трубочек, обеспечивая обслуживающие и модуляторные функции для сосудов; такие клетки включают в себя перициты для малых капилляров, клетки гладких мышц для сосудов большего размера и клетки миокарда в сердце. Hanahan, Science 277:48-50 (1997); Hogan & Kolodziej, Nat. Rev. Genet. 3:513-23 (2002); Lubarsky & Krasnow, Cell 112:19-28 (2003).
Сейчас хорошо установлено, что ангиогенез осуществляется при патогенезе разнообразных расстройств. Они включают в себя твердые опухоли и метастазы, атеросклероз, ретролентальную фиброплазию, гемангиомы, хроническое воспаление, интраокулярные неоваскулярные заболевания, такие как пролиферативные ретинопатии, например, диабетическая ретинопатия, возрастная макулярная дегенерация (AMD), неоваскулярная глаукома, иммунное отторжение трансплантированной корнеальной ткани и других тканей, ревматоидный артрит и псориаз. Folkman et al., J. Biol Chem. 267:10931-34 (1992); Klagsbrun et al., Annu. Rev. Physiol. 53:217-39 (1991); и Garner A., "Vascular diseases," In: Pathobiology of Ocular Disease. A Dynamic Approach, Garner A., Klintworth GK, eds., 2nd Edition (Marcel Dekker, NY, 1994), pp. 1625-1710.
В случае роста опухоли, ангиогенез видимо является критичным для перехода от гиперплазии к неоплазии, и для обеспечения питания для роста и метастазирования опухоли. Folkman et al., Nature 339:58 (1989). Неоваскуляризация дает возможность опухолевым клеткам для получения преимущества в росте и пролиферативной автономии, по сравнению с нормальными клетками. Опухоль обычно начинается как отдельная аберрантная клетка, которая может пролиферировать только до размера в несколько кубических миллиметров, из-за дистанции до ближайшего доступного капиллярного русла, и она может оставаться 'пассивной' без дальнейшего роста и распространения в течение продолжительного периода времени. Некоторые опухолевые клетки затем переключается на ангиогенный фенотип с активированием эндотелиальных клеток, которые пролиферируют и созревают в виде новых капиллярных кровеносных сосудов. Эти вновь сформированные кровеносные сосуды не только делают возможным непрерывный рост первичной опухоли, но также служат для распространения и реколонизации метастатических опухолевых клеток. Соответственно, наблюдается корреляция между плотностью микрососудов в срезах опухоли и выживаемостью пациентов при раке груди, а также при некоторых других опухолях. Weidner et al., N. Engl. J. Med. 324:1-6 (1991); Horak et al., Lancet 340:1120-24 (1992); Macchiarini et al., Lancet 340:145-46 (1992). Точные механизмы, которые контролируют ангиогенное переключение, как следует, непонятны, но предполагается, что неоваскуляризация массы опухоли возникает в результате общего баланса действия множества стимуляторов и ингибиторов ангиогенеза (Folkman, Nat. Med. 1(1):27-31 (1995)).
Процесс развития сосудов плотно регулируется. К настоящему времени, значительное количество молекул, в основном, секретируемых факторов, продуцируемых окружающими клетками, как показано, регулируют дифференциацию, пролиферацию, миграцию и коалесценцию EC в виде тяжеобразных структур. Например, фактор васкулярного эндотелиального роста (VEGF) идентифицирован в качестве ключевого фактора, вовлеченного в стимулирование ангиогенеза и в индуцирование проницаемости сосудов. Ferrara et al., Endocr. Rev. 18:4-25 (1997). Обнаружение того, что потеря даже одной аллели VEGF приводит к летальности эмбриона, показывает незаменимую роль, которую играет этот фактор при развитии и дифференцировании сосудистой системы. Кроме того, VEGF, как показано, является ключевым медиатором неоваскуляризации, связанной с опухолями и интраокулярными расстройствами. Ferrara et al., Endocr. Rev, выше. мРНК VEGF сверхэкспрессируется в большинстве исследуемых опухолях человека. Berkman et al., J. Clin. Invest. 91:153-59 (1993); Brown et al., Human Pathol. 26:86-91 (1995); Brown et al., Cancer Res. 53:4727-35 (1993); Mattern et al., Brit. J. Cancer 73:931-34 (1996); Dvorak et al., Am. J. Pathol. 146:1029-39 (1995).
Также, уровни концентрации VEGF в глазных жидкостях сильно коррелируют с присутствием активной пролиферации кровеносных сосудов у пациентов с диабетическими и другими связанными с ишемией ретинопатиями. Aiello et al., N. Engl. J. Med. 331:1480-87 (1994). Кроме того, исследования продемонстрировали локализацию VEGF в хороидальных неоваскулярных мембранах у пациентов, подверженных AMD. Lopez et al., Invest. Ophthalmol. Vis. Sci. 37:855-68 (1996).
Нейтрализующие антитела анти-VEGF подавляют рост разнообразных линий клеток опухолей человека у голых мышей (Kim et al., Nature 362:841-44 (1993); Warren et al., J. Clin. Invest. 95:1789-97 (1995); Borgstrom et al., Cancer Res. 56:4032-39 (1996); Melnyk et al., Cancer Res. 56:921-24 (1996)), а также ингибируют интраокулярный ангиогенез у моделей ишемических ретинальных расстройств (Adamis et al., Arch. Ophthalmol. 114:66-71 (1996)). По этой причине, моноклональные антитела анти-VEGF или другие ингибиторы действия VEGF являются обещающими кандидатами для лечения опухолей и различных интраокулярных неоваскулярных расстройств. Такие антитела описываются, например, в EP 817648, опубликованном 14 января, 1998 года; и в заявках WO 98/45331 и WO 98/45332, обе они опубликованы 15 октября, 1998 года. Одно из антител анти-VEGF, бевацизумаб, было одобрено FDA (администрация по лекарственным средствам и пищевым продуктам США) для использования в сочетании с режимом химиотерапии для лечения метастатического рака толстой и прямой кишки (CRC). И еще бевацизумаб исследуется во множестве осуществляемых клинических исследований для лечения различных проявлений рака.
Известно, что внеклеточный матрикс (ECM) играет важную роль в процессе ангиогенеза. Madri, Transpl. Immunol. 5:179-83 (1997). EC окружены временным ECM во время их миграции, и они прилипают в вновь синтезированным базальным мембранам сосуда после формирования просвета. В дополнение к созданию скелета во время морфогенеза капилляра, ECM, как показано, осуществляет комплексный локальный контроль функционирования EC. Например, ECM способен регулировать доступность растворимых ангиогенных медиаторов для EC и конкретизировать природу и тип взаимодействий с интегрином и клеточную адгезию молекул. Предполагается также, что выживаемость EC регулируется кооперацией между рецепторами фактора роста и интегринами, которые, в свою очередь, управляются композицией локального ECM. Stupack & Cheresh, Oncogene 22:9022-29 (2003).
Несмотря на множество достижений в области ангиогенеза, некоторые из стадий во время формирования трубки сосуда по-прежнему определены плохо. Особенно мало известно относительно того, как регулируется тубулогенез - как сосудистые тяжи развиваются, становясь трубочками, и какие факторы регулируют этот переход. Имея ввиду роль ангиогенеза во многих заболеваниях и расстройствах, является желательным создание средств уменьшения или ингибирования одного или нескольких биологических воздействий, вызывающих эти процессы. Является также желательным создание средств анализа присутствия патогенных полипептидов в нормальных и болезненных состояниях, в особенности при раке. Существует также необходимость в композициях и способах, которые могут повысить эффективность существующих терапевтических подходов против ангиогенеза.
Сущность изобретения
Настоящее изобретение частично основывается на идентификации антител против EGFL7 со свойствами, которые показывают, что они являются особенно полезными для терапии.
В одном из аспектов, настоящее изобретение предоставляет антитела, продуцируемые гибридомами анти-EGFL7 mumab 4F11.1.8, анти-EGFL7 mumab 10G9.1.6 и анти-EGFL7 mumab 18F7.1.8.
В одном из аспектов, настоящее изобретение предоставляет антитело анти-EGFL7, содержащее одну или несколько комплементарно определяемых областей (CDR), выбранных из группы, состоящей из: (a) последовательности 4F11 CDR-L1 KASQSVDYDGDSYMS (SEQ ID NO: 5); (b) последовательности 4F11 CDR-L2 GASNLES (SEQ ID NO: 6); (c) последовательности 4F11 CDR-L3 QQNNEDPYT (SEQ ID NO: 7); (d) последовательности 4F11 CDR-H1 TYGMS (SEQ ID NO: 8); (e) последовательности 4F11 CDR-H2 WINTHSGVPTYADDFKG (SEQ ID NO: 9); и (f) последовательности 4F11 CDR-H3 LGSSA (SEQ ID NO: 10). В некоторых вариантах осуществления, легкая цепь указанного антитела содержит, по меньшей мере, одну, по меньшей мере, две или все три последовательности CDR, выбранных из: KASQSVDYDGDSYMS (SEQ ID NO: 5), GASNLES (SEQ ID NO: 6), и QQNNEDPYT (SEQ ID NO: 7). В некоторых вариантах осуществления, тяжелая цепь указанного антитела содержит, по меньшей мере, одну, по меньшей мере, две или все три последовательности CDR, выбранных из: TYGMS (SEQ ID NO: 8), WINTHSGVPTYADDFKG (SEQ ID NO: 9), и LGSSA (SEQ ID NO: 10). В некоторых вариантах осуществления, легкая цепь указанного антитела содержит, по меньшей мере, одну, по меньшей мере, две или все три последовательности CDR, выбранных из: KASQSVDYDGDSYMS (SEQ ID NO: 5), GASNLES (SEQ ID NO: 6), и QQNNEDPYT (SEQ ID NO: 7); и тяжелая цепь указанного антитела содержит, по меньшей мере, одну, по меньшей мере, две или все три последовательности CDR, выбранных из: TYGMS (SEQ ID NO: 8), WINTHSGVPTYADDFKG (SEQ ID NO: 9), и LGSSA (SEQ ID NO: 10). В некоторых вариантах осуществления, легкая цепь антитела содержит последовательность: DIVLTQSPASLAVSLGQRATISCKASQSVDYDGDSYMSWYQQKPGQPPKLLIYGASNLESGIPARFSGSGSGTDFTLNIHPVEEEDAATYYCQQNNEDPYTFGGGTKVEIKR (SEQ ID NO: 1). В некоторых вариантах осуществления, тяжелая цепь антитела содержит последовательность: QIQLVQSGPELKKPGETVKISCKASGHTFTTYGMSWVKQAPGKGLKWMGWINTHSGVPTYADDFKGRFAFSLETSASTAHLQINNLKNEDTATYFCARLGSSAVDYWGQGTTVTVSS (SEQ ID NO: 2).
В одном из аспектов, настоящее изобретение предоставляет антитело анти-EGFL7, содержащее одну или несколько комплементарно определяемых областей (CDR), выбранных из группы, состоящей из: (a) последовательности 10G9 CDR-L1 RSSQSLVIITNGITYLH(SEQIDNO: 11); (b) последовательности 10G9 CDR-L2 KVSNRFS (SEQ ID NO: 12); (c) последовательности 10G9 CDR-L3 SQSTHVPLT (SEQ ID NO: 13); (d) последовательности 10G9 CDR-H1 DYYMNSDYYMN (SEQ ID NO: 14); (e) последовательности 10G9 CDR-H2 DINPKNGGTTYNQKFKG (SEQ ID NO: 15) и (f) последовательности 10G9 CDR-H3 ALGVFDY (SEQ ID NO: 16). В некоторых вариантах осуществления, легкая цепь указанного антитела содержит, по меньшей мере, одну, по меньшей мере, две или все три последовательности CDR, выбранных из: RSSQSLVHTNGITYLH (SEQ ID NO: 11), KVSNRFS (SEQ ID NO: 12), и SQSTHVPLT (SEQ ID NO: 13). В некоторых вариантах осуществления, тяжелая цепь указанного антитела содержит, по меньшей мере, одну, по меньшей мере, две или все три последовательности CDR, выбранных из: DYYMNSDYYMN (SEQ ID NO: 14), DINPKNGGTTYNQKFKG (SEQ ID NO: 15) и ALGVFDY (SEQ ID NO: 16). В некоторых вариантах осуществления, легкая цепь указанного антитела содержит, по меньшей мере, одну, по меньшей мере, две или все три последовательности CDR, выбранных из: RSSQSLVHTNGITYLH (SEQ ID NO: 11), KVSNRFS (SEQ ID NO: 12), и SQSTHVPLT (SEQ ID NO: 13); и тяжелая цепь указанного антитела содержит, по меньшей мере, одну, по меньшей мере, две или все три последовательности CDR, выбранных из: DYYMNSDYYMN (SEQ ID NO: 14), DINPKNGGTTYNQKFKG (SEQ ID NO: 15), и ALGVFDY (SEQ ID NO: 16). В некоторых вариантах осуществления, легкая цепь антитела содержит последовательность: DIVMTQTPLSLPVSLGDQASISCRSSQSLVHTNGITYLHWYLQKPGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDLGVYFCSQSTHVPLTFGAGTKVEIKR (SEQ ID NO: 3). В некоторых вариантах осуществления, тяжелая цепь антитела содержит последовательность: EVQLQQSGPELVKPGASVKISCKASGYTFSDYYMNSDYYMNWVKQSHGKSLEWIGDINPKNGGTTYNQKFKGKATLTVDKSSSTAYMELRSLTSEDSAVYYCAREADYDPIYYAMDYWGQGTTLTVSA (SEQ ID NO: 4).
В некоторых вариантах осуществления, настоящее изобретение предоставляет антитела анти-EGFL7, которые специфично связываются с полипептидом, содержащим одну из следующих последовательностей аминокислот: CCP, TIY и ACS. В некоторых вариантах осуществления, настоящее изобретение предоставляет выделенные антитела, которые связываются с таким же эпитопом EGFL7 человека, как и другие антитела по настоящему изобретению. В некоторых вариантах осуществления, настоящее изобретение предоставляет выделенные антитела, которые конкурируют за связывание EGFL7 с другими антителами по настоящему изобретению.
В некоторых вариантах осуществления, антитело по настоящему изобретению представляет собой моноклональное антитело. В некоторых вариантах осуществления, антитело по настоящему изобретению представляет собой химерное антитело, гуманизированное антитело, антитело с созревшим сродством, антитело человека или биспецифичное антитело. В некоторых вариантах осуществления, антитело представляет собой фрагмент антитела.
В некоторых вариантах осуществления, настоящее изобретение предоставляет фармацевтическую композицию, содержащую антитело анти-EGFL7 по настоящему изобретению. В некоторых вариантах осуществления, фармацевтическая композиция дополнительно содержит антиангиогенный агент. В некоторых вариантах осуществления, антиангиогенный агент представляет собой бевацизумаб или ранибизумаб.
В некоторых вариантах осуществления, настоящее изобретение предоставляет полинуклеотид, кодирующий антитело по настоящему изобретению. В некоторых вариантах осуществления, настоящее изобретение предоставляет векторы, содержащие эти полинуклеотиды. В некоторых вариантах осуществления, векторы представляют собой векторы экспрессии. В некоторых вариантах осуществления, настоящее изобретение предоставляет клетки-хозяева, включая прокариотические и эукариотические клетки (включая клетки млекопитающих), содержащие такие векторы. В некоторых вариантах осуществления, настоящее изобретение предоставляет способ получения антитела анти-EGFL7, включающий в себя (a) экспрессирование вектора экспрессии в соответствующей клетке-хозяине и (b) извлечение антитела.
В некоторых вариантах осуществления, настоящее изобретение предоставляет способ уменьшения или ингибирования ангиогенеза у субъекта, имеющего патологическое состояние, связанное с ангиогенезом, включающий в себя введение субъекту эффективного количества антитела анти-EGFL7 по настоящему изобретению или фармацевтической композиции, содержащей антитело анти-EGFL7 по настоящему изобретению. В некоторых вариантах осуществления, патологическое состояние представляет собой неоплазму, например, карциному. В некоторых вариантах осуществления, патологическое состояние связано с глазом, например, представляет собой интраокулярное неоваскулярное заболевание. В некоторых вариантах осуществления, антиангиогенный агент вводится субъекту в дополнение к антителу анти-EGFL7 по настоящему изобретению. В некоторых вариантах осуществления, антиангиогенный агент представляет собой антагонист фактора васкулярного эндотелиального роста (VEGF), например, антитело анти-VEGF (включая бевацизумаб и ранибизумаб). В некоторых вариантах осуществления, антиангиогенный агент вводится до или после введения антитела анти-EGFL7. В некоторых вариантах осуществления, антиангиогенный агент вводится одновременно с антителом анти-EGFL7.
В некоторых вариантах осуществления, настоящее изобретение предоставляет способ повышения эффективности антиангиогенного агента у субъекта, имеющего патологическое состояние, связанное с ангиогенезом, включающий в себя введение субъекту антитела анти-EGFL7 по настоящему изобретению или фармацевтической композиции, содержащей антитело анти-EGFL7 по настоящему изобретению. В некоторых вариантах осуществления, патологическое состояние представляет собой неоплазму, например карциному. В некоторых вариантах осуществления, патологическое состояние связано с глазом, например, представляет собой интраокулярное неоваскулярное заболевание. В некоторых вариантах осуществления, антиангиогенный агент вводится субъекту в дополнение к антителу анти-EGFL7 по настоящему изобретению. В некоторых вариантах осуществления, антиангиогенный агент представляет собой антагонист фактора васкулярного эндотелиального роста (VEGF), например антитело анти-VEGF (включая бевацизумаб и ранибизумаб). В некоторых вариантах осуществления, антиангиогенный агент вводится до или после введения антитела анти-EGFL7. В некоторых вариантах осуществления, антиангиогенный агент вводится одновременно с антителом анти-EGFL7. В некоторых вариантах осуществления, применяются также другие средства лечения, например, кортикостероиды или фотодинамическая терапия.
Краткое описание чертежей
Фиг. 1 показывает последовательность аминокислот вариабельного домена легкой цепи Mab 4F11 (SEQ ID NO: 1) и HuKI (SEQ ID NO: 17).
Фиг. 2 показывает последовательность аминокислот вариабельного домена тяжелой цепи Mab 4F11 (SEQ ID NO: 2) и HuIII (SEQ ID NO: 18).
Фиг. 3 показывает последовательность аминокислот вариабельного домена легкой цепи Mab 10G9 (SEQ ID NO: 3) и HuKI (SEQ ID NO: 17).
Фиг. 4 показывает последовательность аминокислот вариабельного домена тяжелой цепи Mab 10G9 (SEQ ID NO: 4) и HuIII (SEQ ID NO: 18).
Фиг. 5 иллюстрирует домены полноразмерного EGFL7 и его усеченных форм, используемых для картирования сайтов связывания антитела.
Фиг. 6 показывает объем опухоли in vivo у моделей Xenomouse, трансфицированных раком легких человека (NSCLC; HI 299), в течение курса лечения антителами анти-VEGF и анти-EGFL7 по настоящему изобретению.
Фиг. 7 показывает выживаемость у моделей Xenomouse, трансфицированных in vivo раком легких человека (NSCLC; H1299), в течение курса лечения антителами анти-VEGF и анти-EGFL7 по настоящему изобретению.
Фиг. 8 показывает объем опухоли in vivo у моделей Xenomouse, трансфицированных раком груди человека (MDA-MB231), в течение курса лечения антителами анти-VEGF и анти-EGFL7 по настоящему изобретению.
Фиг. 9 показывает объем опухоли in vivo у моделей Xenomouse, трансфицированных раком груди человека (MDA-MB231), в течение курса лечения антителом анти-VEGF и анти-EGFL7 Mab 18F7 по настоящему изобретению.
Подробное описание предпочтительных вариантов осуществления
Настоящее изобретение предоставляет антитела анти-EGFL7, которые являются пригодными для использования, например, для лечения или предотвращения болезненных состояний, связанных с экспрессированием и/или активностью EGFL7, например, с повышенным экспрессированием и/или активностью или нежелательным экспрессированием и/или активностью. В некоторых вариантах осуществления, антитела по настоящему изобретению используются для лечения опухоли, рака и/или пролиферативного расстройства клеток.
В другом аспекте, антитела анти-EGFL7 по настоящему изобретению являются пригодными для использования в качестве реагентов для детектирования и/или выделения EGFL7, например для детектирования EGFL7 в различных тканях и типах клеток.
Настоящее изобретение дополнительно предоставляет способы получения антител анти-EGFL7, полинуклеотидов, кодирующих антитела анти-EGFL7, и клеток, содержащих полинуклеотиды, кодирующие антитела анти-EGFL7.
Общие технологии
Технологии и процедуры, описываемые или упоминаемые здесь, как правило, хорошо понятны и повсеместно применяются с использованием обычной методологии специалистами в данной области, такие, например, как широко используемые методологии, описанные в Sambrook et al., Molecular Cloning: A Laboratory Manual 3rd. edition (2001) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. CURRENT PROTOCOLS IN MOLECULAR BIOLOGY (F.M. Ausubel, et al. eds., (2003)); в серии METHODS IN ENZYMOLOGY (Academic Press, Inc.): PCR 2: A PRACTICAL APPROACH (M.J. MacPherson, B.D. Hames and G.R. Taylor eds. (1995)), Harlow and Lane, eds. (1988) ANTIBODIES, A LABORATORY MANUAL., and ANIMAL CELL CULTURE (R.I. Freshney, ed. (1987)).
Определения
"Выделенное" антитело представляет собой такое, которое идентифицируется и выделяется и/или извлекается из компонента его природно-окружающей среды. Загрязняющие компоненты его природно-окружающей среды представляют собой материалы, которые могут влиять отрицательно на диагностические или терапевтические применения антитела, и могут включать в себя ферменты, гормоны и другие белковые или небелковые растворимые вещества. В некоторых вариантах осуществления, антитело будет очищаться (1) до получения более чем 95 мас.% антитела, как определяется посредством метода Лаури, а иногда, до более чем 99 мас.%, (2) до уровня, достаточного для получения, по меньшей мере, 15 остатков N-концевой или внутренней последовательности аминокислот посредством использования секвенатора с вращающейся чашкой, или (3) до гомогенности в соответствии с анализом SDS-PAGE при восстанавливающих или невосстанавливающих условиях, с использованием окрашивания с помощью кумаси голубого или серебряного окрашивания. Выделенное антитело включает в себя антитело in situ в рекомбинантных клетках, поскольку, по меньшей мере, один компонент природной окружающей среды антитела не будет присутствовать. Обычно, однако, выделенное антитело будет приготавливаться посредством, по меньшей мере, одной стадии очистки.
"Выделенная" молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, которая идентифицируется и отделяется, по меньшей мере, от одной загрязняющей молекулы нуклеиновой кислоты, с которой она обычно связывается в природном источнике нуклеиновых кислот антитела. Выделенная молекула нуклеиновой кислоты находится в другой форме или имеет параметры иные, чем те, с которыми она находится в природе. По этой причине выделенные молекулы нуклеиновых кислот отличаются от молекулы нуклеиновой кислоты, как она существует в природных клетках. Однако выделенная молекула нуклеиновой кислоты включает в себя молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно экспрессируют антитело, где, например, молекула нуклеиновой кислоты находится в хромосомальном положении, отличном от природных клеток.
Термины "остаток вариабельного домена, пронумерованный согласно Kabat" или "положение аминокислоты, пронумерованное согласно Kabat" и их вариации относятся к системе нумерации, используемой для вариабельных доменов тяжелых цепей или вариабельных доменов легких цепей компиляции антител в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991). При использовании этой системы нумерации, реальная линейная последовательность аминокислот может содержать меньшее количество аминокислот или дополнительные аминокислоты, соответствующие сокращению или инсерции в FR или CDR вариабельного домена. Например, вариабельный домен тяжелой цепи может содержать одну вставку аминокислоты (остаток 52a в соответствии с Kabat) после остатка 52 H2 и вставленные остатки (например, остатки 82a, 82b, и 82c и тому подобное, в соответствии с Kabat) после остатка 82 FR тяжелой цепи. Нумерация остатков согласно Kabat может определяться для данного антитела посредством совмещения в областях гомологии последовательности антитела со "стандартной" последовательностью, пронумерованной по Kabat.
Фраза "по существу сходные" или "по существу одинаковые", как здесь используется, означает достаточно высокой уровень сходства между двумя численными значениями (как правило, одно из них связано с антителом по настоящему изобретению, а другое связано с эталонным/сравнительным антителом) так что специалист в данной области посчитал бы различие между двумя значениями малым или не имеющим биологической и/или статистической значимости в контексте биологической характеристики, измеряемой с помощью указанных значений (например, значений Kd). Разность между указанными двумя значениями, как правило, является меньшей, примерно, чем 50%, примерно 40%, примерно 30%, примерно 20% или примерно 10%, как функция значения для эталонного/сравнительного антитела.
"Сродство связывания", как правило, относится к прочности общей суммы нековалентных взаимодействий между отдельным сайтом связывания молекулы (например, антитела) и ее партнером по связыванию (например, антигеном). Если не указывается иного, как здесь используется, "сродство связывания" относится к собственному сродству связывания, которое отражает взаимодействие 1:1 между элементами связывающейся пары (например, антитела и антигена). Сродство молекулы X к ее партнеру Y может, как правило, быть представлено посредством константы диссоциации (Kd). Сродство может измеряться посредством обычных способов, известных в данной области, включая те, которые описываются здесь. Антитела с малым сродством, как правило, связывают антиген медленно и имеют тенденцию к легкой диссоциации, в то время антитела с высоким сродством, как правило, связывают антиген быстрее и имеют тенденцию к сохранению связи в течение более продолжительного времени. Разнообразные способы измерения сродства связывания известны в данной области, и любой из них может использоваться для целей настоящего изобретения. Конкретные иллюстративные варианты осуществления описываются далее.
В одном из вариантов осуществления, "Kd" или "значение Kd" в соответствии с настоящим изобретением измеряется посредством анализа связывания радиоактивно меченого антигена (RIA), осуществляемого с помощью версии Fab антитела, представляющего интерес, и его антигена, как описывается с помощью следующего анализа, который измеряет сродство связывания в растворе Fab к антигену посредством уравновешивания Fab с минимальной концентрацией (125I)-меченого антигена в присутствии ряда титрований немеченого антигена, а затем захвата связанного антигена с помощью планшета, покрытого антителом анти-Fab (Chen, et al., J. Mol. Biol. 293:865-81 (1999)). Для установления условий для анализа, планшеты для микротитрования (Dynex) покрываются в течение ночи 5 мкг/мл захватывающего антитела анти-Fab (Cappel Labs) в 50 мМ растворе карбоната натрия (pH 9,6), а затем блокируются с помощью 2% (масс/объем) раствора сывороточного бычьего альбумина в PBS в течение от двух до пяти часов при комнатной температуре (приблизительно 23°C). В планшете без адсорбента (Nunc #269620), 100 пМ или 26 пМ [I25I]-антигена смешивают с последовательными разбавлениями Fab, представляющего интерес (например, в соответствии с оценками антитела анти-VEGF, Fab-12, у Presta et al., Cancer Res. 57:4593-99 (1997)). Затем Fab, представляющие интерес, инкубируют в течение ночи; однако инкубирование может продолжаться в течение более длительного периода (например, 65 часов), чтобы обеспечить достижение равновесия. После этого смеси переносят на планшет для захвата, для инкубирования при комнатной температуре (например, в течение одного часа). Затем раствор удаляют, и планшет промывают восемь раз 0,1% раствором Tween-20 в PBS. Затем планшеты сушат, добавляют 150 мкл/лунка сцинтиллянта (MicroScint-20; Packard) и планшеты считают на гамма-счетчике Topcount (Packard) в течение десяти минут. Концентрации каждого из Fab, которые дают 20% или меньше от максимального связывания, выбирают для использования в анализах конкурентного связывания. В соответствии с другим вариантом осуществления Kd или значение Kd измеряют посредством анализов с использованием поверхностного плазмонного резонанса, с использованием BIAcoreTM-2000 или BIAcoreTM-3000 (BIAcore, Inc., Piscataway, NJ) при 25°C, с помощью чипов с иммобилизованным антигеном CM5, при ~10 единицах отклика (RU). Вкратце, биосенсорные чипы с карбоксиметилированным декстраном (CM5, BIAcore Inc.) активируют с помощью N-этил-N'-(3-диметиламинопропил)карбодиимида гидрохлорида (EDC) и N-гидроксисукцинимида (NHS) в соответствии с инструкциями производителя. Антиген разбавляют в 10 мМ раствора ацетата натрия, pH 4,8, как 5 мкг/мл (~0,2 мкМ) перед инжекцией при скорости потока 5 мкл/минута, для достижения приблизительно 10 единиц отклика (RU) связанного белка. После инжекции антигена, инжектируют 1M этаноламина для блокировки непрореагировавших групп. Для измерений кинетики, двукратное последовательное разбавление Fab (от 0,78 нМ до 500 нМ) инжектируют в PBS с 0,05% Tween 20 (PBST) при 25°C, при скорости потока приблизительно 25 мкл/мин. Скорости ассоциации (kon) и скорости диссоциации (koff) вычисляют с использованием простой модели связывания Лангмюра один к одному (BIAcore Evaluation Software version 3.2) посредством одновременной подгонки сенсограммы ассоциации и диссоциации. Равновесную константу диссоциации (Kd) вычисляют как отношение koff/kon. См., например, Chen, Y., et al., J. Mol. Biol. 293:865-881 (1999). Если скорость ассоциации превышает 106 M-1 сек-1, согласно анализу с использованием поверхностного плазмонного резонанса, описанного выше, тогда скорость ассоциации может определяться посредством использования методики гашения флюоресценции, где измеряют увеличение или уменьшение интенсивности испускания флюоресценции (возбуждение=295 нм; испускание=340 нм, полосовой фильтр, 16 нм) при 25°C для 20 нМ антитела анти-антиген (форма Fab) в PBS, pH 7,2, в присутствии увеличивающихся концентраций антигена, что измеряют на спектрометре, таком как спектрофотометр, с остановкой потока (Aviv Instruments) или спектрофотометр 8000-series SLM-Aminco (ThermoSpectronic) с красной кюветой с перемешиванием.
"Скорость прямой реакции" или "скорость ассоциации" или "скорость связывания", или "kon" в соответствии с настоящим изобретением может также определяться с помощью той же технологии поверхностного плазмонного резонанса, описанной выше, с использованием BIAcoreTM-2000 или BIAcoreTM-3000 (BIAcore, Inc., Piscataway, NJ) при 25°C, с помощью чипов с иммобилизованным антигеном CM5 при ~10 единиц оклика (RU). Вкратце, биосенсорные чипы с карбоксиметилированным декстраном (CM5, BIAcore Inc.) активируют с помощью N-этил-N'-(3-диметиламинопропил)карбодиимида гидрохлорида (EDC) и N-гидроксисукцинимида (NHS) в соответствии с инструкциями производителей. Антиген разбавляют 10 мМ ацетата натрия, pH 4,8, при 5 мг/мл (~0,2 мкМ), перед инжекцией при скорости потока 5 мкл/минут для достижения приблизительно 10 единиц оклика (RU) связанного белка. После инжекции антигена, инжектируют 1M этаноламина для блокировки непрореагировавших групп. Для измерений кинетики, двукратные последовательные разбавления Fab (от 0,78 нМ до 500 нМ) инжектируют в PBS с 0,05% Tween 20 (PBST) при 25°C, при скорости потока приблизительно 25 мкл/мин. Скорости ассоциации (kon) и скорости диссоциации (koff) вычисляют с использованием простой модели связывания Лангмюра один к одному (BIAcore Evaluation Software version 3.2) посредством одновременной подгонки сенсограмм ассоциации и диссоциации. Равновесная константа диссоциации (Kd) вычисляется как отношение koff/kon. См., например, Chen, Y., et al., J. Mol. Biol. 293:865-81 (1999). Однако, если скорость ассоциации превышает 106 M-1 сек-1, согласно анализу с помощью поверхностного плазмонного резонанса, описанного выше, тогда скорость ассоциации, как правило, определяется посредством использования методики гашения флюоресценции, где измеряют увеличение или уменьшение интенсивности испускаемой флюоресценции (возбуждение=295 нм; испускание=340 нм, полосовой фильтр, 16 нм) при 25°C для 20 нМ антитела анти-антиген (форма Fab) в PBS, pH 7,2, в присутствии увеличивающихся концентраций антигена, что измеряют на спектрометре, таком как спектрофотометр, с системой остановки потока (Aviv Instruments), или спектрофотометр 8000-series SLM-Aminco (ThermoSpectronic) с перемешиваемой кюветой.
Термин "вектор", как здесь используется, предназначен для упоминания молекулы нуклеиновой кислоты, способной переносить другую нуклеиновую кислоту, с которой она связана. Один из типов вектора представляет собой "плазмиду, которая относится к круговой петле двухнитевой ДНК, с которой могут лигироваться дополнительные сегменты ДНК. Другой тип вектора представляет собой фаговый вектор. Другой тип вектора представляет собой вирусный вектор, где дополнительные сегменты ДНК могут лигироваться в виде вирусного генома. Определенные векторы способны к автономной репликации в клетке-хозяине, в которую они вводятся (например, бактериальные векторы, имеющие бактериальный источник репликации, и эписомальные векторы млекопитающих). Другие векторы (например, неэписомальные векторы млекопитающих) могут встраиваться в геном клетки-хозяина при введении в клетку-хозяина, и при этом они реплицируются вместе с геномом хозяина. Кроме того, определенные векторы способны направлять экспрессию генов, с которыми они функционально связаны. Такие векторы упоминаются здесь как "рекомбинантные векторы экспрессии" (или просто, "рекомбинантные векторы"). Как правило, векторы экспрессии, используемые в технологиях рекомбинантной ДНК, часто находятся в форме плазмид. В настоящем описании, "плазмида" и "вектор" могут использоваться взаимозаменяемо, поскольку пазмида представляет собой наиболее часто используемую форму вектора.
"Полинуклеотид" или "нуклеиновая кислота", как здесь используются взаимозаменяемо, относятся к полимерам из нуклеотидов любой длины и включают в себя ДНК и РНК. Нуклеотиды могут представлять собой дезоксирибонуклеотиды, рибонуклеотиды, модифицированные нуклеотиды или основания, и/или их аналоги, или любой субстрат, который может включаться в полимер с помощью ДНК или РНК полимеразы или посредством реакции синтеза. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. Если она присутствует, модификация структуры нуклеотидов может придаваться до или после сборки полимера. Последовательность нуклеотидов может прерываться ненуклеотидными компонентами. Полинуклеотид может дополнительно модифицироваться после синтеза, например, посредством конъюгирования с меткой. Другие типы модификаций включают в себя, например, "кэппирование", замещение одного или нескольких существующих в природе нуклеотидов аналогом, внутринуклеотидные модификации, такие, например, как модификации с незаряженными связями (например, метилфосфонаты, сложные фосфотриэфиры, фосфоамидаты, карбаматы и тому подобное) и с заряженными связями (например, фосфоротиоаты, фосфородитиоаты и тому подобное), замещения, содержащие боковые остатки, такие, например, как белки (например, нуклеазы, токсины, антитела, сигнальные пептиды, поли-L-лизин и тому подобное), замещения интеркаляторами (например, акридин, псорален и тому подобное), замещения, содержащие хелатирующие агенты (например, металлы, радиоактивные металлы, бор, окислительные металлы и тому подобное), замещения, содержащие алкиляторы, замещения с модифицированными связями (например, альфа аномерные нуклеиновые кислоты и тому подобное), а также немодифицированные формы полинуклеотида (полинуклеотидов). Кроме того, любые из гидроксильных групп, обычно присутствующих в сахарах, могут заменяться, например, фосфонатными группами, фосфатными группами, защищенными с помощью стандартных защитных групп или активированными для получения дополнительных связей с дополнительными нуклеотидами, или могут конъюгироваться с твердыми или полутвердыми подложками. Группа OH на 5' и 3' концах может быть фосфорилированной или замещенной аминами или органическими остатками кэппирующих групп из 1-20 атомов углерода. Другие гидроксилы могут также дериватизовываться до стандартных защитных групп. Полинуклеотиды могут также содержать аналогичные формы рибозных или дезоксирибозных сахаров, которые являются, как правило, известными в данной области, включая, например, 2'-O-метил-, 2'-O-аллил, 2'-фтор- или 2'-азидо-рибозу, карбоциклические аналоги сахаров, альфа-аномерные сахара, эпимерные сахара, такие как арабиноза, ксилозы или ликсозы, пиранозные сахара, фуранозные сахара, седогептулозы, ациклические аналоги и аналоги с основными нуклеозидами, такие как метилрибозид. Одна или несколько связей сложных фосфодиэфиров может заменяться альтернативными связывающими группами. Эти альтернативные связывающие группы включают в себя, но, не ограничиваясь этим, варианты осуществления, где фосфат заменяется P(O)S ("тиоат"), P(S)S ("дитиоат"), (O)NR2 ("амидат"), P(O)R, P(O)OR', CO или CH2 ("формацеталь"), в котором каждый из R или R' независимо представляет собой H или замещенный или незамещенный алкил (1-20 C), необязательно содержащий эфирную (-O-) связь, арил, алкенил, циклоалкил, циклоалкенил или аральдил. Не все связи в полинуклеотиде должны быть одинаковыми. Предыдущее описание относится ко всем полинуклеотидам, упоминаемым здесь, включая РНК и ДНК.
"Олигонуклеотид", как здесь используется, как правило, относится к коротким, как правило, однонитевым, как правило, синтетическим полинуклеотидам, которые имеют, как правило, но, необязательно, менее, примерно, чем 200 нуклеотидов в длину. Термины "олигонуклеотид" и "полинуклеотид" не являются взаимно исключающими. Описание выше для полинуклеотидов является в такой же степени применимым и полностью применимым к олигонуклеотидам.
Термин "EGFL7" (взаимозаменяемо определяемый как "подобный эпедирмальному фактору роста 7"), как здесь используется, относится, если только конкретно или контекстуально, не указывается иного, к любому нативному или вариантному (либо нативному, либо синтетическому) полипептиду EGFL7, как описывается, например, в заявке WO 2005/117968, описание которой включается сюда во всей ее полноте для всех целей. Термин "нативная последовательность" конкретно охватывает существующие в природе усеченные или секретируемые формы (например, последовательность внеклеточного домена), существующие в природе вариантные формы (например, альтернативно сплайсированные формы) и существующие в природе аллельные варианты. Термин "EGFL7 дикого типа", как правило, относится к полипептиду, содержащему последовательность аминокислот существующего в природе белка EGFL7. Термин "последовательность EGFL7 дикого типа" как правило, относится к последовательности аминокислот, обнаруживаемой у существующих в природе EGFL7.
Термины "антитело" и "иммуноглобулин", используются взаимозаменяемо в самом широком смысле и включают в себя моноклональные антитела (например, полноразмерные или интактные моноклональные антитела), поликлональные антитела, многовалентные антитела, мультиспецифичные антитела (например, биспецифичные антитела, поскольку они проявляют желаемую биологическую активность) и могут также содержать определенные фрагменты антител (как здесь описывается более подробно). Антитело может быть человеческим, гуманизированным и/или представлять собой антитело с созревшим сродством.
Термин "вариабельный" относится к тому факту, что определенные части вариабельных доменов сильно различаются по последовательности для различных антител и используются при связывании и специфичности каждого конкретного антитела к его конкретному антигену. Однако вариабельность не является равномерно распространенной по вариабельным доменам антител. Она концентрируется в трех сегментах, называемых областями, определяющими комплементарность (CDR) или гипервариабельными областями, в вариабельных доменах, как легких цепей, так и тяжелых цепей. В большей степени сохраняющиеся части вариабельных доменов называются рамкой (FR). Вариабельные домены нативных тяжелых и легких цепей содержат, каждый, по четыре FR области, по большей части, принимающие β-складчатую конфигурацию, соединяемые тремя CDR, которые образуют петли, соединяющие β-складчатую структуру, а в некоторых случаях, образующие часть β-складчатой структуры. CDR в каждой цепи удерживаются вместе, в тесной близости с помощью FR областей, и вместе с CDR от другой цепи, внося вклад в образование сайта связывания антигена у антитела (см. Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, National Institute of Health, Bethesda, MD (1991)). Константные домены не вовлекаются непосредственно в связывание антитела с антигеном, но демонстрируют разнообразные эффекторные функции, такие как участие антитела в зависимой от антитела клеточной токсичности.
Папаиновый гидролиз антител дает два идентичных фрагмента связывания антигена, называемые фрагментами "Fab", каждый, с одним сайтом связывания антигена, и остаточный фрагмент "Fc", наименование которого отражает его способность к легкой кристаллизации. Обработка пепсином дает фрагмент F(ab')2, который имеет два сайта, объединяющих антигены, и является по-прежнему способным к поперечному связыванию антигенов.
"Fv" представляет собой минимальный фрагмент антитела, который содержит полный сайт распознавания и связывания антигена. В двухцепочечных частицах Fv, эта область состоит из димера вариабельных доменов одной тяжелой и одной легкой цепи в прочной, нековалентной ассоциации. В одноцепочечных частицах Fv, вариабельные домены одной тяжелой и одной легкой цепи могут ковалентно связываться с помощью гибкого пептидного линкера, так что легкая и тяжелая цепи могут ассоциироваться в "димерную" структуру, аналогичную структуре двухцепочечных частиц Fv. В этой конфигурации осуществляется то, что три CDR каждого вариабельного домена взаимодействуют с определением сайта связывания антигена на поверхности димера VH-VL. Коллективно, шесть CDR определяют специфичность антитела при связывании антигена. Однако, даже отдельный вариабельный домен (или половина Fv, содержащая только три CDR, специфичных к антигену) обладает способностью распознавать и связывать антиген, хотя и с более низким сродством, чем сайт связывания в целом.
Фрагмент Fab также содержит константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. Фрагменты Fab' отличаются от фрагментов Fab добавлением нескольких остатков на карбокси-конце домена CH1 тяжелой цепи, содержащем один или несколько цистеинов из шарнирной области антитела. Здесь Fab'-SH представляет собой обозначение для Fab', в котором цистеиновый остаток (остатки) константных доменов несет свободную тиольную группу. Фрагменты F(ab')2 антитела изначально продуцируются как пары фрагментов Fab', которые имеют шарнирные цистеины между ними. Известны также другие химические связи фрагментов антител.
"Легкие цепи" антител (иммуноглобулинов) из любых сочлененных частиц могут быть приписаны к одному из двух четко различаемых типов, называемых каппа (Κ) и лямбда (λ), на основе последовательностей аминокислот их константных доменов.
В зависимости от последовательности аминокислот константного домена их тяжелых цепей, иммуноглобулины могут быть приписаны к различным классам. Имеются пять главных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и несколько из них могут быть дополнительно разделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелых цепей, которые соответствуют различным классам иммуноглобулинов, называются α, δ, ε, γ и µ соответственно. Структуры субъединиц и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны.
"Фрагменты антитела" содержат только часть интактного антитела, где часть предпочтительно сохраняет, по меньшей мере, одну, предпочтительно, большинство или все функции, обычно связанные с этой частью, когда она присутствует в интактном антителе. Примеры фрагментов антител включают в себя фрагменты Fab, Fab', F(ab')2, и Fv; двойные антитела; линейные антитела; одноцепочечные молекулы антител и мультиспецифичные антитела, сформированные из фрагментов антител. В одном из вариантов осуществления, фрагмент антитела содержит сайт связывания антигена интактного антитела и таким образом сохраняет способность к связыванию антигена. В другом варианте осуществления, фрагмент антитела, например, такой, который содержит область Fc, сохраняет, по меньшей мере, одну из биологических функций, обычно связываемых с областью Fc, когда она присутствует в интактном антителе, такую как связывание FcRn, модулирование половинного времени жизни антитела, функция ADCC и комплементарное связывание. В одном из вариантов осуществления, фрагмент антитела представляет собой одновалентное антитело, которое имеет половинное время жизни in vivo по существу сходное с интактным антителом. Например, такой фрагмент антитела может содержать плечо для связывания антигена, связанное с последовательностью Fc, способной обеспечить стабильность фрагмента in vivo.
Термин "гипервариабельная область", "HVR" или "HV", когда здесь используются, относится к областям вариабельного домена антитела, которые являются гипервариабельными в последовательности и/или образуют структурно определенные петли. Как правило, антитела содержат шесть гипервариабельных областей; три в VH (H1, H2, H3), и три в VL (L1, L2, L3). Ряд трансдифференцировок гипервариабельных областей используется и охватывается здесь. Области, определяющие комплементарность (CDR) согласно Kabat основываются на вариабельности последовательности и используются чаще всего (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Chothia упоминает, вместо этого, положение структурных петель (Chothia & Lesk J. Mol. Biol. 196:901-17 (1987)). Гипервариабельные области AbM представляют собой некий компромисс между CDR согласно Kabat и структурными петлями согласно Chothia, и используются программным обеспечением для моделирования антител Oxford Molecular's AbM. "Контактные" гипервариабельные области основываются на анализе доступных сложных кристаллических структур.
Гипервариабельные области могут содержать "расширенные гипервариабельные области" следующим образом: 24-36 (L1), 46-56 (L2) и 89-97 (L3) в VL и 26-35 (H1), 49-65 или 50-65 (H2) и 93-102 (H3) в VH. Остатки вариабельных доменов нумеруются в соответствии с Kabat et al., выше, для каждого из этих определений.
Остатки "рамок" или "FR" представляют собой остатки вариабельных доменов, иные, чем остатки гипервариабельных областей, как здесь определяется.
"Гуманизированные" формы антител не-человека (например, мышей) представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из иммуноглобулина не-человека. По большей части, гуманизированные антитела представляют собой иммуноглобулины человека (антитела реципиента), в которых остатки из гипервариабельной области реципиента заменяются остатками из гипервариабельной области видов не-человека (донорные антитела), например, мыши, крысы, кролика или примата не-человека, имеющие желаемую специфичность, сродство и емкость. В некоторых случаях, остатки области рамки (FR) иммуноглобулина человека заменяют соответствующими остатками не-человека. Кроме того, гуманизированные антитела могут содержать остатки, которые не обнаруживаются в антителе реципиента или в антителе донора. Эти модификации делаются для дополнительного ограничения рабочих характеристик антитела. Как правило, гуманизированное антитело будет содержать по существу все, по меньшей мере, из одного, а, как правило, двух вариабельных доменов, в которых все или по существу все гипервариабельные петли соответствуют иммуноглобулину не-человека и все или по существу все FR соответствуют последовательности иммуноглобулина человека. Гуманизированное антитело необязательно будет также содержать, по меньшей мере, часть константной области иммуноглобулина (Fc), как правило, иммуноглобулина человека. Относительно дополнительных деталей см. Jones et al., Nature 321:522-25 (1986); Riechmann et al., Nature 332:323-29 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-96 (1992). См. также следующие обзорные статьи и источники, цитируемые в них: Vaswani & Hamilton, Ann. Allergy, Asthma & Immunol. 1:105-15 (1998); Harris, Biochem. Soc. Transactions 23:1035-38 (1995); Hurle & Gross, Curr. Op. Biotech. 5:428-33 (1994).
"Химерные" антитела (иммуноглобулины) имеют часть тяжелой и/или легкой цепи, идентичную или гомологичную соответствующим последовательностям в антителах, полученных от конкретных видов или принадлежащих к конкретному классу или подклассу антител, в то время как оставшаяся часть цепи (цепей) является идентичной или гомологичной соответствующим последовательностям в антителах, полученных от других видов или принадлежащих другому классу или подклассу антител, а также фрагменты таких антител, поскольку они проявляют желаемую биологическую активность (заявка на патент США № 4816567; и Morrison et al., Proc Natl. Acad. Sci. USA 81:6851-6855 (1984)). Гуманизированное антитело, как здесь используется, представляет собой подкласс химерных антител.
Фрагменты "одноцепочечных Fv" или "scFv" антител содержат домены VH и VL антитела, где эти домены присутствуют в одной полипептидной цепи. Как правило, полипептид scFv дополнительно содержит полипептидный линкер между доменами VH и VL, который позволяет scFv формировать желаемую структуру для связывания антигена. Относительно обзора по scFv см. Pluckthun, The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994).
"Антиген" представляет собой заданный антиген, с которым антитело может селективно связываться. Целевой антиген может представлять собой полипептид, углевод, нуклеиновую кислоту, липид, гаптен или другое существующее в природе или синтетическое соединение. Как правило, целевой антиген представляет собой полипептид.
"Эпитоп" представляет собой часть антигена, с которым селективно связывается антитело. Для полипептидного антигена, эпитоп, как правило, представляет собой часть пептида примерно из 4-10 аминокислот.
Термин "двойные антитела" относится к малым фрагментам антител с двумя сайтами связывания антигенов, эти фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в одну и ту же полипептидную цепь (VH-VL). Если использовать линкер, который является слишком коротким, чтобы сделать возможным спаривание между двумя доменами на одной и той же цепи, домены принуждаются к спариванию с комплементарными доменами другой цепи и образуют два сайта связывания с антигенами. Двойные антитела описываются более подробно, например, в EP 404097; в заявке WO 93/11161 и Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-48 (1993).
"Антитело человека" представляет собой такое антитело, которое имеет последовательность аминокислот, которая соответствует последовательности антитела, продуцируемого человеком, и/или приготавливается с использованием любой из технологий для получения антител человека, как здесь описывается. Это определение антитела человека конкретно исключает гуманизированное антитело, содержащее остатки связывания с антигеном не-человека.
Антитело "с cозревшим сродством" представляет собой антитело с одним или несколькими изменениями в одном или нескольких его CDR, что приводит к улучшению сродства антитела к антигену, по сравнению с исходным антителом, которое не обладает таким изменением (изменениями). Предпочтительные антитела с созревшим сродством будут иметь наномолярное или даже пикомолярное сродство к целевому антигену. Созревшие для сродства антитела получают с помощью процедур, известных в данной области. Marks et al. Bio/Technology 10:779-83 (1992) описывает созревание сродства с помощью шаффлинга VH и VL доменов. Неупорядоченный мутагенез CDR и/или рамочных остатков описывается: Barbas et al., Proc. Nat. Acad. Sci. USA 91:3809-13 (1994); Schier et al., Gene 169:147-55 (1995); Yelton et al., J. Immunol. 155:1994-2004 (1995); Jackson et al., J. Immunol. 154(7):3310-19 (1995); и Hawkins et al., J. Mol. Biol. 226:889-96 (1992).
"Эффекторные функции" антитела относятся к его биологическим активностям, приписываемым области Fc (нативной области с последовательностью Fc или области Fc с вариантом последовательности аминокислот) антитела, и они изменяеются с изотипом антитела. Примеры эффекторных функций антитела включают в себя: связывание C1q и комплементзависимую цитотоксичность; связывание рецепторов Fc; антителозависимую клеточно-обусловленную цитотоксичность (ADCC); фагоцитоз; отрицательную регуляцию поверхностных рецепторов клетки (например, рецепторов B лимфоцитов) и активирование B лимфоцитов.
"Антителозависимая клеточно-обусловленная цитотоксичность" или "ADCC" относится к форме цитотоксичности, при которой секретируемый Ig связывается с рецепторами Fc (FcR), присутствующих на определенных цитотоксичных клетках (например, на клетках природных киллеров (NK), нейтрофилах и макрофагах), они дают возможность этим цитотоксичным эффекторным клеткам специфично связываться с несущей антиген целевой клеткой, а впоследствии уничтожать целевую клетку с помощью цитотоксинов. Антитела образуют "плечо" цитотоксичных клеток и являются абсолютно необходимыми для такого уничтожения. Первичные клетки для опосредования ADCC, клетки NK, экспрессируют только FcγRIII, в то время как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессирование FcR на гематопоэтических клетках приводится в Таблице 3 на странице 464, в Ravetch & Kinet, Annu. Rev. Immunol. 9:457-92 (1991). Для оценки активности ADCC молекулы, представляющей интерес, может осуществляться анализ ADCC in vitro, такой как описывается в заявке на патент США № 5500362 или 5821337 или в заявке на патент США № 6737056, Presta. Полезные эффекторные клетки для таких анализов включают в себя мононуклеарные клетки периферической крови (PBMC) и клетки природных киллеров (NK). Альтернативно, или в дополнение к этому, активность при ADCC у молекул, представляющих интерес, может оцениваться in vivo, например, на животной модели, такой как, та, которая описывается в Clynes et al., Proc. Natl. Acad. Sci. USA 95:652-56 (1998).
"Эффекторные клетки человека" представляют собой лейкоциты, которые экспрессируют один или несколько FcR и осуществляют эффекторные функции. Предпочтительно, клетки экспрессируют, по меньшей мере, FcγRIII и осуществляют эффекторную функцию ADCC. Примеры лейкоцитов человека, которые медиируют ADCC, включают в себя мононуклеарные клетки периферической крови (PBMC), клетки природных киллеров (NK), моноциты, цитотоксические T лимфоциты и нейтрофилы; при этом PBMC и клетки NK являются предпочтительными. Эффекторные клетки могут выделяться из нативного источника, например из крови.
"Рецептор Fc" или "FcR" описывает рецептор, который связывается с областью Fc антитела. Предпочтительный FcR представляет собой FcR человека с нативной последовательностью. Кроме того, предпочтительный FcR представляет собой такой, который связывается с антителом IgG (гамма рецептором) и включает в себя рецепторы подклассов FcγRI, FcγRII и FcγRIII, включая аллельные варианты и альтернативно сплайсированные формы этих рецепторов. Рецепторы FcγRII включают в себя FcγRIIA ("активирующий рецептор") и FcγRIIB ("ингибирующий рецептор"), которые имеют сходные последовательности аминокислот, которые различаются в основном в своих цитоплазматических доменах. Активирующий рецептор FcγRIIA содержит мотив активирования иммунорецептора на основе тирозина (ITAM) в своем цитоплазматическом домене. Ингибирующий рецептор FcγRIIB содержит мотив ингибирования иммунорецептора на основе тирозина (ITIM) в своем цитоплазматическом домене, (см. обзор в M. Daeron, Annu. Rev. Immunol. 15:203-34 (1997)). FcR обсуждаются в Ravetch and Kinet, Annu. Rev. Immunol. 9:457-92 (1991); Capel et al., Immunomethods 4:25-34 (1994); и в de Haas et al., J. Lab. Clin. Med. 126:330-41 (1995). Другие FcR, включая те, которые должны быть идентифицированы в будущем, охватываются здесь термином "FcR". Термин также включает в себя неонатальный рецептор, FcRn, который является ответственным за перенос IgG от матери к зародышу (Guyer et al., J. Immunol. 117:587 (1976) и Kim et al., J. Immunol. 24:249 (1994)) и регулирует гомеостаз иммуноглобулинов.
Заявка WO 00/42072 (Presta) описывает варианты антител с улучшенным или уменьшенным связыванием FcR. Содержание этой публикации конкретно включается сюда в виде ссылки. См., также, Shields et al. J. Biol. Chem. 9(2): 6591-6604 (2001).
Способы измерения связывания с FcRn известны (см., например, Ghetie and Ward Immunol. Today 18:592-8 (1997)). Связывание с FcRn человека in vivo и половинное время жизни в сыворотке полипептидов с высоким сродством связывания с FcRn человека может анализироваться, например, на трансгенных мышах или на трансфицированных линиях клеток человека, экспрессирующих FcRn человека, или на приматах, которым вводят полипептиды вариантов Fc.
"Комплементзависимая цитотоксичность" или "CDC" относится к лизису целевой клетки в присутствии комплемента. Активирование классического пути комплемента инициируется посредством связывания первого компонента системы комплемента (C1q) с антителами (из соответствующего подкласса), которые связываются с их когнатным антигеном. Для оценки активирования комплемента, может осуществляться анализ CDC, например, как описывается в J. Immunol. Methods 202:163 (1996), Gazzano-Santoro et al.
Варианты полипептидов с измененными последовательностями аминокислот области Fc и увеличенной или уменьшенной способностью к связыванию C1q описываются в заявке на патент США № 6194551B1 и в заявке WO 99/51642. Содержание этих патентных публикаций конкретно включается сюда в виде ссылок. См., также, Idusogie et al., J. Immunol. 164: 4178-84 (2000).
"Блокирующее" антитело или антитело "антагонист" представляет собой такое антитело, которое ингибирует или уменьшает биологическую активность антигена, с которым оно связывается. Предпочтительные блокирующие антитела или антитела антагонисты по существу или полностью ингибируют биологическую активность антигена.
"Расстройство" или "заболевание" представляют собой любое состояние, которое может улучшаться от лечения с помощью вещества/молекулы или способа по настоящему изобретению. Оно включает в себя хронические и острые расстройства или заболевания, включая в себя такие патологические состояния, которые предрасполагают млекопитающее к рассматриваемому расстройству. Неограничивающие примеры расстройств, которые могут лечиться здесь, включают в себя злокачественные и доброкачественные опухоли; карциному, бластому и саркому.
Термины "клеточное пролиферативное расстройство" и "пролиферативное расстройство" относятся к расстройствам, которые связаны с некоторым уровнем аномальной пролиферации клеток. В одном из вариантов осуществления, клеточное пролиферативное расстройство представляет собой рак.
"Опухоль", как здесь используется, относится к любому росту клеток новообразования и их пролиферации, злокачественной или доброкачественной, и ко всем предраковым и раковым клеткам и тканям. Термины "рак", "раковый", "клеточное пролиферативное расстройство", "пролиферативное расстройство" и "опухоль" не являются взаимоисключающими при упоминании здесь.
Термины "рак" и "раковый" упоминают или описывают физиологическое состояние у млекопитающих, которые, как правило, отличается нерегулируемым ростом/пролиферацией клеток. Примеры рака включают в себя, но, не ограничиваясь этим, карциному, лимфому, бластому, саркому и лейкемию. Более конкретные примеры таких раковых заболеваний включают в себя сквамозно-клеточный рак, мелкоклеточный рак легких, рак гипофиза, рак пищевода, астроцитому, саркому мягких тканей, немелкоклеточный рак легких, аденокарциному легких, сквамозную карциному легких, рак брюшной полости, рак клеток печени, рак желудочно-кишечных органов, рак поджелудочной железы, глиобластому, рак шейки матки, рак яичников, рак печени, рак мочевого пузыря, гепатому, рак груди, рак толстой кишки, рак толстой и прямой кишки, эндометриальную карциному или карциному матки, карциному слюнных желез, рак почек, рак печени, рак простаты, рак женских половых органов, рак щитовидной железы, карциному печени, рак мозга, эндометриальный рак, рак яичка, холангиокарциному, карциному желчного пузыря, рак желудка, меланому и различные типы рака головы и шеи. Дизрегуляция ангиогенеза может привести к тому, что многие расстройства могут лечиться с помощью композиций и способов по настоящему изобретению. Эти расстройства включают в себя состояния, как не связанные с новообразованиями, так и связанные с ними. Связанные с новообразованиями состояния включают в себя, но, не ограничиваясь этим, те, которые описываются выше. Не связанные с новообразованиями расстройства включают в себя, но, не ограничиваясь этим, нежелательную или аберрантную гипертрофию, артрит, ревматоидный артрит (RA), псориаз, псориатические бляшки, саркоидоз, атеросклероз, атеросклеротические бляшки, диабетические и другие пролиферативные ретинопатии, включая преждевременную ретинопатию, ретролентальную фиброплазию, неоваскулярную глаукому, возрастную макулярную дегенерацию, диабетический макулярный отек, корнеальную неоваскуляризацию, неоваскуляризацию корнеального трансплантата, отторжение корнеального трансплантата, ретинальную/хороидальную неоваскуляризацию, неоваскуляризацию радужно-роговичного угла (рубеоз), окулярное неоваскулярное заболевание, сосудистый рестиноз, артериовенозные мальформации (AVM), менингиому, гемангиому, ангиофиброму, гиперплазию щитовидной железы (включая болезнь Граве), трансплантацию корнеальных и других тканей, хроническое воспаление, воспаление легких, острое повреждение легких/ARDS, сепсис, первичную пульмонарную гипертонию, злокачественные пульмонарные эффузии, отек мозга (например, связанный с острым инсультом/закрытым повреждением головы/травмой), синовиальное воспаление, образование паннуса при RA, оссификантный миозит, гипертрофическое образование костей, остеоартрит (OA), резистентые асциты, полицистозное заболевание яичников, эндометриоз, заболевания, ассоциированные с проникновением жидкостей в объемы (панкреатит, компартмент-синдром, ожоги, заболевание желудка), маточные фиброиды, преждевременные роды, хроническое воспаление, такое как IBD (болезнь Крона и язвенный колит), отторжение пересаженной почки, воспалительное заболевание желудка, нефротический синдром, нежелательный или аберрантный рост массы тканей (не раковый), гемофильные суставы, гипертрофированные рубцы, замедление роста волос, синдром Ослера-Вебера, пиогенную гранулему, ретролентальные фиброплазии, склеродерму, трахому, сосудистые адгезии, синовит, дерматит, преэклампсию, асциты, перикардиальную эффузию (такую как связана с перикардитом) и плевральную эффузию.
Термин "истощающее" расстройство (например, истощающий синдром, кахеция, саркопения) относится к расстройству, вызывающему нежелательную и/или нездоровую потерю массы или потерю массы клеток тела. У пожилых людей, а также при СПИДе и у больных раком, истощающее заболевание может приводить к нежелательной потери массы тела, включая как жировые, так и не содержащие жира отделы. Истощающие заболевания могут являться результатом неадекватного приема пищи и/или метаболических изменений, связанных с болезненными и/или возрастными процессами. Больные раком и больные СПИДом, а также пациенты после обширного хирургического вмешательства или имеющие хронические инфекции, иммунологические заболевания, гипертироидизм, болезнь Крона, психогенное заболевание, хроническую сердечную недостаточность или какую-либо серьезную травму, часто страдают от истощающего заболевания, которое иногда также упоминается как кахеция, метаболическое, а иногда, связанное с приемом пищи, расстройство. Кахеция дополнительно отличается гиперметаболизмом и гиперкатаболизмом. Хотя термины кахеция и истощающее заболевание часто используются взаимозаменяемо для упоминания состояний истощения, имеется, по меньшей мере, один набор исследований, которые отличают кахецию от синдрома истощения как потери жировой массы, в частности массы клеток тела (Mayer, J. Nutr. 129 (1S Suppl.):256S-59S (1999)). Саркопения, еще одно такое расстройство, которое может влиять на пожилых индивидуумов, как правило, характеризуется потерей мышечной массы. Истощающее заболевание в конечной стадии, как описывается выше, может развиться у индивидуумов, страдающих либо от кахеции, либо от саркопении.
Как здесь используется, "лечение" относится к клиническому вмешательству в попытке изменить естественный ход событий для индивидуума или клетки, которых лечат, и может осуществляться либо для профилактики, либо в ходе клинической патологии. Желаемые эффекты лечения включают в себя предотвращение появления или повторного появления заболевания, ослабление симптомов, уменьшение любых прямых или косвенных патологических последствий заболевания, уменьшение скорости развития заболевания, ослабление или улучшение состояния заболевания и ремиссию или улучшение прогноза. В некоторых вариантах осуществления, антитела по настоящему изобретению используются для продления развития заболевания или расстройства.
"Индивидуум", "субъект" или "пациент" представляет собой позвоночное, например млекопитающее, включая в частности человека. Млекопитающие включают в себя, но, не ограничиваясь этим, людей, домашних и сельскохозяйственных животных, животных из зоопарков, спортивных учреждений, комнатных животных, таких как собаки, лошади, кошки, коровы и тому подобное.
"Эффективное количество" относится к количеству, эффективному при таких дозировках и в течение таких периодов времени, которые необходимы для достижения желаемого терапевтического или профилактического результата.
"Терапевтически эффективное количество" вещества/молекул по настоящему изобретению, агониста или антагониста, может изменяться в соответствии с такими факторами, как состояние заболевания, возраст, пол и масса индивидуума, способность вещества/молекулы, агониста или антагониста, вызывать желаемую реакцию у индивидуума. Терапевтически эффективное количество также представляет собой такое, при котором любые токсические или вредные воздействия вещества/молекулы, агониста или антагониста, перевешиваются терапевтически полезными воздействиями.
"Профилактически эффективное количество" относится к количеству, эффективному при таких дозировках и в течение таких периодов времени, которые необходимы для достижения желаемого профилактического результата. Когда профилактическая доза используется для субъектов до заболевания или на его ранней стадии, профилактически эффективное количество, как правило, но необязательно, будет меньшим, чем терапевтически эффективное количество.
Термин "цитотоксический агент", как здесь используется, относится к веществу, которое ингибирует или предотвращает функционирование клеток и/или вызывает разрушение клеток. Термин предназначен для включения в него радиоактивных изотопов (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32 и радиоактивных изотопов Lu), химиотерапевтических агентов, например метотрексата, адриамицина, алкалоидов винка (винкристина, винбластина, этопозида), доксорубицина, мелфалана, митомицина C, хлорамбуцила, даунорубицина или других интеркалирующих агентов, ферментов и их фрагментов, таких как нуклеотические ферменты, антибиотики и токсины, такие как низкомолекулярные токсины или ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения, включая их фрагменты и/или варианты, и разнообразные противоопухолевые или противораковые агенты, описанные ниже. Другие цитотоксические агенты описываются ниже. Туморицидный агент вызывает разрушение клеток опухоли.
"Химиотерапевтический агент" представляет собой химическое соединение, пригодное для использования при лечении рака. Примеры химиотерапевтических агентов включают в себя алкилирующие агенты, такие как тиотепа и циклофосфамид CYTOXAN®; алкилсульфонаты, такие как бисульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбохон, метуредопа и уредопа; этиленамины и метиламеламины, включающие в себя альтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилоломеламин; ацетогенины (в частности, буллатацин и буллатацинон); дельта-9-тетрагидроканнабинол (дронабинол, MARINOL®); бета-лапахон; лапахол; колхицины; бетулиновую кислоту; камптотецин (включающий в себя синтетический аналог топотекана (HYCAMTIN®), CPT-11 (иринотекан, CAMPTOSAR®), ацетилкаптотецин, скополектин и 9-аминокаптотецин); бриостатин; каллистатин; CC-1065 (включающий в себя его синтетические аналоги адозелезин, карзелезин и безелезин); подофиллотоксин; подофиллиновую кислоту; тенипозид; криптофицины (в частности, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включающий в себя синтетические аналоги, KW-2189 и CB1-TM1); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; азотные иприты, такие как хлорамбуцил, хлорнафазин, холофосфамид, эстрамустин, ифостамид, мехлорэтамин, мехлорэтамин оксид гидрохлорид, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урациловый иприт; нитрозоамочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин и ранимнустин; антибиотики, такие как энедииновые антибиотики (например, калихеамицин, в частности калихеамицин гамма 1I и калихеамицин омега I1 (см., например, Agnew, Chem Intl. Ed. Engl. 33: 183-186 (1994)); динемицин, включающий в себя динемицин A; эсперамицин; а также неокарзиностатиновый хромофор и родственные хромопротеиновые хромофоры на основе антибиотика энедиина), аклацитомизины, актиномицин, атрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карзинофилин, хромомицин, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, ADRIAMYCIN® доксорубицин (включающий в себя морфолино-доксорубицин, цианоморфолино-доксорубицин, 2-пирролино-доксорубицин и деоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин C, микофенольную кислоту, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пурина, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидина, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидеоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, дромостанолон пропионат, эпитиостанол, мепитиостан, тестолактон; антиадренальные препараты, такие как аминоглютетимид, митота, трилостан; пополнитель фолиевой кислоты, такие как фролиновая кислота; ацеглатон; альдофосфамид гликозид; аминолевулиновую кислоту; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатраксат; дефофамин; демеколцин; диазихон; элфорнитин; эллиптиний ацетат; эпотилон; этоглюцид; нитрат галлия; гидроксимочевину; лентинан; лонидаинин; майтанзиноиды, такие как майтансин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитраерин; пентостатин; фенамет; пирарубицин; лозоксантрон; 2-этилгидразид; прокарбазин; комплекс полисахаридов PSK® (JHS Natural Products, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновую кислоту; триазихон; 2,2',2"-трихлортриэтиламин; трихотены (в частности, T-2 токсин, веррацурин A, роридин A и ангуидин); уретан; виндезин (ELDISINE®, FILDESIN®); дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид ("Ara-C"); тиотепа; таксоиды, например, TAXOL® паклитаксел (Bristol-Myers Squibb Oncology, Princeton, N.J.), Не содержащий Cremophor от ABRAXANETM, препарат паклитаксела на основе наночастиц альбумина (American Pharmaceutical Partners, Schaumberg, Illinois) и доксетаксел TAXOTERE® (Rhone-Poulenc Rorer, Antony, France); хлоранбуцил; гемцитабин (GEMZAR®); 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин (VELBAN®); платину; этопозид (VP-16); ифосфамид; митоксантрон; винкристин (ONCOVIN®); оксалиплатин; лейкововин; винорельбин (NAVELBINE®); новантрон; эдатрексат; дауномицин; аминоптерин; ибандронат; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин (XELODA®); фармацевтически приемлемые соли, кислоты или производные любого из веществ, указанных выше; а также сочетания двух или более из веществ, указанных выше, такие как CHOP, аббревиатура для сочетанной терапии из циклофосфамида, доксорубицина, винкристина и преднизолона, и FOLFOX, аббревиатура для режима лечения с помощью оксалиплатина (ELOXATINTM), сочетанного с 5-FU и лекововином.
Также включенными в это определение являются антигормональные агенты, которые действуют, регулируя, уменьшая, блокируя или ингибируя действия гормонов, которые могут способствовать росту рака, и часто присутствуют в форме системного или предназначенного для организма в целом лечения. Они могут представлять собой гормоны сами по себе. Примеры включают в себя антиэстрогены и селективные модуляторы рецепторов эстрогенов (SERMs), включающие в себя, например, тамоксифен (включающий в себя тамоксифен NOLVADEX®), ралоксифен EVISTA®, дролоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и торемифен FARESTON®; антипрогестероны; отрицательные регуляторы рецепторов эстрогенов (ERD); агенты, которые функционируют, подавляя или выключая яичники, например, агонисты гормона, лютеинизирующего релизинг-гормона (LHRH), такие как LUPRON® и лейпролид ацетат ELIGARD®, гозерелин ацетат, бузерелин ацетат и триптерелин; другие антиандрогены, такие как флютамид, нилютамид и бикалютамид; и ингибиторы ароматазы, которые ингибируют фермент ароматазу, который регулирует продуцирование эстрогенов в надпочечниках, такие как, например, 4(5)-имидазолы, аминоглютетимид, мегестроол ацетат MEGASE®, эксеместан AROMASIN®, форместани, фадрозол, ворозол RIVISOR®, летрозол FEMARA® и анастрозол ARIMIDEX®. В дополнение к этому, такое определение химиотерапевтических агентов включает в себя бисфосфонаты, такие как клодронат (например, BONEFOS® или OSTAC®), этидронат DIDROCAL®, NE-58095, золендроновую кислоту/золедронат ZOMETA®, алендронат FOSAMAX®, памидронат AREDIA®, тилудронат SKELID® или ризедронат ACTONEL®; а также троксацитабин (аналог 1,3-диоксолан-нуклеозида цитозина); антисмысловые олигонуклеотиды, в частности такие, которые ингибируют экспрессию генов в путях передачи сигналов, участвующих в пролиферации аберрантных клеток, такие, например, как PKC-альфа, Raf, H-Ras и рецептор эпидермального фактора роста (EGF-R); вакцины, такие как вакцина THERATOPE® и вакцины для генной терапии, например вакцина ALLOVECTIN®, вакцина LEUVECTIN® и вакцина VAXID®; ингибитор топоизомеразы 1 LURTOTECAN®; ABARELIX® rmRH; лапатиниб дитосилат (ErbB-2 и низкомолекулярный двойной ингибитор тирозинкиназы EGFR, также известный как GW572016); и фармацевтически приемлемые соли, кислоты или производные любого соединения, указанного выше.
"Агент, ингибирующий рост", когда здесь используется, относится к соединению или композиции, которая ингибирует рост клеток (таких как клетки, экспрессирующие EGFL7) либо in vitro, либо in vivo. Таким образом, агент, ингибирующий рост, может представлять собой ингибитор, который значительно понижает процент клеток (таких как клетки, экспрессирующие EGFL7) в S-фазе. Примеры агентов, ингибирующих рост, включают в себя агенты, которые блокируют развитие клеточного цикла (в месте, ином, чем S-фаза), такие как агенты, которые индуцируют приостановку G1 и приостановку M-фазы. Классические блокаторы M-фазы включают в себя алколоиды барвинка (винкристин и винбластин), таксаны и ингибиторы тоизомеразы II, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Те агенты, которые приостанавливают G1, также вызывают попутно приостановку в S-фазе, например, агенты, алкилирующие ДНК, такие как тамоксифен, преднизон, дакарбазин, мехлорэтамин, цисплатин, метотрексат, 5-фторурацил и ara-C. Кроме того, информацию можно найти в The Molecular Basis of Cancer, Mendelsohn and Israel, eds., Chapter 1, entitled "Cell cycle regulation, oncogenes, and antineoplastic drugs" by Murakami el al. (WB Saunders: Philadelphia, 1995), в частности, на p. 13. Таксаны (паклитаксел и доцетаксел) представляют собой противораковые лекарственные средства, оба они получены из тиссового дерева. Доцетаксел (TAXOTERE®, Rhone-Poulenc Rorer), полученный из европейского тисса, является полусинтетическим аналогом паклитаксела (TAXOL®, Bristol-Myers Squibb). Паклитаксел и доцетаксел способствуют сборке микротрубочек из тубулиновых димеров и стабилизируют микротрубочки посредством предотвращения деполимеризации, которая приводит к ингибированию митоза в клетках.
"Доксорубицин" представляет собой антрациклиновый антибиотик. Полное химическое наименование доксорубицина представляет собой (8S-цис)-10-[(3-амино-2,3,6-тридеокси-α-L-ликсогексапиранозил)окси]-7,8,9,10-тетрагидро-6,8,11-тригидрокси-8-(гидроксиацетил)-1-метокси-5,12-нафтацендион.
Термин "область полипептида, содержащая Fc" относится к полипептиду, такому как антитело или иммуноадгезин (см. определения здесь), который содержит область Fc. C-концевой лизин (остаток 447 в соответствии с системой нумерации ЕС) области Fc может быть удален, например, во время очистки полипептида или посредством рекомбинантного получения с помощью генной инженерии нуклеиновой кислоты, кодирующей полипептид. Соответственно, композиция, содержащая полипептид, имеющая область Fc в соответствии с настоящим изобретением, может содержать полипептиды с K447, где все K447 удаляются, или смесь полипептидов с остатком K447 и без него.
Композиции по настоящему изобретению и способы их получения
Настоящее изобретение охватывает композиции, включающие в себя фармацевтические композиции, содержащие антитело анти-EGFL7; и полинуклеотиды, содержащие последовательности, кодирующие антитело анти-EGFL7. Как здесь используется, композиции содержат одно или несколько антител, которые связываются с EGFL7, и/или один или несколько полинуклеотидов, содержащих последовательности, кодирующие одно или несколько антител, которые связываются с EGFL7. Эти композиции могут дополнительно содержать соответствующие носители, такие как фармацевтически приемлемые эксципиенты, включающие в себя буферы, которые хорошо известны в данной области.
Настоящее изобретение также охватывает варианты осуществления выделенного антитела и полинуклеотида. Настоящее изобретение также охватывает по существу чистое антитело и варианты осуществления полинуклеотидов.
Антитела анти-EGFL7 по настоящему изобретению являются предпочтительно моноклональными. Также охваченными в рамках настоящего изобретения являются фрагменты Fab, Fab', Fab'-SH и F(ab')2 антител анти-EGFL7, представленные здесь. Эти фрагменты антитела могут создаваться с помощью традиционных средств, таких как ферментативный гидролиз, или могут генерироваться с помощью рекомбинантных технологий. Такие фрагменты антител могут быть химерными или гуманизированными. Эти фрагменты являются пригодными для диагностических и терапевтических целей, приведенных ниже.
Моноклональные антитела получают из популяций по существу гомогенных антител, то есть индивидуальные антитела, составляющие популяцию, являются идентичными за исключением возможных существующих в природе мутаций, которые могут присутствовать в небольших количествах. Таким образом, прилагательное "моноклональный" указывает на характер антитела, которое не является смесью дискретных антител.
Моноклональные антитела анти-EGFL7 по настоящему изобретению могут быть получены с использованием гибридомного способа, впервые описанного Kohler et al., Nature 256:495 (1975), или могут быть получены с помощью способов рекомбинантной ДНК (заявка на патент США № 4816567).
В гибридомном способе, мышь или другое соответствующее животное хозяин, такое как хомячок, иммунизируется для получения лимфоцитов, которые продуцируют или могут продуцировать антитела, которые будут специфично связываться с белком, используемым для иммунизации. Антитела к EGFL7, как правило, возникают у животных под действием множества подкожных (sc) или внутрибрюшинных (ip) инъекций EGFL7 и адъюванта. EGFL7 могут быть получены с использованием способов, хорошо известных в данной области, некоторые из них дополнительно описываются здесь. Например, рекомбинантное продуцирование EGFL7 описывается ниже. В одном из вариантов осуществления, животные иммунизируются с помощью производного EGFL7, которое содержит внеклеточный домен (ECD) EGFL7, слитый с частью Fc тяжелой цепи иммуноглобулина. В одном из вариантов осуществления, животные иммунизируются с помощью белка слияния EGFL7-IgGl. Животные обычно иммунизируются против иммуногенных конъюгатов или производных EGFL7 с помощью монофосфориллипида A (MPL)/трегалоза дикриномиколята (TDM) (Ribi Immunochem. Research, Inc., Hamilton, MT), и раствор употребляют для интрадермальной инъекции во множестве мест. Через две недели животные подвергаются повторной стимуляции. Через 7-14 дней у животных отбирают кровь и анализируют сыворотку на титры анти-EGFL7. Животных подвергают повторной стимуляции до достижения плато титрования.
Альтернативно, лимфоциты могут иммунизироваться in vitro. Лимфоциты затем сливают с клетками миеломы с использованием соответствующего агента для слияния, такого как полиэтиленгликоль, с формированием гибридомных клеток (Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103 (Academic Press, 1986)).
Гибридомные клетки, полученные таким образом, высеваются и выращиваются в соответствующей культурной среде, которая предпочтительно содержит одно или несколько веществ, которые ингибируют рост или выживаемость не слитых, исходных клеток миеломы. Например, если в исходных клетках миеломы отсутствует фермент гипоксантин гуанин фосфорибозил трансфераза (HGPRT или HPRT), культурная среда для гибридом, как правило, будет содержать гипоксантин, аминоптерин и тимидин (среда HAT), эти вещества предотвращают рост HGPRT-дефицитных клеток.
Предпочтительные клетки миеломы представляют собой такие клетки, которые эффективно сливаются, поддерживают на стабильно высоком уровне продуцирование антитела выбранными антителопродуцирующими клетками и являются чувствительными к среде, такой как среда HAT. Среди них, предпочтительные линии клеток миеломы представляют собой линии миеломы мышей, таких как те, которые получены от опухолей мышей MOPC-21 и MPC-11, доступных от Salk Institute Cell Distribution Center, San Diego, California USA, и клетки SP-2 или X63-Ag8-653, доступные от American Type Culture Collection, Rockville, Maryland USA. Линии клеток миеломы человека и гетеромиеломы мыши-человека также описываются относительно продуцирования моноклональных антител человека (Kozbor, J. Immunol., 133:3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987)).
Культурная среда, в которой выращивают клетки гибридомы, оценивается на продуцирование моноклональных антител, направленных против EGFL7. Предпочтительно, специфичность связывания моноклональных антител, продуцируемых клетками гибридомы, определяется посредством иммунной преципитации или посредством анализа связывания in vitro, такого как радиоиммунологический анализ (RIA) или иммуноадсорбционный анализ на связанном ферменте (ELISA).
Сродство к связыванию моноклонального антитела может определяться, например, посредством анализа Скатчарда, Munson et al., Anal. Biochem. 107:220 (1980).
После идентификации гибридомных клеток, которые продуцируют антитела с желаемой специфичностью, сродством и/или активностью, клоны могут субклонироваться посредством процедур серийного разведения и выращиваться с помощью стандартных способов (Goding, Monoclonal Antibodies: Principles and Practice, pp. 59-103 (Academic Press, 1986)). Соответствующие культурные среды для этой цели включают в себя, например, среду D-MEM или RPMI-1640. В дополнение к этому, гибридомные клетки могут выращиваться in vivo как асцитные опухоли у животных.
Моноклональные антитела, секретируемые субклонами, соответствующим образом выделяются из культурной среды, асцитной жидкости или сыворотки с помощью обычных процедур очистки иммуноглобулинов, таких, например, как сорбент белок A-Sepharose, гидроксилапатитная хроматография, гель-электрофорез, диализ или аффинная хроматография.
Антитела анти-EGFL7 по настоящему изобретению могут быть получены с использованием комбинаторных библиотек для скрининга клонов синтетических антител с желаемой активностью или активностями. В принципе, клоны синтетических антител выбираются с помощью библиотек скрининга фагов, содержащих фаг, который выявляет различные фрагменты вариабельных областей антитела (Fv), слитого с белком покрытия фага. Такие библиотеки фагов сортируются с помощью аффинной хроматографии относительно желаемого антигена. Клоны, экспрессирующие фрагменты Fv, способные связывать желаемый антиген, адсорбируются на антигене и таким образом отделяются от несвязывающихся клонов в библиотеке. Связывающиеся клоны затем элюируются с антигена и могут дополнительно обогащаться с помощью дополнительных циклов адсорбции/элюирования по отношению к антигену. Некоторые антитела анти-EGFL7 по настоящему изобретению могут быть получены посредством конструирования процедуры скрининга соответствующего антигена для выбора клона фага, представляющего интерес, с последующим конструированием полноразмерного клона антитела анти-EGFL7 с использованием последовательностей Fv от клона фага, представляющего интерес, и соответствующих последовательностей константной области (Fc), описанных в Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD (1991), vols. 1-3.
Домен связывания с антигеном у антитела формируется из двух вариабельных (V) областей примерно из 110 аминокислот, по одной из легкой (VL) и тяжелой (VH) цепей, обе представляют собой три гипервариабельные петли или области, определяющие комплементарность (CDR). Вариабельные домены могут функционально проявляться на фаге, либо как одноцепочечные фрагменты Fv (scFv), в которых VH и VL ковалентно связываются через короткий, гибкий пептид, либо как фрагменты Fab, в которых они слиты, каждый, с константным доменом и взаимодействуют нековалентно, как описывается в Winter et al., Ann. Rev. Immunol. 12: 433-55 (1994). Как здесь используется, scFv, кодирующие клоны фагов и Fab, кодирующие клоны фагов, коллективно упоминаются как "клоны фагов Fv " или "клоны Fv".
Популяции генов VH и VL могут клонироваться по отдельности с помощью полимеразной цепной реакции (PCR) и рекомбинировать случайным образом в библиотеках фагов, в которых затем может производиться поиск клонов, связывающих антиген, как описывается в Winter et al., Ann. Rev. Immunol. 12: 433-55 (1994). Библиотеки из иммунизированных источников дают антитела с высоким сродством к иммуногену без выполнения требования конструирования гибридом. Альтернативно, наивная популяция может клонироваться для создания единого источника антител человека для несобственных антигенов и собственных антигенов без какой-либо иммунизации, как описывает Griffiths et al., EMBO J. 12: 725-34 (1993). Наконец, наивные библиотеки могут также быть получены синтетически посредством клонирования неперестроившихся сегментов V-генов от стволовых клеток, и использования праймеров PCR, содержащих случайную последовательность, для кодирования очень вариабельных областей CDR3 и получения перестройки in vitro, как описывается Hoogenboom & Winter, J. Mol. Biol. 227:381-88 (1992).
Нитевидный фаг используется для выявления фрагментов антител посредством слияния с минорным белком покрытия pIII. Фрагменты антител могут быть выявлены как одноцепочечные фрагменты Fv, в которых домены VH и VL соединяются на одной и той же полипептидной цепи с помощью гибкого полипептидного спейсера, например, как описывается в Marks et al., J. Mol. Biol. 222:581-597 (1991), или как фрагменты Fab, в которых одна цепь сливается с pIII, а другая секретируется в периплазму бактериальной клетки-хозяина, где собирает структуру белков покрытия Fab, которое становится видна на поверхности фага, посредством замещения некоторых белков покрытия дикого типа, например, как описывается в Hoogenboom et al., Nucl. Acids Res. 19:4133-37 (1991).
Как правило, нуклеиновые кислоты, кодирующие фрагменты генов антитела, получают из иммунных клеток, собранных у людей или животных. Если желаемой является библиотека, сдвинутая в сторону клонов анти-EGFL7, субъект иммунизируется с помощью EGFL7 для генерирования реакций антител, и клетки селезенки и/или циркулирующие B лимфоциты или другие лимфоциты периферической крови (PBL) извлекаются для конструирования библиотеки. В предпочтительном варианте осуществления, библиотека фрагментов генов антител человека, смещенная в сторону клонов EGFL7 анти-человек, получается посредством генерирования реакции антител EGFL7 анти-человек в трансгенных мышах, несущих матрицу генов функциональных иммуноглобулинов человека (и у которых отсутствует система продуцирования функциональных эндогенных антител), так что иммунизация EGFL7 приводит к появлению B лимфоцитов, продуцирующих антитела человека против EGFL7. Генерирование трансгенных мышей, продуцирующих антитела человека, описывается ниже.
Дополнительное обогащение популяций клеток, реагирующих на анти-EGFL7, может быть получено посредством использования соответствующей процедуры скрининга для выделения B лимфоцитов, экспрессирующих связанное с мембраной антитело, специфичное к EGFL7, например, посредством разделения клеток с помощью аффинной хроматографии на EGFL7 или адсорбции клеток на меченном флюорохромом EGFL7, с последующей проточной сортировкой активированных клеток (FACS).
Альтернативно, использование клеток селезенки и/или B лимфоцитов, или других PBL от иммунизированного донора дает лучшую репрезентацию возможной популяции антител, а также делает возможным конструирование библиотеки антител с использованием видов животных (человека или не-человека), в которых EGFL7 не является антигенным. Для библиотек, включающих в себя конструирование генов антител in vitro, стволовые клетки собираются у субъекта для получения нуклеиновых кислот, кодирующих неперестроившиеся сегменты генов антител. Иммунные клетки, представляющие интерес, могут быть получены от разнообразных видов животных, таких как человек, мышь, крыса, зайцеобразные, волки, псовые, кошачьи, свиньи, жвачные, лошади и птицы и тому подобное.
Нуклеиновые кислоты, кодирующие сегменты вариабельных генов антител (включая сегменты VH и VL), извлекаются из клеток, представляющих интерес, и амплифицируются. В случае перестроенных библиотек генов VH и VL, желаемая ДНК может быть получена посредством выделения геномной ДНК или мРНК из лимфоцитов, с последующей полимеразной цепной реакцией (PCR) с праймерами, согласующимися с 5' и 3' концами перестроенных генов VH и VL, как описывается в Orlandi et al., Proc. Natl. Acad. Sci. USA 86:3833-37 (1989), тем самым, получая различные популяции V генов для экспрессирования. V гены могут амплифицироваться посредством цДНК и геномной ДНК, с помощью обратных праймеров на 5' конце экзона, кодирующего зрелый V-домен, и прямых праймеров, базирующихся в J-сегменте, как описывается в Orlandi et al. (1989) и в Ward et al., Nature 341:544-46 (1989). Однако для амплифицирования с помощью цДНК, обратные праймеры также могут базироваться в лидерном экзоне, как описывается в Jones et al., Biotechnol. 9:88-89 (1991), а прямые праймеры, в константной области, как описывается в Sastry et al., Proc. Natl. Acad. Sci. USA 86:5728-32 (1989). Для доведения до максимума комплементарности, дегенерация может быть включена в праймеры, как описывается в Orlandi et al. (1989) или Sastry et al. (1989). Предпочтительно, разнообразие библиотеки доводится до максимума посредством использования праймеров PCR, направленных на каждое семейство V-генов для амплифицирования всех доступных расположений VH и VL, присутствующих в образце нуклеиновой кислоты иммунной клетки, например, как описывается в способе Marks et al., J. Mol. Biol. 222:581-97 (1991), или как описывается в способе Orum et al., Nucl. Acids Res. 21:4491-4498 (1993). Для клонирования амплифицированной ДНК в вектор экспрессии, редкие сайты рестрикции могут вводиться в праймер PCR как тэг на одном конце, как описывается в Orlandi et al. (1989), или с помощью дополнительного амплифицирования PCR с праймером с тэгом, как описывается в Clackson et al., Nature 352:624-628 (1991).
Популяции синтетически перестроенных V генов могут быть получены in vitro из сегментов V генов. Большинство сегментов VH-генов человека клонированы и секвенированы (как сообщается в Tomlinson et al., J. Mol. Biol. 227:776-98 (1992)), и картированы (как сообщается в Matsuda et al., Nature Genet. 3:88-94 (1993); эти клонированные сегменты (включая все главные конформации петли H1 и H2) могут использоваться для генерирования разнообразных популяций VH генов с PCR праймерами, кодирующими петли H3 с различными последовательностями и длинами, как описывается в Hoogenboom & Winter, J. Mol. Biol. 227:381-88 (1992). Популяции VH могут также быть получены со всем разнообразием последовательностей, сфокусированным в длинной петле H3 одной длины, как описывается в Barbas et al., Proc. Natl. Acad. Sci. USA 89:4457-61 (1992). Сегменты VΚ и Vλ человека клонированы и секвенированы (как сообщается в Williams & Winter, Eur. J. Immunol. 23:1456-61 (1993)) и могут использоваться для получения популяций синтетических легких цепей. Популяции синтетических V генов, на основе диапазона складок VH и VL, и длин L3 и H3, будут кодировать антитела со значительным структурным разнообразием. После амплификации ДНК, кодирующих V-ген, сегменты гермлайн V-гена могут перестраиваться in vitro в соответствии со способами Hoogenboom & Winter, J. Mol. Biol. 227:381-88 (1992).
Популяции фрагментов антител могут конструироваться посредством объединения вместе популяций VH и VL генов несколькими путями. Каждая популяция может создаваться в различных векторах, и векторы рекомбинируют in vitro, например, как описывается в Hogrefe et al., Gene 128:119-26 (1993), или in vivo посредством комбинаторной инфекции, например, системы loxP, описанной в Waterhouse et al., Nucl. Acids Res. 21:2265-66 (1993). Подход с рекомбинацией in vivo использует двухцепочечную природу фрагментов Fab для преодоления предела размера библиотеки, накладываемого эффективностью трансформирования E.coli. Наивные популяции VH и VL клонируются по отдельности, один в фагемид, а другой в фаговый вектор. Затем эти две библиотеки объединяются посредством инфицирования фагом содержащей фагемид бактерии, так что каждая клетка содержит иное сочетание, и размер библиотеки ограничивается только количеством присутствующих клеток (примерно 1012 клонов). Сигналы обоих векторов содержат рекомбинации in vivo, так что гены VH и VL рекомбинируют в единый репликон и совместно упаковываются в вирионы фагов. Эти мощные библиотеки дают большие количества разнообразных антител с хорошим сродством (Kd-1 примерно 10-8 M).
Альтернативно, популяции могут клонироваться последовательно в один и тот же вектор, например, как описывается в Barbas et al., Proc. Natl Acad. Sci. USA 88:7978-82 (1991), или собираться вместе с помощью PCR, а затем клонироваться, например, как описывается в Clackson et al., Nature 352:624-28 (1991). Сборка с помощью PCR может также использоваться для соединения VH и VL ДНК с ДНК, кодирующей гибкий пептидный спейсер, с формированием одноцепочечных популяций Fv (scFv). Еще в одной технологии, "клеточная сборка с помощью PCR" используется для объединения генов VH и VL внутри лимфоцитов с помощью PCR, а затем клонирования популяций связанных генов, как описывается в Emblazon et al., Nucl. Acids Res. 20:3831-37 (1992).
Антитела, продуцируемые наивными библиотеками (либо природными, либо синтетическими), могут иметь умеренное сродство (Kd-1 от примерно 106 до 107 M-1), но созревание сродства может также копироваться in vitro посредством конструирования и повторного выбора из вторичных библиотек, как описывается в Winter et al. (1994), выше. Например, мутация может вводиться случайным образом in vitro посредством использования ошибочно-направленной полимеразы (как сообщается в Leung et al., Technique 1:11-15 (1989)) в способе Hawkins et al., J. Mol. Biol. 226:889-96 (1992) или в способе Gram et al., Proc. Natl. Acad. Sci. USA 89:3576-80 (1992). В дополнение к этому, созревание сродства может осуществляться посредством случайных мутаций одной или нескольких CDR, например, с использованием PCR с праймерами, несущими случайную последовательность, расширяющую CDR, представляющую интерес, в выбранных индивидуальных клонах Fv и скрининга клонов с более высоким сродством. Заявка WO 96/07754 (опубликованная 14 марта 1996 года) описывает способ индуцирования мутагенеза в комплементарно определяемой области легкой цепи иммуноглобулина для создания библиотеки генов легких цепей. Другой эффективный подход заключается в рекомбинации доменов VH или VL, выбранных с помощью фагового дисплея, с популяциями существующих в природе вариантов домена V, полученных от неиммунизированных доноров, и скрининга на более высокое сродство в нескольких заходах решаффлинга цепи, как описывается в Marks et al., Biotechnol. 10:779-83 (1992). Это технология делает возможным продуцирование антител и фрагментов антител со сродством в диапазоне 10-9 M.
Последовательность нуклеиновых кислот, кодирующая EGFL7, может конструироваться с использованием последовательности аминокислот желаемой области EGFL7. Альтернативно, может использоваться последовательность цДНК (или ее фрагменты). Дополнительные последовательности EGFL7 дополнительно описываются, например, в NM_022963 и Xie et al., Cytokine 11:729-35 (1999). ДНК, кодирующие EGFL7, могут быть получены посредством различных способов, известных в данной области. Эти способы включают в себя, но, не ограничиваясь этим, химический синтез с помощью любого из способов, описанных в Engels et al., Agnew. Chem. Int. Ed. Engl. 28:716-34 (1989), таких как способы со сложным триэфиром, фосфитом, фосфорамидитом и H-фосфонатом. В одном из вариантов осуществления, кодоны, предпочитаемые экспрессирующими клетками-хозяевами, используются для конструирования ДНК, кодирующей EGFL7. Альтернативно, ДНК, кодирующая EGFL7, может выделяться из геномной библиотеки или библиотеки цДНК.
После конструирования молекулы ДНК, кодирующей EGFL7, молекула ДНК функционально соединяется с контрольной последовательностью экспрессии в векторе экспрессии, таком как плазмида, где контрольная последовательность распознается в клетке-хозяине, трансформированной с помощью вектора. Как правило, векторы плазмид содержат последовательности репликации и контроля, которые получают от видов, совместимых с клеткой-хозяином. Вектор обычно несет сайт репликации, а также последовательности, которые кодируют белки, которые способны обеспечивать фенотипическую селекцию в трансформированных клетках. Соответствующие векторы для экспрессии в прокариотических и эукариотических клетках-хозяевах, известны в данной области и некоторые из них дополнительно описываются здесь. Могут использоваться эукариотические организмы, такие как дрожжи, или клетки, полученные от многоклеточных организмов, таких как млекопитающие.
Необязательно, ДНК, кодирующая EGFL7, функционально соединяется с секреторной лидерной последовательностью, что приводит к секреции продукта экспрессии клеток-хозяев в культурной среде. Примеры секреторных лидерных последовательностей включают в себя stII, экотин, lamB, герпес GD, lpp, щелочную фосфатазу, инвертазу и альфа фактор. Также пригодной для использования здесь является лидерная последовательность из 36 аминокислот белка A (Abrahmsen et al., EMBO J. 4:3901 (1985)).
Клетки-хозяева трансфицируются и предпочтительно трансформируются с помощью описанных выше векторов экспрессии или клонирования по настоящему изобретению и культивируются в обычных питательных средах, модифицированных соответствующим образом, для индуцирования промоторов, селекции трансформантов, или амплификации генов, кодирующих желаемые последовательности.
Трансфекция относится к поглощению вектора экспрессии клеткой-хозяином, независимо от того, экспрессируются ли на самом деле кодирующие последовательности или нет. Многочисленные способы трансфицирования известны специалистам в данной области, например, преципитация с помощью CaPO4 и электропорообразование. Успешная трансфекция, как правило, наблюдается, когда осуществляется любая индикация функционирования этого вектора внутри клетки-хозяина. Методы трансфицирования хорошо известны в данной области, и некоторые из них дополнительно описываются здесь.
Трансформация означает введение ДНК в организм так, что ДНК является реплицируемой, либо как экстрахромосомальный элемент, либо посредством хромосомального интегранта. В зависимости от используемой клетки-хозяина, трансформирование осуществляется с использованием стандартных технологий, соответствующих таким клеткам. Способы трансформирования хорошо известны в данной области, и некоторые из них дополнительно описываются здесь.
Прокариотические клетки-хозяева, используемые для продуцирования EGFL7, могут культивироваться, как описывается, в целом, в Sambrook et al., выше.
Клетки-хозяева млекопитающих, используемые для продуцирования EGFL7, могут культивироваться в разнообразных средах, которые хорошо известны в данной области, и некоторые из которых описываются здесь.
Клетки-хозяева, упоминаемые в настоящем описании, охватывают как клетки в культуре in vitro, так и клетки, которые находятся внутри животного-хозяина.
Очистка EGFL7 может осуществляться с использованием способов, известных в данной области.
Очищенные EGFL7 могут присоединяться к соответствующей матрице, такой как агарозные шарики, акриламидные шарики, стелянные шарики, целлюлоза, различные акриловые сополимеры, гидроксил метакрилатные гели, полиакриловые и полиметакриловые сополимеры, нейлон, нейтральные и ионные носители и тому подобное, для использования при разделении с помощью аффинной хроматографии клонов фаговых дисплеев. Прикрепление белка EGFL7 к матрице может осуществляться с помощью способов, описанных в Meth. Enzymol. vol. 44 (1976). Повсеместно используемая технология для прикрепления белковых лигандов к полисахаридным матрицам, например, агарозе, декстрану или целлюлозе, включает в себя активирование носителя с помощью цианоген галогенидов и последующее связывание первичных алифатических или ароматических аминов пептидных лигандов с активированной матрицей.
Альтернативно, EGFL7 может использоваться для покрытия лунок адсорбционных планшетов, экспрессироваться в клетках-хозяевах, зафиксированных на адсорбционных планшетах, или использоваться при сортировке клеток, или конъюгироваться с биотином для захвата с помощью шариков, покрытых стрептавидином, или использоваться в любом другом способе, известном в данной области для пэннинга библиотек фаговых дисплеев.
Образцы библиотек фагов приводятся в контакт с иммобилизованной EGFL7 при условиях, пригодных для связывания, по меньшей мере, части частиц фагов с адсорбентом. Обычно, условия, включающие в себя pH, ионную силу, температуру и тому подобное, выбираются с копированием физиологических условий. Фаги, связанные с твердой фазой, промываются, а затем элюируются с помощью кислоты, например, как описывается в Barbas et al., Proc. Natl. Acad. Sci USA 88:7978-82 (1991), или щелочи, например, как описывается в Marks et al., J. Mol. Biol. 222:581-97 (1991), или посредством конкурирования с антигеном EGFL7, например, в процедуре, сходной со способом конкурирования антигенов Clackson et al., Nature 352:624-28 (1991). Фаги могут обогащаться в 20-1000-раз за одну стадию селекции. Кроме того, обогащенные фаги могут выращиваться в бактериальной культуре и подвергаться дополнительным заходам селекции.
Эффективность селекции зависит от многих факторов, включая кинетику диссоциации во время промывки, или от того могут ли множество фрагментов антител на одном фаге одновременно соединяться с антигеном. Антитела с быстрой кинетикой диссоциации (и со слабым сродством связывания) могут удерживаться посредством использования коротких промывок, многовалентного фагового дисплея и высокой плотности покрытия из антигена в твердой фазе. Высокая плотность не только стабилизирует фаг во время многовалентных взаимодействий, но и способствует повторному связыванию с фагом, который диссоциирует. Селекция антител с медленной кинетикой диссоциации (и с хорошим сродством связывания) может ускоряться посредством использования длительных промывок и одновалентного фагового дисплея, как описывается в Bass et al., Proteins 8:309-14 (1990) и в заявке WO 92/09690, и низкой плотности покрытия антигена, как описывается в Marks et al., Biotechnol. 10:779-83 (1992).
Является возможной селекция антител фагов с различным сродством, даже если сродство различается мало, к EGFL7. Однако случайная мутация выбранного антитела (например, как делается в некоторых из технологий созревания сродства, описанных выше), вероятно, даст множество мутантов, в большинстве своем связывающимся с антигеном, и несколько из них с более высоким сродством. При ограничении количества EGFL7, редкие фаги с высоким сродством будут отсеиваться в конкуренции. Для удерживания всех мутантов с более высоким сродством, фаги могут инкубироваться вместе с избытком биотинилированных EGFL7, но при концентрации биотинилированных EGFL7 с более низкой молярностью, чем целевое молярное сродство для EGFL7. Фаги с высоким сродством связывания могут затем захватываться с помощью покрытых стрептавидином парамагнитных шариков. Такой "равновесный захват" делает возможным селекцию антител, в соответствии с их сродством связывания, с чувствительностью, которая делает возможным выделение мутантных клонов всего лишь со сродством, в два раза большим, из большого избытка фагов с более низким сродством. Условия, используемые для промывки фагов, связанных с твердой фазой, могут также подбираться для дискриминации на основе кинетики диссоциации. Клоны анти-EGFL7 могут также выбираться по активности.
ДНК, кодирующая моноклональные антитела, полученные из гибридом, или клоны фагового дисплея Fv по настоящему изобретению, легко выделяется и секвенируется с использованием обычных процедур (например, посредством использования олигонуклеотидных праймеров, сконструированных для специфичной амплификации областей кодирования тяжелых и легких цепей, представляющих интерес, из матрицы ДНК гибридомы или фага). После выделения, ДНК может помещаться в векторы экспрессии, которые затем трансфицируются в клетки-хозяева, такие как клетки E.coli, клетки COS обезьян, клетки яичников китайского хомячка (CHO) или клетки миеломы, которые в ином случае не продуцируют белков иммуноглобулинов, для получения синтеза желаемых моноклональных антител в рекомбинантных клетках-хозяевах. Обзорные статьи по рекомбинантной экспрессии в бактериях ДНК, кодирующих антитела, включают в себя Skerra et al., Curr. Opinion in Immunol 5: 256 (1993) и Plilckthun, Immunol. Rev. 130:151 (1992).
ДНК, кодирующие клоны Fv по настоящему изобретению, могут объединяться с известными последовательностями ДНК, кодирующими константные области тяжелых цепей и/или легких цепей (например, соответствующие последовательности ДНК могут быть получены согласно Kabat et al., выше), для формирования клонов, кодирующих полную или частичную длину тяжелых и/или легких цепей. Будет понятно, что константные области любого изотипа могут использоваться для этой цели, включая константные области IgG, IgM, IgA, IgD и IgE, и что такие константные области могут быть получены от любого человека или видов животных. Клон Fv, полученный от вариабельного домена ДНК одного вида животного (такого как человека), а затем слитой с константной областью ДНК других видов животных, с образованием кодирующей последовательности (последовательностей) для "гибридной" полноразмерной тяжелой цепи и/или легкой цепи включается в определение "химерного" и "гибридного" антитела, как здесь используется. В предпочтительном варианте осуществления, клон Fv, полученный от вариабельной ДНК человека, сливается с константной областью ДНК человека, с формированием кодирующей последовательности (последовательностей) для всех тяжелых и/или легких цепей с полной или частичной длиной для человека.
ДНК, кодирующая антитело анти-EGFL7, полученная из гибридомы по настоящему изобретению, также может модифицироваться, например, посредством замещения кодирующей последовательности для константных доменов тяжелых и легких цепей для человека вместо гомологических последовательностей мышей, полученных из клона гибридомы (например, как в способе Morrison et al., Proc. Natl. Acad. Sci. USA 81:6851-55 (1984)). ДНК, кодирующая антитело, полученное из гибридомы или клона Fv, или его фрагмент, может дополнительно модифицироваться путем ковалентного присоединения к кодирующей последовательности иммуноглобулина всей кодирующей последовательности или ее части, для полипептида, отличного от иммуноглобулина. Таким образом, получают "химерные" или "гибридные" антитела, которые имеют специфичность связывания полученных от клона Fv или клона гибридомы антител по настоящему изобретению.
Фрагменты антител
Настоящее изобретение охватывает фрагменты антител. В определенных обстоятельствах имеются преимущества использования фрагментов антител, вместо целых антител. Меньший размер фрагментов делает возможным быстрое выведение и может привести к улучшению доступа к твердым опухолям.
Различные технологии разработаны для продуцирования фрагментов антитела. Традиционно эти фрагменты получают посредством протеолитического гидролиза антител (см., например, Morimoto et al., J. Biochem. Biophys. Meth. 24:107-17 (1992); и Brennan et al., Science 229:81 (1985)). Однако эти фрагменты сегодня могут продуцироваться непосредственно рекомбинантными клетками-хозяевами. Все фрагменты Fab, Fv и ScFv антитела могут экспрессироваться и секретироваться из E.coli, делая, таким образом, возможным простое продуцирование больших количеств этих фрагментов. Фрагменты антител могут выделяться из фаговых библиотек антител, обсуждаемых выше. Альтернативно, фрагменты Fab'-SH могут непосредственно извлекаться из E.coli и химически соединяться с формированием фрагментов F(ab')2 (Carter et al., Bio/Technology 10:163-67 (1992)). В соответствии с другим подходом, фрагменты F(ab')2 могут выделяться непосредственно из культуры рекомбинантных клеток-хозяев. Фрагмент Fab и F(ab')2 с увеличенным половинным временем жизни in vivo, содержащий остатки эпитопов связывания рецепторов реутилизации, описывается в заявке на патент США № 5869046. Другие технологии для продуцирования фрагментов антитела будут понятны специалисту в данной области. В других вариантах осуществления, выбранное антитело представляет собой одноцепочечный фрагмент Fv (scFv). См. заявку WO 93/16185; заявки на патенты США № 5571894; и 5587458. Fv и sFv представляют собой единственные частицы с объединением интактных сайтов, которые не содержат константных областей; таким образом, они являются пригодными для уменьшения неспецифичного связывания при использовании in vivo. Белки слияния sFv могут конструироваться для получения эффекторного белка слияния, либо на амино, либо на карбокси-конце sFv. См. Antibody Engineering, ed. Borrebaeck, W.H. Freeman and Company (1992). Фрагмент антитела также может представлять собой "линейное антитело", как описывается, например, в заявке на патент США № 5641870. Такие линейные фрагменты антитела могут быть моноспецифичными или биспецифичными.
Гуманизированные антитела
Настоящее изобретение охватывает гуманизированные антитела. Различные способы гуманизации антител не-человека известны в данной области. Например, гуманизированное антитело может иметь один или несколько остатков аминокислот, введенных в него из источника, который представляет собой не-человека. Эти остатки аминокислот не-человека часто упоминаются как "импортные" остатки, которые обычно берутся от "импортного" вариабельного домена. Гуманизация может осуществляться по существу, следуя способу Winter и сотрудников (Jones et al., Nature 321:522-25; Riechmann et al., Nature 332:323-27 (1988); Verhoeyen et al.. Science 239:1534-36 (1988)), посредством замещения последовательностями гипервариабельной области соответствующих последовательностей в антителе человека. Соответственно, такие "гуманизированные" антитела представляют собой химерные антитела (заявка на патент США № 4816567), в которых по существу менее чем интактный вариабельный домен человека замещается соответствующей последовательностью от видов не-человека. На практике, гуманизированные антитела представляют собой, как правило, антитела человека, с некоторыми остатками гипервариабельной области и, возможно, некоторыми остатками FR, замещенными остатками из аналогичных сайтов антител грызунов.
Выбор вариабельных доменов человека, как легких, так и тяжелых, которые должны использоваться при получении гуманизированных антител, является очень важным для уменьшения антигенности. В соответствии с так называемым способом "наилучшей подгонки", последовательность вариабельного домена антитела грызуна просматривается в сравнении со всей библиотекой известных последовательностей вариабельных доменов человека. Последовательность человека, которая ближе всех к последовательности грызуна, принимается затем в качестве рамки человека для гуманизированного антитела (Sims et al., J. Immunol 151:2296 (1993); Chothia et al., J. Mol. Biol. 196:901 (1987). Другой способ использует конкретную рамку, полученную от консенсусной последовательности всех антител человека для конкретной подгруппы легких или тяжелых цепей. Такая же рамка может использоваться для нескольких различных гуманизированных антител (Carter et al., Proc. Natl. Acad. Sci. USA 89:4285 (1992); Presta et al., J. Immunol. 151:2623(1993).
Является важным, кроме того, чтобы антитела гуманизировались с сохранением высокого сродства к антигену и других благоприятных биологических свойств. Для достижения этой цели, в соответствии с одним из способов, гуманизированные антитела приготавливают с помощью способа анализа исходных последовательностей и различных концептуальных гуманизированных продуктов с использованием трехмерных моделей исходных и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов являются широко доступными и хорошо знакомы специалистам в данной области. Доступны компьютерные программы, которые иллюстрируют и отображают возможные трехмерные конформационные структуры выбранных кандидатов в иммуноглобулиновые последовательности. Их просмотр делает возможным анализ вероятной роли остатков при функционировании кандидатов в иммуноглобулиновые последовательности, то есть анализ остатков, которые воздействуют на способность кандидата в иммуноглобулины к связыванию его антигена. Таким образом, остатки FR могут выбираться и объединяться из последовательностей реципиента и импортных последовательностей, так что достигаются желаемые характеристики антитела, такие как повышенное сродство к целевому антигену (антигенам). Как правило, остатки гипервариабельных областей являются непосредственно и наиболее существенно вовлеченными во влияние на связывание антигена.
Антитела человека
Антитела человека анти-EGFL7 по настоящему изобретению могут конструироваться посредством объединения последовательности (последовательностей) вариабельного домена клона Fv, выбранной из полученных от человека библиотек фаговых дисплеев с известной последовательностью (последовательностями), и константного домена человека, как описывается выше. Альтернативно, моноклональные антитела человека анти-EGFL7 по настоящему изобретению могут быть получены с помощью гибридомного способа. Линии клеток миеломы человека и гетеромиеломы мыши-человека для продуцирования моноклональных антител человека описываются, например, в Kozbor J. Immunol. 133:3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987); и Boerner et al., J. Immunol. 147:86 (1991).
Сегодня возможно получение трансгенных животных (например, мышей), которые способны, при иммунизации, продуцировать полную популяцию антител человека в отсутствии продуцирования эндогенного иммуноглобулина. Например, описывалось, что гомозиготная делеция соединительной области тяжелой цепи гена антитела (JH) у химерных и гермлайн мутантных мышей приводит к полному ингибированию продуцирования эндогенного антитела. Перенос матрицы гермлайн генов иммуноглобулинов человека в таких гермлайн мутантных мышей будет приводить к продуцированию антител человека при провоцировании антигеном. См., например, Jakobovits et al., Proc. Natl. Acad. Sci USA 90:2551 (1993); Jakobovits et al., Nature 362:255 (1993); Bruggermann et al., Year in Immunol. 7:33 (1993).
Шаффлинг генов также может использоваться для получения антител человека из антител не-человека, например грызуна, где антитело человека имеет сходное сродство и специфичность с исходным антителом не-человека. В соответствии с этим способом, который называется также "импринтинг эпитопа", вариабельная область либо тяжелой, либо легкой цепи фрагмента антитела не-человека, полученного с помощью технологий фагового дисплея, как описывается выше, заменяется с помощью популяции генов V домена человека, создавая популяцию химер scFv или Fab, цепь не-человека/цепь человека. Селекция с помощью антигена приводит к выделению химерных scFv или Fab цепь не-человека/цепь человека, где цепь человека восстанавливает сайт связывания с антигеном, разрушенный при удалении соответствующей цепи не-человека в первичном клоне фагового дисплея, то есть эпитоп определяет (отпечатывает) выбор партнера цепи человека. Когда этот процесс повторяется для замены оставшейся цепи не-человека, получается антитело человека (см. публикацию PCT заявки WO 93/06213, опубликованную 1 апреля 1993 года). В отличие от традиционной гуманизации антител не-человека с помощью имплантации CDR, эта технология создает полностью человеческие антитела, которые не имеют остатков FR или CDR нечеловеческого происхождения.
Биспецифичные антитела
Биспецифичные антитела представляют собой моноклональные антитела, предпочтительно, антитела человека или гуманизированные антитела, которые имеют специфичность связывания, по меньшей мере, к двум различным антигенам. В настоящем случае, одна из специфичностей связывания предназначена для EGFL7, а другая для любого другого антигена. Примерные биспецифичные антитела могут связываться с двумя различными эпитопами белка EGFL7. Биспецифичные антитела могут также использоваться для локализации цитотоксичных агентов в клетках, которые экспрессируют EGFL7. Эти антитела имеют плечо связывания для EGFL7 и плечо, которое связывает цитотоксичный агент (например, сапорин, анти-интерферон-α, алколоид барвинка, цепь A рицина, метотрексат или гаптен с радиоактивным изотопом). Биспецифичные антитела могут быть получены в виде полноразмерных антител или фрагментов антител (например, биспецифичные антитела F(ab')2).
Способы получения биспецифичных антител известны в данной области. Традиционно, рекомбинантное получение биспецифичных антител основывается на совместной экспрессии двух пар тяжелая цепь:легкая цепь иммуноглобулина, где две тяжелых цепи имеют различные специфичности (Milstein & Cuello, Nature 305:537 (1983)). Благодаря случайному ассортименту тяжелых и легких цепей иммуноглобулинов, эти гибридомы (квадромы) дают потенциальную смесь из 10 различных молекул антител, из которых только одна имеет правильную биспецифичную структуру. Очистка правильной молекулы, которая обычно осуществляется с помощью стадий аффинной хроматографии, является скорее трудоемкой, и выход продуктов низкий. Сходные процедуры описываются в заявке WO 93/08829, опубликованной 13 мая, 1993 года, и в Traunecker et al., EMBO J. 10:3655 (1991).
В соответствии с другим и более предпочтительным подходом, вариабельные домены антител с желаемыми специфичностями связывания (сайтами объединения антитело-антиген) сливаются с последовательностями константных доменов иммуноглобулинов. Слияние предпочтительно осуществляется с помощью константного домена тяжелой цепи иммуноглобулина, содержащего, по меньшей мере, часть шарнирной области, областей CH2 и CH3. Предпочтительно иметь первую константную область тяжелой цепи (CH1), содержащую сайт, необходимый для связывания легкой цепи, присутствующей, по меньшей мере, в одном из слияний. ДНК, кодирующие слияния тяжелых цепей иммуноглобулина и, если это желательно, легкой цепи иммуноглобулина, вставляются в отдельные векторы экспрессии и совместно трансфицируются в соответствующий организм хозяин. Это обеспечивает большую гибкость при подборе взаимных пропорций трех полипептидных фрагментов в вариантах осуществления, когда неравные отношения трех полипептидных цепей, используемых при конструировании, дают оптимальные выходы. Однако является возможной вставка кодирующих последовательностей для двух или для всех трех полипептидных цепей в один вектор экспрессии, когда экспрессирование, по меньшей мере, двух полипептидных цепей при равных отношениях дает в результате высокий выход, или когда отношения не имеют особого значения.
В некоторых вариантах осуществления этого подхода, биспецифичные антитела состоят из тяжелой цепи гибридного иммуноглобулина с первой специфичностью связывания в одном плече и с парой тяжелая цепь - легкая цепь гибридного иммуноглобулина (обеспечивающей вторую специфичность связывания) в другом плече. Обнаружено, что эта асимметричная структура облегчает отделение желаемого биспецифичного соединения от нежелательных сочетаний цепей иммуноглобулинов, поскольку присутствие легкой цепи иммуноглобулина только в одной половине биспецифичной молекулы обеспечивает удобный способ разделения. Этот подход описывается в заявке WO 94/04690. Относительно дополнительных деталей генерирования биспецифичных антител см., например, Suresh et al., Meth. Enzymol. 121:210 (1986).
В соответствии с другим подходом, область контакта между парой молекул антител может быть получена с помощью генной инженерии для доведения до максимума процента гетеродимеров, которые извлекаются из рекомбинантной культуры клеток. Предпочтительная область контакта содержит, по меньшей мере, часть домена CH3 из константного домена антитела. В этом способе, одна или несколько боковых цепей малых аминокислот из области контакта молекулы первого антитела заменяются боковыми цепями большего размера (например, тирозина или триптофана). Компенсаторные "полости" размера, идентичного или сходного с большой боковой цепью (цепями), создаются в области контакта молекулы второго антитела посредством замены больших боковых цепей аминокислот меньшими (например, аланином или треонином). Это создает механизм увеличения выхода гетеродимеров по сравнению с другими нежелательными конечными продуктами, такими как гомодимеры.
Биспецифичные антитела включают в себя поперечно связанные или "гетероконъюгированные" антитела. Например, одно из антител в гетероконъюгате может быть связано с авидином, другое с биотином. Такие антитела были, например, предложены для нацеливания клеток иммунной системы на нежелательные клетки (заявка на патент США № 4676980) и для лечения ВИЧ инфекции (заявки WO 91/00360 и WO 92/00373). Гетероконъюгированные антитела могут быть получены с использованием любых обычных способов поперечного связывания. Пригодные для использования агенты для поперечного связывания хорошо известны в данной области и описываются в заявке на патент США № 4676980, вместе с рядом технологий поперечного связывания.
Технологии генерирования биспецифичных антител из фрагментов антител также описываются в литературе. Например, биспецифичные антитела могут быть получены с использованием химических связей. Brennan et al., Science 229:81 (1985) описывают процедуру, где интактные антитела протеолитически расщепляются с генерированием фрагментов F(ab')2. Эти фрагменты восстанавливаются в присутствии дитиолового комплексообразующего агента арсенита натрия для стабилизации вицинальных дитиолов и предотвращения образования внутримолекулярных дисульфидов. Генерируемые фрагменты Fab' затем преобразуются в тионитробензоатные (TNB) производные. После этого производные Fab'-TNB преобразуются затем обратно в Fab'-тиол посредством восстановления с помощью меркаптоэтиламина и смешиваются с эквимолярным количеством другого производного Fab'-TNB с формированием биспецифичных антител. Полученные биспецифичные антитела могут использоваться в качестве агентов для селективной иммобилизации ферментов.
Недавний прогресс сделал возможным непосредственное извлечение фрагментов Fab'-SH из E.coli, которые могут химически связываться с образованием биспецифичных антител. Shalaby et al., J. Exp. Med 175:217-25 (1992) описывает получение молекулы полностью гуманизированного биспецифичного антитела F(ab')2. Каждый фрагмент Fab' отдельно секретируется из E.coli и подвергается направленному химическому связыванию in vitro с формированием биспецифичного антитела. Биспецифичное антитело, сформированное таким образом, способно связывать клетки, сверхэкспрессирующие рецептор HER2 и нормальные T-лимфоциты человека, а также запускать литическую активность цитотоксичных лимфоцитов человека против целевых опухолей груди человека.
Различные технологии получения и выделения биспецифичных фрагментов антител непосредственно из рекомбинантной культуры клеток также описываются. Например, биспецифичные антитела получают с использованием лейциновой «застежки». Kostelny et al., J. Immunol. 148(5):1547-53 (1992). Пептиды лейциновой «застежки» из белков Fos и Jun связываются с частями Fab' двух различных антител посредством слияния генов. Гомодимеры антител восстанавливаются в шарнирной области с формированием мономеров, а затем повторно окисляются с формированием гетеродимеров антител. Этот способ может также использоваться для получения гомодимеров антител. Технология "диабоди", описанная Hollinger et al., Proc. Natl. Acad. Sci. USA 90:6444-48 (1993), дает альтернативный механизм для получения биспецифичных фрагментов антител. Фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) с помощью линкера, который слишком короткий, чтобы сделать возможным спаривание между двумя доменами в одной и той же цепи. Соответственно, домены VH и VL одного фрагмента принуждаются к спариванию с комплементарными VL и VH доменами другого фрагмента, тем самым формируя два сайта связывания с антигеном. Другая стратегия получения биспецифичных фрагментов антитела посредством использования одноцепочечных димеров Fv (sFv) также описывается. См. Gruber et al., J. Immunol. 152:5368 (1994).
Также рассматриваются антитела, имеющие более двух валентностей. Например, могут быть получены триспецифичные антитела. Tutt et al. J. Immunol. 147:60 (1991).
Многовалентные антитела
Многовалентное антитело может быть интернализовано (и/или катаболизировано) быстрее, чем двухвалентное антитело, клеткой, экспрессирующей антиген, с которым связываются антитела. Антитела по настоящему изобретению могут представлять собой многовалентные антитела (которые не принадлежат к классу IgM) с тремя или более сайтами связывания антигена (например, четырехвалентные антитела), которые могут легко быть получены посредством рекомбинантной экспрессии нуклеиновой кислоты, кодирующей полипептидные цепи антитела. Многовалентное антитело может содержать домен димеризации и три или более сайтов связывания антигена. Предпочтительный домен димеризации содержит область Fc или шарнирную область (или состоит из нее). В этом сценарии, антитело будет содержать область Fc и три или более сайтов связывания антигена, амино-концевых по отношению к области Fc. Предпочтительное многовалентное антитело здесь содержит из трех до примерно восьми, но предпочтительно четыре сайта связывания антигена (или состоит из них). Многовалентное антитело содержит, по меньшей мере, одну полипептидную цепь (а предпочтительно две полипептидных цепи), где полипептидная цепь (цепи) содержит два или более вариабельных доменов. Например, полипептидная цепь (цепи) может содержать VD1-(X1)n-VD2-(X2)n-Fc, где VD1 представляет собой первый вариабельный домен, VD2 представляет собой второй вариабельный домен, Fc представляет собой одну полипептидную цепь из области Fc, X1 и X2 представляют аминокислоту или полипептид, и n равен 0 или 1. Например, полипептидная цепь (цепи) может содержать: VH-CH1-гибкий линкер-VH-CH1-цепь области Fc; или VH-CH1-VH-CH1-цепь области Fc. Многовалентное антитело здесь предпочтительно дополнительно содержит, по меньшей мере, два (а предпочтительно четыре) полипептида вариабельного домена легкой цепи. Многовалентное антитело здесь может, например, содержать от примерно двух до примерно восьми полипептидов вариабельного домена легкой цепи. Полипептиды вариабельного домена легкой цепи, рассматриваемые здесь, содержат вариабельный домен легкой цепи и, необязательно, дополнительно содержат домен CL.
Варианты антител
В некоторых вариантах осуществления рассматривается модификация (модификации) последовательностей аминокислот антител, описанных здесь. Например, может быть желательным улучшение сродства связывания и/или других биологических свойства антитела. Варианты последовательностей аминокислот антитела получают посредством введения соответствующих изменений нуклеотидов в нуклеиновую кислоту антитела или посредством синтеза пептидов. Такие модификации включают в себя, например, делеции и/или инсерции, и/или замещения остатков в последовательностях аминокислот антитела. Любое сочетание делеций, инсерций и замещений осуществляется для получения конечного продукта, при условии, что конечный продукт имеет желаемые характеристики. Изменения аминокислот могут вводиться в последовательность аминокислот рассматриваемого антитела в то время, когда получается последовательность.
Полезный способ идентификации определенных остатков или областей антитела, которые представляют собой предпочтительные положения для мутагенеза, называется "аланиновый сканирующий мутагенез", как описывается в Cunningham & Wells, Science 244:1081-85 (1989). Здесь остаток или группа целевых остатков идентифицируется (например, заряженные остатки, такие как arg, asp, his, lys и glu) и заменяется нейтральными или отрицательно заряженными аминокислотами (наиболее предпочтительно, аланином или полиаланином) для осуществления взаимодействия аминокислот с антигеном. Те положения аминокислот, которые демонстрируют функциональную чувствительность к замещениям, затем улучшаются посредством введения дополнительных или других вариантов на сайтах замещения или вместо них. Таким образом, хотя сайт для введения вариации последовательности аминокислот является заданным, природа мутации сама по себе не должна быть заданной. Например, для анализа параметров мутации на данном сайте, осуществляют ala сканирование или случайный мутагенез на целевом кодоне или области, и экспрессируемые иммуноглобулины просматриваются на желаемую активность.
Инсерции последовательности аминокислот включают в себя амино- и/или карбоксил-концевые слияния, находящиеся в диапазоне длин от одного остатка до полипептидов, содержащих сто или боле остатков, а также инсерции внутри последовательности одного или множества остатков аминокислот. Примеры конечных инсерций включают в себя антитело с N-концевым метионильным остатком или антитело, слитое с цитотоксичным полипептидом. Другие инсерционные варианты молекул антитела включают в себя слияние с помощью N- или C-конца антитела с ферментом (например, ADEPT) или полипептидом, который увеличивает половинное время жизни в сыворотке для антитела.
Другой тип варианта аминокислот антитела изменяет исходный паттерн гликозилирования антитела. Такое изменение включает в себя делецию одного или нескольких углеводных остатков, находящихся в антителе, и/или добавление одного или нескольких сайтов гликозилирования, которые не присутствуют в антителе.
Гликозилирование полипептидов, как правило, является либо N-связанным, либо O-связанным. N-связанное гликозилирование относится к присоединению углеводного остатка к боковой цепи аспарагинового остатка. Трипептидные последовательности аспарагин-X-серин и аспарагин-X-треонин, где X представляет собой любую аминокислоту за исключением пролина, представляют собой распознающие последовательности для ферментативного присоединения углеводного остатка к аспарагиновой боковой цепи. Таким образом, присутствие любой из этих трипептидных последовательностей в полипептиде создает потенциальный сайт гликозилирования. O-связанное гликозилирование относится к присоединению одного из сахаров N-ацетилгалоктозамина, галоктозы или ксилозы к гидроксиаминокислоте, чаще всего, к серину или треонину, хотя 5-гидроксипролин или 5-гидроксилизин также могут использоваться.
Добавление сайтов гликозилирования к антителу удобно осуществлять посредством изменения последовательности аминокислот, так чтобы она содержала одну или несколько из описанных выше трипептидных последовательностей (для N-связанных сайтов гликозилирования). Изменение может также осуществляться посредством добавления, или замещения с их помощью, одного или нескольких сериновых или треониновых остатков к последовательности исходного антитела (для O-связанных сайтов гликозилирования).
Когда антитело содержит область Fc, углевод, присоединенный к ней, может быть изменен. Например, антитела со зрелой углеводной структурой, в которой отсутствует фукоза, присоединенная к области Fc антитела, описываются в заявке на патент США № 2003/0157108 (Presta, L.). См. также заявку США № 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). Антитела с биссекторным N-ацетилглюкозамином (GlcNAc) в углеводе, присоединенном к области Fc антитела, упоминаются в заявке WO 2003/011878, Jean-Mairet et al. и в заявке на патент США № 6602684, Umafia et al. Об антителах, по меньшей мере, с одним галактозным остатком в олигосахариде, присоединенном к области Fc антитела, сообщается в заявке WO 97/30087, Patel et al. См., также, заявку WO 98/58964 (Raju, S.) и заявку WO99/22764 (Raju, S.) относительно антител с измененным углеводом, присоединенным к их области Fc. См. также заявку на патент США № 2005/0123546 (Umana et al.) относительно связывающихся с антигеном молекул с модифицированным гликозилированием.
Предпочтительный вариант гликозилирования здесь включает в себя область Fc, где углеводная структура, присоединенная к области Fc, не содержит фукозы. Такие варианты имеют улучшенную функцию ADCC. Необязательно, область Fc дополнительно содержит одно или несколько замещений аминокислот в ней, что дополнительно улучшает ADCC, например, замещения в положениях 298, 333 и/или 334 области Fc (нумерация остатков согласно EU). Примеры публикаций, относящихся к "дефукозилированным" или "фукоза-дефицитным" антителам, включают в себя: заявку на патент США № 2003/0157108; заявку WO 2000/61739; заявку WO 2001/29246; заявку на патент США № 2003/0115614; заявку на патент США № 2002/0164328; заявку на патент США № 2004/0093621; заявку на патент США № 2004/0132140; заявку на патент США № 2004/0110704; заявку на патент США № 2004/0110282; заявку на патент США № 2004/0109865; заявку WO 2003/085119; заявку WO 2003/084570; заявку WO 2005/035586; заявку WO 2005/035778; заявку WO 2005/053742; Okazaki et al., J. Mol. Biol. 336:1239-49 (2004); Yamane-Ohnuki et al., Biotech. Bioeng. 87: 614 (2004). Примеры линий клеток, продуцирующих дефукозилированные антитела, включают в себя клетки Lec13 CHO с дефицитом фукозилирования белков (Ripka et al., Arch. Biochem. Biophys. 249:533-45 (1986); заявка на патент США № 2003/0157108 Al., Presta, L; и заявка WO 2004/056312 A1, Adams et al., в частности, в Примере 11), и линии нокаутированных клеток, таких как имеющие ген альфа-1,6-фукозилтрансферазы, нокаутированные клетки CHO FUT8 (Yamane-Ohnuki et al. Biotech. Bioeng. 87:614 (2004)).
Другой тип варианта представляет собой вариант с замещением аминокислот. Эти варианты имеют, по меньшей мере, один остаток аминокислоты в молекуле антитела, замененный другим остатком. Сайты, представляющие наибольший интерес для замещающего мутагенеза, включают в себя гипервариабельные области, но изменения FR также рассматриваются. Консервативные замещения показаны в Таблице 1 под заголовком "предпочтительные замещения". Если такие замещения приводят к изменениям биологической активности, тогда более существенные изменения, обозначенные "примерные замещения" в Таблице 1, или как дополнительно описывается ниже в ссылке на классы аминокислот, могут вводиться, а продукты подвергаются скринингу.
Существенные модификации биологических свойств антитела получаются посредством выбора замещений, которые значительно отличаются по своим воздействиям на поддержание (a) структуры полипептидной основной цепи в области замещения, например, в виде конформации складок или спиралей, (b) заряда или гидрофобности молекулы на целевом сайте, или (c) объема боковой цепи. Существующие в природе остатки делятся на группы на основе их общих свойств боковых цепей:
(1) гидрофобные: норлейцин, met, ala, val, leu, ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислотные: asp, glu;
(4) основные: his, lys, arg;
(5) остатки, которые влияют на ориентацию цепи: gly, pro; и
(6) ароматические: trp, tyr, phe.
Неконсервативные замещения будут содержать замену элемента из одного из этих классов на другой класс.
Один из типов замещающего варианта содержит замещение одного или нескольких остатков гипервариабельной области исходного антитела (например, гуманизированного антитела или антитела человека). Как правило, полученный вариант (варианты), выбранный для дальнейшей разработки, будет иметь улучшенные биологические свойства по сравнению с исходным антителом, из которых они генерируются. Удобный способ генерирования таких замещающих вариантов включает в себя созревание сродства с использованием фагового дисплея. Вкратце, несколько сайтов гипервариабельной области (например, 6-7 сайтов) мутируют с генерированием всех возможных замещений аминокислот на каждом сайте. Антитела, генерируемые таким образом, проявляются из нитеобразных частиц фагов, как слияние с продуктом гена III M13, упакованного внутри каждой частицы. Проявляемые с помощью фагов варианты затем просматриваются на их биологическую активность (например, сродство связывания), как здесь описывается. Для идентификации кандидатов на модификацию среди сайтов гипервариабельной области, может осуществляться мутагенез с аланиновым сканированием для идентификации остатков гипервариабельной области, вносящей значительный вклад в связывание антигена. Альтернативно, или в дополнение к этому, может быть полезным анализ кристаллической структуры комплекса антиген-антитело для идентификации точек контакта между антителом и антигеном. Такие контактные остатки и соседние остатки являются кандидатами на замещение в соответствии с технологиями, разработанными здесь. После того как такие варианты генерируются, панель вариантов подвергается скринингу, как здесь описывается, и антитела с превосходными свойствами при одном или нескольких релевантных анализах могут быть выбраны для дальнейшей разработки.
Молекулы нуклеиновых кислот, кодирующие варианты последовательностей аминокислот антитела, получают посредством разнообразных способов, известных в данной области. Эти способы включают в себя, но, не ограничиваясь этим, выделение из природного источника (в случае существующих в природе вариантов последовательностей аминокислот) или приготовление с помощью опосредуемого олигонуклеотидами (или сайт-направленного) мутагенеза, PCR мутагенеза и кассетного мутагенеза полученного ранее варианта или невариантной версии антитела.
Может быть желательным введение одной или нескольких модификаций аминокислот в области Fc полипептидов иммуноглобулина по настоящему изобретению, тем самым генерируя вариант области Fc. Вариант области Fc может содержать последовательность области Fc человека (например, область Fc IgG1, IgG2, IgG3 или IgG4 человека), содержащую модификацию аминокислот (например, замещения) в одном или нескольких положениях аминокислот, включая положения шарнирного цистеина.
В соответствии с настоящим описанием и концепциями, известными в данной области, предполагается, что в некоторых вариантах осуществления, антитело, используемое в способах по настоящему изобретению, может содержать одно или несколько изменений, по сравнению с эквивалентным антителом дикого типа, например, в области Fc. Эти антитела, тем не менее, должны сохранять по существу такие же характеристики, необходимые для терапевтического применения, по сравнению с их эквивалентом дикого типа. Например, считается, что определенные изменения могут осуществляться в области Fc, что должно привести к изменению (то есть, либо повышению, либо понижению) связывания C1q и/или комплемент-зависимой цитотоксичности (CDC), например, как описывается в заявке WO 99/51642. См. также Duncan & Winter, Nature 322:738-40 (1988); заявку на патент США № 5648260; заявку на патент США № 5624821 и заявку WO94/29351, относительно других примеров вариантов области Fc. Заявка WO 00/42072 (Presta) и заявка WO 2004/056312 (Lowman) описывают варианты антитела с повышенным или пониженным связыванием с FcR. Содержание этих патентных публикаций конкретно включается сюда в виде ссылок. См., также, Shields et al. J. Biol. Chem. 9(2):6591-6604 (2001). Антитела с повышенными половинными временами жизни и улучшенным связыванием с неонатальным рецептором Fc (FcRn), который отвечает за перенос IgG от матери к эмбриону (Guyer et al., J. Immunol. 117:587 (1976) и Kim et al., J. Immunol. 24:249 (1994)), описаны в заявке на патент США № 2005/0014934A1 (Hinton et al.). Эти антитела содержат область Fc с одним или несколькими замещениями в ней, которые улучшают связывание области Fc с FcRn. Полипептидные варианты с измененными последовательностями аминокислот области Fc и повышенной или пониженной способностью к связыванию C1q описываются в заявке на патент США № 6194551B1, в заявке WO99/51642. Содержание этих патентных публикаций конкретно включаются сюда в виде ссылок. См., также, Idusogie et al. J. Immunol. 164:4178-84 (2000).
Производные антител
Антитела по настоящему изобретению могут дополнительно модифицироваться, чтобы они содержали дополнительные небелковые остатки, которые известны в данной области и являются легкодоступными. Предпочтительно, остатки, пригодные для дериватизации антитела, представляют собой водорастворимые полимеры. Неограничивающие примеры водорастворимых полимеров включают в себя, но, не ограничиваясь этим, полиэтиленгликоль (PEG), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпиролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (либо гомополимеры, либо неупорядоченные сополимеры) и декстран или поли(н-винилпиролидон)полиэтиленгликоль, гомополимеры пропиленгликоля, сополимеры полипропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Полиэтиленгликоль пропиональдегид может иметь преимущества при производстве, благодаря его стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Количество полимеров, присоединенных к антителу, может изменяться, и если присоединяется более одного полимера, они могут представлять собой одинаковые или различные молекулы. Как правило, количество и/или тип полимеров, используемых для дериватизации, может определяться на основе соображений, включающих в себя, но, не ограничиваясь этим, конкретные свойства или функции антитела, которые должны улучшаться, будет ли производное антитела использоваться в терапии при определенных условиях и тому подобное.
Скрининг антител с желаемыми свойствами
Антитела по настоящему изобретению связывают EGFL7 и в некоторых вариантах осуществления могут модулировать один или несколько аспектов эффектов, связанных с EGFL7, включая, но, не ограничиваясь этим, разрушение любого биологически релевантного биологического пути EGFL7 и/или лечение и/или предотвращение появления опухоли, пролиферативного клеточного расстройства или рака; и/или лечение или предотвращение расстройства, связанного с экспрессированием и/или активностью EGFL7 (такого как повышенного экспрессирования и/или повышенной активности EGFL7). Например, антитела по настоящему изобретению могут просматриваться на их способность к блокированию адгезии клеток HUVEC и EGFL7 и миграции HUVEC на планшетах, покрытых белком EGFL7, как здесь описывается.
Очищенные антитела могут дополнительно характеризоваться с помощью ряда анализов, включая, но, не ограничиваясь этим, N-концевое секвенирование, анализ аминокислот, неденатурирующую эксклюзионную высокоэффективную жидкостную хроматографию (ВЭЖХ), масс-спектрометрию, ионообменную хроматографию и папаиновый гидролиз.
В определенных вариантах осуществления настоящего изобретения, антитела, продуцируемые здесь, анализируются на их биологическую активность. В некоторых вариантах осуществления, антитела по настоящему изобретению исследуются на их активность при связывании антигена. Анализы связывания антигена, которые известны в данной области и могут использоваться здесь, включают в себя, но, не ограничиваясь этим, любые анализы прямого или конкурентного связывания с использованием таких технологий, как вестерн блот, радиоиммуннологические анализы, ELISA (иммуносорбентный анализ с помощью связанного фермента), иммунные "сэндвич" анализы, анализы иммунопреципитации, флюоресцентные иммунные анализы и иммунные анализы с помощью белка A. Иллюстративные анализы связывания антигенов приводятся ниже в секции Примеры.
В некоторых вариантах осуществления, настоящее изобретение предоставляет антитело анти-EGFL7, которое конкурирует с антителом, содержащим вариабельный домен легкой цепи, содержащей последовательность, выбранную из SEQ ID NO: 1 и SEQ ID NO: 3, и вариабельный домен тяжелой цепи, содержащей последовательность, выбранную из SEQ ID NO: 2 и SEQ ID NO: 4, за связывание EGFL7. Такие конкурентные антитела могут быть получены посредством скрининга супернатантов гибридомы анти-EGFL7 на связывание с иммобилизованным EGFL7 при конкуренции с меченым антителом, содержащим вариабельный домен легкой цепи, содержащей последовательность, выбранную из SEQ ID NO: 1 и SEQ ID NO: 3, и вариабельный домен тяжелой цепи, содержащей последовательность, выбранную из SEQ ID NO: 2 и SEQ ID NO: 4. Такие конкурентные антитела включают в себя антитела, которые распознают эпитоп EGFL7, который является таким же, как эпитоп EGFL7, распознаваемый антителом, или перекрывается с ним. Супернатант гибридомы, содержащей конкурентное антитело, будет уменьшать количество связанного, меченого антитела, детектируемого в рассматриваемой смеси с конкурентным связыванием, по сравнению с количеством связанного меченого антитела, детектируемого в контрольной смеси со связыванием, содержащей иррелевантное антитело (или не содержащей ничего). Любые анализы конкурентного связывания, описанные здесь, являются пригодными для использования в указанной выше процедуре.
Антитела анти-EGFL7 по настоящему изобретению, обладающие свойствами, описанными здесь, могут быть получены посредством скрининга клонов гибридомы анти-EGFL7 на желаемые свойства с помощью любого обычного способа. Например, если является желаемым моноклональное антитело анти-EGFL7, которое конкурирует или не конкурирует за связывание EGFL7 с антителом, содержащим вариабельный домен легкой цепи, содержащей последовательность, выбранную из SEQ ID NO: 1 и SEQ ID NO: 3, и вариабельный домен тяжелой цепи, содержащей последовательность, выбранную из SEQ ID NO: 2 и SEQ ID NO: 4, антитело-кандидат может исследоваться в анализе конкурентного связывания. Конкурентные анализы хорошо известны в данной области.
Другие функциональные анализы для определения ингибиторной способности антитела анти-EGFL7 известны в данной области, некоторые из них иллюстрируются здесь.
В некоторых вариантах осуществления, настоящее изобретение предполагает измененные антитела, которые обладают некоторыми, но не всеми эффекторными функциями, которые делают их желательными кандидатами для множества применений, в которых является важным половинное время жизни антитела in vivo, а некоторые эффекторные функции (такие как комплементы и ADCC) являются необязательными или вредными. В некоторых вариантах осуществления, активности Fc полученного иммуноглобулина измеряются, чтобы быть уверенным, что поддерживаются только желаемые свойства. Анализы цитотоксичности in vitro и/или in vivo могут осуществляться для подтверждения уменьшения/обеднения активностей CDC и/или ADCC. Например, могут осуществляться анализы связывания рецептора Fc (FcR), чтобы быть уверенным, что у антитела нет связывания FcγR (следовательно, вероятно, отсутствует активность ADCC), но оно сохраняет способность к связыванию FcRn. Первичные клетки для опосредования ADCC, клетки NK экспрессируют только FcRIII, в то время как моноциты экспрессируют FcRI, FcRII и FcRIII. Экспрессия FcR в гематопоэтических клетках приводится в Таблице 3 на странице 464 Ravetch & Kinet, Annu. Rev. Immunol. 9:457-92 (1991). Пример анализа in vitro для оценки активности ADCC молекулы, представляющей интерес, описывается в заявке на патент США № 5500362 или 5821337. Эффекторные клетки, пригодных для таких анализов, включают в себя мононуклеарные клетки периферической крови (PBMC) и клетки природных киллеров (NK). Альтернативно, или в дополнение к этому, активность ADCC молекулы, представляющей интерес, может оцениваться in vivo, например, на животной модели, такой как та, которая описывается в Clynes et al. Proc. Natl. Acad. Sci. USA 95:652-656 (1998). Анализы связывания C1q также могут осуществляться для подтверждения того, что антитело неспособно связывать C1q и, следовательно, оно не имеет активности CDC. Для оценки активирования комплемента может осуществляться анализ CDC, например, как описывается в Gazzano-Santoro et al., J. Immunol. Meth. 202:163 (1996). Определения связывания FcRn и выведения/половинного времени жизни также могут осуществляться in vivo с использованием способов, известных в данной области.
В некоторых вариантах осуществления, настоящее изобретение предоставляет измененные антитела, которые обладают повышенными эффекторными функциями и/или увеличенным половинным временем жизни.
Векторы, клетки-хозяева и рекомбинантные способы
Для рекомбинантного продуцирования антитела по настоящему изобретению, нуклеиновая кислота, кодирующая его, выделяется и вставляется в реплицируемый вектор для дальнейшего клонирования (амплификации ДНК) или для экспрессирования. ДНК, кодирующая антитело, легко выделяется и секвенируется с использованием обычных процедур (например, посредством использования олигонуклеотидных зондов, которые способны специфично связывать гены, кодирующие тяжелые и легкие цепи антитела). Доступно множество векторов. Выбор вектора частично зависит от клетки-хозяина, которая должна использоваться. Как правило, предпочтительные клетки-хозяева имеют либо прокариотическое, либо эукариотическое происхождение (как правило, от млекопитающих). Будет ясно, что константные области любого изотипа могут использоваться для этой цели, включая константные области IgG, IgM, IgA, IgD и IgE, и, что такие константные области могут быть получены от человека или любых видов животных.
a. генерирование антител с использованием прокариотических клеток-хозяев:
i. Конструирование вектора
Полинуклеотидные последовательности, кодирующие полипептидные компоненты антитела по настоящему изобретению, могут быть получены с использованием стандартных рекомбинантных технологий. Желаемые полинуклеотидные последовательности могут выделяться и секвенироваться из клеток, продуцирующих антитела, таких как клетки гибридомы. Альтернативно, полинуклеотиды могут синтезироваться с использованием технологий синтезатора нуклеотидов или PCR. После получения, последовательности, кодирующие полипептиды, вставляются в рекомбинантный вектор, способный реплицировать и экспрессировать гетерологичные полинуклеотиды в прокариотических хозяевах. Множество векторов, которые являются доступными и известными в данной области, может использоваться для целей настоящего изобретения. Выбор соответствующего вектора будет зависеть в основном от размера нуклеиновых кислот, которые должны вставляться в вектор, и от конкретной клетки-хозяина, которая должна трансформироваться с помощью вектора. Каждый вектор содержит различные компоненты, в зависимости от его функции (амплификации или экспрессирования гетерологичного полинуклеотида, или как того, так и другого) и от его совместимости с конкретной клеткой-хозяином, в которой он находится. Компоненты вектора, как правило, включают в себя, но, не ограничиваясь этим: источник реплицирования, маркерный ген селекции, промотор, сайт связывания рибосом (RBS), сигнальную последовательность, вставку гетерологичной нуклеиновой кислоты и последовательность окончания транскрипции.
Как правило, векторы плазмиды, содержащие репликон и контрольные последовательности, которые получают из видов, совместимых с клеткой-хозяином, используются в связи с этими хозяевами. Вектор обычно несет сайт репликации, а также маркирующие последовательности, которые способны обеспечить фенотипическую селекцию в трансформируемых клетках. Например, E.coli обычно трансформируется с использованием pBR322, плазмиды, полученной от видов E.coli. pBR322 содержит гены, кодирующие стойкость к ампициллину (Amp) и тетрациклину (Tet), и таким образом обеспечивает простые средства для идентификации трансформированных клеток. pBR322, ее производные или другие микробные плазмиды или бактериофаги могут также содержать промоторы или модифицироваться так, чтобы они содержали промоторы, которые могут использоваться организмом микроба для экспрессирования эндогенных белков. Примеры производных pBR322, используемых для экспрессии конкретных антител, подробно описаны в Carter et al., заявке на патент США № 5648237.
В дополнение к этому, фаговые векторы, содержащие репликон и контрольные последовательности, которые совместимы с микроорганизмом хозяином, могут использоваться в качестве трансформирующих векторов в соединении с этими хозяевами. Например, бактериофаг, такой как λGEMTM-11 может использоваться при получении рекомбинантного вектора, который может использоваться для преобразования чувствительных клеток-хозяев, таких как LE392 E.coli.
Вектор экспрессии по настоящему изобретению может содержать две или более пар промотор-цистрон, кодирующих каждый из полипептидных компонентов. Промотор представляет собой нетранслируемую регуляторную последовательность, расположенную перед (5') цистроном, который модулирует ее экспрессию. Прокариотические промоторы, как правило, попадают в два класса, индуцируемые и конститутивные. Индуцируемый промотор представляет собой промотор, который инициирует увеличение уровней транскрипции цистрона под его контролем в ответ на изменения в состоянии культуры, например присутствие или отсутствие питания или изменение температуры.
Большое количество промоторов, распознаваемых разнообразными потенциальными клетками-хозяевами, хорошо известно. Выбранный промотор может функционально связываться с ДНК цистрона, кодирующего легкую или тяжелую цепь, посредством удаления промотора из источника ДНК посредством гидролиза фермента рестрикции и вставки выделенной промоторной последовательности в вектор по настоящему изобретению. Как нативная промоторная последовательность, так и множество гетерологичных промоторов могут использоваться для прямой амплификации и/или экспрессии целевых генов. В некоторых вариантах осуществления, используются гетерологичные промоторы, поскольку они, как правило, делают возможной большую транскрипцию и более высокие выходы экспрессируемого целевого гена по сравнению с нативным целевым полипептидным промотором.
Промоторы, пригодные для использования вместе с прокариотическими хозяевами, включают в себя промоторные системы на основе промотора phoA, β-галактамазы и лактозы, промоторную систему на основе триптофана (trp) и гибридные промоторы, такие как промотор tac или trc. Однако другие промоторы, которые являются функциональными в бактериях (такие как другие известные бактериальные или фаговые промоторы) также являются пригодными для использования. Их нуклеотидные последовательности опубликованы, что дает возможность специалисту в данной области функционально лигировать их с цистронами, кодирующих целевые легкие и тяжелые цепи (Siebenlist et al., Cell 20:269 (1980)), с использованием линкеров или адапторов для получения любых необходимых сайтов рестрикции.
В одном из аспектов настоящего изобретения, каждый цистрон в рекомбинантном векторе содержит компонент сигнальной последовательности секреции, который направляет транслокацию экспрессируемых полипептидов по мембране. Как правило, сигнальная последовательность может представлять собой компонент вектора или может представлять собой часть ДНК целевого полипептида, которая вставляется в вектор. Сигнальная последовательность, выбранная для цели настоящего изобретения, должна быть такой, которая распознается и процессируется (то есть расщепляется с помощью сигнальной петидазы) клеткой-хозяином. Для прокариотических клеток-хозяев, которые не распознают и не процессируют сигнальные последовательности, нативные для гетерологичных полипептидов, сигнальная последовательность замещается прокариотической сигнальной последовательностью, выбранной, например, из группы, состоящей из щелочной фосфатазы, пенициллиназы, Ipp или термически стабильных лидеров на основе энтеротоксина II (STII), LamB, PhoE, PelB, OmpA и MBP.
В одном из вариантов осуществления настоящего изобретения, сигнальные последовательности, используемые в обоих цистронах системы экспрессии, представляют собой сигнальные последовательности STII или их варианты.
В другом аспекте, продуцирование иммуноглобулинов в соответствии с настоящим изобретением может осуществляться в цитоплазме клетки-хозяина, и по этой причине оно не требует присутствия сигнальных последовательностей секреции в каждом цистроне. В этой связи, легкие и тяжелые цепи иммуноглобулина экспрессируются, складываются и собираются с образованием функциональных иммуноглобулинов в цитоплазме. Определенные штаммы хозяев (например, штаммы E.coli trxB') обеспечивают в цитоплазме условия, которые являются благоприятными для формирования дисульфидных связей, тем самым делая возможным соответствующее складывание и сборку экспрессируемых субъединиц белка. Proba & Pluckthun, Gene 159:203 (1995).
Прокариотические клетки-хозяева, пригодные для экспрессирования антител по настоящему изобретению, включают в себя Archaebacteria и Eubacteria, такие как грамотрицательные или грамположительные организмы. Примеры пригодных для использования бактерий включают в себя Escherichia (например, E.coli), Bacilli (например, B.subtilis), Enterobacteria, виды Pseudomonas (например, P.aeruginosa), Salmonella typhimurium, Serratia marcescans, Klebsiella, Proteus, Shigella, Rhizobia, Vitreoscilla или Paracoccus. В некоторых вариантах осуществления, используются грамотрицательные клетки. В некоторых вариантах осуществления, клетки E.coli используются в качестве хозяев для настоящего изобретения. Примеры штаммов E.coli включают в себя штамм W3110 (Bachmann, Cellular and Molecular Biology, vol. 2 (Washington, D.C.: American Society for Microbiology, 1987), pp. 1190-1219; ATCC® Deposit No. 27,325) и его производные, включая штамм 33D3, имеющий генотип W3110 ΔfhuA (ΔtonA) ptr3 lac Iq lacL8 ΔompTA (nmpc-fepE) degP41 kanR (заявка на патент США № 5639635). Другие штаммы и их производные, такие как E.coli 294 (ATCC® 31,446), E.coli B, E.coli л 1776 (ATCC® 31,537) и E.coli RV308(ATCC® 31,608) также являются пригодными. Эти примеры являются скорее иллюстративными, чем ограничивающими. Способы конструирования производных любой из рассмотренных выше бактерий, имеющих определенные генотипы, известны в данной области и описываются, например, в Bass et al., Proteins 8:309-14 (1990). Как правило, необходимо выбирать соответствующие бактерии, принимая во внимание реплицируемость репликона в клетках бактерий. Например, виды E.coli, Serratia или Salmonella могут соответствующим образом использоваться в качестве хозяев, когда хорошо известные плазмиды, такие как pBR322, pBR325, pACYC177, или pKN410, используются для получения репликона. Как правило, клетка-хозяин должна секретировать минимальные количества протеолитических ферментов, и желательно, чтобы дополнительные ингибиторы протеазы включались в клеточную культуру.
ii. Продуцирование антител
Клетки-хозяева трансформируются с помощью описанных выше векторов экспрессии и культивируются в обычных питательных средах, модифицированных соответствующим образом для индуцирования промоторов, селекции трансформантов или амплификации генов, кодирующих желаемые последовательности.
Средства трансформации вводят ДНК в прокариотического хозяина, так что ДНК может реплицироваться, либо как экстрахромосомальный элемент, либо с помощью хромосомального интегранта. В зависимости от используемой клетки-хозяина, трансформирование осуществляется с использованием стандартных технологий, соответствующих таким клеткам. Обработка кальцием с использованием хлорида кальция, как правило, используется для бактериальных клеток, которые содержат достаточные барьеры клеточных стенок. Другой способ трансформирования использует полиэтиленгликоль/ДМСО. Еще одна используемая технология представляет собой электропорацию.
Прокариотические клетки, используемые для получения полипептидов по настоящему изобретению, выращиваются в средах, известных в данной области и пригодных для культивирования выбранных клеток-хозяев. Примеры соответствующих сред включают в бульон Луриа (LB) плюс необходимые питательные добавки. В некоторых вариантах осуществления, среды также содержат агент селекции, выбранный на основе конструкции вектора экспрессии, чтобы селективно давать возможность для роста прокариотических клеток, содержащих вектор экспрессии. Например, ампициллин добавляют в среды для роста клеток, экспрессирующих ген, устойчивый к ампициллину.
Любые необходимые добавки, кроме источников углерода, азота и неорганических фосфатов, также могут включаться при соответствующей концентрации, вводимые по одной или как смесь с другой добавкой или средой, такой как комплексный источник азота. Необязательно, среда для культивирования может содержать один или несколько восстанавливающих агентов, выбранных из группы, состоящей из глютатиона, цистеина, цистамина, тиогликолята, дитиоэритритола и дитиотреитола.
Прокариотические клетки-хозяева культивируют при соответствующих температурах. Для роста E.coli, например, как правило, используют температуру в пределах от примерно 20°C до примерно 39°C, как правило, от примерно 25°C до примерно 37°C, например, примерно 30°C. pH среды может представлять собой любой pH, находящийся в пределах от примерно 5 до примерно 9, он зависит в основном от организма-хозяина. Для E.coli, pH составляет, как правило, от примерно 6,8 до примерно 7,4, а, как правило, примерно, 7,0.
Если в векторе экспрессии по настоящему изобретению используется индуцируемый промотор, экспрессия белка индуцируется при условиях, пригодных для активирования промотора. В одном из аспектов настоящего изобретения, промоторы phoA используют для контроля транскрипции полипептидов. Соответственно, трансформируемые клетки-хозяева культивируют в среде с ограничением фосфатного субстрата для индуцирования. Среда с ограничением фосфатного субстрата представляет собой, как правило, среду C.R.A.P (см., например, Simmons et al., J. Immunol. Meth. 263:133-47 (2002)). Могут использоваться разнообразные другие индукторы, в соответствии с используемым конструктом вектора, как известно в данной области.
В одном из вариантов осуществления, экспрессируемые полипептиды по настоящему изобретению секретируются в периплазме клеток-хозяев и извлекаются из нее. Извлечение белков, как правило, включает в себя разрушение микроорганизма, как правило, с помощью таких средств как осмотический шок, озвучивание или лизис. После разрушения клеток, остатки клеток или цельные клетки могут удаляться с помощью центрифугирования или фильтрования. Белки могут дополнительно очищаться, например, с помощью аффинной хроматографии на смоле. Альтернативно, белки могут переноситься в культурные среды и выделяться из них. Клетки могут удаляться из культуры, и супернатант культуры фильтруется и концентрируется для дальнейшей очистки полученных белков. Экспрессируемые полипептиды могут дополнительно выделяться и идентифицироваться с использованием повсеместно известных способов, таких как электрофорез на полиакриламидном геле (PAGE) и анализ вестерн блот.
В одном из аспектов настоящего изобретения, получение антител осуществляется в больших количествах с помощью способа ферментирования. Различные промышленные загрузочные процедуры ферментирования являются доступными для получения рекомбинантных белков. Промышленные ферментеры имеют емкость, по меньшей мере, 1000 литров, предпочтительно, емкость от примерно 1000 до 100000 литров. Эти ферментеры используют перемешивающие лопасти для распределения кислорода и питательных веществ, в частности, глюкозы (обычного источника углерода/энергии). Ферментирование малого масштаба, как правило, относится к ферментированию в ферментере, который имеет емкость не больше приблизительно 100 литров по объему, и она может находиться в пределах от примерно 1 литра до примерно 100 литров.
В способе ферментирования, индуцирование экспрессии белков, как правило, инициируется после того, как клетки выращиваются при соответствующих условиях до получения желаемой плотности, например, до значения OD550 примерно 180-220, на этой стадии клетки находятся в ранней стационарной фазе. Могут использоваться разнообразные индукторы, в соответствии с используемым конструктом вектора, как известно в данной области и описывается выше. Клетки могут выращиваться в течение более коротких периодов перед индуцированием. Клетки обычно индуцируются примерно в течение 12-50 часов, хотя может использоваться более продолжительное или более короткое время индуцирования.
Для улучшения промышленного выхода и качества полипептидов по настоящему изобретению, могут модифицироваться различные условия ферментирования. Например, для улучшения правильной сборки и складывания секретируемых полипептидов антитела, могут использоваться дополнительные векторы, осуществляющие сверхэкспрессию белков шаперонов, таких как белки Dsb (DsbA, DsbB, DsbC, DsbD и/или DsbG) или FkpA (пептидилпролил цис, транс изомераза с шаперонной активностью) для совместного трансформирования прокариотических клеток-хозяев. Белки шапероны, как продемонстрировано, облегчают правильное складывание и растворимость гетерологичных белков, производимых в бактериальных клетках-хозяевах. Chen et al., J. Biol Chem. 274:19601-05 (1999); Georgiou et al., заявка на патент США № 6083715; Georgiou et al., заявка на патент США № 6027888; Bothmann & Pluckthun, J. Biol Chem. 275:17100-05 (2000); Ramm & Pluckthun, J. Biol. Chem. 275:17106-13 (2000); Arie et al., Molec. Microbiol. 39:199-210 (2001).
Для сведения к минимуму протеолиза экспрессируемых гетерологичных белков (в особенности, тех, которые являются протеолитически чувствительными), определенные штаммы хозяев, дефицитных относительно протеолитических ферментов, могут использоваться для настоящего изобретения. Например, штаммы клеток-хозяев могут модифицироваться для осуществления генетической мутации (мутаций) в генах, кодирующих известные бактериальные протеазы, такие как Протеаза III, OmpT, DegP, Tsp, Протеаза I, Протеаза Мi, Протеаза V, Протеаза VI и их сочетания. Некоторые протеаза-дефицитные штаммы E.coli являются доступными и описываются, например, в Joly et al. (1998), выше; Georgiou et al., в заявке на патент США № 5264365; Georgiou et al., в заявке на патент США № 5508192; Hara et al., Microbial Drug Resistance 2:63-72 (1996).
В одном из вариантов осуществления, штаммы E.coli, дефицитные относительно протеолитических ферментов и трансформированные с помощью плазмид, осуществляющих сверхэкспрессию одного или нескольких белков шаперонов, используют как клетки-хозяева для системы экспрессии по настоящему изобретению.
iii. Очистка антител
Могут использоваться стандартные способы очистки белков, известные в данной области. Следующие процедуры представляют собой примеры соответствующих процедур очистки: фракционирование на иммуноаффинных или ионообменных колонках, преципитация в этаноле, ВЭЖХ с обращенной фазой, хроматография на силикагеле или на катионообменной смоле, такой как DEAE, хроматофокусирование, SDS-PAGE, аммонийсульфатная преципитация и гель-фильтрация с использованием, например, Sephadex G-75.
В одном из аспектов, белок A, иммобилизованный на твердой фазе, используется для иммунноаффинной очистки продуктов полноразмерных антител по настоящему изобретению. Белок A представляет собой белок клеточной стенки с молекулярной массой 41 кДа из Staphylococcus aureas, который связывается с высоким сродством с областью Fc антител. Lindmark et al., J. Immunol. Meth. 62:1-13 (1983). Твердая фаза, на которой иммобилизуется белок A, предпочтительно представляет собой колонку, содержащую поверхность стекла или окиси кремния, более предпочтительно, стеклянную колонку с контролируемыми размерами пор или колонку из кремниевой кислоты. В некоторых применениях, колонку покрывают реагентом, таким как глицерин, в попытке предотвратить неспецифичную адгезию примесей.
В качестве первой стадии очистки, препарат, полученный из культуры клеток, как описывается выше, наносится на твердую фазу с иммобилизованным белком A, чтобы сделать возможным специфичное связывание антитела, представляющего интерес, с белком A. Затем твердую фазу промывают для удаления примесей, неспецифически связанных с твердой фазой. Наконец, антитело, представляющее интерес, извлекается с твердой фазы посредством элюирования.
b. Генерирование антител с использованием эукариотических клеток-хозяев:
Компоненты вектора, как правило, включают в себя, но, не ограничиваясь этим, один или несколько из следующих компонентов: сигнальную последовательность, источник репликации, один или несколько маркерных генов, энхансерный элемент, промотор и последовательность прерывания транскрипции.
(i) Компонент сигнальной последовательности
Вектор для использования в эукариотической клетке-хозяине может также содержать сигнальную последовательность или другой полипептид, имеющий специфичный сайт расщепления на N-конце зрелого белка или полипептида, представляющего интерес. Гетерологичная сигнальная последовательность, выбираемая предпочтительно, представляет собой последовательность, которая распознается и процессируется (то есть расщепляется под действием сигнальной петидазы) клеткой-хозяином. При экспрессии в клетках млекопитающих, являются доступными сигнальные последовательности млекопитающих, а также вирусные секреторные лидеры, например сигнал gD от простого герпеса.
ДНК для такой предшествующей области лигируется в рамке считывания с ДНК, кодирующей антитело.
(ii) Источник репликации
Как правило, источник компонента репликации не является необходимым для векторов экспрессии млекопитающих. Например, источник SV40, как правило, может использоваться только потому, что он содержит ранний промотор.
(iii) Компонент селекции гена
Векторы экспрессии и клонирования могут содержать ген селекции, также называемый селектируемый маркер. Типичные гены селекции кодируют белки, которые (a) придают стойкость к антибиотикам или другим токсинам, например ампициллину, неомицину, метотрексату или тетрациклину, (b) дополняют ауксотрофные дефициты, где это необходимо, или (c) доставляют критические питательные вещества, недоступные из сложных питательных сред.
Один из примеров схемы селекции использует лекарственное средство для приостановки роста клетки-хозяина. Те клетки, которые успешно трансформируются с помощью гетерологичного гена, производят белок, придающий стойкость к лекарственному средству, и таким образом переживают режим селекции. Примеры такой доминантной селекции используют лекарственные средства неомицин, микофенольную кислоту и гигромицин.
Другой пример пригодных для использования маркеров селекции для клеток млекопитающих представляют собой те маркеры, которые делают возможной идентификацию клеток, компетентных для поглощения нуклеиновой кислоты антитела, такие как DHFR, тимидин киназа, металлотионеин-I и -II, предпочтительно, гены металлотионеина приматов, аденозин деаминаза, орнитин декарбоксилаза и тому подобное.
Например, клетки, трансформируемые с помощью гена селекции DHFR, сначала идентифицируют посредством культивирования всех трансформантов в культурной среде, которая содержит метотрексат (Mtx), конкурентный антагонист DHFR. Соответствующая клетка-хозяин, когда используется DHFR дикого типа, представляет собой линию клеток яичников китайского хомячка (CHO) с дефицитом активности DHFR (например, ATCC® CRL-9096).
Альтернативно, клетки-хозяева (в частности, хозяева дикого типа, которые содержат эндогенные DHFR), трансформируемые или совместно трансформируемые с помощью последовательностей ДНК, кодирущей антитело, белка DHFR дикого типа и другого маркера селекции, такого как аминогликозид 3'-фосфотрансфераза (APH), могут выбираться посредством роста клеток в среде, содержащей агент селекции для маркера селекции, такого как аминогликозидный антибиотик, например канамицин, неомицин или G418. См. заявку на патент США № 4965199.
(iv) Компонент промотора
Векторы экспрессии и клонирования обычно содержат промотор, который распознается организмом хозяином и функционально связан с нуклеиновой кислотой полипептида антитела. Промоторные последовательности известны для эукариотов. По существу все эукариотические гены имеют обогащенную AT область, расположенную приблизительно на 25-30 оснований впереди сайта, где инициируется транскрипция. Другая последовательность находится на 70-80 оснований впереди начала транскрипции множества генов, представляет собой область CNCAAT (SEQ ID NO: 19), где N может представлять собой любой нуклеотид. На 3' конце большинства эукариотических генов находится последовательность AATAAA (SEQ ID NO: 20), которая может представлять собой сигнал для добавления хвоста из поли-A к 3' концу кодирующей последовательности. Все эти последовательности соответствующим образом вставляются в эукариотические векторы экспрессии.
Транскрипция полипептидов антитела из векторов в клетках-хозяевах млекопитающих контролируется, например, посредством промоторов, полученных из геномов вирусов, таких как вирус полиомы, вирус птичьей оспы, аденовирус (такой как аденовирус 2), вирус бычьей папилломы, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита-B и вирус обезьян 40 (SV40), от гетерологичных промоторов млекопитающих, например промотора актина или промотора иммуноглобулина, от промоторов теплового шока, при условии, что такие промоторы являются совместимыми с системами клеток-хозяев.
Ранние и поздние промоторы вируса SV40 получают соответствующим образом, как фрагмент рестрикции SV40, который также содержит вирусный источник репликации SV40. Непосредственный ранний промотор цитомегаловируса человека соответствующим образом получают как фрагмент рестрикции HindIII E. Система для экспрессирования ДНК у хозяев млекопитающих с использованием вируса бычьей папилломы как вектора описывается в заявке на патент США № 4419446. Модификация этой системы описывается в заявке на патент США № 4601978. Альтернативно, повторяющаяся единица длинного окончания вируса саркомы Роуза может использоваться как промотор.
(v) Компонент элемента энхансера
Транскрипция ДНК, кодирующей полипептид антитела по настоящему изобретению, с помощью высших эукариотов часто повышается при вставке последовательности энхансера в вектор. Сегодня известно множество последовательностей энхансеров из генов млекопитающих (глобин, эластаза, альбумин, α-фетопротеин и инсулин). Как правило, однако, будут использоваться энхансер из вируса эукариотической клетки. Примеры включают в себя энхансер SV40 на поздней стороне источника репликации (bp 100-270), энхансер раннего промотора цитамегаловируса, энхансер полиомы на поздней стороне источника репликации и энхансеры аденовируса. См. также Yaniv, Nature 297:17-18 (1982) относительно элементов энхансеров для активирования эукариотических промоторов. Энхансер может сплайсироваться в вектор в положении 5' или 3' по отношению к последовательности, кодирующей полипептиды антитела, но предпочтительно находится на сайте 5' промотора.
(vi) Компонент прерывания транскрипции
Векторы экспрессии, используемые в эукариотических клетках-хозяевах, как правило, будут также содержать последовательности, необходимые для прерывания транскрипции и для стабилизации мРНК. Такие последовательности являются повсеместно доступными из 5', а при случае, и 3' нетранслируемых областей эукариотических или вирусных ДНК или цДНК. Эти области содержат сегменты нуклеотидов, транскриптируемые как полиаденилированные фрагменты в нетранслируемой части мРНК, кодирующей антитело. Один из пригодных для использования компонентов прерывания транскрипции представляет собой область полиаденилирования бычьего гормона роста. См. заявку WO 94/11026 и вектор экспрессии, описанный там.
(vii) Селекция и трансформация клеток-хозяев
Соответствующие клетки-хозяева для клонирования или экспрессирования ДНК в векторах включают в себя здесь клетки высших эукариотов, описанных здесь, включая клетки-хозяева позвоночных. Размножение клеток позвоночных в культуре (культуре тканей) становится рутинной процедурой. Примеры пригодных для использования линий клеток-хозяев млекопитающих представляют собой линию CV1 почек обезьян, трансформированную с помощью SV40 (COS-7, ATCC® CRL 1651); линию клеток почки эмбриона человека (клетки 293 или клетки 293, субклонированные для роста в культуре суспензии, Graham et al., J. Gen. Virol. 36:59 (1977)); клетки почки детеныша хомячка (BHK, ATCC® CCL 10); клетки яичников китайского хомячка/-DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77:4216 (1980)); клетки Сертоли мыши (TM4, Mather, Biol. Reprod. 23:243-51 (1980)); клетки почки обезьяны (CV1 ATCC® CCL 70); клетки почки Африканской зеленой мартышки (VERO-76, ATCC® CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC® CCL 2); клетки почек собаки (MDCK, ATCC® CCL 34); клетки печени крысы Buffalo (BRL 3A, ATCC® CRL 1442); клетки легких человека (W138, ATCC® CCL 75); клетки печени человек (Hep G2, HB 8065); клетки опухоли грудных желез мыши (MMT 060562, ATCC® CCL51); клетки TRI (Mather et al., Annals N. Y. Acad. Sci. 383:44-68 (1982)); клетки MRC 5; клетки FS4; и линия клеток гепатомы человека (Hep G2).
Клетка-хозяева трансформируются с помощью описанных выше векторов экспрессии или клонирования для продуцирования антител и культивируются в обычных питательных средах, модифицированных соответствующим образом, для индуцирования промоторов, селекции трансформантов или амплификации генов, кодирующих желательные последовательности.
(viii) Культивирование клеток-хозяев
Клетки-хозяева, используемые для продуцирования антитела по настоящему изобретению, могут культивироваться в разнообразных средах. Коммерчески доступные среды, такие как среда Хэма F10 (Sigma), минимальная эссенциальная среда ((MEM), (Sigma), RPMI-1640 (Sigma) и модифицированная по Дульбекко среда Игла ((DMEM), Sigma) являются пригодными для культивирования клеток-хозяев. В дополнение к этому, любая из сред, описанных в Ham et al., Meth. Enzymol 58:44 (1979), Barnes et al., Anal. Biochem. 102:255 (1980), в заявках на патенты США № 4767704; 4657866; 4927762; 4560655; или 5122469; в заявках WO 90/03430; WO 87/00195; или заявке на патент США Re. 30985 могут использоваться в качестве культурных сред для клеток-хозяев. Любая из этих сред может быть дополнена, если это необходимо, гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста), солями (такими как хлорид и фосфат натрия, кальция, магния), буферами (такими как HEPES), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как лекарственное средство GENTAMYCINTM), микроэлементами (определяемыми как неорганические соединения, присутствующие обычно при конечных концентрациях в микромолярном диапазоне) и глюкозой или эквивалентным источником энергии. Любые другие необходимые добавки могут также включаться при соответствующих концентрациях, которые должны быть известны специалистам в данной области. Условия культивирования, такие как температура, pH и тому подобное, являются такими же, как использовалось ранее для клеток-хозяев, выбранных для экспрессии, и будут понятны специалистам в данной области.
(ix) Очистка антитела
При использовании рекомбинантных технологий, антитело может продуцироваться внутриклеточно или непосредственно секретироваться в среду. Если антитело продуцируется внутриклеточно, на первой стадии, взвешенные частицы, либо клетки-хозяева, либо их лизированные фрагменты, удаляются, например, посредством центрифугирования или ультрафильтрации. Когда антитело секретируется в среду, супернатанты от таких систем экспрессии, как правило, сначала концентрируются с использованием коммерчески доступного фильтра для концентрирования белков, например, узла ультрафильтрации Amicon или Millipore Pellicon. Ингибитор протеазы, такой как PMSF, может включаться на одной из указанных выше стадий для ингибирования протеолиза, и антибиотики могут включаться для предотвращения роста случайных примесей.
Композиция антител, полученная из клеток, может очищаться с использованием, например, гидроксилапатитной хроматографии, гель-электрофореза, диализа и аффинной хроматографии, при этом аффинная хроматография является предпочтительной технологией очистки. Пригодность белка A как аффинного лиганда зависит от видов и изотипа любого домена Fc иммуноглобулина, который присутствует в антителе. Белок A может использоваться для очистки антител, которые базируются на тяжелых цепях γ1, γ2 или γ4 человека (Lindmark et al., J. Immunol. Meth. 62:1-13 (1983)). Белок G рекомендуется для всех изотипов мышей и для γ3 человека (Guss et al., ЕMBO J. 5:1567-75 (1986)). Матрица, к которой присоединяется аффинный лиганд, чаще всего представляет собой агарозу, но и другие матрицы являются доступными. Механически стабильные матрицы, такие как стекло с контролируемыми порами или поли(стиролдивинил)бензол, делают возможными более высокие скорости потока и более короткие времена процесса, чем можно достигнуть с помощью агарозы. Когда антитело содержит домен CH3, смола Bakerbond ABXTM (J.T. Baker, Phillipsburg, NJ) является пригодной для очистки. Другие технологии для очистки белка, такие как фракционирование на ионообменной колонке, преципитация в этаноле, ВЭЖХ с обращенной фазой, хроматография на силикагеле, хроматография на гепаринизированной SEPHAROSETM, хроматография на анионо- или катионообменной смоле (например, на колонке с полиаспрагиновой кислотой), хроматофокусировка, SDS-PAGE и аммонийсульфатная преципитация, также являются доступными, в зависимости от антитела, которое должно извлекаться.
После любой предварительной стадии (стадий) очистки, смесь, содержащая антитело, представляющее интерес, и примеси, может подвергаться хроматографии гидрофобных взаимодействий при низких pH с использованием буфера для элюирования, при pH в пределах примерно 2,5-4,5, предпочтительно осуществляемой при низких концентрациях соли (например, от примерно 0 до 0,25M соли).
Иммуноконъюгаты
Настоящее изобретение также предоставляет иммуноконъюгаты (также называемые взаимозаменяемо "конъюгаты антитело-лекарственное средство" или "ADC"), содержащие любое из антител анти-EGFL7, описанных здесь, конъюгированное с цитотоксичным агентом, таким как химиотерапевтический агент, с лекарственным средством, агентом для ингибирования роста, токсином (например, ферментативно активным токсином бактериального, грибкового, растительного или животного происхождения или с его фрагментами), или с радиоактивным изотопом (то есть, радиоактивный конъюгат).
Использование конъюгатов антитело-лекарственное средство для локальной доставки цитотоксических или цитостатических агентов, то есть лекарственных средств для уничтожения или ингибирования роста опухолевых клеток при лечении рака (Syrigos & Epenetos, Anticancer Research 19:605-14 (1999); Niculescu-Duvaz & Springer, Adv. Drug Del. Rev. 26:151-72 (1997); заявка на патент США № 4975278) делает возможной нацеленную доставку остатка лекарственного средства внутрь опухоли и внутриклеточную аккумуляцию в ней, где системное введение таких неконъюгированных лекарственных агентов, которое может привести к возникновению неприемлемых уровней токсичности для нормальных клеток, также как и для клеток опухоли, считается исключенным (Baldwin et al., Lancet pp. (Mar. 15, 1986):603-05 (1986); Thorpe, "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review," in Monoclonal Antibody '84: Biological and Clinical Applications, A. Pinchera et al. (eds.), pp. 475-506 (1985). При этом рассматривается максимальная эффективность при минимальной токсичности. Как поликлональные антитела, так и моноклональные антитела, как сообщается, являются пригодными для этих стратегий (Rowland et al., Cancer Immunol. Immunother. 21:183-87 (1986)). Лекарственные средства, используемые в этих способах, включают в себя дауномицин, доксорубицин, метотрексат и виндезин (Rowland et al., (1986), выше). Токсины, используемые в конъюгатах антитело-токсин, включают в себя бактериальные токсины, такие как токсин дифтерии, растительные токсины, такие как рицин, низкомолекулярные токсины, такие как гельданамицин (Mandler et al., J. Nat. Cancer Inst. 92(19):1573-81 (2000); Mandler et al., Bioorganic &Med. Chem. Letters 10:1025-28 (2000); Mandler et al., Bioconjugate Chem. 13:786-91 (2002)), майтанзиноиды (EP 1391213; Liu et al., Proc. Natl. Acad. Sci. USA 93:8618-8623 (1996)) и калихеамицин (Lode et al., Cancer Res. 58:2928 (1998); Hinman et al., Cancer Res. 53:3336-3342 (1993)). Токсины могут осуществлять свои цитотоксические и цитостатические воздействия посредством механизмов, включающих в себя связывание тубулинов, связывание ДНК или ингибирование топоизомеразы. Некоторые цитотоксические лекарственные средства имеют тенденцию к тому, чтобы становиться неактивными или менее активными, когда конъюгируются с большими антителами или с лигандами белков рецепторов.
ZEVALINTM (ибритумомаб тиуксетан, Biogen/Idec) представляет собой конъюгат антитело - радиоактивный изотоп, состоящий из моноклонального антитела мышей IgG1 каппа, направленного против антигена CD20, находящегося на поверхности нормальных и злокачественных B-лимфоцитов, и радиоактивного изотопа 111In или 90Y, связанного с помощью линкера - хелатирующего агента тиомочевины (Wiseman et al., Eur. Jour. Nucl. Med. 21(1):166-11 (2000); Wiseman et al., Blood 99(12):4336-42 (2002); Witzig et al., J. Clin. Oncol. 20(10):2453-63 (2002); Witzig et al., J. Clin. Oncol. 20(15):3262-69 (2002)). Хотя ZEVALIN имеет активность против неходжкинской лимфомы B-лимфоцитов (NHL), его введение приводит к острым и продолжительным цитопениям у большинства пациентов. MYLOTARGTM (гемтузумаб озогамицин, Wyeth Pharmaceuticals), конъюгат антитела и лекарственного средства, состоящий из антитела huCD33, связанного с калихеамицином, был одобрен в 2000 для лечения острой миелоидной лейкемии посредством инъекции (Drugs of Future 25(7):686 (2000); заявки на патенты США № 4970198; 5079233; 5585089; 5606040; 5693762; 5739116; 5767285; 5773001). Кантузумаб мертанзин (Immunogen, Inc.), конъюгат антитела и лекарственного средства, состоящий из антитела huC242, связанного через дисульфидный линкер SPP с остатком лекарственного средства майтанзиноида, DM1, перешел в испытание Фазы II для лечения раковых заболеваний, которые экспрессируют CanAg, таких как заболевания толстой кишки, поджелудочной железы, желудка и других. MLN-2704 (Millennium Pharm., BZL Biologies, Immunogen Inc.), конъюгат антитела и лекарственного средства, состоящий из моноклонального антитела против антигена, специфичного к мембранам простаты (PSMA), связанного с остатком лекарственного средства майтанзиноида, DM1, разрабатывается для возможного лечения опухолей простаты. Пептиды ауристатина, ауристатин E (AE) и монометилауристатин (MMAE), синтетические аналоги доластатина, конъюгируются с химерными моноклональными антителами cBR96 (специфичными к Lewis Y на карциномах) и cAC10 (специфичными к CD30 при гематологических злокачественных заболеваниях) (Doronina et al., Nature Biotech. 21(7):778-784 (2003)) и находятся в терапевтической разработке.
Химиотерапевтические агенты, пригодные при генерировании иммуноконъюгатов, описываются здесь (например, выше). Ферментативно активные токсины и их фрагменты, которые могут использоваться, включают в себя цепь A дифтерии, несвязывающиеся активные фрагменты токсина дифтерии, цепь A экзотоксина (от Pseudomonas aeruginosa), цепь A рицина, цепь A абрина, цепь A модеццина, альфа-сарцин, белки Aleurites fordii, белки диантины, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор Momordica charantia, курцин, кротин, ингибитор sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены. См., например, заявку WO 93/21232, опубликованную 28 октября, 1993 года. Разнообразные радионуклиды являются доступными для получения радиоактивно конъюгированных антител. Примеры включают в себя 212Bi, 131I, 131In, 90Y и 186Re. Конъюгаты антитела и цитотоксичного агента получают с использованием разнообразных бифункциональных агентов связывания белков, таких как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), иминотиолан (IT), бифункциональные производные сложных имидоэфиров (таких как диметиладипимидат HCl), сложные активные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глютаральдегид), бис-азидо соединения (такие как бис(п-азидобензоил)гександиамин), бис-диазониевые производные (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол 2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицина может быть получен так, как описывается в Vitetta et al., Science 238: 1098 (1987). Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) представляет собой пример хелатирующего агента для конъюгирования радиоактивно меченого нуклеотида с антителом. См. заявку WO 94/11026.
Конъюгаты антитела и одного или нескольких низкомолекулярных токсинов, таких как калихеамицин, майтанзиноиды, доластатины, ауростатины, трихотецен и CC1065, и производные этих токсинов, которые имеют активность токсинов, также рассматриваются здесь.
i. Майтанзин и майтанзиноиды
В некоторых вариантах осуществления, иммунноконъюгат содержит антитело (полноразмерное антитело или его фрагменты) по настоящему изобретению, конъюгированное с одной или несколькими молекулами майтанзиноидов.
Майтанзиноиды представляют собой митотические ингибиторы, которые действуют, ингибируя полимеризацию тубулина. Майтанзин сначала выделяют из Восточноафриканского кустарника Maytenus serrata (заявка на патент США № 3896111). Впоследствии было обнаружено, что некоторые микробы также продуцируют майтанзиноиды, такие как майтанзинол и сложные C-3 эфиры майтанзинолов (заявка на патент США № 4151042). Синтетический майтанзинол и его производные и аналоги описываются, например, в заявках на патенты США № 4137230; 4248870; 4256746; 4260608; 4265814; 4294757; 4307016; 4308268; 4308269; 4309428; 4313946; 4315929; 4317821; 4322348; 4331598; 4361650; 4364866; 4424219; 4450254; 4362663; и 4371533.
Лекарственные остатки майтанзиноидов представляют собой привлекательные лекарственные остатки в конъюгатах антитела и лекарственного средства, поскольку они являются: (i) относительно доступными для получения посредством ферментирования или химической модификации, дериватизации продуктов ферментирования, (ii) доступными для дериватизации с помощью функциональных групп, пригодных для конъюгирования через не-дисульфидные линкеры с антителами, (iii) стабильными в плазме крови и (iv) эффективными против разнообразных линий опухолевых клеток.
Иммуноконъюгаты, содержащие майтанзиноиды, способы их получения, и их терапевтическое использование описываются, например, в заявках на патенты США № 5208020, 5416064 и EP 0 425 235 B1, описание которых тем самым явным образом включаются сюда в виде ссылок. Liu et al., Proc. Natl. Acad. Sci. USA 93:8618-23 (1996) описывает иммуноконъюгаты, содержащие майтанзиноид, обозначенный DM1, связанный с моноклональным антителом C242, направленные против рака толстой и прямой кишки человека. Конъюгат, как обнаружено, является сильно цитотоксичным против культивируемых клеток рака толстой кишки и показывает противоопухолевую активность в анализах роста опухоли in vivo. Chari et al., Cancer Research 52:127-31 (1992) описывают иммуноконъюгаты, в которых майтанзиноид конъюгируется через дисульфидный линкер с антителом мышей A7, связанным с антигеном к линиям клеток рака толстой кишки человека, или с другим моноклональным антителом мышей TA.1, которой связывает онкоген HER-2/neu. Цитотоксичность конъюгата TA.1-майтанзиноид исследовалась in vitro на линии клеток рака груди человека SK-BR-3, которые экспрессируют 3Ч105 поверхностных антигенов HER-2 на клетку. Конъюгат лекарственного средства достигает степени цитотоксичности, сходной с лекарственным средством на основе свободного майтанзиноида, она может быть повышена посредством увеличения количества молекул майтанзиноида на молекулу антитела. Конъюгат A7-майтанзиноид показывает низкую системную цитотоксичность у мышей.
Конъюгаты антитело-майтанзиноид получают посредством химического связывания антитела с молекулой майтанзиноида без значительного уменьшения биологической активности либо антитела, либо молекулы майтанзиноида. См., например, заявку на патент США № 5208020 (описание которого тем самым в явном виде включается в виде ссылки). В среднем 3-4 молекулы майтанзиноида, конъюгируемые на молекулу антитела, показывают эффективность при увеличении цитотоксичности целевых клеток без отрицательного влияния на функции или растворимость антитела, хотя даже одна молекула токсина/антитела, как ожидается, должна увеличивать цитотоксичность по сравнению с использованием голого антитела. Майтанзиноиды хорошо известны в данной области и могут синтезироваться с помощью известных технологий или выделяться из природных источников. Пригодные для использования майтанзиноиды описываются, например, в заявке на патент США № 5208020 и в других патентных и непатентных публикациях, упоминаемых выше. Предпочтительные майтанзиноиды представляют собой майтанзинол и аналоги майтанзинола, модифицированные на ароматическом кольце или на других положениях молекулы майтанзинола, такие как различные сложные эфиры майтанзинола.
Существуют множество связывающих групп, известных в данной области, для получения конъюгатов антитело-майтанзиноид, включая, например, группы, описанные в заявке на патент США № 5208020 или в EP 0425235 B1, Chari et al., Cancer Research 52:127-131 (1992), и в заявке на патент US2005/0169933A1, описания которых тем самым в явном виде включаются сюда в виде ссылок. Конъюгаты антитело-майтанзиноид, содержащие компонент линкера SMCC, могут быть получены, как описывается в заявке на патент US2005/0169933Al. Связывающие группы включают в себя дисульфидные группы, тиоэфирные группы, группы лабильных кислот, фотолабильные группы, лабильные группы петидазы или лабильные группы эстеразы, как описывается в указанных выше патентах, дисульфидные и тиоэфирные группы являются предпочтительными. Дополнительные связывающие группы описываются и иллюстрируются здесь.
Конъюгаты антитела и майтанзиноида могут быть получены с использованием разнообразных бифункциональных агентов связывания белков, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные сложных имидоэфиров (таких как диметиладипимидат HCl), сложных активных эфиров (такие как дисукцинимидил суберат), альдегидов (таких как глютаральдегид), бис-азидо соединений (таких как бис(п-азидобензоил)гександиамин), бис-диазониевых производных (таких как бис-(п-диазонийбензоил)этилендиамин), диизоцианатов (таких как толуол 2,6-диизоцианат) и бис-активных соединений фтора (таких как 1,5-дифтор-2,4-динитробензол). Особенно предпочтительные связывающие агенты включают в себя N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) (Carlsson et al., Biochem. J. 173:723-37 (1978)) и N-сукцинимидил-4-(2-пиридилтио)пентаноат (SPP) для получения дисульфидной связи.
Линкер может присоединяться к молекуле майтанзиноида в различных положениях, в зависимости от типа связи. Например, сложноэфирная связь может быть получена посредством реакции с гидроксильной группой, с использованием обычных технологий связывания. Реакция может осуществляться в C-3 положении, имеющем гидроксильную группу, в C-14 положении, модифицированном с помощью гидроксиметила, в C-15 положении, модифицированном с помощью гидроксильной группы, и в C-20 положении, имеющем гидроксильную группу. В предпочтительном варианте осуществления, связь образуется в C-3 положении майтанзинола или аналога майтанзинола.
ii. Ауристатины и доластатины
В некоторых вариантах осуществления, иммуноконъюгат содержит антитело по настоящему изобретению, конъюгированное с долостатинами или пептидными аналогами и производными доластатинов, ауристатинами (заявки на патенты США № 5635483; 5780588). Доластатины и ауристатины, как показано, отрицательно влияют на динамику микротрубочек, гидролиз GTP и ядерное и клеточное деление (Woyke et al., Antimicrob. Agents Chemother. 45(12):3580-3584 (2001)) и имеют противораковую (заявка на патент США 5663149) и противогрибковую активность (Pettit et al., Antimicrob. Agents Chemother. 42:2961-2965(1998)). Лекарственный остаток доластатина или ауристатина может присоединяться к антителу через N (амино) конец или C (карбоксильный) конец пептидного лекарственного остатка (заявка WO 02/88172).
Примерные варианты осуществления ауристатина включают в себя соединенные с N-концом лекарственные остатки монометилауристатина DE и DF, описанные в заявке на патент США US2005/0238649, "Monomethylvaline Compounds Capable of Conjugation to Ligands", описание которой в явном виде включается в виде ссылки во всей его полноте.
Как правило, лекарственные остатки на основе пептидов могут быть получены посредством формирования пептидной связи между двумя или более фрагментами аминокислот и/или пептидов. Такие пептидные связи могут быть получены, например, в соответствии со способом жидкофазного синтеза (см. E. Schroder и K. Lϋbke, "The Peptides," volume 1, pp 76-136, 1965, Academic Press), который хорошо известен в области химии пептидов. Лекарственные остатки ауристатина/доластатина могут быть получены в соответствии со способами: заявка на патент США № 5635483; заявка на патент США № 5780588; Pettit et al., J. Am. Chem. Soc. 111:5463-65 (1989); Pettit et al., Аnti-Cancer Drug Design 13:243-77 (1998); Pettit et al., Synthesis 719-25 (1996); и Pettit et al., J. Chem. Soc. Perkin Trans. 15:859-863 (1996). См. также Doronina, Nature Biotechnol. 21(7):778-84 (2003); "Monomethylvaline Compounds Capable of Conjugation to Ligands", заявка на патент США US2005/0238649, которые тем самым включается сюда в виде ссылок во всей их полноте (описывая, например, линкеры и способы получения монометилвалиновых соединений, таких как MMAE и MMAF, конъюгированных с линкерами).
iii. Калихеамицин
В других вариантах осуществления, иммуноконъюгат содержит антитело по настоящему изобретению, конъюгированное с одной или несколькими молекулами калихеамицина. Семейство антибиотиков калихеамицинов способно осуществлять разрывы двухнитевой ДНК при субпикомолярных концентрациях. Для получения конъюгатов семейства калихеамицинов, см. заявки на патенты США № 5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001, 5877296 (все American Cyanamid Company). Структурные аналоги калихеамицина, которые могут использоваться, включают в себя, но, не ограничиваясь этим, γ1I, α2I, α3I, N-ацетил-γ1I, PSAG и θI1 (Hinman et al., Cancer Research 53:3336-42 (1993), Lode et al., Cancer Research 58:2925-28 (1998) и указанные выше заявки на патенты США, принадлежащие American Cyanamid). Другое противоопухолевое лекарственное средство, с которым может конъюгироваться антитело, может представлять собой QFA, который представляет собой антифолиат. Как калихеамицин, так и QFA имеют внутриклеточные сайты действия и не пересекают легко плазматическую мембрану. По этой причине, клеточное поглощение этих агентов посредством обусловленной антителом интернализации сильно увеличивает их цитотоксические эффекты.
iv. Другие цитотоксические агенты
Другие противоопухолевые агенты, которые могут конъюгироваться с антителами по настоящему изобретению, включают в себя BCNU, стрептозоицин, винкристин и 5-фторурацил, семейство агентов, известных коллективно как комплекс LL-E33288, описанный в заявках патенты США № 5053394, 5770710, а также эсперамицины (заявка на патент США № 5877296).
Ферментативно активные токсины и их фрагменты, которые могут использоваться, включают в себя цепь A дифтерии, несвязывающиеся активные фрагменты токсина дифтерии, цепь A экзотоксина (от Pseudomonas aeruginosa), цепь A рицина, цепь A абрина, цепь A модеццина, альфа-сарцин, белки Aleurites fordii, белки диантины, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор Momordica charantia, курцин, кротин, ингибитор sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены. См., например, заявку WO 93/21232, опубликованную 28 октября, 1993 года.
Настоящее изобретение дополнительно предполагает иммуноконъюгат, сформированный между антителом и соединением, которое имеет нуклеолитическую активность (например, рибонуклеазой или эндонуклеазой ДНК, такой как дезоксирибонуклеаза; ДНКаза).
Для селективного разрушения опухоли, антитело может содержать сильно радиоактивный атом. Ряд радиоактивных изотопов доступен для получения радиоактивно конъюгированных антител. Примеры включают в себя At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. Когда конъюгат используется для детектирования, он может содержать радиоактивный атом для сцинтиграфических исследований, например Tc99m или I123, или спиновую метку для получения изображения с помощью ядерного магнитного резонанса (ЯМР) (известного также как получение изображения с помощью ядерного магнитного резонанса, MRI), опять, такие как йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Радиоактивные или другие метки могут включаться в конъюгат известными способами. Например, пептид может синтезироваться биологически или может синтезироваться посредством химического синтеза аминокислот с использованием соответствующих предшественников аминокислот, содержащих, например, фтор-19 вместо водорода. Метки, такие как Tc99m или I123, Re186, Re188 и In111, могут присоединяться через цистеиновый остаток в пептиде. Иттрий-90 может присоединяться через лизиновый остаток. Метод IODOGEN (Fraker et al., Biochem. Biophys. Res. Commun. 80: 49-57 (1978) может использоваться для включения йода-123. "Monoclonal Antibodies in Immunoscintigraphy" (Chatal., CRC Press 1989) подробно описывает другие способы.
Конъюгаты антитела и цитотоксического агента могут быть получены с использованием разнообразных бифункциональных агентов связывания белков, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональных производных сложных имидоэфиров (таких как диметиладипимидат HCl), сложных активных эфиров (таких как дисукцинимидил суберат), альдегидов (таких как глютаральдегид), бис-азидо соединений (таких как бис(п-азидобензоил)гександиамин), бис-диазониевых производных (таких как бис-(п-диазонийбензоил)этилендиамин), диизоцианатов (таких как толуол 2,6-диизоцианат) и бис-активных соединений фтора (таких как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицина может быть получен так, как описывается в Vitetta et al., Science 238:1098 (1987). Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилен триаминпентауксусная кислота (MX-DTPA) представляет собой пример хелатирующего агента для конъюгирования радиоактивного нуклеотида и антитела. См. заявку WO 94/11026. Линкер может представлять собой "расщепляемый линкер", облегчающий высвобождение цитотоксического лекарственного средства в клетке. Например, может использоваться кислотно-лабильный линкер, линкер, чувствительный к пептидазе, фотолабильный линкер, диметиловый линкер или дисульфидсодержащий линкер (Chari et al., Cancer Research 52:127-31 (1992); заявка на патент США № 5208020).
Соединения по настоящему изобретению предположительно включают, но, не ограничиваясь этим, ADC, полученные с помощью реагентов для поперечного связывания; BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, MPBH, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB, и SVSB (сукцинимидил-(4-винилсульфон)бензоат), которые являются коммерчески доступными (например, от Pierce Biotechnology, Inc., Rockford, IL., U.S.A). См. страницы 467-498, 2003-2004 Applications Handbook and Catalog.
v. Получение конъюгатов антитела и лекарственного средства
В конъюгатах антитела и лекарственного средства (ADC) по настоящему изобретению, антитело (Ab) конъюгируется с одним или несколькими лекарственными остатками (D), например примерно с 1 - примерно с 20 лекарственными остатками на одно антитело, через линкер (L). ADC Формулы I могут быть получены с помощью нескольких способов, с использованием реакций, условий и реагентов органической химии, известных специалистам в данной области, включая: (1) реакцию нуклеофильной группы антитела с двухвалентным реагентом связывания, с образованием Ab-L, с помощью ковалентной связи, с последующей реакцией с лекарственным остатком D; и (2) реакцию нуклеофильной группы лекарственного остатка с двухвалентным реагентом связывания, с образованием D-L, через ковалентную связь, с последующей реакцией с нуклеофильной группой антитела. Дополнительные способы получения ADC описываются здесь.
Ab(LD)p I
Линкер может состоять из одного или нескольких компонентов линкера. Примеры компонентов линкера включают в себя 6-малеимидокапроил ("MC"), малеимидопропаноил ("MP"), валин-цитруллин ("val-cit"), аланин-фенилаланин ("ala-phe"), п-аминобензилоксикарбонил ("PAB"), N-сукцинимидил 4-(2-пиридилтио) пентаноат ("SPP"), N-сукцинимидил 4-(N-малеимидометил) циклогексан-1-карбоксилат ("SMCC) и N-сукцинимидил (4-йод-ацетил)аминобензоат ("SLAB"). Дополнительные компоненты линкеров известны в данной области и некоторые из них описываются здесь. См. также "Monomethylvaline Compounds Capable of Conjugation to Ligands", заявку на патент США US2005/0238649, содержание которых тем самым включается сюда в виде ссылок во всей их полноте.
В некоторых вариантах осуществления, линкер может содержать остатки аминокислот. Примерные компоненты аминокислот линкера включают в себя дипептид, трипептид, тетрапептид или пентапептид. Примеры дипептидов включают в себя: валин-цитруллин (vc или val-cit), аланин-фенилаланин (af или ala-phe). Примеры трипептидов включают в себя: глицин-валин-цитруллин (gly-val-cit) и глицин-глицин-глицин (gly-gly-gly). Остатки аминокислот, которые составляют компонент аминокислот линкера, включают в себя те, которые существуют в природе, а также минорные аминокислоты и аналоги аминокислот, не существующие в природе, такие как цитруллин. Компоненты аминокислот линкера могут конструироваться и оптимизироваться по их селективности к ферментативному расщеплению с помощью конкретных ферментов, например протеазы, связанной с опухолью, катепсина B, C и D или плазминпротеазы.
Нуклеофильные группы на антителах включают в себя, но, не ограничиваясь этим: (i) N-концевые аминогруппы, (ii) аминогруппы боковых цепей, например лизин, (iii) тиольные группы боковых цепей, например цистеин, и (iv) гидроксильные или аминогруппы сахаров, когда антитело является гликозилированным. Амино, тиольные и гидроксильные группы являются нуклеофильными и способны взаимодействовать, с формированием ковалентных связей, с электрофильными группами на остатках линкеров и с реагентами связывания, включая: (i) сложные активные эфиры, такие как сложные эфиры NHS, сложные эфиры HOBt, галогенформиаты и гидрогалогениды; (ii) алкил и бензил галогениды, такие как галогенацетамиды; (iii) альдегиды, кетоны, карбоксильные и малеимидные группы. Некоторые антитела имеют восстанавливаемые дисульфиды внутри цепей, то есть цистеиновые мостики. Антитела могут делаться химически активными для конъюгирования с реагентами связывания посредством обработки восстанавливающим агентом, таким как DTT (дитиотреитол). Каждый цистеиновый мостик будет таким образом формировать, теоретически, два химически активных тиольных нуклеофила. Дополнительные нуклеофильные группы могут вводиться в антитела посредством реакции лизинов с 2-иминотиоланом (реагент Траута), приводя к преобразованию амина в тиол. Химически активные тиольные группы могут вводиться в антитело (или его фрагмент) посредством введения одного, двух, трех, четырех или более цистеиновых остатков (например, для получения мутантных антител, содержащих один или несколько не-нативных цистеиновых остатков аминокислот).
Конъюгаты антитела и лекарственного средства по настоящему изобретению могут также быть получены посредством модификации антитела для введения электрофильных остатков, которые могут взаимодействовать с нуклеофильными заместителями на реагенте связывания или лекарственном средстве. Сахара гликозилированных антител могут быть окислены, например, с помощью периодатных окислительных реагентов, с формированием групп альдегидов или кетонов, которые могут взаимодействовать с аминовой группой реагентов связывания или лекарственных остатков. Получаемые в результате иминовые группы оснований Шиффа могут образовывать стабильную связь или могут восстанавливаться, например, с помощью боргидридных реагентов с формированием стабильных аминосвязей. В одном из вариантов осуществления, реакция углеводной части гликозилированного антитела либо с галактоза оксидазой, либо с мета-периодатом натрия может давать карбонильные (альдегидные и кетоновые) группы в белке, которые могут взаимодействовать с соответствующими группами на лекарственном средстве (Hermanson, Bioconjugate Techniques). В другом варианте осуществления, белки, содержащие N-концевые сериновые или треониновые остатки, могут взаимодействовать с мета-периодатом натрия, приводя к получению альдегида вместо первой аминокислоты (Geoghegan & Stroh, Bioconjugate Chem. 3:138-46 (1992); заявка на патент США № 5362852). Такой альдегид может взаимодействовать с лекарственным остатком или нуклеофилом связывания.
Подобным же образом, нуклеофильные группы на лекарственном остатке включают в себя, но, не ограничиваясь этим: амино, тиольные, гидроксильные, гидразидные, оксимовые, гидразиновые, тиосемикарбазоновые, гидразинкарбоксилатные и арилгидразидные группы, способные взаимодействовать с образованием ковалентных связей с электрофильными группами на линкерных остатках и реагентах связывания, включая: (i) сложные активные эфиры, такие как сложные эфиры NHS, сложные эфиры HOBt, галогенформиаты и гидрогалогениды; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; (iii) альдегиды, кетоны, карбоксильные и малеимидные группы.
Альтернативно, белок слияния, содержащий антитело и цитотоксический агент, может быть получен, например, с помощью рекомбинантных технологий или пептидного синтеза. Отрезок ДНК может содержать соответствующие области, кодирующие две части конъюгата, либо расположенные рядом друг с другом, либо разделенные областью, кодирующей линкерный пептид, который не разрушает желаемых свойств конъюгата.
Еще в одном варианте осуществления, антитело может конъюгироваться с "рецептором" (таким как стрептавидин) для использования с предварительным нацеливанием на опухоль, где конъюгат антитело-рецептор вводится пациенту, с последующим удалением несвязанного конъюгата из циркуляции с использованием выводящего агента, а затем введения "лиганда" (например, авидина), который конъюгируется с цитотоксичным агентом (например, радиоактивным нуклеотидом).
Фармацевтические препараты
Терапевтические препараты, содержащие антитело по настоящему изобретению, получают для хранения посредством смешивания антитела, имеющего желаемую степень чистоты, с необязательными физиологически приемлемыми носителями, наполнителями или стабилизаторами (Remington: The Science and Practice of Pharmacy 20th edition (2000)), в форме водных растворов, лиофилизированных или других высушенных препаратов. Приемлемые носители, наполнители или стабилизаторы являются нетоксичными для пациента при используемых дозировках и концентрациях и включают в себя буферы, такие как фосфатный, цитратный, гистидин и другие органические кислоты; антиоксиданты, включающие в себя аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензиламмоний хлорид; гексаметоний хлорид; бензалконий хлорид, бензэтоний хлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (меньше, примерно, чем 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин, или иммуноглобулины; гидрофильные полимеры, такие как поливинилпиролидон; аминокислоты, такие как глицин, глютамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахара, такие как сахароза, маннитол, трегалоза или сорбитол; образующие соль противоионы, такие как натрий; комплексы с металлами (например, комплексы Zn-белок) и/или неионные поверхностно-активные вещества, такие как TWEENTM, PLURONICSTM или полиэтиленгликоль (PEG).
Препараты здесь могут также содержать более одного активного соединения, если это необходимо для конкретного показания, которое лечится, предпочтительно, соединения с комплементарными активностями, которые не влияют отрицательно друг на друга. Такие молекулы присутствуют соответствующим образом в сочетании, в количествах, которые являются эффективными для предполагаемой цели.
Активные ингредиенты могут также захватываться в микрокапсуле, приготовленной, например, с помощью технологии коацервации или межфазной полимеризации, например, в микрокапсуле из гидроксиметилцеллюлозы или желатина и в поли(метилметацилатной) микрокапсуле, соответственно, в коллоидных системах доставки лекарственных средств (например, в липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) или в макроэмульсиях. Такие технологии описываются в Remington: The Science and Practice of Pharmacy 20th edition (2000).
Препараты, которые должны использоваться при введении in vivo, должны быть стерильными. Это легко достигается посредством фильтрования через стерильные фильтрационные мембраны.
Могут быть получены препараты с замедленным высвобождением. Соответствующие примеры препаратов с замедленным высвобождением включают в себя полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие иммуноглобулин по настоящему изобретению, где матрицы находятся в форме сформированных изделий, например пленок или микрокапсул. Примеры матрицы с замедленным высвобождением включают в себя сложные полиэфиры, гидрогели (например, поли(2-гидроксиэтил-метакрилат) или поли(виниловый спирт)), полилактиды (заявка на патент США № 3773919), сополимеры L-глютаминовой кислоты и γ-этил-L-глютамата, недеградируемые сополимеры этилена-винилацетата, деградируемые сополимеры молочной кислоты - гликолевой кислоты, такие как LUPRON DEPOTTM (микросферы для инъекций, состоящие из сополимера молочной кислоты - гликолевой кислоты и лейпролидацетата), и поли-D-(-)-3-гидроксимасляную кислоту. Хотя полимеры, такие как этилен-винилацетат и молочная кислота - гликолевая кислота, делают возможным высвобождение молекул в течение более 100 дней, определенные гидрогели высвобождают белки в течение более коротких периодов времени. Когда инкапсулируемые иммуноглобулины удерживаются в организме в течение длительного времени, они могут денатурировать или агрегировать в результате для экспонирования для влажности при 37°C, в результате чего происходит потеря биологической активности и возможные изменения в иммуногенности. Рациональные стратегии могут создаваться для стабилизации, в зависимости от осуществляемого механизма. Например, если механизм агрегации, как обнаружено, представляет собой внутримолекулярное образование S-S связей посредством взаимного тио-дисульфидного обмена, стабилизация может достигаться посредством модификации сульфгидрильных остатков, лиофилизации из кислотных растворов, контроля содержания влажности, использования соответствующих добавок и разработки конкретных композиций полимерной матрицы.
Применения
Антитело по настоящему изобретению может использоваться in vitro, ex vivo и in vivo, например, в терапевтических методах.
В некоторых вариантах осуществления, настоящее изобретение предоставляет способы понижения или ингибирования ангиогенеза у субъекта, имеющего патологическое состояние, связанное с ангиогенезом, включающие в себя введение субъекту эффективного количества антитела анти-EGFL7 по настоящему изобретению. Эти состояния включают в себя, например, неоплазмы, (включая карциномы) и определенные состояния глаз.
Раковое заболевание, которое может лечиться с помощью настоящего изобретения, включает, но, не ограничиваясь этим, злокачественную карциному, лимфому, бластому, саркому и лейкемию или лимфоидные злокачественные заболевания. Более конкретные примеры таких раковых заболеваний включают рак груди, рак толстой кишки, рак прямой кишки, рак толстой и прямой кишки, рак почек или надпочечников, рак легких, включая мелкоклеточный рак легких, немелкоклеточный рак легких, аденокарциному легких и сквамозно-клеточную карциному легких, сквамозно-клеточный рак (например, эпителиальный сквамозный рак клеток), рак шейки матки, рак яичников, рак простаты, рак печени, рак мочевого пузыря, рак брюшной полости, рак клеток печени, рак желудка или пищеварительной системы, включая рак желудочно-кишечных органов, рак поджелудочной железы, рак головы и шеи, глиобластому, ретинобластому, астроцитому, текомы, арренобластомы, гепатому, гематологические злокачественные заболевания, включая неходжкинскую лимфому (NHL), множественную миелому и острые гематологические злокачественные заболевания, карциному эндометрия или матки, эндометриоз, фибросаркому, хориокарциному, карциному слюнных желез, рак женских половых органов, рак щитовидной железы, карциному пищевода, карциному печени, карциному заднего прохода, карциному мужского полового члена, карциному носоглотки, ларингеальные карциномы, саркому Капоши, меланому, карциномы кожи, шванному, олигодендроглиому, нейробластому, рабдомиосаркому, остеогенную саркому, лейомиосаркомы, карциномы мочевого тракта, карциномы щитовидной железы, опухоль Вильма, а также аномальную васкулярную пролиферацию, связанную с факоматозами, отеком (такие как связанные с опухолями мозга) и синдромом Мейга. Предпочтительно, рак выбирается из группы, состоящей из рака груди, рака толстой и прямой кишки, немелкоклеточного рака легких, неходжкинской лимфомы (NHL), рака надпочечников, рака простаты, рака печени, рака головы и шеи, меланомы, рака яичников, мезотелиомы и множественной миеломы. Более предпочтительно, рак представляет собой рак толстой и прямой кишки. Раковые состояния, которые могут лечиться по настоящему изобретению, включают в себя метастатические раковые заболевания. Настоящее изобретение является особенно пригодным для лечения васкуляризированных опухолей.
Состояния глаз, злокачественные заболевания по настоящему изобретению включают глазные неоваскулярные заболевания, включая, но, не ограничиваясь этим, возрастную макулярную дегенерацию, диабетический макулярный отек, пролиферативную диабетическую ретинопатию, закупорку центральной ретинальной вены с цистоидным макулярным отеком, закупорку ветви ретинальной вены с цистоидным макулярным отеком, рубеоз радужной оболочки, патологическую миопию/CNV, синдром фон Хиппеля-Линдау, птеригий, POHS (гистоплазмоз)/CNV, хороидальную гемангиому, преждевременную ретинопатию (ROP), радиационную ретинопатию, внутриглазные опухоли (например, меланому, ретинобластому, метастазы и кавернозные гемангиомы глазной впадины), полипоидную хороидопатию, идиопатическую юкстафовеальную телангиектазию, болезнь Илза, кавернозные гемангиомы глазной впадины, лимфангиому глазной полости, капиллярные гемангиомы век, васкуляризацию корнеального трансплантата, неоваскуляризацию корнеального трансплантата, болезнь Котса и проблемы с заживлением ран, связанных с хирургическим лечением глаукомы.
Кроме того, по меньшей мере, некоторые из антител по настоящему изобретению могут связывать антиген от других видов. Соответственно, антитела по настоящему изобретению могут использоваться для связывания специфичной активности антигена, например, в клеточной культуре, содержащей антиген, у субъекта человека или у субъектов других млекопитающих, имеющих антиген, с которым антитело по настоящему изобретению перекрестно взаимодействует (например, шимпанзе, бабуин, мартышка, циномолгус и резус, свинья или мышь). В некоторых вариантах осуществления, антитело по настоящему изобретению может использоваться для ингибирования активности антигена посредством приведения в контакт антитела с антигеном, так что активность антигена ингибируется. Предпочтительно, антиген представляет собой молекулу белка человека.
В некоторых вариантах осуществления, антитело по настоящему изобретению может использоваться в способе связывания антигена у субъекта, страдающего от расстройства, связанного с повышенной экспрессией и/или активностью антигена, включающем в себя введение субъекту антитела по настоящему изобретению, так что антиген в субъекте связывается. Предпочтительно, антиген представляет собой молекулу белка человека, и субъект представляет собой субъект человека. Альтернативно, субъект может представлять собой млекопитающее, экспрессирующее антиген, с которым связывается антитело по настоящему изобретению. Кроме того, субъект может представлять собой млекопитающее, в которое вводится антиген (например, посредством введения антигена или посредством экспрессирования трансгена антигена). Антитело по настоящему изобретению может вводиться субъекту человека для терапевтических целей. Кроме того, антитело по настоящему изобретению может вводиться млекопитающему не-человеку, экспрессирующему антиген, с которым иммуноглобулин перекрестно взаимодействует (например, примату, свинье или мыши) для ветеринарных целей или в качестве животной модели заболевания человека. Относительно последней, такие животные модели могут быть пригодными для использования при оценке терапевтической эффективности антител по настоящему изобретению (например, при исследовании дозировок и временных графиков введения).
Антитела по настоящему изобретению могут использоваться для лечения, ингибирования, замедления развития, предотвращения/замедления рецидивов, облегчения или предотвращения заболеваний, расстройств или состояний, связанных с экспрессированием и/или активностью молекул одного или нескольких антигенов.
В определенных вариантах осуществления, иммуноконъюгат, содержащий антитело, конъюгированное с одним или несколькими цитотоксичными агентами, вводится пациенту. В некоторых вариантах осуществления, иммуноконъюгат и/или антиген, с которым он связан, интернализуется (интернализуются) клеткой, что приводит к повышению терапевтической эффективности иммуноконъюгата при уничтожении целевых клеток, с которыми он связывается. В одном из вариантов осуществления, цитотоксический агент нацеливает нуклеиновую кислоту в целевой клетке или взаимодействует с ней. В одном из вариантов осуществления, цитотоксический агент нацелен на полимеризацию микротрубочек или влияет на нее отрицательно. Примеры таких цитотоксических агентов включают в себя любой из химиотерапевтических агентов, отмеченных здесь (такие как майтанзиноид, ауристатин, доластатин или калихеамицин), радиоактивный изотоп или рибонуклеазу, или эндонуклеазу ДНК.
Антитела по настоящему изобретению могут использоваться либо сам по себе, либо в сочетании с другими композициями, при терапии. Например, антитело по настоящему изобретению может вводиться совместно с химиотерапевтическим агентом (агентами) (включая коктейли химиотерапевтических агентов), другим цитотоксическим агентом (агентами), антиангиогенным агентом (агентами), цитокинами и/или агентом (агентами) ингибитором роста. Такие сочетанные терапии, отмеченные выше, включают в себя сочетанное введение (где два или более агентов включаются в один и тот же или в отдельные препараты) и раздельное введение, в этом случае, введение антитела по настоящему изобретению может осуществляться до и/или после введения вспомогательной терапии или терапий.
Антитело по настоящему изобретению (и вспомогательный терапевтический агент) вводится (вводятся) с помощью любых пригодных для использования средств, включая парентеральное, подкожное, внутрибрюшинное, внутрилегочное и интраназальное, и, если это желательно для местного лечения, внутрираневое введение. Парентеральные вливания включают в себя внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. В дополнение к этому, антитело удобно вводится с помощью импульсного вливания, в частности, при уменьшающихся дозах антитела. Дозирование может осуществляться с помощью любого пригодного для этого способа, например, с помощью инъекций, таких как внутривенные или подкожные инъекции, в зависимости, частично, от того, является ли введение кратким или хроническим.
Композиция антитела по настоящему изобретению будет приготавливаться, дозироваться и вводиться способом, совместимым с хорошей медицинской практикой. Факторы для рассмотрения в этом контексте включают в себя конкретное расстройство, которое лечится, конкретное млекопитающее, которое лечится, клиническое состояние индивидуального пациента, причину расстройства, место доставки агента, способ введения, временной график введения и другие факторы, известные практикующим врачам. Хотя это и необязательно для антитела, оно необязательно приготавливается вместе с одним или несколькими агентами, используемыми в настоящее время для предотвращения или лечения рассматриваемого расстройства. Эффективное количество таких дополнительных агентов зависит от количества антител по настоящему изобретению, присутствующих в препарате, типа расстройства или лечения и других факторов, обсуждаемых выше. Они, как правило, используются при таких же дозировках и с помощью способа введения, как использовалось ранее, или от примерно 1 до 99% от дозировок, используемых ранее.
Для предотвращения или лечения заболевания, соответствующая дозировка антитела по настоящему изобретению (когда оно используется само по себе или в сочетании с другими агентами) будет зависеть от типа заболевания, которое должно лечиться, типа антитела, тяжести и хода заболевания, от того, вводится ли антитело для превентивных или терапевтических целей, от предыдущей терапии, анамнеза пациента и реакций на антитело, и от опыта наблюдающего врача. Антитело удобно вводить пациенту за один раз или в виде ряда сеансов лечения. В зависимости от типа и тяжести заболевания, от примерно 1 мкг/кг до 15 мг/кг (например, 0,1 мг/кг-10 мг/кг) антитела представляет собой начальную дозу кандидата для введения пациенту, например, с помощью одного или нескольких отдельных введений, или посредством непрерывного вливания. Типичная ежедневная доза может находиться в пределах от примерно 1 мкг/кг до 100 мг/кг или более, в зависимости от факторов, рассмотренных выше. Для повторяющихся введений в течение нескольких дней или дольше, в зависимости от состояния, лечение поддерживается до тех пор, пока не произойдет желаемого подавления симптомов заболевания. Примерная доза антитела должна находиться в пределах от примерно 0,05 мг/кг до примерно 10 мг/кг. Таким образом, одна или несколько доз, составляющих примерно 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любое их сочетание) может вводиться пациенту. Такие дозы могут вводиться с перерывами, например, каждую неделю или каждые три недели (например, так что пациент принимает от примерно двух до примерно двадцати, например, примерно шесть доз антитела). Может вводиться начальная, более высокая нагружающая доза, а затем одна или несколько более низких доз. Примерный режим дозирования включает в себя введение начальной нагружающей дозы примерно 4 мг/кг, с последующей еженедельной поддерживающей дозой примерно 2 мг/кг антитела. Однако и другие режимы дозирования могут быть пригодными для использования. Ход этой терапии легко отслеживается с помощью обычных методик и анализов.
Антитела анти-EGFL7 по настоящему изобретению являются пригодными при анализах, детектирующих экспрессию EGFL7 (таких как диагностические или прогностические анализы), в конкретных клетках или тканях, где антитела метятся, как описывается ниже, и/или иммобилизуются на нерастворимой матрице.
Настоящее изобретение предоставляет способы детектирования EGFL7, способы включают в себя детектирование комплекса EGFL7 и антитела анти-EGFL7 в образце. Термин "детектирование", как здесь используется, включает в себя качественное и/или количественное детектирование (измерения уровней) со ссылками на контроль или без них.
Настоящее изобретение предоставляет способы диагностирования расстройств, связанных с экспрессией и/или активностью EGFL7, способы включают в себя детектирование комплекса EGFL7-антитело анти-EGFL7 в биологическом образце от пациента, имеющего или предположительно имеющего расстройство. В некоторых вариантах осуществления, экспрессия EGFL7 представляет собой повышенную экспрессию или аномальную (нежелательную) экспрессию.
Настоящее изобретение предоставляет любое антитело анти-EGFL7, описываемое здесь, где антитело анти-EGFL7 содержит детектируемую метку.
Настоящее изобретение предоставляет комплекс любого антитела анти-EGFL7, описываемого здесь, и EGFL7. В некоторых вариантах осуществления, комплекс находится in vivo или in vitro. В некоторых вариантах осуществления, комплекс содержит раковые клетки. В некоторых вариантах осуществления, антитело анти-EGFL7 метится с возможностью детектирования.
Антитела анти-EGFL7 могут использоваться для детектирования EGFL7 в любом из ряда хорошо известных способов детектирования при анализе. Например, биологический образец может анализироваться на EGFL7 посредством получения образца от желаемого источника, смешивания образца с антителом анти-EGFL7, чтобы сделать возможным получение антитела с формированием комплекса антитело/EGFL7 с любым из EGFL7, присутствующих в смеси, и детектирования любого комплекса антитело/EGFL7, присутствующего в смеси. Биологический образец может быть получен для анализа с помощью способов, известных в данной области, которые являются пригодными для конкретного образца. Способы смешивания образца с антителом и способы детектирования комплекса антитело/EGFL7 выбираются в соответствии с типом используемого анализа. Такой анализ включает в себя иммуногистохимический, конкурентный и сэндвич анализ, и анализ стерического ингибирования.
Аналитические способы для EGFL7, все, используют один или несколько из следующих реагентов: меченый аналог EGFL7, иммобилизированный аналог EGFL7, меченое антитело анти-EGFL7, иммобилизированное антитело анти-EGFL7 и стерические конъюгаты. Меченые реагенты также известны как "метки".
Используемая метка представляет собой любую детектируемую функциональную группу, которая не влияет отрицательно на связывание EGFL7 и антитела анти-EGFL7. Многочисленные метки известны для использования в иммунном анализе, примеры включают в себя остатки, которые могут детектироваться непосредственно, такие как флюорохром, хемилюминесцентные и радиоактивные метки, а также остатки, такие как ферменты, которые должны взаимодействовать или дериватизироваться для детектирования.
Используемая метка представляет собой любую детектируемую функциональную группу, которая не влияет отрицательно на связывание EGFL7 и антитела анти-EGFL7. Многочисленные метки известны для использования в иммунном анализе, примеры включают в себя остатки, которые могут детектироваться непосредственно, такие как флюорохром, хемилюминесцентные и радиоактивные метки, а также остатки, такие как ферменты, которые должны взаимодействовать или дериватизироваться для детектирования. Примеры таких меток включают в себя радиоизотопы 32P, 14C, 125I, 3H и 131I, флюорофоры, такие как хелаты редкоземельных металлов или флюоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люцериферазы, например, люциферазу светлячков и бактериальную люциферазу (заявка на патент США № 4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидазу хрена (HRP), щелочную фосфатазу, β-галактозидазу, глюкоамилазу, лизоцим, сахарид оксидазы, например глюкоза оксидазу, галоктоза оксидазу и глюкоза-6-фосфатдегидрогеназу, гетероциклические оксидазы, такие как уриказа и ксантин оксидаза, в сочетании с ферментом, который использует перекись водорода для окисления предшественника красителя, такого как HRP, лактопероксидазау или микропероксидазу, биотин/авидин, спиновые метки, метки бактериофаги, стабильные свободные радикалы и тому подобное.
Обычные способы являются доступными для ковалентного связывания этих меток с белками или полипептидами. Например, связывающие агенты, такие как диальдегиды, карбодиимиды, дималеимиды, бис-имидаты, бис-диазобензидин и тому подобное, могут использоваться для прикрепления к антителам, описанным выше, флуоресцентных, хемилюминесцентных и ферментативных меток. См., например, заявки на патенты США № 3940475 (флуориметрия) и 3645090 (ферменты); Hunter et al., Nature 144: 945 (1962); David et al., Biochemistry 13: 1014-21 (1974); Pain et al., J. Immunol. Meth. 40: 219-30 (1981); и Nygren, J. Histochem. and Cytochem. 30: 407-12 (1982). Предпочтительные метки здесь представляют собой ферменты, такие как пероксидаза хрена и щелочная фосфатаза. Конъюгирование такой метки, включая ферменты, с антителом представляет собой стандартную манипуляционную процедуру для специалиста в области методик иммунных анализов. См., например, O'Sullivan et al., "Methods for Preparation of Enzyme-antibody Conjugates for Use in Enzyme Immunoassay," in Methods Enzymol., ed. J.J. Langone and H. Van Vunakis, Vol. 73 (Academic Press, New York, New York, 1981), pp. 147-166.
Иммобилизация реагентов требуется для некоторых способов анализа. Иммобилизация включает в себя отделение антитела анти-EGFL7 от любых EGFL7, которые остаются свободными в растворе. Обычно это осуществляется либо посредством инсолюбилизации, либо антитела анти-EGFL7, либо аналога EGFL7 перед процедурой анализа, например, с помощью адсорбции на водонерастворимой матрице или поверхности (Bennich et al., заявка на патент США № 3720760), посредством ковалентного связывания (например, с использованием поперечного связывания с помощью глютаральдегида), либо посредством инсолюбилизации антитела анти-EGFL7 или аналога EGFL7 после этого, например, с помощью иммунной преципитации.
Экспрессия белков в образце может исследоваться с использованием иммуногистохимии и протоколов окрашивания. Иммунное гистохимическое окрашивание срезов тканей, как показано, является надежным способом оценки или детектирования присутствия белков в образце. Методики иммуногистохимии ("IHC") используют антитело для зондирования и визуализации клеточных антигенов in situ, как правило, с помощью хромогенных или флуоресцентных способов. Для приготовления образца может использоваться образец ткани или клеток от млекопитающего (как правило, пациента человека). Образец может быть получен с помощью разнообразных процедур, известных в данной области, включая, но, не ограничиваясь этим, хирургическое иссечение, отсос или биопсию. Ткань может быть свежей или замороженной. В одном из вариантов осуществления, образец фиксируется и погружается в парафин или что-либо подобное. Образец ткани может фиксироваться (то есть консервироваться) с помощью обычной методологии. Специалист в данной области поймет, что выбор фиксатива определяется целью, для которой образец должен гистологически окрашиваться или анализироваться иным образом. Специалист в данной области также поймет, что продолжительность фиксации зависит от размера образца ткани и используемого фиксатива.
IHC может осуществляться в сочетании с дополнительными методиками, такими как морфологическое окрашивание и/или флуоресцентная гибридизация in situ. Доступны два общих способа IHC; прямые и опосредованные анализы. В соответствии с первым анализом, связывание антитела с целевым антигеном (например, EGFL7) определяется непосредственно. Этот непосредственный анализ использует меченый реагент, такой как флуоресцентная метка или ферментативно меченое первичное антитело, которое может визуализироваться без дополнительного взаимодействия антитела. В типичном опосредованном анализе, неконъюгированное первичное антитело связывается с антигеном, а затем меченое вторичное антитело связывается с первичным антителом. Когда вторичное антитело конъюгируется с ферментативной меткой, добавляется хромогенный или флюорогенный субстрат для обеспечения визуализации антигена. Усиление сигнала происходит потому, что несколько вторичных антител могут взаимодействовать с различными эпитопами на первичном антителе.
Первичное и/или вторичное антитело, используемое для иммуногистохимии, как правило, метится с помощью детектируемого остатка. Многочисленные метки являются доступными, и они могут в целом быть сгруппированы в следующие категории.
Наряду с процедурами приготовления образца, обсуждаемыми выше, может быть желательной дополнительная обработка среза ткани до, после или во время IHC. Например, могут осуществляться способы извлечения эпитопа, такие как нагрев образца ткани в цитратном буфере (см., например, Leong et al. Appl. Immunohistochem. 4(3):201 (1996)).
После необязательной стадии блокирования, срез ткани экспонируется для первичного антитела в течение достаточного периода времени и при соответствующих условиях, так что первичное антитело связывается с целевым белком антигеном в образце ткани. Соответствующие условия для достижения этого могут быть определены с помощью рутинных экспериментов. Степень связывания антитела с образцом определяется посредством использования любой из детектируемых меток, обсуждаемых выше. Предпочтительно, метка представляет собой ферментативную метку (например, HRPO), которая катализирует химическое изменение хромогенного субстрата, такого как 3,3'-диаминобензидиновый хромоген. Предпочтительно, ферментативная метка конъюгируется с антителом, которое специфично связывается с первичным антителом (например, первичное антитело представляет собой поликлональное антитело кролика, а вторичное антитело представляет собой антитело козы против кролика).
Образцы, приготовленные таким образом, могут закрепляться и помещаться между стеклами. Оценка слайда затем определяется, например, с использованием микроскопа, и могут использоваться критерии интенсивности окрашивания, рутинно используемые в данной области.
Другие способы анализов, известные как конкурентные или сэндвич анализы, хорошо установлены и широко используются в коммерческой диагностической промышленности.
Конкурентные анализы основываются на способности метки аналога EGFL7 к конкуренции с исследуемым образцом EGFL7 за ограниченное количество сайтов связывания антитела анти-EGFL7 с антигеном. Антитело анти-EGFL7, как правило, инсолюбилизируется до или после конкуренции, а затем метка и EGFL7, связанные с антителом анти-EGFL7, отделяются от несвязанной метки и EGFL7. Это разделение осуществляется посредством декантирования (когда партнер по связыванию предварительно инсолюбилизируется) или посредством центрифугирования (когда партнер по связыванию преципитирует после конкурентной реакции). Количество исследуемого образца EGFL7 является обратно пропорциональным количеству связанной метки, как измеряется по количеству вещества маркера. Получают кривые доза-отклик для известных количеств EGFL7 и сравнивают их с результатами исследований для количественного определения количества EGFL7, присутствующего в исследуемом образце. Эти анализы называют системами ELISA, когда ферменты используются в качестве детектируемых маркеров.
Другие виды конкурентного анализа, называемого "гомогенный" анализ, не требуют разделения фаз. Здесь, конъюгат фермента с EGFL7 получают и используют таким образом, что, когда антитело анти-EGFL7 связывается с EGFL7, присутствие антитела анти-EGFL7 модифицирует активность фермента. В этом случае, EGFL7 или его иммунологически активные фрагменты конъюгируют с помощью бифункционального органического мостика с ферментом, таким как пероксидаза. Конъюгаты выбираются для использования вместе с антителом анти-EGFL7, так что связывание антитела анти-EGFL7 ингибирует или усиливает ферментативную активность метки. Этот способ сам по себе широко осуществляется под наименованием EMIT.
Стерические конъюгаты используются в способах стерического затруднения для гомогенного анализа. Эти конъюгаты синтезируют с помощью ковалентного связывания низкомолекулярного гаптена с малым фрагментом EGFL7, так что антитело к гаптену по существу неспособно связывать конъюгат одновременно с антителом анти-EGFL7. При этой процедуре анализа, EGFL7, присутствующий в исследуемом образце, будет связывать антитело анти-EGFL7, тем самым давая возможность анти-гаптену для связывания конъюгата, что приводит к изменению характера конъюгата с гаптеном, например, к изменению флюоресценции, когда гаптен представляет собой флуорофор.
Сэндвич-анализы, в частности, являются пригодными для определения EGFL7 или антитела анти-EGFL7. В последовательных сэндвич-анализах иммобилизованное антитело анти-EGFL7 используют для адсорбции исследуемого образца EGFL7, исследуемый образец удаляют, например, с помощью отмывки, связанный EGFL7 используют для адсорбции второго, меченого антитела анти-EGFL7 и связанный материал затем отделяют от остатка метки. Количество связанного метки прямо пропорционально количеству исследуемого образца EGFL7. При "одновременных" сэндвич-анализах исследуемый образец не разделяется перед добавлением меченого анти-EGFL7. Последовательный сэндвич-анализ с использованием моноклонального антитела анти-EGFL7 в качестве одного антитела и поликлонального антитела анти-EGFL7 в качестве другого является пригодным при исследовании образцов на EGFL7.
Указанное выше описание представляет собой только примеры анализов для детектирования EGFL7. Существуют другие методы в настоящем или будут существовать в будущем, которые используют антитело анти-EGFL7 для определения EGFL7, и они включаются в рамки настоящего изобретения, включая биологические анализы, описанные здесь.
Промышленные изделия
В другом аспекте настоящего изобретения предоставляется промышленное изделие, содержащее материалы, пригодные для лечения, предотвращения и/или диагностики расстройств, описанных выше. Промышленное изделие содержит контейнер и метку или вставку в упаковку, или описание, связанное с контейнером. Пригодные для использования контейнеры включают, например, бутылки, флаконы, шприцы и тому подобное. Контейнеры могут формироваться из различных материалов, таких как стекло или пластик. Контейнер содержит композицию, которая сама по себе или при объединении с другой композицией (композициями) является эффективной при лечении, предотвращении и/или диагностике состояния и может иметь стерильный узел для доступа (например, контейнер может представлять собой мешок или флакон для внутривенного раствора, имеющий пробку, протыкаемую с помощью иглы для подкожных инъекций). По меньшей мере, один активный агент композиции представляет собой антитело по настоящему изобретению. Метка или вставка в упаковку указывает, что композиция используется для лечения выбранного состояния, такого как астма. Кроме того, промышленное изделие может содержать (a) первый контейнер с композицией, содержащейся в нем, где композиция содержит антитело по настоящему изобретению; и (b) второй контейнер с композицией, содержащейся в нем. Промышленное изделие в этом варианте осуществления настоящего изобретения может дополнительно содержать вставку в упаковку, указывающую, что первая и вторая композиция антител могут использоваться для лечения конкретного состояния, например астмы. Альтернативно или в дополнение к этому, промышленное изделие может дополнительно содержать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI), фосфатный буфер, раствор Рингера и раствор декстрозы. Он может, кроме того, содержать другие материалы, желательные с точки зрения коммерции и потребителя, включая другие буферы, разбавители, фильтры, иглы и шприцы.
Далее следуют примеры способов и композиций по настоящему изобретению. Очевидно, могут осуществляться многочисленные другие варианты осуществления, дающиеся общим описанием, приведенным выше.
Примеры
Пример 1. Продуцирование и характеризация моноклональных антител к EGFL7
Продуцирование моноклональных антител
EGFL7 идентифицируют и клонируют в попытке обнаружения новых секретируемых и трансмебранных белков человека, в частности тех, которые вовлечены в регулирование развития сосудов. Подробности клонирования и экспрессирования человека EGFL7 описываются, например, в заявке на патент США US2003/0224948A1 (в которой EGFL7 идентифицируется как PRO1449). Код доступа GenBank® для EGFL7 человека представляет собой NM_016215.
Гомозиготные по EGFL7 нокаутированные мыши (получаемые в Genentech) иммунизируются с помощью продуцируемых E.coli рекомбинантных белков EGFL7 человека и мыши с тэгами His6, разбавленными в адъюванте Риби (Corixia, Hamilton, MT), дважды в неделю, через подошву лапы, восемь доз. Собирают B-лимфоциты из лимфатических узлов от десяти мышей, демонстрирующих высокие титры в сыворотке, а затем сливают с клетками миеломы мышей (X63.Ag8.653; American Type Culture Collection® (ATCC®)). Через 10-14 дней, супернатанты просматривают на продуцирование антител с помощью прямого ELISA, с использованием рекомбинантного белка EGFL7. Позитивы субклонируют дважды для получения моноклональности. Для широкомасштабного производства очищенного антитела, клетки гибридомы вводят как внутривенную инъекцию праймированным пристином мышам BALB/c или культивируют в биологическом реакторе Integra. Асцитные жидкости или супернатанты культур очищают с помощью аффинной хроматографии на белке A (Pharmacia Fast Protein Liquid Chromatography; Pharmacia, Uppsala, Sweden). Три антитела из этих моноклональных антител, обозначенные 4F11, 10G9 и 18F7, отбирают для полного анализа. Эти моноклональные антитела внесены в ATCC® в январе 2006 года.
Моноклональные антитела блокируют адгезию и миграцию клеток HUVEC
Ранее было известно, что EGFL7, нанесенные в виде покрытия на планшеты для культивирования ускоряют адгезию эндотелиальных клеток умбиликальной вены человека (HUVEC), хотя прочность адгезии значительно ниже, чем у других молекул с адгезией клеток, таких как фибронектин и коллаген (Parker et al., Nature 428:754-58 (2004)). Соответственно, авторы осуществили эксперименты, чтобы определить, могут ли Mab 4F11, 10G9 и 18F7 блокировать адгезию клеток на планшетах, покрытых EGFL7. Планшеты покрывают 5 мкг/мл фибронектина (Roche) и рекомбинантным EGFL7 человека, продуцируемым в E.coli. После промывок PBS, HUVEC (Cambrex) размещают на планшетах с плотностью 5Ч105/см2 в среде EGM2 (Cambrex) и центрифугируют в течение 5 мин при 140 g для синхронизации присоединения клеток, а затем инкубируют. Для анализа активности антител, HUVEG в среде EGM2 предварительно инкубируют при концентрациях антитела 0,5, 5 или 50 мкг/мл в 50 мМ Tris/125 мМ NaCl, pH 8,6 перед размещением на планшетах. Каждое из Mab 4F11, 10G9 и 18F7 блокирует адгезию клеток на планшетах, покрытых белком EGFL7 человека или мыши, зависимым от концентрации образом и ни одно из Mab не блокирует адгезию клеток на планшетах, покрытых фибронектином, подтверждая, что блокада является специфичной для EGFL7.
Авторы также проверили, могут ли эти антитела блокировать миграцию HUVEC на планшетах, покрытых EGFL7. Планшеты покрывают 5 мкг/мл одного из следующих белков: BSA (Sigma), коллагена (Upstate), фибронектина (Roche) и рекомбинантного EGFL7 человека (продуцируемого в E.coli на Genentech). После промывок PBS, HUVEC (Cambrex) размещают на планшетах при плотности 5x105/см2 в EGM2 (Cambrex). Клеткам дают возможность для присоединения в течение двух часов и монослой расцарапывают с помощью кончика пипетки. Лунки дважды промывают EGM2 и добавляют свежую среду, содержащую 50 мкг/мл контрольного mab, или 4F11, 10G9, 18F7 соответственно. Лунки фотографируют через несколько временных интервалов в течение 24 часов для мониторинга затягивания царапин. Каждое из Mab 4F11, 10G9 и 18F7 блокирует миграцию клеток на планшетах, покрытых белком EGFL7, но, не на планшетах, покрытых другими белками. Контрольное mab не имеет блокирующей активности по отношению ни к одному из белков. Эти результаты показывают, что все три mab анти-EGFL7 специфично блокируют миграцию HUVEC на EGFL7.
Интересно, что авторы обнаружили, что блокирование с помощью Mab 4F11 зависит от препарата. Конкретно, антитело является очень эффективным при блокировании адгезии клеток HUVEC, когда бульон с антителом приготавливают в 50 мМ Tris/125 мМ NaCl, но проявляет минимальную эффективность, когда его приготавливают в PBS. Это различие не наблюдается для двух других Mab.
Определение последовательности Mab 4F11 и 10G9
РНК в целом извлекают из клеток гибридомы, продуцирующих моноклональные антитела 4F11 и 10G9 анти-EGFL7 человека мыши, с использованием RNeasy® Mini Kit (Qiagen, Germany). Вариабельные легкие (VL) и вариабельные тяжелые (VH) домены 4F11 и 10g9 амплифицируют с использованием RT-PCR со следующими дегенеративными праймерами:
Для 4Fll:
Легкая цепь (LC), прямой: 5'-GTCAGATATCGTKCTSACMCARTCTCCWGC-3' (SEQ ID NO: 21)
Тяжелая цепь (HC), прямой: 5'-GATCGACGTACGCTCAGATHCARYTGGTGCARTCTGGGATCGACGTACGCTCAGATHCARYTGGTGCARTCTGG-3' (SEQ ID NO: 22)
Для 10G9:
Легкая цепь (LC), прямой: 5'-GATCGATATCGTGATGACBCARACTCCACT-3' (SEQ ID NO: 23)
Тяжелая цепь (HC), прямой: 5'-GATCGACGTACGCTGAGGTYCAGCTSCAGCAGTCTGG-3' (SEQ ID NO: 24)
Как для 4F11, так и для 10G9:
Легкая цепь, обратный: 5 '-TTTDAKYTCCAGCTTGGTACC-3' (SEQ ID NO: 25)
Тяжелая цепь, обратный: 5'-ACAGTGGGCCCTTGGTGGAGGCTGMRGAGACDGTGASHRDRGT-3' (SEQ ID NO: 26)
Прямые праймеры являются специфичными для N-концевых последовательностей аминокислот областей VL и VH обоих антител. Соответственно, обратные праймеры LC и HC конструируют для отжига до области в константном домене 1 легкой цепи (CL) и в константном домене 1 тяжелой цепи (CH1), который является идентичным для двух антител и очень консервативным среди видов. Амплифицированный VL клонируют в вектор экспрессии клеток pRK млекопитающих (Shields et al., J. Biol. Chem. 276:659-04 (2000)). Амплифицированный VH вставляют в вектор экспрессии клеток pRK млекопитающих. Последовательность полинуклеотидов вставок определяют с использованием рутинных способов секвенирования. Последовательности легких и тяжелых цепей 4F11 (SEQ ID NO: 1 и 2 соответственно) и легких и тяжелых цепей 10G9 (SEQ ID NO: 3 и 4 соответственно) показаны на фиг. 1-4.
Исследование изотипа и сродства связывания
Mab 4F11, 10G9, и 18F7 определяются как изотип IgG2b с использованием стандартных способов.
Сродство связывания Mab к EGFL7 как человека, так и мыши определяют с помощью поверхностного плазмонного резонанса, с использованием Pharmacia BIAcore® 3000 (BIAcore AB, Uppsala, Sweden) при комнатной температуре (см., например, Morton et al., Meth. Enzymol 295:268-94 (1998)). Антитела анти-EGFL7 иммобилизуют на сенсорном чипе (CM5) через первичные аминогруппы. Карбоксиметилированная матрица на поверхности сенсорного чипа активируется посредством инжектирования 20 мкл смеси 0,025 M N-гидроксисукцинимида и 0,1 M N-этил-N'-(диметиламинопропил)карбодиимида при 5 мкл/мин. 5-10 мкл раствора рекомбинантных белков EGFL7 человека или мыши, 10 мкг/мл, в 10 мМ ацетата натрия, pH 4,5, инжектируют при 5 мкл/мин. После связывания, незанятые сайты на чипе блокируют посредством инжекции 20 мкл 1M этаноламина, pH 8,5. Проточный буфер представляет собой PBS, содержащий 0,05% полисорбата 20. Для кинетических измерений двукратные последовательные разбавления EGFL7 с тэгом поли-His в проточном буфере инжектируют поверх потока клеток в течение 3 минут при скорости потока 30 мкл/мин, и связанные EGFL7 с тэгом поли-His получают возможность для диссоциации в течение 20 минут. Поверхностные связывания регенерируют посредством инжекции 20 мкл 10 мМ глицин·HCl (pH 1,5). Проточная ячейка один, которая активируется, но, не имеет иммобилизованного антитела, используется в качестве ячейки сравнения. Значительное неспецифичное связывание EGFL7 с тэгом поли-His в проточной ячейке один отсутствует. Для вычисления видимого сродства связывания, данные анализируют с использованием модели связывания 1:1 с использованием глобальной подгонки. Константы скорости ассоциации и диссоциации подгоняются одновременно (программное обеспечение BIAevaluation). Результаты этих экспериментов с использованием бульонов Mab в 50 мМ Tris/125 мМ NaCl показаны в Таблице 2.
KD Mab для EGFL7 человека и мыши
Определение эпитопа EGFL7, распознаваемого Mab
Авторы определили эпитоп, распознаваемый моноклональными антителами, авторы впервые картировали область EGFL7, связываемую каждым из Mab. Клетки 293 трансфицируют векторами экспрессии, содержащими либо полноразмерные EGFL7, либо усеченные формы белка, как показано на фиг.5. Анализы вестерн блот лизатов клеток из трансфицированных клеток осуществляли затем с помощью зондов Mab 4F11, 10G9 и 18F7. Авторы наблюдали, что каждое из Mab связывается с частью EMI EGFL7.
Для сужения конкретного эпитопа, распознаваемого каждым Mab, авторы синтезировали перекрывающиеся полипептиды, расширяющие часть домена EMI, и исследовали их способность к конкурированию с полноразмерным EGFL7 за связывание Mab. Например, последовательности полипептидов, используемых для Mab 4F11, являются следующими:
Авторы смешивали Mab 4F11 с белком EGFL7 и 10-кратным молярным избытком каждого из полипептидов p1, p2 и p3, иммунно преципитировали полученные комплексы и визуализировали их с помощью SDS-PAGE. Для Mab 4F11, авторы наблюдали, что только p2 конкурирует с полноразмерным EGFL7 за связывание с Mab 4F11. Результаты показывают, что Mab 4F11 распознает эпитоп EGFL7, который содержит последовательность CCP.
Авторы осуществляли сходные эксперименты с двумя другими Mab с использованием полипептидов, имеющих следующие последовательности:
Авторы определили, что Mab, как 10G9, так и 18F7, распознают эпитоп, который содержит последовательность RTIY (SEQ ID NO 33).
Пример 2. Mab анти-EGFL7 ингибирует рост опухоли in vivo
В этом примере, Mab анти-EGFL7 исследуют на их способность к ингибированию роста опухоли in vivo в нескольких моделях. Авторы сначала исследовали Mab в PBS на модели Colo205 (рак толстой и прямой кишки человека) и на модели A673 (модель рабдомиосаркомы человека). Авторы не наблюдали воздействия в этих моделях с Mab в PBS в качестве единственных агентов.
Затем авторы исследовали Mab сами по себе и/или в сочетании с антителом анти-VEGF, B20.4.1 (описывается в заявке WO 2005/012359). Авторы исследовали антитела на трех моделях: на модели рака груди человека Her2 ("модель Fo5"), на модели рака легких человека (NSCLC) ("H1299") и на другой модели рака груди человека ("MDA-MB231"). Эти модели опухолей хорошо установлены и описываются, например, в Lee et al., Clin Cancer Res. 11(16):6065-74 (2005); Cameron et al., Cancer Cell Int.5:23 (2005); Finkle et al., Clinical Cancer Res. 10:2499-251(2004). Каждое животное лечат Mab против пыльцы амброзии (контрольное); только B20.4; только 18F7; или B20.4 плюс 4F11, 10G9, или 18F7. Вкратце, для модели H1299, самкам мышей nu/nu HRLN делают инъекцию 1×107 клеток опухоли H1299, подкожно в бок; и для модели MDA-MB231, самкам мышей nu/nu HRLN делают инъекцию 5×106 клеток опухоли MDA-MB231 подкожно в бок. Для каждой модели, лечение антителом начинают, когда средний размер опухоли достигает 100 мм3 (соответствует дню 0 на фиг. 6-9). Контрольное Mab против пыльцы амброзии и B20.4 вводят при 10 мг/кг, раз в неделю и 4F11, 10G9 и 18F7 вводят при 10 мг/кг, дважды в неделю (показано стрелками под осью x на фиг. 6, 8 и 9).
Авторы не наблюдали воздействия на модели Fo5. Авторы наблюдают значительные ингибиторные воздействия на опухоли в других двух моделях. Как показано на фиг. 6-7, на модели H1299, Mab 4F11 в сочетании с B20.4.1 является значительно более эффективным, чем либо только контроль, либо только B20.4.1. Как показано на фиг.8, на модели MDA-MB231, либо Mab 4F11, либо Mab 10G9 в сочетании с B20.4.1 является более эффективным, чем либо только контроль, либо только B20.4.1. Как показано на фиг.9, на модели MDA-MB231, только Mab 18F7 является более эффективным, чем только контроль, и в сочетании с B20.4.1 является значительно более эффективным, чем либо только Mab 18F7, либо только B20.4.1.
Интересно, что авторы также обнаружили, что лечение с помощью Mab 4F11 на модели HI299 предотвращает полное восстановление сосудов после прекращения терапии анти-VEGF (B20.4.1). Когда сосудистые структуры опухолей сравниваются между опухолями, леченными только B20.4.1, и теми, которые лечатся B20.4.1 и 4F11, после прекращения лечения, авторы наблюдали значительные замедления реваскуляризации опухоли. Эти результаты четко доказывают, что терапия анти-EGFL7 может обеспечить дополнительную или даже синергическую эффективность, когда она объединяется с терапией анти-VEGF.
Авторы также использовали другие модели опухолей, доступные в данной области, для исследования противоопухолевых активностей антител анти-EGFL7. Они включают в себя, но, не ограничиваясь этим: LS174T (толстая кишка), BXPC3 (простата), HCT116 (толстая кишка), MV-522 (NSCLC), SKMES (NSCLC), Colon26 (толстая кишка), MDA-MB231 (грудь), MCF7 (грудь), H1299 (NSCLC), SW620 (толстая кишка), LL (легкие), Fo5 (грудь), 4T1 (грудь), HT29 (толстая кишка), SW480 (толстая кишка), 786-0 (надпочечники).
Следующие клетки гибридомы внесены в American Type Culture Collection®, PO Box 1549, Manassas, VA, 20108, USA (ATCC®):
Эти депозиты сделаются согласно условиям Budapest Treaty on International Recognition of Deposit of Microorganisms for the Purpose of Patent Procedure (Будапештский договор). Это обеспечивает поддержание жизнеспособного вклада в течение 30 лет от даты внесения. Эти линии клеток будут доступны с помощью ATCC на условиях Будапештского договора и по соглашению между Genentech, Inc. и ATCC, которое обеспечивает константную и неограниченную доступность линий клеток для публики при выдаче соответствующего патента США или при выкладывании для публики любой заявки на патент США или иностранный патент, в зависимости от того, что приходит первым, и обеспечивает доступность линий клеток тем субъектам, которые определяются U.S. Commissioner of Patents and Trademarks, и должны перечисляться в них в соответствии с 35 USC §122 и Commissioner's rules, соответствующими им (включая 37 CFR §1.14 с конкретной ссылкой на 886 OG 638).
Заявитель настоящей заявки согласен с тем, что если внесенные линии клеток будут потеряны или разрушены при культивировании при соответствующих условиях, они будут быстро заменены образцом такой же линии клеток. Доступность внесенных линий клеток не должна рассматриваться как разрешение для осуществления изобретения в противоположность правам, предоставляемым любым правительством в соответствии с его патентными законами.
Хотя указанное выше изобретение описывается в некоторых деталях для иллюстрации и примера, в целях ясности понимания, описания и примеры не должны рассматриваться как ограничивающие рамки настоящего изобретения.











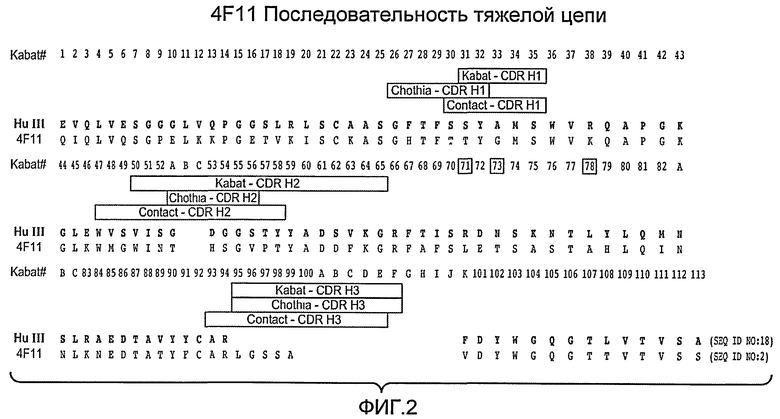

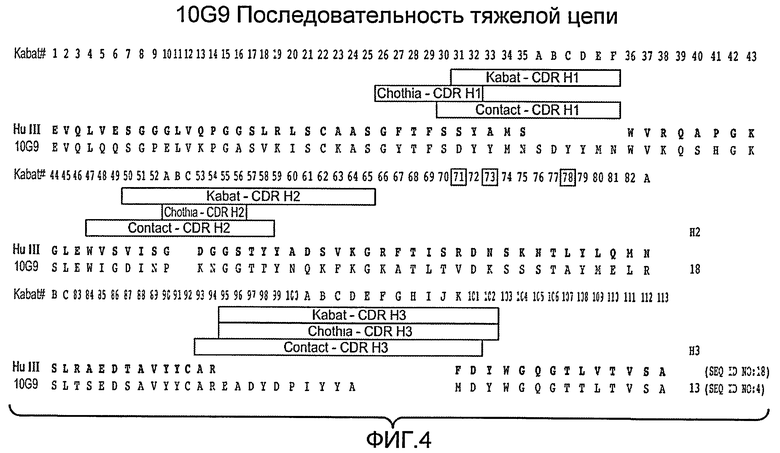
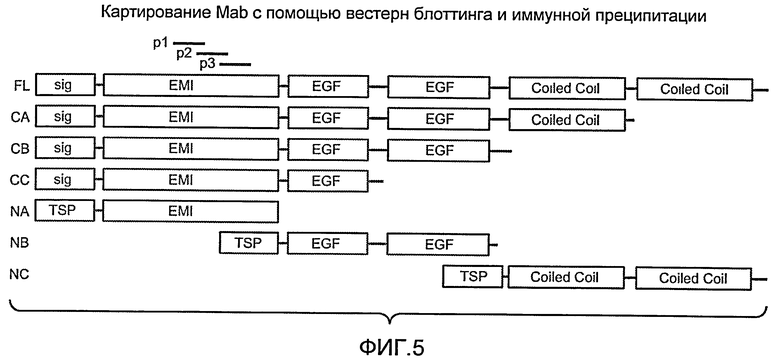
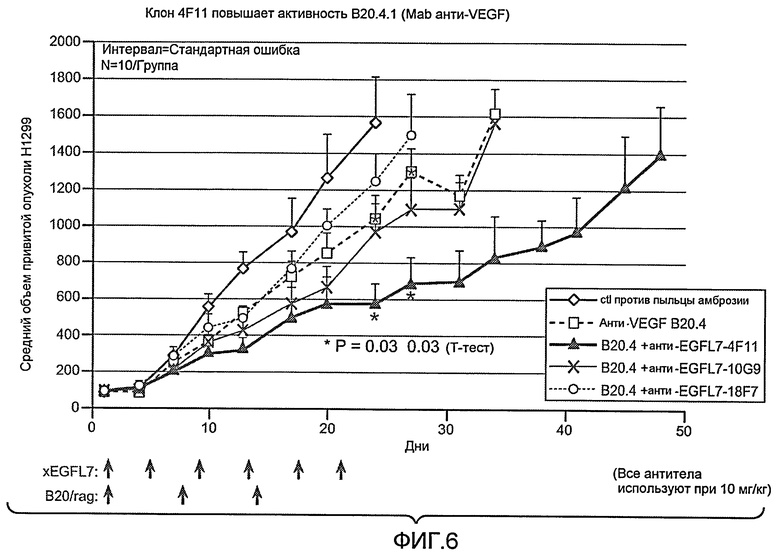
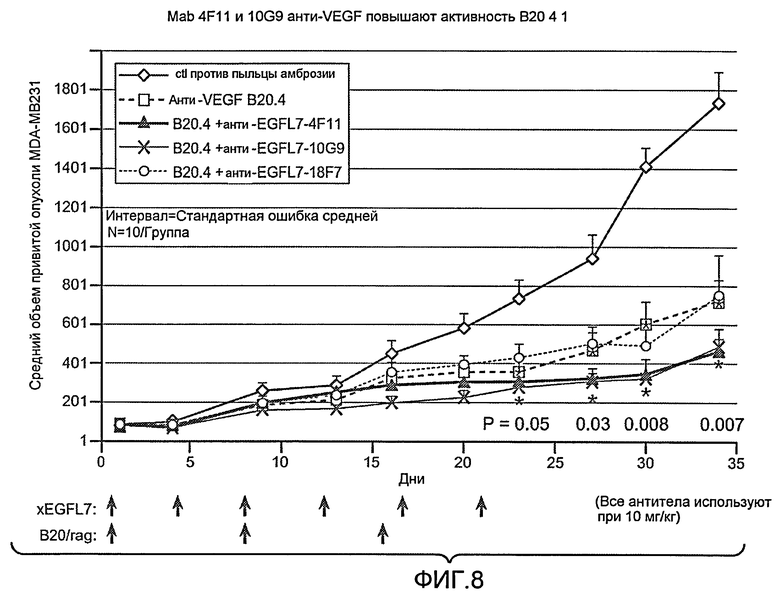

Изобретение относится к биотехнологии. Описано антитело к EGFL7. Предложена фармацевтическая композиция для модулирования развития сосудов, содержащая эффективное количество антитела к EGFL7. Представлены полинуклеотид, кодирующий антитело к EGFL7, вектор экспрессии, содержащий указанный полинуклеотид, клетка-хозяин, способная экспрессировать экзогенный генетический материал, содержащая указанный вектор. Описан способ уменьшения или ингибирования ангиогенеза у субъекта, имеющего патологическое состояние, связанное с ангиогенезом, включающий введение субъекту эффективного количества антитела к EGFL7 или фармацевтической композиции, содержащей указанное антитело. Предложен способ повышения эффективности антиангиогенного агента у субъекта, имеющего патологическое состояние, связанное с ангиогенезом, включающий введение субъекту антитела к EGFL7 или фармацевтической композиции, содержащей указанное антитело. Изобретение расширяет арсенал средств, пригодных для модулирования развития сосудов. 9 н. и 37 з.п. ф-лы, 9 ил, 2 табл.
1. Антитело к EGFL7, содержащее нижеследующие шесть областей, определяющих комплементарность (CDR):
(a) последовательность 4F11 CDR-L1 KASQSVDYDGDSYMS (SEQ ID NO:5);
(b) последовательность 4F11 CDR-L2 GASNLES (SEQ ID NO:6);
(c) последовательность 4F11 CDR-L3 QQNNEDPYT (SEQ ID NO:7);
(d) последовательность 4F11 CDR-H1 TYGMS (SEQ ID NO:8);
(e) последовательность 4F11 CDR-H2 WINTHSGVPTYADDFKG (SEQ ID NO:9) и
(f) последовательность 4F11 CDR-H3 LGSSA (SEQ ID NO:10).
2. Антитело к EGFL7 по п.1, где легкая цепь указанного антитела содержит последовательность:
DIVLTQSPASLAVSLGQRATISCKASQSVDYDGDSYMSWYQQKPGQPPKLLIYGASNLESGIPA
RFSGSGSGTDFTLNIHPVEEEDAATYYCQQNNEDFYTFGGGTKVEIKR (SEQ ID NO:1).
3. Антитело к EGFL7 по п.1, где тяжелая цепь указанного антитела содержит последовательность:
QIQLVQSGPELKKPGETVKISCKASGHTFTTYGMSWVKQAPGKGLKWMGWTNTHSGVPTYADDF
KGRFAFSLETSASTAHLQINNLKNEDTATYFCARLGSSAVDYWGQGTTVTVSS (SEQ ID NO:2).
4. Антитело к EGFL7, содержащее нижеследующие шесть областей, определяющих комплементарность (CDR):
(a) последовательность 10G9 CDR-L1 RSSQSLVHTNGITYLH (SEQ ID NO: 11);
(b) последовательность 10G9 CDR-L2 KVSNRFS (SEQ ID NO:12);
(c) последовательность 10G9 CDR-L3 SQSTHVPLT (SEQ ID NO:13);
(d) последовательность 10G9 CDR-H1 DYYMNSDYYMN (SEQ ID NO:14);
(e) последовательность 10G9 CDR-H2 DINPKNGGTTYNQKFKG (SEQ ID NO: 15) и
(f) последовательность 10G9 CDR-H3 ALGVFDY (SEQ ID NO:16).
5. Антитело к EGFL7 по п.4, где легкая цепь указанного антитела содержит последовательность:
DIVMTQTPLSLPVSLGDQASISCRSSQSLVHTNGITYLHWYLQKPGQSPKLLTYKVSNRFSGVP
DRFSGSGSGTDFTLKISRVEAEDLGVYFCSQSTHVPLTFGAGTKVEIKR (SEQ ID NO:3).
6. Антитело к EGFL7 по п.4, где тяжелая цепь указанного антитела содержит последовательность:
EVQLQQSGPELVKPGASVKISCKASGYTFSDYYMNSDYYMNWVKQSHGKSLEWIGDINPKNGGT
TYNQKFKGKATLTVDKSSSTAYMELRSLTSEDSAVYYCAREADYDPIYYAMDYWGQGTTLTVSA (SEQID NO:4).
7. Антитело по п.1 или 4, где антитело представляет собой моноклональное антитело.
8. Антитело по п.1 или 4, где антитело выбирается из группы, состоящей из химерного антитела, гуманизированного антитела, антитела с созревшим сродством, антитела человека и биспецифичного антитела.
9. Антитело по п.1 или 4, где антитело представляет собой фрагмент антитела.
10. Фармацевтическая композиция для модулирования развития сосудов, содержащая эффективное количество антитела к EGFL7 по любому из пп.1-9.
11. Фармацевтическая композиция по п.10, дополнительно содержащая антиангиогенный агент.
12. Фармацевтическая композиция по п.11, где антиангиогенный агент выбирается из группы, состоящей из бевацизумаба и ранибизумаба.
13. Полинуклеотид, кодирующий антитело по любому из пп.1-9.
14. Вектор экспрессии, содержащий полинуклеотид по п.13.
15. Клетка-хозяин, способная экспрессировать экзогенный генетический материал, содержащая вектор по п.14.
16. Клетка-хозяин по п.15, где клетка-хозяин представляет собой прокариотическую клетку.
17. Клетка-хозяин по п.15, где клетка-хозяин представляет собой эукариотическую клетку.
18. Клетка-хозяин по п.15, где клетка-хозяин представляет собой клетку млекопитающего.
19. Способ получения антитела к EGFL7, где указанный способ включает
(a) экспрессирование вектора по п.14 в соответствующей клетке-хозяине и
(b) извлечение антитела.
20. Способ по п.19, где клетка-хозяин представляет собой прокариотическую клетку.
21. Способ по п.19, где клетка-хозяин представляет собой эукариотическую клетку.
22. Способ уменьшения или ингибирования ангиогенеза у субъекта, имеющего патологическое состояние, связанное с ангиогенезом, включающий введение субъекту эффективного количества антитела к EGFL7 по любому из пп.1-9 или фармацевтической композиции по п.10.
23. Способ по п.22, где патологическое состояние представляет собой неоплазму.
24. Способ по п.23, где неоплазма представляет собой карциному.
25. Способ по п.22, где патологическое состояние связано с глазами.
26. Способ по п.25, где патологическое состояние представляет собой интраокулярное неоваскулярное заболевание.
27. Способ по любому из пп.22-24, дополнительно включающий введение субъекту антиангиогенного агента.
28. Способ по п.27, где антиангиогенный агент представляет собой антагонист сосудистого эндотелиального фактора роста (VEGF).
29. Способ по п.28, где антагонист представляет собой антитело к VEGF.
30. Способ по п.29, где антитело к VEGF представляет собой бевацизумаб.
31. Способ по п.25 или 26, дополнительно включающий введение субъекту антиангиогенного агента.
32. Способ по п.31, где антиангиогенный агент представляет собой антагонист сосудистого эндотелиального фактора роста (VEGF).
33. Способ по п.32, где антагонист представляет собой антитело к VEGF.
34. Способ по п.33, где антитело к VEGF представляет собой ранибизумаб.
35. Способ по п.27, где антиангиогенный агент вводится до или после введения антитела к EGFL7.
36. Способ по п.27, где антиангиогенный агент вводится одновременно с антителом к EGFL7.
37. Способ повышения эффективности антиангиогенного агента у субъекта, имеющего патологическое состояние, связанное с ангиогенезом, включающий введение субъекту антитела по любому из пп.1-9 или фармацевтической композиции по п.10 или 11.
38. Способ по п.27, где патологическое состояние представляет собой неоплазму.
39. Способ по п.28, где неоплазма представляет собой карциному.
40. Способ по п.37, где антиангиогенный агент представляет собой бевацизумаб.
41. Способ по любому из пп.37-40, дополнительно включающий в себя введение химиотерапевтического агента.
42. Способ по п.37, где патологическое состояние связано с глазами.
43. Способ по п.42, где патологическое состояние представляет собой интраокулярное неоваскулярное заболевание.
44. Способ по п.42 или 43, где антиангиогенный агент представляет собой ранибизумаб.
45. Способ по п.42, дополнительно включающий в себя введение кортикостероида.
46. Способ по п.42, дополнительно включающий в себя введение фотодинамической терапии.
| WO 2005117968 А2, 15.12.2005 | |||
| РЫБАЛЬСКИЙ Н.Г., СЕРОВА М | |||
| А., ИГНАТЬЕВА Г.А., СТАРЧЕУС А.П | |||
| «Моноклональные антитела и гибридомы» | |||
| - М.: ВАСХНИЛ, 1989. |

